
Hippocampal and cortical sensory gating in rats: effects of quinpirole microinjections in nucleus...
12
HIPPOCAMPAL AND CORTICAL SENSORY GATING IN RATS : EFFECTS OF QUINPIROLE MICROINJECTIONS IN NUCLEUS ACCUMBENS CORE AND SHELL N. M. W. J. DE BRUIN, a;b * B. A. ELLENBROEK, a E. L. J. M. VAN LUIJTELAAR, b A. R. COOLS a and K. E. STEVENS c a NICI Department of Psychoneuropharmacology, University of Nijmegen, P.O. Box 9104, 6500 HE Nijmegen, The Netherlands b NICI Department of Comparative and Physiological Psychology, University of Nijmegen, P.O. Box 9104, 6500 HE Nijmegen, The Netherlands c University of Colorado, Health Sciences Center/Department of Psychiatry, C268-71, 4200 East 9th Avenue, Denver, CO 80262, USA AbstractöSensory processing disturbances, as measured in the P50/sensory gating paradigm, have been linked to aber- rant auditory information processing and sensory overload in schizophrenic patients. In this paradigm, the response to the second of paired-click stimuli is attenuated by an inhibitory e¡ect of the ¢rst stimulus. Sensory gating has been observed in most healthy human subjects and normal laboratory rats. Because mesolimbic dopamine has been implicated in other ¢ltering disturbances such as prepulse inhibition of the acoustic startle response and given the fact that amphet- amine and apomorphine have been shown to disrupt gating, this study was performed to investigate the role of meso- limbic dopamine in sensory gating. The dopamine D2 receptor agonist quinpirole (10 Wg/0.5 Wl) was injected bilaterally in nucleus accumbens core and shell and e¡ects on cortical and hippocampal sensory gating were investigated. Also, e¡ects of the dopamine D2 receptor antagonist haloperidol (0.1 mg/kg, subcutaneously) as pretreatment were studied. First, quinpirole signi¢cantly reduced both the amplitude to the ¢rst click and gating as measured in the cortex and in the hippocampus. There was a tendency for the quinpirole e¡ects on hippocampal gating to be more pronounced in rats injected in the shell. Secondly, haloperidol did not antagonize e¡ects of quinpirole on hippocampal parameters, whereas haloperidol pretreatment fully antagonized quinpirole e¡ects on cortical parameters. In conclusion, gating can be signi¢cantly reduced when a dopamine agonist is speci¢cally targeted at mesolimbic dopamine D2 receptors. However, an important consideration is that the dopaminergic e¡ects in the present study on gating are predominantly mediated by the e¡ects on the amplitude to the ¢rst click. This has also been suggested for systemic amphetamine injections in rats and schizophrenic patients. This casts doubt on whether dopamine receptor activation a¡ects the putative inhibitory process between the ¢rst and the second stimulus. ß 2001 IBRO. Published by Elsevier Science Ltd. All rights reserved. Key words: auditory evoked potentials, dopamine, D2 receptors, haloperidol, schizophrenia. Aberrant auditory information processing and sensory overload exhibited by schizophrenic patients have been linked to ¢ltering disturbances, as measured in the P50 or sensory gating paradigm. Sensory gating can be de¢ned as the reduced auditory midlatency evoked potential response in the electroencephalogram (EEG) to the second (or test) stimulus compared to the ¢rst (or conditioning) stimulus of a click pair presented 500 ms apart. Sensory gating has been observed in most healthy human subjects and normal laboratory rats. This also holds true for another type of ¢ltering, prepulse inhibition (PPI), the decrease in startle response that occurs when a startling stimulus is preceded by a weaker stimulus (or prepulse) at intervals between 60 and 120 ms. Overactive dopamine (DA) neurotransmission in the nucleus accumbens (NAC) has been proposed to be an important pathology in schizophrenia (Bardgett et al., 1993, 1995; Carlsson, 1988; Poirier, 1998; Snyder, 1976). DA release in the NAC has been regarded as primarily modulating motor output pathways through the connections of the NAC to the basal ganglia (Mogenson et al., 1982). However, it has been proposed that NAC DA also plays a role in sensory information processing de¢cits in schizophrenia (Young et al., 1998). For example, it has become apparent that speci¢cally the mesolimbic DA terminal region in the NAC and pre- 169 *Correspondence to : N.M.W.J. de Bruin, NICI Department of Comparative and Physiological Psychology, University of Nijmegen, P.O. Box 9104, 6500 HB Nijmegen, The Netherlands. Tel.: +31-24- 3615566; fax: +31-24-3616066. E-mail address : [email protected] or [email protected] (N. M. W. J. de Bruin). Abbreviations : AP, anterioposterior ; CAMP, conditioning ampli- tude; CLAT, conditioning latency; DA, dopamine; DV, dorso- ventral; EEG, electroencephalogram; EP, evoked potential; L, lateral; MANOVA, multivariate analysis of variance; mPFC, medial prefrontal cortex; NAC, nucleus accumbens; PPI, pre- pulse inhibition ; SNr, substantia nigra pars reticulata ; TAMP, test amplitude ; TC ratio, test/condition ratio; TLAT, test latency; VP, ventral pallidum; VTA, ventral tegmental area. NSC 5028 16-7-01 www.elsevier.com/locate/neuroscience Neuroscience Vol. 105, No. 1, pp. 169^180, 2001 ß 2001 IBRO. Published by Elsevier Science Ltd Printed in Great Britain. All rights reserved PII:S0306-4522(01)00183-X 0306-4522 / 01 $20.00+0.00
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Hippocampal and cortical sensory gating in rats: effects of quinpirole microinjections in nucleus...

HIPPOCAMPAL AND CORTICAL SENSORY GATING IN RATS:EFFECTS OF QUINPIROLE MICROINJECTIONS IN NUCLEUS
ACCUMBENS CORE AND SHELL
N. M. W. J. DE BRUIN,a;b* B. A. ELLENBROEK,a E. L. J. M. VAN LUIJTELAAR,b A. R. COOLSa
and K. E. STEVENSc
aNICI Department of Psychoneuropharmacology, University of Nijmegen, P.O. Box 9104, 6500 HE Nijmegen, The NetherlandsbNICI Department of Comparative and Physiological Psychology, University of Nijmegen, P.O. Box 9104, 6500 HE Nijmegen,
The NetherlandscUniversity of Colorado, Health Sciences Center/Department of Psychiatry, C268-71, 4200 East 9th Avenue, Denver, CO 80262,
USA
AbstractöSensory processing disturbances, as measured in the P50/sensory gating paradigm, have been linked to aber-rant auditory information processing and sensory overload in schizophrenic patients. In this paradigm, the response tothe second of paired-click stimuli is attenuated by an inhibitory e¡ect of the ¢rst stimulus. Sensory gating has beenobserved in most healthy human subjects and normal laboratory rats. Because mesolimbic dopamine has been implicatedin other ¢ltering disturbances such as prepulse inhibition of the acoustic startle response and given the fact that amphet-amine and apomorphine have been shown to disrupt gating, this study was performed to investigate the role of meso-limbic dopamine in sensory gating. The dopamine D2 receptor agonist quinpirole (10 Wg/0.5 Wl) was injected bilaterally innucleus accumbens core and shell and e¡ects on cortical and hippocampal sensory gating were investigated. Also, e¡ectsof the dopamine D2 receptor antagonist haloperidol (0.1 mg/kg, subcutaneously) as pretreatment were studied. First,quinpirole signi¢cantly reduced both the amplitude to the ¢rst click and gating as measured in the cortex and in thehippocampus. There was a tendency for the quinpirole e¡ects on hippocampal gating to be more pronounced in ratsinjected in the shell. Secondly, haloperidol did not antagonize e¡ects of quinpirole on hippocampal parameters, whereashaloperidol pretreatment fully antagonized quinpirole e¡ects on cortical parameters.
In conclusion, gating can be signi¢cantly reduced when a dopamine agonist is speci¢cally targeted at mesolimbicdopamine D2 receptors. However, an important consideration is that the dopaminergic e¡ects in the present study ongating are predominantly mediated by the e¡ects on the amplitude to the ¢rst click. This has also been suggested forsystemic amphetamine injections in rats and schizophrenic patients. This casts doubt on whether dopamine receptoractivation a¡ects the putative inhibitory process between the ¢rst and the second stimulus. ß 2001 IBRO. Published byElsevier Science Ltd. All rights reserved.
Key words: auditory evoked potentials, dopamine, D2 receptors, haloperidol, schizophrenia.
Aberrant auditory information processing and sensoryoverload exhibited by schizophrenic patients have beenlinked to ¢ltering disturbances, as measured in the P50or sensory gating paradigm. Sensory gating can bede¢ned as the reduced auditory midlatency evokedpotential response in the electroencephalogram (EEG)
to the second (or test) stimulus compared to the ¢rst(or conditioning) stimulus of a click pair presented 500ms apart. Sensory gating has been observed in mosthealthy human subjects and normal laboratory rats.This also holds true for another type of ¢ltering, prepulseinhibition (PPI), the decrease in startle response thatoccurs when a startling stimulus is preceded by a weakerstimulus (or prepulse) at intervals between 60 and 120ms.
