
Redox and Antioxidant Modulation of Circadian Rhythms - MDPI
Upload
independentCategory
view
0download
0
Cortisol circadian rhythms and responseto stress in children with autism*
Blythe A. Corbetta,b,*, Sally Mendozac, Maryam Abdullahb,Jacob A. Wegelind, Seymour Levinea
aDepartment of Psychiatry and Behavioral Sciences, University of California at Davis,2825 50th Street, Sacramento, CA 95817, USAbThe M.I.N.D. Institute, University of California at Davis, Sacramento, CA 95817, USAcDepartment of Psychology, University of California at Davis, Sacramento, CA 95817, USAdDivision of Biostatistics, University of California at Davis, Sacramento, CA 95817, USA
Received 18 October 2004; received in revised form 12 May 2005; accepted 17 May 2005
03do
*
Da
KEYWORDSAutism;Cortisol;Circadian variations;Stress;LHPA
06-4530/$ - see front matter Q 200i:10.1016/j.psyneuen.2005.05.011
National Institute of Health MHO65* Corresponding author. Address: Devis, 2825 50th Street, Sacramento,E-mail address: blythe.corbett@uc
Summary Background: Autism is a severe neurodevelopmental disorder charac-terized by impairment in communication, social interaction, repetitive behaviorsand difficulty adapting to novel experiences. The Hypothalamic-Pituitary-Adreno-cortical (HPA) system responds consistently to perceived novel or unfamiliarsituations and can serve as an important biomarker of the response to a variety ofdifferent stimuli. Previous research has suggested that children with autism mayexhibit dysfunction of the HPA system, but it is not clear whether alteredneuroendrocrine regulation or altered responsiveness underlies the differencesbetween children with and without autism. In order to provide preliminary dataconcerning HPA regulation and responsiveness, we compared circadian rhythms andresponse to a non-social, environmental stressor in children with and without autism.
Methods: Circadian rhythms of cortisol were estimated in children with (NZ12)and without (NZ10) autism via analysis of salivary samples collected in the morning,afternoon and evening on 2 consecutive days. HPA responsiveness was assessed byexamining the time course of changes in salivary cortisol in response to a mock MRI.
Results: Both groups showed expected circadian variation with higher cortisolconcentration in morning than in the evening samples. The children with autism, butnot typical children, showed a more variable circadian rhythm as well as statisticallysignificant elevations in cortisol following exposure to a novel, nonsocial stimulus.
Conclusions: The results suggest that children with autism process and respondidiosyncratically to novel and threatening events resulting in an exaggerated cortisolresponse.Q 2005 Elsevier Ltd. All rights reserved.
Psychoneuroendocrinology (2006) 31, 59–68
www.elsevier.com/locate/psyneuen
5 Elsevier Ltd. All rights reserved.
672 (P.I. David G. Amaral).partment of Psychiatry and Behavioral Sciences, M.I.N.D. Institute, University of California atCA 95817, USA.dmc.ucdavis.edu (B.A. Corbett).

B.A. Corbett et al.60
1. Introduction
Autism is a severe neurodevelopmental disordercharacterized by qualitative impairment before theage of three in verbal and nonverbal communi-cation, reciprocal social interaction, and a mark-edly restricted repertoire of activities and interests(APA, 1994). In addition to these features, childrenwith autism have been described as experiencingdifficulty tolerating novelty and environmentalstressors (Kanner, 1943). Amongst the two mostfrequently used indices of the response to stresshave been observable changes in the behavior of anindividual experiencing stress and the markedincrease in the stress related hormones. When thebehavior and the endocrine markers of stress areconcordant there is little problem in interpretingthe outcome. However, there are examples ofdissociation between these behavioral and endo-crine measures (e.g. Wiener et al., 1988). Inchildren with autism, there are excessive beha-vioral reactions to stressful circumstances, but it isless clear whether or not this is accompanied by acorresponding increase in neuroendocrine activity.
The hypothalamic-pituitary-adrenal (HPA) axis isthe classical endocrine stress system. Although thelabel HPA is still used to describe this system, it isabundantly clear that the regulation of the HPA axisinvolves a complex neural system involving manydifferent anatomical structures and neurochemicalevents that are required to activate and/or inhibitthe HPA axis. Adrenocorticotropic hormone (ACTH)is regulated by two primary molecules at opposingends of the system, which are corticotropinreleasing hormone (CRH) in the brain, whichactivates both behavioral and hormonal stressresponses and glucocorticoids (cortisol in humans)secreted from the adrenal which acts primarily toinhibit the release of CRH, ACTH and cortisolthrough its action on the glucocorticoids receptorsin the brain and the pituitary. Thus, one of the mostwidely used biological markers of stress is therelease of cortisol from the adrenal since elevationsoften occur in response to novel and unpredictablesituations (Gunnar and Donzella, 2002; Hennesseyand Levine, 1979).
It has been established that the regulation of theHPA axis is different depending on the type of stressthe organism is exposed to. The most recentdescriptions of these differences have been elabo-rated by Herman and Cullinan (1997). Thus,although seemingly disparate stimuli activate theHPA axis they utilize different pathways. Systemicstressors are physical and context-independent andare capable of activating the HPA system in
unconscious animals. In general, systemic stimuliusually involve a life-threatening event. In the caseof systemic stimuli, the information required toactivate the HPA system is relayed to the periven-tricular nuclei (PVN) of the hypothalamus via thebrain stem. In contrast, processive stimuli arecontext dependent, requiring the comparison ofcurrent information with past experience and theassignment of emotional meaning. Limbic systemstructures are the primary mediators of processivestimuli. Thus the bed nucleus of the stria terminalis(BNST) preoptic nucleus, lateral and medialseptum, the amygdala, prefrontal cortex andhippocampus all are involved in either activationor inhibition of the stress response to processivestimuli. Given the importance of the limbic systemin the regulation of the stress response it hasbecome more common to refer to the stressresponsive system as the LHPA axis. The ‘L’ refersto the limbic system.
