
Cyanobacterial Composition and Microcystin Accumulation in ...

Environmental Toxicology and Pharmacology 19 (2005) 297–303
Comparison of the structure of key variants of microcystin to vasopressin
Michelle M. Gehringera,∗, Pieter Milneb, Franco Luciettob, Tim G. Downingc
a School of Biotechnology and Biomolecular Sciences, University of New South Wales, Sydney 2052, Australiab Department of Pharmacy, University of Port Elizabeth, P.O. Box 1600, Port Elizabeth 6000, South Africa
c Department of Biochemistry and Microbiology, University of Port Elizabeth, P.O. Box 1600, Port Elizabeth 6000, South Africa
Received 13 March 2004; accepted 3 August 2004Available online 7 October 2004
Abstract
Microcystins (MCs) are cyclic heptapeptide compounds [where X2 (position 2) and Z4 (position 4) are variablel-aminoacids] produced bycyanobacteria and responsible for severe liver damage in animals ingesting acute doses of the toxic compounds. Certain variants of microcystinsare more toxic than others, the differences being commonly ascribed to the hydrophobic nature of the variant. Microcystin-LR (MCLR) [X= l-leucine (L); Z =l-arginine (R); R1 = R2 = CH] is the most toxic of all the microcystins investigated to date. This study investigatest mpiled inH areas andm mso©
K
1
heem1tgatattaX
arecefer-atu-
9mino
o
fars in9eirand-si-
rhag-r-as
1d
3
he similarity of the structures of MCLR and selected MC variants to the liver specific hormone vasopressin. Structures were coyperChem® (professional version 5.1). Initial comparisons of the MCLR and vasopressin indicated comparable volumes, surfaceasses. Further studies using RMS overlays show that the microcystin derivative MCLR(Dha7) is comparably similar to vasopressin in terf tertiary structure.2004 Elsevier B.V. All rights reserved.
eywords:Microcystin; Vasopressin; Polak-Ribiere gradient; RMS overlay
. Introduction
Microcystins are recognized as potent liver toxins inumans and animals (Falconer, 2001; Kuiper-Goodmant al., 1999). They are produced by different gen-ra of cyanobacteria in freshwater bodies, most com-only Microcystis, Anabaenaand Oscillatoria (Sivonen,996). Microcystin consists of a small, cyclic hep-
apeptide: cyclo-(d-alanine1-X2-d-MeAsp3-Z4-ADDA5-d-lutamate6-Mdha7) shown in Fig. 1. d-MeAsp, the thirdmino acid is d-erythro-�-methylaspartic acid. Mdha,
he seventh amino acid isN-methyldehydroalanine andn unusual moiety, (2S,3S,8S,9S)-3-amino-9-methoxy-2,6,8-
rimethyl-10-phenyl-deca-4,6-dienoic acid (ADDA), at posi-ion 5, forms the distinctive side arm of this toxin (Botes etl., 1985). The two variablel-amino acids are denoted as2 and Z4 in Fig. 1. The microcystin nomenclature rests on
∗ Corresponding author. Tel.: +61 2 9385 3235; fax: +61 2 9385 1591.E-mail address:[email protected] (M.M. Gehringer).
the composition of these two variablel-amino acids withmicrocystin-LR, in which the two variable amino acidsleucine2 (L) and arginine4 (R), being the most toxic and henmost commonly studied microcystin. More than 60 difent analogues of microcystin have been identified from nral blooms or laboratory cultures (Sivonen and Jones, 199).Structural variations have been reported in all seven aacids, but most frequently with substitition ofl-amino acidsat positions 2 (X2) and 4 (Z4) and demethylation of aminacids at position 3 and/or 7 (Ito et al., 2002, Vesterkvist andMeriluoto, 2003). Minor changes in structure may havereaching implications in toxicity as reflected by differencethe intraperitoneal LD50 in mice (Sivonen and Jones, 199).
