
Clines in clock genes: fine-tuning circadian rhythms to the environment
9
Clines in clock genes: fine-tuning circadian rhythms to the environment Charalambos P. Kyriacou 1 , Alexandre A. Peixoto 2 , Federica Sandrelli 3 , Rodolfo Costa 3 and Eran Tauber 1 1 Department of Genetics, University of Leicester, Leicester LE1 7RH, UK 2 Laborato ´ rio de Biologia Molecular de Insetos, IOC, Fundacao Oswaldo Cruz, Rio de Janeiro CEP 21040-900, Brazil 3 Department of Biology, University of Padova, 35131 Padova, Italy The dissection of the circadian clock into its molecular components represents the most striking and well-stu- died example of a gene regulatory network underlying a complex behavioural trait. By contrast, the evolutionary analysis of the clock has developed more slowly. Here we review studies that have surveyed intraspecific clock gene variation over large geographical areas and have discovered latitudinal clines in gene frequencies. Such spatial patterns traditionally suggest that natural selec- tion shapes genetic variation, but it is equally possible that population history, or a mixture of demography and selection, could contribute to the clines. We discuss how population genetics, together with functional assays, can illuminate these possible cases of natural selection in Drosophila clock genes. Evolution of the circadian clock The genetic analysis of the endogenous circadian, 24-h clock represents a spectacular example of the molecular dissection of a gene regulatory network underlying a com- plex behavioural trait [1]. Recent transcriptome [2] and proteome studies [3] in several model organisms have shown how a large proportion of both ‘omes’ cycles in abundance with 24-h periods, revealing that most bio- logical processeshave a rhythmic component. This reflects the four billion year evolution of life on a rotating planet with its cycles of day and night and warmth and cold. Phylogenetic analysis of clock genes in Cyanobacteria, one of the earliest photosynthetic organisms, reveals that the kaiC gene might have originated 3000 million years ago (Mya) [4]. The molecular analysis of clocks proceeds at an unprecedented pace, with most studies focusing on identi- fying new clock genes, dissecting clock gene networks and investigating how the clock works at molecular, bio- chemical and physiological levels [5–8] (see Figure I in Box 1). How has the circadian clock evolved? The different model organisms studied by circadian biologists provide the immediate comparative perspective to clock research, and this is buttressed by further studies in non-model organisms. For example, in bees and butterflies, clock genes not only promote circadian functions but are also involved in related processes such as the development of social behaviour and navigation, respectively [9,10]. Furthermore, studies of intraspecific clock gene variation can reveal whether natural selection is involved in maintaining these polymorphisms. [11]. Within this con- text, geographical surveys of genetic variation are useful, particularly if an association with latitude is observed [12,13]. Such ‘clines’ (see Glossary) indicate that environ- mental factors might shape the genetic variation at that particular locus, although it can become difficult to disen- tangle the effects of selection from population history [14– 16]. This review focuses on the geographical variation in clock genes, particularly in Drosophila melanogaster, where we are beginning to understand the evolutionary and functional implications of such spatial patterns. Environmental challenges to the circadian clock The molecular basis for the circadian clock is similar between flies and mice: several conserved positive and negative regulatory molecules are shared, most of which are rhythmically expressed and form a feedback loop (see Figure I in Box 1) [6,8]. Obligatory delays in the feedback Review Glossary Balancing selection: Selection for more than one allele at a locus at frequencies too high to be caused by recurrent mutation. Circadian photosensitivity: The sensitivity of the circadian clock to light, which is usually studied by giving a brief light pulse to an organism that is free- running in darkness. A light pulse early at night gives a phase delay (usually of a few hours) in the circadian cycle, whereas a pulse late at night generates an advance. The size of the advances and delays provides a measure of the light sensitivity of the clock. As an example, in D. melanogaster, ls-tim gives smaller phase delays and advances than s-tim [51]. Clines: A gradual change in a character over the geographical range of a species or population. Latitudinal clines are often associated with correlated changes in environmental variables. Darwinian fitness: The capacity of an individual to reproduce, which can be measured as the relative proportion of that individual’s genes in the genes of succeeding generations. Directional selection: Selection for a phenotype at an extreme of the phenotypic range, or in the case of a specific allele, selection for that allele over alternative alleles because it determines a more extreme phenotype. Effective population size: The average number of individuals in a population that contribute to the gene pool of succeeding generations. Entrain: An organism placed in an environmental cycle, for example 12 h of light, 12 h of dark (LD12:12), will display an ‘entrained’ circadian rhythm to the imposed 24-h oscillation, which usually differs from its ‘free-running’ pattern. Free-running: An organism that is entrained to a light-dark cycle and then placed in constant conditions (for animals, usually constant darkness, DD) will display its own endogenous free-running rhythm with a period of 24 h. Resonance: The free-running period of most organisms is close to 24 h, thereby ‘resonating’ with the environmental cycle. Selection coefficient: A relative measure of the rate of transmission of a given allele compared with another allele, usually the wild-type. Temperature compensation: The ability of the circadian clock to keep a stable 24-h period under different temperatures. Corresponding author: Kyriacou, C.P. ([email protected]). 124 0168-9525/$ – see front matter ß 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.tig.2007.12.003
Transcript of Clines in clock genes: fine-tuning circadian rhythms to the environment

Review
Clines in clock genes: fine-tuningcircadian rhythms to the environmentCharalambos P. Kyriacou1, Alexandre A. Peixoto2, Federica Sandrelli3, Rodolfo Costa3
and Eran Tauber1
1 Department of Genetics, University of Leicester, Leicester LE1 7RH, UK2 Laboratorio de Biologia Molecular de Insetos, IOC, Fundacao Oswaldo Cruz, Rio de Janeiro CEP 21040-900, Brazil3 Department of Biology, University of Padova, 35131 Padova, Italy
Glossary
Balancing selection: Selection for more than one allele at a locus at frequencies
too high to be caused by recurrent mutation.
Circadian photosensitivity: The sensitivity of the circadian clock to light, which
is usually studied by giving a brief light pulse to an organism that is free-
running in darkness. A light pulse early at night gives a phase delay (usually of
a few hours) in the circadian cycle, whereas a pulse late at night generates an
advance. The size of the advances and delays provides a measure of the light
sensitivity of the clock. As an example, in D. melanogaster, ls-tim gives smaller
phase delays and advances than s-tim [51].
Clines: A gradual change in a character over the geographical range of a
species or population. Latitudinal clines are often associated with correlated
changes in environmental variables.
Darwinian fitness: The capacity of an individual to reproduce, which can be
measured as the relative proportion of that individual’s genes in the genes of
succeeding generations.
Directional selection: Selection for a phenotype at an extreme of the
phenotypic range, or in the case of a specific allele, selection for that allele
over alternative alleles because it determines a more extreme phenotype.
Effective population size: The average number of individuals in a population
that contribute to the gene pool of succeeding generations.
Entrain: An organism placed in an environmental cycle, for example 12 h of
light, 12 h of dark (LD12:12), will display an ‘entrained’ circadian rhythm to the
imposed 24-h oscillation, which usually differs from its ‘free-running’ pattern.
Free-running: An organism that is entrained to a light-dark cycle and then
placed in constant conditions (for animals, usually constant darkness, DD) will
display its own endogenous free-running rhythm with a period of �24 h.
Resonance: The free-running period of most organisms is close to 24 h,
thereby ‘resonating’ with the environmental cycle.
Selection coefficient: A relative measure of the rate of transmission of a given
allele compared with another allele, usually the wild-type.
