
A STAT4Dependent Th1 Response Is Required for Resistance to the Helminth Parasite Taenia crassiceps
10
10.1128/IAI.72.8.4552-4560.2004. 2004, 72(8):4552. DOI: Infect. Immun. Lucia E. Rosas, Abhay R. Satoskar and Luis I. Terrazas Miriam Rodríguez-Sosa, Rafael Saavedra, Eda P. Tenorio, Taenia crassiceps Parasite Required for Resistance to the Helminth A STAT4-Dependent Th1 Response Is http://iai.asm.org/content/72/8/4552 Updated information and services can be found at: These include: REFERENCES http://iai.asm.org/content/72/8/4552#ref-list-1 at: This article cites 51 articles, 21 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on June 2, 2014 by guest http://iai.asm.org/ Downloaded from on June 2, 2014 by guest http://iai.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of A STAT4Dependent Th1 Response Is Required for Resistance to the Helminth Parasite Taenia crassiceps

10.1128/IAI.72.8.4552-4560.2004.
2004, 72(8):4552. DOI:Infect. Immun. Lucia E. Rosas, Abhay R. Satoskar and Luis I. TerrazasMiriam Rodríguez-Sosa, Rafael Saavedra, Eda P. Tenorio,
Taenia crassicepsParasite Required for Resistance to the Helminth A STAT4-Dependent Th1 Response Is
http://iai.asm.org/content/72/8/4552Updated information and services can be found at:
These include:
REFERENCEShttp://iai.asm.org/content/72/8/4552#ref-list-1at:
This article cites 51 articles, 21 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on June 2, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
on June 2, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

INFECTION AND IMMUNITY, Aug. 2004, p. 4552–4560 Vol. 72, No. 80019-9567/04/$08.00�0 DOI: 10.1128/IAI.72.8.4552–4560.2004Copyright © 2004, American Society for Microbiology. All Rights Reserved.
A STAT4-Dependent Th1 Response Is Required for Resistance to theHelminth Parasite Taenia crassiceps
Miriam Rodríguez-Sosa,1,2 Rafael Saavedra,3 Eda P. Tenorio,3 Lucia E. Rosas,4Abhay R. Satoskar,4* and Luis I. Terrazas1,2*
Department of Immunology, Instituto Nacional de Cardiología “Ignacio Chavez,” Mexico D.F. 14080,1 Department of Immunology,Instituto de Investigaciones Biomedicas, Universidad Nacional Autonoma de Mexico, Mexico D.F. 04510,3
and Unidad de Biomedicina, FES-Iztacala, Universidad Nacional Autonoma de Mexico,Estado de Mexico 54090,2 Mexico, and Department of Microbiology, The Ohio
State University, Columbus, Ohio 432104
Received 3 September 2003/Returned for modification 15 December 2003/Accepted 28 April 2004
To determine the role of STAT4-dependent Th1 responses in the regulation of immunity to the helminthparasite Taenia crassiceps, we monitored infections with this parasite in resistant mice lacking the STAT4 gene.While T. crassiceps-infected STAT4�/� mice rapidly resolved the infection, STAT4�/� mice were highly sus-ceptible to infection and displayed large parasite loads. Moreover, the inability of STAT4�/� mice to controlthe infection was associated with the induction of an antigen-specific Th2-type response characterized bysignificantly higher levels of Th2-associated immunoglobulin G1 (IgG1) and total IgE as well as interleukin-4(IL-4), IL-10, and IL-13 than those in STAT4�/� mice, who produced significantly more gamma interferon.Furthermore, early after infection, macrophages from STAT4�/� mice produced lower levels of the pro-inflammatory cytokines IL-12, tumor necrosis factor alpha, IL-1�, and nitric oxide (NO) than those fromSTAT4�/� mice, suggesting a pivotal role for macrophages in mediating protection against cysticercosis. Thesefindings demonstrate a critical role for the STAT4 signaling pathway in the development of a Th1-type immuneresponse that is essential for mediating protection against the larval stage of T. crassiceps infection.
Neurocysticercosis is a life-threatening helminth infectioncaused by the larvae of the cestode Taenia solium, which infectshumans and pigs. Although cysticercosis is considered an im-portant public health problem in South America and Asia,recent studies have shown that this disease can also affectpopulations in developed countries (36, 38). Cysticercosiscaused by Taenia crassiceps usually affects rodents, with ca-nines as final hosts (10), although there are reports that im-munocompromised humans can be infected with this parasite(21). Nevertheless, experimental murine cysticercosis has beena useful model for studying the immune mechanisms involvedin determining the disease outcome of cysticercosis (30, 47).
It has been widely accepted that a Th2-mediated responseplays a critical role in the host defense against helminth infec-tions (11, 22), primarily those caused by gastrointestinal nem-atodes (48). However, several reports have shown that somestages of helminth parasites are not efficiently controlled byspecific Th2-type responses. For example, one study found thata Th2 response was not required for controlling Brugia malayiinfection (16). Furthermore, others reported that an early inter-leukin-12 (IL-12)-dependent Th1 response is necessary to medi-ate vaccine-induced protection against schistosomiasis (25).
A series of studies found that although T. crassiceps-infected
BALB/c mice develop an initial but brief Th1-like response,it is replaced by a strong Th2-biased response that is in turnassociated with an increase in the parasite load (46, 51).Members of our laboratory showed previously that the ad-ministration of anti-gamma interferon (IFN-�) neutralizingantibodies to T. crassiceps-infected mice during the earlyphase of infection renders them more susceptible to cysti-cercosis (43). Similarly, it was also found that IL-12 p35�/�
BALB/c mice are more susceptible to the larval stage of T.crassiceps (33). Conversely, T. crassiceps-infected STAT6�/�
mice mounted a strong Th1 response in the absence of Th2development and controlled the infection (31). Taken to-gether, these observations indicate that while a Th1-typeresponse and IFN-� are essential for the development ofimmunity against experimental cysticercosis, Th2-type re-sponses may have a limited role in the control of this par-asitic infection.
Several studies have shown that IFN-� production, which isrequired for immunity against T. crassiceps, can be induced viaboth STAT4-dependent and STAT4-independent signalingpathways (6, 27). Therefore, to determine the relative roles ofSTAT4-dependent and STAT4-independent signaling path-ways in the development of protective immunity against cys-ticercosis, we compared the course of T. crassiceps infection ingenetically resistant C57BL/6 � 129Sv/Ev mice lacking theSTAT4 gene with that in their age- and sex-matched wild-type(STAT4�/�) counterparts. In addition, we analyzed the anti-gen-specific antibody profiles in sera, the cellular immune re-sponses, and cytokine profiles in both spleen cells and perito-neal macrophages as well as the kinetics of cellular recruitmentat the site of infection.
* Corresponding author. Mailing address for Luis I. Terrazas: De-partment of Immunology, Instituto Nacional de Cardiología “IgnacioChavez,” J. Badiano # 1, Mexico, D.F. 14080, Mexico. Phone: (5255)5573-2911. Fax: (5255) 5573-0994. E-mail: [email protected] address for Abhay R. Satoskar: Department of Microbiology,The Ohio State University, 484 West 12th Ave., Columbus, OH 43221.Phone: (614) 292-3243. Fax: (614) 292-8120. E-mail: [email protected].
4552
on June 2, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

Our data demonstrate that the STAT4-dependent IL-12 sig-naling pathway is essential for the development of immunityagainst cysticercosis.
