
CXCL5 Drives Neutrophil Recruitment in TH17-Mediated GN
12
BASIC RESEARCH www.jasn.org CXCL5 Drives Neutrophil Recruitment in T H 17-Mediated GN Erik M. Disteldorf,* Christian F. Krebs,* Hans-Joachim Paust,* Jan-Eric Turner,* Geraldine Nouailles, † André Tittel, ‡ Catherine Meyer-Schwesinger,* Gesa Stege,* Silke Brix,* Joachim Velden, § Thorsten Wiech, | Udo Helmchen, | Oliver M. Steinmetz,* Anett Peters,* Sabrina B. Bennstein,* Anna Kaffke,* Chrystel Llanto, ‡ Sergio A. Lira, ¶ Hans-Willi Mittrücker,** Rolf A.K. Stahl,* Christian Kurts, ‡ Stefan H.E. Kaufmann, † and Ulf Panzer* *III. Medical Clinic and Institutes of | Pathology and **Immunology, Hamburg-Eppendorf University Hospital, Hamburg-Eppendorf, Germany; † Max Planck Institute for Infection Biology, Berlin, Germany; ‡ Institute of Molecular Medicine and Experimental Immunology, Bonn, Germany; § Department of Nephropathology, Erlangen University Hospital, Erlangen, Germany; and ¶ Immunology Institute, Icahn School of Medicine at Mount Sinai, New York, New York ABSTRACT Neutrophil trafficking to sites of inflammation is essential for the defense against bacterial and fungal infections, but also contributes to tissue damage in T H 17-mediated autoimmunity. This process is regu- lated by chemokines, which often show an overlapping expression pattern and function in pathogen- and autoimmune-induced inflammatory reactions. Using a murine model of crescentic GN, we show that the pathogenic T H 17/IL-17 immune response induces chemokine (C-X-C motif) ligand 5 (CXCL5) expression in kidney tubular cells, which recruits destructive neutrophils that contribute to renal tissue injury. By con- trast, CXCL5 was dispensable for neutrophil recruitment and effective bacterial clearance in a murine model of acute bacterial pyelonephritis. In line with these findings, CXCL5 expression was highly upregu- lated in the kidneys of patients with ANCA-associated crescentic GN as opposed to patients with acute bacterial pyelonephritis. Our data therefore identify CXCL5 as a potential therapeutic target for the re- striction of pathogenic neutrophil infiltration in T H 17-mediated autoimmune diseases while leaving intact the neutrophil function in protective immunity against invading pathogens. J Am Soc Nephrol 26: 55–66, 2015. doi: 10.1681/ASN.2013101061 Neutrophils are the most abundant type of leuko- cytes in the blood and form an indispensable part of the innate immune system. Their trafficking into peripheral tissues is pivotal in the defense against invading bacterial and fungal pathogens. 1 To ensure that neutrophils reach the sites of tissue injury, their recruitment is regulated by the local expression of chemoattractants, including chemokines. However, the infiltration of neutrophils also significantly con- tributes to end-organ damage in autoimmune diseases mediated by T helper (T H ) cell T H 17, 2 including human and experimental crescentic GN. 3,4 Chemokines are a large family of small (8–12 kD) secreted proteins that are identified as attractants of different types of leukocytes, including neutrophils, to sites of infection and inflammation. 5 They are produced locally in tissues and act through interaction with specific G protein–coupled receptors that are predominantly expressed on leukocytes. Neutrophil infiltration is mainly mediated by chemo- kines that have a glutamate-leucine-arginine motif (ELR + chemokines). In humans, there are seven ELR chemokine ligands with a C-X-C motif (CXCL1, CXCL2, CXCL3, CXCL5, CXCL6, Received October 10, 2013. Accepted April 4, 2014. E.M.D. and C.F.K. contributed equally to this work. Published online ahead of print. Publication date available at www.jasn.org. Correspondence: Dr. Ulf Panzer, Third Medical Clinic, Hamburg- Eppendorf University Hospital, Martinistrasse 52, 20246 Hamburg, Germany. Email: [email protected] Copyright © 2015 by the American Society of Nephrology J Am Soc Nephrol 26: 55–66, 2015 ISSN : 1046-6673/2601-55 55 BASIC RESEARCH
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of CXCL5 Drives Neutrophil Recruitment in TH17-Mediated GN

BASIC RESEARCH www.jasn.org
CXCL5 Drives Neutrophil Recruitment inTH17-Mediated GN
Erik M. Disteldorf,* Christian F. Krebs,* Hans-Joachim Paust,* Jan-Eric Turner,*Geraldine Nouailles,† André Tittel,‡ Catherine Meyer-Schwesinger,* Gesa Stege,* Silke Brix,*Joachim Velden,§ Thorsten Wiech,| Udo Helmchen,| Oliver M. Steinmetz,* Anett Peters,*Sabrina B. Bennstein,* Anna Kaffke,* Chrystel Llanto,‡ Sergio A. Lira,¶ Hans-Willi Mittrücker,**Rolf A.K. Stahl,* Christian Kurts,‡ Stefan H.E. Kaufmann,† and Ulf Panzer*
*III. Medical Clinic and Institutes of |Pathology and **Immunology, Hamburg-Eppendorf University Hospital,Hamburg-Eppendorf, Germany; †Max Planck Institute for Infection Biology, Berlin, Germany; ‡Institute of MolecularMedicine and Experimental Immunology, Bonn, Germany; §Department of Nephropathology, Erlangen UniversityHospital, Erlangen, Germany; and ¶Immunology Institute, Icahn School of Medicine at Mount Sinai, New York, New York
ABSTRACTNeutrophil trafficking to sites of inflammation is essential for the defense against bacterial and fungalinfections, but also contributes to tissue damage in TH17-mediated autoimmunity. This process is regu-lated by chemokines, which often show an overlapping expression pattern and function in pathogen- andautoimmune-induced inflammatory reactions. Using a murine model of crescentic GN, we show that thepathogenic TH17/IL-17 immune response induces chemokine (C-X-Cmotif) ligand 5 (CXCL5) expression inkidney tubular cells, which recruits destructive neutrophils that contribute to renal tissue injury. By con-trast, CXCL5 was dispensable for neutrophil recruitment and effective bacterial clearance in a murinemodel of acute bacterial pyelonephritis. In line with these findings, CXCL5 expression was highly upregu-lated in the kidneys of patients with ANCA-associated crescentic GN as opposed to patients with acutebacterial pyelonephritis. Our data therefore identify CXCL5 as a potential therapeutic target for the re-striction of pathogenic neutrophil infiltration in TH17-mediated autoimmune diseases while leaving intactthe neutrophil function in protective immunity against invading pathogens.
J Am Soc Nephrol 26: 55–66, 2015. doi: 10.1681/ASN.2013101061
Neutrophils are the most abundant type of leuko-cytes in the blood and form an indispensable part ofthe innate immune system. Their trafficking intoperipheral tissues is pivotal in the defense againstinvading bacterial and fungal pathogens.1 To ensurethat neutrophils reach the sites of tissue injury, theirrecruitment is regulated by the local expression ofchemoattractants, including chemokines. However,the infiltration of neutrophils also significantly con-tributes to end-organ damage in autoimmune diseasesmediated by T helper (TH) cell TH17,2 includinghuman and experimental crescentic GN.3,4
Chemokines are a large familyof small (8–12 kD)secreted proteins that are identified as attractants ofdifferent types of leukocytes, including neutrophils,to sites of infection and inflammation.5 They areproduced locally in tissues and act through
interactionwith specificGprotein–coupled receptorsthat are predominantly expressed on leukocytes.Neutrophil infiltration ismainlymediated by chemo-kines that have a glutamate-leucine-arginine motif(ELR+ chemokines). In humans, there are sevenELR chemokine ligands with a C-X-C motif(CXCL1, CXCL2, CXCL3, CXCL5, CXCL6,
Received October 10, 2013. Accepted April 4, 2014.
E.M.D. and C.F.K. contributed equally to this work.
Published online ahead of print. Publication date available atwww.jasn.org.
