
Morphogenesis and Differentiation in Rhodomicrobium ... - NCBI
55
BAcTERIowGicAL Rzvizws, Sept. 1977, p. 754-808 Vol. 41, No. 2 Copyright 0 1977 American Society for Microbiology Printed in U.S.A. Morphogenesis and Differentiation in Rhodomicrobium vannielii and Other Budding and Prosthecate Bacteria R. WHITTENBURY* AND C. S. DOW Department of Biological Sciences, University of Warwick, Coventry, CV4 7AL, England PREVIEW ...................................... 754 PART A-INTRODUCTION ...................................... 755 Cellular Events ...................................... 755 Cell Cycle Types ...................................... 755 Levels of Organization .......................................................-756 Budding-What It Means in Bacteria ...................................... 756 Intercalation .......................................---------------.--.760 Polar growth ...............................760 Budding Bacteria as Models for the Study of Morphogenesis and Differentiation 762 PART B -BIOLOGY OF THE CELL CYCLES OF R. VANNIELII ..... ........ 762 Review of Published Information on R. vannielii .............................. 762 Enrichment, Isolation, and Cultivation .................................... 763 Physiological and Biochemical Properties ..................................... 764 Characterization of DNA ................................................... 764 Photopigments .....................................-------------...764 Cytochromes ......................................------------.-.---.--.764 Growth physiology.764........................... ... 764 Cell Cycle Morphology of R. vannielii ..............-.-.---.-...---.....--..765 Synchronous cultures ......................................................-766 Synchrony by filtration . .................................. 766 Synchronous Growth of Rhodomicrobium Swarm Cells . 767 Assay of "landmark" events in liquid culture ................................ 767 Vegetative Cell Cycle ........................................................-769 Maturation sequence .................................... 769 Reproductive sequence .....................................................-769 Subsequent development .................................... 770 Other Cell Types Formed ...................-.-.-.-.-.-.-.772 "Double-yolk" cells ......................................--.-.------ 772 Exospore formation ......................--.-.---.........772 Exospores .................. 772 Occurrence and collection ...................----------------772 Morphology and formation ....................--.-..----772 Germination and outgrowth .................. 774 Physiological properties .................................................... 774 Fine Structure of Vegetative Cells .................. 777 Fine structure .................. 777 Fine Structure of Exospores ...................-.-.-.-.-.-.-.-.-.-.782 Synthesis of Chlorophyll, Protein, DNA, and RNA in Synchronized Cells . 788 Chlorophyll synthesis ............... 788 Protein synthesis and inhibition ............................------789 RNA synthesis............................................................. 794 DNA synthesis............................................................. 794 Reproduction ................................------------------800 CONCLUSION.805................................................. 805 LITERATURE CITED ........................... 805 PREVIEW models for the study of development in procar- yotes are highlighted in this comparative treat- This monograph is in two parts. Part A sets ment, which also underlines the meaning of out the essential nature of budding and pros- morphogenesis and differentiation when used thecate bacteria in terms of their cell cycle and in a procaryote context. reproductive processes and contrasts them with Rhodomicrobium vannielii seems the most other eubacteria such as Escherichia coli. Ad- promising of the budding bacteria for the study vantages budding bacteria may possess as of morphogenesis and differentiation and is the 754
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Morphogenesis and Differentiation in Rhodomicrobium ... - NCBI

BAcTERIowGicAL Rzvizws, Sept. 1977, p. 754-808 Vol. 41, No. 2Copyright 0 1977 American Society for Microbiology Printed in U.S.A.
Morphogenesis and Differentiation in Rhodomicrobiumvannielii and Other Budding and Prosthecate Bacteria
R. WHITTENBURY* AND C. S. DOWDepartment ofBiological Sciences, University of Warwick, Coventry, CV4 7AL, England
PREVIEW ...................................... 754PART A-INTRODUCTION ...................................... 755
Cellular Events ...................................... 755Cell Cycle Types ...................................... 755Levels of Organization .......................................................-756Budding-What It Means in Bacteria ...................................... 756
Intercalation .......................................---------------.--.760Polar growth...............................760
Budding Bacteria as Models for the Study of Morphogenesis and Differentiation 762PART B-BIOLOGY OF THE CELL CYCLES OF R. VANNIELII ..... ........ 762Review of Published Information on R. vannielii .............................. 762Enrichment, Isolation, and Cultivation .................................... 763Physiological and Biochemical Properties ..................................... 764
Characterization of DNA ................................................... 764Photopigments .....................................-------------...764Cytochromes ......................................------------.-.---.--.764Growth physiology.764.............................. 764
Cell Cycle Morphology of R. vannielii ..............-.-.---.-...---.....--..765Synchronous cultures ......................................................-766Synchrony by filtration................................... 766
Synchronous Growth of Rhodomicrobium Swarm Cells. 767Assay of "landmark" events in liquid culture ................................ 767
Vegetative Cell Cycle ........................................................-769Maturation sequence .................................... 769Reproductive sequence .....................................................-769Subsequent development .................................... 770
Other Cell Types Formed ...................-.-.-.-.-.-.-.772"Double-yolk" cells ......................................--.-.------ 772Exospore formation......................--.-.---.........772
Exospores.................. 772Occurrence and collection...................----------------772Morphology and formation ....................--.-..----772Germination and outgrowth .................. 774Physiological properties .................................................... 774
Fine Structure of Vegetative Cells .................. 777Fine structure.................. 777
Fine Structure of Exospores ...................-.-.-.-.-.-.-.-.-.-.782Synthesis of Chlorophyll, Protein, DNA, and RNA in Synchronized Cells. 788
Chlorophyll synthesis............... 788Protein synthesis and inhibition ............................------789RNA synthesis............................................................. 794DNA synthesis............................................................. 794
Reproduction................................------------------800CONCLUSION.805.................................................805LITERATURE CITED ........................... 805
PREVIEW models for the study of development in procar-yotes are highlighted in this comparative treat-
This monograph is in two parts. Part A sets ment, which also underlines the meaning ofout the essential nature of budding and pros- morphogenesis and differentiation when usedthecate bacteria in terms of their cell cycle and in a procaryote context.reproductive processes and contrasts them with Rhodomicrobium vannielii seems the mostother eubacteria such as Escherichia coli. Ad- promising of the budding bacteria for the studyvantages budding bacteria may possess as of morphogenesis and differentiation and is the
754

VOL. 41, 1977
central subject of this monograph. Certain sec-tions of Part A draw upon information in PartB, which begins with a brief review of pub-lished information on R. vannielii but is princi-pally an account of our studies on the biology ofthe cell cycles of this organism, with particularreference to morphogenesis and differentiation.
PART A-INTRODUCTIONGrowth and development of higher plants
and animals is a composite expression of thegrowth and differentiation of their componentcells. Consequently, it has been argued that astudy of molecular events taking place duringthe cell cycle, especially those events related todifferentiation and morphogenesis, is directlyrelevant to the understanding of growth anddevelopment of the whole organism.
Eucaryotic cells, obviously, are the cells ofchoice in which to explore these events. How-ever, so much more is known about molecularprocesses and their genetic control in procar-yotes that it is plausible to suppose that perti-nent information about the basic principles, ifnot the precise processes, of morphogenesis anddifferentiation in higher organisms might beunraveled more rapidly by studying procar-yotes.The ideal choice of bacterial species for such
studies would be Escherichia coli, simply be-cause of the vast bank of genetic and biochemi-cal information that already exists about thisspecies. However, E. coli lacks significant dif-ferentiation; it has a simple morphogenetic cellcycle, cell division being the only clearly micro-scopically observable "landmark" event. Pro-caryotes with complex fine structures and mor-phologically distinctive cell cycles would seembetter choices and have proved to be so.
Procaryotes that offer possibilities for suchexploitation include species of Bacillus, Strep-tomyces, Arthrobacter, Myxobacter, the pros-thecate bacteria (including Caulobacter andthe budding bacteria), and filamentous cyano-bacteria (blue-green bacteria). Some of theseorganisms, ofcourse, have already been studiedin this context. Endosporeforming Bacillus spe-cies (26, 30, 49, 82) are the most prominentexamples, followed more recently by Caulobac-ter (43, 74, 76).
Before pursuing the discussion on morpho-genesis, differentiation, and development inbudding bacteria, we think it necessary to at-tempt to be more precise than is presently thecase about the meaning and use of these termsas applied to procaryotes. We spell out belowour notions ofthese terms and then use them infurther descriptions of categories of cell cycles,
MORPHOGENESIS OF R. VANNIELUI 755
intercellular relationships, and levels of orga-nization of cells.
Cellular EventsMorphogenesis -changes in external mor-
phology of the cell and in internal architectureduring the cell cycle.
Differentiation -events initiated by a"switch" in the cell cycle leading to the forma-tion of a new type of cell. Such differentiatedcells may revert to the original form (e.g., sporegermination), or the differentiation event maybe permanent (e.g., heterocysts of cyanobac-teria and cells ofcertain methane oxidizers thathave budded off spores [95]).Development-a composite event involving
morphogenesis and differentiation under inter-cellular influence, as in some myxobacters andcyanobacteria when cells are modified to per-form a particular function necessary to the ac-tivity of the multicellular complex.
Cell Cycle TypesMonomorphic vegetative cell cycle type -this
phrase describes species of procaryote existingalways as vegetative cells and in which there isonly one distinct morphological type of cellformed under what are considered to be normalnutrient and physiological regimes (e.g., E.coli).Dimorphic cell cycle types -this phrase de-
scribes those procaryotes which, at division,form two cells that differ from each other inshape (e.g., Caulobacter), or in size (e.g., cer-tain filamentous blue-green bacteria), or in thepossession by one of the cells of an appendage(e.g., the swarm cell of Rhodopseudomonasacidophila possesses a flagellum not possessedby the mother cell), or in a combination ofthesefeatures.Polymorphic vegetative cell cycle types -this
phrase describes species that may produce un-der different nutrient and physiological re-gimes (but not so bizarre as to cause malfinc-tion in the cell cycle) two or more morphologi-cally distinct types of cell, each of which under-goes a distinctive and constant cell cycle (e.g.,Arthrobacter species [24, 42], Sphaerotilus spe-cies [54, 66] certain cyanobacteria [9, 51], Geo-dermatophilus [36], Hyphomicrobium species[88], aO Rhodomicrobium species; all thesehave two- or more vegetative cell cycles). Cellcycle changes iti-response to nutrient conditionsin Geodermatophilus are shown in Fig. 1. Twoother species with su"h cell cycles have beenstudied by us, a Hyph8iicrobium species (3)(Fig. 2 and 3) and a prosth ate bacterium (Fig.4 and 5).

756 WHITTENBURY AND DOW
,"---- I., the polymorphic cell cycle type (e.g., Rhodomi-, A crobium species. as described in Part B).
Levels of OrganizationC-form~ ~ ~~
o
C - form8tj
I I
%- ad'nAJ, tVIMY Differentiation to R-form
Differentiation toC-form
ok0 0 &§FIG. 1. The polymorphic growth cycle
matophilus strain 22-68 (after Ishigurc[36D). Growth and division in the C-formpresence ofa factor found in tryptose. Abfactor induces differentiation to the R-fotion of the tryptose will induce differenthe R-form to the C-form.
f+ Methanol
Bud It / Motfomaotion % % cel
Obligate As I
Differentiation-
FIG. 2. Polymorphic variation evidphomicrobium species with substitutionsource. The basic medium contained, peiof mineral base (NH4Cl, 0.5 g; MgSO,NaCl, 0.1 g; CaCl2 -2H20, 0.05 g; traceml [61]), distilled water to 1 liter, 10 nKH2PO4, 100 mM methanol, and/or 100amine. Cultures were incubated aerobic
K- torm Unicellular -the organism is normally inde-J pendent of its fellows in terms of function and
reproduction.Occasional intercellular cooperation -de-
scribes bacteria able to lead a unicellular exis-tence but which may express intercellular coop-eration to form a multicellular structure (e.g.,fruiting-body formation by some species ofMyxobacter [23]).Multicellular procaryotes-this expressiondescribes those procaryotes with the capacity
- (not necessarily obligate) to differentiate anddevelop into a multicellular complex that
'eof Geoder- clearly has (i) survival potential superior too and Wolfe that of a single cell of that species in the same
requires the environment, and (ii) an intercellular activityisence ofthis that results in selected cells of that complexarm. Readdi- undergoing differentiation.ltiation from Conventional wisdom holds that eubacteria
are unicellular entities that do not form differ-entiated complexes, as is the case with manytypes of eucaryotic cells. That there are in-
+ Methylanine stances of cellular interdependence in procar-cMoixx yotes has been recognized, as in the case offormation myxobacter and related organisms, but what
seems to have escaped general recognition isthe existence of multicellular units ofcyanobac-teria (Fig. 6) where the evidence (97, 98) indi-cates to us that the filament of cells should beregarded as a whole organism. That the cells
/+Methoaol are interdependent, in the sense that differen-I tiation (the formation of heterocysts, end cells,
1*sw.,,. and akinetes) is a function of cell-cell relation-Cell ships, is now clearly established (9, 51). The
advantage of such a multicelled organism overa single-celled cyanobacterium of the same spe-cies is indicated by the gliding movements ofthe trichome, the N2-fixing powers under aero-bic conditions of the heterocyst, and the forma-tion of one resting cell per filament (9). Ob-viously, a multicellular organism made up of
rent in Hy- procaryotic cells is potentially an excellentofthe carbon model for studies on differentiation and mor-r liter: 20 ml phogenesis.7H20, 0.4 g; In addition, studies with the cyanobacteriumelements, 10 Chlorogloea fritschii (25) have shown that com-mM methyl- plex but predictable morphological variationsally at 30hC can be induced by a variety of environmental
stimuli (Fig. 7).
Spore and cyst cycles -this describes speciesthat may undergo a cell cycle in addition to thevegetative cell cycle. Such species may be ofthemonomorphic vegetative cell cycle variety(e.g., Bacillus and Methylosinus species) or of
Budding-What It Means in Bacteria
Before we can examine the proposition thatbudding bacteria can serve as good models forthe study of cell development, it is necessary to
Mo phogenetic(phenotypoc)C
voriation < a
Ha_ IA
Methylominek exponentialphase)
-= -I
BACTECRIOL. REV.
,' AII
4
II
n I- ---I
¾
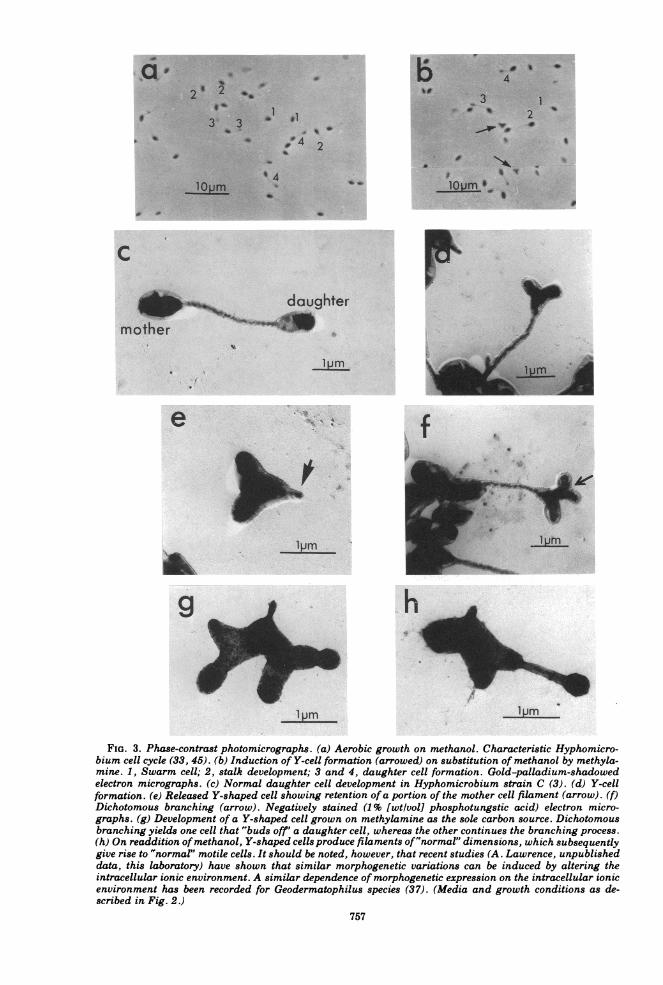
FIG. 3. Phase-contrast photomicrographs. (a) Aerobic growth on methanol. Characteristic Hyphomicro-bium cell cycle (33, 45). (b) Induction of Y-cell formation (arrowed) on substitution of methanol by methyla-mine. 1, Swarm cell; 2, stalk development; 3 and 4, daughter cell formation. Gold-palladium-shadowedelectron micrographs. (c) Normal daughter cell development in Hyphomicrobium strain C (3). (d) Y-cellformation. (e) Released Y-shaped cell showing retention ofa portion of the mother cell filament (arrow). (f)Dichotomous branching (arrow). Negatively stained (1% [wtlvol] phosphotungstic acid) electron micro-graphs. (g) Development of a Y-shaped cell grown on methylamine as the sole carbon source. Dichotomousbranching yields one cell that "buds off' a daughter cell, whereas the other continues the branching process.(h) On readdition ofmethanol, Y-shaped cells produce filaments of"normal" dimensions, which subsequentlygive rise to "normal" motile cells. It should be noted, however, that recent studies (A. Lawrence, unpublisheddata, this laboratory) have shown that similar morphogenetic variations can be induced by altering theintracellular ionic environment. A similar dependence of morphogenetic expression on the intracellular ionicenvironment has been recorded for Geodermatophilus species (37). (Media and growth conditions as de-scribed in Fig. 2.)
757
-.MWOMP111)M- --,
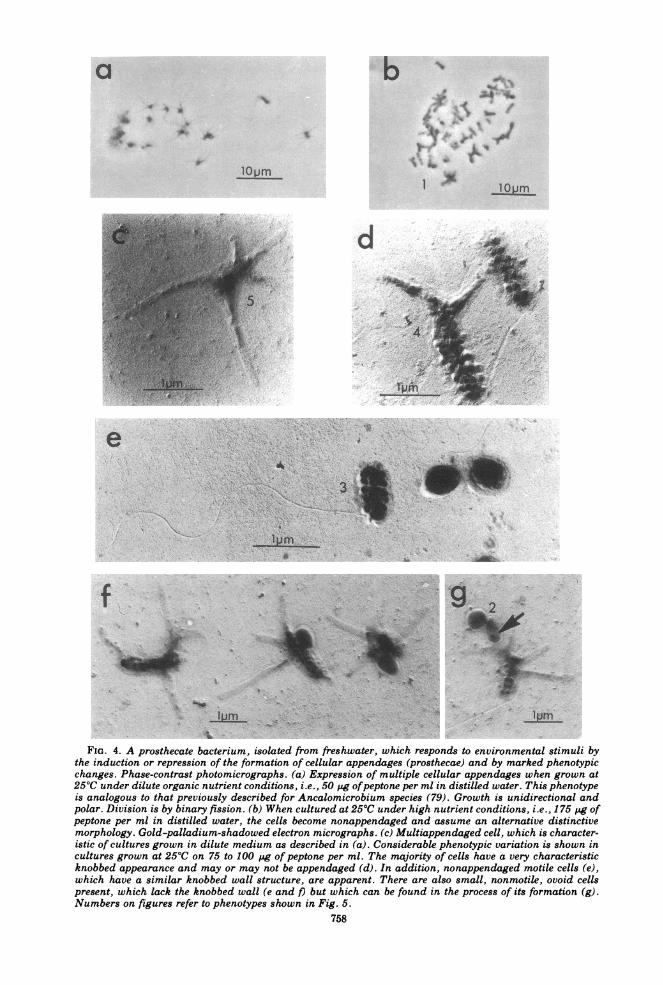
FIG. 4. A prosthecate bacterium, isolated from freshwater, which responds to environmental stimuli bythe induction or repression of the formation of cellular appendages (prosthecae) and by marked phenotypicchanges. Phase-contrast photomicrographs. (a) Expression of multiple cellular appendages when grown at250C under dilute organic nutrient conditions, i.e., 50 pg ofpeptone per ml in distilled water. This phenotypeis analogous to that previously described for Ancalomicrobium species (79). Growth is unidirectional andpolar. Division is by binary fission. (b) When cultured at 250C under high nutrient conditions, i.e., 175 Pg ofpeptone per ml in distilled water, the cells become nonappendaged and assume an alternative distinctivemorphology. Gold-palladium-shadowed electron micrographs. (c) Multiappendaged cell, which is character-istic of cultures grown in dilute medium as described in (a). Considerable phenotypic variation is shown incultures grown at 250C on 75 to 100 pg of peptone per ml. The majority of cells have a very characteristicknobbed appearance and may or may not be appendaged (d). In addition, nonappendaged motile cells (e),which have a similar knobbed wall structure, are apparent. There are also small, nonmotile, ovoid cellspresent, which lack the knobbed wall (e and /) but which can be found in the process of its formation (g).Numbers on figures refer to phenotypes shown in Fig. 5.
758

MORPHOGENESIS OF R. VANNIELII 759
E) 2
Organic nutrientconcentration>175 pgIml
Organic ^nutrient concentration
<50 lig/ml
/ 3
1/ 4FIG. 5. Summary of the phenotypic variations in-
duced in a freshwater prosthecate bacterium by vari-ation of the organic nutrient concentration. The celltypes have been correlated with those in Fig. 4.
endheterocyst_ n cell
-W w ineteC-s~~~~etero eeroyst
..roc st
encell
FIG. 6. Diagrammatic representation of an Ana-baena cylindrica filament showing the constituentcell types. Asymmetric division of the vegetative cellsyield a large (L) and a small (S) cell, which differ indivision time (51).
define what budding really means in the con-text of bacterial reproductive processes.Budding bacteria are currently assumed to
differ fundamentally from other bacteria intheir mode of reproduction. Such an assump-tion stems from a comparison of their cell cycleswith those of rod-shaped bacteria such as E.coli (Fig. 8). In budding bacteria, cell separa-tion usually results in two asymmetric cells(one cell with a filament, one without) whichare not siblings, whereas in E. coli divisionusually results in two symmetrical siblings.Division processes, however -binary in bothcases -are fundamentally similar.A second point of significance concerns a
characteristic that has been consistently over-looked and is probably the most important sin-gle feature characterizing budding bacteria:wall growth is always polar whatever the me-dium in which the organism is grown. Wallgrowth, or cell extension, may be either unidi-rectional, from only one pole (always the samepole) as in Rhodopseudomonas palustris (96),or unidirectional but from either pole, e.g., Hy-phomicrobium species (A. Lawrence, unpub-lished data, this laboratory), or bidirectional,that is to say, both poles of the bacterium (e.g.,
R. vannielii [see Part B]) may give rise to cells.E. coli and other rod-shaped bacteria, on theother hand, usually grow from multiple growthpoints along the envelope approximating to in-tercalary growth (4, 5, 17).That even this process is not always the case
with rod-shaped cells has been demonstratedwith E. coli (17, 18). In minimal medium with ageneration time of 60 min, E. coli will extendonly from one pole. The only difference betweenbudding bacteria and E. coli in this respect isthat in budding bacteria division is asymmetric(Fig. 8). Consequently, no fundamentally im-
type C
type)AA0 ot13h1o sub
FIG. 7. Diagrammatic summary of the morpho-logical variations induced in Chlorogloea fritschii bydifferent environmental stimuli (after E. H. Evans, I.Foulds, and N. G. Carr [25D). Morphological celltypes. Type A -large (2 by 3 pm) granulated cellsexisting either singly or as clumps containing two ormore cells which arise from division in up to threeplanes. Type B-found in clumps which combinelarger groups of cells surrounded by a mucilaginoussheath, which is termed "aseriate" (44). Type C -small (1 ,gm) cells found in short filaments. Type D -larger than type C (1.5 gm) and found in filaments inthe process of dividing.
CHfll DA-
a b cFIG. 8. Diagrammatic representation of: (a) inter-
calary growth of Escherichia coli with a generationtime of less than 40 min (4, 17); (b) polar growthshown by E. coli with a generation time in excess of60 min (17); (c) obligate polar growth of a buddingbacterium, e.g., Hyphomicrobium species (33, 45).
VOL. 41, 1977

760 WHITTENBURY AND DOW
portant feature in cell growth and division proc-esses can be singled out which separates abso-lutely the budding bacteria from the rod-shapedbacteria.By adopting this mode of envelope growth as
a basis of distinction and disregarding the termbudding, more useful concepts can be evolvedabout cell growth and development in buddingbacteria. Once bacteria have adopted a "uni-point" mode of growth, certain constraints asregards shape and internal architecture whichgovern such bacteria as E. coli appear no longerto apply, and it is possible for morphogeneticevolution to take place. That such an idea maybe reasonable is illustrated in Fig. 9, in whichsome of the budding bacteria are classified ac-cording to degree of morphological complexity.Three major features of this particular exam-
ple of morphogenetic evolution, pertinent to R.vannielii in particular, are worth emphasizing.(i) The membranous unit of lamellar structureis no longer symmetrical. A "horseshoe" shapehas evolved (e.g., Nitrobacter winogradsky andR. palustris [56, 93, 96]), which does not giverise to complications at division, since the fis-sion point is beyond the "horseshoe" structure,
maturationI fr
1 2 3 4 5
RhodoP seudomonos
R poiustrisR viridi,
r Hyphomicrobium sps
d 4W_ =
Rhodomicrobium Sps
FIG. 9. Budding bacteria ordered in degrees ofmorphological/cell cycle complexity. In all cases mo-tility (1), flagella shedding (2), maturation ofthe cell(3), daughter cell synthesis by obligate polar growth(4), and asymmetric cell division (5) give rise to an
immature daughter cell and a mature mother cell.(a) Rhodopseudomonas acidophila (60). Simple di-morphic life cycle not involving tube/filament synthe-sis. (b) Rhodopseudomonas palustris and R. viridis(96). Involvement of tube/filament synthesis prior todaughter cell formation. (c) Hyphomicrobium species(33, 45). Reduction in width coupled with an in-crease in length of the tubelfilament compared withR. palustris. (d) Rhodomicrobium species (see PartB). Environmentally induced life cycle variationsranging from a simple dimorphic cell cycle throughcomplex matrix formation to exospore production.
which is formed de novo in the daughter cell.(ii) Multicellular arrays are formed wherebycells remain permanently linked to one an-other. (iii) A second mode of cell separationother than binary fission has evolved in R.vannielii, the plugging of filaments or the lay-ing down of cross walls in the filaments. Theseseparation sites, part way along the filament,are not those at which binary fission wouldhave taken place; fission sites are directly adja-cent to the daughter cells.The significance of the adoption by a bacte-
rium of a polar or unipoint mode of growth asopposed to multipoint, or what approximates toan intercalary mode of growth, is far-reaching.The dramatic consequences of such as appar-ently simple change in growth style are out-lined below in a comparison of intercalary withobligate polar growth processes.
