
contributions of nucleus accumbens circuitry to aspects of
235
CONTRIBUTIONS OF NUCLEUS ACCUMBENS CIRCUITRY TO ASPECTS OF AVERSIVELY-MOTIVATED BEHAVIORS by Patrick T. Piantadosi B.A., St. Mary’s College of Maryland, 2010 M.A., The University of British Columbia, 2013 A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY in THE FACULTY OF GRADUATE AND POSTDOCTORAL STUDIES (Psychology) THE UNIVERSITY OF BRITISH COLUMBIA (Vancouver) December 2017 © Patrick T. Piantadosi, 2017
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of contributions of nucleus accumbens circuitry to aspects of

CONTRIBUTIONS OF NUCLEUS ACCUMBENS CIRCUITRY TO ASPECTS OF
AVERSIVELY-MOTIVATED BEHAVIORS
by
Patrick T. Piantadosi
B.A., St. Mary’s College of Maryland, 2010
M.A., The University of British Columbia, 2013
A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF
THE REQUIREMENTS FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
in
THE FACULTY OF GRADUATE AND POSTDOCTORAL STUDIES
(Psychology)
THE UNIVERSITY OF BRITISH COLUMBIA
(Vancouver)
December 2017
© Patrick T. Piantadosi, 2017

ii
Abstract
The nucleus accumbens is a heterogeneous brain structure involved in the integration of limbic
and cortical input and the coordination of motor output during behavior. Made up primarily of
two major subregions, the nucleus accumbens core (NAcC) and shell (NAcS), this region has
been suggested to contribute to dissociable aspects of appetitive behavior on the basis of
differential functions localized within these subregions. Briefly, the NAcC may promote states of
behavioral action during reward-seeking, while the NAcS may refine such behavior by actively
inhibiting inappropriate or irrelevant actions. In Chapter 1, we discuss relevant research related
to the dissociability of the NAcC and NAcS at the circuit and behavioral levels. In Chapters 2, 3,
and 4, we examine the contribution of these two NAc subregions, as well as associated cortical
and limbic structures, to Pavlovian and instrumental suppression. Results suggested that the
NAcC acted to promote behavioral indices of reward-seeking vigor, while the NAcS was
necessary for the appropriate instantiation and expression of conditioned suppression. In Chapter
5, we probed the relevance of these NAc subregions to the performance of a novel active/passive
avoidance behavior. On this task, rats had to dynamically promote or inhibit their responding,
guided by discrete cues, to avoid a painful stimulus. While both NAc subregions were necessary
for promoting behavior during active avoidance trials, only the NAcS was required for inhibiting
responding during presentations of the passive avoidance stimulus. A control study suggested
that neither NAc subregion was necessary for unconditioned responding to foot-shock, indicating
that the previous results could not be explained by changes in pain sensitivity. We also probed
the role of monoaminergic transmission to motivational conflict and active/passive avoidance by
systemically administering d-amphetamine (AMPH) to a subset of animals in Chapter 3 and 4.
These results suggested that AMPH promoted punishment induced inhibition of behavior during
motivational conflict, but had the opposite effect during passive avoidance trials, inducing

iii
pressing despite punishment. Chapter 5 discusses these results in the framework of a dichotomy
between response-promotion and response-inhibition, relating these findings to extant literature
in the appetitive and aversive domains.

iv
Lay Summary
The ability to inhibit actions that are potentially harmful is an integral part of an organism’s
behavioral repertoire. Dysfunction of this behavior has been suggested to contribute to the
compulsive actions that characterize disorders such as addiction and obsessive-compulsive
disorder. A region within the ventral striatum, the nucleus accumbens, is composed of two
subnuclei, the nucleus accumbens core and shell, that may differentially contribute to aspects of
response-inhibition. Specifically, the accumbens core promotes reward-seeking, while the
accumbens shell acts to inhibit irrelevant information or actions. Whether these two regions
contribute to response-inhibition enforced by an aversive stimulus is unknown. Here, we
examined the contribution of these subregions to such behavior by using small infusions of
pharmacological agents to inhibit neuronal activity. Results suggested that the accumbens shell
contributes to aversively-motivated response-suppression, while the accumbens core promotes
action in the appetitive and aversive domains.

v
Preface
Experimental chapters (2-4) were conducted in the laboratory of Dr. Stan B. Floresco at the
University of British Columbia, within the Department of Psychology. Experiments were
designed by Patrick T. Piantadosi (P.T. Piantadosi) and Dr. Stan B. Floresco (S.B. Floresco). All
data collection was conducted by P.T. Piantadosi and undergraduate students under his direction.
Data were analyzed and written by P.T. Piantadosi, with assistance from S.B. Floresco.
- A version of Chapter 4 has been published in the following form:
Piantadosi, P. T., Yeates, D. C. M. M., Wilkins, M., & Floresco, S. B. (2017).
Contributions of basolateral amygdala and nucleus accumbens subregions to mediating
motivational conflict during punished reward-seeking. Neurobiology of Learning and
Memory, 140, 92–105. https://doi.org/10.1016/j.nlm.2017.02.017
P.T. Piantadosi performed all surgeries, and conducted behavioral training and testing
with assistance from D.C.M Yeates, M. Wikins, and K. Pezarro (undergraduate
volunteers). P.T. Piantadosi wrote the dissertation, with input from S.B. Floresco.
All experimental protocols were approved by the Animal Care Committee (ACC), University of
British Columbia, and conducted in compliance with guidelines provided by the Canadian
Council on Animal Care (CCAC).
ACC certificate numbers: A10-0197 or A14-021

vi
Table of Contents
Abstract ......................................................................................................................................................... ii
Lay summary ............................................................................................................................................... iv
Preface .......................................................................................................................................................... v
Table of Contents ......................................................................................................................................... vi
List of Tables ............................................................................................................................................. viii
List of Figures .............................................................................................................................................. ix
List of Abbreviations .................................................................................................................................... x
Acknowledgements ...................................................................................................................................... xi
Chapter 1: Introduction ................................................................................................................................. 1
1.1 The NAc: A heterogenous interface between affect and action .................................................... 3
1.2 NAc subregion-specific control of action and inhibition .............................................................. 6
1.3 Models of aversive learning and related circuitry ....................................................................... 12
Chapter 2: Cortico-striatal contributions to the acquisition and expression of discriminative conditioned
suppression .................................................................................................................................................. 28
2.1 Introduction ................................................................................................................................. 28
2.2 Methods....................................................................................................................................... 33
2.3 Results ......................................................................................................................................... 40
2.4 Discussion ................................................................................................................................... 48
Chapter 3: Investigating functional cortico-striatal or limbic-striatal circuits contributing to the acquisition
and expression of discriminative conditioned suppression ......................................................................... 72
3.1 Introduction ................................................................................................................................. 72
3.2 Methods....................................................................................................................................... 75
3.3 Results ......................................................................................................................................... 80
3.4 Discussion ................................................................................................................................... 82
3.5 Conclusion .................................................................................................................................. 86
Chapter 4: The role of NAc core and shell in motivational conflict during reward and punishment ......... 91
4.1 Introduction ................................................................................................................................. 91
4.2 Methods....................................................................................................................................... 94
4.3 Results ....................................................................................................................................... 100
4.4 Discussion ................................................................................................................................. 108
4.5 Conclusion ................................................................................................................................ 120
Chapter 5: Dissociable contributions of NAc core and shell during active/passive avoidance ................ 128
5.1 Introduction ............................................................................................................................... 128
5.2 Methods..................................................................................................................................... 133

vii
5.3 Results ....................................................................................................................................... 143
5.4 Discussion ................................................................................................................................. 148
5.5 Conclusion ................................................................................................................................ 164
Chapter 6: General discussion .................................................................................................................. 171
6.1 Dissociable contributions of NAc subregions to the inhibition and promotion of behavior ..... 172
6.2 AMPH induces task-dependent bidirectional changes in instrumental punishment ................. 178
6.3 Experimental merits and future directions ................................................................................ 181
6.4 Relevance to neuropsychiatric disease ...................................................................................... 186
6.5 Conclusion ................................................................................................................................ 188
References ................................................................................................................................................. 190

viii
List of Tables
Table 1. Mean (±SEM) values for overall locomotion, and the change in locomotor activity during CS+
versus CS- presentations within the conditioning session, for animals manipulated prior to
conditioning…………………………………………………………………………………………….64
Table 2. Mean (±SEM) values for total locomotion, rate of lever-pressing, and total lever-presses during
the discriminative fear expression test session………………………………………………………....65
Table 3. Mean (±SEM) values for ancillary measures during the conditioning session, induced by BLA-
NAcS manipulation prior to conditioning……………………………………………………………...87
Table 4. Mean (±SEM) values for ancillary measures induced by BLA-NAcS or PL-NAcS
manipulation.…………………………………………………………………………………………...87
Table 5. Mean (±SEM) values for ancillary measures on the Conflict or No-Conflict
task……………………………………………………………………………………………………..122
Table 6. Mean (± SEM) values for ancillary measures during the active/passive avoidance
task……………………………………………………………………………………………………..166

ix
List of Figures
Figure 1. Discriminative fear task diagram and histology. ......................................................................... 66
Figure 2. Inactivation of mPFC does not impact the acquisition of conditioned suppression .................... 67
Figure 3. Pre-conditioning NAcS, but not NAcC, inactivation diminishes conditioned suppression. ....... 68
Figure 4. Both mPFC subregions control the expression of conditioned suppression. ............................... 69
Figure 5. IL inactivation has no impact on conditioned suppression expression conducted using a
standard, single-stimulus design. ................................................................................................................ 70
Figure 6. NAcS, but not NAcC, mediates the expression of conditioned suppression. .............................. 71
Figure 7. Disconnection methodology diagram. ......................................................................................... 88
Figure 8. A BLA-NAcS disconnection does not mediate the acquisition of conditioned fear. .................. 89
Figure 9. A PL-NAcS projection contributes to the expression of conditioned suppression. ..................... 90
Figure 10. Histology schematic for Conflict and No-Conflict task animals ............................................. 123
Figure 11. Task diagram and data from pharmacological manipulation on the Conflict task .................. 124
Figure 12. Task diagram and data from inactivations on the No-Conflict task ........................................ 125
Figure 13. Baseline analysis suggests NAcS and BLA promote reward seeking as a function of task
history. ...................................................................................................................................................... 126
Figure 14. Trial structure and survival plot of training for the active/passive avoidance task. ................ 167
Figure 15. NAcC activity is necessary for active, but not passive, avoidance performance. ................... 168
Figure 16. NAcS activity is necessary for active and passive avoidance performance. ........................... 168
Figure 17. AMPH administration selectively provokes passive avoidance failure. .................................. 169
Figure 18. Neither NAc subregion is necessary for foot-shock sensitivity. .............................................. 170

x
List of Abbreviations
ACC Anterior cingulate cortex
ANOVA Analysis of variance
AP Anteriorposterior
BA# Broca’s area
B/M Baclofen/Muscimol
BLA Basolateral amygdala
CaMKII Calcium calmodulin-dependent kinase II
CO2 Carbon dioxide
CREB cAMP response element binding protein
CS (or CS+) Conditioned stimulus
CS- Neutral stimulus
dACC Dorsal anterior cingulate cortex
DV Dorsoventral
FR Fixed ratio
IL Infralimbic cortex
ML Mediolateral
mPFC Medial prefrontal cortex
NAc Nucleus accumbens
NAcC Nucleus accumbens core
NAcS Nucleus accumbens shell
OCD Obsessive compulsive disorder
OFC Orbitofrontal cortex
PFC Prefrontal cortex
PIT Pavlovian-to-instrumental transfer
PL Prelimbic cortex
PTSD Post-traumatic stress disorder
SAL Saline
US Unconditioned stimulus
VI Variable interval
VTA Ventral tegmental area

xi
Acknowledgements
This thesis would not have been possible but for the outstanding mentorship of my advisor, Dr.
Stan B. Floresco. Throughout my time in the lab, he has provided excellent guidance and
mentorship, providing me with the opportunity to probe questions that have not traditionally
been the central focus of the laboratory. His curiosity regarding the brain is contagious, and
many of the questions answered in this thesis are a direct testament to that. During my time in
the lab, I believe I have grown tremendously, both as a person and an academic. I credit Dr.
Floresco with enabling this growth, and cannot thank him enough.
I am additionally grateful to the other members of my supervisory committee, Dr. Todd
Handy and Dr. Catharine Winstanley, who have provided valuable insights into the construction
of this thesis. In particular, Dr. Handy and Dr. Winstanley encouraged me to connect this series
of experiments to a broad literature, which I believe strengthens the conclusions drawn within.
Many members of the Floresco lab have helped this thesis come to fruition over the past
four years. Special thanks to Dr. Colin Stopper and Maric Tse for their input on these projects
during their formative stages, provided during long hours of surgery and other animal work.
Other members of the laboratory, including Meagan Auger, Debra Bercovici, Courtney Bryce,
Gemma Floresco, Nicole Jenni, Josh Larkin, Ryan Tomm, and Mieke van Holstein, have
provided advice and camaraderie without which I would be at a loss. Other members of the
behavioral neuroscience department, including Lucille Hoover, Alice Chan, and Anne Cheng,
provided invaluable structural support for the animal work conducted in this thesis.
I am forever grateful to my family, in particular my parents and brother. My parents
raised me to be inquisitive and persevering, both qualities that I believe are apparent in the work
conducted throughout my time at UBC. I am grateful to my brother, a fellow neuroscientist, for

xii
his valuable input on these projects, as well as his friendship. Finally, I cannot express enough
gratitude towards my girlfriend, Joyce Miranda, for everything over the past six years. She has
sacrificed more for me than I’d care to admit, and without her patience and love, I’m not sure
how my Canadian experience might have turned out.

1
Chapter 1: Introduction
Aversive events and the cues that predict them have a tremendous ability to alter animal behavior
(Estes & Skinner, 1941; N. E. Miller, 1948). Depending on the particular contextual or
situational variables encountered, fearful events may inhibit or invigorate activity. In many
cases, such aversively-motivated behaviors are adaptive; for a foraging rodent, hearing a sound
within the frequency range of a predator vocalization will elicit a defensive response that may
protect it from harm. Survival is predicated on the ability of an animal to both attend and react to
predictive cues in the environment that signal when one action (e.g. foraging or approach
behavior) is favored over another (e.g. seeking shelter, or suppressing foraging).
This type of ethological situation has been suggested to have real-world implications for
modern-day humans (Hagenaars, Oitzl, & Roelofs, 2014; McNaughton, 1982; M. A. Miller,
Thomé, & Cowen, 2013; Pearson, Watson, & Platt, 2014; Pellman & Kim, 2016). Although
considerations regarding survival during the pursuit of such needs no longer applies to many
individuals, other costs of which we are afraid, such as losing wealth, status, employment, etc.,
weigh against potential benefits in a similar way as primary punishment. This parallel is
exemplified by the aberrant approach/avoidance behavior observed in neuropsychiatric
conditions. For example, negative consequences such as punishment are less effective at
inhibiting behavior in individuals with substance abuse or obsessive compulsive disorder
(Everitt, 2014; Feil et al., 2010; Wood & Ahmari, 2015), suggesting a potential deficit in
processing or utilizing negative consequences resulting from behavior. In other disorders,
aversive events have an inappropriately extreme impact on behavior, such as the elevated and
persistent levels of fear and anxiety expressed towards ambiguous or non-threatening stimuli in

2
individuals suffering from anxiety or post-traumatic stress disorders (Duits, Cath, Lissek, Hox,
Hamm, Engelhard, Van Den Hout, et al., 2015; Grillon & Morgan, 1999; Lissek et al., 2014).
Given the notable burden placed on individuals, families, and economies by these and
other neuropsychiatric conditions (Hjärthag, Helldin, Karilampi, & Norlander, 2010; Ohaeri,
2003; Whiteford et al., 2013; Whiteford, Ferrari, Degenhardt, Feigin, & Vos, 2015), developing
a better understanding of the neurobiological bases of aversively-mediated behavior is necessary.
As such, the brain mechanisms by which these events are learned about, maintained, and come to
alter behavior are a major focus of modern neuroscience. This interest has led researchers to
probe the brains of relatively simple model organisms, such as rodents, using increasingly
nuanced techniques during situations that provoke fear, or a competition between bivalent
motivations.
The aim of this thesis was to examine a potential role for the rodent nucleus accumbens
(NAc), as well as associated cortico-limbic afferents, in aversively-motivated behavior. Here, we
use the term aversive motivation to refer to any situation during which behavior is altered by the
potential delivery of an aversive stimulus. Although the NAc is commonly considered a
“reward” nucleus, given its established role in reinforcement learning and appetitive behavior, a
bivalent role for this region has been proposed and demonstrated (Aberman & Salamone, 1999;
Kim et al., 2017; Levita et al., 2009; Roitman, Wheeler, & Carelli, 2005; Schoenbaum & Setlow,
2003; Setlow, Schoenbaum, & Gallagher, 2003; Soares-Cunha, Coimbra, Sousa, & Rodrigues,
2016). In the following experiments, we examined the contributions of the NAc, specifically its
subregions, the shell and core, to situations where motivational drives conflict. Parallel findings
from the appetitive conditioning literature implicate these two subregions in partially dissociable
aspects of behavior. Such data suggest that, although both subnuclei may mediate some degree of

3
behavioral approach, the NAcS is uniquely responsible for the refinement of behavior by
inhibiting inappropriate actions. To date, few studies have examined whether such a functional
dichotomy of NAc function exists when response-inhibition or promotion are enforced by an
aversive stimulus, rather than by factors relating to reinforcer availability. We evaluate this
question using established Pavlovian and instrumental aversive conditioning methods, as well as
a novel avoidance paradigm, combined with local pharmacological inactivation of cortico-
limbic-striatal regions of interest.
1.1 The NAc: A heterogenous interface between affect and action
Prior to delving into the specific functions of the NAc related to aversively-motivated behavior, a
discussion of the region’s hodological complexity is necessary. The NAc is a neuroanatomically
and functionally heterogeneous structure, made up primarily of two main subregions, a lateral
core (NAcC) which surrounds the rostral portions of the anterior commisure, and a shell which
borders the core medially and ventrally (NAcS). These two subnuclei are anatomically
dissociable based on their expression of various proteins and neuroactive peptides. For example,
the calcium binding protein calbindin is enriched in the NAcC (similar to the dorsal striatum),
but relatively absent from the medial aspect of the NAcS (Jongen-Rêlo, Voorn, Groenewegen,
Voom, & Groenewegen, 1994; Meredith, Pattiselanno, Groenewegen, & Haber, 1996). In
comparison, expression of the peptide substance P is higher in the medial NAcS than in the
NAcC (Brog, Salyapongse, Deutch, & Zahm, 1993; Jongen-Rêlo et al., 1994). Primarily useful
for distinguishing between these two subregions in situ, such neurochemical distinctions hint at
potential differences in the functions controlled by the two subnuclei.
Although both the NAcS and NAcC receive afferent input from many of the same limbic
and cortical regions, the topographic nature of these projections are largely distinct. Generally,

4
the medial NAcS receives afferent input from ventral regions of the medial prefrontal cortex
(mPFC), as well as caudal or ventral sections of the basolateral amygdala (BLA) and
hippocampus/subiculum, respectively (Berendse, Galis-de Graaf, & Groenewegen, 1992; Brog et
al., 1993; French & Totterdell, 2002; Groenewegen, Wright, Beijer, & Voorn, 1999; Sesack,
Deutch, Roth, & Bunney, 1989; Vertes, 2004). In comparison, the NAcC receives input from
more dorsal regions of the mPFC, as well as a diffuse projection from basolateral amygdala
(BLA) and ventral hippocampus/subiculum. Midbrain dopamine neurons make a substantial
projection to the NAc, although the particular cell groups that project to each structure are
different. The medial A10 neurons in the ventral tegmental area (VTA) project prominently to
the medial NAcS, while the more lateral A10 neurons project predominantly to the NAcC
(Ikemoto, 2007). Thus, afferent projections to NAc subregions are often oriented
topographically, which suggests that behavioral dissociations may be mediated in part by these
partially segregated circuits.
In addition to the heterogeneity of afferent input received by these regions, the NAcS and
NAcC make efferent projections to divergent regions. The NAcC is typically considered to be
more tightly linked to motor output, projecting primarily to lateral ventral pallidum, substantia
nigra pars compacta (as well as the reticulata), and other motor affector sites (Berendse,
Groenewegen, & Lohman, 1992; Heimer, Zahm, Churchill, Kalivas, & Wohltmann, 1991;
Pennartz, Groenewegen, & Lopes Da Silva, 1994). In contrast, the NAcS projects to
dopaminergic cells in the ventral tegmental area, hypothalamic sites, and medial ventral pallidum
to control a diverse array of behavioral functions (Heimer et al., 1991; Pennartz et al., 1994).
Although many projections from these subnuclei are segregated, both regions share overlapping
inputs to the bed nucleus of the stria terminalis, lateral septum, and lateral habenula. NAc

5
subregions also project throughout the basal ganglia, including intrinsic reciprocal connections
between the NAcC and NAcS, which are more extensive from NAcC to NAcS than vice versa
(Van Dongen et al., 2005).
Within each structure, the inputs from limbic and cortical regions converge on inhibitory
GABAergic, medium spiny projection neurons (French & Totterdell, 2002, 2003), which make
up approximately 90% of cells in this nucleus (Meredith, 1999). Physiologically, these medium
spiny neurons have a bistable membrane potential, resting at a relatively hyperpolarized
membrane potential (“down-state”), and oscillating between this resting potential and a more
depolarized potential “up-state” (O’Donnell & Grace, 1995; O’Donnell, Greene, Pabello, Lewis,
& Grace, 1999). These up-states can be driven by strong afferent input from limbic (primarily
ventral subiculum) or prefrontal regions (Calhoon & O’Donnell, 2013a; Goto & O’Donnell,
2002; Gruber & O’Donnell, 2009; O’Donnell & Grace, 1995; O’Donnell et al., 1999). Once in
an upstate, action potential firing can be elicited by activity in critical limbic or cortical afferents,
suggesting that the NAc may effectively act as a gate, allowing task-relevant inputs to control
NAc output (Gruber, Hussain, & O’Donnell, 2009; Mogenson, Jones, & Yim, 1980). When
foraging in an operant environment, for example, coherence between structures mediating spatial
navigation, such as the hippocampus, and the NAcC increases, while exploiting an instrumental
operant response to receive reward increases coherence with a mPFC to NAcC circuit (Gruber et
al., 2009). Results such as these provide support for the hypothesis that the NAc integrates
competing input from limbic and cortical afferents, with the specific circuit most relevant for
task performance being recruited on demand. Given that differences exist in the specific efferent
and afferent projections of each NAc subregion, it is possible that the integration of these inputs
may lead to differences in function.

6
1.2 NAc subregion-specific control of action and inhibition
In fact, the dissociability of these subregions has been demonstrated across a variety of
experimental paradigms, primarily within the appetitive domain (for review, see Floresco, 2015).
Although a comprehensive review of these functions will not be undertaken here, findings from
the appetitive conditioning literature that may be of direct relevance to action selection following
aversive conditioning will be discussed. These functions include the ability of cues to act as
incentive stimuli, the refinement of cue-directed action selection, and the regulation of
impulsivity.
Incentive salience is a construct that describes the process by which discrete
environmental stimuli become imbued with the motivational properties of antecedent primary
reinforcers. This process seeks to explain how, in some animals, discrete cues predictive of
reward can come to control approach behavior (Berridge, 2012; Dickinson & Balleine, 1994).
One way to assess the incentive properties of a cue is by examining the Pavlovian-instrumental
transfer (PIT) effect. Following Pavlovian pairing of a stimulus (CS+; e.g., light, lever) with
reinforcement (e.g., sucrose), some rats learn to approach and engage the CS+, but not an
equivalently presented CS- (similar modality cue, never paired with reinforcement), reflecting a
shift in the incentive value of that cue. Although the CS+ is occasionally a manipulanda such as
an operant lever, this procedure is purely Pavlovian, with no instrumental response required for
reward delivery. During the transfer phase of the PIT procedure, presentation of the CS+ can
invigorate instrumental responding if the instrumental response is reinforced with the same
outcome as the CS+ (outcome-specific) or a novel substance (outcome-general). Lesions or
inactivations of NAc subregions differentially impacts these two types of PIT (Corbit & Balleine,
2011; Corbit, Muir, & Balleine, 2001). Generally, inhibiting activity within the NAcS impairs

7
the outcome-specific form of PIT, while the same manipulation of the NAcC impairs the
outcome-general form (Corbit & Balleine, 2011; Corbit et al., 2001). Consistent with an
integrative role of the NAc as a limbic-motor interface, this dissociation between the regional
specificity of outcome-specific versus general PIT is mediated by BLA-NAcS and BLA-NAcC
projections, respectively (Corbit & Balleine, 2005; Shiflett & Balleine, 2010). Recent reports
suggest a parallel functional circuit between the ventromedial PFC (vmPFC) and the NAcS that
may also mediate outcome-specific PIT (Keistler, Barker, & Taylor, 2015). Thus, NAcS may be
particularly sensitive to specific cue-outcome relationships, while NAcC may act more generally
to increase motivated output, as a function of differential cortico-limbic input.
Further support for such an incentive-motivational account of NAc function comes from
a series of elegant studies examining the meso-cortico-limbic-striatal regulation of response
selection, using a discriminative stimulus (DS) appetitive task (Ambroggi, Ghazizadeh, Nicola,
& Fields, 2011; Ghazizadeh, Ambroggi, Odean, & Fields, 2012; Ishikawa, Ambroggi, Nicola, &
Fields, 2008, 2010; Nicola, Yun, Wakabayashi, & Fields, 2004). This task requires rats to
discriminate between a DS that signals reward availability, which can be obtained by pressing an
active lever, and another stimulus that is never reinforced (NS). In addition, lever-presses on
another, inactive lever are never reinforced. Over the course of training, rats come to both
discriminate well between the DS and NS, as well as allocate their instrumental activity towards
the active lever exclusively during DS presentations. Thus, appropriate action selection results
from the promotion of reinforcement-seeking behavior during the DS, and an inhibition of this
same response during all other task phases.
The neural correlates of this behavior are observed both in the NAc, as well as the mPFC
and BLA (Ambroggi et al., 2011; Ghazizadeh et al., 2012; Ishikawa et al., 2008, 2010). When

8
first acquiring the task, animals learn to refine their behavior by encoding both the relevance of
the DS, and the irrelevance of the NS and the inactive lever, as well as other non-rewarded task
epochs, such as during the inter-stimulus interval. This acquisition is related to phasic activity
within the NAcS that correlates with the inhibition of irrelevant task actions, such as neural
responses to the NS (Ghazizadeh et al., 2012). In addition, a separate mechanism promotes the
tonic activity of neurons that act to oppose reward-seeking, further supporting a response-
inhibitory account of NAcS function. These two inhibitory processes during learning appear to
be mediated by a projection from the vmPFC (Ghazizadeh et al., 2012). In parallel, another
circuit mediated by the NAcC acts to promote approach behavior during DS presentations
(Ambroggi, Ishikawa, Fields, & Nicola, 2008; Ishikawa et al., 2008, 2010). BLA neurons
respond to a DS with short latencies, occurring earlier than do responses in the NAcC (Ambroggi
et al., 2008). Such results suggest that the BLA drives neuronal responses in the NAcC,
contributing to DS-evoked approach activity. The promotion of DS-evoked activity is also driven
by a possible circuit involving the dorsomedial PFC (dmPFC) and NAcC (Ishikawa et al., 2008,
2010). Single unit activity related to cue presentation or operant behavior is often larger in
magnitude when preceded by a DS, as compared to NS, suggesting that the DS-evoked behavior
and neural activity reflect an incentive motivational process.
These electrophysiological studies provide correlative evidence that NAc subregions, in
concert with cortico-limbic afferents, dissociably contribute to action selection. To causally
identify a role for these regions in response promotion and inhibition, pharmacological
compounds can be infused directly into discrete brain regions to affect neuronal activity. When
key regions of the PFC, BLA, or NAc are pharmacologically inhibited during performance,
behavioral impairments suggestive of deficient response-promotion and response-inhibition are

9
observed (Ambroggi et al., 2011, 2008; Ghazizadeh et al., 2012; Ishikawa et al., 2008, 2010;
Nicola et al., 2004; Yun, Wakabayashi, Fields, & Nicola, 2004). For example, the infusion of
GABAB and GABAA receptor agonists, baclofen/muscimol (B/M) into the vmPFC unmasks
activity within the NAcS that encodes previously inhibited task events, including reward-seeking
activity following NS presentation and during lever-presses on the never-reinforced lever
(Ambroggi et al., 2011; Ghazizadeh et al., 2012). The same manipulation of the dmPFC or BLA
preferentially impacts neuronal activity and behavior in response to the DS (Ishikawa et al.,
2008, 2010). When considering the NAc, the inhibition of activity within each subregion
produces differential results on DS-evoked reward-seeking, and NS-evoked behavioral
inhibition. Infusing B/M into the NAcC selectively decreases motivated reward-seeking behavior
driven by presentations of the DS (Ambroggi et al., 2011). Inactivation of the NAcS, in
comparison, makes a relatively specific contribution to the suppression of inappropriate or non-
rewarded behavior. Taking this subregion offline temporarily disinhibits lever-pressing during
the NS, as well as pressing of the inactive (never-reinforced) lever (Ambroggi et al., 2011). Thus,
these regions of the ventral striatum integrate afferent input to refine behavior, consistent with its
hypothesized role as a limbic-motor interface. However, the manner in which action selection is
refined differs by each subregion, with the NAcC allowing for response-promotion in response to
an incentive cue, and the NAcS inhibiting task-irrelevant or inappropriate reward-seeking.
These finding are paralleled by studies examining the reinstatement of reward-seeking
following extinction, which is often exaggerated in animals seeking food, cocaine, or alcohol
following NAcS inactivation (Di Ciano, Robbins, & Everitt, 2008; Floresco, McLaughlin, &
Haluk, 2008; Millan, Furlong, & McNally, 2010; Peters, LaLumiere, & Kalivas, 2008).
Eliminating neural activity within the NAcC, in contrast, typically produces the opposite effect,

10
inhibiting reinstatement (Di Ciano et al., 2008; Floresco et al., 2008). Extinction is a form of
behavioral flexibility that is thought to involve the formation of a new, inhibitory association
between a stimulus or action that previously produced an outcome, and the diminished incentive
value following omission of the outcome (Bouton & Moody, 2004). Inactivation of NAcS during
reinstatement may hamper the usage of this inhibitory memory, subsequently reinstating
behavior to a level comparable to animals that never underwent extinction. On the other hand,
NAcC-inactivation could eliminate phasic activity related to incentive cue presentation,
diminishing reward-seeking. Taken together, these results suggest that, while the NAcC is
relevant for general motivational drive in response to discrete stimuli, the NAcS may be
particularly important for suppressing inappropriate or inefficient response-strategies.
The NAc has also been implicated in impulsivity, which is a multifaceted construct that
reflects an inability to withhold a response when required (for review, see Basar et al., 2010). Of
particular relevance to response-inhibition as conceptualized here are impulsive actions, often
operationalized as premature motor responses that occur without foresight. This sort of
suppression can be indexed by Go/No-Go or five-choice serial reaction time tasks (5-CSRTT). In
a typical Go/No-Go task, discrete cues require either the production (a “Go” response) or
inhibition (a “No-Go” response) of a particular instrumental behavior in order to trigger reward
delivery. Thus, animals must flexibly and bi-directionally alter their behavior depending on the
particular cue presented. Unit recordings within the NAc illustrate that individual neurons
encode Go or No-Go stimuli, increasing or decreasing their activity during cue presentation
(Roitman & Loriaux, 2014; Setlow et al., 2003). Interestingly, response-suppression during
successful No-Go or unsuccessful Go trials has been shown to produce increases in NAc activity
that were greater in magnitude than were decreases, implying that elevations in accumbens

11
activity may allow for response-inhibition (Roitman & Loriaux, 2014). Although no studies have
examined whether the neural correlates of Go or No-Go performance differ across accumbens
subregions, data from other assays of impulsive action, such as the 5-CSRTT, provide insight
into the relative contributions of the NAcS and NAcC. The 5-CSRTT requires rats to wait a set
period of time prior to the brief illumination of a stimulus light, during which a nosepoke in the
illuminated port delivers reward. Responses prior to illumination of the stimulus light provide a
measure of impulsive action, known as premature responses, which delay the possibility of
reward receipt by restarting the waiting period. Inactivation of NAcS has been shown to increase
premature responses, while inactivation of NAcC simply diminishes attentional accuracy on this
task (Feja, Hayn, & Koch, 2014). Consistent with the aforementioned vmPFC-NAcS circuit
mediating response-suppression, vmPFC inactivation produces the same sort of impulsive
actions (Feja & Koch, 2014), which is recapitulated following pharmacological disconnection of
this circuit, but not a vmPFC-NAcC projection (Feja & Koch, 2015). While impulsive actions
may be particularly within the purview of the NAcS, the NAcC has been shown to contribute to
aspects of inhibitory control including impulsive choice (Cardinal, Pennicott, Sugathapala,
Robbins, & Everitt, 2001a; Pothuizen, Jongen-Rêlo, Feldon, & Yee, 2005). Impulsive choice
represents a more cognitive aspect of impulsivity, where animals shift their choice away from a
large reward as the delay associated with reward delivery increases. Such results suggest that the
NAcC may incorporate the costs associated with intertemporal choices, while being less
responsible for the relatively more rapid impulse control deficits associated with impulsive
actions. Therefore, the contribution of the NAcS to impulsive actions seems relatively consistent,
however NAcC may also contribute to aspects of response inhibition depending on the type of
response required.

12
Taken together, these results implicate accumbens subregions in dissociable aspects of
appetitive behavior. In particular, the NAcS mediates the impact that cues have on behavior
reinforced by a specific incentive, while actively inhibiting task-irrelevant information and
actions to refine action-selection. In contrast, the NAcC drives motivated behavior both generally
and in the presence of discrete motivational cues, without a prominent role in behavioral
suppression. Similarly, the NAcS may control the inhibition of impulsive actions, while the
NAcC promotes response accuracy, as well as the arguably more cognitive facets of waiting
impulsivity. That these same psychological principles of NAc function may apply not only to
appetitive behavior, but also to aversively-motivated response-inhibition and promotion has
received less empirical scrutiny.
1.3 Models of aversive learning and related circuitry
The emphasis on action selection evident across studies of NAc function suggests that
aversively-motivated behaviors which require response-promotion or inhibition may similarly
depend upon this region. In the appetitive domain, these two poles of behavior can be provoked
by reward availability versus reward unavailability or the risk of reward omission. In the
aversive domain, response-inhibition results from the presentation of an aversive stimulus, such
as a minor foot-shock, ocular air-puff, or loud acoustic startle stimulus. Depending on the
experimental conditions, response-promotion can also be observed during aversive conditioning,
particularly if an animal is given the ability to escape or avoid potential danger. These two poles
of aversively-motivated behavior, termed defensive reactions and defensive actions (Moscarello
& Ledoux, 2014), make up an essential part of an animals defensive repertoire, and may be
differentially regulated by the NAc. To better understand these two functions, and how NAc

13
subregions may contribute to them, a brief review of their psychological and neurobiological
underpinnings is necessary.
A variety of methods have been devised to evaluate defensive behaviors, built upon two
primary associative learning theories. The first borrows from the tenets of classical conditioning
put forth by Pavlov (1926) and others. Commonly termed Pavlovian fear conditioning, this
procedure involves the pairing of an initially neutral stimulus (e.g., light, auditory tone, context,
etc.) with an aversive unconditioned stimulus (US; e.g., minor foot-shock, ocular air-puff, loud
acoustic startle stimulus, etc.) Following repeated pairings of these stimuli, the neutral stimulus
becomes a conditioned stimulus (CS+), capable of eliciting a conditioned fear response when
presented in the absence of the US. In some designs, presentations of the CS+ can be
intermingled with the presentation of an explicitly neutral stimulus (CS-). Such discriminative
fear paradigms serve to control for baseline levels of fear and examine potential generalization of
the fear response (Likhtik & Paz, 2015; Piantadosi & Floresco, 2014). Importantly, during
Pavlovian fear learning, the behavior of an animal has no consequence on the probability of the
delivery of the aversive US.
In contrast to Pavlovian methods, the second model, based upon the Skinnerian principle
of instrumental conditioning (Skinner, 1938), results when an action is reinforced or punished,
depending on the affective valence of outcome itself. Using this methodology, an animal controls
the probability of US delivery via the production or inhibition of a particular instrumental
response. In the case of punishment, an instrumental action, such as pressing a lever for
reinforcement, can be paired with a contingent aversive unconditioned stimulus, such as foot-
shock. This pairing results in the expression of fear or anxiety during future situations in which
the punished instrumental action is available (Estes & Skinner, 1941; Vogel, Beer, & Clody,

14
1971). In most cases, this procedure is conducted in animals that are in a deprived state, typically
from a primary reinforcer such as food or water. Deprivation ensures that motivational conflict is
produced during punishment, as animals are highly motivated to seek reinforcement due to
deprivation, but also to avoid the aversive punishment that is concurrently delivered.
Whether conducted in a purely Pavlovian or instrumental manner, one can immediately
see that the fear produced by either procedure will have a qualitatively similar impact on
behavior: ongoing activity is inhibited due to the potential delivery of an aversive stimulus.
Despite the inherent difficulty in inferring emotional states in non-verbal species (Ledoux, 2014;
Panksepp, 2011), reliable measures of fear during aversive conditioning have been developed
based upon the innate defensive reactions expressed by mammals (Bolles, 1970; Moscarello &
Ledoux, 2014). The most commonly measured of these defensive reaction is freezing, defined as
the cessation of all movement (except respiration) (Blanchard & Blanchard, 1969; Campbell &
Teghtsoonian, 1958). Freezing reflects an attempt to evade predator detection (Bouton & Bolles,
1980), and provides researchers with a relatively unambiguous index of fear that can be scored
with ease. A secondary measure, which can be used in Pavlovian or instrumental scenarios, is the
conditioned suppression of reinforcement-seeking. Animals innately suppress their foraging
behavior in the presence of threat (Fanselow & Lester, 1988; Whishaw & Dringenberg, 1991).
Similar to conditioned freezing, this behavior is likely caused by a desire to minimize exposure
to danger that may occur during foraging. By utilizing these (and other) behavioral indices of
fear, one can begin to examine the neural correlates of such affective conditioning.
While these defensive reactions predominate in standard, Pavlovian situations where the
behavioral repertoire of an animal is severely curtailed, other, active responses prevail when
animals are provided with control over their environment (Berger & Brush, 1975; Mowrer &

15
Lamoreaux, 1946; Whishaw & Dringenberg, 1991). So called avoidance conditioning
incorporates Pavlovian and instrumental mechanisms, consisting of an early stage where CS
presentations evoke fear following pairing with an aversive US, and a later stage where the
performance of an instrumental response (e.g., lever-press, shuttling response) terminates the CS
and eliminates the potential delivery of the aversive US (Maia, 2010). Thus, animals can learn to
elicit an active approach response, overcoming the initial defensive reactions evoked by CS
presentation, to control the probability of receiving a foot-shock or other aversive stimulus.
Investigation of the neural circuitry underlying aversive learning has leaned heavily on
basic, Pavlovian fear conditioning. Predominantly using freezing as a readout of fear, a central
fear circuit encompassing nodes within the amygdala and prefrontal cortex, as well as midbrain
nuclei, has been identified. Briefly, the sensory properties of the CS+ and foot-shock US
converge on the lateral segment of the basolateral amygdala (BLA), allowing for the acquisition
and expression of conditioned fear (Iwata, LeDoux, Meeley, Arneric, & Reis, 1986; LeDoux,
Cicchetti, Xagoraris, & Romanski, 1990; Wilensky, Schafe, & LeDoux, 1999). Projections from
basal amygdala to the central nucleus of the amygdala (CeA) trigger freezing (as well as
neuroendocrine and autonomic) responses upon re-exposure to the CS+ alone (no foot-shock),
via projections to midbrain nuclei (e.g. periaqueductal gray) (Amorapanth, 1999; Fanselow,
1994).
While the initial acquisition of Pavlovian fear is predicated on amygdala integrity, fear
expression and extinction appear to require mPFC circuitry (Courtin, Bienvenu, Einarsson, &
Herry, 2013; Maren & Quirk, 2004). Generally, the two main subregions of the rodent mPFC,
the more dorsal prelimbic (PL) and the more ventral infralimbic (IL), are suggested to play
dissociable roles in the expression and extinction of fear conditioning. Specifically, PL mPFC

16
activity promotes the expression of conditioned fear, whereas IL activity inhibits fear, as occurs
during extinction (Burgos-Robles, Vidal-Gonzalez, & Quirk, 2009; Corcoran & Quirk, 2007;
Milad, Vidal-Gonzalez, & Quirk, 2004; Quirk, Russo, Barron, & Lebron, 2000). Stimulation of
the PL enhances, whereas pharmacological inactivation or lesion decreases, freezing behavior in
response to an aversively conditioned cue (Quirk et al., 2000; Sierra-Mercado, Padilla-Coreano,
& Quirk, 2011; Vidal-Gonzalez, Vidal-Gonzalez, Rauch, & Quirk, 2006). In contrast, IL
stimulation diminishes conditioned freezing, enhancing extinction, while the opposite occurs
following pharmacological or optogenetic silencing of this subregion (Bukalo et al., 2015;
Sierra-Mercado et al., 2011; Vidal-Gonzalez et al., 2006). Single unit activity in these regions
faithfully tracks their apparent opposite roles in fear expression. PL activity occurs during tone
presentations, and aberrantly elevated PL activity during extinction is correlated with extinction
failure (Burgos-Robles et al., 2009). In contrast, IL excitability decreases during conditioning,
and increases during extinction learning (Santini, Quirk, & Porter, 2008). It is important to
emphasize that, although understood in greater anatomical detail in the rodent, evidence
suggesting that the human amygdala and PFC (subdivisions homologous to PL/IL mPFC in
rodents) perform similar functions to their rodent counterparts has been reported (Adolphs,
Tranel, Damasio, & Damasio, 1995; Bechara et al., 1995; Büchel, Dolan, Armony, & Friston,
1999; Delgado, Nearing, LeDoux, & Phelps, 2008; Hariri et al., 2009; LaBar, Gatenby, Gore,
LeDoux, & Phelps, 1998; Milad et al., 2005; Motzkin, Philippi, Wolf, Baskaya, & Koenigs,
2014).
Reliance on the relatively simple, Pavlovian assessment of fear has left a comparative
imbalance in the understanding of the circuitry relevant to instrumental punishment. However,
recent work has identified structures involved in punishment, including some classically related

17
to fear such as the BLA and mPFC (Bressel & McNally, 2014; Jean-Richard-Dit-Bressel &
McNally, 2015; Pascoli, Terrier, Hiver, & Lüscher, 2015; Vento, Burnham, Rowley, & Jhou,
2017). Inactivation of the BLA disinhibits reward-seeking during punishment, consistent with a
native role for this region in response-inhibition (Jean-Richard-Dit-Bressel & McNally, 2015).
Interestingly, this effect is specific to the manipulation of the caudal aspect of the BLA, as rostral
inactivations had no effect on behavior (Jean-Richard-Dit-Bressel & McNally, 2015). Caudal
BLA projects more strongly to the NAcS than the NAcC (Berendse, Galis-de Graaf, et al., 1992;
Brog et al., 1993; Groenewegen et al., 1999; Kita & Kitai, 1990; Wright, Beijer, &
Groenewegen, 1996), implying that a BLA to NAcS projection may be relevant to punishment-
induced response-inhibition. The BLA likely encodes the value associated with a particular
event, whether positive or negative, and allows for the appropriate modification of behavior in
response. In the case of an aversively conditioned Pavlovian stimulus, the adaptive response
would be to freeze, while in an instrumental punishment setting, the conditioned suppression of
reward-seeking would be expected. Unlike the general assessment of freezing, conditioned
suppression requires the integration of multiple affective signals (e.g., fear, hunger, etc.), for
which an interface between the limbic and motor systems, perhaps the nucleus accumbens, is
likely required.
Unlike the consistency between Pavlovian and instrumental fear responses requiring the
BLA, the dissociation between the function of PL and IL cortex is less reliable. For example,
pharmacological inactivation of PL or IL has been shown to dramatically disinhibit shocked
water-spout licking in thirsty rats (Resstel, Souza, & Guimarães, 2008). Animals become less
sensitive to punishment following inactivation of either prefrontal subregion, persevering in
reward-seeking despite negative consequences. In contrast, other studies have suggested that

18
lateral segments of the PFC, including the orbitofrontal cortex (OFC) and insula, contribute more
to instrumental punishment than do either subregion of the mPFC (Jean-Richard-Dit-Bressel &
McNally, 2016). Importantly, both the insular cortex and OFC project to the ventral striatum,
including the NAc (Brog et al., 1993; Heilbronner, Rodriguez-Romaguera, Quirk, Groenewegen,
& Haber, 2016). The lack of coherence regarding PL/IL cortex function from studies
investigating instrumental punishment and conditioned freezing suggests that separable
mechanisms may underlie each behavior. Specifically, when inhibiting reward-seeking,
subregions of the mPFC may play qualitatively similar roles in the top-down regulation of such
behavior.
This suggestion has also been illustrated in studies examining the neural correlates of
addiction-like compulsive reward seeking, defined as drug-seeking despite foot-shock
punishment (Deroche-Gamonet, Belin, & Piazza, 2004; Everitt et al., 2008). For example,
prolonged access to cocaine produces punishment-resistant drug seeking in some animals
(Vanderschuren & Everitt, 2004), concomitant with hypofunction of medial prefrontal cortex
(mPFC) (Chen et al., 2013). Optogenetic inhibition or activation of mPFC decreases or increases,
respectively, the impact of punishment on cocaine seeking (Chen et al., 2013, but see Pelloux,
Murray, Everitt, 2013), suggesting that mPFC activity may be causally related to the
punishment-mediated inhibition of seeking. Similarly, pharmacological inactivations of the
mPFC produce operant responding for both cocaine and sucrose that is insensitive to potential
punishment (Limpens, Damsteegt, Broekhoven, Voorn, & Vanderschuren, 2015; Resstel et al.,
2008). Thus, prefrontal regions seem to perform a top-down inhibitory function, acting as a
break when responding is directly punished, or in the presence of a fear-inducing stimulus. Like
the BLA, mPFC projects strongly to regions of the ventral striatum, with dorsal regions of the

19
mPFC projecting to the NAcC, and more ventral regions projecting to the NAcS (Berendse,
Galis-de Graaf, et al., 1992; Brog et al., 1993; Sesack et al., 1989; Vertes, 2004).
On the periphery of this fear circuitry is the NAc, a ventral-striatal structure at the nexus
of affective, cognitive, and spatial information arriving from numerous cortico-limbic afferents.
Long considered a “reward” nucleus based in large part upon the necessity of this region for the
production of appetitive motivation (Cardinal, Parkinson, Hall, & Everitt, 2002; Parkinson,
Cardinal, & Everitt, 2000; Stopper & Floresco, 2011), numerous re-conceptualizations have
attempted to reconcile data suggesting that aversive events are also processed and influenced by
NAc activity (Berridge & Kringelbach, 2013; Carlezon & Thomas, 2009; Levita et al., 2009;
Reynolds & Berridge, 2002; Roitman et al., 2005; Salamone, 1994; Schoenbaum & Setlow,
2003; Setlow et al., 2003; Soares-Cunha et al., 2016). These later studies illustrate that single
neurons in the NAc respond to primary aversive stimuli (e.g. quinine taste), as well as the cues
that predict them (Roitman et al., 2005), and are necessary for the ability of such cues to alter
behavior (Schoenbaum & Setlow, 2003; Setlow et al., 2003). Although defensive reactions such
as freezing are equivocally-related to NAc activity, this nucleus may be more relevant for the
modification of reward-seeking behavior by fear (Kim et al., 2017). Finally, the NAc is
implicated directly in the avoidance of harm, a function critical to appropriate navigation of
approach/avoidance scenarios (Ramirez, Moscarello, LeDoux, & Sears, 2015; Salamone, 1994).
1.4 The NAc and aversively-motivated behavior
To postulate that the NAc is responsible for aspects of aversion, an expectation that neurons
within this region process aversive stimuli must be met. In fact, unconditioned aversive stimuli
have been shown to modulate NAc activity and neuromodulator release (Badrinarayan et al.,
2012; Baliki et al., 2013; Budygin et al., 2012; Roitman et al., 2005). For example, neurons

20
within the NAc increase their firing rate to infusion of an aversive quinine taste (Roitman et al.,
2005). This is coupled with a decrease in dopamine signaling during the quinine infusion, which
may be directly related to encoding of the motivational properties of the substance itself
(Roitman, Wheeler, Wightman, & Carelli, 2008). Interestingly, dopamine release may be
differentially affected as a function of subregional differences between the NAcC and NAcS in
response to primary aversive stimulus delivery (Budygin et al., 2012). Voltammetric recordings
from anesthetized rats subject to tail pinch suggests that, while release in the NAcC is time-
locked to the delivery of the tail pinch, dopamine release in the NAcS occurs immediately
following the cessation of the pinch. This result implies that NAcS may be relatively more
important for safety or relief learning, consistent with a variety of findings from animals and
humans (Baliki et al., 2013; Fernando, Urcelay, Mar, Dickinson, & Robbins, 2013; Mohammadi,
Bergado-Acosta, & Fendt, 2014).
Research has also demonstrated that the NAcS in particular can generate bivalent
motivational states via input from cortical subregions (Reynolds & Berridge, 2002; Richard &
Berridge, 2013). Infusions of the GABAA receptor agonist muscimol instigates ingenstive
behavior when infused into the rostral NAcS, but biases behavior towards defensive reactions
when infused into the caudal NAcS (Reynolds & Berridge, 2002). Interestingly, IL cortex acts to
put a break on either of these processes instigated by the NAcS, as activation of this structure
decreases feeding or defensive behaviors induced by rostro-caudal disruption of excitatory
activity within the NAcS (Richard & Berridge, 2013). By potentiating inhibitory signaling in the
NAcS, behaviors that are normally curtailed (e.g., voracious eating, anti-predator behavior when
there is no immediate threat) become unmasked. These findings are in general agreement with a
response-inhibitory circuit that is mediated by cortico-striatal activity and can bias motivational

21
states. Importantly, these effects on feeding only occur in the rostral portion of the NAcS, and do
not generally impact reward-seeking in an operant environment (Hanlon, Baldo, Sadeghian, &
Kelley, 2004; Stratford & Kelley, 1997; Zhang, Balmadrid, & Kelley, 2003, but see Wirtshafter
& Stratford, 2010).
In addition to unconditioned responses, other studies have demonstrated that
physiological and neurochemical indices of aversive learning occur in the NAc. For example,
when a CS is paired with an aversive event, dissociations have been observed between the
release of dopamine within each subnuclei of the accumbens in response to CS delivery
(Badrinarayan et al., 2012; Oleson, Gentry, Chioma, & Cheer, 2012). Badrinarayan and
colleagues (2012) reported that the presentation of an aversive CS decreases dopamine release
probability in the NAcC, while increasing the magnitude of release in the NAcS. Findings
regarding the NAcC have be corroborated by Oleson and colleagues (2012), showing that NAcC
dopamine decreases during CS presentations following fear conditioning (Oleson et al., 2012).
These neurochemical results suggest that NAc subregions differentially encode conditioned
stimuli predicting an aversive consequence. Specifically, decreases in NAcC dopamine release
during CS presentations may induce a state of hypoactivity during fear (Kelley, Baldo, Pratt, &
Will, 2005), while increases observed within the NAcS may signal salience or relief. Direct
electrophysiological recordings in the NAc illustrate that fear conditioning potentiates mPFC to
NAc afferents to CS+, but not CS-, deliveries, in a manner that is dependent on BLA input
(McGinty & Grace, 2008). Interestingly, the majority of recordings conducted by McGinty and
colleagues (2008) were localized in the NAcS, coherent with the suggestion that this region may
be particularly sensitive to aversive conditioning. Although no studies in humans have assessed
the differential contributions of accumbens subregions, activity within the whole NAc does

22
appear to track the valence of aversive cues (Delgado, Jou, Ledoux, & Phelps, 2009; Delgado,
Li, Schiller, & Phelps, 2008; Delgado, Nearing, et al., 2008; Jensen et al., 2003; Klucken et al.,
2009; Pohlack, Nees, Ruttorf, Schad, & Flor, 2012). Presentations of an aversive CS+ increases
activity within the NAc, while CS- presentations result in a smaller change in activity (Jensen et
al., 2008; Levita et al., 2009; Romaniuk et al., 2010). Taken together, these findings imply that
the NAc may play an integral role in the learning and expression of aversive conditioning.
Despite evidence that the NAc is involved in unconditioned and conditioned responses to
aversive stimuli, studies investigating the functional contribution of this nucleus to defensive
behaviors are essentially equivocal. Some studies implicate the NAc in the acquisition (but not
expression) of contextual fear conditioning, while sparing freezing induced by presentation of an
aversive cue (Haralambous & Westbrook, 1999; Riedel, Harrington, Hall, & Macphail, 1997).
This specific effect on contextual fear conditioning has been suggested to be mediated by the
prominent role of the ventral hippocampus/subiculum, which projects strongly to the NAc (Britt
et al., 2012; Brog et al., 1993; French & Totterdell, 2002, 2003), in contextual declarative
memory. However, still others report that inactivation of the NAcC impairs both the acquisition
and expression of fear-potentiated startle towards discrete cues (Schwienbacher, Fendt,
Richardson, & Schnitzler, 2004). Finally, recent studies suggest that the ventral striatum,
including the NAc, is critical for the extinction of fear (Correia, McGrath, Lee, Graybiel, &
Goosens, 2016; Rodriguez-Romaguera, Monte, & Quirk, 2012). Given the diversity of input
reaching the NAc, such discrepant results may not be particularly surprising. As outlined
previously, the NAc receives dense projections from the BLA, mPFC, and ventral hippocampus,
as well as neuromodulatory signals from the midbrain, all of which have been hypothesized to
regulate different aspects of aversive and appetitive conditioning (Cardinal et al., 2002;

23
Carlezon Jr. & Thomas, 2009). Adding to this complexity is that few studies have evaluated the
contribution of individual NAc subregions to aversive conditioning. Of the studies separately
considering the NAcC and NAcS, the majority have utilized permanent lesions which likely
affect multiple aspects of behavior, including learning, consolidation, and expression (Parkinson,
Robbins, & Everitt, 1999; Riedel et al., 1997; Wendler et al., 2013).
In addition, none of these previous studies have evaluated the contribution of these
regions to the aversion-induced suppression of reward-seeking. This is particularly relevant
given that appetitive conditioning studies show that the learned inhibition of behavior may be
uniquely under the control of the NAcS, via input from critical cortico-limbic afferents
(Ambroggi et al., 2011; Floresco et al., 2008; Ghazizadeh et al., 2012; Peters et al., 2008). The
mPFC and BLA have separately been linked to the conditioned inhibition of reward-seeking
(Chen et al., 2013; Jean-Richard-Dit-Bressel & McNally, 2015, 2016; Limpens et al., 2015;
Resstel et al., 2008), which they may enforce by direction projections to the NAc. Until recently,
this hypothesis had not been empirically tested. Kim and colleagues (2017) utilized molecular
and optogenetic techniques to interrogate a mPFC to lateral NAcS circuit during conditioned
suppression. They found a subset of mPFC neurons projecting to the lateral NAcS that were
activated by foot-shock, and whose activity was inversely related to reward-seeking. These
neurons were active during suppression, consistent with a role for the mPFC in top-down
inhibitory control, while hypoactivity within this projection was related to reward-seeking
despite potential punishment. Thus, the NAcS may be a striatal subregion particularly sensitive
to the influence of aversive stimuli on reward-seeking. Still, this prior study examined the lateral
NAcS, which receives less input from regions previously suggested to be relevant for response-

24
inhibition such as the mPFC and caudal BLA. Thus, investigation of the medial NAcS during the
conditioned inhibition of reward-seeking is warranted.
While the contribution of the NAc to Pavlovian and instrumental forms of response
suppression is uncertain, active behaviors designed to escape predation have been shown to rely
upon this nucleus. The learning and expression of active avoidance depends upon intact function
and dopaminergic innervation of the NAc. Dopamine release in the NAc increases during active
avoidance learning, and depleting dopamine in this region subsequently impairs the learning and
expression of this behavior (Boschen, Wietzikoski, Winn, & Cunha, 2011; Gentry, Lee, &
Roesch, 2016; McCullough, Sokolowski, & Salamone, 1993; Oleson et al., 2012; Wadenberg,
Ericson, Magnusson, & Ahlenius, 1990; Wietzikoski et al., 2012). During a successful
avoidance, phasic dopamine release occurs in the NAcC upon avoidance-cue presentation
(Oleson et al., 2012). Consistent with a bivalent role for this nucleus, dopamine release is
provoked by both reward cues and avoidance cues during performance of a well-trained
approach/avoidance task (Gentry et al., 2016). Performance on this task is correlated with cue-
selective dopamine release, as poor performing animals show a pattern of dopamine release that
is non-specifically higher and less selective for relevant cues (Gentry et al., 2016). Neural
activity within the NAcS has also been shown to be necessary for active avoidance performance.
Specifically, temporary inactivation of NAcS, or reversible disconnection of the NAcS from its
efferent BLA projection, impairs the ability of rats to produce an active avoidance (Fernando et
al., 2013; Ramirez et al., 2015). The NAcS may facilitate avoidance by encoding the salience of
signaled periods of safety during avoidance, as inactivation of this structure has been shown to
impair avoidance in situations where safety signals are not presented (Fernando et al., 2013).

