
Co-existence of Echinococcus granulosus infection and cancer metastasis in the liver correlates with...
7
Co-existence of Echinococcus granulosus infection and cancer metastasis in the liver correlates with reduced Th1 immune responses N. TURHAN, 1, * G. ESENDAGLI, 2, * O. OZKAYAR, 3 G. TUNALI, 2 C. SOKMENSUER 3 & O. ABBASOGLU 1 1 Department of General Surgery, Medical Faculty, Hacettepe University, Ankara, Turkey, 2 Department of Basic Oncology, Hacettepe University Cancer Institute, Ankara, Turkey, 3 Department of Pathology, Medical Faculty, Hacettepe University, Ankara, Turkey SUMMARY A possible relationship between cancer and Echinococcus granulosus infection has been postulated. As T cells are crit- ical players in immune responses against both infections and malignancies, in an experimental model of secondary echino- coccosis and breast cancer, this study aims to observe the progression of cancer and to determine the characters of T- cell responses. 4T1 breast tumour cells were subcutaneously injected into mammary region, whereas protoscoleces were intraperitoneally inoculated into the mice. Hydatid cysts, tumours and metastases were determined with macroscopic and histopathological evaluation. T cells found in spleen, liver and tumour were characterised by flow cytometric analysis of CD3, CD4, CD8, CD25, CCR5, CCR3, IL-4 and IFN-c. In the mice inoculated both with protoscoleces and with breast tumour cells, increased frequency of cancer metastasis was observed in the liver. The amount of CD4 + T cells was increased in the liver and in the spleen of mice infected with E. granulosus. However, co-existence of echi- nococcosis and metastatic lesions in the liver was associated with significant reduction in the IFN-c + and CCR5 + Th1 cells and increase in the CD25 + T cells. Our results may indicate an immunological link between cystic echinococcosis and cancer that allows tumour metastasis to flourish in the liver. Keywords breast cancer, Echinococcus , hydatid cyst, parasite INTRODUCTION The character and strength of immune responses within infected organs direct the elimination of parasites or mediate the reactions limiting the spread of infection (1, 2). In early cystic echinococcosis, immune reactions are dominated by type 2 helper T (Th2) cells and eosinoph- ils, whereas immunity induced by Th1-associated cytokines (e.g. IFN-c) can lower the parasitic load (1, 3–5). It has been evidenced that individuals (or animal strains) that can exhibit strong Th1 responses can be resistant to Echinococcus (i.e. Echinococcus multilocularis) infections (1). However, especially in the chronic phases of hydatid disease, both Th1 and Th2 products remain highly expressed (4, 6). In addition, the establishment of hydatid cysts is accompanied by increased numbers of immune suppressive cells such as regulatory T (Treg) cells and myeloid-derived suppressor cells (MDSCs) (6, 7). To survive the immune reactions, cancer cells may employ strategies similar to that of used by infectious organisms, especially parasites and viruses (5). Immune reactions mediated by Th2 or by Treg cells are commonly observed in the tumours (8, 9). Th1-mediated inflamma- tory responses that are required for the elimination of transformed cells are generally impaired in the tumour microenvironment (8, 9). Thus, immune suppression is implicated in tumourigenesis, tumour invasion, metastasis and poor prognosis (5, 8, 9). Co-existence of cancers (e.g. hepatocellular carcinoma, myeloid leukaemia, ovarian tumours and gastric carci- noma) and hydatid disease has been reported by several groups (10–12). Correspondingly, immune deficiencies and immunosuppressive therapies may favour the recur- rence of echinococcosis (13–15). Alternatively, it has been postulated that Echinococcus granulosus infection may have protective effect against cancer development Correspondence: Gunes Esendagli, Department of Basic Oncology, Hacettepe University Cancer Institute, 06100 Sihhiye, Ankara, Tur- key (e-mail: [email protected]). *These authors contributed equally to the study. Received: 15 April 2014 Accepted for publication: 12 October 2014 Parasite Immunology, 2015, 37, 16–22 DOI: 10.1111/pim.12152 © 2014 John Wiley & Sons Ltd 16
Transcript of Co-existence of Echinococcus granulosus infection and cancer metastasis in the liver correlates with...