There is evidence that regulation of the HPAsystem may be dysfunctional in children withautism (Nir et al., 1995; Richdale and Prior, 1992;Yamazaki et al., 1975). Most of the publishedreports examining HPA responsiveness in childrenwith autism have serious methodological flaws,which include a lack of appropriate controls,invasive sampling techniques, mixed diagnosis andvery small samples. Children with autism have beenshown to exhibit an exaggerated stress response asevidenced by increases in ACTH and beta-endorphinfollowing an injection of insulin (Maher et al.,1975). In this investigation, there were no controlsfor the effects of venipuncture per se. Tordjmanet al. (1997) and Curin et al. (2003) report higherlevels of ACTH in autism. These data wereinterpreted as indicating that individuals withautism have a chronic level of anxiety. However,in these studies cortisol was not different or lowerthan controls. Insofar as the release of ACTH ismuch more rapid than cortisol, it appears morelikely that the higher ACTH levels observed in thesubjects with autism was a result of the response tovenipuncture and restraint required to obtain bloodsamples. Richdale and Prior (1992) provideadditional support for the hypothesis that childrenwith autism may be hyperresponsive to environ-mental stress. Subjects with autism who wereintegrated into the regular school system showedhypersecretion of cortisol suggesting an environ-mental stress response in the school environment.In contrast, a recent report showed a lack ofsalivary cortisol elevation or ‘hyporesponsiveness’in a group of autistic-like children diagnosed withMultiple Complex Developmental Disorder (MCDD;Jansen et al., 2000). These investigators exposed

Cortisol circadian rhythms and response to stress in children with autism 61
children diagnosed as MCDD to a stressor involvingpublic speaking (a modified Trier Social Stress Test)and hypothesized that these children may beprocessing social stimuli differently and, therefore,tend to show a diminished salivary cortisolresponse. In a more recent investigation, Jansenet al. (2003) demonstrated that unlike the reducedcortisol response shown in the MCDD children, thechildren with autism showed a relatively elevatedcortisol response to psychosocial stress but werenot significantly different from neurotypicalchildren.
Although the data are by no means consistent,alterations in the normal circadian patterns ofcortisol have been reported in children with autism(Aihara and Hashimoto, 1989; Hill et al., 1977;Hoshino et al., 1987; Richdale and Prior, 1992;Tordjman et al., 1997; Yamazaki et al., 1975). Ingeneral, where alterations in circadian rhythms arereported in children with autism, they seem to bemore apparent in children identified as being lowfunctioning (Jensen et al., 1985). It has also beenreported that children with autism show poornegative feedback regulation. In one study, Hoshinoand colleagues (Hoshino et al., 1987) indicated thatsome children with autism showed abnormal diurnalrhythm and/or a failure to suppress cortisol inresponse to dexamethasone, a synthetic glucocor-ticoid that serves as a potent negative feedbacksignal (Jensen et al., 1985). The results suggest thatnegative feedback mechanisms of the HPA axis maybe inefficient in children with autism, which wouldbe expected to result in prolonged cortisolelevations following activation of the stressresponse. It is possible, therefore, that some claimsof increased HPA responsivity in children withautism are actually due to decreased negativefeedback.
The current study was undertaken to evaluatethe responsiveness and regulation of the HPA axis ina group of relatively high functioning children withautism. In order to avoid stress associated withcollection of blood samples, we measured cortisolin saliva, a noninvasive method for obtainingmultiple samples (Kirschbaum and Hellhammer,1994). It has been determined that cortisol issequestered in saliva, exists in the unbound formand shows detectable changes after about 20 min.Resting levels of cortisol vary predictably as afunction of the diurnal cycle. Thus, in using cortisolas a measure of stress responsiveness we deemed itnecessary to examine the basic regulatory aspectsof the system by obtaining diurnal cortisol samplesfor comparison. Samples obtained over a 48-h cyclewere used to characterize circadian rhythms. Thisalso provided a stable basal level by which to
compare the response to arrival at the laboratory.The response to stress was examined by exposingthe children for twenty minutes to a mock MRIscanner. We reasoned that this manipulation wouldinclude exposure to novelty as well as mild restraintthat would result in the activation of the HPA axis.Finally, cortisol measures were obtained for anextended period of time following the terminationof the stressor to evaluate negative feedback. Thisdesign also makes it possible to study therelationship between presumed basal cortisol levelsobtained in the home to the adrenocorticalresponse to stress in the laboratory. Based uponthe results of previous experiments we predictedthat children with autism would: (1) exhibit normalcircadian patterns of cortisol, (2) show an exagger-ated response to stress reflecting enhanced acti-vation, and (3) would return to basal levels moreslowly reflecting diminished negative feedback.