Intercalation. If a rod-shaped bacteriumgrowing exponentially or in a steady state incontinuous culture lengthens by ideal interca-lary processes (i.e., old and new structures andcytoplasmic material are evenly distributedthroughout the expanding cell), then at divi-sion the two siblings will be both quantitativelyand qualitatively similar to each other. Notonly will the siblings be to all intents and pur-poses indistinguishable, but they will also besimilar physiologically and structurally (i.e., inthe distribution and ratio of old to new compo-nents) to the cell that gave rise to them at themoment it was formed by division. From thisdescription of events, four major featuresemerge which characterize the ideal interca-lary-type cell: (i) such cells are potentially im-mortal; (ii) the cells have an age span that doesnot exceed the division time; (iii) at division,siblings are equivalent in all respects; (iv) in-ternal structures, such as membranous com-plexes, are likely to be symmetrical (for uni-form distribution at division between the twosibling cells) or to be small structures, such asmesosomes, which do not interfere with divi-sion.Polar growth. Taking a Hyphomicrobium
species as a typical example of a bacteriumgrowing by polar processes (Fig. 8 and 9), thefollowing features characterize its growth cy-cle, observed in ideal circumstances similar tothe previous example of an intercalary growingcell.At division, the cell divides asymmetrically,
yielding a mother cell and a daughter cell. Themother cell is a filamented cell composed ofstructural material present in the cell when itwas formed at the previous division (this mate-rial will be confined mainly to the nonfila-mented part of the cell), and of structural mate-
BACTERIOL. REV.
reproduc t ion
MORPHOGENESIS OF R. VANNIELII 761
rial synthesized during the recent cell cycle(this will be restricted to the budding filament).The flagellated daughter will be composedmainly of new material synthesized in the pre-ceding cell cycle.
In subsequent rounds of division, little if anynew structural material will be added to themother cell: in this sense the mother cell ages.The implications of polar growth in contrast
to intercalary growth are as follows.(i) Aging is a real process that extends
through all the division cycles of a particularmother cell's life-span. At division, theswarmer (daughter cell) is structurally youngerthan the mother cell.The aging concept is best illustrated at pres-
ent in procaryotes by the Caulobacter stalkedcells, which have been reported to acquire anadditional cross band in the stalk after eachdivision (80), a possible consequence of a newpiece of stalk being synthesized in each divisioncycle. However, recent evidence summarizedbelow suggests that this interpretation ofevents may need modifying (U. Retnasabapa-thy, unpublished data, this laboratory). A syn-chronized Caulobacter (NCIB 9083) swarm cellpopulation was gently filtered onto a cellulosetriacetate membrane, the membrane was in-verted, and preconditioned medium was passedthrough at a flow rate of 2 ml/min. As theattached population grew and differentiated(Fig. 1Oa), the effluent was monitored for totalcell numbers against time. The data obtained(Fig. lOb) quantified the number of swarm cellsproduced per attached mother cell and gave thetime required for each turn ofthe cycle; i.e., theprecise history of the attached, synchronizedmother cell population was known.
Electron microscopic examination of the at-tached mother cells at various times showedthat in Caulobacter (NCIB 9083) cross bandsdid not appear until after the third or fourthgeneration and that there was no subsequentcorrelation between swarm cell production andcross band formation.
(ii) Cells are probably mortal; they can un-dergo only a limited number of reproductivecycles (e.g., R. vannielii in one phenotypic form[see Part B] only gives rise to four daughtercells).
(iii) Cell division may be asymmetrical.(iv) Complex internal membranous bodies of
obvious asymmetry may be formed, as con-straints on pattern are not imposed by interca-lary growth and subsequent division processes(e.g., Nitrobacter agilis [62] and Rhodopseu-domonas palustris [21]). These membranousbodies are formed de novo in the swarmer ordaughter cell.
a
15
b
CI0
6C5
1 2 3
0 2 3 4 5 6
Time (hours)
FIG. 10. (a) Schematic illustration of the succes-sive stages in the life cycle ofan individual Caulobac-ter cell attached to a cellulose triacetate membrane,leading to the elution of the daughter cell. With asynchronized population, sequential waves ofswarmcells appear in the effluent. (b) Cell number permilliliter of effluent plotted against time. The inter-peak distances indicate the time taken for successivewaves ofswarm cell production. The number ofpeaksgives the number ofswarm cells produced per mothercell (U. Retnasabapathy, unpublished, this labora-tory).
(v) The swarmer or daughter cell, at division,is immature; it has to form internal structuresand filament or budding structures to achievephysiological and structural parity with themother cell that gave rise to it in the first place.The term "siblings" for such cells is seen to beinappropriate. The parent-offspring terminol-ogy used is considered to reflect more accu-rately relationships of the cells at division.With such bacteria, the term sibling now de-scribes the relationship between the daughtercells produced by one mother cell.
(vi) The adoption of a polar growth mecha-nism means that the cell is polarized. In otherwords, such bacteria now have a "front" endand a "back" end. This orientation can beplainly discerned by looking at an R. palustrismother and daughter cell (Fig. 9). Reproductionin this instance always takes place from the one(anterior) pole; holdfast and flagellum are al-
VOL. 41, 1977

762 WHITTENBURY AND DOW
ways at the posterior pole, and each daughtercell formed proves to be a mirror image of themother cell-in other words, the pole of thedaughter cell adjacent to the mother cell's re-productive pole is the pole that eventually givesrise to that cell's first daughter cell.Organisms growing by obligate polar proc-
esses clearly have a potential for morphoge-netic evolution not afforded to cells that grow,for the most part, by intercalary processes. Thiscan be claimed to have led to the evolution of acomplex morphogenesis in some bacteria (anincreasing pattern of morphological complexityin budding bacteria is outlined in Fig. 9). R.vannielii (see Part B) is probably the most spe-cialized example of this form of morphologicaldevelopment.
Budding Bacteria as Models for the Study ofMorphogenesis and Differentiation
All of these features, polar growth, polariza-tion of the cell, and the mother/daughter cellcycles, involve the regulation of temporally re-lated sequences of morphogenetic events, thusunderlining the suitability of polarly growingbacteria as models for studies on morphogene-sis and differentiation.Dimorphic bacteria, such as Caulobacter, are
already being explored as models of differentia-tion and morphogenesis because they undergowell-defined, microscopically distinct eventsthroughout the cell cycle (Fig. 11 and 12) whichcan be linked with distinct molecular eventsand control systems relevant to morphogenesisand differentiation (15, 16, 43, 57, 74). Variousmarker systems have already been employed tofollow these events in Caulobacter, e.g., fla-gella shedding and bacteriophage adsorption,both of which accurately mark stages in the cellcycle (1, 75, 77). Budding bacteria offer similarpossibilities for study. However, they have sev-eral unique advantages, and these we wish toemphasize in this monograph with particularreference to R. vannielii.
PART B-BIOLOGY OF THE CELLCYCLES OF R. VANNIELH
Review of Published Information on R.vannielii
A brief synopsis is given of the informationaccumulated on R. vannielii since its isolationby Duchow and Douglas (22). As will be evi-dent, most of the information refers to physio-logical and biochemical activities and not toproperties directly relevant to development.R. vannielii was shown to be a photohetero-
trophic bacterium of unusual morphology con-
l
4
A
it
w ^daughtercell
Imother
cell
i, -
cross stal
band I
5
holdfast
2
3,4
FIG. 11. Diagrammatic representation of the cellcycle of Caulobacter species. Starting from the motileswarm cell stage, the temporal sequence of morpho-logical events is as follows. Maturation - (1) Loss ofmotility, flagella shedding (77); (2) stalk synthesis;(3) swarm cell formation; (4) formation of the mem-branous organelle at the pole opposite the stalk.Daughter cell synthesis -(5) flagella and holdfastsynthesis; (6) cell division to yield a stalked mothercell and a motile swarm cell. On release, the swarmcell follows the above cycle, while the stalked motherimmediately initiates daughter cell formation; i.e.,there is no requirement for the maturation phase. Ithas recently been reported (80) that the mother cellsynthesizes a cross band after each cycle of replica-tion, cross band formation acting, therefore, as ageneration marker.
sisting of ovoid cells (2 to 3 am by 1 ,um) joinedby filaments (0.3 ,um by 2 to >30 /.m), whichmay be branched, thus forming ramifyinggroups of cells. Subsequently, Murray andDouglas (55) revealed that reproduction was bya system that resembled budding, the cellsbeing separated from one another by a crosswall laid down in the filament. Nonstalked,peritrichously flagellated cells have been re-ported to occur in early exponential cultures(20), but their mode of formation and subse-quent development was not reported.More recently, angular, heat-resistant bodies
(termed, by us, "exospores") have been ob-served in several freshwater isolates (27). Proc-esses leading to their formation and germina-tion have not been documented.The biochemical and physiological properties
of R. vannielii are given in Tables 1 to 6. Theresulting impression is that this organism isbiochemically similar in many respects to other
BACTERIOL. REV.
I
k
VOL. 41, 1977
photoheterotrophs. The obvious gap in informa-tion concerns the biology ofthe cell cycle, whichis the subject of our studies described below.
Enrichment, Isolation, and CultivationIn devising growth conditions and isolation
procedures for strains of R. vannielii, proper-ties consistent with the use of the species as amodel for morphogenesis and differentiation
MORPHOGENESIS OF R. VANNIELII 763
were taken into account. This meant thatgrowth on defined media was considered to beessential and that the carbon source also had tobe the energy source when the organism was tobe grown chemoorganotrophically under aero-bic conditions. Selective enrichment by sub-strate variation (90) was employed and, of thevarious enrichment media devised, malatesalts medium (NH4Cl, 0.5 g; MgSO4 -7H2O, 0.4g; CaCl2 2H2O, 0.05 g; NaCl, 0.4 g; sodiumhydrogen malate, 1.5 g per liter of distilledwater, adjusted with KOH; and 50 ml of 0.1 Mphosphate buffer per liter, added aseptically) atpH 5.5 or 6.0 was the most successful. Incuba-tion was in glass bottles (100 ml) at 300C withan incident light intensity of 1,000 lx (94); 50 mlof medium was added, and the gas atmospherewas dinitrogen (oxygen free). Approximately
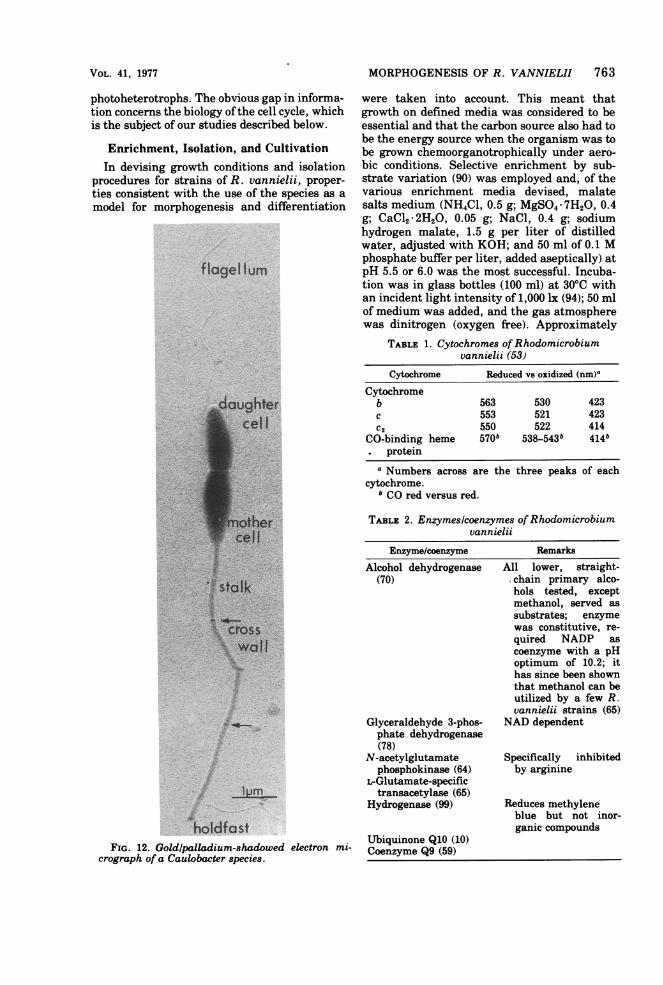
TABLE 1. Cytochromes of Rhodomicrobiumvannielii (53)
Cytochrome Reduced vs oxidized (nm)0Cytochrome
b 563 530 423c 553 521 423C2 550 522 414
CO-binding heme 570b 538-543b 414bprotein
a Numbers across are the three peaks of eachcytochrome.
I CO red versus red.
TABLz 2. Enzymes/coenzymes of Rhodomicrobiumvannielii
Enzyme/coenzyme Remarks
Alcohol dehydrogenase(70)
Glyceraldehyde 3-phos-phate dehydrogenase(78)
N-acetylglutamatephosphokinase (64)
i-Glutamate-specifictransacetylase (65)
Hydrogenase (99)
FIG. 12. Goldlpalladium-shadowed electron mi- Coequionme Q91(059()crograph of a Caulobacter species.
All lower, straight-chain primary alco-hols tested, exceptmethanol, served assubstrates; enzymewas constitutive, re-quired NADP ascoenzyme with a pHoptimum of 10.2; ithas since been shownthat methanol can beutilized by a few R.vannielii strains (65)
NAD dependent
Specifically inhibitedby arginine
Reduces methyleneblue but not inor-ganic compounds

764 WHITTENBURY AND DOW
TABLE 3. Lipid composition ofRhodomicrobiumvannielii (58, 59, 70)
% of dry wt in:Compound
Cells P-lipids
Total P-lipids 4.2Sulfolipids 0.01Steroids NDaLipo amino acid (o-ornithine 46.5
ester of phosphatidylglyc-erol)
Phosphatidylcholine 26.5Phosphatidylglycerol 9.7bis-Phosphatidic acid 6.7Phosphatidic acid 1.8Phosphatidylethanolamine 4.5Lysophosphatidyl glycerol-o- 3.2ornithine ester
Phosphatidyl-N-methyl- NDethanolamine
Phosphatidyl-N,N'-dimethyl- NDethanolamine
Cardiolipin NDN,N-ornithine amide of an 0.95
unidentified fatty acida ND, Not detected.
TABLE 4. Fatty acids ofRhodomicrobiumvannielii (59)
Fatty acid Remarks
Vaccenic acid (11-oc- Largest single componenttadecenoic acid) (90%) of both simple and
complex lipidsExtractable lipids Only straight-chain satu-
(room tempera- rated and unsaturatedture, organic sol- even-numbered fattyvents) acids
Nonextractable Even- and odd-numberedbound lipids saturated and unsatu-
rated straight-chainfatty acids, cyclopro-pane-, branched-, and a-and 8-hydroxy fattyacids
50% of all enrichment cultures inoculated withfreshwater sediment (surface layer) proved tocontain Rhodomicrobium species (detectablewithin 1 to 2 weeks). Subsequent isolation andpurification was by dilution of the enrichmentmedium into agar deeps.The routine culture medium was the enrich-
ment medium plus 1.5 g (wt/vol) sodium pyru-
vate, the pH being adjusted to 6.8. This me-
dium was chosen on the basis of growth rateand cell density achieved. Ofthe various carbonsources, tested individually and as mixtures,malate and pyruvate were the best individ-ually, whereas a mixture of the two surpassedeither singly (Fig. 13).
BACTERIOL. REV.
Pure cultures were grown in a variety ofglass vessels, which were sealed with rubberserum caps (Suba-Seals, William Freeman andCo. Ltd., Barnsley, Yorkshire) flushed with oxy-gen-free nitrogen and incubated with shakingat 30'C under constant illumination from tung-sten lamps (incident light intensity of 2,000 lx).The gaseous environment was assayed chroma-tographically (32).Several of the isolated strains were compared
with the neotype culture (ATCC 17100) bothdirectly and with published results. Rhodomi-crobium strain 5 (Rm 5) was used in all subse-quent experiments.
Physiological and Biochemical PropertiesCharacterization of DNA. Determination of
the guanine-plus-cytosine mole percents by an-alytical ultracentrifuge techniques using neu-tral CsCl density gradients (48) gave results of62.5% for strain Rm 5 and 63% for R. vannieliiATCC 17100. Thermal denaturation curves (47)were characteristic of a single deoxyribonucleicacid (DNA) population and gave results of62.5% in both cases. No plasmid or other satel-lite DNA was detected.Photopigments. In the exponential growth
phase, all strains had similar absorption char-acteristics (see Fig. 40). Isolation and analysisof the photopigments confirmed the presence ofbacteriochlorophyll a and carotenoids of thenormal spirilloxanthin series, with rhodopin asthe major constituent. 8-Carotene is also pres-ent in considerable quantities (11, 39, 46, 69,92).Cytochromes. The cytochrome content of
both strains (strain Rm 5 and R. vannieliiATCC 17100) grown photosynthetically wasidentical and correlated closely with publishedobservations (Table 1).Growth physiology. Rhodomicrobium spe-
cies growing photosynthetically require anelectron donor, a terminal electron acceptor,and a source of cell carbon. Table 5 lists thesubstrates used by strain Rm 5. There was norequirement for growth factors. However, ribo-flavin has been reported to be a growth stimu-lant (86).The addition of C02, furnished as bicarbon-
ate, to the culture medium is only essential inthose instances where CO2 cannot be generatedfrom the organic electron donor; e.g., malateand pyruvate yield considerable amounts ofCO2 during growth and, consequently, the ad-dition of bicarbonate is unnecessary. However,if CO2 is removed from such a culture by vigor-ous gassing with oxygen-free N2, growth onlyproceeds when carbon dioxide is also added; thiswas shown not to be a pH effect.
MORPHOGENESIS OF R. VANNIELII 765
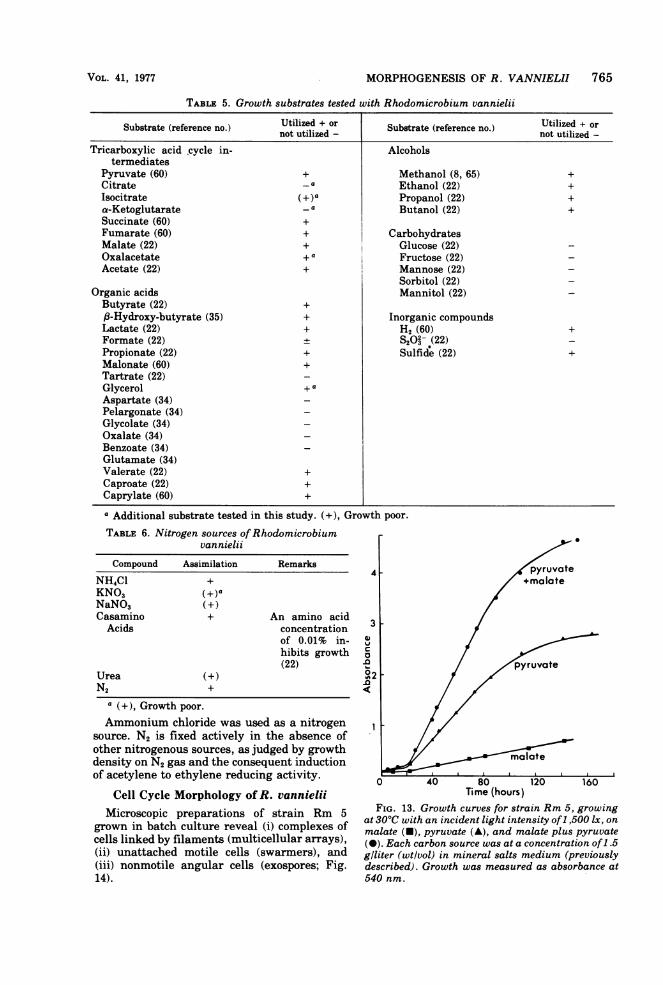
TABLE 5. Growth substrates tested with Rhodomicrobium vannielii
Substrate (reference no.) Utilized + or Substrate (reference no.) Utilized + ornot utilized - not utilized -
Tricarboxylic acid cycle in- Alcoholstermediates
Pyruvate (60) + Methanol (8, 65) +Citrate - a Ethanol (22) +Isocitrate (+)a Propanol (22) +a-Ketoglutarate -a Butanol (22) +Succinate (60) +Fumarate (60) + CarbohydratesMalate (22) + Glucose (22)Oxalacetate + a Fructose (22)Acetate (22) + Mannose (22)
Sorbitol (22)Organic acids Mannitol (22)Butyrate (22) +,f-Hydroxy-butyrate (35) + Inorganic compoundsLactate (22) + H2 (60) +Formate (22) S2O3 (22)Propionate (22) + Sulfide (22) +Malonate (60) +Tartrate (22)Glycerol + a
Aspartate (34)Pelargonate (34)Glycolate (34)Oxalate (34)Benzoate (34)Glutamate (34)Valerate (22) +Caproate (22) +Caprylate (60) +a Additional substrate tested in this study. (+), Growth poor.TABLE 6. Nitrogen sources ofRhodomicrobium
vannielii
Compound Assimilation RemarksNH4Cl +KNO3 (+ )aNaNO3 (+)Casamino + An amino acidAcids concentration
of 0.01% in-hibits growth(22)
Urea (+)N2 +
a (+), Growth poor.Ammonium chloride was used as a nitrogen
source. N2 is fixed actively in the absence ofother nitrogenous sources, as judged by growthdensity on N2 gas and the consequent inductionof acetylene to ethylene reducing activity.
Cell Cycle Morphology of R. vannieliiMicroscopic preparations of strain Rm 5
grown in batch culture reveal (i) complexes ofcells linked by filaments (multicellular arrays),(ii) unattached motile cells (swarmers), and(iii) nonmotile angular cells (exospores; Fig.14).
41
3
v
.0.0
.0
Time (hours)FIG. 13. Growth curves for strain Rm 5, growing
at 30°C with an incident light intensity of1,500 lx, onmalate (U), pyruvate (A), and malate plus pyruvate(-). Each carbon source was at a concentration of1.5giliter (wtlvol) in mineral salts medium (previouslydescribed). Growth was measured as absorbance at540 nm.
VOL. 41, 1977
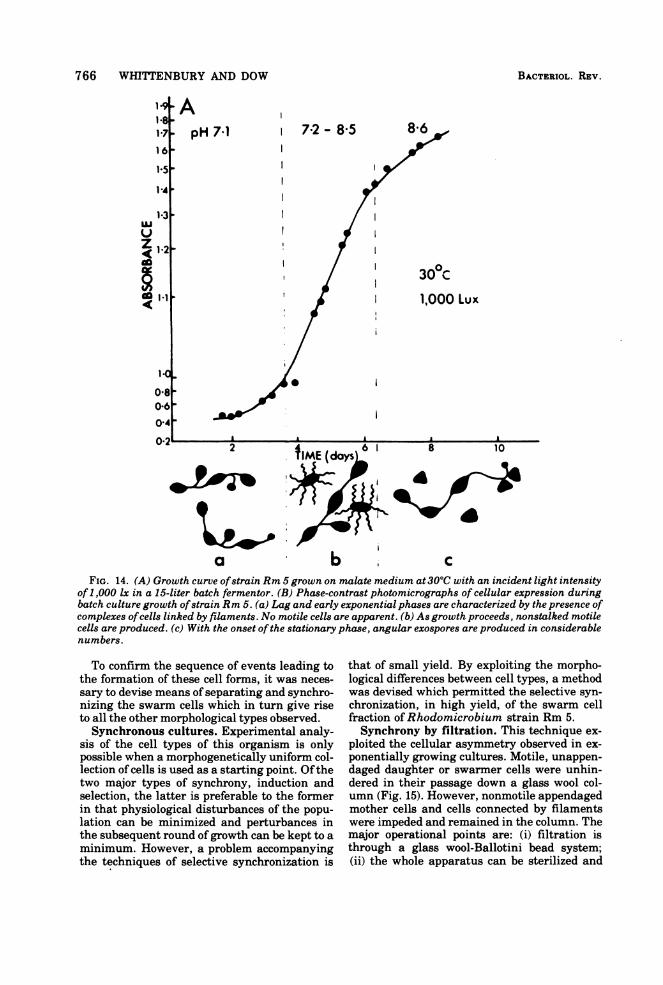
766 WHITTENBURY AND DOW
ApH 7-1 7-2 - 8*5 8-6
V
Z~~12I ~~~~~0
~~~I.1~~~~~30C1,000 Lux
080-6040; I . I.
fIME (dayS 8 10
a b cFIG. 14. (A) Growth curve ofstrain Rm 5 grown on malate medium at 300C with an incident light intensity
of1,000 lx in a 15-liter batch fermentor. (B) Phase-contrast photomicrographs of cellular expression duringbatch culture growth ofstrain Rm 5. (a) Lag and early exponential phases are characterized by the presence ofcomplexes ofcells linked by filaments. No motile cells are apparent. (b) As growth proceeds, nonstalked motilecells are produced. (c) With the onset ofthe stationary phase, angular exospores are produced in considerablenumbers.