25
In humans, active avoidance is also associated with neural activity in the NAc, suggesting
that a conserved avoidance circuit may exist across mammalian species (Delgado et al., 2009;
Levita, Hoskin, & Champi, 2012). Activity within the NAc increases during the learning of an
active avoidance response, in a manner that is correlated with amygdala activity (Delgado et al.,
2009). Thus, similar limbic-striatal interactions may underlie human active avoidance. Human
research has also provided insight into the accumbal regulation the opposite pole of avoidance,
passive avoidance (Levita et al., 2012). During this behavior, animals must withhold an
instrumental response to avoid an aversive stimulus. Levita and colleagues (2012) required
participants to make a button press to avoid an aversive consequence during the presentation of
one stimulus (active avoidance), and to withhold a button press to avoid an aversive consequence
during the presentation of another stimulus (passive avoidance). Participants completed this task
within an fMRI, revealing that BOLD activity within the NAc was differentially modulated by
active versus passive avoidance cues. Active avoidance provoked an increase in BOLD activity
within the NAc, while passive avoidance produced a deactivation in the same region.
Methodological limitations prevented this study from evaluating potential subregional-specificity
of this effect. Still, it is possible that the NAc and NAcS are differentially required on such a
task, in keeping with a potential role for the NAcS in response-inhibition (passive avoidance)
and the NAcC in response-promotion (active avoidance).
1.5 Objectives
Due to the present ambiguity regarding the necessity of NAc subregions to aversively-motivated
behavior, we examined the contribution of these nuclei to three distinct, yet related, behaviors.
These experiments were predicated on the general hypothesis that the NAcS may control aspects
of aversion-mediated response-inhibition, while the NAcC primarily contributes to approach

26
behavior. One behavioral ramification of Pavlovian fear cue presentation is the rapid
reorganization of ongoing behavior, such as during performance of an appetitive task (Estes &
Skinner, 1941; Kamin, Brimer, & Black, 1963). Such conditioned suppression of reward-seeking
has been proposed to reflect a type of aversive PIT, for which the NAc is necessary (as outline
above) in the appetitive domain (Cardinal et al., 2002; Everitt, Cardinal, Parkinson, & Robbins,
2003). A second manifestation of fear on behavior can be examined during instrumentally
delivered punishment, such that rats are fearful of approaching a desired stimulus or reinforcer.
Assessment of such motivational conflict has revealed roles for major NAc afferents, including
regions of the prefrontal cortex (Broersen et al., 1995; Jean-Richard-Dit-Bressel & McNally,
2016; Resstel et al., 2008) and BLA (Jean-Richard-Dit-Bressel & McNally, 2015), suggesting
that NAc itself may be integral. Finally, fear can, in certain situations, invigorate behavior, as
occurs during avoidance. Such active-avoidance is known to be dependent on NAc circuitry
(Delgado et al., 2009; Levita et al., 2012; Ramirez et al., 2015; Wendler et al., 2013). However,
another pole of avoidance behavior is passive-avoidance, whereby animals must inhibit
responding to avoid punishment. In humans, this behavior has been shown to involve activations
or deactivations of the NAc during active and passive avoidance, respectively (Levita et al.,
2012). Thus, we aimed to more specifically examine the circuitry involved in these related, but
distinct, avoidance behaviors, at the level of the NAcS and NAcC.
Chapter 2: Examined the role of NAc and prefrontal subregions to the acquisition and
expression of discriminative Pavlovian conditioned suppression. During these experiments,
animals were subjected to discriminative fear conditioning, where one conditioned stimulus
terminated with a mild foot-shock (CS+), while another had no consequence (CS-). Fear was
assessed by examining the conditioned suppression of reinforcement-seeking during presentation

27
of each CS type. Subregions of the medial PFC and NAc were pharmacologically inactivated
prior to acquiring fear, or prior to the expression of fear. This experiment was designed to
provide evidence that Pavlovian mechanisms of fear are regulated differentially by the NAcC
and NAcS, as well as the PL and IL cortices.
Chapter 3: Examined the role of two potential circuits mediating the acquisition and expression
of discriminative Pavlovian conditioned suppression. Based on the results of Chapter 2, we
utilized a pharmacological disconnection procedure to probe whether a BLA-NAcS circuit
mediates the acquisition of conditioned fear, and whether a PL-NAcS circuit mediates its
expression.
Chapter 4: Examined the role of the NAcS and NAcC in the expression of instrumental
punishment during conflict. During this task, rats were enticed to seek reward by a shift in
reinforcement from a lean to a rich schedule, however, lever-press responses were concurrently
punished by a mild foot-shock. After acquiring this behavior, these two accumbens subregions
were pharmacologically inactivated. This experiment was designed to provide evidence that
response-suppression mediated by instrumental punishment is sensitive to manipulation of the
NAcS, but not NAcC.
Chapter 5: Examined the role of the NAcS and NAcC in active versus passive avoidance. After
extensive training, each subregion was pharmacologically inactivated to examine potentially
dissociable contributions of the NAcS to response-inhibition (passive avoidance trials) and
response-promotion (active avoidance trials). This experiment allowed for the neurobiological
dissection of cue-driven instrumental actions, at the level of the NAc.

28
Chapter 2: Cortico-striatal contributions to the acquisition and expression of
discriminative conditioned suppression
2.1 Introduction
Fear is a powerfully motivating emotion with the ability to have an enduring effect on behavior.
For example, fear-inducing stimuli are capable of suppressing reward-seeking, which, in an
ethological setting, allows animals to go unnoticed by predators during foraging (Estes &
Skinner, 1941; Kamin et al., 1963; Whishaw & Dringenberg, 1991). In modern humans, the
maladaptive expression of such suppression has been suggested to underlie psychiatric disorders
characterized by compulsions or impulse control deficits, including substance abuse and
obsessive compulsive disorder (OCD) (American Psychiatric Association, 2013; Belin-Rauscent,
Fouyssac, Bonci, & Belin, 2016; Everitt, 2014; Feil et al., 2010; Figee et al., 2016; Jentsch &
Taylor, 1999; Limpens, Schut, Voorn, & Vanderschuren, 2014; Lubman, Yücel, & Pantelis,
2004; Perry & Carroll, 2008). A hallmark of substance abuse, for example, is the seeking of the
addictive substance despite adverse consequences, which often include negative effects on
physical and mental health, or the loss of occupational or social relationships. These
ramifications typically induce fear or anxiety in healthy individuals, curtailing such maladaptive
behaviors, but are less effective in these psychiatric populations. Thus, the neural basis of fear-
induced response-inhibition may have important implications for our understanding of behavior
from both an ethological and translational perspective.
Fear conditioning, based upon the associative learning principles outlined by Pavlov
(1926), is the most common method used in the interrogation of these circuits. During a typical
Pavlovian fear conditioning procedure, a brief, unexpected foot-shock (US) is rapidly associated
with co-occurring discrete (elemental) conditioned stimuli (CS). Subsequent re-exposure to these
Pavlovian cues will cause a rat to elicit a variety of defensive behaviors, including defensive

29
reactions (Bouton & Bolles, 1980; Fanselow, 1994; Moscarello & Ledoux, 2014). The most
commonly measured index of defensive behavior during Pavlovian fear is freezing, typically
defined as the cessation of all movement not related to respiration. A second, often
complementary measure is the conditioned suppression of reinforcement-seeking, which indexes
the withholding of an instrumental, reinforcement-seeking response during the presentation of an
aversive CS (Estes & Skinner, 1941; Kamin et al., 1963). This suppression enables animals to
minimize potential exposure to danger while foraging during an event that has been
unambiguously associated with an aversive consequence (Whishaw & Dringenberg, 1991).
Defensive reactions such as conditioned suppression amount to a response-inhibitory mechanism
acting to suppress behavior during a potentially dangerous event.
Investigations of these and related behaviors have helped to delineate a central fear
circuit encompassing distinct subnuclei of the amygdala (for review, see Fanselow & LeDoux,
1999) and prefrontal cortex (for review, see Courtin, Bienvenu, et al., 2013 and Sotres-Bayon &
Quirk, 2010), amongst other regions. Briefly, the sensory components of the aversive US and CS
converge in the lateral and basal amygdala, rendering this basolateral (BLA) complex critical for
fear acquisition, consolidation, and expression (for review, see Fendt & Fanselow, 1999).
Emerging evidence suggests that the two major subregions of the mPFC, the prelimbic (PL) and
infralimbic (IL) cortices, may perform opposing functions during fear expression and extinction
(for review, see Maren & Quirk, 2004). PL activity appears to promote, while IL activity
inhibits, the expression of conditioned freezing, with IL activity mediating the extinction of this
behavior (Corcoran & Quirk, 2007; Sierra-Mercado et al., 2011; Vidal-Gonzalez et al., 2006),
although it is relatively unclear if this distinction applies to other defensive reactions, such as
conditioned suppression. In fact, studies of conditioned suppression following instrumental

30
punishment suggest that PL and IL similarly promote the inhibition of seeking under threat of
danger (Resstel et al., 2008; but see Jean-Richard-Dit-Bressel & McNally, 2016), suggesting that
the suppression of reinforcement-seeking may be regulated differently than freezing at the level
of the prefrontal cortex.
Notably absent from this canonical fear circuitry is the NAc, a region of the ventral
striatum that receives convergent input from prefrontal and amygdala subregions necessary for
both appetitive and aversive affective conditioning (Berendse, Galis-de Graaf, et al., 1992; Brog
et al., 1993; Groenewegen et al., 1999; Vertes, 2004; Wright et al., 1996). This nucleus is
positioned to act as a limbic-motor interface, gating the impact of cortico-limbic input on action
selection via its downstream inputs to motor effector sites (Mogenson et al., 1980). Despite the
prime anatomical and physiological arrangement of this nucleus relevant to fear conditioning,
experimental data is essentially equivocal regarding its involvement in conditioned fear
(McDannald & Galarce, 2011; Parkinson et al., 1999; Riedel et al., 1997; Rodriguez-Romaguera
et al., 2012; Schwienbacher et al., 2004). For example, previous work suggests that lesions or
inactivations of the entire NAc leave instrumental or Pavlovian conditioned suppression intact
(McDannald & Galarce, 2011; Rodriguez-Romaguera et al., 2012). Some of this ambiguity may
relate to a lack of appreciation for the heterogeneous nature of the NAc itself. Like the
aforementioned mPFC, the NAc is composed of at least two distinct subregions, the nucleus
accumbens shell (NAcS) and nucleus accumbens core (NAcC), that are often anatomically as
well as functionally dissociable (Brog et al., 1993; Floresco, 2015; Zahm & Brog, 1992). The
NAcS, which is located on the medial and ventral aspect of the anterior commissure, receives
input from ventromedial mPFC, including PL and IL cortex, as well as the caudal aspect of the
basolateral amygdala. In contrast, the NAcC, a more lateral nucleus encircling the anterior

31
commissure, receives input from the dorsal mPFC, particularly the anterior cingulate and PL
cortex, as well as the full extent of the basolateral amygdala (Berendse, Galis-de Graaf, et al.,
1992; Brog et al., 1993; Vertes, 2004; Wright et al., 1996).
Recent descriptions of the dichotomous nature of these subnuclei suggests that, although
both NAcC and NAcS may be critical for approach behavior, the NAcS makes a unique
contribution to response-suppression (Ambroggi et al., 2011; Floresco, 2015; Ishikawa et al.,
2008; Peters et al., 2008; Piantadosi, Yeates, Wilkins, & Floresco, 2017). For example, neurons
within the NAcC encode the motivational relevance of an appetitive CS, while those within the
NAcS more often encode the appetitive CS as well as unrewarded task events, such as the
presentation of a neutral stimulus (Ambroggi et al., 2011). Consistent with this preferential
encoding of task-irrelevant events, inactivation of the NAcS disinhibits seeking behavior during
portions of the task that are explicitly unrewarded (Ambroggi et al., 2011; Ghazizadeh et al.,
2012; Ishikawa et al., 2008). In comparison, the same manipulation of the NAcC decreases
responding during presentation of the appetitive CS. Similarly, the reinstatement of
reinforcement-seeking following the formation of an inhibitory extinction memory is often
exaggerated in animals following inactivation of the NAcS, but not the NAcC (Floresco et al.,
2008; Millan et al., 2010; Peters et al., 2008). In many cases, the control of action selection has
been shown to involve interactions between the NAc and its key prefrontal (Ghazizadeh et al.,
2012; Ishikawa et al., 2008, 2010; Peters et al., 2008) and BLA (Ambroggi et al., 2008; Millan &
McNally, 2011) afferents. These regions of the NAc have also been differentially associated with
impulsive actions, which occur due to a failure of response-inhibition (Feja et al., 2014; Feja &
Koch, 2015; Murphy, Robinson, Theobald, Dalley, & Robbins, 2008). In the context of
impulsive action, these studies illustrate that the NAcS, via interactions with the vmPFC,

32
promotes response-inhibition, while NAcC is necessary for task performance (Feja et al., 2014;
Feja & Koch, 2015). Such results support a hypothesis that NAcS and NAcC contribute
relatively specifically to the promotion or inhibition of actions, respectively, when examining
reward-seeking.
Until recently, the possibility that the suppression of reward-seeking induced by aversive
consequences relies upon the NAcS has not been experimentally examined. One recent study by
Kim and colleagues (2017) used precise genetic targeting and calcium imaging to illustrate that a
subset of neurons within the mPFC project to the lateral NAcS to promote suppression following
punishment. Activity within this projection was suppressed when an animal sought reward
previously associated with foot-shock, but increased when such reward-seeking was inhibited
(Kim et al., 2017). Optogenetic activation of this pathway inhibited seeking when under risk of
punishment, suggesting a causal role for this projection in response-inhibition mediated by an
aversive event. However, the task utilized by Kim and colleagues (2017) delivered the aversive
stimulus in an instrumental fashion, leaving open the question of whether the NAcS mediates
response-inhibition in response to aversive Pavlovian cues. Moreover, whether medial NAcS and
the NAcC perform dissociable roles during the conditioned suppression of reward-seeking is
unknown.
This chapter aimed to examine whether individual subnuclei of the mPFC and NAc
differentially contribute to conditioned suppression, using a discriminative conditioning protocol.
Temporary pharmacological inactivations of the PL, IL, NAcC, or NAcS were conducted to
probe the involvement of these regions in the acquisition or expression of the discriminative
conditioned suppression of sucrose-seeking. All rats were trained to lever-press for sucrose
reward, and then were subjected to two critical fear conditioning days. During acquisition, two

33
conditioned stimuli were delivered, one that co-terminated with a mild foot-shock (CS+), and
one that was never associated with any consequence (CS-). Following fear learning, rats were
given an expression test day where the influence of each type of CS on lever-press suppression
was evaluated. We hypothesized that, although none of the subnuclei tested would be necessary
for the acquisition of discriminative conditioned suppression, inactivation of either the PL cortex
or NAcS prior to the fear expression test would disinhibit sucrose-seeking during the
presentation of an aversive CS+, consistent with a role for these regions in generating
suppression in response to aversive stimuli. In contrast, we anticipated that inactivation of the IL
cortex, which has previously been linked to the extinction of Pavlovian fear, would enhance fear
expression, while the same manipulation of the NAcC would simply promote response vigor.
2.2 Methods
2.2.1 Animals
All procedures were approved by the Animal Care Committee at the University of British
Columbia, in accordance with the Canadian Council on Animal Care guidelines. Separate groups
of Long Evans rats (Charles River) arrived weighing 250-300g. Rats were initially housed in
groups (4-5 rats/cage) with ad libitum access to food and water. After 5-10 d of acclimatization
to the colony, rats were stereotaxically implanted with bilateral stainless-steel guide cannula,
described in detail below. During the remainder of the experiment (approximately 4 wks), rats
were singly-housed and food-restricted to approximately 90% of their free-feeding weight. Rats
were allowed to gain weight following this initial period of restriction, such that they were
maintained on a delayed growth curve. Each experimental cohort was composed of 16 rats. To
avoid potential cohort effects, care was taken to assign a comparable number of rats to each

34
experimental Treatment condition (B/M vs. SAL), based primarily on matching for the average
number of lever-presses made during baseline sessions.
2.2.2 Apparatus
Behavior was assessed using eight standard Med Associates operant chambers, enclosed in a
sound attenuating chamber (30.5 X 24 X 21 cm; Med Associates, St. Albans, VT, USA). Each
operant chamber was assembled in an identical fashion. Two levers, separated by a food
receptacle where sucrose reinforcement was delivered (45 mg pellet; BioServ, Frenchtown, NJ,
USA), were situated on the right wall of the chamber (as viewed from the open chamber door).
Above each lever was a 100 mA cue light, used as part of the compound CS+. On the opposite
wall of the chamber (left wall), a single 100 mA house light illuminated the chamber and served
as part of a compound CS-. An auditory speaker, which allowed for the delivery of
discriminative auditory stimuli via a programmable generator (ANL-926, Med Associates), was
located next to the house light. Locomotor activity was measured by four infrared photobeams
located just above the grid floor, which was comprised of 19 stainless steel rods spaced 1.5 cm
apart. These rods were wired to a shock source and solid-state grid scrambler to allow for foot-
shock delivery.
2.2.3 Surgery
Due to changes in institutional policies regarding anesthesia, rats were anesthetized either with a
combination of ketamine/xylazine (100/10 mg/ml at 100/10 mg/kg, i.p.) or a half dose of
ketamine/xylazine (same mg/ml, i.p) followed by maintenance using Isoflurane anesthetic (2-3%
Isoflurane concentration) throughout surgery. Twenty-three gauge bilateral stainless-steel guide

35
cannula were implanted aimed at the PL, IL, NAcS or NAcC according to the following
stereotaxic coordinates (in mm):
PL – from bregma: AP +3.2; ML: ±0.7; from dura: DV: -2.8
IL – from bregma: AP: +2.8; ML: ±0.7; from dura: DV: -4.1
NAcS – from bregma, AP: +1.3, ML: ±1.0, from dura, DV: -6.3
NAcC – from bregma, AP: +1.6, ML: ±1.8, from dura, DV: -6.3
Four stainless-steel skull screws were inundated with dental acrylic to secure cannula in place.
Stainless-steel obturators flush with the end of the guide cannula were inserted after surgery.
Rats were given 5-10 d to recover from surgery before beginning behavioral training.
2.2.4 Lever training
The day before their initial operant training session, all rats were provided with ~30 sucrose
pellets in their home cage, to reduce neophobia to the reinforcer. All training was conducted at a
consistent time each day. Rats were initially trained to press the left lever (only lever available
during any portion of training/testing) on a fixed ratio 1 (FR1) schedule of reinforcement to a
criterion of 40 total presses during the 30 min session. After reaching criterion, rats were trained
over three consecutive days on increasing variable interval (VI) schedules, whereby reward was
provided after approximately 15 (VI15), 30 (VI30), or 60 (VI60) seconds of pressing (one
session at a particular schedule, per day). Rats were then trained on the VI60 schedule for 10-13
d, after which aversive conditioning was conducted. A VI60 schedule engenders a high rate of
lever-pressing in rats, while allowing reward rate to remain relatively consistent, allowing for the

36
accurate assessment of conditioned suppression as a proxy for fear (Kamin et al., 1963;
McAllister, 1997; Piantadosi & Floresco, 2014; Quirk et al., 2000).
2.2.5 Discriminative fear conditioning
2.2.5.1 Conditioning session
Following VI60 training, rats underwent discriminative fear conditioning in an identical fashion
as we have reported previously (Piantadosi & Floresco, 2014), based off of discriminative assays
used in rodents and humans (Antunes & Moita, 2010; Balog, Somlai, & Kéri, 2013; Jensen et al.,
2008). During this protocol, rats received 8 presentations each of a neutral conditioned stimulus
(CS-) and an aversive conditioned stimulus (CS+), with an average inter-stimulus interval of 180
s (min: 100 s, max: 240 s). Rats were placed into a chamber and initially received two
presentations of a 30 s CS- (1 kHz, 80 dB tone and flashing house-light). Following these two
presentations, rats received six more CS- presentations, and seven presentations of the 30 s CS+
(9 kHz, 80 dB tone and flashing house-light co-terminating with a 0.5 mA foot-shock delivered
over 0.5 s) in a pseudorandom order. The session ended following one additional CS+ delivery.
Previous work in our laboratory suggests that this combination of visual stimuli and order of
presentation produces robust and reliable discriminative conditioned suppression in control
animals (Piantadosi & Floresco, 2014). The day after this conditioning session, animals were
given a baseline VI60 session (no shocks or conditioned stimuli).
2.2.5.2 Expression test session
The day after the baseline VI60 session (48 hrs post-conditioning), rats were given a fear
expression test session. Rats initially experienced a 5 min period identical to their normal VI60
session, during which they lever-pressed for sucrose reward. Immediately following this period,

37
presentations of the CSs began, initially with four 30 s CS- presentations (five min inter-stimulus
interval), followed by four 30 s presentations of the CS+ (no foot-shock; five min inter-stimulus
interval). The suppression of lever-pressing during each CS presentation served as an index of
fear, as rats suppress seeking behavior in the presence of an aversive CS+ (Kamin et al., 1963;
Piantadosi & Floresco, 2014; Quirk et al., 2000; Sierra-Mercado et al., 2011). Suppression was
calculated using the formula [(A-B)/(A+B)], where A was the number of lever-presses made in
the 30 s epoch prior to CS presentation, and B was the number of lever-presses made during the
30 s CS presentation. Calculated this way, complete suppression is indicated by a value of 1,
while a values at 0 or below indicate no suppression or facilitation, respectively. Rarely, rats did
not press during a pre-tone and tone period; a suppression value of 1 was applied to all such
instances, as in previous reports (Quirk et al., 2000). To ensure that suppression ratios were
accurate, an a priori inclusion criteria of greater than 200 presses made during the test session
was established. Across all experimental cohorts, data from n = 3 rats were eliminated as a result
of this criterion.
2.2.6 Single-stimulus fear conditioning: Pre-test IL inactivation
As the impact of IL cortex inactivation during the expression test was unexpected, we conducted
an additional experiment to ascertain whether conditioned suppression expression differentially
requires the IL as a function of the discriminative versus single-stimulus nature of the design.
Thus, animals were implanted with cannula into the IL cortex, and given an identical lever
training protocol as described above.
However, during the conditioning session, animals received eight presentations of a
single, 30 s CS+ (identical to the CS+ used in the discriminative protocol) only, similar to
conditioning procedures used in prior studies examining IL function during fear (Akirav, Raizel,

38
& Maroun, 2006; Sierra-Mercado et al., 2011). Forty-eight hrs later, rats were given a test
session that was initially identical to a normal VI60 day. Beginning five min into the session,
they received 12 presentations of the 30 s CS+ (no foot-shock), each separated by a three min
interstimulus interval.
2.2.7 Microinfusion
To examine the acquisition or expression of discriminative fear, separate cohorts of rats were
given microinfusion before either the conditioning or expression test sessions. Initially, all rats
were given a mock infusion 10 min prior to their final VI60 session before discriminative
conditioning. During this session, obturators were removed, mock injectors flush with the
indwelling guide cannula were inserted, and animals were allowed to freely move in the infusion
enclosure for approximately two min. On the infusion day, obturators were removed and
stainless-steel injectors extending 0.8 mm beyond the guide cannula were lowered into the region
of interest. Through this injector, rats received bilateral infusion of 0.9% saline (SAL; 0.3
μl/side) or a solution of the GABAB-receptor agonist baclofen and the GABAA-receptor agonist
muscimol (B/M; 75 ng/μl of each drug at a volume of 0.3 μl/side). Infusions were conducted
over 45, with injectors left in place for an additional 60 s to allow for diffusion of solution from
cannula tips. The dose and volume of B/M selected has been used previously to dissociate
between the NAcS and NAcC on a wide variety of behavioral measures (Dalton, Phillips, &
Floresco, 2014; Floresco et al., 2008; Ghods-Sharifi & Floresco, 2010; Stopper & Floresco,
2011). We have also used the same or larger volumes of a B/M solution to dissociate PFC
subregions (Dalton, Wang, Phillips, & Floresco, 2016; St. Onge & Floresco, 2010). We chose to
use the same, smaller volume as infused into NAc subregions to limit the potential for diffusion
across the dorsoventral axis of the PFC.

39
2.2.8 Histology
All rats were euthanized with CO2 and brains were removed and fixed in a 4% phosphate
buffered formalin solution. Brains were sectioned at 50 μm, following which tissue was mounted
and Nissl stained using Cresyl Violet. Placements were examined under a light microscope, and
the ventral extent of each infusion is indicated in Fig. 1B and C.
2.2.9 Data analysis
Because there was no lever available during the conditioning session itself, the only behavioral
measure available for the assessment of conditioning was locomotion, as assessed by
photobeam-breaks/epoch. For each CS delivery, the change in locomotor activity during the CS
presentation was calculated, as compared to the overall locomotor baseline (average of all 16
pre-tone periods). These change in locomotion values were then averaged for each CS type, and
analyzed using two-way between/within ANOVAs with Treatment group (SAL vs. B/M) as the
between-subjects factor, and CS Type (CS+ vs. CS-) as the within-subjects factor. This analysis
attempted to clarify the efficacy of conditioning, and its potential alteration by drug-treatment
during the conditioning session itself.
During the expression test session, the suppression ratio during each CS presentation was
analyzed using between/within-subjects three-way ANOVAs with Treatment group (SAL vs.
B/M) as the between-subjects variable, and CS Type (CS+ vs. CS-) and CS Number (1-4) as the
within-subjects variables. Separate ANOVAs were conducted on data from animals infused pre-
conditioning or pre-test for each brain region (PL, IL, NAcC, and NAcS). Follow-up simple
main effects analyses were conducted using one-way ANOVAs or t-tests, where appropriate.
Locomotion (photobeam breaks/session) during the conditioning session or expression test were

40
analyzed using separate independent samples t-tests. The rate of lever-pressing (presses/min) in
the first 5 min of the expression test session and the total number of lever-presses made during
the session were analyzed in an identical fashion.
2.3 Results
2.3.1 PL cortex inactivation pre-conditioning
To assess the contribution of PL activity during the acquisition of discriminative fear
conditioning, this region was inactivated immediately prior to the conditioning phase of the task.
During this phase, overall locomotor activity was unchanged by Treatment (t(16)=0.26,p>0.79)
(Table 1). Rats increased their locomotion significantly more during CS+ presentations, as
compared to CS- (F(1,16)=32.67,p<0.001), suggesting that animals behaviorally differentiated
between the two stimuli. Treatment had no effect on the change in locomotion induced by either
stimulus, as shown by a non-significant main effect of Treatment (F(1,16)=0.39,p>0.54), and a
non-significant CS Type x Treatment interaction (F(1,16)=0.03,p>0.86) (Table 1). Thus, rats
appeared to respond comparably during conditioning, regardless of treatment condition.
During the expression test, the level of conditioned suppression expressed by rats that had
their PL inactivated (n = 7) prior to the conditioning phase rats did not differ from those infused
with SAL (n = 11), as illustrated by a non-significant main effect of Treatment
(F(1,16)=0.08,p>0.77), as well as a non-significant CS Type x Treatment interaction
(F(1,16)=0.13,p>0.72) (Fig. 2A). A significant main effect of CS Type (F(1,16)=57.21,p<0.001),
indicated that both groups discriminated between the CS+ and CS- accurately during the
expression test, however there was no CS Type x Treatment interaction (F(1,16)=0.13,p>0.72).
Additionally, there was no CS Number x Treatment interaction (F(3,48)=2.36,p>0.08). Neither
the overall number of lever-presses made throughout the test session (t(16)=0.94,p>0.36), nor the

41
rate of lever-pressing during the first 5 min of the expression test were affected by Treatment
(t(16)=0.48,p>0.63) (Table 2). Similarly, overall locomotor activity during the test day did not
differ as a result of Treatment (t(16)=-0.46,p>0.65) (Table 2). Thus, PL cortex activity was not
necessary for the appropriate acquisition of discriminative fear conditioning, and did not impact
general indices of motivated behavior.
2.3.2 IL cortex inactivation pre-conditioning
Temporary inactivation of IL cortex immediately prior to conditioning had no effect on the
change in locomotion in response to CS- or CS+ presentations during the conditioning phase.
The change in locomotor activity was nearly identical across Treatment conditions
(F(1,23)=0.001,p>0.99), and both groups expressed a greater change in locomotion during CS+
presentations, as compared to CS- presentations, indicated by a main effect of CS Type
(F(1,21)=11.57,p<0.005), and no CS Type x Treatment interaction (F(1,21)=0.77,p>0.39) (Table
1). The overall level of locomotion during the conditioning session did not differ between saline
and IL-inactivated groups (t(21)=-0.46,p>0.65) (Table 1). This pattern of results suggests that IL
inactivation does not affect within-session changes gross locomotor output, or the CS-specific
modulation of behavior.
When tested drug-free during the fear expression test session, control rats (n = 10)
expressed similar levels of discriminative conditioned suppression as did rats that underwent IL
inactivation (n = 13) prior to the conditioning session (Fig. 2B). Although there was a significant
effect of CS Type (F(1,21)=75.37,p<0.001), there was no main effect of Treatment,
(F(1,21)=0.17,p>0.68), and no CS Type x Treatment interaction, (F(1,21)=0.66,p>0.42),
indicative of intact conditioned suppression. There was no significant CS Type x CS Number
interaction, and no three-way interaction, (all F-values < 1.2, all p-values > 0.33). No change

42
was observed in locomotor activity throughout the session, (t(21)=-0.15,p>0.88), or the rate of
pressing during the first 5 min of the test session, (t(21)=-0.10,p>0.92), as a function of treatment
(Table 2). Finally, the number of presses made during the entirety of the expression test was not
different in IL-inactivated animals, as compared to controls (t(21)=0.15,p>0.88) (Table 2). Like
the PL cortex, IL activity during fear conditioning acquisition was not necessary for the
appropriate expression of discriminative conditioned suppression during the test session.
2.3.3 NAcS inactivation pre-conditioning
During conditioning, control and NAcS-inactivated rats made similar CS-induced changes in
locomotor activity. There was no main effect of Treatment (F(1,20)=0.004,p>0.94), suggesting
that locomotor activity was comparable across drug conditions, and there was no CS Type x
Treatment interaction (F(1,20)=2.24,p>0.15) (Table 1). On average, rats locomoted more during
CS+ presentations than during CS- presentations, regardless of inactivation status, as evidenced
by a significant main effect of CS Type (F(1,20)=6.91,p<0.02). Consistent with this, there was
no change in the amount of locomotion across the entire session (t(20)=-0.89,p>0.38) (Table 1).
Thus, gross locomotor and cue-induced locomotor activity during the conditioning session were
comparable across treatment conditions, with both groups appearing to acquire the CS
associations without issue.
Interestingly, inactivation of NAcS (n = 11) during the conditioning session diminished
lever-press suppression during the subsequent drug-free expression test, as compared to SAL
animals (n = 11) (Fig. 3A). A main effect of Treatment (F(1,20)=7.55,p<0.02), suggested that
response inhibition produced by CS presentation was decreased as a result of NAcS inactivation
during conditioning. Despite the overall decrease in suppression during CS presentations, rats in
the inactivation group still discriminated accurately between the CS- and CS+, as illustrated by a

43
significant main effect of CS Type (F(1,20)=109.99,p<0.001), but no significant CS Type x
Treatment interaction (F(1,20)=2.60,p>0.12). Although there was no three-way interaction
(F(3,60)=0.98,p>0.40), inspection of the data suggested that suppression allocated towards the
CS+ was particularly diminished by inactivation, indicative of a decrease in strength of the fear
memory. Neither the rate of lever pressing prior to the first CS presentation (t(20)=0.15,p>0.88),
nor the total number of lever-presses made during the expression test session (t(20)=0.22,p>0.83)
were altered by treatment (Table 2). Finally, there was no evidence that inactivation of the NAcS
prior to conditioning impacted locomotor activity during the expression test (t(20)=0.62,p>0.55)
(Table 2). Taken altogether, these results imply that the amount of suppression produced during
the expression test session is reduced by inactivation prior to conditioning, suggesting that the
fear memory established during learning is less enduring, and more labile. The subsequent
impact of this memory on behavior is thus less pronounced, resulting in less conditioned
suppression.
2.3.4 NAcC inactivation pre-conditioning
Inactivation of NAcC immediately prior to the conditioning session slightly altered locomotor
activity during CS presentations. CS presentations (collapsed across CS+ and CS-) tended to
cause less of a change in locomotion in NAcC-inactivated animals than it did in control animals,
as shown by a trend level Treatment effect (F(1,20)=4.22,p=0.053). Consistent with this, overall
locomotor activity throughout the session was decreased in NAcC-inactivated rats
(t(20)=3.06,p<0.007) (Table 1). Still, both treatment groups had a greater increase in locomotion
during CS+ presentations, as compared to CS- presentations, as shown by a significant main
effect of CS Type (F(1,20)=14.36,p<0.002), but no CS Type x Treatment interaction
(F(1,20)=0.02,p>0.88) (Table 1). These findings suggest that, despite NAcC animals being less

44
active, they behaviorally distinguished between each CS type, as measured by their change in
locomotor activity.
On the drug-free expression test day, rats that received NAcC inactivation (n = 10) prior
to conditioning performed similarly to those that received saline (n = 12) (Fig. 3B). The overall
level of conditioned suppression was comparable across treatment conditions
(F(1,20)=0.14,p>0.71), and both groups distinguished between the CS+ and CS- in a similar
manner, as shown by a non-significant CS Type x Treatment interaction (F(1,20)=0.003,p>0.95),
and a significant effect of CS Type (F(1,20)=79.72,p<0.001). Additionally, there was no three-
way interaction (F(3,60)=0.33,p>0.80). The total number of presses made throughout the session
was unchanged by NAcC inactivation (t(20)=-1.62,p>0.12), as was the total amount of
locomotor activity (t(20)=-1.38,p>0.18) (Table 2). Finally, the rate of lever-pressing during the
first 5 min of the test session was unchanged by previous inactivation of the NAcC (t(20)=-
1.21,p>0.24) (Table 2). Like the prefrontal cortex, NAcC activity during conditioning was not
necessary for the subsequent expression of discriminative fear conditioning.
2.3.5 PL cortex inactivation pre-expression test
In contrast to the null effect of pre-conditioning inactivation, PL cortex activity proved necessary
for the appropriate expression of conditioned suppression during the test session (Fig. 4A). These
animals were given discriminative conditioning in a drug-free state, and then subjected to
inactivation of the PL (n = 13) or saline (n = 12) infusion immediately prior to the expression test
session. Here, a significant main effect of Treatment (F(1,23)=13.09,p<0.001), was observed,
suggesting that PL inactivation altered conditioned suppression. This was accompanied by a CS
Type x Treatment interaction (F(1,23)=11.68,p<0.005), with simple main effects analysis
showing that that control rats expressed more fear during the CS+ than the CS-

45
(F(1,11)=26.43,p<0.001), while PL inactivated rats did not (F(1,12)=0.42,p>0.53). There was no
CS Number x Treatment interaction (F(3,69)=1.37,p>0.25). Additionally, there was no change in
the rate of lever-pressing prior to the first CS presentation (t(23)=0.32,p>0.75), suggesting that
the disinhibition of pressing during the CS+ in PL-inactivated animals was not a result of general
behavioral activation (Table 2). Further supporting this, locomotor activity throughout the
session was not altered by PL inactivation (t(23)=-1.76,p>0.09), nor was the total number of
lever-presses made during the session (t(23)=1.28,p>0.20) (Table 2). Thus, PL activity was
necessary for the appropriate expression of fear towards a discriminative CS+, with inactivation
markedly reducing the suppression of activity typically observed during its presentation.
2.3.6 IL cortex inactivation pre-expression test
Temporary inactivation of IL prior to the expression test session had a qualitatively similar effect
on discriminative conditioned suppression than did inactivation of the more dorsal PL cortex
(Fig. 4B). A main effect of Treatment was observed (F(1,20)=5.60,p<0.03), suggesting that the
overall level of suppression across both tone types was lower in IL inactivated rats (n = 12), as
compared to controls (n = 10). However, unlike PL cortex, there was no significant CS Type x
Treatment interaction (F(1,20)=0.16,p>0.69). There was a significant main effect of CS Type
(F(1,20)=4.92,p<0.04), suggesting that, collapsed across treatment conditions, presentation of
the CS+ caused more suppression than did presentation of the CS-. As with PL inactivation,
there was no significant there-way interaction (F(3,60)=0.95,p>0.42). Locomotion was
unchanged following IL inactivation (t(20)=0.75,p>0.46), as was the rate of lever-pressing
during the first 5 min of the session (t(20)=-0.09,p>0.92), and the total number of lever-presses
made (t(20)=-0.74,p>0.46), suggesting that the impact of IL inactivation was specific to

46
behavioral suppression induced by the conditioned stimuli, and not a general effect of behavioral
disinhibition (Table 2).
As the decrease in conditioned suppression following IL manipulation was unexpected,
we chose to perform a control experiment aimed at determining whether the suppression-
reducing impact of IL inactivation was specific to a discriminative context. When a separate
group of rats underwent fear conditioning using a single CS, IL inactivation (n = 8) did not have
a significant effect on conditioned suppression, as compared to control animals (n = 8) (Fig. 5B).
There was no main effect of Treatment (F(1,14)=1.65,p>0.22), with both groups extinguishing at
a comparable rate as indicated by a significant effect of CS Block (F(5,70)=10.02,p<0.001), but
no significant Treatment x CS Block interaction (F(5,70)=1.57,p>0.18). Although the rate of
pressing at the beginning of the session was the same regardless of Treatment
(t(14)=0.14,p>0.89), inactivated animals made more lever presses throughout the session
(t(14)=2.84,p<0.013) (Table 2). However, treatment had no impact on overall locomotor activity
(t(14)=0.17,p>0.87) (Table 2). This pattern of results suggests that conditioned suppression was
not significantly altered by IL inactivation when assessed using a single stimulus, which
contrasts with the significant reduction of conditioned suppression observed in the discriminative
context. This effect may be mediated in part by a general disinhibition of lever-pressing that
appears to have occurred throughout the session, as evidenced by the elevated number of lever
presses made by the IL inactivated animals.
2.3.7 NAcS inactivation pre-expression test
Like the PL cortex, inactivation of NAcS (n = 13) eliminated the appropriate expression of
discriminative conditioned suppression, as compared to control rats (n = 14) (Fig. 6A). There
was a significant CS Type x Treatment interaction (F(1,25)=5.02,p<0.035), indicative of a

47
differential pattern of fear expression induced by NAcS inactivation, as compared to control rats.
This was driven by less suppression during presentation of the CS+ for animals in the NAcS
inactivation group (F(1,25)=4.24,p=0.05). In contrast, lever-pressing during the CS- did not
change as a function of treatment, (F(1,25)=0.20,p>0.66). There were no other significant two-
way interactions, and no significant three-way interaction (all F-values < 1.3, all p-values >
0.25). NAcS inactivation did not alter the total number of lever-presses made during the session
(t(25)=-1.18,p>0.24), nor the rate of pressing during the initial portion of the session
(t(25)=0.15,p>0.88) (Table 2). Similarly, there was no change in overall locomotion during the
expression test session (t(25)=-1.21,p>0.23) (Table 2). Thus, the NAcS can be shown to play a
relatively specific role in producing response-suppression during the presentation of a potentially
aversive CS+, without affecting non-specific indices of motivation such as total lever-press rate
or locomotion.
2.3.8 NAcC inactivation pre-expression test
In contrast to the disinhibitory impact of NAcS inactivation, the same manipulation of the NAcC
had no impact on fear expression (Fig. 6B). Following inactivation of NAcC (n = 9) prior to the
expression test, no main effect of Treatment was observed (F(1,19)=0.05,p>0.84), indicating that
these animals expressed levels of fear comparable to control rats (n = 12). A main effect of CS
Type (F(1,19)=102.36,p<0.001), combined with no CS Type x Treatment interaction
(F(1,19)=0.54,p>0.47), suggested that animals discriminated between the CS- and CS+
regardless of treatment condition. Additionally, there was no significant three-way interaction
(F(3,57)=0.80,p>0.50). Despite the lack of overt effect on suppression during each CS
presentations, the overall number of lever-presses was decreased in NAcC-inactivated animals
(t(19)=2.23,p<0.04), although the rate of lever-pressing during the first five min of the session

48
was not significantly different from control animals (t(19)=1.74,p>0.09) (Table 2). Similarly,
NAcC inactivation decreased locomotion (t(19)=2.80, p<0.02) (Table 2). These results suggest
that NAcC promotes behavioral activation, without a particular role in modulating actions based
on cues predicting safety or an aversive consequence.
2.4 Discussion
Using pharmacological inactivations, we showed that separate subregions of the PFC and NAc
uniquely contribute to the acquisition and expression of discriminative Pavlovian fear, as
measured by conditioned suppression. Under control conditions, presentation of an aversive CS+
in the absence of foot-shock caused a marked suppression of ongoing reward-seeking, while
presentation of a neutral CS- did not alter behavior. Although neither subregion of the mPFC was
necessary for the acquisition of discriminative fear, both subregions regulated the expression of
acquired suppression, in keeping with a top-down, inhibitory function of the mPFC. The
involvement of the NAc, a striatal structure known to integrate cortico-limbic input during
response-selection, was dependent on the particular subregion targeted. Inactivation of the NAcC
left the acquisition and expression of conditioned suppression intact, but tended to diminish
indices of behavioral activation, including locomotion and total lever-presses. In contrast,
inactivation of the NAcS diminished conditioned suppression regardless of whether the
manipulation was conducted prior to the acquisition or expression phase of the task, implicating
this structure in the plasticity associated with fear acquisition, as well as the activity necessary
for response-inhibition during subsequent expression.
2.4.1 Discriminative fear acquisition: Prefrontal and accumbal contributions
Of the prefrontal and accumbal subregions tested, only the NAcS was necessary to
acquire normal levels of suppression towards a discriminative conditioned stimulus, when

49
assessed during a later expression test. Importantly, none of the regions tested affected the
change in locomotion induced by CS- and CS+ presentations during conditioning, indicating that
all animals maintained the ability to discriminate between the two conditioned stimuli.
Furthermore, locomotion in response to the CS+ in part reflects the burst of activity induced by
US delivery, suggesting that unconditioned responses to the foot-shock were not altered by
regional inactivations. The finding that the prefrontal cortex is not required for the acquisition of
conditioned fear to a CS+ is in keeping with much previous research using single stimulus,
Pavlovian designs, and assessing freezing. For example, inactivations or lesions of PL cortex
leaves the acquisition of conditioned fear intact (Corcoran & Quirk, 2007; Morgan, Romanski, &
LeDoux, 1993; Quirk et al., 2000). The complementary finding reported here that IL cortex is
also not involved in the acquisition of conditioned fear is more novel. Still, this result is
consistent with the study of a similar defensive behavior, the acquisition of conditioned place
aversion following intraplantar formalin injection, which is not affected by IL manipulation
(Jiang et al., 2015). These results point to the involvement of other regions in the plasticity
associated with fear learning. They also suggest that, when examining fear acquisition, the
irrelevance of mPFC activity is comparable to that when examining other defensive reactions,
such as freezing.
Despite this apparent lack of necessity during fear acquisition, it is important to recognize
that electrophysiological signatures related to fear discrimination learning have been observed in
mPFC (Laviolette, Lipski, & Grace, 2005; Orona & Gabriel, 1983). During conditioning, the
frequency and burst firing of mPFC neurons increases in response to CS+, but not CS-,
presentations (Laviolette et al., 2005). In addition, dopaminergic modulation of the PL cortex
during the acquisition of discriminative fear conditioning is capable of altering subsequent neural

50
and defensive responses during presentations of an olfactory CS+ and CS- (Lauzon, Ahmad, &
Laviolette, 2012; Lauzon, Bishop, & Laviolette, 2009; Laviolette et al., 2005). Laviolette and
colleagues have suggested that dopamine-receptor mediated activity of calcium calmodulin-
dependent kinase II (CaMKII), a protein critical for memory formation, can bias the salience of
the CS+/CS- depending on the strength of the conditioning procedure (Lauzon et al., 2012, 2009;
Laviolette et al., 2005). Here, we utilized temporary inactivations that served to hyperpolarize
affected neurons, an effect that fundamentally differs from the targeting of specific dopamine-
receptor subtypes, and generally does not impact fear acquisition (Corcoran & Quirk, 2007).
Manipulation of specific neuromodulatory targets within the mPFC may alter salience encoding
in a fashion that does not directly depend upon changes in neuronal excitability, although the
parameters under which this is the case remain to be identified.
Like the two prefrontal subregions tested, NAcC inactivation had no impact on
discriminative fear acquisition. This nucleus may be particularly relevant for contextual, but not
cued, fear conditioning (Levita, Dalley, & Robbins, 2002; Wendler et al., 2013). Yet, one
previous study assessing the conditioned suppression of licking has implicated the NAcC in the
formation of a fear memory in response to a discrete cue (Parkinson et al., 1999). Important
methodological differences may explain this apparent discrepancy. First, Parkinson and
colleagues (1999) utilized permanent lesions, which may impact other processes related to the
acquisition of fear or the instrumental licking behavior. In addition, these researchers employed a
trace conditioning protocol, which involves a short delay between the delivery of the CS and US.
Trace conditioning contrasts with the delay conditioning (CS co-terminates with the US) method
employed here, and has been suggested to rely on partially segregated circuitry (Raybuck &
Lattal, 2014), for example requiring activity within the PL cortex during acquisition (Gilmartin

51
& McEchron, 2005; Gilmartin, Miyawaki, Helmstetter, & Diba, 2013). As neither PL nor NAcC
are generally necessary for the acquisition of delay fear conditioning, the results of Parkinson
and colleagues (1999) may relate to the presence of the trace interval between CS presentation
and US delivery. Still, the present results continue to support an account of the NAcC in
promoting behavioral activation, as inactivation of this nucleus decreased locomotor activity
within the fear conditioning session.
Surprisingly, NAcS inactivation during fear acquisition diminished the subsequent
expression of conditioned suppression. Although these rats maintained the ability to discriminate
between the CS+ and CS-, overall suppression was lower as a result of NAcS inactivation during
learning. At first glance, this result appears to contradict previous findings suggesting that the
NAcS is not a critical structure for the acquisition of cued fear in rodents (Jongen-Rêlo,
Kaufmann, & Feldon, 2003; Parkinson et al., 1999; Riedel et al., 1997). Despite these previous
null findings, the NAcS has been shown to control fear learning in some situations, such as when
learning a new fear association in the presence of an already established fear-predictive cue
(Bradfield & McNally, 2010). In the present design, animals must form two divergent
associations during the conditioning phase, one between CS+ and foot-shock, and one between
the CS- and nothing. If the NAcS is necessary for updating fear based upon the status of
individual cues as fear-predictors, eliminating activity in this subnuclei could subsequently alter
the fear expressed towards the CS+ versus CS-. In addition, electrophysiological signatures of
discriminative fear learning have been reported to occur in the NAcS. Neurons projecting from
mPFC to the NAc (mostly NAcS) encode the aversive nature of an olfactory CS+ (but not a CS-
), in a BLA-dependent manner (McGinty & Grace, 2008).

