
Evaluation of three enzyme-linked immunosorbent assays (ELISAs) for the detection of serum...
20
Veterinary Parasitology 110 (2002) 57–76 Evaluation of three enzyme-linked immunosorbent assays (ELISAs) for the detection of serum antibodies in sheep infected with Echinococcus granulosus Reinhold Kittelberger a,∗ , Michael P. Reichel b , Judy Jenner a , David D. Heath c , Marshall W. Lightowlers d , Pedro Moro e , Mohamed M. Ibrahem f , Philip S. Craig f , Joseph S. O’Keefe a a National Centre for Disease Investigation, Ministry of Agriculture and Forestry, P.O. Box 40742, Ward Street, Upper Hutt, New Zealand b Novartis Animal Health Australasia Pty Ltd., 245 Western Road, Kemps Creek, NSW 2171, Australia c AgResearch, Wallaceville Animal Research Centre, Ward Street, Upper Hutt, New Zealand d University of Melbourne, Veterinary Clinical Centre, Princess Highway, Werribee, Vic. 3030, Australia e Department of Pathology, Universidad Peruana Cayetano Heredia, P.O. Box 5045, Lima, Peru f Cestode Zoonoses Research Group, Biosciences Research Institute, University of Salford, Salford M5 4WT, UK Received 11 April 2002; received in revised form 25 July 2002; accepted 8 August 2002 Abstract The aim of this study was to develop an immunological method for the identification of sheep infected with Echinococcus granulosus which would allow the monitoring of animals imported into countries free from hydatidosis and as an aid to countries where control schemes for the disease are in operation. Three enzyme-linked immunosorbent assays (ELISAs) were developed and validated, using as antigen either a purified 8 kDa hydatid cyst fluid protein (8kDaELISA), a recombinant EG95 oncosphere protein (OncELISA) or a crude protoscolex preparation (ProtELISA). Sera used for the assay validations were obtained from 249 sheep infected either naturally or experimentally with E. granulosus and from 1012 non-infected sheep. The highest diagnostic sensitivity was obtained using the ProtELISA at 62.7 and 51.4%, depending on the cut-off. Assay sensitivities were lower for the 8kDaELISA and the OncELISA. Diagnostic specificities were high, ranging from 95.8 to 99.5%, depending on the ELISA type and cut-off level chosen. A few sera from 39 sheep infected with T. hydatigena and from 19 sheep infected with T. ovis were recorded as positive. Western immunoblot analysis revealed that the dominant antigenic components in the ∗ Corresponding author. Tel.: +64-4-526-5629; fax: +64-4-526-5601. E-mail address: [email protected] (R. Kittelberger). 0304-4017/02/$ – see front matter © 2002 Published by Elsevier Science B.V. PII:S0304-4017(02)00308-4
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Evaluation of three enzyme-linked immunosorbent assays (ELISAs) for the detection of serum...

Veterinary Parasitology 110 (2002) 57–76
Evaluation of three enzyme-linked immunosorbentassays (ELISAs) for the detection of serum
antibodies in sheep infected withEchinococcus granulosus
Reinhold Kittelbergera,∗, Michael P. Reichelb, Judy Jennera,David D. Heathc, Marshall W. Lightowlersd, Pedro Moroe,
Mohamed M. Ibrahemf, Philip S. Craigf, Joseph S. O’Keefeaa National Centre for Disease Investigation, Ministry of Agriculture and Forestry,
P.O. Box 40742, Ward Street, Upper Hutt, New Zealandb Novartis Animal Health Australasia Pty Ltd., 245 Western Road, Kemps Creek, NSW 2171, Australia
c AgResearch, Wallaceville Animal Research Centre, Ward Street, Upper Hutt, New Zealandd University of Melbourne, Veterinary Clinical Centre, Princess Highway, Werribee, Vic. 3030, Australia
e Department of Pathology, Universidad Peruana Cayetano Heredia, P.O. Box 5045, Lima, Peruf Cestode Zoonoses Research Group, Biosciences Research Institute,
University of Salford, Salford M5 4WT, UK
Received 11 April 2002; received in revised form 25 July 2002; accepted 8 August 2002
Abstract
The aim of this study was to develop an immunological method for the identification of sheepinfected withEchinococcus granulosuswhich would allow the monitoring of animals imported intocountries free from hydatidosis and as an aid to countries where control schemes for the diseaseare in operation.
Three enzyme-linked immunosorbent assays (ELISAs) were developed and validated, usingas antigen either a purified 8 kDa hydatid cyst fluid protein (8kDaELISA), a recombinant EG95oncosphere protein (OncELISA) or a crude protoscolex preparation (ProtELISA). Sera used forthe assay validations were obtained from 249 sheep infected either naturally or experimentallywith E. granulosusand from 1012 non-infected sheep. The highest diagnostic sensitivity wasobtained using the ProtELISA at 62.7 and 51.4%, depending on the cut-off. Assay sensitivitieswere lower for the 8kDaELISA and the OncELISA. Diagnostic specificities were high, rangingfrom 95.8 to 99.5%, depending on the ELISA type and cut-off level chosen. A few sera from39 sheep infected withT. hydatigenaand from 19 sheep infected withT. oviswere recorded aspositive. Western immunoblot analysis revealed that the dominant antigenic components in the
∗ Corresponding author. Tel.:+64-4-526-5629; fax:+64-4-526-5601.E-mail address:[email protected] (R. Kittelberger).
0304-4017/02/$ – see front matter © 2002 Published by Elsevier Science B.V.PII: S0304-4017(02)00308-4

58 R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76
crude protoscolex antigen preparation were macromolecules of about 70–150 kDa, most likelyrepresenting polysaccharides.
This study demonstrated that the ProtELISA was the most effective immunological methodof those assessed for detection of infection withE. granulosusin sheep. Because of its limiteddiagnostic sensitivity of about 50–60%, it should be useful for the detection of the presence ofinfected sheep on a flock basis and cannot be used for reliable identification of individual animalsinfected withE. granulosus.© 2002 Published by Elsevier Science B.V.
Keywords: Echinococcus granulosus; Hydatid; ELISA; Protoscolex; Oncosphere; Hydatid cyst fluid
1. Introduction
Cystic echinococcosis (CE) in humans and ruminants is caused by infection with the larvalstage of the dog tapeworm,Echinococcus granulosus. The parasite has a wide geographicaldistribution, affecting many countries on all continents (Eckert et al., 2000). While CEpersists in many parts of the world and is re-emerging in some regions (Brehm et al.,1999; Eckert et al., 2000; Jenkins, 1998), New Zealand has operated a national eradicationcampaign since 1959 and has successfully eradicated the disease (Anon., 1999; Heath, 1999;Kasper, 1990; Montgomery and van der Logt, 1994). However, live animals imported intothe country will have the potential to re-establish the disease and need to be monitored(Mason and Orr, 1993). Such cases have recently occurred in imported cattle and weredetected post-mortem at meat inspection (Anon., 2000; Bosson, 2000). The availability ofimmunological tests with the ability to detect the majority of ruminants infected withE.granulosuswould be desirable for animal import monitoring and also in countries wherecontrol schemes for the disease are operating.
In order to develop useful screening tests forE. granulosusinfections, one has to considerthe host–parasite interactions that take place during the infections. Initially, immunologicalresponses toE. granulosusin the intermediate host (humans, ruminants) will be directedagainst the invading oncospheres. Later, such responses may develop against components ofthe immature cysts and finally against components of the fertile metacestodes and the proto-scoleces (Lightowlers and Gottstein, 1995). Immune responses can be humoral, leading tothe formation of parasite-specific serum antibodies, or they may be cell-mediated, causingthe proliferation of cytokine-producing T-cells. For comprehensive immunological assaydevelopment, antigenic macromolecules from every stage of the infectious agent may haveto be applied. The readily available hydatid cyst fluid (HCF) has been used most frequentlyas a source for parasite antigens. Its components have been comprehensively investigatedfor their applicability in serological tests (Barbieri et al., 1998; Ibrahem et al., 1996; Itoet al., 1999; Leggatt et al., 1992; Leggatt and McManus, 1994; Lightowlers et al., 1984,1989; Lightowlers and Gottstein, 1995; Maddison et al., 1989; Moro et al., 1997; Younget al., 1984).
The so-called antigen 5 or antigen B was identified as a major antigenic component of thehydatids metacestode (Lightowlers and Gottstein, 1995). It comprises a regularly spacedgroup of molecules, of which the smallest subunit has a molecular weight of approximately