Temperature compensation: The ability of the circadian clock to keep a stable
The dissection of the circadian clock into its molecularcomponents represents the most striking and well-stu-died example of a gene regulatory network underlying acomplex behavioural trait. By contrast, the evolutionaryanalysis of the clock has developed more slowly. Herewe review studies that have surveyed intraspecific clockgene variation over large geographical areas and havediscovered latitudinal clines in gene frequencies. Suchspatial patterns traditionally suggest that natural selec-tion shapes genetic variation, but it is equally possiblethat population history, or a mixture of demography andselection, could contribute to the clines. We discuss howpopulation genetics, together with functional assays,can illuminate these possible cases of natural selectionin Drosophila clock genes.
Evolution of the circadian clockThe genetic analysis of the endogenous circadian, 24-hclock represents a spectacular example of the moleculardissection of a gene regulatory network underlying a com-plex behavioural trait [1]. Recent transcriptome [2] andproteome studies [3] in several model organisms haveshown how a large proportion of both ‘omes’ cycles inabundance with 24-h periods, revealing that most bio-logical processeshave a rhythmic component. This reflectsthe four billion year evolution of life on a rotating planetwith its cycles of day and night and warmth and cold.Phylogenetic analysis of clock genes in Cyanobacteria, oneof the earliest photosynthetic organisms, reveals that thekaiC gene might have originated �3000 million years ago(Mya) [4]. The molecular analysis of clocks proceeds at anunprecedented pace, with most studies focusing on identi-fying new clock genes, dissecting clock gene networks andinvestigating how the clock works at molecular, bio-chemical and physiological levels [5–8] (see Figure I inBox 1).
How has the circadian clock evolved? The differentmodel organisms studied by circadian biologists providethe immediate comparative perspective to clock research,and this is buttressed by further studies in non-modelorganisms. For example, in bees and butterflies, clockgenes not only promote circadian functions but are alsoinvolved in related processes such as the development ofsocial behaviour and navigation, respectively [9,10].Furthermore, studies of intraspecific clock gene variation
Corresponding author: Kyriacou, C.P. ([email protected]).
124 0168-9525/
can reveal whether natural selection is involved inmaintaining these polymorphisms. [11]. Within this con-text, geographical surveys of genetic variation are useful,particularly if an association with latitude is observed[12,13]. Such ‘clines’ (see Glossary) indicate that environ-mental factors might shape the genetic variation at thatparticular locus, although it can become difficult to disen-tangle the effects of selection from population history [14–16]. This review focuses on the geographical variation inclock genes, particularly in Drosophila melanogaster,where we are beginning to understand the evolutionaryand functional implications of such spatial patterns.
Environmental challenges to the circadian clockThe molecular basis for the circadian clock is similarbetween flies and mice: several conserved positive andnegative regulatory molecules are shared, most of whichare rhythmically expressed and form a feedback loop (seeFigure I in Box 1) [6,8]. Obligatory delays in the feedback
�24-h period under different temperatures.
$ – see front matter � 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.tig.2007.12.003

Box 1. The molecular circadian clock in D. melanogaster
The D. melanogaster circadian clock is generated by a series of
negative and positive feedback loops (Figure I). The expression of
the negative transcriptional regulators TIMELESS (TIM) and
PERIOD (PER), which make up the PER–TIM loop, and CLOCKWORK
ORANGE (CWO) is mediated through binding of their E-boxes
(hexameric sequence CACGTG) by the positive transcription
factors CLOCK (CLK) and CYCLE (CYC), which form a heterodimer
[6,68]. The negative regulators feed back, after a time delay, to
inhibit CLK and CYC activity. These delays are imposed by the
kinases SHAGGY (SGG), casein kinase 2 (CK2) and DOUBLETIME
(DBT) [8] and by phosphatases including protein phosphatases 2A
(PP2A) and 1 (PP1), which modulate the stability of the negative
regulators [6,8,69]. A second feedback loop interlocks with the
PER–TIM loop; Clk negative regulation by VRILLE (VRI) and positive
regulation by PAR domain protein 1e (PDP1e) [6]. However, more
recent data have shed some doubt on the role of PDP1e in this
process [70].
Cytoplasmic PER and TIM levels start to rise at the beginning of
the night, and late at night PER and TIM translocate to the
nucleus where they negatively regulate (through CLOCK-CYCLE)
their own genes. At dawn, the blue-light circadian photoreceptor
cryptochrome (CRY) is activated and interacts with TIM. This
interaction stimulates TIM phosphorylation, which recruits the F-
box protein JETLAG (JET), a component of a SKP1-CULLIN1-F-
box (SCF)-E3 ubiquitin ligase, which polyubiquitylates TIM,
thereby marking it for degradation [8]. Under conditions of
constant darkness, TIM is degraded by the proteasome at
subjective dawn by a CRY-independent pathway. PER monomers
are strong transcriptional repressors, but in the absence of TIM,
PER is vulnerable to phosphorylation by DBT kinase, and
phosphorylated PER is recognized by the F-box protein SUPER-
NUMARY-LIMBS (SLIMB) and marked for proteasomal degrada-
tion [6,8]. The negative regulation of per and tim is now lifted,
and CLK–CYC begins the cycle again by binding the per and tim
E-boxes. In the brain, the PER and TIM proteins are expressed in
�75 neurons on each side of the brain, as well as in glia and
photoreceptors [71]. Of the two main groups of neurons, the
dorsal and the lateral neurons, the lateral neurons are indis-
pensable for rhythmic behavioural expression [72,73]. However,
many peripheral tissues of the fly are also rhythmic [74], and in
peripheral tissues, CRY seems to function as a clock component
and a photoreceptor [75].
Figure I. Simplified view of the molecular circadian clock in Drosophila. The
expression of negative regulators TIMELESS (TIM, blue), PERIOD (PER, red)
and CLOCKWORK ORANGE (CWO, orange) is mediated by positive
transcription factors CLOCK (CLK, pink) and CYCLE (CYC, purple) binding to
E-boxes (pale blue) on their promoters. The kinases SGG, CK2 and DBT and
phosphatases such as PP2A and PP1, which modulate the stabilities of the
negative regulators are also shown. The second negative feedback loop that
interlocks with the PER–TIM loop has VRILLE (VRI, green) negatively and PDP1e(brown) positively regulating Clock (but see Box 1 for the role of PDP1e). The
circadian photoreceptor CRY is shown in yellow.
Review Trends in Genetics Vol.24 No.3
mechanisms are induced by modulating the stability ofthese canonical clock proteins through kinases, phospha-tases and the cellular degradation machinery [8,17]. Theenvironmental inputs that ‘entrain’ the circadian clockinclude light, temperature, social stimuli and food avail-ability, but light is the most potent ‘Zeitgeber’ (time-giver)[1]. Consequently, latitude differences challenge the clockbecause of associated changes in temperature and photo-period. For example, in light–dark cycles, D. melanogasteradults show a morning and late-afternoon/evening burst ofmovement. Under hotter conditions, evening activity isdelayed, so flies avoid desiccation [18]. This is achievedby repressing the splicing of a 30 period (per) intron,resulting in a delayed increase in per transcript andPER protein levels (Box 1) and a retardation of eveninglocomotion (with a longer afternoon ‘siesta’) [18]. Wemighttherefore expect higher levels of intron retention and lesssplicing in the lower latitudes of both hemispheres, wheretemperature is generally greater. Such amechanismwouldallow the fly to adjust its behaviour in response to changingtemperature. Clock gene polymorphisms provide anothermechanism by which flies can adapt to different environ-ments. Here we discuss two well-understood examples ofpolymorphism in D. melanogaster.
