
Dynamic regulatory network controlling TH17 cell differentiation
Upload
independentCategory
view
0download
0
1 23
In Vitro Cellular & DevelopmentalBiology - Animal ISSN 1071-2690Volume 47Number 8 In Vitro Cell.Dev.Biol.-Animal (2011)47:581-592DOI 10.1007/s11626-011-9444-1
Optimization of human Th17 celldifferentiation in vitro: evaluatingdifferent polarizing factors
Mazdak Ganjalikhani Hakemi, KamranGhaedi, Alireza Andalib, Mohsen Hosseini& Abbas Rezaei

1 23
Your article is protected by copyright and all
rights are held exclusively by The Society for
In Vitro Biology. This e-offprint is for personal
use only and shall not be self-archived in
electronic repositories. If you wish to self-
archive your work, please use the accepted
author’s version for posting to your own
website or your institution’s repository. You
may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

Optimization of human Th17 cell differentiation in vitro:evaluating different polarizing factors
Mazdak Ganjalikhani Hakemi & Kamran Ghaedi &Alireza Andalib & Mohsen Hosseini & Abbas Rezaei
# The Society for In Vitro Biology 2011
Abstract Regarding discrepancies that exist among differ-ent studies which have tried to clarify critical factors inhuman Th17 cell differentiation, the aim of this study wasto identify the best condition for human Th17 differentia-tion and to clarify the possible role of TGF-β indifferentiation of these cells. Naïve CD4+ T cells wereisolated from cord blood samples and cultured either in X-VIVO 15 serum-free medium or RPMI 1640 containing10% FBS. Purified cells were treated with differentcombinations of polarizing cytokines (TGF-β, IL-1β, IL-6, IL-23 and IL-21) followed by analysis of the expressionof characteristic genes and their relevant cytokines by real-time quantitative RT-PCR and ELISA method, respectively.Our data indicate that a combination of TGF-β plus IL-6and IL-23 cytokines in X-VIVO 15 serum-free mediumcould be applied as the best condition for developinghuman Th17 cells in compare with other studied cytokine
treatments. It is shown that TGF-β could be considered as apositive regulator for human Th17 cell differentiation onlyif applied in average concentrations. Interestingly, polariz-ing treatments in absence of TGF-β, induced double-secreting Th17 cells which co-express IL-17 and IFN-γwhereas polarization in presence of TGF-β-induced single-secreting (only IL-17 expressing) Th17 cells.
Keywords IL-17 . RORC2 . TGF-β . Th17
Introduction
Classically, helper T cells had been grouped simply intoTh1 and Th2 cells due to their cytokines expression profile.More recently, IL-17 producing helper T cell (Th17) isidentified as a new subtype different from Th1 and Th2(Volpe et al. 2008; Cardoso et al. 2009). Beyond their rolesin host defense, Th17 cells are known to be associated withseveral inflammatory and autoimmune diseases, the rolethat was previously assigned to Th1 and IFN-γ (Acosta-Rodriguez et al. 2007a, 2007b; Wilson et al. 2007; Streecket al. 2008). IL-17 producing T cells induce production ofchemokines and antimicrobial peptides by tissue cells andcauses the recruitment of neutrophils and induction ofinflammation (Aggarwal et al. 2003; Park et al. 2005;Ivanov et al. 2006; Acosta-Rodriguez et al. 2007a, b;Ghilardi and Ouyang 2007; Streeck et al. 2008; Zhu andPaul 2008). Several studies in human and mouse modelshave shown an association between severity in inflamma-tory diseases and IL-17 concentrations in serum and tissuefluids (Harrington et al. 2005; Acosta-Rodriguez et al.2007a, b; McGeachy and Cua 2008; Ouyang et al. 2008;Yang et al. 2008a).Thus, identification of the factorsresponsible for human Th17 cells differentiation is of
M. Ganjalikhani Hakemi :A. Andalib :A. Rezaei (*)Immunology Dept., Faculty of Medicine,Isfahan University of Medical Sciences,Isfahan, Irane-mail: [email protected]
M. Ganjalikhani Hakemi :A. RezaeiApplied Physiology Research Center, Faculty of Medicine,Isfahan University of Medical Sciences,Isfahan, Iran
K. GhaediBiology Dept., Faculty of Sciences, University of Isfahan,Isfahan, Iran
M. HosseiniEpidemiology Dept., Faculty of Health,Isfahan University of Medical Sciences,Isfahan, Iran
In Vitro Cell.Dev.Biol.—Animal (2011) 47:581–592DOI 10.1007/s11626-011-9444-1
Received: 21 May 2011 /Accepted: 11 July 2011 /Published online: 19 August 2011 / Editor: T. Okamoto
Author's personal copy

special interest due to importance of these cells inphysiologic and pathologic aspects in human.
Since discovery of Th17 cells, increasing efforts havebeen made to characterize their properties and to identifythe factors affecting Th17 cells differentiation and functions(Acosta-Rodriguez et al. 2007a, b). The most data aboutthese cells are derived from experimental mouse modelsand in comparison, there is little knowledge about essentialfactors in differentiation pathways of human Th17 cells(Streeck et al. 2008). At first, IL-23 was thought to becritical in Th17 differentiation. However, subsequentstudies have demonstrated that IL-23 is more importantfor effector functions and survival of Th17, not fordifferentiation toward Th17cells (Ivanov et al. 2006; Gockeet al. 2007; Laurence et al. 2007). Studies on mouse modelshave clarified that combination of TGF-β and IL-6 issuitable for differentiation of mouse naïve CD4+ T cells toTh17 cells (Bettelli et al. 2006; Mangan et al. 2006;Veldhoen et al. 2006; McGeachy and Cua 2008; Yang et al.2008a; Zhu and Paul 2008). Conversely, these twocytokines are not able to drive differentiation to Th17 cellsin human, and it has even been suggested that TGF-β mayconstrain human Th17 development (Acosta-Rodriguez etal. 2007a, b; Wilson et al. 2007). Nevertheless, thecontroversy of whether TGF-β has any role to play inhuman Th17 cell development has not yet been fully laid torest (McGeachy and Cua 2008).
