
Mathematical Modelling of Circadian Signalling in Arabidopsis
Upload
independentCategory
view
3download
0
Appetite, 1998, 30, 255–267
Article No. ap970134
Circadian Rhythms in the Zucker Obese Rat:
Assessment and Intervention
R. E. MISTLBERGER, H. LUKMAN and B. G. NADEAUDepartment of Psychology, Simon Fraser University, Burnaby
Body temperature (Tb) and activity were recorded by telemetry in obese and leanZucker rats in light–dark (LD), constant dark (DD) and constant light (LL). InLD, obese rats, by comparison with lean rats, exhibited a 2–4-h phase advanceand attenuated amplitude of Tb and activity rhythms. These differences persistedon the first day of DD, and thus were not due to differential sensitivity to maskingeffects of light. In LL, obese and lean rats exhibited similar free-running periods,thus the phase advance in LD was also not due to a short intrinsic period. InLD, obese rats exhibited more diurnal food intake and a reduced LD intakeratio. To assess the role of diurnal feeding in weight gain, one group of obeserats was fed ad libitum, and another fed only at night. Food intake did not differsignificantly between groups, but ad libitum fed rats gained 23% more weight(60 g) over 60 days, suggesting that excessive diurnal feeding may contributeadversely to body weight regulation in this animal model of obesity.
1998 Academic Press Limited
I
The Zucker (fa/fa) rat is a widely-used animal model of obesity caused by amutation in a single recessive gene encoding the receptor for leptin, a hormonesecreted by white adipose tissue that contributes to energy homeostasis by suppressingfood intake and increasing energy expenditure in proportion to body-fat levels(Zhang et al., 1994; Frederich et al., 1995; Maffei et al., 1995; Chua et al., 1996).The Zucker fatty is consequently relatively insensitive to leptin (Seeley et al., 1996;Cusin et al., 1996), and exhibits a phenotype characterized by excessive food intake,weight gain and adiposity (Bray, 1977). Altered circadian rhythms have also beendescribed in the Zucker fatty, although there are inconsistencies in the literature, assome studies report apparently normal feeding, activity or body temperature (Tb)rhythms (Demes et al., 1991; Enns & Grinker, 1983; Ens & Grinker, 1982; Grinkeret al., 1979; Keesey et al., 1990; McLaughlin & Baile, 1981; Prins et al., 1986)whereas others describe attenuated amplitude or advanced phase of feeding, Tb,activity or sleep rhythms under standard daily light–dark (LD) cycles (Becker &Grinker, 1977; Danguir, 1989; Fukagawa et al., 1988; Fukagawa et al., 1992; Martinet al., 1979; Murakami et al., 1995; Wangsness et al., 1978). A reduced amplitude
Supported by equipment and operating grants from NSERC, Canada, to R. E. Mistlberger. Thisresearch was presented in preliminary form in the Society for Neuroscience Abstracts, 20, 1219, 1994.H. Lukman is now at the Department of Psychology, University of Leeds, Leeds, U.K.
Address correspondence to: Dr Ralph Mistlberger, Department of Psychology, Simon Fraser Uni-versity, Burnaby, BC Canada, V5A 1S6.
0195–6663/98/030255+13 $25.00/0 1998 Academic Press Limited

256 R. E. MISTLBERGER ET AL.
or altered phase of various daily rhythms has been described in other animal modelsof obesity (e.g. Balagura & Devenport, 1970; Gold et al., 1974; Ho & Chin, 1988)and may also characterize obesity in humans (Belisle et al., 1988; Ferrari et al., 1990;Fricker et al., 1990; Lee et al., 1992; Van Cauter et al., 1994). However, the natureof the circadian disturbance, and its contribution to the development and maintenanceof excessive weight gain and obesity are unclear.
To gain further insight into the nature of the circadian disturbance in obeseZucker rats, and its possible significance for excessive weight gain, two studies wereconducted. In the first study, circadian rhythms of Tb, activity and food intake wererecorded in lean and obese Zucker rats under LD, constant dark (DD) and constantdim light (LL). This sequence of lighting conditions enabled us to confirm andquantify a phase advance and reduced amplitude of circadian rhythms in obese ratsin LD, and to assess whether the phase advance was due to reduced masking effectsof light or to differences in the periodicity of the circadian pacemaker.
In the second study, the importance of the daily timing of food intake for thedevelopment of excessive weight gain was assessed by comparing weight gain over60 days in two groups of obese rats, one fed ad libitum and the second fed only atnight, to prevent the enhanced diurnal feeding that was evident in the obese rats ofthe first study. This experiment served to test a hypothesis that some portion ofexcessive weight gain in Zucker rats is due to the abnormal phase relation of feedingrhythms with respect to the LD cycle, and possibly with respect to metabolic rhythmsthat may remain in phase with the LD cycle. The results provide modest supportfor a general principle that the timing of food intake can impact on the regulationof body weight in metabolically obese animals.
M
General
SubjectsFemale Zucker fa/fa (N=17) or Fa/? (N=10) rats were obtained from Charles
River (Montreal) at 5 weeks of age. All rats were housed in a climate-controlledcolony room for 7 days prior to assignment to one of two experiments.