R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76 59
8 kDa, sometimes referred to as 12 kDa, and the other components appearing to be multimersof 16, 24 and 32 kDa (González et al., 1996; Lightowlers et al., 1989). For humans infectedwith E. granulosus, it has been shown that the diagnostic sensitivity of assays using the8 kDa subunit can be high, reaching up to 90%. Reports on the specificity of this antigenare contradictory and it seems that infections with other taenidae will give positive resultsas well (Barbieri et al., 1998; Ito et al., 1999; Leggatt et al., 1992; Leggatt and McManus,1994; Lightowlers et al., 1989; Maddison et al., 1989). While published data on the useof the 8 kDa antigen in serological tests for ruminants are limited (Ibrahem et al., 1996;Moro et al., 1997), they indicate that the 8 kDa protein is potentially a useful antigen forthe sero-diagnosis ofE. granulosusinfections in intermediate host animals.
Detailed investigations in the canine definite host have shown that proteins of theE. granu-losusprotoscoleces are antigenic. They have been successfully applied in the sero-diagnosisof dogs with high sensitivities and specificities (Ahmad and Nizami, 1998; Craig et al., 1995;Gasser et al., 1988, 1993; Lightowlers and Gottstein, 1995). Studies are rare for the applica-tion of such antigens in the sero-diagnosis of sheep, despite reported diagnostic sensitivitiesof up to 80% (Ris et al., 1987).
Antigens of theE. granulosusoncospheres can be expected to cause immune responses inthe early stages of infection in intermediate hosts. Purified oncosphere proteins caused strongantibody responses in experimentally immunised sheep (Heath and Lawrence, 1996) and arecombinant oncosphere protein EG95 conferred a high degree of protection to sheep, whenused as a vaccine (Lightowlers et al., 1996, 1999, 2000; Woollard et al., 1998). Therefore,oncosphere protein antigens may be potentially useful in sero-diagnostic assays for theruminant intermediate host of this parasite.
The cell-mediated immune response toE. granulosusinfections has been investigatedto a limited extend in humans (Hernández-Pomi et al., 1997; Lightowlers and Gottstein,1995) and mice (Dematteis et al., 1999; Rogan, 1998) but little in sheep. In humans, HCFantigen stimulated the proliferation of peripheral blood mononuclear cells (PMBC) in hy-datid patients and increased levels of cytokines, such as interleukin-5 (IL-5) and gammainterferon (IFN-�) were observed (Hernández-Pomi et al., 1997). Therefore, it could beworthwhile to investigate the antigen-dependent release of cytokines in blood fromE.granulosus-infected ruminants as a potential immunological assay for the identification ofinfected animals.
In this study, we have investigated the usefulness of three differentE. granulosusantigenpreparations with samples fromE. granulosus-infected or uninfected sheep for the detectionof serum antibodies againstE. granulosusand for the stimulation of antigen-dependentrelease of the cytokines IL-5 and IFN-� from blood samples obtained fromE. granulosus-infected and control sheep.
2. Materials, methods and techniques
2.1. Sheep sera:
Group 1. Seventy-nine sera were derived from sheep naturally infected withE. granu-losusfrom slaughterhouses in Tasmania, Australia, as reported before (Lightowlers et al.,

60 R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76
1984). Sera were stored at−20◦C. Cyst numbers, locations and volumes, as well as thepresence of other parasites (T. hydatigena, T. ovis, Fasciola hepatica) were recorded post-mortem.
Group 2. Serum samples were obtained from 89 sheep (Junin breed) naturally infectedwith hydatidosis at an abattoir in an endemic region of Peru (Moro et al., 1997). Sera werestored at−20◦C. The number and location of cysts as well as the presence of other parasiteswere recorded post-mortem.
Group 3. Sera were collected from 58 naturally infected sheep withE. granulosushydatidcysts from abattoirs in Tripoli—Libya, Manchester—UK and Iran (Ibrahem et al., 1996).Positive infection was determined at meat inspection by the gross identification of hydatidcysts in the liver and/or lungs. Sera were stored at−20◦C. For these sera, data on cystnumbers, sizes and volumes were not available.
Group 4. Sera were obtained from 23 experimentallyE. granulosus-infected sheep fromNew Zealand, confirmed by necropsy. Sheep which were reared from birth on cestodeegg-free pastures were infected orally withE. granulosuseggs, varying from 10, 100 or1000 eggs and blood was taken regularly until necropsy (Young et al., 1984). Sera werefreeze-dried and stored at−20◦C.
Group 5. Sera from 1012 sheep from all over New Zealand, submitted in 1998 for otherpurposes, were used as negative serum group. New Zealand is considered free fromE.granulosus(Anon., 1999). Sera were stored at−20◦C.
Others. Sera from sheep with other taenid infections were derived as follows: 18 serafrom experimentally, orallyT. ovis-infected sheep and 19 sera from experimentally, orallyT. hydatigena-infected sheep from New Zealand (Young et al., 1984), and 20 sera fromnaturallyT. hydatigena-infected sheep (Ibrahem et al., 1996). Sera were either freeze-driedand stored at−20◦C (Young et al., 1984) or stored at−20◦C (Ibrahem et al., 1996).
2.2. Preparation of protoscolex antigen
Protoscolex antigen was prepared from protoscoleces derived from hydatid cysts fromlivers from experimentally infected sheep in New Zealand by procedures described else-where (Gasser et al., 1988). Using aseptic procedures, brood capsules and protoscoleceswere aspirated in cyst fluid, centrifuged and frozen at−80◦C until further used.
After thawing, the protoscoleces pellet was washed three times with 20 ml of ice-coldTBS (50 mM Tris–HCl, 150 mM NaCl, 0.02% sodium azide, pH 7.5). About 1.8 g of washedprotoscoleces pellet was suspended in 2 ml TBS to which 40�l 0.2 M phenyl methyl sulpho-nyl fluoride (PMSF) in isopropanol were added. The mixture was sonicated under ice/watercooling for 3× 5 min at high, 18�m peak to peak. After centrifugation for 10 min at5000 rpm, 1.7 ml of supernatant were obtained, further referred to as protoscoleces antigenpreparation (ProtAG).
2.3. Preparation and purification of an 8 kDa HCF protein
Sheep HCFs were aspirated from hydatid cysts of experimentally infected sheep atpost-mortem examinations of the animals. Clear cyst fluids were frozen at−20◦C untilused further. Following procedures described previously (Lightowlers et al., 1989), the