During the past 3 years, a number of independent invitro studies have reported distinct polarizing conditionsrequired for human Th17 cells development. These resultsare somewhat controversial as some of these studiesproposed different combinations of inflammatory cytokinessuch as IL-1β, IL-6, IL-21, and IL-23 to promotedifferentiation of human Th17 lymphocytes in short-termhuman CD4+ Tcell culture, while they found that TGF-β isnot appropriate for this purpose and actually suppressesTh17 development (Acosta-Rodriguez et al. 2007a, b;Wilson et al. 2007). In contrast, some other reports haveshown that TGF-β is critical for human Th17 celldifferentiation and controversy is attributed to the cytokinedosage and the type of culture media used (Volpe et al.2008; Yang et al. 2008a).
Thus, the purpose of the present study was to comparedifferent cytokine combinations to find the most effectivetreatments for development of human Th17 cells in vitroand to clarify the probable role of TGF-β in this process.There are some controversies regarding to the effect ofparticular cytokines on Th17 cells differentiation which arepresumably due to the type of culture media and/ordifferent cytokine concentrations. Hence, we decided toevaluate the effect of various polarizing cytokine combina-tions in two concentrations (dose I: low and dose II: high)and two types of culture media (X-VIVO 15 serum-free and
RPMI 1640 plus 10% fetal bovine serum (FBS)) whichhave been most frequently but separately utilized in otherstudies so far.
Material and Methods
Purification of naïve CD4+ Tcells. Mononuclear cells wereseparated from 100-ml cord blood sample using of Ficoll–Hypaque density gradient (biosera, East Sussex, UK).Naïve CD4+ T cells were isolated using human naïveCD4+ T-cell isolation kit II (Miltenyi Biotec, BergischGladbach, Germany) according to manufacturer’s instructionbriefly as follows: CD45RO+ activated/memory T cells andnon-CD4+ T cells were magnetically labeled applying acocktail of biotin-conjugated antibodies against CD8, CD14,CD15, CD16, CD19, CD25, CD34, CD36, CD45RO, CD56,CD123, TCRγ/δ, HLA-DR, CD235a (Glycophorine A), andanti-biotin micro-beads. Then, highly pure naive CD4+ Tcellswere isolated following depletion of magnetically labeledcells (Acosta-Rodriguez et al. 2007a, b; Naive CD4+ cellisolation kit II human 2008; Volpe et al. 2008; Yang et al.2008a, b). Isolation of highly pure naive CD4+ T cells wasconfirmed after immune staining with FITC-conjugated anti-CD4 and PE-conjugated anti-CD45RA antibodies (BDBiosciences, Franklin Lakes, NJ) and analysis with FACSCa-libur (Becton Dickinson) instrument.
Cell culture and differentiation assay. Each well of 24-wellplates (Orange, Alleud, Belgium) was treated by 200 μlPBS including 5 μg/ml anti-CD3 antibody and 2 μg/mlanti-CD28 antibody (eBioscience, San Diego, CA) andincubated in 4°C over night. Naïve CD4+ T cells were thencultured in these plates at a density of 4–5×105 cells perwell either in X-VIVO 15 serum-free medium (Lonza,Basel, Switzerland) or RPMI 1640 containing 10% (v/v)FBS (Gibco, Billings, MT) in presence of followingcytokine treatments in two doses. Dose I: IL-6 (15 ng/ml),TGF-β (5 ng/ml), IL-23 (25 ng/ml), IL-1β (25 ng/ml), IL-21 (25 ng/ml; eBioscience). Dose II: IL-6 (30 ng/ml), TGF-β (10 ng/ml), IL-23 (100 ng/ml), IL-1β (100 ng/ml), IL-21(100 ng/ml; eBioscience); 10 μg/ml anti-IFN-γ and 10μg/ml anti-IL-4 (eBioscience) per well were added to inhibitunwanted deviation to Th1 or Th2 subsets. Culture mediahad been refreshed after 3 d. After 6 d, cells were washedand their viability was checked by trypan blue exclusion. Inorder to analysis of cytokine production and transcriptlevels, cells were re-stimulated with anti-CD3 (1 μg/ml) foranother 24 h (Vyakarnam 2000; Acosta-Rodriguez et al.2007a, b; Volpe et al. 2008; Yang et al. 2008a).
Analysis of cytokine production. Cytokine contents ofsupernatant culture media were measured by ELISA kits
582 HAKEMI ET AL.
Author's personal copy

for IL-17 (RayBiotech, Norcross, GA) and for IL-22 andIFN-γ (BenderMed Systems, Vienna, Austria) according tothe manufacturer’s instruction. Results were read by micro-plate reader (ELX 800) at 450 nm.
Real time quantitative RT-PCR. Total RNA from culturedCD4+ Tcells was extracted using RNeasy mini kit (Qiagen,Venlo, The Netherlands) and cDNA was synthesized byQuantiTect reverse transcription kit (Qiagen) as instructedby manufacturer. The resulting transcripts were thenquantified by real-time quantitative PCR on a Rotor-Gene6000 real-time DNA amplification system (Corbett lifescience) with QuantiFast SYBR Green PCR kit (Qiagen).Pre-designed primers (QuantiTact primer Assay; Qiagen)specific for IL-17, IL-22, IL-23R, RORC2, and IFN-γ havebeen used. For each sample, transcript quantity wasnormalized to the amount of beta-actin (ACTB) expression.The obtained results have been analyzed by relativequantification method (Kenneth and Thomas 2001; Michael2001).
Statistical analysis. Independent sample t test was used forpair-wise comparisons and one-way ANOVA was utilizedfor comparison among treated groups. The partial correla-tion test was selected for evaluation of significance of theprobable correlation among studied markers of Th17 cells.P values equal to or less than 0.05 were consideredstatistically significant. All experiments carried out intriplicate and in order to summarizing the data, replicateshave been aggregated and their mean and standarddeviation (SD) values have been shown in graphs usinglogarithmic scaling.
Results
Highly pure naïve CD4+ T cells were isolated from cordblood samples and more than 95% purity was confirmedvia flow cytometric analysis (Fig. 1).
The effects of two doses for five different cytokinecombinations were evaluated on differentiation of the naïveCD4+ T cells, cultured in two types of media. For thispurpose, transcript levels of IL-17, IL-22, IL-23R, RORC2,and IFN-γ genes were measured by qRT-PCR and thecytokine levels of IL-17, IL-22, and IFN-γ were alsomeasured using ELISA method.
