
Microcystin diversity in a Planktothrix rubescens population from Lake Albano (Central Italy)
15
Toxicon 48 (2006) 160–174 Microcystin diversity in a Planktothrix rubescens population from Lake Albano (Central Italy) V. Messineo a , D. Mattei a , S. Melchiorre a , G. Salvatore a , S. Bogialli b , R. Salzano c , R. Mazza c , G. Capelli c , M. Bruno a, a Department of Environment and Primary Prevention, National Institute of Health, viale Regina Elena, 299– 00161, Rome, Italy b Department of Chemistry, University ‘‘La Sapienza’’, P.le Aldo Moro 5, 00185, Rome, Italy c Department of Geology, University of Rome ‘‘Roma 3’’, Largo San Leonardo Murialdo , 1-00100, Rome, Italy Received 19 December 2005; received in revised form 3 April 2006; accepted 7 April 2006 Available online 25 April 2006 Abstract The cyanobacterium Planktothrix rubescens Anagnostidis & Komarek (previously Oscillatoria rubescens DC ex Gomont) is present in several Italian lakes and it is known to produce cyanotoxins. The dynamics and toxin production of P. rubescens population in Lake Albano, a volcanic crater lake in Central Italy, has been studied for 5 years (January 2001–April 2005). Winter–spring superficial blooms with frequent scums were observed every year. Total microcystin (MC) levels were measured from April 2004 to October 2005 by liquid chromatography-tandem mass spectrometry. MC levels up to 14.2 mg/l were measured, with high concentrations found in summer at a 20–25 m depth. The intracellular toxin content varied between 1.5 (surface, January 2004) and 0.21 pg/cell (surface, May 2004). Six different MCs were detected, the most abundant being two desmethyl-MC-RR isomers. Of the 13 water wells monitored in the Lake Albano area, two of them showed MC contamination during winter, confirming the ability of these toxins to migrate through groundwater towards public water sources. These results highlight the need for further studies on the mobility and fate of these pervasive cyanobacterial toxins. r 2006 Elsevier Ltd. All rights reserved. Keywords: Lake Albano; Cyanotoxins; Microcystins; Planktothrix rubescens 1. Introduction Cyanobacterial blooms occur worldwide from USA to Australia, causing human health risks and animal deaths as well. In Europe, they have been detected in Belgium, Germany, France, Finland, Norway, United Kingdom, Hungary, Portugal, Spain and Italy (Sivonen and Jones, 1999; Barco et al., 2004). The most common cyanobacterium in deep European prealpine lakes is the filamentous specie Planktothrix rubescens Anagnostidis & Komarek (previously Oscillatoria rubescens DC ex Gomont) (Salmaso, 2000). Like the closely related specie Planktothrix aghardii, P. rubescens efficiently gen- erates microcystins (MCs), which are hepatotoxins acting as specific inhibitors of protein phosphatases 1 and 2A. They dephosphorylate serine and ARTICLE IN PRESS www.elsevier.com/locate/toxicon 0041-0101/$ - see front matter r 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.toxicon.2006.04.006 Corresponding author. Tel.: +390649902173; fax: +390649387083. E-mail address: [email protected] (M. Bruno).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Microcystin diversity in a Planktothrix rubescens population from Lake Albano (Central Italy)

ARTICLE IN PRESS
0041-0101/$ - se
doi:10.1016/j.to
�Correspondifax: +39064938
E-mail addre
Toxicon 48 (2006) 160–174
www.elsevier.com/locate/toxicon
Microcystin diversity in a Planktothrix rubescenspopulation from Lake Albano (Central Italy)
V. Messineoa, D. Matteia, S. Melchiorrea, G. Salvatorea, S. Bogiallib, R. Salzanoc,R. Mazzac, G. Capellic, M. Brunoa,�
aDepartment of Environment and Primary Prevention, National Institute of Health, viale Regina Elena, 299– 00161, Rome, ItalybDepartment of Chemistry, University ‘‘La Sapienza’’, P.le Aldo Moro 5, 00185, Rome, Italy
cDepartment of Geology, University of Rome ‘‘Roma 3’’, Largo San Leonardo Murialdo , 1-00100, Rome, Italy
Received 19 December 2005; received in revised form 3 April 2006; accepted 7 April 2006
Available online 25 April 2006
Abstract
The cyanobacterium Planktothrix rubescens Anagnostidis & Komarek (previously Oscillatoria rubescens DC ex
Gomont) is present in several Italian lakes and it is known to produce cyanotoxins. The dynamics and toxin production of
P. rubescens population in Lake Albano, a volcanic crater lake in Central Italy, has been studied for 5 years (January
2001–April 2005). Winter–spring superficial blooms with frequent scums were observed every year. Total microcystin
(MC) levels were measured from April 2004 to October 2005 by liquid chromatography-tandem mass spectrometry. MC
levels up to 14.2mg/l were measured, with high concentrations found in summer at a 20–25m depth. The intracellular toxin
content varied between 1.5 (surface, January 2004) and 0.21 pg/cell (surface, May 2004). Six different MCs were detected,
the most abundant being two desmethyl-MC-RR isomers. Of the 13 water wells monitored in the Lake Albano area, two of
them showed MC contamination during winter, confirming the ability of these toxins to migrate through groundwater
towards public water sources. These results highlight the need for further studies on the mobility and fate of these pervasive
cyanobacterial toxins.
r 2006 Elsevier Ltd. All rights reserved.
Keywords: Lake Albano; Cyanotoxins; Microcystins; Planktothrix rubescens
1. Introduction
Cyanobacterial blooms occur worldwide fromUSA to Australia, causing human health risks andanimal deaths as well. In Europe, they have beendetected in Belgium, Germany, France, Finland,Norway, United Kingdom, Hungary, Portugal,
e front matter r 2006 Elsevier Ltd. All rights reserved
xicon.2006.04.006
ng author. Tel.: +390649902173;
7083.
ss: [email protected] (M. Bruno).
Spain and Italy (Sivonen and Jones, 1999; Barcoet al., 2004).
The most common cyanobacterium in deepEuropean prealpine lakes is the filamentous speciePlanktothrix rubescens Anagnostidis & Komarek(previously Oscillatoria rubescens DC ex Gomont)(Salmaso, 2000). Like the closely related speciePlanktothrix aghardii, P. rubescens efficiently gen-erates microcystins (MCs), which are hepatotoxinsacting as specific inhibitors of protein phosphatases1 and 2A. They dephosphorylate serine and
.

ARTICLE IN PRESSV. Messineo et al. / Toxicon 48 (2006) 160–174 161
threonine residues in animals and plants. Theinhibition of PP1 and PP2A results in an increaseof the phosphorylation of proteins in the liver cells,affecting several processes like metabolism, cellcontractility, membrane transport, secretion, celldivision and gene transcription and translation.
From a structural point of view, MCs are a classof monocyclic heptapeptides consisting of D-alanineat position 1, two variable L-amino acids atpositions 2 and 4, g-linked D-glutamic acid atposition 6, and 3 unusual amino acids; b-linkedD-erythro-b-methylaspartic acid (MeAsp) at posi-tion 3; (2S, 3S, 8S, 9S)-3-amino-9- methoxy- 2, 6, 8trimethyl-10-phenyldeca-4, 6-dienoic acid (Adda) atposition 5 and N-methyl dehydroalanine (MDha) atposition 7. So far, about 75 variant MCs are known(Sivonen and Jones, 1999). Besides being hepato-toxic compounds (Falconer et al., 1983; Teixeira daGloria Lima Crux et al., 1993; Harada and Teuji,1998; Jochimsen et al., 1998), MCs are tumourpromoters (Falconer and Buckley, 1989; Nishiwaki-Matsushima et al., 1992, Ito et al., 1997; Ueno et al.,1996; Humpage et al., 2000; Fleming et al., 2002;Zhou et al., 2002), endocrine-disruptors (Leierset al., 2000; Ford et al., 1996; Sayed et al., 1997;Rojas et al., 1990; Hernandez et al., 2000) andimmunotoxicants (Lankoff et al., 2004).
MCs have therefore a synergistic effect, welldocumented in experimental studies on mice(Fitzgeorge et al., 1994). In these studies, dailysubacute doses of MCs, intranasally administred tomice for 7-day periods, caused cumulative patholo-gic effects two times stronger than those producedby the sum of the MCs administered as a singledose. Bioaccumulation has been proposed to beresponsible for this effect (Solter et al., 1998).Recently, the need for a better understanding ofcongener-specific MC kinetics in general and inhumans, has been recognized in order to assess therisk in a chronic exposure setting (Dietrich andHoeger, 2005).
The increase in occurrence of cyanobacterial toxicblooms during the last 30 years led the ItalianNational Institute of Health to perform preliminaryscreenings in selected areas of the country in orderto assess human risk.
During these screenings, some volcanic lakes inthe neighbourhood of Rome were found to hostdominant populations of P. rubescens. The dy-namics and toxicity of populations in one of theselakes, Lake Albano, were studied and are presentedin this report along with their relationship with
water quality parameters such as temperature, pH,dissolved oxygen (DO), conductivity, total phos-phorus (TP), total nitrogen (TN) and chlorophyll a.This information will be used to identify restorationstrategies and to evaluate the health hazard inhuman uses.