The toxic effects of MC can be attributed to thinhibition of protein phosphatases 1 and 2A (PP1PP2A) (MacKintosh et al., 1990) resulting in hyperphosphorylation of the cellular cytoskeleton. The resultantnusoidal collapse leads to severe intrahepatic heamoing and in severe cases, death (Falconer, 2001; KuipeGoodman et al., 1999). Protein phosphatase inhibition w
382-6689/$ – see front matter © 2004 Elsevier B.V. All rights reserved.oi:10.1016/j.etap.2004.08.006

298 M.M. Gehringer et al. / Environmental Toxicology and Pharmacology 19 (2005) 297–303
Fig. 1. The generic structure for microcystins. Microcystin-LR has anl-leucine at position X andl-arginine at position Z (Ito et al., 2002). R1 and R2 representmethyl groups ford-erythro-�-methylaspartic acid andN-methyldehydroalanine, respectively. = anchor molecules.
demonstrated in vitro for MCLR, MCYR (tyrosine–arginine)and MCRR (arginine–arginine) in mouse liver lysates(Yoshizawa et al., 1990) and in vivo for MCLR and MCYM(tyrosine–methionine) (Runnegar et al., 1993). [125I]MCYRbinds both PP1 and PP2A covalently at Cys-273 and Cys-266, respectively (MacKintosh et al., 1995; Runnegar et al.,1995a,b). Western blotting of liver homogenates of mice ex-posed to MCLR confirmed the covalent binding of MCLRto nuclear PPs (Guzman and Solter, 2002). There is consid-erable flexibility in the active site of protein phosphatase 1,which would allow a potentially wide range of MC variants tobind to Cys-273 on PP1, thereby allowing for the wide rangeof MC showing inhibition of protein phosphatases (Lavigneet al., 2000).
MCRR (arginine–arginine) is less toxic than MCLR withan LD50 in mice of 600�g/kg (Watanabe et al., 1988) com-pared to the 50�g/kg of MCLR (Botes et al., 1985). MCRRis 1/12 as toxic as MCLR when administered intratracheallyto mice and yet, both toxins inhibit PP1 and PP2A to a similardegree (Ito et al., 2002). It is apparent that the lethality of aMC analogue appears to be more closely related to uptakemechanisms.
Uptake of MC into the liver occurs via a multispecific biletransport system and has been demonstrated for both MCLRand MCYM (tyrosine–methionine) (Runnegar et al., 1995b;E orh ed byeM a-t -a Thef gestsa cellm
orh the
often present arginine at the variable amino acid in posi-tion 2 or 4 (Kuiper-Goodman et al., 1999). Their hydropho-bicity varies depending on variations to the ADDA moietyand incorporation of hydrophobic amino acids in the vari-able positions X and Z. MCLR, the most commonly oc-curring MC and the most toxic, is more hydrophobic thanMCRR, which is less toxic, leading to suggestions that thehydrophobicity of microcystins may enhance their penetra-tion capacity into lipid bilayers and hence their toxic ef-fect in the cell (Vesterkvist and Meriluoto, 2003). Wardand Codd (1999)demonstrated increased toxicity of MCLF(leucine–phenylalanine) and MCLW (leucine–tryptophan) toMCLR in cells of the protozoa,Tetrahymena pyriformis.MCLW and particularly MCLF showed better membranepenetrating capacity than observed for MCLR (Vesterkvistand Meriluoto, 2003). Both of these studies suggested thattoxins with greater hydrophobicity demonstrated greatermembrane penetration ability and, hence increased toxicity.
Vasopressin is a nonapeptide neurohyphoseal hormonesynthesised in the hypothalamus. Its major functions are reg-ulation of readsorption of water in the kidneys and contrac-tion of the smooth muscles in arteries, thereby regulatingvasodilation (Barberis et al., 1998). Vasopressin receptor isfound in the liver, smooth muscle cells in arteries and mostperipheral tissues (Mouillac et al., 1995). Binding of va-s bal-a lecu-l nr s orho nistb ptorae tesc osi-t IP3
riksson et al., 1990) however the identity of the receptas not been identified. Non-hepatocyte cells are affectxposure to MC at higher concentrations of toxin (100�MCLR) than required to elicit similar toxic effects in hep
ocytes (0.8�M MCLR) (McDermott et al., 1998), presumbly due to the lack of an active bile transport system.