Overactive dopamine (DA) neurotransmission in thenucleus accumbens (NAC) has been proposed to be animportant pathology in schizophrenia (Bardgett et al.,1993, 1995; Carlsson, 1988; Poirier, 1998; Snyder,1976). DA release in the NAC has been regarded asprimarily modulating motor output pathways throughthe connections of the NAC to the basal ganglia(Mogenson et al., 1982). However, it has been proposedthat NAC DA also plays a role in sensory informationprocessing de¢cits in schizophrenia (Young et al., 1998).For example, it has become apparent that speci¢cally themesolimbic DA terminal region in the NAC and pre-
169
*Correspondence to: N.M.W.J. de Bruin, NICI Department ofComparative and Physiological Psychology, University of Nijmegen,P.O. Box 9104, 6500 HB Nijmegen, The Netherlands. Tel. : +31-24-3615566; fax: +31-24-3616066.E-mail address: [email protected] or [email protected](N. M. W. J. de Bruin).Abbreviations: AP, anterioposterior ; CAMP, conditioning ampli-
tude; CLAT, conditioning latency; DA, dopamine; DV, dorso-ventral ; EEG, electroencephalogram; EP, evoked potential ; L,lateral; MANOVA, multivariate analysis of variance; mPFC,medial prefrontal cortex; NAC, nucleus accumbens; PPI, pre-pulse inhibition; SNr, substantia nigra pars reticulata; TAMP,test amplitude; TC ratio, test/condition ratio; TLAT, testlatency; VP, ventral pallidum; VTA, ventral tegmental area.
NSC 5028 16-7-01
www.elsevier.com/locate/neuroscience
Neuroscience Vol. 105, No. 1, pp. 169^180, 2001ß 2001 IBRO. Published by Elsevier Science Ltd
Printed in Great Britain. All rights reservedPII: S 0 3 0 6 - 4 5 2 2 ( 0 1 ) 0 0 1 8 3 - X 0306-4522 / 01 $20.00+0.00

dominantly DA D2 receptors regulate PPI (Ho¡man andDonovan, 1994; Peng et al., 1990; Ralph et al., 1999;Schwarzkopf et al., 1993; Swerdlow et al., 1991; Wanand Swerdlow, 1993). Local infusion of DA and quinpir-ole in the NAC has been shown to signi¢cantly reducePPI (Reijmers et al., 1995; Swerdlow et al., 1990, 1992;Wan and Swerdlow, 1993). Furthermore, Wan andSwerdlow (1993) have found that the PPI disruptioninduced by intra-NAC quinpirole (10 Wg/0.5 Wl) couldbe antagonized by pretreatment with haloperidol (0.05^0.1 mg/kg, s.c.). Finally, Wan et al. (1994) have focusedon DA D2 receptor modulation of PPI within lateralcore versus medial shell accumbens subregions. Theyhave found a tendency for quinpirole to be more e¡ectivein the core and central accumbens compared to the shelland anteromedial accumbens. Also, injector distancefrom midline was positively correlated with the PPI-dis-ruptive e¡ect of quinpirole (for the 10 Wg/0.5 Wl dose).
Evidence underlining the role of DA in sensory gatingcomes from studies in which DA receptor agonists wereadministered to induce gating de¢cits. DA receptor ago-nists such as bromocryptine and amphetamine (0.3 mg/kg, per oral) have been found to reduce P50 gating inhealthy human subjects (Adler et al., 1994; Light et al.,1999). In rats, amphetamine (1^1.83 mg/kg, i.p.) alsointerferes with the suppression of cortical and hippocam-pal midlatency components, primarily by decreasing theamplitude to the ¢rst stimulus (Adler et al., 1986;Bickford-Wimer et al., 1990; De Bruin et al., 1999;Johnson et al., 1998; Stevens et al., 1991, 1995, 1996),which parallels cortical gating data in schizophrenicpatients (Adler et al., 1982; Boutros et al., 1991; Cullumet al., 1993; Freedman et al., 1983, 1987; Jin et al., 1997;Judd et al., 1992; Patterson et al., 2000; Zouridakis etal., 1997). It is not yet clear whether stimulation of me-solimbic DA receptors plays a role in sensory gating.
Therefore, the purpose of the present study was todetermine whether cortical and hippocampal sensory gat-ing can be disrupted by injecting the DA receptor agonistquinpirole (10 Wg/0.5 Wl) bilaterally into the NAC. Fur-thermore, the possible antagonistic e¡ect of the DA D2antagonist haloperidol (0.1 mg/kg, s.c.) was studied.Considering that the NAC shell and core have beenfound to di¡er anatomically as well as functionally, itwas decided to separately analyze the e¡ects of quinpir-ole on gating in both these NAC sites (Cools et al., 1993,1995; Groenewegen et al., 1999a,b; Koshikawa et al.,1996; Prinssen et al., 1994; Zahm and Brog, 1992).
EXPERIMENTAL PROCEDURES
Animals and surgery
Eighteen male Wistar rats (Charles Rivers, Wilmington, MA,USA) (weight: mean 412.50 þ S.E.M. 16.5 g) were maintainedon a 12-h light^dark cycle with lights on at 7:00 a.m. They hadad libitum access to standard food and tap water. Animals weretreated in accordance with the `Principals of laboratory animalcare' (NIH) and institutional guidelines. All e¡orts were madeto minimize animal su¡ering and to reduce the number of ani-mals used for the present experiment.
At the time of surgery, rats were anaesthetized with sodium
pentobarbital (Nembutal, 50 mg/kg, i.p.) and placed in a Kopfstereotaxic apparatus (Kopf Instruments, Tujunga, CA, USA).Atropine sulphate (0.4 mg/kg, i.p.) was given pre-operatively.
For the electrophysiological measurements, a surface elec-trode was placed on the vertex; a depth electrode was implantedunilaterally into the hippocampus aimed at the CA3 region anda reference electrode was placed on the frontal cortex. The sur-face electrode consisted of a stainless steel screw (00-09U1/8Q)soldered to a length of 0.005Q diameter Te£on-coated stainlesssteel wire terminated with a WirePro gold pin. This surfaceelectrode was placed 4.1 mm posterior to bregma, just to theright of the midline (`vertex') with the end in contact with dura.The depth electrode consisted of a Te£on-coated stainless steelwire (0.005Q diameter) terminated with a WirePro gold pin. Thiselectrode, targeted to the hippocampus, was placed from 3.3 to4.5 mm posterior to bregma, 4.4 mm lateral, and 3.6 mm deep(Paxinos and Watson, 1997). The depth of the electrode wasestimated from the brain surface. The reference electrode con-sisted of a pair of 0.010Q diameter Te£on-coated stainless steelwires crimped to a single WirePro gold pin at one bent to a 90³angle at the other end. The angled ends were placed in contactwith frontal cortex through burr holes drilled þ 3.0 mm anteriorto bregma and þ 1.5 mm lateral to midline. Pins from the sur-face, depth and reference electrodes were inserted into a malehead-plug assembly (Carlton University, Ottawa, Canada) thatwas secured to the skull with dental acrylic cement. Four screwswere secured to the skull around the periphery to serve asanchor points.
For the microinjections, rats were equipped with 23-gaugestainless steel guide cannulae at a length of 17 mm (Smallparts, Inc., Miami Lakes, FL, USA), bilaterally implanted intothe NAC core or shell, according to a stereotaxic atlas (Paxinosand Watson, 1997): core (lateral/central NAC), anterioposterior(AP)+1.1 (bregma), lateral (L) þ 1.6, dorsoventral (DV)36.8(measured from skull); shell (medial NAC), AP+1.2, L þ 0.9,DV37.0. Cannulae were anchored to the skull with cementand screws. The lumen of the cannulae was occluded with awire stylet. Animals were allowed at least 10 days to recoverfrom surgery before electrophysiological recording commenced.
Apparatus and procedures
EEG recording took place in a Plexiglas chamber (21U22 cm£oor), housed in a sound-attenuated enclosure. The head-plugassembly on the rat was connected to a female head-plug andlocking ring assembly. This assembly was, in turn, attached bycable to a slip-ring commutator at the top of the recordingchamber, thus giving the rat freedom of movement.
Auditory stimuli were delivered through a speaker located inthe ceiling of the recording chamber enclosure, 45 cm above thebase of the chamber. The stimuli consisted of paired 500-Wsduration clicks, presented 0.5 s apart, at 87 dB (SPL) thatwere computer controlled (EPMax, Eclectic Engineering Studio,Canton, MA, USA). Click pairs were automatically presented at15-s intervals. Low-level background noise provided by the ven-tilator fan was continuously present (V60 dB).
The electrical activity was ampli¢ed 1000 times with bandpass1^500 Hz. Evoked potentials (EP) were digitized at 1 kHz over512 m in epochs, beginning 100 ms preceding stimulus onset andstored on a computer for later averaging.
Animals were handled for several minutes prior to connectionto the recording system and were allowed several minutes accli-matization once in the chamber. As trials were presented, thebehavioral state of the animal was noted. Only trials occurringwhile the animal was motionless and alert were accepted foranalysis. Pairs of clicks (a conditioning click followed by testclick) were presented until 30 alert-still trials per recording ses-sion had been stored for subsequent averaging and analysis(EPMax, Eclectic Engineering Studio). At least 10 baselinerecordings were performed on each animal over several days,one recording session per day. The still-alert trials from a singlesession were averaged together, and the latency and amplitudeof waveforms evoked by the conditioning and test clicks weredetermined from the average. For the vertex and hippocampal
NSC 5028 16-7-01
N. M. W. J. de Bruin et al.170
electrode EP, the maximum negativity between 20 and 60 msafter the ¢rst stimulus was selected as the N40 and N35 compo-nents, respectively. The amplitude was calculated relative to thebaseline, de¢ned as the average activity for 100 ms prior tostimulus onset. A test/condition ratio (TC ratio) was calculatedfor each averaged recording by dividing the amplitude of thetest EP by the amplitude of the condition EP. A ratio of lessthan 1.0 indicated that the response to the test click was sup-pressed compared to the conditioning click. If the ratio was lessthan 0.4, the animal was determined to be `gating' its responseto the test click (Stevens et al., 1991). A ratio higher than 1.0indicated that the response to the test click was enhanced com-pared to the conditioning click. Drugs were administered afterstable baseline gating measurements were obtained. Post-drugtrials were acquired at 0^20 min after each drug treatment.