2. Materials and methods
2.1. Site description
Lake Albano is a volcanic basin inside a craterdepression with an ellipsoid shape whose longeraxis, NW–SE oriented, is 3.5 km and the shorter oneis 2.3 km (Chondrogianni et al., 1996). The lakeperimeter is 12 km long and his depth reaches 175m.The mean renewal time is more than 67 years.Lacking outlet tributaries, the lake is characterizedby an ancient Roman channel carved in the lavarocks through the crater, which is used to regulatethe lake level. The lake is eutrophic due to humanactivities, that in the past caused development ofalgal blooms; the first evidence of this was high-lighted by Cannicci (1954). Since 1960 the lake levellowered almost four meters under the artificialoutflow, leading to changing from an oligotrophiccondition to an eutrophic one.
2.2. Field measurements, sample collection and
microscopic observations
Water samples for chemical, toxicological andbiological analysis were collected monthly, using a2.5 l Ruttner bottle, from four surface stationssituated close to the lake shore and from one singlestation at the centre of the lake where the samplingdepths were 0, 5, 10, 15, 20, 25 and 30m (Fig. 1). Inorder to investigate the persistence of MCs ingroundwater, 12 well water samples were alsoanalysed. These wells are used for drinking watersupply inside and outside the volcanic crater.Temperature, conductivity, DO, and pH weremeasured on field, using a multiparametric probe(Mettler Toledo InLabs 781), and chlorophyll a,TP, TN, microscopic analysis, toxin extraction,Vibrio fischeri bioassay and toxin quantificationwere performed in the laboratory. The samples werestored in ice chests and transported to the labora-tory. For microscopic observations subsamples werepreserved in 1% Lugol’s solution and analysed byan inverted microscope (Leitz Labovert FS), ac-cording to Utermohl (1931) and Lund et al. (1958),

ARTICLE IN PRESS
Fig. 1. Map of Lake Albano, with the sampling stations (1–5)
and the contaminated wells (6–7).
V. Messineo et al. / Toxicon 48 (2006) 160–174162
using 25ml sedimentation chambers for phyto-plankton identification and cell density estimation.
2.3. Cell culture
Following a procedure developed by Stein (1973),single trichomes of P. rubescens were isolated fromwinter blooms and grown in non-axenic cultures in250ml Erlenmeyer flasks with 200ml of BG 11medium (Sigma-Aldrich, Munchen, Germany), at13 1C, with a 2.000 lux 16:8 h LD cycles. Thecultures were grown for 2 months, to producesufficient biomasses for the analysis.
2.4. Phosphorus and nitrogen analyses
Analyses were carried out within 24 h aftersampling, using a Perkin Elmer Lambda 25 UV/VS spectrophotometer, supported by a UV-WinLabsoftware. Samples were digested in persulphate todissolve organic and non-organic particulate matterin suspension, as described by Lambert and Maher(1995), in order to allow the simultaneous detectionof both TP and TN.
TP was measured following the method ofMurphy and Riley (1962). TN was determined asdescribed by Ferree and Shannon (2001). Allreagents were high-purity grade.
2.5. Toxin extraction
Toxins were extracted from algae following theprocedure described by Meriluoto and Eriksson(1988). Fresh algal aliquots (10–50mg) obtained bycentrifugation of water samples, were extracted twotimes with 2ml portions of sterile bidistilled H2O.The solution was stirred, sonicated (5min at30–40 1C) (Vibra-Cell, Sonics & Materials Inc.),and then centrifuged for 10min at 11.000 rpm(Beckman, LT-55 Ultracentrifuge) to eliminatedebris. The supernatant was then collected and thewhole process repeated twice. Water samples filteredthrough G/C filters were analysed for extracellularMCs.
2.6. V. fischeri bioassay
The Microtox system (Azur Environmental Inc.,USA), working with the luminescent bacteriumV. fischeri (US Environmental Protection Agency,1979), was developed for detecting aquatic toxicants(McFeters et al., 1983) and adapted to algal toxindetection (Bruno et al., 1992), by using non-acidic,non-hydrophobic extractions (Meriluoto and Eriks-son, 1988; Gjolme and Utkilen, 1996), due to thevery high sensitivity of this bacterium to even traceamounts of solvents.
The use of simple sterile bidistilled water assolvent allows high-efficiency extraction of toxins(Gjolme and Utkilen, 1996) without any risk offalse-positive results from Microtox.
The algal extract in four progressive dilutions wasadded to known concentrations of V. fischeri andtested to give the EC50 (effective concentrationvalue producing 50% reduction of light emissionfrom bacteria, after 5 and 15min of addition to theextract). The value is statistically calculated bysoftware (Microtox Omni Software) for capturingand analysing test data.
2.7. Analysis of MCs by liquid chromatography
(LC) tandem mass spectrometry
MC-LR, MC-RR, MC-YR, MC-LA and MC-LWwere purchased from Calbiochem-Novabiochem,Nottingham, UK. The internal standard (IS), anobsolete carbamate insecticide (trimethacarb), wasobtained from Sigma-Aldrich. A composite workingstandard solution of the MCs was prepared bymixing the stock solutions and diluting with a water/methanol (50:50, v/v) solution to obtain analyte

ARTICLE IN PRESSV. Messineo et al. / Toxicon 48 (2006) 160–174 163
concentrations of 5mg/ml, while a 1.5mg/ml of the ISsolution was prepared by diluting with methanol.When unused, the analytes solution was storedat �251.
Acetonitrile of gradient grade was obtained fromCarlo Erba (Milano, Italy). Trifluoroacetic acid(TFA) was from Aldrich.
Extraction cartridges filled with 0.5 g Carbograph4, and drilled cylindrical Teflon pistons withindented bases and Luer tips for analyte elution inthe back-flushing mode (Di Corcia et al., 1994) weresupplied by LARA (Rome, Italy). Carbograph 4 isan example of the graphitised carbon black sorbentfamily having a surface area of �200m2/g. The SPEcartridge was fitted into a side-arm filtration flask,and liquids were forced through the cartridge byvacuum (water pump). Before processing aqueoussamples, the cartridge was washed with 20ml ofwater acidified with HCl (pH 2), followed by 5ml ofdistilled water.
For MC analyses, 1 l of lake water was passedthrough the SPE cartridge at a flow rate of 20ml/min. The cartridge was then washed by passingthrough it sequentially 20ml of distilled water and0.4ml of methanol. Then, a Teflon piston wasforced into the cartridge until it touched the upperfrit, the cartridge was reversed, and MCs were backeluted with 1ml of methanol followed by 4ml ofCH2Cl2/CH3OH (80:20, v/v) solution containing10mmol/l TFA. Elution in the back-flushing modewas necessary to re-extract MC-LW quantitativelywith a moderate volume of the eluent phase.Analytes were collected in a 1.4-cm i.d. glass vialwith a conical bottom. Before removing solvents ina water bath at 50 1C under a nitrogen stream, 15 mlof the solution containing the IS were added to theeluate. To avoid some analyte loss, solvent removalwas stopped to a volume of �50 ml of the eluate. Theresidue was reconstituted with 200 ml of wateracidified with 10mmol/l formic acid and 50 ml ofthis solution was injected into the LC column.
The LC instrument consisted of a Waters pump(Model 1525m, Milford, USA), a 50 ml-injectionloop, and Alltima 5-mm C-18 guard (7.5� 4.6mmi.d.) and analytical (250� 4.6mm i.d.) columns(Alltech) thermostated at 35 1C and was interfacedto a benchtop triple-quadrupole mass spectrometer(Model Micromass 4 MICRO API, Waters). Forfractionating the analytes, phase A was acetonitrileand phase B was water. Both phases contained10mmol/l formic acid. At 1.0ml/min, the mobilephase linear gradient profile was as follows (t in
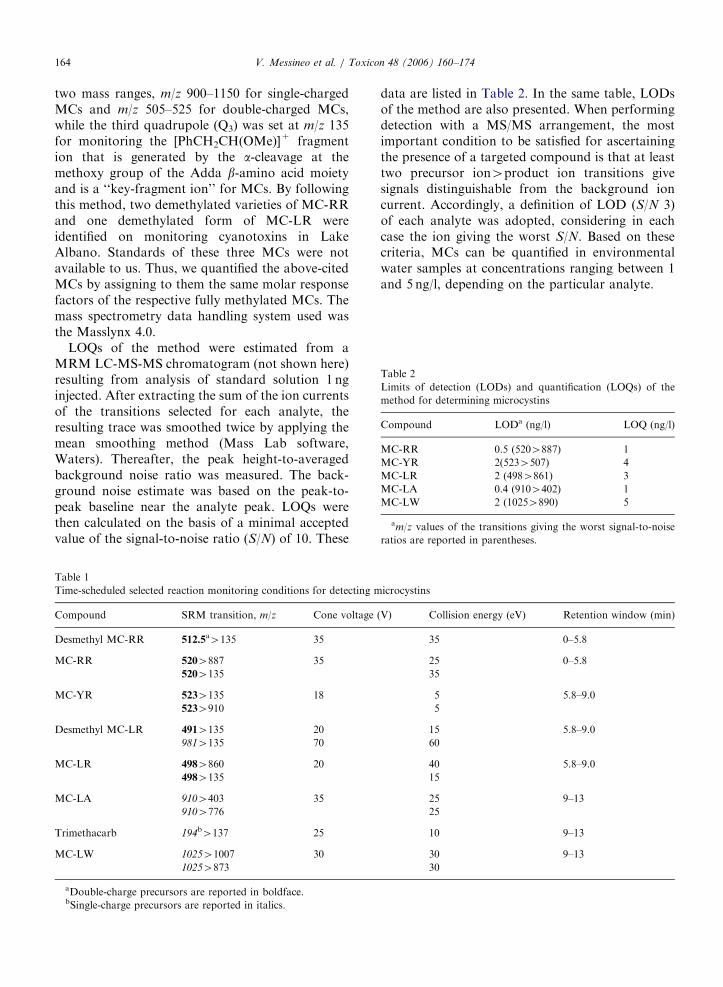
min): t0, A ¼ 30%; t15, A ¼ 60%; t16, A ¼ 100%;t19, A ¼ 100%; t20, A ¼ 30%; t29, A ¼ 20%. Ana-lyte retention times varied p0.5% over two weeks.A diverter valve led the effluent into the electrosprayion source (ESI), operating in the positive mode,with a flow-rate of 300 ml/min only between 4 and17min of the chromatographic run. High-puritynitrogen was used as drying and curtain gases; high-purity argon was used as collision gas. Desolvationgas was set at 650 l/h, while the cone gas was set at50 l/h. The probe and desolvation temperatureswere maintained respectively at 100 and 350 1C.The gas pressure in the collision cell was set at3mbar. Capillary voltage was 3000V, extractorvoltage was 0V. Cone voltage, collision energy andothers transmission parameters were optimized foreach analyte (data are reported in Table 1). Massaxis calibration of each mass-resolving quadrupoleQ1 and Q3 was performed by infusion of a sodiumand cesium iodide solution at 10 ml/min. Unit massresolution was established and maintained in eachmass-resolving quadrupole by keeping a full-widthat half-maximum of approximately 0.7 amu. All thesource and instrument parameters for monitoringeach MC were optimized by water/methanol (50:50,v/v) solution acidified with 10mmol/l formic acidcontaining the analyte at 1 mg/ml levels, infused at10 ml/min by a syringe pump. The multi reaction-monitoring (MRM) mode was used for quantitationby selecting the best two fragmentation reactions foreach analyte, except for the internal standard(trimethacarb) and desmethyl MCs (Table 1).
