
Comparative toxicity evaluation of cyanobacterial cyclic peptide toxin microcystin variants (LR, RR,...
12
Comparative toxicity evaluation of cyanobacterial cyclic peptide toxin microcystin variants (LR, RR, YR) in mice Nidhi Gupta, S.C. Pant, R. Vijayaraghavan, P.V. Lakshmana Rao * Division of Pharmacology and Toxicology, Defence Research and Development Establishment, Jhansi Road, Gwalior 474002, India Received 8 January 2003; received in revised form 10 March 2003; accepted 10 March 2003 Abstract The cyclic peptide toxins microcystins and nodularins are the most common and abundant cyanotoxins present in diverse water systems. They have been the cause of human and animal health hazards and even death. Over 60 microcystin variants have been reported so far. We report here the results of our study on comparative toxicity evaluation of three most predominant microcystins, MC-LR, MC-RR and MC-YR in mice. The mice were administered one LD 50 dose of MC-LR, RR and YR (43, 235.4 and 110.6 mg/kg body weight, respectively), and biochemical and histological variables were determined at 30 min post-treatment and mean time to death (MTD). Significant increase in liver body weight index was induced by all three variants. There was marginal increase in serum levels of hepatic enzymes viz. AST, ALT and g-GT at 30 min post-treatment but 3 /4 fold increase was observed at MTD. In contrast, enhanced LDH leakage, DNA fragmentation and depletion of hepatic glutathione was observed at 30 min post treatment in all three variants. There was no change in levels of serum protein, albumin and albumin/ globulin ratio. Liver histology showed time dependent severe pathological lesions like congestion, haemorrhage, portal mononuclear cell infiltration and obliteration of chromatin material. Lung lesions were predominantly in bronchi and parenchyma. Though qualitatively lesions were identical in all three microcystin variants, degree of liver and lung lesions varied quantitatively with the toxin. The breathing pattern and respiratory frequency of the mice after i.p. administration of the toxin showed uniform pattern for 90 min followed by abrupt change in the respiratory pattern and instantaneous death. Based on biochemical and histological studies, MC-LR was found to be the most potent toxin followed by MC-YR and MC-RR. # 2003 Elsevier Science Ireland Ltd. All rights reserved. Keywords: Cyclic peptide; Cyanotoxins; Microcystin-LR; Microcystin-RR; Microcystin-YR; Microcystin variants; Hepatotoxin 1. Introduction The occurrence of toxic freshwater blooms of cyanobacteria (blue /green algae) has been re- ported in many countries. Toxic cyanobacteria found in eutrophic, freshwater, municipal and residential water supplies are increasing environ- * Corresponding author. Tel.: /91-751-234-1980; fax: /91- 751-234-1148. E-mail address: pv[email protected] (P.V.L. Rao). Toxicology 188 (2003) 285 /296 www.elsevier.com/locate/toxicol 0300-483X/03/$ - see front matter # 2003 Elsevier Science Ireland Ltd. All rights reserved. doi:10.1016/S0300-483X(03)00112-4
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Comparative toxicity evaluation of cyanobacterial cyclic peptide toxin microcystin variants (LR, RR,...

Comparative toxicity evaluation of cyanobacterial cyclicpeptide toxin microcystin variants (LR, RR, YR) in mice
Nidhi Gupta, S.C. Pant, R. Vijayaraghavan, P.V. Lakshmana Rao *
Division of Pharmacology and Toxicology, Defence Research and Development Establishment, Jhansi Road, Gwalior 474002, India
Received 8 January 2003; received in revised form 10 March 2003; accepted 10 March 2003
Abstract
The cyclic peptide toxins microcystins and nodularins are the most common and abundant cyanotoxins present in
diverse water systems. They have been the cause of human and animal health hazards and even death. Over 60
microcystin variants have been reported so far. We report here the results of our study on comparative toxicity
evaluation of three most predominant microcystins, MC-LR, MC-RR and MC-YR in mice. The mice were
administered one LD50 dose of MC-LR, RR and YR (43, 235.4 and 110.6 mg/kg body weight, respectively), and
biochemical and histological variables were determined at 30 min post-treatment and mean time to death (MTD).
Significant increase in liver body weight index was induced by all three variants. There was marginal increase in serum
levels of hepatic enzymes viz. AST, ALT and g-GT at 30 min post-treatment but 3�/4 fold increase was observed at
MTD. In contrast, enhanced LDH leakage, DNA fragmentation and depletion of hepatic glutathione was observed at
30 min post treatment in all three variants. There was no change in levels of serum protein, albumin and albumin/
globulin ratio. Liver histology showed time dependent severe pathological lesions like congestion, haemorrhage, portal
mononuclear cell infiltration and obliteration of chromatin material. Lung lesions were predominantly in bronchi and
parenchyma. Though qualitatively lesions were identical in all three microcystin variants, degree of liver and lung
lesions varied quantitatively with the toxin. The breathing pattern and respiratory frequency of the mice after i.p.
administration of the toxin showed uniform pattern for 90 min followed by abrupt change in the respiratory pattern
and instantaneous death. Based on biochemical and histological studies, MC-LR was found to be the most potent toxin
followed by MC-YR and MC-RR.
# 2003 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Cyclic peptide; Cyanotoxins; Microcystin-LR; Microcystin-RR; Microcystin-YR; Microcystin variants; Hepatotoxin
1. Introduction
The occurrence of toxic freshwater blooms of
cyanobacteria (blue�/green algae) has been re-
ported in many countries. Toxic cyanobacteria
found in eutrophic, freshwater, municipal and
residential water supplies are increasing environ-
* Corresponding author. Tel.: �/91-751-234-1980; fax: �/91-
751-234-1148.
E-mail address: [email protected] (P.V.L. Rao).
Toxicology 188 (2003) 285�/296
www.elsevier.com/locate/toxicol
0300-483X/03/$ - see front matter # 2003 Elsevier Science Ireland Ltd. All rights reserved.
doi:10.1016/S0300-483X(03)00112-4

mental hazard in several parts of the world (Coddet al., 1989; Dawson, 1998). The toxins of fresh-
water cyanobacteria are classified into two groups,
neurotoxins and hepatotoxins, which include cyc-
lic-peptide microcystins and nodularin. Microcys-
tins and related polypeptides are selectively
hepatotoxic to fish, birds and mammals. The
consequence of acute poisoning by these com-
pounds is rapid disorganization of the hepaticarchitecture, breakdown of sinusoidal structures
and, in mammals, pooling of blood in the liver
(Carmichael, 1992; Rao et al., 1995). Chronic
uptake of microcystins results in generalised he-
patocyte degeneration with necrosis, progressive
fibrosis and mononuclear leukocyte infiltration.