To confirm the sequence of events leading tothe formation of these cell forms, it was neces-sary to devise means of separating and synchro-nizing the swarm cells which in turn give riseto all the other morphological types observed.Synchronous cultures. Experimental analy-
sis of the cell types of this organism is onlypossible when a morphogenetically uniform col-lection of cells is used as a starting point. Ofthetwo major types of synchrony, induction andselection, the latter is preferable to the formerin that physiological disturbances of the popu-lation can be minimized and perturbances inthe subsequent round ofgrowth can be kept to aminimum. However, a problem accompanyingthe techniques of selective synchronization is
that of small yield. By exploiting the morpho-logical differences between cell types, a methodwas devised which permitted the selective syn-chronization, in high yield, of the swarm cellfraction of Rhodomicrobium strain Rm 5.Synchrony by filtration. This technique ex-
ploited the cellular asymmetry observed in ex-ponentially growing cultures. Motile, unappen-daged daughter or swarmer cells were unhin-dered in their passage down a glass wool col-umn (Fig. 15). However, nonmotile appendagedmother cells and cells connected by filamentswere impeded and remained in the column. Themajor operational points are: (i) filtration isthrough a glass wool-Ballotini bead system;(ii) the whole apparatus can be sterilized and
BACTERIOL. REV.
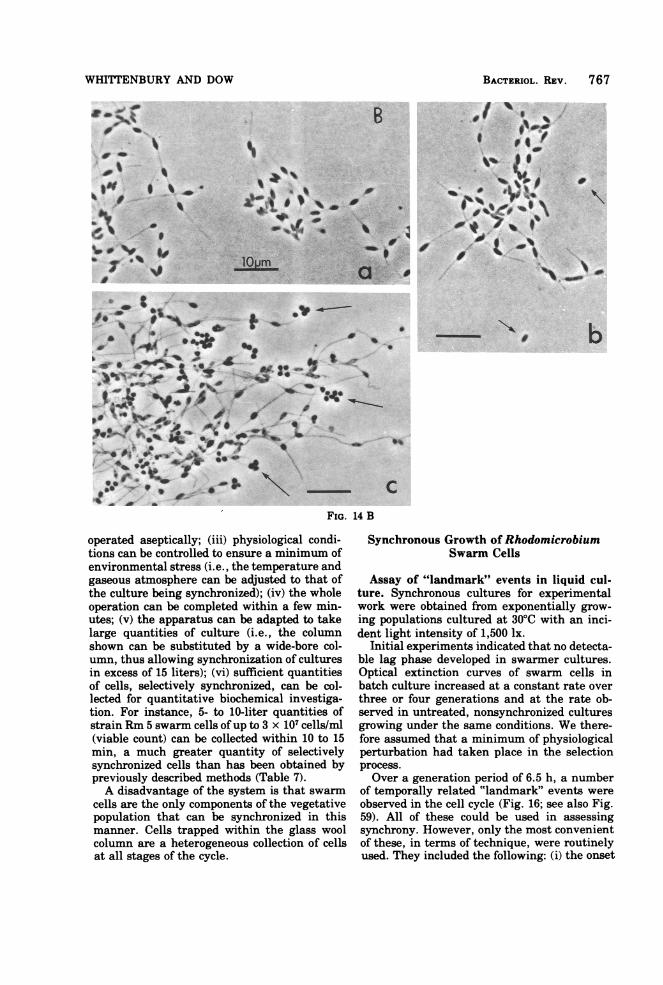
WHITTENBURY AND DOW
FIG. 14 B
operated aseptically; (iii) physiological condi-tions can be controlled to ensure a minimum ofenvironmental stress (i.e., the temperature andgaseous atmosphere can be adjusted to that ofthe culture being synchronized); (iv) the wholeoperation can be completed within a few min-utes; (v) the apparatus can be adapted to takelarge quantities of culture (i.e., the columnshown can be substituted by a wide-bore col-umn, thus allowing synchronization of culturesin excess of 15 liters); (vi) sufficient quantitiesof cells, selectively synchronized, can be col-lected for quantitative biochemical investiga-tion. For instance, 5- to 10-liter quantities ofstrain Rm 5 swarm cells ofup to 3 x 107 cells/ml(viable count) can be collected within 10 to 15min, a much greater quantity of selectivelysynchronized cells than has been obtained bypreviously described methods (Table 7).A disadvantage of the system is that swarm
cells are the only components of the vegetativepopulation that can be synchronized in thismanner. Cells trapped within the glass woolcolumn are a heterogeneous collection of cellsat all stages of the cycle.
Synchronous Growth of RhodomicrobiumSwarm Cells
Assay of "landmark" events in liquid cul-ture. Synchronous cultures for experimentalwork were obtained from exponentially grow-ing populations cultured at 30TC with an inci-dent light intensity of 1,500 lx.
Initial experiments indicated that no detecta-ble lag phase developed in swarmer cultures.Optical extinction curves of swarm cells inbatch culture increased at a constant rate overthree or four generations and at the rate ob-served in untreated, nonsynchronized culturesgrowing under the same conditions. We there-fore assumed that a minimum of physiologicalperturbation had taken place in the selectionprocess.Over a generation period of 6.5 h, a number
of temporally related "landmark" events wereobserved in the cell cycle (Fig. 16; see also Fig.59). All of these could be used in assessingsynchrony. However, only the most convenientof these, in terms of technique, were routinelyused. They included the following: (i) the onset
BACTERIOL. Rim 767
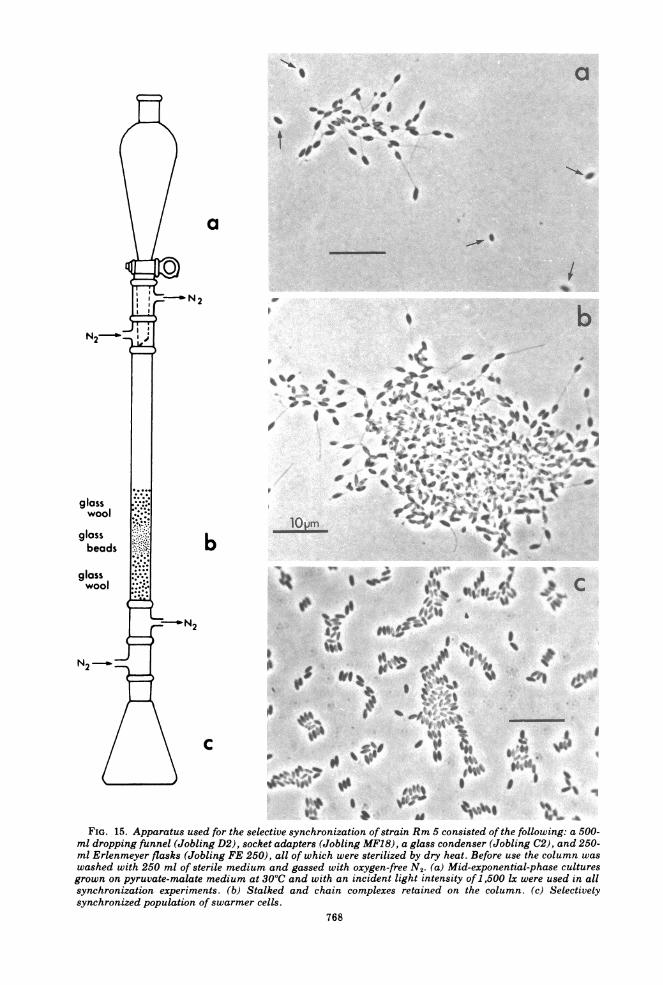
a
ki
N 2
wool~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~l
N2 -aX:'
gloss b
beads
wool 1~~~~~~~~~~~~~..Ktt,M aiD;< 5 thN2 J -w-Aae#E
FIG. 15. Apparatus used for the selective synchronization of strain Rm 5 consisted of the following: a 500-ml dropping funnel (Jobling D2), socket adapters (Jobling MF18), a glass condenser (Jobling C2), and 250-ml Erlenmeyer flasks (Jobling FE 250), all of which were sterilized by dry heat. Before use the column waswashed with 250 ml of sterile medium and gassed with oxygen-free N2. (a) Mid-exponential-phase culturesgrown on pyruvate-malate medium at 30°C and with an incident light intensity of 1,500 Ix were used in allsynchronization experiments. (b) Stalked and chain complexes retained on the column. (c) Selectivelysynchronized population of swarmer cells.
768

MORPHOGENESIS OF R. VANNIELII 769
TABLE 7. Cell yields by selective synchronization techniques
Organism Synchronization procedure Volume of synchronized Viable cells/ml ReferencecultureCaulobacter (i) Filtration Not given 72
(ii) Repeated centrif- 2-3 ml Not given 81ugation
(iii) Adhesion of 25 ml 1 X 107-3 x 107 15"mother" cell
(iv) Adhesion of 150 ml 5.9 x 106 80"mother" cell
Hyphomicrobium Centrifugation and 30 ml 2 X 107 52filtration
Rhodopseudomo- Density gradient cen- 2-3 ml 1.4 x 109 93nas palustris trifugation
Rhodomicrobium Filtration 100 ml-15 liters 1 x 107-2.5 x 107Bacillus subtilis Filtration 1-2% of initial 71
populationEscherichia coli Density gradient cen- 1-2 ml from a 15- 4 x 108 50
trifugation ml gradient
and period of tube formation as reflected in achange of slope when following the absorptioncharacteristics of a synchronized populationwith time (Fig. 16). (ii) The constant pattern ofevents recorded in Fig. 16 were confirmed byplotting histograms of cell shape distribution ina large number of microscopic fields duringsynchronous growth (Fig. 17). (iii) Cell volumechanges during synchronous growth added fur-ther confirmation to the above (Fig. 18). One-step growth curves cannot be used with cul-tures in this morphogenetic form, since daugh-ter cells are not released at division. Viablecounts, consequently, do not double at cell divi-sion.
Vegetative Cell CycleSwarm cell growth leading to the formation
of a first-generation cell was classified into twomajor sequences-maturation and reproduc-tion.Maturation sequence. This occurred (under
the growth conditions described) over a periodof 130 to 150 min and is subdivided into threedistinctive phases.
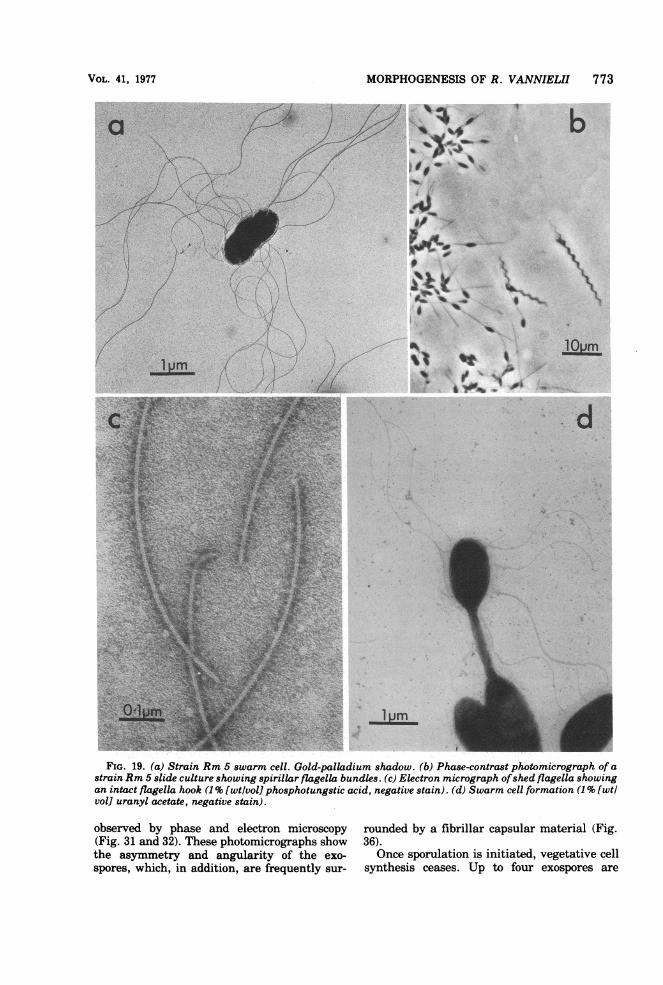
(i) Motile period. This period lasts for 20 to 30min. The swarm cell is peritrichously flagel-lated and has dimensions of 1.5 by 0.8 ,4m (Fig.19a).
(ii) Flagella shedding period. The flagellaare lost over a 15-min period, and all cells be-come permanently immotile. Flagella are shedintact; i.e., they retain the flagella hook (Fig.19c). Only newly formed daughter cells bearflagella (Fig. 19d). When cells are grown inslide cultures, flagella shedding can lead to theappearance of flagella bundles (Fig. 19b) analo-gous to those described for Vibrio alginolyticus(89).
(iii) Terminal maturation period. During
this phase, no obvious external morphologicalchanges take place. This period lasts for a fur-ther 100 to 120 min.
Reproductive sequence. This takes placeover 200 to 240 min (Fig. 16 and 21 to 24) and isdivisible into three phases.
(i) Filament formation phase. During thisphase, filament synthesis occurs at one or, occa-sionally, at both poles ofthe cell and requires 40to 50 min for completion (Fig. 16, 21, 22, 24).
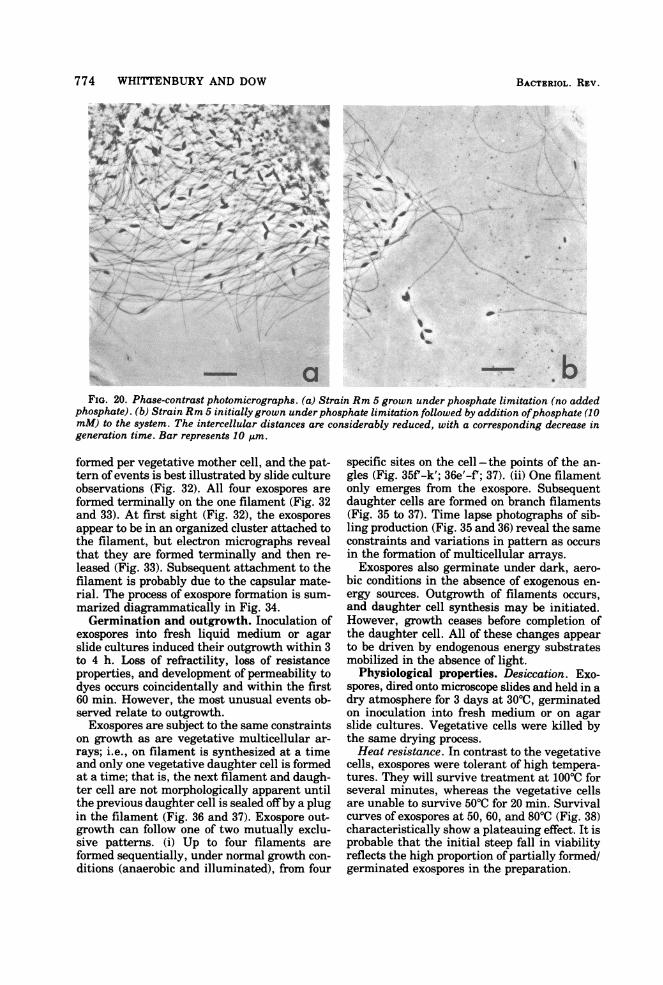
Both filament length and the proportion oftime of the cell cycle occupied in filament syn-thesis can be lengthened or shortened bychanges in the nutrient status of the medium.This effect is particularly obvious when phos-phate is limiting. Medium that has no addedphosphate results in intercell distances in ex-cess of 30 um with a cell generation time of 18to 20 h. Subsequent addition of phosphate (10mM) to such cultures results in a burst of cellsynthesis and replication with intercell dis-tances drastically reduced (2 to 3 ,um) and areduced generation time of 5 to 6 h (Fig. 20).
(ii) Bud synthesis. After 220 to 250 min frominitiation of the cycle, bud synthesis begins.The distal end of the filament begins to swelldetectably, and the process lasts for 150 to 180min. Only one bud is formed at a time, irrespec-tive ofthe number of filaments possessed by thecell.
(iii) Daughter cell completion. This is theperiod when no increase in daughter cell sizetakes place, but physiological separation fromthe mother cell occurs. This may be either bybinary fission at the junction of filament anddaughter cell, or by the synthesis of a plugwithin the filament a short distance from thedaughter cell (Fig. 21i and j, and 25). Thisdistance is reasonably constant and is not afunction of overall filament length (Fig. 26).
VOL. 41, 1977
770 WHITTENBURY AND DOW
A
-p - . ~~~~~~~~~~~~~~~~~culture,E_ v- .^}
G0 ~~~~W_0 2 3 4 5 6 7
TIME (hours)
_<~~~~~~~~~~I
46
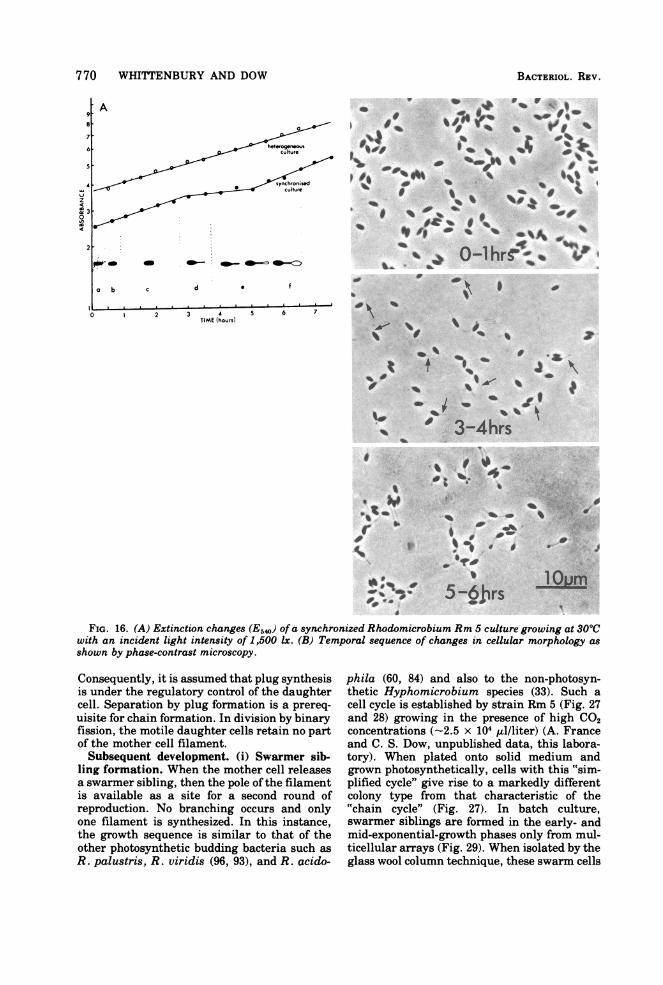
FIG. 16. (A) Extinction changes (E5,) ofa synchronized Rhodomicrobium Rm 5 culture growing at 30'Cwith an incident light intensity of 1,500 Ix. (B) Temporal sequence of changes in cellular morphology asshown by phase-contrast microscopy.
Consequently, it is assumed that plug synthesisis under the regulatory control of the daughtercell. Separation by plug formation is a prereq-uisite for chain formation. In division by binaryfission, the motile daughter cells retain no partof the mother cell filament.Subsequent development. (i) Swarmer sib-
ling formation. When the mother cell releasesa swarmer sibling, then the pole of the filamentis available as a site for a second round ofreproduction. No branching occurs and onlyone filament is synthesized. In this instance,the growth sequence is similar to that of theother photosynthetic budding bacteria such asR. palustris, R. viridis (96, 93), and R. acido-
phila (60, 84) and also to the non-photosyn-thetic Hyphomicrobium species (33). Such acell cycle is established by strain Rm 5 (Fig. 27and 28) growing in the presence of high C02concentrations (-2.5 x 104 1.l/liter) (A. Franceand C. S. Dow, unpublished data, this labora-tory). When plated onto solid medium andgrown photosynthetically, cells with this "sim-plified cycle" give rise to a markedly differentcolony type from that characteristic of the"chain cycle" (Fig. 27). In batch culture,swarmer siblings are formed in the early- andmid-exponential-growth phases only from mul-ticellular arrays (Fig. 29). When isolated by theglass wool column technique, these swarm cells
BACTERIOL. REV.
MORPHOGENESIS OF R. VANNIELII 771
reproduce by chain formation; i.e., they giverise to rough colonies. However, a low percent-age (1 to 2%) of swarmer siblings isolated fromthe late-exponential-growth phase, when theC02 concentration is highest (2.5 x 104pl/liter), follow a "simplified cycle"; i.e., theygive rise to "smooth" colonies.
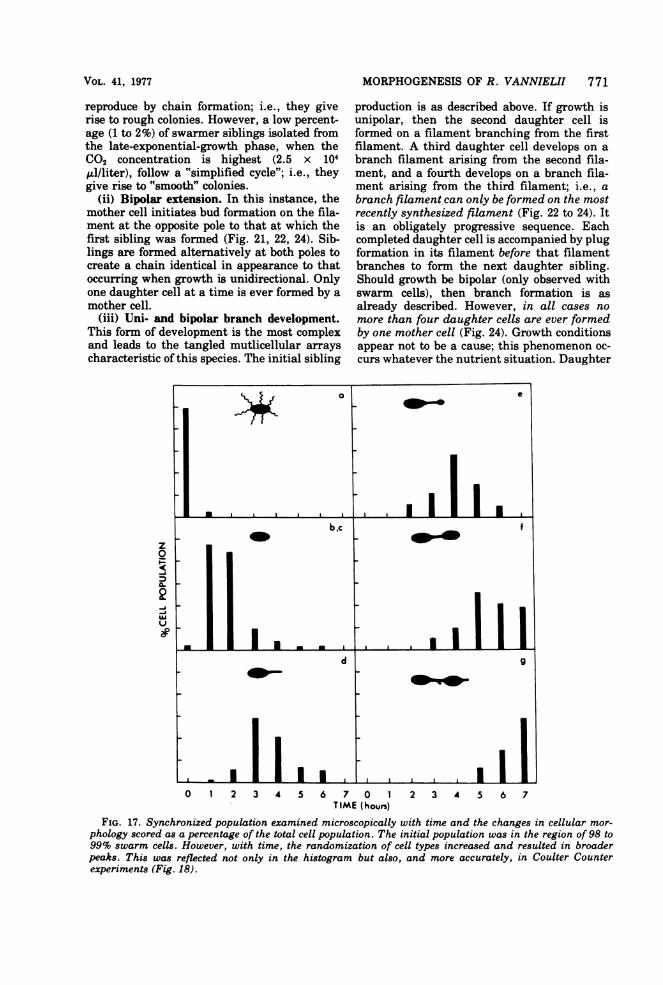
(ii) Bipolar extension. In this instance, themother cell initiates bud formation on the fila-ment at the opposite pole to that at which thefirst sibling was formed (Fig. 21, 22, 24). Sib-lings are formed alternatively at both poles tocreate a chain identical in appearance to thatoccurring when growth is unidirectional. Onlyone daughter cell at a time is ever formed by amother cell.
(iii) Uni- and bipolar branch development.This form of development is the most complexand leads to the tangled mutlicellular arrayscharacteristic of this species. The initial sibling
production is as described above. If growth isunipolar, then the second daughter cell isformed on a filament branching from the firstfilament. A third daughter cell develops on abranch filament arising from the second fila-ment, and a fourth develops on a branch fila-ment arising from the third filament; i.e., abranch filament can only be formed on the mostrecently synthesized filament (Fig. 22 to 24). Itis an obligately progressive sequence. Eachcompleted daughter cell is accompanied by plugformation in its filament before that filamentbranches to form the next daughter sibling.Should growth be bipolar (only observed withswarm cells), then branch formation is asalready described. However, in all cases nomore than four daughter cells are ever formedby one mother cell (Fig. 24). Growth conditionsappear not to be a cause; this phenomenon oc-curs whatever the nutrient situation. Daughter
0
2
-
. . I UVd g
0 1 2 3 4 5 6 7 0 1 2 3 4 5 6 7TIME (hours)
FIG. 17. Synchronized population examined microscopically with time and the changes in cellular mor-phology scored as a percentage of the total cell population. The initial population was in the region of 98 to99% swarm cells. However, with time, the randomization of cell types increased and resulted in broaderpeaks. This was reflected not only in the histogram but also, and more accurately, in Coulter Counterexperiments (Fig. 18).
VOL. 41, 1977
772 WHITTENBURY AND DOW
90
50
10
90
z
3>50
I.-
10
90
50
10
a ohrs f 6-5
AI sAI I
0o6 0-9 1-2 1-5 1-8 0o6 09 1.2 1-5 1 8CELL VOLUMEm"3
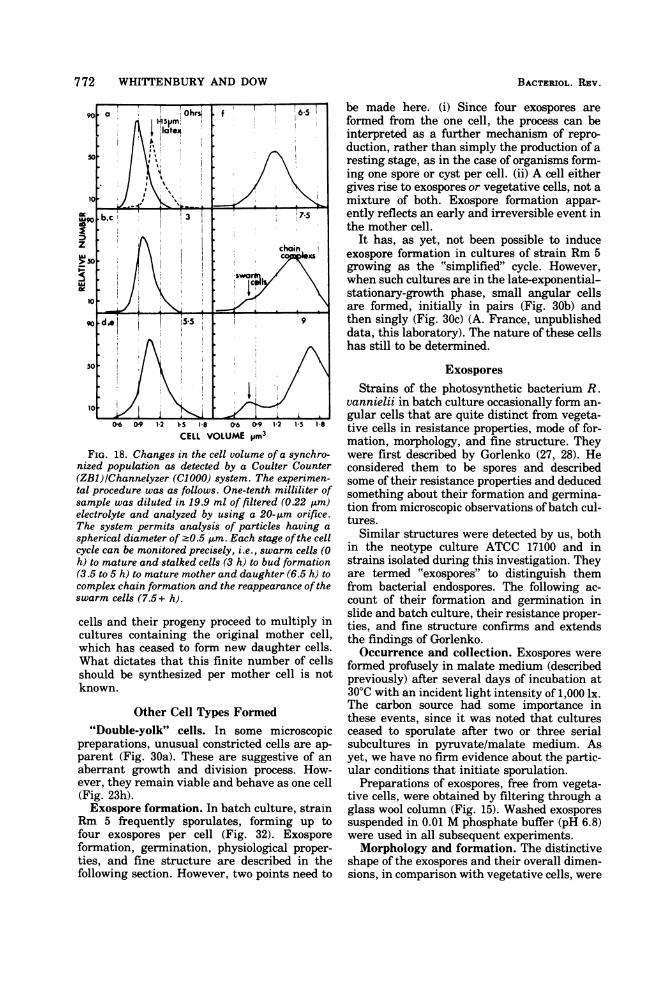
FIG. 18. Changes in the cell volume of a synchro-nized population as detected by a Coulter Counter(ZB1)/Channelyzer (C1000) system. The experimen-tal procedure was as follows. One-tenth milliliter ofsample was diluted in 19.9 ml of filtered (022 gm)electrolyte and analyzed by using a 20-gm orifice.The system permits analysis of particles having a
spherical diameter of -0.5 ,gm. Each stage ofthe cellcycle can be monitored precisely, i.e., swarm cells (0h) to mature and stalked cells (3 h) to bud formation(3.5 to 5 h) to mature mother and daughter (6.5 h) tocomplex chain formation and the reappearance of theswarm cells (7.5+ h).
cells and their progeny proceed to multiply incultures containing the original mother cell,which has ceased to form new daughter cells.What dictates that this finite number of cellsshould be synthesized per mother cell is notknown.