2. Methods
2.1. Subjects
Twenty-four male children entered the study andparticipants met criteria for categorization intothe two groups: Autism or typically developing(Typical). Two typically developing children weredropped from the study due to parental noncom-pliance with procedures. Thus, the subjectsconsisted of 22 male children between 6- and11-years of age (mean age 8.8 years), 12 diagnosedwith autism (mean ageZ8.5) and 10 neurotypicalchildren (mean ageZ9.2 years). The children withautism were diagnosed using previous records orconcurrent testing with the Autism DiagnosticObservation Schedule (ADOS; Lord et al., 1999)and clinical judgment based on DSM-IV criteria(APA, 1994). The ADOS is comprised of semi-structured interactive activities conducted with achild and designed to assess specific currentbehaviors indicative of autism. The activitiesfocus upon the three areas of impairmentassociated with autism: social functioning, commu-nicative functioning, and restricted activities. TheADOS provides an algorithm with cut-offs for autismspectrum disorders (Lord et al., 1999). The typicallydeveloping children were screened via parentinterview for the absence of neurodevelopmentaldisorders, including autism. All subjects were alsoscreened for current and past physical illness.Children with known endocrine, cardiovascular,pulmonary, liver or kidney disease or neurologicaldisorders (e.g. seizures) were excluded from

B.A. Corbett et al.62
enrollment in the study. The groups were matchedon age and gender but not intellectual functioning.The children with autism had an average IQ of 77and the typically developing children had anaverage IQ of 114. A consistent intelligencemeasure was not used across subjects; thus, IQestimates were based on previous records usingdifferent instruments. All subjects were recruitedfrom the University of California, Davis, M.I.N.D.Institute, local school districts and communitycenters. Informed written consent was obtainedfrom parents and verbal assent was obtained fromall research subjects prior to inclusion in the study.The Institutional Review Board of the University ofCalifornia, Davis approved the study.
2.2. Psychological measures
The parents of the participants completed thefollowing questionnaires to ascertain the child’sbehavioral responses to stress and sensorysensitivity across a variety of situations.
The Stress Survey Schedule (SSS; Groden et al.,2001) is a parent-report measure of stress designedfor individuals with autism and other developmen-tal disabilities. The SSS consists of 60 daily stress-related items rated on a five-point likert scale andincludes eight dimensions of stress. Internalconsistency correlations range from .70 to .87. Inthe current study, only the total SSS score was used,as an index of the subject’s observed response tostress across a variety of situations.
The Short Sensory Profile (SSP; Dunn, 1999) is aparent questionnaire related to sensory sensitivityacross several domains, including auditory, visual,vestibular, tactile, oral and multisensoryprocessing. The internal reliability correlations forthe total test and section range from .70 to .90. Theinternal validity correlations for the sensorysections range from .25 to .76 suggesting that thesections represent different constructs and supportthe measure’s factor structure. In the currentstudy, only the total score was used.
2.3. Cortisol sampling methods
We used the current preferred procedure in humanstress research by collecting salivary samples forsubsequent assay of cortisol concentrations. A totalof 10 salivary samples were collected from eachresearch subject, which included six samplescollected from the home to obtain the cortisoldiurnal rhythm and four samples collected toevalute the subject’s response to stress. Parentswere trained on the collection procedures and they
were provided with sample tubes and preprintedlabels. Tubes were stored in a container fitted witha Trackcape (distributed by Aprexw, Union City,CA), which recorded each time the container wasopened and a tube removed. The participant wasgiven Tridentw Original Sugarless chewing gum toact as a saliva stimulant. After 30 s, the child wasasked to spit the gum out into a tissue. The testtube was removed from the Trackcape container.The child was given a large straw and asked to put itin his mouth while the lower end of the straw wasplaced into the collection tube. The participant wasthen asked to deposit saliva into the tube byspitting. This method of direct sampling into acollection container does not have any measurableeffects on the results of the assays (Schwartz et al.,1998). Once completed, a pre-typed, peel-off label(coded with the date and time of collection) wasplaced on the tube. For home collection, thesamples were placed in the home refrigerator fortemporary storage.
2.3.1. TrackCapeA major complication in these studies is the dangerof parental noncompliance with collection ofmultiple salivary samples. We attempted to addressthis concern by monitoring compliance via carefulinstruction and parental diaries. As an additionalmeans of measuring parental compliance to theoutlined experimental procedures, the emptycollection tubes were contained within a bottlefitted with a TrackCape (distributed by Aprexw,Union City, CA). A TrackCap is a bottle capcontaining microelectronics that records the dateand time for each occasion that the bottle isopened. This record was compared with the parentrecord of each sample collection time.
2.4. Circadian rhythmicity
Basal levels of salivary cortisol were collected fortwo diurnal cycles. Research assistants instructedparents on the basic saliva sampling proceduredescribed above for in-home collection. Parentswere provided with a supply packet containing aTrackCape—fitted container with collection tubes,Tridentw Original Sugarless gum, large straws,storage bag, diary forms and a calendar with pre-numbered, peel-off labels that corresponded to thecollection schedule. Research staff regularly calledparents to ensure adherence and to answerquestions regarding the sampling procedures.Within 48 h of the final saliva sample, the testtube kits were collected by the research assistant,

Cortisol circadian rhythms and response to stress in children with autism 63
placed in a cooler and brought to the EndocrineLaboratory.
Home samples were collected by parents 3 timesper day for 2 consecutive days beginning with themid-afternoon sample on the first day, followed byan evening and a morning sample. For each subject,the samples were collected at approximately thesame time each day. Specifically, the morningsample was collected within a half-hour of wakingprior to the participant eating, drinking or brushinghis teeth. The afternoon sample was collectedbetween 13:00 and 15:00 h avoiding eating for aminimum of 1 h prior to sampling. The eveningsample was collected within a half-hour of bedtimeavoiding eating for an hour before and prior tobrushing teeth. The final morning sample wascollected on the day of the stress procedure.
2.5. Nonsocial environmental stress
This portion of the study was conducted at the UCDavis Imaging Research Center (IRC) which has anMRI stimulator (mock scanner). To evaluateresponse to environmental challenges to a non-social environmental stressor, we obtained salivarysamples from the participants before and afterexposure to the mock scanner. The exposure isconceptualized as a model of environmental stressinsofar as it is a novel event that involves mildrestraint and exposure to the computer-simulatedunpleasant noises generated by the MRI scanner. Itis important to note that the children exposed tothis procedure were not provided with additionalstrategies (e.g. watching a videotape of an MRI) toassist them in habituating to the mock scanningprocedures. All mock scanning sessions occuredbetween 13:00 and 15:00 h. A total of four salivasamples were collected which included: immediatecollection upon arrival to the IRC, 20 min post-exposure to the stressor, 40 min post-exposure and2 h post-exposure sampled from home.