52
In addition, most of the previous studies examining the contribution of the NAcS to fear
learning have used a single, discrete stimulus and measured freezing as their dependent measure
of fear. Comparison of the present study with these archival reports suggests that the circuitry
relevant for freezing may diverge from those necessary for conditioned suppression when
considering the involvement of the NAc. Although speculative, as we did not measure freezing
in the present study, this dissociation would be in keeping with the role of the NAc in controlling
motivated behavior as a function of affective input (Mogenson et al., 1980). Unlike freezing,
conditioned suppression requires the integration of affective information with a competing drive
(instrumental action leading to reinforcement), which may induce a state of motivational conflict
that could require activity within prefrontal and striatal structures (Friedman et al., 2015; Kim et
al., 2017; Resstel et al., 2008).
Given that the effect of NAcS inactivation prior to conditioning was observed during a
later fear expression test, fear conditioning may induce plasticity within the NAcS as a result of
input from efferent regions that encode fear conditioning. One candidate afferent region is the
BLA, which projects monosynaptically to the NAcS (Kita & Kitai, 1990; Phillipson & Griffiths,
1985; Wright et al., 1996), and is critical for the encoding of fear conditioning (Fanselow &
LeDoux, 1999). The projection from BLA-NAcS has been shown to mediate related aspects of
aversive-motivation, including the consolidation of inhibitory avoidance, an assay of passive
defensive behavior similar to conditioned suppression (LaLumiere, Nawar, & McGaugh, 2005),
as well as the performance of active avoidance, a defensive action employed to remove a
potentially aversive stimulus (Ramirez et al., 2015). At the molecular level, foot-shock induces
cAMP response element binding protein (CREB) expression in the NAcS, which has been shown
to subsequently decreases motivation and impair the extinction of conditioned fear (Muschamp

53
et al., 2011). A similar induction of CREB occurs during fear conditioning within the lateral
segment of the amygdala (Yiu et al., 2014), suggesting a potentially common mechanism for
fear-ensemble formation during aversive learning. Here, inactivation may prevent the plasticity
associated with CREB expression in this nucleus during discriminative fear acquisition, altering
the expression of conditioned suppression, and accelerating within-session extinction during the
test session.
2.4.2 Discriminative fear expression: Prefrontal and accumbal contributions
Separate animals were tested to examine subregional contributions to the expression of
discriminative conditioned suppression. In these experiments, we observed that inactivation of
either prefrontal subregion disinhibited lever-pressing during CS+ presentations, indicative of a
loss of conditioned fear. The observation that the PL mPFC acts to promote Pavlovian fear
during an expression test, as illustrated here, is concordant with previous literature (Corcoran &
Quirk, 2007; Limpens et al., 2015; Piantadosi & Floresco, 2014; Sangha, Robinson, Greba,
Davies, & Howland, 2014; Sierra-Mercado et al., 2011; Vidal-Gonzalez et al., 2006).
Inactivation of PL prior to an expression test session resulted in rats engaging in lever-pressing,
despite the impending threat posed by the CS+. This effect was apparent from the first CS+
presentation, implying that this alteration was not a product of accelerated extinction. Although
the result of this behavioral change was a loss of discriminative conditioned suppression, this
effect was driven entirely by a loss of fear towards the aversive cue, suggesting that the
irrelevance associated with the neutral CS- remained intact. A model of PL cortex function
during the early stages of fear expression and extinction posits that activity within this subregion
promotes the expression of defensive reactions such as freezing and conditioned suppression
(Pendyam et al., 2013; Sierra-Mercado et al., 2011). Given that the expression of freezing is

54
incompatible with lever-pressing, it is possible that a decrease in freezing explains in part the
loss of conditioned suppression. Some evidence against this suggestion comes from our
assessment of locomotion, which was not altered by PL inactivation. If PL-inactivated animals
froze significantly less than their control counterparts, locomotion may be expected to be higher,
which was not the case. In keeping with a particular role in conditioned suppression, PL cortex
has been shown to regulate aversion-induced response-inhibition when seeking cocaine (Chen et
al., 2013; Limpens et al., 2015) or alcohol (Seif et al., 2013). Similarly, PL (and potentially IL)
cortex appear to mediate the response-inhibition enforced during periods of learned cocaine
unavailability (Gutman, Ewald, Cosme, Worth, & Lalumiere, 2014; Mihindou, Guillem,
Navailles, Vouillac, & Ahmed, 2013). The present study supports these findings, and illustrates
that the fear promoting aspect of the PL cortex is specific to a CS+ in a discriminative context.
Unlike the unambiguous parallel between the findings of this study and previous studies
regarding the PL cortex and fear expression, our observation that IL cortex inactivation
decreased conditioned suppression is somewhat surprising. One critical consideration regarding
this result is the overall lower level of suppression observed following pre-test IL manipulation
(Fig. 4B), when compared to the same manipulation of the more dorsal PL cortex (Fig. 4A). One
possible explanation for this difference is that, because our IL cannula were not lowered at an
angle during surgery, damage caused to the overlying PL cortex diminished fear expression
(Sierra-Mercado et al., 2011). However, this explanation is unlikely, as surgery was conducted in
an identical fashion for animals in the pre-conditioning infusion experiments (Fig. 2A and B),
where control levels of fear were comparable across prefrontal subregions. Moreover, animals
used in the pre-test PL versus IL experiments did not differ in other measures that could have
potentially contributed to the difference in baseline conditioned suppression, such as locomotion,

55
overall lever pressing, or the rate of lever pressing (Table 2). Thus, the diminished overall
conditioned suppression observed in the IL-manipulated (as compared to PL-manipulated)
animals must relate to the infusion being conducted immediately (10 min) prior to the test
session. A review of previous studies manipulating IL cortex function during fear expression or
extinction shows that animals are typically tested upwards of 30-45 min from the time of
infusion, which may abrogate such technical confounds (Akirav et al., 2006; Bravo-Rivera,
Roman-Ortiz, Brignoni-Perez, Sotres-Bayon, & Quirk, 2014; Sierra-Mercado et al., 2011). Here,
animals were tested 10 min post-infusion to maintain both internal and external consistency, as
we have utilized this approach without observing such baseline differences (e.g., Dalton, Wang,
Phillips, & Floresco, 2016; Piantadosi & Floresco, 2014; Stopper & Floresco, 2011). However,
this may have artificially reduced the amount of conditioned suppression, even under control
conditions.
Despite this caveat, we observed a further significant reduction in conditioned
suppression induced by inactivation of the IL, as compared to control rats. Using a single-
stimulus approach, Sierra-Mercado, Quirk and colleagues (2011) have shown that
pharmacological inactivation of IL prolongs conditioned freezing, an effect opposite to that of
PL cortex inactivation. Conversely, stimulation of this region has been shown to decrease fear,
enhancing extinction either within-session or across sessions (Bukalo et al., 2015; Milad et al.,
2004; Vidal-Gonzalez et al., 2006). Here we were interested in the acute impact of each region
on fear expression, and did not formally examine the possibility that IL manipulation may affect
between-session extinction, which has been shown to depend on IL projections to the amygdala
(Bukalo et al., 2015; Do-Monte, Manzano-Nieves, Quinones-Laracuente, Ramos-Medina, &
Quirk, 2015). In these previous experiments, conditioned freezing served as the primary

56
dependent measure of fear. Limited experimental evidence suggests that the expression of
conditioned suppression, unlike conditioned freezing, is either decreased or not affected by IL
inactivation (Jean-Richard-Dit-Bressel & McNally, 2016; Resstel et al., 2008; Sierra-Mercado et
al., 2011). For example, inactivation of either PL or IL cortex reduces lever-press suppression
induced by instrumental punishment (Resstel et al., 2008), a qualitatively similar effect to that
observed here. In addition, a pronounced strain difference in the expression of defensive
reactions following IL manipulation has been reported. Lesions of the IL cortex in Long Evans
rats (as used here) did not affect freezing in response to a CS+, while the same manipulation in
Sprague Dawley rats (used in most previous studies of IL function) abnormally elevated the level
of conditioned freezing, delaying extinction (Chang & Maren, 2010). Thus, differences in fear
expression circuitry across inbred and outbred rat strains, as well as fundamental differences
between the regions necessary for particular defensive reactions, may explain the lack of
consistency between the function of the IL observed here and in previous studies.
Although these explanations may shed light on why PL and IL function are not always
dissociable, they beg the question as to why we observed an effect, a decrease in conditioned
suppression expression following discriminative fear conditioning, of IL inactivation at all. In
fact, when we conducted a single-stimulus assessment of conditioned suppression, IL
inactivation did not significantly impact fear expression (Fig. 5). These data suggest that the
comparable function of PL and IL observed here may additionally relate to the discriminative
nature of our task. In support of this, Sangha, Howland and colleagues (2014) have shown that
these subregions are not functionally dissociable during performance of a similar Pavlovian
discriminative task. In their study, inactivation of PL or IL altered discriminative fear expression
in the same manner, decreasing conditioned freezing during the presentation of an aversive cue,

57
while leaving intact the ability of a neutral, safe cue to ameliorate fear (Sangha et al., 2014).
Thus, IL cortex may also promote fear during situations that produce a conflict between
representations evoked by stimuli encoding safety and fear.
Within the NAc, only the NAcS was relevant for fear expression, with inactivation
decreasing conditioned suppression in a manner similar to the PL cortex. NAcC-inactivated rats
had no difficulty discriminating between the CS+ and CS-, expressing levels of fear and
indifference comparable to control rats. NAcC manipulation was not entirely without effect, as
inactivation resulted in rats locomoting less and performing fewer lever-presses than control rats,
although their rates of lever-pressing were comparable (Table 2). Such an effect is consistent
with previous reports from our and other laboratories suggesting that this nucleus is involved in
the invigoration of behavior (Ghods-Sharifi & Floresco, 2010; Nicola, 2010; Stopper & Floresco,
2011). Outside of the aversive domain, the NAcC is known to be involved in the ability of an
appetitive Pavlovian conditioned stimuli to invigorate behavior (Ambroggi et al., 2011;
Parkinson, Willoughby, Robbins, & Everitt, 2000; Yun et al., 2004). For example, activity and
dopamine release within this nucleus is necessary for a cue predicting reward availability to
efficiently promote instrumental reinforcement-seeking (Ambroggi et al., 2011; McGinty,
Lardeux, Taha, Kim, & Nicola, 2013; Nicola, 2010). Here, NAcC activity was not required for
essentially the opposite pattern of behavior, the inhibition of reinforcement-seeking by an
aversive Pavlovian conditioned stimulus. Thus, the mechanisms through which the NAc
modulate behavior may be biased towards response-promotion, instead of response-inhibition.
On the other hand, NAcS activity proved necessary for rats to appropriately suppress
reinforcement-seeking during the presentation of an aversive Pavlovian CS+. This was not the
result of general behavioral disinhibition, as indices of general behavioral activation, including

58
the rate of pressing early in the session, the total number of lever-presses made, and locomotor
activity were not different than in controls (Table 2). This dissociation points to a more nuanced
role for the NAcS, whereby instrumental reward-seeking is impacted specifically by a Pavlovian
stimulus previously associated with a negative event. Outside of the negative valence, the NAcS
has been suggested to fulfil an inhibitory function during extinction learning for reinforcers
including food (Floresco et al., 2008), alcohol (Millan et al., 2010), and cocaine (Peters et al.,
2008). Similarly, refining behavior through the learned cessation of instrumental responding
during periods of reward unavailability or non-reinforcement is believed to be mediated by an
inhibitory NAcS function (Ambroggi et al., 2011; Blaiss & Janak, 2009; Floresco et al., 2008).
Populations of neurons that encode task-irrelevant stimuli and behaviors during reward-seeking
are more numerous in the NAcS, as compared to the NAcC (Ambroggi et al., 2011), which may
provide a neuronal mechanism for the NAcS-specific impact on fear-induced response-
inhibition. Moreover, the NAcS is necessary for Pavlovian cues to invigorate instrumental
behavior, as assessed by the Pavlovian-to-instrumental transfer (PIT) effect (Corbit & Balleine,
2011; Corbit et al., 2001). Conditioned suppression, which has been described as an aversively-
motivated form of PIT (Cardinal et al., 2002), may also depend on this subregions of the NAc.
Given the dense projection from ventromedial PFC, including the ventral PL and IL
cortex, to the NAcS (Brog et al., 1993; Sesack et al., 1989; Vertes, 2004), it is important to
comment on the qualitative similarity between each region’s effect on conditioned suppression.
Based on the present results, the native role for PL cortex during conditioned suppression
appears to promote the top-down inhibition of seeking behavior under threat. NAcS may
function in a similar manner, although the time-course of inhibition may be somewhat distinct,
given that behavioral disinhibition was apparent from the first stimulus presentation following

59
PL inactivation, while the effect of NAcS inactivation did not appear until after the first CS+
delivery (all trials conducted in extinction). It is possible that this function of the mPFC is
mediated by its projection to downstream targets, including the NAcS. This hypothesis is
supported by previous work suggesting that some neurons projecting from mPFC to NAc
(mostly terminating within the NAcS) encode the behavioral relevance of an aversive CS+ and a
neutral CS- in a BLA-dependent manner (McGinty & Grace, 2008). Similarly, a recent study
identified a microcircuit originating in the mPFC and terminating in the lateral NAcS that
promotes suppression following foot-shock (Kim et al., 2017). Activity within this circuit
decreased when animals made a seeking response during risk of foot-shock, and activation of
this projection inhibited such behavior. Although we targeted the medial NAcS, it is possible that
homologous functions are controlled by these topographically adjacent areas. Pharmacological
disconnection of these two structures would allow for this hypothesis to be tested.
2.4.3 Relevance to fear circuitry in humans, and psychiatric populations
Here, we utilized a discriminative fear conditioning design that is similar to those employed in a
translational setting, where CS- presentations serve as a baseline index of fear, and CS+
presentations induce fear. Using such designs, a relatively conserved fear circuit encompassing
the amygdala, prefrontal cortex, and ventral striatum has been identified in the human brain (for
review, see Adolphs, 2013; Delgado, Nearing, et al., 2008; Milad & Quirk, 2012; Peters et al.,
2009). Within the PFC, the dorsal anterior cingulate cortex (dACC; BA32) and ventromedial
PFC (vmPFC; BA25) have been suggested to be functionally and anatomically homologous to
the rodent PL and IL cortex, respectively (Heilbronner et al., 2016; Milad & Quirk, 2012).
Activity in the dACC occurs in response to CS+ presentations, and this activity (as well as the
overall thickness of the region) correlates positively with physiological measures of fear in

60
humans (Milad, Quirk, et al., 2007). On the other hand, vmPFC activity appears to track
extinction learning in humans, as this region displays patterns of activity consistent with
deactivation during conditioning, but activation during extinction (Milad, Wright, et al., 2007;
Phelps, Delgado, Nearing, & Ledoux, 2004). Here, we provide tentative support for the dACC-
PL homology suggested by these previous studies, as they apply to the expression of conditioned
fear. However, our results seem to suggest that IL cortex performs a similar function, promoting
conditioned suppression, in a manner inconsistent with human vmPFC activity. This may again
stem from the nature of the defensive reaction measured, as freezing (in rats) and skin
conductance or verbal scoring (in humans) do not produce a state of motivational conflict similar
to that induced by the conditioned suppression of reinforcement-seeking. Although conditioned
suppression paradigms exist in humans (Allcoat, Greville, Newton, & Dymond, 2015; Greville,
Newton, Roche, & Dymond, 2013), to date, the relevant functional imaging studies have not
been performed to evaluate this hypothesis.
In addition to prefrontal homology, discriminative aversive conditioning produces
activity in the ventral striatum of humans (Delgado et al., 2009; Delgado, Li, et al., 2008;
Delgado, Nearing, et al., 2008; Jensen et al., 2003; Klucken et al., 2009; Pohlack et al., 2012).
This activity is generally differential, with activity increasing in response to a CS+ to a greater
degree than a CS-, a pattern which develops over the course of the conditioning session (Klucken
et al., 2009). In addition, activity in this nucleus has been shown to translate fear into motivated
action, as learning to avoid an aversive CS+ also recruits the NAc (Delgado et al., 2009). In the
present study, NAcS activity was necessary for the appropriate acquisition and expression of
discriminative conditioned suppression. Thus, it is possible that the NAc activity observed in
human imaging studies of fear learning may reflect preferential activation of the NAcS.

61
Interestingly, only one study has examined subnuclei of the NAc in humans. In this study,
diffusion tractography was used to differentiate the NAcS and NAcC in the human brain, with
results indicating that the putative NAcS responds in anticipation of thermal pain, while NAcC
responds particularly to the offset of a painful stimulus (Baliki et al., 2013). Whether this
anticipatory activity relates to behavior is currently unknown, but may partially explain the
anticipatory activity observed in NAc prior to presentation of a conditioned aversive stimulus
(Jensen et al., 2003).
A number of neuropsychiatric disorders are characterized by the maladaptive influence of
affect on decision-making processes. Meta-analytic studies have consistently shown that patients
with anxiety disorders express more fear to a CS- than do control individuals (Duits, Cath,
Lissek, Hox, Hamm, Engelhard, Van Den Hout, et al., 2015; Lissek et al., 2005). This deficit
may be related to aberrant function of prefrontal circuitry, as trait anxiety is associated with
diminished coupling between the amygdala and the vmPFC and a heightened coupling between
the amygdala and the dorsomedial PFC, patterns that were opposite that observed in healthy
comparison subjects (Kim, Gee, Loucks, Davis, & Whalen, 2011). Specifically, vmPFC activity
is negatively modulated by similarity to a CS+, while dorsomedial PFC activity is positively
modulated by the CS+ similarity. This effect has recently been reported to be disturbed in
individuals with PTSD, suggesting that imbalanced prefrontal discrimination mechanisms may
contribute to anxiety (Kaczkurkin et al., 2017). In the present study, the fear expressed towards a
CS- was normal regardless of treatment. Thus, other regions, such as the BLA, which has been
shown to encode the valence of discriminative stimuli in rats, non-human primates, and humans
(Genud-Gabai, Klavir, & Paz, 2013; McHugh et al., 2013; Sangha, Chadick, & Janak, 2013;
Schiller, Levy, Niv, LeDoux, & Phelps, 2008), may be causally-related to fear generalization.

62
In addition, prefrontal hypofunction appears to be related to inhibitory control deficits in
substance abuse (for review, see Goldstein & Volkow, 2011). In cocaine users, deficits in
inhibitory control are known to correlate with reduced dACC activity, the same region suggested
to promote fear expression previously (Goldstein et al., 2009; Hester & Garavan, 2004;
Kaufman, Ross, Stein, & Garavan, 2003; Li et al., 2008). In rats, hypofunction of the
functionally homologous PL cortex recapitulates key aspects of addictive behavior, including
seeking drug under threat of punishment (Chen et al., 2013; Limpens et al., 2015). Such a deficit
may be related to the loss of response-inhibitory function within the PL or dACC, as a function
of addiction progression. Moreover, obsessive-compulsive disorder is characterized by aberrant
cortico-striatal connectivity, which centers around projections from the orbitofrontal cortex to the
ventral striatum (Figee et al., 2016; Wood & Ahmari, 2015). While involving a partially
overlapping circuit, hyperactivity (not hypoactivity) of this orbitofrontal-ventral striatal
projection appears to mediate compulsive aspects of obsessive-compulsive disorder (Ahmari et
al., 2013). Deep brain stimulation of the ventral striatum can improve OCD symptoms
(Greenberg et al., 2010; Sturm et al., 2003), possibly due to a normalization of oscillatory
activity between ventral striatum and cortex (Figee et al., 2013). The results of the present study
suggest that PL cortex or NAcS activation natively promotes aversively-mediated response
inhibition, which may not be consistent with an OCD-like phenotype. Further investigation of
the cortico-striatal regulation of compulsive-like behaviors relevant to addiction and OCD are
necessary to clarify this distinction.
2.5 Conclusion
Investigation of the cortico-striatal basis of conditioned suppression revealed distinct
roles for particular subnuclei of these regions. NAcC activity was not necessary for the

63
acquisition or expression of discriminative conditioned suppression, yet this subregion promoted
behavioral activation. In contrast, NAcS activity was required for the appropriate acquisition and
expression of conditioned suppression, suggesting that this region is critical for aversively-
motivated response suppression. Although neither region of the mPFC was involved in fear
acquisition, both PL and IL similarly disinhibited reward-seeking during CS+ presentations.
These results provide evidence that particular subregions of the NAc dissociably affect
conditioned suppression, and implicate a possible mPFC (particularly PL) to NAcS circuit in this
effect. In addition, NAcS was shown to play a novel role in fear acquisition, suggesting that an
efferent projection known to be involved in aversive learning, possibly BLA, to this region may
prime plasticity related to fear learning. Thus, these findings logically lead to circuit-based
hypotheses of fear acquisition and expression, involving the NAcS and another structure during
acquisition, and a separate PL to NAcS circuit during expression.

64
Table 1. Mean (±SEM) values for overall locomotion, and the change in locomotor activity
during CS+ versus CS- presentations within the conditioning session, for animals
manipulated prior to conditioning. *: main effect of CS Type during conditioning, p < 0.05.
†: p < 0.05 vs SAL.
Cannula
placement Treatment
Locomotion
(photobeam breaks)
∆ in activity
during CS+
presentations
∆ in activity
during CS-
presentations
PL SAL 1434.82 (±145.92) 0.20 (±0.13) -0.002 (±0.11)*
B/M 1522.29 (±361.36) 0.25 (±0.12) 0.03 (±0.12)*
IL SAL 1581.10 (±250.46) 0.19 (±0.12) 0.06 (±0.12)*
B/M 1444.15 (±180.19) 0.23 (±0.13) 0.02 (±0.10)*
NAcS SAL 1981.73 (±222.79) 0.09 (±0.13) 0.03 (±0.11)*
B/M 2353.82 (±353.55) 0.17 (±0.12) -0.04 (±0.09)*
NAcC SAL 1523.08 (±126.96) 0.20 (±0.15) 0.06 (±0.15)*
B/M 998.40 (±109.63)† 0.08 (±0.09) -0.07 (±0.07)*

65
Table 2. Mean (±SEM) values for total locomotion, rate of lever-pressing, and total lever-presses
during the discriminative fear expression test session. *: p < 0.05 vs SAL. #: p = 0.09.
Infusion
timeline
Cannula
placement Treatment
Locomotion
(photobeam
breaks)
Lever-press
rate
(presses/min)
Total lever-
presses
Pre-
conditioning
PL SAL 1554 (±208) 17.4 (±2.5) 709.6 (±67.7)
B/M 1699 (±212) 19.5 (±4.0) 836.9 (±134.8)
IL SAL 1465 (±274) 18.7 (±2.5) 794.4 (±98.7)
B/M 1603 (±198) 18.3 (±3.4) 773.2 (±111.5)
NAcS SAL 1764 (±251) 18.9 (±2.1) 807.8 (±81.5)
B/M 1554 (±232) 20.2 (±2.5) 777.9 (±105.9)
NAcC SAL 1975 (±163) 16.4 (±1.3) 688.3 (±49.2)
B/M 2335 (±208) 19.3 (±2.1) 846.3 (±89.8)
Pre-test PL SAL 1557 (±188) 22.2 (±3.8) 1003.5 (±140.9)
B/M 1176 (±115) 23.8 (±3.0) 1245.9 (±126.7)
IL SAL 1560 (±165) 21.5 (±2.8) 1029.6 (±147.0)
B/M 1365 (±195) 21.9 (±3.0) 1173.2 (±127.9)
NAcS SAL 1762 (±218) 21.0 (±3.6) 760.6 (±135.8)
B/M 2285 (±388) 20.3 (±2.9) 1028.7 (±189.2)
NAcC SAL 1709 (±199) 21.9 (±2.9) 751.1 (±93.3)
B/M 989 (±131)* 15.6 (±1.8)# 483.0 (±60.9)*
Single-
stimulus
(Pre-test)
IL SAL 2304 (±333) 24.4 (±5.4) 1450.9 (±242.2)
B/M 2412 (±518) 25.4 (±4.6)
2587.9
(±318.2)*

66
Figure 1. Discriminative fear task diagram and histology. (A) Discriminative fear task diagram. Note that separate groups of animals were infused prior to
conditioning and the expression test. (B) Representative histology figure for the pre-conditioning infused,
or (C) pre-test infused animals. Blue filled circles represent PL placements, yellow filled circles represent
IL placements, red triangles represent NAcS placements, while orange pentagons indicate NAcC
placements. Each dot represents the most ventral extent of the infusion, as observed in Nissl stained
sections.

67
Figure 2. Inactivation of mPFC does not impact the acquisition of conditioned suppression (A) PL cortex inactivation (B/M) prior to the conditioning session has no impact on the subsequent
expression of conditioned suppression during the expression test. Both SAL and B/M treated animals
expressed higher levels of conditioned suppression towards the CS+ than the CS-. (B) The same
manipulation of IL cortex had no impact on conditioned suppression. Open star represents a main effect
of CS Type, p < 0.05.

68
Figure 3. Pre-conditioning NAcS, but not NAcC, inactivation diminishes conditioned
suppression. (A) Inactivation (B/M) of the NAcS prior to the conditioning session reduced the amount of conditioned
suppression expressed during the expression test, as compared to SAL-infused animals. (B) Pre-
conditioning inactivation of NAcC was without effect on the level of discriminative conditioned
suppression. Open star represents a main effect of CS Type, p < 0.05, or a simple-main effect analysis of
the difference in CS+ expression between B/M and SAL groups.

69
Figure 4. Both mPFC subregions control the expression of conditioned suppression. (A) PL cortex was necessary for the appropriate expression of discriminative suppression, as B/M
treatment diminished the degree of suppression to the CS+, as compared to SAL treatment. (B)
Inactivation of IL produced a qualitatively similar effect, diminishing overall suppression. Open star
represents a main effect of CS Type or Treatment, p < 0.05. n.s.: non-significant.

70
Figure 5. IL inactivation has no impact on conditioned suppression expression conducted
using a standard, single-stimulus design. (A) Histology schematic for animals in the single-stimulus fear conditioning experiment. Yellow circles
represent the ventral extent of infusion into the IL cortex. (B) Infusion of B/M into the IL had no impact
on the expression of conditioned suppression when evaluated using a single-stimulus approach. Open
star: Simple main effect breakdown of the CS Block effect, p < 0.05 as compared to the first block of CS
presentations.
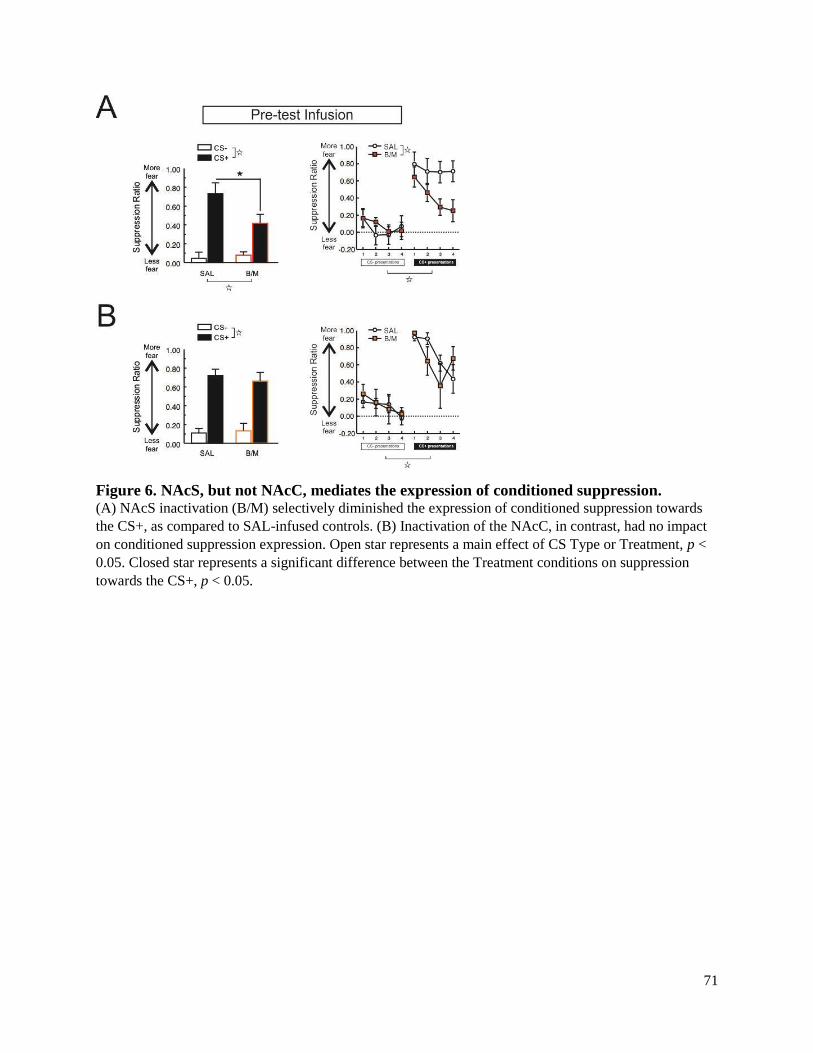
71
Figure 6. NAcS, but not NAcC, mediates the expression of conditioned suppression. (A) NAcS inactivation (B/M) selectively diminished the expression of conditioned suppression towards
the CS+, as compared to SAL-infused controls. (B) Inactivation of the NAcC, in contrast, had no impact
on conditioned suppression expression. Open star represents a main effect of CS Type or Treatment, p <
0.05. Closed star represents a significant difference between the Treatment conditions on suppression
towards the CS+, p < 0.05.

72
Chapter 3: Investigating functional cortico-striatal or limbic-striatal circuits contributing
to the acquisition and expression of discriminative conditioned suppression
3.1 Introduction
In the previous chapter, we established that activity within the NAcS during discriminative
Pavlovian fear conditioning was necessary for the appropriate conditioned suppression of
reinforcement-seeking during a subsequent expression test. In addition, NAcS and PL partially
mediated the expression of conditioned suppression, suggesting that these regions may act in
concert to inhibit action during fear. Investigation of the neural circuits that mediate the
acquisition and expression of conditioned suppression may provide clinically-relevant insight
into the etiology of disorders characterized by affective disturbances such as punishment
insensitivity (Deroche-Gamonet et al., 2004; Figee et al., 2016; Limpens et al., 2014).
To better understand the circuit mechanisms contributing to these effects,
pharmacological disconnections can be utilized to prevent communication between multiple
brain regions during particular task events. By inserting cannula into each region of interest in an
asymmetric fashion (one cannula in each region, in contralateral hemispheres), infusions can be
made to completely abolish communication between the two structures. In contrast, inserting
cannula into each region symmetrically (one cannula in each region, in the ipsilateral
hemisphere) partially disrupts communication, leaving both structures in one hemisphere intact
and capable of maintaining normal behavior. Disruption of behavior following a contralateral
disconnection, combined with a null effect of ipsilateral manipulation, implies that the targeted
regions form a functional circuit. Our research group has consistently utilized this method to
examine the cortico-limbic-striatal regulation of decision-making and executive function (Block,
Dhanji, Thompson-Tardif, & Floresco, 2007; Floresco & Ghods-Sharifi, 2007; Jenni, Larkin, &
Floresco, 2017; St Onge, Stopper, Zahm, & Floresco, 2012),

73
A number of candidate regions relevant to the acquisition of fear conditioning may exert
their effects in part through a projection to the NAcS. Foremost among these is the BLA, a
structure vital to fear acquisition (Fanselow & LeDoux, 1999). Lesions or inactivations of this
region dramatically diminish the acquisition of defensive reactions and physiological indices of
fear (Goosens & Maren, 2001; Helmstetter & Bellgowan, 1994; Koo, Han, & Kim, 2004;
LeDoux et al., 1990; Wilensky et al., 1999), consistent with a role for this region in integrating
the sensory properties of the CS and US for fear memory formation. Synaptic plasticity within
this BLA complex, mediated by glutamate NMDA-receptors, is necessary for fear memory
formation (for review, see Johansen, Cain, Ostroff, & Ledoux, 2011; Orsini & Maren, 2012).
Blocking these receptors within the BLA using the specific NMDA-receptor antagonist AP-V,
for example, has been shown to prevent the normal acquisition of Pavlovian conditioned fear
(Maren, Aharonov, Stote, & Fanselow, 1996).
The BLA mediates many of the behavioral and autonomic manifestations of fear through
projections to the central amygdala, an output nucleus that gates fear expression through
downstream interactions with structures including the periaqueductal gray (for review, see Fendt
& Fanselow, 1999). However, the BLA also regulates other aspects of aversively-motivated
behavior via projections to striatal nuclei, including the NAcS. The BLA projects
monosynaptically and primarily ipsilaterally to the NAcS (Brog et al., 1993; Groenewegen et al.,
1999; Kita & Kitai, 1990; Shinonaga, Takada, & Mizuno, 1994), forming a functional circuit that
is known to mediate active avoidance (Ramirez et al., 2015) as well as the consolidation of
inhibitory avoidance (LaLumiere et al., 2005). These types of avoidance conditioning rely in part
upon Pavlovian mechanisms, suggesting that the associations made during Pavlovian fear
acquisition, and their subsequent effect on conditioned suppression, may be similarly mediated.

74
Additionally, olfactory fear learning modulates mPFC to NAcS activity, an effect which has
been shown to depend upon BLA input to the NAcS (McGinty & Grace, 2008).
Because the impact of NAcS inactivation during conditioning was apparent 48 hrs later
during a fear expression test (Chapter 2), it is possible that BLA neurons projecting to the NAcS
produce glutamate-mediated plasticity (similar to that which occurs in the BLA during
conditioning) within this ventral striatal subnucleus. BLA stimulation has previously been shown
to increases dopamine and glutamate release in the NAc (Floresco, Yang, Phillips, & Blaha,
1998; Jackson & Moghaddam, 2001), as well as induce plasticity within NAc neurons (Floresco,
Blaha, Yang, & Phillips, 2001). Thus, contralateral disconnection of these structures utilizing
traditional pharmacological inactivation of the BLA, and a NMDA-R antagonist to eliminate
plasticity in the NAcS, may recapitulate the effect of bilateral NAcS inactivation prior to the
conditioning session.
The results of the previous chapter also implicated a possible circuit between the PL and
NAcS that may mediate the expression of conditioned suppression. The mPFC, including the PL
and IL cortex (particularly its ventral aspect), projects strongly to the medial NAcS (Brog et al.,
1993; Sesack et al., 1989; Vertes, 2004), providing a candidate circuit for this effect.
Behaviorally, both PL and NAcS make substantial contributions to other types of response
inhibition, including forming a functional circuit mediating impulsive action (Feja et al., 2014;
Feja & Koch, 2014, 2015; Resstel et al., 2008). Additionally, interactions between glutamatergic
activity in the PL and dopaminergic activity in the NAcS mediates the expression of anxiety
(Ahmadi, Nasehi, Rostami, & Zarrindast, 2013). Of direct relevance to fear, a recent study has
illustrated that mPFC neurons projecting to the lateral segment of the NAcS promote the
conditioned suppression of reward-seeking in an instrumental punishment paradigm (Kim et al.,

75
2017). It remains possible that a PL to medial NAcS circuit similarly regulates response
inhibition, which can be investigated by performing contralateral disconnections using
pharmacological inactivation to eliminate neural activity in each structure, asymmetrically.
Two hypotheses were tested, based on our initial regional inactivation experiments
(Chapter 2). The first was whether BLA input to the NAcS during acquisition is necessary for the
appropriate expression of discriminative conditioned suppression. The second was whether the
PL and NAcS form a functional circuit mediating the expression of conditioned suppression.
3.2 Methods
Most experimental procedures were identical to those from Chapter 2. Thus, only notable
deviations from the previous procedure will be described here.
3.2.1 Pharmacological disconnection rationale and surgery
To establish whether a functional circuit between two regions mediates a particular behavior, one
can employ an asymmetric disconnection procedure (Fig. 7A & B). Generally, this technique
relies upon the disruption of neural activity within a brain region of interest in one hemisphere,
and the disruption of activity in a different region in the contralateral hemisphere. When neural
activity in one region (for example, the BLA) is perturbed within one hemisphere, the
transmission of task-relevant information to another region of interest (for example, the NAcS) is
prevented. In the contralateral hemisphere, neural activity can be disrupted in the efferent target
(the NAcS), but not the upstream region (the BLA), such that neither hemisphere has a complete
functional circuit with which to regulate the behavior if interest.
Interpretation of this procedure relies upon the assumption that the disruption of activity
between these structures in the ipsilateral hemisphere should be without effect, as the intact

76
circuit in the contralateral hemisphere should be able to maintain normal function. To control for
this, symmetric disconnections can be performed, where activity is disrupted within each region
in the ipsilateral hemisphere. In the event that an ipsilateral disconnection produces an effect,
unilateral manipulation of each region individually can be performed to see whether the effect of
the symmetric disconnection was due to the partial loss of a functional circuit, or whether the
effect is mediated by a single node within this putative circuit.
The first disconnection experiment was designed to probe the circuit basis of the role of
the NAcS during the conditioning session. In the previous chapter, we illustrated that
inactivation of the NAcS during conditioning resulted in a more labile fear memory during the
expression test (Figure 7A). Here, we examined whether this effect may be mediated by a
projection from BLA to the NAcS. Because the effect of pre-conditioning NAsS inactivation
illustrated in the previous chapter was observed days later (during the expression test), we
suspected that BLA may trigger glutamate-mediated plasticity within the NAcS. To eliminate
this plasticity, we infused a dose of the glutamate NMDA-receptor antagonist AP-5 into the
NAcS, combined with B/M into the BLA to inactivate this region. AP-V has been shown to
block long-term potentiation (Davis, Butcher, & Morris, 1992; Morris, 1989), and has previously
been used to impair the acquisition of a spatial working memory task when infused into the NAc
at this dose (Smith-Roe, Sadeghian, & Kelley, 1999).
The second experiment examined the possibility that PL cortex may drive fear expression
in part through a projection to NAcS, in keeping with their qualitatively similar effects on the
expression of conditioned suppression (Figure 7B). To disconnect the PL and NAcS, rats
received infusion of B/M into the contralateral PL and NAcS (asymmetric disconnection), or
ipsilateral PL and NAcS (symmetric disconnection). In addition, we conducted unilateral

77
infusions into the PL or NAcS to control for the possibility that the disconnection effects were
due to the impact of a single infusion into one hemisphere. A separate group of rats received
infusion of saline into the contralateral PL and NAcS.
Twenty-three gauge stainless steel guide cannula were implanted aimed at the BLA and
NAcS, PL and NAcS, or unilaterally in the PL or NAcS, according to the following stereotaxic
coordinates (in mm):
BLA – from bregma: AP: -3.2 ML: ±5.3; from dura: DV: -6.3
PL – from bregma: AP +3.2; ML: ±0.7; from dura: DV: -2.8
NAcS – from bregma, AP: +1.3, ML: ±1.0, from dura, DV: -6.3
The particular hemisphere selected for each placement was counterbalanced across
experimental conditions, such that roughly equivalent numbers of rats received cannula in each
combination of hemispheres. Four stainless-steel skull screws were inundated with dental acrylic
to secure cannula in place. Stainless-steel obturators flush with the end of the guide cannula were
inserted after surgery. Rats were given 5-10 d to recover from surgery before beginning
behavioral training.
3.2.2 Microinfusion
For the pre-conditioning disconnection experiment, the glutamate NMDA-receptor
antagonist AP-5 (1 μg /0.3 μl saline) was infused into the NAcS, combined with a standard dose
(75 ng/μl of each drug at a volume of 0.3 μl/side) of B/M into the BLA to inactivate this region.
Some rats received these infusions into the contralateral BLA and NAcS (asymmetric
disconnection), while others received infusion into the ipsilateral BLA and NAcS (symmetric

78
disconnection). A separate group of rats received infusion of 0.9% saline (0.3 μl/side) into the
contralateral BLA and NAcS. Each infusion was conducted over 45 s, with the microinjector left
in place for an additional 1 min to allow for diffusion. Separate groups of animals received
contralateral BLANAcS disconnection (n = 13), ipsilateral BLANAcS disconnection (n =
12), or saline infusion (n = 10).
For the pre-test disconnection experiment, rats received infusion of B/M (75 ng/μl of
each drug at a volume of 0.3 μl/side) into the contralateral PL and NAcS (asymmetric
disconnection), or ipsilateral PL and NAcS (symmetric disconnection). A separate group was
given unilateral inactivations of the PL or NAcS (same infusion parameters), to control for the
possibility that the disconnection effects were due to the impact of a single infusion into one
hemisphere. Control rats received infusion of 0.9% saline (0.3 μl/side) into the contralateral PL
and NAcS. Each infusion was conducted over 45 s, with the microinjector left in place for an
additional 1 min to allow for diffusion. Separate groups of animals received contralateral
PLNAcS disconnection (n = 9), ipsilateral PLNAcS disconnection (n = 9), saline infusion (n
= 10), or Uni-PFC (n = 5) and Uni-NAcS (n = 5) infusion.
3.2.3 Histology
All rats were euthanized with CO2 and brains were removed and fixed in a 4% phosphate
buffered formalin solution. Brains were sectioned at 50 μm, following which tissue was mounted
and Nissl stained using Cresyl Violet. Placements were examined under a light microscope, and
the ventral extent of each infusion is indicated in Figure 8A (BLANAcS disconnection) and
Figure 9A (PLNAcS disconnection).

79
3.2.4 Data analysis
Data analysis was conducted in a nearly identical fashion to the previous chapter. For the pre-
conditioning BLANAcS disconnection experiment, analyses were identical with the exception
that the between-subjects Treatment factor had three levels: saline, contralateral BLANAcS
disconnection, and ipsilateral BLANAcS disconnection. Overall locomotor activity during the
session was analyzed with a one-way ANOVA, with Treatment as the between-subjects factor.
Analysis of conditioned suppression during the expression test was conducted in an
identical manner to the previous chapter, with the exception that the between-subjects Treatment
factor for each experiment included more levels. For the pre-conditioning experiment, the
Treatment factor include three levels: saline, contralateral BLANAcS disconnection, and
ipsilateral BLANAcS disconnection. For the expression test experiment, the Treatment factor
was made up of four levels: saline, contralateral PLNAcS disconnection, ipsilateral
PLNAcS disconnection, unilateral inactivation (combined across PL and NAcS placements,
see Results). Follow-up simple main effects analyses were conducted using one-way ANOVAs,
where appropriate. Locomotion (photobeam-breaks/session) during the conditioning session or
expression test were analyzed using separate independent samples t-tests. The rate of lever-
pressing in the first 5 min of the session and the total number of lever-presses made during the
session were analyzed in an identical fashion.

80
3.3 Results
3.3.1 BLA-NAcS disconnection during the acquisition of discriminative conditioned
suppression
Disconnection of BLANAcS had no effect on CS-induced changes in locomotor activity
during the conditioning session, as compared to animals that underwent ipsilateral disconnection
or saline treatment (Table 3). There was no main effect of Treatment (F(2,32)=0.59,p>0.56), and
there was no CS Type x Treatment interaction (F(2,32)=0.23,p>0.79). There was a main effect of
CS Type (F(1,32)=15.29,p<0.001), which indicated that the locomotor increase from baseline
was greater during CS+ presentations than CS- presentation, regardless of treatment.
Disconnection tended to increase overall locomotor activity, although this effect only approached
significance (F(2,32)=3.13,p<0.06). Regardless, disconnection did not affect the differential
change in locomotion from baseline caused by CS+ and CS- presentation.
Surprisingly, disconnection of the BLA from the NAcS during conditioning had no
impact on the subsequent expression of discriminative conditioned suppression (Figure 8B).
There was no effect of Treatment (F(2,32)=0.74,p>0.48), no CS Type x Treatment interaction
(F(2,32)=1.06,p>0.35), and no three-way interaction (F(6,96)=0.52,p>0.79). Thus, all rats
suppressed their reinforcement-seeking more during presentations of the CS+ than the CS-, as
indicated by a main effect of CS Type (F(1,32)=101.68,p<0.001). The total number of lever
presses made during the expression test session did not differ as a function of Treatment
(F(2,32)=2.41,p>0.10), nor did the rate of lever-pressing during the first 5 min of the test session
(F(2,32)=2.19,p>0.12) (Table 4). Similarly, total locomotion was not altered by BLANAcS
disconnection (F(2,32)=0.91,p>0.41) (Table 4). Thus, it is unlikely that a BLANAcS pathway
mediates the role of the NAcS in the acquisition of discriminative conditioned fear.

81
3.3.2 PL-NAcS disconnection during the expression of discriminative conditioned
suppression
As there was no significant difference between the mean suppression ratio during the CS- and
CS+ for animals in the Uni-PFC group (CS-: 0.07±0.06 SEM, CS+: 0.82±0.10 SEM) versus the
Uni-NAcS group (CS-: 0.14±0.06 SEM, CS+: 0.85± 0.08 SEM) (F(1,8)=0.39,p>0.55), these
groups were combined into a singular unilateral inactivation group for all subsequent analyses.
Disconnection of the PL cortex from the NAcS diminished the expression of conditioned
suppression, indicated by a significant main effect of Treatment (F(3,34)=3.66,p<0.022), as well
as a CS Type x Treatment interaction (F(3,34)=6.46,p<0.001) (Figure 9B). There was no three-
way interaction (F(9,102)=1.31,p>0.24). Follow up simple-main effects analyses on the two-way
interaction revealed that this effect was due to a difference between the treatment conditions on
CS+ trials (F(4,33)=8.42,p<0.001), but not CS- trials (F(4,33)=0.66,p>0.58). Further analysis
indicated that suppression during the CS+ was similar between saline animals and the
unilaterally inactivated group (F(1,18)=0.88,p>0.36). In contrast, animals in the contralateral
PLNAcS disconnection group (F(1,17)=24.48,p<0.001), or ipsilateral PLNAcS
disconnection group (F(1,17)=10.07,p<0.006), expressed less conditioned suppression during
CS+ presentations, when compared to saline-infused control animals. Contralateral PLNAcS
disconnection animals also expressed less conditioned suppression during the CS+ than did
unilaterally inactivated animals (F(1,17)=12.16,p<0.003), but the comparison between ipsilateral
PLNAcS disconnection and unilateral infusion only approached significance
(F(1,17)=3.94,p>0.06).
None of the treatments had an effect the number of lever presses made during the
expression test (F(3,34)=0.12,p>0.94), or the rate of lever-pressing made during the initial

82
portion of the test session (F(3,34)=0.30,p>0.82) (Table 4). However, locomotor activity did
differ as a function of treatment (F(3,34)=7.02,p<0.001). This was driven by a significant
increase in locomotor activity in the ipsilateral disconnection group, as compared to all other
groups (all p-values < 0.025).
3.4 Discussion
Here, we attempted to identify two functional circuits involving the NAcS that mediate the
acquisition or expression of Pavlovian conditioned suppression. Contralateral or ipsilateral
disconnection of the NAcS and BLA performed prior to the acquisition of fear conditioning had
no impact on subsequent expression, suggesting that the NAcS may interact with another
structure during this critical task epoch to mediate fear acquisition. In contrast, disconnection of
the PL cortex from the NAcS, whether conducted in a contralateral or ipsilateral manner,
decreased the expression of conditioned suppression. Importantly, unilateral inactivation of
either structure had no impact on performance. The effect of PLNAcS disconnection was
qualitatively similar to the effect of bilateral inactivation of either structure alone, providing
evidence that a functional circuit between these structures controls fear-mediated response-
inhibition.
3.4.1 A BLA-NAcS circuit does not mediate fear acquisition
First, we chose to investigate a possible BLANAcS circuit mediating discriminative fear
acquisition, based in large part on the accepted role for this amygdalar region in fear learning
(Fanselow & LeDoux, 1999). This projection also mediates other aspects of aversively-mediated
behavior, including active and passive avoidance (LaLumiere et al., 2005; Ramirez et al., 2015).
By infusing the NMDA-receptor antagonist AP-V into the NAcS in one hemisphere, and

83
reversibly inactivating the BLA in the opposite hemisphere, we aimed to eliminate the relevant
communication and plasticity that may occur during fear learning. However, animals that
received this contralateral disconnection expressed similar levels of conditioned suppression as
did ipsilateral disconnection and saline-infused controls, suggesting that this pathway is not
involved in fear acquisition.
Although the BLA is consistently involved in the acquisition of conditioned freezing and
other physiological changes, recent data indicate that this effect may be mediated by an intra-
amygdala, rather than accumbens, projections. Targeting specific subsets of neurons that project
from the BLA to the central nucleus of the amygdala (CeA) or the NAc, Namburi, Tye and
colleagues (2015) demonstrated that optogenetic inhibition of the BLA to CeA projecting cells
inhibited fear learning, while the same manipulation of the BLA to NAc projectors was without
effect. Thus, the BLA may mediate the acquisition of Pavlovian fear exclusively through
intrinsic connections with the amygdala. Additionally, there has been some suggestion that
conditioned suppression can persist despite damage to the BLA that eliminates conditioned
freezing reactions (Lee, Dickinson, & Everitt, 2005; McDannald & Galarce, 2011). For example,
after multiple days of conditioning, BLA-lesioned rats never express normal levels of
discriminative freezing towards a CS+, but not a CS-, while conditioned suppression develops
normally (McDannald & Galarce, 2011). Thus, it is possible that the BLA does not consistently
contribute to the acquisition of conditioned suppression, providing a potential theoretical
explanation for the null effect of our disconnection procedure.
This lack of effect leads to the question of what region may mediate the observed effect
of bilateral NAcS inactivation on fear acquisition. One possible region is the ventral
hippocampus (vHPC), which projects strongly to the medial NAcS (Brog et al., 1993; French &

84
Totterdell, 2002, 2003). In fact, the density of the vHPC projection to the medial NAcS has been
shown to be larger than that of the BLA or mPFC (Britt et al., 2012). This region has also been
shown to be involved in fear acquisition (Bast, Zhang, & Feldon, 2001; Esclassan, Coutureau, Di
Scala, & Marchand, 2009; Maren & Holt, 2004). For example, inactivation of vHPC prior to
discriminative fear acquisition disrupts the expression of freezing towards a CS+, without
altering fear expressed towards a CS- (Chen, Foilb, & Christianson, 2016). This effect is
qualitatively similar to that observed in the previous chapter following NAcS inactivation,
implicating a potential serial circuit between these two structures. Previous research suggests that
a projection from the vHPC to NAcS is critical for other form of learning, including spatially-
guided conditioned place preference and foraging (Floresco, Seamans, & Phillips, 1997; Ito,
Robbins, Pennartz, & Everitt, 2008). Thus, future studies investigating the relevant efferent
projection to NAcS should target this vHPC projection.
3.4.2 Interactions between the PL and NAcS mediate discriminative conditioned
suppression
Disconnection of a PLNAcS circuit diminished conditioned suppression (Chapter 2). This
finding supports the contention that information regarding the aversive nature of the CS+ is
serially transmitted from the PL cortex to the NAcS, promoting the top-down regulation of
conditioned suppression. Decreased PL activity has previously been associated with reward-
seeking despite the threat of punishment in a Pavlovian or instrumental context (Chapter 2; Chen
et al., 2013; Limpens et al., 2015; Resstel et al., 2008). This pattern of findings reinforces the
notion that the mPFC inhibits behavior during threat, in a top-down manner. Consistent with this,
a similar circuit has recently been demonstrated between the mPFC and lateral NAcS which
causally enforces response inhibition during periods of reward-seeking under threat of shock

85
(Kim et al., 2017). In that study, a subset of mPFC projection neurons to the NAcS are active
during the decision to inhibit reward-seeking behavior, while hypoactivity within this projection
is typical of seeking during punishment. Exciting this aversion-sensitive projection decreases the
probability of seeking during threat of punishment, consistent with a causal role in response-
selection mediated by this mPFClateral NAcS projection (Kim et al., 2017). Here, we illustrate
that such a PLmedial NAcS projection is similarly involved in the influence of an aversively-
conditioned Pavlovian cue on reward-seeking.
One key point of contention arising from these data is that both contralateral and
ipsilateral disconnections of PL and NAcS resulted in a similar behavioral phenotype, decreasing
conditioned suppression. A parsimonious interpretation of data arising from disconnection
procedures relies upon on the assumption that projections are primarily ipsilateral, such that
removal of a single node within each circuit in different hemispheres effectively prevents all
communication within the circuit. However, if contralateral projections between these two
structures exist, they may remain partially intact and able to impact behavior. In fact, PL cortex
projects both ipsilaterally and contralaterally to the NAcS (Brog et al., 1993; Vertes, 2004). A
common way to eliminate these ipsilateral connections is to perform a partial callosotomy,
severing the contralateral communication between the two structures. Here, our NAcS cannula
transected the corpus callosum, which should limit the contralateral connection between these
two regions, making this explanation unlikely. Importantly, when we inactivated either structure
in isolation, there was no effect on conditioned suppression. Taken together, these results imply
that parallel PLNAcS projections within each hemisphere are necessary for normal
conditioned suppression. Unilateral nodes within this circuit are neither necessary nor sufficient

86
to produce behavior, providing evidence that interactions between these subnuclei are of critical
importance.
3.5 Conclusion
Here, we attempted to answer the question of whether particular circuits involving the NAcS
were involved in the acquisition or expression of discriminative fear. In the first experiment, we
attempted to recapitulate the impact of bilateral NAcS inactivation during the acquisition of
discriminative fear by blocking plasticity within the NAcS while the BLA was inactivated in
contralateral or ipsilateral hemispheres. Contrary to our expectation, disconnection of this
BLANAcS circuit had no impact on the ability to acquire discriminative fear, as measured by
conditioned suppression during an expression test. This result suggests that another afferent
region, possibly the vHPC, may prime NAcS during acquisition, allowing for subsequent
discriminative fear expression. In a separate experiment, we examined whether a direct
PLNAcS circuit may mediate the impact of bilateral inactivation of either structure on the
expression of conditioned suppression. In fact, we observed that disconnection (contralateral or
ipsilateral) of this cortico-striatal circuit recapitulated a qualitatively similar effect, diminishing
the expression of conditioned suppression. This result suggests a top-down role for the PL cortex
in the regulation of Pavlovian conditioned suppression, mediated via the NAcS. Overall, these
studies provide further clarification of how defensive behavior is mediated by discrete cortico-
limbic-striatal circuits, implicating regions of the mPFC and NAcS in the inhibition of
responding during Pavlovian threat, and the NAcC in the promotion of appetitive vigor.

87
Table 3. Mean (±SEM) values for ancillary measures during the conditioning session, induced
by BLA-NAcS manipulation prior to conditioning. Measures included the change in
locomotor activity during CS+ versus CS- presentations and total locomotion within the
conditioning session. *: main effect of CS Type during conditioning, p < 0.05.
Cannula
placement Treatment
Locomotion
(photobeam
breaks)
∆ in activity
during CS+
presentation
∆ in activity
during CS-
presentation
BLA-NAcS SAL 1314.70 (±183.70) 0.19 (±0.19) -0.003 (±0.12)*
Contra-Disc 1359.23 (±115.75) 0.19 (±0.12) 0.04 (±0.13)*
Ipsi-Disc 2287.17 (±485.26) 0.11 (±0.16) -0.02 (±0.14)*
Table 4. Mean (±SEM) values for ancillary measures induced by BLA-NAcS or PL-NAcS
manipulation. Total locomotion, the rate of pressing (first 5 min of the test session), and
total lever-presses are reported during the expression test session. *: p < 0.025 versus all
other PL-NAcS treatment conditions.
Cannula
placement Treatment
Locomotion
(photobeam breaks)
Lever-press rate
(presses/min)
Total lever-
presses
BLA-NAcS SAL 2099 (±187) 26.2 (±2.9)
1069.9 (±87.6)
Contra-Disc 2046 (±192) 27.6 (±3.4) 1079.4 (±123.4)
Ipsi-Disc 2464 (±314) 39.2 (±6.8) 1486.3 (±209.3)
PL-NAcS SAL 1651 (±205) 24.3 (±3.3) 968.2 (±114.7)
Contra-Disc 2155 (±272) 23.6 (±4.0) 1056.2 (±217.1)
Ipsi-Disc 3384 (±357)* 23.5 (±3.9) 938.6 (±104.1)
Uni-Inact 2455 (±371) 19.8 (±2.9) 947.5 (±144.5)

88
Figure 7. Disconnection methodology diagram. (A) Cartoon depicting the functional disconnection employed to examine a potential BLA-NAcS circuit
mediating the acquisition of conditioned suppression, or a (B) PL-NAcS circuit relevant for the
expression of conditioned suppression. The red cartoon structure represents the NAcS, while the afferents
are indicated in green (BLA) or blue (PL). The white X in a filled circle represents the pharmacological
manipulation of a particular structure. Solid black lines with an arrow indicate intact projections, while
broken lines with an interrupted end represent the effect of pharmacological intervention. Note that these
diagrams are overly simplified, and do not depict potentially relevant projections to 3rd brain regions.

89
Figure 8. A BLA-NAcS disconnection does not mediate the acquisition of conditioned fear. (A) Histology schematic illustrating the ventral extent of each infusion in the NAcS (left) or BLA (right)
Closed circles represent contralateral infusions (SAL or Contra-Disc), and closed triangles represent
ipsilateral disconnections (Ipsi-Disc). (B) Contralateral disconnection and ipsilateral disconnection of a
BLA-NAcS pathway had no impact on fear expression, as these animals did not differ from saline-infused
controls. Open star: Main effect of CS Type, p < 0.05.

90
Figure 9. A PL-NAcS projection contributes to the expression of conditioned suppression. (A) Histology schematic illustrating the ventral extent of each infusion in the NAcS (left) or PL cortex
(right). Closed circles represent contralateral infusions (SAL or Contra-Disc), closed triangles represent
ipsilateral disconnections (Ipsi-Disc), and grey pentagons represent unilateral inactivation (Uni-Inact). (B)
Animals that underwent contralateral disconnection or ipsilateral disconnection expressed less fear
towards the CS+, as compared to control animals. Unilateral inactivation was significantly different from
contralateral disconnection animals, but comparison between this group and ipsilateral disconnection
animals only approached significance. Open star: main effect of CS-type, p < 0.05. Closed star:
comparison of suppression to the CS+, p < 0.05 between the SAL group and the Contra-Disc or Ipsi-Disc
group, or the Contra-Disc and the Uni-Inact group. #: p = 0.06 between the Ipsi-Disc and Uni-Inact group.

