
Can LASSBio 596 and dexamethasone treat acute lung and liver inflammation induced by microcystin-LR
9
Can LASSBio 596 and dexamethasone treat acute lung and liver inflammation induced by microcystin-LR? Giovanna M.C. Carvalho a , Vinícius R. Oliveira a , Raquel M. Soares a , Sandra M.F.O. Azevedo a , Lidia M. Lima b , Eliezer J. Barreiro b , Samuel S. Valença c , Paulo H.N. Saldiva d , Débora S. Faffe a , Walter A. Zin a, * a Carlos Chagas Filho Institute of Biophysics, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil b LASSBio Ò , Faculty of Pharmacy, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil 1 c Institute of Biomedical Sciences, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil d Department of Pathology, University of São Paulo, São Paulo, Brazil article info Article history: Received 5 March 2010 Received in revised form 1 June 2010 Accepted 5 June 2010 Available online 12 June 2010 Keywords: Acute lung inflammation Anti-inflammatory Corticosteroid Cyanobacteria Lung mechanics Microcystin-LR abstract The treatment of microcystin-LR (MCYST-LR)-induced lung inflammation has never been reported. Hence, LASSBio 596, an anti-inflammatory drug candidate, designed as symbiotic agent that modulates TNF-a levels and inhibits phosphodiesterase types 4 and 5, or dexamethasone were tested in this condition. Swiss mice were intraperitoneally (i.p.) injected with 60 ml of saline (CTRL) or a sub-lethal dose of MCYST-LR (40 mg/kg). 6 h later they were treated (i.p.) with saline (TOX), LASSBio 596 (10 mg/kg, L596), or dexametha- sone (1 mg/kg, 0.1 mL, DEXA). 8 h after MCYST-LR injection, pulmonary mechanics were determined, and lungs and livers prepared for histopathology, biochemical analysis and quantification of MCYST-LR. TOX showed significantly higher lung impedance than CTRL and L596, which were similar. DEXA could only partially block the mechanical alterations. In both TOX and DEXA alveolar collapse and inflammatory cell influx were higher than in CTRL and L596, being LASSBio 596 more effective than dexamethasone. TOX showed oxidative stress that was not present in CTRL and L596, while DEXA was partially efficient. MCYST-LR was detected in the livers of all mice receiving MCYST-LR and no recovery was apparent. In conclusion, LASSBio 596 was more efficient than dexamethasone in reducing the pulmonary functional impairment induced by MCYST-LR. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction In the last years cyanobacterial blooms increased all over the world. Because this phenomenon also takes place in aquatic ecosystems used as drinking water supplies and these cyanobacteria produce cyanotoxins (e.g., micro- cystins), a public health concern results (Chorus and Bartram, 1999). Humans may be exposed to cyanotoxins through several routes: the oral one is by far the most important. However, dermal, inhalation and parenteral routes may also occur (Funari and Testai, 2008). In 1988 a possible association between a cyanobacterial bloom and 88 deaths among 2000 intoxicated people was described (Teixeira et al., 1993). In 1990 two recruits in England developed severe pneumonia after contact with water containing toxic Microcystis aeruginosa (Turner et al., 1990). Microcystins can reach the lung, and their distri- bution after oral, intraperitoneal and intratracheal admin- istrations has been described (Ito et al., 2000, 2001). Finally, in February 1996 a death was recorded among patients undergoing routine renal dialysis with water contaminated with microcystins; the remaining patients * Corresponding author at: Universidade Federal do Rio de Janeiro, Instituto de Biofísica Carlos Chagas Filho, C.C.S., Av. Carlos Chagas Filho 373, Ilha do Fundão, 21941-902 Rio de Janeiro, RJ, Brazil. Tel.: þ5521 2562-6557; fax: þ5521 2280-8193. E-mail address: [email protected] (W.A. Zin). 1 http://www.farmacia.ufrj.br/lassbio/ Contents lists available at ScienceDirect Toxicon journal homepage: www.elsevier.com/locate/toxicon 0041-0101/$ – see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.toxicon.2010.06.005 Toxicon 56 (2010) 604–612
-
Upload
ussacademy -
Category
Documents
-
view
0 -
download
0
Transcript of Can LASSBio 596 and dexamethasone treat acute lung and liver inflammation induced by microcystin-LR

ilable at ScienceDirect
Toxicon 56 (2010) 604–612
Contents lists ava
Toxicon
journal homepage: www.elsevier .com/locate/ toxicon
Can LASSBio 596 and dexamethasone treat acute lung and liverinflammation induced by microcystin-LR?
Giovanna M.C. Carvalho a, Vinícius R. Oliveira a, Raquel M. Soares a, Sandra M.F.O. Azevedo a,Lidia M. Lima b, Eliezer J. Barreiro b, Samuel S. Valença c, Paulo H.N. Saldiva d,Débora S. Faffe a, Walter A. Zin a,*
aCarlos Chagas Filho Institute of Biophysics, Federal University of Rio de Janeiro, Rio de Janeiro, Brazilb LASSBio�, Faculty of Pharmacy, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil1c Institute of Biomedical Sciences, Federal University of Rio de Janeiro, Rio de Janeiro, BrazildDepartment of Pathology, University of São Paulo, São Paulo, Brazil
a r t i c l e i n f o
Article history:Received 5 March 2010Received in revised form 1 June 2010Accepted 5 June 2010Available online 12 June 2010
Keywords:Acute lung inflammationAnti-inflammatoryCorticosteroidCyanobacteriaLung mechanicsMicrocystin-LR
* Corresponding author at: Universidade FederaInstituto de Biofísica Carlos Chagas Filho, C.C.S., Av373, Ilha do Fundão, 21941-902 Rio de Janeiro, R2562-6557; fax: þ5521 2280-8193.
E-mail address: [email protected] (W.A. Zin).1 http://www.farmacia.ufrj.br/lassbio/
0041-0101/$ – see front matter � 2010 Elsevier Ltddoi:10.1016/j.toxicon.2010.06.005
a b s t r a c t
The treatment of microcystin-LR (MCYST-LR)-induced lung inflammation has never beenreported. Hence, LASSBio 596, an anti-inflammatory drug candidate, designed as symbioticagent that modulates TNF-a levels and inhibits phosphodiesterase types 4 and 5, ordexamethasone were tested in this condition. Swiss mice were intraperitoneally (i.p.)injected with 60 ml of saline (CTRL) or a sub-lethal dose of MCYST-LR (40 mg/kg). 6 h laterthey were treated (i.p.) with saline (TOX), LASSBio 596 (10 mg/kg, L596), or dexametha-sone (1 mg/kg, 0.1 mL, DEXA). 8 h after MCYST-LR injection, pulmonary mechanics weredetermined, and lungs and livers prepared for histopathology, biochemical analysis andquantification of MCYST-LR. TOX showed significantly higher lung impedance than CTRLand L596, which were similar. DEXA could only partially block the mechanical alterations.In both TOX and DEXA alveolar collapse and inflammatory cell influx were higher than inCTRL and L596, being LASSBio 596 more effective than dexamethasone. TOX showedoxidative stress that was not present in CTRL and L596, while DEXA was partially efficient.MCYST-LR was detected in the livers of all mice receiving MCYST-LR and no recovery wasapparent. In conclusion, LASSBio 596 was more efficient than dexamethasone in reducingthe pulmonary functional impairment induced by MCYST-LR.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
In the last years cyanobacterial blooms increased allover the world. Because this phenomenon also takes placein aquatic ecosystems used as drinking water supplies andthese cyanobacteria produce cyanotoxins (e.g., micro-cystins), a public health concern results (Chorus andBartram, 1999). Humans may be exposed to cyanotoxins
l do Rio de Janeiro,. Carlos Chagas FilhoJ, Brazil. Tel.: þ5521
. All rights reserved.
through several routes: the oral one is by far the mostimportant. However, dermal, inhalation and parenteralroutes may also occur (Funari and Testai, 2008).
