
Dopamine receptor sub-types involvement in nucleus accumbens and ventral tegmentum but not in medial...
9
Behavioural Brain Research 86 (1997) 171 – 179 Dopamine receptor sub-types involvement in nucleus accumbens and ventral tegmentum but not in medial prefrontal cortex: on self-stimulation of lateral hypothalamus and ventral mesencephalon Jitendra Singh, T. Desiraju, T.R. Raju * Department of Neurophysiology, National Institute of Mental Health and Neuro Sciences, Bangalore -560 029, India Accepted 26 November 1996 Abstract The involvement of different sub-types of dopamine receptors in the electrical self-stimulation behaviour was investigated using DA receptor subtype specific agonists viz R( +) SKF 38393 and LY 171555 (quinipirole). Rats were chronically implanted with bipolar electrodes in lateral hypothalamus-medial forebrain bundles (LH-MFB) and ventral tegmental area-substantia nigra (VTA-SN) or guide cannula in nucleus accumbens (nACB) or medial prefrontal cortex (mPFRCx) or cannula-cum-electrode in VTA-SN. Combining these, it was possible to inject a given receptor ligand in either nACB or mPFRCx or VTA-SN and assess the changes in intracranial self-stimulation (ICSS) of LH-MFB or VTA-SN. The experimental subjects were trained to press a pedal to obtain 0.25 s trains of cathodal rectangular pulses of 0.1 ms pulse duration (for LH-MFB electrode) or 0.3 ms pulse duration (for VTA-SN electrode). The rate-frequency curve for each subject and each placement were obtained in order to determine M 50 (50% of maximum asymptotic response rate). Microinjection of SKF 38393 (a D 1 agonist) or LY 171555 (a D 2 agonist) into nACB caused facilitatory and suppressive effects on the ICSS of VTA-SN and LH-MFB respectively. On the contrary, intra VTA-SN injection of 10 m g/0.5 m l of SKF 38393 or of LY 171555, reduced its ICSS to half as compared to vehicle injections. Under the effect of these injections, there was no change in the ICSS of LH-MFB. The combined administration of SKF 38393 (5 m g) and LY 171555(10 m g) into nACB produced suppression of ICSS of VTA-SN to greater extent than the LY 171555 (10 m g/0.5 m l). This indictes that coactivation of D 1 and D 2 leads to suppression of ICSS of VTA-SN. The activation of D 1 receptors in nACB facilitates the ICSS of both VTA-SN and LH-MFB by increasing the reinforcement and/locomotor activity, while the activation of D 2 receptors in it has opposite effect. The coactivation of D 1 and D 2 receptors in VTA-SN leads to the suppression of ICSS of both VTA-SN and LH-MFB. Similar injections of these receptor ligands into mPFRCx did not alter the ICSS of either LH-MFB or VTA-SN. The pre- or post-synaptic DA receptors of mPFRCx do not appear to influence the ICSS of either VTA-SN or LH-MFB in any significant manner. © 1997 Elsevier Science B.V. Keywords: Dopamine receptor subtypes D 1 ,D 2 and D 3 ; Nucleus accumbens; Medial prefrontal cortex; Lateral hypothalamus; Ventral mesencephalon; Intracranial self-stimulation 1. Introduction Considerable evidences have been documented from several lines of approaches including anatomical, neu- rophysiological, neurochemical, psychopharmacological and behavioural studies that dopaminergic neurotrans- mission plays a critical role in reinforcement/motivation process [8,41,49]. Abbre6iations: ICSS, Intracranial self-stimulation; LH-MFB, Lateral hypothalamus-medial forebrain bundle; mPFRCx, Medial prefrontal cortex; nACB, Nucleus accumbens; VTA-SN, Ventral teg- mental area-substantia nigra. * Corresponding author. Tel.: +91 80 6642121, ext.207; fax: +91 8 6631830. 0166-4328/97/$17.00 © 1997 Elsevier Science B.V. All rights reserved. PII S0166-4328(96)02263-2
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Dopamine receptor sub-types involvement in nucleus accumbens and ventral tegmentum but not in medial...

Behavioural Brain Research 86 (1997) 171–179
Dopamine receptor sub-types involvement in nucleus accumbens andventral tegmentum but not in medial prefrontal cortex: on
self-stimulation of lateral hypothalamus and ventral mesencephalon
Jitendra Singh, T. Desiraju, T.R. Raju *
Department of Neurophysiology, National Institute of Mental Health and Neuro Sciences, Bangalore-560 029, India
Accepted 26 November 1996
Abstract
The involvement of different sub-types of dopamine receptors in the electrical self-stimulation behaviour was investigated usingDA receptor subtype specific agonists viz R(+ ) SKF 38393 and LY 171555 (quinipirole). Rats were chronically implanted withbipolar electrodes in lateral hypothalamus-medial forebrain bundles (LH-MFB) and ventral tegmental area-substantia nigra(VTA-SN) or guide cannula in nucleus accumbens (nACB) or medial prefrontal cortex (mPFRCx) or cannula-cum-electrode inVTA-SN. Combining these, it was possible to inject a given receptor ligand in either nACB or mPFRCx or VTA-SN and assessthe changes in intracranial self-stimulation (ICSS) of LH-MFB or VTA-SN. The experimental subjects were trained to press apedal to obtain 0.25 s trains of cathodal rectangular pulses of 0.1 ms pulse duration (for LH-MFB electrode) or 0.3 ms pulseduration (for VTA-SN electrode). The rate-frequency curve for each subject and each placement were obtained in order todetermine M50 (50% of maximum asymptotic response rate). Microinjection of SKF 38393 (a D1 agonist) or LY 171555 (a D2
agonist) into nACB caused facilitatory and suppressive effects on the ICSS of VTA-SN and LH-MFB respectively. On thecontrary, intra VTA-SN injection of 10 mg/0.5 m l of SKF 38393 or of LY 171555, reduced its ICSS to half as compared to vehicleinjections. Under the effect of these injections, there was no change in the ICSS of LH-MFB. The combined administration ofSKF 38393 (5 mg) and LY 171555(10 mg) into nACB produced suppression of ICSS of VTA-SN to greater extent than the LY171555 (10 mg/0.5 m l). This indictes that coactivation of D1 and D2 leads to suppression of ICSS of VTA-SN. The activation ofD1 receptors in nACB facilitates the ICSS of both VTA-SN and LH-MFB by increasing the reinforcement and/locomotor activity,while the activation of D2 receptors in it has opposite effect. The coactivation of D1 and D2 receptors in VTA-SN leads to thesuppression of ICSS of both VTA-SN and LH-MFB. Similar injections of these receptor ligands into mPFRCx did not alter theICSS of either LH-MFB or VTA-SN. The pre- or post-synaptic DA receptors of mPFRCx do not appear to influence the ICSSof either VTA-SN or LH-MFB in any significant manner. © 1997 Elsevier Science B.V.
