
2 3 Genetic variations among Echinococcus granulosus isolates in Egypt using RAPD- PCR
10
1 23 Parasitology Research Founded as Zeitschrift für Parasitenkunde ISSN 0932-0113 Volume 111 Number 5 Parasitol Res (2012) 111:1993-2000 DOI 10.1007/s00436-012-3046-z Genetic variations among Echinococcus granulosus isolates in Egypt using RAPD- PCR Hoda A. Taha
Transcript of 2 3 Genetic variations among Echinococcus granulosus isolates in Egypt using RAPD- PCR

1 23
Parasitology ResearchFounded as Zeitschrift fürParasitenkunde ISSN 0932-0113Volume 111Number 5 Parasitol Res (2012) 111:1993-2000DOI 10.1007/s00436-012-3046-z
Genetic variations among Echinococcusgranulosus isolates in Egypt using RAPD-PCR
Hoda A. Taha

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

ORIGINAL PAPER
Genetic variations among Echinococcus granulosus isolatesin Egypt using RAPD-PCR
Hoda A. Taha
Received: 20 May 2012 /Accepted: 10 July 2012 /Published online: 24 July 2012# Springer-Verlag 2012
Abstract Cystic echinococcosis (CE), caused by hydatidcysts, is a widespread and hazardous disease in humansand animals worldwide. The aim of the current study wasto investigate the genetic variations among Echinococcusgranulosus cyst strains isolated from sheep, camel, pig, anddonkey using RAPD-PCR analysis. Seven primers of arbi-trary sequences were used in the PCR reactions. Thescreened primers gave total patterns ranging from 27 to 39reproducible bands for each isolate. Each population isolategave its specific pattern. Although distinct polymorphicpatterns were obtained among the four isolates, there wereseveral shared bands among them in each primer used. Acomparison of the different RAPD-PCR patterns showedthat primers P1, P3, and OPH 04 yielded band patterns thatrevealed a high degree of divergence among the four isolatesof E. granulosus that allowed easy distinction betweenthem. The remaining primers (P2, P4, P5, and OPH14)amplified DNA fragments that were common to two or moreisolates but diversified in the other two or three isolates. Thestudy revealed that the most closely related isolates were ofdonkey and camel where the similarity coefficent betweenthem ranging from 53 % to 78 %, followed by isolates of pigand sheep (sc040 % to 68 %), while the similarity coeffi-cent between isolates of camel and sheep was 33–45 %,between camel and pig was 36 to 57 %, between donkey andpig was 37 to 52 %, and between donkey and sheep was 35to 54 % which means that they more or distant from eachother. In conclusion, hydatid cysts isolated from camel mayhave the genotypic characters of donkey strain.
Introduction
Cystic echinococcosis (CE), caused by Echinococcus gran-ulosus, is an important zoonotic infection causing morbidityand mortality in humans and significant economic losses inlivestock (Budke et al. 2006). Although the definitive host isalmost always a canid, a great many intraspecific variants orstrains of E. granulosus have been described from differentintermediate host species or geographical areas (Thompsonand Lymbery 1988; Eckert and Thompson 1997). E. gran-ulosus is known to exist as biologically distinct subspecificvariants or strains which may vary in their infectivity todomestic animals and man (McManus and Smyth 1986;Thompson 1986).
Evidence is being accumulated that genetic heterogeneityis common within species of the genus Echinococcus andthese intraspecific variants or ‘strains’ may play an impor-tant role with regard to epidemiology, control, pathogenici-ty, and some other aspects (Thompson 1986, 1988;Thompson and Lymbery 1988, 1991; Bowles and McManus1993a; Bowles et al. 1995; Gasser and Chilton 1995;Thompson et al. 1995; Lymbery and Thompson 1996).According to Thompson and Lymbery (1988), a strain ofEchinococcus is a group of individuals which differs statis-tically from other groups of the same species in gene fre-quencies; therefore, this has a significant impact on theepidemiology and control of hydatid disease.
Although morphological and biological studies have pro-vided extremely useful information for strain identification,it has to be considered that these features are variable. Theymay be influenced by host and environmental factors andmay not necessarily reflect distinctness at the genetic level(Hobbs et al. 1990; Bowles and McManus 1993b). Molec-ular techniques allow a direct characterization of the ge-nome of the parasite and have the advantage that they arenot confounded by variability induced by the host or theenvironment (Bowles and McManus 1993b).
H. A. TahaBiology Department, Faculty of Science, Taibah University,KSA, P.O. Box 344, Medina, Saudi Arabia
H. A. Taha (*)Department of Zoology, Faculty of Science, Ain Shams University,Cairo, Egypte-mail: [email protected]
Parasitol Res (2012) 111:1993–2000DOI 10.1007/s00436-012-3046-z
Author's personal copy