Haloperidol pretreatment and quinpirole microinjection
A within-subject design was applied, so that each rat receivedall four treatments with at least 48 h between each treatment(order of treatments was randomized): VEH-SAL = vehicle(1 ml/kg, s.c.) and saline (0.5 Wl, intra-NAC); VEH-QUIN = ve-hicle (1 ml/kg, s.c.) and quinpirole (10 Wg/0.5 Wl, intra-NAC);HAL-SAL = haloperidol (0.1 mg/kg, s.c.) and saline (0.5 Wl,intra-NAC); HAL-QUIN = haloperidol (0.1 mg/kg, s.c.) andquinpirole (10 Wg/0.5 Wl, intra-NAC). Ten minutes before theintracranial injection, rats were pretreated with the dopamineantagonist haloperidol (0.1 mg/kg, s.c., diluted with salinefrom a stock 5 mg/ml water/lactic acid solution to an injectionvolume of 1 ml/kg) or vehicle alone. Immediately prior to therecording session, the stylet wires were removed from the intra-NAC cannulae. A 30-gauge stainless steel injection cannula,connected to a microliter syringe with micro-bore polyethylenetubing, was inserted into the guide cannula. Each rat received abilateral intra-NAC infusion (either core or shell) of the D2/D3agonist quinpirole (10 Wg/0.5 Wl saline per side) or vehicle (0.5 Wlper side) over a period of 30 s. The injection cannula was left inplace for an additional 30 s to prevent back£ow up the guidecannula and to allow for di¡usion of the drug.
Statistical analyses
Five parameters were assessed in each session: conditioningamplitude (CAMP); test amplitude (TAMP); conditioninglatency (CLAT); test latency (TLAT); gating parameter (TCratio, TAMP/CAMP). Data were analyzed with multivariateanalysis of variance (MANOVA) repeated measures (SPSS 8.0Statistical Package) to determine the drug, NAC site and inter-action e¡ects on the EP parameters. The general linear model(GLM) procedure was used with haloperidol (vehicle/haloperi-dol pretreatment) and quinpirole (saline/quinpirole NAC micro-
injection) as within-subject factors and NAC site (core/shell) asbetween-subject factor. Statistical signi¢cance of main e¡ectsand interactions was set at the probability level of P6 0.05.
Histology
Following the completion of the experiment, rats were deeplyanaesthetized with sodium pentobarbital. A current (1 V deliv-ered every 2 s, 15 times) was passed through the depth hippo-campal electrode to induce a small electrolytic lesion in order toverify electrode location. Each rat received a bilateral intra-NAC infusion of a solvent blue 38 S-3382 solution (0.5 Wl) forhistological veri¢cation of injection site. Subsequently, rats wereperfused transcardially with 30 ml of 4% formalin solution con-taining 10% potassium ferrocyanide. The brains were stored in a4% formalin solution containing 10% potassium ferrocyanidefor at least 3 days, after which they were transferred to vialscontaining a 30% sucrose solution as a cryoprotectant, wherethey remained until they sank (1 day). Ferric ions depositedfollowing the electrolytic lesion reacted with the potassium fer-rocyanide and produced a blue color to aid in locating theelectrode tip. The brains were frozen at 324³C, cut on a cryo-stat into 40-W slices, placed on gelatin-coated slides, allowed todry and stained with Cresyl Violet for determination of theinjection sites and the localization of the hippocampal electrode.Data from rats with misplaced cannulae were discarded.
Drugs
Nembutal brand of sodium pentobarbital (50 mg/ml) wasobtained from Abbott laboratories (North Chicago, IL, USA).( þ )-Quinpirole dihydrochloride (LY-141865) was obtainedfrom Research Biochemicals International (RBI, Natick, MA,USA); Haldol brand haloperidol (5 mg/ml) was obtained fromMcNeilab, Inc. (Spring House, PA, USA). Formalin, sucrose,Cresyl Violet, solvent blue 38 (S-3382), atropine sulphate andpotassium ferrocyanide were purchased from Sigma ChemicalCompany (St. Louis, MO, USA). All drugs were dissolved in0.9% saline with the exception of haloperidol.
RESULTS
Histology
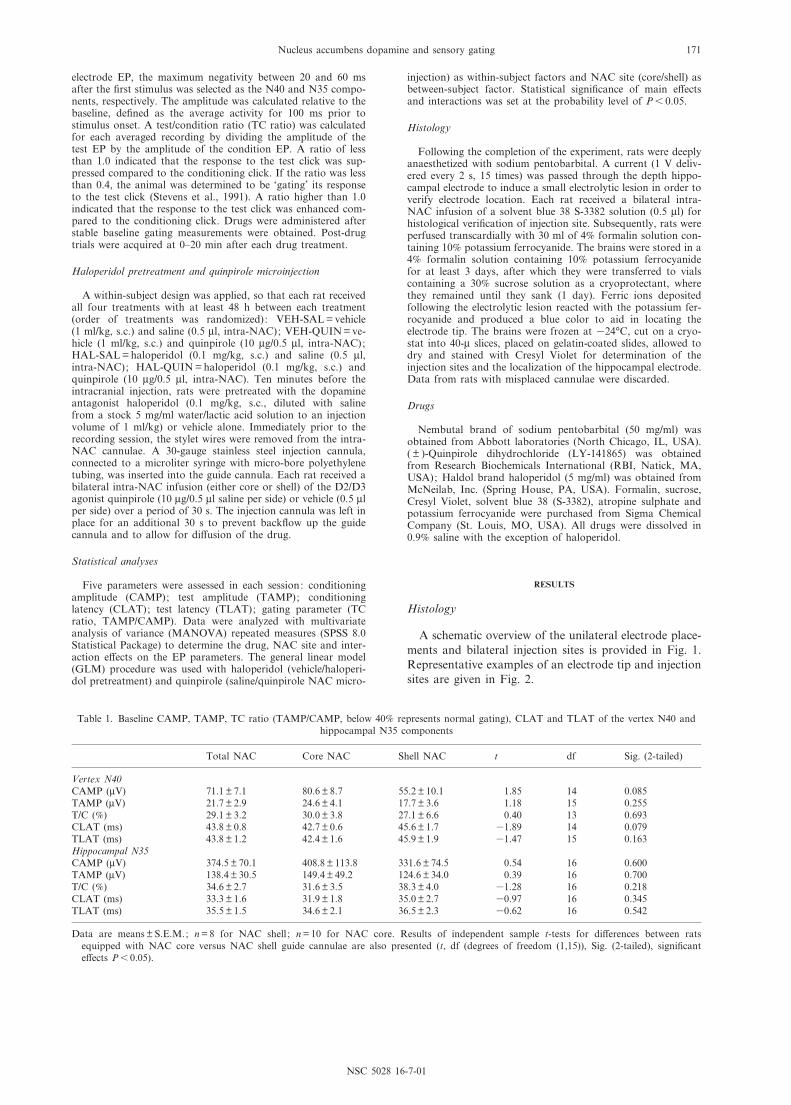
A schematic overview of the unilateral electrode place-ments and bilateral injection sites is provided in Fig. 1.Representative examples of an electrode tip and injectionsites are given in Fig. 2.
Table 1. Baseline CAMP, TAMP, TC ratio (TAMP/CAMP, below 40% represents normal gating), CLAT and TLAT of the vertex N40 andhippocampal N35 components
Total NAC Core NAC Shell NAC t df Sig. (2-tailed)
Vertex N40CAMP (WV) 71.1 þ 7.1 80.6 þ 8.7 55.2 þ 10.1 1.85 14 0.085TAMP (WV) 21.7 þ 2.9 24.6 þ 4.1 17.7 þ 3.6 1.18 15 0.255T/C (%) 29.1 þ 3.2 30.0 þ 3.8 27.1 þ 6.6 0.40 13 0.693CLAT (ms) 43.8 þ 0.8 42.7 þ 0.6 45.6 þ 1.7 31.89 14 0.079TLAT (ms) 43.8 þ 1.2 42.4 þ 1.6 45.9 þ 1.9 31.47 15 0.163Hippocampal N35CAMP (WV) 374.5 þ 70.1 408.8 þ 113.8 331.6 þ 74.5 0.54 16 0.600TAMP (WV) 138.4 þ 30.5 149.4 þ 49.2 124.6 þ 34.0 0.39 16 0.700T/C (%) 34.6 þ 2.7 31.6 þ 3.5 38.3 þ 4.0 31.28 16 0.218CLAT (ms) 33.3 þ 1.6 31.9 þ 1.8 35.0 þ 2.7 30.97 16 0.345TLAT (ms) 35.5 þ 1.5 34.6 þ 2.1 36.5 þ 2.3 30.62 16 0.542
Data are means þ S.E.M.; n = 8 for NAC shell ; n = 10 for NAC core. Results of independent sample t-tests for di¡erences between ratsequipped with NAC core versus NAC shell guide cannulae are also presented (t, df (degrees of freedom (1,15)), Sig. (2-tailed), signi¢cante¡ects P6 0.05).