Co-existence of Echinococcus granulosus infection and cancer
metastasis in the liver correlates with reduced Th1 immune
responses
N. TURHAN,1,* G. ESENDAGLI,2,* O. OZKAYAR,3 G. TUNALI,2 C. SOKMENSUER3 & O. ABBASOGLU1
1Department of General Surgery, Medical Faculty, Hacettepe University, Ankara, Turkey, 2Department of Basic Oncology, HacettepeUniversity Cancer Institute, Ankara, Turkey, 3Department of Pathology, Medical Faculty, Hacettepe University, Ankara, Turkey
SUMMARY
A possible relationship between cancer and Echinococcusgranulosus infection has been postulated. As T cells are crit-ical players in immune responses against both infections andmalignancies, in an experimental model of secondary echino-coccosis and breast cancer, this study aims to observe theprogression of cancer and to determine the characters of T-cell responses. 4T1 breast tumour cells were subcutaneouslyinjected into mammary region, whereas protoscoleces wereintraperitoneally inoculated into the mice. Hydatid cysts,tumours and metastases were determined with macroscopicand histopathological evaluation. T cells found in spleen,liver and tumour were characterised by flow cytometricanalysis of CD3, CD4, CD8, CD25, CCR5, CCR3, IL-4and IFN-c. In the mice inoculated both with protoscolecesand with breast tumour cells, increased frequency of cancermetastasis was observed in the liver. The amount of CD4+ Tcells was increased in the liver and in the spleen of miceinfected with E. granulosus. However, co-existence of echi-nococcosis and metastatic lesions in the liver was associatedwith significant reduction in the IFN-c+ and CCR5+ Th1cells and increase in the CD25+ T cells. Our results mayindicate an immunological link between cystic echinococcosisand cancer that allows tumour metastasis to flourish in theliver.
Keywords breast cancer, Echinococcus, hydatid cyst,parasite
INTRODUCTION
The character and strength of immune responses withininfected organs direct the elimination of parasites ormediate the reactions limiting the spread of infection (1,2). In early cystic echinococcosis, immune reactions aredominated by type 2 helper T (Th2) cells and eosinoph-ils, whereas immunity induced by Th1-associatedcytokines (e.g. IFN-c) can lower the parasitic load (1,3–5). It has been evidenced that individuals (or animalstrains) that can exhibit strong Th1 responses can beresistant to Echinococcus (i.e. Echinococcus multilocularis)infections (1). However, especially in the chronic phasesof hydatid disease, both Th1 and Th2 products remainhighly expressed (4, 6). In addition, the establishment ofhydatid cysts is accompanied by increased numbers ofimmune suppressive cells such as regulatory T (Treg)cells and myeloid-derived suppressor cells (MDSCs) (6,7).To survive the immune reactions, cancer cells may
employ strategies similar to that of used by infectiousorganisms, especially parasites and viruses (5). Immunereactions mediated by Th2 or by Treg cells are commonlyobserved in the tumours (8, 9). Th1-mediated inflamma-tory responses that are required for the elimination oftransformed cells are generally impaired in the tumourmicroenvironment (8, 9). Thus, immune suppression isimplicated in tumourigenesis, tumour invasion, metastasisand poor prognosis (5, 8, 9).Co-existence of cancers (e.g. hepatocellular carcinoma,
myeloid leukaemia, ovarian tumours and gastric carci-noma) and hydatid disease has been reported by severalgroups (10–12). Correspondingly, immune deficienciesand immunosuppressive therapies may favour the recur-rence of echinococcosis (13–15). Alternatively, it hasbeen postulated that Echinococcus granulosus infectionmay have protective effect against cancer development
Correspondence: Gunes Esendagli, Department of Basic Oncology,Hacettepe University Cancer Institute, 06100 Sihhiye, Ankara, Tur-key (e-mail: [email protected]).*These authors contributed equally to the study.Received: 15 April 2014Accepted for publication: 12 October 2014
Parasite Immunology, 2015, 37, 16–22 DOI: 10.1111/pim.12152
© 2014 John Wiley & Sons Ltd16

(16). This assumption relies on the observationsregarding the antigenic similarities between E. granulosusand certain types of cancers and on the rarity of inci-dental liver tumours in the patients operated for hydatidcysts (16–18).Here, in an experimental cystic echinococcosis and
breast cancer model, the traits of immune modulationrelated to the secondary E. granulosus infection and thecancer metastasis were investigated. In the miceinoculated both with protoscoleces and with breasttumour cells, increased frequency of cancer metastasiswas observed in the liver. Accordingly, the co-existenceof E. granulosus infection and cancer metastases wasassociated with significant reduction in the amount ofIFN-c+ and CCR5+ Th1 cells and increase in CD25+ Tcells.