Experiment 1
ProceduresMiniature, radiofrequency transmitters (Minimitter Inc., Sunriver, OR) were
implanted in the peritoneal cavities of seven obese and five lean rats anesthetizedwith halothane. The rats were then housed individually in sawdust-lined plasticcages (45×25×20 cm) inside sound-attenuating, electrically shielded isolation boxesequipped with fans and incandescent lighting controlled by computer. Tb andmovement were monitored continuously via telemetry and Dataquest III software(Minimitter Inc.) for 25 days under LD (12:12, 10 lux during lights-on), 3 days ofDD (0 lux) and 7 weeks in LL (10 lux). Tb and activity data were stored and averagedin 10-min intervals. Food intake was measured hourly for 24 h in LD by manuallyweighing and replacing rat chow pellets (Richmond Standard 5001 rat chow, PMIFeeds, BC, Canada).

257CIRCADIAN RHYTHMS IN ZUCKER OBESE RATS
Data analysisTemperature and activity data were converted to text files compatible with the
Circadia data-analysis program for the Macintosh. The data were first normalizedwith respect to daily mean values for each animal. Group means were obtainedusing Excel and then subject to a weighted running average (1:3:1). Average waveformsand best-fitting cosine functions for group mean and individual data were thencalculated and plotted using Circadia. Acrophase (expressed in hours after lights-on), amplitude (arbitrary units) and mean level of the cosine functions for Tb andactivity rhythms during 10 days of LD and the first day of DD were quantified. Theperiod of rhythms in LL was quantified by periodogram (Dorrscheidt & Beck, 1975)and the resulting Q statistic was used as an estimation of amplitude. Significance ofdifferences were assessed by repeated measures ANOVA and Bonferroni t-tests.Group mean data reported ±SEM.
Experiment 2
ProceduresTen obese and five lean Zucker rats were housed individually in stainless steel
hanging cages in a climate-controlled colony room under LD 10:14 (100 lux duringlights-on). Rat chow intake and body weight were measured daily for 1 week. Twogroups of five obese rats were matched for weight. For the next 60 days, one groupreceived food ad libitum and the other received food only during the 14-h darkperiod each day. The use of a 14-h night enabled night-fed rats to maintain foodintake at levels equivalent to rats fed ad libitum. Food and body weight weremeasured 30 min before dark onset each day. Group differences in body weight andfood intake were evaluated by ANOVA and Tukey HSD tests.
R
Experiment 1
Circadian rhythms in LDGroup mean acrophases, amplitudes and mean levels are presented in Table 1.
There were significant group differences in the acrophase of Tb [F (1,10)=36·84,p<0·001] and activity [F (1,10)=43·13, p=43·13, p<0·001], in the amplitude ofactivity [F (1,10)=24·48, p=0·001], and in the mean level of activity [F (1,10)=29·99, p<0·001]. In LD, the acrophase of best-fitting cosine functions was significantlyphase advanced in obese rats, by comparison with lean rats, for both Tb (2·2 h; t=9·71, p<0·001; Fig. 1a,b) and activity (2·5 h; t=4·38, p=0·001; Fig. 1e,f). Obeserats also showed significantly reduced amplitude (t=4·1, p=0·01) and mean level(t=4·95, p=0·002) of the cosine functions for activity. Amplitude and mean levelfor Tb were also lower in obese rats, but these differences did not reach significance.
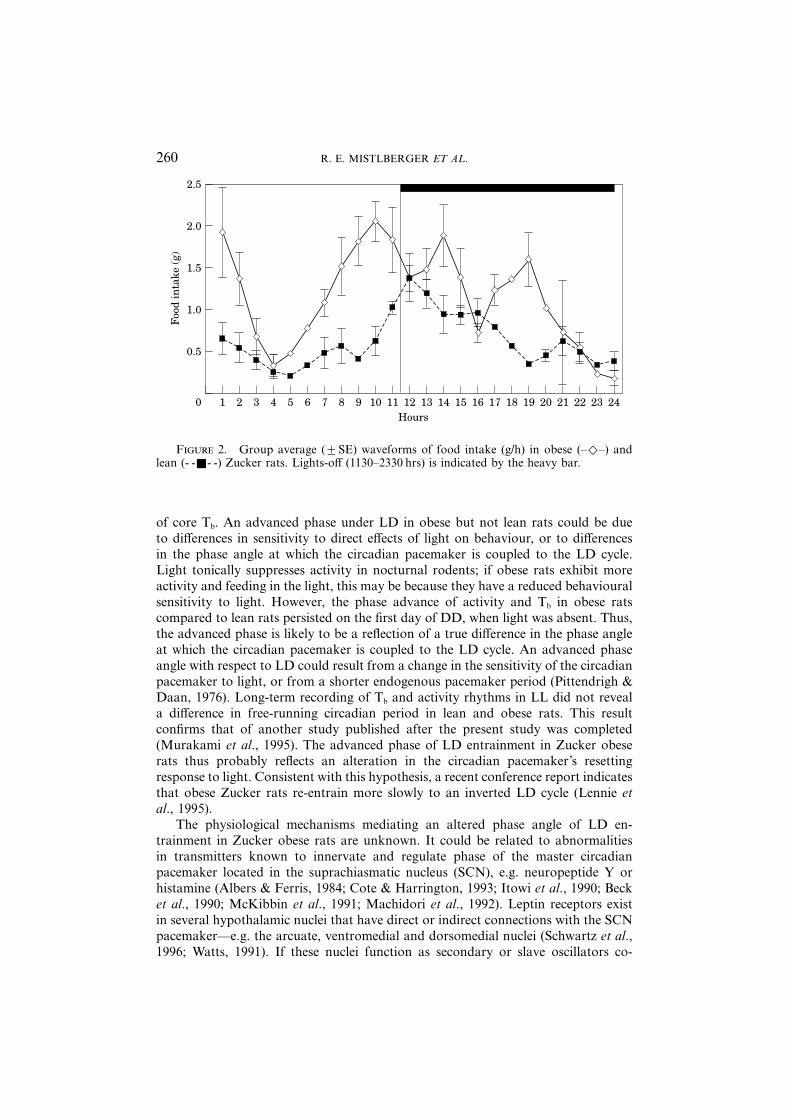
Total 24-h food intake, measured hourly over one full day in LD, averaged25·9±2 g in the obese rats and 13·4±1 g in the lean rats (t=5·14, p<0·001; Fig.2). The average waveform of food intake did not closely approximate a sine wavein the obese rats, so curve fitting was not attempted. However, visual inspection ofthe waveforms indicates that the peak level of food intake occurred about 2 h beforedark onset in the obese rats, as opposed to during the first hour of dark in the lean

