
A male-specific effect of dominant-negative Fos
12
PATTERNS & PHENOTYPES A Male-Specific Effect of Dominant-Negative Fos Wooly Pierre, Rosa Morra, John Lucchesi, and Barry Yedvobnick * The transcription factor Fos contains a basic DNA binding domain combined with a leucine zipper (bZip). Expression of a truncated form of Fos in Drosophila that contains only the bZip region (Fos bZip) elicits phenotypes resembling fos mutations. These effects presumably derive from competition between wild-type and truncated forms for dimerization partners, with the truncation acting in a dominant-negative manner. We found that expression of Fos bZip elicits male-specific phenotypes. Moreover, genetic interactions occur between Fos bZip and mutations in loci encoding the X chromosome dosage compensation complex. Fos bZip effects are correlated with aberrant male X chromosome structure and depressed signaling through the X-linked Notch locus. Unexpectedly, the male-specific effects are not reproduced with Fos RNAi, suggesting that Fos bZip can be neomorphic in nature. These results provide insight into how mutations in bZip proteins can exhibit gain of function activity. Developmental Dynamics 237:3361–3372, 2008. © 2008 Wiley-Liss, Inc. Key words: Drosophila; dominant-negative; Fos; Notch; dosage compensation Accepted 20 August 2008 INTRODUCTION The Fos and Jun proteins dimerize and form the AP-1 transcription fac- tor, which mediates gene regulatory responses to a wide array of signals, including those affecting growth, pat- terning and apoptosis in multicellular organisms (Hess et al., 2004). In Dro- sophila, these proteins have key roles in several well described embryonic and postembryonic processes, such as tissue closure and patterning in the eye and wing (Ciapponi et al., 2001; Kockel et al., 2001). Consistent with these observations, Fos and Jun func- tion has been associated with several signal transduction pathways, includ- ing MAP kinase, TGF, and Notch, among others (Kockel et al., 2001). In a prior genetic screen for modifiers of Notch function in the wing margin, we identified a transposon insert up- stream of the Fos-encoding locus kayak that suppresses a Notch path- way loss of function phenotype (Hall et al., 2004). To investigate the Fos- Notch relationship, we performed ad- ditional genetic analyses using a transgene encoding a truncated, dom- inant-negative form of Fos (Eresh et al., 1997). During these studies, we observed a sex-differential effect of the transgene, with males being more sensitive to the effects of dominant- negative Fos than females in a variety of tissues. The phenotypes produced by the transgene in males synergize strongly with Notch pathway muta- tions. These data suggest that an ab- errant form of Fos protein is capable of eliciting both loss of function and gain of function (neomorphic) phenotypes, the latter interfering with the mecha- nism of dosage compensation (Luc- chesi et al., 2005). RESULTS AND DISCUSSION A Dominant-Negative Form of Fos Preferentially Elicits Male Phenotypes During an analysis of candidates for Notch pathway modifiers, we used a dominant-negative form of Fos named Fos bZip (UAS-Fbz, Eresh et al., 1997) for genetic interaction studies. This Gal4-UAS-dependent transgenic con- struct expresses the bZip region of Fos, involved in DNA binding and pro- tein dimerization, and it has been Department of Biology, Emory University, Atlanta, Georgia Grant sponsor: NSF; Grant number: IBN-0415453. *Correspondence to: Barry Yedvobnick, Department of Biology, Emory University, OW Rollins Research Building, 1510 Clifton Road, Atlanta, GA. 30322. E-mail: [email protected] DOI 10.1002/dvdy.21751 Published online 10 October 2008 in Wiley InterScience (www.interscience.wiley.com). DEVELOPMENTAL DYNAMICS 237:3361–3372, 2008 © 2008 Wiley-Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of A male-specific effect of dominant-negative Fos

PATTERNS & PHENOTYPES
A Male-Specific Effect of Dominant-NegativeFosWooly Pierre, Rosa Morra, John Lucchesi, and Barry Yedvobnick*
The transcription factor Fos contains a basic DNA binding domain combined with a leucine zipper (bZip).Expression of a truncated form of Fos in Drosophila that contains only the bZip region (Fos bZip) elicitsphenotypes resembling fos mutations. These effects presumably derive from competition between wild-typeand truncated forms for dimerization partners, with the truncation acting in a dominant-negative manner.We found that expression of Fos bZip elicits male-specific phenotypes. Moreover, genetic interactions occurbetween Fos bZip and mutations in loci encoding the X chromosome dosage compensation complex. FosbZip effects are correlated with aberrant male X chromosome structure and depressed signaling throughthe X-linked Notch locus. Unexpectedly, the male-specific effects are not reproduced with Fos RNAi,suggesting that Fos bZip can be neomorphic in nature. These results provide insight into how mutations inbZip proteins can exhibit gain of function activity. Developmental Dynamics 237:3361–3372, 2008.© 2008 Wiley-Liss, Inc.
Key words: Drosophila; dominant-negative; Fos; Notch; dosage compensation
Accepted 20 August 2008
INTRODUCTION
The Fos and Jun proteins dimerizeand form the AP-1 transcription fac-tor, which mediates gene regulatoryresponses to a wide array of signals,including those affecting growth, pat-terning and apoptosis in multicellularorganisms (Hess et al., 2004). In Dro-sophila, these proteins have key rolesin several well described embryonicand postembryonic processes, such astissue closure and patterning in theeye and wing (Ciapponi et al., 2001;Kockel et al., 2001). Consistent withthese observations, Fos and Jun func-tion has been associated with severalsignal transduction pathways, includ-ing MAP kinase, TGF�, and Notch,among others (Kockel et al., 2001). Ina prior genetic screen for modifiers of
Notch function in the wing margin, weidentified a transposon insert up-stream of the Fos-encoding locuskayak that suppresses a Notch path-way loss of function phenotype (Hallet al., 2004). To investigate the Fos-Notch relationship, we performed ad-ditional genetic analyses using atransgene encoding a truncated, dom-inant-negative form of Fos (Eresh etal., 1997). During these studies, weobserved a sex-differential effect ofthe transgene, with males being moresensitive to the effects of dominant-negative Fos than females in a varietyof tissues. The phenotypes producedby the transgene in males synergizestrongly with Notch pathway muta-tions. These data suggest that an ab-errant form of Fos protein is capable of
eliciting both loss of function and gainof function (neomorphic) phenotypes,the latter interfering with the mecha-nism of dosage compensation (Luc-chesi et al., 2005).
RESULTS AND DISCUSSION
A Dominant-Negative Formof Fos Preferentially ElicitsMale Phenotypes
During an analysis of candidates forNotch pathway modifiers, we used adominant-negative form of Fos namedFos bZip (UAS-Fbz, Eresh et al., 1997)for genetic interaction studies. ThisGal4-UAS-dependent transgenic con-struct expresses the bZip region ofFos, involved in DNA binding and pro-tein dimerization, and it has been
Department of Biology, Emory University, Atlanta, GeorgiaGrant sponsor: NSF; Grant number: IBN-0415453.*Correspondence to: Barry Yedvobnick, Department of Biology, Emory University, OW Rollins Research Building, 1510Clifton Road, Atlanta, GA. 30322. E-mail: [email protected]
DOI 10.1002/dvdy.21751Published online 10 October 2008 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 237:3361–3372, 2008
© 2008 Wiley-Liss, Inc.