R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76 61
HCF was centrifuged at 15,000 rpm (Sorvall SS34 rotor) for 15 min at 4◦C and dialysed(3.5 kDa cut-off of membrane) against 50 volumes (five changes) of distilled water. Thedialysed HCF was frozen and then freeze-dried. Freeze-dried material was re-dissolved in1/10 of the original volume in TBS and centrifuged at 18,000 rpm (Sorvall SS34 rotor) for20 min at 4◦C, resulting in a clear supernatant (conc. HCF). Aliquots were frozen at−20◦Cfor further use.
An aliquot of 10 ml of the conc. HCF was heated in a boiling water bath (Ibrahem et al.,1996) and centrifuged at 18,000 rpm (Sorvall SS34) for 20 min at 4◦C. The supernatant(conc. HCF-boiled) was stored frozen at−20◦C. The pellet was suspended in 1 ml ofsodium dodecyl sulphate (SDS)-polyacrylamide gel electrophoresis (PAGE) sample buffer(SB) containing mercaptoethanol (SB/ME= 62 mM Tris–HCl of pH 6.9, 5% SDS, 1% ME,10% glycerol), boiled for 5 min and then centrifuged for 5 min at 5000×g. The supernatantwas used for SDS-PAGE.
2.4. Purification of 8 kDa protein
SDS-gel filtration chromatography was performed as described before (Kittelberger et al.,1998). A volume of 10 ml of ice-cold conc. HCF-boiled was mixed with 40 ml ice-coldethanol and centrifuged at 3000 rpm for 10 min. The supernatant was discarded and thepellet was air-dried at 37◦C overnight. The pellet was then dissolved in 1 ml SB/ME andapplied to gel filtration chromatography on a Sephacryl S-300 column (diameter 2.5 cm,length 77 cm) at 0.5 ml/h in TBS containing 0.1% SDS. Fractions of 5 ml were collected.Aliquots of 100�l of the fractions were mixed with equal volumes of SB/ME, boiled for5 min and applied to SDS-PAGE.
2.5. Preparation of a recombinant oncosphere antigen
A recombinant 16.5 kDa oncosphere antigen, expressed inE. coli as polyHIS fusionprotein (EG95), was prepared and purified as described before (Lightowlers et al., 1996).
2.6. Polyacrylamide gel electrophoresis (PAGE)
Antigen preparations were diluted in SB/ME, boiled for 5 min then centrifuged for 5 minat 5000× g and the supernatants used for gel electrophoresis. Electrophoresis was per-formed on homogeneous polyacrylamide gels (15%T, 2.7%C) of 1.5 mm thickness usinga 5% stacking gel with a discontinuous Tris–glycine buffer system (Laemmli, 1970) in aMini-Protean II electrophoresis cell (Bio-Rad, Richmond, CA). Gels were run at 200 V for45 min and then either Coomassie-stained, silver-stained (Kittelberger and Hilbink, 1993)or used for Western immunoblotting.
2.7. Immunoblotting
Electrophoretic transfer of antigens from polyacrylamide gels onto membranes (Im-mobilon P membrane, Millipore, Bedford, MA) was carried out in a Trans-Blot SD cell

62 R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76
(Bio-Rad, Richmond, CA) at 15 V for 30 min. Membranes were blocked overnight at roomtemperature in TBS containing 3% non-fat dry milk powder (Anchor, Auckland, NewZealand; TBS-MP), then incubated for 2 h at room temperature with serum samples of50�l, diluted 1:50 in TBS-MP in a Cassette Miniblot System (Immunetics, Cambridge,MA), followed by three washes with TBS-T. Membranes were subsequently incubatedfor 1 h at room temperature with alkaline phosphatase (AP)-conjugated recombinant pro-tein G (Zymed, San Francisco, CA), diluted 1:2000 in TBS-MP and followed by threewashes with TBS-T. Finally, they were incubated for 10 min at room temperature with sub-strate solution (0.1 M Tris–HCl, 0.1 M NaCl, pH 9.8, 60 mM nitro blue tetrazolium and30 mM 5-bromo-4-chloro indolyl phosphate) followed by two 10 min washes with distilledwater.
2.8. Enzyme-linked immunosorbent assay (ELISA) procedure optimisation
Conditions for the three ELISAs were optimised by performing checkerboard titrationsfor every antigen in combination with a positive and a negative control serum. Two typesof microtitre plates were trialled, a high binding plate (Maxisorp, 96-well, Nunc, Kam-strup, Denmark) and a low binding plate (Linbro/Titretek 96-well, microtitration plate, ICNBiomedicals, Aurora, OH). Various coating buffers (50 mM phosphate buffer, pH 7.5 and50 mM carbonate buffer, pH 9.6) and a number of blocking reagents (Tween-20, bovineserum albumin, non-fat dry milk powder) were also tested. All washing steps were per-formed with distilled water containing 0.05% Tween-20. The optimised ELISA procedureswere as follows.
2.9. 8 kDa ELISA and ProtELISA
For the 8 kDa ELISA, microtitration plates (96-well, Linbro/Titertek) were coated byadding 150�l per well of fraction 50 from the gel filtration run, diluted 1/400 in phosphatebuffer (0.05�g protein per well), and incubating at 4◦C overnight in a moisture chamber.Fraction 50 contained pure 8 kDa protein. Then the plates were washed five times, tappeddry and frozen at−20◦C for later use.
For the ProtELISA, microtitration plates (96-well, Linbro/Titertek) were coated by adding150�l per well of ProtAG, diluted 1/50,000 in phosphate buffer (0.005�g protein per well),and incubating at 4◦C overnight in a moisture chamber. Then the plates were washed fivetimes, tapped dry and frozen at−20◦C for later use.
For both ELISAs, the procedure continued as follows: after thawing, the plates wereblocked by incubating with 150�l per well of PBS-T (phosphate-buffered saline: 50 mMphosphate, pH 7.4, 0.15 M sodium chloride, 0.05% Tween-20) for 1 h at 37◦C, followedby five washes. Serum incubations were performed by adding per well 100�l serum, di-luted 1/100 in PBS-T, and incubating at 37◦C for 3 h. After five washes the plates wereincubated with 100�l per well of peroxidase-conjugated Protein G (Zymed, San Fran-cisco, CA) diluted 1/10,000 in PBS-T, for 1 h at 37◦C. After five washes, 100�l per wellTMB substrate (Kirkegaard and Perry Laboratories, Gaithersburg, MD) was added, incu-bated at room temperature for 10 min and the reaction stopped by adding 50�l per well of

