
Compressive Strength Forecasting of Air-Entrained ... - MDPI
Upload
independentCategory
view
0download
0
Behavioural Brain Research 158 (2005) 133–142
Research report
Differential role of the accumbens Shell and Core subterritoriesin food-entrained rhythms of rats
Jorge Mendoza, Manuel Angeles-Castellanos, Carolina Escobar∗
Departamento de Anatom´ıa, Facultad de Medicina, Edificio “B” 4◦ Piso,Universidad Nacional Aut´onoma de M´exico, Mexico DF 04510, M´exico
Received 27 May 2004; received in revised form 12 August 2004; accepted 25 August 2004Available online 30 September 2004
Abstract
Restricted feeding schedules (RFS) entrain behavioral and physiological rhythms even in suprachiasmatic nucleus ablated animals, suggest-ing the existence of a food-entrained oscillator. The nucleus accumbens is an important structure for the expression of motivational behaviorsand because its anatomical subterritories, Shell (Acc-Sh) and Core (Acc-Co) establish connections with different functional systems, theym rainmentu differentiale tty acids( d-entrainedr ting meal-t ticipatorya data indi-c le betweenb©
K
1
dattaaocoe
thealtionswithr the
dulestheyvi-vity,chesns
sandtence
0d
ay participate in a differential way in food-entrainment. A first experiment, explored the role of Acc-Sh and Acc-Co in food-entsing the immunohistochemical detection of the protein c-Fos as a transcriptional activation marker. Experiment 2 tested theffect of Acc-Sh and Acc-Co, NMDA excitotoxic lesions. Lesioned rats were entrained to RFS and locomotor activity and free faFFA) concentrations were evaluated. Results data show that in the Acc-Sh there is an increase of c-Fos immunoreactivity in fooats principally during feeding, whereas c-Fos expression in the Acc-Co region was increased during feeding and also anticipaime. FFA were entrained in both lesioned groups, but the basal level was lower in Core-lesion rats. All rats exhibited food anctivity (FAA). However, FAA was increased in Shell-lesioned animals and was almost abolished in the Core-lesion rats. Theseate that the accumbens nucleus is involved with behavioral and metabolic food-entrainment, and that there is a differential rooth subregions.2004 Elsevier B.V. All rights reserved.
eywords:Circadian; Food-anticipatory activity; Motivational; c-Fos; Free fatty acids
. Introduction
Circadian rhythms in behavior and physiology are un-er the control of internal biological clocks. The suprachi-smatic nucleus (SCN) of the hypothalamus in mammals is
he master circadian clock which is entrained principally byhe light–dark cycle[18]. In the SCN, a set of clock genesnd their protein products are driven by interlocking positivend negative feedback loops in a circadian way to generatescillations[28]. Likewise, several peripheral tissues exhibitircadian oscillations of clock genes[2,3,6,13,37]. Ablationsf the SCN render an animal arrhythmic[25,31], and alsoliminate the oscillations of clock genes in peripheral oscil-
∗ Corresponding author. Tel.: +52 55 5623 2422; fax: +52 55 5623 2425.E-mail address:[email protected] (C. Escobar).
lators, indicating that peripheral oscillators depend onintegrity of the SCN[3,29]. However, this is controversibecause recently it was reported that clock gene oscillain peripheral tissues persist up to 20 cycles in animalsSCN lesions, suggesting that SCN is not necessary foperipheral tissues oscillations[38].
When animals are exposed to restricted feeding sche(RFS), allowing them to feed for a few hours every day,develop food anticipatory activity (FAA) starting 2–3 h preous to mealtime. FAA can be observed in locomotor actiwheel running, drinking behavior and food cup approa[24,32]. In addition, physiological and metabolic functioare entrained by food availability[10,12,19]. SCN lesiondo not prevent entrainment of behavioral, physiologicalmolecular parameters by daily RFS, suggesting the exisof a food-entrainable oscillator (FEO)[8,15,23,32]. Diverse
166-4328/$ – see front matter © 2004 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2004.08.016

134 J. Mendoza et al. / Behavioural Brain Research 158 (2005) 133–142
studies have tried to identify the anatomical and functionalsubstrate of FEO by producing lesions in several nuclei inthe central nervous system (CNS)[22,23,32], however, theselesions do not prevent FAA. Only lesions of the parabraquialnucleus (PBN) attenuated FAA, probably because it is a relaystructure that mediates the entraining signal from the periph-ery to the CNS[9]. Expression ofmPer1andmPer2in thecerebral cortex and hippocampus, and c-Fos in the hypotha-lamus are in phase with FAA, suggesting the possibility thatthese regions are involved in the behavioral manifestationof FEO[4,36]. For the study of the anatomical substrate ofFEO, main attention has been focused in neural structuresinvolved with energy balance and regulation of hunger andsatiation. Although in FAA there is an evident increase of themotivational state, the mechanisms for increased motivationin the expression of food-entrained circadian rhythms havebeen poorly studied. There is evidence that restricted feedingproviding a palatable diet entrains locomotor activity in ham-sters, suggesting that the hedonic value of food is importantfor entrainment[1,21].
In the brain, the nucleus accumbens has been consideredan interface between limbic and motor systems and is rel-evant for motivated behaviors and reward-related processes[24]. The nucleus accumbens is innervated by dopaminergicneurons from the ventral tegmental area (VTA) and ablationo otora to alc bensi -Sh)w bensCE icals e dif-f ingb ot s tot thala-m sys-t blest hi erentp uress (LH)[ ele-v
othA cesso thee ter-m ood-e ren-t eenb le oft d cir-c as a
marker of transcriptional activity. In a second experiment weevaluated behavioral and metabolic entrainment in rats withspecific excitotoxic lesions of either one of the accumbenssubregions.