NSC 5028 16-7-01
Nucleus accumbens dopamine and sensory gating 171
Baseline values of hippocampal N35 and corticalN40 EP parameters
Both the hippocampal N35 and vertex N40 showedsigni¢cant gating at baseline with the mean TC ratioless than 0.4 (see Table 1). No di¡erences were foundat baseline between rats equipped with cannulae in theNAC core and rats equipped with cannulae in the NACshell with respect to all ¢ve parameters (see results ofindependent t-tests in Table 1).
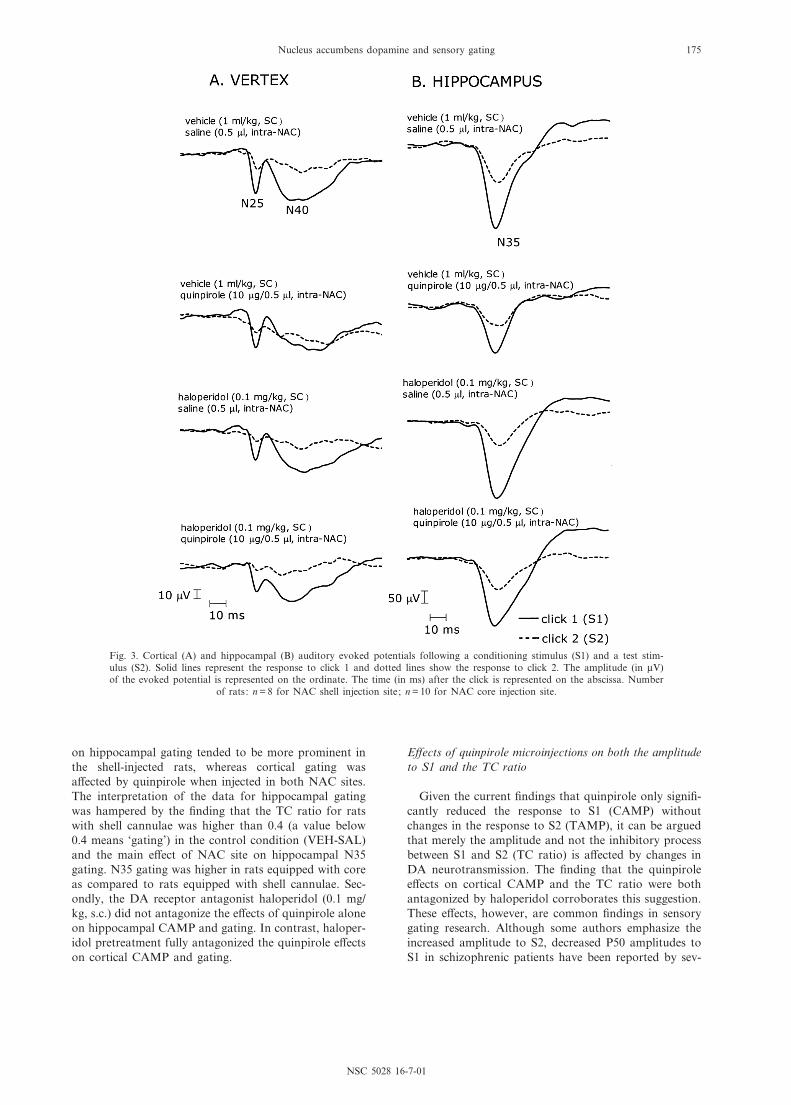
Drug e¡ects on cortical N40
Grand-average vertex auditory evoked potential wave-forms per drug condition are presented in Fig. 3A.E¡ects of bilateral NAC quinpirole injections, haloperi-dol alone, haloperidol pretreatment and interactions on
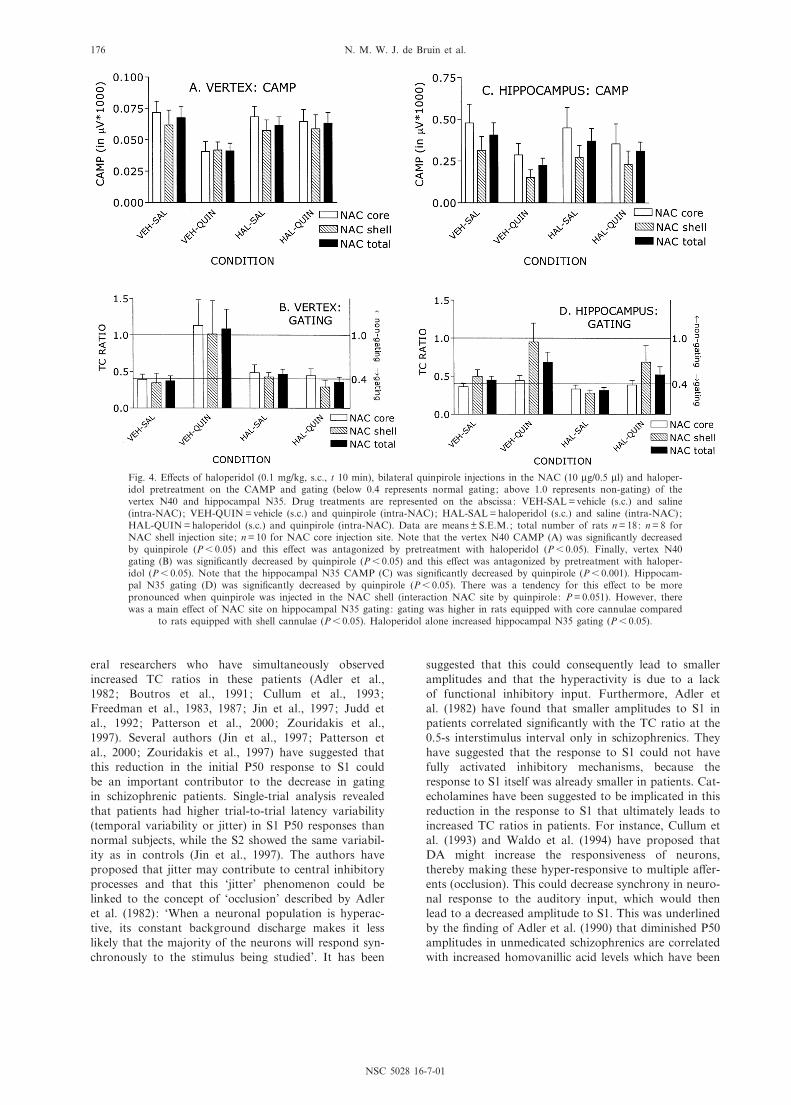
vertex N40 parameters are presented in Fig. 4A,B andTable 2.
No signi¢cant di¡erences were found between ratsinjected in the core and rats injected in the shell (therewere no main e¡ects of NAC site, see Table 2). Also,interactions between the between-subject factor NAC site(shell/core) and the within-subject factors (haloperidoland quinpirole) were not signi¢cant for vertex parame-ters (see Table 2). Quinpirole signi¢cantly reducedCAMP (main e¡ect of quinpirole: F1;15 = 5.3, P6 0.05).There was no e¡ect of haloperidol alone on CAMP(main e¡ect of haloperidol: F1;15 = 0.0, not signi¢cant).Haloperidol, co-administered with quinpirole, fullyreversed the reduction of CAMP observed with quinpir-ole alone (haloperidol by quinpirole interaction:F1;15 = 6.0, P6 0.05). Finally, no signi¢cant drug e¡ectswere found on vertex N40 TAMP, CLAT or TLAT (seeTable 2).
Fig. 1. Schematic drawings (adapted from Paxinos and Watson, 1997) of coronal sections, showing the locations of electrodetips aimed at the CA3 region of the hippocampus (A) and of injection sites in the lateral/central (core, B) part and medial(shell, C) part of the NAC. (CA3: AP 3.3^4.5 mm posterior to bregma, L 34.4 mm, DV 33.6 mm (measured from brainsurface) ; core/lateral-central NAC: AP 0.7^1.7 mm anterior to bregma, L þ 1.6 mm, DV 36.8 (measured from skull) ; shell/medial NAC: AP 0.7^1.7 mm anterior to bregma, L þ 0.9 mm, DV 37.0 (measured from skull)). Number of rats: n = 18 for
unilateral hippocampal electrode; n = 10 for bilateral NAC core injection site; n = 8 for bilateral NAC shell injection site.