Correspondence: Dr. Ulf Panzer, Third Medical Clinic, Hamburg-Eppendorf University Hospital, Martinistrasse 52, 20246 Hamburg,Germany. Email: [email protected]
Copyright © 2015 by the American Society of Nephrology
J Am Soc Nephrol 26: 55–66, 2015 ISSN : 1046-6673/2601-55 55
BASICRESE
ARCH

CXCL7, and CXCL8/IL-8). Their chemotactic effects are me-diated via binding to the chemokine receptors CXCR1 andCXCR2. Mice lack complete homologs of the seven humanELR chemokines and have only five members (CXCL1,CXCL2, CXCL3, CXCL5, and CXCL7, which all bind to themurine CXCR2).6 Interestingly, previous reports show that IL-17A, the master effector cytokine of TH17 cells, induces theexpression of the ELR+ chemokines CXCL1, CXCL2, andCXCL57,8 and thereby might drive the recruitment of patho-genic neutrophils in autoimmunity. The development of atherapeutic strategy targeting ELR+ neutrophil–attracting che-mokines or their receptors is complicated by an often overlap-ping expression pattern and function of these molecules inpathogen- and autoimmune-induced inflammatory reac-tions.9
Herewe describe for the first time a nonredundant functionof the chemokines CXCL1 and CXCL5 in murine models ofcrescentic GN and acute bacterial pyelonephritis. CXCL1mediated early glomerular neutrophil recruitment in the non-T cell–dependent initiation phase of GN, whereas CXCL5was responsible for the infiltration of pathogenic neutrophilsinto sites of inflammation in later TH17-dependent phases ofthe disease. Of note, CXCL5 did not affect neutrophil infil-tration and bacterial clearance in a murine model of acutebacterial pyelonephritis, one of the most prevalent kidneyinfections in humans. These findings suggest that CXCL5has a unique function in the trafficking of neutrophils inTH17 cell–mediated autoimmunity, but not in the innateimmune response. CXCL5 therefore represents an attractivetherapeutic target for the restriction of pathogenic neutrophilinfiltration in TH17-driven autoimmune diseases without af-fecting the vital functions of neutrophils in the defense againstacute bacterial infections.
RESULTS
Time- and Compartment-Specific Infiltration ofNeutrophils in Murine Crescentic GNNephrotoxic nephritis (NTN) is a well characterized model ofmurine crescentic GN, which is induced by the injection ofsheep antiserum raised against kidney cortical components.During the early heterologous phase of the disease, thedeposited antibodies result in glomerular complement acti-vation and neutrophil recruitment, which cause substantialglomerular injury and renal dysfunction.10 An adaptive im-mune response against the foreign sheep protein develops inthe subsequent autologous phase (starting from days 3 to 5),resulting in the activation of nephritogenic CD4+ TH17 andTH1 cells in lymphatic organs. First, TH17 cells and, sub-sequently, TH1 cells migrate into the kidney and promoterenal tissue injury.11–13 The role of neutrophils in the T cell–mediated phase (starting from day 5) is largely unknown. Wetherefore assessed the time course of renal neutrophil infiltra-tion using immunohistochemical staining for the neutrophil
marker Gr1 (Ly6C/Ly6G) (Figure 1A). In the early stage ofnephritis (until day 3), neutrophils were mainly found in theglomerulus (Figure 1, A and B). The infiltration of neutrophilsinto the tubulointerstitial area started at day 5, peaked aroundday 10, and then declined (Figure 1, A and B). Thisdemonstrates a previously unknown time- and compart-ment-specific recruitment of neutrophils into the kidney.
CXCL1 and CXCL5 Have Unique Functions in theRecruitment of Neutrophils in Crescentic GNOne important prerequisite for the infiltration of neutrophilsinto sites of inflammation is the expression of ELR+ chemo-kines, namely CXCL1, CXCL2, CXCL3, CXCL5, and CXCL7,which act via the chemokine receptor CXCR2. QuantitativeRT-PCR analysis of the renal cortex revealed that CXCL1,CXCL2, and CXCL3 mRNA expression was strongly upregu-lated in the early stages of nephritis (12 and 24 hours) (Figure1C). Renal CXCL7 expression was low throughout the diseasecourse (data not shown). CXCL1 expression rapidly declinedto the baseline level by day 3. By contrast, CXCL5 mRNAexpressionwas not increased at early time points but markedlyincreased from day 5 onward, with maximum expression lev-els at day 10 (detectable after approximately 25 PCR cycles).Chemokines can be modified post-translationally by proteo-lytic cleavage in order to achieve the active form and storedintracellular, therefore, a discrepancy between local mRNAlevel and protein level/activity might occur. However, our at-tempts to quantify renal CXCL5 protein levels in nephriticmice by ELISA failed because of high background signalswhen using CXCL5 knockout (KO) mice as negative controls(data not shown).
Renal mRNA expression of CXCR2 was characterized by abimodal course reaching two maxima after 24 hours and 10days (Figure 1C). In line with this, glomerular and tubuloin-terstitial neutrophil recruitment was reduced in nephriticCXCR22/2 mice at days 3 and 14 (Figure 1D). This showedthat CXCR2 drives both the “early” and “late” waves of neu-trophil recruitment.
To elucidate whether the expression patterns of CXCL1 andCXCL5might result in different functions of these chemokinesin the disease course, we performed early (until day 3) and late(days 9 to 14) in vivo neutralization with specific anti-CXCL1or anti-CXCL514 antibodies. In line with their temporal expressionprofile, neutralization of CXCL1 resulted in ameliorated glo-merular neutrophil infiltration at day 3, whereas anti-CXCL5antibody treatment at day 9 resulted in significantly reducedtubulointerstitial neutrophil recruitment at day 14 (Figure 1E).Accordingly, renal FACS analysis revealed a reduction inCD45+CD11b+Ly6G+ neutrophils in nephritic CXCL52/2
mice at day 14 (Figure 1, F and G), but not at day 3 (Supple-mental Figure 1A). Likewise, tubulointerstitial neutrophil accu-mulation was significantly decreased in nephritic CXCL52/2
mice at day 14 (Figure 1H), but not at day 3 (SupplementalFigure 1B). Immunofluorescence staining revealed that neutro-phils in nephritic CXCL52/2mice had a reduced capacity to exit
56 Journal of the American Society of Nephrology J Am Soc Nephrol 26: 55–66, 2015
BASIC RESEARCH www.jasn.org

the peritubular capillaries and to enter thetubulointerstitial space (Figure 1I).
By comparison, the infiltration of T cellsand macrophages/dendritic cells was notsignificantly affected by CXCL5 deficiencyor anti-CXCL5 treatment (SupplementalFigure 2, A–H, J). Furthermore, neutro-phil, monocyte, CD4+, and CD8+ T cellabundance in the blood, spleen, and kidneyunder homeostatic and nephritic condi-tions was not significant different betweenwild-type (WT) and CXCL52/2 mice (Sup-plemental Figure 3). In addition, kidneymorphology and function were not affectedby CXCL5 deficiency under basal conditions(Supplemental Figure 4). This indicates thatreduced neutrophil abundance in the kidneyof nephritic CXCL52/2 animals was not aconsequence of a developmental defect inthe hematopoietic system or the kidney, butrather represents reduced recruitment tothe inflamed tissue in CXCL52/2 mice.
To analyze the role of CXCL1 in the earlyinfiltration of neutrophils into the kidneyusing a second, independent approach, weinduced NTN in WT and CXCL1-deficientmice. In accordance with our anti-CXCL1neutralization data, neutrophil recruitmentwas reduced in nephritic CXCL12/2 mice atday 3, thus underscoring the importance ofthis chemokine in the early course of nephri-tis (Figure 1, J and K).
Figure 1. CXCL1 and CXCL5 have unique functions in the time- and compartment-specific infiltration of neutrophils in crescentic GN. (A) Immunohistochemistry of kidneysections stained for the neutrophil marker GR1 at indicated time points after nephritisinduction. Inserts demonstrate the polymorphonuclear morphology of GR1+ cells(neutrophils). (B) Quantification of tubulointerstitial (per low-power field [lpf]) andglomerular (per glomerular cross-section [gcs]) PMN infiltration in the course of ne-phritis (n=3–6 per time point). (C) RT-PCR analysis of renal CXCL1, CXCL2, CXCL3,CXCL5, and CXCR2 mRNA expression (n=4–6 per time point). (D) Quantification oftubulointerstitial and glomerular PMNs in nephritic WT and CXCR22/2 mice at day 3or day 14 after induction of the disease (n=3–5 per time point). (E) Quantification oftubulointerstitial and glomerular PMNs in nephritic WT mice at day 3 or 14 of nephritisafter administration of anti-CXCL1, anti-CXCL5, or isotype rat-IgG2B antibodies (n=3–
5 per time point). (F) FACS analysis of renalleukocytes from nephritic WT and CXCL52/2
mice stained for the PMNmarker Ly6G at day14. Plots are representative of three in-dependent experiments. (G and H) Quantifi-cation of neutrophils from FACS analysis (G)and quantification of tubulointerstitial PMNsfrom GR1 stained kidney sections (H) in ne-phritic WT and CXCL52/2 mice 14 days afternephritis induction. (I) Immunofluorescencestaining of nephritic kidney section (day 14) ofWT and CXCL52/2 mice using anti-CD31(green/endothelial cells), anti-Ly6G (red/PMNs), and collagen type (blue/basementmembrane). (J and K) Quantification of renalneutrophils by FACS analysis (J) and quanti-fication of glomerular PMNs fromGR1 stainedkidney sections (K) in nephritic WT andCXCL12/2mice 3days after nephritis induction.Symbols/bars represent means6SDs. *P,0.05;**P,0.01; ***P,0.001. Original magnification,3200. Con, control.