MATERIALS AND METHODS
Animals and cell culture
Five-week-old inbred female BALB/c mice (Kobay As.,Ankara, Turkey) were housed under environmentally con-trolled standard conditions. The experimental procedureswere approved by the Institutional Animal Care and UseCommittee of Hacettepe University, Ankara, Turkey(approval number: 2011/37-2).4T1 breast cancer cell line was purchased from the
American Type Culture Collection (ATCC; LGC Promo-chem, Rockville, MD, USA). The cells were cultured inappropriate media and under standard conditions asinstructed by the provider.
Isolation of Echinococcus granulosus protoscoleces
Fertile hydatid cysts were obtained from the liver of asheep (Ovis aries) infected with E. granulosus. The liverwas freshly recovered from an abattoir, and its surface wasdisinfected with 70% isopropyl alcohol and iodine solu-tion. Cyst fluid and protoscoleces were collected into atube containing 19 phosphate-buffered saline (PBS) solu-tion supplemented with 1% penicillin and streptomycin.Following the washing steps, the viability of protoscoleceswas analysed with trypan blue exclusion test and countedunder a light microscope.
Experimental model
Four groups of BALB/c mice were inoculated on day 0and 91 of the experiment according to the followingprotocol: one group of mice (n = 10) (E. granulosusinfection and breast cancer group) was first (on day 0)
intraperitoneally (i.p.) inoculated with a suspension of3000 viable protoscoleces in physiological saline (0�09%NaCl solution) (500 lL/mouse). Then, on day 91, themice were subcutaneously (s.c.) inoculated with 4T1 cells(5 9 104 cells/100 lL) into right-inguinal mammary fatpad. A second group of mice (n = 6) (breast cancergroup) was first i.p. inoculated with 500 lL of physiolog-ical saline/mouse. Then, the mice were s.c. inoculatedwith 4T1 cells (5 9 104 cells/100 lL) into right-inguinalmammary fat pad. A third group of mice (n = 8)(E. granulosus infection group) was first intraperitoneally(i.p.) inoculated with a suspension of 3000 viable protos-coleces in physiological saline (500 lL/mouse). Then, themice were s.c. inoculated with 100 lL physiological sal-ine into right-inguinal mammary fat pad. Finally, afourth group of mice (n = 5) (control group) was firsti.p. injected with 500 lL physiological saline/mouse, andthen, the mice received 100 lL of physiological salines.c. into right-inguinal mammary fat pad. On day 105,all of the mice were sacrificed.
Histopathological evaluation
The tumour, lung and liver samples were fixed in 10%formalin and then embedded in paraffin. Histopathologi-cal evaluation of the paraffin sections was performedunder conventional light microscopy after haematoxylin–eosin staining. Areas of metastasis were determinedwithin the 4-lm serial sections cut across the liver orthe lung.