NSC 5028 16-7-01
N. M. W. J. de Bruin et al.172
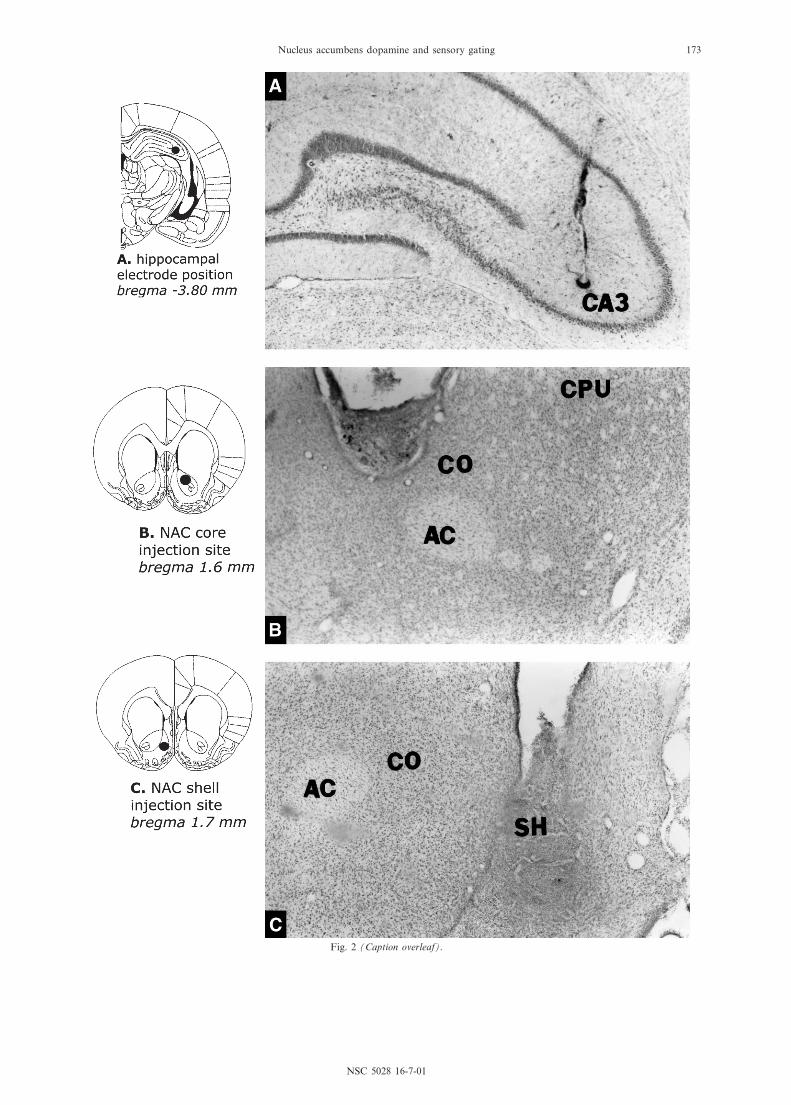
Fig. 2 (Caption overleaf).
NSC 5028 16-7-01
Nucleus accumbens dopamine and sensory gating 173
Signi¢cant gating of the N40 component occurred inrats in the control condition (VEH-SAL), since the val-ues of the TC ratio were less than 0.4. The TC ratio wassigni¢cantly increased following quinpirole alone,thereby decreasing gating (main e¡ect of quinpirole:F1;15 = 4.7, P6 0.05), an e¡ect that was fully antagonizedby haloperidol pretreatment (haloperidol by quinpiroleinteraction: F1;15 = 5.7, P6 0.05). Haloperidol did notchange the vertex TC ratio (F1;15 = 2.7, not signi¢cant).
Drug e¡ects on hippocampal N35
Grand-average (average for all animals) hippocampalauditory evoked potential waveforms per drug conditionare presented in Fig. 3B. E¡ects of bilateral NAC quin-pirole injections, haloperidol alone, haloperidol pretreat-ment and interactions on hippocampal N35 parametersare presented in Fig. 4C, D and Table 2.
Hippocampal N35 CAMP was signi¢cantly reduced byquinpirole (main e¡ect of quinpirole: F1;15 = 25.6,P6 0.001), irrespective of NAC site (NAC site by quin-pirole interaction: F1;15 = 0.8, not signi¢cant). There wasno e¡ect of haloperidol alone on CAMP (main e¡ect ofhaloperidol: F1;15 = 0.6, not signi¢cant) and also no e¡ectof haloperidol co-administered with quinpirole onCAMP (haloperidol by quinpirole interaction:F1;15 = 3.1, not signi¢cant). CLAT was not signi¢cantlya¡ected when quinpirole was administered alone (maine¡ect of quinpirole: F1;15 = 0.0, not signi¢cant), whereasa signi¢cant increase in CLAT was observed followinginjection of haloperidol alone (main e¡ect of haloperi-
dol: F1;15 = 8.7, P6 0.05, VEH-SAL (31.1 þ 1.9 ms) andVEH-HAL (33.0 þ 2.0 ms)). Finally, no drug or injectionsite (core/shell) e¡ects were found on TAMP or TLAT(see Table 2).
Hippocampal N35 gating was signi¢cantly decreasedby quinpirole (main e¡ect of quinpirole: F1;15 = 8.1,P6 0.05). There was a tendency for this e¡ect to bemore pronounced when quinpirole was injected in theNAC shell (interaction NAC site by quinpirole:F1;15 = 4.5, P = 0.051). However, there was also a maine¡ect of NAC site on hippocampal N35 gating: gatingwas higher in rats equipped with core cannulae com-pared to rats equipped with shell cannulae (main e¡ectof NAC site: F1;15 = 7.5, P6 0.05). Moreover, the valueof N35 TC ratio was 0.49 þ 0.084 in shell-injected rats inthe control condition (VEH-SAL) and therefore alreadyhigher than 0.4 (a value below 0.4 represents `gating').Haloperidol alone increased hippocampal N35 gating(main e¡ect of haloperidol: F1;15 = 5.2, P6 0.05). Finally,haloperidol, co-administered with quinpirole, did notreverse the reduction of gating observed with quinpirolealone (haloperidol by quinpirole interaction: F1;15 = 0.0,not signi¢cant).
DISCUSSION
First, microinjections of quinpirole in the NAC (10 Wg/0.5 Wl) signi¢cantly reduced both the amplitude to the¢rst click and gating of the hippocampal N35 and corti-cal N40 components. However, the e¡ect of quinpirole
Table 2. Statistical results of MANOVA repeated measures : e¡ects of haloperidol (within-subject factor), quinpirole (within-subject factor),NAC site (between-subject factor) and interactions on gating (TC ratio), CAMP, TAMP, CLAT and TLAT of hippocampal N35 and cortical
N40
CAMP TAMP TC CLAT TLAT
VERTEXH F = 0.0, P = 0.871 F = 0.2, P = 0.670 F = 2.7, P = 0.118 F = 1.0, P = 0.331 F = 0.1, P = 0.811HxN F = 1.0, P = 0.331 F = 0.0, P = 0.879 F = 0.1, P = 0.798 F = 1.2, P = 0.302 F = 0.2, P = 0.640Q F = 5.3, P = 0.040 F = 0.2, P = 0.632 F = 4.7, P = 0.046 F = 0.0, P = 0.970 F = 0.0, P = 0.999QxN F = 0.1, P = 0.709 F = 2.5, P = 0.131 F = 0.1, P = 0.816 F = 0.9, P = 0.367 F = 0.0, P = 0.917HxQ F = 6.0, P = 0.027 F = 0.4, P = 0.555 F = 5.7, P = 0.033 F = 1.9, P = 0.186 F = 2.3, P = 0.147HxQxN F = 0.5, P = 0.481 F = 1.1, P = 0.305 F = 0.1, P = 0.814 F = 1.2, P = 0.289 F = 2.3, P = 0.153N F = 2.1, P = 0.169 F = 1.3, P = 0.276 F = 0.3, P = 0.579 F = 0.0, P = 0.857 F = 1.5, P = 0.245CA3H F = 0.6, P = 0.458 F = 1.0, P = 0.345 F = 5.2, P = 0.041 F = 8.7, P = 0.010 F = 0.1, P = 0.794HxN F = 0.1, P = 0.752 F = 2.5, P = 0.180 F = 1.8, P = 0.205 F = 2.0, P = 0.178 F = 0.4, P = 0.534Q F = 25.6, P = 0.000 F = 1.0, P = 0.324 F = 8.1, P = 0.012 F = 0.0, P = 0.889 F = 0.4, P = 0.458QxN F = 0.8, P = 0.391 F = 2.3, P = 0.149 F = 4.5, P = 0.051 F = 0.2, P = 0.682 F = 2.9, P = 0.108HxQ F = 3.1, P = 0.098 F = 4.4, P = 0.054 F = 0.0, P = 0.976 F = 0.0, P = 0.983 F = 1.1, P = 0.296HxQxN F = 0.0, P = 0.862 F = 0.3, P = 0.588 F = 0.1, P = 0.718 F = 0.1, P = 0.729 F = 0.1, P = 0.794N F = 1.6, P = 0.227 F = 0.8, P = 0.384 F = 7.5, P = 0.015 F = 1.2, P = 0.299 F = 2.1, P = 0.171
Main e¡ects : H, haloperidol ; Q, quinpirole ; N, NAC site. Interactions : HxN, haloperidol by NAC site ; QxN, quinpirole by NAC site ;HxQxN, haloperidol by quinpirole by NAC site; F, F-value; P, P-value; statistical signi¢cance of main e¡ects and interactions was set atthe probability level of P6 0.05.
6Fig. 2. Photomicrographs of representative coronal sections showing the location of an electrode tip aimed at the CA3 regionof the hippocampus (A) and of injection sites in the lateral (core, B) part and medial (shell, C) part of the NAC. Schematicdrawings of coronal sections are adapted from Paxinos and Watson (1997). Abbreviations: AC, anterior commissure; CO,
core; CPu, caudate^putamen; NAC, nucleus accumbens; SH, shell.
NSC 5028 16-7-01
N. M. W. J. de Bruin et al.174
on hippocampal gating tended to be more prominent inthe shell-injected rats, whereas cortical gating wasa¡ected by quinpirole when injected in both NAC sites.The interpretation of the data for hippocampal gatingwas hampered by the ¢nding that the TC ratio for ratswith shell cannulae was higher than 0.4 (a value below0.4 means `gating') in the control condition (VEH-SAL)and the main e¡ect of NAC site on hippocampal N35gating. N35 gating was higher in rats equipped with coreas compared to rats equipped with shell cannulae. Sec-ondly, the DA receptor antagonist haloperidol (0.1 mg/kg, s.c.) did not antagonize the e¡ects of quinpirole aloneon hippocampal CAMP and gating. In contrast, haloper-idol pretreatment fully antagonized the quinpirole e¡ectson cortical CAMP and gating.