2. Materials and methods
2.1. Animals and housing
Male Wistar rats weighing 230–250 g at the beginning ofthe experiment were housed in groups of five individuals intransparent acrylic cages (40 cm× 50 cm × 20 cm). Ratswere maintained in a 12:12 h light–dark cycle (lights on at0700 designated as zeitgeber time ZT0) and constant temper-ature (22± 1◦C) with free access to food (Rodent LaboratoryChow 5001, IL, USA) and water unless otherwise stated. Ex-periments were approved by the national guide of care anduse of animal experimentation of Mexican laws (Decreto deLey de Proteccion a los Animales del Distrito Federal, GacetaOficial del DF, 26/02/02).
2.2. Experiment 1
2.2.1. Experimental designoth
A ran-d trolg eeksw ghtp ex-p d atd (= val-u thm,a ersis-t thena d for3 essw ivedf thel s thef
2nto-
b lines %p PB;p eredp ces-s PB.F ta ainedw 72 ha ch-
f dopaminergic neurons results in complex sensoriomlterations and hypophagia, which has been attributed
ack of reward associated with food ingestion[30,35]. Ac-ording to its anatomical projections, the nucleus accums subdivided in two regions, the accumbens Shell (Acchich represents the ventromedial part and the accumore (Acc-Co) which represents the dorsolateral area[16].vidence of pharmacological, anatomical and physiologtudies suggests that the Acc-Sh and Acc-Co regions arerentially involved in specific behavioral states, like feedehavior and motor activity[5,20]. The Acc-Co projects t
he dorso-lateral ventral pallidum, which in turn projecthe motor system through the substantia nigra and subic nucleus, and is linked to the extrapiramidal motor
em. The pattern of its efferent connections closely resemhose of the caudate-putamen[16]. In contrast, the Acc-Ss suggested to be more limbic in nature, because its effrojections are closely connected to limbic output structuch as the VTA, amygdala and lateral hypothalamus16]. This later relation between the Acc-Sh and LH is rant for the control of feeding behavior[33,34].
In view of the anatomical and functional relations, bcc-Sh and Acc-Co regions may be involved in the prof food-entrainment but may participate differentially inxpression of FAA. The aim of this study was first to deine whether the nucleus accumbens is involved with fntrainment, and second to determine if there is a diffe
ial role in the regulation of food–circadian rhythms betwoth Acc-Sh and Acc-Co regions. We examined the ro
he nucleus accumbens subterritories in food-entraineadian rhythms with c-Fos immunoreactivity (c-Fos-IR)
To evaluate the c-Fos-IR during food-entrainment in bcc-Sh and Acc-Co subregions, animals were assignedomly to an ad libitum feeding condition (AL) as a conroup, or to a restricted feeding schedule (RF) for 3 with food access only for 2 h daily at the middle of the lihase from ZT6-ZT8 (1300–1500 h). On the last day oferimental manipulation, animals were randomly sacrifieifferent time points ZT4, ZT5, ZT6, ZT8, ZT12, ZT14n5 per group and time) for immunohistochemistry. To eate the persistence of the food-entrained c-Fos-IR rhynother set of rats was assigned to two groups. The p
ence group (RF-F) was entrained to RFS for 3 weeks,llowed free food access for 5 days and left food deprivedays. In the control fasting group (AL-F) free food accas available for 3 weeks and then was left food depr
or 3 days. For both groups, animals were perfused onast day of fasting conditions at the same time points aood-entrained animals.
.2.2. ImmunohistochemistryAnimals were deeply anaesthetized with sodium pe
arbital and were perfused transcardially with 250 ml saolution (0.9%) followed immediately by 250 ml of 4araformaldehyde solution in 0.1 M phosphate buffer (H 7.2). Brains were removed and post-fixed in the buffaraformaldehyde solution for 1 h, then transferred sucively to 10, 20 and 30% sucrose solutions in 0.1 Mrozen coronal sections of 40�m were cut with a cryostand collected in four series. One series of tissue was stith cresyl violet, and another set was incubated duringt 4◦C in c-Fos antibody made in rabbit (Santa Cruz Biote

J. Mendoza et al. / Behavioural Brain Research 158 (2005) 133–142 135
nology, USA) diluted 1:2500 in PB saline goat serum triton(PBSGT; PBS, 1% goat serum and 0.3% Triton X-100). Sec-tions were rinsed three times for 10 min in phosphate buffersaline (PBS), then were incubated in biotinylated secondaryantibody (goat anti-rabbit; Vector laboratories) 1:200 in PB-SGT for 2 h. The tissues were rinsed again and then were incu-bated in avidin–biotin–peroxidase complex (Vectastain EliteABC Kit; Vector Labs) for 2 h and washed again three timesfor 10 min in PBS. The tissue was reacted in diaminobenzi-dine (0.5 mg/ml, in trizma buffer 7.2) with hydrogen peroxide(35�l, 30% H2O2). Sections were mounted in gelatin-coatedslides, intensified with osmium tetroxide 0.1% for 30 s anddehydrated through a series of alcohol, cleared with xylene,and coverslipped with Permount (Fisher Chemicals).
2.2.3. Quantification of cellsThe number of c-Fos-positive nuclei were counted unilat-
erally in matched coronal sections from all groups. Sectionswere selected in accordance with the stereotaxic atlas fromPaxinos and Watson (Acc-Co, Interaural 10.20 mm; Acc-ShInteraural 10.60 mm)[27]. Images of the corresponding cre-syl violet sections were acquired to define the area of eachregion of the nucleus. Images were examined under a lightmicroscope (Nikon Eclipse E600) and captured at a 10×mag-nification using a computerized image analysis system (MetaV 108s gea gridw ves,b arbyr d ors eredp dis-c s cellsp
2
2ps:
S . Form daysa ZT4,Z
ss tof inedt att aysi e ofF her1
2ate
( axici e
induced by infusing 0.4�l of NMDA per side (N-methyl-d-aspartate, Sigma; 10 mg/ml solution in PB 0.1 M, pH 7.4)with the following stereotaxic coordinates: Shell, AP +1.6, L±0.4, DV−7.0; Core, AP +1.2, L±1.8, DV−7.2[27]. Eachlesioned group had a sham operated control which received0.4�l of PB (0.1M, pH 7.4) instead of NMDA.