91
Chapter 4: The role of NAc core and shell in motivational conflict during reward and
punishment
4.1 Introduction
In the previous chapters, we identified that the NAc is functionally heterogeneous when
considering Pavlovian fear expression. Instrumental punishment may similarly rely upon discrete
NAc subregions, however this hypothesis has not been empirically tested. Considerable research
has been dedicated to clarifying the influence of positive reinforcement on decision-making,
implicating the NAc and associated cortico-limbic afferents in such reinforcement learning
(Cardinal et al., 2002; Floresco, 2015; Parkinson, Cardinal, et al., 2000). In contrast, less is
known about how this system guides behavior in response to punishment, a process by which an
instrumental action co-occurs with a negative consequence, such as a lever-press-contingent
foot-shock in rodents. In a majority of individuals, punishment serves to suppress the
instrumental action with which it occurs. However, neuropsychiatric conditions such as
substance abuse and obsessive compulsive disorder are characterized by compulsivity, whereby
punishment is less effective in curtailing detrimental behavioral patterns (Everitt, 2014; Feil et
al., 2010; Figee et al., 2016; Jentsch & Taylor, 1999; Lubman et al., 2004; Perry & Carroll,
2008). As such, investigation of the circuitry underlying punishment-induced inhibitory control
may provide insight into the pathophysiological underpinnings of these symptoms in various
disease states.
Compulsivity in the face of punishment is recognized by the DSM-5 as a core symptom
of substance abuse and other disorders, and pre-clinical findings suggest that these symptoms
may be driven by alterations within cortico-limbic circuitry (Chen et al., 2013; Limpens et al.,
2014; Pelloux, Murray, & Everitt, 2013; Radke, Jury, et al., 2015; Radke, Nakazawa, & Holmes,
2015). Prolonged access to cocaine produces punishment-resistant drug seeking, concomitant

92
with hypofunction of medial prefrontal cortex (mPFC) (Chen et al., 2013). Optogenetic
inhibition or activation of mPFC decreases or increases, respectively, the impact of punishment
on cocaine seeking (but see Pelloux, Murray, Everitt, 2013), suggesting that mPFC activity may
be causally related to the punishment-mediated inhibition of seeking. Similarly, pharmacological
inactivation or lesion of the mPFC produces operant responding for both cocaine and sucrose
that is insensitive to potential punishment, whether presented in a Pavlovian or instrumental
fashion (Limpens et al., 2015; Resstel et al., 2008). Prefrontal regions seem to perform a top-
down inhibitory function, acting as a break when responding is directly punished, or in the
presence of a fear-inducing stimulus. Likewise, the basolateral amygdala (BLA) promotes
behavioral suppression during punishment. Jean-Richard-Dit-Bressel and McNally (2015)
recently showed that inactivation of caudal (but not rostral) BLA eliminated the inhibition of
lever-pressing produced by contingent foot-shock. Inactivated rats made more lever-presses
during punishment, and did not display the typical increase in latency to press caused by
punishment. Thus, both mPFC and BLA may contribute in a similar manner to punishment
avoidance during appetitively-motivated behavior.
Although the BLA and PFC appear to subserve complementary roles in punishment
avoidance, the downstream structure mediating this effect is currently unknown. The nucleus
accumbens (NAc) receives dense glutamatergic input from both mPFC and BLA, and is known
to regulate various forms of appetitive conditioning via its meso-cortico-limbic efferents
(Cardinal et al., 2002; Floresco, 2015; Sesack & Grace, 2010). The NAc is primarily composed
of two functionally and anatomically distinct subregions, the more lateral core (NAcC) and more
medial shell (NAcS) (Heimer et al., 1997; Zahm & Brog, 1992). These two subregions have been
suggested to serve dissociable yet complementary functions during reward-seeking, with the

93
NAcC driving approach towards motivationally-relevant stimuli, and the NAcS facilitating
inhibition of inappropriate behaviors (Ambroggi et al., 2011; Floresco, 2015). In this regard, the
ventral regions of the mPFC and caudal BLA project strongly to the medial NAcS (Berendse,
Galis-de Graaf, et al., 1992; Brog et al., 1993; Groenewegen et al., 1999; Heilbronner et al.,
2016; Kita & Kitai, 1990; Wright et al., 1996), suggesting that this nucleus may facilitate
inhibition of punished behavior regulated by these upstream cortical and limbic regions. A recent
experimental report supports this contention, suggesting that a projection from the mPFC to
lateral NAcS is active when suppressing punished reward-seeking, and inhibited when promoting
seeking regardless of punishment (Kim et al., 2017). However, whether the medial NAcS or
more lateral NAcC contributes to such behavior is unknown. It is therefore possible that NAc
subregions may differentially contribute to adjusting behavior in response to punishment, with
NAcS suppressing reward-seeking in the face of punishment in a manner similar to the BLA or
PFC, and NAcC generally promoting action.
The present series of experiments were designed to both confirm a role for BLA in
mediating reward/punishment conflict, and explore the potential differential contribution of
NAcS versus NAcC to the same behavior. To this end, separate groups of well-trained rats
received reversible inactivation of BLA, NAcS, or NAcC while performing an operant-based
“Conflict” task. We also examined the potential monoaminergic contribution to punished
reward-seeking by administering the monoamine releaser d-amphetamine (AMPH) in a subset of
animals on the Conflict task, as previous studies have suggested that elevations in monoamine
activity promotes the punishment-induced suppression of behavior (Killcross, Everitt, &
Robbins, 1997; Lazareno, 1979; Leone, de Aguiar, & Graeff, 1983). During the Conflict task,
sucrose reward was available on a lean reinforcement schedule, without punishment, during two

94
“Safe/Reward” periods. Interspersed between these periods was a separate “Conflict” period,
wherein sucrose was available on a richer schedule, but 50% of lever-presses triggered a foot-
shock punishment. Results using this Conflict task, and a “No-Conflict” (identical schedules of
reinforcement, but no punishment) control variant, suggested that BLA and NAcS promote
punishment-induced behavioral suppression, while NAcC plays a more general role in driving
reward-seeking.
4.2 Methods
4.2.1 Animals
All experimental protocols were approved by the Animal Care Committee, University of
British Columbia, and conducted in compliance with guidelines provided by the Canadian
Council on Animal Care. All reasonable efforts were made to minimize the number and suffering
of animals used. Male Long-Evans rats arrived weighing 225-350g (Charles River) and were
group housed (4-5 per cage) and allowed 6-7 d of acclimation to the colony. Colony temperature
(21° C) and light cycle (12-h light/dark) were kept constant. Prior to operant training, all rats
were individually housed and food-restricted to approximately 90% of their free-feeding weight,
and allowed to gain weight throughout the course of the experiment on a delayed-growth curve.
4.2.2 Apparatus
Behavioral testing was conducted in eight Med Associates (St Albans, VT, USA) operant
conditioning chambers. Each chamber (30.5 cm x 24 cm x 21 cm) was contained in a sound-
attenuating enclosure, ventilated by a fan that also served to mask external noise. Within each
chamber were two retractable levers along one wall, separated by a food receptacle from which
sucrose pellet reinforcement was delivered (45 mg pellet, BioServ, Frenchtown, NJ). For all
experiments, only the left lever was extended into the chamber. Each box was outfitted with

95
three 100 mA cue lights, one over each retractable lever, and one over the food receptacle. A
single 100 mA house light was situated on the wall opposite the food receptacle. Four infrared
photobeans located just above the grid floors were used to index locomotor activity. The
chamber floor consisted of 19 stainless steel rods spaced 1.5 cm apart. The rods were wired to a
shock source and solid-state grid scrambler for the delivery of foot-shock.
4.2.3 Surgery
Rodent anesthesia was conducted slightly differently for BLA and NAc placements, due to
changes in institutional policies regarding anesthetic techniques. Animals receiving BLA cannula
were anesthetized with a combination of ketamine/xylazine (100 and 20 mg/ml at 100 and 10
mg/kg, i.p.), exclusively. Animals receiving NAc cannula were first anesthetized with a half-
dose of ketamine/xylazine (same mg/ml, i.p.), and then maintained on Isoflurane anesthetic (2-
3% Isoflurane concentration) throughout surgery. Twenty-three gauge bilateral stainless-steel
guide cannula were aimed at the BLA, NAcS, or NAcC according to the following stereotaxic
coordinates (in mm):
BLA – from bregma, AP: -2.7, ML: ±5.3, from dura, DV: -7.0
NAcS – from bregma, AP: +1.3, ML: ±1.0, from dura, DV: -6.3
NAcC – from bregma, AP: +1.6, ML: ±1.8, from dura, DV: -6.3
Dental acrylic adhered to four stainless-steel skull screws held cannula in place. Stainless-steel
obturators flush with the end of the guide cannula were inserted immediately following surgery,
and remained in place throughout the experiment. Rats were given approximately 1 wk to
recover from surgery before beginning behavioral training.

96
4.2.4 Training
Twenty-four hours before their first operant training session, rats were provided with ~30
sucrose pellets in their home cage, to reduce potential food neophobia. Subsequently, 15 min
training sessions were conducted at a consistent time each day, 5-7 days per week. Rats were
initially trained for 3 d on a fixed-response 1 (FR1) schedule, such that each press of the lever
was rewarded with a sucrose pellet. Animals were then trained for 4-5 d on a variable-interval 15
s/FR1 (VI15/FR1) reinforcement schedule, whereby a lever-press after a 15 s interval was
rewarded with a single sucrose pellet. The final portion of training was conducted over 3 days,
on a VI15/FR5 schedule, identical to VI15/FR1 except that the 5th press after the variable-
interval was rewarded with a single pellet.
The Conflict task was based on procedures used by Broersen et al. (1995), and consisted
of three discrete 5 min phases (15 min total session length). During these sessions, the lever
remained inserted into the chamber for the entire session. During the first and the third phases,
the house-light was illuminated and rats were reinforced for lever-presses on a VI15/FR5
schedule. There was no danger of punishment during these two phases, and thus they were called
Safe/Reward phases. In contrast, during the second (middle) block, the house light was turned
off, and the left cue-light was illuminated, signaling the 5 min Conflict period. Here, reward was
delivered on a FR1 schedule, but, in addition, lever-presses resulted in foot-shock punishment
delivered on a random ratio-2 schedule (i.e., 50% of responses were shocked), with no time-out
restricting the number of presses or foot-shocks received. This produces a state of anxiety, akin
to that in the Vogel conflict task (Vogel et al., 1971).
Shock intensity was individually titrated over the course of training, such that each rat
eventually achieved criterion performance of receiving less than 20 shocks per session for two

97
consecutive days. The range of shock intensity across all experimental cohorts was: 0.35 – 0.75
mA. To achieve criterion during the Conflict period, rats in the BLA group required a mean
shock intensity of 0.48 mA (±0.02 SEM), those in the NAcS required a mean shock intensity of
0.53 mA (±0.03 SEM), and those in the NAcC of 0.60 mA (±0.02 SEM). Across the different
experimental groups, rats required 13.9 (±0.4 SEM; range 10-19) training sessions on the
Conflict task to achieve criterion performance.
Separate groups of rats were trained on a No-Conflict control version of the task, for
which the initial training was the same as the Conflict version. This task was identical to the
Conflict task, with the notable exception that no foot-shocks were delivered at any point. This
No-Conflict task was designed to assess whether any potential alterations in responding induced
by regional inactivation could be attributed to changes in the reinforcement schedules (VI15/FR5
vs. FR1) that occurred during the Conflict task. On the No-Conflict task alone, rats with BLA
cannula required 12 days of training, while those with NAc cannula required 15, until they
displayed asymptotic levels of responding as a group during the three phases, defined as two
consecutive days with < 20% variation in lever-pressing across phases.
4.2.5 Microinfusion and systemic pharmacology
Once an individual rat displayed stable behavioral performance, it received a mock infusion 10
min prior to the daily training session. This procedure consisted of removal of the obturators,
insertion of a mock injector flush with the end of the guide cannula, and placement in the
infusion enclosure for approximately 2 min. All microinfusions were conducted 10 min before
animals were placed in their operant chamber. On the infusion day, microinjectors extending 0.8
mm beyond the guide cannula were lowered into the brain and animals received bilateral
infusion of 0.9% saline (0.3 μl/side for NAc infusion and 0.5 μl/side for BLA infusion) or a 75

98
ng (NAc) or 125 ng (BLA) dose of the GABA agonists muscimol and baclofen (B/M; same
volume/side as saline). Each infusion was conducted over 45 s (NAcS) or 75 s (BLA), with
injectors left in place for an additional 60 s to allow for diffusion of solution from cannula tips.
This dose and volume of B/M in the NAc has been used previously to dissociate between the
NAcS and NAcC on a wide variety of behavioral measures (Dalton et al., 2014; Floresco et al.,
2008; Ghods-Sharifi & Floresco, 2010; Stopper & Floresco, 2011). In addition, this dose and
volume of B/M infused into the BLA is similar or identical to other studies examining the role of
the BLA individually (Ghods-Sharifi, St Onge, & Floresco, 2009), or dissociating between
adjacent amygdala subregions (Jean-Richard-Dit-Bressel & McNally, 2015; Millan, Reese,
Grossman, Chaudhri, & Janak, 2015). Order of infusion was counterbalanced across animals,
such that some rats received SAL prior to B/M, while others received infusion in the opposite
order. All animals were re-trained for a minimum of two days prior to receiving their second
infusion.
After receiving counterbalanced infusion test days, rats with NAc placements were given
two additional test days (at least two days after their final microinfusion test). On the first test
day, rats received an injection of saline (SAL; 1 ml/kg, i.p.) 10 min prior to the operant session.
The next day, rats were given an injection of d-amphetamine (AMPH; 1 mg/kg at a concentration
of 1 mg/ml, i.p.). This experiment did not include animals with BLA placements, as it was
conceived of following the completion of that experimental cohort.
4.2.6 Histology
Following the completion of behavioral testing, all rats were euthanized with CO2 and brains
were removed and fixed in a 4% formalin buffered saline solution. Once fixed, brains were
sliced at 50 μm and mounted on glass slides for placement analysis. The ventral extent of the

99
infusion in each region is displayed in Fig. 10. On the Conflict task, 11 animals completed the
experiment with accurate BLA placements (Fig. 10A), 13 with NAcS placements, and another 13
with NAcC placements (Fig. 10B). For the No-Conflict control task, 7 animals were included for
each of the NAcS and NAcC (Fig. 10C). Six animals with BLA cannula were tested on the No-
Conflict control task. The brains from these animals were sectioned, mounted, stained and
initially confirmed to be within the BLA. However, these sections were unfortunately lost prior
to plotting them in a figure. Twenty-nine animals were included in the systemic AMPH study,
including n = 13 from the NAcS and n = 16 from the NAcC (some animals in the NAcC group
that were excluded from microinfusion analysis due to inaccurate cannula placement were
included in this analysis as drug was administered systemically).
4.2.7 Data analysis
Choice behavior during the Conflict and No-Conflict tasks was analyzed using two-way within-
subjects ANOVAs with Treatment (B/M or SAL; AMPH or SAL) and Phase (Safe/Reward 1,
Conflict, and Safe/Reward 2) as the two within-subjects factors. For the inactivation
experiments, we also conducted a supplementary analysis to compare baseline performance
(lever-pressing and pellets received) across the two tasks using a two-way, between/within
subjects ANOVA, with the between-subjects factor of Task (Conflict vs. No-Conflict control)
and the within-subjects factor Phase (Safe/Reward 1, Conflict, and Safe/Reward 2). In these
analyses, baseline data were computed by averaging data obtained on the day prior to each
infusion test day (two days total). In addition, due to similarities in the level of responding during
the Safe/Reward (VI15/FR5) phases between saline-infused rats trained on the control task, and
rats that received BLA or NAcS inactivations on the Conflict task, we also conducted a series of
two-way ANOVAs within these two brain regions, with Condition (SAL No-Conflict vs. B/M

100
Conflict) as the between-subjects factor, and Phase (Safe/Reward 1 and 2) as the within-subjects
factor. Follow-up or exploratory comparisons were conducted using one-way ANOVAs or
paired-samples t-tests, where appropriate. Analysis of supplementary measures (locomotion,
pellets received, etc.) were also conducted using one-way ANOVAs or paired-samples t-tests. To
examine any potential relationship between foot-shock intensity and Conflict period responding,
the Pearson correlation between the mA shock intensity value and lever-presses during the
Conflict period was also analyzed.
4.3 Results
4.3.1 Experiment 1: Conflict task
4.3.1.1 BLA inactivation
Under control conditions, rats that were well-trained on the Conflict task (n = 11) apportioned
their lever-pressing in an adaptive manner across the three, 5-min phases (Fig. 11A), as they had
done during the later phases of training. These animals displayed robust levels of responding
during the un-punished, but less frequently reinforced Safe/Reward phases, whereas during the
punished Conflict phase, rats showed a dramatic reduction in lever-pressing. BLA inactivation
markedly altered this profile (Fig. 11B). Analysis of the lever-pressing data did not reveal a main
effect of Treatment (F(1,10)=2.38, not significant (n.s.)), indicating that the total number of
responses made during a session did not differ after B/M relative to saline infusions. However,
this analysis did produce a significant Treatment x Phase interaction (F(2, 20)=7.59,p<0.05).
Subsequent simple-main effects analyses revealed that BLA inactivation produced a dramatic
disinhibition of responding during the Conflict phase (F(1,10)=16.42, p<0.05). The degree of
disinhibition during Conflict was not significantly correlated with shock intensity (r=-0.07,
p>0.8). Yet, this effect was accompanied by a reduction in responding during the first

101
(F(1,10)=6.28,p<0.05), but not second (F(1,10)=2.70, n.s.), Safe/Reward phases. In keeping with
this effect on responding, rats obtained more pellets following BLA inactivation (main effect of
Treatment: (F(1,10)=16.53, p<0.01), Treatment by Phase interaction: (F(2,20)=15.80, p<0.01)
(Table 5). This was driven by an increase in food received during the Conflict phase
(F(1,10)=16.36, p<0.005), whereas there was no difference across treatment in terms of pellets
obtained during the Safe/Reward phases (both F-values < 1.0, both p-values > 0.40). Locomotor
activity did not differ across treatment conditions (t(10)=0.84,p>0.05) (Table 5). Thus, BLA
inactivation induced a substantial increase in punished reward-seeking, but simultaneously
attenuated responding during unpunished periods during which the effort requirement to obtain
these rewards was greater.
4.3.1.2 NAcS inactivation
Inactivation of the NAcS (n = 13) produced a profile that was qualitatively similar to that
induced by BLA inactivation in some respects (Fig. 11C). Here, data analysis again produced a
significant Treatment x Task Phase interaction (F(2,24)=14.01,p<0.001), with this effect driven
by an increase in responding during the punished Conflict phase (F(1,12)=7.56, p<0.05), and a
reduction in lever-pressing during the first (F(1,12)=14.81,p<0.005), and second
(F(1,12)=11.20,p<0.05) Safe/Reward phases. Notably, the disinhibition of responding during the
Conflict phase following NAcS inactivation was smaller in magnitude relative to that induced by
BLA inactivation. Like BLA inactivation, there was no statistically significant relationship
between shock intensity and the degree of disinhibition during Conflict (r=0.16, p>0.6). NAcS
inactivation reduced overall lever-pressing measured across the entire session, as revealed by a
significant main effect of Treatment (F(1,12)=11.82, p<0.001). Despite this effect, there were no
differences in the number of pellets obtained over the session (main effect of Treatment:

102
(F(1,12)=1.62, n.s.), although there was a significant Treatment x Phase interaction
(F(2,24)=11.63, p<0.001), meaning that rats received fewer pellets during Safe/Reward phase 1
(F(1,12)=9.46,p<0.01) and 2 (F(1,12)=6.05,p<0.05), and more pellets during the Conflict phase
following B/M infusions vs saline (F(1,12)=7.90,p<0.05) (Table 5). NAcS inactivation had no
significant impact on locomotion (t(12)=1.44, n.s.) (Table 5). Thus, suppression of neural
activity within the NAcS, like BLA, disinhibited Conflict responding and attenuated lever-
pressing during the Safe/Reward phases.
4.3.1.3 NAcC inactivation
In contrast to the effects of BLA or NAcS inactivation, infusions of B/M into the NAcC (n = 13)
produced a substantial decrease in responding across all task phases (main effect of Treatment:
(F(2,24)=35.55,p<0.001); Treatment x Task Phase interaction, (F(2,24)=20.45,p<0.001) (Fig.
11D). Simple main-effects analyses confirmed that inactivation reduced responding during
Safe/Reward phase 1 (F(1,12)=29.25,p<0.001) and 2 (F(1,12)=29.77,p<0.001), as well as the
Conflict phase (F(1,12)=7.23, p<0.05). The reduction in lever-pressing during Conflict in
particular was not correlated with shock intensity (r=-0.45,p=.12). Accordingly, animals received
fewer rewards over the entire session after NAcC inactivation (main effect of Treatment:
(F(1,12)=15.14 ,p<0.001) (Table 5). Inactivation also reduced locomotor activity during the
session (t(12)=4.04, p<0.05) (Table 5). Thus, inactivation of the NAcC resulted in a general
suppression of reward-seeking, irrespective of whether responding was punished or not.
4.3.1.4 AMPH administration
Previous research suggests that AMPH administration will suppress punished instrumental
behavior (Killcross et al., 1997; Lazareno, 1979; Leone et al., 1983). In the present study, AMPH

103
(1mg/kg) was administered systemically to animals following the completion of microinfusion
test days (n = 29 total; 13 with NAcS cannula, 16 with NAcC cannula). Here, we did not observe
a main effect of Treatment (F(1,28)=0.31,p>0.58), or a significant Treatment x Task Phase
interaction (F(2,56)=0.22,p>0.80), although there was a significant effect of Task Phase
(F(2,56)=207.61,p<0.001) (Fig. 11E). Due to our a priori prediction that AMPH would decrease
punished seeking, we ran a series of exploratory t-tests to analyze whether AMPH produced the
hypothesized effect. In fact, although AMPH had no impact on responding during the
Safe/Reward phases (both t-values > 0.5, both p-values > 0.65), AMPH significantly decreased
punished responding during the Conflict phase (t(28)=2.62, Bonferroni corrected p<0.014). This
exploratory analysis provided validation of the experimental protocol, and supported previous
findings regarding the role of monoamines in punished reward-seeking behavior. AMPH
administration caused rats to receive fewer sugar pellets, as indicated by a significant main effect
of Treatment (F(1,28)=6.22,p<0.02) and a significant Treatment x Task Phase interaction
(F(2,56)=6.23,p<0.005). Direct comparison suggests that the number of pellets received during
the Safe/Reward phases were identical (both F-values < 1, and p-values > 0.5), the number of
pellets received during the Conflict phase was decreased by AMPH treatment
(F(1,28)=6.88,p<0.02) (Table 5). AMPH treatment did not change the total number of lever-
presses made throughout the session (F(1,28)=0.31,p>0.58), but did significantly increase
locomotion (t(28)=2.34,p<0.03) (Table 5). These results point to a dissociation between the
behaviorally activating impact of AMPH on motoric behavior, and its behaviorally suppressing
impact on reward-seeking during punishment.

104
4.3.2 Experiment 2: No-Conflict control task
In Experiment 1, inactivation of either the BLA or NAcS (but not NAcC) increased responding
during the Conflict period, where lever-presses delivered food on a FR1 schedule but also
delivered foot-shocks after 50% of responses. Inactivation of each of these three target nuclei
reduced responding during the unpunished Safe/Reward phases, where food was delivered on a
leaner, VI15/FR5 schedule. Notably, alterations in responding induced by either BLA or NAcS
inactivation did not cause an overall reduction in the amount of reward obtained over the session,
with BLA inactivation actually increasing the number of pellets received. This latter observation
prompted us to explore whether the reduction in responding during the Safe/Reward periods
observed in Experiment 1 was driven by changes in the manner in which animals allocated the
relative vigor of responding during the different task phases, or merely by reduced motivation to
lever-press for rewards. Thus, separate groups of rats were trained on a No-Conflict control task
that used identical schedules of reinforcement as the Conflict task, but no foot-shock punishment
was delivered during training (Fig. 12A).
4.3.2.1 Experiment 2: BLA, NAcS and NAcC inactivation
In contrast to the profound alterations in response profiles on the Conflict task induced by BLA
inactivation, similar treatments did not significantly alter behavior in animals trained on the No-
Conflict control task (n = 6) (Fig. 12B). BLA inactivation was without effect on either lever-
pressing or pellets received (all F-values<1.73, n.s) or locomotion (t(5)=1.60, n.s.) (Table 5).
Similarly, NAcS inactivation (n = 7) did not significantly impact lever-pressing or rewards
received (all F-values<2.36, n.s.) (Fig. 12C and Table 5), and also did not affect locomotor
activity (t(6)=0.19, n.s.) (Table 5).

105
On the other hand, inactivation of NAcC (n = 7) diminished motivated output during
performance of the No-Conflict task (Fig. 12D). Under control conditions, animals in this group
displayed noticeably lower rates of responding during the Safe/Reward phases of the task,
compared to rats receiving saline infusions into either the BLA or NAcS (Fig. 12B, C, and D).
Nevertheless, infusions of B/M into this nucleus reduced responding across all phases of the task
(main effect of Treatment: (F(1,6)=9.46,p<0.05); Treatment x Phase interaction (F(1,6)=0.94,
n.s.). Accordingly, rats received fewer reward pellets after NAcC inactivation
(F(1,6)=24.24,p<0.01) (Table 5). Although NAcC inactivation decreased locomotor activity on
the Conflict task (Table 5), the same treatment administered prior to performance on the control
task did not significantly affect locomotion (t(6)=1.71, n.s.) (Table 5). Collectively, these results
lend support to the idea that NAcC promotes appetitively-motivated responding. In comparison,
the lack of effect of BLA or NAcS inactivation on this task implies that alterations in behavior
on the Conflict task induced by these treatments are unlikely to be attributed to changes in
arousal or motivation for food reward.
4.3.2.2 Experiment 2: Baseline analysis and cross-task comparison during Safe/Reward
responding
Inactivation of BLA and NAcS differentially affected responding during the V115/FR5 reward
phases for rats trained on the Conflict versus No-Conflict tasks. Inactivation of either structure
during the Conflict task reduced responding during the Safe/Reward periods, whereas, for rats
trained on the No-Conflict task, these same manipulations did not affect performance during
these phases. A closer inspection of the data obtained from the two experiments revealed that,
under control conditions, rats trained on the two tasks appeared to show different rates of
responding during these reward phases (e.g., compare the Safe/Reward panels in Figure 11B & C

106
with those of Figure 12B & C). We also noticed that, by the end of training on the No-Conflict
task, baseline rates of responding (collapsed across all three regions of interest) displayed by
animals trained on the control task (n = 21) differed considerably from those trained on the
Conflict task (n = 37) across all three task phases. To investigate this further, we compared the
number of lever-presses made on the days prior to each infusion treatment (baseline) by all rats
trained on the two tasks (Fig. 13A), using a two-way between/within subjects ANOVA with
Task and Phase as between and within-subjects factors, respectively. Comparison of the lever-
pressing data yielded a significant Task x Phase interaction (F(2,112)=59.69, p<0.001).
Predictably, rats trained on the control task made many more responses during the middle, FR1
phase (F(1,56)=713.50, p<0.001) compared to those trained on the Conflict task, where lever-
presses delivered both food and foot-shocks (Fig. 13B, middle panel). However, rats trained on
the No-Conflict task made fewer responses during the first (F(1,56)=22.53,p<0.001) and third
(F(1,56)=35.44,p<0.001) Safe/Reward phases relative to those trained on the Conflict version
(Fig. 13B, left and right panels). As a consequence, rats trained on the No-Conflict task obtained
fewer pellets during the first (F(1,56)=25.37,p<0.001) and second (F(1,56)=26.89, p<0.001)
Safe/Reward phases compared to those in the Conflict condition (full Task x Phase interaction
(F(2,112)=891.17, p<0.001) (Fig. 13C). Notably, this pattern of results was still observed when
only data from a subset of rats trained on the Conflict task for a comparable number of days (<
13) to those trained on the control task (n = 14) were included, thereby equating the relative
amount of training received by rats in both groups. Here, we again observed significant Task x
Phase interactions for the number of lever-presses (F(2,66)=47.71,p<0.001) and pellets received
per phase (F(2,66)=418.712, p<0.001; data not shown). This further suggests that any alteration
in baseline behavior displayed by rats trained on the two tasks was driven by their experience

107
with punishment, and not a difference in the amount of instrumental training. Thus, rats trained
on the Conflict task appeared to maximize their rates of responding during the unpunished
Safe/Reward phases of the task, presumably to accommodate for their suppression of responding
during the Conflict period. In comparison, during the No-Conflict control task, where reward
was available on identical schedules but in the absence of punishment, rats varied their relative
rates of responding over the session in a different manner. Here, they were more lackadaisical
during the Safe/Reward phases where the effort requirements were higher, and instead responded
more vigorously and obtained more food during the middle, FR1 phase.
We also observed that the lower levels of responding during the Safe/Reward phases by
rats trained on the No-Conflict task were comparable to the rates of responding during the same
two phases of the Conflict task following inactivation of either the BLA or NAcS. In light of
this, we formally compared response rates during the two Safe/Reward phases for saline-infused
rats on the No-Conflict task, and animals that received inactivation of BLA or NAcS on the
Conflict task, using two separate two-way, between/within subjects ANOVA, with Condition
(SAL No-Conflict vs. B/M Conflict) as the between-subjects factor, and Safe/Reward Phase as
the within-subjects factor. Within the BLA, there was no main effect of Condition (F(1,15)=0.14,
n.s.) or Condition x Phase interaction (F(1,15)=2.99, n.s.) (Fig. 13D, left panel). Similarly,
within the NAcS, there was no impact of Condition (F(1,18)=0.16, n.s.), nor a Condition x Phase
interaction (F(1,18)=1.38, n.s.) (Fig. 13D, right panel). Thus, for rats trained on the Conflict task,
inactivation of either the BLA or NAcS attenuated responding during the unpunished period such
that rats behaved in a manner similar to those that received saline infusions into the same brain
region, and had never experienced punishment during training.

108
4.4 Discussion
4.4.1 Summary
The present findings reveal complementary roles for the BLA and NAcS in mediating
responding in situations involving motivational conflict where actions may yield both reward and
potential punishment. Neural activity in these two nuclei facilitated response-suppression when
lever-presses yielded both food and shock, while in the same context, these regions invigorated
responding when the effort requirements to obtain unpunished rewards were greater. These
effects on the Conflict task did not appear to be related to shifts in the reinforcement schedule
from Safe/Reward (VI15/FR5) to Conflict (FR1) periods, as performance on a control task using
the same schedules of reinforcement in the absence of punishment was unaffected by
inactivation of these regions. In contrast, the NAcC appears to more generally promote appetitive
motivation, irrespective of motivational conflict, as inactivation of this region diminished
seeking behavior, including locomotion, across both task conditions. These data suggest that the
NAcS, but not NAcC, mediates aversion-mediated response-inhibition in an instrumental setting,
similar to their respective roles in Pavlovian conditioned suppression and behavioral activation,
as revealed in the previous chapters.
4.4.2 Cooperative roles for the BLA and NAcS in modulating punished reward-seeking
The finding that inactivation of the BLA disinhibited responding during the Conflict period is in
keeping with a vast literature implicating this nucleus in influencing behavioral responses to
aversive or threatening situations (Adolphs, 2013; Fanselow & LeDoux, 1999). Classically, BLA
lesions eliminate conditioned fear responses in both humans and rodents (Adolphs et al., 1995;
Erlich, Bush, & Ledoux, 2012; LaBar et al., 1998; LeDoux et al., 1990). Recent work suggests
that these lesions also shift preference away from smaller, unpunished rewards and towards

109
larger rewards that may also be punished, a finding that is complemented by the results of the
present study (Orsini, Trotta, Bizon, & Setlow, 2015). These studies suggest that BLA activity is
necessary to appropriately recall and utilize the memory of an aversive event, and subsequently
modify behavior. Jean-Richard-Dit-Bressel and McNally (2015) illustrated that the BLA
processes aversive consequences in the context of instrumental punishment, independent of its
role in Pavlovian fear learning. In their study, inactivation of BLA disrupted the suppression of
punished responding, causing rats to approach and engage the lever more often and with faster
latencies compared to control animals, similar to what was observed in the present study.
Interestingly, this effect on punished responding was subject to a pronounced rostral-caudal
dissociation, whereby the caudal (posterior to AP: -2.6), but not rostral (anterior to AP: -2.6),
BLA appeared to play a more prominent role in suppressing behavior under these conditions.
Although our sample size precluded the examination of whether punishment-resistance was
differentially affected by inactivation across the rostral-caudal extent of the BLA, it is notable
that our targeted BLA coordinate fell within the caudal range (AP: -2.7) used by Jean-Richard-
Dit-Bressel and McNally (2015). Taken together, these findings suggest that the BLA,
particularly its caudal aspect, contributes to the modification of responding in situations where
actions are either directly punished or no longer rewarded.
One neuroanatomical feature that distinguishes the caudal BLA from the more rostral
portion is that it sends a relatively dense projection to the medial NAcS (Brog et al., 1993;
Groenewegen et al., 1999; Kita & Kitai, 1990; Shinonaga et al., 1994). As such, BLA may
confer the appropriate inhibition of reward-seeking in the face of potential punishment in part
through its projection to the NAcS. This supposition is supported by the present findings,
whereby NAcS inactivation produced an increase in punished responding similar to that

110
produced by BLA inactivation. These findings complement a burgeoning literature implicating
the NAcS in avoiding potential aversive consequences, with these functions mediated through
direct interactions with the BLA (Fernando et al., 2013; Ramirez et al., 2015). The disinhibition
of punished responding following BLA or NAcS inactivation is also coherent with data
suggesting that the NAcS, via interactions with BLA, promotes the appropriate attenuation of
responding in appetitively-motivated situations. One such construct is the extinction of a
conditioned association, which produces a new inhibitory memory that acts to suppress the now
irrelevant response. Previous work suggests that inactivation of NAcS can disinhibit responding
during the reinstatement of extinguished food, alcohol, or cocaine seeking (Floresco et al., 2008;
Millan et al., 2010; Peters et al., 2008). Inactivation of NAcS in these and similar situations also
typically releases non-rewarded behaviors from inhibition, with rats producing more operant
responses in situations that are never reinforced following inactivation (Ambroggi et al., 2011;
Blaiss & Janak, 2009; Floresco et al., 2008). Interestingly, caudal (but not rostral) BLA
inactivation produces the same type of exaggerated reinstatement response following extinction
of food-seeking (McLaughlin & Floresco, 2007). A similarity in function of the NAcS and
caudal BLA has also been demonstrated by Millan and colleagues (2015), who showed that
inactivation of either structure disinhibits reward-seeking during periods of reward
unavailability. A BLA to NAcS pathway has also been shown to directly mediate the inhibition
of alcohol seeking through extinction, as contralateral disconnection of these two structures
disinhibits extinguished seeking behavior (Millan & McNally, 2011). Here we extend these
observations, illustrating that neural activity within both the BLA and NAcS are crucial for
inhibiting reward-seeking that may also yield aversive consequences.

111
It is notable that the effect of NAcS inactivation on responding during the Conflict period
was comparatively smaller to that induced by BLA inactivation, suggesting that other, parallel
output pathways from the BLA (e.g., the central amygdala or mPFC) may also contribute to
behavioral suppression in situations involving punishment. On the other hand, the NAc has been
suggested to act as a “limbic-motor interface” (Mogenson et al., 1980), integrating input from
cortico-limbic afferents to allow for appropriate action selection (Calhoon & O’Donnell, 2013b;
Gruber et al., 2009; O’Donnell & Grace, 1995), suggesting that other inputs to this nucleus may
also refine behavior in these situations. For example, the NAcS receives efferent input from
mPFC (Berendse, Galis-de Graaf, et al., 1992; Brog et al., 1993; Heilbronner et al., 2016), a
region that additionally contributes to punishment-induced suppression of behavior. Previous
studies have utilized lesions, dopamine antagonism, or optogenetic silencing of the mPFC to
cause persistent instrumental responding for reward despite the potential for punishment
(Broersen et al., 1995; Chen et al., 2013; Resstel et al., 2008). Similarly, in situations where an
aversively conditioned Pavlovian stimulus is presented during reward-seeking, inactivation of
this same region disrupts the typically observed conditioned suppression of lever-pressing, even
when punishment is omitted (Limpens et al., 2015). In Chapter 3, we illustrated that a PL mPFC
to NAcS projection mediates the expression of Pavlovian conditioned suppression, a process that
is likely related to instrumental suppression. A recent study provided additional support for this
hypothesis, illustrating that a mPFC neurons projecting to the lateral NAcS are active during, and
partially responsible for, the suppression of reward-seeking during potentially punished task
phases (Kim et al., 2017). Given this framework, it is plausible that the BLA and mPFC may
provide affective and contextual information to the NAcS, which activates neuronal ensembles to
execute motor programs that inhibit punished responding. Alternatively, the mPFC may provide

112
top-down control over the NAcS and/or BLA (St Onge et al., 2012), aiding in the refinement of
action selection when reward-seeking may result in aversive consequences.
Still, there exists debate on the precise role of mPFC in punishment, as excitotoxic
lesions of mPFC prior to cocaine seeking have been shown to not affect punished seeking
(Pelloux et al., 2013). In that study, permanent lesions of the mPFC were induced prior to self-
administration training, which may have allowed for some degree of long-term compensation
from other brain regions. However, a more recent study by Jean-Richard-Dit-Bressel & McNally
(2016) used reversible inactivations to illustrate that lateral, but not medial PFC contributes to
punishment-induced suppression. Specifically, inactivation of the lateral orbitofrontal cortex
(OFC) disinhibited punished lever-pressing, while inactivation of rostral agranular insular cortex
(RAIC) spared punished seeking, but increased the choice of the previously punished lever in a
shock-free choice test. These lateral frontal regions have been implicated in numerous functions
that may be relevant to punishment, including value encoding, interoception, and response-
inhibition in animals and humans (Bari & Robbins, 2013; Bechara, Damasio, & Damasio, 2000;
Bryden & Roesch, 2015; Clark et al., 2008; Craig, 2009; Morein-Zamir & Robbins, 2015), and
may contribute to punishment through their direct, sometimes reciprocal projections with the
extended amygdala and NAc (Heilbronner et al., 2016; Reynolds & Zahm, 2005; Shinonaga et
al., 1994). Future studies employing pharmacological or pharmacogenetic disconnection of these
structures may help determine the directionality of communication between these proposed
circuits underlying the punishment-induced inhibition of behavior.
There are a number of potential alternative explanations for the observed disinhibition of
punished responding following BLA and NAcS inactivation. For example, disinhibition during
Conflict may reflect a simple decrease in the expression of Pavlovian fear. Upon commencement

113
of the Conflict period, internal (e.g., timing) and external (e.g., cue light illumination) cues may
act to inhibit responding via the production of conditioned fear behaviors such as freezing.
However, we find this explanation unlikely for several reasons. Although we did not measure
conditioned fear in the present study, performance on a similar task was found to be
uncontaminated by conditioned fear, as freezing levels steadily declined across 5 days of
training, and fear-related freezing was unaffected by BLA inactivation (Jean-Richard-Dit-Bressel
& McNally, 2015). Given that training on our task was substantially longer (10-20 days), it is
likely that any conditioned fear produced by the Conflict period was eliminated over the course
of training. We also did not observe any change in locomotor activity following inactivation of
the BLA or NAcS, which may have been expected had we affected the expression of long epochs
of behavioral arrest. Additionally, freezing in response to shock-associated cues is not dependent
on the integrity of the NAcS (Haralambous & Westbrook, 1999; Thomas, Hall, & Everitt, 2002).
It is possible that indices of conditioned fear which incorporate a reward-seeking component,
such as conditioned suppression, may differentially depend on accumbens subregions, as
illustrated in Chapter 3. It is also unlikely that inactivations affected the unconditioned response
to foot-shock, as lesions of the NAc or BLA do not generally alter foot-shock-induced changes in
locomotion or lever-pressing (Levita et al., 2002; McDannald & Galarce, 2011; Schwienbacher
et al., 2004). Finally, the disinhibition of pressing observed following BLA or NAcS inactivation
may have resulted from rats simply being hungrier, or otherwise more motivated to seek food.
Muscimol infusion into the NAcS has been shown to produce orexigenic behavior in rats, but
only when food is freely available (Hanlon et al., 2004; Stratford & Kelley, 1997). However,
infusion of muscimol into the NAcS (at doses similar to those used here) does not impact
instrumental responding for food delivered on a progressive ratio schedule (Zhang et al., 2003).

114
Additional evidence against a simple, hunger-based explanation comes from the finding that
BLA and NAcS inactivation actually decreased responding during the Safe/Reward periods of
the Conflict task, and did not alter responding during any phase of the No-Conflict control task.
Given these considerations, we find it unlikely that the increase in punished responding for food
induced by BLA or NAcS inactivation was attributable to alterations in Pavlovian fear
mechanisms, foot-shock sensitivity, or enhanced motivation to obtain food. Rather, the present
findings suggest that these nuclei work in a cooperative manner to reorganize behavior and
suppress ongoing reward-seeking when these actions may also yield aversive outcomes.
4.4.3 Differential effects of BLA and NAcS inactivation on unpunished reward-seeking.
In addition to increasing punished reward-seeking, inactivation of the BLA and NAcS reduced
lever-pressing during the Safe/Reward phases of the Conflict task, when food was delivered on a
leaner, VI15/FR5 schedule. Yet, in a separate No-Conflict control experiment, inactivation of
these nuclei did not affect responding during the Safe/Reward or FR1 epochs, where rats pressed
for food on identical, shifting schedules of reinforcement, but did not experience foot-shocks at
any point during training. This lack of effect suggests that the reduction in responding during the
Safe/Reward phases of the Conflict task induced by BLA and NAcS inactivation may stem in
part from the history of punishment in this context differentially recruiting these regions during
appetitive behavior.
In an attempt to clarify the seemingly discrepant effects of BLA/NAcS inactivation on
reward-seeking, baseline responses of rats trained on the two tasks were analyzed, revealing
noticeable differences in how these groups allocated their relative response rates across the task
epochs. Those trained on the Conflict task displayed higher rates of responding during the
Safe/Reward phases compared to animals trained on the control task, presumably as a

115
compensatory measure for their reduced responding during the punished Conflict period.
Conversely, rats performing the No-Conflict task obtained considerably more food during the
middle, FR1 phase than those on the Conflict task, which may explain why their response rates
were lower during the Safe/Reward phases. This pattern of results suggests that a history of
punished reward-seeking alters the manner in which animals adjust their response rates to
changes in schedules of positive and negative reinforcement. Inactivation of the BLA or NAcS
prior to the Conflict task altered response profiles, so that behavior over the session resembled
that of animals performing the control task that never experienced punishment. Therefore, the
impact of BLA or NAcS inactivation on responding during the Conflict task may not simply
reflect the involvement of these regions in the suppression of punished responding and/or
invigoration of responding when the effort requirements are high. Rather, neural activity within
these nuclei may mediate a broader perception of the task context that enables appropriate
adjustments in behavioral output to reduce the occurrence of aversive consequences, while at the
same time attempting to maximize the amount of reward that may be obtained.
4.4.4 NAcC inactivation and motivated responding - comparisons with NAcS.
In contrast to the differential effects on responding induced by BLA or NAcS inactivation,
similar infusion of GABA agonists into the NAcC diminished reward-seeking across all phases
of both tasks, concomitant with a decrease in locomotion and other indices of motivated output.
These observations are perhaps unsurprising, as the NAcC has been shown to be necessary for
motivated behavior and flexible approach during appetitive reward-seeking across a variety of
experimental paradigms (Ambroggi et al., 2011; Di Ciano et al., 2008; Ghods-Sharifi &
Floresco, 2010; Ishikawa et al., 2008, 2010; Nicola, 2010; Parkinson, Willoughby, et al., 2000;
Stopper & Floresco, 2011). For example, behavioral responding to a discriminative incentive cue

116
which predicts reward availability is dependent on the NAcC and its cortico-limbic afferents
(Ambroggi et al., 2008; Ishikawa et al., 2008, 2010). Relatedly, flexible approach behavior is
governed by dopaminergic activity in the NAcC (McGinty et al., 2013; Nicola, 2010). Blockade
of dopamine receptors in the NAcC decreases the likelihood that rats trained on a cued FR8
schedule would respond for reward, as a function of spending more time off task. Dopamine
activity in the NAcC also appears critical for the ability of a cue to act as an incentive stimulus,
becoming imbued with the motivational properties of the reinforcer itself (Saunders & Robinson,
2012). Diminished locomotor activity may also contribute to the decrease in lever-pressing
observed following NAcC inactivation. However, locomotor activity was only significantly
decreased on the Conflict task, and not the No-Conflict control task, yet lever-pressing was
substantially decreased by NAcC inactivation on both tasks. This suggests that the impact of
NAcC inactivation on locomotion and lever-press behavior are partially dissociable based on
task history, and thus may be mediated by potentially separable mechanisms. Finally, an intact
NAcC has been shown to be necessary for appropriate effort expenditure during appetitive
behavior, as dopamine blockade (Nunes, Randall, Podurgiel, Correa, & Salamone, 2013;
Salamone, Correa, Farrar, & Mingote, 2007) or inactivation (Ghods-Sharifi & Floresco, 2010) of
this region decreases the amount of physical effort animals are willing to expend to receive a
larger reward. Therefore, the lower degree of task engagement observed following NAcC
inactivation in the present study likely reflects a decrease in willingness to exert effort to obtain
rewards and/or the impact of incentive stimuli on behavior.
Of particular interest is the dramatic contrast between the effects of NAcC versus NAcS
inactivation on these tasks. As described previously, the NAcS appears to be critical for
inhibiting punished responding, consistent with a broad literature implicating this region of the

117
NAc in facilitating optimal foraging behavior by inhibiting task-irrelevant behaviors (Ambroggi
et al., 2011; Floresco et al., 2008; Ishikawa et al., 2010). In contrast, the NAcC plays a key role
in promoting approach behavior towards motivationally relevant stimuli. Inactivation of NAcC
profoundly decreased reward-seeking during both tasks, while NAcS inactivation disinhibited
punished seeking behavior, and only affected safe reward-seeking in rats that had a history of
punishment during training. Furthermore, inactivation of the NAcC tended to cause
hypolocomotion and reduced the number of rewards received by rats on both tasks, while neither
effect was observed following NAcS inactivation. These findings complement a growing
literature that suggests that the NAcC and NAcS may play somewhat opposing, yet
complementary, roles in enabling an organism to obtain its goals (Floresco, 2015). Both nuclei,
via input from their upstream corticolimbic afferents, act in concert to optimize goal directed
behavior, although they appear to do so in distinct manners.
4.4.5 AMPH tends to promote punishment-sensitivity during conflict
Previous research has suggested that pharmacological enhancement of monoamine release
increases punishment susceptibility, biasing behavior away from an instrumental response that is
concurrently rewarded and punished (Broersen et al., 1995; Killcross et al., 1997; Lazareno,
1979; Leone et al., 1983). Here, results of an exploratory analysis provided support for this
account, with AMPH decreasing reward-seeking specifically during the Conflict phase. AMPH
is a potent releaser of the monoamines, including dopamine and serotonin, both of which have
been implicated in response-inhibition (Crockett, Clark, & Robbins, 2009; Killcross et al., 1997;
Pascoli et al., 2015; Simon et al., 2011). For example, AMPH alters performance on a task where
foot-shock is probabilistically associated with one instrumental response that delivers are large
amount of reward, but not another that delivers a small reward (Mitchell, Vokes, Blankenship,

118
Simon, & Setlow, 2011; Orsini, Trotta, et al., 2015; Simon et al., 2011; Simon, Gilbert, Mayse,
Bizon, & Setlow, 2009). On this task, AMPH induces a risk-averse phenotype, biasing choice
away from the instrumental response that probabilistically delivers large reward and punishment,
and towards the instrumental option that is safe, but objectively less rewarding. This potentiation
of risk aversion induced by AMPH has been shown to be mediated in part by dopamine, as
blockade of the D2 receptor reduces the impact of AMPH on choice (Simon et al., 2011).
Although the present task did not allow animals to choose between multiple options during the
Conflict period, animals were more likely to withhold responding during this phase, as a function
of AMPH treatment.
The risk-aversion induced by AMPH in studies using foot-shock punishment contrasts
with other studies that operationalize risk and punishment as reward omission, during which
AMPH promotes risk-seeking (St. Onge & Floresco, 2009). This discrepancy has been suggested
to relate to the ability of AMPH to enhance the salience of relevant task events (Orsini,
Moorman, Young, Setlow, & Floresco, 2015). On tasks employing foot-shock punishment, the
delivery of this aversive stimulus is more salient than is the difference in reward magnitude, and
thus AMPH induces risk-aversion. On tasks where reward omission serves as punishment, the
receipt of a large reward may be the more salient factor, such that AMPH biases choice towards
instrumental actions that may result in that reward, which will manifest as risk aversion. In
support of this, other tasks with arguably more salient omission periods, such as the rodent
gambling task, show the inverse effect of AMPH, with animals becoming more sensitive to
reward omission punishment following treatment (Zeeb, Robbins, & Winstanley, 2009). The
results of the present study are broadly consistent with this dissociation, as punishment is clearly
more salient than the relatively richer schedule of reinforcement during the Conflict period, as

119
evidenced by the large disparity in lever-presses made during the Safe/Reward phases versus the
Conflict phase. Under these conditions, AMPH would be expected to produce risk aversion,
which was broadly confirmed here.
4.4.6 Relevance for psychiatric disorders
The findings that the BLA and NAcS both contribute to suppressing punished reward-seeking
may provide insight into how dysfunction of these circuits contributes to the compulsive
behaviors observed in a variety of psychiatric disorders. Compulsivity in the face of punishment
is a hallmark of drug addiction and obsessive-compulsive disorder (OCD) (Figee et al., 2016;
Morein-Zamir & Robbins, 2015). Structures that promote punishment-induced behavioral
suppression, such as mPFC and BLA, project directly to NAc and are central to the pathology of
both disorders (Wood & Ahmari, 2015). mPFC hypoactivity contributes to deficient top-down
inhibition of drug seeking in rodents (Chen et al., 2013; Limpens et al., 2015), and is correlated
with inhibitory control deficits in cocaine users (Morein-Zamir, Simon Jones, Bullmore,
Robbins, & Ersche, 2013). Furthermore, abstinence from cocaine use is related to improvements
in prefrontal cortical function, suggesting that the successful cessation of drug use is either
predicated on or causally related to normalized cortical activity (Connolly, Foxe, Nierenberg,
Shpaner, & Garavan, 2012). Self-administration of most addictive substances induces
dysregulation of the dopaminergic projections to the NAc, combined with altered NAc plasticity
(Britt & Bonci, 2013; Grueter, Rothwell, & Malenka, 2012; Russo et al., 2010). Prolonged drug
exposure can downregulate dopamine D2 receptor levels in the ventral striatum, of which the
nucleus accumbens is a large part, which is thought to produce impulsive behavior (Lee et al.,
2009; Volkow, Fowler, Wang, Baler, & Telang, 2009). In OCD, the ventral striatum receives
abnormally-elevated afferent input from the orbitofrontal cortex (Abe et al., 2015). Evidence

120
from preclinical models suggests that activity in this pathway may underlie the compulsions
observed in individuals with OCD (Figee et al., 2016; Wood & Ahmari, 2015). Although the
meso-cortico-limbic-striatal circuit overlap between the two disorders is apparent, more work is
required to determine the direction of change and relation to punishment-induced inhibition of
responding. Additional exploration of different nodes within meso-cortico-limbic-striatal
circuitry that contribute to these aspects of behavior may allow for a better understanding of
underlying neuropathophysiology of these disease states. To this end, the present results suggest
that abnormal functioning of the BLA and NAcS may be a contributing factor to compulsive
behaviors associated with these conditions.
4.5 Conclusion
These findings point to complementary roles for the BLA and NAcS in suppressing appetitively-
motivated behaviors in the face of punishment. This form of response-suppression mechanism is
adaptive, with survival often predicated on weighing potential benefits against punishments
when seeking food, or other primary rewards. In addition, all of the regions investigated here
played an important role during safe reward-seeking, although NAcS and BLA were selectively
recruited following a history of punishment, as performance was spared on a punishment-free
control task. We also observed that promoting monoamine release, via systemic treatment with
AMPH, resulted in less reward-seeking during punishment, consistent with previous findings.
Overall, these results may be relevant for neuropsychiatric disorders where compulsive behavior
is resistant to punishment, including substance abuse and obsessive-compulsive disorder (Everitt,
2014; Figee et al., 2016; Morein-Zamir & Robbins, 2015; Wood & Ahmari, 2015). They also
provide novel insights into a subregion-specific bivalent function of the rodent NAc, and suggest
a possible circuit basis for this divergent effect. In summary, our work suggests that BLA and the

121
NAcS are recruited during punishment-induced inhibition of behavior, while the NAcC is
recruited to actively promote seeking behavior, irrespective of punishment.
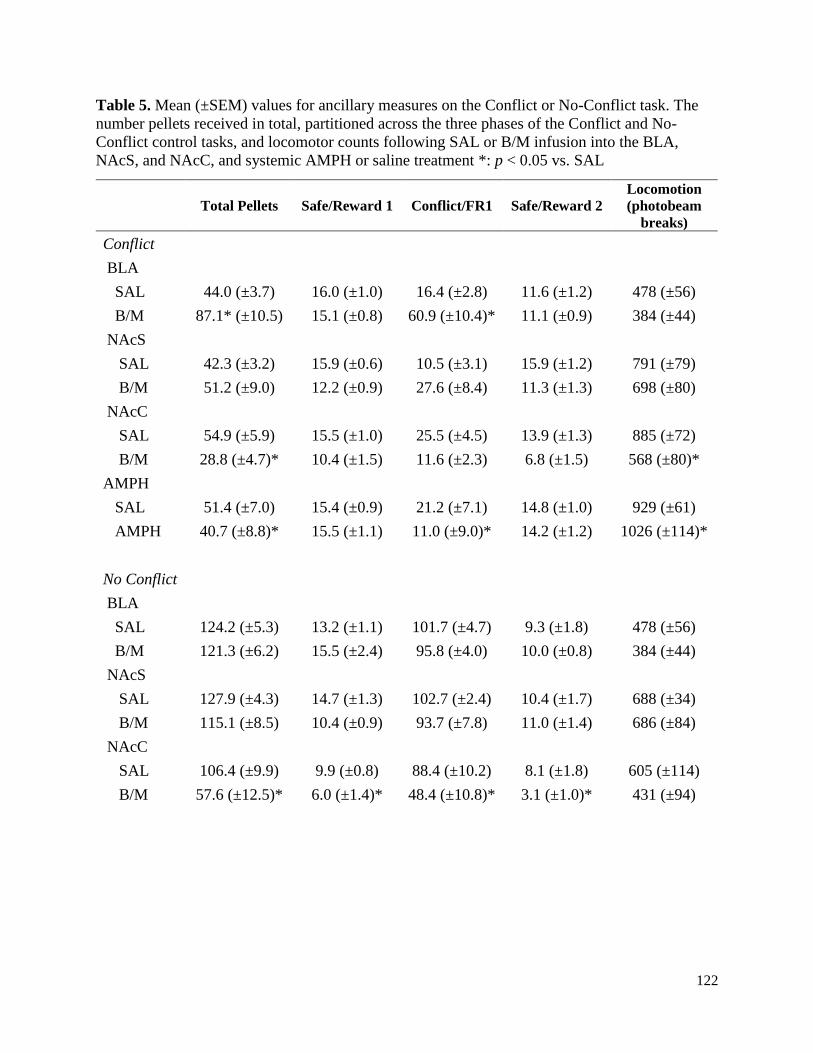
122
Table 5. Mean (±SEM) values for ancillary measures on the Conflict or No-Conflict task. The
number pellets received in total, partitioned across the three phases of the Conflict and No-
Conflict control tasks, and locomotor counts following SAL or B/M infusion into the BLA,
NAcS, and NAcC, and systemic AMPH or saline treatment *: p < 0.05 vs. SAL
Total Pellets Safe/Reward 1 Conflict/FR1 Safe/Reward 2
Locomotion
(photobeam
breaks)
Conflict
BLA
SAL 44.0 (±3.7) 16.0 (±1.0) 16.4 (±2.8) 11.6 (±1.2) 478 (±56)
B/M 87.1* (±10.5) 15.1 (±0.8) 60.9 (±10.4)* 11.1 (±0.9) 384 (±44)
NAcS
SAL 42.3 (±3.2) 15.9 (±0.6) 10.5 (±3.1) 15.9 (±1.2) 791 (±79)
B/M 51.2 (±9.0) 12.2 (±0.9) 27.6 (±8.4) 11.3 (±1.3) 698 (±80)
NAcC
SAL 54.9 (±5.9) 15.5 (±1.0) 25.5 (±4.5) 13.9 (±1.3) 885 (±72)
B/M 28.8 (±4.7)* 10.4 (±1.5) 11.6 (±2.3) 6.8 (±1.5) 568 (±80)*
AMPH
SAL 51.4 (±7.0) 15.4 (±0.9) 21.2 (±7.1) 14.8 (±1.0) 929 (±61)
AMPH 40.7 (±8.8)* 15.5 (±1.1) 11.0 (±9.0)* 14.2 (±1.2) 1026 (±114)*
No Conflict
BLA
SAL 124.2 (±5.3) 13.2 (±1.1) 101.7 (±4.7) 9.3 (±1.8) 478 (±56)
B/M 121.3 (±6.2) 15.5 (±2.4) 95.8 (±4.0) 10.0 (±0.8) 384 (±44)
NAcS
SAL 127.9 (±4.3) 14.7 (±1.3) 102.7 (±2.4) 10.4 (±1.7) 688 (±34)
B/M 115.1 (±8.5) 10.4 (±0.9) 93.7 (±7.8) 11.0 (±1.4) 686 (±84)
NAcC
SAL 106.4 (±9.9) 9.9 (±0.8) 88.4 (±10.2) 8.1 (±1.8) 605 (±114)
B/M 57.6 (±12.5)* 6.0 (±1.4)* 48.4 (±10.8)* 3.1 (±1.0)* 431 (±94)

123
Figure 10. Histology schematic for Conflict and No-Conflict task animals
Histology schematic for Conflict task animals with cannula located in the BLA (A), or NAc
subregions (B), as well as on the No-Conflict control task (C). All symbols indicate the most
ventral point of infusion in the BLA (A; black squares) or NAc (B, C; black triangles = NAcS
placement, grey circles = NAcC placement). Numbers to the left of each representative atlas
section indicate distance (mm) from bregma.