Keywords: Dopamine receptor subtypes D1, D2 and D3; Nucleus accumbens; Medial prefrontal cortex; Lateral hypothalamus;Ventral mesencephalon; Intracranial self-stimulation
1. Introduction
Considerable evidences have been documented fromseveral lines of approaches including anatomical, neu-rophysiological, neurochemical, psychopharmacologicaland behavioural studies that dopaminergic neurotrans-mission plays a critical role in reinforcement/motivationprocess [8,41,49].
Abbre6iations: ICSS, Intracranial self-stimulation; LH-MFB,Lateral hypothalamus-medial forebrain bundle; mPFRCx, Medialprefrontal cortex; nACB, Nucleus accumbens; VTA-SN, Ventral teg-mental area-substantia nigra.
* Corresponding author. Tel.: +91 80 6642121, ext.207; fax: +918 6631830.
0166-4328/97/$17.00 © 1997 Elsevier Science B.V. All rights reserved.PII S 0166 -4328 (96 )02263 -2

J. Singh et al. / Beha6ioural Brain Research 86 (1997) 171–179172
Table 1
A–P (in mm referring to D–V (in mm fromM–L (in mm referring to saggitalBrain regionline)Bregma) dura)
7.290.3, (30°1.3–1.61.5–2.0Nucleus accumbens (nACB)inclination)
0.8–1.2 3.0–3.590.2Medial prefrontal cortex (mPFRCx) 3.2–3.78.490.2−1.8–−3.3Lateral hypothalamus–medial forebrain bundle 1.6–2.0
(LH-MFB)8.690.21.1–1.8−4.8–−6.3Ventral tegmental area–substantia nigra (VTA-SN)
There were three types of placement combinations:1. nACB guide cannula; ipsilateral LH-MFB bipolar electorde and ipsilateral VTA-SN bipolar electrode (n=47).2. mPFRCx guide cannula; ipsilateral LH-MFB bipolar electrode and ipsiateral VTA-SN bipolar electrode (n=9).3. LH-MFB bipolar electrode and ipsilateral VTA-SN cannula-cum-electrode (n=14).
The nucleus accumbens (nACB) [15] and medial pre-frontal cortex (mPRFCx) [22,23,35,38] terminal sites ofmesolimbic and mesocortical projections respectivelyhave been shown to evoke intracranial self-stimulation(ICSS), besides the ventral tegmental area (VTA)-sub-stantia nigra (SN) and interconnecting medial forebrainbundle (MFB) lateral hypothalamus (LH) [19,49].
Increased turnover of dopamine (DA) has been ob-served in both the nACB and caudate-putamen (CPu)after stimulation at the level of LH [11] or VTA [45].Using in vivo microdialysis methodology, it has beenshown that DA levels and their metabolites 3,4-dihy-droxyphenylacetic acid (DOPAC) and homovanillicacid (HVA) increase in both ipsilateral and contralat-eral mPFRCx and nACB to greater extent but in CPuto lesser extent produced by ICSS of MFB innomifensin pre-treated rats [26,27,32,33]. Complimen-tary to these findings, intra-nACB injections of am-phetamine enhance the reward [2] and conversely,injections of neuroleptics into it attenuate reward [39] inreward versus performance measurement method. Elec-trophysiological recordings have demonstrated thatneurons in the nACB exhibit vigrous activation inresponse to electrical self-stimulation of ipsilateralMFB at the level of VTA [50]. DA transmission in thenACB also plays an important role in the self-adminis-tration of amphetamine, cocaine, nicotine, opiates andethanol [4,16,49].
D1 receptors as compared to D2 receptors have beenimplicated in the ICSS of mPFRCx [6,34]. The correla-tion of the D2 receptors mediation in LH has beenpredicted [10]. Contrary to it, the involvement of D1
receptors in the modulation of the ICSS of LH has alsobeen demonstrated [13,17]. The systemic administrationof mixed D1/D2 agonist (apomorphine) or specific D1
receptor agonist (SKF 38393) or D2 receptor agonist(LY 171555) on ICSS has yielded inconsistent resultsthat do not conform to a coherent scheme [9,13,28,29].
In order to resolve the inconsistencies of the effectsof DA agonists on ICSS, this study was designed,
employing intracerebral microinjections into eithernACB or mPFRCx or VTA-SN, since it tends to avoidthe global actions on multiple systems. For this pur-pose, relatively DA receptor subtype specific agonists,R(+ ) SKF 38393 (D1\D3\D2) and LY 171555(D3\D2 and D1) were used.