Other Cell Types Formed"Double-yolk" cells. In some microscopic
preparations, unusual constricted cells are ap-parent (Fig. 30a). These are suggestive of anaberrant growth and division process. How-ever, they remain viable and behave as one cell(Fig. 23h).Exospore formation. In batch culture, strain
Rm 5 frequently sporulates, forming up tofour exospores per cell (Fig. 32). Exosporeformation, germination, physiological proper-ties, and fine structure are described in thefollowing section. However, two points need to
be made here. (i) Since four exospores areformed from the one cell, the process can beinterpreted as a further mechanism of repro-duction, rather than simply the production of aresting stage, as in the case of organisms form-ing one spore or cyst per cell. (ii) A cell eithergives rise to exospores or vegetative cells, not amixture of both. Exospore formation appar-ently reflects an early and irreversible event inthe mother cell.
It has, as yet, not been possible to induceexospore formation in cultures of strain Rm 5growing as the "simplified" cycle. However,when such cultures are in the late-exponential-stationary-growth phase, small angular cellsare formed, initially in pairs (Fig. 30b)- andthen singly (Fig. 30c) (A. France, unpublisheddata, this laboratory). The nature of these cellshas still to be determined.
ExosporesStrains of the photosynthetic bacterium R.
vannielii in batch culture occasionally form an-gular cells that are quite distinct from vegeta-tive cells in resistance properties, mode of for-mation, morphology, and fine structure. Theywere first described by Gorlenko (27, 28). Heconsidered them to be spores and describedsome of their resistance properties and deducedsomething about their formation and germina-tion from microscopic observations ofbatch cul-tures.
Similar structures were detected by us, bothin the neotype culture ATCC 17100 and instrains isolated during this investigation. Theyare termed "exospores" to distinguish themfrom bacterial endospores. The following ac-count of their formation and germination inslide and batch culture, their resistance proper-ties, and fine structure confirms and extendsthe findings of Gorlenko.Occurrence and collection. Exospores were
formed profusely in malate medium (describedpreviously) after several days of incubation at30'C with an incident light intensity of 1,000 lx.The carbon source had some importance inthese events, since it was noted that culturesceased to sporulate after two or three serialsubcultures in pyruvate/malate medium. Asyet, we have no firm evidence about the partic-ular conditions that initiate sporulation.
Preparations of exospores, free from vegeta-tive cells, were obtained by filtering through aglass wool column (Fig. 15). Washed exosporessuspended in 0.01 M phosphate buffer (pH 6.8)were used in all subsequent experiments.Morphology and formation. The distinctive
shape of the exospores and their overall dimen-sions, in comparison with vegetative cells, were
BACTERIOL. REV.
MORPHOGENESIS OF R. VANNIELII 773
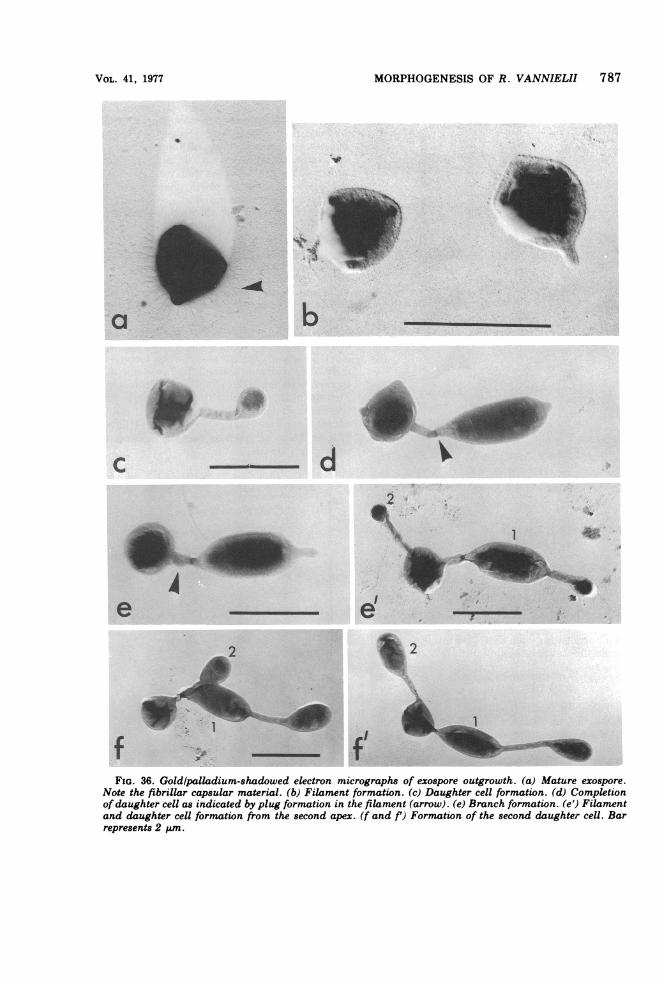
FIG. 19. (a) Strain Rm 5 swarm cell. Gold-palladium shadow. (b) Phase-contrast photomicrograph of astrain Rm 5 slide culture showing spirillar flagella bundles. (c) Electron micrograph ofshed flagella showingan intact flagella hook (1% [wtlvol] phosphotungstic acid, negative stain). (d) Swarm cell formation (1% [wtlvol] uranyl acetate, negative stain).
observed by phase and electron microscopy(Fig. 31 and 32). These photomicrographs showthe asymmetry and angularity of the exo-spores, which, in addition, are frequently sur-
rounded by a fibrillar capsular material (Fig.36).Once sporulation is initiated, vegetative cell
synthesis ceases. Up to four exospores are
VOL. 41, 1977
774 WHITTENBURY AND DOW
FIG. 20. Phase-contrast photomicrographs. (a) Strain Rm 5 grown under phosphate limitation (no addedphosphate). (b) Strain Rm 5 initially grown under phosphate limitation followed by addition ofphosphate (10mM) to the system. The intercellular distances are considerably reduced, with a corresponding decrease ingeneration time. Bar represents 10 pim.
formed per vegetative mother cell, and the pat-tern of events is best illustrated by slide cultureobservations (Fig. 32). All four exospores areformed terminally on the one filament (Fig. 32and 33). At first sight (Fig. 32), the exosporesappear to be in an organized cluster attached tothe filament, but electron micrographs revealthat they are formed terminally and then re-leased (Fig. 33). Subsequent attachment to thefilament is probably due to the capsular mate-rial. The process of exospore formation is sum-marized diagrammatically in Fig. 34.Germination and outgrowth. Inoculation of
exospores into fresh liquid medium or agarslide cultures induced their outgrowth within 3to 4 h. Loss of refractility, loss of resistanceproperties, and development of permeability todyes occurs coincidentally and within the first60 min. However, the most unusual events ob-served relate to outgrowth.Exospores are subject to the same constraints
on growth as are vegetative multicellular ar-rays; i.e., on filament is synthesized at a timeand only one vegetative daughter cell is formedat a time; that is, the next filament and daugh-ter cell are not morphologically apparent untilthe previous daughter cell is sealed offby a plugin the filament (Fig. 36 and 37). Exospore out-growth can follow one of two mutually exclu-sive patterns. (i) Up to four filaments areformed sequentially, under normal growth con-ditions (anaerobic and illuminated), from four
specific sites on the cell -the points of the an-gles (Fig. 35f-k'; 36e'-f; 37). (ii) One filamentonly emerges from the exospore. Subsequentdaughter cells are formed on branch filaments(Fig. 35 to 37). Time lapse photographs of sib-ling production (Fig. 35 and 36) reveal the sameconstraints and variations in pattern as occursin the formation of multicellular arrays.Exospores also germinate under dark, aero-
bic conditions in the absence of exogenous en-ergy sources. Outgrowth of filaments occurs,and daughter cell synthesis may be initiated.However, growth ceases before completion ofthe daughter cell. All of these changes appearto be driven by endogenous energy substratesmobilized in the absence of light.
Physiological properties. Desiccation. Exo-spores, dired onto microscope slides and held in adry atmosphere for 3 days at 300C, germinatedon inoculation into fresh medium or on agarslide cultures. Vegetative cells were killed bythe same drying process.Heat resistance. In contrast to the vegetative
cells, exospores were tolerant of high tempera-tures. They will survive treatment at 1000C forseveral minutes, whereas the vegetative cellsare unable to survive 50'C for 20 min. Survivalcurves of exospores at 50, 60, and 800C (Fig. 38)characteristically show a plateauing effect. It isprobable that the initial steep fall in viabilityreflects the high proportion of partially formed/germinated exospores in the preparation.
BACTERIOL. REV.
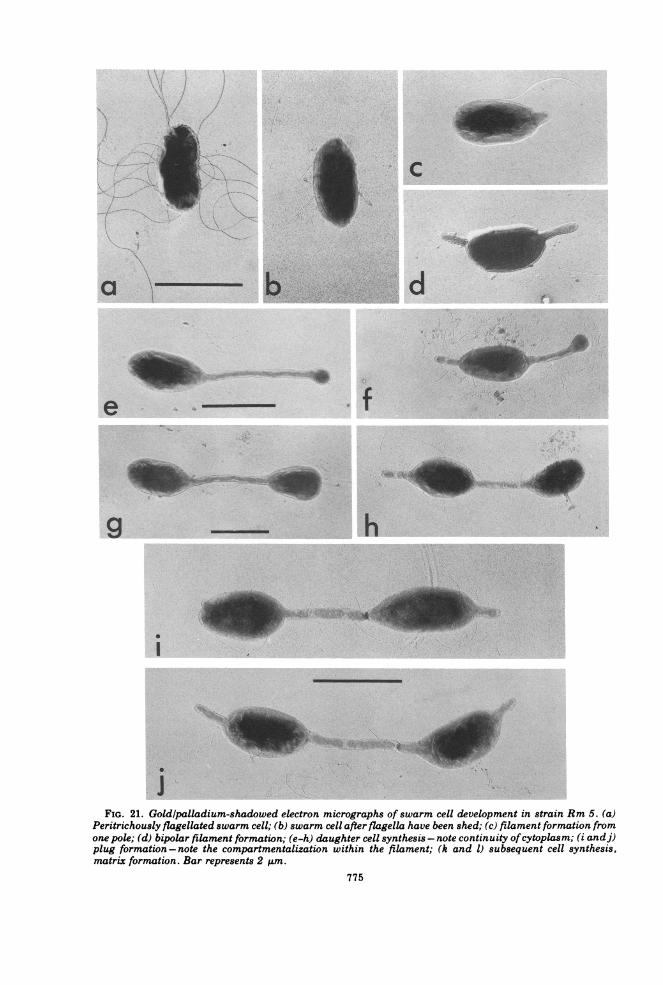
FIG. 21. Goldlpalladium-shadowed electron micrographs of swarm cell development in strain Rm 5. (a)Peritrichously flagellated swarm cell; (b) swarm cell after flagella have been shed; (c) filament formation fromone pole; (d) bipolar filament formation; (e-h) daughter cell synthesis - note continuity ofcytoplasm; (i andj)plug formation - note the compartmentalization within the filament; (k and 1) subsequent cell synthesis,matrix formation. Bar represents 2 ALm.
775
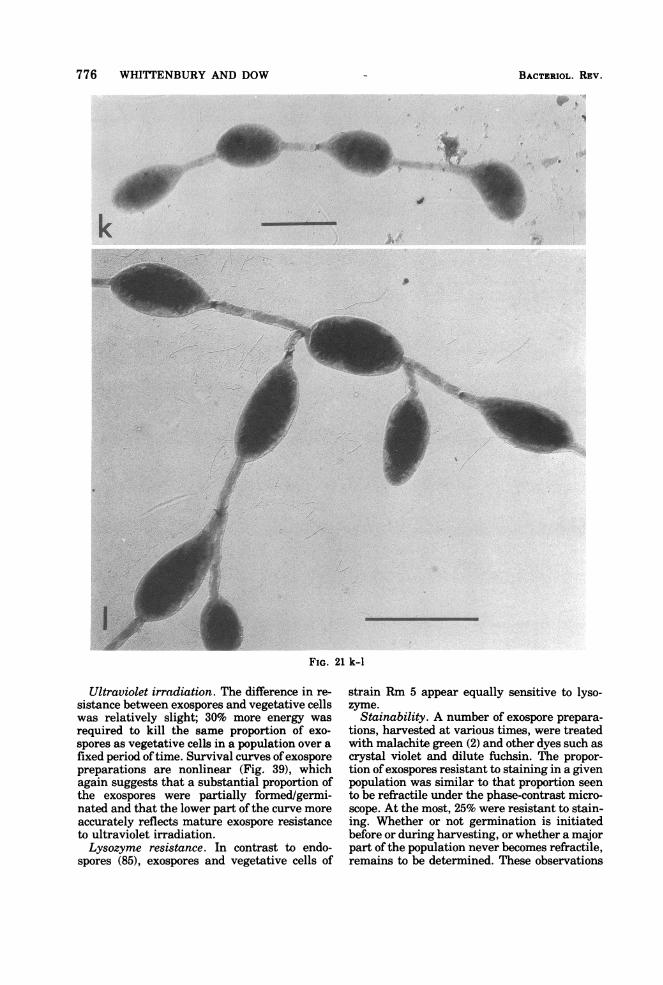
776 WHITTENBURY AND DOW
FIG. 21 k-l
Ultraviolet irradiation. The difference in re-sistance between exospores and vegetative cellswas relatively slight; 30% more energy wasrequired to kill the same proportion of exo-spores as vegetative cells in a population over afixed period oftime. Survival curves ofexosporepreparations are nonlinear (Fig. 39), whichagain suggests that a substantial proportion ofthe exospores were partially formed/germi-nated and that the lower part of the curve moreaccurately reflects mature exospore resistanceto ultraviolet irradiation.Lysozyme resistance. In contrast to endo-
spores (85), exospores and vegetative cells of
strain Rm 5 appear equally sensitive to lyso-zyme.
Stainability. A number of exospore prepara-tions, harvested at various times, were treatedwith malachite green (2) and other dyes such ascrystal violet and dilute fuchsin. The propor-tion ofexospores resistant to staining in a givenpopulation was similar to that proportion seento be retractile under the phase-contrast micro-scope. At the most, 25% were resistant to stain-ing. Whether or not germination is initiatedbefore or during harvesting, or whether a majorpart of the population never becomes refractile,remains to be determined. These observations
BACTIERIOL. REV.
MORPHOGENESIS OF R. VANNIELII 777
3
5
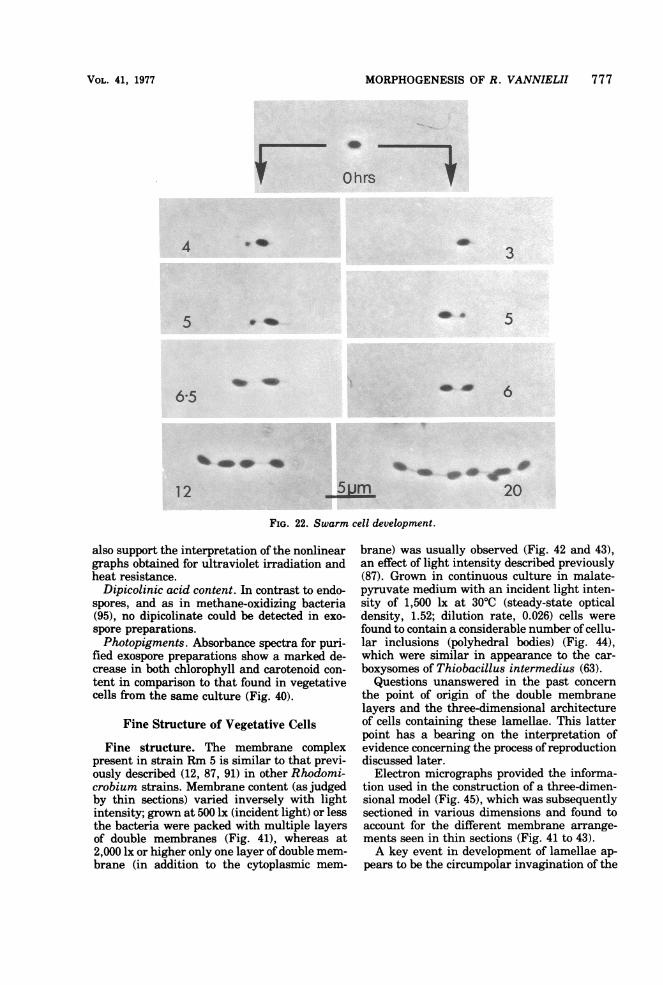
FIG. 22. Swarm cell development.
'ts-{., Et,@
'8- -or > {._r It ' <. s t ; v ¢-t < - B
aa - s >, .:, -ao > s W k; Ad .a o*s I,/ ,1 - 'I A, +,
,.w.,i:' t' ',7s'''' ,s _-*
i:$',jv '.';-'; 7'DBiw ' 7,v > M. _ Halso support the interpretation of the nonlineargraphs obtained for ultraviolet irradiation andheat resistance.
Dipicolinic acid content. In contrast to endo-spores, and as in methane-oxidizing bacteria(95), no dipicolinate could be detected in exo-spore preparations.Photopigments. Absorbance spectra for puri-
fied exospore preparations show a marked de-crease in both chlorophyll and carotenoid con-tent in comparison to that found in vegetativecells from the same culture (Fig. 40).
Fine Structure of Vegetative Cells
Fine structure. The membrane complexpresent in strain Rm 5 is similar to that previ-ously described (12, 87, 91) in other Rhodomi-crobium strains. Membrane content (as judgedby thin sections) varied inversely with lightintensity; grown at 500 lx (incident light) or lessthe bacteria were packed with multiple layersof double membranes (Fig. 41), whereas at2,000 lx or higher only one layer of double mem-brane (in addition to the cytoplasmic mem-
brane) was usually observed (Fig. 42 and 43),an effect of light intensity described previously(87). Grown in continuous culture in malate-pyruvate medium with an incident light inten-sity of 1,500 lx at 30TC (steady-state opticaldensity, 1.52; dilution rate, 0.026) cells werefound to contain a considerable number of cellu-lar inclusions (polyhedral bodies) (Fig. 44),which were similar in appearance to the car-boxysomes of Thiobacillus intermedius (63).
Questions unanswered in the past concernthe point of origin of the double membranelayers and the three-dimensional architectureof cells containing these lamellae. This latterpoint has a bearing on the interpretation ofevidence concerning the process ofreproductiondiscussed later.
Electron micrographs provided the informa-tion used in the construction of a three-dimen-sional model (Fig. 45), which was subsequentlysectioned in various dimensions and found toaccount for the different membrane arrange-ments seen in thin sections (Fig. 41 to 43).A key event in development of lamellae ap-
pears to be the circumpolar invagination of the
VOL. 41, 1977
.7-r
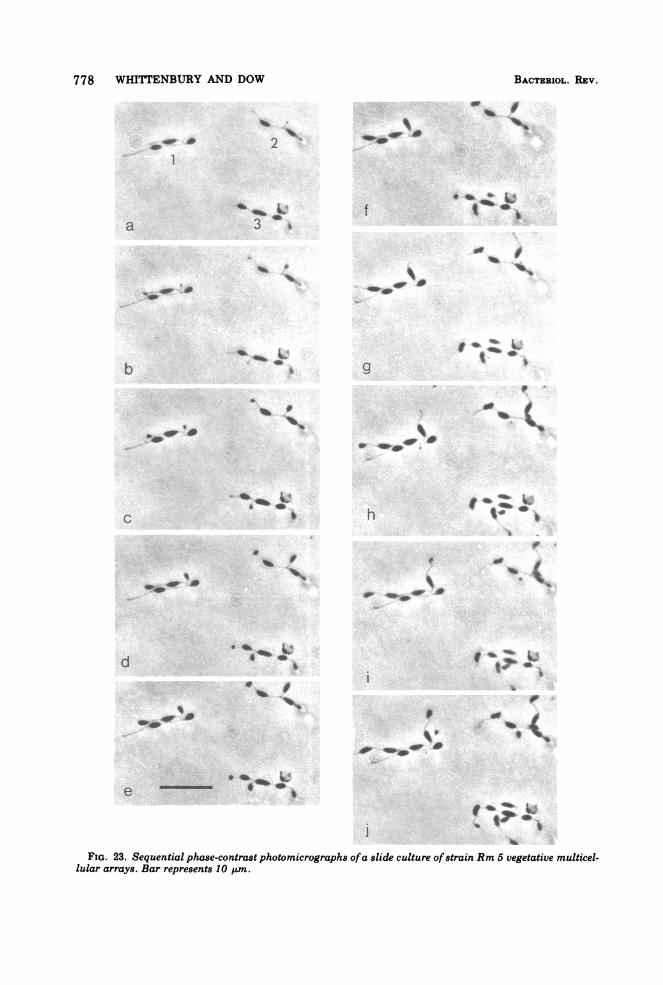
778 WHITTENBURY AND DOW
FIG. 23. Sequential phase-contrast photomicrographs ofa slide culture ofstrain Rm 5 vegetative multicel-lular arrays. Bar represents 10 Aum.
BACTERIOL. REV.
i. lk- I'. v
,!t "I 't -- .2A
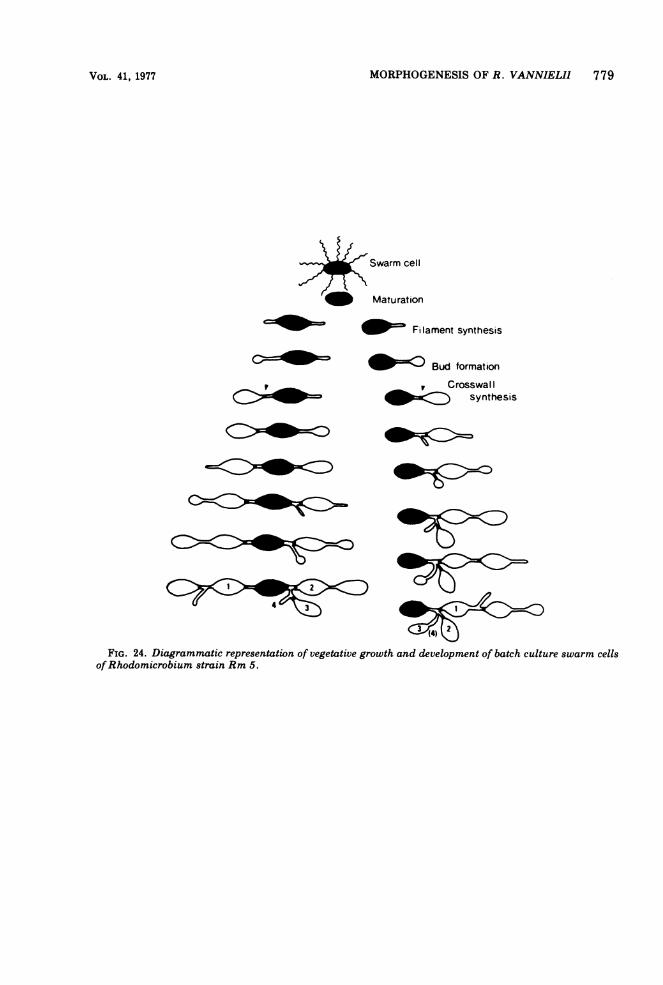
MORPHOGENESIS OF R. VANNIELII 779
C_'D
r K
Swarm cell
Maturation
_ > Filament synthesis
_ °-C) Bud formation
I Crosswallsynthesis
_<KD
FIG. 24. Diagrammatic representation of vegetative growth and development of batch culture swarm cellsofRhodomicrobium strain Rm 5.