MCs for which standards were available werequantified by the external standard quantificationprocedure. Standard solutions were prepared at 8levels by using appropriate volumes of the workingstandard solution. For each analyte, the peak areaversus injected amount chart was obtained bymeasuring at any injected amount the resultingpeak area and relating this area to that of theinternal standard. The response of the ESI-MS/MSsystem was linearly related to injected amounts ofthe analytes up to 300 ng. Sometimes, amounts ofMCs injected from final extracts of lake watersamples exceeded the upper limit of the lineardynamic range of the mass detector response. Inthese cases, extracts were suitably diluted and re-injected. Hummert et al. (2001) demonstrated theusefulness of LC-MS equipped with a triple quad-rupole in identifying those MCs for which standardsare unavailable. To do this, the first quadrupole(Q1) was operated in the scan mode after selecting
ARTICLE IN PRESS
Table 2
Limits of detection (LODs) and quantification (LOQs) of the
method for determining microcystins
Compound LODa (ng/l) LOQ (ng/l)
MC-RR 0.5 (5204887) 1
MC-YR 2(5234507) 4
MC-LR 2 (4984861) 3
MC-LA 0.4 (9104402) 1
MC-LW 2 (10254890) 5
am/z values of the transitions giving the worst signal-to-noise
ratios are reported in parentheses.
V. Messineo et al. / Toxicon 48 (2006) 160–174164
two mass ranges, m/z 900–1150 for single-chargedMCs and m/z 505–525 for double-charged MCs,while the third quadrupole (Q3) was set at m/z 135for monitoring the [PhCH2CH(OMe)]+ fragmention that is generated by the a-cleavage at themethoxy group of the Adda b-amino acid moietyand is a ‘‘key-fragment ion’’ for MCs. By followingthis method, two demethylated varieties of MC-RRand one demethylated form of MC-LR wereidentified on monitoring cyanotoxins in LakeAlbano. Standards of these three MCs were notavailable to us. Thus, we quantified the above-citedMCs by assigning to them the same molar responsefactors of the respective fully methylated MCs. Themass spectrometry data handling system used wasthe Masslynx 4.0.
LOQs of the method were estimated from aMRM LC-MS-MS chromatogram (not shown here)resulting from analysis of standard solution 1 nginjected. After extracting the sum of the ion currentsof the transitions selected for each analyte, theresulting trace was smoothed twice by applying themean smoothing method (Mass Lab software,Waters). Thereafter, the peak height-to-averagedbackground noise ratio was measured. The back-ground noise estimate was based on the peak-to-peak baseline near the analyte peak. LOQs werethen calculated on the basis of a minimal acceptedvalue of the signal-to-noise ratio (S/N) of 10. These
Table 1
Time-scheduled selected reaction monitoring conditions for detecting m
Compound SRM transition, m/z Cone voltage
Desmethyl MC-RR 512.5a4135 35
MC-RR 5204887 35
5204135
MC-YR 5234135 18
5234910
Desmethyl MC-LR 4914135 20
9814135 70
MC-LR 4984860 20
4984135
MC-LA 9104403 35
9104776
Trimethacarb 194b4137 25
MC-LW 102541007 30
10254873
aDouble-charge precursors are reported in boldface.bSingle-charge precursors are reported in italics.
data are listed in Table 2. In the same table, LODsof the method are also presented. When performingdetection with a MS/MS arrangement, the mostimportant condition to be satisfied for ascertainingthe presence of a targeted compound is that at leasttwo precursor ion4product ion transitions givesignals distinguishable from the background ioncurrent. Accordingly, a definition of LOD (S/N 3)of each analyte was adopted, considering in eachcase the ion giving the worst S/N. Based on thesecriteria, MCs can be quantified in environmentalwater samples at concentrations ranging between 1and 5 ng/l, depending on the particular analyte.
icrocystins
(V) Collision energy (eV) Retention window (min)
35 0–5.8
25 0–5.8
35
5 5.8–9.0
5
15 5.8–9.0
60
40 5.8–9.0
15
25 9–13
25
10 9–13
30 9–13
30

ARTICLE IN PRESSV. Messineo et al. / Toxicon 48 (2006) 160–174 165
3. Results
Five hundred and eighty three water samples werecollected from Lake Albano during a 5-year surveystarted in 2001. A series of depth samples weretaken monthly on 46 occasions over the period of 1January 2001 to 31 May 2005.
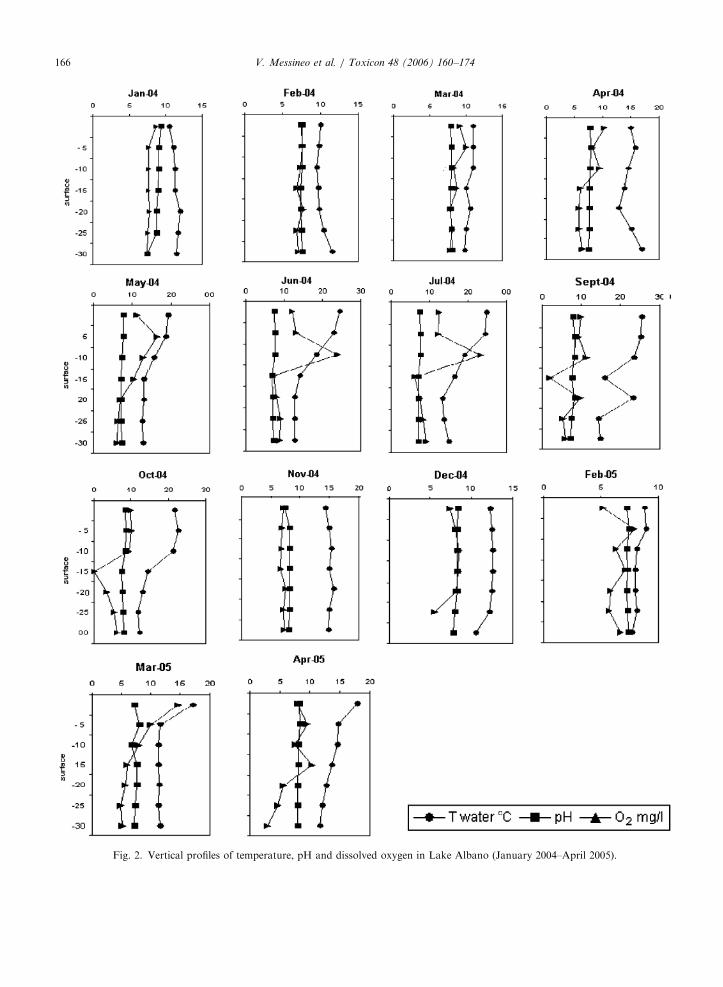
The vertical profiles of DO, temperature and pHin 2004 are shown in Fig. 2. These field measure-ments show variations from April 2004 to May2005, corresponding to the distribution of P.
rubescens (Fig. 3). Lake Albano was thermallystratified from April to October (thermocline from10 to 20m), with the maximum temperaturegenerally reached in August (28 1C at the surface).pH values were between 7 and 9.4, the mean DOvalue was 6.6mg/l in the metalimnic layer and themean annual conductivity value was 502 mS (from434 to 528 mS).
In the epilimnion, TP in the water varied from63 mg/l in July to less than 3 mg/l in April, Novemberand December, while in the hypolimnion the TPdecreased from 146 mg/l in April to less than 3 mg/lin December and November. TN varied from642 mg/l in May (Castello station) to 14 mg/l inNovember in the epilimnion, while in the hypolim-nion varied from 356 mg/l in June (�30m) to 32 mg/l(�30m) in December. TN/TP rates in 2004–2005were calculated along the water column and variedbetween 1 (�30m, July 2004) and 184 (�5m, June2004).