2. Materials and methods
2.1. Experimental subjects
Wistar adult male rats weighing in the range of260–320 g that were about 5–6 1/2 months old at thetime of surgery/implantation, were used. The implantedanimals were housed individually in polypropylenecages (35.5×22.5×15 cm) in a temperature and hu-midity controlled room with access to 18% caesin–wheat–lipids–vitamins–minerals semi-synthetic dietand water ad libitum except during the experimentalsessions.
2.2. Surgery/implantation, beha6ioural testingprocedure and histology
The procedure of surgery/implantation, behaviouraltesting and histology is as followed in our previouspublication [37]. In each rat, following standardstereotaxic procedures, the chronically indwelling bipo-lar electrode, guide cannula or cannula–cum–electrodewere implanted. The bipolar electrode was made from apair of insulated stainless steel wires 220–250 mm andwere fitted to the miniplastic collar. The guide cannulawas made of 24 gauge stainless tubing. The cannulaguide tubing–curn–electrode was made from 24 gaugestainless steel tubing and wire of 30 gauge. These wereinsulated and juxtaposed. The miniature metal collarwas soldered to the guide cannula for inserting andcoupling injector needle at the time of microinjectionwith least restraining. The pre-determined flat skull

J. Singh et al. / Beha6ioural Brain Research 86 (1997) 171–179 173
stereotaxic co-ordinates were adopted from Paxinosand Watson rat atlas (1982) [31] (Table 1).
Each rat was trained to press the pedal for ICSS toobtain reliable, sustainable and reproducible seizure–free rates of ICSS pedal press responses from therespective sites on a continuous reinforcement schedule(CRF=1:1). The stimulus obtained by the animal dur-ing each pedal pressing consisted of 0.25 s train ofcathodal rectangular pulses, 0.1 ms pulse duration (forLH-MFB placements), 0.3 ms pulse duration (for SN-VTA placements). The response/reinforement data wasrecorded on an automated digital counter. The be-havioural criterion was minimum of 60 pedal pressresponses/mm for LH-MFB and minimum of 90 pedalpress responses/min for VTA-SN at optimal currentintensity and frequency. An optimal current was deter-mined as that yielded the criterion rate of respondingwithout any signs of aversiness i.e. retreat from pedal,vocalisation, jerking, etc.
2.3. Rate-frequency functions
To generate a rate-frequency curve with at least sixpoints, out of stimulation frequencies i.e. 10, 25, 40, 50,60, 100, 125, 160 and 200 Hz at the current intensity asdetermined with 100 Hz to obtain criterion rate wererepeated for five sessions, so that animals could dis-criminate accurately changes in frequency of stimula-tion. The data for each frequency point was obtainedfor 4 min, out of which the ICSS score of the first (1)and last (4) min were discarded and mean of mid (2)min score was taken to express response rate/min at agiven frequency.
2.4. Determination of M50 (50% of the maximumasymptotic response rate)
The current intensity and frequency was adjusted tosuch a level so as to obtain 50% of the maximumasymptotic pedal pressing rate designated as (M50). Themean (9S.E.M) stimulation frequency (Hz) for M50,LH-MFB and VTA-SN were 51.994.09 and 61.895.27 respectively. The mean (9S.E.M) pedal pressingrates/mm for LH-MFB and VTA-SN were 41.4395.72and 57.9694.58 respectively (n=24). The representa-tive rate-frequency curve for LH-MFB and VTA-SN(Fig. 1).
2.5. Microinjection
Microinjection was made by replacing the inner styletwith a 30 gauge injection needle which protruded 0.5mm beyond the outer guide cannula. The injectionneedle was connected via polythene tubing to a 5.0 m lHamilton microsyringe. A volume of 0.5 m l was in-jected slowly into each site in a period of about 1 min
in the unrestrained rat without causing stress. Afterwaiting for an additional 1 min to allow diffusion, theinjection needle was slowly removed and the styletreplaced. The whole microinjection procedure tookabout 5 min per subject.
2.6. Statistical analysis
The results were analysed using ‘F ’ ratio to ascertainthe homogenity of variance and if there were no statis-tical difference between the variance then accordinglycomparisons of means of vehicle (control) and drug(experimental) and also before the drug microinjectionwas made and after it on the alternate site employing‘matched paired’ t-test (2-tailed). Two-way ANOVARwas used to find the effect of different doses on the siteof ICSS.