The period Thr-Gly polymorphismAn uninterrupted stretch of alternating threonine-glycine(Thr-Gly) pairs is encoded within the per gene (Figure 1a).D. melanogaster natural populations in Europe and NorthAfrica differ in the length of Thr-Gly region: two majoralleles, per (Thr-Gly)17 and per (Thr-Gly)20, account for�90% of the variation, per (Thr-Gly)23 making up �8%,and per (Thr-Gly)14 at �1% (Figure 1b) [12]. Rare allelesencoding 15, 18, 21 and 24 Thr-Gly pairs constitute thefinal 1%, and each allele can be derived from another bysimple insertions and deletions, with all variants coales-cing back to a per (Thr-Gly)23 sequence [12,19]. In northernEurope, the frequency of per (Thr-Gly)20 increases,whereas that of per (Thr-Gly)17 decreases, generating arobust latitudinal cline (Figure 1b) [12]. A parallel cline inper (Thr-Gly)20 frequency is observed in Australia,although it is neither as steep nor as robust as in Europe.Furthermore, it also seems to be affected by yearly localgenetic drift or climatic changes (Figure 1b) [20–23].Nevertheless, the observation of similar clines in twocontinents suggests the action of selection rather thancoincidentally similar historical or migratory events. Dis-equilibrium pattern analysis, a technique for detectingselection among tightly linked loci, of European and NorthAfrican per sequences suggested that balancing selectionacts on the Thr-Gly region [24].
Functional analysis of Thr-Gly repeat length variationThe ability of the circadian oscillator to maintain 24-hperiods over a wide range of temperatures was examinedin a large sample ofD. melanogaster per (Thr-Gly) variantsfrom Europe and North Africa [25]. The northern per (Thr-Gly)20 allele provided the most thermally stable periods,with an endogenous period that barely deviated from�15–20 min less than 24 h (Figure 1c). The per (Thr-Gly)17allele provided a more precise 24-h free-running period
125
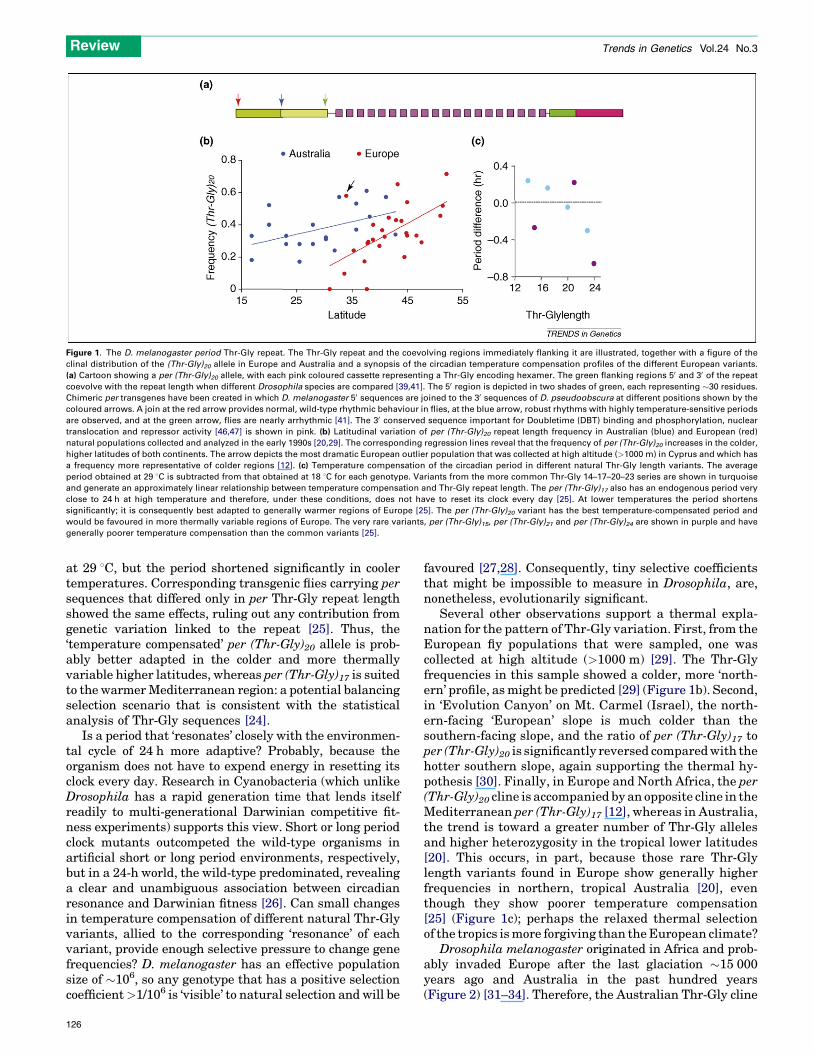
Figure 1. The D. melanogaster period Thr-Gly repeat. The Thr-Gly repeat and the coevolving regions immediately flanking it are illustrated, together with a figure of the
clinal distribution of the (Thr-Gly)20 allele in Europe and Australia and a synopsis of the circadian temperature compensation profiles of the different European variants.
(a) Cartoon showing a per (Thr-Gly)20 allele, with each pink coloured cassette representing a Thr-Gly encoding hexamer. The green flanking regions 50 and 30 of the repeat
coevolve with the repeat length when different Drosophila species are compared [39,41]. The 50 region is depicted in two shades of green, each representing �30 residues.
Chimeric per transgenes have been created in which D. melanogaster 50 sequences are joined to the 30 sequences of D. pseudoobscura at different positions shown by the
coloured arrows. A join at the red arrow provides normal, wild-type rhythmic behaviour in flies, at the blue arrow, robust rhythms with highly temperature-sensitive periods
are observed, and at the green arrow, flies are nearly arrhythmic [41]. The 30 conserved sequence important for Doubletime (DBT) binding and phosphorylation, nuclear
translocation and repressor activity [46,47] is shown in pink. (b) Latitudinal variation of per (Thr-Gly)20 repeat length frequency in Australian (blue) and European (red)
natural populations collected and analyzed in the early 1990s [20,29]. The corresponding regression lines reveal that the frequency of per (Thr-Gly)20 increases in the colder,
higher latitudes of both continents. The arrow depicts the most dramatic European outlier population that was collected at high altitude (>1000 m) in Cyprus and which has
a frequency more representative of colder regions [12]. (c) Temperature compensation of the circadian period in different natural Thr-Gly length variants. The average
period obtained at 29 8C is subtracted from that obtained at 18 8C for each genotype. Variants from the more common Thr-Gly 14–17–20–23 series are shown in turquoise
and generate an approximately linear relationship between temperature compensation and Thr-Gly repeat length. The per (Thr-Gly)17 also has an endogenous period very
close to 24 h at high temperature and therefore, under these conditions, does not have to reset its clock every day [25]. At lower temperatures the period shortens
significantly; it is consequently best adapted to generally warmer regions of Europe [25]. The per (Thr-Gly)20 variant has the best temperature-compensated period and
would be favoured in more thermally variable regions of Europe. The very rare variants, per (Thr-Gly)15, per (Thr-Gly)21 and per (Thr-Gly)24 are shown in purple and have
generally poorer temperature compensation than the common variants [25].
Review Trends in Genetics Vol.24 No.3
at 29 8C, but the period shortened significantly in coolertemperatures. Corresponding transgenic flies carrying persequences that differed only in per Thr-Gly repeat lengthshowed the same effects, ruling out any contribution fromgenetic variation linked to the repeat [25]. Thus, the‘temperature compensated’ per (Thr-Gly)20 allele is prob-ably better adapted in the colder and more thermallyvariable higher latitudes, whereas per (Thr-Gly)17 is suitedto the warmerMediterranean region: a potential balancingselection scenario that is consistent with the statisticalanalysis of Thr-Gly sequences [24].