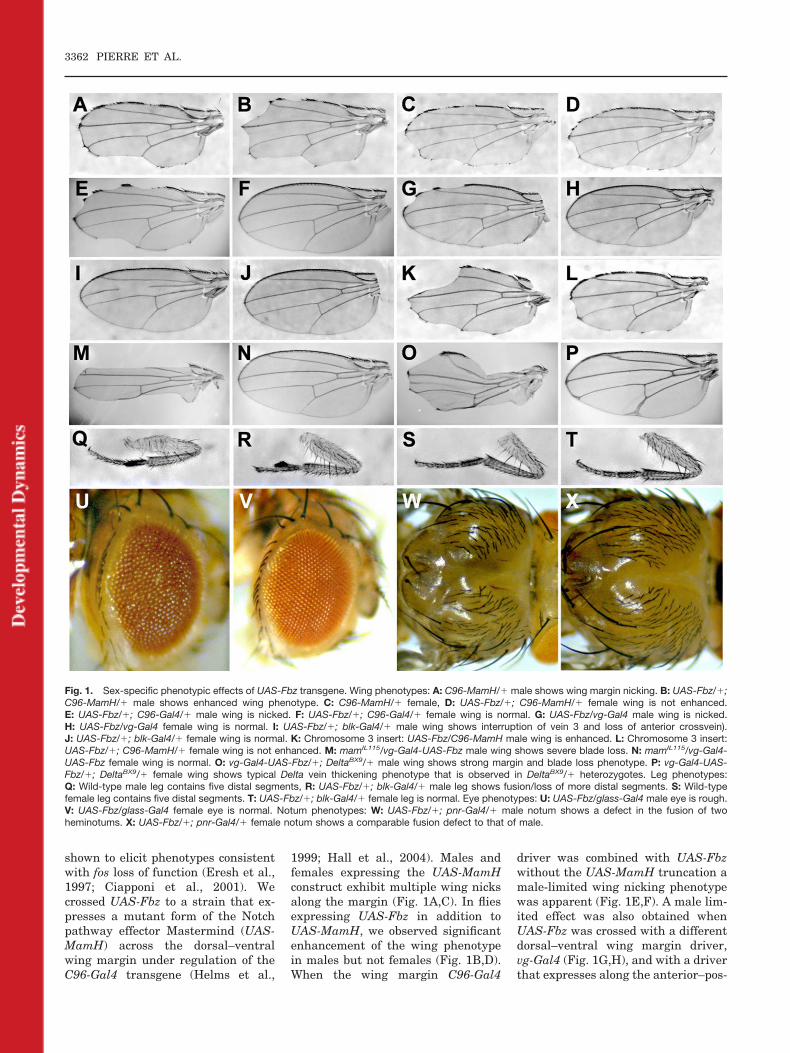
shown to elicit phenotypes consistentwith fos loss of function (Eresh et al.,1997; Ciapponi et al., 2001). Wecrossed UAS-Fbz to a strain that ex-presses a mutant form of the Notchpathway effector Mastermind (UAS-MamH) across the dorsal–ventralwing margin under regulation of theC96-Gal4 transgene (Helms et al.,
1999; Hall et al., 2004). Males andfemales expressing the UAS-MamHconstruct exhibit multiple wing nicksalong the margin (Fig. 1A,C). In fliesexpressing UAS-Fbz in addition toUAS-MamH, we observed significantenhancement of the wing phenotypein males but not females (Fig. 1B,D).When the wing margin C96-Gal4
driver was combined with UAS-Fbzwithout the UAS-MamH truncation amale-limited wing nicking phenotypewas apparent (Fig. 1E,F). A male lim-ited effect was also obtained whenUAS-Fbz was crossed with a differentdorsal–ventral wing margin driver,vg-Gal4 (Fig. 1G,H), and with a driverthat expresses along the anterior–pos-
Fig. 1. Sex-specific phenotypic effects of UAS-Fbz transgene. Wing phenotypes: A: C96-MamH/� male shows wing margin nicking. B: UAS-Fbz/�;C96-MamH/� male shows enhanced wing phenotype. C: C96-MamH/� female, D: UAS-Fbz/�; C96-MamH/� female wing is not enhanced.E: UAS-Fbz/�; C96-Gal4/� male wing is nicked. F: UAS-Fbz/�; C96-Gal4/� female wing is normal. G: UAS-Fbz/vg-Gal4 male wing is nicked.H: UAS-Fbz/vg-Gal4 female wing is normal. I: UAS-Fbz/�; blk-Gal4/� male wing shows interruption of vein 3 and loss of anterior crossvein).J: UAS-Fbz/�; blk-Gal4/� female wing is normal. K: Chromosome 3 insert: UAS-Fbz/C96-MamH male wing is enhanced. L: Chromosome 3 insert:UAS-Fbz/�; C96-MamH/� female wing is not enhanced. M: mamIL115/vg-Gal4-UAS-Fbz male wing shows severe blade loss. N: mamIL115/vg-Gal4-UAS-Fbz female wing is normal. O: vg-Gal4-UAS-Fbz/�; DeltaBX9/� male wing shows strong margin and blade loss phenotype. P: vg-Gal4-UAS-Fbz/�; DeltaBX9/� female wing shows typical Delta vein thickening phenotype that is observed in DeltaBX9/� heterozygotes. Leg phenotypes:Q: Wild-type male leg contains five distal segments, R: UAS-Fbz/�; blk-Gal4/� male leg shows fusion/loss of more distal segments. S: Wild-typefemale leg contains five distal segments. T: UAS-Fbz/�; blk-Gal4/� female leg is normal. Eye phenotypes: U: UAS-Fbz/glass-Gal4 male eye is rough.V: UAS-Fbz/glass-Gal4 female eye is normal. Notum phenotypes: W: UAS-Fbz/�; pnr-Gal4/� male notum shows a defect in the fusion of twoheminotums. X: UAS-Fbz/�; pnr-Gal4/� female notum shows a comparable fusion defect to that of male.
3362 PIERRE ET AL.

terior boundary (vein 3 and anteriorcrossvein) of the wing, blk-Gal4 (Fig.1I,J). The phenotypic effect of thetransgene mapped to chromosome 2,and was produced from either male orfemale parents heterozygous for thetransgene (not shown). When UAS-Fbz was mobilized from its site of res-idence on chromosome 2 to chromo-some 3, the male-specific effectpersisted (Fig. 1K,L), suggesting thata sex-specific chromosomal position-effect on transgene expression did notunderlie the phenotypes. This wascorroborated by expression studies ofUAS-Fbz (described below).
Based on the interactions of UAS-Fbzwith a mutant form of Mam (Fig. 1A–D), we tested an EMS-induced allele ofthe locus, mamIL115, a severe mutation(Schmid et al., 1996). When vg-Gal4driving UAS-Fbz (vg-Fbz) is combinedgenotypically with mamIL115 a signifi-cant loss of wing material is observed inmales, but not females (Fig. 1M,N). Asimilar result was observed when vg-Fbz was combined with another Notchpathway mutation, DeltaBX9 (Fig.1O,P). Thus, the sex-specific effects ofUAS-Fbz expression in the wing are en-hanced when Notch pathway activity isdepressed.
The male-specific effects are notlimited to wing tissue. Expression ofUAS-Fbz under blk-Gal4 regulationperturbed leg segment formation inmales (Fig. 1Q,R), but not females(Fig. 1S,T), and its expression underglass-Gal4 regulation led to rougheyes in males but not females (Fig.1U,V). Additionally, adult offspringfrom matings to ptc-Gal4 were limitedto females (data not shown). In con-trast, we observed two cases wheremale and female effects were indistin-guishable. When UAS-Fbz is driven inthe notum under control of pnr-Gal4there is a thorax fusion defect evidentin both sexes (Fig. 1W,X), and whenthe transgene is driven by means of da-Gal4 both males and females are lethal.As reported previously (Zeitlinger andBohmann, 1999), the thorax fusion de-fect of UAS-Fbz driven by pnr-Gal4 isconsistent with the normal role of Fos inthorax closure. Therefore, the UAS-Fbztransgene can function in females, andit appears to be an effective inhibitor ofprocesses regulated by the wild-typeFos protein. However, the male-specific
phenotypes indicate an effect on otherprocesses as well.