2.7. Materials
R(+ )SKF 38393 and LY 171555 (Quinpirole-hy-drochloride) were obtained from Smith Kline and Lilly,respectively as gift. These were dissolved in isotonicphysiological saline (0.9%)
3. Results
3.1. Microinjections into nucleus accumbens
3.1.1. Dose-response functionAt M50 criterion, the dose-response functions were
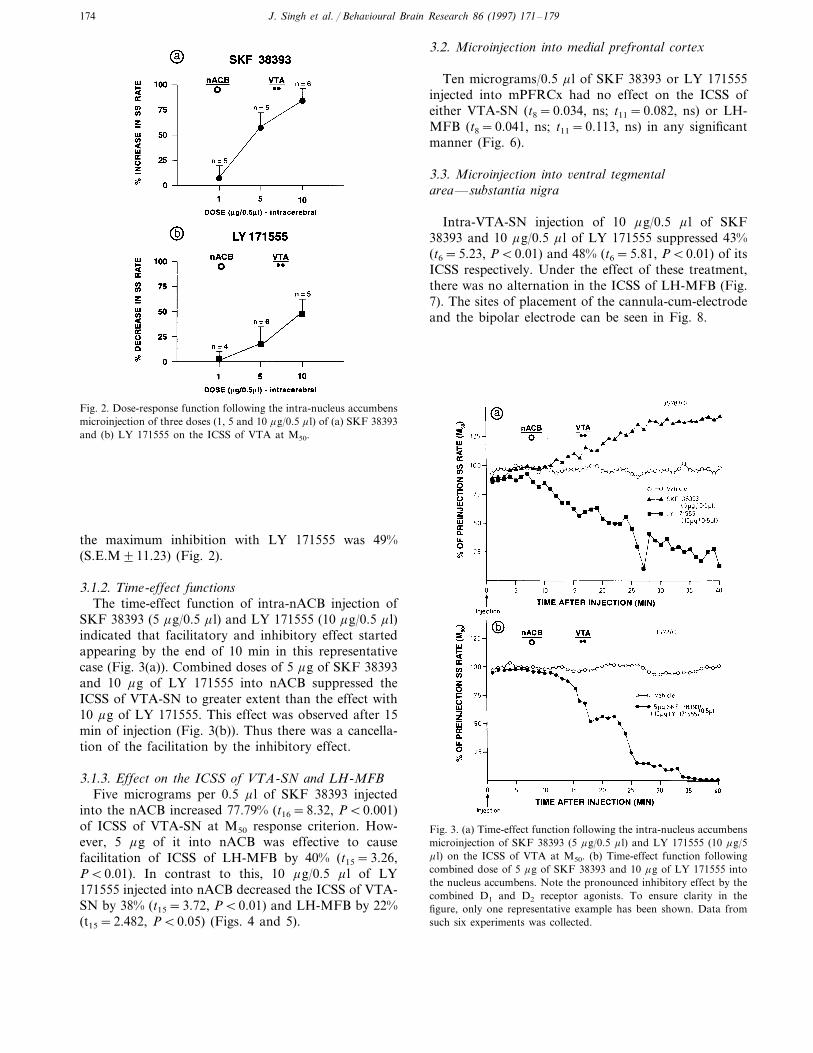
obtained with 1, 5 and 10 mg/0.5 m l of either SKF38393 or LY 171555. SKF 38393 showed dose-depen-dent increase in VTA-ICSS rates. The maximum facili-tation with SKF 38393 was 82.3% (S.E.M96.83) and
Fig. 1. A representative example of the rate-frequency curve todetermine M50 (50% of the maximum asymptotic response rate) forthe ICSS of LH-MFB and VTA-SN. Note that the locus of rise(LOR) and maximum asymptotic response rate (Max) are differentfor these sites of self-stimulation.
J. Singh et al. / Beha6ioural Brain Research 86 (1997) 171–179174
Fig. 2. Dose-response function following the intra-nucleus accumbensmicroinjection of three doses (1, 5 and 10 mg/0.5 m l) of (a) SKF 38393and (b) LY 171555 on the ICSS of VTA at M50.
3.2. Microinjection into medial prefrontal cortex
Ten micrograms/0.5 m l of SKF 38393 or LY 171555injected into mPFRCx had no effect on the ICSS ofeither VTA-SN (t8=0.034, ns; t11=0.082, ns) or LH-MFB (t8=0.041, ns; t11=0.113, ns) in any significantmanner (Fig. 6).
3.3. Microinjection into 6entral tegmentalarea—substantia nigra
Intra-VTA-SN injection of 10 mg/0.5 m l of SKF38393 and 10 mg/0.5 m l of LY 171555 suppressed 43%(t6=5.23, PB0.01) and 48% (t6=5.81, PB0.01) of itsICSS respectively. Under the effect of these treatment,there was no alternation in the ICSS of LH-MFB (Fig.7). The sites of placement of the cannula-cum-electrodeand the bipolar electrode can be seen in Fig. 8.
Fig. 3. (a) Time-effect function following the intra-nucleus accumbensmicroinjection of SKF 38393 (5 mg/0.5 m l) and LY 171555 (10 mg/5m l) on the ICSS of VTA at M50. (b) Time-effect function followingcombined dose of 5 mg of SKF 38393 and 10 mg of LY 171555 intothe nucleus accumbens. Note the pronounced inhibitory effect by thecombined D1 and D2 receptor agonists. To ensure clarity in thefigure, only one representative example has been shown. Data fromsuch six experiments was collected.
the maximum inhibition with LY 171555 was 49%(S.E.M911.23) (Fig. 2).
3.1.2. Time-effect functionsThe time-effect function of intra-nACB injection of
SKF 38393 (5 mg/0.5 m l) and LY 171555 (10 mg/0.5 m l)indicated that facilitatory and inhibitory effect startedappearing by the end of 10 min in this representativecase (Fig. 3(a)). Combined doses of 5 mg of SKF 38393and 10 mg of LY 171555 into nACB suppressed theICSS of VTA-SN to greater extent than the effect with10 mg of LY 171555. This effect was observed after 15min of injection (Fig. 3(b)). Thus there was a cancella-tion of the facilitation by the inhibitory effect.
3.1.3. Effect on the ICSS of VTA-SN and LH-MFBFive micrograms per 0.5 m l of SKF 38393 injected
into the nACB increased 77.79% (t16=8.32, PB0.001)of ICSS of VTA-SN at M50 response criterion. How-ever, 5 mg of it into nACB was effective to causefacilitation of ICSS of LH-MFB by 40% (t15=3.26,PB0.01). In contrast to this, 10 mg/0.5 m l of LY171555 injected into nACB decreased the ICSS of VTA-SN by 38% (t15=3.72, PB0.01) and LH-MFB by 22%(t15=2.482, PB0.05) (Figs. 4 and 5).