2.3.3. Body weight and food intakeBody weight of all animals was registered before the
surgery and during ad libitum conditions before starting RFS.In addition, to verify possible effects of the lesion on foodingestion, food intake for a 24 h interval was monitored oncefor each animal under ad libitum conditions.
2.3.4. Determination of free fatty acidsOn day 15 of food-entrainment, animals were cannu-
lated in the internal jugular vein with a polyethylene silicontube (0.025 in. i.d. and 0.047 in. o.d.; Silastic Brand) filledwith heparine (500 U/ml) as anti-coagulant. Blood samples(600�l) were collected in Eppendorff (1.8 ml) tubes con-taining a clot-activator gel and were centrifuged at 2500 rpmduring 15 min. Aliquots of 100�l were processed with spec-trophotometric methods for determination of free fatty acids(FFA) by using a colorimetric method as previously described[
2ere
b rentp d-p tantt terb wasc o thel mati-c le-p tivityp ac-t sisw ientoy lab-o
2ere
k sedi mlo pro-t weref onsw ainedw entw m-p
ue series 4.5, Universal imaging corporation). A grid ofquares each 40�m× 40�m was superimposed on the imand immunopositive cells for c-Fos included under theere counted manually. To avoid counting false positiackground optical density (OD) was established in a neegion lacking c-Fos-IR, only stained cells that reacheurpassed three fold the OD background were considositive, whereas cells under this staining threshold werearded. Cell number was transformed and is expressed aer square millimeter.
.3. Experiment 2
.3.1. Experimental designAnimals were randomly assigned to one of four grou
hell-lesion, Shell-sham, Core-lesion and Core-shametabolic analysis animals were entrained to RFS for 17nd samples were obtained at several time points: ZT3,T5, ZT6, ZT8 and ZT9 (mealtime at ZT6).
For behavioral analysis, the four groups had free acceood for 15 days (ad libitum), on day 16 they were entrao RFS for 3 weeks with food access only for 2 h dailyhe middle of the light phase (ZT6–ZT8), followed by 3 dn total food deprivation in order to evaluate persistencAA. Finally, rats were left with free food access for ot5 days.
.3.2. SurgeryAnimals were anaesthetized with chloral hydr
10 mg/kg body weight) and secured in a Kopf stereotnstrument with the incisor bar set at−3.3 mm. Lesions wer
12].
.3.5. Behavioral recordingsOne week after metabolic sampling, cannulae w
locked and animals were housed individually in transpalastic cages (30× 45 × 20) and maintained in a sounroof room under a 12:12 h light–dark cycle and cons
emperature (22± 1◦C), with free access to food and waefore any experimental manipulation. General activityontinuously monitored with infrared sensors attached tateral walls of the cages. Behavioral events were autoally stored in 10 min intervals for further analysis. Doublotted actograms were obtained for each animal and acrofiles were constructed for each group with the mean
ivity for each feeding condition (AL, RF, fasting). Analyas performed with the SPAD1 (Sistema de ProcesamAdquisicion de Datos. Version 1.1.1) designed for ourratory.
.3.6. HistologyAt the end of the behavioral experiments, animals w
illed with an overdose of sodium pentobarbital and perfuntracardially with 250 ml of 0.9% saline followed by 250f 4% paraformaldehyde in a 0.1 M PB and then cryo
ected overnight in 10, 20 and 30% of sucrose. Brainsrozen and cut in 40�m coronal sections, alternate sectiere mounted on gelatin-coated glass slides, then stith cresyl violet. Cellular damage and lesion placemere verified with a light microscope. Animals with incolete lesions were discarded from the study.

136 J. Mendoza et al. / Behavioural Brain Research 158 (2005) 133–142
Fig. 1. Mean number± S.E.M. of Fos-IR cells in Acc-Sh (A) and Acc-Co (B) in RF (gray bars) and AL rats (white bars). (C) and (D) Represent number ofFos-IR cells on the Acc-Sh and Acc-Co, respectively, during fasting after RFS (gray bars) and after ad libitum (white bars). Horizontal bars represent mealtimeor expected mealtime (stripes).*p < 0.05 Post-hoc entrainment vs. ad libitum.
2.3.7. Statistical analysesData were classified by groups and time and are repre-
sented as mean± S.E.M. Number of c-Fos-IR cells werecompared with a two-way ANOVA for the main factors groupand time. Body weight as well as food intake were analyzedwith a one-way ANOVA for independent measures for thefactor group (lesion versus sham). Metabolic variations in
F s under AL (left) and RFS (right) at ZT6. LV, Lateral ventricle; aca, anterior comisure.D
serum concentration of FFA, and locomotor activity, underRFS and during fasting conditions, were evaluated with a two-way ANOVA for the factors group (lesion versus sham) andtime, as a factor of repeated measures. All analyses were fol-lowed by a Tukey multiple-comparations post-hoc test with∝ set atp < 0.05. Statistical analysis was performed withStatistica for Windows version 4.5 (StatSoft, 1993).
ig. 2. Photomicrographs of Fos-IR in Acc-Sh and Acc-Co of animal
otted lines represent the counted area. Scale bar: 120�m.