J Am Soc Nephrol 26: 55–66, 2015 CXCL5 Recruits Neutrophils in TH17 Response 57
www.jasn.org BASIC RESEARCH

Kidney Tubular Cells Produce CXCL5 in Late CrescenticGNTo localize renal CXCL5 expression, we performed in situ hy-bridization experiments. CXCL5 expression was clearly de-tectable in tubular cells 14 days after nephritis induction,but not at day 3 or in control mice (Figure 2A).
Replacement of the CXCL5 gene by a b-galactosidase(b-Gal) gene in CXCL5-deficient mice enabled us to indirectlyassess the renal CXCL5 protein expression by b-Gal detection.Nuclear b-Gal staining was observed exclusively in tissues
collected from nephritic CXCL52/2 mice(Figure 2B). In line with the CXCL5mRNA expression pattern, b-Gal–positivecells were detected in the tubular epithe-lium of nephritic mice (Figure 2B).
Quantitative RT-PCR analysis of glo-merular and tubulointerstitial tissue sam-ples isolated from nephritic mice usinglasermicrodissection at days 3 and 14 afterinduction of NTN revealed that CXCL5mRNA expression was predominantlyupregulated at day 14 in the tubulointer-stitial compartment, as indicated by thetubulointerstitial to glomerular expres-sion ratio (Figure 2C). CXCL1mRNAwasmainly expressed in glomeruli at day 3,whereas glomerular and tubulointersti-tial expression were at control levels atday 14 (Figure 2C).
To study whether CXCL5 is mainly pro-duced by renal tissue cells, we generatedbonemarrow (BM) chimeras usingWTandCXCL52/2mice as donors (Figure 2D). In-dependent of the BM transplant used, 14days after induction of nephritis, highCXCL5 mRNA expression levels wereonly present in animals with WT renal tis-sue cells. An approximately 500-fold in-crease in CXCL5 mRNA expression wasfound in both WT recipient mice trans-planted with WT BM or CXCL52/2 BMcompared with control mice. By contrast,CXCL5 mRNA expression in CXCL52/2
mice receiving WT BM or CXCL52/2 BMwas not elevated (Figure 2D). As control,we used WTmice carrying the CD45.2 al-lele as recipients. Six weeks after transplan-tation, .90% of blood leukocytes, .98%of BM leukocytes, and .90% of splenicleukocytes expressed the phenotypicmarker of the transplanted BM (n=6).
The TH17/IL-17 Immune ResponseMediates Renal CXCL5 Expression inCrescentic GN
The striking temporal association between CXCL5 expression,tubulointerstitial neutrophil infiltration (Figure 1, A–H), andthe recruitment of CD4+ TH17 cells into the kidney at day 10(Figure 3A, Supplemental Figure 5) suggest a functional re-lationship. To determine whether IL-17A drives CXCL5 ex-pression in vivo, we induced nephritis in IL-17A2/2mice. WTanimals showed strong upregulation of CXCL5 mRNA at day10 after nephritis induction, whereas IL-17A2/2 mice dis-played reduced upregulation (WT versus IL-17A2 /2 ,P,0.01) (Figure 3B). By contrast, CXCL1 and CXCL2
Figure 2. CXCL5 is expressed by kidney tubular cells. (A) Representative CXCL5 in situhybridizations of nephritic mice show that CXCL5 expression colocalizes with kidneytubular cells (arrow) at day 14. (B) Kidney sections of nephritic WT and CXCL5b-Gal arestained for b-Gal with specific antibodies (AF488; green), wheat germ agglutinin(TexasRed; red), and nuclear marker draq5 (blue) (arrow, b-Gal–positive cells; asterisk,tubulus lumen). (C) RT-PCR analysis of compartment-specific CXCL1 and CXCL5 ex-pression (laser-microdissected glomeruli versus tubulointerstitium) at 24 hours and 14days after induction of nephritis. Data are presented as the ratio of glomerular totubulointerstitial expression and vice versa (n=3 per group). (D) RT-PCR analysis ofrenal CXCL5 expression in BM chimeric nephritic mice at day 14 (n=4–6 per group).Bars represent means6SDs.
58 Journal of the American Society of Nephrology J Am Soc Nephrol 26: 55–66, 2015
BASIC RESEARCH www.jasn.org

expression was not significantly altered in nephritic IL-17A2/2
mice (Figure 3B).
IL-17A and TNF-a Synergistically Induce CXCL5Expression in Mouse Tubular CellsGiven the high renal expression of CXCL5, predominantly intubular cells (Figure 2, A and B), we investigatedwhether IL-17Adrives CXCL5 expression in kidney tubular cells. Because thebiologic effects of IL-17A are mediated by activation of theIL-17 receptors A and C, we demonstrated the presence ofthese receptors in mouse tubular cells by RT-PCR (Supple-mental Figure 6A). Next, we demonstrated that IL-17A in-duces CXCL5 mRNA and protein production in tubularcells in a dose-dependent manner (Supplemental Figure6B), whereas IL-17F and IFN-g had no effect (SupplementalFigure 6C).
The signal transduction pathway mediated by IL-17A is stillincompletely characterized but involvesNF-kBand extracellular-signal regulated kinase (ERK) activation.15 Therefore, it is ofinterest that the combination of IL-17A and TNF-a synergis-tically amplified the RNA expression and protein secretionof CXCL5 (Figure 3, C and D), and synergistically inducedERK activation but not NF-kB signaling in tubular cells
(Supplemental Figure 6D). Although the invivo importance of this finding remains to beelucidated, the time kinetic of renal IL-17A(Figure 3A) and TNF-a expression in NTN(Supplemental Figure 6E) is consistent withCXCL5 formation in the inflamed kidney.
Tubulointerstitial Infiltration ofCXCR2+ Neutrophils Promotes RenalTissue InjuryTo investigate the functional importance ofthe observed tubulointerstitial neutrophilrecruitment, we depleted neutrophils innephritic mice from days 9 to 14 by using aLy6G-specificmAb16 (Supplemental Figure7).Neutrophil depletion significantly reducedtubulointerstitial tissue injury and to alesser degree glomerular crescent forma-tion, demonstrating the pathogenic role ofneutrophils during the TH17 cell–mediatedphase of the disease (Figure 4, A and B).Levels of BUN, a marker inversely corre-lated with renal function, were slightly butnot significantly reduced (Figure 4B).
Next, we showed that CXCR2-deficientmice developed less severe nephritis, intermsof renal tissuedamageandBUNlevelsat day 14 (Figure 4C). This is in line withreduced neutrophil infiltration in nephriticCXCR22/2 mice (Figure 1D). The recruit-ment of CD3+ T cells and mononuclearphagocytes was not significantly affected
by CXCR2 deficiency (Supplemental Figure 2I). Mouse anti-sheep globulin–specific IgG, subclass IgG1, subclass IgG2a,and IgG2b titers were unaffected in nephritic CXCR22/2
mice compared with theWT group (Supplemental Figure 8F).
Targeting of CXCL5 Reduces Renal Tissue Damage inthe T Cell–Mediated Stage of Crescentic GNTo investigate whether CXCL5 contributes to renal tissueinjury, nephritis was induced in CXCL52/2 and WTmice. Atday 14, CXCL52/2mice showed significantly less tubulointer-stitial and glomerular injury compared with controls (Figure4, D and E). In addition, BUN levels were reduced in nephriticCXCL52/2 mice compared with nephritic WTmice at day 14(P,0.001; Figure 4E). Albuminuria was upregulated in allnephritic groups, but was not significantly different betweennephritic WTand CXCL5 KO mice at day 14 (urine albumin/creatinine ratio: WT control [n=6], 0.160.1; WTNTN [n=8],15.864.7; CXCL52/2 NTN [n=10], 9.768.9).