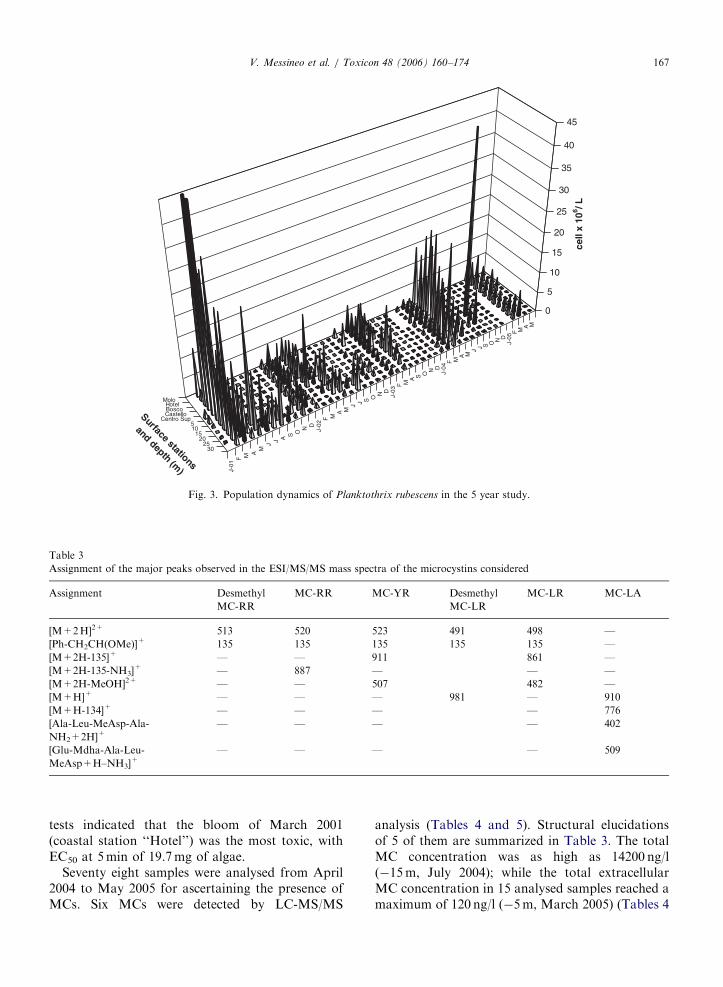
The P. rubescens densities over the 5 years (Fig. 3)reached the annual peak in winter (February/March), with superficial blooms always exceedingthe tolerance limit laid down in Italy, which is5� 106 cell/l, with maxima of 138� 106 cell/l(March 2001, coastal station ‘‘Molo’’) and super-ficial scums of 106� 106 cell/l (April 2005, coastalstation ‘‘Castello’’), and the minimum in September2004 (800 cell/l, coastal station ‘‘Castello’’). Threesuperficial scums were collected (January 2004, May2004 and April 2005) and analysed for toxin content(Table 6). It has to be noted that, during periods ofthermal stratification, the main peak of the popula-tion could be present at depths below the maximumone considered in this study, thus causing a possibleunderestimation of the population size. The densityrapidly increased over the period from January upto early February, when the population begun toaccumulate on the surface, and the ability of thetrichomes to fluctuate or rapidly reach the surfacelayer in few hours, according to the degree of cloudy
or sunny weather, was observed. During our survey,the mean surface temperature of Lake Albano inFebruary ranged between 9.5 and 10.5 1C, and itwas also the lowest mean water temperaturethroughout the year.
During blooms, P. rubescens represented 70–95%of the total phytoplankton. In early spring, the celldensity of P. rubescens decreased and the popula-tion peaks slowly deepened towards the hypolimniclayers from �20 to �30m, where they remainedduring the warmest seasons. In the last part ofApril, when the surface temperature in Lake Albanowas 18 1C, the superficial blooms generally disap-peared, with the exception of 2004, where they werestill present in May (surface temperature from 19 to21 1C). In the June to September period, P.
rubescens abundance in the surface samples wasalways much lower than those in the thermoclineand hypolimnion. Subsurface maximum cyanobac-teria abundance was measured in September 2004,between �10 and �20m (maximum concentrationat �10m). In the September–December period, thepopulation was uniformly distributed along thewater column, the gradual surface concentrationincrease generally beginning in January.
A strange occurrence, an apparently colder (7 1Cof difference) and poorly oxygenated (1–2mg/l)layer was detected in September and October 2004at �15m in the water column; its influenceon the population density and the subsurfacemaximum of September can be noted (Fig. 2). Thiseffect was unclear to us and was evidenced only in2004.
Water samples of wild blooms and pure culturesof P. rubescens were analysed for geosmin and 2-methyl-isoborneol according to Mindrup and Shirey(2000). These musty odour compounds, main causesof odour pollution in waters, are produced byvarious yeasts and cyanobacteria, among which areseveral Oscillatoriaceae (van Breemen et al., 1992).No presence of geosmin and 2-methyl-isoborneol inscums and samples of wild populations collectedfrom the blooms, as well as in P. rubescens purecultures was detected by SPME/GC/MS analysis.All cultures from 40 single trichomes selected fromfour surface water samples grew successfully. Thissuggests that 100% trichomes in these samples wereviable.
During the 5-year survey, all the superficialwater samples and the scums were collectedduring the blooming periods and tested in theMicrotox System, giving positive results. Toxicity
ARTICLE IN PRESS
Fig. 2. Vertical profiles of temperature, pH and dissolved oxygen in Lake Albano (January 2004–April 2005).
V. Messineo et al. / Toxicon 48 (2006) 160–174166
ARTICLE IN PRESS
MoloHotelBoscoCastello
Centro Sup5 10
1520
25 30
J-01
F M A M
J J A S ON DJ-
02F M
A MJ J S O
N DJ-
03F M
A S O N DJ-
04F M
A MJ J S O N D
J-05
F MA M
0
5
10
15
20
25
30
35
40
45
cell
x 10
6 / L
Surface stations
and depth (m)
Fig. 3. Population dynamics of Planktothrix rubescens in the 5 year study.
Table 3
Assignment of the major peaks observed in the ESI/MS/MS mass spectra of the microcystins considered
Assignment Desmethyl
MC-RR
MC-RR MC-YR Desmethyl
MC-LR
MC-LR MC-LA
[M+2H]2+ 513 520 523 491 498 —
[Ph-CH2CH(OMe)]+ 135 135 135 135 135 —
[M+2H-135]+ — — 911 861 —
[M+2H-135-NH3]+ — 887 — — —
[M+2H-MeOH]2+ — — 507 482 —
[M+H]+ — — — 981 — 910
[M+H-134]+ — — — — 776
[Ala-Leu-MeAsp-Ala-
NH2+2H]+— — — — 402
[Glu-Mdha-Ala-Leu-
MeAsp+H–NH3]+
— — — — 509
V. Messineo et al. / Toxicon 48 (2006) 160–174 167
tests indicated that the bloom of March 2001(coastal station ‘‘Hotel’’) was the most toxic, withEC50 at 5min of 19.7mg of algae.