Human illnesses attributed to cyanobacterial tox-
ins can be categorised into gastroenteritis andrelated diseases, allergic and irritation reactions,
and liver diseases (Bell and Codd, 1994; Hitzfeld et
al., 2000; Chorus et al., 2000). Micrcoystins and
nodularin have been found to be potent inhibitors
of protein phosphatase type 1 and 2A as well as
skin and liver tumour promoters in laboratory
animals (Nishiwaki-Matsushima et al., 1992).
They are also suspected to be involved withpromotion of primary liver cancer in humans
exposed to long-term doses of these peptide toxins
through drinking water (Yu, 1995). The bioaccu-
mulation of cyanotoxins by aquatic animals in-
cluding fish, mollusc and zooplankton has been
reported (Amorin and Vasconcelos, 1999), which
indicates that oral consumption by animal tissues
containing cyanotoxins is possible and can lead tohuman toxicity. Tragic death of 60 haemodialysis
patients was reported in Brazil due to the presence
of cyanobacterial toxins in the water supply used
in haemodialysis unit (Azevedo et al., 2002).
Over 60 variants of microcystin have been
reported so far which conform to the following
generalised format: Cyclo (D-Ala1-X2-D-MeAsp3-
Y4-Adda-Arg5-D-Glu6-Mdha7-) where, X and Yare variable amino acids, D-MeAsp is D-erythro-b-
methylaspartic acid, Adda is (2S, 3S, 8S, 9S)-3-
amino-9-methoxy-2-6-8-trimethyl 10-phenyldeca-
4.6-dienoic acid and Mdha is N -methyldehydroa-
lanine. The unusual amino acid Adda is essential
for expression of biological activity. Combinations
of the two variable L-amino acids, X and Y,
account for many of the microcystin variants andare used in the nomenclature of the toxins. The
XY variable amino acids for MC-LR, MC-RR
and MC-YR are leucine (L), arginine (R) and
tyrosine (Y). There is very little information on the
toxicity profile of microcystin variants other than
MC-LR. The objective of the present study was
comparative evaluation of biochemical and histo-
logical effects of three commonly occurring micro-cystin variants, viz. microcystin-LR, RR and YR
in mice.
2. Materials and methods
2.1. Chemicals
The cyanobacterial toxin microcystin-LR, RR
and YR were obtained from Professor W.W.
Carmichael, Wright State University, OH, USA.
All other chemicals were obtained from Sigma
Chemical Co. (St Louis, USA) unless otherwise
specified.
2.2. Animals
Swiss albino female mice weighing between 24
and 26 g body weight from Establishment’s animal
facility were used for the study. The animals were
housed in polypropylene cages with dust-free rice
husk as bedding material, and were provided with
pellet food (Amrut Laboratory Feeds, Maharash-
tra, India) and water ad libitum.
2.3. Experimental protocol
Microcystin variants LR, RR and YR were
dissolved in minimum amount of methanol (0.1%)
and diluted to required concentration in PBS. The
mice were housed four to a cage and allowed to
acclimatise 7 days prior to dosing. Acute 24 h,
LD50 was determined for all the toxins by Dixon’sup and down method for small samples (Dixon,
1965). Animals were divided into four groups of
four animals each and administered as below:
Group Treatment
I PBS (0.2 ml)
N. Gupta et al. / Toxicology 188 (2003) 285�/296286

II One LD50 MC-LRIII One LD50 MC-RR
IV One LD50 MC-YR
All the experiments were repeated at least two
times. The study has the approval of Institute’s
ethical committee on animal experimentation.
2.4. Biochemical parameters
Blood was collected from retro-orbital plexus of
mice at 30 min and at mean time to death (MTD).
Serum harvested from each mice at specified time
points was used to determine serum activity of
lactate dehdydrogenase (LDH), g-glutamyl trans-
peptidase (g-GT) by commercial diagnostic kits of
E. Merck India Ltd. LDH and (g-GT) were
estimated by kinetic method using NADH and g-glutamyl-p -nitroanilide, respectively, as substrates
at 340 nm (LDH) and 405 (g-GT) using Milton
Roy 1201 Spectrophotometer. Serum albumin
concentrations and total protein were measured
and the albumin/globulin (A/G) ratio was deter-
mined. Soon after blood sampling at 30 min and
MTD animals were sacrificed by cervical disloca-
tion, and liver and lung were excised quickly. Liversamples were washed free of adhering extraneous
material, blotted and weighed to determine organ/
body weight index (OBI�/organ weight�/100/
body weight). Protein was estimated by Lowry’s
method (Lowry et al., 1951). GSH was estimated
fluorimetrically using o -pthaladehyde (OPT) as a
reagent (Hisin and Hilf, 1976) in a Shimadzu RF
5000 spectro-fluorophotometer.
2.5. DNA fragmentation assay
DNA fragmentation was assayed as previously
described (Rao et al., 1999). Briefly, the liver from
control and treated animals were quickly excised
and frozen. The frozen tissues were homogenised
in ice-cold lysis buffer (10 mM Tris, 20 mM
EDTA, 0.5% Triton X-100, pH 8.0) and thencentrifuged at 27 000�/g for 30 min. Both pellet
(intact chromatin) and supernatant (DNA frag-
ments) were assayed for DNA content fluorime-
trically by using fluorescent dye 4?,6-diamidino-2-
phenylindole (DAPI). Briefly, to 2 ml of the
reagent 100 ng/ml NaCl, 20 ml of a sample were
added and then fluorescence intensity was mea-sured at 450 nm with excitation at 362 nm. The
percentage of fragmented DNA was defined as the
ratio of the DNA content of the supernatant at
27 000�/g to the total DNA in the lysate (Wyllie,
1980).
2.6. Measurement of respiratory variables
For each toxin four animals were used. They
were restrained in body plethysmograph for the
recording of respiratory signals. A volumetric
pressure transducer (model PT5, Grass Instru-
ment, USA) was used for sensing respiratory
flow signals. A continuous airflow of 170 ml/min
was maintained into each body plethysmograph
using a critical orifice (27 gauge needle). Thesignals from the individual transducer were am-
plified using universal amplifiers (Gould, USA).