Is a period that ‘resonates’ closely with the environmen-tal cycle of 24 h more adaptive? Probably, because theorganism does not have to expend energy in resetting itsclock every day. Research in Cyanobacteria (which unlikeDrosophila has a rapid generation time that lends itselfreadily to multi-generational Darwinian competitive fit-ness experiments) supports this view. Short or long periodclock mutants outcompeted the wild-type organisms inartificial short or long period environments, respectively,but in a 24-h world, the wild-type predominated, revealinga clear and unambiguous association between circadianresonance and Darwinian fitness [26]. Can small changesin temperature compensation of different natural Thr-Glyvariants, allied to the corresponding ‘resonance’ of eachvariant, provide enough selective pressure to change genefrequencies? D. melanogaster has an effective populationsize of �106, so any genotype that has a positive selectioncoefficient>1/106 is ‘visible’ to natural selection and will be
126
favoured [27,28]. Consequently, tiny selective coefficientsthat might be impossible to measure in Drosophila, are,nonetheless, evolutionarily significant.
Several other observations support a thermal expla-nation for the pattern of Thr-Gly variation. First, from theEuropean fly populations that were sampled, one wascollected at high altitude (>1000 m) [29]. The Thr-Glyfrequencies in this sample showed a colder, more ‘north-ern’ profile, asmight be predicted [29] (Figure 1b). Second,in ‘Evolution Canyon’ on Mt. Carmel (Israel), the north-ern-facing ‘European’ slope is much colder than thesouthern-facing slope, and the ratio of per (Thr-Gly)17 toper (Thr-Gly)20 is significantly reversed comparedwith thehotter southern slope, again supporting the thermal hy-pothesis [30]. Finally, in Europe and North Africa, the per(Thr-Gly)20 cline is accompanied by an opposite cline in theMediterranean per (Thr-Gly)17 [12], whereas in Australia,the trend is toward a greater number of Thr-Gly allelesand higher heterozygosity in the tropical lower latitudes[20]. This occurs, in part, because those rare Thr-Glylength variants found in Europe show generally higherfrequencies in northern, tropical Australia [20], eventhough they show poorer temperature compensation[25] (Figure 1c); perhaps the relaxed thermal selectionof the tropics ismore forgiving than theEuropean climate?
Drosophila melanogaster originated in Africa and prob-ably invaded Europe after the last glaciation �15 000years ago and Australia in the past hundred years(Figure 2) [31–34]. Therefore, the Australian Thr-Gly cline

Figure 2. Drosophila melanogaster colonization of Europe, Asia and the New Worlds from Africa. Ancestral populations are shown in red and orange, ancient populations
in green and more recent dispersal in the past few centuries in blue. White areas show regions that are either inhospitable to D. melanogaster or areas where flies have not
been reported. D. melanogaster probably invaded Europe at the end of the last glaciation, �15 000 years ago, and have spread to the Americas and to Australia since the
colonization by Europeans in the past few centuries. Flies in the Caribbean islands have genetic variation that seems to have originated from West Africa, most likely
through the slave trade. Figure redrawn from David and Capy (1998) [31].
Review Trends in Genetics Vol.24 No.3
is much younger than the European cline, suggesting thatselection on the Thr-Gly polymorphism is not as strong ason loci such as alcohol dehydrogensase (Adh) or esterase-6(Est6) that show robust Australian clines [35,36]. Popu-lations from ‘ancestral’ east Africa show enormous vari-ation in Thr-Gly length, and their sequences do notconform to the major European classes of 14–17–20–23repeats [20]. Nevertheless, all Thr-Gly sequences coalesceback to the same per (Thr-Gly)23 allele present in the threecontinents. This variation in sub-Saharan Thr-Gly allelesmight also reflect the relaxation of thermal selection inthese tropical environments allied to a large ancestralpopulation size. As with many different fly genes, a con-siderable bottleneck and loss of variation occurred duringthe colonization of Europe and the New Worlds (Figure 3)[14,15,37,38].
Interspecific analysis of Thr-Gly regionsThere is considerable variation in length of the Thr-Glyregion amongDrosophila species. [39]. InD. pseudoobscura,the basic Thr-Gly repeat has been replaced with�35 copiesof a related degenerate pentapeptide repeat, increasing thelength of the repetitive region to �200 residues comparedwith D. melanogaster’s �50 [40]. Statistical analyses ofrepeat region length variation among 20 Drosophilid andnon-Drosophilid flies suggested that repeat length coe-volved with coding changes in the immediate 50 and 30
flanking nonrepetitive regions [39,40]. This was confirmedexperimentally by using interspecific chimeric per trans-genes between D. melanogaster and D. pseudoobscura(Figure 1a). Depending on the position of the boundarybetween the two species’ coding regions, it is possible toproduce both completely arrhythmic transformants
and robustly rhythmic transformants with extremelytemperature-sensitive periods (Figure 1a) [41]. These inter-specific results may explain why changes in Thr-Gly lengthwithinD.melanogaster gently alter temperature responses,because here, these length changes are not associated withany compensating flanking alterations [25].
A similar argument has been proposed to explain thetemperature compensation of the three major D. simulansThr-Gly variants, persim (Thr-Gly)23, persim (Thr-Gly)24 andpersim (Thr-Gly)25, which each have a different flankinghaplotype [42,43]. In Europe, these three variants aredistributed in the ratio 2:2:1, respectively, but not in aclinal pattern [42]. The temperature compensation of per-
sim (Thr-Gly)25 is more stable than that of the other var-iants, but the period of persim (Thr-Gly)25 is �45 minshorter than 24 h, in contrast to the other variants whoseperiod ‘resonates’ better with 24 h but is more tempera-ture-sensitive [43]. This provides another possible balan-cing selection scenario in which the persim (Thr-Gly)25might have the advantage of a better compensated clockbut the disadvantage of a period offset from 24 h. Neutral-ity tests revealed a balancing selection signature in thisregion [42]. Intriguingly, a natural D. simulans chimericvariant from Africa, carrying the per (Thr-Gly)24 repeatwith a per (Thr-Gly)23 flanking haplotype, was found tosegregate with a complex temperature-sensitive longperiod phenotype [44]. This phenotype mapped to gene(s)on the X chromosome and was modified by factor(s) on theautosomes [44]. Thus, disturbing the usual linkage dise-quilibrium between the repeat and its immediate flankingregion in D. simulans provided a ‘sensitized’ circadianbackground, reminiscent of the interspecific chimericgenes described above [41].