2.6. Mock scanning procedure
The research assistant (RA) demonstrated theprocedure by placing the head coil on her ownhead and lying down on the scanning bed. Weemployed ‘reinforcement sampling’ procedures byexposing the child to a cabinet containing a varietyof non-edible reinforcers for them to choose andearn at the completion of the session. The RA thengently placed earplugs in the ears of the participantexplaining that they would be used to reducethe noise experienced in the scanner. Simulatednoise was generated using various MRI sounds.
The participant was asked to lie on the scanningbed and, once situated, he was subsequently movedinto the mock scanner. The participant wasinstructed to remain still and to not move hishead or extremities during the simulator session.Throughout the simulation, the participants wereasked frequently if they were comfortable. A 1-minmock scan session was initiated to familiarize thechild with the sights and sounds of the procedure.The RA then sat behind the scanner and monitoredhead and extremity movement. Reminders toremain still (and praise for doing so) was providedas needed. If the child’s anxiety became intoler-able, the child was removed from the simulatorupon request of the child, parent or determinationof the research personnel. Two children withautism were unable to tolerate being placed allthe way in the scanner and subsequently wereallowed to simply lie on the bed without full entryinto the scanner. Twenty minutes after initialexposure to the mock scanner the child wasremoved from the scanner and provided a salivarysample. The child was then escorted to the waitingarea and samples were collected 40-min post-exposure at the imaging center. In order to evaluatethe child’s recovery from stress (negative feed-back), salivary samples were collected 2 h post-exposure from the participant’s home.
2.7. Cortisol storage and assays
Cortisol assays were conducted in the EndocrineLaboratory of the University of California, Davis,California National Primate Research Center underthe direction of Dr. Sally Mendoza. Once all of thesamples were collected, they were logged in andstored in a K20 8C freezer until analyzed. Cortisolassays were performed using coated-tube radio-immunoassay (RIA) kits (Diagnostic ProductsCorporation, Los Angeles). Assay procedures wereadjusted to account for the overall lower concen-tration of steroids in saliva as compared to blood.The salivary stimulant (i.e. Tridentw gum) wasanalyzed to ensure that it did not contain cortisol.Prior to assay, the saliva samples were centrifugedat 6000 rpm for 10 min to separate aqueous com-ponent from mucins and other suspended particles.
2.8. Analysis
Statistical analysis was performed with thefollowing objectives: to characterize circadianpatterns of variation, response to stress, andrecovery; to compare these patterns betweendiagnostic groups (Autism/Typical); to examine

B.A. Corbett et al.64
the association between circadian patterns andstress response; and to assess the relationshipbetween age and parent-report measures on theone hand and cortisol patterns on the other. Linearmixed-effects models were fit by restricted maxi-mum likelihood (REML) using R software version2.0.1 (R Development Core Team, 2005). Allavailable measurements were used in each model,including those measuring diurnal rhythm and thosemeasuring stress response and recovery. Thismethod has been used in a comparable studyinvolving longitudinal measurements of salivarycortisol on children, Hessl et al. (2002). The linearmixed-effects approach allows us to estimateindividual (random) effects for each child, whilealso producing average estimates of these quan-tities for each group (Autism/Typical). In addition,estimates of the covariances between randomeffects are available, and it is possible to comparevariances of the random effects between groups(Laird and Ware, 1982).
The response (outcome) variable was log salivarycortisol concentration. Salivary cortisol measure-ments are positive and skewed toward large values,and therefore they were log transformed beforeanalysis to achieve approximate normality. Sub-sequently, diagnostic plots revealed no departuresfrom normality. For graphical presentation, theoriginal scale (nmol/l) is displayed on the verticalaxis.
A single cortisol measurement was missing, viz.,the last post-stress measurement for one child inthe Autism group. In the current family of models,however, parameters can be estimated even ifmeasurements are missing for a small number oftime points. In such situations, it is not necessary todiscard the same time point for all subjects or todiscard an entire subject from the analysis (Mal-linckrodt et al., 2003).
Fixed effects were group (Autism or Typical),time of day (morning, afternoon, or evening)1,sample point post-stressor, and age. It was notpossible, however, to include fixed effects for allthree sample points post-stress because the first andsecond points (20 and 40 min) were nearly pro-portional to each other (rZ0.995). Consequently,these two time points were included as a single fixedeffect (called Stress), linked by a constant ofproportionality that was determined by profilelikelihood (Murphy et al., 2000). The sample pointat 120 min post-stress was included as a fixed effect
1 With Helmert contrasts, the first effect for time of day wasthe difference between morning and afternoon, and the secondeffect was the difference between the evening and the mean ofthe other 2 times of day (morning and afternoon).
(called Recovery). The Stress and Recovery effectsmeasured differences from the afternoon baselinelevel, since this is when the mock scanningprocedure and subsequent cortisol measurementstook place. Interactions of group with time of day,Stress, and Recovery, and the interaction betweenStress and age, were included in the model.