258 R. E. MISTLBERGER ET AL.
T 1Parameters of best-fitting cosine functions for temperature and activity rhythms in LD
and DD, and of free-running rhythms in LL
Light–dark 12:12 Constant dark, day 1 Constant light
Group ACR AMP Mean ACR AMP Mean TAU AMP Mean
Tb lean 17·7 52·2 38·0 17·9 60·8 38·0 24·8 0·46 37·3±0·04 ±2·4 ±0·11 ±0·83 ±3·2 ±0·23 ±0·04 ±0·12 ±0·10
Tb obese 15·6 44·0 37·5 14·2 43·2 37·4 24·6 0·45 37·0±0·12∗∗ ±2·7 ±0·11 ±0·48∗∗ ±8·3 ±0·14 ±0·05 ±0·10 ±0·26
Activity 17·6 29·7 62·9 17·7 37·0 62·2 24·9 0·41 51·2lean ±0·20 ±2·5 ±3·5 ±0·87 ±6·7 ±6·7 ±0·04 ±0·01 ±2·3Activity 15·1 8·2 21·5 13·2 9·6 24·7 24·7 0·43 22·5obese ±0·24∗∗ ±0·92∗ ±2·5∗∗ ±0·72∗∗ ±3·2∗∗ ±4·2∗∗ ±0·06 ±0·01 ±2·2∗∗
All means±SE. Tb=body temperature; ACR=acrophase of cosine function in light–dark (LD)and constant dark (DD), in hours after lights-on; AMP=amplitude of cosine function in LD and DD,of periodogram Q statistic in LL; TAU=free-running period in hours.
Lean vs. obese ∗p<0·01, ∗∗p<0·005.
rats. In addition, the percentage of food consumed at night was significantlyattenuated in the obese rats (obese 51±5·1%, lean 64±4·0%; t=2·45, p<0·05).
Circadian rhythms in DDDifferences between lean and obese rats evident in LD persisted on the first day
of DD; obese rats exhibited significantly phase-advanced circadian rhythms of Tb
(3·7 h; t=4·28, p=0·004; Fig. 1c,d) and activity (4·5 h, t=4·92, p=0·001; Fig. 1g,h), and a significantly lower amplitude (t=4·08, p=0·006) and mean level (t=5·06,p=0·001) for activity. There was a non-significant trend for lower amplitude andmean level for Tb in obese rats. Although the magnitude of the acrophase differencesbetween groups appeared to be greater in DD than in LD, the omnibus F tests didnot support a significant effect of lighting condition or interaction between groupand lighting condition.
Circadian rhythms in LLTb and activity rhythms free-ran in LL, in all cases with a period greater than
24 h. Lean and obese rats did not differ in average period or amplitude, but obeserats continued to express significantly lower levels of activity (t=4·87, p=0·001).The Tb rhythm of an obese rat that is representative of both groups is illustrated inFig. 3.