Treatmentofnephriticmicewith aneutralizinganti-CXCL5antibody at day 9 significantly reduced renal tissue injury at day14 (Figure 4F). The more pronounced protection of nephriticCXCL52/2mice compared with anti-CXCL5–treated animals,in terms of BUN levels, is likely the result of a complete
Figure 3. IL-17A drives renal CXCL5 expression. (A) RT-PCR analysis of renal IL-17Aexpression in the course of nephritis (n=5–6 per time point). RT-PCR analysis of IL-17AmRNA expression in FACS-sorted renal CD4+ T cells isolated from nephritic kidneys atdays 3, 5, 10, and 20. (B) RT-PCR analysis of chemokine mRNA expression in nephriticWT (n=6) and nephritic IL-17A2/2 mice (n=6) at day 10. (C) RT-PCR for CXCL5 ex-pression in murine kidney tubular cells upon stimulation with IL-17A (10 ng/ml), TNF-a(10 ng/ml), and IL-17A+TNF-a for 4 hours. (D) CXCL5-specific ELISA from supernatantof stimulated tubular cells after 24 hours. Bars represent means6SDs. *P,0.05;**P,0.01; ***P,0.001. Con, control.
J Am Soc Nephrol 26: 55–66, 2015 CXCL5 Recruits Neutrophils in TH17 Response 59
www.jasn.org BASIC RESEARCH
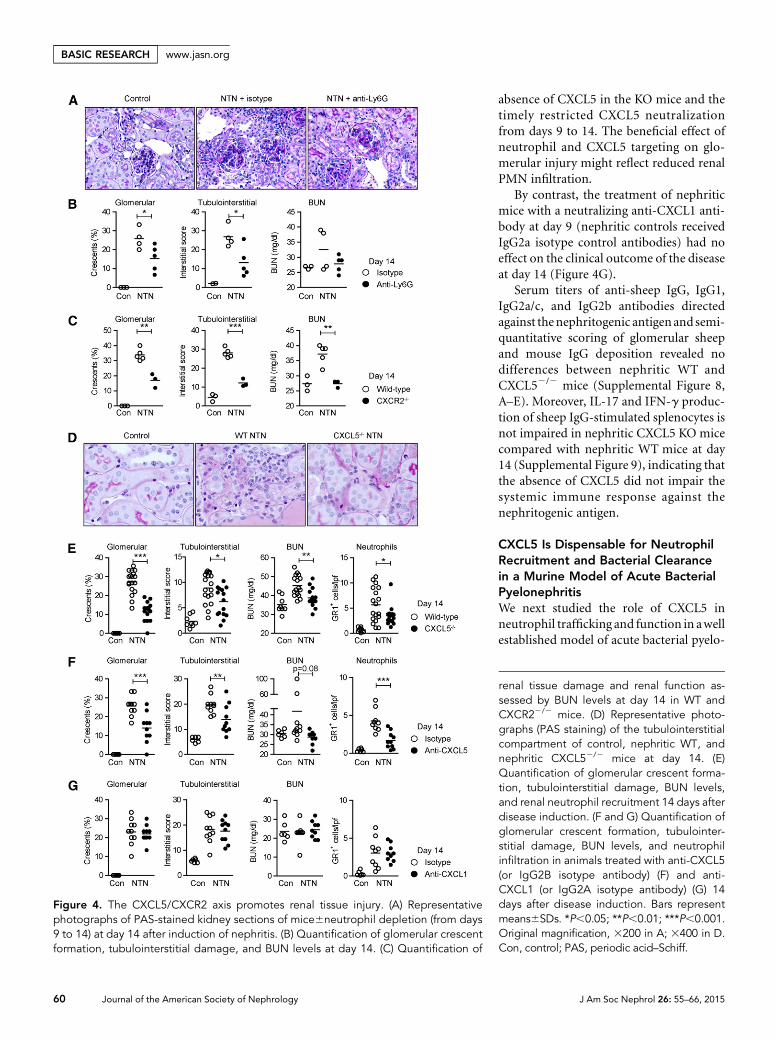
absence of CXCL5 in the KO mice and thetimely restricted CXCL5 neutralizationfrom days 9 to 14. The beneficial effect ofneutrophil and CXCL5 targeting on glo-merular injury might reflect reduced renalPMN infiltration.
By contrast, the treatment of nephriticmice with a neutralizing anti-CXCL1 anti-body at day 9 (nephritic controls receivedIgG2a isotype control antibodies) had noeffect on the clinical outcome of the diseaseat day 14 (Figure 4G).
Serum titers of anti-sheep IgG, IgG1,IgG2a/c, and IgG2b antibodies directedagainst thenephritogenic antigen and semi-quantitative scoring of glomerular sheepand mouse IgG deposition revealed nodifferences between nephritic WT andCXCL52/2 mice (Supplemental Figure 8,A–E). Moreover, IL-17 and IFN-g produc-tion of sheep IgG-stimulated splenocytes isnot impaired in nephritic CXCL5 KO micecompared with nephritic WT mice at day14 (Supplemental Figure 9), indicating thatthe absence of CXCL5 did not impair thesystemic immune response against thenephritogenic antigen.
CXCL5 Is Dispensable for NeutrophilRecruitment and Bacterial Clearancein a Murine Model of Acute BacterialPyelonephritisWe next studied the role of CXCL5 inneutrophil trafficking and function in awellestablished model of acute bacterial pyelo-
Figure 4. The CXCL5/CXCR2 axis promotes renal tissue injury. (A) Representativephotographs of PAS-stained kidney sections of mice6neutrophil depletion (from days9 to 14) at day 14 after induction of nephritis. (B) Quantification of glomerular crescentformation, tubulointerstitial damage, and BUN levels at day 14. (C) Quantification of
renal tissue damage and renal function as-sessed by BUN levels at day 14 in WT andCXCR22/2 mice. (D) Representative photo-graphs (PAS staining) of the tubulointerstitialcompartment of control, nephritic WT, andnephritic CXCL52/2 mice at day 14. (E)Quantification of glomerular crescent forma-tion, tubulointerstitial damage, BUN levels,and renal neutrophil recruitment 14 days afterdisease induction. (F and G) Quantification ofglomerular crescent formation, tubulointer-stitial damage, BUN levels, and neutrophilinfiltration in animals treated with anti-CXCL5(or IgG2B isotype antibody) (F) and anti-CXCL1 (or IgG2A isotype antibody) (G) 14days after disease induction. Bars representmeans6SDs. *P,0.05; **P,0.01; ***P,0.001.Original magnification, 3200 in A; 3400 in D.Con, control; PAS, periodic acid–Schiff.
60 Journal of the American Society of Nephrology J Am Soc Nephrol 26: 55–66, 2015
BASIC RESEARCH www.jasn.org

nephritis. Thismodel is induced by transurethral instillation ofthe uropathogenic Escherichia coli strain 536 (UPEC),17 whichinvades the kidney and is subsequently cleared by recruitedneutrophils. A massive infiltration of neutrophils into the kid-ney (assessed by immunohistochemistry and flow cytometry)
was detected in WT mice 24 hours afterUPEC infection (Figure 5, A and B). Ofnote, neutrophil recruitment was not af-fected by CXCL5 deficiency, whereas neu-trophils were strongly reduced in infectedCXCR2-deficient mice. In line with thepivotal role of neutrophils in bacterialclearance, the numbers of CFUs in the kid-neys were significantly higher in CXCR22/2
mice comparedwithWTandCXCL52/2 an-imals (Figure 5C). Renal expression ofCXCL1, and to a lesser degree of CXCL2,was markedly enhanced, whereas CXCL5was only slightly upregulated 24 hours afterinfection (Figure 5D).
Renal CXCL5 Expression in Patientswith ANCA-Associated GN and AcuteBacterial PyelonephritisWe isolated RNA from renal biopsies andperformed real-time PCR to analyze therenal expression of neutrophil-attractingchemokines in patients with acute ANCA-associated GN, the most common form ofcrescentic GN in humans, and in patientswith acute bacterial pyelonephritis. Al-though neutrophil recruitment is commonto ANCA-associated GN but even strongerin acute pyelonephritis (Figure 6A), upre-gulation of renal tissue CXCL5 expressionwas more profound in ANCA-associatedGN (1013-fold; Figure 6B) comparedwith acute pyelonephritis (16-fold; Figure6B). Similar results were obtained forCXCL6, which is a close homolog to humanCXCL5.6 These results suggest a predomi-nant role of CXCL5 in human autoimmunedisease, but not in acute bacterial infection.Table 1 provides the clinical characteristics ofthe included patients.