The amplified signals were digitised using an
analogue to digital converter (Metrabyte, Taun-
ton, USA) and stored and analysed in a personal
computer. The amplified signals from the universal
amplifier were also fed into an oscillograph for
recording of breathing pattern (WindoGraf,Gould, USA). The animals were acclimatised in
the body plethysmograph for 30 min. After the
acclimatization a control recording of respiratory
variables was carried out for 30 min. One LD50
dose of the toxins MC-LR, MC-RR or MC-YR
was administered intraperitoneally after 30 min.
The respiratory variables were recorded for a
period of 4 h after administration. All the respira-tory variables, viz. tidal volume (VT), inspiratory
time (TI), expiratory time (TE) and respiratory
frequency (f) were measured. A computer program
capable of recognising the effects of chemicals as
sensory irritation, airway obstruction or pulmon-
ary irritation, and a combination of these effects,
was used for the collection and analysis (Vijayar-
aghavan et al., 1994).
2.7. Histology
Mice were treated with one LD50 dose of the
toxin. The animals were sacrificed by cervical
dislocation at specified time points (30 min and
MTD). Tissue samples of liver and lung were
N. Gupta et al. / Toxicology 188 (2003) 285�/296 287

obtained from both control and treated animals.Complete necropsy was performed in tissues from
liver and lung. For pulmonary tissue, trachea was
tied before opening the thoracic cage, lung with
the trachea was excised and fixed in 10% buffered
formalin solution. After proper fixation, small
pieces of lung and liver were processed by dehy-
dration and embedded in paraffin. Multiple sec-
tions from each block were prepared at 5�/6 mmthickness and stained with haematoxylin and eosin
for examination under light microscope. The
severity of the lesions were characterised and
scored using Leica Q Win-500 1W image analyser.
2.8. Statistical analysis
Data were expressed as mean9/S.E. from four
animals per each treatment. Data were analysed by
one-way Analysis of Variance (ANOVA) followed
by multiple mean comparison by Student�/New-
man�/Keul’s test. The level of significance was set
at P 5/0.05.
3. Results
The acute 24 h LD50 (i.p.) of three microcystins
was determined by Dixon’s up and down methodfor small samples to minimise the number of
animals (Dixon, 1965). Table 1 summarises the
molecular formulae and LD50 of the three toxins.
MC-LR was most toxic followed by MC-YR and
MC-RR. The time to death varied considerably. In
MC-LR and MC-YR, MTD was similar com-
pared with higher survival time of MC-RR. For all
subsequent studies on biochemical and histologicalparameters animals were sacrificed at 30 min post
treatment and at MTD when animals were mor-
ibund (referred hereafter as MTD) in order to
examine immediate and delayed effects of the
toxin. Clinical signs of toxicity observed within 1
h of treatment included restlessness, laboured
ventilation, in coordination of movement, spas-modic leaping and splaying of hind limbs. Death
was preceded by severe convulsions. Upon ne-
cropsy the animals showed grossly distended liver
engorged with blood. The liver body weight index
(LBI) of the mice treated with three microcystin
variants at 30 min and before death is shown in
Table 2. At 30 min post-treatment there is no
significant increase in LBI compared with control.However, all three toxins showed significant
increase in LBI at MTD compared with control
group.
The serum biochemical variables measured in-
clude alanine amino transferase (ALT), aspartate
amino transferase (AST), g-glutamyl transpepti-
dase (g-GT), total protein, albumin, globulin,
albumin/globulin ratio (A/G). In addition, hepaticglutathione content (GSH) and hepatic DNA
Table 1
The chemical formulae of microcystin variants, their LD50 values and mean survival time in mice
Toxin Formula MW (mg/kg, i.p.) LD50 Fiducial limits MTD (min)a
MC-LR C49H74N10O12 994 43.0 (37.5�/49.4) 97.59/12.9
MC-RR C49H75N13O12 1037 235.4 (202.3�/272.8) 158.79/9.6
MC-YR C52H72N10O13 1044 110.6 (81.7�/149.6) 76.29/9.8
a Values are mean9/S.E. of four animals for each treatment.
Table 2
Liver body weight index of mice treated with LD50 dose of
microcystin-LR, RR and YR
Toxin Dose (mg/kg) Liver body weight index*
30 min MTD
Control �/ 5.569/0.09a 5.569/0.09a
MC-LR 43.0 6.979/0.47a 7.299/0.12b
MC-RR 235.4 5.799/0.47a 7.259/0.50b
MC-YR 110.6 5.649/0.71a 7.549/0.25b
Values are mean9/S.E. of four animals per treatment.
* Means followed by different alphabet are significantly
different at P 5/0.05 by Student�/Newman�/Keul’s test.
N. Gupta et al. / Toxicology 188 (2003) 285�/296288

fragmentation were estimated. All these biochem-
ical variables were determined at 30 min post-
treatment and MTD. Effect of microcystin var-
iants on plasma enzymes of hepatic origin AST,
ALT and g-GT are shown in Fig. 1A�/C. At 30
min post-treatment there was significant change in
levels of these enzymes from control group. But at
MTD a further significant increase in the levels of
AST, ALT and g-GT was observed. There was no
significant difference among the three variants at
30 min post-treatment for the three enzymes AST,
ALT and g-GT. At MTD, MC-LR induced
significant increase in AST compared with MC-
RR and MC-YR. Similar increased leakage was
observed with respect to ALT and g-GT at MTD
in MC-LR group than MC-RR (Fig. 1B, C). There
is no significant difference in LDH leakage at 30
min among the three variants but more than 2-fold
Fig. 1. Effect of microcystin variants LR, RR and YR on
serum levels of (A) AST, (B) ALT, and (C) g-GT after i.p.
administration of one LD50 dose of toxin. Values are mean9/
S.E. of four animals each. * Means followed by different
alphabet(s) are significantly different at P 5/0.05 by Student�/
Newman�/Keul’s test.
Fig. 2. Effect of microcystin variants LR, RR and YR on (A)
LDH leakage, (B) hepatic GSH content, and (C) hepatic DNA
fragmentation after i.p. administration of one LD50 dose of
toxin. Values are mean9/S.E. of four animals each. * Means
followed by different alphabet(s) are significantly different at
P B/0.05 by Student�/Newman�/Keul’s test.