Experiment 2
Food intake and body weightInitial body weights, average daily weight gains and average daily food intake
over 60 days are presented in Table 2 and illustrated in Fig. 4. Body weights differedsignificantly across groups [F (2,12)=155·06, p<0·001]. Free-feeding obese ratsgained 23% more weight than did nocturnal-fed obese rats (p<0·05) and 200% morethan free-feeding lean rats (p<0·05). Average daily food intake also differed between

259CIRCADIAN RHYTHMS IN ZUCKER OBESE RATS
7.00
198
187.00
Time of day (hrs)
Nor
mal
ized
act
ivit
y
19.00
138
78
13.00 1.00
(g)
Constant dark, Day 1
Fa/xx Leanfa/fa Obese
7.00
198
187.00
Time of day (hrs)
Nor
mal
ized
act
ivit
y
19.00
138
78
13.00 1.00
(h)
Constant dark, Day 1
Fa/xx Leanfa/fa Obese
7.00
200
327.00N
orm
aliz
ed a
ctiv
ity
19.00
116
74
13.00 1.00
(e)
LD12:12
Fa/xx Leanfa/fa Obese
7.00
200
327.00N
orm
aliz
ed a
ctiv
ity
19.00
116
74
13.00 1.00
(f)
LD12:12
Fa/xx Leanfa/fa Obese
7.00
101
977.00
Nor
mal
ized
tem
pera
ture
19.00
99
98
13.00 1.00
(c)
Constant dark, Day 1
Fa/xx Leanfa/fa Obese
7.00
101
977.00
Nor
mal
ized
tem
pera
ture
19.00
99
98
13.00 1.00
(d)
Constant dark, Day 1
Fa/xx Leanfa/fa Obese
7.00
101
977.00
Nor
mal
ized
tem
pera
ture
19.00
99
98
13.00 1.00
(a)
LD12:12
Fa/xx Leanfa/fa Obese
7.00
101
977.00
Nor
mal
ized
tem
pera
ture
19.00
99
98
13.00 1.00
(b)
LD12:12
Fa/xx Leanfa/fa Obese
100
100
158 158
100
100
F 1. Group average waveforms and best fitting cosine functions for temperatureand activity in obese (bold lines) and lean Zucker rats in light–dark (LD) (a,b,e,f) and thefirst day of constant dark (DD) (c,d,g,h). Data for each animal were normalized before groupaverages were calculated and subject to a 1:4:1 running average. Due to low variability,normalized temperature in LD is presented unsmoothed. During LD, lights were on from0700 to 1900 hrs.
lean and obese rats [F (2,12)=99·7, p<0·0001], but neither average daily food intakenor cumulative food intake differed significantly between free-feeding and nocturnal-fed obese rats (p>0·05).
D
Circadian Rhythms in Zucker Rats: Phase, Period and Amplitude
This study confirms that circadian rhythms of activity and feeding are phaseadvanced in obese Zucker rats, and extends this observation to the circadian rhythm
260 R. E. MISTLBERGER ET AL.
24
2.5
Hours
Foo
d in
take
(g)
5
2.0
1.5
1.0
0.5
1 2 3 4 6 7 8 90 2322212019181716151413121110
F 2. Group average (±SE) waveforms of food intake (g/h) in obese (–Η–) andlean (- -Ε- -) Zucker rats. Lights-off (1130–2330 hrs) is indicated by the heavy bar.
of core Tb. An advanced phase under LD in obese but not lean rats could be dueto differences in sensitivity to direct effects of light on behaviour, or to differencesin the phase angle at which the circadian pacemaker is coupled to the LD cycle.Light tonically suppresses activity in nocturnal rodents; if obese rats exhibit moreactivity and feeding in the light, this may be because they have a reduced behaviouralsensitivity to light. However, the phase advance of activity and Tb in obese ratscompared to lean rats persisted on the first day of DD, when light was absent. Thus,the advanced phase is likely to be a reflection of a true difference in the phase angleat which the circadian pacemaker is coupled to the LD cycle. An advanced phaseangle with respect to LD could result from a change in the sensitivity of the circadianpacemaker to light, or from a shorter endogenous pacemaker period (Pittendrigh &Daan, 1976). Long-term recording of Tb and activity rhythms in LL did not reveala difference in free-running circadian period in lean and obese rats. This resultconfirms that of another study published after the present study was completed(Murakami et al., 1995). The advanced phase of LD entrainment in Zucker obeserats thus probably reflects an alteration in the circadian pacemaker’s resettingresponse to light. Consistent with this hypothesis, a recent conference report indicatesthat obese Zucker rats re-entrain more slowly to an inverted LD cycle (Lennie etal., 1995).
The physiological mechanisms mediating an altered phase angle of LD en-trainment in Zucker obese rats are unknown. It could be related to abnormalitiesin transmitters known to innervate and regulate phase of the master circadianpacemaker located in the suprachiasmatic nucleus (SCN), e.g. neuropeptide Y orhistamine (Albers & Ferris, 1984; Cote & Harrington, 1993; Itowi et al., 1990; Becket al., 1990; McKibbin et al., 1991; Machidori et al., 1992). Leptin receptors existin several hypothalamic nuclei that have direct or indirect connections with the SCNpacemaker—e.g. the arcuate, ventromedial and dorsomedial nuclei (Schwartz et al.,1996; Watts, 1991). If these nuclei function as secondary or slave oscillators co-

261CIRCADIAN RHYTHMS IN ZUCKER OBESE RATS
Time of day (hrs)
8:00 20:00 8:00 20:00 8:00
Day
s
LL
DD
LD
F 3. Double raster plot of body temperature in obese Zucker rat in light–dark(LD), constant dark (DD) and constant light (LL). Consecutive days are plotted left-to-rightand top-to-bottom. Ten-minute bins during which temperature was above the mean for thatday are indicated by vertical deflections on the time line for each day. Missing data (transmittersignal out of range or hardware failure) are indicated by blanks. The data for this animal aretypical of the obese rats in all lighting conditions, and of lean and obese rats in LL.