Treatment 1: TGF-β plus IL-6. Applying dose I of TGF-βplus IL-6 did not induce any increase in RORC2, IL-17and IL-23R transcript levels in CD4+ T cells cultured inany of studied media. However, using dose II of thistreatment for T cells cultured in X-VIVO 15 serum-free
medium was shown to be significantly inducible (p<0.05)for RORC2 and IL-23R genes (Fig. 2A). T cells cultured inX-VIVO 15 medium showed an increase in transcript andprotein levels of IL-17 following treatment with dose II;however, this increase was not significant. Following thistreatment, no increase in IL-22 and IFN-γ production wasfound (Fig. 2A, B).
Treatment 2: TGF-β plus IL-6 and IL-23. The mostincrease in RORC2 transcript level was seen followingusage of this treatment for T cells cultured in X-VIVO 15medium (p<0.05). Interestingly, the dose II of the samecytokine combination in RPMI 1640 with 10% FBS had asuppressive effect on this gene (Fig. 3A).The effect of thiscombination on IL-17 and IL-23R expression was signifi-cant (p<0.05) on T cells seeded in X-VIVO 15 serum-freemedium. In presence of RPMI 1640, only dose II waseffective on these markers (p<0.05). Utilizing the dose IIled to significant (p<0.05) increase of IL-22 cytokine in Tcells cultured in X-VIVO 15. However, this treatment wasnot inducible for IFN-γ expression (Fig. 3A, B).
Treatment 3: IL-1β plus IL-6. Although some degree ofincrease appeared in RORC2 mRNA level in every studiedsituation, the only significant increase (p<0.05) resultedfollowing use of dose II of the treatment in X-VIVO 15medium (Fig. 4A). Applying this treatment in all studiedconditions was strongly inducible for IL-17, IFN-γ, and IL-23R expression (p<0.05). In contrast to X-VIVO 15,applying dose II led decrease of IL-17, IFN-γ, and IL-23R in presence of RPMI 1640 (Fig. 4A, B). Combinationof IL-1β plus IL-6 was strongly inducible (p<0.05) for IL-22 in T cells cultured in X-VIVO 15 serum-free medium.This effect was more significant (p<0.05) following use ofdose I (Fig. 4A, B).
Treatment 4: IL-1β plus IL-23. This treatment was shownto be significantly inducible for RORC2 gene expressiononly for T cells cultured in X-VIVO 15 serum free mediaand following use of dose II (p<0.05) (Fig. 5A). Althoughthe effect of treatment with IL-1β plus IL-23 was highlyinducible for IL-17 production in presence of RPMI 1640containing 10% serum (p<0.05), this effect was higherwhile X-VIVO 15 serum-free medium had been applied.Applying the dose II of this treatment led to the samepattern of increase in expression of aforementioned genebut significantly (p<0.05) in lower scale (Fig. 5A, B).Applying dose I for inducing IL-23R gene expression led tosignificant increase especially in presence of RPMI 1640with 10% FBS (p<0.05). Using the dose II, the effect of X-VIVO 15 medium on this expression was strongly higherthan other conditions (p<0.05) (Fig. 5A).Using of IL-1βplus IL-23 with dose I showed a significant inducing effect
OPTIMIZATION OF HUMAN TH17 CELL DIFFERENTIATION 583
Author's personal copy

(p<0.05) on IL-22 transcript and protein levels in T cellshad been cultured in X-VIVO 15 serum-free medium. Thiseffect was more potent (p<0.05) when dose II had beenused (Fig. 5A, B). Evaluation of IFN-γ production and itsgene expression revealed that especially in X-VIVO 15serum free media, dose I of this treatment was the mostinducible condition for IFN-γ expression (p<0.05). In-creased concentration (dose II) of the treatment resulted inconsiderable reduction (p<0.05) in IFN-γ expression level(Fig. 5A, B).
Treatment 5: TGF-β plus IL-21. Combination of TGF-βplus IL-21 had a significant (p<0.05) positive effect onRORC2 and IL-17 expression, only in Tcells cultured in X-VIVO 15 medium. This effect has not changed by alteringthe dose of treatment. The same treatment was notsignificantly effective for IL-17 expression for T cellscultured in RPMI 1640 containing 10% FBS whereas inthis medium, it had a negative effect on RORC2 geneexpression (Fig. 6A, B). This treatment, especially with thedose I, had a strong inducible effect on IL-23R gene whenapplied with X-VIVO 15 serum-free medium (p<0.05).This effect was greatly diminished (p<0.05) when RPMI1640 with 10% FBS had been used; however, it was stillsignificant (p<0.05) (Fig. 6A). In comparison with control,this treatment was not able to increase IL-22 geneexpression and its cytokine production (Fig. 6A, B).Treatment of T cells cultured in X-VIVO 15 with the doseI of these cytokines had a significant effect on IFN-γgeneexpression (p<0.05). However, increase in protein level
was not significant. Dose II of this treatment was absolutelyineffective on this gene (Fig. 6A, B).
Statistical correlations among expression levels of studiedmarkers. Among the studied markers, some were expressedin correlation with each other while no or even negativecorrelation among the others has been found (Table 1).
Discussion
Since discovery of Th17 cells, increasing efforts have beendone to clarify more aspects of these cells and criticalfactors involving in regulation of their differentiationprocess (Acosta-Rodriguez et al. 2007a, 2007b). The mostof our knowledge is derived from mouse models whilethere are controversial issues about human Th17 cellsdifferentiation and functional aspects (Streeck et al. 2008).
To address this issue, current study was designed toexamine different combinations of polarizing cytokineswith different concentrations in two types of culture media(with and without serum) and compare their effects onhuman Th17 cell differentiation. Our data revealed that themost polarizing condition for human Th17 cells develop-ment from naïve CD4+ Tcells was treatment with dose II ofTGF-β plus IL-6 and IL-23 (10, 30, and 100 ng/ml,respectively), in X-VIVO 15 serum-free culture medium.Interestingly, applying the same condition in RPMI1640supplied with 10% FBS did not induce IL-17, RORC2, and
Figure 1. Flow cytometric analysis of naïve CD4 T cells populationisolated from cord blood samples. A, Dot plot diagram shows threedistinct cell populations exist in cord blood mononuclear cellsseparated via ficoll hypaque density gradient, before applying human
naïve CD4+ T cell isolation kit. B, Dot plot diagram shows more than95% purity of CD4+/CD45RA+ T cells in cell samples obtained fromhuman naïve CD4+ T cell isolation column.