In 1988 a possible association between a cyanobacterialbloom and 88 deaths among 2000 intoxicated people wasdescribed (Teixeira et al., 1993). In 1990 two recruits inEngland developed severe pneumonia after contact withwater containing toxicMicrocystis aeruginosa (Turner et al.,1990). Microcystins can reach the lung, and their distri-bution after oral, intraperitoneal and intratracheal admin-istrations has been described (Ito et al., 2000, 2001).Finally, in February 1996 a death was recorded amongpatients undergoing routine renal dialysis with watercontaminated with microcystins; the remaining patients

G.M.C. Carvalho et al. / Toxicon 56 (2010) 604–612 605
developed acute liver failure, and 52 died by December1996. These were the first confirmed human deaths relatedto microcystin intoxication (Barreto et al., 1996; Azevedoet al., 2002).
Microcystins are the cyanotoxins most frequently foundall over theworld. In vertebrate animals, themain targets ofmicrocystins are the hepatic cells, where they irreversiblyinhibit protein phosphatases (PP) of the serine/threoninefamily, especially PP1 and PP2A. Acutely, cytoskeletondisarrangement of hepatocytes follows, leading to hepaticfailure. We previously reported a rapid and continued lunginflammatory process with interstitial edema, alveolarcollapse and recruitment of inflammatory cells in Swissmice injected with a sub-lethal intraperitoneal dose of anaqueous extract containing MCYST-LR, the most potenttoxin (Picanço et al., 2004). Recently we observed pulmo-nary mechanical impairment in Swiss mice that receivedpurified MCYST-LR (Soares et al., 2007). Additionally, theinflammatory process can damage the lung tissue bymeansof reactive oxygen species (ROS) produced by activateddefense cells, such as neutrophils, monocytes and macro-phages (Moreno et al., 2005). There are no reports in theliterature concerning the treatment of the pulmonarydamage induced by microcystin-LR.
Thus, we aimed to evaluate whether LASSBio 596 ordexamethasone could protect the lung and liver against thetoxic actions of microcystin-LR. For such purpose lungmechanics and biochemical analyses, as well as lung andliver histology were evaluated in Swiss mice.
2. Materials and methods
2.1. Animals
Swiss male mice (6–7 weeks of age) were purchasedfrom FIOCRUZ (Fundação Oswaldo Cruz, Rio de Janeiro,Brazil). The animals were housed in plastic cages withabsorbent bedding material and maintained on a 12 hdaylight cycle. Food and water were provided ad libitum.
All animals received humane care in compliance withthe “Principles of Laboratory Animal Care” formulated bythe National Society for Medical Research and the “Guidefor the Care and Use of Laboratory Animals” prepared bythe National Academy of Sciences, USA. The experimentswere approved by the Ethics Committee of the HealthSciences Center, Federal University of Rio de Janeiro(Protocol IBCCF 012).
2.2. Preparation
Forty mice (35–40 g) were randomly divided into4 groups. Theywere initially intraperitoneally (i.p.) injectedwith 60 ml of either sterile saline solution (0.9% NaCl, CTRL,n ¼ 9) or a sub-lethal dose of MCYST-LR (40 mg/kg i.p.,purified material kindly provided by Professor WayneCarmichael, Wright State University, Dayton, OH, USA).After 6 h CTRL animals received 200 ml (i.p.) of sterile salinesolution, while those injected with MCYST-LR were injec-ted with 200 ml (i.p.) of either sterile saline solution (TOX,n ¼ 9), LASSBio 596 (10 mg/kg, L596, n ¼ 8), or dexa-methasone (1 mg/kg, DEXA, n ¼ 10). Initially each group
comprised 10 animals; four animals died along theexperiment.
2.3. Pulmonary mechanics
Eight hours after MCYST-LR injection, i.e., 2 h aftertreatment, the animals were sedated with diazepam (1 mgi.p.), anesthetized with pentobarbital sodium (20 mg kgbody weight�1 i.p.), tracheotomized, and a snugly fittingcannula (0.8 mm id) was introduced into the trachea. Then,the animals were paralyzed with pancuronium bromide(0.1 mg/kg i.v.), and a constant-flow ventilator (SamayVR15, Universidad de la Republica, Montevideo, Uruguay)provided artificial ventilation with a frequency of100 breaths/min, a tidal volume of 0.2 ml, flow of 1 ml/s,and positive end-expiratory pressure amounting to2 cm H2O. The anterior chest wall was surgically removed.
A pneumotachograph (1.5 mm ID, length ¼ 4.2 cm,distance between side ports ¼ 2.1 cm) (Mortola andNoworaj, 1983) was connected to the tracheal cannula forthe measurements of airflow (V0). Lung volume (VT) wasdetermined by digital integration of the flow signal.Tracheal pressure was measured with a Validyne MP-45differential pressure transducer (Engineering Corp, North-ridge, CA, USA). The flow resistance of the equipment (Req),tracheal cannula included, was constant up to flow rates of26mL s�1 and amounted to 0.12 cmH2OmL�1 s. Equipmentresistive pressure (¼Req.V0) was subtracted from pulmo-nary resistive pressure so that the present results representintrinsic values. All signals were conditioned and amplifiedin a Beckman type R Dynograph (Schiller Park, IL, USA).Flow and pressure signals were then passed through 8-poleBessel filters (902LPF, Frequency Devices, Haverhill, MA,USA) with the corner frequency set at 100 Hz, sampled at200 Hz with a 12-bit analog-to-digital converter (DT2801A,Data Translation, Marlboro, MA, USA), and stored ona microcomputer. All data were collected using LABDATsoftware (RHT-InfoData Inc., Montreal, QC, Canada).