262 R. E. MISTLBERGER ET AL.
T 2Average food intake and body weight change of Zucker lean and obese rats during 60
days of recording
Group Intake/day (g) Increment/day (g)
Lean 18·6±1·7a 1·6±0·2a
Obese night-fed 29·8±1·9b 3·9±0·3b
Obese free-fed 31·3±0·8b 4·9±0·4c
Note: Means are±SE. Groups with different letters are significantly different from each other atp<0·05 (Tukey HSD).
60
45
100
Days
Foo
d in
take
(g/
day)
30
35
30
25
20
15
10 15 20 25 40 50 55
40(a)
45355 60
550
100BL
Days
Bod
y w
eigh
t (g
)
30
350300250200150
10 15 20 25 40 50 55
450
(b)
45355
500
400
F 4. Average food intake/day (a) and body weight (b) over 60 days of recordingin two groups of obese and one group of lean rats (N=5 each). Standard error bars areomitted in panel (a) for clarity, and are smaller than the means symbols in most of panel (b).Ο, obese night-fed; Η, obese free-fed; Β, lean free-fed.
ordinated by pacemaking signals from the SCN, it is possible that the phase angleof coupling among these oscillating structures, and thus the apparent phase of LDentrainment, could be altered as a consequence of changes in neural processingassociated with the leptin receptor mutation.
In addition to an altered phase of circadian rhythms in LD, a relatively reducedamplitude of activity and feeding rhythms and a trend toward a reduced amplitudeof Tb rhythms in obese Zucker rats was also observed, confirming several previousreports (e.g. Fukagawa et al., 1988; Murakami et al., 1995). However, a reducedamplitude is difficult to interpret. It is not explained by a weaker masking effect oflight, as it was evident in both LD and DD. It could reflect a reduced amplitude ofthe circadian pacemaker, or it could be due to masking effects of metabolic processesdownstream from the clock, i.e. homeostatic factors driving hyperphagia maysupersede clock control of ingestive behaviour. Feeding behaviour could, in turn,increase both activity and Tb at circadian phases when the clock specifies lowerlevels. This could also contribute to the early rise of Tb prior to the dark period;however, a homeostatic factor would likely compete with the circadian factor at allphases, and thus would not be likely to account for phase-advanced rhythms detectedby curve fitting. Total food deprivation could be used to assess an effect of feedingon the amplitude and phase of the Tb and activity rhythms, although this approachis potentially confounded by deprivation-induced increases in activity.

263CIRCADIAN RHYTHMS IN ZUCKER OBESE RATS
Assessments of circadian function in obese Zucker rats have produced somewhatvariable results across studies. Only two studies have explicitly quantified phase, andreported similar abnormalities in fat animals compared to age-matched lean animals(Fukagawa et al., 1992, and present study). These two studies used different sexes,indicating that variability within the literature is not related to sex differences orestrous cycling, if present. A large number of studies have reported amplitude andmean level measures. Many of these reported attenuated amplitude, usually expressedin the form of LD ratios (e.g. Becker & Grinker, 1977; Danguir, 1989; Fukagawaet al., 1988; Fukagawa et al., 1992; Martin et al., 1979; Murakami et al., 1995;Wangsness et al., 1978), but others reported normal amplitudes (e.g. Demes et al.,1991; Enns & Grinker, 1983; Enns et al., 1982; Grinker et al., 1979; Keesey et al.,1990; McLaughlin & Baile, 1981). However, inspection of the data indicates thatLD ratios of feeding or activity (excluding wheel running) in these studies are in the58–70% nocturnal range, which is considerably below the degree of nocturnalityreported for lean rats of other strains—generally 75–85% (e.g. Rietveld et al., 1980;Spiteri et al., 1982; Vachon & Savoi, 1987; Velasco Plaza et al., 1993; Zucker, 1971).Thus, the literature on obese rats may be more consistent than apparent; studiesdiffer more in what is accepted as normal values for the lean comparison group.
Differences in reported mean levels are also evident across studies, but thesedifferences are more difficult to assess because of the range of measurement devicesutilized (e.g. running wheels, stabilometers, radio-frequency transmitters).
A Role for Circadian Timing of Food Intake in Weight Gain
It has been widely observed that circadian rhythms of food intake are altered inanimal models of obesity (e.g. Zucker rats, ventromedial hypothalamic ablated rats,ob/ob mice). Abnormalities in circadian rhythms of metabolic and hormone systemshave also been noted in obese humans (Ferrari et al., 1990; Lee et al., 1992; VanCauter et al., 1994), as has a tendency for daily food intake to be concentrated laterin the day, by comparison with lean age cohorts (e.g. Belisle et al., 1988; Fricker etal., 1990). The role of these abnormal rhythms in weight gain is unknown. In thepresent study, a simple hypothesis was tested that high levels of food intake duringthe light period might contribute to excessive weight gain in nocturnal Zucker obeserats. Our approach was to prevent phase-advanced feeding by restricting food intaketo a 14-h night. Over a period of 60 days, nocturnal-fed obese Zucker rats consumedthe same amount of food as did free-feeding obese rats, yet gained 23% less weight.On average, this amounted to a significant, albeit modest 60 g, suggesting that anabnormal concentration of food intake in the day may be a minor contributor toweight gain in the Zucker rat. Additional work will be necessary to determine whatproportion of the weight difference is accounted for by reduced adiposity; however,given that the rats were not food deprived, there is little reason to suspect thatweight loss was primarily in lean body tissue. Previous studies have failed to detectbenefits of restricted food intake on excessive weight gain in Zucker rats (Cleary etal., 1980), but the present study is the first to restrict the timing rather than theamount of food in this animal model. A study of the development of obesity inventromedial hypothalamic-ablated rats found no beneficial effect of restricting foodintake to the day or the night (Gold et al., 1974), suggesting that the timing of foodintake, while of measurable importance in Zucker rats, may have little or nosignificance for obesitys that are not characterized by a mutation in the leptin system.