584 HAKEMI ET AL.
Author's personal copy

IL-22 gene expression. Indeed, this situation was ratherinhibitory for Th17 cell differentiation. Applying lowconcentrations of same treatment in X-VIVO 15 mediumfailed to induce IL-22 expression, unlike its significantinducible effect on IL-17, IL-23R, and RORC2. Our dataare in consistent with several studies reported that a
combination of TGF-β and IL-23 with inflammatorycytokines such as IL-6 and IL-1β in serum-free mediumis effective to induce human Th17 cells differentiation(Veldhoen et al. 2006; Volpe et al. 2008; Mus et al. 2010;Hirota et al. 2010; Valmori et al. 2010). Nevertheless,Wilson et al. reported that using TGF-β plus IL-6 neither in
Figure 2. Expression analysisof Th17 cell characteristicmarkers after treatment withTGF-β plus IL-6. A, qRT-PCRanalysis of mRNA levels ofselected genes in human naïveCD4+ T cells after 6 d incuba-tion in either in X-VIVO 15 orRPMI 1640 media. Data areshown in logarithmic scale andare the mean and S.D. of threesimilar experiments. Dose II ofthis treatment in X-VIVO 15medium was more efficient forIL-17 expression (a, p<0.05)even though, it is not significant.Using the dose II of this treat-ment for T cells cultured in X-VIVO 15 serum-free mediumwas inducible for RORC2 andIL-23R gene expression(*a, p<0.05). Culture in X-VIVO wassignificantly different fromRPMI 1640 (¤, p<0.05). B,Measurement of protein levelsof selected cytokines usingELISA method. Data are themean and S.D. of three similarexperiments and are shown inlogarithmic scale. The produc-tion patterns of IL-17, IL-22,and IFN-γ cytokine were corre-lated with their mRNA levels.
OPTIMIZATION OF HUMAN TH17 CELL DIFFERENTIATION 585
Author's personal copy
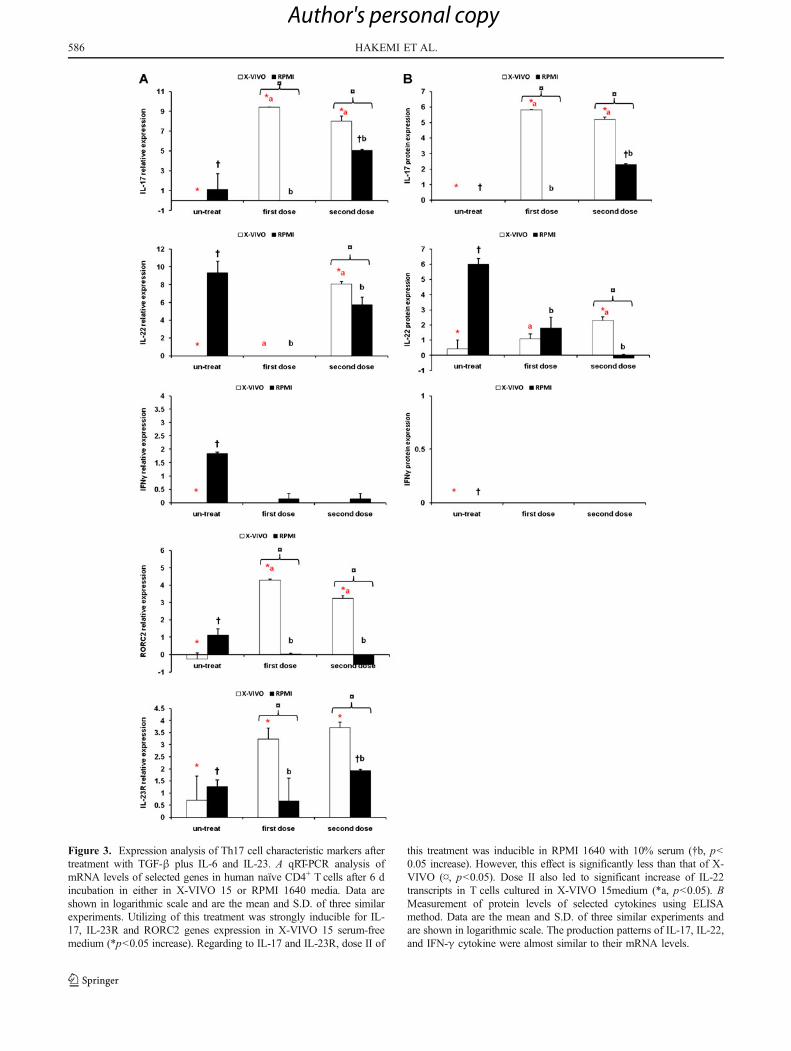
Figure 3. Expression analysis of Th17 cell characteristic markers aftertreatment with TGF-β plus IL-6 and IL-23. A qRT-PCR analysis ofmRNA levels of selected genes in human naïve CD4+ T cells after 6 dincubation in either in X-VIVO 15 or RPMI 1640 media. Data areshown in logarithmic scale and are the mean and S.D. of three similarexperiments. Utilizing of this treatment was strongly inducible for IL-17, IL-23R and RORC2 genes expression in X-VIVO 15 serum-freemedium (*p<0.05 increase). Regarding to IL-17 and IL-23R, dose II of
this treatment was inducible in RPMI 1640 with 10% serum (†b, p<0.05 increase). However, this effect is significantly less than that of X-VIVO (¤, p<0.05). Dose II also led to significant increase of IL-22transcripts in T cells cultured in X-VIVO 15medium (*a, p<0.05). BMeasurement of protein levels of selected cytokines using ELISAmethod. Data are the mean and S.D. of three similar experiments andare shown in logarithmic scale. The production patterns of IL-17, IL-22,and IFN-γ cytokine were almost similar to their mRNA levels.