Flow cytometry
The spleen, liver and tumour samples were minced, sus-pended in 19 PBS and passed through a mesh with47 lm pore size. The cells were washed and were sepa-rated by Biocoll (Biochrom, Berlin, Germany) densitygradient centrifugation. For the immunophenotypic analy-ses, cells were labelled with anti-mouse CD3 (17A2)(R&D Systems, Minneapolis, MN, USA); anti-mouseCCR3 (83103), anti-mouse CD25 (PC61.5), anti-mouseCD4 (GK1.5), anti-mouse CD8 (53-6.7), anti-mouse IL-4(11B11), anti-mouse IFN-c (XMG1.2) (eBioscience, SanDiego, CA, USA); anti-mouse CCR5 (HM-CCR5 (7A4))(Becton Dickinson, San Jose, CA, USA) monoclonalantibodies. Intracellular staining of IFN-c and IL-4 wasperformed with intracellular staining kit (Cytofix/Cyto-permTM Plus; Becton Dickinson) according to the manu-facturer’s recommendations. The percentage of positivecells was calculated by comparison with the appropriateisotype-matched antibody controls. The flow cytometrygating strategy commonly used in this study is shown in
Volume 37, Number 1, January 2015 E. granulosus and cancer metastasis
© 2014 John Wiley & Sons Ltd, Parasite Immunology, 37, 16–22 17
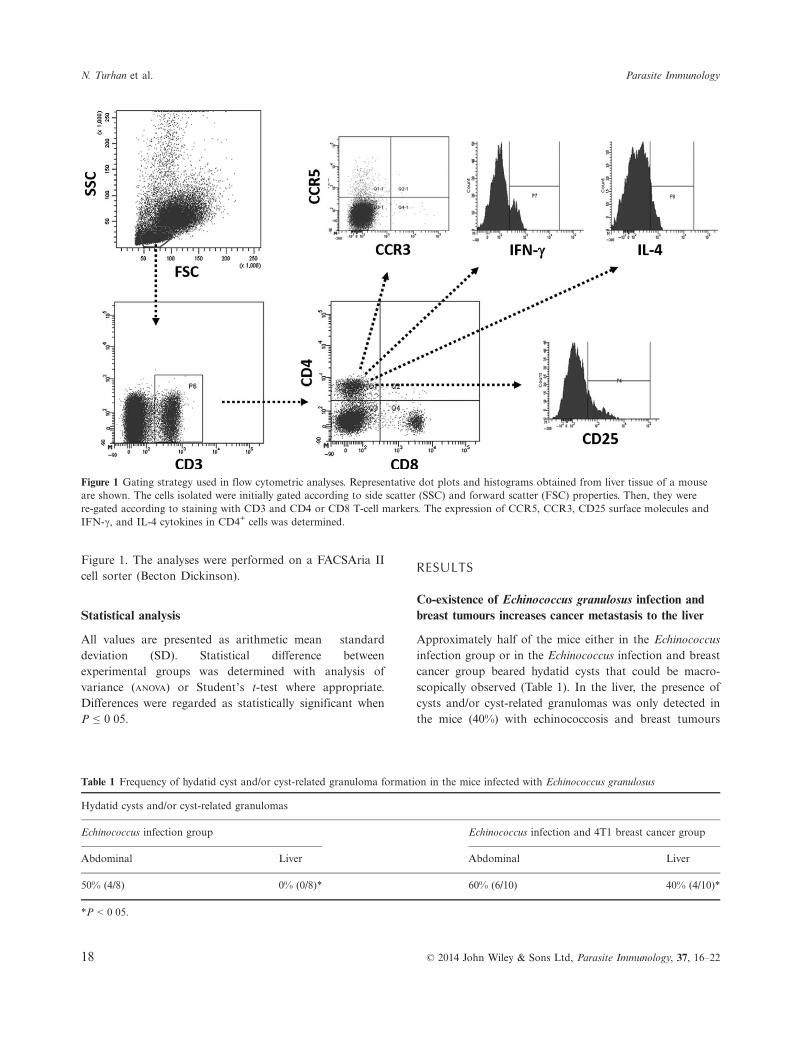
Figure 1. The analyses were performed on a FACSAria IIcell sorter (Becton Dickinson).
Statistical analysis
All values are presented as arithmetic mean � standarddeviation (SD). Statistical difference betweenexperimental groups was determined with analysis ofvariance (ANOVA) or Student’s t-test where appropriate.Differences were regarded as statistically significant whenP ≤ 0�05.
RESULTS
Co-existence of Echinococcus granulosus infection andbreast tumours increases cancer metastasis to the liver
Approximately half of the mice either in the Echinococcusinfection group or in the Echinococcus infection and breastcancer group beared hydatid cysts that could be macro-scopically observed (Table 1). In the liver, the presence ofcysts and/or cyst-related granulomas was only detected inthe mice (40%) with echinococcosis and breast tumours
Figure 1 Gating strategy used in flow cytometric analyses. Representative dot plots and histograms obtained from liver tissue of a mouseare shown. The cells isolated were initially gated according to side scatter (SSC) and forward scatter (FSC) properties. Then, they werere-gated according to staining with CD3 and CD4 or CD8 T-cell markers. The expression of CCR5, CCR3, CD25 surface molecules andIFN-c, and IL-4 cytokines in CD4+ cells was determined.