Lung resistive (DP1) and viscoelastic/inhomogeneous(DP2) pressures, total pressure drop (DPtot ¼ DP1 þ DP2),static elastance (Est), and elastic componentof viscoelasticity(DE) were computed by the end-inflation occlusion method(Bates et al., 1985, 1988). Briefly, DP1 selectively reflectsairway resistance in normal animals and humans and DP2reflects stress relaxation, or viscoelastic properties of thelung, together with a tiny contribution of time constantinequalities (Bates et al.,1988; Saldiva et al.,1992). Lung static(Est) elastance was calculated by dividing Pel (elastic recoilpressureof the lung)byVT.DEwascalculatedas thedifferencebetween static and dynamic elastances (Bates et al., 1985,1988). Pulmonary mechanics were measured 10–15 timesin each animal.All datawere analyzed using ANADAT data analysis software(RHT-InfoData Inc., Montreal, QC, Canada). The duration ofthe experiments approximated 30 min.
2.4. Histological analysis
A lower longitudinal laparotomy was done immediatelyafter the determination of pulmonary mechanics, andheparin (1000 IU) was intravenously injected. The trachea
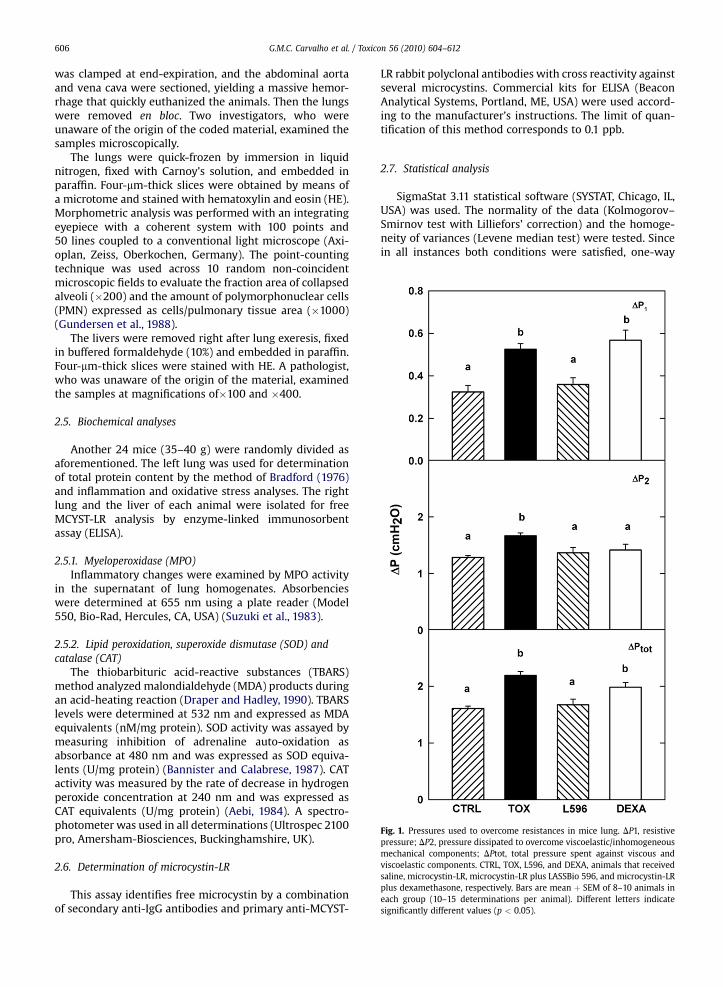
Fig. 1. Pressures used to overcome resistances in mice lung. DP1, resistivepressure; DP2, pressure dissipated to overcome viscoelastic/inhomogeneousmechanical components; DPtot, total pressure spent against viscous andviscoelastic components. CTRL, TOX, L596, and DEXA, animals that receivedsaline, microcystin-LR, microcystin-LR plus LASSBio 596, and microcystin-LRplus dexamethasone, respectively. Bars are mean þ SEM of 8–10 animals ineach group (10–15 determinations per animal). Different letters indicatesignificantly different values (p < 0.05).
G.M.C. Carvalho et al. / Toxicon 56 (2010) 604–612606
was clamped at end-expiration, and the abdominal aortaand vena cava were sectioned, yielding a massive hemor-rhage that quickly euthanized the animals. Then the lungswere removed en bloc. Two investigators, who wereunaware of the origin of the coded material, examined thesamples microscopically.
The lungs were quick-frozen by immersion in liquidnitrogen, fixed with Carnoy’s solution, and embedded inparaffin. Four-mm-thick slices were obtained by means ofa microtome and stained with hematoxylin and eosin (HE).Morphometric analysis was performed with an integratingeyepiece with a coherent system with 100 points and50 lines coupled to a conventional light microscope (Axi-oplan, Zeiss, Oberkochen, Germany). The point-countingtechnique was used across 10 random non-coincidentmicroscopic fields to evaluate the fraction area of collapsedalveoli (�200) and the amount of polymorphonuclear cells(PMN) expressed as cells/pulmonary tissue area (�1000)(Gundersen et al., 1988).
The livers were removed right after lung exeresis, fixedin buffered formaldehyde (10%) and embedded in paraffin.Four-mm-thick slices were stained with HE. A pathologist,who was unaware of the origin of the material, examinedthe samples at magnifications of�100 and �400.
2.5. Biochemical analyses
Another 24 mice (35–40 g) were randomly divided asaforementioned. The left lung was used for determinationof total protein content by the method of Bradford (1976)and inflammation and oxidative stress analyses. The rightlung and the liver of each animal were isolated for freeMCYST-LR analysis by enzyme-linked immunosorbentassay (ELISA).
2.5.1. Myeloperoxidase (MPO)Inflammatory changes were examined by MPO activity
in the supernatant of lung homogenates. Absorbencieswere determined at 655 nm using a plate reader (Model550, Bio-Rad, Hercules, CA, USA) (Suzuki et al., 1983).
2.5.2. Lipid peroxidation, superoxide dismutase (SOD) andcatalase (CAT)
The thiobarbituric acid-reactive substances (TBARS)method analyzed malondialdehyde (MDA) products duringan acid-heating reaction (Draper and Hadley, 1990). TBARSlevels were determined at 532 nm and expressed as MDAequivalents (nM/mg protein). SOD activity was assayed bymeasuring inhibition of adrenaline auto-oxidation asabsorbance at 480 nm and was expressed as SOD equiva-lents (U/mg protein) (Bannister and Calabrese, 1987). CATactivity was measured by the rate of decrease in hydrogenperoxide concentration at 240 nm and was expressed asCAT equivalents (U/mg protein) (Aebi, 1984). A spectro-photometer was used in all determinations (Ultrospec 2100pro, Amersham-Biosciences, Buckinghamshire, UK).
2.6. Determination of microcystin-LR
This assay identifies free microcystin by a combinationof secondary anti-IgG antibodies and primary anti-MCYST-
LR rabbit polyclonal antibodies with cross reactivity againstseveral microcystins. Commercial kits for ELISA (BeaconAnalytical Systems, Portland, ME, USA) were used accord-ing to the manufacturer’s instructions. The limit of quan-tification of this method corresponds to 0.1 ppb.