MATERIALS AND METHODS
Animals. STAT4�/� mice were generated by gene disruption as previouslydescribed (44). The STAT4-KO mice used for our experiments were obtainedfrom homozygous inbreeding in the F2 generation (129Sv � C57BL/6). Addi-tionally, we also performed an experiment with STAT4�/� and STAT4�/� lit-termates derived by intercrossing of the third-generation STAT4�/� mice back-crossed to the C57BL/6 strain, and we used C57BL/6 mice (Harlan, Mexico) asan additional control (Fig. 1, inset). The mice in all experiments were 8 to 10weeks old and were bred and maintained in the specific-pathogen-free facility atthe Instituto Nacional de Cardiología “Ignacio Chavez” according to the Insti-tutional and National Guidelines for Animal Research.
Parasites and infection protocols. Metacestodes of T. crassiceps were har-vested from the peritoneal cavity of female BALB/c mice after 2 to 4 months ofinfection. The cysticerci were washed four times in sterile phosphate-bufferedsaline (PBS) (0.15 M, pH 7.2). Experimental infection was achieved by intra-peritoneal (i.p.) injection with 20 small (ca. 2 mm in diameter) nonbuddingcysticerci of T. crassiceps suspended in 0.3 ml of PBS per mouse, and mice weresacrificed at weeks 2, 4, 8, and 16 postinfection. The parasite load was evaluatedby counting all parasites found in the peritoneal cavity after extensive washeswith PBS.
T. crassiceps soluble antigen (TcAg) was obtained from freshly and sterilelyisolated cysticerci from BALB/c female mice after 2 to 4 months of infection. Theparasites were extensively washed with PBS and homogenized with a TissueTearor (Dremel, Racine, Wis.) for cycles of 2 to 3 min on ice. Homogenizedcysticerci were centrifuged at 10,000 rpm for 1 h at 4°C, the supernatant was
collected, and protein levels were determined by the Bradford method. TcAg forcell cultures was sterilized by filtration.
Cell preparations, culture conditions, and cytokine assays. Proliferation as-says were performed on spleen cells obtained from T. crassiceps-infected mice atdifferent time points after infection. Briefly, single-cell suspensions were pre-pared in RPMI 1640 supplemented with 10% fetal bovine serum, 100 U ofpenicillin-streptomycin, 2 mM glutamine, 25 mM HEPES buffer, and 1% non-essential amino acids (complete medium) (all from GIBCO BRL, Grand Island,N.Y.). The erythrocytes were lysed, and viable cells were adjusted to 3 � 106
cells/ml. The cell suspension (100 �l/well) was placed into 96-well flat-bottomedculture plates (Costar, Cambridge, Mass.), stimulated with TcAg (50 �g/ml) in atotal volume of 200 �l, and incubated at 37°C for 96 h. Eighteen hours prior toculture termination, 0.5 �Ci of [3H]thymidine (185 GBq/mmol) (Amersham,Buckinghamshire, England) per well was added. The cells were harvested andthymidine uptake was measured with a Betaplate counter (Wallac, Turku, Fin-land). Values are expressed as mean counts per minute for triplicate wells andare the results after subtracting the counts per minute for cultures in the absenceof antigen. The supernatants from these cultures were analyzed for IFN-�, IL-4,IL-10 (PharMingen, San Diego, Calif.), and IL-13 (R&D Systems, Minneapolis,Minn.) production by enzyme-linked immunosorbent assays (ELISAs).
Isolation of CD4� T cells and cultures with CD4-depleted splenocytes. Spleencells were depleted of CD4� T cells (�95% by fluorescence-activated cell sorting[FACS] analysis) by the use of CD4 magnetic cell sorter beads (MACS; MiltenyiBiotec, Bergisch, Germany) according to the manufacturer’s instructions. CD4�
splenocytes (�5% CD4�) (3 � 106 cells/ml; 100 �l/well) were plated in 96-wellflat-bottomed plates (Costar) as described above. The cultures were maintainedat 37°C in 5% CO2 for 4 days, and then [3H]thymidine (Amersham) (0.5 �Ci/well) was added and the cells were incubated for a further 18 h. The cells wereharvested on a 96-well harvester (Tomtec, Toku, Finland) and counted with aBetaplate counter. Similar cultures were performed with enriched CD4� cells (2
FIG. 1. STAT4�/� mice are highly susceptible while STAT4�/� mice are highly resistant to T. crassiceps infection. The graph shows the courseof T. crassiceps infection in STAT4�/� and STAT4�/� mice after i.p. infection with 20 cysticerci. Data are expressed as mean parasite loads �standard errors (SE) of six mice per group. *, P � 0.01 for STAT4�/� versus STAT4�/� data at the same time point. The data shown arerepresentative of two independent experiments. Inset, C57BL/6 and STAT4�/� littermates displayed similar resistance patterns, while partiallybackcrossed STAT4�/� mice were highly susceptible to T. crassiceps infection. *, P � 0.05 for STAT4�/� versus STAT4�/� and C57BL/6 mice (fiveto six mice per group).
VOL. 72, 2004 ROLE OF STAT4 IN CYSTICERCOSIS 4553
on June 2, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

� 106 cells/ml; 100 �l/well) from the same mice, using irradiated splenocytesfrom healthy STAT4�/� and STAT4�/� mice as antigen-presenting cells (APC)(106 cells/ml; 100 �l/well), and the cultures were processed as described above.
Isolation of peritoneal macrophages and analysis of response to LPS-plus-IFN-� stimulation. Peritoneal exudate cells (PECs) were obtained from theperitoneal cavity of mice infected with T. crassiceps at 2, 4, 8, and 16 weekspostinfection. The cells were washed twice with Hanks balanced salt solution,and erythrocytes were lysed by resuspending the cells in Boyle’s solution (0.17 MTris and 0.16 M ammonium chloride). After two more washes, viable cells werecounted by trypan blue exclusion. PECs were adjusted to 5 � 106/ml in completeRPMI and were cultured in six-well plates (Costar). After 2 h at 37°C in 5% CO2,nonadherent cells were removed by washing with warm supplemented RPMImedium. Cold Ca2�- and Mg2�-free PBS was added, the cells were incubated for5 min, and adherent cells were gently detached with a sterile rubber policeman.The plates were rinsed twice with Ca2�- and Mg2�-free PBS for the collection ofresidual cells. These cells were centrifuged and readjusted to 106/ml. Viabilitywas determined by trypan blue exclusion and was usually �95%. These cellsconstituted �90% macrophages according to FACS analysis with the F4/80monoclonal antibody. One milliliter of cell suspension was then plated, and cellactivation was performed in 24-well plates (Costar) with lipopolysaccharide(LPS) (1 �g/ml, from Escherichia coli 111:B4; Sigma, St Louis, Mo.) plus 2 ng ofrecombinant murine IFN-� (BD Pharmingen, San Diego, Calif.)/ml followed byincubation for 24 h at 37°C in 5% CO2. The supernatants were harvested,centrifuged, and examined by ELISAs for IL-1-, IL-12, IL-18, and tumor ne-crosis factor alpha (TNF-) production (antibodies and cytokines were obtainedfrom BD Pharmingen) and for nitric oxide (Griess reaction) production. TotalPECs were also analyzed by a cytospin preparation stained with Wright-Giemsastain (Sigma), and 300 cells were counted per slide.