Table 1 Frequency of hydatid cyst and/or cyst-related granuloma formation in the mice infected with Echinococcus granulosus
Hydatid cysts and/or cyst-related granulomas
Echinococcus infection group Echinococcus infection and 4T1 breast cancer group
Abdominal Liver Abdominal Liver
50% (4/8) 0% (0/8)* 60% (6/10) 40% (4/10)*
*P < 0�05.
N. Turhan et al. Parasite Immunology
18 © 2014 John Wiley & Sons Ltd, Parasite Immunology, 37, 16–22

(Table 1). These hepatic hydatid cysts were also confirmedwith histopathological analysis (data not shown).All mice inoculated with 4T1 breast cancer cells devel-
oped tumours. There was no significant difference in thesize and the growth rate of tumours between the breastcancer group and the Echinococcus infection and breastcancer group. The lung was primary target for metastasisin both of the groups. However, in the Echinococcus infec-tion and breast cancer group, the liver was more fre-quently invaded by the tumour cells compared to that ofthe breast cancer group (60% vs. 33�3%, P < 0�05)(Table 2). Collectively, the presence of cystic echinococco-sis prior to tumour cells’ inoculation increased cancermetastasis to the liver.
The percentage of CD4+ T cells was increased in theliver of tumour-bearing mice infected with Echinococcusgranulosus
In the spleen, percentages of both CD4+ helper(42�73 � 5�7%) and CD8+ cytotoxic (22�23 � 3�2%) Tcells were significantly increased in the Echinococcus infec-tion group, whereas the lowest levels (CD4+, 25�6 � 3�5%;CD8+, 13 � 1�8%) were obtained from the breast cancergroup (Figure 2a).In the liver, compared to the control (CD4+,
15�53 � 2�4%) and to the breast cancer (CD4+,14�76 � 1�8%) groups, the percentage of helper T cells wassignificantly higher both in the Echinococcus infection group(25�06 � 5%, P < 0�05) and in the Echinococcus infectionand breast cancer group (21�53 � 3�3%, P < 0�05) (Fig-ure 2b). There was no significant difference in the amountof hepatic CD8+ T cells among the groups (Figure 2b).
In the liver of tumour-bearing mice with cysticechinococcosis, the ratio of CD25+ T cells tends tooverwhelm Th1 subtype
In all groups, splenic helper T cells displayed a na€ıveimmunophenotype characterized with very low percentagesof IFN-c, IL-4, CCR3, CCR5 and CD25 positivity (range,1�41–3�2%).
Compared to the other groups, in the liver of mice fromthe Echinococcus infection and breast cancer group, IFN-c+ or CCR5+ helper T cells, that are regarded as of Th1subset (19), were considerably reduced (Figure 3a,b).Intriguingly, the percentage of Th2 cells identified with IL-4 cytokine was explicitly low in all groups (range, 5�86–2�36%). These cells were almost absent in the liver of micewith echinococcosis and cancer (Figure 3c). CCR3,another marker for Th2 subtype (19), was barely detectedon CD4+ T cells (data not shown). In contrast, the per-centage of hepatic CD4+CD25+ T cells were increased inthe tumour-bearing mice, but this increment could onlyreach to the level of statistical significance in the Echino-coccus infection and breast cancer group (Figure 3d).Of the interest, the presence of CD25 or CCR5 positive
helper T cells in the tumour tissue was also investigated.Irrespective of the Echinococcus infection, the tumourswere highly infiltrated by CD25+CD4+ and CCR5+CD4+
cells (Figure 4). In addition, the liver was determined as aspecific compartment in which helper T cell subsets weresignificantly modulated in the presence of cancer and echi-nococcosis (Figure 4).
DISCUSSION
The relation between inflammation and cancer or chronicinfections is a well-established phenomenon. The magni-tude and character of immunity can be directly correlatedwith progression or eradication of the malignant or infec-tious diseases (2). Individuals under immune suppressionmay have an increased incidence for opportunistic infec-tions (including echinococcosis in the endemic areas) orfor developing certain cancers (1, 9, 20). Moreover, boththe tumour cells and the E. granulosus parasites can divertimmune reactions and maintain a favourable, that isimmune suppressive, microenvironment to survive (1, 2).Intriguingly, certain strategies are shared by cancer cellsand Echinococcus species to avoid or to mitigate immuneresponses. Here, our results indicate an immunological linkbetween cystic echinococcosis and cancer that allowsmetastases to flourish in the liver upon reduction ofTh1-mediated immunity.