127

Figure 3. The timeless polymorphism. The 50 tim sequence in which the G insertion that gives rise to the natural ls-tim variant is illustratedn, together with the spatial
patterns of ls-tim in Europe. The diapause of ls-tim and s-tim females from southern and northern European populations are shown, as is the strength of the molecular
interactions between L- and S-TIM with CRYPTOCROME in yeast. (a) The ls-tim sequence is shown in blue, ATG start codons in red, and the G insertion (black) is indicated
by an arrow [50]. Both L-TIM1421 and S-TIM1398 isoforms are produced [51]. In the s-tim sequence (green), the G deletion is denoted by an asterisk and hence a premature
stop codon, TGA (red), is produced, potentially generating a 19-residue peptide, S-TIM19 (bold, italics) from the upstream ATG, hence the major product from s-tim is the
shorter S-TIM1398 isoform. The amino acid sequences are shown beneath the gene sequences. (b) The latitudinal cline in ls-tim frequency (blue) in Europe, from
southeastern Italy to Sweden. The two outlier populations are from Crete and Haifa, Israel. If the ls-tim frequencies are plotted as an overland distance from southeastern
Italy, a linear relationship is generated. Redrawn from Tauber et al., (2007) [13]. (c) Diapause incidence of ls-tim (blue) and s-tim (green) females at different photoperiods at
13 8C from a southern Italian (South) and a Dutch (North) population. The y-axis represents the proportion of females (arcsin transformed, degrees) in diapause. Adapted,
with permission, from Tauber et al., (2007) [13]. (d) In light conditions, CRY interacts more strongly with S-TIM (note darker blue colonies in plate assay) than L-TIM in the
yeast two hybrid system [51]. (Figure provided by F. Sandrelli).
Review Trends in Genetics Vol.24 No.3
Possible mechanisms for Thr-Gly circadianmodulationA PER-LACZ fusion protein (in which the central 230residue fragment containing the Thr-Gly repeat and itscoevolved regions was removed) is hypophosphorylatedand stable compared with a protein with an intact centralregion [45]. The key sequence within this fragment is ashort motif that binds Doubletime (DBT) kinase and isconserved among flies and mammals (Box 1; Figure 1a). Adeletion of this motif generates behaviourally arrhythmictransformants [46,47]. This sequence lies immediatelydownstream of the 30 coevolved region that flanks theThr-Gly repeat [41]; therefore, the phosphorylation,nuclear localization and repressor activities associatedwith this fragment might be modulated by modificationsto the Thr-Gly region (Figure 1a). In addition, Nuclearmagnetic resonance (NMR) studies of poly(Thr-Gly)peptides revealed that (Thr-Gly)3 has a b-turn
128
conformation, as does D. pseudoobscura’s correspondingpentapeptide repeat [48,49]. This (Thr-Gly)3 conformation-al monomer is particularly interesting because the majorallelic European variants are in a (Thr-Gly)3 interval series(14–17–20–23) that have a linear relationship withtemperature compensation (Figure 1c). The rarer variantsnot included in this series have a poorer thermal response,which does not follow a predictable linear thermal pattern(Figure 1c) [25]. The major variants are not divisible by(Thr-Gly)3 because the degenerate Thr-Gly repeats oneither side of the uninterrupted repeat also contribute tothe b-turn conformation [40,48].
Natural polymorphisms in timeless
The 50 coding region of tim in D. melanogaster is charac-terized by a single nucleotide insertion (Figure 3a) [50].The insertion haplotype (ls-tim) has two in-frame initiationcodons, producing a long TIM isoform from the upstream

Review Trends in Genetics Vol.24 No.3
methionine (L-TIM1421) and a shorter isoform from thedownstream ATG. By contrast, the major products of thedeletion haplotype is the short isoform, S-TIM1398, and aputative truncated 19-residue peptide, S-TIM19, from theupstream ATG (Figure 3a) [51]. Natural EuropeanD. melanogaster populations show a latitudinal cline forthis polymorphism, with ls-tim being most frequent insoutheastern Italy, whereas s-tim prevails in northernEurope (Figure 3b) [13]. A battery of neutrality tests wereperformed on the tim polymorphism and on two intergenicregions�15- and 30-kb downstream. The null hypothesis ofneutrality was rejected for the tim polymorphic region, butnot for the two downstreamregions [13]. In addition, the lowlevel of genetic variation in the polymorphic region com-pared with the downstream regions suggested that the 50
polymorphism is driven by directional rather than balan-cing selection. Although a bottleneck might give a similarresult, we would expect it to affect all nearby sites equally,and this was not the case in the neighbouring 30 loci [13].
Phylogenetic analysis, rooted and calibrated with theorthologousD. simulans sequence, revealed that ls-timwasa recent mutation �10 000 years old [13] and thereforeyounger than the 15 000 years since the last glaciation,when D. melanogaster likely colonized Europe (Figure 2)[31–34]. This ‘modern’ view is reinforced because the ls-timmutation has not been identified in a survey of naturalpopulations from the ancestral homelands of sub-SaharanEast Africa [13]. However, if ls-tim is so recently derived,enough time might not have elapsed for new neutral poly-morphisms to become linked to ls-tim, a requirement forsome neutrality tests to detect the signature of balancingselection [16].
Functional implications of the tim polymorphism:circadian and photoperiodic phenotypesUnlike theperpolymorphism, the tim variants donot showany systematic differences in temperature compensation[13]. Because TIM is the light-sensitive clock molecule(Box 1), seasonal day length,which co-varieswith latitude,is a potential selective agent. In light pulse experiments,circadian photosensitivity is compromised in homozygousls-timflies, and this findingwas recapitulated using trans-formants that mimicked the natural variants (i.e. expres-sing only S-TIM or LS-TIM) [51]. Transformantsexpressing only L-TIM showed a more severe defect thanls-tim flies [51]. These transgenic experiments also ruledout the possibility of linkage disequilibrium contributingto the attenuated circadian photosensitivity phenotype, aswell as of any major role for S-TIM19 in these phenotypes.Changes in circadian photosensitivity have been docu-mented in D. auraria Japanese populations, where fliesfrom the northern islands (438N) have a reduced circadianlight response compared with those in the southernislands (348N) [52]. This cline was interpreted as beingan adaptation of the circadian system to the long summerday lengths that can disrupt circadian behaviour andphysiology in the higher latitudes [52]. For example, inLeicester (528N) on June 21–22, 2007, we measured theabsolute light-dark (LD) cycle as 19.5 h light, 4.5 h dark,whereas on the equator, the photoperiod would be LD12:12. It is therefore puzzling that ls-tim, whose circadian
photosensitivity is attenuated, predominates in southernrather than northern Europe [13].
Like many insects in temperate regions, Drosophilaanticipate the oncoming winter; under short photoperiodsand colder temperatures, females respond with a protec-tive physiological diapause, which includes an ovariandevelopmental arrest. Studies of different insect species,including Drosophilids, have, as expected, revealed thatpopulations from higher latitudes show enhanced levels ofdiapause [53]. D. melanogaster lines with high levels ofdiapause are more resistant to environmental stressessuch as cold or starvation and display enhancedfitness under these conditions [54–56]. Natural ls-timfemales (and their respective transgenics), show signifi-cantly higher diapause levels even under relatively longday lengths than do s-tim flies, reflecting again an atte-nuated photoresponse (Figure 3c) [13]. These results, likethose on circadian photosensitivity, do not fit with thespatial distribution of these alleles, because the southernls-tim variants would seem better adapted to northernconditions. However, when the diapause of northern andsouthern European populations is compared (irrespectiveof tim haplotype), the northern population has a generallyenhanced response (Figure 3c) [13], suggesting the possib-ility of at least a shallow cline in diapause in Europe tomatch one that has been reported in North America[55,57]. Thus, other loci, in addition to tim,might generatea European latitudinal cline for diapause, althoughthe geographic variation in tim would seemingly act inthe opposite direction. One candidate gene encodes theinsulin-regulated phosphoinositide-3 kinase, which isassociatedwith diapause incidence in natural North Amer-ican populations [58].