Three random effects were fit for each subject:overall level (intercept), stress level, and level atrecovery. In addition to a standard deviation for allthe levels, separate standard deviations for thelevels in each group were estimated and the ratio ofthese standard deviations (Autism to Typical) wascomputed. In addition to the p-values for the fixedeffects returned by the regression software, wecomputed p-values by the bootstrap (Efron andTibshirani, 1993) and compared these with theformer p-values. To examine the sensitivity of theresults to influential subjects or multivariate out-liers, each subject was removed in turn from thedata, creating each time a data set with only 21subjects. For each such abridged sample, themodel was recomputed, and the p-values werecompared with those obtained from the full dataset. In a linear mixed-effects model, likelihood-based estimates of variances and covariances ofrandom effects may be somewhat unstable. Accord-ingly, we used the bootstrap to perform inferencesregarding these parameters, as follows. To assesswhether the standard deviations of the randomoverall levels (intercepts) differed between theAutism and Typical groups, a 95% confidenceinterval for the ratio of the standard deviations,and a two-sided p-value to test the null hypothesisthat the ratio was one, were computed by thebootstrap. To test for a relationship betweenchild’s overall level and stress response, a 95%confidence interval was computed for the corre-lation between child’s overall level and child’sstress response. Similarly, a bootstrap confidenceinterval was computed for the correlation betweenchild’s overall level and child’s recovery level.
To test for a relationship between parent reportmeasures and cortisol patterns, an additional modelwas fit similar to the main model, but with theStress Survey Schedule total score and the ShortSensory Profile total score, and their interactionswith Stress, added as fixed effects. One participantin the Autism group did not have these measures,and consequently these measures were notincluded in the main model.
To test for the possibility of increased ordecreased cortisol levels on the morning of thestressor, an additional model was fit with a dummyvariable for that sample point and the interaction ofthat variable with the group variable.
Time (minutes)
Sal
ivar
y C
ortis
ol (
nmol
/l)
Mean Stress Response
2.5
3.1
3.9
4.8
6
0 20 40 120
AutismTypical
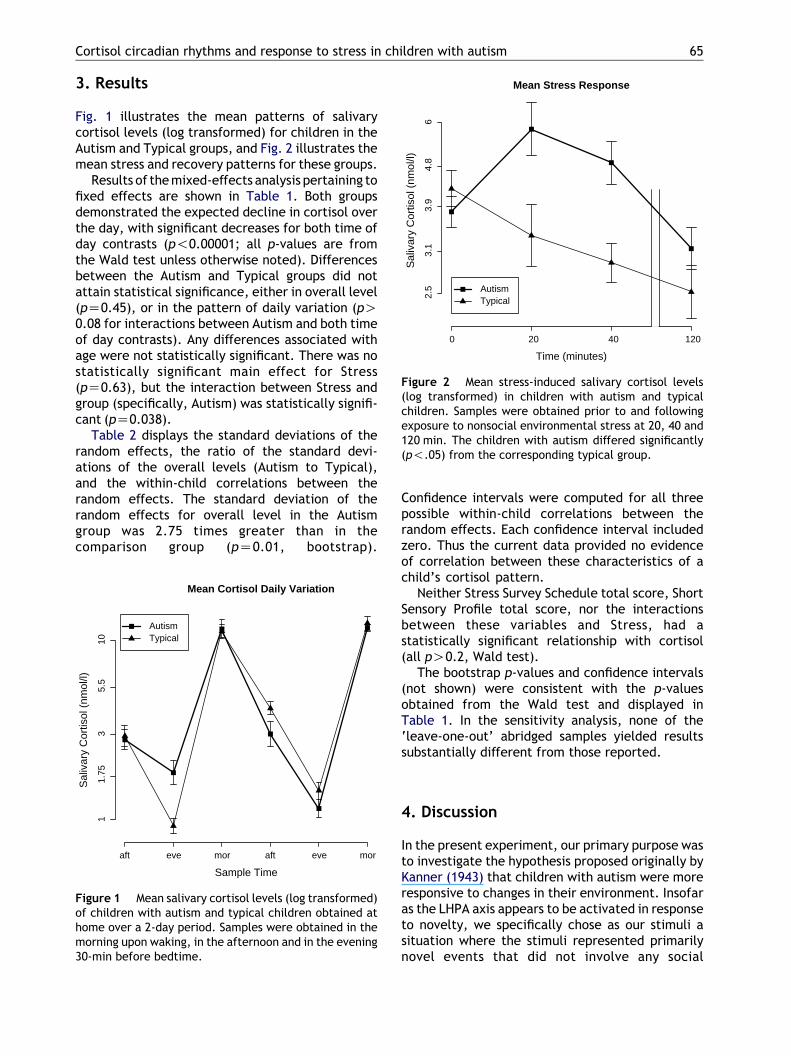
Figure 2 Mean stress-induced salivary cortisol levels(log transformed) in children with autism and typicalchildren. Samples were obtained prior to and followingexposure to nonsocial environmental stress at 20, 40 and120 min. The children with autism differed significantly(p!.05) from the corresponding typical group.
Cortisol circadian rhythms and response to stress in children with autism 65
3. Results
Fig. 1 illustrates the mean patterns of salivarycortisol levels (log transformed) for children in theAutism and Typical groups, and Fig. 2 illustrates themean stress and recovery patterns for these groups.
Results of the mixed-effects analysis pertaining tofixed effects are shown in Table 1. Both groupsdemonstrated the expected decline in cortisol overthe day, with significant decreases for both time ofday contrasts (p!0.00001; all p-values are fromthe Wald test unless otherwise noted). Differencesbetween the Autism and Typical groups did notattain statistical significance, either in overall level(pZ0.45), or in the pattern of daily variation (pO0.08 for interactions between Autism and both timeof day contrasts). Any differences associated withage were not statistically significant. There was nostatistically significant main effect for Stress(pZ0.63), but the interaction between Stress andgroup (specifically, Autism) was statistically signifi-cant (pZ0.038).
Table 2 displays the standard deviations of therandom effects, the ratio of the standard devi-ations of the overall levels (Autism to Typical),and the within-child correlations between therandom effects. The standard deviation of therandom effects for overall level in the Autismgroup was 2.75 times greater than in thecomparison group (pZ0.01, bootstrap).