Determination of proliferation of CD4 and CD8 T cells by CFSE staining.Spleen cells (107 cells) were stained with 0.5 �M CFSE (5,6-carboxyfluoresceindiacetate succinimidyl ester) as previously described (34). One million cells werestimulated with TcAg (50 �g/ml) in a total volume of 2 ml in 24-well plates andwere incubated at 37°C in 5% CO2 for 4 days. The cells were harvested, washedwith Dulbecco’s PBS–1% fetal calf serum–0.1% NaN3, and stained for 30 min(4°C) with a phycoerythrin (PE)-labeled anti-CD4 monoclonal antibody (1 �g/106 cells) from clone RM4-5 (BD Biosciences) or with a PE-labeled anti-CD8monoclonal antibody (0.25 �g/106 cells) from clone 53-6.7 (BioLegend, SanDiego, Calif.). The cells were washed twice in the same buffer, resuspended inDulbecco’s PBS, and analyzed by flow cytometry. Events were captured as pre-viously described (23).
Flow cytometric analysis. The expression of membrane markers on peritonealadherent cells was analyzed by flow cytometry. Adherent PECs were blockedwith an anti-mouse Fc�R antibody (CD16/CD32) and stained with fluoresceinisothiocyanate-conjugated monoclonal antibodies against F4/80 (Serotec, Ox-ford, United Kingdom) or PE-conjugated antibodies against CD23 or CCR5 (BDPharmingen). Stained cells were analyzed on a FACSCalibur cytometer (BectonDickinson, Mountain View, Calif.).
Antibody ELISA. Blood was collected from the tails of T. crassiceps-infectedSTAT4�/� and STAT4�/� mice at different times after infection. Specific end-point titers of immunoglobulin G1 (IgG1) and IgG2a were determined by ELISAas previously described (42). Total IgE production (serum dilution, 1:10) wasdetected by Opt-ELISA (BD Pharmingen).
Statistical analysis. Comparisons between groups were made with Student’s ttest. P values of �0.05 were considered significant. The statistical significance ofthe titers in sera was determined by a nonparametric Mann-Whitney U–Wil-coxon rank test.
RESULTS AND DISCUSSION
It is widely accepted that the Th2-like response plays acritical role in mediating protective immunity against mosthelminthic infections (11). For example, either IL-4 or IL-13 isnecessary to expulse the gastrointestinal nematode Trichurismuris (5) and the STAT6-mediated signaling pathway has alsobeen shown to promote protective immunity against Trichinellaspiralis (50) and Nippostrongylus brasiliensis (49). Nevertheless,some studies have reported that the Th2-type response maynot be effective against certain stages of helminth parasites (3,39). In fact, some investigators, including us, have shown thatan IL-12-induced Th1-like response is necessary for the suc-
cessful control of helminths such as Schistosoma mansoni (4)and T. crassiceps (33). For the present study, we usedSTAT4�/� mice to investigate the potential roles of theSTAT4-dependent and STAT4-independent signaling path-ways in the development of a protective Th1 response duringT. crassiceps infection. As early as 2 weeks after i.p. inoculationwith 20 nonbudding cysticerci, striking differences in parasitenumbers were observed between STAT4�/� mice (20 � 4parasites) and STAT4�/� (1 � 1 parasites) mice (Fig. 1).Furthermore, as infection progressed, the parasite burdensincreased dramatically in STAT4�/� mice compared to inSTAT4�/� mice, which successfully controlled parasite growthat 2 weeks postinfection and contained only a few parasites intheir peritoneal cavities (Fig. 1). Similar results were also ob-served in experiments that were performed with 10 and 25cysticerci infecting the mice (data not shown). Together, thesefindings indicate that a STAT4-mediated IL-12 signaling path-way plays a critical role in the development of protective im-munity against cysticercosis caused by T. crassiceps. Further-more, in an independent experiment we also found thatSTAT4�/� mice derived by the intercrossing of STAT4�/�
mice were significantly more susceptible to T. crassiceps thanwere their STAT4�/� littermates, suggesting that the pheno-type observed for STAT4�/� mice is most likely due to theselective lack of STAT4 rather than other genes.
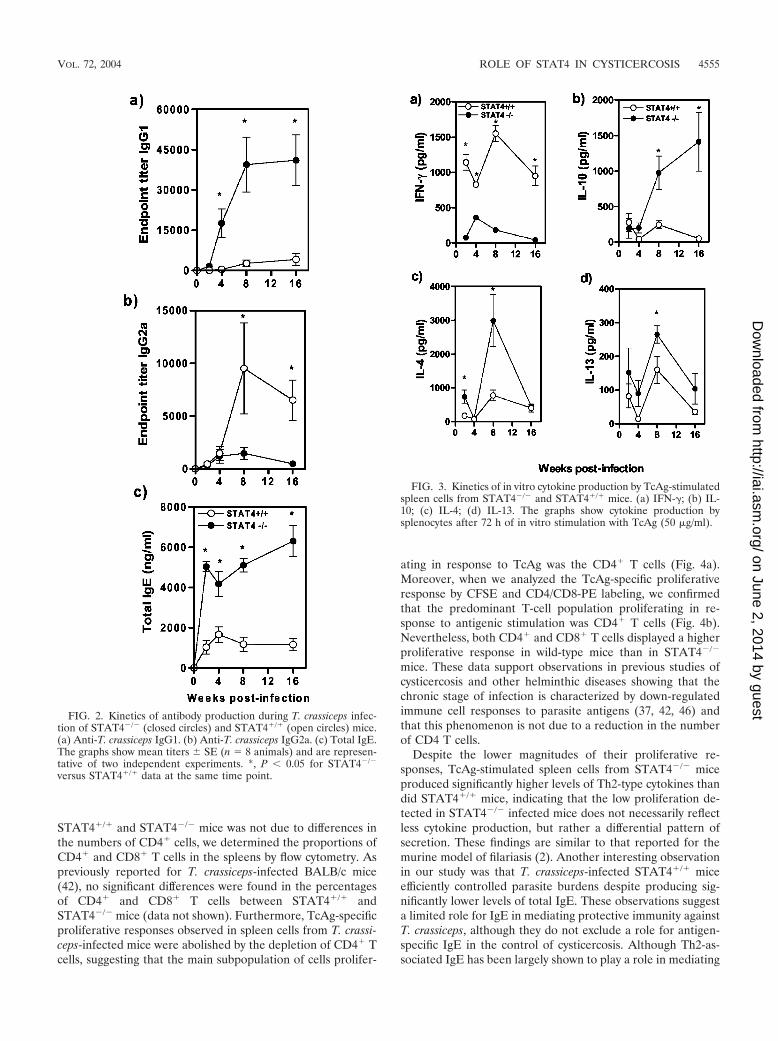
Several studies have demonstrated that a STAT4-mediatedIL-12 signaling pathway mediates protective immunity againstintracellular protozoan parasites by favoring Th1 developmentand simultaneously inhibiting the development of a detrimen-tal Th2 response (7, 40, 41). In contrast, only one study on therole of STAT4 during helminth infections has been performedwhich demonstrated that STAT4�/� mice develop smaller pul-monary granulomas after schistosome egg injection (14). Wehad previously shown that susceptible mice treated with IFN-�plus IL-2 during the early course of T. crassiceps infectionrestricted parasite growth, suggesting that the Th1 responsemediates protective immunity against this parasite (43). In thepresent study, by week 4 postinfection and thereafter, T. cras-siceps-infected STAT4�/� mice displayed significantly highertiters of Th2-associated TcAg-specific IgG1 (Fig. 2a) and totalIgE (Fig. 2c) but significantly less Th1-associated TcAg-specificIgG2a (at 8 and 16 weeks postinfection) than similarly infectedSTAT4�/� mice (Fig. 2b). Furthermore, at weeks 2, 4, 8, and16 postinfection, TcAg-stimulated spleen cells from STAT4�/�
mice produced markedly elevated and sustained levels ofIFN-� compared to those from STAT4�/� mice, who producedsignificantly more IL-10 at week 4 postinfection and thereafter(Fig. 3a and b). At all of these time points, TcAg-stimulatedspleen cells from T. crassiceps-infected STAT4�/� mice se-creted persistently higher levels of IL-4 and IL-13 whichpeaked at week 8 postinfection (Fig. 3c and d).