2.7. Statistical analysis
SigmaStat 3.11 statistical software (SYSTAT, Chicago, IL,USA) was used. The normality of the data (Kolmogorov–Smirnov test with Lilliefors’ correction) and the homoge-neity of variances (Levene median test) were tested. Sincein all instances both conditions were satisfied, one-way
G.M.C. Carvalho et al. / Toxicon 56 (2010) 604–612 607
ANOVA followed by Student–Newman–Keuls test wasused to assess differences among groups, when required.The morphometric parameters underwent an arcsinetransformation and, then, were compared. In all instancesthe significance level was set at 5% (p < 0.05).
3. Results
3.1. Pulmonary mechanics
There was no statistically significant difference in flowand inspired volume among the groups. All mechanicalparameters were higher in TOX than in CTRL. L596 andCTRL produced similar data, but in DEXA increases in DP1and DPtot like those in TOX were detected (Figs. 1 and 2).
3.2. Lung histology
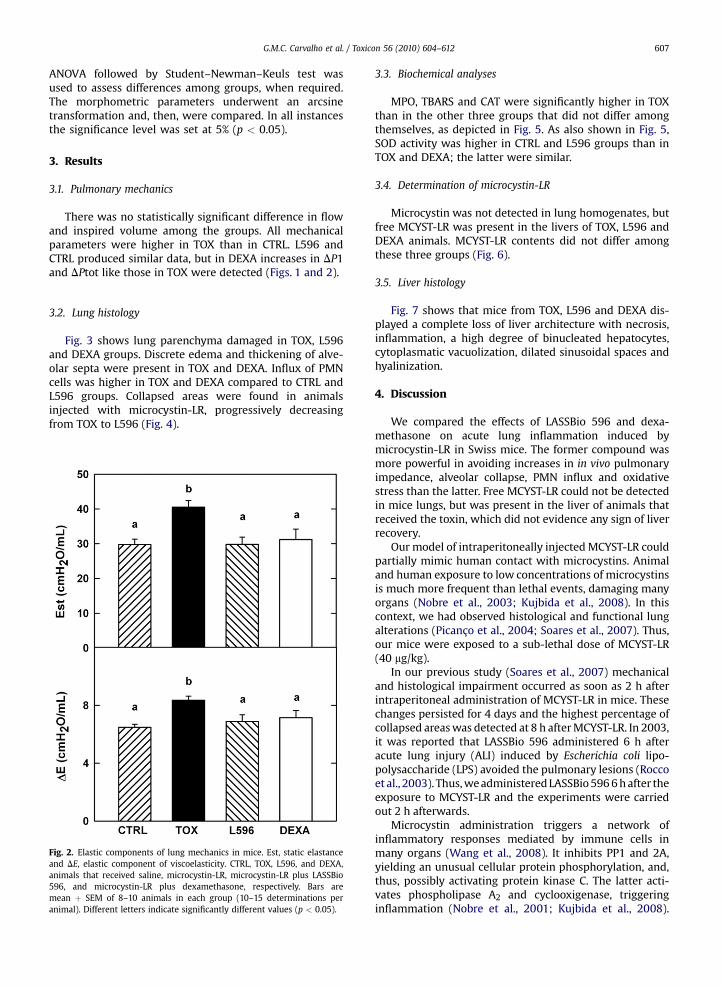
Fig. 3 shows lung parenchyma damaged in TOX, L596and DEXA groups. Discrete edema and thickening of alve-olar septa were present in TOX and DEXA. Influx of PMNcells was higher in TOX and DEXA compared to CTRL andL596 groups. Collapsed areas were found in animalsinjected with microcystin-LR, progressively decreasingfrom TOX to L596 (Fig. 4).
Fig. 2. Elastic components of lung mechanics in mice. Est, static elastanceand DE, elastic component of viscoelasticity. CTRL, TOX, L596, and DEXA,animals that received saline, microcystin-LR, microcystin-LR plus LASSBio596, and microcystin-LR plus dexamethasone, respectively. Bars aremean þ SEM of 8–10 animals in each group (10–15 determinations peranimal). Different letters indicate significantly different values (p < 0.05).
3.3. Biochemical analyses
MPO, TBARS and CAT were significantly higher in TOXthan in the other three groups that did not differ amongthemselves, as depicted in Fig. 5. As also shown in Fig. 5,SOD activity was higher in CTRL and L596 groups than inTOX and DEXA; the latter were similar.
3.4. Determination of microcystin-LR
Microcystin was not detected in lung homogenates, butfree MCYST-LR was present in the livers of TOX, L596 andDEXA animals. MCYST-LR contents did not differ amongthese three groups (Fig. 6).
3.5. Liver histology
Fig. 7 shows that mice from TOX, L596 and DEXA dis-played a complete loss of liver architecture with necrosis,inflammation, a high degree of binucleated hepatocytes,cytoplasmatic vacuolization, dilated sinusoidal spaces andhyalinization.
4. Discussion
We compared the effects of LASSBio 596 and dexa-methasone on acute lung inflammation induced bymicrocystin-LR in Swiss mice. The former compound wasmore powerful in avoiding increases in in vivo pulmonaryimpedance, alveolar collapse, PMN influx and oxidativestress than the latter. Free MCYST-LR could not be detectedin mice lungs, but was present in the liver of animals thatreceived the toxin, which did not evidence any sign of liverrecovery.
Our model of intraperitoneally injectedMCYST-LR couldpartially mimic human contact with microcystins. Animaland human exposure to low concentrations of microcystinsis much more frequent than lethal events, damaging manyorgans (Nobre et al., 2003; Kujbida et al., 2008). In thiscontext, we had observed histological and functional lungalterations (Picanço et al., 2004; Soares et al., 2007). Thus,our mice were exposed to a sub-lethal dose of MCYST-LR(40 mg/kg).
In our previous study (Soares et al., 2007) mechanicaland histological impairment occurred as soon as 2 h afterintraperitoneal administration of MCYST-LR in mice. Thesechanges persisted for 4 days and the highest percentage ofcollapsed areaswas detected at 8 h afterMCYST-LR. In 2003,it was reported that LASSBio 596 administered 6 h afteracute lung injury (ALI) induced by Escherichia coli lipo-polysaccharide (LPS) avoided the pulmonary lesions (Roccoet al., 2003). Thus,weadministeredLASSBio5966hafter theexposure to MCYST-LR and the experiments were carriedout 2 h afterwards.
Microcystin administration triggers a network ofinflammatory responses mediated by immune cells inmany organs (Wang et al., 2008). It inhibits PP1 and 2A,yielding an unusual cellular protein phosphorylation, and,thus, possibly activating protein kinase C. The latter acti-vates phospholipase A2 and cyclooxigenase, triggeringinflammation (Nobre et al., 2001; Kujbida et al., 2008).