J. Mendoza et al. / Behavioural Brain Research 158 (2005) 133–142 137
3. Results
3.1. Experiment 1
3.1.1. c-Fos expressionThe number of c-Fos-IR cells was increased in both Acc-
Sh and Acc-Co in food-entrained rats. The Acc-Sh of theRF group had significantly more c-Fos-IR cells than the ALgroup at ZT6, mealtime, and at ZT12, 6 h after feeding. Al-though not statistical significant, a marginal increase was ob-served at ZT8 (Figs. 1A and 2). In the Acc-Co a marginalincrease started in c-Fos–IR of RF rats at ZT5 and a statisti-cal increase was observed at ZT6 (Figs. 1B and 2). Becauseanimals for the ZT6 point in the RF group remained unfeduntil sacrifice, the c-Fos-IR at this time point was mainly as-sociated with the anticipatory activation. Although there wasnot statistical difference, there was also an increase in ZT12(Fig. 1B). The two-way ANOVA confirmed a main effect be-tween AL and RF in the Acc-Sh (F(1,42) = 25.78;p< 0.001)and in Acc-Co (F(1,41) = 21.04;p< 0.001). In the time factor,statistical analysis did not show a significant effect (Acc-Sh,F(5,41) = 1.081; NS; Acc-Co,F(5,42) = 2.06; NS). Further-more, there was a significant group× time interaction in theAcc-Sh (F(5,42) = 4.76;p< 0.001), but not interaction effectfor Acc-Co (F(5,41) = 1.52; NS).
A general effect during fasting was a lower number ofc rsis-t ow-e ceb -Cor= edo
3
3osis)
o werem omeS terala ry oft les.E s andi ctions eriorc ofm hella
TB
sion (n = 9) Core-sham (n = 7) Core-lesion (n = 7)
B 6± 3.24a 281.28± 4.13 252.28± 4.42a
F ± 0.62a 24.85± 1.03 12.00± 0.53a
Aspective control (Tukey post-hoc test).
Fig. 3. Schematic representation of lesions extent (maximum, light shading;minimum, dark shading) of the Acc-Sh (left) and Acc-Co (right). Represen-tative reconstruction of serial coronal sections are +1.7 mm anterior through+1.0 mm posterior to bregma (based on the Atlas of Paxinos and Watson,1998).
3.2.2. Body weight and food intakeBoth Shell-sham and Shell-lesion animals increased their
weight throughout the experiment. Shell-lesion animalsgained more weight than Shell-sham under ad libitum condi-tions (F(1,15) = 4.51;p< 0.05) (Table 1). Core-sham gainedweight in a similar proportion as the previous groups, how-ever, the Core-lesion group gained weight in a lower propor-tion and when compared to the Core-sham group the weightwas significantly lower (F(1,12) = 11.70;p< 0.005) (Table 1).The effect of the lesion on food ingestion during the 24 h in-terval was also different between the Shell and Core groups.Food intake was enhanced in Shell-lesion animals and thiseffect was significant compared with the Shell-sham group(F(1,14) = 9.60;p< 0.0078). In contrast, Core-lesion animalsshowed a strong decrease in ingested food during the exper-iment (F(1,12) = 122.11;p < 0.0001) (Table 1), in fact, forsome animals food had to be placed in their proximity.
-Fos-IR cells in both groups. There was not a clear peence of the food-entrained pattern in the RF-F group. Hver, the two-way ANOVA indicated a significant differenetween RF-F and AL-F groups in both Acc-Sh and Accegions (Acc-Sh,F(1,40) = 9.335;p< 0.003; Acc-Co,F(1,40)9.781;p< 0.003;Fig. 1C and D), post-hoc analysis shownly difference between groups in Acc-Co at ZT8 (p< 0.05).
.2. Experiment 2
.2.1. HistologyIn the sham-lesioned animals, no signs (cell loss or gli
f lesion-induced damage were observed. Shell lesionsainly restricted to the medial and ventral regions. In shell-lesioned animals, gliosis was observed in the land medial septum apparently affected by the trajecto
he injection needle or distension of the lateral ventricuvaluation of the Core-lesions revealed a marked cell los
ntense gliosis in a discrete area surrounding the injeite and shrinkage of the Core region around the antomissure.Fig. 3 shows an illustration of the extensionaximal and minimal lesions in representative rats with Snd Core lesions.
able 1ody weight and food intake in sham and lesioned animals
Shell-sham (n = 8) Shell-le
ody weight (g) 280.12± 5.02 293.6ood intake (g) 23.68± 0.86 27.00
ll values are mean± S.E.M.a p < 0.05 Difference between NMDA lesioned animals and their re

138 J. Mendoza et al. / Behavioural Brain Research 158 (2005) 133–142
Fig. 4. Temporal profile (mean± S.E.M.) of free fatty acids of rats exposedfor 3 weeks to RFS (A) for the Shell-lesions rats (white circles) and theircontrols (black circles) and for (B) Core-lesions rats (white triangles) andtheir controls (black triangles) The horizontal black bar indicates mealtime.*p < 0.05 Lesion vs. sham.
3.2.3. Serum free fatty acidsThe diurnal rhythm of FFA was entrained to RFS. Serum
levels of FFA were increased before meal time and declinedafter feeding in the Shell-sham and Shell-lesioned animals
F ity of raar
(Fig. 4A). The ANOVA indicated a significant difference inthe time factor (F(5,50) = 47.15;p < 0.0001), but there wasnot a difference between groups (F(1,10) = 1.24; NS). Also,both Core-sham and Core-lesion groups showed an increasein FFA plasma concentrations anticipating meal time (F(5,50)= 11.43;p < 0.0001), however, mean FFA levels were lowerin the Core-lesion animals (Fig. 4B). The ANOVA indicateda significant difference between groups (F(1,10) = 123.26;p< 0.0001).