264 R. E. MISTLBERGER ET AL.
The mechanism by which nocturnal feeding attenuates weight gain in Zuckerrats is unknown. One possibility is that the obese, nocturnal-fed rats were moreactive during the day, owing to short-term food deprivation. However, a previousstudy has reported no significant reductions in body weight of obese Zucker ratsdespite a ten-fold increase in wheel running (Enns et al., 1982), suggesting thatincreases in activity would have to be quite substantial (and unlikely in the smallholding cages that we used) before an impact on body weight could be realized.
Alternatively, nocturnal feeding might attenuate weight gain by normalizing thephase relation between circadian rhythms of food intake and metabolic processesresponsible for the utilization and storage of ingested fuels. This “circadian de-synchronization” hypothesis attributes obesity, at least in minor part, to an un-favourable phase relation between food intake and metabolism. A mechanism bywhich time of feeding might affect weight has been demonstrated in humans; thethermogenic response to food intake is reduced late in the day and at night, i.e. foodintake at the wrong circadian phase should predispose toward weight gain (Romonet al., 1993). The Zucker fatty rat may be susceptible to benefits of restrictednocturnal feeding because of the leptin receptor defect. Leptin normally increasesenergy expenditure at the minimum of the daily cycle of metabolic rate (Stehling etal., 1996), which occurs during the mid-to-late lights-on period in rats. Leptininsensitivity should thus reduce diurnal basal energy expenditure while increasingappetite. Excessive diurnal feeding would therefore be expected to contribute dis-proportionately to total daily weight gain, and preventing feeding at this time, evenwithout reducing total daily food intake, would thus be expected to attenuate weightgain, as was observed in the present study. Concurrent measurements of dailyrhythms of ingestive behaviour, metabolism and thermogenic responses to foodintake will be necessary to evaluate this hypothesis more directly.
C
It remains unclear whether alterations in circadian organization are causal orepiphenomenal to corpulence status. The present results provide modest support fora hypothesis that the timing of food intake impacts on the development of excessiveweight gain in rats bearing a mutation in the leptin receptor, and, taken togetherwith several studies relating ingestive rhythms to obesity in humans (Belisle et al.,1988; Fricker et al., 1990; Spaggiari et al., 1994), suggest that more attention tocircadian factors in weight regulation may be warranted. The recent observationthat there is a circadian variation in serum leptin in humans (Sinha et al., 1996),and that the nocturnal rise is inversely related to body fat gain in early adolescence,adds support to this argument (Matkovic et al., 1997).
R
Albers, H. E. & Ferris, C. F. (1984). Neuropeptide Y: role in light–dark cycle entrainment ofhamster circadian rhythms. Neuroscience Letters, 50, 163–168.
Balagura, S. & Devenport, L. D. (1970). Feeding patterns of normal and ventromedialhypothalamic lesioned male and female rats. Journal of Comparative and PhysiologicalPsychology, 71, 357–364.

265CIRCADIAN RHYTHMS IN ZUCKER OBESE RATS
Beck, B., Burlet, A., Nicolas, P. C. & Burlet, C. (1990). Hyperphagia in obesity is associatedwith a central peptidergic dysregulation in rats. Journal of Nutrition, 120, 806–811.
Becker, E. E. & Grinker, J. A. (1977). Meal patterns in the genetically obese Zucker rat.Physiology and Behavior, 18, 685–692.
Belisle, F., Rolland-Cachera, M. F., Deheeger, M. & Guilloud-Bataille, M. (1988). Obesityand food intake in children: evidence for a role of metabolic and/or behavioral dailyrhythms. Appetite, 11, 111–118.
Bray, G. A. (1977). The Zucker fatty rat: a review. Federation Proceedings, 36, 148–153.Castonguay, T. W., Upton, D. E., Leung, P. M. B. & Stern, J. S. (1982). Meal patterns in the
genetically obese Zucker rat: a reexamination. Physiology and Behavior, 28, 911–916.Chua, S. C., Chung, W. K., Wu-Peng, X. S., Zhang, S., Liu, L., Tartaglia, L. & Leibel, R.
L. (1996). Phenotypes of mouse diabetes and rat fatty due to mutations in the OB (leptin)receptor. Science, 271, 994–996.
Cleary, M. P., Vasselli, J. R. & Greenwood, M. R. C. (1980). Development of obesity inZucker obese (fafa) rat in absence of hyperphagia. American Journal of Physiology, 238,E284–292.
Cote, N. K. & Harrington, M. E. (1993). Histamine phase shifts the circadian clock in amanner similar to light. Brain Research, 613, 149–151.