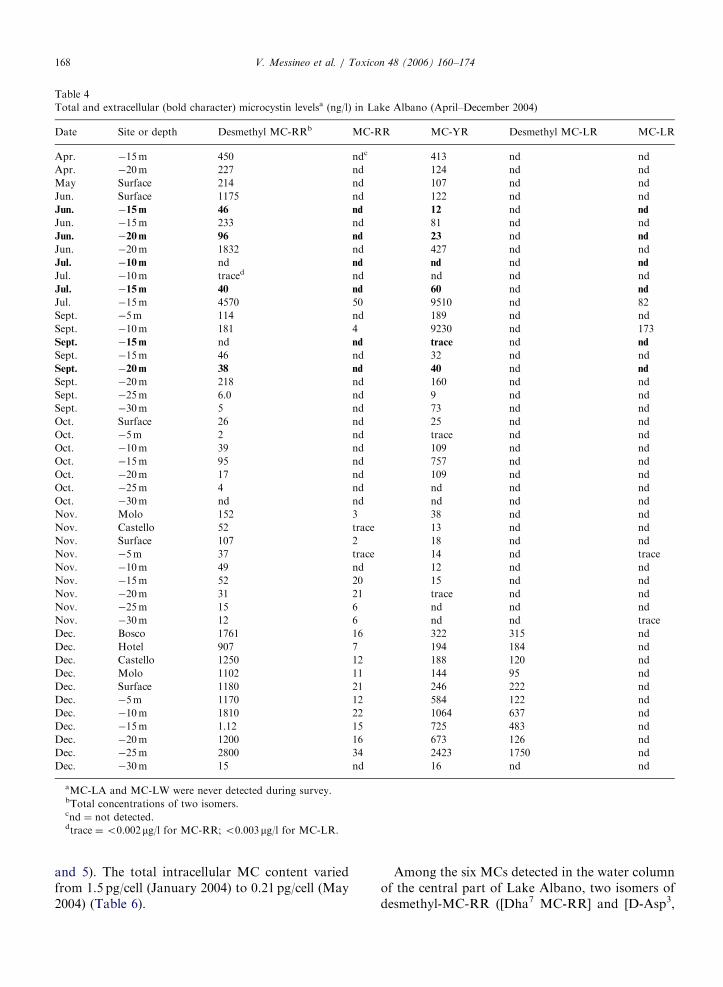
Seventy eight samples were analysed from April2004 to May 2005 for ascertaining the presence ofMCs. Six MCs were detected by LC-MS/MS
analysis (Tables 4 and 5). Structural elucidationsof 5 of them are summarized in Table 3. The totalMC concentration was as high as 14200 ng/l(�15m, July 2004); while the total extracellularMC concentration in 15 analysed samples reached amaximum of 120 ng/l (�5m, March 2005) (Tables 4
ARTICLE IN PRESS
Table 4
Total and extracellular (bold character) microcystin levelsa (ng/l) in Lake Albano (April–December 2004)
Date Site or depth Desmethyl MC-RRb MC-RR MC-YR Desmethyl MC-LR MC-LR
Apr. �15m 450 ndc 413 nd nd
Apr. �20m 227 nd 124 nd nd
May Surface 214 nd 107 nd nd
Jun. Surface 1175 nd 122 nd nd
Jun. �15m 46 nd 12 nd nd
Jun. �15m 233 nd 81 nd nd
Jun. �20m 96 nd 23 nd nd
Jun. �20m 1832 nd 427 nd nd
Jul. �10m nd nd nd nd nd
Jul. �10m traced nd nd nd nd
Jul. �15m 40 nd 60 nd nd
Jul. �15m 4570 50 9510 nd 82
Sept. �5m 114 nd 189 nd nd
Sept. �10m 181 4 9230 nd 173
Sept. �15m nd nd trace nd nd
Sept. �15m 46 nd 32 nd nd
Sept. �20m 38 nd 40 nd nd
Sept. �20m 218 nd 160 nd nd
Sept. �25m 6.0 nd 9 nd nd
Sept. �30m 5 nd 73 nd nd
Oct. Surface 26 nd 25 nd nd
Oct. �5m 2 nd trace nd nd
Oct. �10m 39 nd 109 nd nd
Oct. �15m 95 nd 757 nd nd
Oct. �20m 17 nd 109 nd nd
Oct. �25m 4 nd nd nd nd
Oct. �30m nd nd nd nd nd
Nov. Molo 152 3 38 nd nd
Nov. Castello 52 trace 13 nd nd
Nov. Surface 107 2 18 nd nd
Nov. �5m 37 trace 14 nd trace
Nov. �10m 49 nd 12 nd nd
Nov. �15m 52 20 15 nd nd
Nov. �20m 31 21 trace nd nd
Nov. �25m 15 6 nd nd nd
Nov. �30m 12 6 nd nd trace
Dec. Bosco 1761 16 322 315 nd
Dec. Hotel 907 7 194 184 nd
Dec. Castello 1250 12 188 120 nd
Dec. Molo 1102 11 144 95 nd
Dec. Surface 1180 21 246 222 nd
Dec. �5m 1170 12 584 122 nd
Dec. �10m 1810 22 1064 637 nd
Dec. �15m 1.12 15 725 483 nd
Dec. �20m 1200 16 673 126 nd
Dec. �25m 2800 34 2423 1750 nd
Dec. �30m 15 nd 16 nd nd
aMC-LA and MC-LW were never detected during survey.bTotal concentrations of two isomers.cnd ¼ not detected.dtrace ¼o0.002mg/l for MC-RR; o0.003mg/l for MC-LR.
V. Messineo et al. / Toxicon 48 (2006) 160–174168
and 5). The total intracellular MC content variedfrom 1.5 pg/cell (January 2004) to 0.21 pg/cell (May2004) (Table 6).
Among the six MCs detected in the water columnof the central part of Lake Albano, two isomers ofdesmethyl-MC-RR ([Dha7 MC-RR] and [D-Asp3,
ARTICLE IN PRESS
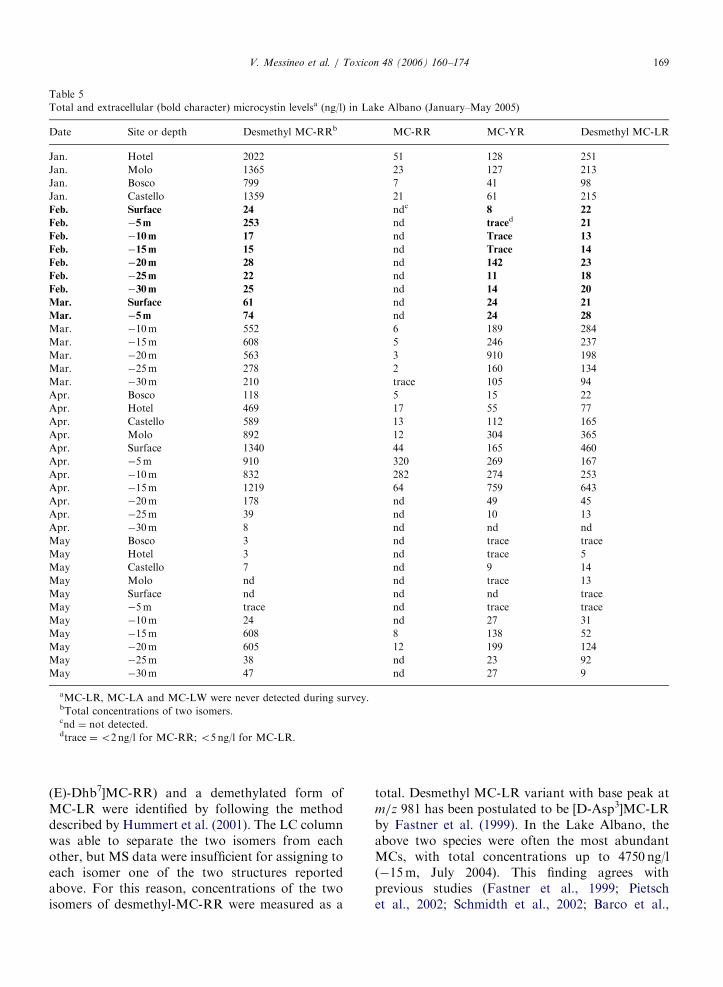
Table 5
Total and extracellular (bold character) microcystin levelsa (ng/l) in Lake Albano (January–May 2005)
Date Site or depth Desmethyl MC-RRb MC-RR MC-YR Desmethyl MC-LR
Jan. Hotel 2022 51 128 251
Jan. Molo 1365 23 127 213
Jan. Bosco 799 7 41 98
Jan. Castello 1359 21 61 215
Feb. Surface 24 ndc 8 22
Feb. �5m 253 nd traced 21
Feb. �10m 17 nd Trace 13
Feb. �15m 15 nd Trace 14
Feb. �20m 28 nd 142 23
Feb. �25m 22 nd 11 18
Feb. �30m 25 nd 14 20
Mar. Surface 61 nd 24 21
Mar. �5m 74 nd 24 28
Mar. �10m 552 6 189 284
Mar. �15m 608 5 246 237
Mar. �20m 563 3 910 198
Mar. �25m 278 2 160 134
Mar. �30m 210 trace 105 94
Apr. Bosco 118 5 15 22
Apr. Hotel 469 17 55 77
Apr. Castello 589 13 112 165
Apr. Molo 892 12 304 365
Apr. Surface 1340 44 165 460
Apr. �5m 910 320 269 167
Apr. �10m 832 282 274 253
Apr. �15m 1219 64 759 643
Apr. �20m 178 nd 49 45
Apr. �25m 39 nd 10 13
Apr. �30m 8 nd nd nd
May Bosco 3 nd trace trace
May Hotel 3 nd trace 5
May Castello 7 nd 9 14
May Molo nd nd trace 13
May Surface nd nd nd trace
May �5m trace nd trace trace
May �10m 24 nd 27 31
May �15m 608 8 138 52
May �20m 605 12 199 124
May �25m 38 nd 23 92
May �30m 47 nd 27 9
aMC-LR, MC-LA and MC-LW were never detected during survey.bTotal concentrations of two isomers.cnd ¼ not detected.dtrace ¼o2 ng/l for MC-RR; o5 ng/l for MC-LR.
V. Messineo et al. / Toxicon 48 (2006) 160–174 169
(E)-Dhb7]MC-RR) and a demethylated form ofMC-LR were identified by following the methoddescribed by Hummert et al. (2001). The LC columnwas able to separate the two isomers from eachother, but MS data were insufficient for assigning toeach isomer one of the two structures reportedabove. For this reason, concentrations of the twoisomers of desmethyl-MC-RR were measured as a
total. Desmethyl MC-LR variant with base peak atm/z 981 has been postulated to be [D-Asp3]MC-LRby Fastner et al. (1999). In the Lake Albano, theabove two species were often the most abundantMCs, with total concentrations up to 4750 ng/l(�15m, July 2004). This finding agrees withprevious studies (Fastner et al., 1999; Pietschet al., 2002; Schmidth et al., 2002; Barco et al.,
ARTICLE IN PRESSV. Messineo et al. / Toxicon 48 (2006) 160–174170
2004) indicating these isomers as the toxic markersof the species P. rubescens. Higher acute toxicitytoward grazers characterizes these compounds, ascompared to MC-LR, MC-YR and MC-RR (Blomet al., 2001). Both desmethyl-MC-RRs are moretoxic than MC-RR (Sano and Kaya, 1998; Sivonenand Jones, 1999). MC-YR and MC-LR were alsodetected in some samples. The total MC concentra-tion in water follows an opposite pattern withrespect to the N/P ratio, and its growth is generallyrelated to the increase of the temperature in thesuperficial layers (Fig. 2) and in the water column.The highest values of MCs were found in spring–midsummer in the water layers under the thermo-cline (20–25m depth, Tables 4 and 5).
The total superficial MC concentration notice-ably increased in June 2004, corresponding to thedecline of the bloom with scum detected in May.Another increase occurred in all the samples inDecember 2004 and January 2005, corresponding to
Table 6
Intracellular microcystin concentrations (mg/g) in three superficial
scums (fresh algae weight)
Compound Scum Jan.