3.2.4. Food-anticipatory activityDuring the first 15 days under food ad libitum condi-
tions, all groups showed a clear diurnal rhythm of activity.Statistical analysis showed that there was not a differencein the intensity of activity between Shell-sham and Shell-lesion groups (F(1,17) = 1.06; NS) and there was a differ-ence in time factor, indicating circadian rhythms entrainedto the light–dark cycle (F(23,391) = 23.40;p < 0.0001)(Figs. 5A, C and 6A). For the Core-sham and Core-lesiongroups there was a clear diurnal rhythm of locomotor activ-ity (F(23,368) = 15.85;p < 0.00001) and there was not adifference in intensity between groups (F(1,16) = 0.0001;NS) (Figs. 5B, D and 6D). During RFS, Shell-sham andShell-lesion animals exhibited a clear FAA. In the Shell-lesion group, FAA started 2 h before the Shell-sham group.G ctiv-i wasa
ig. 5. Representative double-plotted actograms of daily general activ
ccess (ad libitum). Rats were entrained to RFS (vertical gray bar) for 3 weerepresentative samples of Shell-sham and Core-sham rats respectively. (espectively.
ats kept in light–dark cycle 12:12 h. During the first 15 days animals had free food
eneral activity level was increased in the nocturnal aty as well as during FAA and in some animals there
large phase advance in this activity (Figs. 5C and 6B).
ks, followed by 3 days of fasting conditions (vertical white bar). (A) and (B) ShowC) and (D) Are two representative animals with NMDA lesions in Shell and Core,
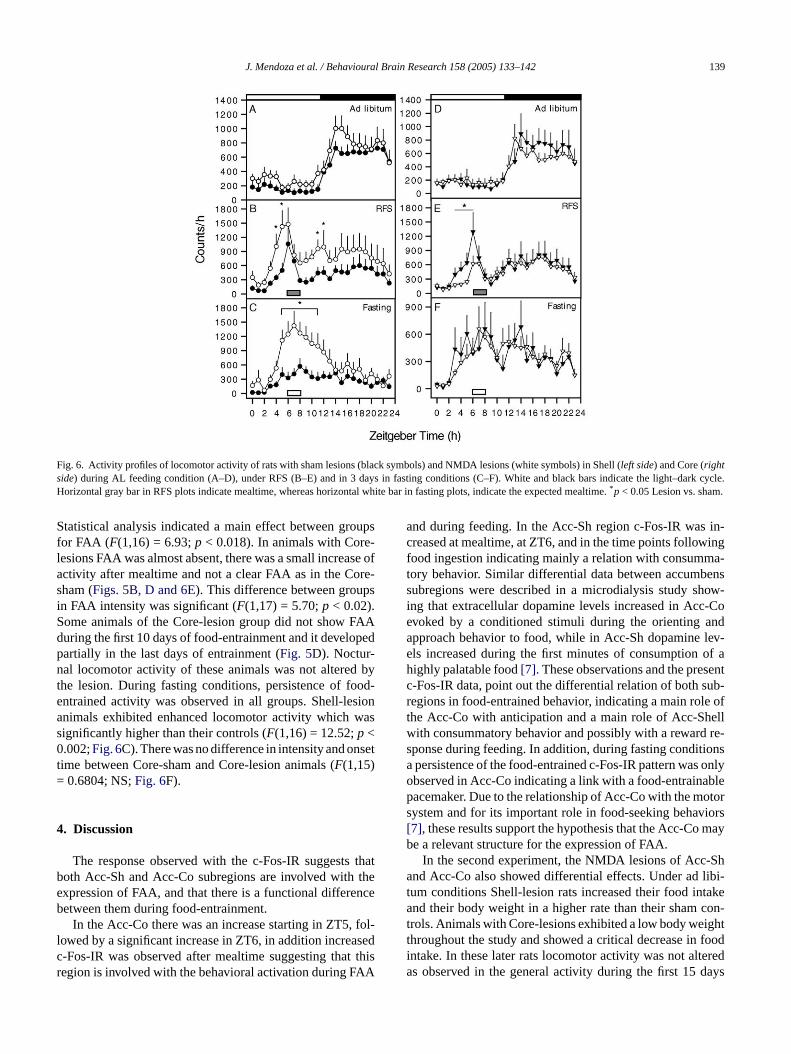
J. Mendoza et al. / Behavioural Brain Research 158 (2005) 133–142 139
Fig. 6. Activity profiles of locomotor activity of rats with sham lesions (black symbols) and NMDA lesions (white symbols) in Shell (left side) and Core (rightside) during AL feeding condition (A–D), under RFS (B–E) and in 3 days in fasting conditions (C–F). White and black bars indicate the light–dark cycle.Horizontal gray bar in RFS plots indicate mealtime, whereas horizontal white bar in fasting plots, indicate the expected mealtime.*p < 0.05 Lesion vs. sham.
Statistical analysis indicated a main effect between groupsfor FAA (F(1,16) = 6.93;p < 0.018). In animals with Core-lesions FAA was almost absent, there was a small increase ofactivity after mealtime and not a clear FAA as in the Core-sham (Figs. 5B, D and 6E). This difference between groupsin FAA intensity was significant (F(1,17) = 5.70;p < 0.02).Some animals of the Core-lesion group did not show FAAduring the first 10 days of food-entrainment and it developedpartially in the last days of entrainment (Fig. 5D). Noctur-nal locomotor activity of these animals was not altered bythe lesion. During fasting conditions, persistence of food-entrained activity was observed in all groups. Shell-lesionanimals exhibited enhanced locomotor activity which wassignificantly higher than their controls (F(1,16) = 12.52;p <0.002;Fig. 6C). There was no difference in intensity and onsettime between Core-sham and Core-lesion animals (F(1,15)= 0.6804; NS;Fig. 6F).