A molecular basis for the altered tim phenotypesTIM regulates the transduction of photic signals to thecircadian clock through its interaction with cryptochrome(CRY), the circadian blue-light receptor (see Figure I inBox 1) [1]. In a yeast two-hybrid assay, CRY interactedmorestrongly with S-TIM than L-TIM (Figure 3d) [51]. If thischange inTIM–CRYdynamics is reflected in vivo, ls-timfliesshould have more stable TIM products. Immunoblotanalysis showed this to be the case [51,59] and is buttressedby the observation that daytime TIM levels do not fall asexpected in L-TIM transformants [51]. Thus, the L-TIMisoform likely provides greater overall TIM stability andcauses the attenuated circadian and seasonal photore-sponses of the ls-tim genotype [13,51].
Solving the tim spatial puzzleThe discrepancy between the geographical distributionof tim alleles and the observed phenotypes can bereconciled with the results from the neutrality tests, whichrevealed that the tim polymorphism might be under direc-tional and not balancing selection, representing an evol-utionary transient for a young allele (ls-tim) presentlyincreasing in frequency [13]. That this new variant seemsto have enhanced cold and photoperiodic adaptation(earlier diapause and reduced circadian photosensitivity)explains why it was favoured by selection in a Europeanseasonal environment. In additional populations from
129

Box 2. A clock gene clines in a non-model vertebrate
Repetitive sequences, as within per, are believed to represent a
major substrate for evolutionary change given their rapid mutation
rates [76]. The positive circadian regulator, Clock (CLK), contains a
poly-glutamine (polyQ) region that serves as a transcriptional
transactivation domain [77]. Variation in the length of the polyQ-
encoding repeat in Clk does not show a cline in Australian D.
melanogaster populations [23]. However, in birds, the polyQ repeat
length shows a latitudinal cline within European blue tit (Cyanistes
caeruleus) populations, with longer repeats found in higher
latitudes [78]. Patterns of length variation within and between
populations do not reflect those of neutral microsatellite markers,
suggesting the possibility of balancing selection, and the authors
speculate that this could be related to the seasonal migratory
polymorphism also observed in this species. The implication of this
study is that changes in polyQ length might alter the transactivation
potential of Clk, which in turn could alter the response of the
circadian mechanism to environmental inputs. However, any direct
relationship between these circadian phenotypes and polyQ varia-
tion in this vertebrate remains to be defined.
Review Trends in Genetics Vol.24 No.3
Crete and Israel, the frequency of ls-tim declined and thelatitudinal cline collapsed (Figure 3b) [13]. Correlating ls-tim frequencies with overland distances from southeast-ern Italy, where ls-tim frequency is highest, restores ahighly predictable linear relationship (Figure 3b). Thisfinding suggests that the ls-tim allele arose in south-eastern Italy and through directional selection is spread-ing outward in every direction, presumably because of theselection imposed by the European seasonal environment[13]. However, it is difficult to explain why, if the newallele emerged several thousand years ago, it has not yetbeen fixed in lower and higher latitudes? D. melanogasteris a human commensal; therefore, ls-tim might have beenmaintained in small isolated populations in southeasternItaly for hundreds or even thousands of years until thelate Bronze age (�3000 ya) and the development of viti-culture. Human migration would have spread the allele,albeit slowly, in every direction (Figure 2) [31,34]. Inaddition, in southern Europe, seasonal photoperiodicselection is expected to be less intense, inevitably slowingthe initial spread of the new allele.
However, although neutrality test results are consist-ent with directional selection, it is still possible that ls-tim is maintained by balancing selection for reasonsstated earlier. One might rationalize that the attenuatedcircadian photoresponsiveness and enhanced diapausemediated by tim variation could be favoured only in anarrow range of southern latitudes. For example, dia-pausing females go into spontaneous remission after afew weeks [60]. As ls-tim females move into diapauseunder relatively long days early in the season, anyspontaneous re-emergence might find them underunfavourable mid-winter conditions, particularly innorthern regions, with a consequently higher fitnesscost. Further south, in Crete or Israel, seasonal selectionmight be more relaxed, leading to reduced ls-tim fre-quencies. This contrived balancing scenario would elim-inate the need to propose a single ‘epicentre’ for wherethe new allele arose. One possibility for clarifying thebalancing versus directional hypotheses might be tosurvey tim variation in another continent. The coloniza-tion of North America by European immigrants, and theflies they brought with them over the last four centuries,suggests there is not a single geographical focus for thels-tim allele, as proposed for Europe, because ls-timwould have been repeatedly reintroduced (Figure 2)[34,61]. If 400 years is long enough for selection toinfluence ls-tim gene frequencies, under balancing selec-tion, a similar European-like distribution of the twovariants at similar latitudes might be observed in theUnited States and Canada. Under directional selection,we would expect a relatively smooth gradient withhigher levels of ls-tim in the northern parts of the con-tinent (Labrador, latitude �538N, assuming flies can befound there; Figure 2), with reduced frequencies insouthern Florida (latitude �258N), which is furthersouth than Greece or Israel.
Concluding remarksA tenet of modern evolution is that unfavourable newmutations are eliminated by selection, neutral mutations
130
can be maintained by drift, and adaptive mutations willspread through the population. The relatively new ls-timallele provides an example of the latter that has beenstudied at several biological levels [13,51]. Several out-standing issues remain; American diapausing flies seem tohave enhanced fitness under stressful environmental con-ditions [54–56], but this has not been shown specifically forEuropean ls-tim. Furthermore, the observation that ls-timis a young allele canmake interpretation of neutrality testsproblematic; therefore, functional data offer additional andimportant contributions to understanding the evolutionarybiology of this locus. Indeed, a recent review outlines thecomplexities of neutrality tests and advocates the use offunctional assays in verifying targets of selection [16].Nevertheless, examination of ls-tim spatial variation onother continents could be helpful in further illuminatingthe selective forces that might be at work here. Moregenerally, the involvement of circadian clock genes inseasonal phenomena has been an area of much speculationand controversy in the insect literature [62], whereas it isgenerally accepted inmammals [63] andArabidopsis thali-ana [64]. In D. melanogaster [13,51], and in a relatedDrosophilid, Chymomyza costata [65,66], it seems thattim plays a role in diapause, but whether tim affectsdiapause through its effects on the clock, rather thanexclusively on a shared light input to both circadian andphotoperiodic mechanisms, is an important question to beanswered. Similarly, a biochemical mechanism has yet tobe shown through which the Thr-Gly repeats affect naturalvariation in temperature compensation.
Nature provides us with long-term experiments whereall the complexities of dynamically changing environmentsare brought to bear on individual loci. Clines suggestselection, but simulating environmental conditions in anattempt to isolate any selective agent, as well as beingdifficult in the laboratory, is perhaps premature unlessindependent evidence for selection can be obtained.Neutrality tests, although sometimes difficult to interpret[16], can detect the signature of selection at the DNAsequence level and provide the evolutionary frameworkfor studying putative selective forces. Natural polymorph-isms also provide less severe phenotypes for genetic

Review Trends in Genetics Vol.24 No.3
analysis compared with mutagenesis-derived variants [67]and, whereas the latter is required to define the locus, theformer firmly places that locus in an evolutionary andecological context. Thus, the study of natural geographicalvariation in clock genes has added a novel dimension torhythm research, bringing into focus the selective value offine tuning the underlying circadian mechanism by thelocal environment. We envisage that, in the future,these types of geographical studies of clock gene variationmight be extended to bothmodel and non-model organisms(Box 2), in an attempt to learn more about their roles inanimal adaptation.