124
Figure 11. Task diagram and data from pharmacological manipulation on the Conflict task
(A) Flow-chart of a daily Conflict task session indicating the schedules of food reinforcement
and punishment. (B, C, D, E) The left, center, and right graphs represent the first Safe/Reward
period, the Conflict period, and the second Safe/Reward period, respectively. Data are presented
as mean ± SEM. (B) BLA inactivation with baclofen/muscimol (B/M) decreased output during
Safe periods (left, right), but dramatically disinhibited punished responding during Conflict
(center), relative to saline (SAL) control treatments. (C) Similarly, NAcS inactivation reduced
lever-pressing during both Safe/Reward periods (left, right), and disinhibited pressing during the
Conflict period (center). (D) NAcC inactivation diminished motivated output, regardless of task
phase. (E) Exploratory analysis revealed that AMPH tended to promote response-inhibition
during the Conflict phase, without affecting performance during the Safe/Reward phases. Closed
star denotes p < 0.05 between SAL and B/M or SAL and AMPH treatment during a particular
task phase.

125
Figure 12. Task diagram and data from inactivations on the No-Conflict task
(A) Flow-chart of a daily No-Conflict control task session. Note that this task differs from the
Conflict task only in the fact that no punishment was ever delivered during the middle epoch
where food was delivered on an FR1 schedule of reinforcement. (B, C, D) The left, center, and
right graphs represent the first Safe/Reward period, the “Conflict” period, and the second
Safe/Reward period, respectively. Neither BLA (B) nor NAcS (C) inactivation (B/M) had any
significant effect on performance. (D) NAcC inactivation decreased reward-seeking across all
phases of the No-Conflict control task, similar to the effect of inactivation during the Conflict
task. Note the difference in scaling during the FR1 period (middle panels) compared to that used
for the conflict period data displayed in Figure 2. Closed star denotes p<0.05 between SAL and
B/M infusion during a particular task phase.

126
Figure 13. Baseline analysis suggests NAcS and BLA promote reward seeking as a function
of task history.
(A) Combined task diagram for the Conflict and No-Conflict control task. Both tasks had
identical Safe/Reward periods (left and right columns), but different during the “Conflict” period
(middle column). (B, C) The left, center, and right graphs represent the first Safe/Reward period,
the “Conflict” period, and the second Safe/Reward period, respectively. (B) Rats trained on the
Conflict task pressed maximally during the Safe/Reward periods, and markedly suppressed their
pressing during the Conflict period, whereas rats trained on the No-Conflict control task
displayed the inverse. (C) The same pattern of results was found when examining pellets
received. (D) Direct comparison of lever-press behavior during the Safe/Reward phases. The left
graph displays the performance of rats that received intra-BLA B/M during the Conflict task to
those that received intra-BLA SAL during the No-Conflict Control task. The right graph displays
the performance of rats that received intra-NAcS B/M during the Conflict task to those that

127
received intra-NAcS SAL during the No-Conflict Control task. There was no difference in the
number of presses made during the Safe/Reward phases between rats that received SAL during
the No-Conflict task and inactivation during the Conflict task for either BLA (left graph) or
NAcS (right graph), suggesting that inactivation of BLA or NAcS may induce a behavioral state
similar to saline-infused rats trained on the No-Conflict task that never encountered foot-shock
during training. Closed star denotes p < 0.05 between task conditions during a particular task
phase. n.s. = not significant.

128
Chapter 5: Dissociable contributions of NAc core and shell during active/passive avoidance
5.1 Introduction
When faced with a potential threat, animals may employ one of two main types of defensive
behaviors: defensive reactions and defensive actions (LeDoux, 2012; Moscarello & Ledoux,
2014). Defensive reactions are designed to evade predator detection and, in rodents, include
forms of behavioral suppression such as freezing. These reactions can facilitate the passive
avoidance of dangerous or threatening stimuli. Conversely, defensive actions are typically
instrumental behaviors which enable the organism to actively avoid or escape threat. Both active
and passive avoidance responses serve adaptive functions, with their flexible application,
conducted in accordance with environmental contingencies, being critical to survival.
This dichotomy of active versus passive defensive strategies may be viewed analogously
to processes that govern appetitive behavior. For example, Go/No-Go conditioning generally
requires an active approach response to receive reward in the presence of one cue (a “Go”
response), while another cue signals that suppressing approach (a “No-Go” response) results in
reward delivery. It is well-established that different aspects of appetitively-motivated behavior
are predicated on activity in meso-cortico-limbic-striatal circuitry. A particularly crucial node in
this network is the nucleus accumbens (NAc), which integrates diverse limbic, cognitive, and
neuromodulatory input to promote flexible action selection (Calhoon & O’Donnell, 2013b;
Floresco, 2015; Gruber et al., 2009; Gruber & O’Donnell, 2009; Mogenson et al., 1980). The
NAc has been further partitioned into lateral core (NAcC) and medial shell (NAcS) regions,
based on neuroanatomical and functional differences (for review, see Heimer et al, 1997; Zahm
and Brog, 1992), with these regions often playing dissociable, yet complementary, roles in
guiding motivated behavior.

129
The NAcC has been proposed to promote active approach behaviors, while the NAcS
may fulfill a dual role, inhibiting inappropriate responses while also aiding in the production of
active behaviors (Ambroggi et al., 2011; Blaiss & Janak, 2009; Floresco, 2015; Floresco et al.,
2008; Ghazizadeh et al., 2012; Ghods-Sharifi & Floresco, 2010; Piantadosi et al., 2017). For
example, neurophysiological studies have shown that neurons in both the NAcC and NAcS
encode a discriminative stimulus that signals reward availability, yet a higher proportion of
neurons in the NAcS (as compared to NAcC) also encode a neutral stimulus that signals reward
unavailability (Ambroggi et al., 2011). Inactivation of the NAcC preferentially affects behavior
elicited by reward-predictive stimuli, while inactivation of NAcS unmasks irrelevant behaviors
such as lever-pressing and Pavlovian approach during presentation of non-rewarded stimuli and
intertrial intervals (Ambroggi et al., 2011; Blaiss & Janak, 2009). The NAcS (but not NAcC) has
also been suggested to actively inhibit extinguished and non-reinforced instrumental behavior
during the reinstatement of food (Floresco et al., 2008), alcohol (Millan et al., 2010), or cocaine
seeking (Peters et al., 2008). Consideration of these data implies that these two nuclei facilitate
reward-seeking in partially distinct ways, with the NAcS enforcing response-inhibition to focus
and constrain behavioral output, and the NAcC promoting approach towards relevant stimuli.
Although the NAc is typically viewed as a “reward” nucleus, it is important to note that
neurons within this region are also responsive to aversive stimuli and the cues that predict them
(Delgado, Li, et al., 2008; Jensen et al., 2003; Roitman, Wheeler, & Carelli, 2005; Schoenbaum
& Setlow, 2003; Setlow, Schoenbaum, & Gallagher, 2003). For example, on a mixed valence
Go/No-Go task, largely separate populations of NAc neurons develop phasic responses to cues
that predict appetitive or aversive outcomes (Setlow et al., 2003). These responses may facilitate
behavioral flexibility in both appetitive and aversive contexts, allowing for active responses to be

130
elicited to obtain rewards, while also enabling the response-suppression necessary to avoid
punishment. Interestingly, data from a similar Go/No-Go task suggests that NAc neurons track
the behavioral response necessitated by a Go or No-Go cue, in keeping with a role for this
nucleus in action selection (Roitman & Loriaux, 2014). Consistent with this idea, we have
recently shown that subregions of the NAc are differentially responsible for the promotion and
inhibition of reward-seeking during punishment (Chapter 4). Specifically, inactivation of the
NAcS disinhibited punished reward-seeking, whereas similar inactivation of the NAcC induced a
general suppression of instrumental responding for reward (Chapter 4; Piantadosi et al., 2017).
Similarly, the NAcS disinhibited reward-seeking despite the presentation of an aversive
Pavlovian conditioned stimulus, while NAcC simply promoted behavioral activation (Chapter 2
& 3). However, it remains unclear whether these two NAc subregions perform dissociable
functions during action-selection motivated exclusively by cues that predict aversive outcomes.
Of particular interest would be whether the NAcS and NAcC are differentially responsible for
the performance of defensive reactions versus actions in response to discrete cues.
Previous work has separately examined the contribution of the NAc to these two types of
defensive behaviors. With respect to defensive actions, the NAc and its dopaminergic input are
integral for the learning and expression of “Go”-like actions such as active avoidance (Fernando
et al., 2013; Gentry et al., 2016; Ilango, Shumake, Wetzel, & Ohl, 2014; Lichtenberg,
Kashtelyan, Burton, Bissonette, & Roesch, 2014; Oleson et al., 2012; Ramirez et al., 2015;
Salamone, 1994). In particular, inactivation of NAcS, or disconnection of amygdalar inputs to
this nucleus impairs the expression of active avoidance (Fernando et al., 2013; Ramirez et al.,
2015). In comparison, avoidance expression does not appear to be affected by NAcC inactivation
(Ramirez et al., 2015). Yet, DA release in the NAcC increases during the presentation of an

131
active avoidance cue, suggesting that transmission in this region may be relevant for the
execution of this behavior (Gentry et al., 2016; Oleson et al., 2012). Thus, both subnuclei of the
accumbens may contribute to aversively-motivated active behaviors that avoid negative
consequences.
In comparison to its role in active avoidance, neurotransmission in the NAc has been
shown to be necessary for the acquisition, but not expression, of defensive reactions such as
passive avoidance, as measured by latency on one-trial step-through tasks (Bracs, Gregory, &
Jackson, 1984; De Leonibus et al., 2003; Lorenzini, Baldi, Bucherelli, & Tassoni, 1995;
Martínez et al., 2002; Shirayama et al., 2015). When conducted prior to learning, manipulations
that perturb NAc functioning cause rats to approach a context previously associated with foot-
shock more rapidly than control rats, although these effects are typically absent when conducted
prior to expression. Unlike active avoidance, this mnemonic test is acute and not amenable to
repeated testing. In addition, the difficulty posed by a No-Go trial during Go/No-Go performance
is enhanced by the necessity to accurately discriminate between discrete Go vs. No-Go stimuli,
and then withhold a prepotent response. These crucial aspects of passive avoidance behavior are
not captured by such one-trial step-through tasks. Thus, development of a task that can
adequately measure the flexibility and repetition associated with fully aversively-motivated “Go”
vs. “No-Go” performance is necessary. In this regard, gerbils have been trained to perform a
two-way active avoidance procedure, whereby two different conditioned stimuli necessitate
either a passive or active avoidance response in order to avoid foot-shock (Schulz, Woldeit,
Gonçalves, Saldeitis, & Ohl, 2015; Stark, Rothe, Wagner, & Scheich, 2004; Wetzel, Ohl, &
Scheich, 2008). During one auditory stimulus, animals were required to make an instrumental
shuttling response to avoid a foot-shock, while presentation of the other auditory stimulus

132
required the inhibition of a shuttling response. Acquisition of this task has been shown to
increase dopamine release in the prefrontal cortex (Stark et al., 2004), similar to other forms of
behavioral flexibility (for review, see Floresco, 2013). Interestingly, coherence between the
auditory cortex and ventral striatum, of which the NAc is a primary component, increases
following presentation of the active avoidance stimulus over the course of training on this task
(Schulz et al., 2015). These later results suggest that the NAc may integrate afferent input to
accurately promote or inhibit responding during complex avoidance performance.
Additional insight into accumbal contributions to active versus passive avoidance comes
from functional imaging studies conducted with human subjects (Levita et al., 2009, 2012). In
one study, participants were trained to discriminate between two visual stimuli that instructed
them to either press a button to make an active avoidance response or passively withhold a
response to avoid an aversive outcome. Performance of an active avoidance response induced an
increase in BOLD signal within the NAc, yet successful passive avoidance trials were associated
a deactivation in this region (Levita et al., 2012). This pattern of activation/deactivation suggests
that the NAc may function to promote active avoidance, while suppression is necessary for
appropriate inhibition during passive avoidance. Unfortunately, the constraints on spatial
resolution imposed by fMRI in that study did not permit a more detailed characterization of how
changes in activation within different subregions of the NAc may be associated with different
types of avoidance responses. Developing a preclinical analog of this task would aid in clarifying
the role of different brain nuclei in the appropriate promotion versus suppression of aversively-
motivated behavior, as well as generally improving our understanding of complex avoidance
behaviors.

133
Here we report on the development of a novel operant task that required rats to use
discriminative cues that informed them of whether an impending foot-shock could be avoided by
either pressing a lever or withholding a lever press, permitting the examination of the neural
basis of the active versus passive poles of avoidance behavior. Using reversible inactivation, we
explored the contribution of the NAcC or NAcS to these different aspects of behavior. We
hypothesized that inactivation of NAcC, which is involved in Pavlovian and instrumental
approach, would impair active avoidance selectively. On the other hand, we expected that
inactivation of the NAcS would not only impair approach-mediated active avoidance, but also
perturb the suppression of behavior during passive avoidance trials. In addition, we probed
potential monoaminergic contributions to active/passive avoidance behavior by investigating the
effect of systemically administered d-amphetamine (AMPH).
5.2 Methods
Active/Passive Avoidance training was adapted from previous reports conducting active
avoidance in an operant setting (Fernando, Mar, Urcelay, Dickinson, & Robbins, 2015; Fernando
et al., 2013; McCullough et al., 1993; Sokolowski, McCullough, & Salamone, 1994), and based
on a paradigm used in humans, as described by Levita et al (2012).
5.2.1 Animals
All experimental protocols were approved by the Animal Care Committee, University of British
Columbia, and conducted in compliance with guidelines provided by the Canadian Council on
Animal Care. All reasonable efforts were made to minimize the number and suffering of animals
used. Male Long-Evans rats arrived weighing 250-275 g for active/passive avoidance training
and 325-350 g for foot-shock sensitivity (Charles River) and were initially group housed (4-5 per

134
cage) and allowed to acclimatize to the vivarium for 6-7 days. The temperature (21° C) and light
cycle (12-h light/dark) were kept constant.
Forty-eight total rats were utilized in the Active/Passive experiments, with three separate
cohorts of n = 16 rats tested experimentally over the course of approximately 18-24 mo. For the
foot-shock sensitivity experiments, one cohort of animals (n = 12) was given surgery and tested
over a 3 wk period. All animals (except those used to test foot-shock sensitivity) were initially
food-restricted to approximately 90% of their free-feeding weight, to promote exploration and
exploitation of the operant environment, even though the task did not use food as a reinforcer.
Throughout the course of the experiment, rats were allowed to gain weight at a rate of
approximately 5 g/wk, maintaining a slightly delayed growth curve.
5.2.2 Apparatus
Eight standard Med Associates (St. Albans VT, USA) operant conditioning chambers were used
for all training and testing, as previously described (Piantadosi et al., 2017). A sound attenuating
enclosure (30.5 cm x 24 cm x 21 cm) surrounded the operant chamber, providing ventilation and
masking external noise via a fan. Each operant chamber contained two retractable levers on one
wall, with a food receptacle in the middle (although no food was delivered in these experiments).
Only the left operant lever was inserted into the chamber during these experiments. Each box
was outfitted with three 100 mA cue lights, situated above the operant levers and the food
receptacle. The opposite wall of the operant chamber contained a centrally located 100 mA
house light, and an audio speaker that allowed for delivery of auditory stimuli via a
programmable generator (ANL-926, Med Associates). Locomotor activity was monitored by
four infrared photobeam sensors located slightly above the stainless-steel grid floor. The grid
floor was wired to a shock source and solid-state grid scrambler for the delivery of foot-shock.

135
5.2.3 Initial lever shaping
After reaching approximately 90% of their free-feeding weight, rats began to receive daily
(conducted 5-7 d per week at a consistent hour) operant sessions. During the first session, rats
were placed in the operant box, which was illuminated by the house light. No lever was extended
for the duration of this session, and rats were simply allowed to locomote inside the chamber for
1 hr. This session served to reduce the neophobia associated with the novel environment, and
allow rats to familiarize themselves with the environment. In turn, the insertion of the lever on
the subsequent training day would be a novel stimulus that would elicit approach.
The day after this habituation session, rats underwent a lever-retraction training session,
which consisted of the left operant lever being extended into the box under constant illumination
of the house-light. During the 60 min lever-retraction training session, a press on the lever
caused it to retract for 1 s, followed by its reinsertion. This procedure allowed the rat to learn the
mechanics of the operant lever. If rats did not respond on this operant lever during the initial
session, a small amount of sucrose powder was placed on the lever to entice the rat to produce an
operant response. Note that this was the only point of the entire training where rats may have
experienced some food in the chamber. All rats completed lever-retract training in 1-3 days. Rats
performed a mean of 155 ± 24 SEM lever-presses during their final lever-retraction training
session (range: 15-968 presses).
5.2.4 Active avoidance training
After progressing from lever-retraction training, rats began the initial phase of active avoidance
training. This task consisted of 20 discrete active avoidance trials, each of which occurred after a
105 (± 30) s ITI. A trial began with the left operant lever being inserted into the chamber and an

136
auditory cue played simultaneously. Across separate rats, the auditory cue varied between three
distinct tones: a white noise cue (0 Hz, 80 dB), a high pitch pure tone (9 kHz, 80 dB), and a low
pitch pure tone (1 kHz, 80 dB). The tone assigned to signal an active avoidance trial remained
consistent throughout the experiment. In the initial portion of training, the signaled active
avoidance period was 20 s. A lever-press during this period terminated the tone and resulted in
the retraction of the lever. The house light was then extinguished, and a 30 s visual safety signal
(illumination of the central cue light, located in between the two retractable levers) was
presented. Presentation of a safety signal reinforces avoidance learning by explicitly signaling
successful avoidance, and thus, safety (Berger & Brush, 1975; Dinsmoor, 2001; Dinsmoor &
Sears, 1973; Fernando et al., 2013; Fernando, Urcelay, Mar, Dickinson, & Robbins, 2014;
Morris, 1975). If rats took longer than 20 s to make a lever-press, the active avoidance auditory
cue terminated and the escape period began, during which rats received a foot-shock at the end
of the 20 s active avoidance period, and then again 5 and 10 s later (i.e. 25 and 30 s post-
cue/lever presentation). As escape behavior typically precedes the development of active
avoidance performance, successful escapes were also reinforced with the delivery of the same 30
s safety signal (Solomon & Wynne, 1953). Responses within the first 20 s were termed
successful active avoidances, while responses made during the subsequent 10 s escape period
(following at least 1 foot-shock) were classified as escapes. A lack of a response during either
the 20 s avoidance or 10 s escape period caused the lever to retract, termination of the tone and
house light, and the trial was scored as an active avoidance failure.
On the first day of training, foot-shock intensity was set to 0.2 mA, and then individually
titrated in 0.05 mA increments throughout training, such that rats ideally remained motivated via
negative reinforcement to perform an active avoidance response (Fernando et al., 2015). Once

137
rats made approximately > 60% active avoidance or escape responses on the initial training task
(i.e. < 40% failures), the avoidance period was decreased to 15 s. Rats were then trained on this
15 s active avoidance task to the same criterion (< 40% failure). A small percentage of rats that
progressed beyond this portion of the task and began performing poorly were given remedial
sessions on active avoidance, in order to rescue performance.
5.2.5 Active/passive avoidance training
Following the initial active avoidance training, rats were trained on a blocked version of the
active/passive avoidance task. During this task, 12 active avoidance trials (identical to those
described previously, 15 s active avoidance period) and 12 passive avoidance trials were
presented (Fig. 14A). During passive avoidance trials, one of the three tones not used for active
avoidance (counterbalanced across rats) was presented at the same time as insertion of the left
operant lever. On these trials, after insertion of the lever, rats were required to withhold a lever
press for 15 s to avoid a food-shock. After a successful passive avoidance trial, the lever was
retracted, and a 30 s safety cue (same as active avoidance training) was presented. In contrast, a
lever press during a passive avoidance trial resulted in the immediate delivery of a foot-shock of
the individually titrated intensity, and was scored as a passive avoidance failure. If a rat made a
press during a passive avoidance trial, the lever remained extended until the 15 s passive
avoidance period elapsed. Thus, rats could make multiple presses during these trials, with each
press resulting in foot-shock. The number of lever presses made during passive avoidance trials
were recorded.
This initial active/passive avoidance training was conducted in a blocked design.
Typically, each session began with 12 active avoidance trials and ended with 12 passive
avoidance trials. In order to familiarize rats with the eventual randomized presentation of active

138
avoidance and passive avoidance trials, rats also received days where trials were presented in the
opposite order, 12 passive avoidance trials followed by 12 active avoidance trials. Typically,
performance on the variant where active avoidance trials preceded passive avoidance trials was
better than performance on the opposite (passive followed by active) variant. Thus, the criteria
for successful acquisition of the active/passive avoidance contingency was < 50% failure on
active avoidance and passive avoidance trials, combined, during the passive followed by active
version of the task.
After acquiring the active/passive avoidance contingency in a blocked design, rats began
daily sessions of a fully randomized final version of the task (Fig. 14A). Each session again
consisted of 12 active avoidance and 12 passive avoidance trials, pseudorandomly presented
according to a programmed sequence. All task parameters were otherwise identical to the
previous training stage. Rats were trained on this intermixed version of the active/passive
avoidance task until reaching a final task criterion of > 50 % success on both active and passive
avoidance trials. As with the previous portions of training, a small percentage of rats that
progressed beyond this phase of the task and began performing poorly were given remedial
sessions on the blocked active avoidance and passive avoidance design, in order to rescue
performance.
Upon reaching the final active/passive avoidance performance criterion, rats underwent
stereotaxic surgery for the implantation of guide cannula into the NAcC or NAcS. Following
post-surgical recovery, rats were retrained to criterion before pharmacological testing.

139
5.2.6 Surgery
Rats were initially anesthetized with ketamine (50 mg/kg, i.p.) and xylazine (5 mg/kg,
i.p.). Following this induction protocol, rats were prepped for surgery, placed in a stereotaxic
frame, and maintained on isoflurane anesthesia (2-3% isoflurane concentration) for the duration
of the procedure. Twenty-three-gauge bilateral stainless-steel guide cannula were aimed at the
NAcS or NAcC, according to the following stereotaxic coordinates (in mm):
NAcS – from bregma, AP: +1.3, ML: ±1.0, from dura, DV: -6.3
NAcC – from bregma, AP: +1.6, ML: ±1.8, from dura, DV: -6.3
Four stainless-steel skull screws were inundated with dental acrylic, holding the cannula in place.
Stainless-steel obturators flush with the end of the guide cannula were inserted into the guide
cannula at the conclusion of surgery. Rats were allowed 7-10 days to recover from surgery prior
to either being re-trained on the active/passive avoidance task, or tested for foot-shock
sensitivity.
5.2.7 Microinfusion and Systemic AMPH Administration
Prior to any mock or microinfusion, all rats (except those used to test foot-shock sensitivity)
were required to perform stably across three straight days, with < 25% variation in the
percentage of active avoidance and passive avoidance successes. These rats initially received a
mock infusion 10 min prior to their regular training session, during which obturators were
removed, and a stainless-steel mock injector flush with the end of the guide cannula was inserted
for approximately 2 min. During this duration, rats were placed into a small enclosure and
allowed to freely move. Following this mock infusion day, rats were subjected to the first of two
microinfusion test days. These test days were counterbalanced, such that roughly half of all

140
animals received bilateral infusion of a solution containing the GABA agonists baclofen and
muscimol (B/M; 75 ng each in 0.3 ul/side), while the others received infusion of 0.9% saline
alone (SAL; 0.3 µl/side). Each infusion took place over 45 s, with the microinfusion injectors left
in place for an additional 60 s to allow for the infusate to diffuse from the injector tip. Following
the initial microinfusion test day, rats were retrained over the course of at least two days until
they again displayed criterion performance, after which they received their second,
counterbalanced microinfusion test. We have previously used this dose/volume to behaviorally
dissociate between the NAcC and NAcS during an approach/avoidance Conflict task (Piantadosi
et al., 2017), as well as a number of other behavioral assays of cognition and motivation (Dalton
et al., 2014; Floresco et al., 2008; Ghods-Sharifi & Floresco, 2010; Stopper & Floresco, 2011).
To ensure that only data from animals that understood the task contingencies were
included in the regional inactivation analyses, a criterion of > 50% successful active avoidance
and < 50% failure on passive avoidance during their saline infusion day was set. Details
regarding the ramification of this exclusion criteria are described below (see Task Acquisition
and Baseline Performance).
Following successful completion of two microinfusion test days, a subset of rats (n = 20;
10 each from NAcC and NAcS) were given an additional two test days, during which they
received an injection (i.p.) of 0.9% saline (1 ml/kg) or a 1 mg/kg (delivered in 1ml/kg of 0.9%
saline) dose of AMPH. These test days were counterbalanced and separated by at least two re-
training sessions. Each injection was given 10 minutes prior to placing the rat in the operant box.
This subset of animals included some rats that were excluded from regional inactivation analysis
due to missed cannula placements (n = 3). Systemic AMPH manipulation was not conducted on

141
the first cohort of rats (n = 16), as this experiment was conceived of following the completion of
this experimental group.
5.2.8 Foot-Shock Sensitivity
Separate groups of animals underwent a foot-shock sensitivity experiment, to examine whether
changes in avoidance behavior following NAc subregion inactivation could be explained by
alterations in pain sensitivity. Procedures were based off of established protocols (Pang et al.,
2010; Quirk et al., 2000; Tian et al., 2011). Animals were initially implanted with guide cannula
aimed at the NAcC or NAcS, and allowed to recover (see Surgery). On the first day, all rats were
placed into the operant chamber for 1 hr under constant illumination of the house light. The door
to the sound attenuating enclosure surrounding the chamber was left open, as animals needed to
be visible during foot-shock delivery on the following test days. The day after this locomotion
session, rats underwent the first of two foot-shock sensitivity test days. Half of the rats within
each region were infused with saline or B/M, in an identical manner as described above (see
Microinfusion and systemic AMPH administration). The rat was then placed into the operant
chamber, under illumination of the house-light. After 15-20 s, a 0.5 s foot-shock was
administered at an intensity of 0.05 mA. An experimenter blind to the experimental treatment
scored the following behaviors during each shock delivery: noticing (any noticeable reaction),
flinching (hind-paws briefly raised off the grid floor), vocalizing. The current was increased by
0.05 mA and delivered every 10 s, until the rat vocalized. Rats were then given 2-3 d without
being placed into the operant box. Following this break, rats were given 2 more locomotion
sessions, conducted in a similar manner to their first day. These sessions were aimed at
eliminating any contextual fear that may have occurred during the first foot-shock session. The

142
next day, rats were infused with the counterbalanced treatment, and foot-shock delivery and
scoring was conducted in an identical manner as above.
5.2.9 Histology
After the completion of all test days, rats were euthanized with CO2 and brains were removed
and fixed in a 4% formalin buffered saline solution. Following adequate fixation, brains were
sliced at 50 µm and mounted on glass slides and Nissl stained using cresyl violet acetate. For
animals in the active/passive avoidance experiment, the ventral extent of the infusion bolus is
displayed in Fig. 15A for the NAcC, and Fig. 16A for the NAcS. For animals in the foot-shock
sensitivity experiment, the location of infusion is displayed in Fig. 18A.
5.2.10 Data analysis
For both active and passive avoidance trials, the number of successful avoidances, escapes, and
failures were converted into a percentage of total trials of each type. For each brain region, the
percentage of successful active avoidances, escapes, and failures were analyzed using one-way
ANOVAs, with Treatment (SAL and B/M, or SAL and AMPH) as the within-subjects factor.
The percentage of successful passive avoidances and the number of presses made during a
passive failure were analyzed using separate one-way ANOVAs, again with Treatment as the
within-subjects factor. We also compared the relative importance of the NAcC and NAcS to
active avoidance success by first calculating the change between the saline and B/M conditions,
and then conducting a one-way ANOVA with Treatment (SAL vs. B/M) as the within-subjects
factor.
Locomotor data were converted into a beam-break/min value, and subsequently analyzed
using separate one-way ANOVAs. For manipulations that caused a change in locomotion,

143
Pearson correlations were conducted between the locomotor values and the % passive avoidance
failure, to determine whether locomotion varied systematically with behavior during passive
avoidance trials. The latency to make a response following cue presentation and lever insertion
were collected for all trial types (active avoidance, escape, and passive avoidance). Due to the
nested nature of this data (multiple and variable numbers of responses for different rats), we
choose to analyze this data using a multilevel modelling approach, using the lme4 package in the
statistical program R (Bates, Mächler, Bolker, & Walker, 2014; R Core Team, 2017).
For the foot-shock sensitivity experiment, potential order effects of Treatment were first
examined using a two-way ANOVA on data from animals infused with SAL on the first or
second day. Thus, this between/within-subjects ANOVA had the between subjects factor of Test
Day (Day 1 versus Day 2), and the within-subjects factor of Response Type (Noticing, Flinching,
and Vocalizing). Then, with separate two-way ANOVAs were conducted across cannula
placement. The within-subjects factors were Treatment (SAL and B/M) and Response Type
(Noticing, Flinching, and Vocalizing).
5.3 Results
5.3.1 Task Acquisition and Baseline Performance
During the initial portion of avoidance training, all rats (n = 48) acquired the active avoidance
contingency. All of the following descriptive statistics regarding training are presented as a mean
±SEM. Rats generally completed active avoidance training rapidly, in a mean of 12.96 ±0.91
days, with a range of 4-28 days. On the final day of their active avoidance training, rats
performed 77.4% ±3.3 active avoidance responses, 11.1% ±2.2 escapes, and 11.5% ±2.5 active
avoidance failures. All rats then progressed to the blocked design, during which they received
active avoidance followed by passive avoidance trials, or vice versa. Of the 48 total rats, n = 43

144
successfully reached criterion on this blocked design after an additional 14.4 ±1.2 sessions, over
a range of 4-36 days. On the final day of training on the blocked design, performance remained
good, with rats making 67.4% ±5.6 active avoidance responses, 8.8% ±2.9 escapes, 23.8% ±5.8
active avoidance failures, and 17.4% ±3.5 passive avoidance failures.
Of the 43 total rats that completed the blocked design, 35 rats reached the final task
criterion on the full active/passive avoidance task. Data from 8 rats that did not reach criterion
were not included in the final analysis. These 35 successful rats achieved criterion performance
after an additional 9.9 ±1.0 training sessions (range: 2-24). Upon reaching criterion, these rats
again demonstrated good levels of performance on both active and passive avoidance trials,
making 70.5% ±5.4 active avoidances, 10.6% ±4.1 escapes, and 18.9% ±4.9 active avoidance
failures, and 23.7% ±5.5 passive avoidance failures. A survival plot of training (Fig. 14B)
displays the total number of pre-surgery avoidance training sessions (all training except the
initial locomotion and lever-retract training sessions) required by all rats (Fig. 14B, black line)
and the high performing rats (Fig. 14B, broken gray line) that reached the final task criterion.
5.3.2 Active/passive avoidance inactivation studies
Of the 35 rats that achieved the final task criteria and were implanted with guide cannula, 14
were excluded from the final analysis due to either cannula placements outside of the region of
interest (n = 7), unexpected mortality following surgery (n = 4), or poor performance following
surgery (n = 3). This resulted in final ns of 10 and 11 for the NAcC (Fig. 15A) and NAcS (Fig.
16A) groups, respectively. The mean shock intensity for NAcC group was 0.30 mA (range: 0.25-
0.35 mA), while for the NAcS it was 0.29 mA (rage: 0.25-0.35 mA). These mean intensities
were not significantly different (F(1,19)=0.06,p>0.80).

145
5.3.3 Active/passive avoidance: NAcC inactivation
Inactivation of NAcC markedly impaired performance on active avoidance trials (F(1,9)=39.51,
p<0.001) (Fig. 15B). Subsequent analysis probing this impairment revealed that, although escape
behavior was left intact (F(1,9)=1.55,p<0.24) (Fig. 15B), the incidence of active avoidance
failure was dramatically increased (F(1,9)=47.09,p<0.001) (Fig. 15C). As a result of poorer
performance on active avoidance trials, rats received more foot-shocks following NAcC
inactivation (F(1,9)=44.73,p<0.001) (Table 6). Thus, animals maintained the ability to escape
foot-shock at a comparable level as under control conditions, but their ability to proactively
utilize the active avoidance cue was potently disrupted.
In contrast to the effect on active avoidance trials, passive avoidance behavior was
unaffected by NAcC inactivation, as there was no difference in the percentage of passive
avoidance failures (F(1,9)=0.19,p=0.67) or the number of lever-presses made during passive
avoidance trials (F(1,9)=0.22,p<0.65) (Fig. 15D and E). Inactivation of NAcC had no impact on
the latency to produce an active avoidance (B=0.66, t = 0.95, p>0.34), escape behavior (B = 1.89,
t = 1.03, p > 0.32), or passive avoidance (B=-0.52, t = -0.36, p > 0.72) (Table 6). Furthermore,
locomotor activity was normal following inactivation of the NAcC (F(1,9)=1.00, p>0.34) (Table
6). These data indicate that activity within the NAcC appears to play a selective role in initiating
active defensive responses instigated by cues signaling an impending aversive outcome.
Similarly, successful passive avoidance does not require the NAcC, as the presentation of a
discriminative stimulus associated with lever-press contingent punishment remained effective at
producing response-inhibition. This latter finding also confirms that inactivation of the NAcC
did not impair the ability to discriminate between the two auditory cues.

146
5.3.4 Active/passive avoidance: NAcS inactivation
Inactivation of the NAcS also resulted in poor performance during active avoidance trials
(F(1,10)=24.38,p<0.001) (Fig. 16B). Again, escape behavior was left intact
(F(1,10)=1.70,p>0.22) (Fig. 16B). Thus, impaired avoidance behavior resulted from a selective
increase in active avoidance failures (F(1,10)=24.40,p<0.001) (Fig. 16C). The increase in active
avoidance failure following NAcS inactivation caused animals to receive more foot-shocks
(F(1,10)=26.12,p<0.001) (Table 6). Visual inspection of these data suggested that the decrease in
active avoidance performance following NAcC inactivation may have been quantitatively more
dramatic than the decrement induced by NAcS inactivation. To investigate this, we calculated
the percent change in the number of active avoidance successes during the inactivation test day,
compared to performance following saline infusion. This analysis revealed a numerically greater
decrement in active avoidance performance induced by inactivation of the NAcC (-55.0% ±7.5)
versus the same manipulation of the NAcS (-35.3 ±7.7). However, this difference only
approached statistical significance (F(1,19)=3.31,p=0.085).
In marked contrast to the effects of NAcC inactivation, similar treatments within the
NAcS disinhibited behavior during passive avoidance trials (F(1,10)=12.86,p<0.005), causing
rats to erroneously respond more on the lever during presentation of the passive avoidance cue
(F(1,10)=8.38,p<0.02) (Fig. 16D and E). This manipulation also significantly increased
locomotion during the session (F(1,10)=8.80,p<0.05) (Table 6). However, this increase in
locomotion was not correlated with the disinhibition of passive responding observed following
inactivation of NAcS (r = -0.23, p>0.49) (Fig. 16F), nor with the decrease in active avoidance (r
= -0.29,p>0.39)(data not shown). NAcS inactivation did not affect the latency to respond during
active avoidance trials (B = -0.01, t = -0.03, p > 0.9), escapes (B = -0.24, t = -0.14, p > 0.89), or

147
passive avoidance trials (B = 0.54, t = 0.39, p > 0.70) (Table 6). Thus, this pattern of results
suggests that NAcS promotes active avoidance while also suppressing inappropriate behavioral
activation during passive avoidance.
5.3.5 Active/passive avoidance: Systemic AMPH administration
Unlike inactivation of either accumbens subregion, systemic treatment with AMPH (1 mg/kg)
had no overt impact on active avoidance performance (F(1,19)=0.79,p>0.38) (Fig. 17A),
although animals were significantly quicker to make an active avoidance response (B = -0.83, t =
-2.38, p < 0.02), but not other responses (both p-values > 0.01) (Table 6). This manipulation also
spared escape behavior (F(1,19)=1.00,p>0.32) (Fig. 17A), and thus did not result in a change in
active avoidance failure (F(1,19)=1.51,p>0.23) (Fig. 17B).
On the other hand, AMPH administration produced a selective increase in the percentage
of passive failures (F(1,19)=10.60,p<0.005) (Fig 17C), without altering the overall number of
passive presses made during these failures (F(1,19)=1.10,p>0.30) (Fig. 17D). Thus, although
AMPH disinhibited behavior during passive avoidance trials, rats remained susceptible to
instrumental punishment, making a comparable number of passive presses during failure as
under control conditions. As expected, locomotion was increased following AMPH
administration (F(1,19)=33.79,p<0.001) (Table 6), and interestingly, this locomotor increase
tended to be positively correlated with passive avoidance failure (r = 0.42, p = 0.06) (Fig. 17E)
5.3.6 Foot-shock sensitivity: NAc inactivations
Of the 12 animals allocated to the foot-shock sensitivity experiment, one animal died during
surgery, and one animal had a cannula placement outside of the region of interest, resulting in
final ns of 6 for the NAcC group, and 4 for the NAcS group (Fig. 18A). First, we examined any

148
potential order effect of infusion, to insure that previous experience with a foot-shock test
session did not impact subsequent performance on the second test session. Analysis of animals
infused with SAL on the first test day versus the second suggested that there was no order effect,
as illustrated by no effect of Test Day (F(1,8)=2.15,p>0.18), and no Test Day x Response Type
interaction (F(2,16)=0.87,p>0.43). Thus, data from both days were combined for further within-
subjects analysis.
Inactivation of the NAcC did not affect foot-shock sensitivity (Fig. 18B). There was no
significant effect of Treatment (F(1,5)=0.46,p>0.52), nor a Treatment x Response Type
interaction (F(2,10)=0.46,p>0.64). There was a pronounced main effect of Response Type,
(F(2,10)=32.60,p<0.001), which suggested that the current intensity required to elicit each
response increased across the three behaviors scored, regardless of treatment (all p-values <
0.025) (Fig. 18B). NAcS inactivation also had no effect on foot-shock sensitivity, as there was
no main effect of Treatment (F(1,3)=0.21,p>0.68), and no Treatment x Response Type
interaction (F(2,6)=1.17,p>0.37) (Fig. 18C). Again, a significant main effect of Response Type
(F(2,6)=22.21,p<0.005), was the result of the current intensity requirement increasing across the
three behaviors scored, independent of treatment (all p-values < 0.05) (Fig. 18C). These results
imply that the sensitivity to the aversive stimulus used here was not affected by manipulation of
either NAc subregion.
5.4 Discussion
Although the NAc has long been known to be a key output nucleus in the production of
appetitive behaviors, a bivalent role for this nucleus is relatively understudied. In the present
experiments, a novel behavioral assay was designed to probe the contribution of NAc subregions
and monoamine function to active and passive avoidance. During this task, discriminative stimuli

149
signaled whether the avoidance of an aversive foot-shock could be achieved by either performing
or withholding an instrumental action. Our findings revealed that both the NAcC and NAcS
contribute to successful active avoidance behavior. However, the NAcS also played a role in
suppressing behavior in response to a cue signaling that a passive strategy will avoid
punishment, as inactivation of this nucleus alone disinhbited behavioral responding during
passive avoidance trials. Furthermore, treatment with the monoamine releaser AMPH selectively
enhanced behavioral activation, increasing locomotion as well as passive avoidance failures,
suggesting that the excessive release of dopamine and other monoamines may impede the
suppression of behaviors that lead to aversive outcomes.
5.4.1 Behavioral considerations
Initial training on the active avoidance task produced relatively rapid learning of a lever-press
avoidance response in nearly all animals, as compared to previous reports using similar training
methodology (Berger & Brush, 1975; Fernando et al., 2015; McCullough et al., 1993; Oleson et
al., 2012). Lever-press active avoidance is notoriously difficult to train in rats, particularly when
compared to more simple, naturalistic behaviors such as shuttling (D’Amato & Schiff, 1964;
Meyer, Cho, & Wesemann, 1960). The enhanced rate of learning observed here may be related
to the lower foot-shock current intensity used, as well as the individual titration of current
intensity during learning, factors that differed from most previous reports (D’Amato & Schiff,
1964; McCullough et al., 1993; Meyer et al., 1960; Oleson et al., 2012). As illustrated by the
foot-shock sensitivity experiment, the shock intensities used to motivate avoidance were able to
induce unconditioned responses indicative of discomfort in all rats tested, suggesting that these
intensities were sufficient to act as aversive-motivators. In addition, the novel lever-retraction
session conducted prior to avoidance training served to establish the instrumental response

150
required for an active avoidance, without any explicit reinforcement contingency associated with
a response. This also prevented the need for experimenter-based shaping of behavior oriented
towards the lever (McCullough et al., 1993; Oleson et al., 2012). These procedural variations
may aid in optimizing lever-press active avoidance procedures for use with rodents in the future.
Following acquisition of active avoidance, a passive avoidance component was added,
initially in separate trial blocks, and eventually as randomly presented trials. A protocol
consisting of active avoidance training preceding passive avoidance (and not vice versa) was
chosen because success on passive trials is indicated by response suppression, for which there
needs to be an established active behavior to inhibit. Although a majority of rats were able to
acquire the blocked design, a fair amount of experimental attrition was observed during training
on the full active/passive avoidance task, where active and passive avoidance trials were
intermixed randomly over a session. This attrition stemmed partially from an a priori inclusion
criterion for rats to perform well on both trial types, to allow for conclusions to be drawn about
the effect of pharmacological manipulation. A small number of rats minimized active avoidance
failure by predominantly performing escape responses, a mediating strategy that allowed rats to
potentially discern between trial types (if a shock is received following a tone, but prior to any
instrumental response, then that trial is an active avoidance). Escape behavior can also represent
an intermediate step in active avoidance learning, which may suggest inadequate acquisition of
the active avoidance response (Solomon & Wynne, 1953). Other rats simply were unable to
maintain active avoidance performance once the discrimination component was introduced,
likely developing a learned helplessness-like phenotype due to the receipt of foot-shock without
the production of an avoidance response for an extended period of training (Seligman &
Beagley, 1975). Although we were not able to include these animals due to our performance

151
criteria, future studies may utilize these poor performing animals in order to provide insight into
mechanisms that oppose avoidance, including the pervasive expression of conditioned fear
(Martinez et al., 2013).
The full version of the active/passive avoidance task was designed to provide insight into
aversively-motivated flexible behavior. In the appetitive (or mixed valence) domain, assays such
as the Go/No-Go task probe the ability of animals to utilize cues that necessitate opposing
behaviors on a flexible basis. Previous research suggests that NAc activity is modulated by the
presentation of Go and No-Go cues (Roitman & Loriaux, 2014; Setlow et al., 2003). This
activity is strongly related to the action necessitated by cue presentation, consistent with a role
for this nucleus in action selection (Roitman & Loriaux, 2014). The results of the present study
are broadly consistent with a parallel role for this nucleus in aversively-motivated flexible
behavior. Perhaps more intriguingly, we observed a dissociation between the impact of NAcC
and NAcS inactivation that may be related to similar mechanisms underlying appetitive
reinforcement-seeking, including a particular role for the NAcS in response-inhibition, and a
dual role for these structures in active approach (Ambroggi et al., 2011, 2008; Floresco, 2015;
Ghazizadeh et al., 2012; Peters et al., 2008).
5.4.2 Regulation of active behaviors by NAcC
Inactivation of the NAcC profoundly impaired the expression of active avoidance, without
affecting passive avoidance performance. The involvement of the NAcC in the production of
active avoidance is in keeping with a number of previous neurochemical studies. During active
avoidance learning, dopamine release within the NAc is positively correlated with successful
performance (Dombrowski et al., 2013; McCullough et al., 1993). Similarly, NAc dopamine
release occurs during the presentation of the active avoidance cue on successful avoidance trials

152
during established active avoidance expression (Gentry et al., 2016; Oleson et al., 2012).
Importantly, these later studies targeted their voltammetric assessment of dopamine release to the
NAcC, providing confirmatory evidence that transmitter release in this subregion mediates
avoidance.
Although neuromodulatory activity within the NAcC is relevant to active avoidance
learning and performance, few studies have investigated how altering neural activity in this
subnucleus may affect such behavior. To our knowledge, only one previous study has separately
examined NAcS and NAcC function during the expression of well-trained active avoidance
(Ramirez et al., 2015). In that study, rats learned a simple two-way active avoidance response
over the course of four days, which was not affected by subsequent NAcC inactivation on day
five (Ramirez et al., 2015). Two main factors distinguish the present study from the one
conducted by Ramirez and colleagues (2015). First, it may be that the auditory stimulus
disambiguation required here recruits brain regions that are not necessary for the simple, single-
stimulus active avoidance behavior. Consistent with this, a previous study has shown that
coherence between auditory cortex and the lateral ventral striatum, which may include the
NAcC, increases when learning about a stimulus that necessitates an active avoidance response,
but not when associating a different stimulus with a passive avoidance response (Schulz et al.,
2015). This finding was interpreted to suggest that plasticity within the auditory cortex and
ventral striatum allows for stimulus discrimination and appropriate behavioral output. Thus, it is
possible that such a mechanism continues to be necessary for the normal expression of active
avoidance, particularly in situations requiring stimulus discrimination. Secondly, the
instrumental response required here (lever-press) is a relatively more complex action to produce
than is shuttling (Bolles, 1970; D’Amato & Schiff, 1964). Given that neurotransmission in the

153
NAcC is critical for re-engagement during bouts of lever-pressing for reward (McGinty et al.,
2013; Nicola, 2010), inactivation of this nucleus could render rats unable to efficiently locate and
engage the lever, although gross locomotor activity may remain intact. Such an effect may
diminish lever-press avoidance, while sparing the more naturalistic (and less localized) shuttling
response required by Ramirez et al. (2015). However, Bravo-Rivera, Quirk and colleagues
(2014) examined NAc function on a platform-based avoidance task where animals had the
concurrent opportunity to lever-press for sucrose reward. Using infusions that primarily targeted
the NAcC, these researchers demonstrated that inactivation impaired avoidance and
concomitantly increased freezing during the avoidance stimulus (Bravo-Rivera et al., 2014).
Thus, the NAcC may also promote avoidance by suppressing Pavlovian defensive reactions such
as freezing.
Taken together, these findings raise the possibility that active behaviors instigated by
Pavlovian or instrumental mechanisms may require the NAcC, regardless of whether the
behavior is aversively or appetitively motivated. Neurotransmission in the NAcC has previously
been shown to control flexible approach towards Pavlovian or instrumental stimuli conditioned
via appetitive reinforcement (McGinty & Grace, 2008; Nicola, 2010; Saunders & Robinson,
2012). For example, NAcC activity is necessary for the acquisition of discriminative Pavlovian
conditioned approach, where rats learn to approach a CS+ that signals reward delivery, but not a
CS- that signals no reward (Di Ciano et al., 2008; Parkinson et al., 1999; Parkinson, Willoughby,
et al., 2000; Saunders & Robinson, 2012). NAcC activity is also required for the ability of a
Pavlovian stimulus to drive appetitively-motivated instrumental behavior (Ambroggi et al., 2011,
2008; Hall, Parkinson, Connor, Dickinson, & Everitt, 2001). Lesions of the NAcC disrupt the
general form of Pavlovian-to-instrumental transfer, where a previously learned appetitive CS+

154
potentiates the vigor with which a novel instrumental response is acquired (Hall et al., 2001).
Similarly, instrumental responding during a discriminative stimulus that signals reward
availability is diminished by NAcC inactivation (Ambroggi et al., 2011, 2008). These previous
findings serve to illustrate that actions motivated by appetitive reinforcement require NAcC
activity. In the present study, the ability of a negatively-reinforced auditory stimulus to elicit
approach and engagement with a lever to actively avoid foot-shock was impaired following
NAcC inactivation. Importantly, this effect was specific to anticipatory behavior, as the number
of escapes, which are motivated by the US directly, remained unchanged following NAcC
inactivation. This result suggests that the NAcC promotes approach behavior mediated by
aversive motivation in a similar manner to this regions role in appetitive motivation.
It is important to note that the effect of NAcC inactivation on active avoidance was not
the result of psychomotor slowing, which could manifest as poor active avoidance performance.
This consideration is particularly relevant given that inactivation of the NAcC often slows
response latencies and decreases locomotor activity during cognitive performance (Ambroggi et
al., 2011; Dalton et al., 2014; Feja et al., 2014; Floresco et al., 2008; Ghods-Sharifi & Floresco,
2010; Stopper & Floresco, 2011). Following NAcC inactivation during this active/passive
avoidance task, response latencies and locomotion were comparable to control data. This
discrepancy illustrates a potential divergence between the mechanisms underlying appetitively
and aversively-motivated behaviors. Such a suggestion has implications for our understanding of
the aforementioned flexible approach hypothesis (McGinty et al., 2013; Nicola, 2010), which
emphasizes that neurons within in this nucleus facilitate locomotor approach and engagement
during reinforcement-seeking, in a dopamine-dependent manner. Thus, although there are
notable similarities between the behavioral ramifications of NAcC inactivation on approach

155
behavior mediated by reinforcement and punishment, underlying processes such as motor
activation and reaction time may be differentially mediated.
That NAcC inactivation spared passive avoidance behavior fits with hypotheses
suggesting that NAcS, but not NAcC, is uniquely responsible for inhibiting inappropriate
behavioral responses (Ambroggi et al., 2011; Blaiss & Janak, 2009; Floresco, 2015). Previous
studies have shown that parameters of response inhibition that are affected by NAcS inactivation,
such as instrumental responding on an inactive lever or during an explicitly non-rewarded period,
are unchanged by NAcC inactivation (Ambroggi et al., 2011, 2008; Blaiss & Janak, 2009;
Floresco et al., 2008). Here, we operationalize response-inhibition as the ability to withhold a
lever press during the presentation of a cue predicting instrumentally-delivered punishment. We
have previously shown that the withholding of a sucrose-seeking response by the presentation of
a Pavlovian aversive cue or by instrumental punishment is intact following NAcC inactivation
(Chapter 2, 3, and 4; Piantadosi et al., 2017). Other tasks assessing impulsivity, a multifaceted
construct that reflects the inability to withhold an action due to motor or cognitive dysfunction,
have produced inconsistent results regarding the requirement of NAc subregions. Some studies
have suggested that the NAcC promotes response-inhibition (Cardinal, Pennicott, Sugathapala,
Robbins, & Everitt, 2001b; Christakou, Robbins, & Everitt, 2004; Pothuizen et al., 2005), while
others implicate the NAcS (Feja et al., 2014), or neither structure (Eagle & Robbins, 2003;
Murphy et al., 2008). Of the studies supporting a role for the NAcC in impulsive responding, the
majority utilized permanent lesions that may severely impact underlying processes, such as the
appropriate timing of responding (Singh et al., 2011). In addition, assays of impulsivity
commonly operationalize punishment as a loss of opportunity for (more) reward, while the
present study enforces response-inhibition via the delivery of foot-shock. Thus, the involvement

156
of the NAcC in response-inhibition may be situationally dependent on the method used to
provoke response-inhibition. Future studies are necessary to determine the extent to which
passive avoidance taxes NAcC-mediated mechanisms similar to those regulating the multifaceted
processes underlying impulsivity.
5.4.3 Dual functions of the NAcS in active and passive behavior
Here, we hypothesized that NAcS would contribute to the performance of both active and
passive avoidance. Consistent with this, inactivation of NAcS produced a qualitatively similar
impairment during active avoidance trials as did NAcC inactivation. One potential explanation
for the comparable impact of inactivation of either subnuclei was the spread of the infusate from
within the NAcS to the NAcC. We have previously demonstrated dissociable roles for these
subregions using similar infusion procedures to great effect (Dalton et al., 2014; Floresco,
Ghods-Sharifi, Vexelman, & Magyar, 2006; Floresco et al., 2008; Piantadosi et al., 2017;
Stopper & Floresco, 2011), and, in the present study, dissociations between each subregion were
observed during passive avoidance trials, as well as locomotor activity. This suggests that
spillover of the infusate from NAcS to NAcC cannot fully account for the similar effect on active
avoidance. A more likely explanation comes from previous work suggesting that NAcS
independently promotes active avoidance, via interactions with the BLA (Ramirez et al., 2015).
Our data extend this observation beyond the realm of two-way active avoidance, illustrating that
NAcS promotes active responding in an operant environment. As a subnucleus within the limbic-
motor interface (Mogenson et al., 1980), the NAcS is positioned to integrate affective
information regarding the avoidance stimulus arriving from BLA, and translate this information
into defensive action, in this case active avoidance (Martinez et al., 2013; Ramirez et al., 2015).
NAcS projects to downstream targets within the ventral pallidum and midbrain dopamine

157
system, pathways which may act to promote the appropriate expression of active avoidance
(Ilango et al., 2014; Ilango, Shumake, Wetzel, Scheich, & Ohl, 2012; Saga et al., 2017). Another
related possible mechanism contributing to the promotion of avoidance by the NAcS is the
invigoration of responding by a safety signal during successful avoidances. Safety signals
reinforce avoidance behavior by explicitly indicating that the instrumental response has been
successful, potentially coming to act as a conditioned reinforcer (Dinsmoor & Sears, 1973;
Fernando et al., 2013, 2014; Morris, 1975). Inactivation or infusion of AMPH into the NAcS (but
not NAcC) has been shown to decrease operant active avoidance only during sessions where a
safety signal was present (Fernando et al., 2013). Suppressing NAcS neural activity via
inactivation may have reduced the motivational impact that the safety signal has on behavior,
causing a decrement in active avoidance responding.
The key dissociation observed in the present study was that activity within the NAcS, but
not the NAcC, is necessary for the appropriate inhibition of punished responding during passive
avoidance trials. Although motor activity was also disinhibited following NAcS inactivation,
hyperlocomotion alone cannot explain the resulting deficit in passive avoidance, as there was no
correlation between these two measures. This effect is also unlikely to be explained by a general
mnemonic impairment, as performance on tasks assessing passive avoidance expression
independently are not affected by NAc manipulation (De Leonibus et al., 2003; Lorenzini et al.,
1995). Thus, this result is likely related to the demands of the task, requiring rats to balance
active versus passive behaviors on a dynamic basis, akin to classic Go/No-Go paradigms.
Relatedly, we have recently shown that this particular accumbens subregion (as well as the
basolateral amygdala) is required when animals inhibit reward-seeking under threat of
punishment on a “conflict” task (Chapter 4; Piantadosi et al., 2017). When seeking sucrose