Sample Time
Sal
ivar
y C
ortis
ol (
nmol
/l)
Mean Cortisol Daily Variation
11.
753
5.5
10
aft eve mor aft eve mor
AutismTypical
Figure 1 Mean salivary cortisol levels (log transformed)of children with autism and typical children obtained athome over a 2-day period. Samples were obtained in themorning upon waking, in the afternoon and in the evening30-min before bedtime.
Confidence intervals were computed for all threepossible within-child correlations between therandom effects. Each confidence interval includedzero. Thus the current data provided no evidenceof correlation between these characteristics of achild’s cortisol pattern.
Neither Stress Survey Schedule total score, ShortSensory Profile total score, nor the interactionsbetween these variables and Stress, had astatistically significant relationship with cortisol(all pO0.2, Wald test).
The bootstrap p-values and confidence intervals(not shown) were consistent with the p-valuesobtained from the Wald test and displayed inTable 1. In the sensitivity analysis, none of the‘leave-one-out’ abridged samples yielded resultssubstantially different from those reported.
4. Discussion
In the present experiment, our primary purpose wasto investigate the hypothesis proposed originally byKanner (1943) that children with autism were moreresponsive to changes in their environment. Insofaras the LHPA axis appears to be activated in responseto novelty, we specifically chose as our stimuli asituation where the stimuli represented primarilynovel events that did not involve any social

Table 1 Fixed effects from mixed-effects linear regression (REML) fit.
Parameter Estimate Std. error DF t p (Wald test)
Intercept 1.29 0.0891 188 14.4 !0.00001Autism 0.146 0.19 19 0.771 0.45Time of day 1 K0.583 0.0706 188 K8.25 !0.00001Time of day 2 K0.581 0.0435 188 K13.4 !0.00001Stress K0.626 1.3 188 K0.482 0.631Recovery K0.384 0.196 188 K1.96 0.0516Age K0.00813 0.0444 19 K0.183 0.857Autism!time ofday 1
K0.0695 0.0956 188 K0.727 0.468
Autism!time ofday 2
0.104 0.0589 188 1.76 0.0803
Autism!stress 3.67 1.75 188 2.09 0.0378Autism!recovery 0.351 0.269 188 1.3 0.194Stress!age K0.44 0.505 188 K0.872 0.384
Model fit: AICZ432.51, BICZ509.16, LogLikelihoodZK193.25.
Table 2 Standard deviations of random effects,ratio of SDs between groups, and correlations betweenrandom effects, with 95% bootstrap confidenceintervals.
Lower Estimate Upper
SDs of random effectsOverall level
Combined 0.245 0.404 0.511Typical 0.0263 0.198 0.310Autism 0.315 0.545 0.662
Stress 0.786 2.05 3.08Recovery 0.0021 0.251 0.437
Ratio of SDs of overall levelsAutism/typical 1.36 2.75 18.8
CorrelationsLevel with stress K0.595 0.134 0.776Level with recovery K0.985 K0.172 0.792Recovery with stress K0.969 K0.283 0.967
B.A. Corbett et al.66
parameters. It has been well established that one ofthe features of children with autism is a deficit inprocessing social stimuli. For this experiment wespecifically wanted to examine the responses toenvironmental changes.
We did not find statistically significant differencesin circadian variation in terms of peak in the morningand trough in the evening between the groups. Thus,our results do not replicate previous reports showingalterations in the normal circadian patterns ofcortisol in individuals with autism (Aihara andHashimoto, 1989; Hill et al., 1977; Hoshino et al.,1987; Yamazaki et al., 1975). On the other hand, thebetween-child variation in cortisol was greater inthe autism group than among the typically develop-ing children suggesting more variability in circadiancortisol. This variability may be the result ofheightened sensitivity to daily changes and unpre-dictable events in the environment.
As hypothesized, the majority of the childrenwith autism (8 out of 12 subjects) demonstrated anincrease in cortisol after exposure to the non-socialstressor. In contrast, the majority of the typicallydeveloping children showed no response or areduction in cortisol after being placed in the mockMRI. Thus, the response of the children with autismis even more profound insofar as the majority of theAutism group respond with increase in cortisol incontrast to the Typical group that show no responseor an actual reduction in cortisol level followingexposure to the mock MRI. This paradoxical result isparticularly intriguing considering that it is difficultto achieve a measurable cortisol response to stressin neurotypical children. Several investigationshave shown that upon arrival for testing in thelaboratory, typically developing children often showsignificantly lower cortisol levels than obtained in
the home at the same time of day (e.g. Gunnar andDonzella, 2002; Gunnar and Vazquez, 2001). Sincethe typical children did not respond to the mock MRIit is difficult to evaluate our hypothesis regardingnegative feedback, but it should be noted thatenhanced negative feedback in neurotypical chil-dren compared to children with autism couldcontribute to the pattern of results we found.
In addition, there was no statistically significantrelationship between parent report measures andchange in cortisol following stress. Thus, it isunclear whether the lack of association betweenthese variables is the result of measuring differentconstructs or the discordance between behavioraland biological responsiveness.
As previously discussed in Section 1, an individ-ual’s response to stress can be measured through

Cortisol circadian rhythms and response to stress in children with autism 67
behavioral observation and report or by numerousbiological variables, such as cortisol. In the currentstudy, the majority of the children with autism didnot show observable behavioral agitation as wouldhave been expected despite the demonstratedcortisol response to the stressor. In fact, of thetwo children with autism that refused to enter allthe way into the scanner only one showedsignificant elevation in cortisol. Conversely, thetwo highest cortisol stress responders did not showany overt behavioral arousal or agitation to themock MRI procedures. Thus, cortisol measurementscan serve as an important biomarker of theresponsivity to a variety of different stimuli.