R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76 63
2 M sulphuric acid solution. The absorbance values were measured in a microplate reader(Bio-Tek Instruments, Winooski, VT) at 450 nm.
2.10. OncELISA
Microtitration plates (96-well, Linbro/Titertek) were coated by adding 150�l per well ofpurified EG95 recombinant oncosphere antigen, diluted 1/2400 in 50 mM carbonate bufferpH 9.6 (0.01�g protein per well) and incubated at 4◦C overnight in a moisture chamber.Then the plates were washed five times, tapped dry and frozen at−20◦C for later use. Afterthawing, the plates were blocked by incubating with 200�l per well of PBS containing 3%non-fat dry milk powder (PBS-MP) for 1 h at 37◦C, followed by five washes. Serum incuba-tions were performed by adding 100�l serum per well, diluted 1/200 in PBS-MP, and incu-bating at 37◦C for 3 h. After five washes, the plates were incubated with 100�l per well ofperoxidase-conjugated Protein G (Zymed, San Francisco, CA) diluted 1/12,000 in PBS-MP,for 1 h at 37◦C. After five washes, 100�l per well TMB substrate (Kirkegaard and PerryLaboratories, Gaithersburg, MD) was added, incubated at room temperature for 10 min andthe reaction stopped by adding 50�l per well of 2 M sulphuric acid solution. The absorbancewas measured in a microplate reader (Bio-Tek Instruments, Winooski, VT) at 450 nm.
2.11. Calculations
All sera were run in the three ELISAs in duplicates (duplicates on two separate microtitreplates) and also a positive and a negative control serum were run on every plate. Calculationswere then performed using the Microsoft Excel Program. Absorbances of duplicates wereaveraged. All the positive and negative control values were averaged over the 32 plates runin total. Absorbances of all sera were normalised against the average of the positive controlvalue for each ELISA type.
From the absorbance values of the 1012 sera from non-infected sheep, means and standarddeviations were calculated and a cut-off values for each ELISA type was established as meanplus two times standard deviation (cut-off 1) and mean plus three times standard deviation(cut-off 2).
Coefficient of variations over 32 plates was calculated as follows: CV(%) = (100×S.D.
of absorbance of positive controls)/(mean of absorbance of positive controls).Diagnostic sensitivities were calculated for both cut-offs as number of positive sera
divided by total number of sera from infected sheep. Diagnostic specificities were calcu-lated for both cut-offs as number of negative sera divided by total number of sera fromnon-infected sheep.
Frequency distributions were calculated by using the histogram function in the MicrosoftExcel Data Analysis Pak. Absorbances of the various groups of sera were sorted by groupsin 0.5 absorbance units. Frequencies were expressed as relative values to the total numberof sera of the groups (n = 1012 for the negative serum group,n = 249 for the infectedserum group).
Statistical analysis to assess the normality of the distribution of negative sera was per-formed by using the Descriptive Statistics Data Analysis tool provided in the programmeMicrosoft Excel 97.

64 R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76
Fig. 1. Fifteen percent polyacrylamide gel, Coomassie-stained, of various antigen preparations used in this study.Lane 1: 10×concentrated sheep HCF; lane 2: heat-stable proteins from boiled HCF; lane 3: re-dissolved precipitatefrom boiled HCF; lane 4: protoscolex antigen preparation; lane 5: recombinant oncosphere protein EG95; lane 6:molecular weight standards. Note the presence of the 8 kDa protein (7–11 kDa) in the HCF preparation (lanes 1and 2).
2.12. Gamma interferon and interleukin-5 assays
Heparinised bloods were collected from 12 sheep, experimentally infected withE. granu-losusand from eight non-infected sheep. Ten of the infected sheep had hydatid cysts, whichwere either detected by ultrasound scanning or confirmed by post-mortem examination.
Aliquots of 1 ml of the fresh blood samples were incubated for 18 h at 37◦C in 5% CO2with various antigens at 50�g protein per 1 ml of blood. The antigens were concentratedHCF (Fig. 1, lane 1), protoscolex antigen (Fig. 1, lane 4) and recombinant oncosphereantigen (Fig. 1, lane 5). All antigen preparations had been dialysed against PBS and sterilefiltered before use. Blood aliquots were also incubated with TBS as control. After centrifuga-tion (500×g) for 10 min the supernatants were collected and stored frozen at−20◦C. Theywere tested in commercial ELISA kits for the presence of IFN-� (Bovigam, CSL, Parkville,Vic., Australia) and for the presence of IL-5 (OptEIA, mouse IL-5 set, BD Pharmingen,San Diego, CA). Both assays are capture ELISAs, using monoclonal antibodies.
For the IFN-� ELISA, a cut-off was calculated using the mean absorbance of the negativecontrol and two standard deviations(n = 6), and for the IL-5 ELISA using the mean

R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76 65
absorbance of all sera from non-infected sheep plus two standard deviations(n = 48).Positive control sera were run with each commercial ELISA as supplied with the kits.
3. Results
3.1. Antigen preparations
The various antigen preparations used in this study are shown inFig. 1. ConcentratedHCF (Fig. 1, lane 1) contained proteins ranging from 7 to about 150 kDa. Boiling of HCFfollowed by centrifugation, resulted in a supernatant (Fig. 1, lane 2) containing a protein ofabout 8 kDa (6.6–10.5 kDa) and proteins of 15 and 26 kDa, which are considered dimers andtrimers of the 8 kDa protein (Lightowlers et al., 1989). Some higher molecular weight bandsof 60–80 kDa were also present. In the re-dissolved precipitate from the heat-treated HCF(Fig. 1, lane 3) most other HCF proteins were found, mainly of higher molecular weight.
Separation of the proteins in the supernatant from heat-treated HCF was performed bySDS-gel filtration chromatography (Fig. 2). The 8 kDa protein and its multimers could becompletely separated from other components of the HCF (Fig. 2, fraction 52 and 53). Thehighest concentrations of the 8 kDa protein were obtained as a mixture of the monomer andthe dimer (Fig. 2, fractions 50 and 51).
In the protoscolex preparation (Fig. 1, lane 4), at least 10 major protein bands were visible,ranging in size from about 15 to more than 100 kDa. The recombinant oncosphere protein(Fig. 1, lane 5) exhibited a major band of about 20 kDa (Lightowlers et al., 1996).
Fig. 2. Fifteen percent polyacrylamide gel, silver-stained, of fractions from the SDS-gel filtration chromatographyof heat-stable concentrated HCF proteins (HCF conc.). Positions of the monomer 8 kDa protein (8) and its dimer(15) and trimer (26) are indicated.