UAS-Fbz ExpressionPreferentially Affects NotchTarget Expression in MaleWing Discs
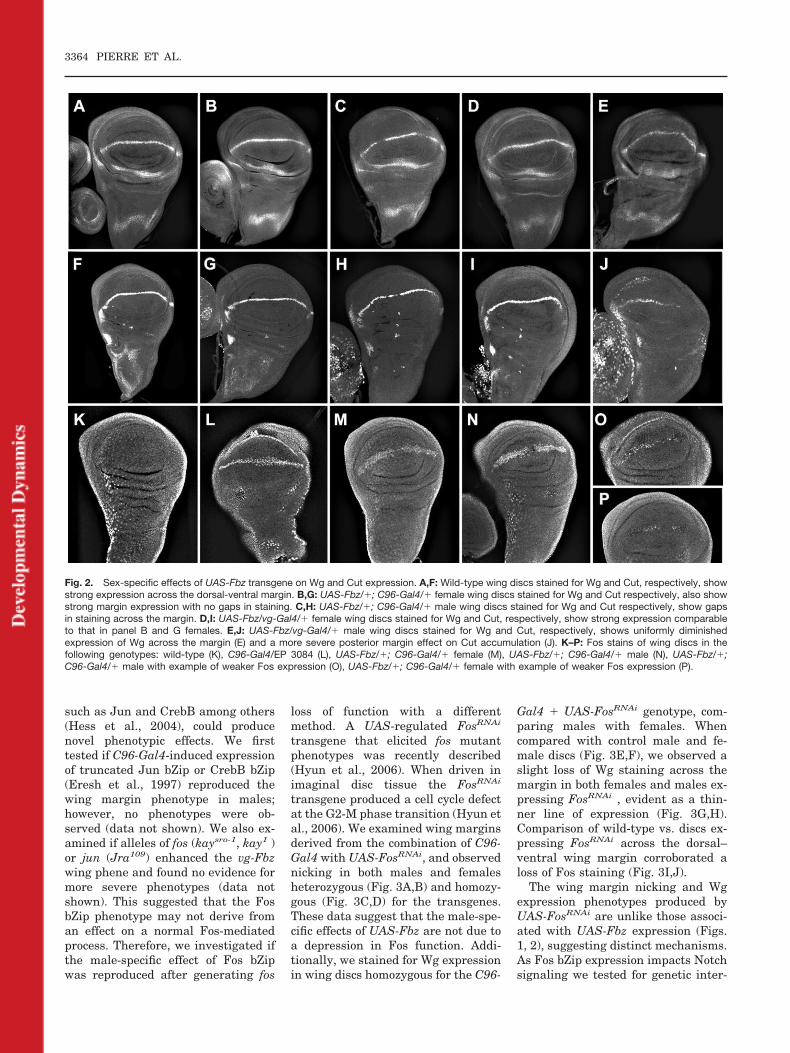
The observation that UAS-Fbz can en-hance a Notch pathway wing marginphenotype only in males or produce awing margin nicking phenotype onlyin males, suggested that expression ofFos bZip may impact Notch targetgene expression in a sex-specific man-ner. We examined this by assaying ex-pression of Wingless (Wg) and Cut ex-pression across the dorsal–ventralmargin in wing discs expressing UAS-Fbz under C96-Gal4 or vg-Gal4 con-trol. Both Wg and Cut are well-de-scribed targets of the Notch pathway(Neumann and Cohen, 1996; Helms etal., 1999). In wild-type wing discs bothWg and Cut are evident at high levelsat the margin (Fig. 2A,F, respec-tively). In female wing discs express-ing C96-Gal4 � UAS-Fbz (Fig. 2B) orvg-Gal4 � UAS-Fbz (Fig. 2D), Wg per-sists at high levels with no gaps inexpression. In contrast, male wingdiscs exhibit diminished levels of Wgwith discrete gaps evident in thestaining (Fig. 2C,E). Analysis of Cutexpression in these genotypes wasconsistent with the Wg analysis. Fe-male discs expressing UAS-Fbz acrossthe wing margin exhibited uniformlyhigh levels of Cut (Fig. 2G,I), whereasthose derived from males showed sig-nificantly lower levels of Cut expres-sion (Fig. 2H,J). Thus, the differentialphenotypic effects of UAS-Fbz expres-sion on wing structure in males re-flects the relative levels of accumula-tion of two proteins required forproper wing margin formation.
One explanation for the male-spe-cific effects could derive from differen-tial expression of the UAS-Fbz trans-gene in males vs. females. We initiallyaddressed this by transposing thetransgene to an alternative site (Fig.1K,L), where we observed a compara-ble phenotypic effect limited to males.However, this experiment did not ruleout a sex-specific expression effect in-herent to the transgene itself. Conse-quently, we assayed expression of theUAS-Fbz transgene by staining wingdiscs for Fos protein. First, we tested
the Fos antisera on control discs toexamine the endogenous Fos protein’sexpression pattern. We observed thatthe expression of Fos in wild-typediscs is widespread, with higher levelsevident in the peripodial membrane(Fig. 2K), as reported previously(Zeitlinger and Bohmann, 1999). As apositive control to assay induced lev-els of wing margin Fos expression, weused the EP 3084 transgene, which isa UAS-controlled P element situatedupstream of the kayak (D Fos) locus(see the Experimental Procedures sec-tion). The EP 3084 strain was crossedwith C96-Gal4 and transheterozygouswing discs were stained for Fos ex-pression. In such discs, a strong pat-tern of Fos accumulation across thedorsal–ventral margin is evident thatis not observed in wild-type discs (Fig.2L). When C96-Gal4 is used to drivethe shortened Fos protein encoded byUAS-Fbz, we could detect Fos stainingacross the wing margin in both female(Fig. 2M) and male (Fig. 2N) discs.When compared with C96-Gal4-EP3084 discs, the staining for Fos wasweaker and much more variable inlevel in both females and males ex-pressing the shorter protein encodedby UAS-Fbz. For example, weak stain-ing in a male and female disc is shownin Figures 2O and P, respectively. Theweaker staining is expected becausethe UAS-Fbz product contains feweramino acid residues recognizable bythe Fos antisera. As we observe com-parable variability in Fos staininglevels of female vs. male, C96-Gal4�UAS-Fbz discs, but 100% pene-trant phenotypic effects limited tomales, we conclude that the sex-spe-cific effects of UAS-Fbz are not due todifferential expression.
The Male-Specific Effect ofDominant-Negative Fos IsNot Recapitulated by FosLoss of Function
The Fos bZip transgene is an effectiveinhibitor of Fos function and its ex-pression perturbs a range of tissues(Eresh et al., 1997; Zeitlinger and Bo-hmann, 1999; Ciapponi et al., 2001).However, the effects of driving thetruncated form of Fos may not be lim-ited to loss of Fos function. For exam-ple, the potential for heterodimeriza-tion with other bZip binding partners,
MALE SPECIFIC PHENOTYPES 3363
such as Jun and CrebB among others(Hess et al., 2004), could producenovel phenotypic effects. We firsttested if C96-Gal4-induced expressionof truncated Jun bZip or CrebB bZip(Eresh et al., 1997) reproduced thewing margin phenotype in males;however, no phenotypes were ob-served (data not shown). We also ex-amined if alleles of fos (kaysro-1, kay1 )or jun (Jra109) enhanced the vg-Fbzwing phene and found no evidence formore severe phenotypes (data notshown). This suggested that the FosbZip phenotype may not derive froman effect on a normal Fos-mediatedprocess. Therefore, we investigated ifthe male-specific effect of Fos bZipwas reproduced after generating fos
loss of function with a differentmethod. A UAS-regulated FosRNAi
transgene that elicited fos mutantphenotypes was recently described(Hyun et al., 2006). When driven inimaginal disc tissue the FosRNAi
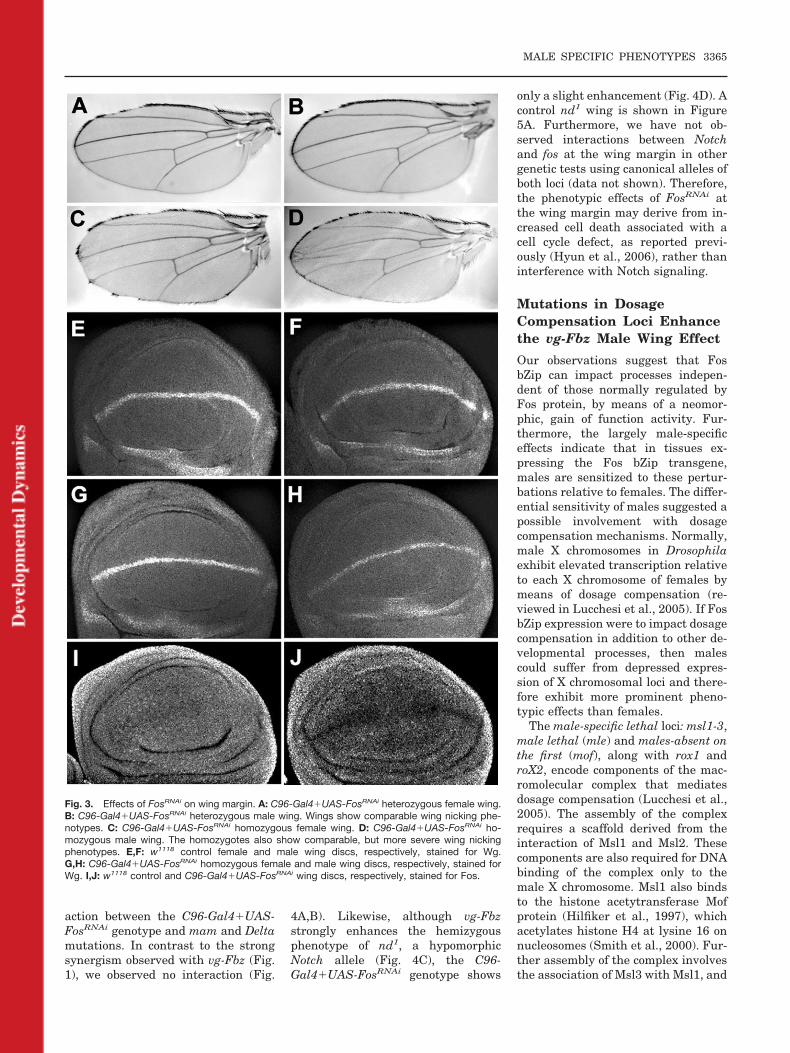
transgene produced a cell cycle defectat the G2-M phase transition (Hyun etal., 2006). We examined wing marginsderived from the combination of C96-Gal4 with UAS-FosRNAi, and observednicking in both males and femalesheterozygous (Fig. 3A,B) and homozy-gous (Fig. 3C,D) for the transgenes.These data suggest that the male-spe-cific effects of UAS-Fbz are not due toa depression in Fos function. Addi-tionally, we stained for Wg expressionin wing discs homozygous for the C96-
Gal4 � UAS-FosRNAi genotype, com-paring males with females. Whencompared with control male and fe-male discs (Fig. 3E,F), we observed aslight loss of Wg staining across themargin in both females and males ex-pressing FosRNAi , evident as a thin-ner line of expression (Fig. 3G,H).Comparison of wild-type vs. discs ex-pressing FosRNAi across the dorsal–ventral wing margin corroborated aloss of Fos staining (Fig. 3I,J).