act that non-hepatic cells are able to take up MC sugnother means of uptake, possibly diffusion through theembrane.Microcystins are amphipathic molecules with polar
ydrophilic carboxylic acids in positions 3 and 6 and
opressin to its receptor is characterised by a delicatence of hydrogen bonds as opposed to a few key mo
ar points of interaction (Mouillac et al., 1995). Vasopressieceptor antagonists virtually all carry bulky sidechainydrophobic ring structures (Barberis et al., 1998). Antag-nist binding is not affected by mutations to key agoinding sites, indicating different subsites of the recere involved in hormone/antagonist interactions (Mouillact al., 1995). Binding of vasopressin to its receptor medialeavage of phosphatidylinositol 4,5-bisphosphate to inol 1,4,5-triphosphate (IP3) and diacylglycerol (DAG).

M.M. Gehringer et al. / Environmental Toxicology and Pharmacology 19 (2005) 297–303 299
subsequently activates the release of Ca2+ from the endo-plasmic reticulum (Michell et al., 1979). Activation of thevasopressin receptor also results in activation of signalingpathways within the hepatocyte, including activation of phos-pholipases A2, C and D (Briley et al., 1994).
Microcystin potently inhibits PP2A thereby affectingthe regulation of several mitogen activated protein kinases(MAPK). Okadaic acid, another inhibitor of protein phos-phatases is known to activate a variety of MAPKs. It wasproposed that this activation proceeded via tyrosine kinase-linked protein receptors, which would in turn activate phos-pholipase C. The ensuing cleavage of phosphatidylinositol4,5-bisphosphate would result in DAG activation of proteinkinase C, while IP3 would induce Ca2+ release from the ER(Gehringer, 2004; Rossini, 2000). Hepatocytes exposed toMC exhibit increased release of Ca2+ into the cytoplasm(Ding et al., 2000). This is accompanied by an increase inactivity of the calcium regulated phospholipase A2 in the kid-neys of exposed animals (Nobre et al., 2001). Fladmark et al.(2002)proved that protein kinase C, an activator of phospho-lipase A2, is not activated after exposure to MCLR, suggest-ing that cleavage of phosphatidylinositol 4,5-bisphosphatedid not occur.Macias-Silva and Garcia-Sainz (1994)alsofound that both protein phosphatase inhibitors, microcystinand okadaic acid inhibited vasopressin induced IP3 produc-t mayp lingr
ss ofv ee ift y.
2
ssinw n5 cy-c m of3 r un-t asd i-e ctedo rmeri n-f r mi-c ed byt culew mostr tudya erlayw sc rlaysw aluew tives
Table 1The table of parameters used to determine the lowest energy conformers forall the microcystin variants and vasopressin
Type of search Usage directed
Acceptance energy criterion Maximum of 50 kcal/mol above the rest
Test options(a) Pre-optimization check Skip if atoms are closer than 0.5A
Skip if torsions are within 5◦ of previous
Duplication testsDuplicates if Energy is within 0.05 kcal/mol
RMS error within 0.25A
Optimization termination RMS gradient = 0.0001 kcal/(A mol−1)Maximum cycles 1000
Limits 10000 iterations or1000 optimizationsDon’t keep more than 10 conformations
The selected conformational isomer is then overlaid withthe lowest energy conformer of vasopressin, in order to viewthe similarities between the two structures, and ultimatelythe likelihood that they would experience similar activity atthe receptor site. This, however, is assuming that the twoagents act at the same active site and not for example at anallosteric one. The conformational search was carried outwith the parameters indicated inTable 1. The RMS errorbetween all the microcystin variants with known LD50 valuesand vasopressin were compared with the LD50 values in orderto determine any correlation or trends between LD50 andsimilarity to vasopressin.
3. Results
The results of the overlays between all the MC variants(lowest energy conformers) in order to find the most repre-sentative of the group of agents are given inTable 2. The LD50values are provided as determined from literature (Sivonenand Jones, 1999). MCYM(O) and MCLR were among themost representative of the group having a lower overall aver-age fit, and higher bioactivity with lower LD50 values of 56and 50�g/kg, respectively (Botes et al., 1985). A similar sit-uation is illustrated with the [(6Z)-ADDA]MCLR stereoiso-mh vingi ;Ka fore -r -t erec ergyc O)(
en-e est
ion at 1�M in rat hepatocytes. They suggested that MCerturbate phospholipase C activation via G-protein coupeactions.