N. Gupta et al. / Toxicology 188 (2003) 285�/296 289
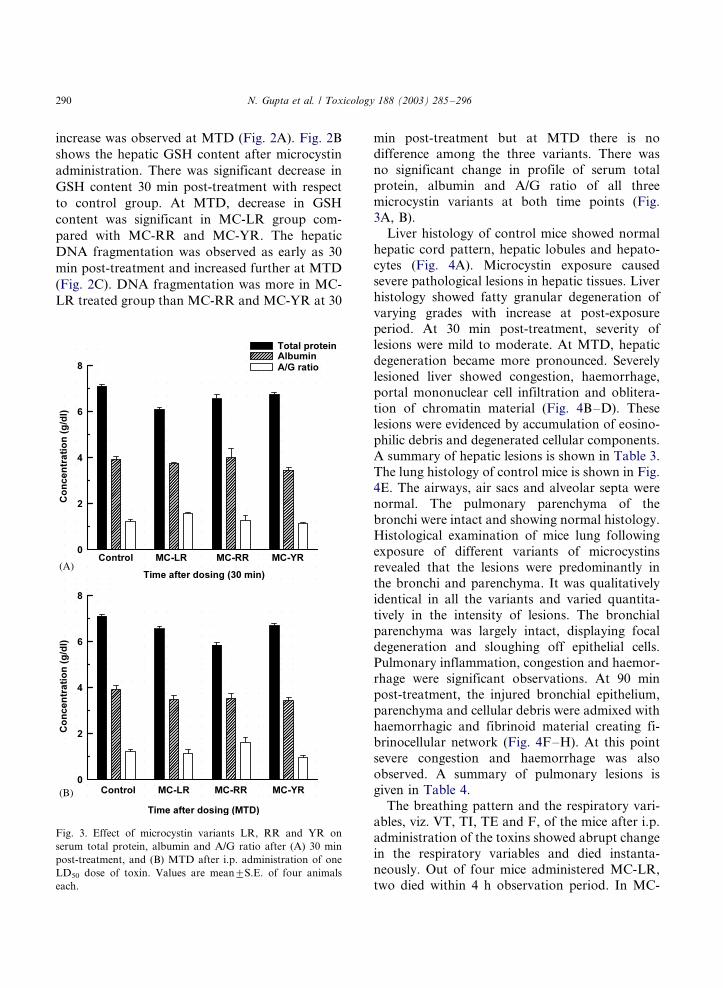
increase was observed at MTD (Fig. 2A). Fig. 2B
shows the hepatic GSH content after microcystin
administration. There was significant decrease in
GSH content 30 min post-treatment with respect
to control group. At MTD, decrease in GSH
content was significant in MC-LR group com-
pared with MC-RR and MC-YR. The hepatic
DNA fragmentation was observed as early as 30
min post-treatment and increased further at MTD
(Fig. 2C). DNA fragmentation was more in MC-
LR treated group than MC-RR and MC-YR at 30
min post-treatment but at MTD there is nodifference among the three variants. There was
no significant change in profile of serum total
protein, albumin and A/G ratio of all three
microcystin variants at both time points (Fig.
3A, B).
Liver histology of control mice showed normal
hepatic cord pattern, hepatic lobules and hepato-
cytes (Fig. 4A). Microcystin exposure causedsevere pathological lesions in hepatic tissues. Liver
histology showed fatty granular degeneration of
varying grades with increase at post-exposure
period. At 30 min post-treatment, severity of
lesions were mild to moderate. At MTD, hepatic
degeneration became more pronounced. Severely
lesioned liver showed congestion, haemorrhage,
portal mononuclear cell infiltration and oblitera-tion of chromatin material (Fig. 4B�/D). These
lesions were evidenced by accumulation of eosino-
philic debris and degenerated cellular components.
A summary of hepatic lesions is shown in Table 3.
The lung histology of control mice is shown in Fig.
4E. The airways, air sacs and alveolar septa were
normal. The pulmonary parenchyma of the
bronchi were intact and showing normal histology.Histological examination of mice lung following
exposure of different variants of microcystins
revealed that the lesions were predominantly in
the bronchi and parenchyma. It was qualitatively
identical in all the variants and varied quantita-
tively in the intensity of lesions. The bronchial
parenchyma was largely intact, displaying focal
degeneration and sloughing off epithelial cells.Pulmonary inflammation, congestion and haemor-
rhage were significant observations. At 90 min
post-treatment, the injured bronchial epithelium,
parenchyma and cellular debris were admixed with
haemorrhagic and fibrinoid material creating fi-
brinocellular network (Fig. 4F�/H). At this point
severe congestion and haemorrhage was also
observed. A summary of pulmonary lesions isgiven in Table 4.
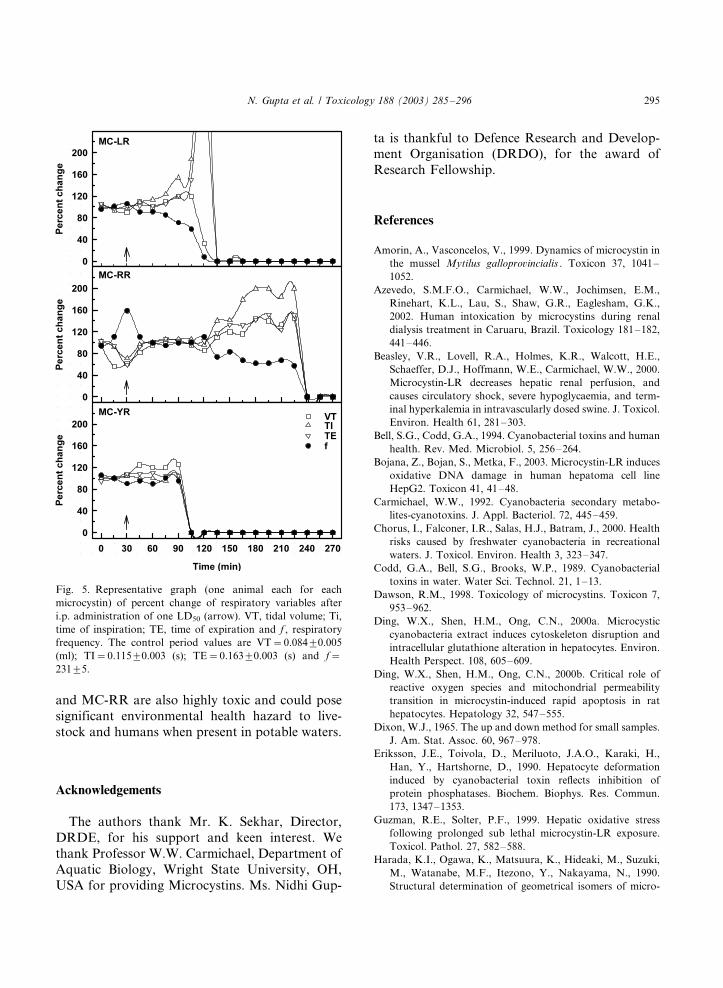
The breathing pattern and the respiratory vari-
ables, viz. VT, TI, TE and F, of the mice after i.p.