Interestingly, the spleen cells from T. crassiceps-infectedSTAT4�/� and STAT4�/� mice displayed differences in themagnitudes of proliferative responses after antigen-specific invitro recall. While TcAg-stimulated splenocytes fromSTAT4�/� and STAT4�/� mice displayed comparable prolif-erative responses during early phases of infection, as infectionprogressed those from STAT4�/� mice displayed significantlyhigher responses than those from STAT4�/� mice (Fig. 4). Toensure that this discrepancy in proliferation between
4554 RODRIGUEZ-SOSA ET AL. INFECT. IMMUN.
on June 2, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
STAT4�/� and STAT4�/� mice was not due to differences inthe numbers of CD4� cells, we determined the proportions ofCD4� and CD8� T cells in the spleens by flow cytometry. Aspreviously reported for T. crassiceps-infected BALB/c mice(42), no significant differences were found in the percentagesof CD4� and CD8� T cells between STAT4�/� andSTAT4�/� mice (data not shown). Furthermore, TcAg-specificproliferative responses observed in spleen cells from T. crassi-ceps-infected mice were abolished by the depletion of CD4� Tcells, suggesting that the main subpopulation of cells prolifer-
ating in response to TcAg was the CD4� T cells (Fig. 4a).Moreover, when we analyzed the TcAg-specific proliferativeresponse by CFSE and CD4/CD8-PE labeling, we confirmedthat the predominant T-cell population proliferating in re-sponse to antigenic stimulation was CD4� T cells (Fig. 4b).Nevertheless, both CD4� and CD8� T cells displayed a higherproliferative response in wild-type mice than in STAT4�/�
mice. These data support observations in previous studies ofcysticercosis and other helminthic diseases showing that thechronic stage of infection is characterized by down-regulatedimmune cell responses to parasite antigens (37, 42, 46) andthat this phenomenon is not due to a reduction in the numberof CD4 T cells.
Despite the lower magnitudes of their proliferative re-sponses, TcAg-stimulated spleen cells from STAT4�/� miceproduced significantly higher levels of Th2-type cytokines thandid STAT4�/� mice, indicating that the low proliferation de-tected in STAT4�/� infected mice does not necessarily reflectless cytokine production, but rather a differential pattern ofsecretion. These findings are similar to that reported for themurine model of filariasis (2). Another interesting observationin our study was that T. crassiceps-infected STAT4�/� miceefficiently controlled parasite burdens despite producing sig-nificantly lower levels of total IgE. These observations suggesta limited role for IgE in mediating protective immunity againstT. crassiceps, although they do not exclude a role for antigen-specific IgE in the control of cysticercosis. Although Th2-as-sociated IgE has been largely shown to play a role in mediating
FIG. 2. Kinetics of antibody production during T. crassiceps infec-tion of STAT4�/� (closed circles) and STAT4�/� (open circles) mice.(a) Anti-T. crassiceps IgG1. (b) Anti-T. crassiceps IgG2a. (c) Total IgE.The graphs show mean titers � SE (n � 8 animals) and are represen-tative of two independent experiments. *, P � 0.05 for STAT4�/�
versus STAT4�/� data at the same time point.
FIG. 3. Kinetics of in vitro cytokine production by TcAg-stimulatedspleen cells from STAT4�/� and STAT4�/� mice. (a) IFN-�; (b) IL-10; (c) IL-4; (d) IL-13. The graphs show cytokine production bysplenocytes after 72 h of in vitro stimulation with TcAg (50 �g/ml).
VOL. 72, 2004 ROLE OF STAT4 IN CYSTICERCOSIS 4555
on June 2, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

FIG. 4. Cell type analysis of proliferative responses to TcAg-spe-cific stimulation. (a) Splenocytes from STAT4�/� and wild-type micetaken 2, 4, 8, and 16 weeks after T. crassiceps infection were stimulatedwith 50 �g of TcAg/ml. In some experiments, CD4 cells were magnet-ically removed and CD4� splenocytes were stimulated in the same wayas total splenocytes. *, P � 0.05 for STAT4�/� versus STAT4�/� dataat the same time points. (b) TcAg-specific proliferation of CD4 andCD8 cells. Spleen cells from infected wild-type and STAT4�/� mice (at2 and 16 weeks postinfection) were labeled with CFSE and culturedwith 50 �g of TcAg/ml for 4 days prior to staining with a PE-conju-gated anti-CD4 or anti-CD8 monoclonal antibody and analysis on aflow cytometer. Five thousand nonproliferating, CFSE� (CD4� orCD8�) cells were collected, and the percentages of proliferating andnonproliferating cells were calculated and are indicated in each dotplot. The results shown are representative of those obtained for threemice per group.
4556 RODRIGUEZ-SOSA ET AL. INFECT. IMMUN.
on June 2, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

immunity to helminths (8, 9), one study has found that the lackof IgE does not alter immunity to S. mansoni (39). Neverthe-less, taken together, these findings demonstrate that the Th1-type response induced via the STAT4-dependent signalingpathway is essential for the control of cysticercosis, whereas aTh2-type response is detrimental and enhances susceptibilityto the disease. Consistent with our observations, other studiesusing a radiation-attenuated vaccine with IL-12 as an adjuvanthave also shown that a Th1-like response can mediate protec-tive immunity against certain parasitic helminths, such asSchistosoma (26).
Importantly, we found that CD4� cells were the main sourceof the cytokines analyzed (Table 1), with the exception of IL-10in STAT4�/� mice, in which CD4-depleted spleen cells main-tained the capacity to produce IL-10, suggesting that cellsother than CD4� T cells may be a possible source of thiscytokine which remain to be identified. A crucial role for IL-10in the regulation of immunity has been reported for otherhelminthic diseases, such as schistosomiasis (35) and filariasis(20). Moreover, we have previously shown that a blockade ofIL-10 in susceptible BALB/c mice improved their ability tocontrol cysticercosis (43). Thus, IL-10 may play a major role inthe pathogenesis of cysticercosis due to its immunomodulatoryactivities, such as the down-regulation of costimulatory mole-cules on APC or the suppression of IL-12, IFN-�, and otherpro-inflammatory cytokines (24).