The wing margin nicking and Wgexpression phenotypes produced byUAS-FosRNAi are unlike those associ-ated with UAS-Fbz expression (Figs.1, 2), suggesting distinct mechanisms.As Fos bZip expression impacts Notchsignaling we tested for genetic inter-
Fig. 2. Sex-specific effects of UAS-Fbz transgene on Wg and Cut expression. A,F: Wild-type wing discs stained for Wg and Cut, respectively, showstrong expression across the dorsal-ventral margin. B,G: UAS-Fbz/�; C96-Gal4/� female wing discs stained for Wg and Cut respectively, also showstrong margin expression with no gaps in staining. C,H: UAS-Fbz/�; C96-Gal4/� male wing discs stained for Wg and Cut respectively, show gapsin staining across the margin. D,I: UAS-Fbz/vg-Gal4/� female wing discs stained for Wg and Cut, respectively, show strong expression comparableto that in panel B and G females. E,J: UAS-Fbz/vg-Gal4/� male wing discs stained for Wg and Cut, respectively, shows uniformly diminishedexpression of Wg across the margin (E) and a more severe posterior margin effect on Cut accumulation (J). K–P: Fos stains of wing discs in thefollowing genotypes: wild-type (K), C96-Gal4/EP 3084 (L), UAS-Fbz/�; C96-Gal4/� female (M), UAS-Fbz/�; C96-Gal4/� male (N), UAS-Fbz/�;C96-Gal4/� male with example of weaker Fos expression (O), UAS-Fbz/�; C96-Gal4/� female with example of weaker Fos expression (P).
3364 PIERRE ET AL.
action between the C96-Gal4�UAS-FosRNAi genotype and mam and Deltamutations. In contrast to the strongsynergism observed with vg-Fbz (Fig.1), we observed no interaction (Fig.
4A,B). Likewise, although vg-Fbzstrongly enhances the hemizygousphenotype of nd1, a hypomorphicNotch allele (Fig. 4C), the C96-Gal4�UAS-FosRNAi genotype shows
only a slight enhancement (Fig. 4D). Acontrol nd1 wing is shown in Figure5A. Furthermore, we have not ob-served interactions between Notchand fos at the wing margin in othergenetic tests using canonical alleles ofboth loci (data not shown). Therefore,the phenotypic effects of FosRNAi atthe wing margin may derive from in-creased cell death associated with acell cycle defect, as reported previ-ously (Hyun et al., 2006), rather thaninterference with Notch signaling.
Mutations in DosageCompensation Loci Enhancethe vg-Fbz Male Wing Effect
Our observations suggest that FosbZip can impact processes indepen-dent of those normally regulated byFos protein, by means of a neomor-phic, gain of function activity. Fur-thermore, the largely male-specificeffects indicate that in tissues ex-pressing the Fos bZip transgene,males are sensitized to these pertur-bations relative to females. The differ-ential sensitivity of males suggested apossible involvement with dosagecompensation mechanisms. Normally,male X chromosomes in Drosophilaexhibit elevated transcription relativeto each X chromosome of females bymeans of dosage compensation (re-viewed in Lucchesi et al., 2005). If FosbZip expression were to impact dosagecompensation in addition to other de-velopmental processes, then malescould suffer from depressed expres-sion of X chromosomal loci and there-fore exhibit more prominent pheno-typic effects than females.
The male-specific lethal loci: msl1-3,male lethal (mle) and males-absent onthe first (mof), along with rox1 androX2, encode components of the mac-romolecular complex that mediatesdosage compensation (Lucchesi et al.,2005). The assembly of the complexrequires a scaffold derived from theinteraction of Msl1 and Msl2. Thesecomponents are also required for DNAbinding of the complex only to themale X chromosome. Msl1 also bindsto the histone acetytransferase Mofprotein (Hilfiker et al., 1997), whichacetylates histone H4 at lysine 16 onnucleosomes (Smith et al., 2000). Fur-ther assembly of the complex involvesthe association of Msl3 with Msl1, and
Fig. 3. Effects of FosRNAi on wing margin. A: C96-Gal4�UAS-FosRNAi heterozygous female wing.B: C96-Gal4�UAS-FosRNAi heterozygous male wing. Wings show comparable wing nicking phe-notypes. C: C96-Gal4�UAS-FosRNAi homozygous female wing. D: C96-Gal4�UAS-FosRNAi ho-mozygous male wing. The homozygotes also show comparable, but more severe wing nickingphenotypes. E,F: w1118 control female and male wing discs, respectively, stained for Wg.G,H: C96-Gal4�UAS-FosRNAi homozygous female and male wing discs, respectively, stained forWg. I,J: w1118 control and C96-Gal4�UAS-FosRNAi wing discs, respectively, stained for Fos.
MALE SPECIFIC PHENOTYPES 3365

the association of Mle, an ATP-depen-dent helicase, with both Msl2 and roXRNAs (Lucchesi et al., 2005; Li et al.,2005, 2008). This complex assemblesonly in males because msl2 RNA is nottranslated in females (Kelley et al.,1997). Acetylation of nucleosomal his-tones on the male X chromosome isthought to facilitate transcriptionalelongation approximately twofold
(Smith et al., 2001). We investigatedgenetic interactions between the msland mle mutations, and vg-Fbz, whichshows a male-limited wing phenotype.We also tested interactions with an Xchromosome containing loss of func-tion for both the rox1 and roX2 redun-dant loci (Meller and Rattner, 2002;Deng et al., 2005).
We first tested heterozygosity of msl
and mle mutations for an effect onwing margin formation independentof dominant-negative Fos. Given thebroad role of these loci in expressionlevel of X-linked genes in males, it waspossible that decreasing their activitycould elicit general phenotypic en-hancement effects. This could be par-ticularly significant for this study, be-cause several Notch pathway loci,
Fig. 4. Notch pathway mutations do not interact with FosRNAi A: mamIL115/�; C96-Gal4�UAS-FosRNAi/� male wing does not show enhancedphenotype. B: DeltaBX9/C96-Gal4�UAS- FosRNAi/� male wing similarly is not enhanced. Compare to strong enhancement by means of these Notchpathway alleles with vg-Gal4-UAS-Fbz (Fig. 1M, O). C: nd1/Y; vg-Gal4-UAS-Fbz /� male wing shows strong enhancement. D: nd1/Y; C96-Gal4�UAS-FosRNAi/� male wing does not show enhanced phenotype. The control nd1/Y wing phenotype is shown in panel A of Figure 5.