The aim of this study was to investigate the relatedneariants of MC structure to vasopressin structure and shere was a link between structural similarity and toxicit
. Materials and methods
The structure of MCLR (and all variants) and vasopreere constructed using HyperChem® (professional versio.1). All agents in the study were minimized through 200les of a steepest descent gradient, followed by a maximu2,650 cycles of a Polak-Ribiere conjugated gradient o
il a minima of 0.001 kcal/(A mol−1) was reached. This wone utilizing an AMBER force field and the AM1 semmpirical method. A conformational search was condun each microcystin variant and the lowest energy confo
dentified (Table 1). Each microcystin lowest energy coormer variant was overlaid in sequence with every otherocystin derivative to obtain a best average fit representhe agent with the lowest mean for the overlays. The moleith the best average fit (lowest mean) is therefore the
epresentative of the entire group of microcystins under snd noted as a potential lead compound. The RMS ovas conducted using HyperChem®. Two anchor moleculeommon to all variants were selected to ensure that oveere not biased in any way. The resultant RMS error vas entered into an Excel® spreadsheet and basic descriptatistics were calculated.
er, MCLR(d-Asp3), MCLR(Dha7) and MCRR(Dha7), allaving less representative overall structures, but pro
n vitro to have higher LD50 values (Harada et al., 1990rishnamyrthy et al., 1989; Sivonen et al., 1992) andlso lower bioactivity. Plotting the mean RMS valuesach microcystin variant versus the LD50 yielded a corelation coefficient of 0.65 withp = 0.003. The quantitaive structure–activity relationship (QSAR) properties wompared for MCLR, vasopressin and the lowest enonformer for the microcystin variants, namely MCYM(Table 3).
The RMS errors obtained from overlaying the lowestrgy conformer of each microcystin variant with the low
300 M.M. Gehringer et al. / Environmental Toxicology and Pharmacology 19 (2005) 297–303
Table 2RMS values for each microcystin variant overlayed with every other microcystin derivative to obtain a best average fit using HyperChem®
MCLR MCLR[(6Z)-ADDA] MCYM(O) MCLR( d-Asp3) MCLR(Dha7) MCRR(Dha7)
MCLR 0.00 0.04 0.17 0.66 0.67 1.85MCLR[(6Z)-ADDA] 0 .04 0.00 0.13 1.76 1.78 0.92MCYM(O) 0.17 0.13 0.00 0.55 0.57 0.12MCLR(d-Asp3) 0.66 1.76 0.55 0.00 0.06 0.84MCLR(Dha7) 0.67 1.78 0.56 0.06 0.00 0.08MCRR(Dha7) 1.85 0.92 0.11 0.84 0.08 0.00Mean 0.565 0.7717 0.255 0.645 0.5267 0.633Standard deviation 0.58 0.71 0.25 0.56 0.64 0.59LD50 (�g/kg) 50 >1200 56 160–300 250 180
Two anchor molecules (Fig. 1) were selected to ensure that overlays were not biased in any way.