E¡ects of quinpirole microinjections on both the amplitudeto S1 and the TC ratio
Given the current ¢ndings that quinpirole only signi¢-cantly reduced the response to S1 (CAMP) withoutchanges in the response to S2 (TAMP), it can be arguedthat merely the amplitude and not the inhibitory processbetween S1 and S2 (TC ratio) is a¡ected by changes inDA neurotransmission. The ¢nding that the quinpirolee¡ects on cortical CAMP and the TC ratio were bothantagonized by haloperidol corroborates this suggestion.These e¡ects, however, are common ¢ndings in sensorygating research. Although some authors emphasize theincreased amplitude to S2, decreased P50 amplitudes toS1 in schizophrenic patients have been reported by sev-
Fig. 3. Cortical (A) and hippocampal (B) auditory evoked potentials following a conditioning stimulus (S1) and a test stim-ulus (S2). Solid lines represent the response to click 1 and dotted lines show the response to click 2. The amplitude (in WV)of the evoked potential is represented on the ordinate. The time (in ms) after the click is represented on the abscissa. Number
of rats: n = 8 for NAC shell injection site; n = 10 for NAC core injection site.
NSC 5028 16-7-01
Nucleus accumbens dopamine and sensory gating 175
eral researchers who have simultaneously observedincreased TC ratios in these patients (Adler et al.,1982; Boutros et al., 1991; Cullum et al., 1993;Freedman et al., 1983, 1987; Jin et al., 1997; Judd etal., 1992; Patterson et al., 2000; Zouridakis et al.,1997). Several authors (Jin et al., 1997; Patterson etal., 2000; Zouridakis et al., 1997) have suggested thatthis reduction in the initial P50 response to S1 couldbe an important contributor to the decrease in gatingin schizophrenic patients. Single-trial analysis revealedthat patients had higher trial-to-trial latency variability(temporal variability or jitter) in S1 P50 responses thannormal subjects, while the S2 showed the same variabil-ity as in controls (Jin et al., 1997). The authors haveproposed that jitter may contribute to central inhibitoryprocesses and that this `jitter' phenomenon could belinked to the concept of `occlusion' described by Adleret al. (1982): `When a neuronal population is hyperac-tive, its constant background discharge makes it lesslikely that the majority of the neurons will respond syn-chronously to the stimulus being studied'. It has been
suggested that this could consequently lead to smalleramplitudes and that the hyperactivity is due to a lackof functional inhibitory input. Furthermore, Adler etal. (1982) have found that smaller amplitudes to S1 inpatients correlated signi¢cantly with the TC ratio at the0.5-s interstimulus interval only in schizophrenics. Theyhave suggested that the response to S1 could not havefully activated inhibitory mechanisms, because theresponse to S1 itself was already smaller in patients. Cat-echolamines have been suggested to be implicated in thisreduction in the response to S1 that ultimately leads toincreased TC ratios in patients. For instance, Cullum etal. (1993) and Waldo et al. (1994) have proposed thatDA might increase the responsiveness of neurons,thereby making these hyper-responsive to multiple a¡er-ents (occlusion). This could decrease synchrony in neuro-nal response to the auditory input, which would thenlead to a decreased amplitude to S1. This was underlinedby the ¢nding of Adler et al. (1990) that diminished P50amplitudes in unmedicated schizophrenics are correlatedwith increased homovanillic acid levels which have been
Fig. 4. E¡ects of haloperidol (0.1 mg/kg, s.c., t 10 min), bilateral quinpirole injections in the NAC (10 Wg/0.5 Wl) and haloper-idol pretreatment on the CAMP and gating (below 0.4 represents normal gating; above 1.0 represents non-gating) of thevertex N40 and hippocampal N35. Drug treatments are represented on the abscissa: VEH-SAL = vehicle (s.c.) and saline(intra-NAC); VEH-QUIN = vehicle (s.c.) and quinpirole (intra-NAC); HAL-SAL = haloperidol (s.c.) and saline (intra-NAC);HAL-QUIN = haloperidol (s.c.) and quinpirole (intra-NAC). Data are means þ S.E.M.; total number of rats n = 18: n = 8 forNAC shell injection site; n = 10 for NAC core injection site. Note that the vertex N40 CAMP (A) was signi¢cantly decreasedby quinpirole (P6 0.05) and this e¡ect was antagonized by pretreatment with haloperidol (P6 0.05). Finally, vertex N40gating (B) was signi¢cantly decreased by quinpirole (P6 0.05) and this e¡ect was antagonized by pretreatment with haloper-idol (P6 0.05). Note that the hippocampal N35 CAMP (C) was signi¢cantly decreased by quinpirole (P6 0.001). Hippocam-pal N35 gating (D) was signi¢cantly decreased by quinpirole (P6 0.05). There was a tendency for this e¡ect to be morepronounced when quinpirole was injected in the NAC shell (interaction NAC site by quinpirole: P = 0.051). However, therewas a main e¡ect of NAC site on hippocampal N35 gating: gating was higher in rats equipped with core cannulae compared
to rats equipped with shell cannulae (P6 0.05). Haloperidol alone increased hippocampal N35 gating (P6 0.05).
NSC 5028 16-7-01
N. M. W. J. de Bruin et al.176

suggested to re£ect the activity of DA projections (Pickaret al., 1984).
In short, it can be concluded that DA primarilyincreases the TC ratio by decreasing the amplitude toS1. So, given the reduction in the amplitude to S1 byquinpirole that for the cortex could be antagonized byhaloperidol in the present study, it can be suggested thatstimulation of DA D2 receptors in the NAC could haveincreased occlusion, thereby increasing the temporal var-iability in the response to S1 (or jitter) that could explainthe reduction in the amplitude to S1 and the increase inthe TC ratio.
The local NAC e¡ects of quinpirole on CAMP andTC ratio in the current study and the ¢ndings in schiz-ophrenic patients have also been found for systemicallyinjected amphetamine (1^1.83 mg/kg) and apomorphine(1 mg/kg) in rats. Both DA receptor agonists have beenfound to reduce cortical CAMP and increase the corticalTC ratio (Adler et al., 1986, 1988; De Bruin et al., 1999;Johnson et al., 1998; Stevens et al., 1991, 1995, 1996).Both e¡ects could be antagonized by haloperidol (1 mg/kg, i.p.) (Adler et al., 1986). Amphetamine (1.0 mg/kg,i.p.) has also been shown to reduce the suppression ofthe hippocampal N40 and decrease the hippocampalresponse to S1 (Bickford-Wimer et al., 1990). In thepresent study, quinpirole e¡ects on CAMP and the TCratio as measured in the hippocampus were not reversedby haloperidol. In contrast, Bickford-Wimer et al. (1990)have shown that the amphetamine-induced reduction ofhippocampal gating was antagonized by haloperidol.However, they have injected a considerably higher doseof haloperidol (1 mg/kg, i.p.) compared to the dose usedin the present study (0.1 mg/kg, s.c.).
Several arguments have been presented that theincrease in TC ratio (re£ecting a reduction in gating)by amphetamine can either be induced by a decrease inCAMP due to DA mechanisms or an increase in TAMPdue to noradrenergic mechanisms. First, DSP-4 was usedto remove e¡ects of amphetamine mediated by norepi-nephrine by selectively lesioning the noradrenergic termi-nals (Adler et al., 1988). Since the amphetamine-inducedreduction in gating was diminished following DSP-4, ithas been suggested that norepinephrine plays an impor-tant role in the amphetamine e¡ects on gating. Theamphetamine-induced reduction in amplitude was stillpresent following DSP-4. Therefore, it has been sug-gested that DA is involved in the modulation by amphet-amine of the amplitude to S1. Secondly, Stevens et al.(1991) have shown that amphetamine reduced gatingthrough its e¡ects on alpha- and beta-adrenergic recep-tors and DA D1 receptors. However, adrenergic e¡ectswere predominantly on the amplitude to S2, whereas DAe¡ects were on the amplitude to S1. The role of DAactivity at D2 receptors was suggested to be less evident,since amphetamine e¡ects were only partially blocked bythe D2 receptor antagonist sulpiride (Stevens et al.,1991).
Results from the present study, in which a speci¢c DAreceptor agonist (quinpirole) was injected, strongly sug-gest that mesolimbic DA D2 receptors regulate sensorygating primarily through their e¡ects on the CAMP,
possibly enhancing occlusion. This holds particularlytrue for cortical parameters, since e¡ects of quinpiroleon both cortical CAMP and TC ratio were fully antag-onized by the DA D2 antagonist haloperidol. A similarmechanism could be implicated in the e¡ects on DA D1receptors, since the D1 receptor antagonist SCH 23390has been shown to normalize gating (or decrease the TCratio) by elevating the amphetamine-reduced amplitudeto S1 (Stevens et al., 1991).
NAC core versus shell injections
The e¡ect of quinpirole on the TC ratio and CAMP ofthe cortical component was signi¢cant following injec-tion in both NAC core and shell. In contrast, in thehippocampus, the e¡ects on the TC ratio tended to bemore prominent when quinpirole was injected in theNAC shell region than in the core. In order to explainthe role of the accumbal subdivisions in the DA modu-lation of the TC ratio and CAMP as measured in thehippocampus and cortex, it is necessary to review thedi¡erential output pathways of the shell and the coreregions. Since there are no direct projections from theNAC to the hippocampus or cortex, the indirect circui-tries described here are merely hypotheses. Therefore,there are no conclusive anatomical explanations for theNAC quinpirole e¡ects on hippocampal and corticalparameters.