Fig. 5. Mutations in msl and mle loci do not consistently affect the X-linked nd1 wing phenotype. A: nd1/Y control wing shows wing margin nicks andslight vein thickening. B: nd1/Y; mle/� wing shows comparable phenotype to controls. C: nd1/Y; msl-3/� wing shows less severe phenotype comparedwith controls. D: nd1/Y; msl-2 msl-1 mle/� wing shows more severe phenotype compared with controls.
3366 PIERRE ET AL.

including Notch itself, are X linked.Therefore, we investigated if muta-tions in msl and mle loci modify thephenotype of a hemizygous viable al-lele of the X chromosome Notch locus,nd1. The mle, msl-3, and msl-2 msl-1mle triple recombinant strains weremated to nd1 females. Hemizygousnd1 males heterozygous for these mu-tant chromosomes were analyzed formodification of the control nd1 wingphenotype. The control nd1 males de-rived from crosses to the w1118 strain.These males exhibit minor wing nick-ing and vein thickening (Fig. 5A) Themle, msl-3, and msl-2 msl-1 mle chro-mosomes, when combined with nd1,differed in the extent of wing nicking;however, in only one case did we ob-serve an increase in severity (Fig. 5B–D). Therefore, heterozygosity for mu-tations in msl and mle loci appears notto act as a universal enhancer of X-linked mutations.
When vg-Fbz was crossed to the con-trol w1118 strain, we observed that57% of the male vg-Fbz/ w1118 wingsshowed defects, whereas 0.2% of fe-male wings exhibited defects (Table1). When vg-Fbz was crossed to muta-tions in mle, msl-3, and the triple re-combinant chromosome containing al-leles of msl-2, msl-1, and mle weobserved a significant increase in thepercentage of defective male wings(Table 1): mle (71%), msl-3 (71.5%),and msl-2 msl-1 mle (73%). We also
tested a UAS-regulated RNAi trans-gene directed against msl-3 RNA (Di-etzl et al., 2007). When this constructwas driven by means of Vg-Gal4, a lowlevel (4.5%) of wing defects was evi-dent, limited to males. However, sig-nificant enhancement of the vg-Fbzwing phenotype (77.4%) was apparentafter coexpression of msl-3 RNAi, cor-roborating the msl allele results (Ta-ble 1). Finally, we examined if vg-Fbzinteracted with an X chromosome de-ficient for roX1 and roX2 functions:roX1ex21A Df(1)52 (Deng et al., 2005).In this experiment, the lethality asso-ciated with roX2 deficiency (Df(1)52)was covered with a rescuing trans-gene (w�4�4.3) that does not containroX2 (Meller and Rattner, 2002). De-spite a loss of function for both roXloci, male escapers are observed inthis genotype, because the roX1ex21A
allele is not null (V. Meller, personalcommunication). Males carrying theroX-deficient X chromosome, w�4�4.3,and vg-Fbz were compared with siblingmales carrying the Binscy balancer,w�4�4.3, and vg-Fbz. Sufficient num-bers of roX-deficient viable males wereobtained to allow us to score an en-hanced and highly penetrant wing phe-notype in this class (Fig. 6). Therefore,reductions in the activity of loci thatcontribute both RNA and protein to thedosage compensation complex enhancethe male-specific effect of Fos bZip inthe wing.
Overexpression of Wild-Typeand Mutant Forms of MslProteins Modify the vg-FbzMale Wing Effect
The Fos bZip phenotypic effects mayreflect novel heterodimerizations be-tween truncated Fos and wild-typeforms of zipper proteins. Importantly,a leucine zipper like-motif withinMsl1 protein was demonstrated to me-diate association with Msl2 (Li et al.,2005). This association appears toform the scaffold for binding othercomponents of the dosage compensa-tion complex, as well as for DNA bind-ing (Lucchesi et al., 2005; Li et al.,2005, 2008). Therefore, one hypothe-sis to explain the male phenotypesfrom Fos bZip expression involves in-terference with the assembly of thecomplex through effects on Msl1 orMsl2. To test this possibility, we de-termined if overexpression of heatshock-regulated wild-type Msl1, Msl2,or the combination Msl 1�Msl2 (Denget al., 2005) could rescue the Fos bZipwing phenotype. After a daily heatshock regimen, we observed partial,but significant rescue of the vg-Fbzphenotype with expression of the Msl1and Msl1�Msl2 constructs (Table 2),indicating an effect of Msl1 overex-pression. Msl1 and Msl2 bind to eachother through interactions of theiramino terminal regions (Li et al., 2005).Therefore, we examined whether over-expression of truncated forms of Msl1 orMsl2 enhanced the Fos bZip pheno-types. If Fos bZip elicits its effectsthrough a perturbation of this interac-tion, then overexpression of Msl1 orMsl2 forms containing only amino ter-minal regions may modify the Fos bZipphenotype. We used heat shock-regu-lated transgenes carrying residues1-265 of Msl1 and 1-193 of Msl2, whichcontain the polypeptide regions impli-cated in the Msl1-Msl2 physical associ-ation, to investigate this idea (Li et al.,2005, 2008). Significant enhancementof the Fos bZip phenotype was ob-served after expression of each trun-cated protein (44% vs. 67%; Table 3).Enhancement was also seen after ex-pression of a carboxy segment ofMsl-2, containing residues 490-773(44% vs. 74%; Table 3). Although thissegment of Msl-2 is not implicated in
TABLE 1. Genetic Interaction of Fos bZip With msl Loci
Genotype % Male % Female
vg-Fbz/w1118 57 (687) 0.2 (1127)vg-Fbz/Gla* 53 (260) 0 (100)vg-Fbz/msl-2,1,mle **73 (226) 0 (208)vg-Fbz/mle **71 (120) 0 (166)vg-Fbz/�; msl-3/� **71.5 (158) 0 (121)vg-Fbz/�; msl-3 RNAi/� **77.4 (297) 0 (582vg-Gal4/�; msl-3 RNAi/� 4.5 (308) 0 (268)
vg-Fbz was crossed to w1118 controls, mutations in msl loci and an msl3 RNAiconstruct, and wings scored for defects. %male and %female represent percentage ofwings exhibiting a defect. The number of wings scored in each class is indicated inparentheses.*vg-Fbz/Gla class is an internal control for the genetic background of mslchromosomes.
**Significant enhancement of wing defects relative to vg-Fbz/w1118 controls P �0.001, �2 test.
MALE SPECIFIC PHENOTYPES 3367

the physical interaction with Msl-1, ithas been shown to bind MLE and in-fluence rox RNA function (Li et al.,2008). In contrast, expression of anMsl-1 protein deleted for the amino-terminal 84 amino acids (Msl-1�84)significantly rescued the Fos bZip phe-notype (31% vs. 44%; Table 3). Be-cause Msl-1�84 retains the region re-quired for association with Msl2 (Scottet al., 2000), this truncation may ti-trate out the Fos bZip polypeptide. Nomale wing effects of Msl truncationexpression were observed in controlcrosses to w1118, or in experimentalfemale offspring.
The Male-specific Effect ofUAS-Fbz Expression IsAssociated With an AberrantMale X ChromosomeMorphology
To investigate further the potentialrelationship between Fos bZip expres-sion and dosage compensation, we de-termined if expression of Fos bZip af-fected formation of the dosagecompensation complex on male poly-tene X chromosomes. UAS-Fbz wasdriven in third instar larval salivaryglands by means of a ptc-Gal4 trans-
gene, and the preparations werestained with antibodies against Msl1and Mle proteins to measure the sta-tus of the complex (Sass et al., 2003).The polytene chromosomes derivedfrom the ptc-Gal4/UAS-Fbz genotype(Fig. 7A,I,J) were smaller than thoseof wild-type or ptc-Gal4/w1118 controls(Fig. 7E,K,L), as revealed throughstains with DAPI (4�,6-diamidine-2-phenylidole-dihydrochloride). More-over, the male X chromosome exhib-ited a particularly diffuse structurewith poor banding characteristics(Fig. 7A,J arrows). Despite this aber-rant structure, formation of the dos-
Fig. 6. Loss of roX function enhances the vg-Fbz wing phenotype. Females heterozygous for an X chromosome partially deficient for roX1 functionand deleted for roX2: roX1ex21A Df(1)52/Binscy were mated to vg-Fbz/Cy; w�4�4.3/ w�4�4.3 males. Non Cy males were scored for wing phenotypes.A: Wings from roX1ex21A Df(1)52/Y; vg-Fbz/�; w�4�4.3/� males show significant nicking along anterior and posterior margin at high penetrance (95%N � 43 wings). B: Control wings from Binscy/Y; vg-Fbz/�; w�4�4.3/� males show minor nicking at lower penetrance (20% N � 69). C: roX1ex21A
Df(1)52/Y; w�4�4.3/�. D: Binscy/Y; w�4�4.3/� from crosses of roX1ex21A Df(1)52/Binscy females to w�4�4.3/ w�4�4.3 males did not exhibit anywing defects.