66 R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76
3.2. Optimisation of ELISA parameters
Parameters for all three ELISAs were optimised initially by using a small number ofsera from sheep experimentally infected withE. granulosusand non-infected sheep. Var-ious blocking reagents, such as Tween-20, bovine serum albumin and milk powder anda number of horse radish peroxidase conjugates (various rabbit-anti-ovine IgGs “Fab, Fc,whole IgG” and recombinant protein G) were also trialled. Checkerboard titrations of anti-gen preparations and sera from infected animals were performed. Once ELISA parameterswere optimised, absorbances of the sera from infected sheep≥1.000 and ratios of sera frominfected to non-infected sheep (signal to noise ratio) at≥5 these conditions were used forfurther validations of the ELISAs.
A problem encountered was the relatively high noise level (absorbance readings for serafrom non-infected sheep). It turned out that recombinant protein G-horse radish peroxidasegave a much lower noise level than other conjugates (data not shown). Of the variousblocking reagents, Tween-20 was best suited for the 8kDaELISA and ProtELISA. Optimisedconditions for each ELISA are as described inSection 2.
3.3. Evaluation of ELISAs
Evaluation of the three ELISAs was performed following recommended procedures(Jacobson, 1998). Evaluation data are listed inTable 1. Coefficients of variation from plateto plate were around 14–15% and signal to noise ratios around 5–7, being within acceptableranges for ELISAs.
Frequency distributions of absorbances of the non-infected sera indicated that for ev-ery ELISA type, these absorbances did not follow a perfect normal distribution (kurto-sis/skewness: ProtELISA= 9.45/3.18; 8kDaELISA= 2.22/1.87; OncELISA= 3.72/2.19). Furthermore, the frequency distributions of the positive sample pool overlappedstrongly with the negative values in every ELISA (Fig. 3). Because of this, two cut-offswere chosen for the ELISA validations. Based on the absorbance values of 1012 sera fromnon-infected sheep, the cut-offs were calculated as mean plus 2× standard deviation andmean plus 3× standard deviations. The cut-off values were relatively high for all threeELISAs at absorbances of around 0.3–0.5. Lowest values were obtained for the ProtELISAat 0.304 and 0.369, respectively.
The diagnostic specificities for the three ELISAs were all above 95%. The 8kDaELISAexhibited highest specificities with up to 99.5%, while the ProtELISA and the OncELISAshowed lower but similar values at 95 and 98% for the two cut-offs. Results with thesera fromT. hydatigena-infected sheep revealed that at both cut-offs the ProtELISA willidentify some sera as positives (87.2 and 92.3% diagnostic specificity) and the 8kDaELISAat cut-off 1 (94.9% diagnostic specificity). With sera fromT. ovis-infected animals, onlythe OncELISA was not 100% specific at cut-off 2 (94.7% diagnostic specificity).
Diagnostic sensitivities were relatively low for the ProtELISA, poor for the 8kDaELISAand very poor for the OncELISA. The ProtELISA showed the highest sensitivities fornaturallyE. granulosus-infected sheep at 64.6 and 53.1% for the two cut-offs, respectively.Interestingly, the diagnostic sensitivities were markedly lower (34.5 and 34.8%) for theexperimentally infected animals. This situation was reversed for the 8kDaELISA, where

R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76 67
Table 1Summary of validation data for three hydatids ELISAsa
ELISA data Value
ProtELISA 8kDaELISA OncELISA
Mean of positive control serum OD over 12 plates (24 values) 1.001 2.196 1.476S.D. of positive control serum OD over 12 plates (24 values) 0.144 0.334 0.205CV (%) between 12 plates based on positive control serum 14.3 15.2 13.9Mean of negative control serum OD over 12 plates (24 values) 0.196 0.326 0.282Cut-off 1, mean of 1012 negative sera+ 2 × S.D. (OD, normalised) 0.304 0.407 0.425Cut-off 2, mean of 1012 negative sera+ 3 × S.D. (OD, normalised) 0.369 0.489 0.532Signal/noise (OD positive control serum/OD negative control serum) 5.1 6.7 7.2
Diagnostic specificity (%), non-infected sheepAt cut-off 1 (n = 1012) 95.8 96.7 95.8At cut-off 2 (n = 1012) 98.2 99.5 98.1
Diagnostic specificity (%),T. hydatigena-infected sheepAt cut-off 1 (n = 39) 87.2 94.9 100.0At cut-off 2 (n = 39) 92.3 100.0 100.0
Diagnostic specificity (%),T. ovis-infected sheepAt cut-off 1 (n = 19) 100.0 100.0 94.7At cut-off 2 (n = 19) 100.0 100.0 100.0
Diagnostic sensitivity (%),E. granulosus-infected sheepAt cut-off 1, all animals(n = 249) 62.7 11.2 5.2At cut-off 2, all animals(n = 249) 51.4 5.2 1.6At cut-off 1, experimentally infected sheep(n = 23) 34.5 26.1 4.3At cut-off 2, experimentally infected sheep(n = 23) 34.8 26.1 0.0At cut-off 1, naturally infected sheep(n = 226) 64.6 9.7 5.3At cut-off 2, naturally infected sheep(n = 226) 53.1 3.1 1.8
a S.D.: standard deviation; OD: optical density (absorbance); CV: coefficient of variation; cut-off 1: mean+2 × S.D.; cut-off 2: mean+ 3 × S.D..
the diagnostic sensitivities were much lower for naturally infected (9.7 and 3.1%) than forexperimentally infected sheep (26.1% for both cut-offs).
3.4. Analysis of antigenic components
In order to identify immuno-dominant antigenic components in the HCF and in the pro-toscolex antigen preparation, seven sera from infected sheep and 14 sera from non-infectedsheep were run in immunoblots with concentrated, boiled HCF and with the protoscolexantigen. Results are shown inFig. 4. With HCF as antigen, sera from infected sheep showeddominant staining of the 8 kDa protein and its multimers. Sera from non-infected sheepexhibited no staining.
Immunoblots with the protoscolex antigen preparation showed strong staining of bandsin the molecular weight range 70–150 kDa with sera from infected sheep and also weakstaining of a band of about 28 kDa. Serum samples from non-infected sheep were notcompletely free from staining. Some exhibited weak staining in the 60–150 kDa range, butalso weak staining around 18–20 kDa. From these immuno-stainings, one can conclude that

68 R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76
Fig. 3. Frequency distributions for ELISA absorbances of sera from non-infected sheep(n = 1012) and sera fromE. granulosus-infected sheep(n = 249).