2004
Scum May
2004
Scum Apr.
2005
Desmethyl MC-
RRa
115 90.3 2.9
MC-RR 0.24 0 0.013
MC-YR 2.2 15.3 1.9
Desmethyl MC-
LR
nd nd 0.68
MC-LR nd nd 0.032
MC-LA 0.02 nd
MC-LW nd nd 0.032
Tot. 117.5 105.6 5.6
pg/cell 1.47 0.21 0.8
aSum of concentrations for two isomers.
Table 7
Microcystin levels (ng/l) in two wells (2004–2005)
Compound Albano
Dec. 2004 Jan. 2005
Dem. MC-RR nda nd
MC-RR 4 27
MC-YR nd 26
Dem. MC-LR nd nd
MC-LR nd 14
MC-LA nd nd
MC-LW nd nd
and ¼ not detected.
the beginning of the winter increase in cell popula-tion. A very noticeable increase in MC-YR con-centration occurred in July October 2004, with amaximum in September at a 10m depth, corre-sponding to the unusual subsurface maximum of P.
rubescens (52� 106 cell/l, at �10m) and to thepresence of a colder and poorly oxygenated waterlayer (see above).
During the highest cellular concentrations insurface layers, directly related to the N/P ratioand inversely related to the temperature, theoccurrence of MCs was quite exclusively due tothe intracellular ones (Table 6).
Among MCs found in scums, the two desmethylMC-RR isomers were by far the most abundant.Maximum concentration of desmethyl MC-RRisomers was observed in winter, which is period ofproliferation of P. rubescens.
Two out of 13 wells (Fig. 1) showed the presenceof MCs, i.e. Albano well (60m deep, an indicatorwell 6 in Fig. 1) during all the blooming period(December 2004–March 2005), and Sforza Cesariniwell (100m deep, a well for drinking water uses, well7 in Fig. 1) in March (final period of the blooms)(Table 7).
4. Discussion
Freshwater cyanobacterial toxic blooms are acommon problem in many Italian lakes. Thisphenomenon, which has been widely reported inthe literature during the last 10 years, involves manygenera, including Anabaena (Bruno et al., 1994;Salmaso et al., 1994), Microcystis, Aphanizomenon
(Bruno et al., 1989), Oscillatoria (Bruno et al., 1992)and Planktothrix spp. (Pomati et al., 2000).
The species Planktothrix rubescens and Micro-
cystis aeruginosa are mainly responsible for toxic
Sforza Cesarini
Feb. 2005 Mar. 2005 Mar. 2005
nd nd 17
13 13 nd
nd nd nd
nd nd nd
nd nd nd
nd nd nd
nd nd nd

ARTICLE IN PRESSV. Messineo et al. / Toxicon 48 (2006) 160–174 171
algal blooms in Italy. P. rubescens in Italy is awinter species, with blooms often extending fromlate autumn to mid-spring. Its presence in LakeAlbano is dated back to 1953 (Cannicci, 1954). Thegenus Planktothrix is able to produce gas vesicleswhich can provide buoyancy, enabling these speciesto regulate their position between metalimnic andepilimnic layers (Reynolds et al., 1987). This is theprincipal differentiating feature existing betweenPlanktothrix and Oscillatoria genera (Anagnostidesand Komarek, 1988). Different strains of P.
rubescens, living in deep lakes, evolved strongerforms of gas vesicles in order to withstand highhydrostatic pressures (Bright and Walsby, 1999).Phosphorus, ammonium or nitrate-limited popula-tions move upward if supplied with the lackingnutrients, due to an increased production of gasvesicles (Klemer et al., 1982).
P. rubescens is able to excrete nitrite viaassimilatory reduction, contributing to the forma-tion of nitrite maxima in lakes from absorption ofnitrate but not from ammonia, even if the latter ismore rapidly assimilated (Ohmori, 1978). Organicnitrogen from arginine, glycine, serine, glutamateand aspartate can be used by this species to supportgrowth at low irradiance (Zotina et al., 2003).Evidence from the study of metalimnetic popula-tions during summer showed that they do notmaintain a real steady state, but experience a seriesof transient states (Konopka, 1982).
The general trend of the population dynamics forP. rubescens in Albano lake shows superficialstratification with blooms in winter and metalimnicstratification with a peak (25–30m) in summer, butanyway with an overall population decrease(80–86%) in the late April to early Novemberperiod. These data are consistent with findings ofWalsby et al. (1998) for Lake Zurich and Jacquet etal. (2005) for Lac du Bourget.
In spring 2004 and spring 2005, with frequentcold and rainy weather, the thermal stratification inthe upper layers was sensitive to climatic fluctua-tions and unusual rises of the P. rubescens popula-tions were observed even in April–May, withsuperficial scums.
According to Paerl et al. (2001) the N/P rule isless applicable to highly eutrophic systems whenboth N and P loadings and inputs exceed theassimilative capacity of phytoplankton. However,Teubner et al. (1999) found that in a hypertrophiclake the species P. aghardii took dominance onAphanizomenon flos-aquae when a TN/TP ratio
higher than 16 was reached, then prolonging thegrowth even with a reduced TN/TP rate. In LakeAlbano, P. rubescens growth and moving upwardsoccurred in the winter months with water tempera-ture of 10–11 1C when a N/P ratio exceeded 10, andwith an annual mean N/P ratio equal to 16.
P. rubescens and the related species P. aghardii
are efficient producers of MC-RR and demethylatedMC-RR forms. These filamentous species are noteasy to manage in the drinking water treatments,because of their ability to pass through even the besttreatment systems, in the case of massive input and/or improper maintenance of the plant. In November1989 and September 1994, toxic blooms of P.
aghardii were responsible for severe outbreaks ofacute illness due to contamination of drinking waterin the town of Taalintehaan in Finland, and inScania, Sweden. MC levels varied between 100 and500 ng/l (drinking water in Taalintehtaan), andamounted to 820 ng/l in Lake Vombsjon, Scania(Annadotter et al., 2001).
In addition to MC-RR and two demethylatedMC-RR isomers, the P. rubescens population ofLake Albano is able to produce MC-LR, defined asthe strongest liver carcinogen known (Dow andSwoboda, 2000).
According to Codd, MCs are not released abovetrace amounts into water until cell lysis and death(Codd, 1995). These data agree with the lowamounts of extracellular MCs found in Lake Albanoduring the maximum bloom development(February, March 2005). The bloom, which oc-curred during winter 2004–2005, was of mean celldensity. In the superficial layers (surface and �5m)in June and December 2004, and in January andApril 2005, the MC total content measured by usexceeded the limit of the acute exposure laid down inItaly for bathing activities (840 ng/l). MC contents incollected scums (Table 6) in May and December2004 and April 2005, have to be considered for thetotal toxin evaluation in the risk assessment forrecreational activities in Lake Albano.
The increase of extracellular MC concentrationsgenerally occurs in Lake Albano when environ-mental factors (mainly physical, like water tempera-ture, rain or light levels, rather than chemical, dueto the abundance of primary nutrients in the year)act as a disturbing agent on the P. rubescens
populations, i.e. the lowered water temperaturebetween �15 and �20m in June 2004.
The downward moving of the trichomes towardsthe metalimnic layers increases the losses in cell

ARTICLE IN PRESSV. Messineo et al. / Toxicon 48 (2006) 160–174172
population. The unusual cold-water layer of Septem-ber–October 2004 favoured a strong subsurface cellmaximum in a period, which is normally not optimalfor growth. This is associated with a high productionof MC-YR, which is not the main variant among theP. rubescens toxins. The peculiar production could beexplained by the prevalence of a specific clone in agenetically heterogeneous population, or with achange in the metabolic toxin production, due tothe specific environmental conditions.
The major surface blooms in February withoptimal environmental conditions, show minimumcell loss with quite no toxic release in water.
Noticeable is the strength of the bloom in the year2001, when the mean annual rainfall during the last10 years reached the minimum value (35mm),probably contributing to the increase of nutrientconcentration in groundwater and in the lake. TheMicrotox system was used as a preliminary screen-ing to detect toxicity in the superficial water samplesduring the blooming seasons (winter–spring), givingno false-negative results. Although the V. fisheri
bioassay is sensitive to MCs (Vezie et al., 1996;Bruno et al., 2006, in press), its results did notcorrelate with those ones by LC-MS/MS. For thisreason and due to the non-specific nature of thisbioassay, the Microtox system could be advanta-geously used only as a screening method.
Groundwater can link lakes across the landscape(Dodds, 2002). MCs from contaminated lakes canpercolate in the groundwater towards the pumpingstations for drinking water supply, with presence inthe water proportional to the duration of the toxicblooms (Eynard et al., 2000). In the district ofHaimen, China, MCs were detected in shallow wellwaters used for drinking purposes (Ueno et al.,1996). In our study, we detected MCs in two wellsfor human use inside the volcanic crater (Table 7),one of them (well 7) being the main source ofdrinking water for a neighbouring town. Thepresence of MCs was limited to MC-RR and MC-YR in well 6 (the closest well to the lake)throughout the blooming season, and to twodemethylated MC-RR isomers in well 7, only atthe end of the 2004–2005 blooming season. Thetotal MC content of the latter well was under thelimit for human acute toxicity (840 ng/l) laid downin Italy, but above the limit of 10 ng/l for chronicrisk suggested by Ueno et al. (1996) and Hernandezet al. (2000). Crop irrigation in fields surroundingLake Albano is done using well waters. The MClevels detected in both wells were lower than those
toxic to cultures (McElhiney et al., 2001). However,in case of more toxic blooms the ability of thesetoxins to reach groundwater could lead to higherconcentrations throughout the blooming season inwells surrounding the lake.