VOL. 41, 1977
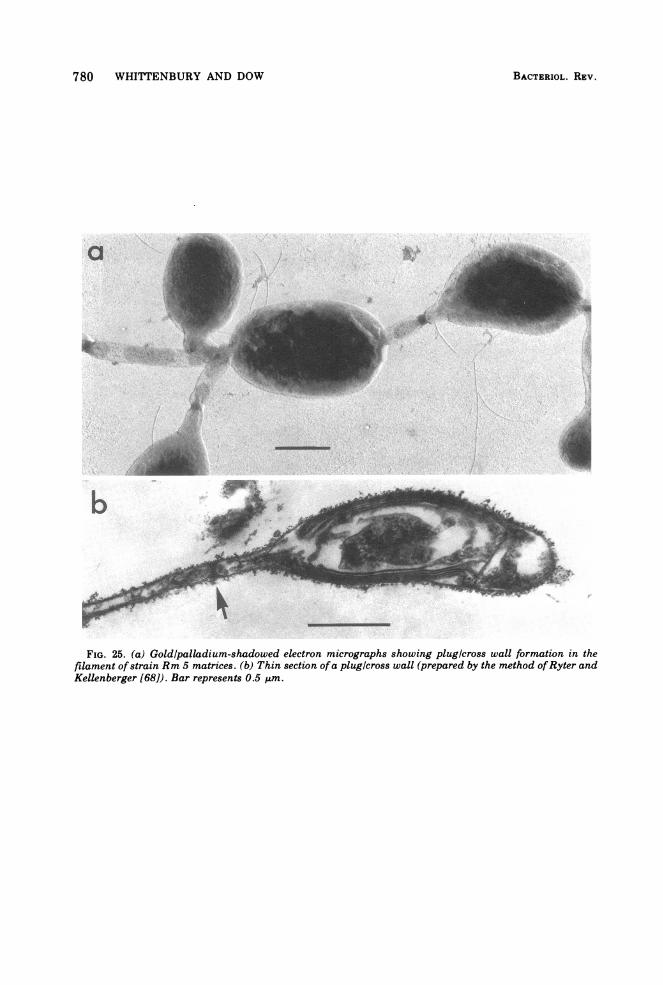
780 WHITTENBURY AND DOW
FIG. 25. (a) Goldlpalladium-shadowed electron micrographs showing plug/cross wall formation in thefilament of strain Rm 5 matrices. (b) Thin section ofa plug/cross wall (prepared by the method ofRyter andKellenberger [68]). Bar represents 0.5 am.
BACTERIOL. REV.

MORPHOGENESIS OF R. VANNIELII 781
:0'-00 :-t.y'
'd; .e,.:fF.t :.aXi, ^
A; .B; a.; i. or I, >.-,
X,, ,,''SL _'IX .. .' _
a. Y .-_Ean* > ?AS. i-,: ,r
! + 'atBrg';^ ."',-'' '.
_ k;d 'he x,¢'s.' '.
ante"="'''" ''A.-F oW0_ ail_ I'm' I,{]' ' -1,.'.' ,lDs')'_ w_Ets; -.e S A- s-:,: A...L- Of;. . i._z _ en _
FIG. 26. Goldlpalladium-shadowed electron micrographs showing sites of plug formation. The distancefrom the mother cell (m) to the plug is variable and dependent on the growth rate. The distance from thecompleted daughter cell to the plug is relatively constant (c). The sequence ofevents leading to the formation ofcellular matrices is shown diagrammatically. Bar represents 2.5 pum.
VOL. 41, 1977
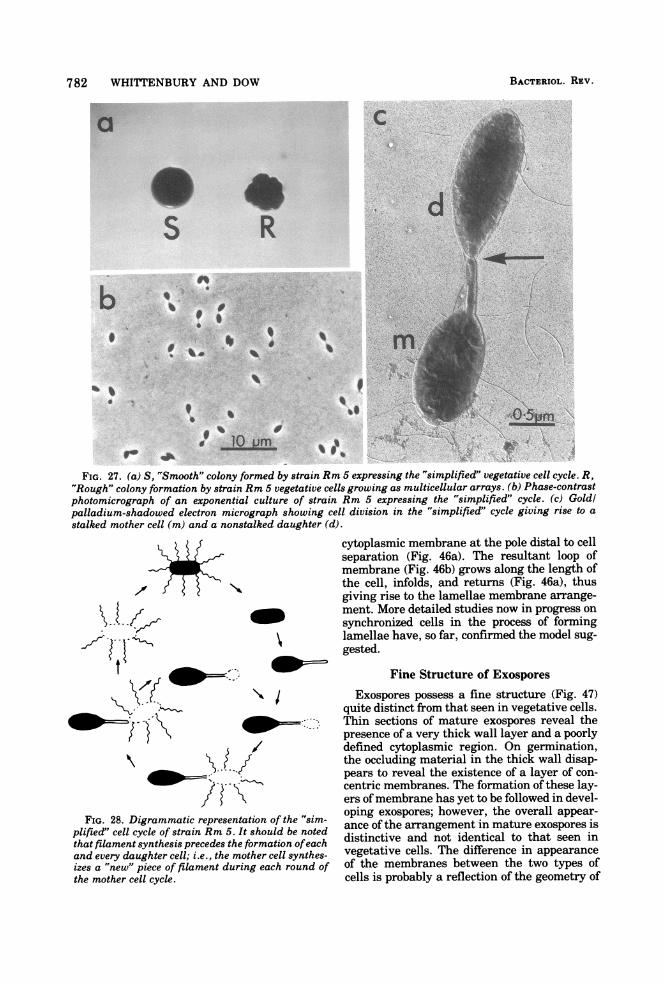
782 WHITTENBURY AND DOW
FIG. 27. (a) S, "Smooth" colony formed by strain Rm 5 expressing the "simplified" vegetative cell cycle. R,"Rough" colony formation by strain Rm 5 vegetative cells growing as multicellular arrays. (b) Phase-contrastphotomicrograph of an exponential culture of strain Rm 5 expressing the "simplified" cycle. (c) Goldlpalladium-shadowed electron micrograph showing cell division in the "simplified" cycle giving rise to astalked mother cell (m) and a nonstalked daughter (d).
FIG. 28. Digrammatic representation of the "sim-plified" cell cycle of strain Rm 5. It should be notedthat filament synthesis precedes the formation ofeachand every daughter cell; i.e., the mother cell synthes-izes a "new" piece of filament during each round ofthe mother cell cycle.
cytoplasmic membrane at the pole distal to cellseparation (Fig. 46a). The resultant loop ofmembrane (Fig. 46b) grows along the length ofthe cell, infolds, and returns (Fig. 46a), thusgiving rise to the lamellae membrane arrange-ment. More detailed studies now in progress onsynchronized cells in the process of forminglamellae have, so far, confirmed the model sug-gested.
Fine Structure of Exospores
Exospores possess a fine structure (Fig. 47)quite distinct from that seen in vegetative cells.Thin sections of mature exospores reveal thepresence of a very thick wall layer and a poorlydefined cytoplasmic region. On germination,the occluding material in the thick wall disap-pears to reveal the existence of a layer of con-centric membranes. The formation of these lay-ers ofmembrane has yet to be followed in devel-oping exospores; however, the overall appear-ance of the arrangement in mature exospores isdistinctive and not identical to that seen invegetative cells. The difference in appearanceof the membranes between the two types ofcells is probably a reflection of the geometry of
BACTERIOL. REV.
I
"I'll, N*
... --X,..n
4I
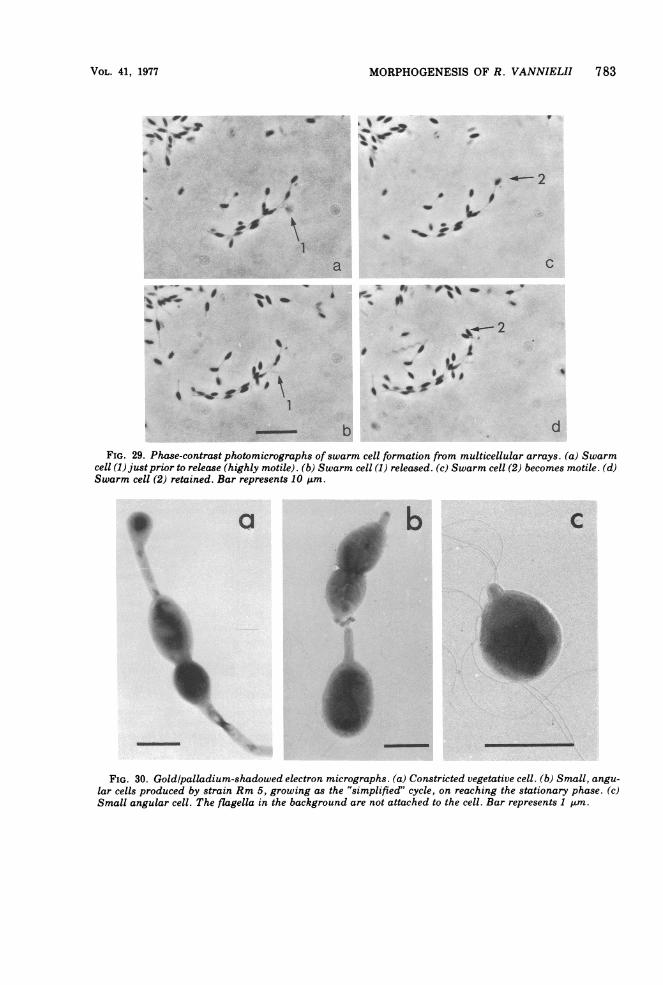
MORPHOGENESIS OF R. VANNIELII 783
I0 -*- 2
_ * *qp 1
a.RP- -f
I~ I
SI
d%
b
% tq~Jr
eo 0rt: V'_!- ^ r
SW 4r2,
_%' % .qI t
bFIG. 29. Phase-contrast photomicrographs ofswarm cell formation from multicellular arrays. (a) Swarm
cell (1) just prior to release (highly motile). (b) Swarm cell (1) released. (c) Swarm cell (2) becomes motile. (d)Swarm cell (2) retained. Bar represents 10 mtm.
)k
a b c
FIG. 30. Goldlpalladium-shadowed electron micrographs. (a) Constricted vegetative cell. (b) Small, angu-lar cells produced by strain Rm 5, growing as the 'simplified" cycle, on reaching the stationary phase. (c)Small angular cell. The flagella in the background are not attached to the cell. Bar represents 1 ttm.
)0,_O p
Io
a
C
d
VOL. 41, 1977
1
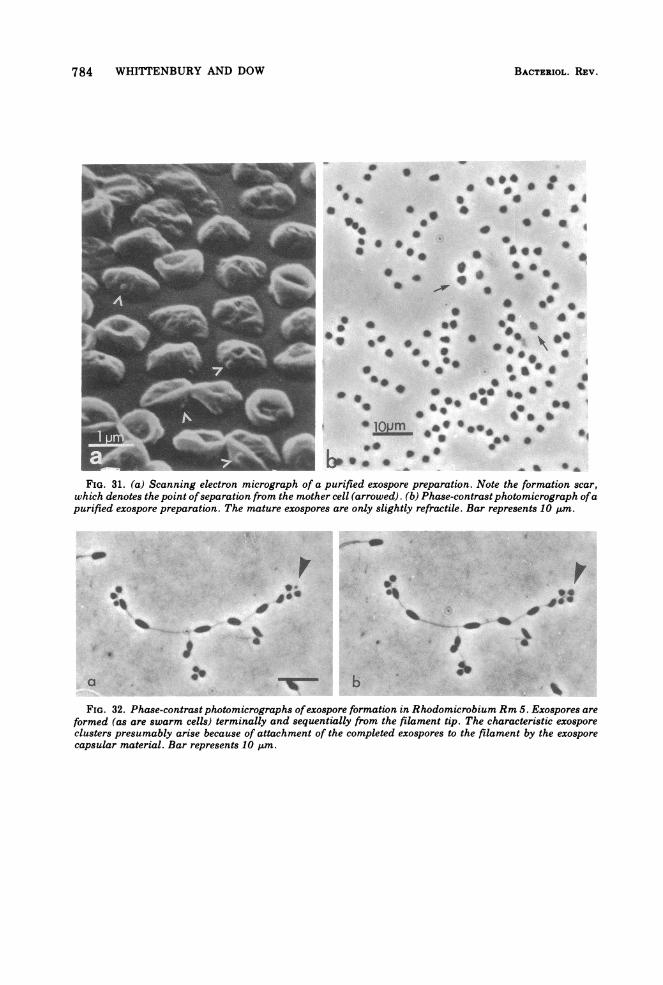
784 WHITTENBURY AND DOW
iIs
-_ 3 *_ 0 - _ ,A ssFIG. 31. (a) Scanning electron micrograph of a purified exospore preparation. Note the formation scar,
which denotes the point ofseparation from the mother cell (arrowed). (b) Phase-contrastphotomicrograph ofapurified exospore preparation. The mature exospores are only slightly refractile. Bar represents 10 ,um.
FIG. 32. Phase-contrast photomicrographs ofexospore formation in Rhodomicrobium Rm 5. Exospores areformed (as are swarm cells) terminally and sequentially from the filament tip. The characteristic exosporeclusters presumably arise because of attachment of the completed exospores to the filament by the exosporecapsular material. Bar represents 10 ktm.
BACTERIOL. REV.
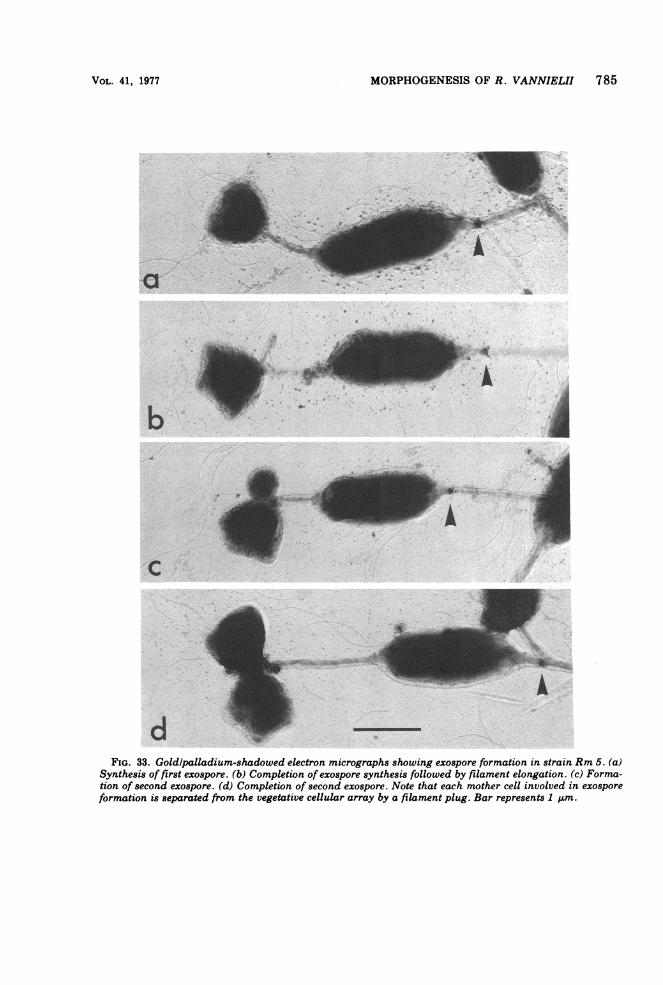
MORPHOGENESIS OF R. VANNIELII 785
FIG. 33. Goldlpalladium-shadowed electron micrographs showing exospore formation in strain Rm 5. (a)Synthesis of first exospore. (b) Completion of exospore synthesis followed by filament elongation. (c) Forma-tion of second exospore. (d) Completion of second exospore. Note that each mother cell involved in exosporeformation is separated from the vegetative cellular array by a filament plug. Bar represents 1 Ium.
VOL. 41, 1977
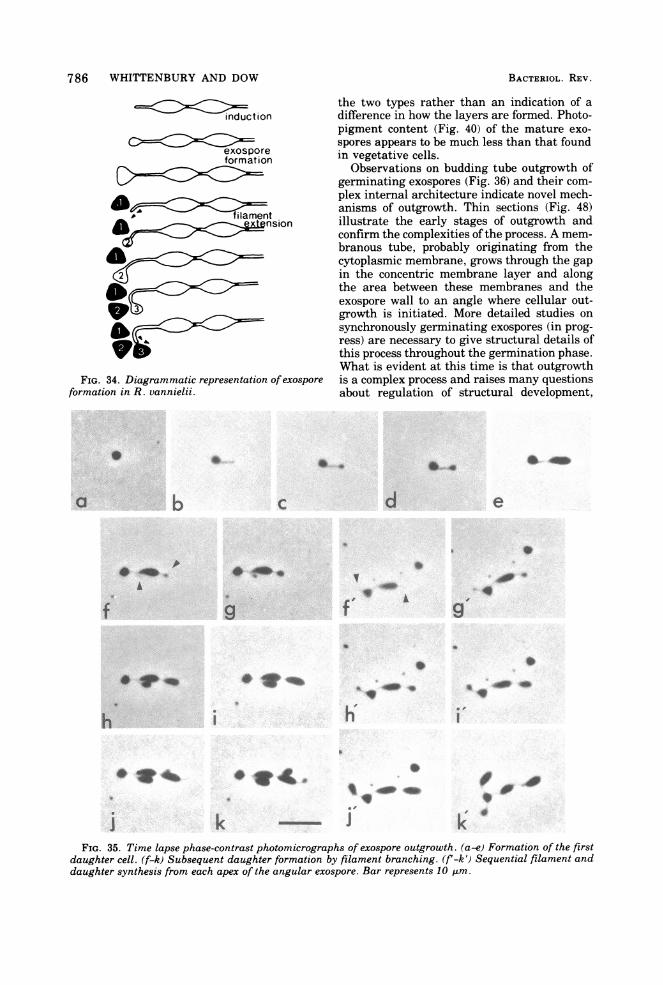
786 WHITTENBURY AND DOW
induction
000exosporeformation
a~~~~~~~~~~~~~~~s ~~~~filament
£^ffinsion
FIG. 34. Diagrammatic representation of exosporeformation in R. vannielii.
i , !
i
the two types rather than an indication of adifference in how the layers are formed. Photo-pigment content (Fig. 40) of the mature exo-spores appears to be much less than that foundin vegetative cells.
Observations on budding tube outgrowth ofgerminating exospores (Fig. 36) and their com-plex internal architecture indicate novel mech-anisms of outgrowth. Thin sections (Fig. 48)illustrate the early stages of outgrowth andconfirm the complexities ofthe process. A mem-branous tube, probably originating from thecytoplasmic membrane, grows through the gapin the concentric membrane layer and alongthe area between these membranes and theexospore wall to an angle where cellular out-growth is initiated. More detailed studies onsynchronously germinating exospores (in prog-ress) are necessary to give structural details ofthis process throughout the germination phase.What is evident at this time is that outgrowthis a complex process and raises many questionsabout regulation of structural development,
.0.s.. i . .. ta . ... . 0., ....... g 8 .-. f d.+. s tts > t ia Sig v 70 FB' J .+ D.:
pA; FLi, 1i !L ti . ri > ,
;s X,
D'.rWi j X_._s s W t.d _.s4.w''S2, X, :,t .'S Li' ) '., we
.+ r 7.*i?;N .- s$E-
;St 'b. 'M=>.'V' '. u.s 0,@X. Bt:
S
IV. _
_o A
4
.i'
h
*
g
**&_04al
* .4.w 4m
J kFIG. 35. Time lapse phase-contrast photomicrographs of exospore outgrowth. (a-e) Formation of the first
daughter cell. (f-k) Subsequent daughter formation by filament branching. (f'-k') Sequential filament anddaughter synthesis from each apex of the angular exospore. Bar represents 10 Am.
BACTERIOL. REV.
op
i
S.-
f 0 AO0
k 4
MORPHOGENESIS OF R. VANNIELII 787
FIG. 36. Goldlpalladium-shadowed electron micrographs of exospore outgrowth. (a) Mature exospore.Note the fibrillar capsular material. (b) Filament formation. (c) Daughter cell formation. (d) Completionof daughter cell as indicated by plug formation in the filament (arrow). (e) Branch formation. (e') Filamentand daughter cell formation from the second apex. (f and f') Formation of the second daughter cell. Barrepresents 2 p1m.
VOL. 41, 1977
4-1
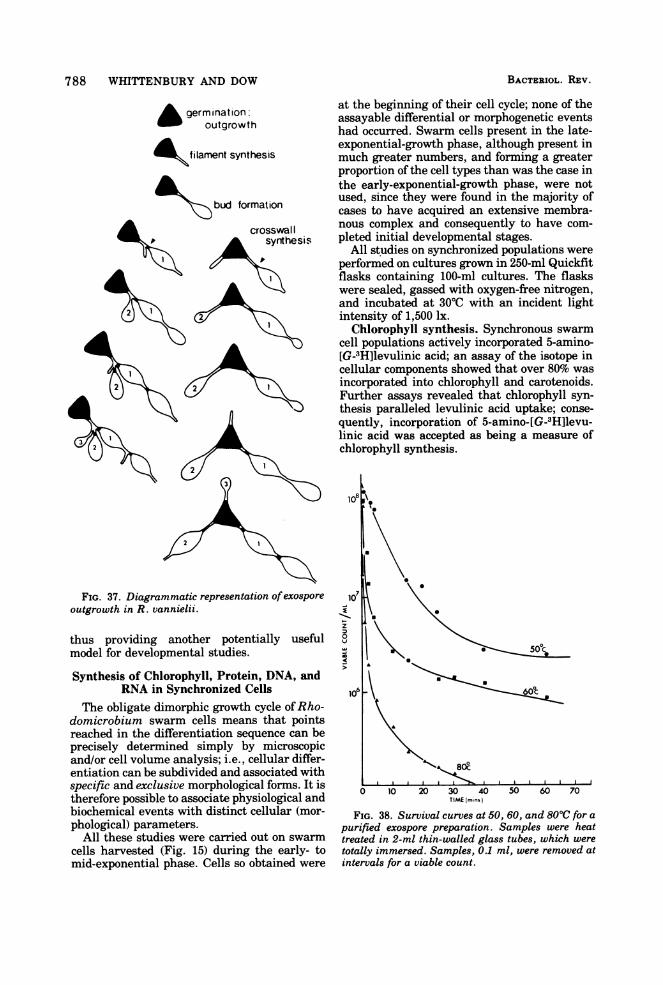
788 WHITTENBURY AND DOW
germination:outgrowth
£ka tlament synthesis
\>bud formation
at the beginning of their cell cycle; none of theassayable differential or morphogenetic eventshad occurred. Swarm cells present in the late-exponential-growth phase, although present inmuch greater numbers, and forming a greaterproportion of the cell types than was the case inthe early-exponential-growth phase, were notused, since they were found in the majority ofcases to have acquired an extensive membra-nous complex and consequently to have com-pleted initial developmental stages.
All studies on synchronized populations wereperformed on cultures grown in 250-ml Quickfitflasks containing 100-ml cultures. The flaskswere sealed, gassed with oxygen-free nitrogen,and incubated at 30TC with an incident lightintensity of 1,500 lx.
Chlorophyll synthesis. Synchronous swarmcell populations actively incorporated 5-amino-[G-3H]levulinic acid; an assay of the isotope incellular components showed that over 80% wasincorporated into chlorophyll and carotenoids.Further assays revealed that chlorophyll syn-thesis paralleled levulinic acid uptake; conse-quently, incorporation of 5-amino-[G-3H]levu-linic acid was accepted as being a measure ofchlorophyll synthesis.
FIG. 37. Diagrammatic representation ofexosporeoutgrowth in R. vannielii.
thus providing another potentially usefulmodel for developmental studies.
Synthesis of Chlorophyll, Protein, DNA, andRNA in Synchronized Cells
The obligate dimorphic growth cycle ofRho-domicrobium swarm cells means that pointsreached in the differentiation sequence can beprecisely determined simply by microscopicand/or cell volume analysis; i.e., cellular differ-entiation can be subdivided and associated withspecific and exclusive morphological forms. It istherefore possible to associate physiological andbiochemical events with distinct cellular (mor-phological) parameters.
All these studies were carried out on swarmcells harvested (Fig. 15) during the early- tomid-exponential phase. Cells so obtained were
0 10 20 30 40 50 60 70TIME (mins)
FIG. 38. Survival curves at 50, 60, and 800C for apurified exospore preparation. Samples were heattreated in 2-ml thin-walled glass tubes, which weretotally immersed. Samples, 0.1 ml, were removed atintervals for a viable count.
BACTERIOL. REV.
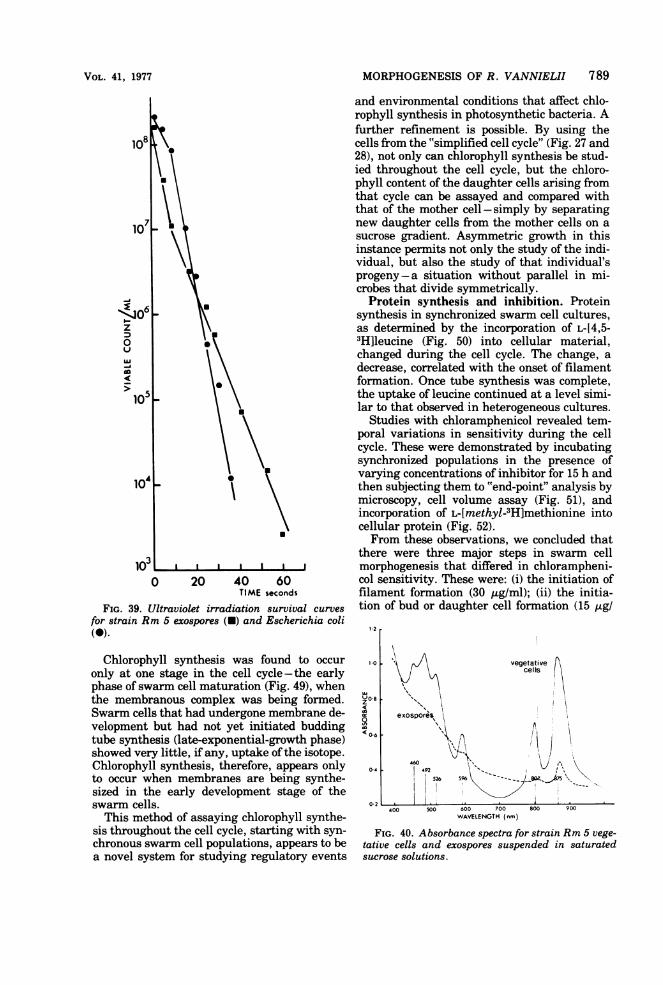
MORPHOGENESIS OF R. VANNIELII 789
Lu
-j
0 20 40 60TIME seconds
FIG. 39. Ultraviolet irradiation survival curvesfor strain Rm 5 exospores (a) and Escherichia coli(a).