R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76 69
Fig. 4. Immunoblots with concentrated, heat-stable HCF antigen (a) and a protoscolex antigen preparation (b).Lanes 1–7: sera fromE. granulosus-infected sheep; lanes 8–21: sera from non-infected sheep. Note the dominant8 kDa protein and its multimers in (a) and the dominant proteins of 70–150 kDa in (b).
70 R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76
the immuno-dominant components of the protoscolex antigen are macromolecules of about100 kDa. Their nature is unclear since no strong protein-staining is visible in this molecularweight range (Fig. 1, lane 4) and therefore they may represent polysaccharides.
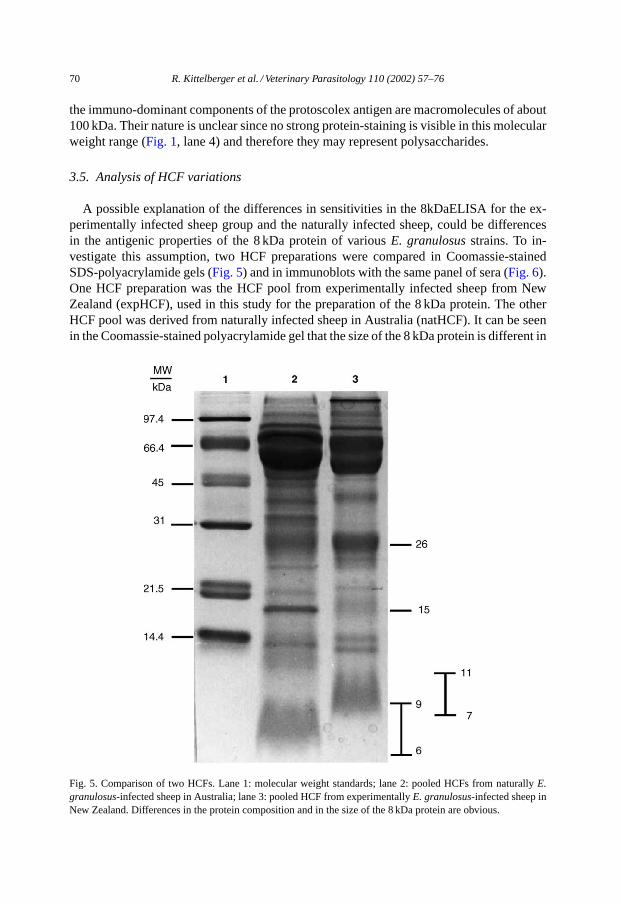
3.5. Analysis of HCF variations
A possible explanation of the differences in sensitivities in the 8kDaELISA for the ex-perimentally infected sheep group and the naturally infected sheep, could be differencesin the antigenic properties of the 8 kDa protein of variousE. granulosusstrains. To in-vestigate this assumption, two HCF preparations were compared in Coomassie-stainedSDS-polyacrylamide gels (Fig. 5) and in immunoblots with the same panel of sera (Fig. 6).One HCF preparation was the HCF pool from experimentally infected sheep from NewZealand (expHCF), used in this study for the preparation of the 8 kDa protein. The otherHCF pool was derived from naturally infected sheep in Australia (natHCF). It can be seenin the Coomassie-stained polyacrylamide gel that the size of the 8 kDa protein is different in
Fig. 5. Comparison of two HCFs. Lane 1: molecular weight standards; lane 2: pooled HCFs from naturallyE.granulosus-infected sheep in Australia; lane 3: pooled HCF from experimentallyE. granulosus-infected sheep inNew Zealand. Differences in the protein composition and in the size of the 8 kDa protein are obvious.

R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76 71
Fig. 6. Western immunoblot results of a panel of sheep sera with two HCF preparations. (a) HCF from experimen-tally infected sheep, (b) HCF from naturally infected sheep. Lanes 1–9: sera from naturallyE. granulosus-infectedPeruvian sheep (8kDaELISA negative), previously reported to show the 8 kDa band in immunoblots; lanes 10–12:sera from experimentally infected sheep (8kDaELISA positive); lanes 13–20: sera from naturally infected Aus-tralian sheep (only sera in lanes 15 and 19 were 8kDaELISA positive); lane 21: pool of sera from non-infectedsheep.

72 R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76
Table 2Antigen-dependent cytokine release in bloods fromE. granulosus-infected and non-infected sheep
Sheep group Number of bloods positive (%)
IFN-� IL-5
ProtAG HCF OncAG ProtAG HCF OncAG
ExperimentallyE. granulosus-infected(n = 12)
3 (25.0) 1 (8.3) 1 (8.3) 3 (25.0) 3 (25.0) 2 (16.7)
Non-infected(n = 8) 4 (50.0) 2 (25.0) 0 (0) 1 (12.5) 1 (12.5) 0 (0)
the natHCF (Fig. 5, lane 2, 5.8–8.5 kDa) from the 8 kDa component in the expHCF (Fig. 5,lane 3, 6.6–10.5 kDa). While many protein bands are identical in both preparations, somedifferences in the protein-banding patterns are obvious.
For immunoblot analysis (Fig. 6), a limited number of sera from experimentally infectedsheep and from naturally infected sheep from Peru and Australia were used. Nine sera werefrom Peruvian sheep (lanes 1–9) previously reported to show staining of the 8 kDa bandin the immunoblot (Moro et al., 1997). All were negative in the 8kDaELISA in this study.Three sera were from experimentally infected sheep (lanes 10–12) which were all strongpositive in the 8kDaELISA. Eight sera were from naturally infected sheep in Australia(lanes 13–20), of which the sera in lanes 15 and 19 were strong positive in the 8kDaELISA,while the other six were negative in the ELISA. In the immunoblot, differences betweenthe two HCF preparations became obvious. While both preparations contained about equalconcentrations of proteins and the 8 kDa protein was present in both in equal amounts (asjudged from the Coomassie-stained gel inFig. 5), pronounced staining of the 8 kDa bandor its multimers was only visible with the expHCF. From the five 8kDaELISA positivesera, four exhibited strong staining of the 8 kDa protein and other proteins, including itsmultimers.
From these results, one cannot necessarily conclude that strain differences are causingthe differences in reactivity of the various sera groups in the 8kDaELISA. Nevertheless,it became clear that there are differences in the size and immunological reactivity of thisprotein in different HCF preparations.
3.6. Analysis of cell-mediated immune response
This experiment was performed with a limited number of bloods from experimentallyinfected and non-infected sheep. NoE. granulosusinfection-specific release of IFN-� orIL-5 in response to any of the three antigens was observed (Table 2).
4. Discussion
In the present study, three ELISAs were evaluated with a large number of serum samplesfor their suitability to detect sheep infected withE. granulosus. The antigens applied inthese ELISAs were derived from various stages of the life cycle of the parasite, relevant to