TABLE 2. Partial Rescue of Fos bZip Phenotype Via Msl1 Overexpressiona
w1118 hs-msl-1 hs-msl-2 hs-msl-1�msl-2
% Male % Female % Male % Female % Male % Female % Male % Femalevg-Fbz 39.1 (621) 0.2 (733) *25.9 (502) 0 (673) 34.6 (292) 0.6 (224) *26.3 (293) 0 (0)
vg-Fbz was crossed to w1118 controls and msl1, msl2, and msl1 � msl2 strains under heat shock regulation. Vials were heat shockeddaily and wings scored for defects. %male and %female represent percentage of wings exhibiting a defect. The number of wingsscored in each class is indicated in parentheses. We observed a decrease in the percentage of control wings that exhibited defectsafter heat shocking, as well as extended development times (Table 1 vs. Table 2 and Table 3 data). No females overexpressinghs-msl-1 � hs-msl-2 were recovered since high levels of Msl2 activate the dosage compensation mechanism on female Xchromosomes (Lucchesi et al., 2005).*Rescue of wing defects was significant for males relative to control: hs-msl-1 and hs-msl-1�hs-msl-2 P � 0.001, �2 test.
3368 PIERRE ET AL.
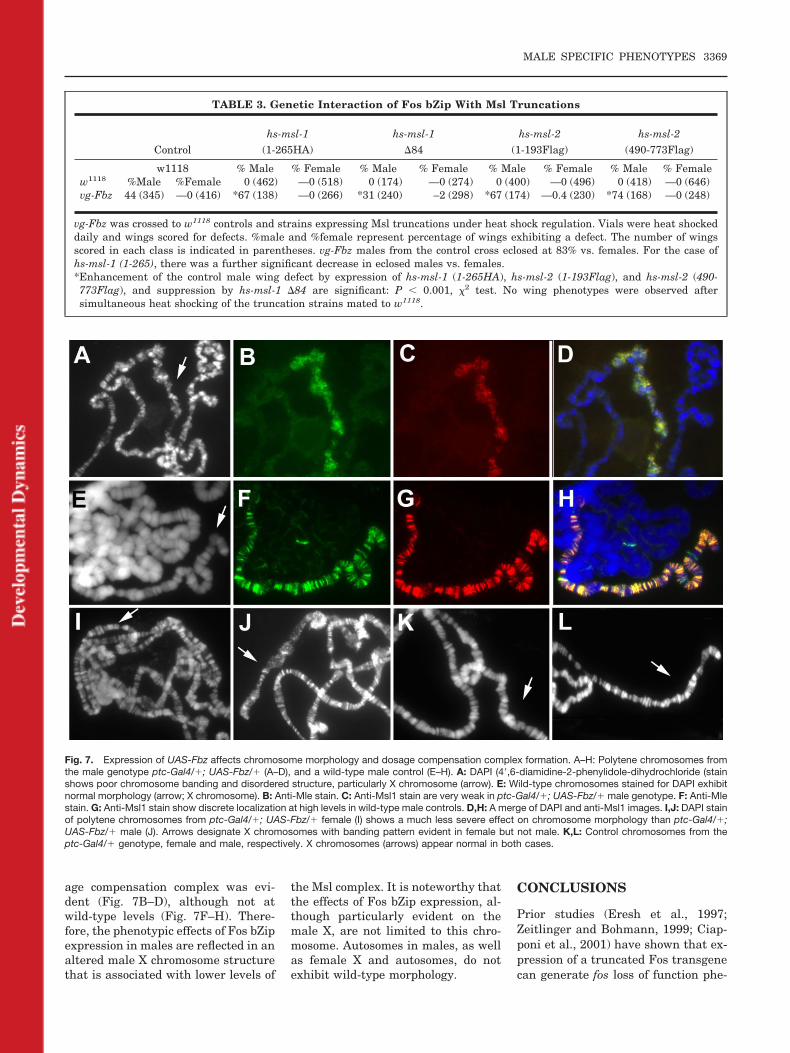
age compensation complex was evi-dent (Fig. 7B–D), although not atwild-type levels (Fig. 7F–H). There-fore, the phenotypic effects of Fos bZipexpression in males are reflected in analtered male X chromosome structurethat is associated with lower levels of
the Msl complex. It is noteworthy thatthe effects of Fos bZip expression, al-though particularly evident on themale X, are not limited to this chro-mosome. Autosomes in males, as wellas female X and autosomes, do notexhibit wild-type morphology.
CONCLUSIONS
Prior studies (Eresh et al., 1997;Zeitlinger and Bohmann, 1999; Ciap-poni et al., 2001) have shown that ex-pression of a truncated Fos transgenecan generate fos loss of function phe-
Fig. 7. Expression of UAS-Fbz affects chromosome morphology and dosage compensation complex formation. A–H: Polytene chromosomes fromthe male genotype ptc-Gal4/�; UAS-Fbz/� (A–D), and a wild-type male control (E–H). A: DAPI (4�,6-diamidine-2-phenylidole-dihydrochloride (stainshows poor chromosome banding and disordered structure, particularly X chromosome (arrow). E: Wild-type chromosomes stained for DAPI exhibitnormal morphology (arrow; X chromosome). B: Anti-Mle stain. C: Anti-Msl1 stain are very weak in ptc-Gal4/�; UAS-Fbz/� male genotype. F: Anti-Mlestain. G: Anti-Msl1 stain show discrete localization at high levels in wild-type male controls. D,H: A merge of DAPI and anti-Msl1 images. I,J: DAPI stainof polytene chromosomes from ptc-Gal4/�; UAS-Fbz/� female (I) shows a much less severe effect on chromosome morphology than ptc-Gal4/�;UAS-Fbz/� male (J). Arrows designate X chromosomes with banding pattern evident in female but not male. K,L: Control chromosomes from theptc-Gal4/� genotype, female and male, respectively. X chromosomes (arrows) appear normal in both cases.
TABLE 3. Genetic Interaction of Fos bZip With Msl Truncations
Controlhs-msl-1
(1-265HA)hs-msl-1
�84hs-msl-2
(1-193Flag)hs-msl-2
(490-773Flag)
w1118 % Male % Female % Male % Female % Male % Female % Male % Femalew1118 %Male %Female 0 (462) —0 (518) 0 (174) —0 (274) 0 (400) —0 (496) 0 (418) —0 (646)vg-Fbz 44 (345) —0 (416) *67 (138) —0 (266) *31 (240) –2 (298) *67 (174) —0.4 (230) *74 (168) —0 (248)
vg-Fbz was crossed to w1118 controls and strains expressing Msl truncations under heat shock regulation. Vials were heat shockeddaily and wings scored for defects. %male and %female represent percentage of wings exhibiting a defect. The number of wingsscored in each class is indicated in parentheses. vg-Fbz males from the control cross eclosed at 83% vs. females. For the case ofhs-msl-1 (1-265), there was a further significant decrease in eclosed males vs. females.*Enhancement of the control male wing defect by expression of hs-msl-1 (1-265HA), hs-msl-2 (1-193Flag), and hs-msl-2 (490-773Flag), and suppression by hs-msl-1 �84 are significant: P � 0.001, �2 test. No wing phenotypes were observed aftersimultaneous heat shocking of the truncation strains mated to w1118.