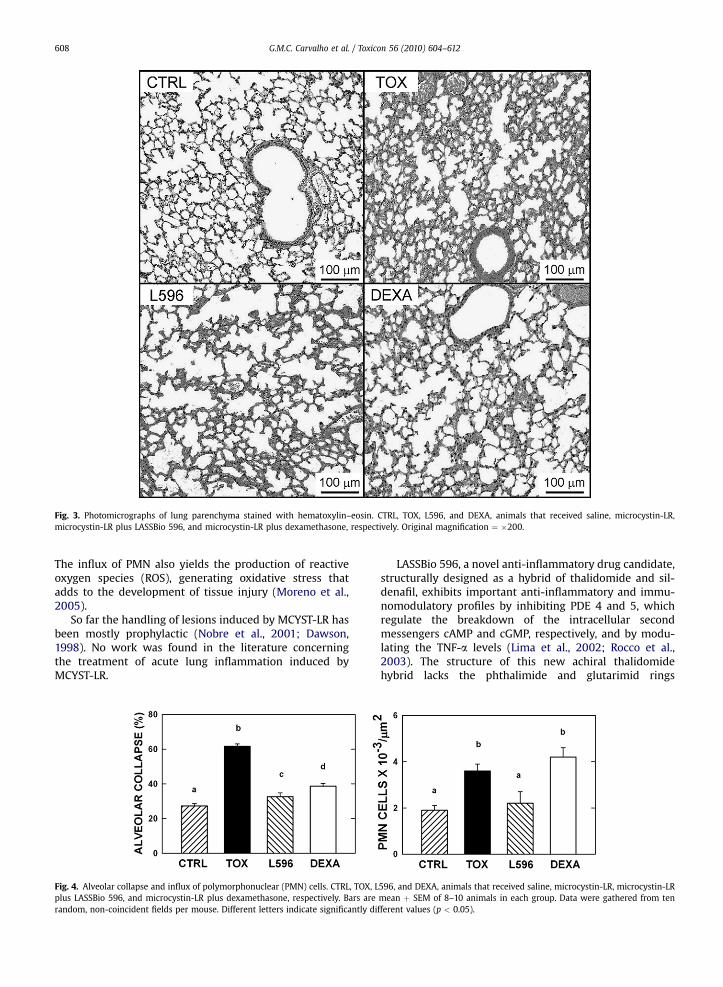
Fig. 3. Photomicrographs of lung parenchyma stained with hematoxylin–eosin. CTRL, TOX, L596, and DEXA, animals that received saline, microcystin-LR,microcystin-LR plus LASSBio 596, and microcystin-LR plus dexamethasone, respectively. Original magnification ¼ �200.
G.M.C. Carvalho et al. / Toxicon 56 (2010) 604–612608
The influx of PMN also yields the production of reactiveoxygen species (ROS), generating oxidative stress thatadds to the development of tissue injury (Moreno et al.,2005).
So far the handling of lesions induced by MCYST-LR hasbeen mostly prophylactic (Nobre et al., 2001; Dawson,1998). No work was found in the literature concerningthe treatment of acute lung inflammation induced byMCYST-LR.
Fig. 4. Alveolar collapse and influx of polymorphonuclear (PMN) cells. CTRL, TOX, Lplus LASSBio 596, and microcystin-LR plus dexamethasone, respectively. Bars arerandom, non-coincident fields per mouse. Different letters indicate significantly di
LASSBio 596, a novel anti-inflammatory drug candidate,structurally designed as a hybrid of thalidomide and sil-denafil, exhibits important anti-inflammatory and immu-nomodulatory profiles by inhibiting PDE 4 and 5, whichregulate the breakdown of the intracellular secondmessengers cAMP and cGMP, respectively, and by modu-lating the TNF-a levels (Lima et al., 2002; Rocco et al.,2003). The structure of this new achiral thalidomidehybrid lacks the phthalimide and glutarimid rings
596, and DEXA, animals that received saline, microcystin-LR, microcystin-LRmean þ SEM of 8–10 animals in each group. Data were gathered from tenfferent values (p < 0.05).
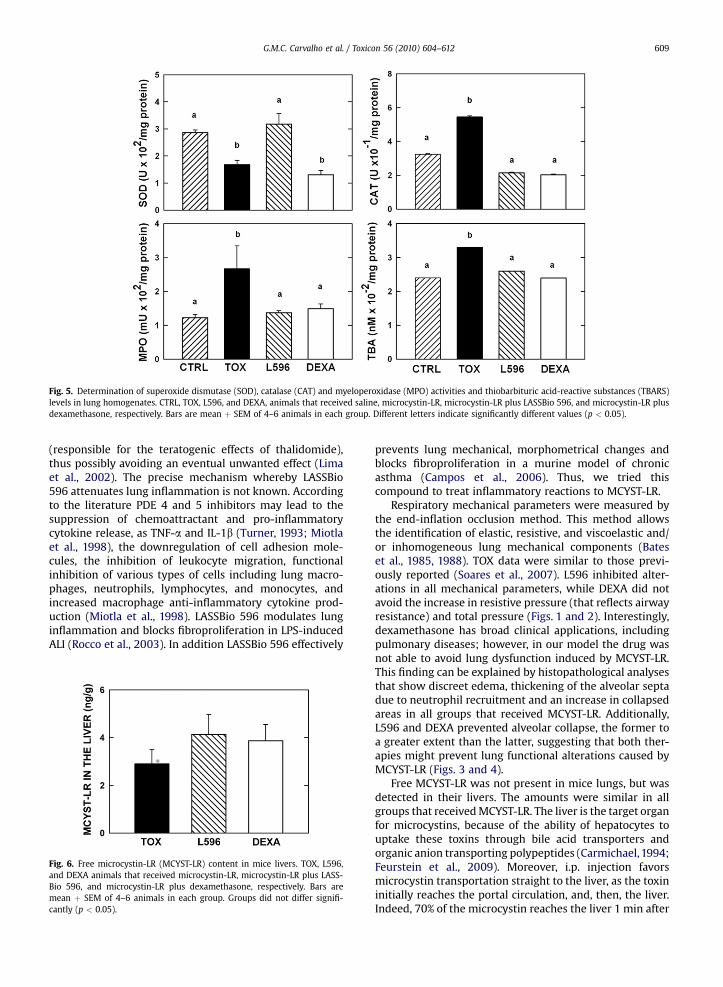
Fig. 5. Determination of superoxide dismutase (SOD), catalase (CAT) and myeloperoxidase (MPO) activities and thiobarbituric acid-reactive substances (TBARS)levels in lung homogenates. CTRL, TOX, L596, and DEXA, animals that received saline, microcystin-LR, microcystin-LR plus LASSBio 596, and microcystin-LR plusdexamethasone, respectively. Bars are mean þ SEM of 4–6 animals in each group. Different letters indicate significantly different values (p < 0.05).