Within the confines of this study it is not possibleto identify any specific neural structure that isimplicated in the activation of the LHPA axis inchildren with autism. However, there is evidencethat in some limbic system structures (i.e. amyg-dala, hippocampus), there are abnormalities instructure and function (Abell et al., 1999; Baumanand Kemper, 1985; Bauman, 1991; Howard et al.,2000; Schumann et al., 2004; Sparks et al., 2002). Inorder to determine which limbic system structuresmay be implicated in the activation of the endocrinestress response it will be necessary to combineendocrine measures with fMRI studies.
It is unclear at this time and beyond the scope ofthis investigation as to whether the findings of thecurrent study have implications for neuroimagingresearch, in particular functional magneticresonance imaging (fMRI) in which the activation ofspecific brain regions is investigated in vivo. What isunknown is whether stress can influence baselinefMRI activity. However, our findings do suggest thatthe child with autism may be experiencing morestress in response to the mock MRI.
There are several limitations to our study, whichinclude the fact that we report a relatively smallsample of children. We also did not use aconsistent standardized measure of intelligence(IQ) across subjects to precisely measure currentlevel of cognitive functioning. Nevertheless, theestimated level of intelligence suggested thatthe majority of the children with autism had IQsin the average to borderline range based onprevious and current report. Thus, it does notappear that our findings concerning circadianrhythms were the result of the children beinglower functioning as reported in previous investi-gations (Jensen et al., 1985). However, there islittle known about the relationship betweenresponse to stress and IQ in children. In thisstudy we did not include a more precisebehavioral measure taken during the actual stresstest. Such a measure may have shed some light on
the apparent discordance between gross obser-vation and cortisol levels. It should be noted thatthe mock MRI is a complex stimulus that containsseveral different elements (i.e. noise, mild physi-cal restraint, novelty). It would be of interest todetermine which of these specific elements isprimarily responsible for the activation of theLHPA axis.
In conclusion, the present study found that mostchildren with autism show more variation in day-time patterns of cortisol and significant elevationsin salivary cortisol in response to a non-socialstressor. We have not found that children withautism constitute a homogeneous group in theirresponse to stress. Cortisol levels of four of thechildren in the Autism group actually went downafter stress similar to that observed in most of thecomparison group. Our current diagnostic tools donot allow us to discriminate the subtleties in autism(i.e. stress responsiveness), but perhaps the furtherstudy of the HPA axis in children with autism maylead to more careful consideration of phenotypicand idiosyncratic responsiveness in these children.There may be alterations in the limbic system inautism, which results in the children processing andresponding idiosyncratically to novel and threaten-ing events resulting in an exaggerated LHPAresponse (e.g. social vs. nonsocial, systemic vs.processive). By examining the endocrine responseto a variety of environmental challenges we canmore closely assess the manner in which childrenwith autism process information concerning theworld about them. Consequently, this finding maylead to the refinement of more discriminatingdiagnostic procedures.
Acknowledgements
The authors wish to thank the children and familieswho participated in this study and assisted in thecollection of home samples. We also express ourgratitude to Christine Brennan who ran the cortisolassays for the project.
References
Abell, F., Krams, M., Ashburner, J., Passingham, R., Friston, K.,Frackowiak, R., Happe, F., Frith, C., Frith, U., 1999. Theneuroanatomy of autism: a voxel-based whole brain analysisof structural scans. Neuroreport 10 (8), 1647–1651.
Aihara, R., Hashimoto, T., 1989. Neuroendocrinologic studies onautism. No To Hattatsu 21 (2), 154–162.
APA, 1994. Diagnostic and Statistical Manual of Mental Disorders,fourth ed. American Psychiatric Association, Washington, DC.

B.A. Corbett et al.68
Bauman, M.L., 1991. Microscopic neuroanatomic abnormalitiesin autism. Pediatrics 87 (5 Pt 2), 791–796.
Bauman, M., Kemper, T.L., 1985. Histoanatomic observations ofthe brain in early infantile autism. Neurology 35 (6), 866–874.
Curin, J.M., Terzic, J., Petkovic, Z.B., Zekan, L., Terzic, I.M.,Susnjara, I.M., 2003. Lower cortisol and higher ACTH levels inindividuals with autism. J. Autism Dev. Disord. 33 (4), 443–448.
Dunn, W., 1999. Short Sensory Profile. The PsychologicalCorporation, USA.
Efron, B., Tibshirani, R.J., 1993. An Introduction to theBootstrap. Chapman and Hall, San Francisco.
Groden, J., Diller, A., Bausman, M., Velicer, W., Norman, G.,Cautela, J., 2001. The development of a stress surveyschedule for persons with autism and other developmentaldisabilities. J. Autism Dev. Disord. 31, 207–217.
Gunnar, M.R., Donzella, B., 2002. Social regulation of thecortisol levels in early human development. Psychoneuroen-docrinology 27 (1–2), 199–220.
Gunnar, M.R., Vazquez, D.M., 2001. Low cortisol and a flatteningof expected daytime rhythm: potential indices of risk inhuman development. Dev. Psychopathol. 13 (3), 515–538.
Hennessey, J.W., Levine, S., 1979. Stress, arousal, and thepituitary-adrenal system: a psychoendocrine hypothesis. In:Sprague, J.M., Epstein, S. (Eds.), Progress in Psychobiologyand Physiological Psychology, vol. 8, pp. 133–178.
Herman, J.P., Cullinan, W.E., 1997. Neurocircuitry of stress:central control of the hypothalamo-pituitary-adrenocorticalaxis. Trends Neurosci. 20 (2), 78–84.