4. Discussion
The response observed with the c-Fos-IR suggests thatboth Acc-Sh and Acc-Co subregions are involved with theexpression of FAA, and that there is a functional differencebetween them during food-entrainment.
fol-l sedc t thisr AA
and during feeding. In the Acc-Sh region c-Fos-IR was in-creased at mealtime, at ZT6, and in the time points followingfood ingestion indicating mainly a relation with consumma-tory behavior. Similar differential data between accumbenssubregions were described in a microdialysis study show-ing that extracellular dopamine levels increased in Acc-Coevoked by a conditioned stimuli during the orienting andapproach behavior to food, while in Acc-Sh dopamine lev-els increased during the first minutes of consumption of ahighly palatable food[7]. These observations and the presentc-Fos-IR data, point out the differential relation of both sub-regions in food-entrained behavior, indicating a main role ofthe Acc-Co with anticipation and a main role of Acc-Shellwith consummatory behavior and possibly with a reward re-sponse during feeding. In addition, during fasting conditionsa persistence of the food-entrained c-Fos-IR pattern was onlyobserved in Acc-Co indicating a link with a food-entrainablepacemaker. Due to the relationship of Acc-Co with the motorsystem and for its important role in food-seeking behaviors[7], these results support the hypothesis that the Acc-Co maybe a relevant structure for the expression of FAA.
In the second experiment, the NMDA lesions of Acc-Shand Acc-Co also showed differential effects. Under ad libi-tum conditions Shell-lesion rats increased their food intakeand their body weight in a higher rate than their sham con-t ghtt foodi ereda ays
In the Acc-Co there was an increase starting in ZT5,owed by a significant increase in ZT6, in addition increa-Fos-IR was observed after mealtime suggesting thaegion is involved with the behavioral activation during F
rols. Animals with Core-lesions exhibited a low body weihroughout the study and showed a critical decrease inntake. In these later rats locomotor activity was not alts observed in the general activity during the first 15 d

140 J. Mendoza et al. / Behavioural Brain Research 158 (2005) 133–142
of baseline monitoring. Thus we assume that the decrease ofingestion was not due to a locomotor impairment. All groupsshowed entrainment of FFA although in Core-lesion rats thegeneral basal level of this metabolite was significantly de-creased. The lower levels of FFA can be explained by thedecrease in food intake and low weight gain in Core-lesionedanimals. All groups showed also FAA, however, in Shell-lesion rats the onset time was advanced and general activa-tion was increased, while Core-lesion rats showed a low ornon-FAA in the first days of entrainment.
The contrasting effect of Core and Shell-lesions may beattributed to specific anatomical connections associated withboth subregions of the nucleus accumbens. Whereas the Acc-Co establishes efferent projections to the motor system, theefferents of the Acc-Sh have more limbic characteristics. Adifferential role between Acc-Sh and Acc-Co in feeding hasbeen evidenced blocking excitatory amino acid input to theAcc-Sh, which resulted in a marked stimulation on feedingbehavior, while the same manipulation in Acc-Co, impairedthe response to food during an instrumental test[5,14].
Evidence obtained in this study suggests a relevant role ofAcc-Sh for the control and inhibition of feeding; first, basalfood ingestion was increased in Acc-Sh lesion animals, andsecond, c-Fos-IR was increase in Acc-Sh during feeding. Thissuggestion finds support in the anatomical relation betweenA ainn nd takeaT e toS duet . Ina thatA s ano FAAd .
AAo ay bei -Coi -Sh,b ate-p c de-s n re-gt hy-p stingt s re-q ssaryf asei thatA FAw ad-d orti-c ons ot odi-fi that
the FEO’s clock network is constituted by a distributed sys-tem[4], for this model it is possible that the Acc-Co is partof this network. In such a case its lesion would produce adecrease of FAA but not its total absense.
A decrease in FAA can be due to weak entraining signal toFEO, as well as due to a modification in the output pathway.Previous studies have described that the nutritious value andthe size of a meal are relevant to attain properties as a zeitge-ber[21], when the meal size is reduced during RFS, no FAAis observed. In addition, a previous study showed a decreasein FAA due to PBN lesions[9]. The most likely interpretationof these data was that the PBN is a relay nucleus for the en-training signal elicited by food and the gut. Thus, the lesionof PBN may decrease the intensity of food related signalsand thus produce a lower intensity of FAA. Present data alsodescribe a decrease in FAA intensity due to Acc-Co lesions,we suggest based on the anatomical relation of this region,that this decrease is mainly due to an alteration in the outputpathway or as described in the previous paragraph, due to analteration in the oscillator’s network itself.