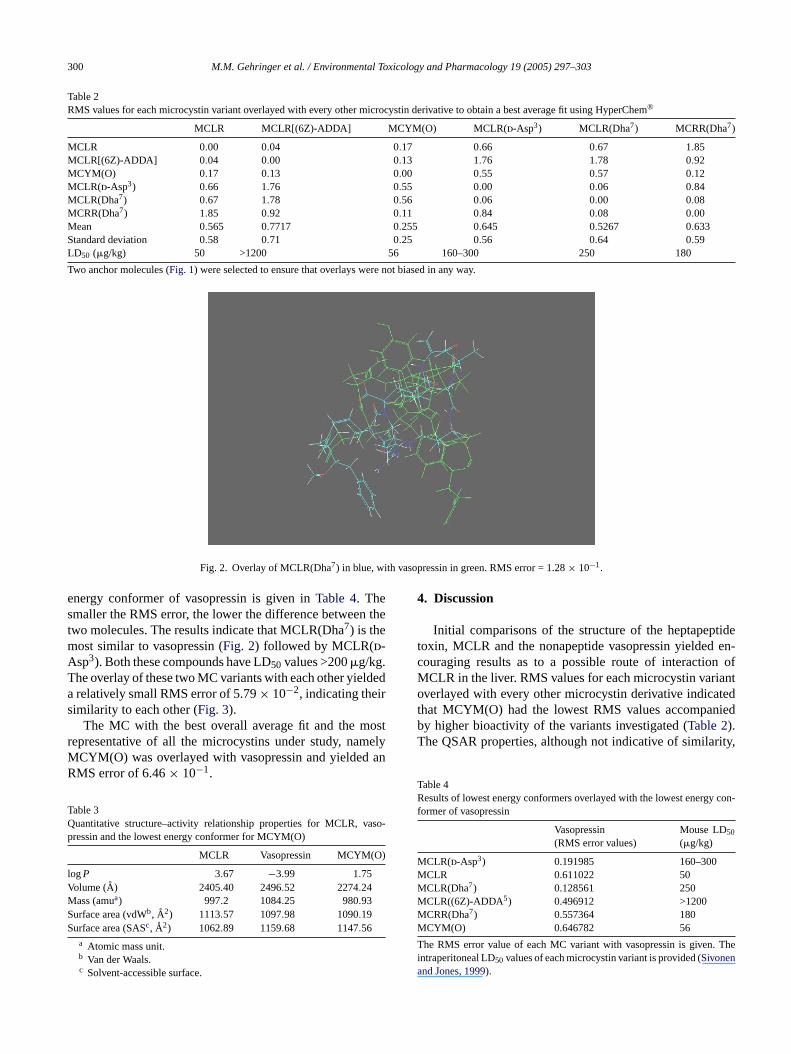
Fig. 2. Overlay of MCLR(Dha7) in blue, with vasopressin in green. RMS error = 1.28× 10−1.
energy conformer of vasopressin is given inTable 4. Thesmaller the RMS error, the lower the difference between thetwo molecules. The results indicate that MCLR(Dha7) is themost similar to vasopressin (Fig. 2) followed by MCLR(d-Asp3). Both these compounds have LD50 values >200�g/kg.The overlay of these two MC variants with each other yieldeda relatively small RMS error of 5.79× 10−2, indicating theirsimilarity to each other (Fig. 3).
The MC with the best overall average fit and the mostrepresentative of all the microcystins under study, namelyMCYM(O) was overlayed with vasopressin and yielded anRMS error of 6.46× 10−1.
Table 3Quantitative structure–activity relationship properties for MCLR, vaso-pressin and the lowest energy conformer for MCYM(O)
MCLR Vasopressin MCYM(O)
logP 3.67 −3.99 1.75Volume (A) 2405.40 2496.52 2274.24Mass (amua) 997.2 1084.25 980.93Surface area (vdWb, A2) 1113.57 1097.98 1090.19Surface area (SASc, A2) 1062.89 1159.68 1147.56
a Atomic mass unit.b Van der Waals.c Solvent-accessible surface.
4. Discussion
Initial comparisons of the structure of the heptapeptidetoxin, MCLR and the nonapeptide vasopressin yielded en-couraging results as to a possible route of interaction ofMCLR in the liver. RMS values for each microcystin variantoverlayed with every other microcystin derivative indicatedthat MCYM(O) had the lowest RMS values accompaniedby higher bioactivity of the variants investigated (Table 2).The QSAR properties, although not indicative of similarity,
Table 4Results of lowest energy conformers overlayed with the lowest energy con-former of vasopressin
Vasopressin(RMS error values)
Mouse LD50
(�g/kg)
MCLR(d-Asp3) 0.191985 160–300MCLR 0.611022 50MCLR(Dha7) 0.128561 250MCLR((6Z)-ADDA5) 0.496912 >1200MCRR(Dha7) 0.557364 180MCYM(O) 0.646782 56
The RMS error value of each MC variant with vasopressin is given. Theintraperitoneal LD50 values of each microcystin variant is provided (Sivonenand Jones, 1999).