AcknowledgementsC.P.K. thanks the Royal Society for a Wolfson Research Merit Award andBBSRC and NERC for grant support. C.P.K. and R.C. acknowledgeEuropean Community grants under Frameworks 2, 4 and 6 (EUCLOCK018741) and Ministero dell’Universita e della Ricerca Scientifica eTecnologica (MURST)/British Council. R.C, also acknowledges a grantfrom the Italian Space Agency, DCMC grant. A.A.P. is supported by theHoward Hughes Medical Institute, Fiocruz and CNPq. E.T. acknowledgesa project grant from NERC.
References1 Hall, J.C. (2003) Genetics and molecular biology of rhythms in
Drosophila and other insects. Adv. Genet. 48, 1–2802 Duffield, G.E. (2003) DNAmicroarray analyses of circadian timing: the
genomic basis of biological time. J. Neuroendocrinol. 15, 991–10023 Reddy, A.B. et al. (2006) Circadian orchestration of the hepatic
proteome. Curr. Biol. 16, 1107–11154 Dvornyk, V. et al. (2003) Origin and evolution of circadian clock genes in
prokaryotes. Proc. Natl. Acad. Sci. U. S. A. 100, 2495–25005 Maywood, E.S. et al. (2006) Circadian timing in health and disease.
Prog. Brain Res. 153, 253–2696 Hardin, P.E. (2005) The circadian timekeeping system of Drosophila.
Curr. Biol. 15, R714–R7227 Liu, A.C. et al. (2007) Mammalian circadian signaling networks and
therapeutic targets. Nat. Chem. Biol. 3, 630–6398 Gallego, M. and Virshup, D.M. (2007) Post-translational modifications
regulate the ticking of the circadian clock. Nat. Rev. Mol. Cell Biol. 8,139–148
9 Toma, D.P. et al. (2000) Changes in periodmRNA levels in the brain anddivision of labor in honey bee colonies. Proc. Natl. Acad. Sci. U. S. A. 97,6914–6919
10 Reppert, S.M. (2006) A colorful model of the circadian clock. Cell 124,233–236
11 Costa, R. and Kyriacou, C.P. (1998) Functional and evolutionaryimplications of natural variation in clock genes. Curr. Opin.Neurobiol. 8, 659–664
12 Costa, R. et al. (1992) A latitudinal cline in a Drosophila clock gene.Proc. Biol. Sci. 250, 43–49
13 Tauber, E. et al. (2007) Natural selection favors a newly derivedtimeless allele in Drosophila melanogaster. Science 316, 1895–1898
14 Glinka, S. et al. (2003) Demography and natural selection have shapedgenetic variation in Drosophila melanogaster: a multi-locus approach.Genetics 165, 1269–1278
15 Ometto, L. et al. (2005) Inferring the effects of demography andselection on Drosophila melanogaster populations from achromosome-wide scan of DNA variation. Mol. Biol. Evol. 22, 2119–2130
16 Jensen, J.D. et al. (2007) Approaches for identifying targets of positiveselection. Trends Genet. 23, 568–577
17 Gatfield, D. and Schibler, U. (2007) Physiology. Proteasomes keep thecircadian clock ticking. Science 316, 1135–1136
18 Majercak, J. et al. (1999) How a circadian clock adapts to seasonaldecreases in temperature and day length. Neuron 24, 219–230
19 Costa, R. et al. (1991) Length polymorphism in the threonine-glycine-encoding repeat region of the period gene in Drosophila. J. Mol. Evol.32, 238–246
20 Sawyer, L.A. et al. (2006) The period gene Thr-Gly polymorphism inAustralian and African Drosophila melanogaster populations:implications for selection. Genetics 174, 465–480
21 Kyriacou, C.P. et al. (2007) A cline in the Drosophila melanogasterperiod gene in Australia: neither down nor under. J. Evol. Biol. 20,1649–1651
22 Weeks, A.R. et al. (2007) Robust clines and robust sampling: a reply toKyriacou et al.. J. Evol. Biol. 20, 1652–1654
23 Weeks, A.R. et al. (2006) In search of clinal variation in the period andclock timing genes in AustralianDrosophila melanogaster populations.J. Evol. Biol. 19, 551–557
24 Rosato, E. et al. (1997) Linkage disequilibrium, mutational analysisand natural selection in the repetitive region of the clock gene, period,in Drosophila melanogaster. Genet. Res. 69, 89–99
25 Sawyer, L.A. et al. (1997) Natural variation in a Drosophila clock geneand temperature compensation. Science 278, 2117–2120
26 Ouyang, Y. et al. (1998) Resonating circadian clocks enhance fitness incyanobacteria. Proc. Natl. Acad. Sci. U. S. A. 95, 8660–8664
27 Aquadro, C.F. (1992) Why is the genome variable? Insights fromDrosophila. Trends Genet. 8, 355–362
28 Kimura, M. (1962) On the probability of fixation of mutant genes in apopulation. Genetics 47, 713–719
29 Peixoto, A.A. (2002) Evolutionary behavioral genetics in Drosophila.Adv. Genet. 47, 117–150
30 Zamorzaeva, I. et al. (2005) Sequence polymorphism of candidatebehavioural genes in Drosophila melanogaster flies from ‘Evolutioncanyon’. Mol. Ecol. 14, 3235–3245
31 David, J.R. and Capy, P. (1988) Genetic variation of Drosophilamelanogaster natural populations. Trends Genet. 4, 106–111
32 Lachaise, D. et al. (1988) Historical biogeography of the Drosophila-melanogaster species subgroup. Evol. Biol. 22, 159–225
33 Stephan, W. and Li, H. (2007) The recent demographic and adaptivehistory of Drosophila melanogaster. Heredity 98, 65–68
34 Keller, A. (2007) Drosophila melanogaster’s history as a humancommensal. Curr. Biol. 17, R77–R81
35 Oakeshott, J.G. et al. (1981) Latitudinal relationships of esterase-6 andphosphoglucomutase gene frequencies in Drosophila melanogaster.Heredity 47, 385–396
36 Anderson, A.J. and Oakeshott, J.G. (1984) Parallel geographicalpatterns of allozyme variation in two sibling Drosophila species.Nature 308, 729–731
37 Haddrill, P.R. et al. (2005) Multilocus patterns of nucleotide variabilityand the demographic and selection history of Drosophila melanogasterpopulations. Genome Res. 15, 790–799
38 Li, H. and Stephan, W. (2006) Inferring the demographic history andrate of adaptive substitution in Drosophila. PLoS Genet. 2, e166
39 Peixoto, A.A. et al. (1993) Molecular evolution of a repetitive regionwithin the per gene of Drosophila. Mol. Biol. Evol. 10, 127–139
40 Nielsen, J. et al. (1994) Big flies, small repeats: the ‘Thr-Gly’ region ofthe period gene in Diptera. Mol. Biol. Evol. 11, 839–853
41 Peixoto, A.A. et al. (1998) Molecular coevolution within a Drosophilaclock gene. Proc. Natl. Acad. Sci. U. S. A. 95, 4475–4480
42 Rosato, E. et al. (1994) Molecular polymorphism in the period gene ofDrosophila simulans. Genetics 138, 693–707
43 Rogers, A.S. et al. (2004) Molecular analysis of circadian clocks inDrosophila simulans. Genetica 120, 213–222
44 Rogers, A.S. et al. (2004) A mutation in Drosophila simulans thatlengthens the circadian period of locomotor activity. Genetica 120,223–232
45 Dembinska, M.E. et al. (1997) Circadian cycling of a PERIOD-beta-galactosidase fusion protein in Drosophila: evidence for cyclicaldegradation. J. Biol. Rhythms 12, 157–172
46 Nawathean, P. et al. (2007) A small conserved domain of DrosophilaPERIOD is important for circadian phosphorylation, nuclearlocalization, and transcriptional repressor activity. Mol. Cell. Biol.27, 5002–5013