It is important to point out that despite a clear decrease ofFAA in Core-lesion rats, no decrease was observed for thenocturnal activity, which showed similar values as the othergroups as can be observed inFigs. 5 and 6. This specific effecton FAA confirms that effects were not due to a locomotor im-p on aso ntalc cor-t te ion int ules[ be ac parto fort ad-d viag withtr ago-n ioni ina vi-d ssiono et-w ichd -p f theN eralp ight[ ven-t ecu-l forf
ionsi e-p ns of
cc-Sh and LH via an inhibitory gabaergic pathway as meurotransmitter[33,34]. Inhibition of the Acc-Sh functioue to an injection of a GABA agonist increases food innd produces an increase of c-Fos expression in LH[34].hus, the increase in body weight and food intake duhell-lesions, can be explained as a desinhibition of LH
o the absence of the GABA projections from the Acc-Shddition, evidence obtained in this study also suggestscc-Sh may be a region that inhibits locomotor activity autput of FEO, because in the Acc-Sh lesioned animals,uring RFS and during fasting conditions was increased
In contrast, the reduced amplitude and intensity of Fbserved in Core-lesion rats suggests that this region m
nvolved with the expression of FEO’s output. The Accs related to a different functional system than the Accecause the Acc-Co is functionally related with the caudutamen. Previous studies have described that specifitruction of dopamine terminals in the caudate-putameion in rats inhibits feeding, but not locomotion[30]. At
he molecular level, dopamine deficient mice becomeophagic and die of starvation by 4 weeks of age, sugge
hat regulated release of dopamine within the striatum iuired for the integration of salient cues that are nece
or feeding[35]. In despite that there was a clear decren FAA of Acc-Co lesioned animals, we can not arguecc-Co is the FEO, because FAA and anticipation of Fere not totally absent. In addition, this study did notress other variables that can be driven by FEO, like coesterone and clock genes expression in several regihe brain, which are entrained with RFS and should be med in animals with Acc-Co lesions. We have suggested
f
airment and suggests a specific role of the Acc-Co regiutput for the behavioral expression of FEO. The prefroortex, including the prelimbic and anterior cingulatedex, project to the Acc-Co, but not to the Acc-Sh[16]. Recenvidence has described a rhythm in clock genes expresshe prefrontal cortex which is entrained by feeding sched36]. These findings have suggested that the cortex mayircadian oscillator, and in this case, the Acc-Co may bef the output pathway to the principal motor structures
he expression of behavioral food-entrained rhythms. Inition, prefrontal cortex and Acc-Co are interconnectedlutamatergic fibers and this circuit has been implicated
he acquisitions of instrumental behaviors[17]. A previouseport described that intraperitoneal injection of an antist of NMDA receptors, MK-801, impaired FAA express
n rats[26]. Our results indicating an impairment of FAAnimals with Acc-Co lesions, in combination with the eence of the study cited above, suggest that the expref FAA may require the integrity of this corticostriatal nork. This possibility is further supported by a study whescribed that in the striatum and cortexNPAS2gene is exressed in a circadian manner and that in knout mice oPAS2gene FAA is deficient. These mice also show sevroblems to adapt to this food regimen and to gain we
11]. Interesting, the nucleus accumbens is part of theral striatum, suggesting that at physiological and molar level, manipulations of this region could be criticalood-entrainment.
A previous study reported that large accumbens lesn rats do not abolish FAA[22]. In that study, authors rorted a decrease of general activity by complete lesio

J. Mendoza et al. / Behavioural Brain Research 158 (2005) 133–142 141
accumbens including several structures like the septum, ven-tral pallidum, vertical limb of the diagonal band, the bednucleus of stria terminalis, anterior and ventral caudato-putamen and paraventricular thalamic nucleus. The weakFAA was attributed to the general state of hypoactivity andthe conclusion was that this structure was not involved withthe FEO. However, the present study does not agree withsuch findings, because we have described a differential roleof accumbens subregions and present data indicate that bothregions play a relevant role for food-entrainment.
At the moment there is not a clear evidence of an anatom-ical location of FEO. The present study adds information ona differential role of the two subregions of the accumbensas two components of the distributed system for the food-entrainable system. For this model Acc-Sh may be involvedwith the consummatory and motivational aspects of ingestionand thus with FEO’s inputs and Acc-Co is involved with theexpression of FAA and may be an output or an element of theclock’s network of FEO.
Acknowledgement
This work was supported by grants DGAPA-UNAMPAPIIT-IN-203803 to C.E. and Ph.D. fellowship (CONA-C
R
ians. Am
akirosc
ianCN-the
ex-m J
tivebens. Be-
Cell
minecom-
F,rs inmatic
irca-ion in304.
[ C.ulin,
glucagon, and corticosterone in food-restricted rats. Am J PhysiolRegul Integr Comp Physiol 2000;279:R2048–56.
[11] Dudley CA, Erbel-Sieler C, Estill SJ, Reick M, Franken P, Pitts S, etal. Altered patterns of sleep and behavioral adaptability in NPAS2-deficient mice. Science 2003;301:379–83.
[12] Escobar C, Dıaz-Munoz M, Encinas F, Aguilar-Roblero R. Persis-tence of metabolic rhythmicity during fasting and its entrainmentby restricted feeding schedules in rats. Am J Physiol Regul IntegrComp Physiol 1998;274:R1309–16.
[13] Granados-Fuentes D, Prolo LM, Abraham U, Herzog ED. TheSuprachiasmatic nucleus entrains, but does not sustain circadianrhythmicity in the olfactory bulb. J Neurosci 2004;24:615–9.
[14] Hanlon EC, Baldo BA, Sadeghian K, Kelley AE. Increases in foodintake or food-seeking behavior induced by GABAergic, opioid, ordopaminergic stimulation of the nucleus accumbens: is it hunger?Psychopharmacology 2004;172:241–7.
[15] Hara R, Wan K, Wakamatsu H, Aida R, Moriya T, Akiyama M, etal. Restricted feeding entrains liver clock without participation ofthe suprachiasmatic nucleus. Genes Cell 2001;6:269–78.
[16] Heimer L, Zham DS, Churchill L, Kalivas PW, Wohltmann C. Speci-ficity in the projection patterns of accumbens core and shell in therat. Neuroscience 1991;41:89–125.
[17] Kelley AE. Neural integrative activities of nucleus accumbenssubregions in relation to learning and motivation. Psychobiology1999;27:198–213.
[18] Klein DC, Moore RY, Reppert SM. The mind’s clock. NY: OxfordUniversity Press; 1991.
[19] Krieger DT. Food and water restriction shifts corticosterone,temperature, activity and brain amine periodicity. Endocrinology1974;95:1195–201.
[ oreisruptull
[ atoryand
[ od-ine
[ alRev
[ unc-tem.
[ oneRes
[ on-K-hav
[ . 4th
[ irca-
[ a H,er2)
achi-–42.
[ mineacol
[ andons.
yT) to J.M.
eferences
[1] Abe H, Rusak B. Anticipatory activity and entrainment of circadrhythms in Syrian hamsters exposed to restricted palatable dietJ Physiol Regul Integr Comp Physiol 1992;263:R116–24.
[2] Abe M, Herzog ED, Yamazaki S, Straume M, Tei H, SakY, et al. Circadian rhythms in isolated brain regions. J Neu2002;22:350–6.