Chlorophyll synthesis was found to occuronly at one stage in the cell cycle-the earlyphase of swarm cell maturation (Fig. 49), whenthe membranous complex was being formed.Swarm cells that had undergone membrane de-velopment but had not yet initiated buddingtube synthesis (late-exponential-growth phase)showed very little, if any, uptake of the isotope.Chlorophyll synthesis, therefore, appears onlyto occur when membranes are being synthe-sized in the early development stage of theswarm cells.
This method of assaying chlorophyll synthe-sis throughout the cell cycle, starting with syn-chronous swarm cell populations, appears to bea novel system for studying regulatory events
and environmental conditions that affect chlo-rophyll synthesis in photosynthetic bacteria. Afurther refinement is possible. By using thecells from the "simplified cell cycle" (Fig. 27 and28), not only can chlorophyll synthesis be stud-ied throughout the cell cycle, but the chloro-phyll content of the daughter cells arising fromthat cycle can be assayed and compared withthat of the mother cell-simply by separatingnew daughter cells from the mother cells on asucrose gradient. Asymmetric growth in thisinstance permits not only the study of the indi-vidual, but also the study of that individual'sprogeny-a situation without parallel in mi-crobes that divide symmetrically.Protein synthesis and inhibition. Protein
synthesis in synchronized swarm cell cultures,as determined by the incorporation of L-[4,5-3H]leucine (Fig. 50) into cellular material,changed during the cell cycle. The change, adecrease, correlated with the onset of filamentformation. Once tube synthesis was complete,the uptake of leucine continued at a level simi-lar to that observed in heterogeneous cultures.
Studies with chloramphenicol revealed tem-poral variations in sensitivity during the cellcycle. These were demonstrated by incubatingsynchronized populations in the presence ofvarying concentrations of inhibitor for 15 h andthen subjecting them to "end-point" analysis bymicroscopy, cell volume assay (Fig. 51), andincorporation of L-[methyl-3H]methionine intocellular protein (Fig. 52).From these observations, we concluded that
there were three major steps in swarm cellmorphogenesis that differed in chlorampheni-col sensitivity. These were: (i) the initiation offilament formation (30 jig/ml); (ii) the initia-tion of bud or daughter cell formation (15 Atg/
z
< 0-6
0-4
400 500 600 700 800 900WAVELENGTH (nm)
FIG. 40. Absorbance spectra for strain Rm 5 vege-tative cells and exospores suspended in saturatedsucrose solutions.
VOL. 41, 1977
1 2 r
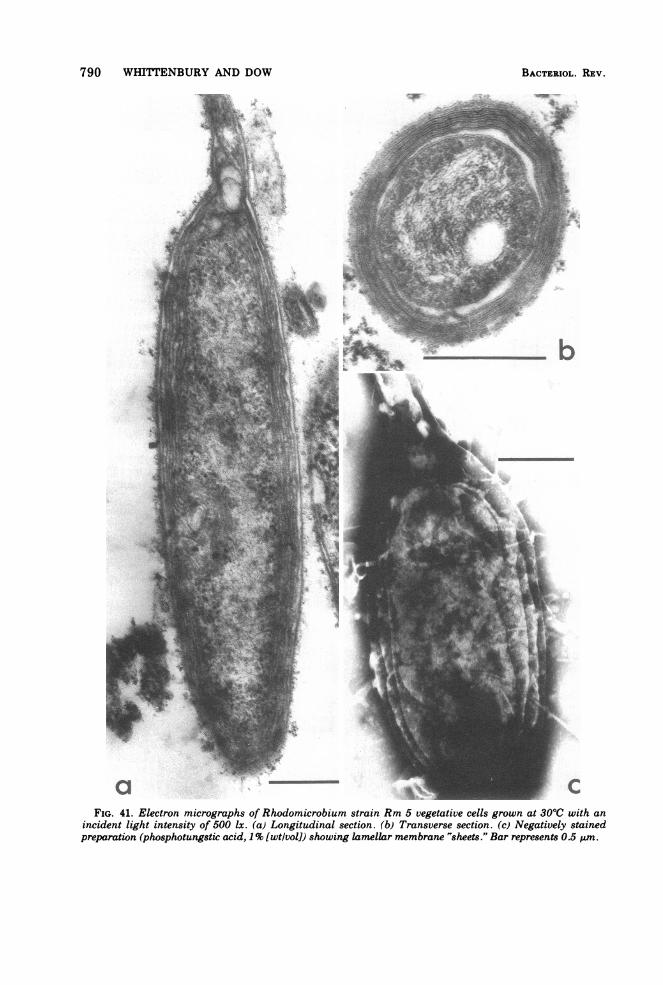
790 WHITTENBURY AND DOW
a 7 CFIG. 41. Electron micrographs of Rhodomicrobium strain Rm 5 vegetative cells grown at 300C with an
incident light intensity of 500 lx. (a) Longitudinal section. (b) Transverse section. (c) Negatively stainedpreparation (phosphotungstic acid, 1% [wt/volD showing lamellar membrane 'sheets." Bar represents 0.5 pAm.
BACTERIOL. REV.
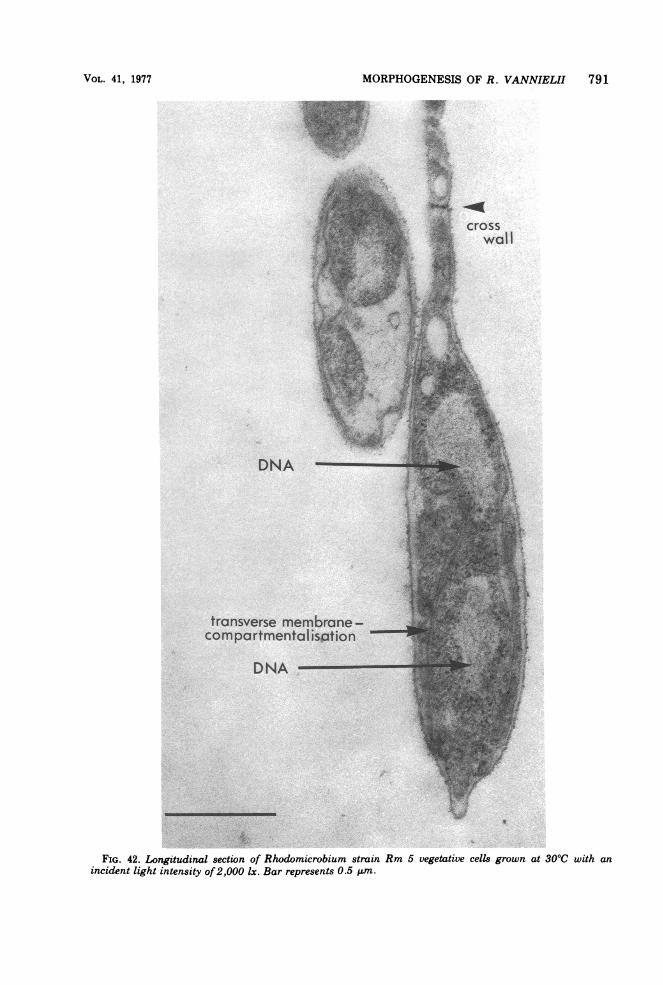
MORPHOGENESIS OF R. VANNIELII 791
-s-
ofa.e g '; '-t_-.o ,, 4- . 7, 0
-- ,, z', # je. Si_t ^_
+. j2o,'vw' ,. +E''-
--'0';' 'e'-'tJto*; u -;
9 .'' t'' ' ;' D ' ,, f , i .',.. . ' ',, '., , (*,' * i0 0 . , ... . ' . P s.* -. ' . . S ,, Ly t ,*j * t ,<
! ;, ,. o', F,,hS ''s 0 C'TraX L;SS' ' -S A;-s , ; 0, ksf - o- ... 6.
f X,' 0- < 74 :,!
FIG. 42. Longitudinal section of Rhodomicrobium strain Rm 5 vegetative cells grown at 30°C with anincident light intensity of2,000 lx. Bar represents 0.5 ttm.
VOL. 41, 1977
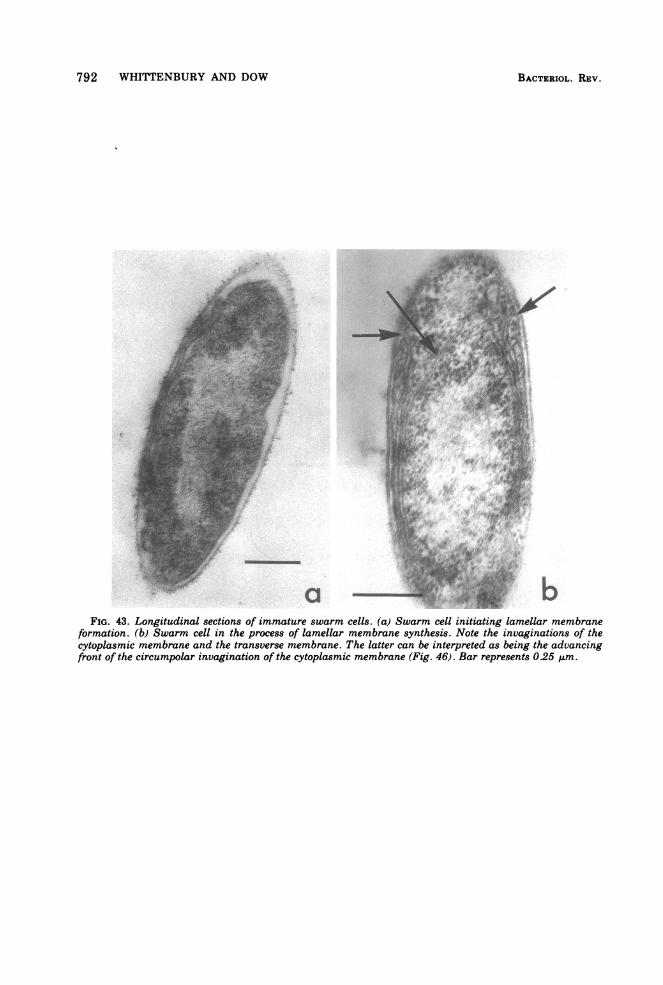
792 WHITTENBURY AND DOW
FIG. 43. Longitudinal sections of immature swarm cells. (a) Swarm cell initiating lamellar membraneformation. (b) Swarm cell in the process of lamellar membrane synthesis. Note the invaginations of thecytoplasmic membrane and the transverse membrane. The latter can be interpreted as being the advancingfr-ont of the circumpolar invagination of the cytoplasmic membrane (Fig. 46). Bar represents 0.25 tim.
BACTERIOL. REV.
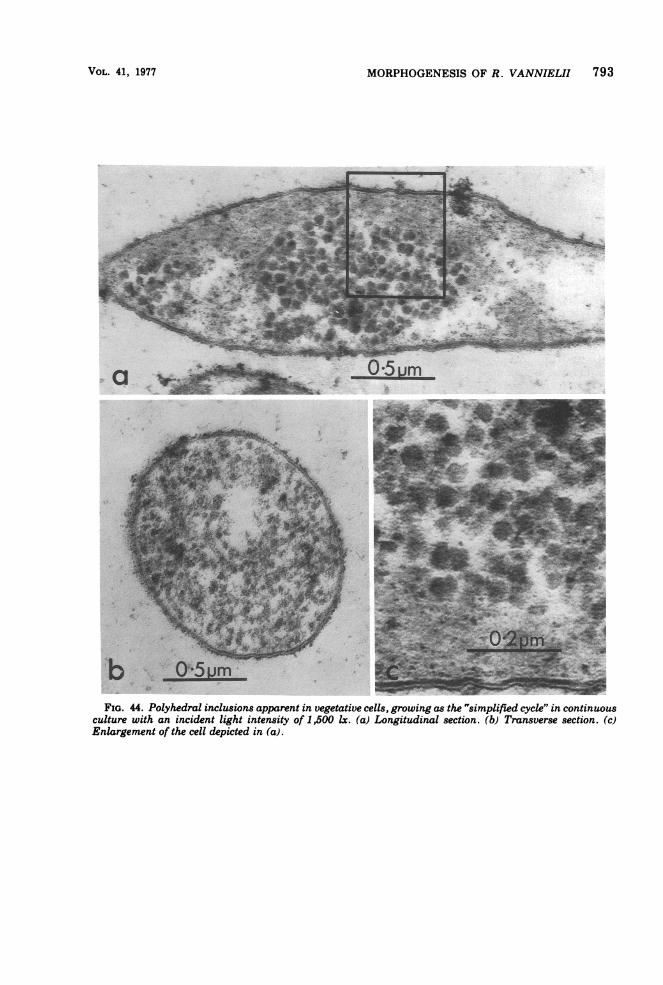
MORPHOGENESIS OF R. VANNIELII 793
FIG. 44. Polyhedral inclusions apparent in vegetative cells, growing as the 'simplified cycle" in continuousculture with an incident light intensity of 1,500 lx. (a) Longitudinal section. (b) Transverse section. (c)Enlargement of the cell depicted in (a).
VOL. 41, 1977

794 WHITTENBURY AND DOW
lSmembraneo _ ~~~'bops'
cytoplasmicmembrane
FIG. 45. Diagrammatic representation of a cut-away three-dimensional model ofa Rhodomicrobiumvegetative cell grown under conditions of low lightintensity (500 lx).
ml); and (iii) the physiological separation ofmother and daughter cell by plug formation (5,ug/ml, an obligate step before a further repro-
ductive cycle. Electron microscopy of cells inthe last category showed a total lack of crosswall synthesis.These observations have been further sub-
stantiated by subjecting synchronized popula-tions to the critical chloramphenicol concentra-tions at differing times during the cycle; e.g., 5,g of chloramphenicol per ml will inhibit crosswall formation (division) provided it is added aminimum of 30 to 40 min before the event. Itwould therefore appear that the data obtainedby end-point analysis does not simply reflectthat the higher the concentration of an inhibi-tor the sooner it exerts an effect; i.e., it isindicative of sequential changes in chloram-phenicol sensitivity.RNA synthesis. The incorporation of [G-
3H]uridine by swarm cells into cellular mate-rial was assumed to reflect ribonucleic acid(RNA) synthesis (Fig. 50) during the cell cycle;RNA synthesis appeared to be at a constantrate throughout the cycle.The effect of different rifampin concentra-
tions on swarm cell development was deter-mined by using the same experimental protocolas was employed to study chloramphenicol sen-sitivity. The data (Fig. 53 and 54) indicated thatthere was a significant change in rifampin sen-sitivity between immature, nonreproductiveswarm cells (50 ,ug/ml) and the mature repro-ductive mother cell (25 /Lg/ml). The distinctplateau occurring between 25 and 50 ,ug of ri-fampin per ml suggested that the RNA polym-erase had been modified in some way. An inter-pretation is that the terminal event in the mat-
uration process is the modification of the RNApolymerase that subsequently permits recogni-tion and transcription of those genes concernedwith mother cell function, i.e., reproduction.Such a modified polymerase would be unable totranscribe those genes associated with swarmcell maturation, a once-only event in the life ofeach and every swarm cell.DNA synthesis. Nucleic acid synthesis in
synchronized Rhodomicrobium swarm cellswas examined by: (i) radioisotope incorpora-tion, (ii) fluorescence, (iii) Giemsa staining,and (iv) specific inhibitors.
(i) Radioisotope incorporation. Attempts tomonitor deoxyribonucleic acid (DNA) synthesisby the uptake of [methyl-3H]thymidine failedpresumably because the thymidine was de-graded to thymine by thymidine phosphorylase(6). It has been shown that in E. coli thymidinephosphorylase can be inhibited or repressed bythe addition of uridine (7). However, when uri-dine (10 mM) was added to Rhodomicrobiumcells, there was no significant change in isotopeincorporation. Similar difficulties were experi-enced with thymidine 5'-monophosphate (73).There was no incorporation of deoxyguanosineand low levels of deoxyadenosine (0.1 mM) in-hibited growth within 30 min.Although we have used several mutation and
isolation procedures, we have as yet been un-able to isolate a thymine-requiring mutant.
(ii) Fluorometric determination of DNAsynthesis. Taking advantage of the large quan-tities of synchronized cells we were able to pre-pare, by the filtration method described earlier,DNA synthesis (Fig. 55) was followed by usingthe fluorometric assay system proposed by Don-kersloot et al. (19), which involves complexingthe nucleic acid with ethidium bromide andmeasuring the resulting fluorescence. The ma-jor finding, when comparing synchronized andheterogeneous populations, was that DNA syn-thesis in the former was discontinuous, beingrepressed up to the midway point in the "matu-ration" period. A similar situation is known toexist for Caulobacter (15) and R. palustrisswarm cells (93). As repression ofDNA synthe-sis was observed only in "immature" swarmcells it was assumed that (i) mother cells syn-thesize DNA continuously, and (ii) first-gener-ation swarm cells must pass part way throughthe maturation period before DNA synthesis isinitiated. RNA synthesis in synchronouslygrowing cells measured by this technique par-alleled the curve recorded for uridine uptake(Fig. 50).
(iii) Giemsa staining. Morphologicalchanges, that is, broad changes in conformation
BACTERIOL. REV.
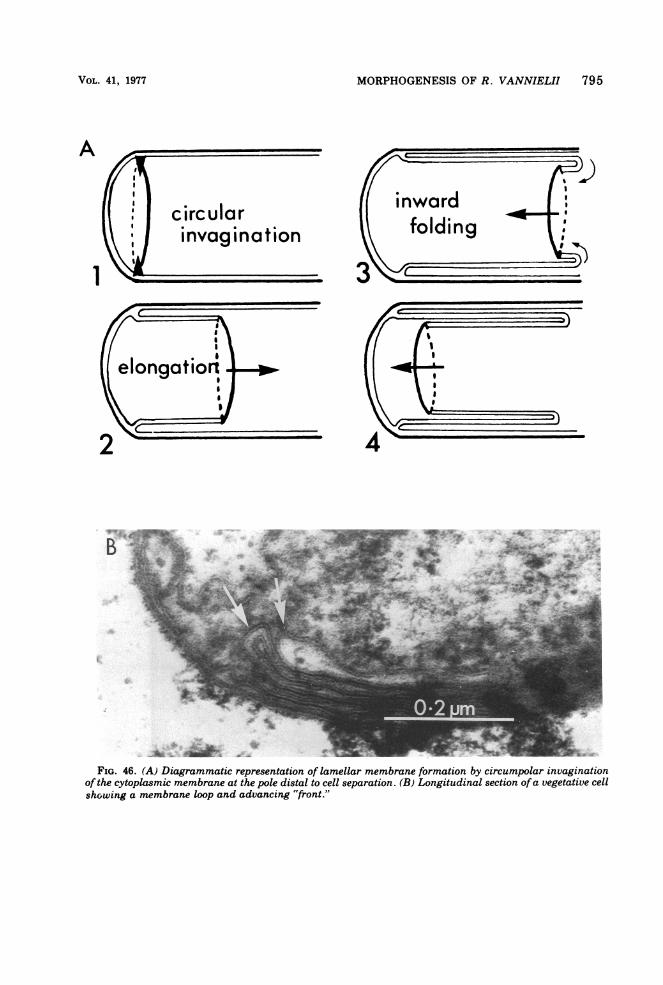
MORPHOGENESIS OF R. VANNIELII 795
circularinvagination
2 - 4
,*- _" v-
H ie"3 Aim f
FIG. 46. (A) Diagrammatic representation of lamellar membrane formation by circumpolar invaginationof the cytoplasmic membrane at the pole distal to cell separation. (B) Longitudinal section ofa vegetative cellshowing a membrane loop and advancing "front."
A
1
VOL. 41, 1977
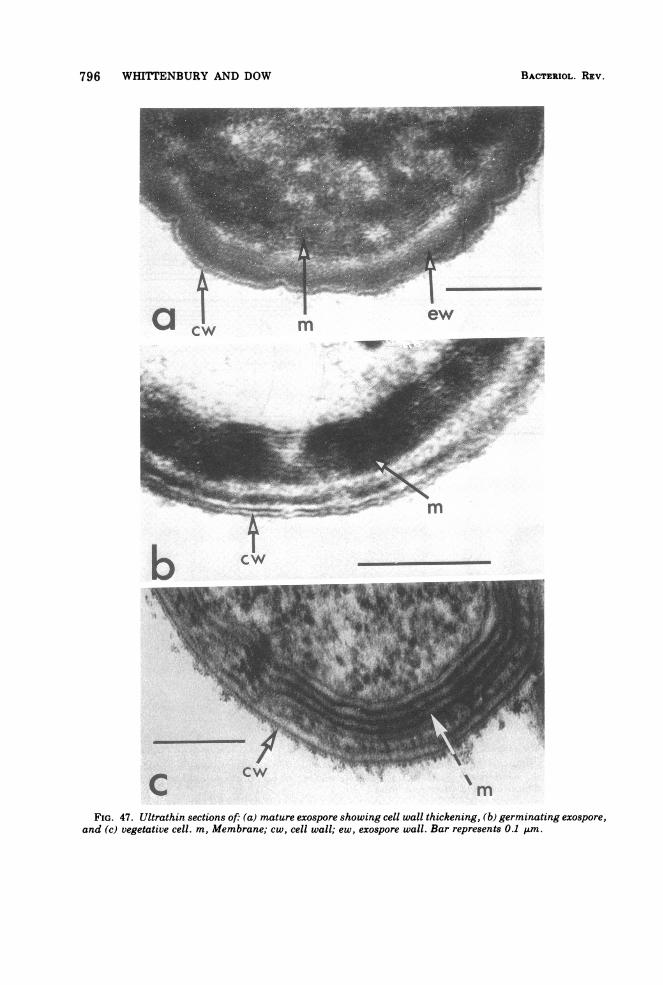
796 WHITTENBURY AND DOW
b
tIw'> 085 f t Vi
X, >.~~~T
FIG. 47. Ultrathin sections of (a) mature exospore showing cell wall thickening, (b) germinating exospore,
and (c) vegetative cell. m, Membrane; cw, cell wall; ew, exospore wall. Bar represents 0.1 ,m.
I
mIlew
CW -
BACTERIOL. REV.
cw
-11 t
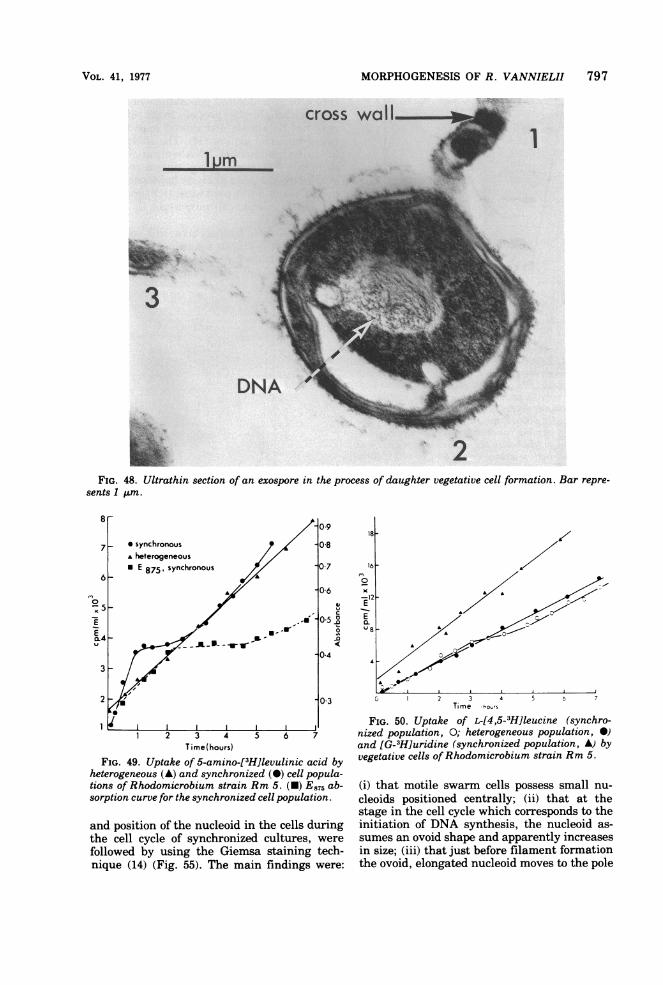
MORPHOGENESIS OF R. VANNIELII 797
3 A.
FIG. 48. Ultrathin section of an exospore in the process of daughter vegetative cell formation. Bar repre-sents 1 um.
Time(hours)
FIG. 49. Uptake of 5-amino-[3H]levulinic acid byheterogeneous (A) and synchronized (A) cell popula-tions of Rhodomicrobium strain Rm 5. (-) E875 ab-sorption curve for the synchronized cell population.
and position of the nucleoid in the cells duringthe cell cycle of synchronized cultures, were
followed by using the Giemsa staining tech-nique (14) (Fig. 55). The main findings were:
0 2 3 4 5 t 7Time hours
FIG. 50. Uptake of L-[4,5-3H]leucine (synchro-nized population, 0; heterogeneous population, *)and [G-3H]uridine (synchronized population, A) byvegetative cells ofRhodomicrobium strain Rm 5.