J. Singh et al. / Beha6ioural Brain Research 86 (1997) 171–179 175
Fig. 4. Cannula sites on the coronal section histological plates ofPaxinos and Watson’s rat atlas (1982). Sites for intra-nucleus accum-bens microinjections of SKF 38393 and LY 171555, for experimentsof Fig. 4. The number in the circle represents the number of animalsand (x) indicates where the injections were not effective to produceany significant change in response rates.
[14,25]. In contrast to it, 6-OHDA (6-hydroxy do-pamine) lesions of nACB attenuate ICSS of LH [39]and VTA [7] and self-administration of amphetamineand cocaine [16] but not of opiates [49]. Excitotoxinlesion studies point in favour of the catecholaminergicfibres for the maintanence of MFB self-stimulation [40]against the local intrinsic neurons in LH [44] and VTA[42].
There are at least five sub-types of DA receptorsbased on their molecular configuration [12]. However,these receptor sub-types can also be separated into twocategories based on their relationship with the linkageof adenylate cyclase. In this categorisation, D1-likereceptors (D1 and D5) are those that are coupled tovariety of signal mechanisms, the activation of whicheither has no effect or inhibition of production of
Fig. 5. The effect of intra-nucleus accumbens microinjection of (a)SKF 38393 (5 mg/0.5 m l) and (b) LY 177555 (10 mg/0.5 m l) on theICSS of VTA and LH at M50 as compared to vehicle injection. Theschedule of microinjection of the vehicle and drug and behaviouraltesting session was as follows: Vehicle: injection of vehicle wasfollowed by waiting for 10 min; then testing of ICSS responses at M50
from site of stimulation for 15 min= (V); after this a gap of 30 minwas given for rest before drug injection. Drug: injection of the drugwas followed by waiting for pre-determined time interval based ontime–effect pattern as observed in preliminary experiments. Testingof ICSS responses at M50 from the site of stimulation for 15 min=(D). (�) represents the cannula and (��) indicates bipolar electrode.*=PB0.05; **=PB0.01 ***=PB0.001.
4. Discussion
The microinjection of SKF 38393 into nACB causedthe increase in the ICSS of VTA-SN as well as LH-MFB at the M50, whereas, a similar injection of it intoVTA-SN decreased its ICSS without changing the ICSSof LH-MFB. In contrast to it, the injections of LY171555 into either nACB or VTA-SN caused the reduc-tion in the ICSS of VTA. No such effects were observedwhen similar injections of it were administered intomPFRCx. These results are consistent with previousreports on the effects of injections of D1/D2agonist [17]or amphetamine [2,30] or neuroleptics [5,20,21,39] intonACB and no effect of these injections into mPFRCxon the ICSS of VTA and MFB.
Lesion studies are not consistent with respect to therole of terminal sites of meso-cortico-limbic pathwaysin self-stimulation. It was argued that this system maynot be necessary for ICSS, as lesions of VTA appear tospare ICSS derived from nACB and mPFRCx [35].Coronal knife-cuts in the rostral MFB-LH at the levelof anterior commissure and lateral preoptic area todisrupt the descending outflow of diagonal band andmedial septal regions had little permanent effect on theICSS of LH [47]. However, anterolateral and largeMFB transections behind the lateral preoptic area wereeffective in degrading the LH/VTA stimulation reward

J. Singh et al. / Beha6ioural Brain Research 86 (1997) 171–179176
Fig. 6. Sites for intra-medial prefrontal cortex injections. Only data ofseven rats has been shown. The placement of one rat could not beverified.
could not resolve how DA agonists influence ICSS [9].D1 and D2 receptors mediation in the ICSS of LH andVTA has also been demonstrated [10,13,17]. D1 recep-tor antagonist-SCH 23390 or D2 receptor antagonist-spiperone decreased fixed- interval (FI) self-stimulation[29]. SKF 38393, a D1 receptor agonist inhibited FIself-stimulation, in reward summation model [13], butquinpirole, a D2 receptor agonist facilitated self-stimu-lation in rate-frequency paradigm [28]. The suppressiveeffect of SCH 23390 on the ICSS of VTA has also beendemonstrated by intra-nACB injections of it in rate-in-tensity paradigm [17]. Thus the effect of D1 receptorantagonist on ICSS was not opposite to the effect of D1
agonist. However, D2 receptor agonist and antagonist
Fig. 7. Effect of intra-ventral tegmental area-substantia nigra mi-croinjection of SKF 38393 (10 mg/0.5 m l) and LY 17155Ss (10 mg/0.5m l) on the ICSS of VTA-SN and LH-MFB at M50. The schedule ofmicroinjection and behavioural testing session: Vehicle: injection ofvehicle was followed by waiting for 10 min; the testing of ICSSresponses at M50 from the injection site was carried out for 15min= (V). This was followed by another waiting for 10 min. Thentesting of ICSS responses from another site for 15 min= (B). Afterthis, a gap of 30 min was given for rest before drug injection. Drug:injection of the drug was followed by waiting for pre-determined timeintervals based on time-effect pattern; Testing of ICSS responses atM50 from the injection site was carried out for 15 min= (D). A gapof 10 min intervals, testing of ICSS responses from another site for 15min= (A). Animals were tested with the vehicle and one dose of drugon a single day. In a few cases, the testing was done on thenon-injection site after 10 min of microinjection to avoid the timedelay. Drug tests were separated by at least 48 hours. (��) representscannula-cum-electrode, the rest of the notations are as in Fig. 4.