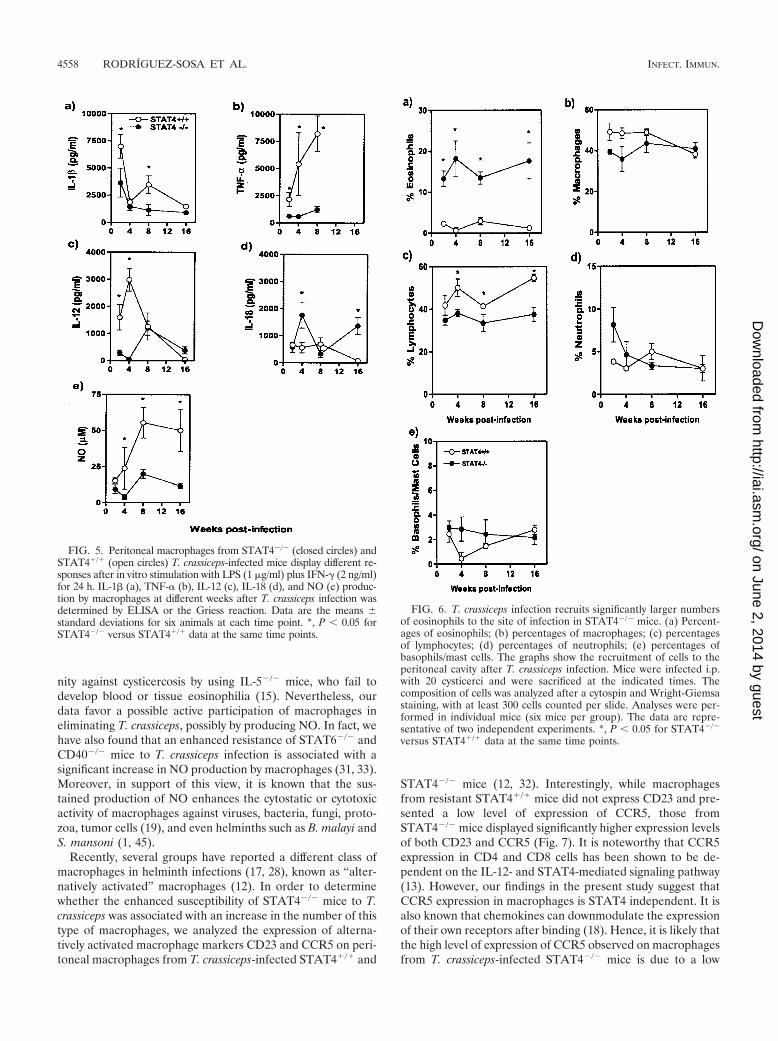
Several studies have demonstrated that macrophages play acritical role in immunity against many intracellular pathogensby their ability to secrete Th1-inducing cytokines such as IL-12and IL-18 and to produce nitric oxide (NO), which is micro-bicidal even to larvae of Schistosoma or Brugia (45). We havehypothesized that macrophages may be involved in mediatingprotective immunity against T. crassiceps metacestodes inSTAT6�/� mice by secreting Th1-inducing cytokines such asIL-12 and/or by releasing NO. Therefore, we analyzed IL-1,IL-12, IL-18, TNF-, and NO production by adherent perito-neal macrophages from T. crassiceps-infected STAT4�/� andSTAT4�/� mice. LPS-plus-IFN-�-activated macrophages fromT. crassiceps-infected STAT4�/� mice obtained during theearly phase of infection (2 weeks) produced higher levels ofIL-1, TNF-, and IL-12 than those from similarly infectedSTAT4�/� mice (Fig. 5a to c). As infection progressed, mac-rophages from chronically infected STAT4�/� mice produced
lower levels of IL-1, IL-18, and IL-12 (Fig. 5a, c, and d), butsignificantly higher levels of TNF- (Fig. 5b). This pattern ofmacrophage response was different from that observed forSTAT4�/� mice, who displayed a low production level of IL-12, IL-1, and TNF- early in infection but had a sustainedproduction of IL-18 during the later phase of infection (Fig. 5ato d). Macrophages from T. crassiceps-infected STAT4�/�
mice also produced significantly lower levels of NO than thosefrom STAT4�/� mice throughout the course of infection (Fig.5e). Taken together, these observations suggest that theSTAT4 signaling pathway mediates resistance to T. crassiceps,at least in part, by favoring macrophage IL-1 and TNF-production, which could be involved directly or indirectly inparasite elimination. It is also likely that the impaired NOproduction observed for STAT4�/� macrophages is due totheir inability to produce TNF-, which is known to activatemacrophages to produce NO. The roles of these factors inregulating resistance to cysticercosis will be the focus of futureinvestigations in our laboratory.
Classically, eosinophils have been considered to be amongthe most efficient effector cells in several helminth parasiticdiseases (8, 9). Therefore, we also evaluated the proportions ofcell populations in inflammatory infiltrates at the site of infec-tion (peritoneal cavity) in STAT4�/� and STAT4�/� mice atdifferent time points after T. crassiceps infection. At 2 weekspostinfection, STAT4�/� mice recruited �12% eosinophilsinto their peritoneal cavities, and this high percentage wasmaintained throughout the course of infection (Fig. 6a). Incontrast, the peritoneal cavities of STAT4�/� mice containedonly a few eosinophils (�2%; P � 0.05) (Fig. 6a). Nonetheless,STAT4�/� mice had an early increased infiltration of macro-phages and lymphocytes compared to STAT4�/� mice (Fig. 6bto c). Additional differences were observed in the recruitmentof neutrophils, which were detected in higher percentages inthe peritoneal cavities of STAT4�/� mice during the earlycourse of infection (Fig. 6d). No significant difference wasobserved in the numbers of basophils/mast cells between thegroups. These results suggest that eosinophils, neutrophils, andbasophils/mast cells do not play a significant role in mediatingresistance against T. crassiceps infection and oppose the dogmathat eosinophils are the key cells that play a critical role ineliminating helminths (8, 9). However, further studies are war-ranted to evaluate the definitive role of eosinophils in immu-
TABLE 1. Antigen-specific cytokine production by CD4-depleted and CD4-enriched splenocytes from STAT4�/� and STAT4�/� miceinfected with T. crassicepsa
Mice
Amt (pg/ml) of cytokine at indicated wk
IL-13 IL-10 IFN-�
2 8 16 2 8 16 2 8 16
STAT4�/�
CD4� �20 ND �20 �20 211 � 211 �20 60 � 25 84 � 41 76 � 105CD4� 38 � 31 ND �20 38 � 70 290 � 173 �20 1,893 � 181 939 � 340 1,215 � 519
STAT4�/�
CD4� �20 ND �20 282 � 60 746 � 352 1,513 � 195 �20 141 � 76 �20CD4� 299 � 86 ND 273 � 163 325 � 191 710 � 190 1,535 � 196 150 � 81 234 � 23 �20
a Splenocytes were recovered after T. crassiceps infection after (2, 8, and 16 weeks) and pooled, and CD4 cells were magnetically removed as described in Materialsand Methods. CD4-depleted splenocytes were stimulated with TcAg (50 �g/ml). Likewise, CD4� cells were replated with splenocytes from naıve STAT4�/� orSTAT4�/� mice as APC and were similarly stimulated. Supernatants were harvested 72 h after stimulation and evaluated for cytokine production by ELISA. Data areexpressed as means � standard deviations and were pooled from two experiments. ND, not determined.
VOL. 72, 2004 ROLE OF STAT4 IN CYSTICERCOSIS 4557
on June 2, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
nity against cysticercosis by using IL-5�/� mice, who fail todevelop blood or tissue eosinophilia (15). Nevertheless, ourdata favor a possible active participation of macrophages ineliminating T. crassiceps, possibly by producing NO. In fact, wehave also found that an enhanced resistance of STAT6�/� andCD40�/� mice to T. crassiceps infection is associated with asignificant increase in NO production by macrophages (31, 33).Moreover, in support of this view, it is known that the sus-tained production of NO enhances the cytostatic or cytotoxicactivity of macrophages against viruses, bacteria, fungi, proto-zoa, tumor cells (19), and even helminths such as B. malayi andS. mansoni (1, 45).