(i) that motile swarm cells possess small nu-cleoids positioned centrally; (ii) that at thestage in the cell cycle which corresponds to theinitiation of DNA synthesis, the nucleoid as-sumes an ovoid shape and apparently increasesin size; (iii) that just before filament formationthe ovoid, elongated nucleoid moves to the pole
VOL. 41, 1977
..I L)mno
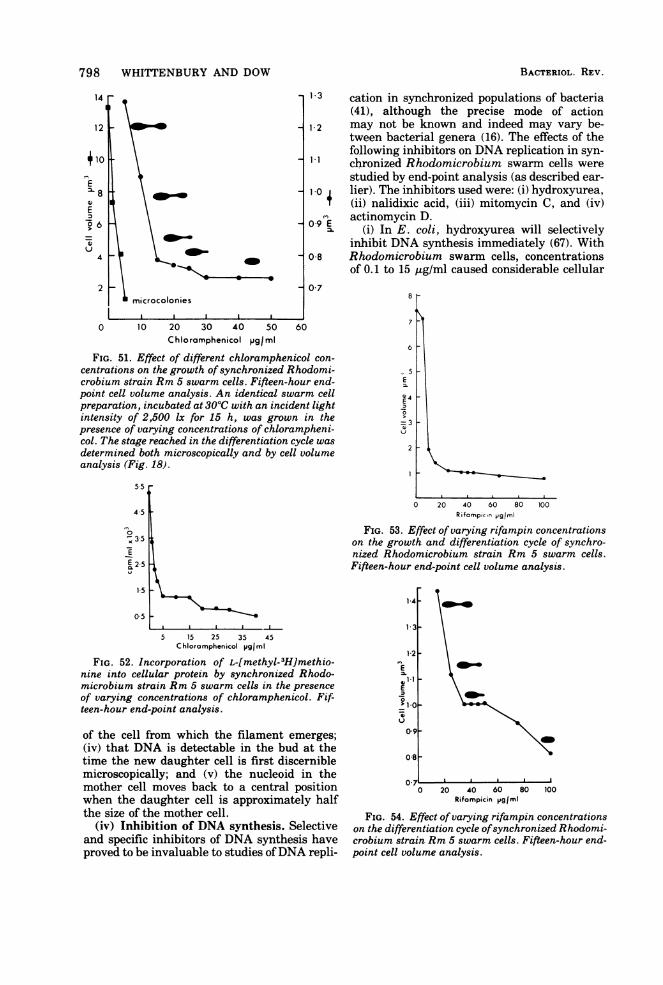
798 WHITTENBURY AND DOW
14
12
t 2
410In
E
E
:1)0 6
4
2
\_
Imicrocolonies
0 10 20 30 40 50Chloramphenicol pg/ml
13 cation in synchronized populations of bacteria(41), although the precise mode of action
1-2 may not be known and indeed may vary be-tween bacterial genera (16). The effects of thefollowing inhibitors on DNA replication in syn-
-1 chronized Rhodomicrobium swarm cells werestudied by end-point analysis (as described ear-
10 lier). The inhibitors used were: (i) hydroxyurea,4 (ii) nalidixic acid, (iii) mitomycin C, and (iv)
actinomycin D.09E (i) In E. coli, hydroxyurea will selectively
inhibit DNA synthesis immediately (67). With08 Rhodomicrobium swarm cells, concentrations
of 0.1 to 15 pug/ml caused considerable cellular07
60
FIG. 51. Effect of different chloramphenicol con-centrations on the growth of synchronized Rhodomi-crobium strain Rm 5 swarm cells. Fifteen-hour end-point cell volume analysis. An identical swarm cellpreparation, incubated at 300C with an incident lightintensity of 2,500 Ix for 15 h, was grown in thepresence of varying concentrations of chlorampheni-col. The stage reached in the differentiation cycle wasdetermined both microscopically and by cell volumeanalysis (Fig. 18).
5.5
4.5
3-35
E2-5
15
05 -
5 15 25 35 45Chloramphenicol V91mI
FIG. 52. Incorporation of L-[methyl-3H]methio-nine into cellular protein by synchronized Rhodo-microbium strain Rm 5 swarm cells in the presenceof varying concentrations of chloramphenicol. Fif-teen-hour end-point analysis.
of the cell from which the filament emerges;(iv) that DNA is detectable in the bud at thetime the new daughter cell is first discerniblemicroscopically; and (v) the nucleoid in themother cell moves back to a central positionwhen the daughter cell is approximately halfthe size of the mother cell.
(iv) Inhibition of DNA synthesis. Selectiveand specific inhibitors of DNA synthesis haveproved to be invaluable to studies ofDNA repli-
6
- 5mEo-p4E
>=3
0 20 40 60 80 100Rifampcon pgiml
FIG. 53. Effect of varying rifampin concentrationson the growth and differentiation cycle of synchro-nized Rhodomicrobium strain Rm 5 swarm cells.Fifteen-hour end-point cell volume analysis.
1-4
1 2-
E.1*1E0
'10
0.8
0 20 40 60 80 100Rifompicin Vg/rmI
FIG. 54. Effect of varying rifampin concentrationson the differentiation cycle ofsynchronized Rhodomi-crobium strain Rm 5 swarm cells. Fifteen-hour end-point cell volume analysis.
BACTERIOL. REV.
1- 3
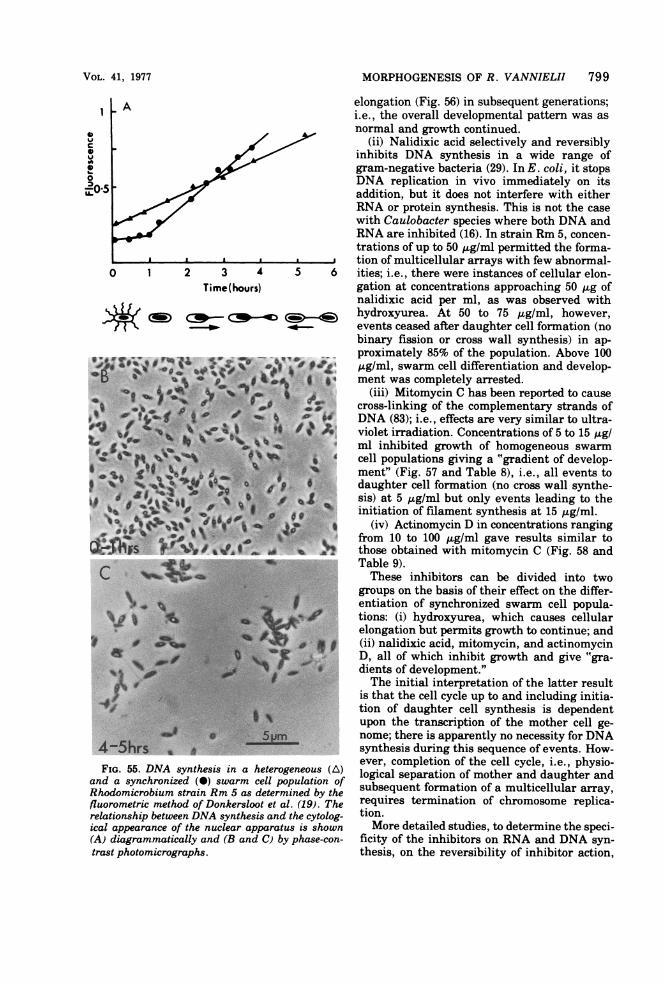
MORPHOGENESIS OF R. VANNIELII 799
CHW
0
0 1 2 3 4Time(hours)
yC0
FIG. 55. DNA synthesis in a heterogeand a synchronized (S) swarm cell popRhodomicrobium strain Rm 5 as determifluorometric method of Donkersloot et al.relationship between DNA synthesis and tical appearance of the nuclear apparatus(A) diagrammatically and (B and C) bytrast photomicrographs.
elongation (Fig. 56) in subsequent generations;i.e., the overall developmental pattern was asnormal and growth continued.
(ii) Nalidixic acid selectively and reversiblyinhibits DNA synthesis in a wide range ofgram-negative bacteria (29). In E. coli, it stopsDNA replication in vivo immediately on itsaddition, but it does not interfere with eitherRNA or protein synthesis. This is not the casewith Caulobacter species where both DNA andRNA are inhibited (16). In strain Rm 5, concen-trations of up to 50 ug/ml permitted the forma-
.........a tion of multicellular arrays with few abnormal-5 6 ities; i.e., there were instances of cellular elon-
gation at concentrations approaching 50 Ag ofnalidixic acid per ml, as was observed with
< 9 hydroxyurea. At 50 to 75 Ag/ml, however,events ceased after daughter cell formation (nobinary fission or cross wall synthesis) in ap-proximately 85% of the population. Above 100
C tg/ml, swarm cell differentiation and develop-_ ment was completely arrested.
(iii) Mitomycin C has been reported to causecross-linking of the complementary strands ofDNA (83); i.e., effects are very similar to ultra-violet irradiation. Concentrations of 5 to 15 tug/ml inhibited growth of homogeneous swarmcell populations giving a "gradient of develop-ment" (Fig. 57 and Table 8), i.e., all events todaughter cell formation (no cross wall synthe-sis) at 5 ,ug/ml but only events leading to theinitiation of filament synthesis at 15 ,tg/ml.
(iv) Actinomycin D in concentrations rangingfrom 10 to 100 ,ug/ml gave results similar tothose obtained with mitomycin C (Fig. 58 andTable 9).These inhibitors can be divided into two
groups on the basis of their effect on the differ-entiation of synchronized swarm cell popula-tions: (i) hydroxyurea, which causes cellularelongation but permits growth to continue; and(ii) nalidixic acid, mitomycin, and actinomycinD, all of which inhibit growth and give "gra-dients of development."The initial interpretation of the latter result
is that the cell cycle up to and including initia-tion of daughter cell synthesis is dependentupon the transcription of the mother cell ge-nome; there is apparently no necessity for DNAsynthesis during this sequence of events. How-ever, completion of the cell cycle, i.e., physio-
mleots (of logical separation of mother and daughter and,ulation ofsubsequent formation of a multicellular array,(19). The requires termination of chromosome replica-
the cytolog- tion.s is shown More detailed studies, to determine the speci-phase-con- ficity of the inhibitors on RNA and DNA syn-
thesis, on the reversibility of inhibitor action,
VOL. 41, 1977
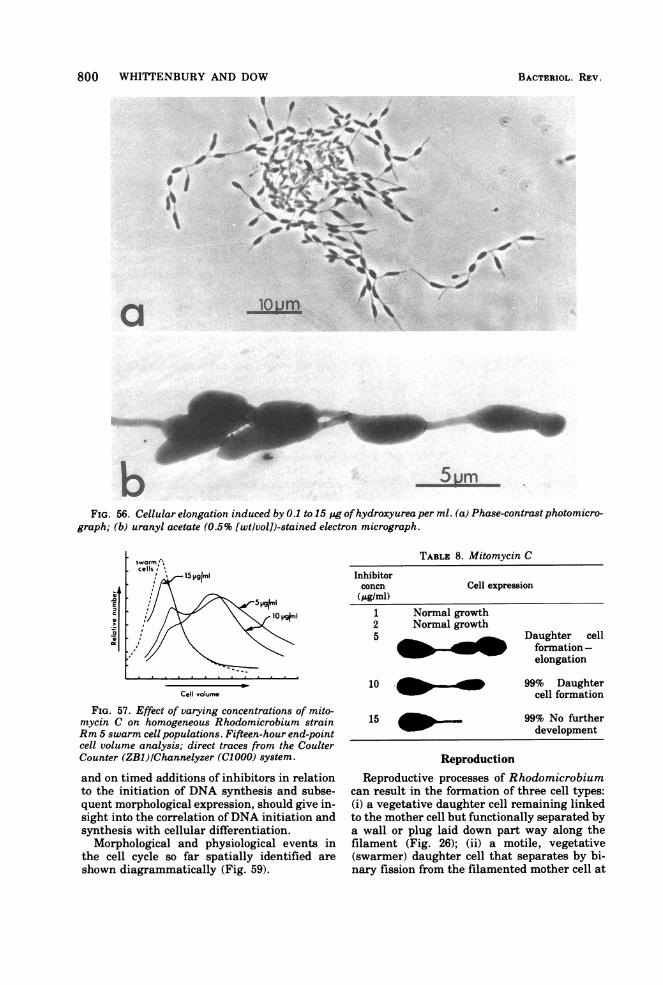
800 WHI[TENBURY AND DOW
bFIG. 56. Cellular elongation induced by 0.1 to 15 pg ofhydroxyurea per ml. (a) Phase-contrast photomicro-
graph; (b) uranyl acetate (0.5% [wt/vol])-stained electron micrograph.
TABLE 8. Mitomycin C
Cell expression
E 5 Vgjm..l
Cell volume
FIG. 57. Effect of varying concentrations of mito-mycin C on homogeneous Rhodomicrobium strainRm 5 swarm cell populations. Fifteen-hour end-pointcell volume analysis; direct traces from the CoulterCounter (ZB1)/Channelyzer (C1000) system.
and on timed additions of inhibitors in relationto the initiation of DNA synthesis and subse-quent morphological expression, should give in-sight into the correlation ofDNA initiation andsynthesis with cellular differentiation.
Morphological and physiological events inthe cell cycle so far spatially identified areshown diagrammatically (Fig. 59).
1 Normal growth2 Normal growth5 Daughter cell
_ _ _ formation -elongation
10 _ 99% Daughtercell formation
15 _ 99% No furtherdevelopment
ReproductionReproductive processes of Rhodomicrobium
can result in the formation of three cell types:(i) a vegetative daughter cell remaining linkedto the mother cell but functionally separated bya wall or plug laid down part way along thefilament (Fig. 26); (ii) a motile, vegetative(swarmer) daughter cell that separates by bi-nary fission from the filamented mother cell at
Inhibitorconcn(pgIml)
BACTERIOL. REV.
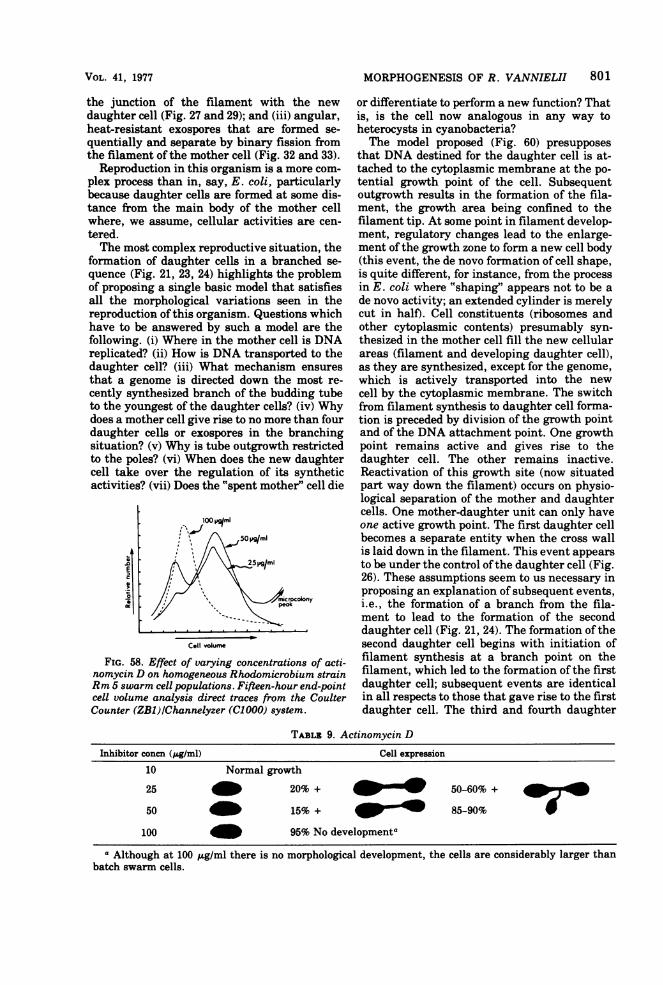
MORPHOGENESIS OF R. VANNIELII 801
the junction of the filament with the newdaughter cell (Fig. 27 and 29); and (iii) angular,heat-resistant exospores that are formed se-quentially and separate by binary fission fromthe filament of the mother cell (Fig. 32 and 33).
Reproduction in this organism is a more com-plex process than in, say, E. coli, particularlybecause daughter cells are formed at some dis-tance from the main body of the mother cellwhere, we assume, cellular activities are cen-tered.The most complex reproductive situation, the
formation of daughter cells in a branched se-quence (Fig. 21, 23, 24) highlights the problemof proposing a single basic model that satisfiesall the morphological variations seen in thereproduction of this organism. Questions whichhave to be answered by such a model are thefollowing. (i) Where in the mother cell is DNAreplicated? (ii) How is DNA transported to thedaughter cell? (iii) What mechanism ensuresthat a genome is directed down the most re-cently synthesized branch of the budding tubeto the youngest of the daughter cells? (iv) Whydoes a mother cell give rise to no more than fourdaughter cells or exospores in the branchingsituation? (v) Why is tube outgrowth restrictedto the poles? (vi) When does the new daughtercell take over the regulation of its syntheticactivities? (vii) Does the "spent mother" cell die
100 Vml
50Ia ml
E ~~~~~2511gml
olony
Cell volume
FIG. 58. Effect of varying concentrations of acti-nomycin D on homogeneous Rhodomicrobium strainRm 5 swarm cell populations. Fifteen-hour end-pointcell volume analysis direct traces from the CoulterCounter (ZB1)/Channelyzer (C1000) system.
or differentiate to perform a new function? Thatis, is the cell now analogous in any way toheterocysts in cyanobacteria?The model proposed (Fig. 60) presupposes
that DNA destined for the daughter cell is at-tached to the cytoplasmic membrane at the po-tential growth point of the cell. Subsequentoutgrowth results in the formation of the fila-ment, the growth area being confined to thefilament tip. At some point in filament develop-ment, regulatory changes lead to the enlarge-ment of the growth zone to form a new cell body(this event, the de novo formation of cell shape,is quite different, for instance, from the processin E. coli where "shaping" appears not to be ade novo activity; an extended cylinder is merelycut in half). Cell constituents (ribosomes andother cytoplasmic contents) presumably syn-thesized in the mother cell fill the new cellularareas (filament and developing daughter cell),as they are synthesized, except for the genome,which is actively transported into the newcell by the cytoplasmic membrane. The switchfrom filament synthesis to daughter cell forma-tion is preceded by division of the growth pointand of the DNA attachment point. One growthpoint remains active and gives rise to thedaughter cell. The other remains inactive.Reactivation of this growth site (now situatedpart way down the filament) occurs on physio-logical separation of the mother and daughtercells. One mother-daughter unit can only haveone active growth point. The first daughter cellbecomes a separate entity when the cross wallis laid down in the filament. This event appears
to be under the control ofthe daughter cell (Fig.26). These assumptions seem to us necessary inproposing an explanation of subsequent events,i.e., the formation of a branch from the fila-ment to lead to the formation of the seconddaughter cell (Fig. 21, 24). The formation of thesecond daughter cell begins with initiation offilament synthesis at a branch point on thefilament, which led to the formation of the firstdaughter cell; subsequent events are identicalin all respects to those that gave rise to the firstdaughter cell. The third and fourth daughter
TABLz 9. Actinomycin DInhibitor concn (pg/ml) Cell expression
10 Normal growth
25 _ 20% + 50-60% +
50 15% + 85-90%
100 _ 95% No developmentaa Although at 100 j.g/ml there is no morphological development, the cells are considerably larger than
batch swarm cells.
VOL. 41, 1977
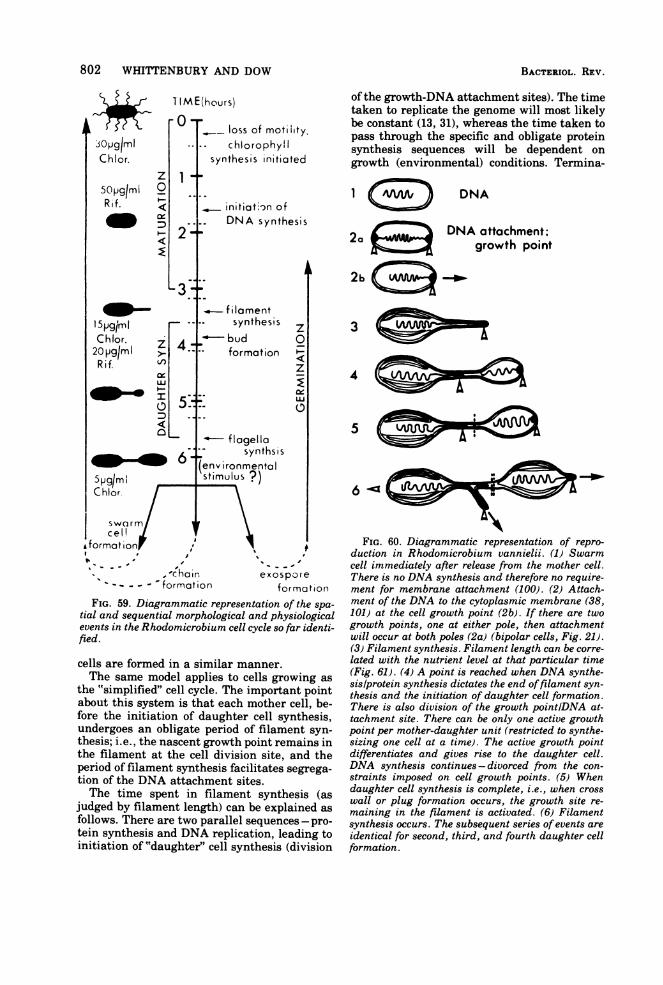
802 WHITTENBURY AND DOW
1I M E(hours)
3Opg/mlChlor.
5opg/miRif.
l5-1gI
z0wo
D_5:E
-0-
1i
2-
Lhlor.iz
_ LI5
5::
6e
5pgIm StifChlor.
swarmscell
A format ion ,
v - - 'iChair
loss of motility.chlorophyl
synthesis initiated
initiation ofDNA synthesis
filamentsynthesis
-budformat ion
C
2
U.
C:
- flagellasynthsis
Ivironmentalimulus 9)
exospo)reformation formation
FIG. 59. Diagrammatic representation of the spa-tial and sequential morphological and physiologicalevents in the Rhodomicrobium cell cycle so far identi-fied.
cells are formed in a similar manner.The same model applies to cells growing as
the "simplified" cell cycle. The important pointabout this system is that each mother cell, be-fore the initiation of daughter cell synthesis,undergoes an obligate period of filament syn-thesis; i.e., the nascent growth point remains inthe filament at the cell division site, and theperiod of filament synthesis facilitates segrega-tion of the DNA attachment sites.The time spent in filament synthesis (as
judged by filament length) can be explained as
follows. There are two parallel sequences - pro-tein synthesis and DNA replication, leading toinitiation of "daughter" cell synthesis (division
of the growth-DNA attachment sites). The timetaken to replicate the genome will most likelybe constant (13, 31), whereas the time taken topass through the specific and obligate proteinsynthesis sequences will be dependent ongrowth (environmental) conditions. Termina-
DNA
DNA attachment;2a growth point
2b
4
S
FIG. 60. Diagrammatic representation of repro-duction in Rhodomicrobium vannielii. (1) Swarmcell immediately after release from the mother cell.There is no DNA synthesis and therefore no require-ment for membrane attachment (100). (2) Attach-ment of the DNA to the cytoplasmic membrane (38,101) at the cell growth point (2b). If there are twogrowth points, one at either pole, then attachmentwill occur at both poles (2a) (bipolar cells, Fig. 21).(3) Filament synthesis. Filament length can be corre-lated with the nutrient level at that particular time(Fig. 61). (4) A point is reached when DNA synthe-sis/protein synthesis dictates the end offilament syn-thesis and the initiation of daughter cell formation.There is also division of the growth point/DNA at-tachment site. There can be only one active growthpoint per mother-daughter unit (restricted to synthe-sizing one cell at a time). The active growth pointdifferentiates and gives rise to the daughter cell.DNA synthesis continues - divorced from the con-straints imposed on cell growth points. (5) Whendaughter cell synthesis is complete, i.e., when crosswall or plug formation occurs, the growth site re-maining in the filament is activated. (6) Filamentsynthesis occurs. The subsequent series of events areidentical for second, third, and fourth daughter cellformation.
BACTIERIOL. REV.
I

MORPHOGENESIS OF R. VANNIELII 803
attachment replication terminationattof DNAc of genome . of genome -of DNA of genome replication
swarmcell A1
A sequential protein synthesis Z
initiationof proteinanllpnep
filamentsynthesis
termination ofprotein sequence
division ofgrowth/attachment
site
nascentgrowth/attachment daughter cell
site oformationactivation of growth site- .filament synthesis, i.e., physiologicalinitiation of protein se- separation ofquence. (Replication of mother anddaughterthe genome and proteinsequence linked as in E.coli by a requirement forprotein synthesis beforetermination of genomereplication)
FIG. 61. Control ofswarm cell development by correlation ofprotein and DNA synthesis.
tion of both pathways will be essential for initi-ation of daughter cell synthesis (Fig. 61). Asomewhat similar model has been proposed toexplain the control of replication and divisionin E. coli (40).
Obviously, the model proposed requires sub-stantial experimental support before it can beclaimed to be more than just a model; this weare now attempting to provide. Circumstantialevidence, in part answer to the original ques-tions posed and in support of the proposedmodel, is as follows.
(i) Thin sections have only ever revealed asingle cytoplasmic membrane in the filament(Fig. 41 and 42); in other words, DNA from thebody of the mother cell is not transported to asecond daughter cell by an outgrowth of aninvagination of the cytoplasmic membranefrom the mother cell.
(ii) The mother and daughter cells are alwayscontinuous up to the point of fission or plugging(Fig. 21).
(iii) Separation and fission processes appearto be under the control of the newly formeddaughter cell. Alteration of filament length of
the mother cell by varying nutrient concentra-tions (Fig. 20) does not materially alter thepoint of fission or the distance ofplug formationfrom the daughter cell.
(iv) The arrangement of lamellae, maskingthe cytoplasmic membrane from the DNAalong the length of the cell, seems to be in parta reason why outgrowth only takes place fromthe poles of the cells where the DNA can makedirect contact with the cytoplasmic membrane.
(v) That there is only a single growth point orgrowth zone is demonstrated by penicillin ex-periments (Fig. 62) and is further substantiatedby autoradiographic experiments concerningthe incorporation of 2,6-diamino-[G-3H]pimelicacid by synchronous cultures of swarm cells.Pulse-chase experiments indicate that label isprimarily incorporated at the filament tip andin the developing daughter cell at the pole dis-tal to the mother cell and filament. That theactive growth point is not located only at thepole of the mother cell is confirmed by branchgrowth.