Due to the risk of human chronic exposure toMC—containing drinking water, measures shouldbe introduced to equip the wells of the volcaniccrater with suitable devices in order to prevent thetoxin passage in drinking water supply during theblooming period.
5. Conclusions
The evidence of this study shows a shift of LakeAlbano from an oligotrophic condition (Stella andSocciarelli, 1950) to a state of mesotrophy tendingto eutrophy (mean chlorophyll a values rangingbetween 12 mg/l in 2001 and 11 mg/l in 2004), causedby the increasing levels of N and P from manuretreatment and sewage inputs in the groundwaterfeeding the lake and in the lake water itself.Contaminant concentration resulting from theprogressive decrease of the water lake level, whichoccurred in decades of uncontrolled suckling ingroundwaters, presumably helped this process.
MCs produced by the toxic species P. rubescens isa permanent situation throughout the year in avaluable and scenic lake near Rome. MCs arepresent with several variants, in concentrationsoften exceeding the tolerance level enacted in Italyand preventing human use.
In conclusion, the findings of the present study pointout the need for more extensive investigations on thepathogenic role and environmental route of this newand important class of risk substances in Italy.
References
Anagnostides, K., Komarek, J., 1988. Modern approaches to the
classification system of cyanophytes. 3. Oscillatoriales. Arch.
Hydrobiol. 80, 327–472.
Annadotter, H., Cronberg, G., Lawton, L., Hansson, H.B.,
Gothe, U., Skulberg, O., 2001. An extensive outbreak of
gastroenteritis associated with the toxic cyanobacterium
Planktothrix agardhii (Oscillatoriales, Cyanophyceae) in
Scania, South Sweden. In: Chorus, I. (Ed.), Cyanotoxins.
Occurrences, Causes, Consequences. Springer, Berlin,
pp. 200–208.
Barco, M., Flores, C., Rivera, J., Caixach, J., 2004. Determina-
tion of microcystin variants and related peptides present in a
water bloom of Planktothrix (Oscillatoria) rubescens in a
Spanish drinking reservoir by LC/ESI-MS. Toxicon 44,
881–886.

ARTICLE IN PRESSV. Messineo et al. / Toxicon 48 (2006) 160–174 173
Blom, J.F., Robinson, J.A., Juttner, F., 2001. High grazer
toxicity of [D-Asp3, (E)-Dhb7] microcystin-RR of P. rubes-
cens as compared to different microcystins. Toxicon 39,
1923–1932.
Bright, D.I., Walsby, A.E., 1999. The relationship between
critical pressure and width of gas vesicles in isolates of
Planktothrix rubescens from Lake Zurich. Microbiology 145,
2769–2775.
Bruno, M., Barbini, D.A., Pierdominici, E., Serse, A.P., Ioppolo,
A., 1994. Anatoxin-a and a previously unknown toxin in
Anabaena planctonica from blooms found in lake Mulargia
(Italy). Toxicon 32, 369–373.
Bruno, M., Gucci, P.M.B., Pierdominici, E., Sestili, P., Ioppolo,
A., Sechi, N., Volterra, L., 1992. Microcystin-like toxins in
different freshwater species of Oscillatoria. Toxicon 30,
1307–1311.
Bruno, M., Gucci, P.M.B., Volterra, L., 1989. Fioriture algali:
rilevabilitadella presenza di biotossine. Ambiente, Risorse,
Salute 91, 6–7.
Bruno, M., Fiori, M., Mattei, D., Melchiorre, S., Messineo, V.,
Volpi, F., Bogialli, S., Nazzari, M., 2006. ELISA and LC-MS/
MS methods for determining cyanobacterial toxins in blue-
green algae food supplements. Nat. Prod. Res. 20 (9),
827–834.
Cannicci, G., 1954. Su una eccezionale fioritura del lago di
Albano. Boll. Pesca, Piscico. Idrobiol. 24, 221–233.
Chondrogianni, C., Ariztegui, D., Guilizzoni, P., Lami, A., 1996.
Lakes Albano and Nemi (central Italy): an overview. In:
Guilizzoni, P., Oldfield, F. (Eds.), Palaeoenvironmental
Analysis of Italian Crater Lakes and Adriatic Sediments
(PALICLAS). Mem. Ist. ital. Idrobiol. 55, 17–22.
Codd, G.A., 1995. Cyanobacterial toxins: occurrence, properties
and biological significance. Water Sci. Technol. 32, 149–156.
Di Corcia, A., Marchese, S., Samperi, R., 1994. Determination of
phenol pollutants in water at trace level: extraction by a
reversibile graphitized carbon black cartridge. J. AOAC Int.
77, 446.
Dietrich, D., Hoeger, S., 2005. Guidance values for microcystins
in water and cyanobacterial supplement products (blue-green
algal supplements): a reasonable or misguided approach?
Toxicol. Appl. Pharmacol. 203, 273–289.
Dodds, W.K., 2002. Freshwater Ecology. Concepts and Envir-
onmental Applications. Academic Press, San Diego, USA.
Dow, C.S., Swoboda, U.K., 2000. Cyanotoxins. In: Whitton,
B.A., Potts, M. (Eds.), The Ecology of Cyanobacteria. Their
diversity in Time and Space. Kluwer Academic, New York.
Eynard, F., Mez, K., Walther, J.L., 2000. Risk of cyanobacterial
toxins in Riga waters (Latvia). Water Res. 34, 2979–2988.
Falconer, I.R., Buckley, T.H., 1989. Tumor promotion by
Microcystis sp., a blue-green alga occurring in water supplies.
Med. J. Aust. 150, 351.
Falconer, I.R., Beresford, A., Runnegar, M.T.C., 1983. Evidence
of liver damage in human populations exposed to toxin from
a bloom of the blue-green alga M. aeruginosa in a drinking
water supply reservoir. Med. J. Aust. 1, 511–515.
Fastner, J., Erhard, M., Carmichael, W.W., Sun, F., Rinehart,
K.L., Ronicke, H., Chorus, I., 1999. Characterization and
diversity of microcystins in natural blooms and strains of the
genera Microcystis and Planktothrix from German fresh-
waters. Arch. Hydrobiol. 145 (2), 147–163.
Ferree, M.A., Shannon, R.D., 2001. Evaluation of a second
derivative UV/Visible spectroscopy technique for nitrate and
total nitrogen analysis of wastewater samples. Water Res. 35,
327–332.
Fitzgeorge, R.B., Clark, I.A., Keevil, C.W., 1994. Routes of
intoxication. In: Codd, G.A., Jefferies, T.M., Keevil, C.W.,
Potter, E. (Eds.), Detection Methods for Cyanobacterial
Toxins. The Royal Society of Chemistry, Cambridge, UK, pp.
69–74.
Fleming, L.E., Rivero, C., Burns, J., Williama, C., Bean, J.A.,
Shea, K.A., Stinn, J., 2002. Blue green algae (cyanobacterial)
toxins, surface drinking water, and liver cancer in Florida.
Harmful Algae 1, 57–168.
Ford, S.L., Abayasekara, D.R.E., Persaud, S.J., Jones, P.M.,
1996. Role of phosphoprotein phosphatases in the corpus
luteum: I Identification and characterization of serine/
threonine phosphoprotein phosphatases in isolated rat luteal
cells. J. Endocrinol. 150, 205–211.
Gjolme, N., Utkilen, H., 1996. The extraction and stability of
microcystin-RR in different solvents. Phycology 35, 80–82.
Harada, K.I., Teuji, K., 1998. Persistence and decomposition of
hepatotoxic microcystins produced by cyanobacteria in
natural environment. J. Toxicol. 17, 385–403.
Hernandez, M., Macia, M., Padilla, C., Del Campo, F., 2000.
Modulation of human polymorphonuclear leukocyte adher-
ence by cyanopeptide toxins. Environ. Res. Sect. A 84, 64–68.
Hummert, C., Dahlmann, J., Reinhardt, K., Dang, H.Ph.H.,
Dang, D.K., Luckas, B., 2001. Liquid chromatography-Mass
spectrometry identification of microcystins in Microcystis
aeruginosa strain from lake Thanh Cong, Hhanoi, Vietnam.
Chromatographia 54, 569.
Humpage, A.R., Hardy, S.J., Moore, E.J., Froscio, S.M.,
Falconer, I.R., 2000. Microcystins (cyanobacterial toxins) in
drinking water enhance the growth of abberrant crypt foci in
the mouse colon. J. Toxicol. Environ. Health Part A. 61,
155–165.
Ito, E., Kondo, F., Terao, K., Harada, K.I., 1997. Neoplastic
nodular formation in mouse liver induced by repeated intraper-
itoneal injections of microcystin-LR. Toxicon 35, 1453–1457.
Jacquet, S., Briand, J.F., Leboulanger, C., Avois-Jacquet, C.,
Oberhaus, L., Tassin, B., Vincon-Leite, B., Paolini, G.,
Druart, J.C., Anneville, O., Humbert, J.F., 2005. The
proliferation of the toxic cyanobacterium Planktothrix
rubescens following restoration of the largest natural French
lake (Lac du Bourget). Harmful Algae 4, 651–672.