administration of the toxins showed abrupt change
in the respiratory variables and died instanta-
neously. Out of four mice administered MC-LR,
two died within 4 h observation period. In MC-
Fig. 3. Effect of microcystin variants LR, RR and YR on
serum total protein, albumin and A/G ratio after (A) 30 min
post-treatment, and (B) MTD after i.p. administration of one
LD50 dose of toxin. Values are mean9/S.E. of four animals
each.
N. Gupta et al. / Toxicology 188 (2003) 285�/296290

RR and MC-YR, three mice died. All the deathwas abrupt with sudden change in respiratory
variables. The mice that did not die during the 4-
h monitoring period did not show any change in
any of the variables, and they also survived.
Representative graph (one animal each for each
toxin) of the respiratory variables measured con-
tinuously after administration of the toxins is
given in Fig. 5.
4. Discussion
Many cyanobacterial species are capable of
producing potent toxins, which have been known
to cause intoxications and fatalities in wildlife,
livestock and humans. Microcystis is among the
most widely distributed genus and are known toproduce cyclic peptide toxins, the microcystins
(Carmichael, 1992; Hitzfeld et al., 2000). These
toxins are retained within the cyanobacterial cells
and are released only upon lysis of the cells during
the decomposition of cyanobacterial blooms. De-
tailed studies have not been carried out on toxic
manifestation of different microcystin variants. In
this study we investigated the comparative effectsof three most commonly and widely distributed
microcystin variants namely LR, RR and YR on
certain biochemical, respiratory and histological
variables in mice.
The acute LD50 determination of the three
variants showed variation in toxicity with MC-
LR being the most toxic followed by MC-YR and
MC-RR. There was significant difference betweentime and death of MC-YR compared with LR and
RR. The potent toxicity of the microcystins is
primarily due to marked inhibition of protein
phosphatase 1 and 2A (Eriksson et al., 1990).
Acute microcystin poisoning in mammals is char-
acterised by disruption of hepatic architecture
leading to massive intra hepatic haemorrhage
and death. In the present study all the three-microcystin variants caused significant increase in
LBI due to pooling of blood in the liver. Increase
in liver weight is a characteristic toxic effect of
microcystins and reported in a number of animal
models. Serum enzymes of hepatic origin have
shown significant increase in all the three toxins.
Serum levels of AST, ALT and g-GT showedsignificant increase compared with control as early
as 30 min post-exposure in all three toxins and was
further enhanced at MTD. Similarly increased
LDH levels were noticed at 30-min post treatment.
Increased serum activity of enzymes considered
indicative of hepatocellular damage has been
previously observed in both acute and chronic
toxicity studies (Rao et al., 1995; Nishiwaki-Matsushima et al., 1992). Increased serum ALT
activity is an important clinical indicator of
hepatocellular damage. It is a cytosolic enzyme
that is released into the blood following hepato-
cellular damage or necrosis. The results of our
study show that all three microcystin variants
induced time dependent increase in serum ALT
levels. The sensitivity of serum ALT activity as anindicator of hepatic damage may be decreased
secondary to decreased hepatic levels (Guzman
and Solter, 1999). It was shown that sub chronic
MC-LR exposure resulted in decreased hepatic
ALT protein and mRNA concentrations indicat-
ing one of the important toxic manifestation of
MC-LR could be decreased hepatic ALT synthesis
(Solter et al., 2000). The effects of down regulationof hepatic enzyme synthesis by microcystins on
hepatic function are not well known, however; it is
possible that the metabolic abilities of the liver
could be compromised.
The three toxins also caused depletion of hepatic
GSH levels with MC-LR causing significant de-
pletion compared with MC-RR and MC-YR. The
hepatic GSH content is critical factor for preser-ving normal cellular redox balance and protecting
the hepatocytes against oxidative stress. Cellular
reduced glutathione is important for the regulation
of cytoskeletal organisation. At present there are
several lines of evidence suggesting that GSH may
play a critical role in the detoxification of micro-
cystins. Microcystins could conjugate with GSH in
cell-free systems or in the liver tissue in vivo, whichsubstantially reduced its cytotoxicity (Ding et al.,
2000a). In the present study DNA fragmentation
was observed at 30 min post treatment in all the
three-microcystin variants. Significant further in-
crease was observed at MTD. The finding indi-
cates that DNA damage is an early event occurring
before necrotic liver damage. Microcystins in-
N. Gupta et al. / Toxicology 188 (2003) 285�/296 291
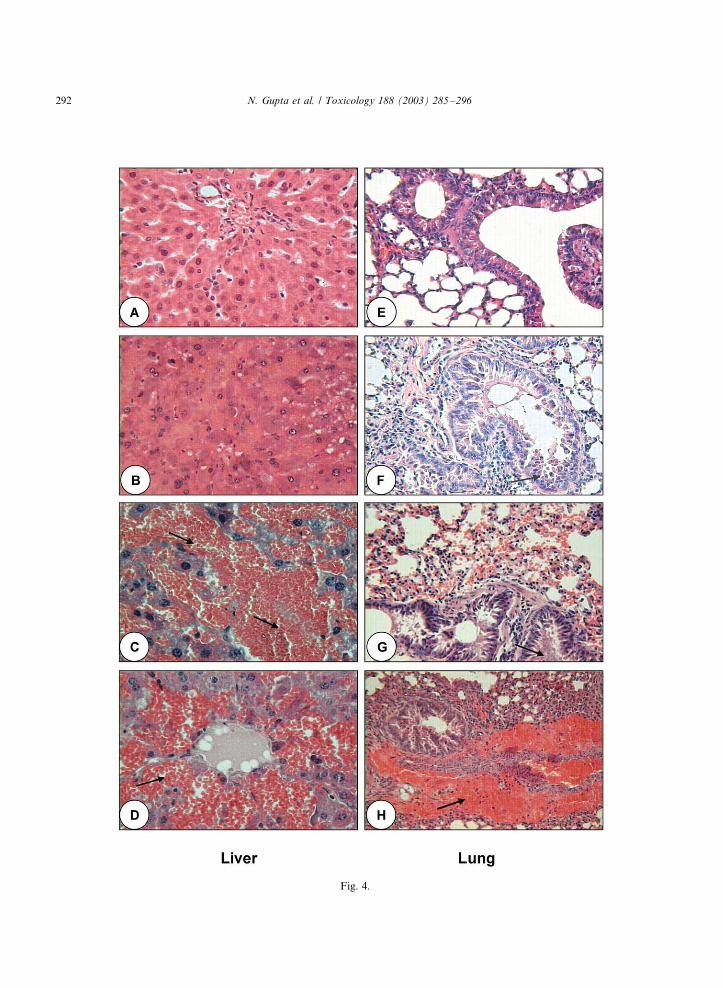
Fig. 4.