Recently, several groups have reported a different class ofmacrophages in helminth infections (17, 28), known as “alter-natively activated” macrophages (12). In order to determinewhether the enhanced susceptibility of STAT4�/� mice to T.crassiceps was associated with an increase in the number of thistype of macrophages, we analyzed the expression of alterna-tively activated macrophage markers CD23 and CCR5 on peri-toneal macrophages from T. crassiceps-infected STAT4�/� and
STAT4�/� mice (12, 32). Interestingly, while macrophagesfrom resistant STAT4�/� mice did not express CD23 and pre-sented a low level of expression of CCR5, those fromSTAT4�/� mice displayed significantly higher expression levelsof both CD23 and CCR5 (Fig. 7). It is noteworthy that CCR5expression in CD4 and CD8 cells has been shown to be de-pendent on the IL-12- and STAT4-mediated signaling pathway(13). However, our findings in the present study suggest thatCCR5 expression in macrophages is STAT4 independent. It isalso known that chemokines can downmodulate the expressionof their own receptors after binding (18). Hence, it is likely thatthe high level of expression of CCR5 observed on macrophagesfrom T. crassiceps-infected STAT4�/� mice is due to a low
FIG. 5. Peritoneal macrophages from STAT4�/� (closed circles) andSTAT4�/� (open circles) T. crassiceps-infected mice display different re-sponses after in vitro stimulation with LPS (1 �g/ml) plus IFN-� (2 ng/ml)for 24 h. IL-1 (a), TNF- (b), IL-12 (c), IL-18 (d), and NO (e) produc-tion by macrophages at different weeks after T. crassiceps infection wasdetermined by ELISA or the Griess reaction. Data are the means �standard deviations for six animals at each time point. *, P � 0.05 forSTAT4�/� versus STAT4�/� data at the same time points.
FIG. 6. T. crassiceps infection recruits significantly larger numbersof eosinophils to the site of infection in STAT4�/� mice. (a) Percent-ages of eosinophils; (b) percentages of macrophages; (c) percentagesof lymphocytes; (d) percentages of neutrophils; (e) percentages ofbasophils/mast cells. The graphs show the recruitment of cells to theperitoneal cavity after T. crassiceps infection. Mice were infected i.p.with 20 cysticerci and were sacrificed at the indicated times. Thecomposition of cells was analyzed after a cytospin and Wright-Giemsastaining, with at least 300 cells counted per slide. Analyses were per-formed in individual mice (six mice per group). The data are repre-sentative of two independent experiments. *, P � 0.05 for STAT4�/�
versus STAT4�/� data at the same time points.
4558 RODRIGUEZ-SOSA ET AL. INFECT. IMMUN.
on June 2, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
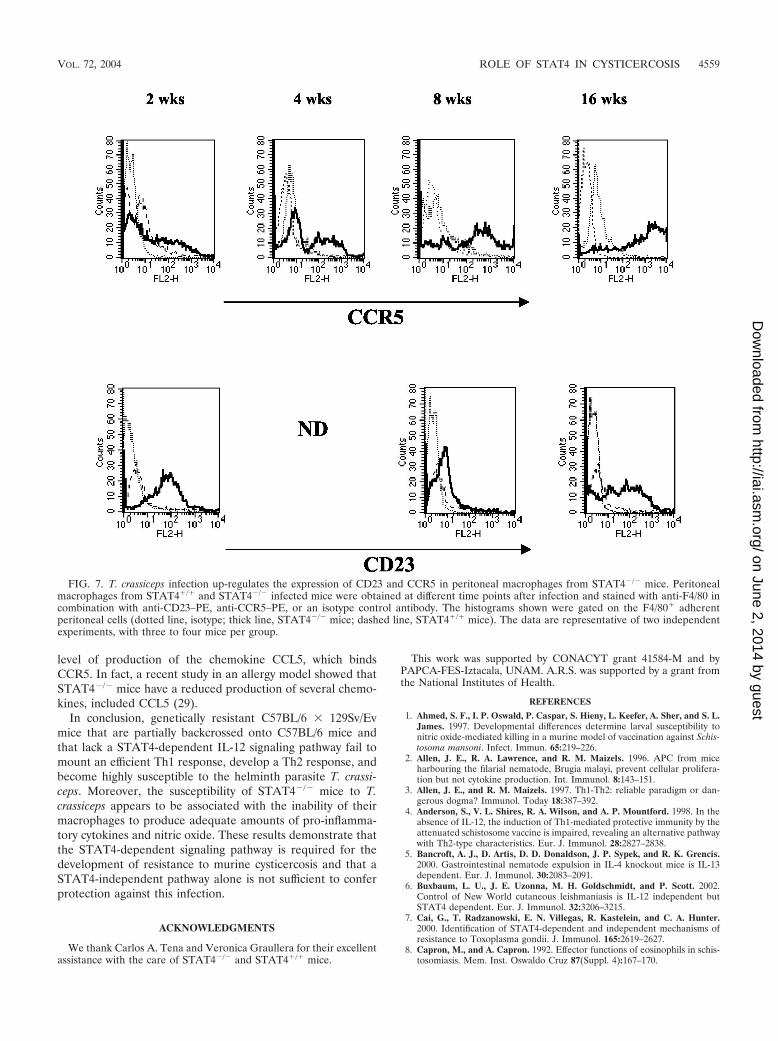
level of production of the chemokine CCL5, which bindsCCR5. In fact, a recent study in an allergy model showed thatSTAT4�/� mice have a reduced production of several chemo-kines, included CCL5 (29).
In conclusion, genetically resistant C57BL/6 � 129Sv/Evmice that are partially backcrossed onto C57BL/6 mice andthat lack a STAT4-dependent IL-12 signaling pathway fail tomount an efficient Th1 response, develop a Th2 response, andbecome highly susceptible to the helminth parasite T. crassi-ceps. Moreover, the susceptibility of STAT4�/� mice to T.crassiceps appears to be associated with the inability of theirmacrophages to produce adequate amounts of pro-inflamma-tory cytokines and nitric oxide. These results demonstrate thatthe STAT4-dependent signaling pathway is required for thedevelopment of resistance to murine cysticercosis and that aSTAT4-independent pathway alone is not sufficient to conferprotection against this infection.
ACKNOWLEDGMENTS
We thank Carlos A. Tena and Veronica Graullera for their excellentassistance with the care of STAT4�/� and STAT4�/� mice.
This work was supported by CONACYT grant 41584-M and byPAPCA-FES-Iztacala, UNAM. A.R.S. was supported by a grant fromthe National Institutes of Health.
REFERENCES
1. Ahmed, S. F., I. P. Oswald, P. Caspar, S. Hieny, L. Keefer, A. Sher, and S. L.James. 1997. Developmental differences determine larval susceptibility tonitric oxide-mediated killing in a murine model of vaccination against Schis-tosoma mansoni. Infect. Immun. 65:219–226.
2. Allen, J. E., R. A. Lawrence, and R. M. Maizels. 1996. APC from miceharbouring the filarial nematode, Brugia malayi, prevent cellular prolifera-tion but not cytokine production. Int. Immunol. 8:143–151.