DISCUSSION
In recent years, a large number of studieshave established the crucial role of TH17cells in the pathogenesis of autoimmunediseases,7,18–20 including proliferative andcrescentic GN.3,11,21,22 Consequently, thefindings from animal models have been
translated into clinical practice, and IL-17A and related cyto-kines have been successfully targeted for treatment of psoria-sis23 and Crohn’s disease.24
However, the mechanisms by which the TH17 immunitydirectly contributes to end-organ damage still remain to be
Figure 5. CXCL5 is dispensable for neutrophil recruitment and bacterial clearance ina murine model of acute bacterial pyelonephritis. (A) Representative immunohisto-chemistry for the neutrophil marker GR-1 24 hours after transurethral instillation ofUPEC. (B) Quantification of FACS analyses of renal PMNs (CD45+, GR-1HI, F4/802,CD11c2), (numbers represent total cells per kidney; n=5–6 per group). (C) CFU perkidney at 24 hours (n=5–6). (D) RT-PCR analysis of renal CXCL1, CXCL2, and CXCL5expression 24 hours after UPEC infection. Bars/symbols represent means6SDs.*P,0.05; **P,0.01; *** P,0.001. Original magnification, 3400.
Figure 6. CXCL5 expression in patients with ANCA-associated GN and acute bacterialpyelonephritis. (A) Representative PMN staining (proteinase 3) in renal biopsies ofpatients with ANCA-associated GN and acute bacterial pyelonephritis. (B) RT-PCRanalysis of renal chemokine mRNA expression (n=6–8 per group). Bars representmeans6SDs. *P,0.05. Original magnification, 3990.
J Am Soc Nephrol 26: 55–66, 2015 CXCL5 Recruits Neutrophils in TH17 Response 61
www.jasn.org BASIC RESEARCH

fully elucidated. TH17 cells produce a variety of proinflamma-tory cytokines, including IL-17A, IL-17F, IL-22, TNF-a, andGM-CSF, which drive the recruitment of neutrophils to sitesof inflammation and thereby promote tissue injury by mech-anisms that are thus far incompletely understood.7 Recentstudies indicate that ELR+ chemokines play a pivotal role inthis process and might therefore represent an attractive targetfor the prevention of neutrophil-induced tissue injury.25–27
This approach is complicated by an often overlapping/redun-dant expression pattern and function of chemokines in auto-immune and infectious disease.5 Targeting of chemokines inorder to reduce pathogenic neutrophil infiltration in TH17-mediated autoimmunity might therefore impair defenseagainst microbial pathogens.
Recent data by the Chou et al. and McDonald et al. suggestthat neutrophil chemoattractants might have less redundantfunctions in the recruitment of neutrophils in vivo than pre-viously thought.28,29 To investigate the role of the TH17 im-mune response for neutrophil recruitment and function indetail, we performed time-kinetic analyses in a well character-ized TH17-dependent model of crescentic GN (NTN).30 Al-though NTN does not represent a human disease in all aspectsof its pathophysiology (e.g., with respect to immune complex–GN, anti-glomerular basement membrane nephritis, orANCA-associated GN), it remains one of the most efficientmouse model of crescentic GN.31,32 Neutrophils showed atime- and compartment-specific migratory behavior duringcrescentic GN, suggesting that the interstitial “second wave” ofneutrophil infiltration is promoted by renal TH17 cells.
It is generally thought that IL-17A, the major effectorcytokine of the TH17 immune response, induces the expres-sion of all neutrophil-attracting ELR+ chemokines in a redun-dant manner.25,26,33 In murine crescentic GN, however, weunexpectedly found that the expression of CXCL1 andCXCL2 was upregulated until day 3 and then declined even
before infiltration of the first TH17 cells. By striking contrast,CXCL5 expression peaked 10 days after induction of the dis-ease and was predominantly expressed by resident tubularcells when the renal TH17 immune response had reached itsmaximum level. In line with this, IL-17A resulted in a strongupregulation of CXCL5 mRNA and protein expression in mu-rine tubular epithelial cells in vitro, which is further enhancedin the presence of TNF-a, potentially as a consequence ofsynergistic activation of the ERK signaling pathway. This find-ing is in accordance with a recent report by Liu et al. showingthat CXCL5 mRNA is induced by IL-17A in alveolar epithelialcells.26 To further support our hypothesis that IL-17A (pro-duced by renal TH17 cells) is directly involved in the inductionof CXCL5 expression in tubular cells in vivo, the generation ofmice that are deficient in the IL-17 receptors A and C or in theIL-17 receptor adaptor protein connection to IkB kinase andstress-activated protein kinases,34 specifically in renal tubularcells, would be of great interest.
IL-17A promoted renal CXCL5 expression and neutrophilinfiltration in NTN, whereas CXCL1 and CXCL2 expressionwas IL-17A independent. These data further point to non-redundant roles of CXCR2 ligands in the attraction ofneutrophils to the kidney, most likely resulting from thetemporal and spatial segregation of their expression. CXCL1and CXCL5 in vivo neutralization experiments indeed con-firmed that early glomerular neutrophil migration is mainlydriven by CXCL1, whereas the second wave of interstitial neu-trophil infiltration is strongly dependent on IL-17A–inducedCXCL5. Our finding is consistent with a recent study in amodel of moderate airway inflammation in CXCL52/2 miceshowing that neutrophil infiltration into the inflamed tissuewas dependent on CXCL5.27 In contrast with a study by Meiet al. that indicates that enterocyte-derived CXCL5 in the gutregulates local IL-17A levels and contributes toCXCR2-dependentneutrophil homeostasis,35 our study did not show any
Table 1. Clinical characteristics of patients with acute ANCA-associated GN and bacterial pyelonephritis
Patient Age (yr) Sex ANCA TypePR3/MPO
ELISA (U/ml)SerumCreatinine
(mg/dl)Proteinuria(mg/24 h)
Immunosuppressionbefore Biopsy
1 70 Man pANCA MPO 32.6 2.7 5550 No2 67 Man pANCA MPO 17.2 1.6 1130 No3 62 Man pANCA MPO 34 2.5 580 Steroid4 83 Man cANCA PR3 109 4.8 Anuric Steroid + CyP5 47 Man cANCA PR3 26.5 1.6 3000 No6 41 Woman cANCA PR3 .200 1.7 1730 No7 18 Woman cANCA PR3 .200 0.6 430 No8 76 Man cANCA PR3 .200 3.2 760 Steroid9 80 Woman Negative Not assessed 5.5 900 Steroid
10 39 Woman Negative Not assessed 1.2 Not assessed No11 64 Woman Negative Not assessed 4.8 Not assessed No12 71 Man Negative Not assessed 5.0 490 No13 50 Man Negative Not assessed 10.2 .300 No14 50 Man Negative Not assessed 6.8 .3000 No
Clinical characteristics of patients with acute ANCA-associated GN (patients 1–8) and acute pyelonephritis (patients 9–14) at the time of renal biopsy. pANCA,perinuclear ANCA; cANCA, cytoplasmic ANCA; PR3, proteinase 3; MPO, myeloperoxidase; CyP, cypionate.
62 Journal of the American Society of Nephrology J Am Soc Nephrol 26: 55–66, 2015
BASIC RESEARCH www.jasn.org

indication of a role for CXCL5 in the kidney under noninflam-matory conditions. The time-dependent role of CXCL5 in GNis further emphasized by the finding that CXCL52/2 mice andanti-CXCL5–treatedWTanimals displayed an ameliorated dis-ease course at day 14, but not at day 3.
To determine whether the blockade of CXCL5 and CXCR2generally interfereswithneutrophil infiltration,we inducedacutebacterial pyelonephritis in WT, CXCL52/2, and CXCR22/2
mice. In this model, the antibacterial defense relies on the rapidrecruitment of neutrophils. As reported, neutrophil traffickingand bacterial clearance critically depend on CXCR2, which ismost likely activated by locally expressed CXCL1 andCXCL2,17,36–39 making CXCR2 an unfavorable candidate fortherapeutic targeting in autoimmunity. By contrast, CXCL5deficiency did not affect early neutrophil infiltration and bac-terial clearance in this model of acute bacterial pyelonephritis.