Table 2 Frequency of breast cancer metastasis in the tumour-bearing mice
Breast cancer metastasis
4T1 breast cancer group Echinococcus infection and 4T1 breast cancer group
Lung Liver Lung Liver
66�6% (4/6) 33�3% (2/6)* 80% (8/10) 60% (6/10)*
*P < 0�05.
Volume 37, Number 1, January 2015 E. granulosus and cancer metastasis
© 2014 John Wiley & Sons Ltd, Parasite Immunology, 37, 16–22 19

In its natural life cycle, E. granulosus embryos can frequentlyarrive to the liver via portal circulation; hence, hydatid cystsare mostly observed in this organ (4). Inoculation of protos-coleces directly into the peritoneal cavity of mice serves as asecondary infection model in an intermediate host (21). Inthis model, cysts are generally formed in the abdominal cav-
ity, whereas the spread of parasitic infection through vari-ous internal organs such as spleen, liver and lungs is alsoobserved (2, 21). In our study, macroscopic cysts wereobserved in the peritoneal cavity and/or in the liver of someanimals, whereas all mice had lymphocytosis (the increasein both CD4+ and CD8+ cells in the spleen, and CD4+ cells
(a) (b)
Figure 2 Distribution of CD4+ and CD8+ lymphocytes (a) in the spleen and (b) in the liver of experimental groups. Data were obtainedfrom at least three independent experiments and are shown as mean � SD (Student’s t-test; *P < 0�05). (Con., control group; Ech.,Echinococcus infection group; Ca., breast cancer group; Ech.+Ca., Echinococcus infection and breast cancer group).
(a) (c)
(b) (d)
Figure 3 IFN-c, CCR5, IL-4 and CD25 expression in helper T cells. Percentages of CD4+ T cells expressing (a) IFN-c, (b) CCR5, (c) IL-4and (d) CD25 in the liver are given. Data were obtained from at least three independent experiments and are shown as mean � SD(Student’s t-test; *P < 0�05). (Con., control group; Ech., Echinococcus infection group; Ca., breast cancer group; Ech.+Ca., Echinococcusinfection and breast cancer group).
N. Turhan et al. Parasite Immunology
20 © 2014 John Wiley & Sons Ltd, Parasite Immunology, 37, 16–22

in the liver) that confirms the systemic effect of infection.Moreover, this observation indicated the influence of echi-nococcosis on helper T-cell responses in the liver.4T1 breast cancer cells are highly malignant and can
spontaneously metastasize to distant organs. In this exper-imental tumour model, the lung is the preferential organfor metastasis, whereas 4T1 cells can also metastasize intoseveral other organs such as liver, lymph nodes and spleen(22). Organ-specific metastasis can be determined by manyfactors including inflammatory mediators that induce tis-sue remodelling and influence the growth and migrationof tumour cells (23, 24). The co-existence of E. granulosusinfection and breast cancer transformed the liver intoanother preferential site for metastasis. In echinococcosis,the expression of proinflammatory cytokines TNF-a, IL-1a, IL-1b and IL-6 is enhanced in the liver (25, 26). Inaddition, cell migration into inflamed areas becomes facili-tated upon exposure to these mediators (27). Thus, theincrease in breast cancer metastasis to the liver might bedue to the inflammatory mediators secreted in response toE. granulosus infection.4T1 tumours can severely suppress the antitumour
immune responses through generation of MDSCs and Tregcells. These cells can accumulate in the spleen and in theliver (23). Tregs and MDSCs are also detected in early peri-toneal immune responses against experimental E. granulo-sus infection (6, 28). While we did not check the status ofMDSCs, high levels of CD4+ lymphocytes, especially theCD25+CD4+ T cells, were observed in the liver of the micefrom the Echinococcus infection and breast cancer group.Tregs are generally determined with CD25 and FoxP3expression; however, in cancer, the majority of CD25+CD4+
T cells are determined to express FoxP3 transcription factorand display Treg characters (29, 30). These cells produceIL-10 and TGF-b suppressive cytokines, which are highlyexpressed in cystic echinococcosis and also in the tumourmicroenvironments (7, 31). In addition to the tumour cells,
Tregs can also hamper differentiation and function of theantitumour Th1 cells (7). Accordingly, while the percentageof CD4+CD25+ T cells was augmented, the amount of Th1cells expressing IFN-c or CCR5 was diminished in the liverof tumour-bearing mice with echinococcosis.The balance between Th1 and Th2 immune responses is
modulated during E. granulosus infection (1–3). In thechronic stages of the disease, Th2-polarized responsesbecome predominant (4). However, in our experimentalsetting, we did not detect high levels of IL-4- or CCR3-expressing helper T cells. Here, the experiments were ter-minated at the day 105 after the inoculation of protoscole-ces, and it was not determined whether the chronicinflammatory responses were induced. In addition, Th2differentiation is also inhibited by Treg cells, which mayexplain the reduction of IL-4+ phenotype in the Echino-coccus infection and breast cancer group (7).The inflammatory responses given to infectious organ-
isms can promote carcinogenesis and influence the metas-tasis to distant tissues. Alternatively, immune suppressionand/or immune dysregulation that prevails in advancedcancers may favour opportunistic infections (5, 9). Hence,our results may indicate an immunological link betweencystic echinococcosis and cancer that influences the meta-static behaviour of tumour cells.