3. Allen, J. E., and R. M. Maizels. 1997. Th1-Th2: reliable paradigm or dan-gerous dogma? Immunol. Today 18:387–392.
4. Anderson, S., V. L. Shires, R. A. Wilson, and A. P. Mountford. 1998. In theabsence of IL-12, the induction of Th1-mediated protective immunity by theattenuated schistosome vaccine is impaired, revealing an alternative pathwaywith Th2-type characteristics. Eur. J. Immunol. 28:2827–2838.
5. Bancroft, A. J., D. Artis, D. D. Donaldson, J. P. Sypek, and R. K. Grencis.2000. Gastrointestinal nematode expulsion in IL-4 knockout mice is IL-13dependent. Eur. J. Immunol. 30:2083–2091.
6. Buxbaum, L. U., J. E. Uzonna, M. H. Goldschmidt, and P. Scott. 2002.Control of New World cutaneous leishmaniasis is IL-12 independent butSTAT4 dependent. Eur. J. Immunol. 32:3206–3215.
7. Cai, G., T. Radzanowski, E. N. Villegas, R. Kastelein, and C. A. Hunter.2000. Identification of STAT4-dependent and independent mechanisms ofresistance to Toxoplasma gondii. J. Immunol. 165:2619–2627.
8. Capron, M., and A. Capron. 1992. Effector functions of eosinophils in schis-tosomiasis. Mem. Inst. Oswaldo Cruz 87(Suppl. 4):167–170.
FIG. 7. T. crassiceps infection up-regulates the expression of CD23 and CCR5 in peritoneal macrophages from STAT4�/� mice. Peritonealmacrophages from STAT4�/� and STAT4�/� infected mice were obtained at different time points after infection and stained with anti-F4/80 incombination with anti-CD23–PE, anti-CCR5–PE, or an isotype control antibody. The histograms shown were gated on the F4/80� adherentperitoneal cells (dotted line, isotype; thick line, STAT4�/� mice; dashed line, STAT4�/� mice). The data are representative of two independentexperiments, with three to four mice per group.
VOL. 72, 2004 ROLE OF STAT4 IN CYSTICERCOSIS 4559
on June 2, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

9. Capron, M., and A. Capron. 1994. Immunoglobulin E and effector cells inschistosomiasis. Science 264:1876–1877.
10. Chau, C. Y., and R. S. Freeman. 1976. Intraperitoneal passage of Taeniacrassiceps in rats. J. Parasitol. 62:837–839.
11. Finkelman, F. D., T. Shea-Donohue, J. Goldhill, C. A. Sullivan, S. C. Morris,K. B. Madden, W. C. Gause, and J. F. Urban, Jr. 1997. Cytokine regulationof host defense against parasitic gastrointestinal nematodes: lessons fromstudies with rodent models. Annu. Rev. Immunol. 15:505–533.
12. Goerdt, S., O. Politz, K. Schledzewski, R. Birk, A. Gratchev, P. Guillot, N.Hakiy, C. D. Klemke, E. Dippel, V. Kodelja, and C. E. Orfanos. 1999.Alternative versus classical activation of macrophages. Pathobiology 67:222–226.
13. Iwasaki, M., T. Mukai, C. Nakajima, Y. F. Yang, P. Gao, N. Yamaguchi, M.Tomura, H. Fujiwara, and T. Hamaoka. 2001. A mandatory role for STAT4in IL-12 induction of mouse T cell CCR5. J. Immunol. 167:6877–6883.
14. Kaplan, M. H., J. R. Whitfield, D. L. Boros, and M. J. Grusby. 1998. Th2cells are required for the Schistosoma mansoni egg-induced granulomatousresponse. J. Immunol. 160:1850–1856.
15. Kopf, M., F. Brombacher, P. D. Hodgkin, A. J. Ramsay, E. A. Milbourne,W. J. Dai, K. S. Ovington, C. A. Behm, G. Kohler, I. G. Young, and K. I.Matthaei. 1996. IL-5-deficient mice have a developmental defect in CD5�B-1 cells and lack eosinophilia but have normal antibody and cytotoxic T cellresponses. Immunity 4:15–24.
16. Lawrence, R. A., J. E. Allen, W. F. Gregory, M. Kopf, and R. M. Maizels.1995. Infection of IL-4-deficient mice with the parasitic nematode Brugiamalayi demonstrates that host resistance is not dependent on a T helper2-dominated immune response. J. Immunol. 154:5995–6001.
17. Loke, P., A. S. MacDonald, A. Robb, R. M. Maizels, and J. E. Allen. 2000.Alternatively activated macrophages induced by nematode infection inhibitproliferation via cell-to-cell contact. Eur. J. Immunol. 30:2669–2678.
18. Mack, M., J. Cihak, C. Simonis, B. Luckow, A. E. Proudfoot, J. Plachy, H.Bruhl, M. Frink, H. J. Anders, V. Vielhauer, J. Pfirstinger, M. Stangas-singer, and D. Schlondorff. 2001. Expression and characterization of thechemokine receptors CCR2 and CCR5 in mice. J. Immunol. 166:4697–4704.
19. MacMicking, J., Q. W. Xie, and C. Nathan. 1997. Nitric oxide and macro-phage function. Annu. Rev. Immunol. 15:323–350.
20. Mahanty, S., S. N. Mollis, M. Ravichandran, J. S. Abrams, V. Kuma-raswami, K. Jayaraman, E. A. Ottesen, and T. B. Nutman. 1996. High levelsof spontaneous and parasite antigen-driven interleukin-10 production areassociated with antigen-specific hyporesponsiveness in human lymphatic fi-lariasis. J. Infect. Dis. 173:769–773.
21. Maillard, H., J. Marionneau, B. Prophette, E. Boyer, and P. Celerier. 1998.Taenia crassiceps cysticercosis and AIDS. AIDS 12:1551–1552.
22. Maizels, R. M., M. J. Holland, F. H. Falcone, X. X. Zang, and M. Yazdan-bakhsh. 1999. Vaccination against helminth parasites—the ultimate chal-lenge for vaccinologists? Immunol. Rev. 171:125–147.
23. Mannering, S. I., J. S. Morris, K. P. Jensen, A. W. Purcell, M. C. Honeyman,P. M. van Endert, and L. C. Harrison. 2003. A sensitive method for detectingproliferation of rare autoantigen-specific human T cells. J. Immunol. Meth-ods 283:173–183.
24. Moore, K. W., R. de Waal Malefyt, R. L. Coffman, and A. O’Garra. 2001.Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 19:683–765.
25. Mountford, A. P., S. Anderson, and R. A. Wilson. 1996. Induction of Th1cell-mediated protective immunity to Schistosoma mansoni by co-adminis-tration of larval antigens and IL-12 as an adjuvant. J. Immunol. 156:4739–4745.
26. Mountford, A. P., and E. Pearlman. 1998. Interleukin-12 and the host re-sponse to parasitic helminths; the paradoxical effect on protective immunityand immunopathology. Parasite Immunol. 20:509–517.
27. Muller, U., G. Kohler, H. Mossmann, G. A. Schaub, G. Alber, J. P. Di Santo,F. Brombacher, and C. Holscher. 2001. IL-12-independent IFN-gamma pro-duction by T cells in experimental Chagas’ disease is mediated by IL-18.J. Immunol. 167:3346–3353.