MALE SPECIFIC PHENOTYPES 3369

notypes. We have observed that thisconstruct also elicits sex-differentialphenotypes when expressed in varioustissues, indicating a neomorphic effecton processes not normally associatedwith fos. At the wing margin, the maleeffect synergizes with Notch pathwaymutations and affects expression ofthe Cut and Wg proteins, which areregulated by Notch signaling. Themale effect is not recapitulated by de-pression of Fos protein mediated by aFosRNAi transgene, despite the abilityof this transgene to induce a wingnicking phenotype. Furthermore, incontrast to Notch pathway mutations,alleles of fos do not modify the FosbZip wing phenotype. These data im-plicate Fos bZip in two anomalous ef-fects. First, it can interfere with Notchsignaling during wing margin forma-tion, and second, the interference isonly detected in the male genotype.Our data linking Fos bZip to dosagecompensation explains both effects.The Notch interaction likely reflects adiminution in the activity of X-linkedgenes, including Notch, mediated byFos bZip. Consistent with this, coex-pression of activated Notch can rescuethe Fos bZip wing phenotype to a sub-stantial extent (data not shown).
We have shown that the male-spe-cific wing phenotype of Fos bZip is en-hanced by mutations in msl and mleloci, an RNAi construct directedagainst msl3 product, simultaneousloss of roX1 and roX2, and truncatedversions of Msl1 and Msl2. Con-versely, the same phenotype is par-tially rescued by overexpression ofwild-type Msl1, and a version of Msl1deleted for a small part of the aminoterminus. One explanation for male-specific effects of Fos bZip is based onthe observation that the Msl1 aminoterminal end contains a leucine zip-per-like domain essential for associa-tion with Msl2 (Li et al., 2005). FosbZip may act as a competitive inhibi-tor of the association and limit forma-tion of the complex. This explanationis supported by our data derived fromMsl truncations and wild-type Msl1.Although expression of Msl1 and Msl2truncations alone produced no wingphenotypes, it greatly sensitizedmales to the effect of Fos bZip.
We also observed a correlation be-tween the male effects of Fos bZipexpression and aberrant morphology
of polytene chromosomes, especiallyX chromosomes in males. Also ofnote was lower than normal stainingfor the dosage compensation complexon the male X. Given the diminishedformation of the complex, our datashowing genetic modification of thevg-Fbz wing phenotype through al-tered levels/function of msl productsis explicable. However, despite thesegenetic data, the Fos bZip effect ondosage compensation may be indi-rect. Rather than interfering with anMsl1-Msl2 physical association, it isconceivable that Fos bZip perturbsthe structure of the X chromosome,and that this sensitizes males by de-pressing the ability of the dosagecompensation complex to function.For example, if the affinity of themale X chromosome for the complexwas lowered by Fos bZip, then de-pressions in the levels of Msl pro-teins would be expected to reducethe level of bound complex even fur-ther. Three observations supportthis idea of a more indirect effect.First, male autosomal polytene chro-mosome structure, and female X andautosomal structure are also abnor-mal in the ptc-Gal4�UAS-Fbz geno-type. This suggests an effect on chro-mosome structure independent ofdosage compensation. Second, virtu-ally all vg-Fbz/� female wings ap-pear wild-type (Table 1). However,homozygous vg-Fbz/vg-Fbz femalessurvive at a low rate, and these sur-vivors can show nicked wings (datanot shown). This demonstrates an ef-fect on female gene expression.Third, mutations shown to affectchromosome structure can impactdosage compensation (Delattre etal., 2004; Spierer et al., 2008), simi-lar to the Fos bZip effect proposedhere.
Finally, the Drosophila genome en-codes numerous proteins with bZip do-mains (Fassler et al., 2002), and addi-tional proteins with zippers that arenot closely flanked by basic residues.Our results provide insight into theneomorphic effects that may derivefrom atypical mutations in this classof proteins. Moreover, such effectscould in principle arise from eithernaturally occurring mutations, or al-ternatively, in vitro-produced trans-genic constructs.
EXPERIMENTALPROCEDURES
Drosophila Strains
C96-Gal4, blk-Gal4, glass-Gal4, pnr-Gal4, C96-MamH, EP 3084, w1118,DeltaBX9/TM6c, mamIL115/Cy, and nd1
were described previously (Schmid etal., 1996; Helms et al., 1999; Hall etal., 2004). vg-Gal4 was obtained fromthe Artavanis-Tsakonas lab (Har-vard). These Gal4 strains were used todrive expression as follows: C96-Gal4(wing margin), blk-Gal4 (wing veinand leg), glass-Gal4 (eye), pnr-Gal4(notum), vg-Gal4 (wing margin). C96-MamH is a recombinant chromosomethat contains the C96-Gal4 driver anda UAS-MamH transgene. The lattertransgene encodes a truncated form ofthe Notch pathway effector Master-mind (Helms et al., 1999), and the re-combinant chromosome produces a100% penetrant wing nicking pheno-type. yw; delta 2-3 Ki was obtainedfrom Kevin Moses (HHMI) and da-Gal4 (embryonic expression) from K.Bhat (UT Galveston). ptc-Gal4 (salivarygland and embryonic expression), FosbZip: y[1] w[1118]; P{w[�mC]�UAS-Fra.Fbz}5, Jun bZip: y[1] w[1118];P{w[�mC]�Jbz1, CrebB bZip: y[1]w[1118]; P{w[�mC]�Cbz4, kaysro-1/TM3 Sb, kay1/TM3 Sb Ser, and Jra109/CyO were obtained from the Blooming-ton stock center. UAS-FosRNAi (FI 35/19III) was obtained from D. Bohmann(Rochester), P[ry� �2-3] (99B) was ob-tained from Kazemi-Esfarjani in theBenzer lab (CalTech). msl-3 red/TM3, eSb Ser, mle[185] cn bw/CyO and msl-2msl-1 mle/CyO have been describedpreviously (Lucchesi et al., 2005). hs-msl-1 �84, hs-msl-1 (1-265HA), hs-msl-2 (1-193Flag) and hs-msl-2 (490-773Flag) were obtained from M. Scott(Massey University). roX1ex21A Df(1)-52/Binscy, w�4�4.3/w�4�4.3, hs-Msl-1-Z1, hs-Msl-2-6I and the recombinanths-Msl-1-Z1�hs-Msl-2-6I were ob-tained from V. Meller (Wayne State). AUAS-regulated RNAi transgene di-rected against msl-3 RNA was obtainedfrom the VDRC (Dietzl et al., 2007). Forgenetic interaction experiments UAS-Fbz was recombined onto chromosome 2with the vg-Gal4 driver, creating thevg-Fbz chromosome. Crosses were per-formed at 22°C.
3370 PIERRE ET AL.

Heat Shock Conditions
Mated flies were allowed to lay eggsfor 24–72 hr and then vials were heattreated daily one to three times in awater bath at 37°C for 1 hr. A 2-hrrecovery period was used betweenheat treatments.
Mobilization of the UAS-FosbZip Transgene FromChromosome 2
The P[ry� �2-3] (99B) strain of trans-posase was mated to a CyO strain andresulting CyO/�; P[ry� �2-3]� weremated with homozygous w� UAS-Fra.Fbz}5. w� UAS-Fra.Fbz}5/CyO;�/ P[ry� �2-3 flies showing mottledeyes were mated to w1118 and the re-sulting F1 were selected for both w�and CyO. We obtained new insert siteson the CyO chromosome as well aschromosome three.
Identification of GenomicSequence Flanking EP 3084Through Inverse PolymeraseChain Reaction
The EP 3084 strain derived from theRorth collection (Rorth, 1996) previ-ously maintained by Exelixis. Weidentified EP 3084 in a prior geneticmodifier screen as a Notch pathwayphenotype suppressor (Hall et al.,2004), however, at that time we hadnot corroborated its genomic locationwhich was designated as chromosome3 heterochromatin. Consequently, weisolated EP 3084 flanking genomic se-quences by means of inverse polymer-ase chain reaction (PCR). GenomicDNA was isolated by the followingprotocol. Flies were ground in lysisbuffer (100 mM NaCl, 25 mM ehty-lenediaminetetraacetic acid, 0.5% so-dium dodecyl sulfate, 10 mM Tris pH8.0), containing 250 �g/ml proteinaseK. The extract was incubated at 50°Cfor 60 min and then phenol-chloro-form extracted until the interface wasclean. The DNA was then ethanol pre-cipitated two times, washed in 80%ethanol, and resuspended in TE. Pro-cedures for restriction digestion, liga-tion, and inverse PCR were derivedfrom the BDGP Web site (www.fruit-fly.org/about/methods/inverse.pcr).PCR products were gel purified, quan-tified and sent to the Iowa State Se-
quencing and Synthesis Facility forsequence determination (http://dna.biotech.iastate.edu). Sequence datawas initially analyzed by means ofFlyBlast (http://flybase.net/blast) andsubsequently through the Apollo Ge-nome Annotation and Curation Tool(http://www.fruitfly.org/annot/apollo).This analysis revealed that EP 3084 issituated approximately 1,200 basepairs upstream of the kayak (D-Fos)locus, with its UAS-promoter orientedto drive expression of the locus in ge-notypes containing a Gal4 driver.