A recent study in a murine model of ANCA-associated GNdemonstrated that IL-17A–producing TH17 effector cells di-rectly induce renal inflammation by effector responses involv-ing neutrophils.40 Moreover, the first studies in humanANCA-associated GN suggest a prominent role for TH17/IL-17 responses in this form of crescentic GN.4,41 To examinewhether our findings are relevant to human disease, we as-sessed the renal CXCL5 mRNA expression in patients withacute ANCA-associated GN and acute bacterial pyelonephri-tis, which are both characterized by neutrophil recruitment. Inline with the results from our animal models, CXCL5 expres-sion was much higher in renal biopsy specimens from patientswith autoimmune-mediated ANCA-associated GN comparedwith those with acute bacterial pyelonephritis. Interestingly,van der Veen et al. recently reported an increased renal CXCL5expression in a murine model of anti-myeloperoxidase IgG/LPS-induced crescentic GN, further supporting the role ofCXCL5 in autoimmune-mediated ANCA disease.42 The sim-ilar expression pattern of CXCL1, CXCL2, and CXCL5 duringthe course of the disease (assessed at two time points) mightbe a consequence of the coapplication of LPS in this model.
In conclusion, our study provides the first evidence for apreviously unknownnonredundant effectormechanism of theTH17 immune response in experimental crescentic GN involv-ing T cell–derived IL-17A–, CXCL5–, and CXCR2–bearingneutrophils that promote renal tissue injury. By contrast,CXCL5 seems to be dispensable for maintaining neutrophil-mediated innate immune surveillance. However, the potentialrenal functional benefit of an intervention to target CXCL5 inestablished crescentic GN remains to be fully elucidated.
CONCISE METHODS
AnimalsCXCR22/2 and IFN-g2/2 mice were purchased from The Jackson
Laboratory (Bar Harbor, ME). IL-17A–deficient mice were provided
by Yoichiro Iwakura (University of Tokyo, Tokyo, Japan), CXCL5-
deficient mice were provided by theMax Plank Institute for Infection
Biology (Berlin, Germany)43 and CXCL1-deficient mice were provided
by S. Lira (Mount Sinai Hospital, New York, NY). All KO mice were
backcrossed to the C57BL/6 background for at least 10 generations. All
mice were raised under specific pathogen-free conditions. Animal ex-
perimentswere performed according tonational animal care and ethical
guidelines, and were approved by local ethical committees.
Induction of NTN and Functional StudiesNTNwas induced in male mice aged 8 to 10 weeks by intraperitoneal
injection of 0.6 ml of nephrotoxic sheep serum per mouse, as
previously described.30 Controls were injected intraperitoneally with
an equal amount of nonspecific sheep IgG. Urinary albumin excre-
tion was determined by standard ELISA analysis (Mice-Albumin Kit;
Bethyl, Montgomery, TX), whereas urinary creatinine and serum
BUN were measured using standard laboratory methods. For neu-
tralization/depletion experiments, anti–Ly6G-Ab (clone 1A8),16
anti-CXCL1 Ab (clone 48415; R&D Systems, Minneapolis, MN),14
or anti-CXCL5 Ab (clone 61905; Leinco Technologies, St. Louis,
MO)14 was used at 100 mg per mouse (intraperitoneally). Nephritic
control mice in interventional studies received 100 mg rat IgG2A
(clone 54447; R&D Systems) or rat IgG2B (clone 61905; Leinco Tech-
nologies) antibodies intraperitoneally. Anti-CXCL1 and anti-CXCL5
antibodies were given either the day beforeNTN induction in the case
of early neutralization studies (analysis was performed at day 3) or at
day 9 in the late stages of the interventional experiments (analysis was
performed at day 14).
Acute Bacterial Pyelonephritis ModelFemale mice aged 8 to 12 weeks were infected by transurethral
instillation of 13109 UPEC. Three hours later, the procedure was
repeated to induce pyelonephritis. Twenty-four hours after infection,
the number of ascended bacteria was quantified by scoring CFUs after
overnight culture of kidney collagenase digest as previously de-
scribed.17
Real-Time RT-PCR AnalysesTotal RNA of the renal cortex was prepared according to standard
laboratory methods. RNA from microdissected tissues was isolated
using the RNA Nano prep kit (PALM, Bernried, Germany) as
previously described.44 Real-time PCR was performed on a StepO-
nePlus Real-Time PCR system (Applied Biosystems, Foster City, CA).
In Situ HybridizationIn situ hybridization was performed as described.45 In brief, the
CXCL5 cRNA probe was labeled with a[35S]UTP (1250 Ci/mmol;
PerkinElmer) of subcloned cDNAcorresponding to nucleotides 208–
532 of cDNA sequence NM_009141.2. In situ hybridization was per-
formed on 16-mm cryosections using the 35S-labeled antisense and
sense RNA probes. Sections were dipped into Kodak NTB nuclear
track emulsion and exposed for 3 weeks; after development, sections
were stained with Mayer’s hemalaun.
Morphologic AnalysesGlomerular crescent formation was assessed in 30 glomeruli per
mouse in a blinded fashion in paraffin sections stained with periodic
J Am Soc Nephrol 26: 55–66, 2015 CXCL5 Recruits Neutrophils in TH17 Response 63
www.jasn.org BASIC RESEARCH

acid–Schiff.45 To assess tubulointerstitial injury, photographs of
four nonoverlapping cortical areas from kidney sections stained
with periodic acid–Schiff were taken per mouse and counted in
low-magnification fields (3200). The interstitial area was then de-
termined by superimposing the photographs with a grid containing
40 points and by counting the matches of points with interstitial
tissue (excluding glomeruli, blood vessels, and tubules) in a blinded
fashion. Dividing the positive matches of four photographs by 160
provided the percentage of positive tissue equal to the interstitial
injury score.11,45
For immunohistochemical stainings, paraffin-embedded sec-
tions were deparaffinized, rehydrated, and stained with the follow-
ing antibodies: GR-1 (1:50, Ly6 G/C, NIMP-R14; Hycult Biotech,
Uden, TheNetherlands), CD3 (1:1000, A0452;Dako,Germany), F4/
80 (1:400, BM8; BMA, Germany), MAC-2 (1:1000, M3/38; Cedarlane,
ON, Canada), sheep IgG/mouse IgG (both Jackson Immunoresearch
Laboratories), and proteinase 3 to detect PMN in humans. Antigen
retrieval was performed by incubation with proteinase type XXIV
(5 mg/ml; Sigma-Aldrich, St. Louis, MO) for 15 minutes at 37°C
(F4/80, GR1, sheep IgG/mouse IgG) or by microwave antigen retriev-
al in citrate buffer (DAKO 52367), pH 6.1, for 25 minutes (CD3,
MAC-2). Nuclear b-Gal was revealed after protease-mediated antigen
retrieval using a polyclonal antibody (Europa Bioproducts, Cam-
bridge, UK). Slides were counterstained with wheat germ agglutinin
(Vector, Auckland, New Zealand) and draq 5 (Molecular Probes, Eu-
gene, OR). Tissue sections were developed with the Vectastain ABC-
AP kit (Vector Laboratories, Burlingame, CA). Glomerular GR1+,
MAC-2+, and CD3+cells in 30 glomerular cross-sections and tubu-
lointerstitial F4/80+ and CD3+ cells in 30 high-power fields (3400)
per kidney were counted in a blinded manner. To quantify tubuloin-
terstitial GR1+ cells, at least 20 low-power fields (3200) were counted.
All slides were evaluated under an Axioskop light microscopy (Zeiss,
Jena, Germany) and photographedwith anAxiocamHRc (Zeiss) or by
confocal microscopy with a LSM 510 meta microscope using the LSM
software.
Isolation of Leukocytes from Kidney and SpleenPreviously described methods for leukocyte isolation from murine
kidneys were used.30 In brief, kidneys were finely minced and diges-
ted for 45 minutes at 37°C with 0.4 mg/ml collagenase D (Roche,
Mannheim, Germany). Spleens were minced to isolate splenocytes.
Both cell suspensions were sequentially filtered through 70-mm and
40-mmnylonmeshes and washed with HBSSwithout Ca2+ andMg2+
(Invitrogen). Cell viability was assessed by trypan blue staining be-
fore flow cytometry or cell transfer experiments.