G.M.C. Carvalho et al. / Toxicon 56 (2010) 604–612 609
(responsible for the teratogenic effects of thalidomide),thus possibly avoiding an eventual unwanted effect (Limaet al., 2002). The precise mechanism whereby LASSBio596 attenuates lung inflammation is not known. Accordingto the literature PDE 4 and 5 inhibitors may lead to thesuppression of chemoattractant and pro-inflammatorycytokine release, as TNF-a and IL-1b (Turner, 1993; Miotlaet al., 1998), the downregulation of cell adhesion mole-cules, the inhibition of leukocyte migration, functionalinhibition of various types of cells including lung macro-phages, neutrophils, lymphocytes, and monocytes, andincreased macrophage anti-inflammatory cytokine prod-uction (Miotla et al., 1998). LASSBio 596 modulates lunginflammation and blocks fibroproliferation in LPS-inducedALI (Rocco et al., 2003). In addition LASSBio 596 effectively
Fig. 6. Free microcystin-LR (MCYST-LR) content in mice livers. TOX, L596,and DEXA animals that received microcystin-LR, microcystin-LR plus LASS-Bio 596, and microcystin-LR plus dexamethasone, respectively. Bars aremean þ SEM of 4–6 animals in each group. Groups did not differ signifi-cantly (p < 0.05).
prevents lung mechanical, morphometrical changes andblocks fibroproliferation in a murine model of chronicasthma (Campos et al., 2006). Thus, we tried thiscompound to treat inflammatory reactions to MCYST-LR.
Respiratory mechanical parameters were measured bythe end-inflation occlusion method. This method allowsthe identification of elastic, resistive, and viscoelastic and/or inhomogeneous lung mechanical components (Bateset al., 1985, 1988). TOX data were similar to those previ-ously reported (Soares et al., 2007). L596 inhibited alter-ations in all mechanical parameters, while DEXA did notavoid the increase in resistive pressure (that reflects airwayresistance) and total pressure (Figs. 1 and 2). Interestingly,dexamethasone has broad clinical applications, includingpulmonary diseases; however, in our model the drug wasnot able to avoid lung dysfunction induced by MCYST-LR.This finding can be explained by histopathological analysesthat show discreet edema, thickening of the alveolar septadue to neutrophil recruitment and an increase in collapsedareas in all groups that received MCYST-LR. Additionally,L596 and DEXA prevented alveolar collapse, the former toa greater extent than the latter, suggesting that both ther-apies might prevent lung functional alterations caused byMCYST-LR (Figs. 3 and 4).
Free MCYST-LR was not present in mice lungs, but wasdetected in their livers. The amounts were similar in allgroups that receivedMCYST-LR. The liver is the target organfor microcystins, because of the ability of hepatocytes touptake these toxins through bile acid transporters andorganic anion transporting polypeptides (Carmichael,1994;Feurstein et al., 2009). Moreover, i.p. injection favorsmicrocystin transportation straight to the liver, as the toxininitially reaches the portal circulation, and, then, the liver.Indeed, 70% of the microcystin reaches the liver 1 min after
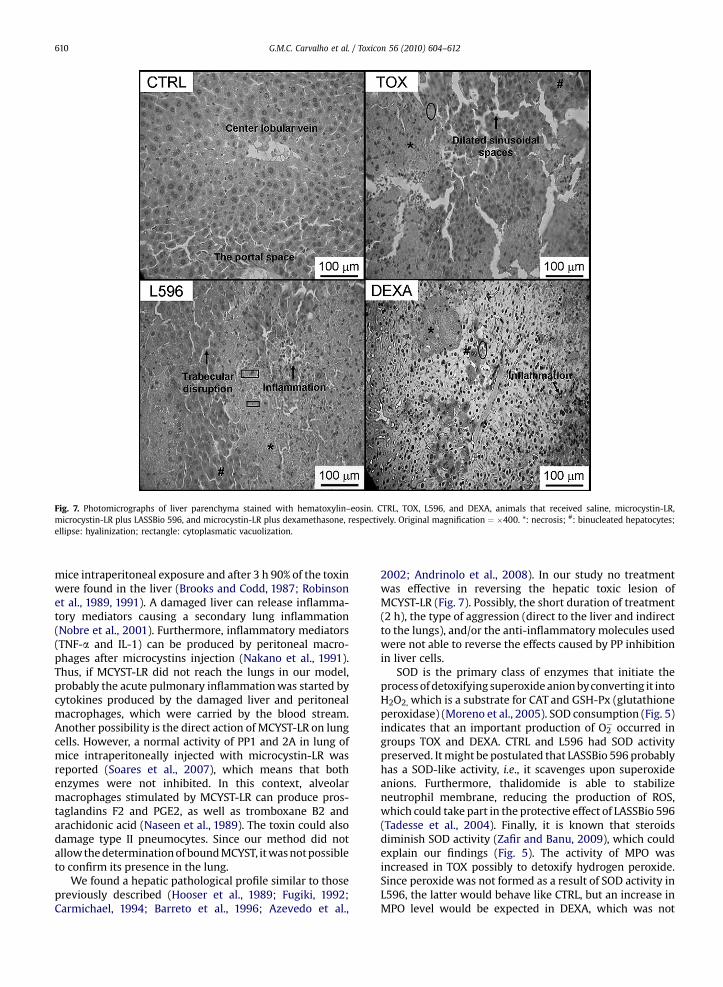
Fig. 7. Photomicrographs of liver parenchyma stained with hematoxylin–eosin. CTRL, TOX, L596, and DEXA, animals that received saline, microcystin-LR,microcystin-LR plus LASSBio 596, and microcystin-LR plus dexamethasone, respectively. Original magnification ¼ �400. *: necrosis; #: binucleated hepatocytes;ellipse: hyalinization; rectangle: cytoplasmatic vacuolization.
G.M.C. Carvalho et al. / Toxicon 56 (2010) 604–612610
mice intraperitoneal exposure and after 3 h 90% of the toxinwere found in the liver (Brooks and Codd, 1987; Robinsonet al., 1989, 1991). A damaged liver can release inflamma-tory mediators causing a secondary lung inflammation(Nobre et al., 2001). Furthermore, inflammatory mediators(TNF-a and IL-1) can be produced by peritoneal macro-phages after microcystins injection (Nakano et al., 1991).Thus, if MCYST-LR did not reach the lungs in our model,probably the acute pulmonary inflammationwas started bycytokines produced by the damaged liver and peritonealmacrophages, which were carried by the blood stream.Another possibility is the direct action of MCYST-LR on lungcells. However, a normal activity of PP1 and 2A in lung ofmice intraperitoneally injected with microcystin-LR wasreported (Soares et al., 2007), which means that bothenzymes were not inhibited. In this context, alveolarmacrophages stimulated by MCYST-LR can produce pros-taglandins F2 and PGE2, as well as tromboxane B2 andarachidonic acid (Naseen et al., 1989). The toxin could alsodamage type II pneumocytes. Since our method did notallowthedeterminationofboundMCYST, itwasnotpossibleto confirm its presence in the lung.
We found a hepatic pathological profile similar to thosepreviously described (Hooser et al., 1989; Fugiki, 1992;Carmichael, 1994; Barreto et al., 1996; Azevedo et al.,
2002; Andrinolo et al., 2008). In our study no treatmentwas effective in reversing the hepatic toxic lesion ofMCYST-LR (Fig. 7). Possibly, the short duration of treatment(2 h), the type of aggression (direct to the liver and indirectto the lungs), and/or the anti-inflammatory molecules usedwere not able to reverse the effects caused by PP inhibitionin liver cells.