N. Gupta et al. / Toxicology 188 (2003) 285�/296292
duced DNA damage in mouse liver in vivo (Rao
and Bhattacharya, 1996) and in vitro in cultured
rat hepatocytes (Ding et al., 2000b), baby hamster
kidney cells and mouse embryo fibroblasts (Rao et
al., 1998), and Hep G2 cells (Bojana et al., 2003)
support the present results. The DNA damage
observed in the present study might be a conse-
quence of endonucleolytic DNA degradation as-
sociated with cytotoxicity, rather than by a direct
toxin�/DNA interaction.The morphological evidence of hepatocellular
injury by MC-LR, RR and YR was characterised
by apoptosis, centrilobular fibrosis, inflammation
and fatty vacuolar change. The induction of free
radical formation and mitochondrial alteration are
two major events found in microcystin treated
hepatocytes (Ding et al., 2000b). Immunohisto-
chemical localisation studies have shown that
positive immunostaining with anti-MC-MAb in
centrilobular hepatocytes in association with the
occurrence of haemorrhage or apoptosis. As
apoptotic hepatocytes would be finally phagocy-
tized and degraded by neighbouring hepatocytes
and macrophages, apoptotic cells/bodies contain-
ing MC-LR may have a role in elimination of the
toxin from the liver (Yoshida et al., 1998).
In the present study, in addition to histological
changes in the liver, all three toxins also showed
significant histological changes in lung like pul-
monary inflammation, congestion and haemor-
rhage. It is a normal practice to monitor
respiratory variables induced by a toxicant after
inhalation exposure. However, considering the
changes observed in lung histology we evaluated
the respiratory variables after i.p. administration
of toxin. Continuous monitoring of respiratory
variables after i.p. or p.o. administration can
reveal pulmonary effects (Sykes et al., 1977). All
three microcystin variants caused abrupt death
without much change in the respiratory variables
Table 3
Hepatic lesions caused by microcystin variants after i.p. administration of one LD50 dose in mice
Lesions MC-LR MC-RR MC-YR
30 min MTD 30 min MTD 30 min MTD
Basophilic stipling �/ �/�/ �/ �/�/ �/�/ �/�/�/
Degeneration and vacuolation of hepatic parenchyma �/�/ �/�/�/ �/ �/�/ �/�/ �/�/�/�/
Portal mononuclear cell infiltration �/ �/�/ �/ �/�/ �/�/�/ �/�/�/�/
Congestion and haemorrhage �/�/ �/�/�/ �/�/ �/�/�/ �/�/�/ �/�/�/�/
Hepatic vacuolation �/ �/�/ �/ �/�/ �/�/ �/�/�/
Granulatory cytoplasm �/ �/�/�/ �/ �/ �/�/ �/�/�/
Eosinophilic debris �/ �/�/ �/ �/ �/�/�/ �/�/�/�/
Hyper activation of Kupffer cells �/�/ �/�/�/ �/ �/�/ �/�/�/ �/�/�/�/
Accumulation of hemosiderin �/�/ �/�/�/ �/ �/�/ �/�/�/ �/�/�/�/
Scoring: �/, None, �/, Minimal (B/12%), �/�/, Mild (B/22%), �/�/�/, Moderate (B/45%), �/�/�/�/, Severe (�/45%). MTD, mean
time to death.
Fig. 4. Liver (A�/D) and lung (E�/H) histology of microcystin variants treated mice. Tissue samples were taken at time point before
death. H&E; �/100. (A) Control liver showing normal hepatic cord pattern of lobules, hepatocytes, central canal and bile duct; (B)
liver from MC-RR treated mice showing mild congestion, haemorrhage, degeneration and vacuolation of hepatic parenchyma; (C)
liver from MC-YR treated mice showing severe vacuolative degeneration, congestion, haemorrhage and accumulation of hemosiderin
(arrow); (D) liver from MC-LR treated mice showing degenerative changes with congestion, haemorrhage (arrow) and granulation of
cytoplasm. (E) Control mice lung showing homogenous parenchyma with normal airways, air sacs, alveolar septa and bronchiole; (F)
lung from MC-RR treated mice showing focal degeneration and sloughing off bronchiole parenchyma (arrow); (G) lung from MC-YR
treated mice showing severe bronchial parenchyma cell degeneration and sloughing off cells leading to occlusion of bronchiole (arrow);
(H) MC-LR treated mice showing inflammatory reaction and accumulation of eosinophilic edematous fluid.
N. Gupta et al. / Toxicology 188 (2003) 285�/296 293
during the observation period, showing that they
do not have any direct effect on the respiratory
system. The chemicals that cause direct effect on
the pulmonary epithelium may induce sensory or
pulmonary irritation, and those that alter the
sympathetic or the parasympathetic nervous sys-
tem may show broncho dilatation or broncho
constriction (Vijayaraghvan et al., 1994). Slatkin
et al. (1983) proposed that atypical pulmonary
thrombi might account for pulmonary congestion
and heart failure, resulting in centrilobular hepatic
necrosis and haemorrhage. Mild pulmonary
edema, peritoneal fluid accumulation, enteritis
with haemorrhage have been reported in response
to microcystin exposure in a range of species. The
lungs of intra vascularly dosed MC-LR in swine
remained effective in gas exchange, as reflected by
high pO2 and low pCO2 values (Beasley et al.,
2000).