R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76 73
the intermediate host, i.e. a recombinant oncosphere protein (EG95), an 8 kDa HCF proteinand a crude protoscolex antigen preparation. From these antigens, the 8 kDa protein wasexpected to be the most promising antigen because of its comprehensively reported use invarious serological assays for human infections and, to a much lesser extent, in animals.
In humans, several reports found high diagnostic sensitivities and specificities for this8 kDa (12 kDa) antigen for patients with hepatic cysts. (Ito et al., 1999; Leggatt et al., 1992;Leggatt and McManus, 1994; Maddison et al., 1989). A Western immunoblot assay forthe 8 kDa monomer and its multimers (Maddison et al., 1989) is also commercially avail-able, with stated 80% diagnostic sensitivity and 100% diagnostic specificity (Immunetics,Cambridge, MA). Nevertheless, results by others could not confirm these high specificities(Lightowlers et al., 1989). Diagnostic sensitivities of serological tests developed for rumi-nants, ranged from 57 to 90% in ELISA, depending on the source of the antigen (Ibrahemet al., 1996), and up to 73% in the Western blot (Moro et al., 1997). Diagnostic specificitieswere high and ranged from 93 to 100%.
In the present study, with larger numbers of sera from infected sheep than used in previousreports, we found quite low sensitivities (11.2 and 5.2%) for the purified 8 kDa antigen inthe ELISA. This is interesting because many of the sheep sera used here were obtained frompanels applied before in these studies (Ibrahem et al., 1996; Moro et al., 1997). Furthermore,sera from sheep, which had been shown to stain the 8 kDa band in another study, did notreact here in the Western blot (seeFig. 6).
These findings are difficult to explain but may be related to theE. granulosusstrain in-volved. The antigen, a purified 8 kDa protein, was obtained from HCF from experimentallyinfected sheep from New Zealand, which were infected with the sameE. granulosusstrainfor many years. A first indication of this difference was the much higher diagnostic sensitiv-ity for the experimentally infected sheep group compared to the naturally infected sheep (seeTable 1). Furthermore, differences in the size of the 8 kDa component from HCF from exper-imentally infected sheep from New Zealand could be demonstrated in Coomassie-stainedgels, when compared to the 8 kDa protein from HCF from Australian, naturally infectedanimals (Fig. 5). Differences of the immunological reactivity in the Western blot werealso observed (Fig. 6). Whatever the reasons are for the low diagnostic sensitivity of the8kDaELISA, it became clear that this assay, using a purified 8 kDa protein, was not suitableas a test method for the identification of the majority of sheep infected withE. granulosus.
Protoscoleces-derived antigens have been successfully applied in ELISAs for the detec-tion of serum antibodies againstE. granulosusin dogs (Craig et al., 1995; Gasser et al., 1988,1993; Reichel et al., 1996). Very little work has been done to date to apply such antigens forthe sero-diagnosis of the intermediate hosts, despite the fact that protoscoleces are parts ofthe maturing hydatid cysts. An ELISA, using polysaccharide antigens from protoscoleces,was reported to be 80% sensitive in sheep, when combined with an ELISA using a mouseHCF membrane antigen (Ris et al., 1987). In the present study, a protoscolex-derived crudeantigen preparation resulted in relatively high diagnostic sensitivities (up to 64.6%) andspecificities (up to 98.2%). Immuno-staining in Western blots revealed that the majorityof serum antibodies were directed against antigens of about 70–150 kDa. Since little pro-tein staining in this molecular weight range was observed in the polyacrylamide gel, theimmuno-dominant antigenic components are most likely polysaccharides. Further purifica-tion of these components for use in ELISA may lead to improved serological test methods.

74 R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76
The recombinant oncosphere antigen EG95, which confers protective immunity to sheepand is used as an effective vaccine (Heath and Holcman, 1997; Lightowlers et al., 1996, 1999;Woollard et al., 1998, 1999), failed in this study as a diagnostic antigen for the sero-diagnosisof E. granulosusin sheep. While the recombinant protein leads to the formation of serumantibodies in vaccinated sheep (Woollard et al., 1998), it seems that little specific serumantibodies are present in the naturally or experimentally infected animal.
This study and many others (Lightowlers and Gottstein, 1995) have shown that sheep doproduce specific immunological responses during naturalE. granulosusinfections. How-ever, it seems that these responses do not lead to raised serum antibody levels in manyanimals or they are not maintained throughout the course of the infection. Another pos-sibility to diagnose the presence of the infection could be the testing for cell-mediatedimmune responses (Hernández-Pomi et al., 1997; Rogan, 1998). The antigen-specific re-lease of the cytokines IFN-� and IL-5, for which standardised tests were available, weretherefore measured in a limited number of whole bloods from infected and non-infectedsheep. The antigens used were concentrated HCF, containing the 8 kDa protein, EG95 andthe protoscolex antigen preparation. The results from this limited study showed that thereis no specific release of IFN-� or IL-5 in the animals investigated. It may be that theE.granulosus-infected sheep were already in a too advanced stage of the infection in order toexhibit TH2/TH1-type responses. It has been shown in mice that TH2-type responses, i.e.the release of IL-10, IL-4 and IL-5 were only stimulated in the first 4 weeks post-infection(Dematteis et al., 1999). Little is known about cell-mediated immunity in animals infectedwith E. granulosus. More work needs to be done in this field to obtain a clearer picture ofthe cell-mediated immune response in sheep.
The major outcome of these studies was the finding that an ELISA using a protoscoleces-derived crude antigen preparation could be successfully applied for the detection of infectedanimals. Its diagnostic sensitivity was up to 62.7% and its diagnostic specificity up to98.2%, depending on the cut-off values used. The ProtELISA should be a useful tool forthe identification of sheep flocks infected withE. granulosus.
Acknowledgements
This work was funded by grants from MAF Policy, New Zealand and the National Healthand Medical Research Council of Australia.
References
Ahmad, G., Nizami, W.A., 1998. Coproantigens: early detection and suitability of an immunodiagnostic methodfor echinococcosis in dogs. Vet. Parasitol. 77, 237–244.
Anon., 1999. Hydatids close to eradication. Vetscript March, 24.Anon., 2000. Hydatid cyst in a cow. Surveillance 27, 14.Barbieri, M., Fernández, C., González, A., Luaces, V.M., Nieto, A., 1998. Diagnostic evaluation of a synthetic
peptide derived from a novel antigen B subunit as related to other available peptides and native antigens usedfor serology of cystic hydatidosis. Parasite Immunol. 29, 51–61.
Bosson, M., 2000. Hydatidosis in New Zealand-a case report. Surveillance 27, 17.Brehm, K., Kern, P., Hubert, K., Frosch, M., 1999. Echinococcosis from every angle. Parasitol. Today 15, 351–352.