58 HAKEMI ET AL.6
Author's personal copy

Figure 4. Expression analysis of Th17 cell characteristic markersafter treatment with IL-1β plus IL-6. A, qRT-PCR analysis of mRNAlevels of selected genes in human naïve CD4+ T cells after 6 dincubation in either in X-VIVO 15 or RPMI 1640 media. Data areshown in logarithmic scale and are the mean and S.D. of three similarexperiments. The efficiency of dose I in both media used was nearlysimilar and showed a significant inducible effect on expression of IL-17 gene (* and †, p<0.05 increase). However, efficiency of dose IIwas considerably more in X-VIVO 15 medium (¤, p<0.05) incompare with RPMI 1640. Increase of RORC2 gene expression was
significant only after treatment with dose II in X-VIVO 15 medium(*a, p<0.05). This treatment was significantly effective on expressionof IL-23R and IL-22 genes only in X-VIVO 15 medium (*, p<0.05increase). Significant increase of IFN-γ transcripts in T cells culturedin X-VIVO (*, p<0.05) and RPMI 1640(†, p<0.05) media resultedfollowing this treatment. B, Measurement of protein levels of studiedcytokines using ELISA method. Data are the mean and S.D. of threesimilar experiments and are shown in logarithmic scale. Theproduction patterns of IL-17, IL-22 and IFN-γ cytokine were almostsimilar to their mRNA levels.
OPTIMIZATION OF HUMAN TH17 CELL DIFFERENTIATION 587
Author's personal copy

combination nor with IL-1β plus IL-23 is able to induceTh17 and is rather inhibitory (Wilson et al. 2007). Weshowed similar inhibitory role for TGF-β by adding 10%FBS to culture medium as well while TGF-β was effectivewhen applied with serum-free media. In study done byWilson et al., such a condition has not been evaluated.
A number of reports indicate that TGF-β is not a polarizingfactor for human Th17 cells and it even concerned as an
inhibitor for this process (Acosta-Rodriguez et al. 2007a, b;Wilson et al. 2007). It is reported that implementation ofTGF-β in absence of inflammatory cytokines promotesinducible Treg (iTreg) differentiation, presumably due toinduction of Foxp3 which antagonizes RORγt functionrequired for Th17 cell development (Martin-Orozco et al.2009). Accordingly, many reports have recently shown theimportance of TGF-β, not alone but accompanied with
Figure 5. Expression analysisof Th17 cell characteristicmarkers after treatment with IL-1β plus IL-23. A, qRT-PCRanalysis of mRNA levels ofselected genes in human naïveCD4+ T cells after 6 d incuba-tion in either in X-VIVO 15 orRPMI 1640 media. Data areshown in logarithmic scale andare the mean and S.D. of threesimilar experiments. Dose II ofthis treatment was shown to besignificantly inducible forRORC2 gene expression, just inT cells cultured in X-VIVO 15serum-free medium (*a, p<0.05). Although the effect oftreatment with IL-1β plus IL-23was highly inducible for IL-17production, this effect washigher with dose I of this treat-ment, either in X-VIVO (*a, p<0.05) or in RPMI 1640 (†b, p<0.05). Dose I of this treatmentwas stronger to induce IL-23Rgene expression in RPMI 1640(¤, p<0.05); however, by usingthe dose II, the effect wasstronger in T cells cultured in X-VIVO 15 medium (¤, p<0.05).Increase in expression of IL-22gene has resulted in T cellscultured in X-VIVO 15 serum-free medium especially follow-ing use of dose II (*a, p<0.05).The effect of dose I on IFN-γexpression was significantlymore, either in X-VIVO (*a, p<0.05) or in RPMI 1640 (†b, p<0.05). B, Measurement of pro-tein levels of studied cytokinesusing ELISA method. Data arethe mean and S.D. of threesimilar experiments and areshown in logarithmic scale. Theproduction patterns of IL-17, IL-22, and IFN-γ cytokine werecorrelated with their mRNAlevels.
58 HAKEMI ET AL.8
Author's personal copy

inflammatory cytokines, in Th17 differentiation as well(Bettelli et al. 2006; Mangan et al. 2006; Veldhoen et al.2006; Volpe et al. 2008; Burgler et al. 2009; Hirota et al.2010; Mus et al. 2010; Valmori et al. 2010). Indeed, morerecent data utilizing serum-free media and naïve T cellsisolated from cord blood have shown that TGF-β is requiredfor the differentiation of human Th17 cells (Manel et al.
2008; Volpe et al. 2008; Yang et al. 2008a; Hirota et al. 2010)as verified by our data.
These discrepancies among different studies could bealso reflected by the amount of TGF-β and/or presence ofserum in culture media (Zhu and Paul 2008). This may beaddressed by the fact that TGF-β may exists in serumapplied in medium which significantly elevates the amount
Figure 6. Expression analysisof Th17 cell characteristicmarkers after treatment withTGF-β plus IL-21. A, qRT-PCRanalysis of mRNA levels ofselected genes in human naïveCD4+ T cells after 6 d incuba-tion in either in X-VIVO 15 orRPMI 1640 media. Data areshown in logarithmic scale andare the mean and S.D. of threesimilar experiments. Combina-tion of TGF-β plus IL-21 had astrong positive effect on RORC2gene expression only in T cellscultured in X-VIVO 15 medium(*p<0.05). This treatment wasalso significant for inducing IL-17 expression in the same me-dium (*p<0.05). For IL-23Rtranscript level, although thetreatment were inducible inRPMI 1640 (†, p<0.05), effi-ciency was more in X-VIVO 15medium (¤, p<0.05) and withthe dose I (*a, p<0.05). Thistreatment only with dose I in Tcells cultured in X-VIVO 15 hada slightly significant effect onIFN-γ expression (*a, p<0.05).B, Measurement of protein lev-els of studied cytokines usingELISA method. Data are themean and S.D. of three similarexperiments and are shown inlogarithmic scale. The produc-tion patterns of IL-17, IL-22,and IFN-γ cytokine were almostsimilar to their mRNA levels.
OPTIMIZATION OF HUMAN TH17 CELL DIFFERENTIATION 589
Author's personal copy

of this cytokine. Therefore, it could be concluded that TGF-β is an inducible element for human Th17 cell differenti-ation only in average concentrations and such a role is notexpected when too low or too high concentrations of thiscytokine are applied.
Our results indicated that two more conditions (combi-nations of IL-1β either with IL-6 or IL-23) induced humanTh17 cell differentiation. However, effectiveness wasincreased, especially following use of IL-1β and IL-6, withdose II in X-VIVO 15 medium. Using a combination of IL-1β with either IL-6 or IL-23, Hirota et al. have shownsynergistic effects of IL-6 and IL-1β in Th17 celldifferentiation and up-regulation of RORC2 and IL-17production (Hirota et al. 2010). Some other reports havealso shown inducible effect of IL-6 and IL-1β in Th17 celldifferentiation and up-regulation of RORC2 as well as IL-17 production (Acosta-Rodriguez et al. 2007a, b; Ghilardiand Ouyang 2007; Wilson et al. 2007; Yang et al. 2008a)which supports our findings in present study.