ACKNOWLEDGEMENTS
This study is funded by The Scientific and TechnologicalResearch Council of Turkey (TUBITAK, project no.:111S329). We thank Dr. Yakut Akyon-Yilmaz and Mrs.Digdem Yoyen-Ermis for their help during conduct of thestudy.
CONFLICT OF INTEREST
None declared.
(a) (b)
Figure 4 Comparison of CD25+ and CCR5+ helper T cell fraction found in spleen, liver and tumour tissues in the breast cancer and in theEchinococcus infection and breast cancer groups. The percentages of (a) CD25+CD4+ and (b) CCR5+CD4+ cells are given. Data wereobtained from at least three independent experiments and are shown as mean � SD (Student’s t-test; *P < 0�05).
Volume 37, Number 1, January 2015 E. granulosus and cancer metastasis
© 2014 John Wiley & Sons Ltd, Parasite Immunology, 37, 16–22 21

REFERENCES
1 Vuitton DA. The ambiguous role of immunityin echinococcosis: protection of the host or ofthe parasite? Acta Trop 2003; 85: 119–132.
2 Zhang W, Li J & McManus DP. Concepts inimmunology and diagnosis of hydatid dis-ease. Clin Microbiol Rev 2003; 16: 18–36.
3 Dematteis S, Baz A, Rottenberg M, Fernan-dez C, Orn A & Nieto A. Antibody andTh1/Th2-type responses in BALB/c miceinoculated with live or dead Echinococcusgranulosus protoscoleces. Parasite Immunol1999; 21: 19–26.
4 Zhang W, Ross AG & McManus DP. Mech-anisms of immunity in hydatid disease:implications for vaccine development. JImmunol 2008; 181: 6679–6685.
5 Abbas AK, Lichtman AH & Pillai S. Effec-tor mechanisms of immune responses. InSchmitt W (ed.): Cellular and MolecularImmunology, 6th edn, updated ed. Philedel-phia, PA, Saunders Elsevier, 2010: 267–350.
6 Pan W, Zhou HJ, Shen YJ, et al. Surveillanceon the status of immune cells after Echinno-coccus granulosus protoscoleces infection inBalb/c mice. PLoS ONE 2013; 8: e59746.
7 Vignali DA, Collison LW & Workman CJ.How regulatory T cells work. Nat Rev Immu-nol 2008; 8: 523–532.
8 Finn OJ. Cancer immunology. N Engl J Med2008; 358: 2704–2715.
9 Dunn GP, Old LJ & Schreiber RD. Thethree Es of cancer immunoediting. Annu RevImmunol 2004; 22: 329–360.
10 Ali R, Ozkalemkas F, Ozkocaman V, et al.Hydatid disease in acute leukemia: effect ofanticancer treatment on echinococcosis.Microbes Infect 2005; 7: 1073–1076.
11 Zold E, Barta Z & Zeher M. Hydatid dis-ease of the liver and associated hepatocellu-lar carcinoma. Clin Gastroenterol Hepatol2005; 3: xxxv.
12 Gungor T, Altinkaya SO, Sirvan L, LafuenteRA & Ceylaner S. Coexistence of borderline
ovarian epithelial tumor, primary pelvichydatid cyst, and lymphoepithelioma-likegastric carcinoma. Taiwan J Obstet Gynecol2011; 50: 201–204.