cAMP or phosphatidylinositol turnover or opening ofK+ channel causing hyperpolarization [43]. The postsynaptic D1 receptors are the larger population that arelinked to DA sensitive adeylate cyclase via Gs (stimulat-ing G protein) [12]. D2 receptors that are linked toadeylate cyclase via Gs (inhibiting G protein) have beenreported either postsynaptic or presynaptic autorecep-tors on the dendrites and soma of DA neurons [24].The presynaptic autoreceptors are impulse regulating inaction or modulating Ca2+-dependent DA release [43].D3 receptors (G-protein coupled) seem to be postsynap-tic and may not be associated with cAMP [12]. Postsy-naptic D1 receptors and presynaptic D2 autoreceptorspredominate in mPFRCx [12,34]. Besides the presenceof D1 and D2 receptors, densities of D3 receptor seem tobe highest in nACB but more abundant in hypothala-mus than in SN [12]. Further, evidence seems to befavouring the view that D1 receptors are distributedpredominantly on axon terminals afferenting onto SN[1], whereas D2 receptors on neurons in SN-VTA [24].Intracellular electrophysiological studies show that DAis inhibitory to DA as well as non-DA neurons in SNand VTA [18]. D1 receptor blockade disables postsy-naptic D2 receptor function, it does so, however, with-out secondary effects on autoreceptor function [46].
A biphasic dose response i.e. increase in response atlower doses and decrease in response at higher doseswas demonstrated by systemic administration of non-selective DA agonist (apomorphine), but this study

J. Singh et al. / Beha6ioural Brain Research 86 (1997) 171–179 177
Fig. 8. Sites of placement of cannula-cum-electrode and bipolar electrode (n=14) for the experiments of Fig. 7.
facilitated and inhibited ICSS respectively, showingsymmetrical effects [29].
The co-activation of both D1 and D2 receptors ap-pears essential for the expression of behaviours involv-ing motor functions [46,48]. The synergistic interactionof D1 and D2 receptors has been proposed in the sameneuronal structure at the cellular level. It could not beresolved whether the locus of this interaction in ICSS isin the nACB or VTA or SN [17,28]. The cancellation offacilitatory effect of D1 receptor agonist by inhibitoryeffect of D2 receptor agonist, following combined ad-ministration of D1 and D2 agonist, lend support for thesynergistic interaction in the nACB. The combinedadministration of apomorphine, an D1/D2 mixed ago-nist and haloperidol, a D2 antagonist into VTA-SNfailed to show any cancellation effect on the ICSS ofVTA-SN [36]. Since both D1 and D2 agonist intoVTA-SN showed similar inhibitory effect on its ICSS,
such synergistic interaction in VTA-SN cannot be in-ferred.
A discrimination between the change in the reward-ing effect of electrical stimulation and a change in theperformance involving motor execution becomesdifficult in many experimental paradigms of ICSS [19].Moreover interpretations also vary as DA agonist mayprovide reward which is not contigent on pedal pressingand therefore, lower response rate by extinguishing theoperant task. Intra VTA-SN injections of SKF 38393(D1\D3\D2) or LY 171555 (D3\D2\D1) sup-pressed its ICSS, but the ICSS of LH-MFB remainedunaffected. These results can be interpreted to infer thatthe observed effects were due to the change in rewardefficacy but not due to the inability of the animal toexecute motor functions. The activation of D1 receptorsin nACB facilitates the ICSS of both VTA-SN andLH-MFB, by increasing the reinforcement and/locomo-

J. Singh et al. / Beha6ioural Brain Research 86 (1997) 171–179178
tor activity. In contrast to it, the activation of D2
receptor in nACB has opposite effect, as this leads tothe inhibition of ICSS of both VTA-SN and LH-MFB.
Systemic administration or intra-mPFRCx micro in-jections of SCH 23390 a D1 agonist produced dose-de-pendent decrease in the ICSS of mPFRCx [34].Contrary to it, systemic administration of cis-flup-enthixol, a mixed D1/D2 antagonist affected the ICSSof MFB, but not that of ICSS of mPFRCx [3]. Thus,the dopaminergic mechanisms of mPFRCx, eitherthrough D1 or D2/D3 receptors do not exert any modu-latory effect on the ICSS of VTA-SN and LH-MFB.
These findings taken together, can be interpreted tosuggest that the meso-limbic link (VTA-nACB), in con-trast to meso-cortico link (VTA-mPFRCx) appears tobe critical for inducing and blocking reinforcement. Itcan be further inferred that the ICSS of a given brainregion may be modulated by the activation of differentreceptor types and sub-types either at the same site oras some other linked region. It is also possible thatreceptors at the site of stimulation and some otherinterconnected region generate a set of conditions in theneural circuitry leading to the rewarding effect [36,37].
Acknowledgements
The financial support of the Indian Council of Medi-cal Research, New Delhi is gratefully acknowledged.We thank Smith Kline French Laboratories and LillyResearch Laboratories, USA for the supply of R(+ )SKF 38393 and LY 171555 respectively.
References
[1] Altar, C.A. and Hauser, K., Topography of substantia nigrainnervation by D1-receptor containing striatal neurons, BrainRes., 410 (1987) 1–11.
[2] Colle, L.M. and Wise, R.A., Effects of nucleus accumbensamphetamine on lateral hypothalamic brain stimulation reward,Brain Res., 459 (1988) 361–368.
[3] Corbett, D., Differences in sensitivity to neuroleptic blockade:medial forebrain bundle versus frontal cortex self-stimulation,Beha6. Brain Res., 36 (1990) 91–96.
[4] DiChiara, G. and Imperato, A., Drugs abused by humans pref-erentially increasing synaptic dopamine concentrations in themesolimbic systems of freely moving rats, Proc. Natl. Acad. Sci.USA., 85 (1988) 5274–5278
[5] DuVauchella, C.L., Levitin, M., MacConell, L.A. and Lee, L.K.Ettenberg, A. Oposite effects of prefrontal cortex and nucleusaccumbens infusions of flupenthixol on stimulation-induced lo-comation and brain stimulation, Brain Res., 576 (1992) 104–110.