R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76 75
Craig, P.S., Gasser, R.B., Parada, L., Cabrera, L., Parietti, S., Borgues, C., Acuttis, A., Agulla, J., Sowden, K.,Paolillo, E., 1995. Diagnosis of canine echinococcosis: comparison of coproantigen and serum antibody testswith arecoline purgation in Uruguay. Vet. Parasitol. 56, 293–301.
Dematteis, S., Baz, A., Rottenberg, M., Fernández, C., Anders, Ö., Nieto, A., 1999. Antibody and Th1/Th2-typeresponses in BALB/c mice inoculated with live or deadEchinococcus granulosusprotoscoleces. ParasiteImmunol. 21, 19–26.
Eckert, J., Conraths, F.J., Tackmann, K., 2000. Echinococcosis: an emerging or re-emerging zoonosis? Int. J.Parasitol. 30, 1283–1294.
Gasser, R.B., Lightowlers, M.W., Obendorf, D.L., Jenkins, D.J., Rickard, M.D., 1988. Evaluation of a serologicaltest system for the diagnosis of naturalEchinococcus granulosusinfection in dogs usingE. granulosusprotoscolex and oncosphere antigens. Aust. Vet. J. 65, 369–373.
Gasser, R.B., Jenkins, D.J., Paolillo, E., Parada, L., Cabrera, L., Craig, P.S., 1993. Serum antibodies in canineechinococcosis. Int. J. Parasitol. 23, 579–586.
González, G., Nieto, A., Fernández, C., Örn, A., Wernstedt, C., Hellman, U., 1996. Two different 8 kDa monomersare involved in the oligomeric organization of the nativeEchinococcus granulosusantigen B. Parasite Immunol.18, 587–596.
Heath, D.D., 1999. Hydatids close to eradication in New Zealand, Abstract 39–40 at the XXXIII Archivos Int.Hidatidosis, San Carlos de Bariloche, Rio Negro, Argentina.
Heath, D.D., Holcman, B., 1997. Vaccination againstEchinococcusin perspective. Acta Trop. 67, 37–41.Heath, D.D., Lawrence, S.B., 1996. Antigenic polypeptides ofEchinococcus granulosusoncospheres and definition
of protective molecules. Parasite Immunol. 18, 347–357.Hernández-Pomi, A., Borras-Salvador, R., Mir-Gisbert, A., 1997. Analysis of cytokine and specific antibody
profiles in hydatid patients with primary infection and relapse of disease. Parasite Immunol. 19, 553–561.Ibrahem, M.M., Craig, P.S., McVie, A., Ersfeld, K., Rogan, M.T., 1996.Echinococcus granulosusantigen B and
seroreactivity in natural ovine hydatidosis. Res. Vet. Sci. 61, 102–106.Ito, A., Liang, M.A., Schanz, P.M., Gottstein, B., Liu, Y.-H., Chai, J.-J., Abdel-Hafez, S.K., Altintas, N., Joshi, D.D.,
Lightowlers, M.W., Pawlowski, Z.S., 1999. Differential serodiagnosis for cystic and alveolar echinococcosisusing fractions ofEchinococcus granulosuscyst fluid (antigen B) andE. multilocularisprotoscolex (EM18).Am. J. Trop. Med. Hyg. 60, 188–192.
Jacobson, R.H., 1998. Validation of serological assays for diagnosis of infectious diseases. Rev. Sci. Tech. Off.Int. Epiz. 17, 469–486.
Jenkins, D.J., 1998. Hydatidosis—a zoonosis of unrecognised increasing importance? J. Med. Microbiol. 47,281–282.
Kasper, K., 1990. Hydatids control in New Zealand. Surveillance 17, 35–36.Kittelberger, R., Hilbink, F., 1993. Sensitive silver-staining detection of bacterial lipopolysaccharides in
polyacrylamide gels. J. Biochem. Biophys. Meth. 26, 81–86.Kittelberger, R., Diack, D.S., Vizcaı́no, N., Cloeckaert, A., Zygmunt, M.S., 1998. Characterization of an
immuno-dominant antigen inBrucella ovisand evaluation of its use in an enzyme-linked immunosorbentassay. Vet. Microbiol. 59, 213–227.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature227, 680–685.
Leggatt, G.R., McManus, D.P., 1994. Identification and diagnostic value of a major antibody epitope on the 12kDa antigen fromEchinococcus granulosus(hydatid disease) cyst fluid. Parasite Immunol. 16, 87–96.
Leggatt, G.R., Yang, W., McManus, D.P., 1992. Serological evaluation of the 12 kDa subunit of antigen B inEchinococcus granulosuscyst fluid by immunoblot analysis. Trans. R. Soc. Trop. Med. Hyg. 86, 189–192.
Lightowlers, M.W., Gottstein, B., 1995. Echinococcosis/hydatidosis: antigens, immunological and moleculardiagnosis. In: Thompson, R.C.A., Lymbery, A.J. (Eds.), Echinococcus and Hydatid Disease. CAB International,Wallingford, UK, pp. 354–410.
Lightowlers, M.W., Rickard, M.D., Honey, R.D., Obendorf, D.L., Mitchell, G.F., 1984. Serological diagnosisof Echinococcus granulosusinfection in sheep using cyst fluid antigen processed by antibody affinitychromatography. Aust. Vet. J. 61, 101–108.
Lightowlers, M.W., Dongyou, L., Haralambous, A., Rickard, M.D., 1989. Subunit composition and specificity ofthe major cyst fluid antigens ofEchinococcus granulosus. Mol. Biochem. Parasitol. 37, 171–182.

76 R. Kittelberger et al. / Veterinary Parasitology 110 (2002) 57–76
Lightowlers, M.W., Lawrence, S.B., Gauci, C.G., Young, J., Ralston, M.J., Maas, D., Health, D.D., 1996.Vaccination against hydatidosis using a defined recombinant antigen. Parasite Immunol. 18, 457–462.
Lightowlers, M.W., Jensen, O., Fernandez, E., Iriarte, J.A., Woollard, D.J., Gauci, C.G., Jenkins, D.J., Heath, D.D.,1999. Vaccination trials in Australia and Argentina confirm the effectiveness of the EG95 hydatid vaccine insheep. Int. J. Parasitol. 29, 531–534.
Lightowlers, M.W., Flisser, A., Gauci, C.G., Heath, D.D., Jensen, O., Rolfe, R., 2000. Vaccination againstcysticercosis and hydatid disease. Parasitol. Today 16, 191–196.
Maddison, S.E., Slemenda, S.B., Schantz, P.M., Fried, J.A., Wilson, M., Tsang, V.C., 1989. A specific diagnosticantigen ofEchinococcus granulosuswith an apparent molecular weight of 8 kDA. Am. J. Trop. Med. Hyg. 40,377–383.
Mason, P., Orr, M., 1993. Sarcocystosis and hydatidosis in lamoids—diseases we can do without. Surveillance 20,14.
Montgomery, R.H., van der Logt, P., 1994. The last hydatids outbreak. Surveillance 23, 26–27.Moro, P., Verastegui, M., Gilman, R.H., Falcon, N., Bernal, T., Gavidia, C., Gonzalez, A., Malqui, V., Moro, M.H.,
Dueger, E., 1997. Enzyme-linked immunoelectrotransfer blot assay for diagnosis of hydatidosis (Echinococcusgranulosus) in sheep. Vet. Rec. 140, 605–606.
Reichel, R., Baber, D.J., Craig, P.S., Gasser, R.B., 1996. Cystic echinococcosis in the Falkland Islands. Prev. Vet.Med. 27, 115–123.
Ris, D.R., Hamel, K.L., Mackle, Z.M., 1987. Use of two polysaccharide antigens in ELISA for the detection ofantibodies toEchinococcus granulosusin sheep sera. Res. Vet. Sci. 43, 257–263.
Rogan, M.T., 1998. T-cell activity associated with secondary infections and implanted cysts ofEchinococcusgranulosusin BALB/c mice. Parasite Immunol. 20, 527–533.
Woollard, D.J., Gauci, C.G., Heath, D.D., Lightowlers, M.W., 1998. Epitope specificities and antibody responsesto the EG95 hydatid vaccine. Parasite Immunol. 20, 535–540.
Woollard, D.J., Gauci, C.G., Lightowlers, M.W., 1999. Synthetic peptides induce antibody against a host-protectiveantigen ofEchinococcus granulosus. Vaccine 18, 785–794.
Young, W.K., Heath, D.D., Van Knapen, F., 1984. Comparison of cestode antigens in an enzyme-linkedimmunosorbent assay for the diagnosis ofEchinococcus granulosus, Taenia hydatigena. Res. Vet. Sci. 36,24–31.




