13 Kim J, Delioukina ML, Lee W, et al. Suc-cessful allogeneic hematopoietic stem celltransplantation for acute myelogenous leuke-mia in a patient with hepatic echinococcalcyst managed by delayed hepatectomy.Transpl Infect Dis 2011; 13: 273–277.
14 Sqalli TH, Buchler M, Al Najjar A, et al.Hydatid cyst disease in a renal allograft reci-pient. Saudi J Kidney Dis Transpl 2009; 20:1057–1060.
15 Bresson-Hadni S, Koch S, Beurton I, et al.Primary disease recurrence after liver trans-plantation for alveolar echinococcosis: long-term evaluation in 15 patients. Hepatology1999; 30: 857–864.
16 Akgul H, Tez M, Unal AE, Keskek M, Say-ek I & Ozcelik T. Echinococcus against can-cer: why not? Cancer 2003; 98: 1999–2000.
17 Noya V, Bay S, Festari MF, et al. Mucin-likepeptides from Echinococcus granulosusinduce antitumor activity. Int J Oncol 2013;43: 775–784.
18 van Knapen F. Echinococcus granulosusinfection and malignancy. Br Med J 1980;281: 195–196.
19 Nakajima H, Fukuda K, Doi Y, et al.Expression of TH1/TH2-related chemokinereceptors on peripheral T cells and correla-tion with clinical disease activity in patientswith multiple sclerosis. Eur Neurol 2004; 52:162–168.
20 Vajdic CM & van Leeuwen MT. Cancer inci-dence and risk factors after solid organtransplantation. Int J Cancer 2009; 125:1747–1754.
21 De Rycke PH, Pennoit-de Cooman E.Experimental secondary echinococcosis ofEchinococcus granulosus. IV. Vaccination ofhost mice. Z Parasitenkd 1973; 42: 49–59.
22 Pulaski BA & Ostrand-Rosenberg S. Mouse4T1 breast tumor model. ed. Coligan JE.Curr Protoc Immunol 2001; Chapter 20: Unit20.22.
23 Nguyen DX, Bos PD & Massague J. Metas-tasis: from dissemination to organ-specificcolonization. Nat Rev Cancer 2009; 9: 274–284.
24 Kim S, Takahashi H, Lin WW, et al. Carci-noma-produced factors activate myeloid cellsthrough TLR2 to stimulate metastasis.Nature 2009; 457: 102–106.
25 Dai WJ & Gottstein B. Nitric oxide-medi-ated immunosuppression following murineEchinococcus multilocularis infection. Immu-nology 1999; 97: 107–116.
26 Haralabidis S, Karagouni E, Frydas S &Dotsika E. Immunoglobulin and cytokineprofile in murine secondary hydatidosis.Parasite Immunol 1995; 17: 625–630.
27 Moro P & Schantz PM. Echinococcosis: areview. Int J Infect Dis 2009; 13: 125–133.
28 Mourglia-Ettlin G, Marques JM, ChabalgoityJA & Dematteis S. Early peritoneal immuneresponse during Echinococcus granulosusestablishment displays a biphasic behavior.PLoS Negl Trop Dis 2011; 5: e1293.
29 Xu L, Xu W, Jiang Z, Zhang F, Chu Y &Xiong S. Depletion of CD4(+)CD25(high)regulatory T cells from tumor infiltratinglymphocytes predominantly induces Th1type immune response in vivo which inhibitstumor growth in adoptive immunotherapy.Cancer Biol Ther 2009; 8: 66–72.
30 Chen L, Huang TG, Meseck M, Mandeli J,Fallon J & Woo SL. Rejection of metastatic4T1 breast cancer by attenuation of Tregcells in combination with immune stimula-tion. Mol Ther 2007; 15: 2194–2202.
31 Wang J, Zhang C, Wei X, et al. TGF-betaand TGF-beta/Smad signaling in the interac-tions between Echinococcus multilocularisand its hosts. PLoS ONE 2013; 8: e55379.
22 © 2014 John Wiley & Sons Ltd, Parasite Immunology, 37, 16–22
N. Turhan et al. Parasite Immunology




