Cusin, I., Rohner-Jeanrenaud, F., Stricker-Krongrad, A. & Jeanrenaud, B. (1996). The weightreducing effect of an intracerebroventricular bolus injection of leptin in genetically obsefa/fa rats: reduced sensitivity compared with lean animals. Diabetes, 45, 1446–1451.
Danguir, J. (1989). Sleep patterns in the genetically obese Zucker rat effect of acarbosetreatment. American Journal of Physiology, 256, R281–283.
Demes, G. L., Buskirk, E. R., Alpert, S. S. & Loomis, J. L. (1991). Energy turnover and heatexchange in mature lean and obese Zucker rats acutely exposed to three environmentaltemperatures for 24 h. International Journal of Obesity, 15, 375–385.
Dorrscheidt, G. T. & Beck, L. (1975). Advanced methods for evaluating characteristicparameters (d,a,r,) of circadian rhythms. Journal of Mathematical Biology, 12, 107–121.
Enns, M. P. & Grinker, J. A. (1983). Dietary self-selection and meal patterns of obese andlean Zucker rats. Appetite, 4, 281–293.
Enns, M. P., Wecker, J. & Grinker, J. A. (1982). Interrelationships among activity, food intakeand weight gain in genetically obese and lean Zucker rats. Physiology and Behavior, 28,1059–1064.
Ferrari, E., Fraschini, F. & Brambilla, F. (1990). Hormonal circadian rhythms in eatingdisorders. Biological Psychiatry, 27, 1007–1020.
Frederich, R. C., Lollmann, B., Hamann, A., Napolitano-Rosen, A., Kahn, B. B., Lowell,B. B. & Flier, J. S. (1995). Expression of OB mRNA and its encoded protein in rodents.Journal of Clinical Investigation, 96, 1658–1663.
Fricker, J., Giroux, S., Fumeron, F. & Apfelbaum, M. (1990). Circadian rhythm of energyintake and corpulance status in adults. International Journal of Obesity, 14, 387–393.
Fukagawa, K., Sakata, T., Yoshimatsu, H., Fujimoto, K. & Shiraishi, T. (1988). Disruptionof light–dark cycle of feeding and drinking behavior, ambulatory activity induced bydevelopment of obesity in the Zucker rat. International Journal of Obesity, 12, 481–490.
Fukagawa, K., Sakata, T., Yoshimatsu, H., Fujimoto, K., Uchimura, K. & Asano, C. (1992).Advance shift of feeding circadian rhythm induced by obesity progression in Zucker rats.American Journal of Physiology, 263, R1169–1175.
Gold, R. M., Sumprer, G., Ueberacher, H. & Kapatos, G. (1974). Hypothalamic hyperphagiadespite imposed diurnal or nocturnal feeding and drinking rhythms. Physiology andBehavior, 14, 861–865.
Grinker, J. A., Drewnowski, A., Enns, M. & Kissileff, H. (1979). Effects of d-amphetamineand fenfluramine on feeding patterns and activity of obese and lean Zucker rats.Pharmacology, Biochemistry and Behavior, 12, 265–275.
Ho, A. & Chin, A. (1988). Circadian feeding and drinking patterns of genetically obese micefed solid chow diet. Physiology & Behavior, 43, 651–656.
Itowi, N., Yamatodani, A., Nagai, K., Nakagawa, H. & Wada, H. (1990). Effects of histamineand alpha-fluoromethylhistidine injections on circadian phase of free-running rhythms.Physiology and Behavior, 47, 59–554.

266 R. E. MISTLBERGER ET AL.
Keesey, R. A., Swiergiel, A. H. & Corbett, S. W. (1990). Contribution of spontaneous activityto daily energy expenditure of adult obese and lean Zucker rats. Physiology and Behavior,48, 327–331.
Lee, A., Ader, M, Bray, G. A. & Bergman, R. N. (1992). Diurnal variation in glucose tolerancecyclic suppression of insulin action and insuling secretion in normal weight, but not obesesubjects. Diabetes, 41, 742–749.
Lennie, T. A., Rowsey, P. J., Cimprich, B. & Metzger, B. L. (1995). Impaired circadian rhythmresponse for body temperature and activity of obese Zucker rats to reversal of thelight–dark cycle. Society for Neuroscience Abstracts, 21, 954.
Machidori, H., Sakata, T., Yoshimatsu, H., Ookuma, K., Fujimoto, K., Kurodawa, M.,Yamatodani, A. & Wada, H. (1992). Zucker obese rats: defect in brain histamine controlof feeding. Brain Research, 590, 180–186.
Maffei, M., Halass, J., Rayussin, E., Pratley, R. E., Lee, G. M., Zhang, Y., Fei, H., Kim, S.,Lallone, R., Ranganathan, S., Kern, P. A. & Friedman, J. M. (1995). Leptin levels inhuman and rodent: measurements of plasma leptin and ob RNA in obese and weight-reduced subjects. Nature Medicine, 1, 1155–1161.
Martin, R. J., Stolz, D. J. & Buck, D. C. (1979). Diurnal changes in adipose and liver tissuemetabolism of lean and obese Zucker rats. Journal of Nutrition, 109, 412–417.