Antibody Staining of ThirdInstar Larval Wing Discs
The following protocol (Brennan et al.,1998) was followed for the fixation,staining, and washing of imaginaldiscs. Discs were dissected in 0.1 Msodium phosphate buffer (pH 7.4),fixed (2% paraformaldehyde, 75 mMlysine, 0.25% sodium periodate, 50mM sodium phosphate pH 7.4) for 45min and washed (0.1 M sodium phos-phate, 0.1% Triton X-100, pH 7.4).They were blocked in wash buffer con-taining 10% normal goat serum andincubated with primary antiserum for60 min at 37°C. The discs were thenwashed twice, blocked, and incubatedwith secondary antibodies overnightat 4°C. After three final washes, thediscs were mounted in Slowfade (Mo-lecular Probes). Images were obtainedusing a Bio-Rad MRC 1024 confocalmicroscope and assembled with AdobePhotoshop. The following antibodies/dilutions were used: Wg (1:20 mousemonoclonal, Developmental StudiesHybridoma Bank), Cut (1:20 mousemonoclonal, Developmental StudiesHybridoma Bank), Fos (1:250 rabbitpolyclonal, from D. Bohmann, Roches-ter). Alexa 488 and Alexa 565 second-ary antibodies were obtained from In-vitrogen.
Mounting of Wings andNotums
Wings were dehydrated in isopropanoland mounted in Euparol. Adult tis-sues were positioned on slides andphotographed under a dissecting mi-croscope (Hall et al., 2004).
Antibody Staining ofPolytene Chromosomes
Salivary glands were dissected fromthird instar larvae. Chromosomeswere fixed and stained with antibod-ies to Mle and Msl-1, and prepared forimmunofluorescence as previously de-scribed (Sass et al., 2003).
ACKNOWLEDGMENTSWe thank Dirk Bohmann for provid-ing reagents, the Bloomington StockCenter and the various fly labs thatprovided strains. Particular thanks toMax Scott and Victoria Meller for sup-plying multiple strains, and for theiradvice. Supported by National ScienceFoundation grant IBN-0415453 (B.Y.)and NIH Grant GM15691 (J.L.).
REFERENCES
Brennan CA, Ashburner M, Moses K. 1998.Ecdysone pathway is required for furrowprogression in the developing Drosophilaeye. Development 125:2653–2664.
Ciapponi L, Jackson DB, Mlodzik M, Bohm-ann D. 2001. Drosophila Fos mediates ERKand JNK signals via distinct phosphoryla-tion sites. Genes Dev 15:1540–1553.
Delattre M, Spierer A, Jaquet Y, Spierer P.2004. Increased expression of DrosophilaSu(var)3-7 triggers Su(var)3-9-depen-dent heterochromatin formation. J CellSci 117:6239–6247.
Deng X, Rattner BP, Souter S, Meller VH.2005. The severity of roX1 mutations ispredicted by MSL localization on the Xchromosome. Mech Dev 1221094–1105.
Dietzl G, Chen D, Schnorrer F, Su KC,Barinova Y, Fellner M, Gasser B, KinseyK, Oppel S, Scheiblauer S, Couto A,Marra V, Keleman K, Dickson BJ. 2007.A genome-wide transgenic RNAi libraryfor conditional gene inactivation in Dro-sophila. Nature 448:151–156.
Eresh S, Riese J, Jackson DB, Bohmann D,Bienz M. 1997. A CREB binding site as atarget for decapentaplegic signaling dur-ing Drosophila endoderm induction.EMBO J 16:2014–2022.
Fassler J, Landsman D, Acharya A, MollJR, Bonovich M, Vinson C. 2002. B-Zipproteins encoded by the Drosophila ge-nome: evaluation of potential dimerizationpartners. Genome Res 12:1190–1200.
Hall LE, Alexander SJ, Chang M, Woo-dling NS, Yedvobnick B. 2004. An EPover expression screen for genetic modi-fiers of Notch pathway function in Dro-sophila. Genet Res 83:71–82.
Helms W, Lee H, Ammerman M, Parks A,Muskavitch M, Yedvobnick B. 1999. En-gineered truncations in the DrosophilaMastermind protein disrupt Notch path-way function. Dev Biol 215:358–374.
Hess J, Angel P, Schorpp-Kistner M. 2004.AP-1 subunits: quarrel and harmonyamong siblings. J Cell Sci 117:5965–5973.
MALE SPECIFIC PHENOTYPES 3371

Hilfiker A, Hilfiker-Kleiner D, Pannuti A,Lucchesi JC. 1997. mof, a putative acetyltransferase gene related to the Tip60and MOZ human genes and to the SASgenes of yeast, is required for dosagecompensation in Drosophila. EMBO J16:2054–2060.
Hyun J, Becam I, Yanicostas C, BohmannD. 2006. Control of the G2/M transitionby Drosophila Fos. Mol Cell Biol26:8293–8302.
Kelley RL, Wang J, Bell L, Kuroda M.1997. Sex lethal controls dosage compen-sation in Drosophila by a non-splicingmechanism. Nature 387:195–199.
Kockel L, Homsy JG, Bohmann D. 2001.Drosophila AP-1: lessons from an inver-tebrate. Oncogene 20:2347–2364.
Li F, Parry DA, Scott MJ. 2005. The amino-terminal region of Drosophila MSL1 con-tains basic, glycine-rich and leucine zip-per-like motifs that promote Xchromosome binding, self-association,and MSL2 binding, respectively. MolCell Biol 25:8913–8924.
Li F, Schiemann AH, Scott MJ. 2008. In-corporation of the noncoding roX RNAs
alters the chromatin-binding specificityof the Drosophila MSL1/MSL2 complex.Mol Cell Biol 28:1252–1264.
Lucchesi JC, Kelly WG, Panning B. 2005.Chromatin remodeling in dosage com-pensation. Annu Rev Genet 39:615–651.
Meller VH, Rattner BP. 2002. The roXgenes encode redundant male-specific le-thal transcripts required for targeting ofthe MSL complex. EMBO J 21:1084–1091.
Neumann CJ, Cohen SM. 1996. A hierar-chy of cross-regulation involving Notch,wingless, vestigial and cut organizes thedorsal/ventral axis of the Drosophilawing. Development 122:3477–3485.
Rorth P. 1996. A modular misexpressionscreen in Drosophila detecting tissue-specific phenotypes. Proc Natl Acad SciU S A 93:12418–12422.
Sass GL, Pannuti A, Lucchesi JC. 2003.Male-specific lethal complex of Drosophilatargets activated regions of the X chromo-some for chromatin remodeling. Proc NatlAcad Sci U S A 100:8287–8291.
Schmid A, Tinley T, Yedvobnick B. 1996.Transcription of the neurogenic gene
mastermind during Drosophila develop-ment. J Exp Zool 274:207–220.
Scott MJ, Pan LL, Cleland SB, Knox AL,Heinrich J. 2000. MSL1 plays a centralrole in assembly of the MSL complex,essential for dosage compensation inDrosophila. EMBO J 19:144–155.
Smith ER, Pannuti A, Gu W, Steurnagel A,Cook RG, Allis CD, Lucchesi JC. 2000.The Drosophila MSL complex acetylateshistone H4 at lysine 16, a chromatinmodification linked to dosage compensa-tion. Mol Cell Biol 1:312–318.
Smith ER, Allis CD, Lucchesi JC. 2001.Linking global histone acetylation to thetranscription enhancement of X-chromo-somal genes in Drosophila males. J BiolChem 276:31483–31486.
Spierer A, Begeot F, Spierer P, Delattre M.2008. SU(VAR)3-7 links heterochroma-tin and dosage compensation in Dro-sophila. PLoS Genet 4:e1000066.
Zeitlinger J, Bohmann D. 1999. Thorax clo-sure in Drosophila: involvement of Fosand the JNK pathway. Development126:3947–3956.
3372 PIERRE ET AL.




