Flow CytometryThe following antibodieswereused forFACSanalysis:CD45 (30-F11),
CD11b (M1/70), CD11c (HL3), Ly6G (1A8), F4/80 (BM8), MHCII
(AF6-120.1), CD3 (506A2), and IL-17A (TC11-18H10) (BD Bio-
sciences, San Jose, CA; eBioscience, San Diego, CA; or R&D Sys-
tems).46 Analyses were performed on a B&D LSRII system using
Diva software and were analyzed using Flowjo software (Treestar,
Ashland, OR).
Culture of Mouse Kidney Epithelial Tubular Cells andStimulationMouse kidney tubular cells47 were cultured in DMEM with 3% FCS
(Gibco, Eggenstein, Germany) and stimulated with varying concen-
trations of IL-17A, IL-17F, TNF-a, and IFN-g, as indicated (all from
Pepro Tech, Hamburg, Germany). CXCL5 mRNA expression levels
were analyzed after 4 hours of incubation. Protein levels were deter-
mined after 8 and 24 hours in the supernatants by specific CXCL5
ELISA (R&D Systems). Antibodies to detect NF-kB p65 (D14E12)
and phospho-NFk-B p65 (93H1) as well as ERK and phospho-ERK
(D13.14.4E) in immunoblots were from Cell Signaling Technology
(Beverly, MA).
Laser MicrodissectionMicrodissection was carried out on mouse kidney tissue using the
PALM MicroBeam IP 230V Z microscope.44 Under direct visual
control, selected tissue compartments (glomeruli or cortical tubu-
lointerstitium) were dissected. RNA from microdissected tissue was
prepared using the PALM RNA extraction kit.
BM TransplantationMale CXCL52/2 or WTmice aged 5 to 6 weeks received 9.5 Gy total
body irradiation. Each recipient mouse (WTor CXCL52/2) was in-
travenously injected with 53106 BM cells (WTor CXCL52/2) within
6 hours of irradiation.48 The efficiency of BM replacement was as-
sessed using congenic CD45 mouse strains (CD45.1 and CD45.2).
After irradiation and transplantation, .90% of circulating leuko-
cytes, .98% of BM leukocytes, and .90% of splenic leukocytes ex-
pressed the phenotypic marker of the transplanted BM (n=6; data not
shown). NTN was induced 4 weeks after BM transplantation.
RNA Isolation and RT-PCR Analysis in Human PatientsRNA was isolated with the RNA Micro Kit (Roche) from paraffin-
embedded renal specimens of patients with ANCA-associated GN or
acute bacterial pyelonephritis. Chemokine expressionwas analyzedby
RT-PCR. Baseline kidney allograft biopsies before transplantation
(without significant pathology) served as controls. All samples were
run in duplicate and were normalized to 18S ribosomal RNA to
account for small RNA and cDNA variability. Human analysis was
approved by the local ethics committee (PV3162).
Statistical AnalysesData were expressed as the means6SDs. All statistical analyses were
performed with the SPSS package and GraphPad Prism 5 software.
P,0.05 was considered statistically significant. The t test was used for
comparison between two groups. In the case of three or more groups,
one-way ANOVA was used, followed by a post hoc analysis with
Bonferroni’s test for multiple comparisons.
ACKNOWLEDGMENTS
This study was supported by grants from the German Research
Foundation (KFO 228: PA 754/7-2 to U.P. and C.F.K.; MI 476/4-2 to
H.-W.M.).
64 Journal of the American Society of Nephrology J Am Soc Nephrol 26: 55–66, 2015
BASIC RESEARCH www.jasn.org

DISCLOSURESNone.
REFERENCES
1. Kolaczkowska E, Kubes P:Neutrophil recruitment and function in healthand inflammation. Nat Rev Immunol 13: 159–175, 2013
2. Eyles JL, Hickey MJ, Norman MU, Croker BA, Roberts AW, Drake SF,James WG, Metcalf D, Campbell IK, Wicks IP: A key role for G-CSF-induced neutrophil production and trafficking during inflammatory ar-thritis. Blood 112: 5193–5201, 2008
3. Summers SA, Steinmetz OM, Li M, Kausman JY, Semple T, Edgtton KL,Borza DB, Braley H, Holdsworth SR, Kitching AR: Th1 and Th17 cellsinduce proliferative glomerulonephritis. J Am Soc Nephrol 20: 2518–2524, 2009
4. Velden J, Paust HJ, Hoxha E, Turner JE, Steinmetz OM, Wolf G, JabsWJ, Özcan F, Beige J, Heering PJ, Schröder S, Kneißler U, Disteldorf E,Mittrücker HW, Stahl RA, Helmchen U, Panzer U: Renal IL-17 expressionin human ANCA-associated glomerulonephritis. Am J Physiol RenalPhysiol 302: F1663–F1673, 2012
5. Sallusto F, Baggiolini M: Chemokines and leukocyte traffic. Nat Im-munol 9: 949–952, 2008
6. Zlotnik A, YoshieO: The chemokine superfamily revisited. Immunity 36:705–716, 2012
7. Korn T, Bettelli E, OukkaM, Kuchroo VK: IL-17 and Th17 cells.Annu RevImmunol 27: 485–517, 2009
8. Fogli LK, Sundrud MS, Goel S, Bajwa S, Jensen K, Derudder E, Sun A,Coffre M, Uyttenhove C, Van Snick J, Schmidt-Supprian M, Rao A,Grunig G, Durbin J, Casola SS, Rajewsky K, Koralov SB: T cell-derivedIL-17 mediates epithelial changes in the airway and drives pulmonaryneutrophilia. J Immunol 191: 3100–3111, 2013
9. Schall TJ, Proudfoot AE: Overcoming hurdles in developing successfuldrugs targeting chemokine receptors. Nat Rev Immunol 11: 355–363,2011
10. Devi S, Li A, Westhorpe CL, Lo CY, Abeynaike LD, Snelgrove SL, Hall P,Ooi JD, Sobey CG, Kitching AR, Hickey MJ: Multiphoton imagingreveals a new leukocyte recruitment paradigm in the glomerulus. NatMed 19: 107–112, 2013
11. Paust HJ, Turner JE, Riedel JH, Disteldorf E, Peters A, Schmidt T, KrebsC, Velden J, Mittrücker HW, Steinmetz OM, Stahl RA, Panzer U: Che-mokines play a critical role in the cross-regulation of Th1 and Th17immune responses in murine crescentic glomerulonephritis. Kidney Int82: 72–83, 2012
12. Odobasic D, Gan PY, Summers SA, Semple TJ, Muljadi RC, Iwakura Y,Kitching AR, Holdsworth SR: Interleukin-17A promotes early but at-tenuates established disease in crescentic glomerulonephritis in mice.Am J Pathol 179: 1188–1198, 2011
13. Heymann F, Meyer-Schwesinger C, Hamilton-Williams EE, HammerichL, Panzer U, Kaden S, Quaggin SE, Floege J, Gröne HJ, Kurts C: Kid-ney dendritic cell activation is required for progression of renal diseasein a mouse model of glomerular injury. J Clin Invest 119: 1286–1297,2009
14. Pickens SR, Chamberlain ND, Volin MV, Gonzalez M, Pope RM,Mandelin AM 2nd, Kolls JK, Shahrara S: Anti-CXCL5 therapy amelio-rates IL-17-induced arthritis by decreasing joint vascularization. An-giogenesis 14: 443–455, 2011
15. Gaffen SL: Recent advances in the IL-17 cytokine family. Curr OpinImmunol 23: 613–619, 2011
16. Daley JM, Thomay AA, Connolly MD, Reichner JS, Albina JE: Use ofLy6G-specific monoclonal antibody to deplete neutrophils in mice. JLeukoc Biol 83: 64–70, 2008
17. Tittel AP, Heuser C, Ohliger C, Knolle PA, Engel DR, Kurts C: Kidneydendritic cells induce innate immunity against bacterial pyelonephritis.J Am Soc Nephrol 22: 1435–1441, 2011
18. Jones SA, Sutton CE, Cua D, Mills KH: Therapeutic potential of tar-geting IL-17. Nat Immunol 13: 1022–1025, 2012
19. Turner JE, Paust HJ, Steinmetz OM, Panzer U: The Th17 immune re-sponse in renal inflammation. Kidney Int 77: 1070–1075, 2010