In contrast to murine Th17 differentiation, application ofTGF-β plus IL-6 was not shown to be effective for humanTh17 cell development (Acosta-Rodriguez et al. 2007a, b;Wilson et al. 2007) as verified by our results. We have shownthat such treatment can only induce RORC2 and, in a lowerdegree, IL-23R gene without any positive effect on IL-22 inhuman T cells. Accordingly, Kryczek et al. reported thatTGF-β exerts an inhibitory role for IL-22 gene activation(Kryczek et al. 2007). In addition, in a report from Li Yang etal., it is demonstrated that the combination of IL-6 and TGF-β can promote RORC2 gene activation in human T cells asin mice but such a combination is not able to induce IL-17expression (Yang et al. 2008a). Hence, unidentified factor(s)may act in accordance with RORC2 to promote IL-17 geneexpression in human Th17 cells (Yang et al. 2008a). As inthe mice that RORα is found to exhibit such a role in
collaboration with RORγt (McGeachy and Cua 2008; Yanget al. 2008a, b; Burgler et al. 2010).
Yang et al. have stated that treating CD4+ T cells withTGF-β plus IL-21 in X-VIVO 15 serum-free medium iseffective to induce gene activation of RORC2 and IL-17(Yang et al. 2008a), which is the same as our results.However, such a combination was shown to be ineffectiveon IL-22 production in our study which was not concernedby Yang et al. This is again consistent with Kryczek et alreport which mentioned so far (Kryczek et al. 2007).
Recently, several independent studies have reported theexistence of a subtype of human Th17 cells that is able toproduce both IFN-γ and IL-17 which is named as Th17/Th1 cells (Acosta-Rodriguez et al. 2007a, b; Annunziato etal. 2007; Cosmi et al. 2008; Shi et al. 2008; McGeachy andCua 2008; Peck and Mellins 2009; Maggi et al. 2010;Valmori et al. 2010). Another study has demonstrated that
Table 1 Correlations among different studied markers of Th17 cells
Variables IL-17gene
IL-22gene
IL-23Rgene
RORC2gene
IFN-γgene
IL-17protein
IL-22protein
IFN-γprotein
IL-17 gene Correlation 1.000 −.109 .183 .675* .307* .814* .015 .279*
Significance (2-tailed) .350 .116 .000 .007 .000 .898 .015
IL-22 gene Correlation −.109 1.000 .123 −.220 .516* .055 .776* .525*
Significance (2-tailed) .350 . .293 .058 .000 .637 .000 .000
IL-23R gene Correlation .183 .123 1.000 .388* .279* .144 −.007 .083
Significance (2-tailed) .116 .293 . .001 .015 .218 .950 .477
RORC2 gene Correlation .675* −.220 .388* 1.000 −.192 .221 −.203 −.223Significance (2-tailed) .000 .058 .001 . .098 .057 .080 .055
IFN-γ gene Correlation .307* .516* .279* −.192 1.000 .658* .399* .929*
Significance (2-tailed) .007 .000 .015 .098 . .000 .000 .000
The effects of treatment, concentration and medium are controlled. For each studied marker, n=81
*p=0.05 level
Figure 7. Schematic view of naïve CD4+ T cell polarization todifferent types of Th17 cells. Polarization in presence of TGF-β led todevelopment of single secreting Th17 whereas, polarization withoutTGF-β resulted in double secreting Th17 cells.
5 HAKEMI ET AL.90
Author's personal copy

the combination of IL-β and IL-23 is able to promote thedifferentiation not only of Th17 but also Th17/Th1phenotype (Annunziato et al. 2009). Accordingly, our dataindicated that developing of such cell type could beachieved by treating with IL-1β and IL-6. In addition,using IL-1β plus IL-23 in low concentrations, especiallywhen applied for T cells cultured in serum-supplementedmedium was highly effective on IFN-γ gene activationwhereas, this treatment has lost their effect on IFN-γ whenutilized with dose II.
Interestingly, adding TGF-β to the combination of IL-6plus IL-23, which is shown as the best treatment fordeveloping Th17 cells in current study, did not induce IFN-γ expression and led to development of single secretingTh17 cells (Fig. 7). This may be the prototype of Th17cells. Accordingly, it has been reported that IL-17 and IFN-γ double-secreting Th17 cells were prevalently polarizedfollowing polarization without TGF-β, while polarizationin presence of TGF-β led to induce single-secreting (onlyIL-17 expressing) Th17 cells (Valmori et al. 2010).Regarding to IL-23, although it is required for optimalexpression of IL-17, it may act as a source for outgrowth ofIFN-γ+ progeny under conditions in which TGF-β isdeficient. However, it is suggested that the IFN-γ produc-ing T cells may arise from Th17 precursors. Since IL-23activates STAT4, although weaker than IL-12, it is likelythat STAT4 activation by IL-23 may also contribute tophenotype shift via downstream activation of T-bet (Lee etal. 2009). Regarding to importance of this pathway in IFN-γ gene expression, more detailed studies are needed.
In current study, we showed a significant correlationbetween RORC2 and IL-17 genes expression. In addition,correlation between RORC2 and IL-23R mRNA levels wasalso significant (Table 1). These correlations have beenconfirmed using specific siRNA against RORC2 gene (datanot shown). This phenomenon was also reported by othergroups (Harrington et al. 2005; Ivanov et al. 2006; Volpe etal. 2008; Mus et al. 2010). Thus, RORC2 can be involvedin human IL-17 and IL-23R gene activation. On the otherhand, no correlation between IL-22 and RORC2 genesexpression was found in this research whereas, a significantcorrelation between IL-22 and IFN-γ was observed. Volpeet al. reported that IL-22 is rather a Th1 cytokine and itsproduction by Th1 is more than Th17 subtype (Volpe et al.2008). They have supposed that Th1 population can bedeveloped along with Th17 cells in same microenviron-ment. However, further studies in this context are required.In addition, two different studies have reported that,although the activation of RORC2 may slightly increasethe induction of IL-22 production, initial factors driving IL-22 gene activation are still undefined.