SOD is the primary class of enzymes that initiate theprocessof detoxifying superoxideanionbyconverting it intoH2O2, which is a substrate for CAT and GSH-Px (glutathioneperoxidase) (Moreno et al., 2005). SOD consumption (Fig. 5)indicates that an important production of O2
� occurred ingroups TOX and DEXA. CTRL and L596 had SOD activitypreserved. Itmight be postulated that LASSBio 596 probablyhas a SOD-like activity, i.e., it scavenges upon superoxideanions. Furthermore, thalidomide is able to stabilizeneutrophil membrane, reducing the production of ROS,which could take part in the protective effect of LASSBio 596(Tadesse et al., 2004). Finally, it is known that steroidsdiminish SOD activity (Zafir and Banu, 2009), which couldexplain our findings (Fig. 5). The activity of MPO wasincreased in TOX possibly to detoxify hydrogen peroxide.Since peroxide was not formed as a result of SOD activity inL596, the latter would behave like CTRL, but an increase inMPO level would be expected in DEXA, which was not

G.M.C. Carvalho et al. / Toxicon 56 (2010) 604–612 611
verified. However, it cannot be forgotten that steroidsstabilize lysossomalmembranes, thus avoidingMPO releaseby PMN (Rugstad, 1988). The same reasoning could beapplied to CAT (Zafir and Banu, 2009). As a final result, TOXpresented higher TBA levels than CTRL, L596 and DEXA thatdid not differ among them. Oxidative damage, as indicatedby TBARS,was present solely inTOX. In this line, exposure toMCYST-LR increases the products of lipid peroxidation, asevidenced by accumulation of TBARS in liver and kidney ofrats and fish (Moreno et al., 2005). Taken together, ourresults indicated that in this model of lung inflammationoxidative stress was avoided principally by LASSBio 596.
5. Conclusion
The results of present study showed that both LASSBio596 and dexamethasone reduced the inflammation andchanges in lung mechanics induced by MCYST-LR, but thebeneficial effects of LASSBio 596 outperformed those ofdexamethasone. LASSBio 596 effectively prevented respi-ratory mechanical changes, minimized lung morpho-metrical alterations, inflammation and oxidative stress ina mouse model of microcystin-induced acute lung injury.Although mouse and man share many basic physiologicalprocesses, the results reported herein cannot be extrapo-lated directly to humans.
Acknowledgements
The authors are grateful to João Luiz Coelho Rosas Alvesand Antonio Carlos de Souza Quaresma for their skillfultechnical assistance in our laboratory. This study was sup-ported by: The Centers of Excellence Program (PRONEX/FAPERJ), The Brazilian Council for Scientific and Techno-logical Development (CNPq), The Carlos Chagas Filho Rio deJaneiro State Research Supporting Foundation (FAPERJ),The National Institute of Science and Technology of Drugsand Medicines (INCT-INOFAR).
Conflict of interest statement
The authors declare that there are no conflicts ofinterest.
References
Aebi, H., 1984. Catalase in vitro. Methods Enzymol. 105, 121–126.Andrinolo, D., Sedan, D., Telese, L., Aura, C., Masera, S., Giannuzzi, L.,
Marra, C.A., Alaniz, M.J.T., 2008. Hepatic recovery after damageproduced by sub-chronic intoxication with the cyanotoxin micro-cystin LR. Toxicon 51, 457–467.
Azevedo, S.M.F.O., Carmichael, W.W., Jochimsen, E.M., Rinehart, K.L.,Lau, S., Shaw, G.R., Eaglesham, G.K., 2002. Human intoxication bymicrocystins during renal dialysis treatment in Caruaru – Brazil.Toxicology 181, 441–446.
Bannister, J.V., Calabrese, L., 1987. Assays for superoxide dismutase.Methods Biochem. Anal. 32, 279–312.
Barreto, V., Lira, V., Figueiredo, J., Fittipaldi, H., Juca, N., Gayotto, L.C.,Raposso, F., Barbosa, J., Holmes, C., Cardo, C., Azevedo, S.M.F.O.,Carmichael, W., 1996. Caruaru Syndrome-a previously undescribedform of acute toxic liver disease in humans caused by microcystin-LRwith high lethality rate. Hepatology 24 (4 Pt 2), 244 (Abstract Am.Assoc. for the Study of Liver Diseases).
Bates, J.H.T., Rossi, A., Milic-Emili, J., 1985. Analysis of the behavior of therespiratory system with constant inspiration flow. J. Appl. Physiol. 58,1840–1848.
Bates, J.H.T., Ludwig,M.S., Sly, P.D., Brown,K.,Martin, J.G., Fredberg, J.J.,1988.Interrupter resistance elucidated by alveolar pressuremeasurement inopen-chest normal dogs. J. Appl. Physiol. 65, 408–414.
Bradford, M.M., 1976. A rapid and sensitive method for the quantificationof microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254.
Brooks, W.P., Codd, G.A., 1987. Distribution of Microcystis aeruginosapeptide toxin and interactions with hepatic microsomes in mice.Pharmacol. Toxicol. 60, 187–191.
Campos, H.S., Xisto, D.G., Oliveira, M.B., Teixeira, I., Negri, E.M., Mauad, T.,Carnielli, D., Lima, L.M., Barreiro, E.J., Faffe, D.S., Zin, W.A., Lapa eSilva, J.R., Rocco, P.R., 2006. Protective effects of phosphodiesteraseinhibitors on lung function and remodeling in a murine model ofchronic asthma. Braz. J. Med. Biol. Res. 39, 283–287.
Carmichael, W.W., 1994. The toxins of cyanobacteria. Sci. Am. 270,78–86.
Chorus, I., Bartram, J., 1999. Toxic Cyanobacteria in Water – A Guide toTheir Public Health Consequences, Monitoring and Management. E &FN Spon, London.
Draper, H.H., Hadley, M., 1990. Malondialdehyde determination as indexof lipid peroxidation. Methods Enzymol. 186, 421–431.
Dawson, R.M., 1998. The toxicology of microcystins. Toxicon 36, 953–962.Feurstein, D., Holst, K., Fischer, A., Dietrich, D.R., 2009. Oatp-associated
uptake and toxicity of microcystin in primary murine whole braincells. Toxicol. Appl. Pharmacol. 234, 247–255.
Fugiki, H., 1992. Is the inhibition of protein phosphatase 1 and 2A activ-ities a general mechanism of tumor promotion in human cancerdevelopment? Mol. Carcinog. 5, 91–94.
Funari, E., Testai, E., 2008. Human health risk assessment related tocyanotoxins exposure. Crit. Rev. Toxicol. 38, 97–125.
Gundersen, H.J.G., Bendtsen, T.F., Korbo, L., Marcussen, N., Moller, A.,Nielsen, K., Nyengaard, J.R., Pakkenberg, B., Sorensen, F.B., Vesterby, A.,West, M.J., 1988. Some new, simple and efficient stereological methodsand their use in pathological research and diagnosis. Acta Pathol.Microbiol. Immunol. Scand. 96, 379–394.