47 Kim, E.Y. et al. (2007) A DOUBLETIME kinase binding domain on theDrosophila PERIOD protein is essential for its hyperphosphorylation,transcriptional repression, and circadian clock function.Mol. Cell. Biol.27, 5014–5028
48 Castiglione-Morelli, M.A. et al. (1995) Conformational study of theThr-Gly repeat in the Drosophila clock protein, PERIOD. Proc. Biol.Sci. 260, 155–163
131

Review Trends in Genetics Vol.24 No.3
49 Guantieri, V. et al. (1999) Different period gene repeats take ’turns’ atfine-tuning the circadian clock. Proc. Biol. Sci. 266, 2283–2288
50 Rosato, E. et al. (1997) Conceptual translation of timeless revealsalternative initiating methionines in Drosophila. Nucleic Acids Res.25, 455–458
51 Sandrelli, F. et al. (2007) A molecular basis for natural selection at thetimeless locus in Drosophila melanogaster. Science 316, 1898–1900
52 Pittendrigh, C.S. and Takamura, T. (1989) Latitudinal clines in theproperties of a circadian pacemaker. J. Biol. Rhythms 4, 217–235
53 Tauber, E. and Kyriacou, B.P. (2001) Insect photoperiodism andcircadian clocks: models andmechanisms. J. Biol. Rhythms 16, 381–390
54 Schmidt, P.S. et al. (2005) Genetic variance for diapause expressionand associated life histories inDrosophila melanogaster. Evolution Int.J. Org. Evolution 59, 2616–2625
55 Schmidt, P.S. et al. (2005) Geographic variation in diapause incidence,life-history traits, and climatic adaptation inDrosophila melanogaster.Evolution Int. J. Org. Evolution 59, 1721–1732
56 Schmidt, P.S. and Conde, D.R. (2006) Environmental heterogeneityand the maintenance of genetic variation for reproductive diapause inDrosophila melanogaster. Evolution Int. J. Org. Evolution 60, 1602–1611
57 Williams, K.D. and Sokolowski, M.B. (1993) Diapause in Drosophilamelanogaster females: a genetic analysis. Heredity 71, 312–317
58 Williams, K.D. et al. (2006) Natural variation in Drosophilamelanogaster diapause due to the insulin-regulated PI3-kinase.Proc. Natl. Acad. Sci. U. S. A. 103, 15911–15915
59 Peschel, N. et al. (2006) Veela defines a molecular link betweenCryptochrome and Timeless in the light-input pathway toDrosophila’s circadian clock. Proc. Natl. Acad. Sci. U. S. A. 103,17313–17318
60 Saunders, D.S. et al. (1989) Induction of diapause in Drosophilamelanogaster: photoperiodic regulation and the impact ofarrhythmic clock mutations on time measurement. Proc. Natl. Acad.Sci. U. S. A. 86, 3748–3752
61 Caracristi, G. and Schlotterer, C. (2003) Genetic differentiationbetween American and European Drosophila melanogasterpopulations could be attributed to admixture of African alleles. Mol.Biol. Evol. 20, 792–799
62 Bradshaw, W.E. et al. (2003) Circadian rhythmicity andphotoperiodism in the pitcher-plant mosquito: adaptive response to
Elsevier.com – linking scientists
Designed for scientists’ information needs, Elsevier
customer-focused navigation and an intuitive arch
greater prod
The easy-to-use navigational tools and structure
from one entry point. Users can perform rapid an
functionality, using the FAST technology of Scirus.
define their searches by any number of criteria to p
specific author or editor, book publication date,
physical sciences and social sciences – or by produ
1800 Elsevier journals, 2200 new books every year
In addition, tailored content for authors, editors an
on new products
Elsevier is proud to be a partner with the scientific
our mission and values at Elsevier.com. Discover
medical communities worldwide through partnershi
awards from The Else
As a world-leading publisher of scientific, technical
linking researchers and professionals to the best t
deepest coverage in a range of media types to
breakthroughs in research and discovery, and t
Elsevier. Building insightswww.elsev
132
the photic environment or correlated response to the seasonalenvironment? Am. Nat. 161, 735–748
63 Hazlerigg, D.G. et al. (2005) Photoperiod differentially regulates geneexpression rhythms in the rostral and caudal SCN. Curr. Biol. 15,R449–R450
64 Yanovsky, M.J. and Kay, S.A. (2002) Molecular basis of seasonal timemeasurement in Arabidopsis. Nature 419, 308–312
65 Pavelka, J. et al. (2003) TIMELESS: a link between the fly’s circadianand photoperiodic clocks? Eur. J. Entomol. 100, 255–265
66 Kostal, V. and Shimada, K. (2001) Malfunction of circadian clock in thenon-photoperiodic-diapause mutants of the drosophilid fly, Chymomyzacostata. J. Insect Physiol. 47, 1269–1274
67 Greenspan, R.J. (1997) A kinder, gentler genetic analysis of behavior:dissection gives way to modulation. Curr. Opin. Neurobiol. 7, 805–811
68 Kadener, S. et al. (2007) Clockwork Orange is a transcriptionalrepressor and a new Drosophila circadian pacemaker component.Genes Dev. 21, 1675–1686
69 Fang, Y. et al. (2007) Post-translational regulation of the Drosophilacircadian clock requres protein phosphatase 1 (PP1). Genes Dev. 21,1506–1518
70 Benito, J. et al. (2007) PDP1epsilon functions downstream of thecircadian oscillator to mediate behavioral rhythms. J. Neurosci. 27,2539–2547
71 Helfrich-Forster, C. (2003) The neuroarchitecture of the circadian clockin the brain ofDrosophila melanogaster.Microsc. Res. Tech. 62, 94–102
72 Stoleru, D. et al. (2004) Coupled oscillators control morning andevening locomotor behaviour of Drosophila. Nature 431, 862–868
73 Grima, B. et al. (2004) Morning and evening peaks of activity rely ondifferent clock neurons of the Drosophila brain. Nature 431, 869–873
74 Plautz, J.D. et al. (1997) Independent photoreceptive circadian clocksthroughout Drosophila. Science 278, 1632–1635
75 Krishnan, B. et al. (2001) A new role for cryptochrome in a Drosophilacircadian oscillator. Nature 411, 313–317
76 Fondon, J.W., III and Garner, H.R. (2004) Molecular origins of rapidand continuous morphological evolution. Proc. Natl. Acad. Sci. U. S. A.101, 18058–18063
77 Gekakis, N. et al. (1998) Role of the CLOCK protein in the mammaliancircadian mechanism. Science 280, 1564–1569
78 Johnsen, A. et al. (2007) AvianClock gene polymorphism: evidence for alatitudinal cline in allele frequencies. Mol. Ecol. 16, 4867–4880
to new research and thinking
.com is powered by the latest technology with
itecture for an improved user experience and
uctivity.
connect scientists with vital information – all
d precise searches with our advanced search
com, the free science search engine. Users can
inpoint information and resources. Search by a
subject area – life sciences, health sciences,
ct type. Elsevier’s portfolio includes more than
and a range of innovative electronic products.
d librarians provides timely news and updates
and services.
and medical community. Find out more about
how we support the scientific, technical and
ps with libraries and other publishers, and grant
vier Foundation.
and health information, Elsevier is dedicated to
hinking in their fields. We offer the widest and
enhance cross-pollination of information,
he sharing and preservation of knowledge.
. Breaking boundaries.ier.com




