It is possible that IL-22 producing cells are the other yetunknown sub-population of effector T cells in human
(Crome et al. 2009; Unutmaz 2009). This can be confirmedby presence of newly identified transcription factor Arylhydrocarbon receptor (AhR), which is uniquely expressedin Th17 cells. It is demonstrated that AhR-deficient Th17cells completely failed to express IL-22 (Veldhoen et al.2009, 2010; Hirota et al. 2010). Therefore, finding nocorrelation between IL-22 and RORC2 is not a surprise.
Collectively, data presented in this study improved ourknowledge about polarization conditions for Th17 celldifferentiation from human naïve helper T cells.
Acknowledgments The main part of this work was supplied byIsfahan University of Medical Sciences (Grant No. 187058) and theremaining was financially supported by Iran National ScienceFoundation (Grant No. 87040290). We wish to thanks Dr. Shahshahaniand Mrs Janghorban for their excellent helps in cord blood collection,Mrs Homayouni and Mrs Esmaeili for their kindly collaborations inflow cytometry and ELISA techniques and Mr. Ghoochani for hisexcellent helps in data interpretation.
References
Acosta-Rodriguez E. V.; Napolitani G.; Lanzavecchia A.; SallustoF. Interleukins 1beta and 6 but not transforming growthfactor-beta are essential for the differentiation of interleukin17-producing human T helper cells. Nat. Immunol. 8: 942–949; 2007a.
Acosta-Rodriguez E. V.; Rivino L.; Geginat J.; Jarossay D.; GattornoM. Surface phenotype and antigenic specificity of humaninterleukin 17-producing T helper memory cells. Nat. Immunol.8: 639–46; 2007b.
Aggarwal S.; Ghilardi N.; Xie M. H.; de Sauvage F. J.; Gurney A. L.Interleukin-23 promotes a distinct CD4 T cell activation statecharacterized by the production of interleukin-17. J. Biol. Chem.278: 1910–1914; 2003.
Annunziato F.; Cosmi L.; Liotta F.; Maggi E.; Romagnani S. HumanTh17 cells: are they different from murine Th17 cells? Eur. J.Immunol. 39: 637–640; 2009.
Annunziato F.; Cosmi L.; Santarlasci V.; Maggi L.; Liotta F.;Mazzinghi B.; Parente E. Phenotypic and functional features ofhuman Th17 cells. J. Exp. Med. 204: 1849–1861; 2007.
Bettelli E.; Carrier Y.; Gao W.; Korn T.; Storm T.; Oukka M.Reciprocal developmental pathways for the generation ofpathogenic effector TH17 and regulatory T cells. Nature 441:235–8; 2006.
Burgler S.; Mantel P. Y.; Bassin C.; Ouaked N.; Akdis C. A.; Schmidt-Weber C. B. RORC2 is involved in T cell polarization throughinteraction with the FOXP3 promoter. J. Immunol. 184: 6161–6169; 2010.
Burgler S.; Ouaked N.; Bassin C.; Basinski T. M.; Mantel P. Y.;Siegmund K.; Meyer N.; Akdis C. A.; Schmidt-Weber C. B.Differentiation and functional analysis of human T(H)17 cells. J.Allergy Clin. Immunol. 123(588–95): 595; 2009.
Cardoso C. R.; Garlet G. P.; Crippa G. E.; Rosa A. L.; Junior W. M.;Rossi M. A.; Silva J. S. Evidence of the presence of T helper type17 cells in chronic lesions of human periodontal disease. OralMicrobiol. Immunol. 24: 1–6; 2009.
Cosmi L.; De Palma R.; Santarlasci V.; Maggi L.; Capone M.; FrosaliF.; Rodolico G. Human interleukin-17-producing cells originatefrom a CD161+ CD4+ T-cell precursor. J. Exp. Med. 205: 1903–1916; 2008.
OPTIMIZATION OF HUMAN TH17 CELL DIFFERENTIATION 591
Author's personal copy

Crome S. Q.; Wang A. Y.; Kang C. Y.; Levings M. K. The role ofretinoic acid-related orphan receptor variant 2 and IL-17 in thedevelopment and function of human CD4+ T cells. Eur. J.Immunol. 39: 1480–1493; 2009.
Ghilardi N.; Ouyang W. Targeting the development and effectorfunctions of TH17 cells. Semin. Immunol. 19: 383–393; 2007.
Gocke A. R.; Cravens P. D.; Ben L. H.; Hussain R. Z.; Northrop S. C.;Racke M. K.; Lovett-Racke A. E. T-bet regulates the fate of Th1and Th17 lymphocytes in autoimmunity. J. Immunol. 178: 1341–1348; 2007.
Harrington L. E.; Hatton R. D.; Mangan P. R.; Turner H.; Murphy T.L.; Murphy K. M.; Weaver C. T. Interleukin 17-producing CD4+effector T cells develop via a lineage distinct from the T helpertype 1 and 2 lineages. Nat. Immunol. 6: 1123–1132; 2005.
Hirota K.;Martin B.; VeldhoenM.Development, regulation and functionalcapacities of Th17 cells. Semin. Immunopathol. 32: 3–16; 2010.
Ivanov I. I.; McKenzie B. S.; Zhou L.; Tadokoro C. E.; Lepelley A.;Lafaille J. J.; Cua D. J.; Littman D. R. The orphan nuclear receptorRORgammat directs the differentiation program of proinflammatoryIL-17+ T helper cells. Cell 126: 1121–1133; 2006.
Kenneth J. L.; Thomas D. S. Analysis of Relative Gene ExpressionData Using Real-Time Quantitative PCR and the 2-ddCt Method.Methods 25: 402–408; 2001.
Kryczek I.; Wei S.; Vatan L.; Escara-Wilke J.; Szeliga W.; Keller E. T.;Zou W. Cutting edge: opposite effects of IL-1 and IL-2 on theregulation of IL-17+ T cell pool IL-1 subverts IL-2-mediatedsuppression. J. Immunol. 179: 1423–1426; 2007.
Laurence A.; Tato C. M.; Davidson T. S.; Kanno Y.; Chen Z.; Yao Z.;Blank R. B.; Meylan F.; Siegel R.; Hennighausen L.; Shevach E.M.; O'Shea J. J. Interleukin-2 signaling via STAT5 constrains Thelper 17 cell generation. Immunity. 26: 371–381; 2007.
Lee Y. K.; Turner H.; Maynard C. L.; Oliver J. R.; Chen D.; Elson C.O.; Weaver C. T. Late developmental plasticity in the T helper 17lineage. Immunity. 30: 92–107; 2009.