M.M. Gehringer et al. / Environmental Toxicology and Pharmacology 19 (2005) 297–303 301
Fig. 3. Overlay of MCLR(d-Asp3) in blue over MCLR(Dha7) in green. RMS error = 5.79× 10−2.
also showed close similarities (Table 3). The logP values,indicative of hydrophobicity, are quite diverse. This couldbe attributed largely to the presence of the ADDA moi-ety of microcystin, which is hydrophobic (Vesterkvist andMeriluoto, 2003). Overlays of MCLR(Dha7), MCYM(O) andMCLR with vasopressin indicate that at all times the ADDAmoiety is external to the overlapping regions showing thelowest RMS values (Figs. 2, 4 and 5, respectively).
Comparisons of several variants of MCLR with knownLD50 values, and their variants were done (Table 4). No linkcould be made between structural representivity and observedLD50 values. A possible reason for this paradox between av-erage fit and activity may be due to specific groups suchas methionine oxide (MCYM(O)) and/or tyrosine–arginineresidues containing essential pharmacophores needed forbioactivity or to enhance bioactivity. Further modeling to de-termine actual pharmacophores needs to be done. This may
th vaso
be best achieved by making use of networking strategies.Further investigations into the receptor action of microcystinvariants with regard to their action as agonists or antagonistsis warranted in order to more accurately describe the resultsobserved. As previously mentioned, the binding of knownantagonists to the vasopressin receptor are not affected bymutations to key agonist binding sites indicating that the an-tagonists identified to date bind at different sites on the re-ceptor than vasopressin itself. This has not been shown formicrocystin, which may bind to the vasopressin receptor site.The literature indicates a role for microcystin inhibition ofvasopressin binding (Macias-Silva and Garcia-Sainz, 1994),which may indicate a potential overlap of binding sites, if notdirect competition. This is supported by the fact that the toxinsshowing greater similarity to vasopressin were the less toxicvariants and would imply therefore a decrease in vasopressin-induced IP3 release.
Fig. 4. Overlay of MCYM(O) in blue, wi
pressin in green. RMS error = 6.46× 10−1.
302 M.M. Gehringer et al. / Environmental Toxicology and Pharmacology 19 (2005) 297–303
Fig. 5. Overlay of MCLR in blue, with vasopressin (green). RMS error = 6.11× 10−1.
Experiments in silico do not attempt to replace wet workin the laboratory, but serve to narrow the range of investiga-tions and exclude hypothesis that clearly have no chemicalgrounding. To date molecular mimicry has not been consid-ered in the toxicology of MCLR. Various hypotheses havebeen presented as to the great range of toxicities of MC vari-ants, however all have proved inconclusive. The results pre-sented in this investigation have highlighted the use of molec-ular modeling techniques in studying the potential bioactiv-ity of compounds as well as emphasizing the need for invitro/in vivo investigations to confirm or disprove the resultsobtained.
References
Barberis, C., Mouillac, B., Durroux, T., 1998. Structural bases of caso-pressin/oxytocin receptor function. J. Endocrinol. 156, 223–229.
Botes, D.P., Wessels, P.L., Kruger, H., 1985. Structural studies oncyanoginosins-LR, -YR, -YA, and -YM, peptide toxins fromMicro-cystis aeruginosa. Chem. Soc. Perkin Trans. 1, 2747–2748.
Briley, E., Lolait, S., Axelrod, J., Felder, C., 1994. The cloned vasopressinreceptor stimulated phospholipase A2, phospholipase C and phospho-lipase D through activation of receptor-operated calcium channels.Neuropeptides 27, 63–74.
Ding, W.-X., Shen, H.-M., Ong, C.-N., 2000. Critical role of reactive oxy-stin-–555.
E A.O.,c
F alian–233.
F øe,02.for
G lular
Guzman, R.E., Solter, P.F., 2002. Characterisation of sublethalmicrocystin-LR in mice. Vet. Pathol. 39, 17–26.