The NAC shell has an inhibitory projection to theventral pallidum (VP) (Groenewegen et al., 1999a,b)that in turn has an inhibitory output to the ventral teg-mental area (VTA) (for review Heimer et al., 1995). TheVTA provides DA inputs to the dendate gyrus, entorhi-nal cortex and medial prefrontal cortex (mPFC) (Bergeret al., 1976; Bjo«rklund and Lindvall, 1984; Descarries etal., 1987; Fuxe et al., 1985; Swanson, 1982). So, quin-pirole injections in the NAC shell could have induced areduction in feedback inhibition via the VP therebyincreasing DA release in the hippocampal and corticalareas. Since DA has been proposed to increase theresponsiveness of neurons leading to occlusion (Cullumet al., 1993; Waldo et al., 1994), DA could have locally(cortex and hippocampus) decreased synchrony inresponse and decreased the amplitude of the evokedpotential. Furthermore, there might be an alternativeexplanation for the e¡ects of quinpirole in the NACshell on cortical parameters. The VP that is predomi-nantly innervated by the NAC shell, also sends GABAer-gic ¢bers directly to the mPFC (for review Groenewegenet al., 1993, 1999b; Heimer et al., 1995). Additionally,the VP projects to the cortex predominantly via excita-tory projections from the thalamus. Thus, it can be pro-posed that DA activation in the NAC shell might lead toa reduction in GABAergic output to the mPFC and dis-inhibition of thalamocortical cells via the VP. This couldhave resulted in cortical neurons to become hyper-responsive and consequently lead to occlusion and areduction in cortical CAMP.
Quinpirole injections in the NAC core decreased bothgating and CAMP as measured on the cortex in thepresent study. Core neurons have been suggested to
NSC 5028 16-7-01
Nucleus accumbens dopamine and sensory gating 177

send inhibitory projections to the substantia nigra parsreticulata (SNr) (Groenewegen et al., 1999a). The SNrsends GABAergic ¢bers to the thalamus that in turninnervates the cortex. DA activity in the NAC core cantherefore be suggested to result in a disinhibition of tha-lamocortical neurons via the SNr. This could haveinduced the cortical neurons to become hyper-responsiveand this could consequently lead to occlusion and areduction in cortical CAMP.
Quinpirole and D3 receptors
Some authors have suggested that DA D3 receptors ofthe mesolimbic system are associated with the patho-physiology of schizophrenia (Gri¡on et al., 1995; Joyceand MeadorWoodru¡, 1997). Since quinpirole is alsoconsidered as a DA D3 agonist, it can also be arguedthat the e¡ects found in this study could have been dueto the action at D3 receptors. However, haloperidol, adrug known to block DA D2 receptors, antagonized thequinpirole-induced cortical CAMP and gating reductionin the present study, which suggests that these e¡ects aremediated by DA D2 receptors. Whether NAC DA D3receptors play a role in hippocampal gating remains tobe investigated in future studies.
Comparison between PPI and auditory evokedpotential gating
It has been extensively shown that mesolimbic DA D2receptors are involved in PPI (Reijmers et al., 1995;Swerdlow et al., 1990, 1992; Wan and Swerdlow,1993), another type of gating de¢cit implicated in schiz-ophrenia. When comparing quinpirole e¡ects in theNAC on cortical gating and PPI, it can be proposed
that mesolimbic DA D2 receptors are involved in boththese gating measures. When comparing the quinpirolee¡ects in the NAC on hippocampal gating and PPI, itcan be argued that both gating parameters tend to bedi¡erentially mediated by DA mechanisms, sincethe e¡ects on the hippocampal TC ratio tended to bemore pronounced with quinpirole injections in theNAC shell, whereas quinpirole e¡ects on PPI weremore pronounced when injected in the NAC core (Wanet al., 1994).
CONCLUSION
It can be concluded from the present study thatCAMP and gating in the cortex as well as in the hippo-campus can be reduced when a DA agonist is speci¢callytargeted at mesolimbic DA D2 receptors. However, theDA D2 involvement is more pronounced for corticalthan for hippocampal parameters, since the quinpirole-induced cortical e¡ects are fully antagonized whereas thequinpirole-induced hippocampal e¡ects are not antago-nized by haloperidol.
An important consideration is that the DA e¡ects ongating in the present study are predominantly mediatedby the e¡ects on the amplitude to S1 (or CAMP). Thishas also been suggested for systemic amphetamine injec-tions in rats and for schizophrenic patients. This castsdoubt on whether dopamine receptor activation a¡ectsthe putative inhibitory process between the ¢rst and thesecond stimulus.
AcknowledgementsöThe authors are indebted to Cathy Adams(Ph.D.) and Heidi O'Neill for their assistance.
REFERENCES
Adler, L.E., Pachtman, E., Franks, R.D., Pecevich, M., Waldo, M.C., Freedman, R., 1982. Neurophysiological evidence for a defect in neuronalmechanisms involved in sensory gating in schizophrenia. Biol. Psychiatry 17, 639.
Adler, L.E., Rose, G., Freedman, R., 1986. Neurophysiological studies of sensory gating in rats: e¡ects of amphetamine, phencyclidine, andhaloperidol. Biol. Psychiatry 21, 787^798.
Adler, L.E., Pang, K., Gerhardt, G., Rose, G.M., 1988. Modulation of the gating of auditory evoked potentials by norepinephrine: pharmaco-logical evidence obtained using a selective neurotoxin. Biol. Psychiatry 24, 179^190.
Adler, L.E., Gerhardt, G.A., Franks, R., Baker, N., Nagamoto, H., Drebing, C., Freedman, R., 1990. Sensory physiology and catecholamines inschizophrenia and mania. Psychiatry Res. 31, 297^309.
Adler, L.E., Hope, C., Ho¡ner, L.D., Stephen, C., Young, D., Gerhardt, G., 1994. Bromocriptine impairs P50 auditory sensory gating in normalcontrol subjects. Biol. Psychiatry 35, 630.
Bardgett, M.E., Olney, J.W., Jackson, J.L., Wrona, C.T., Csernansky, J.G., 1993. Connecting hippocampal neuropathy with dopamine dysfunc-tion in schizophrenia: ICV kainic acid as an animal model. Soc. Neurosci. Abstr. 19, 8.
Bardgett, M.E., Jackson, J.L., Taylor, G.T., Csernasky, J.G., 1995. Kainic acid decreases hippocampal neuronal number and increase in dopaminereceptor binding in the nucleus accumbens: an animal model of schizophrenia. Behav. Brain Res. 70, 153^164.
Berger, B., Thierry, A.M., Tassin, J.P., Moyne, M.A., 1976. Dopaminergic innervation of the rat prefrontal cortex: a £uorescence histochemicalstudy. Brain Res. 106, 133^145.
Bickford-Wimer, P.C., Nagamoto, H., Johnson, R., Adler, L.E., Egan, M., Rose, G.M., Freedman, R., 1990. Auditory sensory gating in hippo-campal neurons: a model system in the rat. Biol. Psychiatry 27, 174^182.
Bjo«rklund, A., Lindvall, O., 1984. Dopamine-containing system in the CNS. In: Bjo«rklund, A., Ho«kfelt, O. (Eds.), Handbook of ChemicalNeuroanatomy, Part I: Classical Transmitters in the CNS. Elsevier, Amsterdam, pp. 55^122.
Boutros, N., Zouridakis, G., Overall, J., 1991. Replication and extension of the P50 ¢ndings of the P50 ¢ndings in schizophrenia. Clin. Electro-encephalogr. 22, 40^45.
Carlsson, A., 1988. The current status of the dopamine hypothesis of schizophrenia. Neuropsychopharmacology 1, 197.Cools, A.R., De Beltran, K.K., Prinssen, E.P.M., Koshikawa, N., 1993. Di¡erential role of core and shell of the nucleus accumbens in jaw
movements of rats. Neurosci. Res. Commun. 14, 55^61.Cools, A.R., Miwa, Y., Koshikawa, N., 1995. Role of dopamine D1 and D2 receptors in the nucleus accumbens in jaw movements of rats:
a critical role of the shell. Eur. J. Pharmacol. 286, 41^47.
NSC 5028 16-7-01
N. M. W. J. de Bruin et al.178

Cullum, C.M., Harris, J.G., Waldo, M.C., Smerno¡, E., Madison, A., Nagamoto, H.T., Gri¤th, J., Adler, L.E., Freedman, R., 1993. Neuro-physiological and neuropsychological evidence for attentional dysfunction in schizophrenia. Schizophr. Res. 10, 131^141.
De Bruin, N.M.W.J., Ellenbroek, B.A., Cools, A.R., Coenen, A.M.L., Van Luijtelaar, E.L.J.M., 1999. Di¡erential e¡ects of ketamine on gating ofauditory evoked potentials and prepulse inhibition in rats. Psychopharmacology 142, 9^17.
Descarries, T.W., Lemay, B., Doucet, G., Berger, B., 1987. Regional and laminar density of the dopamine innervation in adult rat cerebral cortex.Neuroscience 21, 807^824.
Freedman, R., Adler, L.E., Waldo, M.C., Pachtman, E., Franks, R.D., 1983. Neurophysiological evidence for a defect in inhibitory pathways inschizophrenia: comparison of medicated and drug-free patients. Biol. Psychiatry 18, 537^551.
Freedman, R., Adler, L.E., Gerhardt, G.A., Waldo, M., Baker, N., Rose, G.M., Drebing, C., Nagamoto, H., Bickford-Wimer, P., Franks, R.,1987. Neurobiological studies of sensory gating in schizophrenia. Schizophr. Bull. 13, 669^678.