158
during the conflict period, where the reinforcement schedule is rich but concurrently punished,
inactivation of NAcS and BLA, disinhibited lever-pressing despite punishment. The passive
avoidance trials employed here are similar to the punished period on the conflict task, during
which control animals typically make few inhibitory control errors. Removing the influence of
the NAcS eliminates the break on punished responding, causing rats to make passive avoidance
errors. Inactivated animals also became less susceptible to instrumental punishment delivery, as
they made more passive presses during these errors. This later finding indicates that the
impairment in passive avoidance extends beyond an inability to properly respond to predictive
conditioned stimuli, and includes a loss of instrumentally administered aversive-motivation
following the punishing foot-shock itself. The results of our foot-shock control experiment
suggest that this disinhibition of pressing is unlikely to be caused by changes in foot-shock
sensitivity per se, as unconditioned responses were normal following NAcS inactivation. Instead,
the link between foot-shock receipt and the implementation of suppression may be diminished in
the absence of NAcS activity.
The mechanism through which the NAcS regulates punishment-induced response-
inhibition may also relate to the ability of this subregion to refine behavior by encoding the
disadvantageous nature or irrelevance of stimuli and actions (Ambroggi et al., 2011; Blaiss &
Janak, 2009; Floresco et al., 2008; Gal, Schiller, & Weiner, 2005; Millan et al., 2010; Peters et
al., 2008; Pothuizen et al., 2005). In the appetitive domain, inactivation or blockade of dopamine
function within the NAcS releases inappropriate behavior from inhibition, such as operant
responding during task periods that are explicitly not reinforced (Ambroggi et al., 2011; Blaiss &
Janak, 2009; Ghazizadeh et al., 2012). Activity within the NAcS is often necessary for the
inhibition of behavior following extinction learning, as disrupting transmission within this region

159
disinhibits extinguished behavior (Blaiss & Janak, 2009; Floresco et al., 2008; Peters et al.,
2008). Lesions of this subnucleus also prevent the acquisition of aversively-motivated learned
irrelevance, which occurs when numerous non-reinforced presentations of a stimulus retard the
subsequent association of that stimulus with punishment (Gal et al., 2005; Pothuizen et al.,
2005). These results support the contention that the NAcS promotes response-inhibition under
some circumstances, independent of the valence of the motivator.
As discussed previously, the literature implicating the NAc in impulsivity is mixed,
perhaps due in part to a lack of attention to subregional distinctions and an overreliance on
permanent lesions (Basar et al., 2010). One recent study has utilized reversible inactivations to
demonstrate that NAcS activity opposes impulsive actions and choices, while NAcC is more
necessary for general aspects of motivated behavior (Feja et al., 2014). The NAcS may facilitate
response inhibition in concert with dopaminergic input from the midbrain, as blocking dopamine
D2 receptors within the NAcS (but not NAcC) exacerbates the performance of impulsive actions
in highly impulsive rats (Besson et al., 2009). The present data suggest that the ability to inhibit
lever pressing during the presentation of a passive avoidance stimulus dependent in part on
NAcS function, which may relate to a generalizable role for this nucleus in response-inhibition.
5.4.4 Monoaminergic correlates of response promotion and inhibition
We also probed the ability of the monoamine releaser AMPH to alter the expression of well-
learned active/passive avoidance, illustrating that AMPH selectively affected performance on
passive avoidance trials. It is well-established that AMPH administration can potentiate the
acquisition of active avoidance performance (Barrett, Leith, & Ray, 1972; Kulkarni, 1968;
Niemegeers, Verbruggen, & Janssen, 1970). This effect has been suggested to be the result of an
increase in general motoric output, allowing animals to overcome the behavioral suppression

160
induced by fear early in active avoidance training (Kulkarni, 1968). Thus, it is perhaps
unsurprising that AMPH had no impact on performance in the well-trained animals tested here,
as they have already overcome this obstacle to successful active avoidance. Consistent with this,
AMPH administration has been shown to be ineffective at altering active avoidance performance
in animals performing at asymptote (Rosen & La Flore, 1973). Still, dopaminergic signals in the
NAc persist during performance of a similar approach/avoidance task requiring animals to attend
to discrete cues that necessitate one of three instrumental responses, an active avoidance
response, a sucrose-seeking response, or no action at all (Gentry et al., 2016). Taken together,
these results suggest that, although a baseline level of dopamine activity within the NAc is likely
necessary for active avoidance, potentiating this signal via AMPH does not impact performance.
In contrast to the null effect on active avoidance, AMPH administration impaired
performance on passive avoidance trials. The trend-level correlation observed here between
hyperlocomotion and passive avoidance failure suggests that motor disinhibition may contribute
to this effect. Broadly, this finding is consistent with previous studies suggesting that AMPH
administration can provoke impulsive actions (for review, see Robbins, 2002). For example, at
doses similar to those used here, AMPH has been shown to produce premature responses on the
five-choice serial reaction time task, which may reflect a loss of inhibitory control over prepotent
actions (Baarendse & Vanderschuren, 2012; Cole & Robbins, 1989, 1987; Harrison, Everitt, &
Robbins, 1997; Murphy et al., 2008; Pattij, Janssen, Vanderschuren, Schoffelmeer, & Van
Gaalen, 2007; Wiskerke et al., 2011). In addition, AMPH administration selectively impairs
performance on No-Go trials, without affecting Go responses, in an appetitively-motivated
Go/No-Go paradigm (Blackburn & Hevenor, 1996). Interestingly, the loss of inhibitory control
induced by AMPH on assays of impulsive action has been shown to be related to dopamine and

161
µ-opiod receptor activation within the NAcS (Pattij et al., 2007; Wiskerke et al., 2011). These
findings suggest that AMPH may act directly or indirectly within the NAcS to alter response
inhibition, in keeping with the similarity between the two treatments shown here. However,
unlike NAcS inactivation, rats treated with AMPH were able to inhibit their passive responding
upon receipt of the foot-shock, making the same amount of passive presses as under control
conditions. Notably, this finding is unlikely to be due to alterations in foot-shock sensitivity, as
previous studies suggest that unconditioned responses to aversive stimuli are unchanged by
AMPH administration at the doses given here (Conti, Maeiver, Ferkany, & Abreu, 1990;
Mitchell et al., 2011; Orsini, Trotta, et al., 2015; Simon et al., 2009). This discrepancy between
the impact of AMPH and NAcS inactivation suggests some degree of dissociation between the
mechanisms mediating cue-induced suppression, and the suppression induced by receipt of a
punishing unconditioned stimulus.
The suggestion that AMPH produces deficits in response-inhibition during punished trials
may appear to conflict with data demonstrating that AMPH decreases the willingness of animals
to accept punishment during reinforcement-seeking (Geller & Seifter, 1960; Lazareno, 1979;
Mitchell et al., 2011; Orsini, Trotta, et al., 2015; Simon et al., 2009). During these tasks, animals
are more reticent to produce an instrumental reward-seeking response during punishment, an
effect that is potentiated by AMPH administration. One major difference between these studies
and the present work is the presence or absence of response-competition related to the goal of the
instrumental response. In the case of the experiments described here, there is no response-
competition induced by the instrumental response itself. In comparison, during these other
punishment tasks, there is a prominent response-conflict component, as rats are highly motivated
to seek reward, yet simultaneously want to limit exposure to potential harm. This distinction

162
suggests that AMPH may bias behavior towards or away from punishment, as a function of the
presence or absence of response-competition, respectively.
Although the behavioral effects of AMPH are most commonly attributed to its
modulation of dopamine function via blockade of the dopamine transporter, this stimulant also
elevates other monoamines, including serotonin (Kuczenski & Segal, 1989; Seiden, Sabol, &
Ricaurte, 1993; Sitte & Freissmuth, 2015). Evidence supporting a dopamine-mediated account of
the effects of AMPH comes from previous studies on impulsive action, where the effect of
AMPH is dramatically reduced by intra-NAc dopamine lesions (Cole & Robbins, 1989).
Similarly, blockade of dopamine D2/3 receptors in the NAc blocks the impulsigenic impact of
systemic AMPH administration (Pattij et al., 2007). In contrast, depletion of brain serotonin
induces impulsive action and prevents the ability of AMPH to potentiate this aberrant behavior
(Harrison et al., 1997). Taken together, these results suggest that the AMPH-induced passive
avoidance impairment seen here may be mediated by dopamine release, possibly within the NAc.
Still, the relation between impulsivity and passive avoidance as operationalized here remains to
be established, and more work is necessary to probe the particular brain region and transmitters
mediating this effect.
5.4.5 NAc circuitry regulating active/passive avoidance: Relevance for humans
Aberrant avoidance behavior is present across a number of neuropsychiatric conditions (Dymond
& Roche, 2009; Figee et al., 2016; Maner & Schmidt, 2006; Ottenbreit & Dobson, 2004; Trew,
2011). Therefore, identifying the neural substrates underlying the basic behavior may improve
our understanding of disorders characterized by such disturbances. To this end, activity within
the human ventral striatum, which contains the NAc, has been associated with active avoidance
performance (Delgado et al., 2009; Jensen et al., 2003; Levita et al., 2012). Bilateral ventral

163
striatal activation has been reported during the presentation of an avoidance cue, as compared to
a neutral cue. These results indicate that neurons within the ventral striatum are relevant while
learning the association between a particular instrumental action and its avoidance outcome
(Delgado et al., 2009), as well as while performing an active avoidance response (Jensen et al.,
2003; Levita et al., 2012). Although neither study had the spatial resolution necessary to probe
specific contributions of ventral striatal subregions to this behavior, they are broadly consistent
with the present finding suggesting that this region is necessary for instrumental active
avoidance.
Comparatively less attention has been paid to the circuitry underlying passive avoidance
in humans. Here we based our task design on one of the few studies to have used functional
imaging techniques to probe both poles of avoidance behavior using a button-press
active/passive avoidance task (Levita et al., 2012). These researchers illustrated that deactivation
within the NAc is observed during passive avoidance trials, while the aforementioned active
avoidance trials produced activations within this nucleus. In keeping with the clinical relevance
of such data, this pattern of activation/deactivation was correlated with a measure of state
anxiety. Given that our manipulation involved temporarily decreasing neuronal activity within
the NAc, one might have expected an improvement in passive avoidance performance based on
this limited human literature. In fact, we observed the opposite pattern of results, when
manipulating the NAcS specifically. One potential explanation for this discrepancy arises from
research illustrating that deactivations as measured by fMRI may not necessarily correspond to
decreases in neuronal activity, particularly when measured within striatal regions (Hayes &
Huxtable, 2012; Mishra et al., 2011). Specifically, comparisons between functional imaging and
electrophysiological indicators of neuronal activity suggest that concordance between these

164
measures is low when measured in the striatum, but high when measured in cortical and thalamic
regions. These researchers hypothesized that deactivations may instead reflect alterations in
neurovascular coupling, rather than changes in neuronal activity. If this is the case, the
deactivation observed in the NAc by Levita et al. (2012) may not necessarily imply that
diminished activity in this nucleus precedes successful passive avoidance. Additionally, Levita
and colleagues (2012) assessed BOLD activity within the entire window around active/passive
avoidance cue presentation and behavior, which likely includes the outcome phase. Thus, some
of the activity observed in that study may reflect relief or safety processing, functions which may
require the NAc (Baliki et al., 2013; Mohammadi et al., 2014). Another level of ambiguity is
added by the fact that most fMRI research is not capable of dissociating between subregions of
the NAc, which we have shown here to be differentially responsible for aspects of avoidance
behavior. To date, the only functional imaging study to have dissociated NAcS and NAcC in the
human brain has suggested that activity within the NAcS occurs in anticipation of thermal pain,
while NAcC activity occurs following the cessation of a painful stimulus, supporting a role for
these subnuclei in aversion (Baliki et al., 2013). Thus, future studies examining active/passive
avoidance performance should account for possible functional differences within accumbens
subregions.
5.5 Conclusion
Using a novel active/passive avoidance task, we illustrate that the two main subregions of the
rodent NAc, the NAcS and NAcC, differentially regulate aspects of this behavior. Specifically,
both subregions promoted the expression of active avoidance, while only the NAcS contributed
to response-suppression during passive avoidance. Administration of the monoamine releaser
AMPH also selectively impaired passive avoidance responses. These results are in keeping with

165
previous research differentially implicating these subregions in avoidance, as well as processes
that contribute to active approach and impulsive action.

166
Table 6. Mean (± SEM) values for ancillary measures during the active/passive avoidance task.
Overall locomotion, the number of shocks received (during active avoidance failure), and the
response latency for active avoidances, escapes, and passive avoidances following regional
inactivation or AMPH treatment. * = p < 0.05 vs SAL
Locomotion
(photobeam
breaks/min)
Shocks
received
(active
avoidance)
Active
avoidance
response
latency (s)
Escape
response
latency (s)
Passive
avoidance
response
latency (s)
NAcC
SAL 24.4 (±3.1) 7.2 (±1.2) 5.5 (±0.7) 2.5 (±0.6) 6.7 (±1.4)
B/M 21.3 (±2.9) 23.8 (±1.7)* 6.3 (±1.0) 4.4 (±1.2) 6.2 (±0.9)
NAcS
SAL 24.3 (±3.2) 4.3 (±0.9) 6.1 (±0.8) 3.5 (±1.3) 5.3 (±1.2)
B/M 51.6 (±8.1)* 14.4 (±2.2)* 5.7 (±0.8) 4.4 (±1.1) 6.2 (±1.1)
AMPH
SAL 23.9 (±1.7) 3.7 (±0.7) 5.5 (±0.4) 2.9 (±0.5) 6.4 (±0.9)
1 mg/kg 58.0 (±6.2)* 5.4 (±1.8) 4.7 (±0.3)* 1.9 (±0.2) 5.7 (±0.6)

167
Figure 14. Trial structure and survival plot of training for the active/passive avoidance
task. (A) Layout of a single trial on the active/passive avoidance task. Each trial type and potential outcome are
outlined from the branches following Trial Start. LP = lever press, gray outlined lightning bolt = foot-
shock delivery. (B) A survival plot showing all rats that reached the full active/passive avoidance task
(black line), and a subset of rats that reached the criterion on the final version of the full active/passive
avoidance task (broken gray line).

168
Figure 15. NAcC activity is necessary for active, but not passive, avoidance performance. (A) Histology diagram, the ventral extent of each microinfusion within the NAcC is labeled by a filled
triangle. (B) NAcC inactivation decreased the percentage of successful active avoidance trials, without
affecting escape responses. (C) Inactivation induced more failures during active avoidance trials. (D, E)
The percentage of passive avoidance failures and the number of passive presses did not change following
NAcC inactivation. Star denotes p<0.05 between the SAL and B/M conditions.
Figure 16. NAcS activity is necessary for active and passive avoidance performance. (A) Histology diagram, the ventral extent of each microinfusion within the NAcS is indicated by a filled
circle. (B) Inactivation of the NAcS decreased active avoidance success, but left escape behavior intact.
(C) The percentage of active avoidance trials ending in failure was increased by NAcS inactivation. (D)
NAcS inactivation induced passive avoidance failures, and (E) increased the total number of presses made
during passive failure. (F) Scatterplot comparing locomotion (beam breaks/min) against the number of
passive avoidance failures in the NAcS inactivation condition. There was no significant relationship
between these measures. Star denotes p<0.05 between the SAL and B/M conditions.

169
Figure 17. AMPH administration selectively provokes passive avoidance failure. (A) Performance of active avoidance and escape behavior was normal following AMPH (1 mg/kg)
administration. (B) AMPH treatment had no effect on active avoidance failure. (C) AMPH increased the
percentage of passive avoidance failures, without altering (D) the total number of presses made during
passive avoidance failure. (E) Scatterplot comparing locomotion (beam breaks/min) against the number of
passive avoidance failures in the AMPH condition. There was a trend towards a positive correlation
between these two measures. Star denotes p < 0.05 between the SAL and AMPH conditions.

170
Figure 18. Neither NAc subregion is necessary for foot-shock sensitivity. (A) Histology diagram, the ventral extent of each microinfusion within the NAcC (closed triangles) and
NAcS (gray circles) are indicated. (B, C) NAcS inactivation (B) or NAcC inactivation (C) had no effect
on the current threshold required for animals to notice, flinch, or vocalize following foot-shock delivery.
Star denotes simple main effects comparisons at a p < 0.05 level, between each measure of foot-shock
sensitivity (regardless of treatment).

171
Chapter 6: General discussion
The present experiments examined the function of two major NAc subregions, the NAcC and
NAcS, in a variety of related aversively-motivated behaviors. Consistent across these studies was
the necessity to inhibit responding during discrete task epochs, while promoting active behavior
during others. Response-inhibition was motivated either by purely Pavlovian mechanisms, as in
Chapters 2 and 3, or by the potential for instrumental punishment, as in Chapters 4 and 5.
Regardless of the conditioning mechanism, the NAcS was necessary for animals to suppress
responding, as inactivation of this structure disinhibited lever-pressing during threat. In the case
of Pavlovian fear, this effect appeared to be mediated in part by a projection from the PL cortex
to the NAcS. In the next two experiments, NAcS inactivation diminished the impact of
instrumental punishment on reward-seeking and passive avoidance. Critically, neither
accumbens subregion was necessary for normal unconditioned responding to foot-shock alone,
suggesting that these results cannot be explained by alterations in pain sensitivity.
Unlike the NAcS, the NAcC was not responsible for aversive motivation, as inactivation
of this structure instead affected indices of behavioral activation, such as locomotor activity and
response vigor. During the Pavlovian fear task, this region played no role in fear acquisition or
expression. Instead, the NAcC appeared to promote locomotor activity, as well as the vigor with
which animals pressed the operant lever. A similar effect was observed in Chapter 4, with NAcC
inactivation reducing operant reward-seeking, concomitant with a decrease in locomotion,
regardless of whether rats were trained on a task delivering instrumental punishment or not. We
then illustrated that the promotion of responding mediated by the NAcC was not exclusive to the
appetitive context, as active avoidance performance, which required an instrumental response to

172
avoid foot-shock, was powerfully impaired by inactivation. Thus, these results suggest a
fundamental role for the NAcC in the invigoration of behavior.
In addition to the interrogation of these ventral striatal subregions, we probed the
necessity of relevant cortico-limbic regions to aspects of these aversively-motivated behaviors.
In the case of the prefrontal cortex, we observed that the top-down control of Pavlovian
conditioned suppression expression was mediated by the PL cortex, and to a lesser extent the IL
cortex. Pharmacological disconnection illustrated that a direct projection from the PL cortex to
the NAcS was responsible for the former effect. We also evaluated the possibility that a
BLANAcS circuit was necessary for fear acquisition, although a disconnection experiment
demonstrated that this projection was not involved. Although glutamatergic projections from the
BLA to the NAcS did not mediate Pavlovian fear acquisition, intact activity in the BLA was
necessary for the inhibition of reward-seeking during punishment. Finally, the promotion of
catecholaminergic activity was examined for its effect on Conflict performance, as well as
active/passive avoidance, given that both tasks necessitated response-inhibition. Interestingly,
AMPH selectively and bidirectionally affected indices of response-inhibition on both tasks,
promoting suppression during instrumental punishment, but disinhibiting instrumental actions
during punished passive avoidance trials.
6.1 Dissociable contributions of NAc subregions to the inhibition and promotion of
behavior
Here, we designed a series of experiments to probe the hypothesized involvement of the NAcC
and NAcS in aversively-motivated behavior. The dissociability of these regions has primarily
been examined in the appetitive domain, using behavioral tasks that assess Pavlovian and
instrumental mechanisms contributing to action selection. A particularly instructive illustration
of the functional differences (and similarities) between these two regions comes from

173
electrophysiological and pharmacological experiments during the performance of a simple
behavioral assay. During this task, animals learn that the presentation of a discriminative
stimulus indicates that reward is available for a press on an operant lever, while presses during
other epochs, including the presentation of a neutral stimulus, are never reinforced (Ambroggi et
al., 2011, 2008; Ghazizadeh et al., 2012; Ishikawa et al., 2008, 2010; Nicola et al., 2004; Yun et
al., 2004). Neurons within the NAcS encode task events that acquire irrelevance over the course
of training, such as the presentation of a neutral stimulus or pressing a never-reinforced inactive
lever (Ghazizadeh et al., 2012). Learning to inhibit these irrelevant responses recruits the NAcS,
via a projection from the vmPFC, which promotes the activity of tonically active NAcS neurons
that inhibit behavior during non-reinforced task phases (Ghazizadeh et al., 2012).
During performance of this task, the same pattern of results holds true. While NAcS
activity preferentially tracks task-irrelevant events, the NAcC is more likely to be activated by
the rewarded discriminative stimulus (Ambroggi et al., 2011). Inactivation of the NAcS has a
disinhibitory effect on reward-seeking actions during irrelevant task phases (Ambroggi et al.,
2011). In contrast, NAcC inactivation decreases behavior during presentation of the
discriminative stimulus indicating reward availability (Ambroggi et al., 2011, 2008; Ishikawa et
al., 2008). Further evidence for such a dissociation comes from studies illustrating that the NAcS
inhibits the reinstatement of reward-seeking for a variety of substances, while the NAcC
typically promotes such behavior (Di Ciano et al., 2008; Floresco et al., 2008; Millan et al.,
2010; Peters et al., 2008).
Using three separate, but related aversive conditioning paradigms, we observed that the
NAcS subserved a response-inhibitory function, while the NAcC simply promoted actions. The
aversive Pavlovian or instrumental inhibition of reward-seeking was less pronounced when

174
NAcS was taken silenced using reversible inactivations. This was true regardless of whether the
behavior being suppressed was motivated by appetitive reinforcement or by negative
reinforcement. These results suggest a consistent role for the NAcS across these distinct
conditioning paradigms, in keeping with the conceptualization of this region as a limbic-motor
integrator (Mogenson et al., 1980). Such a hypothesis is not incompatible with evidence
suggesting that the NAcS is also able to promote actions that enable avoidance or escape of
danger, as also illustrated here (Fernando et al., 2014; Ramirez et al., 2015). The NAcS may be
recruited to suppress activity in situations where escape or avoidance are not possible, while also
facilitating actions to ensure safety when such opportunities are available. The BLA, which
integrates valence signals to allow for appropriate behavior, likely contributes to such action (or
inaction) selection, as this region is critical for passive defensive responses, as well as active
defensive actions (for which a direct projection to NAcS has been demonstrated) (Correia et al.,
2016; Jean-Richard-Dit-Bressel & McNally, 2015; Ramirez et al., 2015; Sierra-Mercado et al.,
2011).
This later facet of active behavior motivated by aversive consequences also required the
NAcC. This is in keeping with a variety of research from the appetitive conditioning literature
suggesting that the NAcC motivates active behaviors. As discussed, neurons within the NAcC
encode stimuli that signal reward availability, and inactivation of this structure decreases
instrumental reward-seeking behaviors (Ambroggi et al., 2011). In addition, this subnucleus
plays an important role in enacting the behaviorally activating effects of conditioned stimuli. For
example, blocking dopamine activity within the NAcC diminishes the expression of a
conditioned approach response mediated by a cue that predicts reward delivery, without altering
behavior in animals that do not attribute incentive salience to the cue (Saunders & Robinson,

175
2012). A formal conceptualization of this function, known as the flexible approach hypothesis,
suggests that activity (particularly dopamine release) within the NAcC allows animals to
appropriately engage and re-engage with instrumental manipulanda in the environment (McGinty
et al., 2013; Nicola, 2010). Although we did not assess dopaminergic activity within the NAcC,
inactivation of this subregion typically impaired the vigor with which animals engaged in a
particular behavior, regardless of task context. By decreasing neural activity in this region, we
may have provoked a similar state to that induced by hypo-dopaminergia in previous studies.
Relatedly, blockade of neuronal activity or activity at dopamine receptors decreases the amount
of effort rats are willing to expend to receive reward (Ghods-Sharifi & Floresco, 2010; Nunes et
al., 2013; Salamone et al., 2007), which may contribute to the lower rate of pressing during
reward-seeking observed across the two reward-seeking tasks examined.
An open question stemming from these results relates to why the NAcS, in comparison to
the NAcC, preferentially regulates response-inhibition. One likely explanation relates to the
partially segregated pattern of afferent input made to each region. The NAcS receives projections
from regions of the vmPFC and caudal BLA that regulate response-inhibition, while neurons in
the dorsal mPFC and rostral BLA that promote behavioral activation project to the NAcC
(Berendse, Galis-de Graaf, et al., 1992; Kita & Kitai, 1990; Sesack et al., 1989). Thus, when an
animal encounters a cue that predicts punishment, for example, glutamatergic activity from
vmPFC or caudal BLA may enhance activity in a subpopulation of neurons within the NAcS that
regulate response-inhibition. On the other hand, when the promotion of an active behavior is
necessitated, dorsal mPFC and rostral BLA may preferentially be activated to carry out this
function. In fact, these afferent projections have in many cases been borne out experimentally
(Ambroggi et al., 2008; Ghazizadeh et al., 2012; Ishikawa et al., 2008; McGinty & Grace, 2008;

176
Setlow, Roozendaal, & McGaugh, 2000). Still, this circuit description is clearly oversimplified,
as a fully segregated circuit is not supported by physiological or pharmacological analyses, as
illustrated in the present data and previous work (Ambroggi et al., 2011, 2008; Ishikawa et al.,
2008). For example, both NAcC and NAcS promote actions under some circumstances, such as
when performing an active avoidance. It is conceivable that this similarity in effect is mediated
by the extant, but potentially more sparse, overlapping projections from these afferent regions.
Still, the generation of dissociable functions within these two regions can likely be attributed in
part to differential afferent input.
Once these subregions have been activated, they must enact changes in response
promotion or inhibition via downstream projections. In comparison to study of the NAc afferents
that mediate complex forms of action selection, less is known about the downstream mediators
of such effects. Regardless, NAcS and NAcC project to largely distinct target areas, with NAcC
maintaining mostly inter-basal ganglia projections to structures like the substantia nigra and
lateral ventral pallidum, while NAcS projects to limbic associated structures, including the
ventral tegmental area (VTA), lateral hypothalamus, and medial ventral pallidum (Berendse,
Groenewegen, et al., 1992; Groenewegen et al., 1999; Ikemoto, 2007; Pennartz et al., 1994;
Zahm & Brog, 1992; Zahm & Heimer, 1993). Projections from NAcC to the basal ganglia leave
it poised to directly affect motor actions, consistent with the integral role of this nucleus in the
promotion of active behaviors reported here and elsewhere (Ambroggi et al., 2008; Ghods-
Sharifi & Floresco, 2010; Ishikawa et al., 2008; Salamone et al., 2007; Saunders & Robinson,
2012). For example, the ventral pallidum regulates the interaction between cortico-basal ganglia
loops that are necessary for reward-related behavior (for review, see Smith, Tindell, Aldridge, &
Berridge, 2010). These researchers propose that the ventral pallidum acts as a “final common

177
pathway” for limbic input to influence approach behavior, in this case mediating reward-seeking.
The dense projection from NAcC to the ventral pallidum may also allow this region to promote
both appetitive and aversively-motivated actions, as a function of limbic-striatal-pallidal
interactions.
Compared to the intra-basal ganglia projections made by the NAcC, projections from the
NAcS are relatively more diverse, consistent with the notion that this nucleus is a transition zone
between the extended amygdala (Alheid, 2003; Heimer et al., 1997). The projection from the
NAcS to the VTA may have direct relevance to aversively-mediated response-inhibition.
Optogenetic self-stimulation of VTA dopamine induces plasticity in NAcS neurons, increases the
excitability of OFC neurons, and produces punishment-resistant seeking of cocaine (Pascoli et
al., 2015). As NAcS neurons are primarily GABAergic, activity of these neurons would be
expected to inhibit VTA dopamine cells. Such a projection could phasically inhibit VTA
dopamine cells, preventing the activity necessary to produce reward-seeking during danger.
Supporting this dopamine-disinhibition account of punishment resistance is evidence that
silencing a key inhibitory afferent to the VTA, the rostromedial tegmentum, produces reward-
seeking during punishment similar to that which was produced by NAcS (or BLA) inactivation
here (Vento et al., 2017).
In addition, a pathway from the NAcS to the lateral hypothalamus has been directly
linked to the inhibition of drug-seeking (Millan et al., 2010). Following the extinction of alcohol
seeking, inactivation of the NAcS enhances reinstatement, while increasing activity in
neuropeptidergic cells within the lateral hypothalamus. Silencing the lateral hypothalamus
eliminates the effect of NAcS inactivation on reinstatement, suggesting that tonic inhibition of
the lateral hypothalamus by the NAcS enforces the learned inhibition of alcohol seeking (Millan

178
et al., 2010). Evidence implicating the lateral hypothalamus in aversion-mediated response-
inhibition comes from another study examining input from this region to the VTA during
punished sucrose-seeking (Nieh et al., 2015). Stimulation of this lateral hypothalamus to VTA
pathway disinhibits punished seeking, while silencing this circuit inhibits the same behavior, an
opposite pattern that would be expected if this effect was further mediated by the NAcS. Thus,
despite NAcS projecting to the lateral hypothalamus, this target region appears to function in an
opposite manner. Such a paradoxical finding implies that these regions may not function in
parallel during punishment, instead operating in concert with other relevant afferents. Overall,
further work is necessary to identify downstream targets of NAcC and NAcS through which they
can accomplish their respective roles in response-promotion and inhibition.
6.2 AMPH induces task-dependent bidirectional changes in instrumental punishment
In addition to probing NAc function during motivational conflict and active/passive avoidance,
we examined potential monoaminergic contributions to both behaviors by administering a
systemic dose of AMPH. AMPH administration provokes the release dopamine and serotonin
(Kuczenski & Segal, 1989; Seiden et al., 1993; Sitte & Freissmuth, 2015; Sulzer, Sonders,
Poulsen, & Galli, 2005), and has been used extensively to probe constructs such as incentive
salience and impulse control. Here, this manipulation provided valuable insight into
neurochemical targets related to aversively-motivated response inhibition. Previous research has
suggested that AMPH-induced monoamine release may enhance punishment sensitivity,
particularly in situations where punishment is associated with reward-seeking behavior (Broersen
et al., 1995; Geller & Seifter, 1960; Killcross et al., 1997; Lazareno, 1979; Leone et al., 1983).
Thus, when an instrumental action associated with reward-seeking is punished, AMPH or other
monoamine-releasers diminish seeking. Results of an exploratory analysis conducted here

179
suggested that AMPH similarly facilitated response-inhibition during punishment, supporting the
external validity of our Conflict task.
A similar pattern of behavior has been observed during performance on a more complex
decision-making assay, where rats have the option to choose a lever that delivers a small amount
of reward, with no chance of punishment, or another lever that delivers a larger reward with a
probability of punishment that increases across discrete trial blocks. AMPH administration biases
rats away from the lever that delivers a large reward and a probabilistic shock, indicative of
enhanced punishment sensitivity (Mitchell et al., 2011; Orsini, Trotta, et al., 2015; Simon et al.,
2011, 2009). This effect appears to be mediated by the dopamine D2 receptor, as antagonism of
this receptor blocks the impact of AMPH on risky choice (Simon et al., 2011).
These results suggest the intriguing possibility that dopamine D2 receptors within the
NAcS may promote response-inhibition during punished reward-seeking. Activity at these
receptors in the NAcS has previously been shown to oppose impulsive actions (Besson et al.,
2009). Similarly, highly impulsive animals have lower levels of D2 receptor expression within
the ventral striatum, an effect which is predictive of enhanced escalation of seeking of the
psychostimulant cocaine (Dalley et al., 2007). Impulsivity has also been directly related to the
taking of cocaine in a compulsive manner, operationalized as perseverance through foot-shock
punishment (Belin, Mar, Dalley, Robbins, & Everitt, 2008). Thus, dopaminergic activity and
receptor-expression within the NAcS may similarly relate to putative compulsive reward-
seeking, such as perseveration through instrumental punishment.
In contrast to the apparent promotion of suppression mediated by AMPH during conflict,
this same manipulation caused rats trained on an active/passive avoidance task to produce more
passive avoidance failures, indicative of a loss of response-inhibition. Despite this change in

180
passive failure rate, rats maintained the ability to inhibit responding upon receipt of a painful
stimulus, as the total number of passive presses did not differ following AMPH treatment. This
later finding implies that pain sensitivity is intact following AMPH administration, suggesting
that alterations in punishment-induced response-inhibition were not due to changes in pain
threshold.
The effect of AMPH on passive avoidance trials is in keeping with data suggesting that
AMPH administration can cause response-inhibitory deficits on No-Go trials of a Go/No-Go task
(Blackburn & Hevenor, 1996), and induce impulsive actions, a subtype of impulse control deficit
that reflects motor behavior produced without forethought (Pattij et al., 2007). Lesions of the
NAcS block the impact of AMPH on impulsive actions (Murphy et al., 2008), as does intra-
NAcS blockade of D2./3 receptors (Pattij et al., 2007). Given that both the Conflict and
active/passive avoidance tasks assess the withholding of a punished response, it is surprising that
AMPH would produce an opposite pattern of results on each task. As outlined in Chapter 4,
AMPH has been proposed to affect task performance based on the salience of options or
outcomes (Orsini, Moorman, et al., 2015). For example, on the Conflict task, a behavior that
provokes a shock is further inhibited by AMPH because the shock is more salient than the
relatively richer schedule of reinforcement. Given that rats were trained on the active avoidance
portion of the task first, animals apply more salience to the active avoidance cue. This would
lead to a bias towards active avoidance, which may enhance the prepotency of this response.
Support for such an account is provided by the relatively higher levels of passive avoidance
failure, as compared to active avoidance failure observed in rats at baseline. AMPH
administration may further enhance this bias, promoting approach behavior to a pathological
degree, and causing passive avoidance failures.

181
6.3 Experimental merits and future directions
While the present results provide meaningful insight into brain regions that are relevant to
aspects of aversive-motivation, a number of methodological issues bear considering. First, these
studies were conducted primarily using a single methodology, reversible pharmacological
inactivations. This consistency was necessary to facilitate the generalization of findings related
to aversively-motivated response-inhibition across tasks. Additionally, inactivations are a
preferable first pass technique to traditional permanent lesion studies for examining novel
functions of brain nuclei, as they are likely less susceptible to compensatory mechanisms that
may obscure the role of the targeted region (Poulos, Ponnusamy, Dong, & Fanselow, 2010;
Zelikowsky, Bissiere, Hast, Bennett, & Abdipranoto, 2013). Still, the limitations of this
technique warrant discussion. First, we targeted small brain subnuclei, which are often separated
by less than 1 mm. This proximity raises the possibility that our effects may be mediated in part
by diffusion from the targeted region into neighboring regions. Most studies examining the
functional spread of microinfusions conducted in the manner described here have found that
functional spread ranges from between 0.5-3 mm in situ (Allen et al., 2008; Edeline, Hars,
Hennevin, & Cotillon, 2002; Lorenzini et al., 1995). Thus, there is some possibility that
contamination in surrounding regions may explain some of the present observations. Although
we cannot exclude this possibility, the key behavioral dissociations observed in the majority of
studies described here were in opposite directions, which would be difficult to reconcile based
simply on drug diffusion outside of the region of interest. In many of the cases presented here,
results fit into a theoretical framework outlined in directional hypotheses, a fact that would be
inconsistent with a non-specific drug effect. Similarly, we have used these same infusion
parameters to dissociate these two regions on a variety of behavioral tasks, previously (Dalton et
al., 2014; Floresco et al., 2008; Ghods-Sharifi & Floresco, 2010; Stopper & Floresco, 2011).

182
Still, new techniques have been developed that will aid in testing the hypotheses
generated by the present thesis with a higher degree of specificity. The most relevant of these
techniques are optogenetics and chemogenetics, both of which utilize viral or genetic methods to
target cells, enabling the expression of engineered channels that can be manipulated precisely
without relying on drug diffusion (Britt & Bonci, 2013; Johansen, Wolff, Lüthi, & Ledoux,
2012; Roth, 2016; Stuber, Britt, & Bonci, 2012). Broadly, these techniques are extremely well-
suited for the investigation of small brain nuclei. For example, optogenetic manipulations allow
particular brain subnuclei to be infected and then targeted with light, minimizing concerns
regarding drug diffusion. Similarly, chemogenetic techniques allow for the expression of
receptors engineers to respond to a specific, non-bioactive ligand. Once receptors have been
infused into a particular subregion, this ligand can be administered systemically at doses that
cross the blood-brain barrier (but see Gomez et al., 2017), eliminating the need to directly infuse
drug into a brain region, which can potentially impact baseline behavior (see Chapter 2,
infralimbic effect prior to test). Similarly, circuit-based investigations can be conducted with
more confidence regarding the anatomical specificity of the targeted projection. Using the
PLNAcS disconnection experiment from Chapter 3 as an example, virus coding for an
excitatory or inhibitory channel could be infused into the PL, which is eventually trafficked in an
anterograde fashion and expressed in axon terminals in projection regions. Optic fibers can then
be implanted in the terminal region of interest, in this case, the NAcS, allowing for light-based
manipulation of PL axon terminals located in the NAcS. Such an experiment eliminates the
necessity of ipsilateral control groups, for example, as stimulation of terminals eliminates the
possibility that an effect is mediated by projections to the contralateral hemisphere or a third
brain region.

183
In addition to the refinement in anatomical targeting, the temporal specificity afforded by
these modern manipulations is dramatically better than that provided by pharmacological
inactivations. Optogenetic stimulation or inhibition allows for precise, millisecond control over
neuronal activity. On tasks such as those conducted here, the bolus infusion of receptor agonists
eliminates activity for upwards of two hours (Duuren et al., 2007; Edeline et al., 2002). While
pharmacological methods allow for the gross assessment of a region’s contribution to behavior,
they preclude the assessment of which specific task epochs the region is involved in. Given that
neural activity is often time-locked to particular task events (Ambroggi et al., 2011; Burgos-
Robles, Vidal-Gonzalez, & Quirk, 2009; Kim et al., 2017; Nieh et al., 2015), modulating
neuronal activity during such periods could refine our understanding of the contributions that
each brain region makes to specific components of a given behavior. This may help tease apart
effects like those observed on the Conflict task (Chapter 4), where the impact on reward-seeking
during safety may be mediated by functions unrelated to the direct inhibition of reward-seeking
during punishment, for example.
Such precision is necessary as it is becoming increasingly apparent that even within brain
subnuclei, heterogeneous populations of neurons exist that may not have the same impact on
behavior. For example, Kim and colleagues (2017) demonstrated that neural activity within a
projection from mPFC to the NAcS, but not from mPFC to VTA, was correlated with response-
inhibition during potential threat. This circuit was then broken down even further, with only a
subpopulation of shock-activated cells within the mPFC to NAcS projection being of crucial
relevance to the suppression of reward-seeking during danger. Thus, while pharmacological
disconnections may be able to identify the necessity of one projection versus another, the ability
to delve deeper into circuit-based mechanisms requires the use of optogenetic or chemogenetic

184
techniques paired with molecular and activity-based tagging of neurons. Some combination of
these techniques could be used to probe the circuit-basis of the effects observed in Chapters 4
and 5. For example, the downstream region through which BLA enforces the inhibition of
reward-seeking behavior during punishment is currently unknown. Given the results outlined in
the present thesis, the NAcS may be one such output region, which could be confirmed by
infusing virus coding for an excitatory or inhibitory channel into the BLA, and placing optic
fibers into the NAcS to stimulate or inhibit activity within this projection. Similarly, the
active/passive avoidance task is likely mediated by cortico-limbic-striatal circuitry, and contains
numerous time-locked events that would be amenable to interrogation through a combination of
in vivo electrophysiology and optogenetic or chemogenetic manipulation. These experiments
could be accompanied by receptor-specific pharmacological manipulations to examine the
contribution of various neuromodulators, such as dopamine acting at the D2 receptor within the
NAcS, to the behaviors identified in this thesis, as there is no methodological substitute at
present for the investigation of these targets (Jenni et al., 2017).
A final important limitation of the present data set is that our main outcome measure for
Chapters 2-4 was conditioned suppression, which is the absence of a response. This measure was
chosen as our stated interest was in the impact of fear on motivated behavior, emphasizing
aversively-motivated response-inhibition. While this measure often correlates strongly with other
measures of conditioned fear, such as freezing (McDannald & Galarce, 2011; Sierra-Mercado et
al., 2011), we can only speculate on what the animal is doing during these task epochs, as our
operant chambers are not equipped with cameras for the assessment of other defensive reactions.
Given that the expression of conditioned freezing is generally incompatible with ongoing operant
behavior, it is possible that some of our results may be explained by changes in conditioned

185
freezing. With regards to motivational conflict, previous studies using similar methodology have
demonstrated that freezing is essentially eliminated during punishment training, and is not
affected by BLA inactivations that disinhibit instrumental behavior during punishment (Jean-
Richard-Dit-Bressel & McNally, 2015). Thus, it is unlikely that these results could be explained
by changes in the expression of freezing.
Similarly, previous work has shown that freezing diminishes over the course of training
on an active avoidance task where rats concomitantly can lever-press for reward (Bravo-Rivera
et al., 2014; Oleson et al., 2012). While not identical to the present active/passive avoidance task,
one study illustrates that the refinement of behavior when behavioral responses compete (in this
case, active avoidance involves standing on a platform that entirely prevents reward-seeking)
involves a decrease in freezing and an increase in reward-seeking and avoidance. However,
Bravo-Rivera and colleagues (2014) have shown that inactivation of the NAc (mostly targeting
the NAcC) dramatically impairs active avoidance, in part by potentiating freezing during
presentations of the active avoidance stimulus. Thus, a possible explanation for the decrease in
avoidance or motivational conflict performance in NAcC-inactivated animals is an increase in
conditioned freezing. One piece of evidence suggesting that freezing alone may not explain the
diminished active avoidance observed following NAcC inactivation is that locomotor activity
was normal following NAcC inactivation. If NAcC-inactivation caused freezing to predominate
during active/passive avoidance, one might expect that the level of locomotor activity would be
lower, which was not the case. Still, concurrent measurement of freezing during these tasks is the
only way to truly eliminate this possibility in the future.

186
6.4 Relevance to neuropsychiatric disease
While we view this work through the lens of basic science, it is important to consider
what implications the present results have for brain dysfunction, as occurs in numerous
neuropsychiatric conditions. Of the most relevance to the present experiments are disorders
characterized by compulsive or impulsive patterns of behavior. For example, in substance abuse,
reward-seeking often occurs despite negative punishment (American Psychiatric Association,
2013). This phenotype has been suggested to be due to a deficit in the response-inhibition
typically induced by an aversive consequence, and can be assessed pre-clinically by using
conditioned suppression paradigms (Belin-Rauscent et al., 2016; Chen et al., 2013; Limpens et
al., 2014; Nieh et al., 2015; Pascoli et al., 2015). In humans, homologous regions of the
prefrontal cortex to those which we showed are involved in suppressing reward-seeking during
instrumental punishment and Pavlovian fear in rats, have been shown to be hypoactive during
impulse control in cocaine addicts (Goldstein & Volkow, 2011; Morein-Zamir et al., 2013).
Interestingly, these prefrontal deficits are related to decreased dopamine D2 receptor expression
in the NAc of addicted individuals, even following protracted drug-abstinence (Volkow et al.,
2009; Volkow, Wang, Fowler, Tomasi, & Telang, 2011). These D2 receptors are thought to be
inhibitory, suggesting that a loss of signaling at this dopaminergic substrate may contribute to
inhibitory control deficits (Everitt et al., 2008; Volkow & Morales, 2015). Unfortunately,
technological limitations in human imaging have not permitted the subregional assessment of
such effects. Taken together, these findings strongly implicate the prefrontal cortex and NAc in
aspects of response-inhibition of direct relevance to substance use disorders.
Our results are also potentially relevant to disorders of fear or anxiety, which are
characterized by deficits in fear discrimination, extinction, and aberrant avoidance (Duits, Cath,

187
Lissek, Hox, Hamm, Engelhard, van den Hout, et al., 2015; Graham & Milad, 2011; Jovanovic
& Norrholm, 2011; Lissek et al., 2014; Maner & Schmidt, 2006). For example, activity within
human ventromedial PFC promotes extinction (Milad, Wright, et al., 2007), while activity of the
dorsal ACC promotes fear expression (Delgado, Nearing, et al., 2008; Milad, Quirk, et al., 2007).
In the present study, we recapitulated a dorsal ACC-like effect by examining the function of PL
cortex during early fear extinction, inactivation of which potently inhibited fear expression. IL
cortex, which has been described as being functionally homologous to the ventromedial PFC in
humans (Heilbronner et al., 2016; Milad & Quirk, 2012), had a similar effect, in contrast to its
established role in fear extinction. Although methodological concerns clouded the interpretation
of this result, we suggest that ventral regions of the PFC like the IL cortex may promote fear
expression under certain conditions, such as when conflict exists between opposing motivational
drives. Such a function is consistent with the deficits observed in individuals with damage to the
vmPFC on tasks assessing emotion-guided decision-making, such as under conditions of risk
(Bechara, Damasio, Damasio, & Anderson, 1994; Bechara et al., 2000; Bechara, Damasio,
Damasio, & Lee, 1999; Clark et al., 2008).
Our results further suggest that mPFC and NAc are not critical for fear discrimination,
which is characteristically disturbed in individuals suffering from anxiety or post-traumatic stress
(Duits, Cath, Lissek, Hox, Hamm, Engelhard, van den Hout, et al., 2015), as none of our
manipulations impacted the level of fear expressed towards the CS-. This is particularly
interesting given that we have previously shown that mPFC disinhibition can elevate fear
expressed towards a CS-, while simultaneously decreasing fear towards a CS+, indicating a loss
of discrimination (Piantadosi & Floresco, 2014). Such results imply that regions downstream to
the mPFC, such as the basal amygdala, which has been shown to encode the presentation of a

188
neutral CS- in non-human primates (Genud-Gabai et al., 2013), or ventral hippocampus, which is
hyperactive during fear generalization in patients with post-traumatic stress disorder (Kaczkurkin
et al., 2017), may mediate fear discrimination.
Finally, we evaluated response-inhibition and promotion during a fully aversively-mediated
active/passive avoidance task. In humans, active avoidance has been associated with ventral
striatal activity, which includes the NAc (Delgado et al., 2009; Levita et al., 2012). The degree of
NAc activation during active avoidance has been positively correlated with state anxiety,
suggesting that high anxiety may co-occur with high levels of avoidance, consistent with clinical
findings (Dymond & Roche, 2009). To date, only one study has examined the response-
inhibitory pole of passive avoidance, with results suggesting that NAc deactivations may be of
critical importance to this behavior (Levita et al., 2012). In this thesis, we observed that NAcS
inactivation provoked inhibitory control failures, an effect opposite to what would be predicted
from this previous imaging study. Although these results may relate to the difficulty (and
possibly inaccuracy) of interpreting BOLD deactivations , they do provide evidence that neurons
within the NAc are sensitive to passive avoidance performance (Hayes & Huxtable, 2012;
Mishra et al., 2011). Thus, further basic and translational research on this task, ideally utilizing
imaging techniques that can dissociate the major subdivisions of the NAc in humans, and
employing manipulations with improved anatomical and temporal specificity in rats, is
necessary.
6.5 Conclusion
Overall, the present results add to a growing body of literature suggesting that the
heterogeneity within brain regions may have important functional implications. Here, we have
dissociated the two major subregions of the NAc, the shell and core, during aspects of

189
aversively-motivated behavior. Whether assessed in a Pavlovian or instrumental fashion,
response-suppression motivated by a potential aversive consequence was mediated by the NAcS,
while the NAcC simply promoted motivational vigor. In the case of Pavlovian fear expression, a
functional circuit between the PL cortex and NAcS appeared to mediate this effect, while
qualitative similarities existed between the functions of the BLA and NAcS during motivational
conflict. Similarly, performance of a complex avoidance behavior that required response-
promotion and response-inhibition necessitated the function of these subnuclei, with both regions
being necessary for normal response-promotion, and the NAcS being necessary for response-
inhibition.
This thesis represents some of the first evidence for a dissociable function of these
regions in aversively-motivated behavior. These results are generally coherent hypotheses
suggesting a role for the NAcC in approach behavior, and the NAcS in response-suppression.
They also provide more evidence against a reward-specific interpretation of NAc function
(Levita et al., 2009; Salamone, 1994). Instead, these results suggest that bivalent motivational
signals can affect actions via differential activation of NAc subnuclei. Although further work is
necessary to clarify the specific circuits that mediate these effects, it is likely that differential
input from subregions of the PFC and amygdala, and output to downstream projections within
the basal ganglia and mesencephalic structures, helps to produce these distinct mechanism
guiding action selection.

190
References
Abe, Y., Sakai, Y., Nishida, S., Nakamae, T., Yamada, K., Fukui, K., & Narumoto, J. (2015).
Hyper-influence of the orbitofrontal cortex over the ventral striatum in obsessive-
compulsive disorder. European Neuropsychopharmacology, 25(11), 1898–1905.
https://doi.org/10.1016/j.euroneuro.2015.08.017
Aberman, J. E., & Salamone, J. D. (1999). Nucleus Accumbens dopamine depletions make rats
more sensitive to high ratio requirements but do not impair primary food reinforcement.
Neuroscience, 92(2), 545–552. https://doi.org/10.1097/00008877-199908001-00202
Adolphs, R. (2013). The biology of fear. Current Biology, 23(2), R79–R93.
https://doi.org/10.1016/j.cub.2012.11.055
Adolphs, R., Tranel, D., Damasio, H., & Damasio, A. R. (1995). Fear and the human amygdala.
The Journal of Neuroscience, 15(9), 5879–5891. https://doi.org/10.1016/j.conb.2008.06.006
Ahmadi, H., Nasehi, M., Rostami, P., & Zarrindast, M. R. (2013). Involvement of the nucleus
accumbens shell dopaminergic system in prelimbic NMDA-induced anxiolytic-like
behaviors. Neuropharmacology, 71, 112–123.
https://doi.org/10.1016/j.neuropharm.2013.03.017
Ahmari, S. E., Spellman, T., Douglass, N. L., Kheirbek, M. A., Simpson, H. B., Deisseroth, K.,
… Hen, R. (2013). Repeated Cortico-Striatal Stimulation Generates Persistent OCD-Like
Behavior. Science, 340(6137), 1234–1239. https://doi.org/10.1126/science.1234733
Akirav, I., Raizel, H., & Maroun, M. (2006). Enhancement of conditioned fear extinction by
infusion of the GABA(A) agonist muscimol into the rat prefrontal cortex and amygdala. The
European Journal of Neuroscience, 23(3), 758–64. https://doi.org/10.1111/j.1460-
9568.2006.04603.x
Alheid, G. F. (2003). Extended amygdala and basal forebrain. Ann N Y Acad Sci, 985, 185–205.
https://doi.org/10.1111/j.1749-6632.2003.tb07082.x
Allcoat, D., Greville, W. J., Newton, P. M., & Dymond, S. (2015). Frozen with fear: Conditioned
suppression in a virtual reality model of human anxiety. Behavioural Processes, 118, 98–
101. https://doi.org/10.1016/j.beproc.2015.06.011
Allen, T. a., Narayanan, N. S., Kholodar-Smith, D. B., Zhao, Y., Laubach, M., & Brown, T. H.
(2008). Imaging the spread of reversible brain inactivations using fluorescent muscimol.
Journal of Neuroscience Methods, 171(1), 30–38.
https://doi.org/10.1016/j.jneumeth.2008.01.033
Ambroggi, F., Ghazizadeh, A., Nicola, S. M., & Fields, H. L. (2011). Roles of nucleus
accumbens core and shell in incentive-cue responding and behavioral inhibition. Journal of
Neuroscience, 31(18), 6820–6830. https://doi.org/10.1523/JNEUROSCI.6491-10.2011
Ambroggi, F., Ishikawa, A., Fields, H. L., & Nicola, S. M. (2008). Basolateral amygdala neurons
facilitate reward-seeking behavior by exciting nucleus accumbens neurons. Neuron, 59(4),
648–661. https://doi.org/10.1016/j.neuron.2008.07.004
American Psychiatric Association. (2013). The Diagnostic and Statistical Manual of Mental

191
Disorders: DSM 5. Washington, D C: American Psychiatric Press.
Amorapanth, P. (1999). Lesions of Periaqueductal Gray Dissociate-Conditioned Freezing From
Conditioned Suppression Behavior in Rats. Learning & Memory, 6(5), 491–499.
https://doi.org/10.1101/lm.6.5.491
Antunes, R., & Moita, M. A. (2010). Discriminative auditory fear learning requires both tuned
and nontuned auditory pathways to the amygdala. Journal of Neuroscience, 30(29), 9782–
9787. https://doi.org/10.1523/JNEUROSCI.1037-10.2010
Baarendse, P. J. J., & Vanderschuren, L. J. M. J. (2012). Dissociable effects of monoamine
reuptake inhibitors on distinct forms of impulsive behavior in rats. Psychopharmacology,
219(2), 313–326. https://doi.org/10.1007/s00213-011-2576-x
Badrinarayan, A., Wescott, S. A., Vander Weele, C. M., Saunders, B. T., Couturier, B. E.,
Maren, S., & Aragona, B. J. (2012). Aversive stimuli differentially modulate real-time
dopamine transmission dynamics within the nucleus accumbens core and shell. The Journal
of Neuroscience : The Official Journal of the Society for Neuroscience, 32(45), 15779–90.
https://doi.org/10.1523/JNEUROSCI.3557-12.2012
Baliki, M. N., Mansour, A., Baria, A. T., Huang, L., Berger, S. E., Fields, H. L., & Apkarian, A.
V. (2013). Parceling human accumbens into putative core and shell dissociates encoding of
values for reward and pain. The Journal of Neuroscience : The Official Journal of the
Society for Neuroscience, 33(41), 16383–93. https://doi.org/10.1523/JNEUROSCI.1731-
13.2013
Balog, Z., Somlai, Z., & Kéri, S. (2013). Aversive conditioning, schizotypy, and affective
temperament in the framework of the salience hypothesis. Personality and Individual
Differences, 54(1), 109–112. https://doi.org/10.1016/j.paid.2012.08.020
Bari, A., & Robbins, T. W. (2013). Inhibition and impulsivity: Behavioral and neural basis of
response control. Progress in Neurobiology, 108, 44–79.
https://doi.org/10.1016/j.pneurobio.2013.06.005
Barrett, R. J., Leith, N. J., & Ray, O. S. (1972). Permanent facilitation of avoidance behavior by
d-amphetamine and scopolamine. Psychopharmacologia, 25(4), 321–331.
https://doi.org/10.1007/BF00421971
Basar, K., Sesia, T., Groenewegen, H., Steinbusch, H. W. M., Visser-Vandewalle, V., & Temel,
Y. (2010). Nucleus accumbens and impulsivity. Progress in Neurobiology, 92(4), 533–557.
https://doi.org/10.1016/j.pneurobio.2010.08.007
Bast, T., Zhang, W. N., & Feldon, J. (2001). The ventral hippocampus and fear conditioning in
rats: Different anterograde amnesias of fear after tetrodotoxin inactivation and infusion of
the GABAA agonist muscimol. Experimental Brain Research, 139(1), 39–52.
https://doi.org/10.1007/s002210100746
Bates, D., Mächler, M., Bolker, B., & Walker, S. (2014). Fitting Linear Mixed-Effects Models
using lme4. Eprint arXiv:1406.5823, 67(1), 51. https://doi.org/10.18637/jss.v067.i01
Bechara, A., Damasio, A. R., Damasio, H., & Anderson, S. W. (1994). Insensitivity to future
consequences following damage to human prefrontal cortex. Cognition, 50, 7–15.