Harada, K.I., Ogawa, K., Matsuura, K., Murata, H., Suzuki, M., Watan-abe, M.F., Itezono, Y., Nakayma, N., 1990. Structural determinationof geometrical isomers of microcystins-LR and -RR from cyanobac-teria by two dimensional NMR spectroscopic techniques. Chem. Res.Toxicol. 3, 473–481.
Ito, E., Takai, A., Kondo, F., Masui, H., Imanishi, S., Harada, K.,2002. Comparison of protein phosphatase inhibitory activity and ap-parent toxicity of microcystins and related compounds. Toxicon 40,1017–1025.
Kuiper-Goodman, T., Falconer, I., Fitzgerald, J., 1999. Human health as-pects. In: Chorus, I., Bartram, J. (Eds.), Toxic Cyanobacteria in Water:A Guide to Public Health Consequences, Monitoring and Manage-ment. E & FN Spon, London, pp. 114–141.
Krishnamyrthy, T., Szafraniec, L., Hunt, D.F., Shabanowitz, J., Yates,Y.R., Hauer, C.R., Carmichael, W.W., Skulberg, O., Codd, G.A.,Missler, S., 1989. Structural characterization of toxic cyclic peptidesfrom blue-green algae by tandem mass spectrometry. Proc. Natl. Acad.Sci. U.S.A. 86, 770–774.
Lavigne, P., Bagu, J.R., Boyko, R., Willard, L., Holmes, C.F.B., Sykes,B.D., 2000. Structure based thermodynamic analysis of the dissoci-ation of protein phosphatase-1 catalytic subunit and microcystin-LRdocked complexes. Protein Sci. 9, 252–264.
MacKintosh, C., Beattie, K.A., Klumpp, S., Cohen, P., Codd, G.A., 1990.Cyanobacterial microcystin-LR is a potent and specific inhibitor ofprotein phosphatases 1 and 2A from both mammals and higher plants.FEBS Lett. 264, 187–192.
MacKintosh, R.W., Dalby, K.N., Campbell, D.G., Cohen, P.T.W., MacKin-tosh, C., 1995. The cyanobacterial toxin microcystin binds covalently
–240.M ne-
kele-ine,
M hevari-
M ofhep-
M yer,ding
gen species and mitochondrial permeability transition in microcyinduced rapid apoptosis in rat hepatocytes. Hepatology 32, 547
riksson, J.E., Gronberg, L., Nygard, S., Slotte, J.P., Neriloto, J.1990. Hepatocellular uptake of3H-dihydromicrocystin-LR a cyclihepatotoxin. Biochem. Biophys. Acta 1025, 60–66.
alconer, I.R., 2001. Toxic cyanobacterial bloom problems in Austrwaters: risks and impacts on human health. Phycologia 40, 228
ladmark, K.E., Brustugun, O.T., Mellgren, G., Krakstad, C., BR., Vintermeyr, O.K., Schulman, H., Døskeland, O., 20Ca2+/calmodulin-dependent protein kinase II is requiredmicrocystin-induced apoptosis. J. Biol. Chem. 277, 2804–2811.
ehringer, M.M., 2004. Microcystin-LR and okadaic acid-induced celeffects: a dualistic response. FEBS Lett. 557, 1–8.
to cysteine-273 on protein phosphatase 1. FEBS Lett. 371, 236acias-Silva, M., Garcia-Sainz, J.A., 1994. Inhibition of hormo
stimulated inisitol phosphate productions and disruption of cytostal structure. Effects of okadaic acid, microcystin, chlorpromazW7 and nystatin. Toxicon 32, 105–112.
cDermott, C.M., Nho, C.W., Howard, W., Holton, B., 1998. Tcyanobacterial toxin, microcystin-LR, can induce apoptosis in aety of cell types. Toxicon 36, 1981–1996.
ichell, R.H., Kirk, C.J., Billah, M.M., 1979. Hormonal stimulationphosphatidylinositol breakdown with particular reference to theatic effects of vasopressin. Biochem. Soc. Trans. 7, 861–865.
ouillac, B., Chini, B., Balestre, M.N., Elands, J., Trumpp-KallmeS., Hoflack, J., Hibert, M., Jard, S., Barberis, C., 1995. The bin

M.M. Gehringer et al. / Environmental Toxicology and Pharmacology 19 (2005) 297–303 303
site of neuropeptide vasopressin V1a receptor. Evidence for a ma-jor localisation within transmembrane regions. J. Biol. Chem. 270,25771–25777.