[6] Ferrer, J.M.R., Sanguinetti, A.M., Vives, F. and Mora, F.,Effects of agonists and antagonists of D1 and D2 dopaminereceptors on self-stimulation of the medial prefrontal cortex inthe rat, Pharmacol. Biochem. Beha6., 19 (1983) 211–217.
[7] Fibiger, H.C., LePiane, F.G., Jakubovic, A. and Phillips, A.G.,The role of dopamine in intracranial self-stimulation of theventral tegmental area, J. Neurosci., 7 (1987) 3888–3896.
[8] Fibiger, H.C. and Phillips, A.G., Role of Catecholamine trans-mitters in brain reward systems: implications for the neurobiol-ogy of affect. In J. Engel and L. Oreland (Eds.) Brain rewardsystem and abuse, 1987, Raven, New York, pp. 61–74.
[9] Fouriezos, G. and Francis, S., Apomorphine and electrical self-stimulation of rat brain, Beha6. Brain Res., 52 (1992) 73–80.
[10] Gallistel, C.R. and Freyd, G., Quantitative determination of theeffects of catecholaminergic agonist and antagonist on rewardingefficacy of brain stimulation, Pharmacol. Biochem. Beha6., 26(1987) 731–742.
[11] Garrigues, A.M. and Cazala, P., Central catecholaminemetabolism and hypothalamic self-stimulation behaviour in twoinbred strains of mice, Brain Res., 265 (1983) 265–271.
[12] Gingrich, J.A. and Caron, M.G., Recent advances in the molec-ular biology of dopamine receptors, Annu. Re6. Neurosci., 16(1993) 299–321.
[13] Hunt, G.E., Atrens, D.M. and Jackson, D.M., Reward summa-tion and the effects of dopaine D1 and D2 agonists and antago-nists on fixed interval responding for brain stimulation,Pharmacol. Bechem. Beha6., 48 (1994) 853–862.
[14] Janas, J. and Stellar, J.R., Effects of knife-cut lesions on themedial forebrain bundle in self-stimulation rats, Beha6. Neu-rosci., 101 (1987) 832–845.
[15] Jenkins, O.F., Atrens, D.M. and Jackson, D.M., Self-stimulationof the nucleus accumbens and some comparisons with hypotha-lamic self- stimulation, Pharmacol. Biochem. Beha6., 18 (1983)585–591.
[16] Koob, G.F. and Bloom, F.E., Cellular and molecular mecha-nisms of drug dependence, Science 242 (1988) 715–723.
[17] Kurumiya, S. and Nakajima, S., Dopamine D1 receptors in thenucleus accumbens: involvement in the reinforcing effect oftegmental stimulation, Brain Res., 448 (1988) 1–6.
[18] Lacey, M.G. Mercuri, N.B. and North, R.A., Two cell types inrat substantia nigra zona compacta distinguished by membraneproperties and the actions of dopamine and opioids. J. Neurosci.,9 (1989) 1233–1241.
[19] Liebman, J.M., Drug effects on behaviours maintained by elec-trical brain stimulation. In A.A. Boulten, G.B. Baker and A.J.Greenshaw (Eds.) Neuromethods V. 13. Psychopharmacology,The Humana Press, Chifton, N.J. 1989, pp. 447–511.
[20] Mogenson, G.J., Takigawa, M., Robertson, A. and Wu, M.,Self-stimulation of the nucleus accumbens and ventral tegmentalarea of Tsai attenuated by microinjections of spiroperidol intothe nucleus accumbens, Brain Res., 171 (t979) 247–259.
[21] Mora, F. Myers, R.D. and Sanguinetti, A.M., Self-stimulation ofthe MFB or VTA after microinjection of haloperidol into theprefrontal cortex of the rat, Pharmacol. Biochem. Beha6., 6(1977) 239–241.
[22] Mora, F. and Cobo, M., The neurobiological basis of prefrontalcortex self-stimulation. A review and an integrative hypothesis,Prog. Brain Res., 85 (1990) 419–431.
[23] Mora, F. and Ferrer, J.M.R., Neurotransmitters, pathways andcircuits as the neural substrates on self-stimulation of the pre-frontal cortex facts and speculations, Beha6. Brain Res., 22(1986) 127–140.
[24] Morelli, M., Mennin, T. and DiChiara, G., Nigral dopamineautoreceptors are exclusively of the D2 type: quantitative autora-diography of (125I) iodosulpiride and (125I) SCH 23982 in adja-cent brain sections, Neuroscience, 27 (1988) 865–870.
[25] Murray, B. and Shizgal P., Anterolateral lesions of the medialforebrain bundle increase the frequency threshold for self-stimu-lation of the lateral hypothalamus and ventral tegmental area inthe rat, Psychobiology 19 (1990) 135–146.
[26] Nakahara, D., Fuchikami, K., Ozaki, N., Iwasaki, T. and Na-gatsu, T., Differential effect of self-stimulation on dopaminerelase and metabolism in the rat medial frontal cortex, nucleusaccumbens and striatum studied by in vivo microdialysis, BrainRes., 574 (1992) 164–170.

J. Singh et al. / Beha6ioural Brain Research 86 (1997) 171–179 179
[27] Nakahara, D., Ozaki, N., Miura, Y., Miura, H. and Nagatsu, T.,Increased dopamine and serotonin metabolism in rat nucleusaccumbens produced by intracranial self-stimulation of medialforebrain bundle as measured by in vivo microdialysis, BrainRes., 495 (1989) 178–181.