192
https://doi.org/10.1016/0010-0277(94)90018-3
Bechara, A., Damasio, H., & Damasio, A. R. (2000). Emotion, decision making and the
orbitofrontal cortex. Cerebral Cortex (New York, N.Y. : 1991), 10(3), 295–307.
https://doi.org/10.1093/cercor/10.3.295
Bechara, A., Damasio, H., Damasio, A. R., & Lee, G. P. (1999). Different contributions of the
human amygdala and ventromedial prefrontal cortex to decision-making. The Journal of
Neuroscience : The Official Journal of the Society for Neuroscience, 19(13), 5473–5481.
https://doi.org/0270-6474/99/19135473-09$05.00/0
Bechara, A., Tranel, D., Damasio, H., Adolphs, R., Rockland, C., & Damasio, A. R. (1995).
Double dissociation of conditioning and declarative knowledge relative to the amygdala and
hippocampus in humans. Science, 269(25 August), 1115–1118.
https://doi.org/10.1126/science.7652558
Belin-Rauscent, A., Fouyssac, M., Bonci, A., & Belin, D. (2016). How preclinical models
evolved to resemble the diagnostic criteria of drug addiction. Biological Psychiatry, 79(1),
39–46. https://doi.org/10.1016/j.biopsych.2015.01.004
Belin, D., Mar, A. C., Dalley, J. W., Robbins, T. W., & Everitt, B. J. (2008). High Impulsivity
Predicts the Switch to Compulsive Cocaine-Taking. Science, 320(5881), 1352–1355.
https://doi.org/10.1126/science.1158136
Berendse, H. W., Galis-de Graaf, Y., & Groenewegen, H. J. (1992). Topographical organization
and relationship with ventral striatal compartments of prefrontal corticostriatal projections
in the rat. The Journal of Comparative Neurology, 316(3), 314–347.
https://doi.org/10.1002/cne.903160305
Berendse, H. W., Groenewegen, H. J., & Lohman, A. H. (1992). Compartmental distribution of
ventral striatal neurons projecting to the mesencephalon in the rat. The Journal of
Neuroscience : The Official Journal of the Society for Neuroscience, 12(6), 2079–2103.
https://doi.org/http://www.jneurosci.org/content/12/6/2079.long
Berger, D., & Brush, F. (1975). Rapid acquisition of discrete-trial lever-press avoidance: Effects
of signal-shock interval. Journal of the Experimental Analysis of Behavior, 24(2), 227–239.
Berridge, K. C. (2012). From prediction error to incentive salience: mesolimbic computation of
reward motivation. The European Journal of Neuroscience, 35(7), 1124–43.
https://doi.org/10.1111/j.1460-9568.2012.07990.x
Berridge, K. C., & Kringelbach, M. L. (2013). Neuroscience of affect: Brain mechanisms of
pleasure and displeasure. Current Opinion in Neurobiology. Elsevier Ltd.
https://doi.org/10.1016/j.conb.2013.01.017
Besson, M., Belin, D., McNamara, R., Theobald, D. E., Castel, A., Beckett, V. L., … Dalley, J.
W. (2009). Dissociable control of impulsivity in rats by dopamine D2/3 receptors in the
core and shell subregions of the nucleus accumbens. Neuropsychopharmacology, 35(2),
560–569. https://doi.org/10.1038/npp.2009.162
Blackburn, J. R., & Hevenor, S. J. (1996). Amphetamine disrupts negative patterning but does
not produce configural association deficits on an alternative task. Behavioural Brain

193
Research, 80(1–2), 41–49. https://doi.org/10.1016/0166-4328(96)00017-4
Blaiss, C. A., & Janak, P. H. (2009). The nucleus accumbens core and shell are critical for the
expression, but not the consolidation, of Pavlovian conditioned approach. Behavioural
Brain Research, 200(1), 22–32. https://doi.org/10.1016/j.bbr.2008.12.024
Blanchard, R. J., & Blanchard, C. D. (1969). Passive and active reactions to fear-eliciting
stimuli. Journal of Comparative and Physiological Psychology, 68(1), 129–135.
https://doi.org/10.1037/h0027676
Block, A. E., Dhanji, H., Thompson-Tardif, S. F., & Floresco, S. B. (2007). Thalamic-prefrontal
cortical-ventral striatal circuitry mediates dissociable components of strategy set shifting.
Cerebral Cortex (New York, N.Y. : 1991), 17(7), 1625–36.
https://doi.org/10.1093/cercor/bhl073
Bolles, R. C. (1970). Species-specific defense reactions and avoidance learning. Psychological
Review, 77(1), 32–48. https://doi.org/10.1007/BF03001275
Boschen, S. L., Wietzikoski, E. C., Winn, P., & Cunha, C. Da. (2011). The role of nucleus
accumbens and dorsolateral striatal D2 receptors in active avoidance conditioning.
Neurobiology of Learning and Memory, 96(2), 254–262.
https://doi.org/10.1016/j.nlm.2011.05.002
Bouton, M. E., & Bolles, R. C. (1980). Conditioned fear assessed by freezing and by the
suppression of three different baselines. Animal Learning & Behavior, 8(3), 429–434.
https://doi.org/10.3758/BF03199629
Bouton, M. E., & Moody, E. W. (2004). Memory processes in classical conditioning.
Neuroscience and Biobehavioral Reviews, 28(7), 663–674.
https://doi.org/10.1016/j.neubiorev.2004.09.001
Bracs, P., Gregory, P., & Jackson, D. (1984). Passive avoidance in rats: disruption by dopamine
applied to the nucleus accumbens. Psychopharmacology, 83(1), 70–75.
Bradfield, L. a, & McNally, G. P. (2010). The role of nucleus accumbens shell in learning about
neutral versus excitatory stimuli during Pavlovian fear conditioning. Learning & Memory
(Cold Spring Harbor, N.Y.), 17(7), 337–43. https://doi.org/10.1101/lm.1798810
Bravo-Rivera, C., Roman-Ortiz, C., Brignoni-Perez, E., Sotres-Bayon, F., & Quirk, G. J. (2014).
Neural Structures Mediating Expression and Extinction of Platform-Mediated Avoidance.
Journal of Neuroscience, 34(29), 9736–9742. https://doi.org/10.1523/JNEUROSCI.0191-
14.2014
Bressel, P. J. R. D., & McNally, G. P. (2014). The role of the lateral habenula in punishment.
PLoS ONE, 9(11). https://doi.org/10.1371/journal.pone.0111699
Britt, J. P., Benaliouad, F., McDevitt, R. A., Stuber, G. D., Wise, R. A., & Bonci, A. (2012).
Synaptic and behavioral profile of multiple glutamatergic inputs to the nucleus accumbens.
Neuron, 76(4), 790–803. https://doi.org/10.1016/j.neuron.2012.09.040
Britt, J. P., & Bonci, A. (2013). Optogenetic interrogations of the neural circuits underlying
addiction. Current Opinion in Neurobiology, 23(4), 539–545.

194
https://doi.org/10.1016/j.conb.2013.01.010
Broersen, L. M., Heinsbroek, R. P. W., de Bruin, J. P. C., Laan, J.-B., Joosten, R. N. J. M. A., &
Olivier, B. (1995). Local pharmacological manipulations of prefrontal dopamine affect
conflict behaviour in rats. Behavioural Pharmacology, 6(4), 395–404.
https://doi.org/10.1097/00008877-199506000-00010
Brog, J. S., Salyapongse, A., Deutch, A. Y., & Zahm, D. S. (1993). The patterns of afferent
innervation of the core and shell in the “accumbens” part of the rat ventral striatum:
Immunohistochemical detection of retrogradely transported fluoro-gold. Journal of
Comparative Neurology, 338(2), 255–278. https://doi.org/10.1002/cne.903380209
Bryden, D. W., & Roesch, M. R. (2015). Executive Control Signals in Orbitofrontal Cortex
during Response Inhibition, 35(9), 3903–3914. https://doi.org/10.1523/JNEUROSCI.3587-
14.2015
Büchel, C., Dolan, R. J., Armony, J. L., & Friston, K. J. (1999). Amygdala-hippocampal
involvement in human aversive trace conditioning revealed through event-related functional
magnetic resonance imaging. The Journal of Neuroscience : The Official Journal of the
Society for Neuroscience, 19(24), 10869–10876. https://doi.org/Cited By (since 1996)
199\rExport Date 21 February 2012
Budygin, E. A., Park, J., Bass, C. E., Grinevich, V. P., Bonin, K. D., & Wightman, R. M. (2012).
Aversive stimulus differentially triggers subsecond dopamine release in reward regions.
Neuroscience, 201, 331–337. https://doi.org/10.1016/j.neuroscience.2011.10.056
Bukalo, O., Pinard, C. R., Silverstein, S., Brehm, C., Hartley, N. D., Whittle, N., … Holmes, A.
(2015). Prefrontal inputs to the amygdala instruct fear extinction memory formation.
Science Advances, 1(6), e1500251–e1500251. https://doi.org/10.1126/sciadv.1500251
Burgos-Robles, A., Vidal-Gonzalez, I., & Quirk, G. J. (2009). Sustained conditioned responses
in prelimbic prefrontal neurons are correlated with fear expression and extinction failure.
Journal of Neuroscience, 29(26), 8474–8482. https://doi.org/10.1523/JNEUROSCI.0378-
09.2009
Calhoon, G. G., & O’Donnell, P. (2013a). Closing the gate in the limbic striatum: prefrontal
suppression of hippocampal and thalamic inputs. Neuron, 78(1), 181–190.
https://doi.org/10.1016/j.neuron.2013.01.032
Calhoon, G. G., & O’Donnell, P. (2013b). Closing the gate in the limbic striatum: prefrontal
suppression of hippocampal and thalamic inputs. Neuron, 78(1), 181–190.
https://doi.org/10.1016/j.neuron.2013.01.032
Campbell, B. A., & Teghtsoonian, R. (1958). Electrical and behavioral effects of different types
of shock stimuli on the rat. Journal of Comparative and Physiological Psychology, 51(2),
185–192. https://doi.org/10.1037/h0043856
Cardinal, R. N., Parkinson, J. A., Hall, J., & Everitt, B. J. (2002). Emotion and motivation: the
role of the amygdala, ventral striatum, and prefrontal cortex. Neuroscience & Biobehavioral
Reviews, 26(3), 321–352. https://doi.org/10.1016/S0149-7634(02)00007-6
Cardinal, R. N., Pennicott, D. R., Sugathapala, C. L., Robbins, T. W., & Everitt, B. J. (2001a).

195
Impulsive choice induced in rats by lesions of the nucleus accumbens core. Science, 292,
2499–2501. https://doi.org/10.1126/science.1060818
Cardinal, R. N., Pennicott, D. R., Sugathapala, C. L., Robbins, T. W., & Everitt, B. J. (2001b).
Impulsive choice induced in rats by lesions of the nucleus accumbens core. Science, 292,
2499–2501. https://doi.org/10.1126/science.1060818
Carlezon, W. A., & Thomas, M. J. (2009). Biological substrates of reward and aversion: A
nucleus accumbens activity hypothesis. Neuropharmacology, 56(SUPPL. 1), 122–132.
https://doi.org/10.1016/j.neuropharm.2008.06.075
Carlezon Jr., W. A., & Thomas, M. J. (2009). Biological substrates of reward and aversion: a
nucleus accumbens activity hypothesis. Neuropharmacology, 56 Suppl 1, 122–132.
https://doi.org/10.1016/j.neuropharm.2008.06.075
Chang, C., & Maren, S. (2010). Strain difference in the effect of infralimbic cortex lesions on
fear extinction in rats. Behavioral Neuroscience, 124(3), 391–397.
https://doi.org/10.1037/a0019479.Strain
Chen, B. T., Yau, H. J., Hatch, C., Kusumoto-Yoshida, I., Cho, S. L., Hopf, F. W., & Bonci, A.
(2013). Rescuing cocaine-induced prefrontal cortex hypoactivity prevents compulsive
cocaine seeking. Nature, 496(7445), 359–362. https://doi.org/10.1038/nature12024
Chen, V. M., Foilb, A. R., & Christianson, J. P. (2016). Inactivation of ventral hippocampus
interfered with cued-fear acquisition but did not influence later recall or discrimination.
Behavioural Brain Research, 296, 249–253. https://doi.org/10.1016/j.bbr.2015.09.008
Christakou, A., Robbins, T. W., & Everitt, B. J. (2004). Prefrontal cortical-ventral striatal
interactions involved in affective modulation of attentional performance: Implications for
corticostriatal circuit function. Journal of Neuroscience, 24(4), 773–780.
https://doi.org/10.1523/JNEUROSCI.0949-03.2004
Clark, L., Bechara, A., Damasio, H., Aitken, M., Sahakian, B., & Robbins, T. (2008).
Differential effects of insular and ventromedial prefrontal cortex lesions on risky decision-
making. Brain, 131(Pt 5), 1311–22. https://doi.org/10.1093/brain/awn066
Cole, B., & Robbins, T. (1989). Effects of 6-hydroxydopamine lesions of the nucleus accumbens
septi on performance of a 5-chice serial reaction time task in rats: mplications for theories
of selective attention and arousal. Behav Brain Res., 33(2), 165–179.
https://doi.org/10.1016/S0166-4328(89)80048-8
Cole, B., & Robbins, T. W. (1987). Amphetamine impairs the discriminative performance of rats
with dorsal noradrenergic bundle lesions on a 5-choice serial reaction time task: New
evidence for central dopaminergic-noradrenergic interactions. Psychopharmacology, 91(4),
458–466. https://doi.org/10.1007/BF00216011
Connolly, C. G., Foxe, J. J., Nierenberg, J., Shpaner, M., & Garavan, H. (2012). The
neurobiology of cognitive control in successful cocaine abstinence. Drug and Alcohol
Dependence, 121(1–2), 45–53. https://doi.org/10.1016/j.drugalcdep.2011.08.007
Conti, L. H., Maeiver, C. R., Ferkany, J. W., & Abreu, M. E. (1990). Footshock-induced freezing
behavior in rats as a model for assessing anxiolyties. Psychopharmacology, 102(4), 492–

196
497.
Corbit, L. H., & Balleine, B. W. (2005). Double Dissociation of Basolateral and Central
Amygdala Lesions on the General and Outcome-Specific Forms of Pavlovian-Instrumental
Transfer. Journal of Neuroscience, 25(4), 962–970.
https://doi.org/10.1523/JNEUROSCI.4507-04.2005
Corbit, L. H., & Balleine, B. W. (2011). The General and Outcome-Specific Forms of Pavlovian-
Instrumental Transfer Are Differentially Mediated by the Nucleus Accumbens Core and
Shell. Journal of Neuroscience, 31(33), 11786–11794.
https://doi.org/10.1523/JNEUROSCI.2711-11.2011
Corbit, L. H., Muir, J. L., & Balleine, B. W. (2001). The role of the nucleus accumbens in
instrumental conditioning: Evidence of a functional dissociation between accumbens core
and shell. The Journal of Neuroscience : The Official Journal of the Society for
Neuroscience, 21(9), 3251–3260.
https://doi.org/http://www.jneurosci.org/content/21/9/3251
Corcoran, K. A., & Quirk, G. J. (2007). Activity in prelimbic cortex is necessary for the
expression of learned, but not innate, fears. Journal of Neuroscience, 27(4), 840–844.
https://doi.org/10.1523/JNEUROSCI.5327-06.2007
Correia, S. S., McGrath, A. G., Lee, A., Graybiel, A. M., & Goosens, K. A. (2016). Amygdala-
ventral striatum circuit activation decreases long-term fear. eLife, 5, 1–25.
https://doi.org/10.7554/eLife.12669
Courtin, J., Bienvenu, T. C. M., Einarsson, E. Ö., & Herry, C. (2013). Medial prefrontal cortex
neuronal circuits in fear behavior. Neuroscience, 240, 219–42.
https://doi.org/10.1016/j.neuroscience.2013.03.001
Craig, A. D. (2009). How do you feel — now? The anterior insula and human awareness. Nature
Reviews Neuroscience, 10(1), 59–70. https://doi.org/10.1038/nrn2555
Crockett, M. J., Clark, L., & Robbins, T. W. (2009). Reconciling the role of serotonin in
behavioral inhibition and aversion: acute tryptophan depletion abolishes punishment-
induced inhibition in humans. The Journal of Neuroscience: The Official Journal of the
Society for Neuroscience, 29(38), 11993–11999.
https://doi.org/10.1523/JNEUROSCI.2513-09.2009
D’Amato, M. R., & Schiff, D. (1964). Long-term discriminated avoidance performance in the
rat. Journal of Comparative & Physiological Psychology, 57(1), 123–126.
https://doi.org/10.1037/h0046678
Dalley, J. W., Fryer, T. D., Brichard, L., Robinson, E. S. J., Theobald, D. E. H., Laane, K., …
Robbins, T. W. (2007). Nucleus Accumbens D2/3 Receptors Predict Trait Impulsivity and
Cocaine Reinforcement. Science, 315(5816), 1267–1270.
https://doi.org/10.1126/science.1137073
Dalton, G. L., Phillips, A. G., & Floresco, S. B. (2014). Preferential involvement by nucleus
accumbens shell in mediating probabilistic learning and reversal shifts. The Journal of
Neuroscience : The Official Journal of the Society for Neuroscience, 34(13), 4618–4626.

197
https://doi.org/10.1523/JNEUROSCI.5058-13.2014
Dalton, G. L., Wang, N. Y., Phillips, A. G., & Floresco, S. B. (2016). Multifaceted Contributions
by Different Regions of the Orbitofrontal and Medial Prefrontal Cortex to Probabilistic
Reversal Learning. Journal of Neuroscience, 36(6), 1996–2006.
https://doi.org/10.1523/JNEUROSCI.3366-15.2016
Davis, S., Butcher, S. P., & Morris, R. G. (1992). The NMDA receptor antagonist D-2-amino-5-
phosphonopentanoate (D-AP5) impairs spatial learning and LTP in vivo at intracerebral
concentrations comparable to those that block LTP in vitro. The Journal of Neuroscience :
The Official Journal of the Society for Neuroscience, 12(1), 21–34. https://doi.org/1345945
De Leonibus, E., Costantini, V. J. A., Castellano, C., Ferretti, V., Oliverio, A., & Mele, A.
(2003). Distinct roles of the different ionotropic glutamate receptors within the nucleus
accumbens in passive-avoidance learning and memory in mice. European Journal of
Neuroscience, 18(8), 2365–2373. https://doi.org/10.1046/j.1460-9568.2003.02939.x
Delgado, M. R., Jou, R. L., Ledoux, J. E., & Phelps, E. A. (2009). Avoiding negative outcomes:
tracking the mechanisms of avoidance learning in humans during fear conditioning.
Frontiers in Behavioral Neuroscience, 3, 1–9. https://doi.org/10.3389/neuro.08.033.2009
Delgado, M. R., Li, J., Schiller, D., & Phelps, E. A. (2008). The role of the striatum in aversive
learning and aversive prediction errors. Philosophical Transactions of the Royal Society of
London. Series B, Biological Sciences, 363(1511), 3787–3800.
https://doi.org/10.1098/rstb.2008.0161
Delgado, M. R., Nearing, K. I., LeDoux, J. E., & Phelps, E. A. (2008). Neural circuitry
underlying the regulation of conditioned fear and its relation to extinction. Neuron, 59(5),
829–838. https://doi.org/10.1016/j.neuron.2008.06.029
Deroche-Gamonet, V., Belin, D., & Piazza, P. V. (2004). Evidence for addiction-like behavior in
the rat. Science (New York, N.Y.), 305(5686), 1014–7.
https://doi.org/10.1126/science.1099020
Di Ciano, P., Robbins, T. W., & Everitt, B. J. (2008). Differential effects of nucleus accumbens
core, shell, or dorsal striatal inactivations on the persistence, reacquisition, or reinstatement
of responding for a drug-paired conditioned reinforcer. Neuropsychopharmacology :
Official Publication of the American College of Neuropsychopharmacology, 33(6), 1413–
1425. https://doi.org/10.1038/sj.npp.1301522
Dickinson, A., & Balleine, B. (1994). Motivational control of goal-directed action. Animal
Learning & Behavior, 22(1), 1–18. https://doi.org/10.3758/BF03199951
Dinsmoor, J. (2001). Stimuli inevitably generated by behavior that avoids electric shock are
inherently reinforcing. Journal of the Experimental Analysis of Behavior, 3(3), 311–333.
Dinsmoor, J., & Sears, G. W. (1973). Control of avoidance by a response-produced stimulus.
Learning and Motivation, 4, 284–293.
Do-Monte, F. H., Manzano-Nieves, G., Quinones-Laracuente, K., Ramos-Medina, L., & Quirk,
G. J. (2015). Revisiting the Role of Infralimbic Cortex in Fear Extinction with
Optogenetics. Journal of Neuroscience, 35(8), 3607–3615.

198
https://doi.org/10.1523/JNEUROSCI.3137-14.2015
Dombrowski, P. a., Maia, T. V., Boschen, S. L., Bortolanza, M., Wendler, E., Schwarting, R. K.
W., … Da Cunha, C. (2013). Evidence that conditioned avoidance responses are reinforced
by positive prediction errors signaled by tonic striatal dopamine. Behavioural Brain
Research, 241(1), 112–119. https://doi.org/10.1016/j.bbr.2012.06.031
Duits, P., Cath, D. C., Lissek, S., Hox, J. J., Hamm, A. O., Engelhard, I. M., … Baas, J. M. P.
(2015). Updated meta-analysis of classical fear conditioning in the anxiety disorders.
Depression and Anxiety, 32(4), 239–253. https://doi.org/10.1002/da.22353
Duits, P., Cath, D. C., Lissek, S., Hox, J. J., Hamm, A. O., Engelhard, I. M., … Baas, J. M. P.
(2015). Updated meta-analysis of classical fear conditioning in the anxiety disorders.
Depression and Anxiety, 32(4), 239–253. https://doi.org/10.1002/da.22353
Duuren, E. Van, Plasse, G. Van Der, Blom, R. Van Der, Joosten, R. N. J. M. A., Mulder, A. B.,
Pennartz, C. M. A., & Feenstra, M. G. P. (2007). Pharmacological Manipulation of
Neuronal Ensemble Activity by Reverse Microdialysis in Freely Moving Rats : A
Comparative Study of the Effects of Tetrodotoxin , Lidocaine , and Muscimol, 323(1), 61–
69. https://doi.org/10.1124/jpet.107.124784.ulations
Dymond, S., & Roche, B. (2009). A contemporary behavior analysis of anxiety and avoidance.
The Behavior Analyst, 32(1), 7–27.
Eagle, D. M., & Robbins, T. W. (2003). Lesions of the medial prefrontal cortex or nucleus
accumbens core do not impair inhibitory control in rats performing a stop-signal reaction
time task. Behavioural Brain Research, 146(1–2), 131–144.
https://doi.org/10.1016/j.bbr.2003.09.022
Edeline, J.-M., Hars, B., Hennevin, E., & Cotillon, N. (2002). Muscimol diffusion after
intracerebral microinjections: a reevaluation based on electrophysiological and
autoradiographic quantifications. Neurobiology of Learning and Memory, 78(1), 100–124.
https://doi.org/10.1006/nlme.2001.4035
Erlich, J. C., Bush, D. E. A., & Ledoux, J. E. (2012). The role of the lateral amygdala in the
retrieval and maintenance of fear-memories formed by repeated probabilistic reinforcement.
Frontiers in Behavioral Neuroscience, 6(April), 16.
https://doi.org/10.3389/fnbeh.2012.00016
Esclassan, F., Coutureau, E., Di Scala, G., & Marchand, A. R. (2009). Differential contribution
of dorsal and ventral hippocampus to trace and delay fear conditioning. Hippocampus,
19(1), 33–44. https://doi.org/10.1002/hipo.20473
Estes, W. K., & Skinner, B. F. (1941). Some quantitative properties of anxiety. Journal of
Experimental Psychology, 29(5), 390–400. https://doi.org/10.1037/h0062283
Everitt, B. J. (2014). Neural and psychological mechanisms underlying compulsive drug seeking
habits and drug memories - indications for novel treatments of addiction. European Journal
of Neuroscience, 40(1), 2163–2182. https://doi.org/10.1111/ejn.12644
Everitt, B. J., Belin, D., Economidou, D., Pelloux, Y., Dalley, J. W., & Robbins, T. W. (2008).
Neural mechanisms underlying the vulnerability to develop compulsive drug-seeking habits

199
and addiction. Philosophical Transactions of the Royal Society B: Biological Sciences,
363(1507), 3125–3135. https://doi.org/10.1098/rstb.2008.0089
Everitt, B. J., Cardinal, R. N., Parkinson, J. A., & Robbins, T. W. (2003). Appetitive behavior:
impact of amygdala-dependent mechanisms of emotional learning. Annals of the New York
Academy of Sciences, 985, 233–250. https://doi.org/10.1111/j.1749-6632.2003.tb07085.x
Fanselow, M. S. (1994). Neural organization of the defensive behavior system responsible for
fear. Psychonomic Bulletin & Review, 1(4), 429–438. https://doi.org/10.3758/BF03210947
Fanselow, M. S., & LeDoux, J. E. (1999). Why we think plasticity underlying pavlovian fear
conditioning occurs in the basolateral amygdala. Neuron, 23(2), 229–232.
https://doi.org/10.1016/S0896-6273(00)80775-8
Fanselow, M. S., & Lester, L. S. (1988). A functional behavioristic approach to aversively
motivated behavior: Predatory imminence as a determinant of the topography of defensive
behavior. Evolution and Learning, (May), 185–212.
Feil, J., Sheppard, D., Fitzgerald, P. B., Yücel, M., Lubman, D. I., & Bradshaw, J. L. (2010).
Addiction, compulsive drug seeking, and the role of frontostriatal mechanisms in regulating
inhibitory control. Neuroscience and Biobehavioral Reviews, 35(2), 248–275.
https://doi.org/10.1016/j.neubiorev.2010.03.001
Feja, M., Hayn, L., & Koch, M. (2014). Nucleus accumbens core and shell inactivation
differentially affects impulsive behaviours in rats. Progress in Neuro-Psychopharmacology
and Biological Psychiatry, 54, 31–42. https://doi.org/10.1016/j.pnpbp.2014.04.012
Feja, M., & Koch, M. (2014). Ventral medial prefrontal cortex inactivation impairs impulse
control but does not affect delay-discounting in rats. Behavioural Brain Research, 264,
230–9. https://doi.org/10.1016/j.bbr.2014.02.013
Feja, M., & Koch, M. (2015). Frontostriatal systems comprising connections between ventral
medial prefrontal cortex and nucleus accumbens subregions differentially regulate motor
impulse control in rats. Psychopharmacology, 232(7), 1291–1302.
https://doi.org/10.1007/s00213-014-3763-3
Fendt, M., & Fanselow, M. S. (1999). The neuroanatomical and neurochemical basis of
conditioned fear, 23, 743–760.
Fernando, A. B. P., Mar, A. C., Urcelay, G. P., Dickinson, A., & Robbins, T. W. (2015).
Avoidance behavior: A free-operant lever-press avoidance task for the assessment of the
effects of safety signals. Current Protocols in Neuroscience, 70, 8.32.1-8.32.12.
https://doi.org/10.1002/0471142301.ns0832s70
Fernando, A. B. P., Urcelay, G. P., Mar, A. C., Dickinson, A., & Robbins, T. W. (2013). The role
of the nucleus accumbens shell in the mediation of the reinforcing properties of a safety
signal in free-operant avoidance: Dopamine-dependent inhibitory effects of d-amphetamine.
Neuropsychopharmacology, 39(6), 1–11. https://doi.org/10.1038/npp.2013.337
Fernando, A. B. P., Urcelay, G. P., Mar, A. C., Dickinson, A., & Robbins, T. W. (2014). Safety
signals as instrumental reinforcers during free-operant avoidance. Learning & Memory
(Cold Spring Harbor, N.Y.), 21(9), 488–497. https://doi.org/10.1101/lm.034603.114

200
Figee, M., Luigjes, J., Smolders, R., Valencia-Alfonso, C.-E., van Wingen, G., de Kwaasteniet,
B., … Denys, D. (2013). Deep brain stimulation restores frontostriatal network activity in
obsessive-compulsive disorder. Nature Neuroscience, 16(4), 386–387.
https://doi.org/10.1038/nn.3344
Figee, M., Pattij, T., Willuhn, I., Luigjes, J., van den Brink, W., Goudriaan, A., … Denys, D.
(2016). Compulsivity in obsessive-compulsive disorder and addictions. European
Neuropsychopharmacology, 26(5), 856–868.
https://doi.org/10.1016/j.euroneuro.2015.12.003
Floresco, S. B. (2013, January). Prefrontal dopamine and behavioral flexibility: Shifting from an
“inverted-U” toward a family of functions. Frontiers in Neuroscience.
https://doi.org/10.3389/fnins.2013.00062
Floresco, S. B. (2015). The nucleus accumbens: An interface between cognition, emotion, and
action. Annual Review of Psychology, 66, 25–52. https://doi.org/10.1146/annurev-psych-
010213-115159
Floresco, S. B., Blaha, C. D., Yang, C. R., & Phillips, A. G. (2001). Dopamine D1 and NMDA
receptors mediate potentiation of basolateral amygdala-evoked firing of nucleus accumbens
neurons. The Journal of Neuroscience, 21(16), 6370–6376. https://doi.org/21/16/6370 [pii]
Floresco, S. B., & Ghods-Sharifi, S. (2007). Amygdala-prefrontal cortical circuitry regulates
effort-based decision making. Cerebral Cortex, 17(2), 251–260.
https://doi.org/10.1093/cercor/bhj143
Floresco, S. B., Ghods-Sharifi, S., Vexelman, C., & Magyar, O. (2006). Dissociable roles for the
nucleus accumbens core and shell in regulating set shifting. The Journal of Neuroscience :
The Official Journal of the Society for Neuroscience, 26(9), 2449–2457.
https://doi.org/10.1523/JNEUROSCI.4431-05.2006
Floresco, S. B., McLaughlin, R. J., & Haluk, D. M. (2008). Opposing roles for the nucleus
accumbens core and shell in cue-induced reinstatement of food-seeking behavior.
Neuroscience, 154(3), 877–884. https://doi.org/10.1016/j.neuroscience.2008.04.004
Floresco, S. B., Seamans, J. K., & Phillips, A. G. (1997). Selective roles for hippocampal,
prefrontal cortical, and ventral striatal circuits in radial-arm maze tasks with or without a
delay. Journal of Neuroscience, 17(5), 1880–1890. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/9030646
Floresco, S. B., Yang, C. R., Phillips, A. G., & Blaha, C. D. (1998). Basolateral amygdala
stimulation evokes glutamate receptor-dependent dopamine efflux in the nucleus
accumbens of the anaesthetized rat. European Journal of Neuroscience, 10(4), 1241–1251.
https://doi.org/10.1046/j.1460-9568.1998.00133.x
French, S. J., & Totterdell, S. (2002). Hippocampal and prefrontal cortical inputs
monosynaptically converge with individual projection neurons of the nucleus accumbens.
Journal of Comparative Neurology, 446(2), 151–165. https://doi.org/10.1002/cne.10191
French, S. J., & Totterdell, S. (2003). Individual nucleus accumbens-projection neurons receive
both basolateral amygdala and ventral subicular afferents in rats. Neuroscience, 119(1), 19–

201
31. https://doi.org/10.1016/S0306-4522(03)00150-7
Friedman, A., Homma, D., Gibb, L. G., Amemori, K. I., Rubin, S. J., Hood, A. S., … Graybiel,
A. M. (2015). A corticostriatal path targeting striosomes controls decision-making under
conflict. Cell, 161(6), 1320–1333. https://doi.org/10.1016/j.cell.2015.04.049
Gal, G., Schiller, D., & Weiner, I. (2005). Latent inhibition is disrupted by nucleus accumbens
shell lesion but is abnormally persistent following entire nucleus accumbens lesion: The
neural site controlling the expression and disruption of the stimulus preexposure effect.
Behavioural Brain Research, 162(2), 246–55. https://doi.org/10.1016/j.bbr.2005.03.019
Geller, I., & Seifter, J. (1960). The effects of meprobamate, barbiturates, d-amphetamine and
promazine on experimentally induced conflict in the rat. Psychopharmacology, 1(6), 482–
492. https://doi.org/10.1007/BF00429273
Gentry, R. N., Lee, B., & Roesch, M. R. (2016). Phasic dopamine release in the rat nucleus
accumbens predicts approach and avoidance performance. Nature Communications, 7,
13154. https://doi.org/10.1038/ncomms13154
Genud-Gabai, R., Klavir, O., & Paz, R. (2013). Safety signals in the primate amygdala. The
Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 33(46),
17986–94. https://doi.org/10.1523/JNEUROSCI.1539-13.2013
Ghazizadeh, A., Ambroggi, F., Odean, N., & Fields, H. L. (2012). Prefrontal cortex mediates
extinction of responding by two distinct neural mechanisms in accumbens shell. Journal of
Neuroscience, 32(2), 726–737. https://doi.org/10.1523/JNEUROSCI.3891-11.2012
Ghods-Sharifi, S., & Floresco, S. B. (2010). Differential effects on effort discounting induced by
inactivations of the nucleus accumbens core or shell. Behavioral Neuroscience, 124(2),
179–191. https://doi.org/10.1037/a0018932
Ghods-Sharifi, S., St Onge, J. R., & Floresco, S. B. (2009). Fundamental contribution by the
basolateral amygdala to different forms of decision making. The Journal of Neuroscience :
The Official Journal of the Society for Neuroscience, 29(16), 5251–5259.
https://doi.org/10.1523/JNEUROSCI.0315-09.2009
Gilmartin, M. R., & McEchron, M. D. (2005). Single neurons in the medial prefrontal cortex of
the rat exhibit tonic and phasic coding during trace fear conditioning. Behavioral
Neuroscience, 119(6), 1496–1510. https://doi.org/10.1037/0735-7044.119.6.1496
Gilmartin, M. R., Miyawaki, H., Helmstetter, F. J., & Diba, K. (2013). Prefrontal Activity Links
Nonoverlapping Events in Memory. Journal of Neuroscience, 33(26), 10910–10914.
https://doi.org/10.1523/JNEUROSCI.0144-13.2013
Goldstein, R. Z., Alia-Klein, N., Tomasi, D., Carrillo, J. H., Maloney, T., Woicik, P. A., …
Volkow, N. D. (2009). Anterior cingulate cortex hypoactivations to an emotionally salient
task in cocaine addiction. Proceedings of the National Academy of Sciences of the United
States of America, 106(23), 9453–8. https://doi.org/10.1073/pnas.0900491106
Goldstein, R. Z., & Volkow, N. D. (2011). Dysfunction of the prefrontal cortex in addiction:
neuroimaging findings and clinical implications. Nature Reviews Neuroscience, 12(11),
652–669. https://doi.org/10.1038/nrn3119

202
Gomez, J. L., Bonaventura, J., Lesniak, W., Mathews, W. B., Sysa-Shah, P., Rodriguez, L. A.,
… Michaelides, M. (2017). Chemogenetics revealed: DREADD occupancy and activation
via converted clozapine. Science, 357(6350), 503–507.
https://doi.org/10.1126/science.aan2475
Goosens, K. A., & Maren, S. (2001). Contextual and Auditory Fear Conditioning are Mediated
by the Lateral, Basal, and Central Amygdaloid Nuclei in Rats. Learning & Memory, 8(3),
148–155. https://doi.org/10.1101/lm.37601
Goto, Y., & O’Donnell, P. (2002). Timing-dependent limbic-motor synaptic integration in the
nucleus accumbens. Proceedings of the National Academy of Sciences of the United States
of America, 99(20), 13189–93. https://doi.org/10.1073/pnas.202303199
Graham, B. M., & Milad, M. R. (2011). The study of fear extinction: implications for anxiety
disorders. The American Journal of Psychiatry, 168(12), 1255–1265.
https://doi.org/10.1176/appi.ajp.2011.11040557
Greenberg, B. D., Gabriels, L. A., Malone, D. A., Rezai, A. R., Friehs, G. M., Okun, M. S., …
Nuttin, B. J. (2010). Deep brain stimulation of the ventral internal capsule/ventral striatum
for obsessive-compulsive disorder: worldwide experience. Molecular Psychiatry, 15(1), 64–
79. https://doi.org/10.1038/mp.2008.55
Greville, W. J., Newton, P. M., Roche, B., & Dymond, S. (2013). Conditioned suppression in a
virtual environment. Computers in Human Behavior, 29(3), 552–558.
https://doi.org/10.1016/j.chb.2012.11.016
Grillon, C., & Morgan, C. a. (1999). Fear-potentiated startle conditioning to explicit and
contextual cues in Gulf War veterans with posttraumatic stress disorder. Journal of
Abnormal Psychology, 108(1), 134–142. https://doi.org/10.1037/0021-843x.108.1.134
Groenewegen, H. J., Wright, C. I., Beijer, A. V. J., & Voorn, P. (1999). Convergence and
segregation of ventral striatal inputs and outputs. Annals of the New York Academy of
Sciences, 877, 49–63. https://doi.org/10.1111/j.1749-6632.1999.tb09260.x
Gruber, A. J., Hussain, R. J., & O’Donnell, P. (2009). The nucleus accumbens: a switchboard for
goal-directed behaviors. PloS One, 4(4), e5062.
https://doi.org/10.1371/journal.pone.0005062
Gruber, A. J., & O’Donnell, P. (2009). Bursting activation of prefrontal cortex drives sustained
up states in nucleus accumbens spiny neurons in vivo. Synapse (New York, N.Y.), 63(3),
173–180. https://doi.org/10.1002/syn.20593
Grueter, B. A., Rothwell, P. E., & Malenka, R. C. (2012). Integrating synaptic plasticity and
striatal circuit function in addiction. Current Opinion in Neurobiology, 22(3), 545–551.
https://doi.org/10.1016/j.conb.2011.09.009
Gutman, A. L., Ewald, V. A., Cosme, C. V, Worth, W. R., & Lalumiere, R. T. (2014). The
infralimbic and prelimbic cortices contribute to the inhibitory control of cocaine-seeking
behavior during a discriminative stimulus task in rats. https://doi.org/10.1111/adb.12434
Hagenaars, M. A., Oitzl, M., & Roelofs, K. (2014). Updating freeze: Aligning animal and human
research. Neuroscience and Biobehavioral Reviews, 47, 165–176.

203
https://doi.org/10.1016/j.neubiorev.2014.07.021
Hall, J., Parkinson, J. A., Connor, T. M., Dickinson, A., & Everitt, B. J. (2001). Involvement of
the central nucleus of the amygdala and nucleus accumbens core in mediating pavlovian
influences on instrumental behaviour. European Journal of Neuroscience, 13(10), 1984–
1992. https://doi.org/10.1046/j.0953-816X.2001.01577.x
Hanlon, E. C., Baldo, B. A., Sadeghian, K., & Kelley, A. E. (2004). Increases in food intake or
food-seeking behavior induced by GABAergic, opioid, or dopaminergic stimulation of the
nucleus accumbens: Is if hunger? Psychopharmacology, 172(3), 241–247.
https://doi.org/10.1007/s00213-003-1654-0
Haralambous, T., & Westbrook, R. F. (1999). An infusion of bupivacaine into the nucleus
accumbens disrupts the acquisition but not the expression of contextual fear conditioning.
Behavioral Neuroscience, 113(5), 925–940. https://doi.org/10.1037/0735-7044.113.5.925
Hariri, A. R., Gorka, A., Hyde, L. W., Kimak, M., Halder, I., Ducci, F., … Manuck, S. B. (2009).
Divergent Effects of Genetic Variation in Endocannabinoid Signaling on Human Threat-
and Reward-Related Brain Function. Biological Psychiatry, 66(1), 9–16.
https://doi.org/10.1016/j.biopsych.2008.10.047
Harrison, A. A., Everitt, B. J., & Robbins, T. W. (1997). Central 5-HT depletion enhances
impulsive responding without affecting the accuracy of attentional performance:
Interactions with dopaminergic mechanisms. Psychopharmacology, 133(4), 329–342.
https://doi.org/10.1007/s002130050410
Hayes, D. J., & Huxtable, A. G. (2012). Interpreting deactivations in neuroimaging. Frontiers in
Psychology, 3(FEB), 2–4. https://doi.org/10.3389/fpsyg.2012.00027
Heilbronner, S. R., Rodriguez-Romaguera, J., Quirk, G. J., Groenewegen, H. J., & Haber, S. N.
(2016). Circuit-Based Corticostriatal Homologies Between Rat and Primate. Biological
Psychiatry, 80(7), 509–521. https://doi.org/10.1016/j.biopsych.2016.05.012
Heimer, L., Alheid, G. F., de Olmos, J. S., Groenewegen, H. J., Haber, S. N., Harlan, R. E., &
Zahm, D. S. (1997). The accumbens: beyond the core-shell dichotomy. The Journal of
Neuropsychiatry and Clinical Neurosciences, 9(3), 354–381.
https://doi.org/10.1176/jnp.9.3.354
Heimer, L., Zahm, D. S., Churchill, L., Kalivas, P. W., & Wohltmann, C. (1991). Specificity in
the projection patterns of accumbal core and shell in the rat. Neuroscience, 41(1), 89–125.
https://doi.org/10.1016/0306-4522(91)90202-Y
Helmstetter, F. J., & Bellgowan, P. S. (1994). Effects of Muscimol Applied to the Basolateral
Amygdala on Acquisition and Expression of Contextual Fear Conditioning in Rats, 108(5),
1005–1009.
Hester, R., & Garavan, H. (2004). Executive dysfunction in cocaine addiction: evidence for
discordant frontal, cingulate, and cerebellar activity. The Journal of Neuroscience : The
Official Journal of the Society for Neuroscience, 24(49), 11017–22.
https://doi.org/10.1523/JNEUROSCI.3321-04.2004
Hjärthag, F., Helldin, L., Karilampi, U., & Norlander, T. (2010). Illness-related components for

204
the family burden of relatives to patients with psychotic illness. Social Psychiatry and
Psychiatric Epidemiology, 45(2), 275–283. https://doi.org/10.1007/s00127-009-0065-x
Ikemoto, S. (2007). Dopamine reward circuitry: Two projection systems from the ventral
midbrain to the nucleus accumbens-olfactory tubercle complex. Brain Research Reviews,
56(1), 27–78. https://doi.org/10.1016/j.brainresrev.2007.05.004
Ilango, A., Shumake, J., Wetzel, W., & Ohl, F. W. (2014). Contribution of emotional and
motivational neurocircuitry to cue-signaled active avoidance learning. Frontiers in
Behavioral Neuroscience, 8(October), 1–5. https://doi.org/10.3389/fnbeh.2014.00372
Ilango, A., Shumake, J., Wetzel, W., Scheich, H., & Ohl, F. W. (2012). The role of dopamine in
the context of aversive stimuli with particular reference to acoustically signaled avoidance
learning. Frontiers in Neuroscience, 6(SEP), 1–9. https://doi.org/10.3389/fnins.2012.00132
Ishikawa, A., Ambroggi, F., Nicola, S. M., & Fields, H. L. (2008). Dorsomedial prefrontal cortex
contribution to behavioral and nucleus accumbens neuronal responses to incentive cues. The
Journal of Neuroscience, 28(19), 5088–5098. https://doi.org/10.1523/JNEUROSCI.0253-
08.2008
Ishikawa, A., Ambroggi, F., Nicola, S. M., & Fields, H. L. (2010). Contributions of the
amygdala and medial prefrontal cortex to incentive cue responding. Neuroscience, 155(3),
573–584. https://doi.org/10.1016/j.neuroscience.2008.06.037.Contributions
Ito, R., Robbins, T. W., Pennartz, C. M., & Everitt, B. J. (2008). Functional Interaction between
the Hippocampus and Nucleus Accumbens Shell Is Necessary for the Acquisition of
Appetitive Spatial Context Conditioning. Journal of Neuroscience, 28(27), 6950–6959.
https://doi.org/10.1523/JNEUROSCI.1615-08.2008
Iwata, J., LeDoux, J. E., Meeley, M. P., Arneric, S., & Reis, D. J. (1986). Intrinsic neurons in the
amygdaloid field projected to by the medial geniculate body mediate emotional responses
conditioned to acoustic stimuli. Brain Research, 383(1–2), 195–214.
https://doi.org/10.1016/0006-8993(86)90020-X
Jackson, M. E., & Moghaddam, B. (2001). Amygdala regulation of nucleus accumbens
dopamine output is governed by the prefrontal cortex. Journal of Neuroscience, 21(2), 676–
681. https://doi.org/21/2/676 [pii]
Jean-Richard-Dit-Bressel, P., & McNally, G. P. (2015). The role of the basolateral amygdala in
punishment. Learning & Memory, 22, 128–137.
Jean-Richard-Dit-Bressel, P., & McNally, G. P. (2016). Lateral, not medial, prefrontal cortex
contributes to punishment and aversive instrumental learning. Learning & Memory (Cold
Spring Harbor, N.Y.), 23(11), 607–617. https://doi.org/10.1101/LM.042820.116
Jenni, N. L., Larkin, J. D., & Floresco, S. B. (2017). Prefrontal Dopamine D1 and D2 Receptors
Regulate Dissociable Aspects of Decision Making via Distinct Ventral Striatal and
Amygdalar Circuits. The Journal of Neuroscience, 37(26), 6200–6213.
https://doi.org/10.1523/JNEUROSCI.0030-17.2017
Jensen, J., McIntosh, A. R., Crawley, A. P., Mikulis, D. J., Remington, G., & Kapur, S. (2003).
Direct activation of the ventral striatum in anticipation of aversive stimuli. Neuron, 40(6),

205
1251–1257. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/14687557
Jensen, J., Willeit, M., Zipursky, R., Savina, I., Smith, A. J., Menon, M., … Kapur, S. (2008).
The formation of abnormal associations in schizophrenia: neural and behavioral evidence.
Neuropsychopharmacology, 33(3), 473–479. https://doi.org/10.1038/sj.npp.1301437
Jentsch, J. D., & Taylor, J. R. (1999). Impulsivity resulting from frontostriatal dysfunction in
drug abuse: Implications for the control of behavior by reward-related stimuli.
Psychopharmacology, 146(4), 373–390. https://doi.org/10.1007/PL00005483
Jiang, Z.-C., Pan, Q., Zheng, C., Deng, X.-F., Wang, J.-Y., & Luo, F. (2015). Inactivation of the
prelimbic rather than infralimbic cortex impairs acquisition and expression of formalin-
induced conditioned place avoidance. Neuroscience Letters, 569, 89–93.
https://doi.org/10.1016/j.neulet.2014.03.074
Johansen, J. P., Cain, C. K., Ostroff, L. E., & Ledoux, J. E. (2011). Molecular mechanisms of
fear learning and memory. Cell, 147(3), 509–524. https://doi.org/10.1016/j.cell.2011.10.009
Johansen, J. P., Wolff, S. B. E., Lüthi, A., & Ledoux, J. E. (2012). Controlling the elements: An
optogenetic approach to understanding the neural circuits of fear. Biological Psychiatry,
71(12), 1053–1060. https://doi.org/10.1016/j.biopsych.2011.10.023
Jongen-Rêlo, A. L., Kaufmann, S., & Feldon, J. (2003). A differential involvement of the shell
and core subterritories of the nucleus accumbens of the rats in memory processes.
Behavioral Neuroscience, 117(1), 150–168. https://doi.org/10.1037/0735-7044.117.1.150
Jongen-Rêlo, A. L., Voorn, P., Groenewegen, H. J., Voom, P., & Groenewegen, H. J. (1994).
Immunohistochemical characterization of the shell and core territories of the nucleus
accumbens in the rat. European Journal of Neuroscience, 6(8), 1255–1264.
https://doi.org/10.1111/j.1460-9568.1994.tb00315.x
Jovanovic, T., & Norrholm, S. D. (2011). Neural Mechanisms of Impaired Fear Inhibition in
Posttraumatic Stress Disorder. Frontiers in Behavioral Neuroscience, 5(July), 8.
https://doi.org/10.3389/fnbeh.2011.00044
Kaczkurkin, A. N., Ph, D., Burton, P. C., Ph, D., Chazin, S. M., Manbeck, A. B., & Espensen-
sturges, T. (2017). Neural Substrates of Overgeneralized Conditioned Fear in PTSD.
American Journal of Psychiatry, 1742(2), 125–134.
https://doi.org/10.1176/appi.ajp.2016.15121549
Kamin, L. J., Brimer, C. J., & Black, A. H. (1963). Conditioned suppression as a monitor of fear
of the CS in the course of avoidance training. Journal of Comparative and Phyisological
Psychology, 56(3), 497–501.
Kaufman, J. N., Ross, T. J., Stein, E. A., & Garavan, H. (2003). Cingulate hypoactivity in
cocaine users during a GO-NOGO task as revealed by event-related functional magnetic
resonance imaging. Journal of Neuroscience, 23(21), 7839–43. https://doi.org/23/21/7839
[pii]
Keistler, C., Barker, J. M., & Taylor, J. R. (2015). Infralimbic prefrontal cortex interacts with
nucleus accumbens shell to unmask expression of outcome- selective Pavlovian-to-
instrumental transfer. Learning & Memory, 22, 509–514.

206
https://doi.org/10.1101/lm.038810.115
Kelley, A. E., Baldo, B. a., Pratt, W. E., & Will, M. J. (2005). Corticostriatal-hypothalamic
circuitry and food motivation: Integration of energy, action and reward. Physiology and
Behavior, 86(5), 773–795. https://doi.org/10.1016/j.physbeh.2005.08.066
Killcross, S., Everitt, B. J., & Robbins, T. W. (1997). Symmetrical effects of amphetamine and
alpha-flupenthixol on conditioned punishment and conditioned reinforcement: Contrasts
with midazolam. Psychopharmacology, 129(2), 141–152.
https://doi.org/10.1007/s002130050174
Kim, C. K., Ye, L., Jennings, J. H., Pichamoorthy, N., Tang, D. D., Yoo, A. W., … Deisseroth,
K. (2017). Molecular and circuit-dynamical identification of top-down neural mechanisms
for restraint of reward seeking. Cell, 1–15. https://doi.org/10.1016/j.cell.2017.07.020
Kim, M. J., Gee, D. G., Loucks, R. A., Davis, F. C., & Whalen, P. J. (2011). Anxiety Dissociates
Dorsal and Ventral Medial Prefrontal Cortex Functional Connectivity with the Amygdala at
Rest, (July), 1667–1673. https://doi.org/10.1093/cercor/bhq237
Kita, H., & Kitai, S. T. (1990). Amygdaloid projections to the frontal cortex and the striatum in
the rat. Journal of Comparative Neurology, 298(1), 40–49.
https://doi.org/10.1002/cne.902980104
Klucken, T., Tabbert, K., Schweckendiek, J., Merz, C. J., Kagerer, S., Vaitl, D., & Stark, R.
(2009). Contingency learning in human fear conditioning involves the ventral striatum.
Human Brain Mapping, 30(11), 3636–3644. https://doi.org/10.1002/hbm.20791
Koo, J. W., Han, J.-S., & Kim, J. J. (2004). Selective Neurotoxic Lesions of Basolateral and
Central Nuclei of the Amygdala Produce Differential Effects on Fear Conditioning. Journal
of Neuroscience, 24(35), 7654–7662. https://doi.org/10.1523/JNEUROSCI.1644-04.2004
Kuczenski, R., & Segal, D. (1989). Concomitant characterization of behavioral and striatal
neurotransmitter response to amphetamine using in vivo microdialysis. The Journal of
Neuroscience : The Official Journal of the Society for Neuroscience, 9(6), 2051–2065.
Kulkarni, A. S. (1968). Facilitation of instrumental avoidance learning by amphetamine: An
analysis. Psychopharmacologia, 13(5), 418–425. https://doi.org/10.1007/BF00404957
LaBar, K. S., Gatenby, J. C., Gore, J. C., LeDoux, J. E., & Phelps, E. A. (1998). Human
amygdala activation during conditioned fear acquisition and extinction: a mixed-trial fMRI
study. Neuron, 20(5), 937–945. https://doi.org/10.1016/S0896-6273(00)80475-4
LaLumiere, R. T., Nawar, E. M., & McGaugh, J. L. (2005). Modulation of memory
consolidation by the basolateral amygdala or nucleus accumbens shell requires concurrent
dopamine receptor activation in both brain regions. Learning & Memory, 12(3), 296–301.
https://doi.org/10.1101/lm.93205
Lauzon, N. M., Ahmad, T., & Laviolette, S. R. (2012). Dopamine D4 receptor transmission in
the prefrontal cortex controls the salience of emotional memory via modulation of calcium
calmodulin-dependent kinase II. Cerebral Cortex (New York, N.Y. : 1991), 22(11), 2486–
2494. https://doi.org/10.1093/cercor/bhr326

207
Lauzon, N. M., Bishop, S. F., & Laviolette, S. R. (2009). Dopamine D1 versus D4 receptors
differentially modulate the encoding of salient versus nonsalient emotional information in
the medial prefrontal cortex. Journal of Neuroscience, 29(15), 4836–4845.
https://doi.org/10.1523/JNEUROSCI.0178-09.2009
Laviolette, S. R., Lipski, W. J., & Grace, A. A. (2005). A subpopulation of neurons in the medial
prefrontal cortex encodes emotional learning with burst and frequency codes through a
dopamine D4 receptor-dependent basolateral amygdala input. The Journal of
Neuroscience : The Official Journal of the Society for Neuroscience, 25(26), 6066–6075.
https://doi.org/10.1523/JNEUROSCI.1168-05.2005
Lazareno, S. (1979). d-Amphetamine and punished responding: The role of catecholamines and
anorexia. Psychopharmacology, 66(2), 133–142. https://doi.org/10.1007/BF00427620
Ledoux, J. E. (2014). Coming to terms with fear. Proceedings of the National Academy of
Sciences, 111(8), 2871–2878. https://doi.org/10.1073/pnas.1400335111
LeDoux, J. E. (2012). Rethinking the Emotional Brain. Neuron, 73(4), 653–676.
https://doi.org/10.1016/j.neuron.2012.02.004
LeDoux, J. E., Cicchetti, P., Xagoraris, A., & Romanski, L. M. (1990). The lateral amygdaloid
nucleus: sensory interface of the amygdala in fear conditioning. The Journal of
Neuroscience, 10(4), 1062–1069. https://doi.org/2329367
Lee, B., London, E. D., Poldrack, R. A., Farahi, J., Nacca, A., Monterosso, J. R., … Mandelkern,
M. A. (2009). Striatal dopamine d2/d3 receptor availability is reduced in methamphetamine
dependence and is linked to impulsivity. The Journal of Neuroscience, 29(47), 14734–
14740. https://doi.org/10.1523/JNEUROSCI.3765-09.2009
Lee, J. L. C., Dickinson, A., & Everitt, B. J. (2005). Conditioned suppression and freezing as
measures of aversive Pavlovian conditioning: Effects of discrete amygdala lesions and
overtraining. Behavioural Brain Research, 159(2), 221–233.
https://doi.org/10.1016/j.bbr.2004.11.003
Leone, C. M. L., de Aguiar, J. C., & Graeff, F. G. (1983). Role of 5-hydroxytryptamine in
amphetamine effects on punished and unpunished behaviour. Psychopharmacology, 80(1),
78–82. https://doi.org/10.1007/BF00427500
Levita, L., Dalley, J. W., & Robbins, T. W. (2002). Disruption of Pavlovian contextual
conditioning by excitotoxic lesions of the nucleus accumbens core. Behavioral
Neuroscience, 116(4), 539–552. https://doi.org/10.1037/0735-7044.116.4.539
Levita, L., Hare, T. A., Voss, H. U., Glover, G., Ballon, D. J., & Casey, B. J. (2009). The
bivalent side of the nucleus accumbens. NeuroImage, 44(3), 1178–1187.
https://doi.org/10.1016/j.neuroimage.2008.09.039
Levita, L., Hoskin, R., & Champi, S. (2012). Avoidance of harm and anxiety: A role for the
nucleus accumbens. NeuroImage, 62(1), 189–198.
https://doi.org/10.1016/j.neuroimage.2012.04.059
Li, C. S., Huang, C., Yan, P., Bhagwagar, Z., Milivojevic, V., & Sinha, R. (2008). Neural
correlates of impulse control during stop signal inhibition in cocaine-dependent men.