Modifications at different sites of the micro-
cystin molecule were found to produce mild to
marked changes in toxicity. Harada et al. (1990)
described geometrical isomers of microcystin-LR
and RR, which were nonlethal when administered
i.p. to mice at 1.2 mg/kg. Replacement of second
amino acid leucine in MC-LR with tyrosine (MC-
YR) or arginine (MC-RR) still retained hepato-
toxicity. When L-leucine, which is a hydrophobic
amino acid is replaced by other hydrophobic
amino acids, i.e. Phe, Ala or Trp the resulting
microcystin retained LD50 from 171 to 249 mg/kg
(Stotis et al., 2000). The differences between
hydrophilic and hydrophobic microcystins could
result in changes in organotropism, toxicokinetics
and bioaccumulation (Pia and Meriluoto, 2003).
Apart from the protein phosphatase inhibitory
activity of microcystins some reports are available
on other toxic manifestation of microcystin var-
iants. The infusion of macrophages supernatants
stimulated by microcystin-LR caused significant
increase in renal vascular resistance, glomerular
filtration rates and urinary flow (Nobre et al.,
2003). Lankoff and Kolataj (2000) showed inhibi-
tory action of MC-YR on certain proteases that
affects the stability of lysosomal membranes and
provoke autodestruction of the cell and severe
damage of the liver. Mikhailov et al. (2003)
recently identified ATP-synthase as a novel in-
tracellular target for microcystin-LR and observed
that most of the physiological effects of micro-
cystin exposure are still only secondary to protein
phosphatase inhibition. In the present study
though there are minor variations in some of the
biochemical variables and histological effects, the
pronounced hepatotoxicity was observed in all the
three toxins. In conclusion, results of our study
indicate in addition to potent toxicity of micro-
cystin-LR, other two common variants MC-YR
Table 4
Pulmonary lesions caused by microcystin variants after i.p. administration of one LD50 dose in mice
Lesions MC-LR MC-RR MC-YR
30 min MTD 30 min MTD 30 min MTD
Bronchial epithelial hyperplasia �/ �/�/ �/ �/�/ �/�/�/ �/�/�/�/
Bronchial epithelial necrosis �/�/ �/�/�/ �/�/ �/�/�/ �/�/�/ �/�/�/�/
Occlusion bronchiole �/�/ �/�/�/ �/ �/�/ �/�/ �/�/�/
Haemorrhage �/ �/�/ �/ �/�/ �/�/ �/�/�/�/
Congestion �/�/ �/�/ �/ �/�/ �/�/ �/�/�/
Stinosis of bronchial lumen �/ �/�/ �/ �/�/ �/�/ �/�/
BALT necrosis �/ �/�/ �/ �/ �/�/ �/�/�/
Inflammatory reaction �/ �/�/ �/ �/�/ �/�/ �/�/�/
Edema �/ �/ �/ �/ �/ �/�/
Accumulation of fibrinoid debris �/ �/�/ �/ �/ �/�/ �/�/�/
Scoring: �/, None, �/, Minimal (B/12%), �/�/, Mild (B/22%), �/�/�/, Moderate (B/45%), �/�/�/�/, Severe (�/45%); MTD, mean
time to death.
N. Gupta et al. / Toxicology 188 (2003) 285�/296294
and MC-RR are also highly toxic and could pose
significant environmental health hazard to live-
stock and humans when present in potable waters.
Acknowledgements
The authors thank Mr. K. Sekhar, Director,
DRDE, for his support and keen interest. We
thank Professor W.W. Carmichael, Department of
Aquatic Biology, Wright State University, OH,
USA for providing Microcystins. Ms. Nidhi Gup-
ta is thankful to Defence Research and Develop-ment Organisation (DRDO), for the award of
Research Fellowship.
References
Amorin, A., Vasconcelos, V., 1999. Dynamics of microcystin in
the mussel Mytilus galloprovincialis . Toxicon 37, 1041�/
1052.
Azevedo, S.M.F.O., Carmichael, W.W., Jochimsen, E.M.,
Rinehart, K.L., Lau, S., Shaw, G.R., Eaglesham, G.K.,
2002. Human intoxication by microcystins during renal
dialysis treatment in Caruaru, Brazil. Toxicology 181�/182,
441�/446.
Beasley, V.R., Lovell, R.A., Holmes, K.R., Walcott, H.E.,
Schaeffer, D.J., Hoffmann, W.E., Carmichael, W.W., 2000.
Microcystin-LR decreases hepatic renal perfusion, and
causes circulatory shock, severe hypoglycaemia, and term-
inal hyperkalemia in intravascularly dosed swine. J. Toxicol.
Environ. Health 61, 281�/303.
Bell, S.G., Codd, G.A., 1994. Cyanobacterial toxins and human
health. Rev. Med. Microbiol. 5, 256�/264.
Bojana, Z., Bojan, S., Metka, F., 2003. Microcystin-LR induces
oxidative DNA damage in human hepatoma cell line
HepG2. Toxicon 41, 41�/48.
Carmichael, W.W., 1992. Cyanobacteria secondary metabo-
lites-cyanotoxins. J. Appl. Bacteriol. 72, 445�/459.
Chorus, I., Falconer, I.R., Salas, H.J., Batram, J., 2000. Health
risks caused by freshwater cyanobacteria in recreational
waters. J. Toxicol. Environ. Health 3, 323�/347.
Codd, G.A., Bell, S.G., Brooks, W.P., 1989. Cyanobacterial
toxins in water. Water Sci. Technol. 21, 1�/13.
Dawson, R.M., 1998. Toxicology of microcystins. Toxicon 7,
953�/962.
Ding, W.X., Shen, H.M., Ong, C.N., 2000a. Microcystic
cyanobacteria extract induces cytoskeleton disruption and
intracellular glutathione alteration in hepatocytes. Environ.
Health Perspect. 108, 605�/609.
Ding, W.X., Shen, H.M., Ong, C.N., 2000b. Critical role of
reactive oxygen species and mitochondrial permeability
transition in microcystin-induced rapid apoptosis in rat
hepatocytes. Hepatology 32, 547�/555.
Dixon, W.J., 1965. The up and down method for small samples.
J. Am. Stat. Assoc. 60, 967�/978.
Eriksson, J.E., Toivola, D., Meriluoto, J.A.O., Karaki, H.,
Han, Y., Hartshorne, D., 1990. Hepatocyte deformation
induced by cyanobacterial toxin reflects inhibition of
protein phosphatases. Biochem. Biophys. Res. Commun.
173, 1347�/1353.
Guzman, R.E., Solter, P.F., 1999. Hepatic oxidative stress
following prolonged sub lethal microcystin-LR exposure.