Fuxe, K., Agnati, L.F., Kalia, M., Goldstein, M., Andersson, K., Ha«rfstrand, A., 1985. Dopaminergic system in the brain and pituitary. In:Fluckiger, E., Muller, E.E., Thorner, M.O. (Eds.), The Dopaminergic System: Basic and Clinical Aspects of Neuroscience. Springer, Berlin,pp. 11^25.
Gri¡on, N., Sokolo¡, P., Diaz, J., Levesque, D., Sautel, F., Schwartz, J.C., Simon, P., Costentin, J., Garrido, F., Mann, A., Wermuth, C., 1995.The dopamine D-3 receptor and schizophrenia: Pharmacological, anatomical and genetic approaches. Eur. Neuropsychopharmacol. 5 (Suppl.S), 3^9.
Groenewegen, H.J., Berendse, H.W., Haber, S.N., 1993. Organization of the output of the ventral pallidal system in the rat: ventral pallidale¡erents. Neuroscience 57, 113^142.
Groenewegen, H.J., Galis de Graaf, Y., Smeets, W.J., 1999a. Integration and segregation of limbic cortico-striatal loops at the thalamic level : anexperimental tracing study in rats. J. Chem. Neuroanat. 16, 167^185.
Groenewegen, H.J., Wright, C.I., Beijer, A.V., Voorn, P., 1999b. Convergence and segregation of ventral striatal inputs and outputs. Ann. N. Y.Acad. Sci. 877, 49^63.
Heimer, L., Zahm, D.S., Alheid, G.F., 1995. Basal ganglia. In: Paxinos, G. (Ed.), The Rat Nervous System. Academic Press, San Diego, Ch. 23,pp. 579^628.
Ho¡man, D.C., Donovan, H., 1994. D1 and D2 dopamine receptor antagonists reverse PPI de¢cits in an animal model of schizophrenia.Psychopharmacology 115, 447^453.
Jin, Y., Potkin, S.G., Patterson, J.V., Sandman, C.A., Hetrick, W.P., Bunney, W.E., Jr., 1997. E¡ects of P50 temporal variability on sensory gatingin schizophrenia. Psychiatry Res. 70, 71^81.
Johnson, R.G., Stevens, K.E., Rose, G.M., 1998. 5-Hydroxytryptamine-2 receptors modulate auditory ¢ltering in the rat. J. Pharmacol. Exp. Ther.285, 643^650.
Joyce, J.N., MeadorWoodru¡, J.H., 1997. Linking the family of D-2 receptors to neuronal circuits in human brain: Insights into schizophrenia.Neuropsychopharmacology 16, 375^384.
Judd, L.L., McAdams, L., Budnick, N., Bra¡, D.L., 1992. Sensory gating de¢cits in schizophrenia: new results. Am. J. Psychiatry 149, 488^493.Koshikawa, N., Yoshida, Y., Kitamura, M., Saigusa, T., Kobayashi, M., Cools, A.R., 1996. Stimulation of acetylcholine or dopamine receptors in
the nucleus accumbens di¡erentially alters dopamine release in the striatum of freely moving rats. Eur. J. Pharmacol. 303, 13^19.Light, G.A., Malaspina, D., Geyer, M.A., Luber, B.M., Coleman, E.A., Sackeim, H.A., Bra¡, D.L., 1999. Amphetamine disrupts P50 suppression
in normal subjects. Biol. Psychiatry 46, 990^996.Mogenson, G., Jones, D.L., Yim, C.Y., 1982. From motivation to action: functional interface between the limbic system and the motor system.
Prog. Neurobiol. 14, 69^97.Patterson, J.V., Jin, Y., Gierczak, M., Hetrick, W.P., Potkin, S., Bunney, W.E., Sandman, C., 2000. E¡ects of temporal variability on P50 and the
gating ratio in schizophrenia. Arch. Gen. Psychiatry 57, 57^64.Paxinos, G., Watson, C., 1997. The Rat Brain in Stereotaxic Coordinates. Academic Press, London.Peng, R., Mansbach, R.E., Bra¡, D.L., Geyer, M.A., 1990. A D2 dopamine receptor agonist disrupts sensorimotor gating in rats: Implications for
dopaminergic abnormalities in schizophrenia. Neuropsychopharmacology 3, 211^218.Pickar, D., Labarca, R., Linnoila, M., Roy, A., Hommer, D., Everett, D., Paul, S.M., 1984. Neuroleptic-induced decrease in plasma homovanillic
acid and antipsychotic activity in schizophrenic patients. Science 225, 954^957.Poirier, M.F., 1998. Trends in clinical psychiatric research. Presse Med. 27, 2185^2189.Prinssen, E.P.M., Balestra, W., Bemelmans, F.F.J., Cools, A.R., 1994. Evidence for a role of the shell of the nucleus accumbens in oral behavior of
freely moving rats. J. Neurosci. 14, 1555^1562.Ralph, R.J., Varty, G.B., Kelly, M.A., Wang, Y.M., Caron, M.G., Rubinstein, M., Grandy, D.K., Low, M.J., Geyer, M.A., 1999. The dopamine
D2, but not D3 or D4, receptor subtype is essential for the disruption of prepulse inhibition produced by amphetamine in mice. J. Neurosci. 19,4627^4633.
Reijmers, L.G.J.E., Vanderheyden, P.M.L., Peeters, B.W.M.M., 1995. Changes in prepulse inhibition after local administration of NMDAreceptor ligands in the core region of the rat nucleus accumbens. Eur. J. Pharmacol. 272, 131^138.
Schwarzkopf, S.B., Bruno, J.P., Mitra, T., 1993. E¡ects of haloperidol and SCH23390 on acoustic startle and prepulse inhibition under basal andstimulated conditions. Prog. Neuropsychopharmacol. Biol. Psychiatry 17, 1023^1036.
Snyder, S.H., 1976. The dopamine hypothesis of schizophrenia: focus on the dopamine receptor. Am. J. Psychiatry 133, 198.Stevens, K.E., Fuller, L.L., Rose, G.M., 1991. Dopaminergic and noradrenergic modulation of amphetamine-induced changes in auditory gating.
Brain Res. 555, 91^98.Stevens, K.E., Meltzer, J., Rose, G.M., 1995. Nicotinic cholinergic normalization of amphetamine-induced loss of auditory gating in freely moving
rats. Psychopharmacology 119, 163^170.Stevens, K.E., Luthman, J., Lindqvist, E., Jonhson, R.G., Rose, G.M., 1996. E¡ects of neonatal dopamine depletion on sensory inhibition in the
rat. Pharmacol. Biochem. Behav. 53, 817^823.Swanson, L.W., 1982. The projections of the ventral tegmental area and adjacent regions: A combined £uorescent retrograde tracer and immu-
no£uorescence study in the rat. Brain Res. Bull. 9, 321^353.Swerdlow, N.R., Bra¡, D.L., Masten, V.L., Geyer, M.A., 1990. Schizophrenia-like sensorimotor gating abnormalities in rats following dopamine
infusion into the nucleus accumbens. Psychopharmacology 101, 414^420.Swerdlow, N.R., Keith, V.A., Bra¡, D.L., Geyer, M.A., 1991. E¡ects of spiperone, raclopride, SCH 23390 and clozapine on apomorphine
inhibition of sensorimotor gating of the startle response in the rat. J. Pharmacol. Exp. Ther. 256, 230^556.Swerdlow, N.R., Caine, S.B., Geyer, M.A., 1992. Regionally selective e¡ects of intracerebral dopamine infusion on sensorimotor gating of the
startle re£ex in rats. Psychopharmacology 108, 189^195.Waldo, M.C., Cawthry, E., Adler, L.E., Dubester, S., Staunton, M., Nagamoto, H., Baker, N., Madison, A., Simon, J., Scherzinger, A., Drebing,
C., Gerhardt, G., Freedman, R., 1994. Auditory sensory gating, hippocampal volume and catecholamine metabolism in schizophrenics andtheir siblings. Schizophr. Res. 12, 93^106.
NSC 5028 16-7-01
Nucleus accumbens dopamine and sensory gating 179

Wan, F.J., Swerdlow, N.R., 1993. Intra-accumbens infusion of quinpirole impairs sensorimotor gating of acoustic startle in rats. Psychopharma-cology 113, 103^109.
Wan, F.J., Geyer, M.A., Swerdlow, N.R., 1994. Accumbens D2 modulation of sensorimotor gating in rats: Assessing anatomical localization.Pharmacol. Biochem. Behav. 49, 155^163.
Young, A.M.J., Ahier, R.G., Upton, R.L., Joseph, M.H., Gray, J.A., 1998. Increased extracellular dopamine in the nucleus accumbens of the ratduring associative learning of neutral stimuli. Neuroscience 83, 1175^1183.
Zahm, D.S., Brog, J.S., 1992. On the signi¢cance of subterritories in the accumbens part of the ventral striatum. Neuroscience 50, 751^767.Zouridakis, G., Boutros, N.N., Jansen, B.H., 1997. A fuzzy clustering approach to study the auditory P50 component in schizophrenia. Psychiatry
Res. 69, 169^181.
(Accepted 17 April 2001)
NSC 5028 16-7-01
N. M. W. J. de Bruin et al.180










![Dopamine release in nucleus accumbens during rewarded task switching measured by [¹¹C]raclopride](https://static.fdokumen.com/doc/165x107/633ae2b3ea31206fd207b694/dopamine-release-in-nucleus-accumbens-during-rewarded-task-switching-measured-by.jpg)