208
Neuropsychopharmacology, 33(8), 1798–1806. https://doi.org/10.1038/sj.npp.1301568
Lichtenberg, N. T., Kashtelyan, V., Burton, A. C., Bissonette, G. B., & Roesch, M. R. (2014).
Nucleus accumbens core lesions enhance two-way active avoidance. Neuroscience, 258,
340–346. https://doi.org/10.1016/j.neuroscience.2013.11.028
Likhtik, E., & Paz, R. (2015). Amygdala-prefrontal interactions in (mal)adaptive learning.
Trends in Neurosciences, 38(3), 158–166. https://doi.org/10.1016/j.tins.2014.12.007
Limpens, J. H. W., Damsteegt, R., Broekhoven, M. H., Voorn, P., & Vanderschuren, L. J. M. J.
(2015). Pharmacological inactivation of the prelimbic cortex emulates compulsive reward
seeking in rats. Brain Research, 1628, 210–218.
https://doi.org/10.1016/j.brainres.2014.10.045
Limpens, J. H. W., Schut, E. H. S., Voorn, P., & Vanderschuren, L. J. M. J. (2014). Using
conditioned suppression to investigate compulsive drug seeking in rats. Drug and Alcohol
Dependence, 142, 314–324. https://doi.org/10.1016/j.drugalcdep.2014.06.037
Lissek, S., Kaczkurkin, A. N., Rabin, S., Geraci, M., Pine, D. S., & Grillon, C. (2014).
Generalized anxiety disorder is associated with overgeneralization of classically
conditioned fear. Biological Psychiatry, 75(11), 909–915.
https://doi.org/10.1016/j.biopsych.2013.07.025
Lissek, S., Powers, A. S., McClure, E. B., Phelps, E. A., Woldehawariat, G., Grillon, C., & Pine,
D. S. (2005). Classical fear conditioning in the anxiety disorders: A meta-analysis.
Behaviour Research and Therapy, 43(11), 1391–1424.
https://doi.org/10.1016/j.brat.2004.10.007
Lorenzini, C. A., Baldi, E., Bucherelli, C., & Tassoni, G. (1995). Time-dependent deficits of
rat’s memory consolidation induced by tetrodotoxin injections into the caudate-putamen,
nucleus accumbens, and globus pallidus. Neurobiology of Learning and Memory.
https://doi.org/10.1006/nlme.1995.1008
Lubman, D. I., Yücel, M., & Pantelis, C. (2004). Addiction, a condition of compulsive
behaviour? Neuroimaging and neuropsychological evidence of inhibitory dysregulation.
Addiction, 99(12), 1491–1502. https://doi.org/10.1111/j.1360-0443.2004.00808.x
Maia, T. V. (2010). Two-factor theory, the actor-critic model, and conditioned avoidance.
Learning & Behavior, 38(1), 50–67. https://doi.org/10.3758/LB.38.1.50
Maner, J. K., & Schmidt, N. B. (2006). The role of risk avoidance in anxiety. Behavior Therapy,
37(2), 181–189. https://doi.org/10.1016/j.beth.2005.11.003
Maren, S., Aharonov, G., Stote, D. L., & Fanselow, M. S. (1996). N-methyl-D-aspartate
receptors in the basolateral amygdala are required for both acquisition and expression of
conditional fear in rats. Behavioral Neuroscience, 110(6), 1365–1374.
https://doi.org/10.1037/0735-7044.110.6.1365
Maren, S., & Holt, W. G. (2004). Hippocampus and Pavlovian Fear Conditioning in Rats:
Muscimol Infusions Into the Ventral, but Not Dorsal, Hippocampus Impair the Acquisition
of Conditional Freezing to an Auditory Conditional Stimulus. Behavioral Neuroscience,
118(1), 97–110. https://doi.org/10.1037/0735-7044.118.1.97

209
Maren, S., & Quirk, G. J. (2004). Neuronal signalling of fear memory. Nature Reviews
Neuroscience, 5(11), 844–852. https://doi.org/10.1038/nrn1535
Martínez, G., Ropero, C., Funes, A., Flores, E., Landa, A. I., & Gargiulo, P. A. (2002). AP-7 into
the nucleus accumbens disrupts acquisition but does not affect consolidation in a passive
avoidance task. Physiology and Behavior, 76(2), 205–212. https://doi.org/10.1016/S0031-
9384(02)00696-0
Martinez, R. C. R., Gupta, N., Lázaro-Muñoz, G., Sears, R. M., Kim, S., Moscarello, J. M., …
Cain, C. K. (2013). Active vs. reactive threat responding is associated with differential c-
Fos expression in specific regions of amygdala and prefrontal cortex. Learning & Memory,
20(8), 446–452. https://doi.org/10.1101/lm.031047.113
McAllister, K. H. (1997). A single administration of d-amphetamine prior to stimulus pre-
exposure and conditioning attenuates latent inhibition. Psychopharmacology, 130(2), 79–
84. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/9106903
McCullough, L. D., Sokolowski, J. D., & Salamone, J. D. (1993). A neurochemical and
behavioral investigation of the involvement of nucleus accumbens dopamine in instrumental
avoidance. Neuroscience, 52(4), 919–925. https://doi.org/10.1016/0306-4522(93)90538-Q
McDannald, M. A., & Galarce, E. M. (2011). Measuring Pavlovian fear with conditioned
freezing and conditioned suppression reveals different roles for the basolateral amygdala.
Brain Research, 1374, 82–89. https://doi.org/10.1016/j.brainres.2010.12.050
McGinty, V. B., & Grace, A. A. (2008). Selective activation of medial prefrontal-to-accumbens
projection neurons by amygdala stimulation and Pavlovian conditioned stimuli. Cerebral
Cortex (New York, N.Y. : 1991), 18(8), 1961–1972. https://doi.org/10.1093/cercor/bhm223
McGinty, V. B., Lardeux, S., Taha, S. A., Kim, J. J., & Nicola, S. M. (2013). Invigoration of
reward seeking by cue and proximity encoding in the nucleus accumbens. Neuron, 78(5),
910–922. https://doi.org/10.1016/j.neuron.2013.04.010
McHugh, S. B., Marques-Smith, A., Li, J., Rawlins, J. N. P., Lowry, J., Conway, M., …
Bannerman, D. M. (2013). Hemodynamic responses in amygdala and hippocampus
distinguish between aversive and neutral cues during Pavlovian fear conditioning in
behaving rats. The European Journal of Neuroscience, 37(3), 498–507.
https://doi.org/10.1111/ejn.12057
McLaughlin, R. J., & Floresco, S. B. (2007). The role of different subregions of the basolateral
amygdala in cue-induced reinstatement and extinction of food-seeking behavior.
Neuroscience, 146(4), 1484–1494. https://doi.org/10.1016/j.neuroscience.2007.03.025
McNaughton, N. (1982). Gray’s Neuropsychology of anxiety: An enquiry into the functions of
septohippocampal theories. Behavioral and Brain Sciences, 5(3), 492.
https://doi.org/10.1017/S0140525X00013170
Meredith, G. E. (1999). The Synaptic Framework for Chemical Signaling in Nucleus
Accumbens. Annals of the New York Academy of Sciences, 877, 140–156.
Meredith, G. E., Pattiselanno, A., Groenewegen, H. J., & Haber, S. N. (1996). Shell and core in
monkey and human nucleus accumbens identified with antibodies to calbindin-D28k. J

210
Comp Neurol, 365(4), 628–639. https://doi.org/10.1002/(SICI)1096-
9861(19960219)365:4<628::AID-CNE9>3.0.CO;2-6
Meyer, D. R., Cho, C., & Wesemann, A. F. (1960). On problems of conditioning discriminated
lever-press avoidance responses. Psychological Review, 67(4), 224–228.
Mihindou, C., Guillem, K., Navailles, S., Vouillac, C., & Ahmed, S. H. (2013). Discriminative
Inhibitory Control of Cocaine Seeking. Biological Psychiatry, 73(3), 271–279.
https://doi.org/10.1016/j.biopsych.2012.08.011
Milad, M. R., Quinn, B. T., Pitman, R. K., Orr, S. P., Fischl, B., & Rauch, S. L. (2005).
Thickness of ventromedial prefrontal cortex in humans is correlated with extinction
memory. Proceedings of the National Academy of Sciences of the United States of America,
102(30), 10706–10711. https://doi.org/10.1073/pnas.0502441102
Milad, M. R., & Quirk, G. J. (2012). Fear extinction as a model for translational neuroscience:
ten years of progress. Annual Review of Psychology, 63, 129–51.
https://doi.org/10.1146/annurev.psych.121208.131631
Milad, M. R., Quirk, G. J., Pitman, R. K., Orr, S. P., Fischl, B., & Rauch, S. L. (2007). A Role
for the Human Dorsal Anterior Cingulate Cortex in Fear Expression. Biological Psychiatry,
62(10), 1191–1194. https://doi.org/10.1016/j.biopsych.2007.04.032
Milad, M. R., Vidal-Gonzalez, I., & Quirk, G. J. (2004). Electrical stimulation of medial
prefrontal cortex reduces conditioned fear in a temporally specific manner. Behavioral
Neuroscience, 118(2), 389–94. https://doi.org/10.1037/0735-7044.118.2.389
Milad, M. R., Wright, C. I., Orr, S. P., Pitman, R. K., Quirk, G. J., & Rauch, S. L. (2007). Recall
of Fear Extinction in Humans Activates the Ventromedial Prefrontal Cortex and
Hippocampus in Concert. Biological Psychiatry, 62(5), 446–454.
https://doi.org/10.1016/j.biopsych.2006.10.011
Millan, E. Z., Furlong, T. M., & McNally, G. P. (2010). Accumbens shell-hypothalamus
interactions mediate extinction of alcohol seeking. The Journal of Neuroscience, 30(13),
4626–4635. https://doi.org/10.1523/JNEUROSCI.4933-09.2010
Millan, E. Z., & McNally, G. P. (2011). Accumbens shell AMPA receptors mediate expression
of extinguished reward seeking through interactions with basolateral amygdala. Learning &
Memory (Cold Spring Harbor, N.Y.), 18(7), 414–421. https://doi.org/10.1101/lm.2144411
Millan, E. Z., Reese, R. M., Grossman, C. D., Chaudhri, N., & Janak, P. H. (2015). Nucleus
Accumbens and Posterior Amygdala Mediate Cue-Triggered Alcohol Seeking and Suppress
Behavior During the Omission of Alcohol-Predictive Cues. Neuropsychopharmacology,
40(11), 2555–2565. https://doi.org/10.1038/npp.2015.102
Miller, M. A., Thomé, A., & Cowen, S. L. (2013). Intersection of effort and risk: Ethological and
neurobiological perspectives. Frontiers in Neuroscience, 7(7 NOV), 1–11.
https://doi.org/10.3389/fnins.2013.00208
Miller, N. E. (1948). Studies of fear as an acquirable drive: I. Fear as motivation and fear-
reduction as reinforcement in the learning of new responses. Journal of Experimental
Psychology, 38(1), 89–101. https://doi.org/10.1037/h0058455

211
Mishra, A. M., Ellens, D. J., Motelow, J. E., Purcaro, J., Desalvo, M. N., Enev, M., … Blumfeld,
H. (2011). Where fMRI and electrophysiology agree to disagree: corticothalamic and
striatal activity patterns in the WAG/Rij rat. Journal of Neuroscience, 31(42), 15053–
15064. https://doi.org/10.1523/JNEUROSCI.0101-11.2011.Where
Mitchell, M. R., Vokes, C. M., Blankenship, A. L., Simon, N. W., & Setlow, B. (2011). Effects
of acute administration of nicotine, amphetamine, diazepam, morphine, and ethanol on risky
decision-making in rats. Psychopharmacology, 218(4), 703–712.
https://doi.org/10.1007/s00213-011-2363-8
Mogenson, G. J., Jones, D. L., & Yim, C. Y. (1980). From motivation to action: Functional
interface between the limbic system and the motor system. Progress in Neurobiology, 14(2–
3), 69–97. https://doi.org/10.1016/0301-0082(80)90018-0
Mohammadi, M., Bergado-Acosta, J. R., & Fendt, M. (2014). Relief learning is distinguished
from safety learning by the requirement of the nucleus accumbens. Behavioural Brain
Research, 272, 40–45. https://doi.org/10.1016/j.bbr.2014.06.053
Morein-Zamir, S., & Robbins, T. W. (2015). Fronto-striatal circuits in response-inhibition:
Relevance to addiction. Brain Research, 1628, 117–129.
https://doi.org/10.1016/j.brainres.2014.09.012
Morein-Zamir, S., Simon Jones, P., Bullmore, E. T., Robbins, T. W., & Ersche, K. D. (2013).
Prefrontal hypoactivity associated with impaired inhibition in stimulant-dependent
individuals but evidence for hyperactivation in their unaffected siblings.
Neuropsychopharmacology, 38(10), 1945–1953. https://doi.org/10.1038/npp.2013.90
Morgan, M. a, Romanski, L. M., & LeDoux, J. E. (1993). Extinction of emotoinal learning:
contribution of medial prefrontal cortex. Neurosci Lett, 163, 109–113.
Morris, R. G. M. (1975). Preconditioning of reinforcing properties to an exteroceptive feedback
stimulus. Learning and Motivation, 6, 289–298.
Morris, R. G. M. (1989). Synaptic plasticity and learning: selective impairment of learning rats
and blockade of long-term potentiation in vivo by the N-methyl-D-aspartate receptor
antagonist AP5. The Journal of Neuroscience : The Official Journal of the Society for
Neuroscience, 9(9), 3040–3057.
Moscarello, J. M., & Ledoux, J. E. (2014). Diverse effects of conditioned threat stimuli on
behavior. Cold Spring Harbor Symposia on Quantitative Biology, 79, 11–19.
https://doi.org/10.1101/sqb.2014.79.024968
Motzkin, J. C., Philippi, C. L., Wolf, R. C., Baskaya, M. K., & Koenigs, M. (2014).
Ventromedial Prefrontal Cortex Is Critical for the Regulation of Amygdala Activity in
Humans. Biological Psychiatry, 1–9. https://doi.org/10.1016/j.biopsych.2014.02.014
Mowrer, O. H., & Lamoreaux, R. R. (1946). Fear as an intervening variable in avoidance
conditioning. Journal of Comparative Psychology, 39(1), 29–50.
https://doi.org/10.1037/h0060150
Murphy, E. R., Robinson, E. S. J., Theobald, D. E. H., Dalley, J. W., & Robbins, T. W. (2008).
Contrasting effects of selective lesions of nucleus accumbens core or shell on inhibitory

212
control and amphetamine-induced impulsive behaviour. European Journal of Neuroscience,
28(2), 353–363. https://doi.org/10.1111/j.1460-9568.2008.06309.x
Muschamp, J. W., Van’t Veer, A., Parsegian, A., Gallo, M. S., Chen, M., Neve, R. L., …
Carlezon, W. A. (2011). Activation of CREB in the nucleus accumbens shell produces
anhedonia and resistance to extinction of fear in rats. The Journal of Neuroscience : The
Official Journal of the Society for Neuroscience, 31(8), 3095–103.
https://doi.org/10.1523/JNEUROSCI.5973-10.2011
Namburi, P., Beyeler, A., Yorozu, S., Calhoon, G. G., Halbert, S. a., Wichmann, R., … Tye, K.
M. (2015). A circuit mechanism for differentiating positive and negative associations.
Nature, 520(7549), 675–8. https://doi.org/10.1038/nature14366
Nicola, S. M. (2010). The flexible approach hypothesis: unification of effort and cue-responding
hypotheses for the role of nucleus accumbens dopamine in the activation of reward-seeking
behavior. The Journal of Neuroscience, 30(49), 16585–16600.
https://doi.org/10.1523/JNEUROSCI.3958-10.2010
Nicola, S. M., Yun, I. A., Wakabayashi, K. T., & Fields, H. L. (2004). Cue-evoked firing of
nucleus accumbens neurons encodes motivational significance during a discriminative
stimulus task. Journal of Neurophysiology, 91(4), 1840–1865.
https://doi.org/10.1152/jn.00657.2003
Nieh, E. H., Matthews, G. A., Allsop, S. A., Presbrey, K. N., Leppla, C. A., Wichmann, R., …
Tye, K. M. (2015). Decoding neural circuits that control compulsive sucrose seeking. Cell,
160(3), 528–541. https://doi.org/10.1016/j.cell.2015.01.003
Niemegeers, C. J. ., Verbruggen, F. J., & Janssen, P. A. . (1970). The influence of various
neuroleptic drugs on shock avoidance responding in rats. III. Amphetamine antagonism in
the discriminated Sidman avoidance procedure. Psychopharmacologia, 17, 151–159.
Nunes, E. J., Randall, P. A., Podurgiel, S., Correa, M., & Salamone, J. D. (2013). Nucleus
accumbens neurotransmission and effort-related choice behavior in food motivation: Effects
of drugs acting on dopamine, adenosine, and muscarinic acetylcholine receptors.
Neuroscience and Biobehavioral Reviews, 37(9), 2015–2025.
https://doi.org/10.1016/j.neubiorev.2013.04.002
O’Donnell, P., & Grace, A. A. (1995). Synaptic interactions among excitatory afferents to
nucleus accumbens neurons: hippocampal gating of prefrontal cortical input. The Journal of
Neuroscience, 15(5), 3622–3639.
O’Donnell, P., Greene, J., Pabello, N., Lewis, B. L., & Grace, A. A. (1999). Modulation of cell
firing in the nucleus accumbens. Annals of the New York Academy of Sciences, 877, 157–
175. https://doi.org/10.1111/j.1749-6632.1999.tb09267.x
Ohaeri, J. U. (2003). The burden of caregiving in families with a mental illness: a review of
2002. Current Opinion in Psychiatry, 16(4), 457–465.
https://doi.org/10.1097/01.yco.0000079212.36371.c0
Oleson, E. B., Gentry, R. N., Chioma, V. C., & Cheer, J. F. (2012). Subsecond dopamine release
in the nucleus accumbens predicts conditioned punishment and its successful avoidance.

213
The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 32(42),
14804–14808. https://doi.org/10.1523/JNEUROSCI.3087-12.2012
Orona, E., & Gabriel, M. (1983). Multiple-unit activity of the prefrontal cortex and mediodorsal
thalamic nucleus during acquisition of discriminative avoidance behavior in rabbits. Brain
Research, 263(2), 295–312. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/6839177
Orsini, C. A., & Maren, S. (2012). Neural and cellular mechanisms of fear and extinction
memory formation. Neuroscience and Biobehavioral Reviews, 36(7), 1773–1802.
https://doi.org/10.1016/j.neubiorev.2011.12.014
Orsini, C. A., Moorman, D. E., Young, J. W., Setlow, B., & Floresco, S. B. (2015). Neural
mechanisms regulating different forms of risk-related decision-making: Insights from
animal models. Neuroscience and Biobehavioral Reviews, 58, 147–167.
https://doi.org/10.1016/j.neubiorev.2015.04.009
Orsini, C. A., Trotta, R. T., Bizon, J. L., & Setlow, B. (2015). Dissociable Roles for the
Basolateral Amygdala and Orbitofrontal Cortex in Decision-Making under Risk of
Punishment. Journal of Neuroscience, 35(4), 1368–1379.
https://doi.org/10.1523/JNEUROSCI.3586-14.2015
Ottenbreit, N. D., & Dobson, K. S. (2004). Avoidance and depression: The construction of the
Cognitive-Behavioral Avoidance Scale. Behaviour Research and Therapy, 42(3), 293–313.
https://doi.org/10.1016/S0005-7967(03)00140-2
Pang, M. H., Kim, N. S., Kim, I. H., Kim, H., Kim, H. T., & Choi, J. S. (2010). Cholinergic
transmission in the dorsal hippocampus modulates trace but not delay fear conditioning.
Neurobiology of Learning and Memory, 94(2), 206–213.
https://doi.org/10.1016/j.nlm.2010.05.008
Panksepp, J. (2011). Cross-Species affective neuroscience decoding of the primal affective
experiences of humans and related animals. PLoS ONE, 6(9).
https://doi.org/10.1371/journal.pone.0021236
Parkinson, J. A., Cardinal, R. N., & Everitt, B. J. (2000). Limbic cortical-ventral striatal systems
underlying appetitive conditioning. Progress in Brain Research, 126, 263–285.
https://doi.org/10.1016/S0079-6123(00)26019-6
Parkinson, J. A., Robbins, T. W., & Everitt, B. J. (1999). Selective excitotoxic lesions of the
nucleus accumbens core and shell differentially affect aversive Pavlovian conditioning to
discrete and contextual cues, 27(2), 256–266.
Parkinson, J. A., Willoughby, P. J., Robbins, T. W., & Everitt, B. J. (2000). Disconnection of the
anterior cingulate cortex and nucleus accumbens core impairs Pavlovian approach behavior:
Further evidence for limbic cortical-ventral striatopallidal systems. Behavioral
Neuroscience, 114(1), 42–63. https://doi.org/10.1037//0735-7044.114.1.42
Pascoli, V., Terrier, J., Hiver, A., & Lüscher, C. (2015). Sufficiency of Mesolimbic Dopamine
Neuron Stimulation for the Progression to Addiction. Neuron, 88(5), 1054–1066.
https://doi.org/10.1016/j.neuron.2015.10.017
Pattij, T., Janssen, M. C. W., Vanderschuren, L. J. M. J., Schoffelmeer, A. N. M., & Van Gaalen,

214
M. M. (2007). Involvement of dopamine D1 and D2 receptors in the nucleus accumbens
core and shell in inhibitory response control. Psychopharmacology, 191(3), 587–598.
https://doi.org/10.1007/s00213-006-0533-x
Pavlov, I. P. (1926). Conditioned reflexes.
Pearson, J. M., Watson, K. K., & Platt, M. L. (2014). Decision making: The neuroethological
turn. Neuron, 82(5), 950–965. https://doi.org/10.1016/j.neuron.2014.04.037
Pellman, B. A., & Kim, J. J. (2016). What Can Ethobehavioral Studies Tell Us about the Brain’s
Fear System? Trends in Neurosciences. Elsevier Ltd.
https://doi.org/10.1016/j.tins.2016.04.001
Pelloux, Y., Murray, J. E., & Everitt, B. J. (2013). Differential roles of the prefrontal cortical
subregions and basolateral amygdala in compulsive cocaine seeking and relapse after
voluntary abstinence in rats. European Journal of Neuroscience, 38, 3018–3026.
https://doi.org/10.1111/ejn.12289
Pendyam, S., Bravo-Rivera, C., Burgos-Robles, A., Sotres-Bayon, F., Quirk, G. J., & Nair, S. S.
(2013). Fear signaling in the prelimbic-amygdala circuit: a computational modeling and
recording study. Journal of Neurophysiology, 110(4), 844–61.
https://doi.org/10.1152/jn.00961.2012
Pennartz, C. M. A., Groenewegen, H. J., & Lopes Da Silva, F. H. (1994). The nucleus
accumbens as a complex of functionally distinct neuronal ensembles: An integration of
behavioural, electrophysiological and anatomical data. Progress in Neurobiology, 42(6),
719–761. https://doi.org/10.1016/0301-0082(94)90025-6
Perry, J. L., & Carroll, M. E. (2008). The role of impulsive behavior in drug abuse.
Psychopharmacology, 200(1), 1–26. https://doi.org/10.1007/s00213-008-1173-0
Peters, J., Kalivas, P. W., & Quirk, G. J. (2009). Extinction circuits for fear and addiction
overlap in prefrontal cortex. Learning & Memory (Cold Spring Harbor, N.Y.), 16(5), 279–
288. https://doi.org/10.1101/lm.1041309
Peters, J., LaLumiere, R. T., & Kalivas, P. W. (2008). Infralimbic prefrontal cortex is responsible
for inhibiting cocaine seeking in extinguished rats. The Journal of Neuroscience, 28(23),
6046–53. https://doi.org/10.1523/JNEUROSCI.1045-08.2008
Phelps, E. A., Delgado, M. R., Nearing, K. I., & Ledoux, J. E. (2004). Extinction learning in
humans: Role of the amygdala and vmPFC. Neuron, 43(6), 897–905.
https://doi.org/10.1016/j.neuron.2004.08.042
Phillipson, O. T., & Griffiths, A. C. (1985). The topographic order of inputs to nucleus
accumbens in the rat. Neuroscience, 16(2), 275–296. https://doi.org/10.1016/0306-
4522(85)90002-8
Piantadosi, P. T., & Floresco, S. B. (2014). Prefrontal cortical GABA transmission modulates
discrimination and latent inhibition of conditioned fear: Relevance for schizophrenia.
Neuropsychopharmacology : Official Publication of the American College of
Neuropsychopharmacology, 39(10), 2473–2484. https://doi.org/10.1038/npp.2014.99

215
Piantadosi, P. T., Yeates, D. C. M. M., Wilkins, M., & Floresco, S. B. (2017). Contributions of
basolateral amygdala and nucleus accumbens subregions to mediating motivational conflict
during punished reward-seeking. Neurobiology of Learning and Memory, 140, 92–105.
https://doi.org/10.1016/j.nlm.2017.02.017
Pohlack, S. T., Nees, F., Ruttorf, M., Schad, L. R., & Flor, H. (2012). Activation of the ventral
striatum during aversive contextual conditioning in humans. Biological Psychology, 91(1),
74–80. https://doi.org/10.1016/j.biopsycho.2012.04.004
Pothuizen, H. H. J., Jongen-Rêlo, A. L., Feldon, J., & Yee, B. K. (2005). Double dissociation of
the effects of selective nucleus accumbens core and shell lesions on impulsive-choice
behaviour and salience learning in rats. European Journal of Neuroscience, 22(10), 2605–
2616. https://doi.org/10.1111/j.1460-9568.2005.04388.x
Poulos, A. M., Ponnusamy, R., Dong, H., & Fanselow, M. S. (2010). Compensation in the neural
circuitry of fear conditioning awakens learning circuits in the bed nuclei of the stria
terminalis, 107(33), 14881–14886. https://doi.org/10.1073/pnas.1005754107
Quirk, G. J., Russo, G. K., Barron, J. L., & Lebron, K. (2000). The role of ventromedial
prefrontal cortex in the recovery of extinguished fear. Journal of Neuroscience, 20(16),
6225–6231. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/10934272
R Core Team. (2017). R: A Language and Environment for Statistical Computing. Vienna,
Austria. Retrieved from https://www.r-project.org/
Radke, A. K., Jury, N. J., Kocharian, A., Marcinkiewcz, C. A., Lowery-Gionta, E. G., Pleil, K.
E., … Holmes, A. (2015). Chronic EtOH effects on putative measures of compulsive
behavior in mice. Addiction Biology, (July 2016). https://doi.org/10.1111/adb.12342
Radke, A. K., Nakazawa, K., & Holmes, A. (2015). Cortical GluN2B deletion attenuates
punished suppression of food reward-seeking. Psychopharmacology, 232(20), 3753–3761.
https://doi.org/10.1007/s00213-015-4033-8
Ramirez, F., Moscarello, J. M., LeDoux, J. E., & Sears, R. M. (2015). Active avoidance requires
a serial basal amygdala to nucleus accumbens shell circuit. Journal of Neuroscience, 35(8),
3470–3477. https://doi.org/10.1523/JNEUROSCI.1331-14.2015
Raybuck, J. D., & Lattal, K. M. (2014). Bridging the interval: Theory and neurobiology of trace
conditioning. Behavioural Processes, 101, 103–111.
https://doi.org/10.1016/j.beproc.2013.08.016
Resstel, L. B. M., Souza, R. F., & Guimarães, F. S. (2008). Anxiolytic-like effects induced by
medial prefrontal cortex inhibition in rats submitted to the Vogel conflict test. Physiology
and Behavior, 93(1–2), 200–205. https://doi.org/10.1016/j.physbeh.2007.08.009
Reynolds, S. M., & Berridge, K. C. (2002). Positive and negative motivation in nucleus
accumbens shell: bivalent rostrocaudal gradients for GABA-elicited eating, taste
“liking”/“disliking” reactions, place preference/avoidance, and fear. The Journal of
Neuroscience : The Official Journal of the Society for Neuroscience, 22(16), 7308–7320.
https://doi.org/20026734
Reynolds, S. M., & Zahm, D. S. (2005). Specificity in the projections of prefrontal and insular

216
cortex to ventral striatopallidum and the extended amygdala. The Journal of Neuroscience,
25(50), 11757–11767. https://doi.org/10.1523/JNEUROSCI.3432-05.2005
Richard, J. M., & Berridge, K. C. (2013). Prefrontal cortex modulates desire and dread generated
by nucleus accumbens glutamate disruption. Biological Psychiatry, 73(4), 360–370.
https://doi.org/10.1016/j.biopsych.2012.08.009
Riedel, G., Harrington, N. R., Hall, G., & Macphail, E. M. (1997). Nucleus accumbens lesions
impair context, but not cue, conditioning in rats. Neuroreport, 8(11), 2477–2481.
https://doi.org/10.1097/00001756-199707280-00013
Robbins, T. W. (2002). The 5-choice serial reaction time task: behavioural pharmacology and
functional neurochemistry. Psychopharmacology, 163(3–4), 362–380.
https://doi.org/10.1007/s00213-002-1154-7
Rodriguez-Romaguera, J., Monte, F. H. M. Do, & Quirk, G. J. (2012). Deep brain stimulation of
the ventral striatum enhances extinction of conditioned fear, 2012.
https://doi.org/10.1073/pnas.1200782109/-
/DCSupplemental.www.pnas.org/cgi/doi/10.1073/pnas.1200782109
Roitman, J. D., & Loriaux, A. L. (2014). Nucleus accumbens responses differentiate execution
and restraint in reward-directed behavior. Journal of Neurophysiology, 111(2), 350–60.
https://doi.org/10.1152/jn.00350.2013
Roitman, M. F., Wheeler, R. A., & Carelli, R. M. (2005). Nucleus accumbens neurons are
innately tuned for rewarding and aversive taste stimuli, encode their predictors, and are
linked to motor output. Neuron, 45(4), 587–597.
https://doi.org/10.1016/j.neuron.2004.12.055
Roitman, M. F., Wheeler, R. A., Wightman, R. M., & Carelli, R. M. (2008). Real-time chemical
responses in the nucleus accumbens differentiate rewarding and aversive stimuli. Nature
Neuroscience, 11(12), 1376–1377. https://doi.org/10.1038/nn.2219
Romaniuk, L., Honey, G. D., King, J. R. L., Whalley, H. C., McIntosh, A. M., Levita, L., …
Hall, J. (2010). Midbrain activation during Pavlovian conditioning and delusional
symptoms in schizophrenia. Archives of General Psychiatry, 67(12), 1246–1254.
https://doi.org/10.1001/archgenpsychiatry.2010.169
Rosen, A. J., & La Flore, J. E. (1973). Effects of intraperitoneal and intraventricular d-
amphetamine administration on active avoidance performance in the rat. Life Sciences,
13(11), 1573–1580.
Roth, B. L. (2016). Primer DREADDs for Neuroscientists. Neuron, 89(4), 683–694.
https://doi.org/10.1016/j.neuron.2016.01.040
Russo, S. J., Dietz, D. M., Dumitriu, D., Morrison, J. H., Malenka, R. C., & Nestler, E. J. (2010).
The addicted synapse: Mechanisms of synaptic and structural plasticity in nucleus
accumbens. Trends in Neurosciences, 33(6), 267–276.
https://doi.org/10.1016/j.tins.2010.02.002
Saga, Y., Richard, A., Sgambato-Faure, V., Hoshi, E., Tobler, P. N., & Tremblay, L. (2017).
Ventral pallidum encodes contextual information and controls aversive behaviors. Cerebral

217
Cortex (New York, N.Y. : 1991), 27(4), 2528–2543. https://doi.org/10.1093/cercor/bhw107
Salamone, J. D. (1994). The involvement of nucleus accumbens dopamine in appetitive and
aversive motivation. Behavioural Brain Research, 61(2), 117–133.
https://doi.org/10.1016/0166-4328(94)90153-8
Salamone, J. D., Correa, M., Farrar, A., & Mingote, S. M. (2007). Effort-related functions of
nucleus accumbens dopamine and associated forebrain circuits. Psychopharmacology,
191(3), 461–482. https://doi.org/10.1007/s00213-006-0668-9
Sangha, S., Chadick, J. Z., & Janak, P. H. (2013). Safety encoding in the basal amygdala. The
Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 33(9),
3744–51. https://doi.org/10.1523/JNEUROSCI.3302-12.2013
Sangha, S., Robinson, P. D., Greba, Q., Davies, D. a, & Howland, J. G. (2014). Alterations in
reward, fear and safety cue discrimination after inactivation of the rat prelimbic and
infralimbic cortices. Neuropsychopharmacology : Official Publication of the American
College of Neuropsychopharmacology, 39(10), 2405–13.
https://doi.org/10.1038/npp.2014.89
Santini, E., Quirk, G. J., & Porter, J. T. (2008). Fear conditioning and extinction differentially
modify the intrinsic excitability of infralimbic neurons. Journal of Neuroscience, 28(15),
4028–4036. https://doi.org/10.1523/JNEUROSCI.2623-07.2008
Saunders, B. T., & Robinson, T. E. (2012). The role of dopamine in the accumbens core in the
expression of Pavlovian-conditioned responses. The European Journal of Neuroscience,
36(4), 2521–2532. https://doi.org/10.1111/j.1460-9568.2012.08217.x
Schiller, D., Levy, I., Niv, Y., LeDoux, J. E., & Phelps, E. a. (2008). From fear to safety and
back: reversal of fear in the human brain. The Journal of Neuroscience : The Official
Journal of the Society for Neuroscience, 28(45), 11517–25.
https://doi.org/10.1523/JNEUROSCI.2265-08.2008
Schoenbaum, G., & Setlow, B. (2003). Lesions of nucleus accumbens disrupt learning about
aversive outcomes. The Journal of Neuroscience : The Official Journal of the Society for
Neuroscience, 23(30), 9833–9841. https://doi.org/23/30/9833 [pii]
Schulz, A. L., Woldeit, M. L., Gonçalves, A. I., Saldeitis, K., & Ohl, F. W. (2015). Selective
increase of auditory cortico-striatal coherence during auditory-cued Go/NoGo
discrimination learning. Frontiers in Behavioral Neuroscience, 9(368).
https://doi.org/10.3389/fnbeh.2015.00368
Schwienbacher, I., Fendt, M., Richardson, R., & Schnitzler, H. U. (2004). Temporary
inactivation of the nucleus accumbens disrupts acquisition and expression of fear-
potentiated startle in rats. Brain Research, 1027, 87–93.
https://doi.org/10.1016/j.brainres.2004.08.037
Seiden, L. S., Sabol, K. E., & Ricaurte, G. A. (1993). Amphetamine: effects on catecholamine
systems and behavior. Annual Review of Pharmacology and Toxicology, 33, 639–677.
https://doi.org/10.1146/annurev.pa.33.040193.003231
Seif, T., Chang, S.-J., Simms, J. a, Gibb, S. L., Dadgar, J., Chen, B. T., … Hopf, F. W. (2013).

218
Cortical activation of accumbens hyperpolarization-active NMDARs mediates aversion-
resistant alcohol intake. Nature Neuroscience, 16(8), 1094–100.
https://doi.org/10.1038/nn.3445
Seligman, M., & Beagley, G. (1975). Learned helplessness in the rat. Journal of Comparative
and Physiological Psychology, 88(2), 534–541. https://doi.org/10.1037/h0076430
Sesack, S. R., Deutch, a. Y., Roth, R. H., & Bunney, B. S. (1989). Topographical organization
of the efferent projections of the medial prefrontal cortex in the rat: An anterograde tract-
tracing study with Phaseolus vulgaris leucoagglutinin. Journal of Comparative Neurology,
290(2), 213–242. https://doi.org/10.1002/cne.902900205
Sesack, S. R., & Grace, A. A. (2010). Cortico-Basal Ganglia reward network: microcircuitry.
Neuropsychopharmacology : Official Publication of the American College of
Neuropsychopharmacology, 35(1), 27–47. https://doi.org/10.1038/npp.2009.93
Setlow, B., Roozendaal, B., & McGaugh, J. L. (2000). Involvement of a basolateral amygdala
complex-nucleus accumbens pathway in glucocorticoid-induced modulation of memory
consolidation. European Journal of Neuroscience, 12(1), 367–375.
https://doi.org/10.1046/j.1460-9568.2000.00911.x
Setlow, B., Schoenbaum, G., & Gallagher, M. (2003). Neural encoding in ventral striatum during
olfactory discrimination learning. Neuron, 38(4), 625–636. https://doi.org/10.1016/S0896-
6273(03)00264-2
Shiflett, M. W., & Balleine, B. W. (2010). At the limbic-motor interface: Disconnection of
basolateral amygdala from nucleus accumbens core and shell reveals dissociable
components of incentive motivation. European Journal of Neuroscience, 32(10), 1735–
1743. https://doi.org/10.1111/j.1460-9568.2010.07439.x
Shinonaga, Y., Takada, M., & Mizuno, N. (1994). Topographic organization of collateral
projections from the basolateral amygdaloid nucleus to both the prefrontal cortex and
nucleus accumbens in the rat. Neuroscience, 58(2), 389–397. https://doi.org/10.1016/0306-
4522(94)90045-0
Shirayama, Y., Ishima, T., Oda, Y., Okamura, N., Iyo, M., & Hashimoto, K. (2015). Opposite
roles for neuropeptide S in the nucleus accumbens and bed nucleus of the stria terminalis in
learned helplessness rats. Behavioural Brain Research, 291, 67–71.
https://doi.org/10.1016/j.bbr.2015.05.007
Sierra-Mercado, D., Padilla-Coreano, N., & Quirk, G. J. (2011). Dissociable roles of prelimbic
and infralimbic cortices, ventral hippocampus, and basolateral amygdala in the expression
and extinction of conditioned fear. Neuropsychopharmacology, 36(2), 529–538.
https://doi.org/10.1038/npp.2010.184
Simon, N. W., Gilbert, R. J., Mayse, J. D., Bizon, J. L., & Setlow, B. (2009). Balancing risk and
reward: a rat model of risky decision making. Neuropsychopharmacology : Official
Publication of the American College of Neuropsychopharmacology, 34(10), 2208–2217.
https://doi.org/10.1038/npp.2009.48
Simon, N. W., Montgomery, K. S., Beas, B. S., Mitchell, M. R., Lasarge, C. L., Mendez, I. A.,

219
… Setlow, B. (2011). Dopaminergic Modulation of Risky Decision-Making, 31(48),
17460–17470. https://doi.org/10.1523/JNEUROSCI.3772-11.2011
Singh, T., McDannald, M. A., Takahashi, Y. K., Haney, R. Z., Cooch, N. K., Lucantonio, F., &
Schoenbaum, G. (2011). The role of the nucleus accumbens in knowing when to respond.
Learning & Memory (Cold Spring Harbor, N.Y.), 18(2), 85–87.
https://doi.org/10.1101/lm.2008111
Sitte, H. H., & Freissmuth, M. (2015). Amphetamines, new psychoactive drugs and the
monoamine transporter cycle. Trends in Pharmacological Sciences, 36(1), 41–50.
https://doi.org/10.1016/j.tips.2014.11.006
Skinner, B. F. (1938). The Behavior of Organisms: An experimental analysis. The Psychological
Record. https://doi.org/10.1037/h0052216
Smith-Roe, S. L., Sadeghian, K., & Kelley, a E. (1999). Spatial learning and performance in the
radial arm maze is impaired after N-methyl-D-aspartate (NMDA) receptor blockade in
striatal subregions. Behavioral Neuroscience, 113(4), 703–717.
https://doi.org/10.1037/0735-7044.113.4.703
Smith, K. S., Tindell, A. J., Aldridge, J. W., & Berridge, K. C. (2009). Ventral pallidum roles in
reward and motivation. Behavioural Brain Research, 196(2), 155–167.
https://doi.org/10.1016/j.bbr.2008.09.038.Ventral
Soares-Cunha, C., Coimbra, B., Sousa, N., & Rodrigues, A. J. (2016). Reappraising striatal D1-
and D2-neurons in reward and aversion. Neuroscience and Biobehavioral Reviews, 68, 370–
386. https://doi.org/10.1016/j.neubiorev.2016.05.021
Sokolowski, J. D., McCullough, L. D., & Salamone, J. D. (1994). Effects of dopamine depletions
in the medial prefrontal cortex on active avoidance and escape in the rat. Brain Research,
651(1–2), 293–299. https://doi.org/10.1016/0006-8993(94)90709-9
Solomon, R. L., & Wynne, L. C. (1953). Traumatic avoidance learning: Acquisition in normal
dogs. Psychological Monographs: General and Applied, 67(4), 1–19.
https://doi.org/10.1037/h0093649
Sotres-Bayon, F., & Quirk, G. J. (2010). Prefrontal control of fear: more than just extinction.
Current Opinion in Neurobiology, 20(2), 231–235.
https://doi.org/10.1016/j.conb.2010.02.005
St. Onge, J. R., & Floresco, S. B. (2009). Dopaminergic modulation of risk-based decision
making. Neuropsychopharmacology, 34(3), 681–697. https://doi.org/10.1038/npp.2008.121
St. Onge, J. R., & Floresco, S. B. (2010). Prefrontal cortical contribution to risk-based decision
making. Cerebral Cortex, 20(8), 1816–1828. https://doi.org/10.1093/cercor/bhp250
St Onge, J. R., Stopper, C. M., Zahm, D. S., & Floresco, S. B. (2012). Separate Prefrontal-
Subcortical Circuits Mediate Different Components of Risk-Based Decision Making.
Journal of Neuroscience, 32(8), 2886–2899. https://doi.org/10.1523/JNEUROSCI.5625-
11.2012
Stark, H., Rothe, T., Wagner, T., & Scheich, H. (2004). Learning a new behavioral strategy in

220
the shuttle-box increases prefrontal dopamine. Neuroscience, 126(1), 21–29.
https://doi.org/10.1016/j.neuroscience.2004.02.026
Stopper, C. M., & Floresco, S. B. (2011). Contributions of the nucleus accumbens and its
subregions to different aspects of risk-based decision making. Cognitive, Affective &
Behavioral Neuroscience, 11(1), 97–112. https://doi.org/10.3758/s13415-010-0015-9
Stratford, T. R., & Kelley, A. E. (1997). GABA in the nucleus accumbens shell participates in
the central regulation of feeding behavior. The Journal of Neuroscience, 17(11), 4434–
4440.
Stuber, G. D., Britt, J. P., & Bonci, A. (2012). Optogenetic modulation of neural circuits that
underlie reward seeking. Biological Psychiatry, 71(12), 1061–1067.
https://doi.org/10.1016/j.biopsych.2011.11.010.Optogenetic
Sturm, V., Lenartz, D., Koulousakis, A., Treuer, H., Herholz, K., Klein, J. C., & Klosterkötter, J.
(2003). The nucleus accumbens: A target for deep brain stimulation in obsessive-
compulsive- and anxiety-disorders. Journal of Chemical Neuroanatomy, 26(4), 293–299.
https://doi.org/10.1016/j.jchemneu.2003.09.003
Sulzer, D., Sonders, M. S., Poulsen, N. W., & Galli, A. (2005). Mechanisms of neurotransmitter
release by amphetamines: A review. Progress in Neurobiology, 75(6), 406–433.
https://doi.org/10.1016/j.pneurobio.2005.04.003
Thomas, K. L., Hall, J., & Everitt, B. J. (2002). Cellular imaging with zif268 expression in the
rat nucleus accumbens and frontal cortex further dissociates the neural pathways activated
following the retrieval of contextual and cued fear memory. European Journal of
Neuroscience, 16(9), 1789–1796. https://doi.org/10.1046/j.1460-9568.2002.02247.x
Tian, S., Huang, F., Gao, J., Li, P., Ouyang, X., Zhou, S., … Yan, Y. (2011). Ventrolateral
prefrontal cortex is required for fear extinction in a modified delay conditioning paradigm
in rats, 189, 258–268. https://doi.org/10.1016/j.neuroscience.2011.05.002
Trew, J. L. (2011). Exploring the roles of approach and avoidance in depression: An integrative
model. Clinical Psychology Review, 31(7), 1156–1168.
https://doi.org/10.1016/j.cpr.2011.07.007
Van Dongen, Y. C. C., Deniau, J.-M. M., Pennartz, C. M. a. M. A., Galis-De Graaf, Y., Voorn,
P., Thierry, a.-M. M., & Groenewegen, H. J. J. (2005). Anatomical evidence for direct
connections between the shell and core subregions of the rat nucleus accumbens.
Neuroscience, 136(4), 1049–1071. https://doi.org/10.1016/j.neuroscience.2005.08.050
Vanderschuren, L. J. M. J., & Everitt, B. J. (2004). Drug seeking becomes compulsive after
prolonged cocaine self-administration. Science (New York, N.Y.), 305(5686), 1017–9.
https://doi.org/10.1126/science.1098975
Vento, P. J., Burnham, N. W., Rowley, C. S., & Jhou, T. C. (2017). Learning From One’s
Mistakes: A Dual Role for the Rostromedial Tegmental Nucleus in the Encoding and
Expression of Punished Reward Seeking. Biological Psychiatry, 81(12), 1041–1049.
https://doi.org/10.1016/j.biopsych.2016.10.018
Vertes, R. P. (2004). Differential projections of the infralimbic and prelimbic cortex in the rat.

221
Neurobiology of Learning and Memory, 51(1), 32–58. https://doi.org/10.1002/syn.10279
Vidal-Gonzalez, I., Vidal-Gonzalez, B., Rauch, S. L., & Quirk, G. J. (2006). Microstimulation
reveals opposing influences of prelimbic and infralimbic cortex on the expression of
conditioned fear. Learning & Memory, 13(6), 728–733. https://doi.org/10.1101/lm.306106
Vogel, J. R., Beer, B., & Clody, D. E. (1971). A simple and reliable conflict procedure for testing
anti-anxiety agents. Psychopharmacologia, 21(1), 1–7. https://doi.org/10.1007/BF00403989
Volkow, N. D., Fowler, J. S., Wang, G. J., Baler, R., & Telang, F. (2009). Imaging dopamine’s
role in drug abuse and addiction. Neuropharmacology, 56(SUPPL. 1), 3–8.
https://doi.org/10.1016/j.neuropharm.2008.05.022
Volkow, N. D., & Morales, M. (2015). The Brain on Drugs: From Reward to Addiction. Cell,
162(4), 712–725. https://doi.org/10.1016/j.cell.2015.07.046
Volkow, N. D., Wang, G.-J., Fowler, J. S., Tomasi, D., & Telang, F. (2011). Addiction: Beyond
dopamine reward circuitry. Proceedings of the National Academy of Sciences of the United
States of America, 108(37), 15037–15042. https://doi.org/10.1073/pnas.1010654108
Wadenberg, M. L., Ericson, E., Magnusson, O., & Ahlenius, S. (1990). Suppression of
conditioned avoidance behavior by the local application of (-)sulpiride into the ventral, but
not the dorsal, striatum of the rat. Biological Psychiatry, 28(4), 297–307.
https://doi.org/10.1016/0006-3223(90)90657-N
Wendler, E., Gaspar, J. C. C., Ferreira, T. L., Barbiero, J. K., Andreatini, R., Vital, M. A. B. F.,
… Da Cunha, C. (2013). The roles of the nucleus accumbens core, dorsomedial striatum,
and dorsolateral striatum in learning: Performance and extinction of Pavlovian fear-
conditioned responses and instrumental avoidance responses. Neurobiology of Learning and
Memory, 109, 27–36. https://doi.org/10.1016/j.nlm.2013.11.009
Wetzel, W., Ohl, F. W., & Scheich, H. (2008). Global versus local processing of frequency-
modulated tones in gerbils: an animal model of lateralized auditory cortex functions. Proc
Natl Acad Sci USA, 105(18), 6753–6758. https://doi.org/10.1073/pnas.0707844105
Whishaw, I. Q., & Dringenberg, H. C. (1991). How does the rat (Rattus norvegicus) adjust food-
carrying responses to the influences of distance, effort, predatory odor, food size, and food
availability? Psychobiology, 19(3), 251–261. https://doi.org/10.3758/BF03332076
Whiteford, H. A., Degenhardt, L., Rehm, J., Baxter, A. J., Ferrari, A. J., Erskine, H. E., … Vos,
T. (2013). Global burden of disease attributable to mental and substance use disorders:
Findings from the Global Burden of Disease Study 2010. The Lancet, 382(9904), 1575–
1586. https://doi.org/10.1016/S0140-6736(13)61611-6
Whiteford, H. A., Ferrari, A. J., Degenhardt, L., Feigin, V., & Vos, T. (2015). The global burden
of mental, neurological and substance use disorders: An analysis from the global burden of
disease study 2010. PLoS ONE, 10(2), 1–14. https://doi.org/10.1371/journal.pone.0116820
Wietzikoski, E. C., Boschen, S. L., Miyoshi, E., Bortolanza, M., Dos Santos, L. M., Frank, M.,
… Da Cunha, C. (2012). Roles of D1-like dopamine receptors in the nucleus accumbens
and dorsolateral striatum in conditioned avoidance responses. Psychopharmacology, 219(1),
159–169. https://doi.org/10.1007/s00213-011-2384-3

222
Wilensky, A. E., Schafe, G. E., & LeDoux, J. E. (1999). Functional inactivation of the amygdala
before but not after auditory fear conditioning prevents memory formation. The Journal of
Neuroscience, 19(24), RC48.
Wirtshafter, D., & Stratford, T. R. (2010). Evidence for motivational effects elicited by
activation of GABA-A or dopamine receptors in the nucleus accumbens shell.
Pharmacology Biochemistry and Behavior, 96(3), 342–346.
https://doi.org/10.1016/j.pbb.2010.06.004
Wiskerke, J., Schetters, D., van Es, I. E., van Mourik, Y., den Hollander, B. R. O., Schoffelmeer,
A. N. M., & Pattij, T. (2011). μ-Opioid receptors in the nucleus accumbens shell region
mediate the effects of amphetamine on inhibitory control but not impulsive choice. The
Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 31(1), 262–
272. https://doi.org/10.1523/JNEUROSCI.4794-10.2011
Wood, J., & Ahmari, S. (2015). A framework for understanding the emerging role of
corticolimbic-ventral striatal networks in OCD-associated repetitive behaviors. Frontiers in
Systems Neuroscience, 9(December), 1–22. https://doi.org/10.3389/fnsys.2015.00171
Wright, C. I., Beijer, A. V, & Groenewegen, H. J. (1996). Basal amygdaloid complex afferents to
the rat nucleus accumbens are compartmentally organized. The Journal of Neuroscience,
16(5), 1877–1893.
Yiu, A. P., Mercaldo, V., Yan, C., Richards, B., Rashid, A. J., Hsiang, H. L. L., … Josselyn, S.
A. (2014). Neurons Are Recruited to a Memory Trace Based on Relative Neuronal
Excitability Immediately before Training. Neuron, 83(3), 722–735.
https://doi.org/10.1016/j.neuron.2014.07.017
Yun, I. A., Wakabayashi, K. T., Fields, H. L., & Nicola, S. M. (2004). The ventral tegmental
area is required for the behavioral and nucleus accumbens neuronal firing responses to
incentive cues. The Journal of Neuroscience : The Official Journal of the Society for
Neuroscience, 24(12), 2923–2933. https://doi.org/10.1523/JNEUROSCI.5282-03.2004
Zahm, D. S., & Brog, J. S. (1992). On the significance of subterritories in the “accumbens” part
of the rat ventral striatum. Neuroscience, 50(4), 751–767. https://doi.org/10.1016/0306-
4522(92)90202-D
Zahm, D. S., & Heimer, L. (1993). Specificity in the efferent projections of the nucleus
accumbens in the rat: Comparison of the rostral pole projection patterns with those of the
core and shell. Journal of Comparative Neurology, 327(2), 220–232.
https://doi.org/10.1002/cne.903270205
Zeeb, F. D., Robbins, T. W., & Winstanley, C. A. (2009). Serotonergic and Dopaminergic
Modulation of Gambling Behavior as Assessed Using a Novel Rat Gambling Task.
Neuropsychopharmacology, 34(10), 2329–2343. https://doi.org/10.1038/npp.2009.62
Zelikowsky, M., Bissiere, S., Hast, T. A., Bennett, R. Z., & Abdipranoto, A. (2013). Prefrontal
microcircuit underlies contextual learning after hippocampal loss.
https://doi.org/10.1073/pnas.1301691110/-
/DCSupplemental.www.pnas.org/cgi/doi/10.1073/pnas.1301691110

223
Zhang, M., Balmadrid, C., & Kelley, A. E. (2003). Nucleus accumbens opioid, GABaergic, and
dopaminergic modulation of palatable food motivation: contrasting effects revealed by a
progressive ratio study in the rat. Behavioral Neuroscience, 117(2), 202–211.
https://doi.org/10.1037/0735-7044.117.2.202




