Nobre, A.C.L., Coelho, G.R., Coutinho, M.C.M., Silva, M.M.M., Ange-lim, E.V., Menezes, D.B., Fonteles, M.C., Monteiro, H.S.A., 2001.The role of phospholipase A2 and cyclooxygenase in renal toxicityinduced by microcystin-LR. Toxicon 39, 721–724.
Rossini, G.P., 2000. In: Botana, L.M. (Ed.), Seafood and Freshwater Tox-ins. Marcel Dekker, New York, pp. 257–287.
Runnegar, M.T., Kong, S., Bernt, N., 1993. Protein phosphatase inhibi-tion and in vivo hepatotoxicity of microcystins. Am. J. Physiol. 265,G224–G230.
Runnegar, M.T., Bernt, N., Kong, S.-M., Lee, E.Y.C., Zhang, L.,1995a. In vivo and in vitro binding of microcystin to protein phos-phatases 1 and 2A. Biochem. Biophys. Res. Commun. 216, 162–169.
Runnegar, M., Berndt, N., Kaplowitz, N., 1995b. Microcystin uptake andinhibition of protein phosphatases: effects of chemoprotectants andself-inhibition in relation to known hepatic transporters. Toxicol. Appl.Pharmacol. 134, 264–272.
Sivonen, K., Namikoshi, M., Evans, W.R., Carmichael, W.W., Sun, F.,Rouhainene, L., Luukkainen, R., Rhinehart, K.L., 1992. Isolations
and characterization of a variety of microcystins from seven strainsof the cyanobacterial genusAnabaena. Appl. Environ. Microbiol. 58,2495–2500.
Sivonen, K., 1996. Cyanobacterial toxins and toxin production. Phycolo-gia 35 (Suppl. 6), 12–24.
Sivonen, K., Jones, G., 1999. Cyanobacterial toxins. In: Chorus, I., Bar-tram, J. (Eds.), Toxic Cyanobacteria in Water. A Guide to PublicHealth Consequences, Monitoring and Management. E & FN Spon,London, pp. 41–111.
Vesterkvist, P.S.M., Meriluoto, J.A.O., 2003. Interaction between micro-cystins of different hydrophobicities and lipid monolayers. Toxicon41, 349–355.
Ward, C.J., Codd, G.A., 1999. Comparitive toxicity of four microcystinsof different hydrophobicities to the protozoan,Tetrahymena pyri-formis. J. Appl. Microbiol. 86, 874–882.
Watanabe, M.F., Oishi, S., Harada, K.I., Matsuura, K., Kawai, H., Suzuki,M., 1988. Toxins contained in Microcystis species of cyanobacteria(blue-green algae). Toxicon. 26, 1017–1025.
Yoshizawa, S., Matsushima, R., Watanabe, M.F., Harada, K.-I., Ichihara,A., Carmichael, W.W., Fujiki, H., 1990. Inhibition of protein phos-phatases by microcystin and nodularin associated with hepatotoxicity.J. Cancer Res. Clin. Oncol. 116, 609–614.
Copyright © 2022 FDOKUMEN


![Variants of Penney's Game arXiv:2006.13002v1 [math.HO] 19 ...](https://static.fdokumen.com/doc/165x107/6319649f1e5d335f8d0b2299/variants-of-penneys-game-arxiv200613002v1-mathho-19-.jpg)







![Histopathological effects of [D-Leu 1]Microcystin-LR variants on liver, skeletal muscle and intestinal tract of Hypophthalmichthys molitrix (Valenciennes, 1844](https://static.fdokumen.com/doc/165x107/631cac57a1cc32504f0c98d9/histopathological-effects-of-d-leu-1microcystin-lr-variants-on-liver-skeletal.jpg)