Maggi L.; Santarlasci V.; Capone M.; Peired A.; Frosali F.; Crome S.Q.; Querci V.; Fambrini M.; Liotta F.; Levings M.; Maggi E.;Cosmi L.; Romagnani S.; Annunziato F. CD161 is a marker of allhuman IL-17-producing T-cell subsets and is induced by RORC.Eur. J. Immunol. 40: 2174–2181; 2010.
Manel N.; Unutmaz D.; Littman D. R. The differentiation of human T(H)-17 cells requires transforming growth factor-beta andinduction of the nuclear receptor RORgamma. Nat. Immunol. 9:641–49; 2008.
Mangan P. R.; Harrington L. E.; O'quinn D.; Helms W.; Bullard D.;Elson C. Transforming growth factor-ß induces development ofTH17 lineage. Nature 441: 231–4; 2006.
Martin-Orozco N.; Chung Y.; Chang S. H.; Wang Y. H.; Dong C. Th17cells promote pancreatic inflammation but only induce diabetesefficiently in lymphopenic hosts after conversion into Th1 cells.Eur. J. Immunol. 39: 216–224; 2009.
McGeachy M. J.; Cua D. J. Th17 cell differentiation: the long andwinding road. Immunity. 28: 445–453; 2008.
Michael W. P. A new mathematical model for relative quantification inreal-time RT-PCR. Nucleic Acids Res. 29: 2002–2007; 2001.
Mus A. M.; Cornelissen F.; Asmawidjaja P. S.; van Hamburg J. P.;Boon L.; Hendriks R. W.; Lubberts E. Interleukin-23 promotesTh17 differentiation by inhibiting T-bet and FoxP3 and isrequired for elevation of interleukin-22, but not interleukin-21,in autoimmune experimental arthritis. Arthritis Rheum. 62: 1043–1050; 2010.
Naive CD4+ T cell isolation kit II human. [130-094-131]. 2008.Miltenyi Biotec GmbH. Ref Type: Catalog.
Ouyang W.; Kolls J. K.; Zheng Y. The biological functions of T helper17 cell effector cytokines in inflammation. Immunity. 28: 454–467; 2008.
Park H.; Li Z.; Yang X. O.; Chang S. H.; Nurieva R.; Wang Y. H.;Wang Y.; Hood L.; Zhu Z.; Tian Q.; Dong C. A distinct lineage ofCD4 T cells regulates tissue inflammation by producing interleu-kin 17. Nat. Immunol. 6: 1133–1141; 2005.
Peck A.; Mellins E. D. Plasticity of T-cell phenotype and function: theT helper type 17 example. Immunology 129: 147–153; 2009.
Shi G.; Cox C. A.; Vistica B. P.; Tan C.; Wawrousek E. F.; Gery I.Phenotype switching by inflammation-inducing polarized Th17cells, but not by Th1 cells. J. Immunol. 181: 7205–7213; 2008.
Streeck H.; Cohen K. W.; Jolin J. S.; Brockman M. A.; Meier A.;Power K. A.; Waring M. T.; Alter G.; Altfeld M. Rapid ex vivoisolation and long-term culture of human Th17 cells. J. Immunol.Methods 333: 115–125; 2008.
Unutmaz D. RORC2: the master of human Th17 cell programming.Eur. J. Immunol. 39: 1452–1455; 2009.
Valmori D.; Raffin C.; Raimbaud I.; Ayyoub M. Human ROR{gamma}t+TH17 cells preferentially differentiate from naiveFOXP3+Treg in the presence of lineage-specific polarizingfactors. Proc. Natl. Acad. Sci. U. S. A 107: 19402–19407; 2010.
Veldhoen M.; Hirota K.; Christensen J. Natural agonists for arylhydrocarbon receptor in culture medium are essential foroptimal differentiation of Th17 T cells. J. Exp. Med. 206: 43–49; 2009.
Veldhoen M.; Hirota K.; Westendorf A. The aryl hydrocarbon receptorlinks Th17-cell-mediated autoimmunity to invironmental toxins.Nature 453: 106–109; 2010.
Veldhoen M.; Hocking R.; Atkins C.; Locksley R.; Stockinger B.TGF-ß in the context of an inflammatory cytokine milieusupports de novo differentiation of IL-17-producing T cells.Immunity 24: 179–189; 2006.
Volpe E.; Servant N.; Zollinger R.; Bogiatzi S. I.; Hupe P.; Barillot E.;Soumelis V. A critical function for transforming growth factor-beta, interleukin 23 and proinflammatory cytokines in drivingand modulating human T(H)-17 responses. Nat. Immunol. 9:650–657; 2008.
Vyakarnam A. Human CD4 Culture. In: Rowland-Jones S. L.;McMichael A. (eds) Lymphocytes: a practical approach. OxfordUniversity Press, Oxford, pp 136–142; 2000.
Wilson N. J.; Boniface K.; Chan J. R.; McKenzie B. S.; BlumenscheinW. M.; Mattson J. D.; Basham B.; Smith K.; Chen T.; Morel F.;Lecron J. C.; Kastelein R. A.; Cua D. J.; McClanahan T. K.;Bowman E. P.; de Waal M. R. Development, cytokine profile andfunction of human interleukin 17-producing helper T cells. Nat.Immunol. 8: 950–957; 2007.
Yang L.; Anderson D. E.; Baecher-Allan C.; Hastings W. D.; BettelliE.; Oukka M.; Kuchroo V. K.; Hafler D. A. IL-21 and TGF-betaare required for differentiation of human T(H)17 cells. Nature454: 350–352; 2008a.
Yang X. O.; Pappu B. P.; Nurieva R.; Akimzhanov A.; Kang H. S.;Chung Y.; Ma L.; Shah B.; Panopoulos A. D.; Schluns K. S.;Watowich S. S.; Tian Q.; Jetten A. M.; Dong C. T helper 17lineage differentiation is programmed by orphan nuclear recep-tors ROR alpha and ROR gamma. Immunity. 28: 29–39; 2008b.
Zhu J.; Paul W. E. CD4 Tcells: fates, functions, and faults. Blood 112:1557–1569; 2008.
5 2 HAKEMI ET AL.9
Author's personal copy
Copyright © 2022 FDOKUMEN




