28. Noel, W., G. Raes, G. Hassanzadeh Ghassabeh, P. De Baetselier, and A.Beschin. 2004. Alternatively activated macrophages during parasite infec-tions. Trends Parasitol. 20:126–133.
29. Raman, K., M. H. Kaplan, C. M. Hogaboam, A. Berlin, and N. W. Lukacs.2003. STAT4 signal pathways regulate inflammation and airway physiologychanges in allergic airway inflammation locally via alteration of chemokines.J. Immunol. 170:3859–3865.
30. Restrepo, B. I., P. Llaguno, M. A. Sandoval, J. A. Enciso, and J. M. Teale.1998. Analysis of immune lesions in neurocysticercosis patients: central ner-vous system response to helminth appears Th1-like instead of Th2. J. Neu-roimmunol. 89:64–72.
31. Rodriguez-Sosa, M., J. R. David, R. Bojalil, A. R. Satoskar, and L. I. Ter-
razas. 2002. Cutting edge: susceptibility to the larval stage of the helminthparasite Taenia crassiceps is mediated by Th2 response induced via STAT6signaling. J. Immunol. 168:3135–3139.
32. Rodriguez-Sosa, M., A. R. Satoskar, R. Calderon, L. Gomez-Garcia, R.Saavedra, R. Bojalil, and L. I. Terrazas. 2002. Chronic helminth infectioninduces alternatively activated macrophages expressing high levels of CCR5with low interleukin-12 production and Th2-biasing ability. Infect. Immun.70:3656–3664.
33. Rodriguez-Sosa, M., A. R. Satoskar, J. R. David, and L. I. Terrazas. 2003.Altered T helper responses in CD40 and interleukin-12 deficient mice reveala critical role for Th1 responses in eliminating the helminth parasite Taeniacrassiceps. Int. J. Parasitol. 33:703–711.
34. Saavedra, R., E. Segura, R. Leyva, L. A. Esparza, and L. M. Lopez-Marin.2001. Mycobacterial di-O-acyl-trehalose inhibits mitogen- and antigen-in-duced proliferation of murine T cells in vitro. Clin. Diagn. Lab. Immunol.8:1081–1088.
35. Sadler, C. H., L. I. Rutitzky, M. J. Stadecker, and R. A. Wilson. 2003. IL-10is crucial for the transition from acute to chronic disease state during infec-tion of mice with Schistosoma mansoni. Eur. J. Immunol. 33:880–888.
36. Schantz, P. M., and V. C. Tsang. 2003. The US Centers for Disease Controland Prevention (CDC) and research and control of cysticercosis. Acta Trop.87:161–163.
37. Sciutto, E., G. Fragoso, M. Baca, V. De la Cruz, L. Lemus, and E. Lamoyi.1995. Depressed T-cell proliferation associated with susceptibility to exper-imental Taenia crassiceps infection. Infect. Immun. 63:2277–2281.
38. Sciutto, E., G. Fragoso, A. Fleury, J. P. Laclette, J. Sotelo, A. Aluja, L.Vargas, and C. Larralde. 2000. Taenia solium disease in humans and pigs: anancient parasitosis disease rooted in developing countries and emerging as amajor health problem of global dimensions. Microbes Infect. 2:1875–1890.
39. Sher, A., R. L. Coffman, S. Hieny, and A. W. Cheever. 1990. Ablation ofeosinophil and IgE responses with anti-IL-5 or anti-IL-4 antibodies fails toaffect immunity against Schistosoma mansoni in the mouse. J. Immunol.145:3911–3916.
40. Stamm, L. M., A. A. Satoskar, S. K. Ghosh, J. R. David, and A. R. Satoskar.1999. STAT-4 mediated IL-12 signaling pathway is critical for the develop-ment of protective immunity in cutaneous leishmaniasis. Eur. J. Immunol.29:2524–2529.
41. Tarleton, R. L., M. J. Grusby, and L. Zhang. 2000. Increased susceptibilityof Stat4-deficient and enhanced resistance in Stat6-deficient mice to infec-tion with Trypanosoma cruzi. J. Immunol. 165:1520–1525.
42. Terrazas, L. I., R. Bojalil, T. Govezensky, and C. Larralde. 1998. Shift froman early protective Th1-type immune response to a late permissive Th2-typeresponse in murine cysticercosis (Taenia crassiceps). J. Parasitol. 84:74–81.
43. Terrazas, L. I., M. Cruz, M. Rodriguez-Sosa, R. Bojalil, F. Garcia-Tamayo,and C. Larralde. 1999. Th1-type cytokines improve resistance to murinecysticercosis caused by Taenia crassiceps. Parasitol. Res. 85:135–141.
44. Thierfelder, W. E., J. M. van Deursen, K. Yamamoto, R. A. Tripp, S. R.Sarawar, R. T. Carson, M. Y. Sangster, D. A. Vignali, P. C. Doherty, G. C.Grosveld, and J. N. Ihle. 1996. Requirement for Stat4 in interleukin-12-mediated responses of natural killer and T cells. Nature 382:171–174.
45. Thomas, G. R., M. McCrossan, and M. E. Selkirk. 1997. Cytostatic andcytotoxic effects of activated macrophages and nitric oxide donors on Brugiamalayi. Infect. Immun. 65:2732–2739.
46. Toenjes, S. A., R. J. Spolski, K. A. Mooney, and R. E. Kuhn. 1999. Thesystemic immune response of BALB/c mice infected with larval Taeniacrassiceps is a mixed Th1/Th2-type response. Parasitology 118:623–633.
47. Toledo, A., G. Fragoso, G. Rosas, M. Hernandez, G. Gevorkian, F. Lopez-Casillas, B. Hernandez, G. Acero, M. Huerta, C. Larralde, and E. Sciutto.2001. Two epitopes shared by Taenia crassiceps and Taenia solium conferprotection against murine T. crassiceps cysticercosis along with a prominentT1 response. Infect. Immun. 69:1766–1773.
48. Urban, J. F., Jr., K. B. Madden, A. Svetic, A. Cheever, P. P. Trotta, W. C.Gause, I. M. Katona, and F. D. Finkelman. 1992. The importance of Th2cytokines in protective immunity to nematodes. Immunol. Rev. 127:205–220.
49. Urban, J. F., Jr., N. Noben-Trauth, D. D. Donaldson, K. B. Madden, S. C.Morris, M. Collins, and F. D. Finkelman. 1998. IL-13, IL-4Ralpha, and Stat6are required for the expulsion of the gastrointestinal nematode parasiteNippostrongylus brasiliensis. Immunity 8:255–264.
50. Urban, J. F., Jr., L. Schopf, S. C. Morris, T. Orekhova, K. B. Madden, C. J.Betts, H. R. Gamble, C. Byrd, D. Donaldson, K. Else, and F. D. Finkelman.2000. Stat6 signaling promotes protective immunity against Trichinella spi-ralis through a mast cell- and T cell-dependent mechanism. J. Immunol.164:2046–2052.
51. Villa, O. F., and R. E. Kuhn. 1996. Mice infected with the larvae of Taeniacrassiceps exhibit a Th2-like immune response with concomitant anergy anddownregulation of Th1-associated phenomena. Parasitology 112:561–570.
Editor: J. F. Urban, Jr.
4560 RODRIGUEZ-SOSA ET AL. INFECT. IMMUN.
on June 2, 2014 by guesthttp://iai.asm
.org/D
ownloaded from




