[28] Nakajima, S., Liu, X. and Lau, C.L., Synergistic interaction ofD1 and D2 dopamine receptors in the modulation of reinforcingeffect of brain stimulation, Beha6. Neurosci., 107 (1993) 161–165.
[29] Nakajima, S. and O’Regan, N.B., The effects of dopaminergicagonists and antagonists and the frequency-response function forhypothalamic self-stimulation in the rat, Pharmacol. Biochem.Beha6., 39 (1991) 465–468.
[30] Olds, M.E., Enhanced dopamine receptor activation in accum-bens and frontal cortex has opposite effects on medial forebrainbundle, Neuroscience, 35 (1990) 313–325.
[31] Paxinos, G. and Watson, C., The rat brain in stereotaxic co-ordi-nates, Academic press, New York, 1982.
[32] Philips, A.G., Coury, A., Fiorino, D., LePiane, F.G., Brown, E.and Fibiger, H.C., Self-stimulation of the vental tegmental areaenhances dopamine release in the nucleus accumbens: a microdi-alysis study, Ann. N.Y. Acad. Sci., 654 (1992) 199–206.
[33] Phillips, A.G., Jakubovic, A. and Fibiger, H.C., Increased invivo tyrosine hyroxylase activity in rat telencehalon produced byself-stimulation of the ventral tegmental area, Brain Res., 402(1987) 109–116.
[34] Sabater, R., Saez, J.A. and Ferrer, J.M.R., SCH 23390 decreasesself-stimulation of the medial prefrontal cortex in the rat, Eur. J.Phamacol., 242 (1993) 205–208.
[35] Simon, H., Stinus, L., Tassin, J.P., Lavielle, S., Blanc G.,Thierry, A.M., Glowinski, J. and Lemoral, M., Is the dopamin-ergic mesocorticolimbic system necessary for intracranial self-stimulation?, Beha6. Neural Biol., 27 (1979) 125–145.
[36] Singh, J. and Desiraju, T., Differential involvement of dopamin-ergic and noradrenergic receptor types of lateral hypothalamusand ventral midbrain in self-stimulation evoked from these re-gions. In B.N. Dhawan, R.C. Srimal, R. Raghubir and R.S.Rapaka (Eds.) Recent ad6ances in the study of neurotransmitterreceptors, Drug research perspectives: CDRI series, Vol. 17,1994, Central Drug Research Institute, Lucknow, pp. 71–112.
[37] Singh, Desiraju, T., Nagaraja, T.N. and Raju, T.R., Facilitationof self-stimulation of ventral tegmentum by microinjection ofopioid receptor subtypes, Physiol. Beha6., 55 (1994) 627–631.
[38] Spence, S.J., Silverman, J.A., Corbett, D., Cortical and ventraltegmental systems expert opposing influences on self-stimulationfrom the prefrontal cortex, Beha6. Brain Res., 17 (1985) 117–124.
[39] Stellar J.R. and Corbett, D., Regional neuroleptic microinjectionindicate a role for nucleus accumbens in lateral hypothalamicself-stimulation reward, Brain Res., 477 (1989) 126–143.
[40] Stellar, J.R., Hall, F.S. and Waraczynski, M., The effects ofexcitotoxin lesions of the lateral hypothalamus on self-stimula-tion reward, Brain Res., 541 (1991) 29–40.
[41] Snyder, S.H., The dopamine connections. Nature (London) 347(1996) 121–122.
[42] Takeichi, T., Kurumiya S., Umemto, M. and Olds, M.E., Roleof catecholaminergic terminals and intrinsic neurons of the ven-tral tegmentam in self-stimulation investigated in neonatallydopamine-depleted rats, Pharmacol. Biochem. Beha6., 24 (1985)1101–1109.
[43] Vallar, L. and Meldolesi, J., Mechanisms of signal transductionat the dopamine D2 receptor, TIPS, 10 (1989) 74–77.
[44] Valley, L., Verney, C., Kemp, E. and Berger, B., Opposite effectsof ibotenic acid and 6-hydroxy dopamine lesions of the lateralhypothalamus on intracranial self-stimulation and stimulationinduced locomotion, Neuroscience., 25 (1988) 595–603.
[45] Van Heuven-Nolson, D., Van Wolfswinkel, L., Van Ree, J.M..and Versteeg, D.H.G., Electrical stimulation of the ventral teg-mental area and catecholamine metabolism in discrete regions ofthe rat brain, Brain Res., 268 (1983) 362–366.
[46] Walters, J.R., Bergstrom, D.A., Carlson, J.H., Chase, T.H. andBraun, A.R., D1 dopamine receptor activation required forpost-synaptic expression of D2 agonist effects, Science 236 (1987)719–722.
[47] Waraczynski, M.A., Basal forebrain knife-cuts and medial fore-brain bundle self-stimulation, Brain Res., 438 (1989) 8–22.
[48] Weick, B.G. and Walters, J.R., Effects of D1 and D2 dopaminereceptor stimulation on the activity of substantia nigra parsreticulata neurons in 6-hydroxy dopamine lesioned rats: D1/D2
coactivation induces potentiated responses, Brain Res., 405(1987) 234–246.
[49] Wise, R.A. and Rompre, P.-P., Brain dopamine and reward,Annu. Re6. Psychol., 40 (1989) 191–225.
[50] Wolske, M., Rompie, P.-P., Wise, R.A. and West, M.O., Activa-tion of single neurons in the rat nucleus accumbens duringself-stimulation of the ventral tegmental area, J. Neurosci., 13(1993) 1–12.
.
.




