(vi) Movement of DNA to new daughter cellsby a process other than "membrane drag"
VOL. 41, 1977
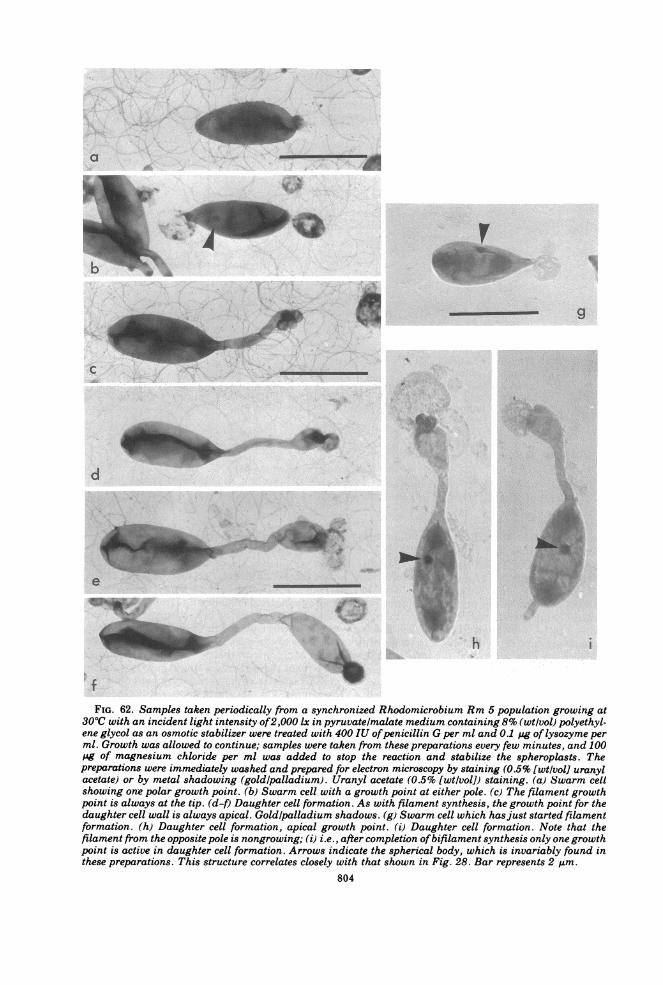
FIG. 62. Samples taken periodically from a synchronized Rhodomicrobium Rm 5 population growing at300C with an incident light intensity of2,000 lx in pyruvate/malate medium containing 8% (wtlvol) polyethyl-ene glycol as an osmotic stabilizer were treated with 400 IU ofpenicillin G per ml and 0.1 Ag oflysozyme perml. Growth was allowed to continue; samples were taken from these preparations every few minutes, and 100pg of magnesium chloride per ml was added to stop the reaction and stabilize the spheroplasts. Thepreparations were immediately washed and prepared for electron microscopy by staining (0.5% [wt/vol] uranylacetate) or by metal shadowing (gold/palladium). Uranyl acetate (0.5% [wt/vol]) staining. (a) Swarm cellshowing one polar growth point. (b) Swarm cell with a growth point at either pole. (c) The filament growthpoint is always at the tip. (d-f) Daughter cell formation. As with filament synthesis, the growth point for thedaughter cell wall is always apical. Goldlpalladium shadows. (g) Swarm cell which hasjust started filamentformation. (h) Daughter cell formation, apical growth point. (i) Daughter cell formation. Note that thefilament from the opposite pole is nongrowing; (i) i.e., after completion ofbifilament synthesis only one growthpoint is active in daughter cell formation. Arrows indicate the spherical body, which is invariably found inthese preparations. This structure correlates closely with that shown in Fig. 28. Bar represents 2 ,Am.
804

MORPHOGENESIS OF R. VANNIELII 805
seems most unlikely in view of daughter cellformation when the tube is over 40 jum long(phosphate limitation, Fig. 20) and whenbranching is involved (Fig. 21 and 24).
(vii) Evidence for parallel pathways of DNAand protein synthesis leading to daughter celldevelopment comes from inhibitor studies.When DNA synthesis is specifically inhibited,there is no physiological separation or binaryfission of mother and daughter cell. This sug-gests that termination of genome replication isessential to separation of cells (40). Similarly,specific inhibition of protein synthesis and thechanges in sensitivity to chloramphenicol indi-cate a temporally related sequence of proteinsyntheses during differentiation, which termi-nate in "division."
CONCLUSIONBudding bacteria-unlike E. coli but like
Caulobacter species -possess dimorphic cell cy-cles and, like Caulobacter species, should beuseful models for the study of morphogenesisand differentiation in procaryotes. This possi-bility has been explored by using R. vannielii.To avoid confusion in comparing different ex-amples of morphogenesis and differentiation asexpressed by different species of bacteria, wehave attempted to define and categorize suchevents. E. coli, for instance, is seen to be amorphologically simple organism with a simplevegetative cell cycle in contrast to Anabaenacylindrical, which we consider to be an exampleofa multicellular procaryote, in which differen-tiated cells of at least three types regularlyoccur, differentiation reflecting intercellularrelationships.The supposed differences between budding
and binary fission- modes of reproduction wereshown to be more apparent than real. The keyfeature distinguishing budding bacteria fromrod-shaped bacteria seemed to be the obligatepolar mode of growth by the former, leadingfrequently to asymmetric cell division, andmore importantly to a mother-daughter cell cy-cle rather than the sibling cell cycle as in E.coli. Aging was shown to be a characteristic ofmother cells, and immaturity was seen to becharacteristic of daughter cells.The uniqueness ofthe daughter cell -in com-
parison with a sibling cell formed by E. coli- isthat a series of events (both morphological andmolecular) occur which are not repeated in sub-sequent rounds of cell cycles of that particularcell. Such a feature, apparently restricted tobacteria with dimorphic cell cycles, is clearly ofrelevance to the study of morphogenesis anddifferentiation in that it is an obligate event inthe cell cycle, not an occasional event, as is
spore formation and other systems presentlystudied in this context.R. vannielii, an exosporeforming, photosyn-
thetic bacterium possessing a complex internalfine structure and external morphology, under-goes complex cell cycles and proved to be auseful example of a budding bacterium for ex-ploring the nature of morphogenesis and differ-entiation.
Selective synchronization-a prime requisitefor work on such organisms- was accomplishedrapidly and simply (by filtration through glasswool) and resulted in yields of cells (daughtercells at the beginning of the cell maturationcycle) far in excess of previously recorded yieldsfor selective synchronization techniques. Suchyields meant that both quantitative and quali-tative methods could be used in the investiga-tions.Changes occurring through the cell cycle
were explored. The pattern of events recordedparallel, to some degree, those published forCaulobacter crescentus but also included addi-tional changes useful for further exploration ofthe molecular biology.ofmorphogenesis and dif-ferentiation.
LITERATURE CITED1. Agabian-Keshishian, N., and L. Shapiro. 1971.
Bacterial differentiation and phage infec-tion. Virology 44:46-53.
2. Ashby, C. 1938. Simplified Shaeffer sporestain. Science 87:443.
3. Attwood, M. M., and W. Harder. 1972. A rapidand specific enrichment procedure for Hy-phomicrobium spp. Antonie van Leeuwen-hoek J. Microbiol Serol. 38:369-378.
4. Autissier, F., A. Jaffe, and A. Kepes. 1971.Segregation of galactoside permease, a mem-brane marker during growth and cell divi-sion in Escherichia coli. Mol. Gen. Genet.112:275-288.
5. Beachey, E. H., and R. M. Cole. 1966. Cellwall replication in Escherichia coli studiedby immunofluorescence and immunoelectronmicroscopy. J. Bacteriol. 92:1245-1251.
6. Bodmer, W. F., and S. Grether. 1965. Uptakeand incorporation ofthymine, thymidine, ur-acil and 5-fluorouracil into the nucleic acidsof Bacillus subtilhs. J. Bacteriol. 89-.1011-1014.
7. Budman, D. R., and A. B. Pardee. 1967. Thy-midine and thymine incorporation intoDNA: inhibition and repression by uridine ofthymidine phosphorylase ofEscherichia coli.J. Bacteriol. 94:1546-1550.
8. Caldwell, D. E., and P. Hirsch. 1973. Growthof micro-organisms in two-dimensionalsteady-state diffusion gradients. Can. J. Mi-crobiol. 19-.53-58.
9. Carr, N. G., and S. Bradley. 1973. Aspects ofdevelopment in blue-green algae, p. 161-188.In J. M. Ashworth and J. E. Smith (ed.),
VOL. 41, 1977

806 WHITTENBURY AND DOW
Microbial differentiation. Symposium of theSociety for General Microbiology 23. Cam-bridge University Press, Cambridge.
10. Carr, N. G., and G. Excell. 1965. Ubiquinoneconcentrations in Athiorhodaceae grown un-der various environmental conditions. Bio-chem. J. 96:688-692.
11. Conti, S. F., and C. R. Benedict. 1962. Carote-noids ofRhodomicrobium vannielii. J. Bacte-riol. 83:929-930.
12. Conti, S. F., and P. Hirsch. 1965. Biology ofbudding bacteria. III. Fine structure ofRho-domicrobium and Hyphomicrobium spp. J.Bacteriol. 89:503-512.
13. Cooper, S., and C. E. Helmstetter. 1968. Chro-mosome replication and the division cycle ofEscherichia coli B/r. J. Mol. Biol. 31:519-540.
14. Cruickshank, R. 1968. Giemsa's stain, p. 667-669. In R. Cruickshank (ed.), Medical micro-biology. E. and S. Livingston, London.
15. Degnen, S. T., and A. Newton. 1972. Chromo-some replication during development in Cau-lobacter crescentus. J. Mol. Biol. 64:671-680.
16. Degnen, S. T., and A. Newton. 1972. Depend-ence of cell division on the completion ofchromosome replication in Caulobacter cres-centus. J. Bacteriol. 110:852-856.
17. Donachie, W. D., and K. J. Begg. 1970. Growthof the bacterial cell. Nature (London)227:1220-1224.
18. Donachie, W. D., N. C. Jones, and R. Teather.1973. The bacterial cell cycle, p. 944. In J.M. Ashworth and J. E. Smith (ed.), Micro-bial differentiation. Symposium of the Soci-ety for General Microbiology 23. CambridgeUniversity Press, Cambridge.
19. Donkersloot, J. A., S. A. Robrish, and M. I.Krichevsky. 1972. Fluorimetric determina-tion of DNA in bacteria with ethidium bro-mide. Appl. Microbiol. 24:179-183.
20. Douglas, H. C., and R. S. Wolfe. 1959. Motilityof Rhodomicrobium vannielii. J. Bacteriol.78:597-598.
21. Dow, C. S., D. Westmacott, and R. Whitten-bury. 1976. Ultrastructure of budding andprosthecate bacteria, p. 187-221. In R. Fullerand D. W. Lovelock (ed.), Microbial ultra-structure. Society for Applied BacteriologyTechnical Series 10. Academic Press Inc.,New York.
22. Duchow, E., and H. C. Douglas. 1949. Rho-domicrobium vannielii, a new photohetero-trophic bacterium. J. Bacteriol. 58:409-416.
23. Dworkin, M. 1973. Cell-cell interactions in themyxobacteria, p. 125-142. In J. M. Ashworthand J. E. Smith (ed.), Microbial differentia-tion. Symposium of the Society for GeneralMicrobiology 23. Cambridge UniversityPress, Cambridge.
24. Ensign, J. C., and R. S. Wolfe. 1964. Nutri-tional control of morphogenesis in Arthro-bacter crystallopoietes. J. Bacteriol. 87:924-932.
25. Evans, E. H., I. Foulds, and N. G. Carr. 1976.Environmental conditions and morphologi-
BACTERIOL. REV.
cal variation in the blue-green alga Chlorog-loea fritschii. J. Gen. Microbiol. 92:147-155.
26. Freese, E. 1972. Sporulation of Bacilli, a modelof cellular differentiation. Curr. Top. Dev.Biol. 7:85-120.
27. Gorlenko, V. M. 1969. Spore formation in abudding photoheterotrophic bacterium (inEnglish). Microbiol. U.S.S.R. 38:106-111.
28. Gorlenko, V. M., N. N. Egorova, and A. N.Puchkov. 1974. Fine structure ofexospores ofnon-sulphur purple bacterium Rhodomicro-bium vannielii. Mikrobiologiya 43:1076-1078.
29. Goss, W. A., W. H. Deitz, and T. M. Cook.1965. Mechanism of action of nalidixic acidon Escherichia coli. II. Inhibition of deoxyri-bonucleic acid synthesis. J. Bacteriol.89:1068-1074.
30. Halvorson, H. 0. 1965. Sequential expressionof biochemical events during intracellulardifferentiation, p. 343-368. In M. R. Pollockand M. H. Richmond (ed.), Function andstructure in micro-organisms. Symposium ofthe Society for General Microbiology 15.Cambridge University Press, Cambridge.
31. Helmstetter, C. E. 1967. Rate of DNA synthe-sis during the division cycle of Escherichiacoli B/r. J. Mol. Biol. 24:417-427.
32. Herbert, R. A., and A. J. Holding. 1972. Rapidseparation and estimation of gases producedor utilized by micro-organisms. J. Chro-matogr. Sci. 10:174-175.
33. Hirsch, P. 1974. Genus HyphomicrobiumStutzer and Hartleb 1898, p. 148-150. In R.E. Buchanan and N. E. Gibbons (ed.), Ber-gey's manual of determinative bacteriology,8th ed. The Williams & Wilkins Co., Balti-more.
34. Hirsch, P. 1974. Budding bacteria. Annu. Rev.Microbiol. 28:392-444.
35. Hoare, D. S., and S. L. Hoare. 1969. Hydrogenmetabolism by Rhodomicrobium vannielii.J. Bacteriol. 100:1124-1125.
36. Ishiguro, E. E., and R. S. Wolfe. 1970. Controlof morphogenesis in Geodermatophilus: ul-trastructural studies. J. Bacteriol. 104:566-580.
37. Ishiguro, E. E., and R. S. Wolfe. 1974. Induc-tion of morphogenesis in Geodermatophilusby inorganic cations and by organic nitroge-nous cations. J. Bacteriol. 117:189-195.
38. Jacob, F., S. Brenner, and F. Cuzin. 1963. Onthe regulation of DNA replication in bacte-ria. Cold Spring Harbor Symp. Quant. Biol.28:329-348.
39. Jensen, A., 0. Aasmundrud, and K. E.Eimhjellen. 1964. Chlorophylls of photosyn-thetic bacteria. Biochim. Biophys. Acta88:466-479.
40. Jones, N. C., and W. D. Donachie. 1973. Chro-mosome replication, transcription and con-trol of cell division in Escherichia coli. Na-ture (London) New Biol. 243:100-103.
41. Kersten, H., and W. Kersten. 1974. Inhibitorsof nucleic acid synthesis. Biophysical andbiochemical aspects. Molecular biology, bio-

VOL. 41, 1977
chemistry and biophysics, 18. Chapman andHall Ltd., London.
42. Krulwich, T. A., and J. C. Ensign. 1969. Alter-ation of glucose metabolism of Arthrobactercrystallopoietes by compounds which inducesphere to rod morphogenesis. J. Bacteriol.97:526-534.
43. Kurn, N., and L. Shapiro. 1975. Regulation ofthe Caulobacter cell cycle. Curr. top. Cell.Regul. 9:41-62.
44. Lazaroff, N. 1973. Photomorphogenesis andNostocacean development, p. 279-319. In N.G. Carr and B. A. Whittan (ed.), The biologyof the blue-green algae. Blackwell Scientific,Oxford.
45. Leifson, E. 1964. Hyphomicrobium neptuniumsp. n. Antonie van Leeuwenhoek J. Micro-biol. Serol. 13:249-256.
46. Liaaen-Jensen, S., and A. G. Andrewes. 1972.Microbial carotenoids. Annu. Rev. Micro-biol. 26:225-244.
47. Mandel, M., and J. Marmur. 1968. Use ofultra-violet absorbance-temperature profile for de-termination ofthe guanine plus cytosine con-tent of DNA. Methods Enzymol. 12B:195-206.
48. Mandel, M., C. L. Schildkraut, and J. Mar-mur. 1968. Use of CsCl density gradientanalysis for determining the guanine pluscytosine content of DNA. Methods Enzymol.12B:184-195.
49. Mandelstam, J. 1969. Regulation of bacterialspore formation, p. 377402. In P. Meadowand S. J. Pirt (ed.), Microbial growth. Sym-posium of the Society for General Microbiol-ogy 19. Cambridge University Press, Cam-bridge.
50. Mitchison, J. M., and W. S. Vincent. 1965.Preparation of synchronous cell cultures bysedimentation. Nature (London) 205:987-989.
51. Mitchison, G. J., and M. Wilcox. 1972. Rulegoverning cell division inAnabaena. Nature(London) 239:110-111.
52. Moore, R. L., and P. Hirsch. 1973. First gener-ation synchrony of isolated Hyphomicrobiumswarmer populations. J. Bacteriol. 116:418-423.
53. Morita, S., and S. F. Conti. 1963. Localizationand nature of the cytochromes of Rhodomi-crobium vannielii. Arch. Biochem. Biophys.100:302-307.
54. Mulder, E. G., and W. L. van Veen. 1963.Investigation on the Sphaerotilus-Leptothrixgroup. Antonie van Leeuwenhoek J. Micro-biol. Serol. 29:121-153.
55. Murray, R. G. E., and H. C. Douglas. 1950.The reproductive mechanism ofRhodomicro-bium vannielii and the accompanying nu-clear changes. J. Bacteriol. 59:157-167.
56. Murray, R. G. E., and S. W. Watson. 1965.Structure of Nitrosocystis oceanus and com-parison with Nitrosomonas and Nitrobacter.J. Bacteriol. 89:1594-1609.
57. Newton, A. 1972. Role of transcription in the
MORPHOGENESIS OF R. VANNIELII 807
temporal control of development in Caulo-bacter crescentus. Proc. Natl. Acad. Sci.U.S.A. 69:447451.
58. Park, C. E., and L. R. Berger. 1967. Complexlipids of Rhodomicrobium vannielii. J. Bac-teriol. 93:221-229.
59. Park, C. E., and L. R. Berger. 1967. Fattyacids of extractable and bound lipids ofRho-domicrobium vannielii. J. Bacteriol. 93:230-236.
60. Pfennig, N. 1969. Rhodopseudomonas acido-phila, sp. n., a new species of the buddingpurple non-sulphur bacteria. J. Bacteriol.99:597-602.
61. Pfennig, N., and K. D. Lippert. 1966. Uber dasVitamin B12-Bedurfnis phototropher SchiveFelbakterien. Arch. Mikrobiol. 55:258-266.
62. Pope, L. M., D. S. Hoare, and A. J. Smith.1969. Ultrastructure of Nitrobacter agilisgrown under autotrophic and heterotrophicconditions. J. Bacteriol. 97:936-939.
63. Purohit, K., B. A. McFadden, and M. M.Shaykh. 1976. D-Ribulose-1,5-bisphosphatecarboxylase and polyhedral inclusion bodiesin Thiobacillus intermedius. J. Bacteriol.127:516-522.
64. Quadri, S. M. H., D. S. Hoare, and S. L.Hoare. 1972. Metabolism of pyruvate by pur-ple nonsulfur bacteria. In Abstr. Annu.Meet. Am. Soc. Microbiol. 1973, P170, p. 164.
65. Quayle, J. R., and N. Pfennig. 1975. Utiliza-tion of methanol by Rhodospirillaceae. Arch.Mikrobiol. 102:193-198.
66. Rogers, S. R., and J. J. Anderson. 1976. Mea-surement of growth and iron deposition inSphaerotilus discophorus. J. Bacteriol.126:257-263.
67. Rosenkranz, H. S., A. J. Garro, J. A. Levy, andH. S. Carr. 1966. Studies with hydroxyurea.I. The reversible inhibition of bacterial DNAsynthesis and the effect of hydroxyurea onbacteriocidal action of streptomycin.Biochim. Biophys. Acta 114:501-515.
68. Ryter, A., and E. Kellenberger. 1958. Etude aumicroscope electronique de plasmas conten-ant de l'acide desoxyribonucleique I. Les nu-cleoides des bacteries en croissance active. Z.Naturforsch. 136:597-605.
69. Ryvarden, L., and S. Liaaen-Jensen. 1964.Bacterial carotenoids. XIV. The carotenoidsof Rhodomicrobium vannielii. Acta Chem.Scand. 18:643-654.
70. Sandhu, G. R., and N. G. Canr. 1970. A novelalcohol dehydrogenase present in Rhodomi-crobium vannielii. Arch. Mikrobiol. 70:340-347.
71. Sargent, M. G. 1973. Synchronous cultures ofBacillus subtilis obtained by filtration withglass fiber filters. J. Bacteriol. 116:736-740.
72. Schmidt, J. M., and R. Y. Stainer. 1966. Thedevelopment of cellular stalks in bacteria. J.Cell Biol. 28:423-436.
73. Schwan, H., and A. W. Holldorf. 1975. Effec-tive utilization of exogenous deoxythymi-dine-5'-mono-phosphate for DNA synthesis

808 WHITTENBURY AND DOW
in enterobacteria. FEBS Lett. 57:179-182.74. Shapiro, L. 1976. Differentiation in the Caulo-
bacter cell cycle. Annu. Rev. Microbiol.30:377-402.
75. Shapiro, L., and N. Agabian-Keshishian. 1970.Specific assay for differentiation in thestalked bacterium Caulobacter crescentus.Proc. Natl. Acad. Sci. U.S.A. 67:200-203.
76. Shapiro, L., N. Agabian-Keshishian, and I.Bendis. 1971. Bacterial differentiation. Sci-ence 173:884-892.
77. Shapiro, L., and J. V. Maizel, Jr. 1973. Synthe-sis and structure of Caulobacter crescentusflagella. J. Bacteriol. 113:478-485.
78. Smillie, R. M., and R. C. Fuller. 1960. Furtherobservations on glyceraldehyde 3-phosphatedehydrogenases in plants and photosyntheticbacteria. Biochem. Biophys. Res. Commun.3:368-372.
79. Staley, J. T. 1968. Prosthecomicrobium andAn-calomicrobium: new prothecate freshwaterbacteria. J. Bacteriol. 95:1921-1942.
80. Staley, J. T., and T. L. Jordan. 1973. Cross-bands of Caulobacter crescentus stalks serveas indicators of cell age. Nature (London)246:155-156.
81. Stove, J. L., and R. Y. Stanier. 1962. Cellulardifferentiation in stalked bacteria. Nature(London) 196:1189-1192.
82. Szulmajster, J. 1973. Initiation of bacterialsporogenesis, p. 45-83. In J. M. Ashworthand J. E. Smith (ed.), Microbial differentia-tion. Symposium of the Society for GeneralMicrobiology 23. Cambridge UniversityPress, Cambridge.
83. Szybalski, W., and V. N. Iyer. 1964. Crosslink-ing of DNA by enzymatically or chemicallyactivated mitomycins and profinomycins, bi-functionally "alkylating" antibiotics. Fed.Proc. 23:946-957.
84. Tauschel, H. D., and J. F. M. Hoeniger. 1974.The fine structure ofRhodopseudomonas aci-dophila. Can. J. Microbiol. 20:13-17.
85. Tomesik, J., and J. B. Baumann-Grace. 1959.Specific exosporium reaction ofBacillus meg-aterium. J. Gen. Microbiol. 21:666-675.
86. Trentini, W. C. 1967. Defined medium allowingmaximal growth ofRhodomicrobium vannie-lii. J. Bacteriol. 94:1260-1261.
87. Trentini, W. C., and M. P. Starr. 1967. Growthand ultrastructure of Rhodomicrobium van-nielii as a function of light intensity. J. Bac-teriol. 93:1699-1704.
88. Tyler, P. A., and K. C. Marshall. 1967. Pleo-
morphy in stalked budding bacteria. J. Bac-teriol. 93:1132-1136.
89. Ulitzur, S., and M. Kessel. 1973. Giant flagel-lar bundles of Vibrio alginolyticus. Arch.Mikrobiol. 94:331-339.
90. Van Niel, C. B. 1944. The culture, generalphysiology, morphology and classification ofthe non-sulphur purple and-brown bacteria.Bacteriol. Rev. 8:1-118.
91. Vatter, A. E., H. C. Douglas, and R. S. Wolfe.1959. Structure of Rhodomicrobium vannie-lii. J. Bacteriol. 77:812-813.
92. Volk, W. A., and D. Pennington. 1949. Thepigments of the photosynthetic bacteriumRhodomicrobium vannielii. J. Bacteriol.59:169-170.
93. Westmacott, D., and S. B. Primrose. 1976.Synchronous growth of Rhodopseudomonaspalustris from the swarmer phase. J. Gen.Microbiol. 94:117-125.
94. Whittenbury, R. 1971. Enrichment and isola-tion of photosynthetic bacteria, p. 241-249.In D. A. Shapton and R. G. Board (ed.), TheSociety for Applied Bacteriology TechnicalSeries 5. Academic Press Inc., New York.
95. Whittenbury, R., S. L. Davies, and J. F.Davey. 1970. Exospores and cysts formed bymethane-utilizing bacteria. J. Gen. Micro-biol. 61:219-226.
96. Whittenbury, R., and A. G. McLee. 1967. Rho-dopseudomonas palustris and R. viridis-photosynthetic budding bacteria. Arch. Mik-robiol. 59:324-334.
97. Wilcox, M., G. J. Mitchison, and R. J. Smith.1973. Pattern formation in the blue-greenalga, Anabaena. I. Basic mechanisms. J.Cell Sci. 12:707-723.
98. Wilcox, M., G. J. Mitchison, and R. J. Smith.1973. Pattern formation in the blue-greenalga, Anabaena. II. Controlled proheterocystregression. J. Cell Sci. 13:627-649.
99. Woodfolk, C. A., and H. R. Whiteley. 1962.Reduction of inorganic compounds with mo-lecular hydrogen by Micrococcus lactilyticus.J. Bacteriol. 84:647-658.
100. Worcel, A., and E. Burgi. 1974. Properties of amembrane-attached form of the folded chro-mosome of Escherichia coli. J. Mol. Biol.82:91-105.
101. Yamaguchi, K., and H. Yoshikawa. 1973. To-pography of chromosome membrane junctionin Bacillus subtilis. Nature (London) NewBiol. 244:204-206.
BACTZRIOL. REV.




