Hessl, D., Glaser, B., Dyer-Friedman, J., Blasey, C., Hastie, T.,Gunnar, M., Reiss, A.L., 2002. Cortisol and behavior in fragileX syndrome. Psychoneuroendocrinology 27 (7), 855–872.
Hill, S.D., Wagner, E.A., Shedlarski Jr., J.G., Sears, S.P., 1977.Diurnal cortisol and temperature variation of normal andautistic children. Dev. Psychobiol. 10 (6), 579–583.
Hoshino, Y., Yokoyama, F., Watanabe, M., Murata, S., Kaneko, M.,Kumashiro, H., 1987. The diurnal variation and response todexamethasone suppression test of saliva cortisol level inautistic children. Jpn. J. Psychiatry Neurol. 41 (2), 227–235.
Howard, M.A., Cowell, P.E., Boucher, J., Broks, P., Mayes, A.,Farrant, A., Roberts, N., 2000. Convergent neuroanatomicaland behavioural evidence of an amygdala hypothesis ofautism. Neuroreport 11 (13), 2931–2935.
Jansen, L.M., Gispen-de Wied, C.C., Van der Gaag, R.J., tenHove, F., Willemsen-Swinkels, S.W., Harteveld, E., VanEngeland, H., 2000. Unresponsiveness to psychosocial stressin a subgroup of autistic-like children, multiple complexdevelopmental disorder. Psychoneuroendocrinology 25 (8),753–764.
Jansen, L.M., Gispen-de Wied, C.C., van der Gaag, R.J., vanEngeland, H., 2003. Differentiation between autism andmultiple complex developmental disorder in response topsychosocial stress.Neuropsychopharmacology28(3), 582–590.
Jensen, J.B., Realmuto, G.M., Garfinkel, B.D., 1985. Thedexamethasone suppression test in infantile autism. J. Am.Acad. Child Psychiatry 24 (3), 263–265.
Kanner, L., 1943. Autistic disturbances of affective contact.Nervous Child 2, 217–250.
Kirschbaum, C., Hellhammer, D.H., 1994. Salivary cortisolin psychoneuroendocrine research: recent developmentsand applications. Psychoneuroendocrinology 19 (4), 313–333.
Laird, N.M., Ware, J.H., 1982. Random-effects models forlongitudinal data. Biometrics 38 (4), 963–974.
Lord, C., Rutter, M., DiLavore, P., Risi, S., 1999. AutismDiagnostic Observation Schedule-WPS. Western PsychologicalServices, Los Angeles, CA.
Maher, K.R., Harper, J.F., Macleay, A., King, M.G., 1975.Peculiarities in the endocrine response to insulin stress inearly infantile autism. J. Nerv. Ment. Dis. 161 (3), 180–184.
Mallinckrodt, C.H., Sanger, T.M., Dube, S., DeBrota, D.J.,Molenberghs, G., Carroll, R.J., Potter, W.Z.,Tollefson, G.D., 2003. Assessing and interpreting treatmenteffects in longitudinal clinical trials with missing data. Biol.Psychiatry 53 (8), 754–760.
Murphy, S.A., Van Der Vaart, A.W., 2000. On profile likelihood.Journal of the American Statistical Association 95, 449–465.
Nir, I., Meir, D., Zilber, N., Knobler, H., Hadjez, J., Lerner, Y.,1995. Brief report: circadian melatonin, thyroid-stimulatinghormone, prolactin, and cortisol levels in serum of youngadults with autism. J. Autism Dev. Disord. 25 (6), 641–654.
R Development Core Team, 2005. R: A Language and Environ-ment for Statistical Computing 2005. Vienna, Austria.
Richdale, A.L., Prior, M.R., 1992. Urinary cortisol circadianrhythm in a group of high-functioning children with autism.J. Autism Dev. Disord. 22 (3), 433–447.
Schumann,C.M.,Hamstra, J.,Goodlin-Jones,B.L.,Lotspeich,L.J.,Kwon, H., Buonocore, M.H., Lammers, C.R., Reiss, A.L.,Amaral, D.G., 2004. The amygdala is enlarged in children butnot adolescents with autism; the hippocampus is enlarged at allages. J. Neurosci. 24 (28), 6392–6401.
Schwartz, E.B., Granger, D.A., Susman, E.J., Gunnar, M.R.,Laird, B., 1998. Assessing salivary cortisol in studies of childdevelopment. Child Dev. 69 (6), 1503–1513.
Sparks, B.F., Friedman, S.D., Shaw, D.W., Aylward, E.H.,Echelard, D., Artru, A.A., Maravilla, K.R., Giedd, J.N.,Munson, J., Dawson, G., Dager, S.R., 2002. Brain structuralabnormalities in young children with autism spectrumdisorder. Neurology 59 (2), 184–192.
Tordjman, S., Anderson, G.M., McBride, P.A., Hertzig, M.E.,Snow, M.E., Hall, L.M., Thompson, S.M., Ferrari, P.,Cohen, D.J., 1997. Plasma beta-endorphin, adrenocortico-tropin hormone, and cortisol in autism. J. Child Psychol.Psychiatry 38 (6), 705–715.
Wiener, S.G., Coe, C.L., Levine, S., 1988. Endocrine andneurochemical sequelae of primate vocalizations. In:Newman, J. (Ed.),, Ed. The Physiological Control of MammalianVocalization, Vol.. Plenum Press, New York, pp. 367–394.
Yamazaki, K., Saito, Y., Okada, F., Fujieda, T., Yamashita, I.,1975. An application of neuroendocrinological studies inautistic children and Heller’s syndrome. J. Autism Child.Schizophr. 5 (4), 323–332.
Copyright © 2022 FDOKUMEN




