Hooser, S.B., Beasley, V.R., Lovell, R.A., Carmichael, W.W., Haschek, W.M.,1989. Toxicity of microcystin-LR, a cyclic heptapeptide hapatotoxinfrom Microcystis aeruginosa, to rats and mice. Vet. Pathol. 26,246–252.
Ito., E., Kondo, F., Harada, K.-I., 2000. First report on the distribution oforally administered microcystin-LR in mouse tissue using an immu-nostaining method. Toxicon 38, 37–48.
Ito, E., Kondo, F., Harada, K.-I., 2001. Intratracheal administration ofmicrocystin-LR and its distribution. Toxicon 39, 265–271.
Kujbida, P., Hatanaka, E., Campa, A., Curi, R., Farsky, S.H.P., Pinto, E., 2008.Analysis of chemokines and reactive oxygen species formation by ratand human neutrophils induced by microcystin-LA, -YR and -LR.Toxicon 51, 1274–1280.
Lima, L.M., Castro, P., Machado, A.L., Fraga, C.A., Lugnier, C., de Moraes, V.L.,Barreiro, E.J., 2002. Synthesis and anti-inflammatory activity ofphthalimide derivatives, designed as new thalidomide analogues.Bioorg. Med. Chem. 10, 3067–3073.
Miotla, J.M., Teixeira, M.M., Hellewell, P.G., 1998. Supression of acute lunginjury in mice by an inhibitor of phosphodiesterase type 4. Am. J.Respir. Cell. Mol. Biol. 18, 411–420.
Moreno, I.M., Pichardo, S., Jos, A., Gomez-Amores, L., Mate, A., Vazquez, C.M., Camean, A.M., 2005. Antioxidant enzyme activity and lipid per-oxidation in liver and kidney of rats exposed to microcystin-LRadministered intraperitoneally. Toxicon 45, 395–402.
Mortola, J.P., Noworaj, J.A., 1983. Two-sidearm tracheal cannula forrespiratory airflow measurements in small animals. J. Appl. Physiol.55, 250–253.
Nakano, Y., Shirai, M., Mori, N., Nakano, M., 1991. Neutralization ofmicrocystin shock in mice by tumor necrosis factor alpha antiserum.Appl. Environ. Microbiol. 57, 327–330.
Naseen, S.M., Hines, H.B., Creasia, D.A., 1989. Effect of toxins on arach-idonic acid metabolism in rat cultured pulmonary alveolar macro-phages. Biochem. Int. 19, 583–592.
Nobre, A.C.L., Coelho, G.R., Coutinho, M.C.M., Silva, M.M.M., Angelim, E.V.,Menezes, D.B., Fonteles, M.C., Monteiro, H.S.A., 2001. The role ofphospholipase A2 and cyclooxygenase in renal toxicity induced bymicrocystin-LR. Toxicon 39, 721–724.
Nobre, A.C.L., Martins, A.M., Hayt, A., Benevides, C., Lima, A.A.,Fonteneles, M.C., Monteiro, H.S., 2003. Renal effects of supernatantfrom rat peritoneal macrophages activated by microcystin-LR: roleprotein mediators. Toxicon 41, 377–381.

G.M.C. Carvalho et al. / Toxicon 56 (2010) 604–612612
Picanço, M.R., Soares, R.M., Cagido, V.R., Azevedo, S.M.F.O., Rocco, P.R.M.,Zin, W.A., 2004. Toxicity of a cyanobacterial extract containingmicrocystins to mouse lungs. Braz. J. Med. Biol. Res. 37, 1225–1229.
Robinson, N.A., Miura, G.A., Matson, C.F., Dinterman, R.E., Pace, J.G., 1989.Characterization of chemically tritiated microcystin-LR and itsdistribution in mice. Toxicon 27, 1035–1042.
Robinson, N.A., Pace, J.G., Matson, C.F., Miura, G.A., Lawrence, W.B., 1991.Tissue distribution, excretion and hepatic biotransformation ofmicrocystin-LR in mice. J. Pharmacol. Exp. Ther. 256, 176–182.
Rocco, P.R., Momesso, D.P., Figueira, R.C., Ferreira, H.C., Cadete, R.A.,Légora-Machado, A., Koatz, V.L., Lima, L.M., Barreiro, E.J., Zin, W.A.,2003. Therapeutic potential of a new phosphodiesterase inhibitor inacute lung injury. Eur. Respir. J. 22, 20–27.
Rugstad, H.E., 1988. Antiinflammatory and immunoregulatory effects ofglucocorticoids:modeofaction. Scand. J. Rheumatol. Suppl. 76,257–264.
Saldiva, P.H., Zin, W.A., Santos, R.L., Eidelman, D.H., Milic-Emili, J., 1992.Alveolar pressure measurement in open-chest rats. J. Appl. Physiol.72, 302–306.
Soares, R.M., Cagido, V.R., Ferraro, R.B., Meyer-Fernandes, J.R., Rocco, R.M.,Zin, W.A., Azevedo, S.M.F.O., 2007. Effects of microcystin-LR on mouselungs. Toxicon 50, 330–338.
Suzuki, K., Ota, H., Sasagawa, S., Sakatani, T., Fujikura, T., 1983. Assaymethod for myeloperoxidase in human polymorphonuclear leuko-cytes. Anal. Biochem. 132, 345–352.
Tadesse, A., Sandoval, F., Shannon, E.J., 2004. Stabilization of red blood cellmembranes by thalidomide in vitro. Immunopharmacol. Immuno-toxicol. 26, 501–509.
Teixeira, Mda G., Costa, Mda C., Carvalho, V.L.P., Pereira, Mdos. S.,Hage, E., 1993. Gastroenteritis epidemic in the area of theItaparica dam, Bahia, Brazil. Bull. Pan Am. Health Organ 27,244–253.
Turner, C.R., 1993. Therapeutic intervention in a rat model of ARDS: IV.Phosphodiesterase IV inhibition. Circ. Shock 39, 237–245.
Turner, P.C., Gamme, A.J., Hollinrake, K., Codd, G.A., 1990. Pneumoniaassociated with cyanobacteria. Br. Med. J. 300, 1400–1414.
Wang, Q., Xie, P., Chen, J., Liang, G., 2008. Distribution of microcystinin various organs (heart, liver, intestine, gonad, brain, kidneyand lung) of Wistar rat via intravenous injection. Toxicon 52,721–727.
Zafir, A., Banu, N., 2009. Modulation of in vivo oxidative statusby exogenous corticosterone and restraint stress in rats. Stress 12,167–177.






![[1990] Tonga LR 99 - Crown Law Tonga](https://static.fdokumen.com/doc/165x107/63221e49887d24588e0416ae/1990-tonga-lr-99-crown-law-tonga.jpg)