[3] Amir S, Waddington-Lamont E, Robinson B, Stewart J. A circadrhythm in the expression of PERIOD 2 protein reveals a novel Scontrolled oscillator in the oval nucleus of the bed nucleus ofstria terminalis. J Neurosc 2004;24:781–90.
[4] Angeles-Castellanos M, Aguilar-Roblero R, Escobar C. c-Fospression in the hypothalamic nuclei of food-entrained rats. APhysiol Regul Integr Comp Physiol 2004;286:R158–65.
[5] Baldo BA, Sadeghian K, Baso AM, Kelley AE. Effects of selecdopamine D1 and D2 receptor blockade within nucleus accumsubregions on ingestive behavior and associated motor activityhav Brain Res 2002;137:165–77.
[6] Balsalobre A. Clock genes in mammalian peripheral tissues.Tissue Res 2002;309:193–9.
[7] Bassaero V, Di Chiara G. Differential responsiveness of dopatransmission to food-stimuli in nucleus accumbens shell–corepartments. Neuroscience 1999;113:324–36.
[8] Damiola F, Le Minh N, Preitner N, Kornmann B, Fleury-OlelaSchibler U, et al. Restricted feeding uncouples circadian oscillatoperipheral tissues from the central pacemaker in the suprachiasnucleus. Genes Dev 2000;14:2950–61.
[9] Davidson AJ, Cappendijk SL, Stephan FK. Feeding-entrained cdian rhythms are attenuated by lesions of the parabrachial regrats. Am J Physiol Regul Integr Comp Physiol 2000;278:R1296–
10] Dıaz-Munoz M, Vazquez-Martınez O, Aguilar-Roblero R, EscobarAnticipatory changes in liver metabolism and entrainment of ins
20] Maldonado-Irizarri CS, Kelley AE. Excitotoxic lesions of the cand shell subregions of the nucleus accumbens differentially dbody weight regulation and motor activity in rat. Brain Res B1995;38:551–9.
21] Mistlberger RE, Rusak B. Palatable daily meals entrain anticipactivity rhythms in free-feeding rats: dependence on meal sizenutrient content. Physiol Behav 1987;41:219–26.
22] Mistlberger RE, Mumby DG. The limbic system and foanticipatory circadian rhythms in the rat: ablation and dopamblocking studies. Behav Brain Res 1992;47:159–68.
23] Mistlberger RE. Circadian food anticipatory activity: formmodels and physiological mechanism. Neurosci Biobehav1994;18:171–95.
24] Mogenson GJ, Jones DJ, Yim CY. From motivation to action: ftional interface between the limbic system and the motor sysProg Neurobiol 1980;14:69–97.
25] Moore RY, Eichler VB. Loss of a circadian adrenal corticosterrhythm following suprachiasmatic nucleus lesions in rats. Brain1972;42:201–6.
26] Ono M, Shibata S, Minamoto Y, Watanabe S. Effect of the ncompetitiveN-methyl-d-aspartate (NMDA) receptor antagonist M801 on food-anticipatory activity rhythm in the rat. Physiol Be1996;59:585–9.
27] Paxinos G, Watson C. The rat brain in stereotaxic coordinatesed. San Diego, USA: Academic Press; 1998.
28] Reppert SM, Weaver DR. Molecular analysis of mammalian cdian rhythms. Annu Rev Physiol 2001;63:647–76.
29] Sakamoto K, Nagase T, Fukio H, Horikawa K, Okada T, Tenaket al. Multitissue circadian expression of rat period homolog (rPmRNA is governed by the mammalian circadian clock, the suprasmatic nucleus in the brain. J Biol Chem 1998;273(42):27039
30] Salamone JD, Mahan K, Rogers S. Ventrolateral striatal dopadepletions impair feeding and food handling in rats. PharmBiochem Behav 1993;44:605–10.
31] Stephan FK, Zucker I. Circadian rhythms in drinking behaviorlocomotor activity of rats are eliminated by hypothalamic lesiProc Natl Acad Sci USA 1972;69:1583–6.

142 J. Mendoza et al. / Behavioural Brain Research 158 (2005) 133–142
[32] Stephan FK. The “other” circadian system: food as a zeitgeber. JBiol Rhythms 2002;17:284–92.
[33] Stratford TR, Kelley AE. GABA in the nucleus accumbens shellparticipates in the central regulation of feeding behavior. J Neurosci1997;17:4434–40.
[34] Stratford TR, Kelley AE. Evidence of a functional relationship be-tween the nucleus accumbens shell and lateral hypothalamus sub-serving the control of feeding behavior. J Neurosci 1999;19:11040–8.
[35] Szczypka MS, Kwok K, Brot MD, Marck BT, Matsumoto AM, Don-ahue BA, et al. Dopamine production in the caudate putamen restoresfeeding in dopamine-deficient mice. Neuron 2001;30:819–28.
[36] Wakamatsu H, Yoishinobu Y, Aida R, Moriya T, Akiyama M, Shi-bata S, et al. Restricted-feeding-induced anticipatory activity rhythmin associated with a phase-shift of the expression in mPer1 andmPer2 mRNA in the cerebral cortex and hippocampus but not in thesuprachiasmatic nucleus of mice. Eur J Neurosci 2001;13:1190–6.
[37] Yamazaki S, Numano R, Abe M, Hida A, Takahashi R, Ueda M, etal. Resseting central and peripheral circadian oscillators in transgenicrats. Science 2000;288:682–5.
[38] Yoo SH, Yamazaki S, Lowrey PL, Shimomura K, Ko CH, BuhrED, et al. PERIOD2::LUCIFERASE real-time reporting of circadiandynamics reveals persistent circadian oscillations in mouse peripheraltissues. Proc Natl Acad Sci USA 101: 5339–46.
Copyright © 2022 FDOKUMEN




