Toxicol. Pathol. 27, 582�/588.
Harada, K.I., Ogawa, K., Matsuura, K., Hideaki, M., Suzuki,
M., Watanabe, M.F., Itezono, Y., Nakayama, N., 1990.
Structural determination of geometrical isomers of micro-
Fig. 5. Representative graph (one animal each for each
microcystin) of percent change of respiratory variables after
i.p. administration of one LD50 (arrow). VT, tidal volume; Ti,
time of inspiration; TE, time of expiration and f , respiratory
frequency. The control period values are VT�/0.0849/0.005
(ml); TI�/0.1159/0.003 (s); TE�/0.1639/0.003 (s) and f�/
2319/5.
N. Gupta et al. / Toxicology 188 (2003) 285�/296 295

cystin LR and RR from cyanobacteria by two-dimensional
NMR spectroscopic techniques. Chem. Res. Toxicol. 3,
473�/481.
Hisin, P.J., Hilf, R., 1976. A fluorometric method for determi-
nation of oxidised and reduced glutathione in tissues. Anal.
Biochem. 74, 214�/226.
Hitzfeld, B.C., Hoger, S.J., Dietrich, R., 2000. Cyanobacterial
toxins: removal during drinking water treatment, and
human risk assessment. Environ. Health Perspect. 10,
113�/122.
Lankoff, A., Kolataj, A., 2000. Influence of microcystin-YR
and nodularin on the activity of some proteolytic enzymes
in mouse liver. Toxicon 39, 419�/423.
Lowry, O.H., Rosenberg, N.J., Farr, A.L., Randall, R.J., 1951.
Protein measurement with the Folin phenol reagent. J. Biol.
Chem. 193, 265�/275.
Mikhailov, A., Harmala-Brasken, A., Hellman, J., Meriluoto,
J., Eriksson, J.E., 2003. Identification of ATP-synthase as a
novel intracellular target for microcystin-LR. Chem.-Biol.
Interact. 142, 223�/237.
Nishiwaki-Matsushima, R., Ohta, T., Nishiwaki, S., Suga-
numa, M., Kohyama, K., Ishiwaka, T., Carmichael,
W.W., Fujiki, H., 1992. Liver tumour promotion by the
cyanobacterial cyclic peptide toxin microcystin-LR. J.
Cancer Res. Clin. Oncol. 118, 420�/424.
Nobre, A.C.L., Martins, A.M.C., Havt, A., Benevides, C.,
Lima, A.A.M., Fonteles, M.C., Monteiro, H.S.A., 2003.
Renal effects of supernatant from rat peritoneal macro-
phages activated by microcystin-LR: role protein mediators.
Toxicon 41, 377�/381.
Pia, S.M., Meriluoto, V., 2003. Interaction between microcys-
tins of different hydrophobicities and lipid monolayers.
Toxicon 41, 349�/355.
Rao, P.V.L., Bhattacharya, R., 1996. The cyanobacterial toxin
microcystin-LR induced DNA damage in mouse liver in
vivo. Toxicology 114, 29�/36.
Rao, P.V.L., Bhattacharya, R., Pant, S.C., Bhaskar, A.S.B.,
1995. Toxicity evaluation of in vitro cultures of freshwater
cyanobacterium Microcystis aeruginosa : I Hepatic and
histopathological effects in rats. Biomed. Environ. Sci. 8,
254�/264.
Rao, P.V.L., Bhattacharya, R., Parida, M.M., Jana, A.M.,
Bhaskar, A.S.B., 1998. Freshwater cyanobacterium Micro-
cystis aeruginosa (UTEX 2385) induced DNA damage in
vivo and in vitro. Environ. Toxicol. Pharmacol. 5, 1�/6.
Rao, P.V.L., Bhaskar, A.S.B., Vijayaraghavan, R., 1999.
Sulphur mustard induced DNA damage in mice after
dermal and inhalation exposure. Toxicology 139, 39�/51.
Slatkin, D.N., Stoner, R.D., Adams, W.H., Kycia, J.H.,
Siegelman, H.W., 1983. Atypical pulmonary thrombosis
caused by a toxic cyanobacterial peptide. Science 220,
1383�/1385.
Solter, P.F., Zonglin, L., Guzman, R., 2000. Decreased hepatic
ALT synthesis is an outcome of sub-chronic microcystin-LR
toxicity. Toxicol. Appl. Pharmacol. 164, 216�/220.
Stotis, R.S., Namikoshi, M., Haschek, W.M., Rinehart, K.L.,
Carmichael, W.W., Dahlem, A.M., Beasley, V.R., 2000.
Structural modifications imparting reduced toxicity in
microcystins from Microcystis spp. Toxicon 31, 783�/789.
Sykes, B.I., Purchase, I.F., Smith, L.L., 1977. Pulmonary
ultrastructure after oral and intravenous dosage of paraquat
to rats. J. Pathol. 121, 233�/241.
Vijayaraghavan, R., Schaper, M., Thompson, R., Stock, M.F.,
Boylstein, L.A., Luo, J.E., Alarie, Y., 1994. Computer
assisted recognition and quantification of the effects of
airborne chemicals acting at different areas of respiratory
tract in mice. Arch. Toxicol. 68, 490�/499.
Wyllie, A.H., 1980. Glucocorticoid induced thymocyte apop-
tosis is associated with endogenous endonuclease activation.
Nature 284, 555�/556.
Yoshida, T., Makita, Y., Tsutsumi, T., Nagata, S., Tashiro, F.,
Yoshida, F., Sekijima, M., Tamura, S.I., Harada, T., Maita,
K., Ueno, Y., 1998. Immunohistochemical localization of
microcystin-LR in the liver of mice: a study on the
pathogenesis of microcystin-LR induced hepatotoxicity.
Environ. Toxicol. Pathol. 26, 411�/418.
Yu, S.Z., 1995. Primary prevention of hepatocellular carci-
noma. J. Gastroenterol. Hepatol. 10, 674�/682.
N. Gupta et al. / Toxicology 188 (2003) 285�/296296










![@_ YR]WhRj ^R[`c 4RSZ_Ve cV[ZX e`URj - Daily Pioneer](https://static.fdokumen.com/doc/165x107/631f50b94573ad0c3e030007/-yrwhrj-rc-4rszve-cvzx-eurj-daily-pioneer.jpg)