20. Dong X, Bachman LA, Miller MN, Nath KA, Griffin MD: Dendritic cellsfacilitate accumulation of IL-17 T cells in the kidney following acuterenal obstruction. Kidney Int 74: 1294–1309, 2008
21. Kitching AR, Holdsworth SR: The emergence of TH17 cells as effectorsof renal injury. J Am Soc Nephrol 22: 235–238, 2011
22. Tulone C, Giorgini A, Freeley S, Coughlan A, Robson MG: Transferredantigen-specific T(H)17 but not T(H)1 cells induce crescentic glomer-ulonephritis in mice. Am J Pathol 179: 2683–2690, 2011
23. Papp KA, Leonardi C, Menter A, Ortonne JP, Krueger JG, Kricorian G,Aras G, Li J, Russell CB, Thompson EH, Baumgartner S: Brodalumab, ananti-interleukin-17-receptor antibody for psoriasis. N Engl J Med 366:1181–1189, 2012
24. Sandborn WJ, Gasink C, Gao LL, Blank MA, Johanns J, Guzzo C, SandsBE, Hanauer SB, Targan S, Rutgeerts P, Ghosh S, de Villiers WJ,Panaccione R, Greenberg G, Schreiber S, Lichtiger S, Feagan BG;CERTIFI Study Group: Ustekinumab induction and maintenancetherapy in refractory Crohn’s disease. N Engl J Med 367: 1519–1528,2012
25. Laan M, Cui ZH, Hoshino H, Lötvall J, Sjöstrand M, Gruenert DC,Skoogh BE, Lindén A: Neutrophil recruitment by human IL-17 via C-X-Cchemokine release in the airways. J Immunol 162: 2347–2352, 1999
26. Liu Y, Mei J, Gonzales L, Yang G, Dai N, Wang P, Zhang P, Favara M,Malcolm KC, Guttentag S, Worthen GS: IL-17A and TNF-a exert syn-ergistic effects on expression of CXCL5 by alveolar type II cells in vivoand in vitro. J Immunol 186: 3197–3205, 2011
27. Mei J, Liu Y, Dai N, FavaraM, Greene T, Jeyaseelan S, Poncz M, Lee JS,Worthen GS: CXCL5 regulates chemokine scavenging and pulmonaryhost defense to bacterial infection. Immunity 33: 106–117, 2010
28. Chou RC, Kim ND, Sadik CD, Seung E, Lan Y, Byrne MH, Haribabu B,Iwakura Y, Luster AD: Lipid-cytokine-chemokine cascade drives neu-trophil recruitment in a murine model of inflammatory arthritis. Immu-
nity 33: 266–278, 201029. McDonald B, Pittman K, Menezes GB, Hirota SA, Slaba I, Waterhouse
CC, Beck PL, Muruve DA, Kubes P: Intravascular danger signals guideneutrophils to sites of sterile inflammation. Science 330: 362–366, 2010
30. Paust HJ, Turner JE, Steinmetz OM, Peters A, Heymann F, Hölscher C,Wolf G, Kurts C,Mittrücker HW, Stahl RA, Panzer U: The IL-23/Th17 axiscontributes to renal injury in experimental glomerulonephritis. J Am
Soc Nephrol 20: 969–979, 200931. Couser WG: Basic and translational concepts of immune-mediated
glomerular diseases. J Am Soc Nephrol 23: 381–399, 201232. Kurts C, Panzer U, Anders HJ, Rees AJ: The immune system and kidney
disease: Basic concepts and clinical implications.Nat Rev Immunol 13:738–753, 2013
33. Sadik CD, KimND, Alekseeva E, Luster AD: IL-17RA signaling amplifiesantibody-induced arthritis. PLoS ONE 6: e26342, 2011
34. Pisitkun P, Ha HL, Wang H, Claudio E, Tivy CC, Zhou H, Mayadas TN,Illei GG, Siebenlist U: Interleukin-17 cytokines are critical in de-velopment of fatal lupus glomerulonephritis. Immunity 37: 1104–1115,2012
35. Mei J, Liu Y, Dai N, Hoffmann C, Hudock KM, Zhang P, Guttentag SH,Kolls JK, Oliver PM, Bushman FD, Worthen GS: Cxcr2 and Cxcl5 reg-ulate the IL-17/G-CSF axis and neutrophil homeostasis in mice. J Clin
Invest 122: 974–986, 201236. Hang L, Haraoka M, Agace WW, Leffler H, Burdick M, Strieter R,
Svanborg C: Macrophage inflammatory protein-2 is required for neu-trophil passage across the epithelial barrier of the infected urinary tract.J Immunol 162: 3037–3044, 1999
37. Svensson M, Irjala H, Svanborg C, Godaly G: Effects of epithelial andneutrophil CXCR2 on innate immunity and resistance to kidney in-fection. Kidney Int 74: 81–90, 2008
J Am Soc Nephrol 26: 55–66, 2015 CXCL5 Recruits Neutrophils in TH17 Response 65
www.jasn.org BASIC RESEARCH

38. Svensson M, Yadav M, Holmqvist B, Lutay N, Svanborg C, Godaly G:Acute pyelonephritis and renal scarring are caused by dysfunctionalinnate immunity in mCxcr2 heterozygous mice. Kidney Int 80: 1064–1072, 2011
39. Frendéus B, GodalyG, Hang L, KarpmanD, Lundstedt AC, Svanborg C:Interleukin 8 receptor deficiency confers susceptibility to acute ex-perimental pyelonephritis and may have a human counterpart. J ExpMed 192: 881–890, 2000
40. Gan PY, Steinmetz OM, Tan DS, O’Sullivan KM, Ooi JD, Iwakura Y,Kitching AR, Holdsworth SR: Th17 cells promote autoimmune anti-myeloperoxidase glomerulonephritis. J Am Soc Nephrol 21: 925–931,2010
41. van Timmeren MM, Heeringa P: Pathogenesis of ANCA-associatedvasculitis: Recent insights from animal models. Curr Opin Rheumatol24: 8–14, 2012
42. van der VeenBS, PetersenAH, Belperio JA, Satchell SC,Mathieson PW,Molema G, Heeringa P: Spatiotemporal expression of chemokines andchemokine receptors in experimental anti-myeloperoxidase antibody-mediated glomerulonephritis. Clin Exp Immunol 158: 143–153, 2009
43. Nouailles G, Dorhoi A, Koch M, Zerrahn J, Weiner J 3rd, Faé KC, ArreyF, Kuhlmann S, Bandermann S, Loewe D, Mollenkopf HJ, Vogelzang A,Meyer-Schwesinger C, Mittrücker HW, McEwen G, Kaufmann SH:CXCL5-secreting pulmonary epithelial cells drive destructive neutro-philic inflammation in tuberculosis. J Clin Invest 124: 1268–1282, 2014
44. Panzer U, Steinmetz OM, Reinking RR, Meyer TN, Fehr S, Schneider A,Zahner G, Wolf G, Helmchen U, Schaerli P, Stahl RA, Thaiss F:
Compartment-specific expression and function of the chemokine IP-10/CXCL10 in a model of renal endothelial microvascular injury. J AmSoc Nephrol 17: 454–464, 2006
45. Riedel JH, Paust HJ, Turner JE, Tittel AP, Krebs C, Disteldorf E,Wegscheid C, Tiegs G, Velden J, Mittrücker HW, Garbi N, Stahl RA,Steinmetz OM, Kurts C, Panzer U: Immature renal dendritic cells recruitregulatory CXCR6(+) invariant natural killer T cells to attenuate cres-centic GN. J Am Soc Nephrol 23: 1987–2000, 2012
46. Turner JE, Krebs C, Tittel AP, Paust HJ, Meyer-Schwesinger C,Bennstein SB, Steinmetz OM, Prinz I, Magnus T, Korn T, Stahl RA,Kurts C, Panzer U: IL-17A production by renal gd T cells promoteskidney injury in crescentic GN. J Am Soc Nephrol 23: 1486–1495,2012
47. Wolf G, Mueller E, Stahl RA, Ziyadeh FN: Angiotensin II-induced hy-pertrophy of cultured murine proximal tubular cells is mediated byendogenous transforming growth factor-beta. J Clin Invest 92: 1366–1372, 1993
48. Kluger MA, Zahner G, Paust HJ, Schaper M, Magnus T, Panzer U, StahlRA: Leukocyte-derived MMP9 is crucial for the recruitment of proin-flammatory macrophages in experimental glomerulonephritis. KidneyInt 83: 865–877, 2013
This article contains supplemental material online at http://jasn.asnjournals.org/lookup/suppl/doi:10.1681/ASN.2013101061/-/DCSupplemental.
66 Journal of the American Society of Nephrology J Am Soc Nephrol 26: 55–66, 2015
BASIC RESEARCH www.jasn.org




