Jochimsen, E.M., Carmichael, W.W., An, J., Cardo, D.M.,
Cookson, S.T., Holmes, C.E.M., Antunes, B.C., Liho, D.,
Lyra, T.M., Spinelli, T., Barreto, V., Azevedo, S.M.F.O.,
Jarvis, W.R., 1998. Liver failure and death after exposure to
microcystins at a hemodialysis center in Brazil. New Engl. J.
Med. 339, 873–878.
Klemer, A.R., Feuillade, J., Feuillade, M., 1982. Cyanobacterial
blooms: carbon and nitrogen limitation have opposite effects
on the buoyancy of Oscillatoria. Science 215, 1629–1631.
Konopka, A., 1982. Physiological ecology of a metalimnetic
Oscillatoria rubescens population. Limnol. Oceanogr. 27,
1154–1161.
Lambert, D., Maher, W., 1995. An evaluation of the efficiency of
the alkaline persulphate digestion method for the determina-
tion of total phosphorus in turbid waters. Water Res. 29, 7–9.
Lankoff, A., Carmichael, W.W., Grasman, K.A., Yuan, M.,
2004. The uptake kinetics and immunotoxic effects of
microcystin-LR in human and chicken peripheral blood
lymphocytes in vitro. Toxicology 204, 23–40.

ARTICLE IN PRESSV. Messineo et al. / Toxicon 48 (2006) 160–174174
Leiers, T., Bihlmayer, A., Ammon, H.P.T., Wahl, M.A., 2000.
½Ca2þ�i- and insulin-stimulating effect of the non-membrane-
permeable phosphatase-inhibitor microcystin-LR in intact
insulin-secreting cells (RINm5F). Brit. J. Pharmacol. 130,
1406–1410.
Lund, J.W.G., Kipling, C., Le Cren, E., 1958. The inverted
microscope method of estimating algal numbers and the
statistical basis of estimations by counting. Hydrobiology 11,
143–170.
McElhiney, J., Lawton, L.A., Leifert, C., 2001. Investigations
into the inhibitory effects of microcystins on plant growth,
and the toxicity of plant tissues following exposure. Toxicon
39, 1411–1420.
McFeters, G.A., Bond, P.J., Olson, S.B., Tchan, Y.T., 1983. A
comparison of microbial bioassays for the detection of
aquatic toxicants. Water Res. 17, 1757–1762.
Mindrup, R., Shirey, R., 2000. Identifying odors. Today’s
Chemist at Work, July, 30–33.
Meriluoto, J., Eriksson, J., 1988. Rapid analysis of peptide toxins
in Cyanobacteria. J. Chromatogr. 438, 93–99.
Murphy, J., Riley, J.P., 1962. A modified single solution method
for the determination of phosphate in natural waters. Anal.
Chim. Acta 27, 31–36.
Nishiwaki-Matsushima, R., Ohta, T., Nishiwaki, S., Suganuma,
M., Kohyama, K., Ishikawa, T., Carmichael, W.W., Fujiki,
H., 1992. Liver tumor promotion by the cyanobacterial cyclic
peptide toxin microcystin-LR. J. Cancer Res. Clin. Oncol.
118, 420–424.
Ohmori, M., 1978. Nitrite excretion by a blue-green alga,
Oscillatoria rubescens D.C. Arch. Hydrobiol. 83, 485–493.
Paerl, H.W., Fulton, R.S., Moisander, P.H., Dyble, J., 2001.
Harmful freshwater algal blooms, with an emphasis on
cyanobacteria. Sci. World 1, 76–113.
Pietsch, J., Bornmann, K., Schmidt, W., 2002. Relevance of
intra–and extracellular cyanotoxins for drinking water treat-
ment. Acta Hydrochim. Hydrobiol. 30, 7–15.
Pomati, F., Sacchi, S., Rossetti, C., Giovannardi, S., 2000. The
freshwater cyanobacterium Planktothrix sp. FPl: molecular
identification and detection of paralytic shellfish poisoning
toxins. Phycol. 36, 553–562.
Reynolds, C.S., Oliver, R.L., Walsby, A.E., 1987. Cyanobacterial
dominance: the role of buoyancy regulation in dynamic lake
environments. N. Z. J. Mar. Freshwater Res. 21, 379–390.
Rojas, M., Nunez, M.T., Zambrano, F., 1990. Inhibitory effect of
a toxic peptide isolated from a waterbloom of Microcystis sp.
(Cyanobacteria) on iron uptake by rabbit reticulocytes.
Toxicon 28, 1325–1332.
Salmaso, N., 2000. Factors affecting the seasonality and
distribution of cyanobacteria and chlorophytes: a case study
from the large lakes south of the Alps, with special reference
to Lake Garda. Hydrobiology 438, 43–63.
Salmaso, N., Cavolo, F., Cordella, P., 1994. Fioriture di
Anabaena e Microcystis nel Lago di Garda. Eventi rilevati e
caratterizzazione dei periodic di sviluppo. Acqua Aria 1,
17–28.
Sayed, S.B., Whitehouse, B.J., Jones, P.M., 1997. Phosphoserine/
threonine phosphatases in the rat adrenal cortex: a role in the
control of steroidogenesis? J. Endocrinol. 154, 449–458.
Sano, T., Kaya, K., 1998. Two new (E)-2-amino-2-butenoic acid
(Dhb)-containing microcystins isolated from Oscillatoria
aghardii. Tetrahedron 54, 463–470.
Schmidth, W., Willmitzer, H., Bornmann, K., Pietsch, J., 2002.
Production of drinking water from raw water containing
cyanobacteria-pilot plant studies for assessing the risk of
microcystin breakthrough. Environ. Toxicol. 17, 375–385.
Sivonen, K., Jones, G., 1999. Cyanobacterial toxins. In: Chorus,
I., Bartram, J. (Eds.), Toxic Cyanobacteria in Water: a Guide
to their public Health Consequences, Monitoring and
Management. World Health Organization, E.&F.N. Spon,
Geneva, pp. 41–111.
Solter, P.H., Wollenberg, G.K., Huang, X., Chu, F.S., Runnegar,
M., 1998. Prolonged sublethal exposure to the protein
phosphatase inhibitor microcystin-LR results in multiple
dose-dependent hepatotoxic effects. Toxicol. Sci. 44, 87–96.
Stein, J., 1973. Culture methods and growth measurements.
Handbook of Phycological Methods. Cambridge University
Press, London and New York.
Stella, E., Socciarelli, M., 1950. Il lago di Albano. Parte I: La
fisiografia e le societa pelagiche. Boll. pesca, pisci. idrobiol. 4,
154–173.
Teixeira da Gloria Lima Crux, M., Da Conceicao Nascimento
Costa, M., Lucia Pires de Carvalho, V., Dos Santos Pereira,
M., Hage, E., 1993. Gastroenteritis epidemic in the area of the
Itaparica dam—Bahia Brazil. Bull. Pan Am. Health Organ.
27, 244–253.
Teubner, K., Feyerabend, M., Henning, M., Nicklisch, A.,
Woitke, P., Kohl, J.G., 1999. Alternative blooming of
Aphanizomenon flos-aquae or P. agardhii induced by the
timing of the critical nitrogen: phosphorus ratio in hyper-
trophic riverine lakes. Arch. Hydrobiol. Spec. Issues Adv.
Limnol. 54, 325–344.
Ueno, Y., Nagata, S., Tsutsumi, T., Hasegawa, A., Watanabe,
M.F., Park, H.D., Chen, G.C., Chen, G., Yu, S.Z., 1996.
Detection of microcystins, a blue-green algal hepatotoxin, in
drinking water sampled in Haimen and Fusui, endemic areas
of primary liver cancer in China, by highly sensitive
immunoassay. Carcinog. 17, 1317–1321.
US Environmental Protection Agency, 1979. Microtox Applica-
tion Notes: M 103–116. Microtox Operation Book, Microbics
Inc.
Utermohl, H., 1931. Neue Wege in der quantitativen Earfassung
des Planktons (mit besonderer Berucksichtigung des Ultra-
planktons). Verh. int. Ver. theor. angew. Limnol. 5, 567–596.
Van Breemen, L.W.C.A., Dits, J.S., Ketelaars, H.A.M., 1992.
Production and reduction of geosmin and 2-methylisoborneol
during storage of river water in deep reservoirs. Water Sci.
Tech. 25, 224–233.
Vezie, C., Benoufella, F., Sivonen, K., Bertru, G., Laplanche, A.,
1996. Detection of toxicity of cyanobacterial strains using
Artemia salina and Microtox assays compared with mouse
bioassay results. Phycologia 35, 198–202.
Walsby, A.E., Avery, A., Schanz, F., 1998. The critical pressure
of gas vesicles in Planktothrix rubescens in relation to the dept
of winter mixing in Lake Zurich, Switzerland. J. Plank. Res.
20, 1357–1375.
Zhou, L., Yu, H., Chen, K., 2002. Relationship between
microcystin in drinking water and colorectal cancer. Biomed.
Env. Sci. 15, 166–171.
Zotina, T., Koster, O., Juttner, F., 2003. Photoeterotrophy and
light-dependent uptake of organic and organic nitrogen
compounds by Planktothrix rubescens under low irradiance.
Freshwater Biol. 48, 1859–1872.
![Histopathological effects of [D-Leu 1]Microcystin-LR variants on liver, skeletal muscle and intestinal tract of Hypophthalmichthys molitrix (Valenciennes, 1844](https://static.fdokumen.com/doc/165x107/631cac57a1cc32504f0c98d9/histopathological-effects-of-d-leu-1microcystin-lr-variants-on-liver-skeletal.jpg)



