Matkovic, V., Ilich, J. Z. Badenhop, N. E., Skugor, M., Clairmont, A., Klisovic, D. & Landoll,J. D. (1997). Gain in body fat is inversely related to the nocturnal rise in serum leptinlevel in young females. Journal of Clinical Endocrinology and Metabolism, 82, 1368–1372.
McLaughlin C. L. & Baile, C. A. (1981). Ontogeny of feeding behavior in the Zucker obeserat. Physiology and Behavior, 36, 607–612.
McKibbin, P. E., Cotton, S. J., Mcmillan, S., Holloway, B., Mayers, R., McCarthy, H. D. &Williams, G. (1991). Altered neuropeptide-Y concentrations in specific hypothalamicregions of obese Zucker rats—possible relationship to obesity and neuroendocrinedisturbances. Diabetes, 40, 1423–1429.
Murakami, D. M., Horwitz, B. A. & Fuller, C. A. (1995). Circadian rhythms of temperature andactivity in obese and lean Zucker rats. American Journal of Physiology, 269, R1038–1043.
Pittendrigh, C. S. & Daan, S. (1976). A functional analysis of circadian pacemakers in nocturnalrodents: IV. Entrainment: pacemaker as clock. Journal of Comparative Physiology A,106, 291–331.
Prins, A. A., de Jong-Nagelsmit, A., Keijser, J. & Strubbe, J. H. (1986). Daily rhythms offeeding in the genetically obese and lean Zucker rats. Physiology and Behavior, 38,423–426.
Rietveld, W. J., Flory, W., Kooij, M. & ten Hoor, F. (1980). Observations on total food intakeand 24 h food intake patterns during long term registration of food intake behavior inrats. Versuchstierk, 22, 156–160.
Romon, M., Edme, J. L., Bouloenguez, C., Lescroart, J. L. & Frimat, P. (1993). Circadianvariation of diet-induced thermogenesis. American Journal of Clinical Nutrition, 57,476–480.
Schwartz, M. W., Seeley, R. J., Campfield, L. A., Burn, P. & Baskin, D. G. (1996). Identificationof targets of leptin action in rat hypothalamus. Journal of Clinical Investigation, 98,1101–1106.
Seeley, R. J., van Dijk, G., Campfield, L. A., Smith, F. J., Burn, P., Nelligan, J. A., Bell,S. M., Baskin, D. G., Woods, S. C. & Schwartz, M. W. (1996). Intraventricular leptinreduces food intake and body weight of lean rats but not obese Zucker rats. Hormoneand Metabolic Research, 28, 664–668.
Sinha, M. K., Ohannesian, J. P., Heiman, M. L., Kriauciunas, A., Stephans, T. W., Magosin,S., Marco, C. & Caro, J. F. (1996). Nocturnal rise of leptin in lean, obese and non-insulin-dependent diabetes mellitus subjects. Journal of Clinical Investigation, 97, 1344–1347.
Spaggiari, M. C., Granella, F., Parrino, L., Marchesi, C., Melli, I. & Terzano, M. G. (1994).Nocturnal eating syndrome in adults. Sleep, 17, 339–344.
Spiteri, N. J., Prins, A. A., Keyser, J. & Strubbe, J. H. (1982). Circadian pacemaker controlof feeding in the rat at dawn. Physiology and Behavior, 29, 1141–1145.
Stehling, O., Doring, H., Ertl, J., Preibisch, G. & Schmidt, I. (1996). Leptin reduces juvenilefat stores by altering the circadian cycle of energy expenditure. American Journal ofPhysiology, 271, R1770–1774.

267CIRCADIAN RHYTHMS IN ZUCKER OBESE RATS
Vachon, C. & Savoi, L. (1987). Circadian variation of food intake and digestive tract contentsin the rat. Physiology and Behavior, 39, 629–632.
Van Cauter, E., Polonsky, K. S., Blackman, J. D., Roland, D., Sturris, J., Byrne, M. M. &Scheen, A. J. (1994). Abnormal temporal patterns of glucose tolerance in obesity:relationship to sleep-related growth hormone secretion and circadian cortisol rhythmicity.Journal of Clinical Endocrinology and Metabolism, 79, 1797–1805.
Velasco Plaza, A., Granda, T. & Cachero, M. T. G. (1993). Circadian rhythms of food andwater intake and urine excretion in diabetic rats. Physiology and Behavior, 54, 665–670.
Wangsness, P. J., Dilettuso, B. A. & Martin, R. J. (1978). Dietary effects on body weight,feed intake and diurnal feeding behavior of genetically obese rats. Journal of Nutrition,108, 256–264.
Watts, A. G. (1991). The efferent projections of the suprachiasmatic nucleus: anatomicalinsights into the control of circadian rhythms. In D. C. Klein, R. Y. Moore & S. M.Reppert (Eds), The suprachiasmatic nucleus: the mind’s clock. Pp. 77–106. New York:Oxford University Press.
Zhang, Y., Proeca, R., Maffie, M., Barone, M., Leopold, L. & Friedman, J. M. (1994).Positional cloning of the mouse obese gene and its human homologue. Nature, 372,425–432.
Zucker, I. (1971). Light–dark rhythms in rat eating and drinking behavior. Physiology andBehavior, 6, 115–126.
Received 6 May 1997, revision 25 August 1997
Copyright © 2022 FDOKUMEN




