Biological and molecular criteria indicate that there aredistinct horse–dog and sheep–dog forms of E. granulo-sus. The sheep strain is genetically uniform and cosmo-politan in its geographical distribution and the horsestrain is genetically similar in several European countries(Ammann and Eckert 1995). In total, nine distinct geno-types or strains of E. granulosus have been describedusing DNA sequence data: G1 (common sheep strain),G2 (Tasmania sheep strain), G3 (buffalo strain), G4(horse strain), G5 (cattle strain), G6 (camel strain), G7(pig strain), G8 (cervid strain), and G9 (human strain)(Bowles et al. 1992; Scott et al. 1997; McManus 2002).Forms in horses and cattle are now considered to representdistinct species E. equinus and E. ortleppi, respectively(Thompson and McManus 2002).
A range of DNA approaches has been used to studygenetic variation within Echinococcus (Thompson et al.1995; McManus and Bowles 1996; Bart et al. 2006; Varcasiaet al. 2007; Šnábel et al. 2009). Traditionally, restrictionfragment length polymorphism (RFLP) analysis of genomicDNA by Southern blotting was used (McManus and Rishi1989; Hope et al. 1991). Moreover, polymerase chain reaction(PCR)-based approaches (Mullis et al. 1986) such as PCR-linked RFLP and arbitrarily primed PCR (AP-PCR) (WelshandMcClelland 1990; Williams et al. 1990) have found broadapplicability because their sensitivity permits the analysis ofgenes from minute amounts of parasite material (e.g., Bowleset al. 1992, 1994; Bowles andMcManus 1993a, c; Siles-Lucaset al. 1993; Gasser and Chilton 1995). The present studyaimed to explore the genetic variability of E. granulosusstrains in Egypt using RAPD-PCR assay to determine thegenetic relationship among and within isolates from camel,donkey, sheep, and pig origin.
Materials and methods
Parasitic material
Cystic samples were obtained from camels, sheep, pigs, anddonkeys. They were sacrificed at a local abattoir (Zagazigabattoir, Al-Sharqueyia and slaughter house of donkey atGiza Zoo, Egypt). Each individual cyst was handled andprocessed as an E. granulosus isolate. Their dissection wascarried out under sterile conditions. The clean, transparenthydatid fluids were aspirated and examined for presence ofprotoscoleces.
Host materials
To avoid cross contamination of parasite DNA with hostDNA, samples of noninfected host tissues were obtainedand DNA extraction was carried out.
DNA extraction
Prior to DNA extraction, the hydatid fluids containing theprotoscoleces (fertile cysts) were precipitated and thenwashed with PBS 1×. Genomic DNA was extracted byCTAB precipitation method (Yap and Thompson 1987;Mostafa et al. 2003). Briefly, tissues were homogenizedindividually in 1.5 ml of lysis buffer (8 % Triton X-100,0.25 M sucrose, 50 mM Tris–HCl, 50 mM EDTA, pH 7.5).Then, freshly prepared proteinase K (1 mg/ml) was addedand the homogenate was incubated at 65 °C in water bathfor 2 h. To precipitate the genomic DNA, 1 ml of a sterile2 % CTAB (cetyltrimethylammonium bromide) solutionwas added to the homogenate and centrifuged at 1,500g.The supernatant was discarded and the precipitate was dis-solved in 0.5 ml of 2.5 M NaCl, 10 mM EDTA, pH 7.7 anddiluted with 1 ml of 40 mM Tris–HCl, 2 mM EDTA, pH 7.7.Two volumes of chloroform were added to the mixture,mixed gently, and centrifuged at 12,000g for 10 min. After-wards, the DNA was precipitated in absolute ethanol, incu-bated at −20 °C overnight and centrifuged at 12,000g for10 min. The DNA pellet was washed with 70 % ethanol anddissolved in 30 μl of TE buffer (10 mM Tris, 1 mM EDTA,pH 8.0). The DNA was stored at −20 °C until used foramplification.
RAPD-PCR
PCR amplifications were performed by the protocol reportedby Williams et al. (1990). The 25 μl mixture containedabout 25 ng of DNA template, 1.5 U of Taq polymerase,10 mM dNTPs, 10 pM primer, and 2.5 μl of 10× PCRbuffer. Amplifications were performed in a T-personalthermal cycler (Biometra, Germany), programmed for 45cycles at 94 °C for 1 min, 35 °C for 1 min, and 72 °Cfor 1 min. An initial denaturation step (3 min at 94 °C)and final extension holding (10 min at 72 °C) wereincluded in the first and last cycles, respectively. Reactionproducts (10 μl) were resolved by 1.5 % agarose gelelectrophoresis at 90 V in 1× TAE buffer. The gel wasstained with ethidium bromide and photographed by adigital camera (Sony, Cyber shot) under UV transillumi-nator. In an initial experiment, 20 decimer oligonucleotideprimers were tested to determine those primers that pro-duced reproducible RAPD patterns. Each primer was test-ed three times. Good and distinct patterns were producedonly using seven arbitrary decimers (as shown in Table 1).
Data analysis
All observed individual bands were scored as present orabsent (1 or 0) for each isolate. Similarity coefficient (S)was calculated according to Nei and Li (1979) and Lynch
1994 Parasitol Res (2012) 111:1993–2000
Author's personal copy
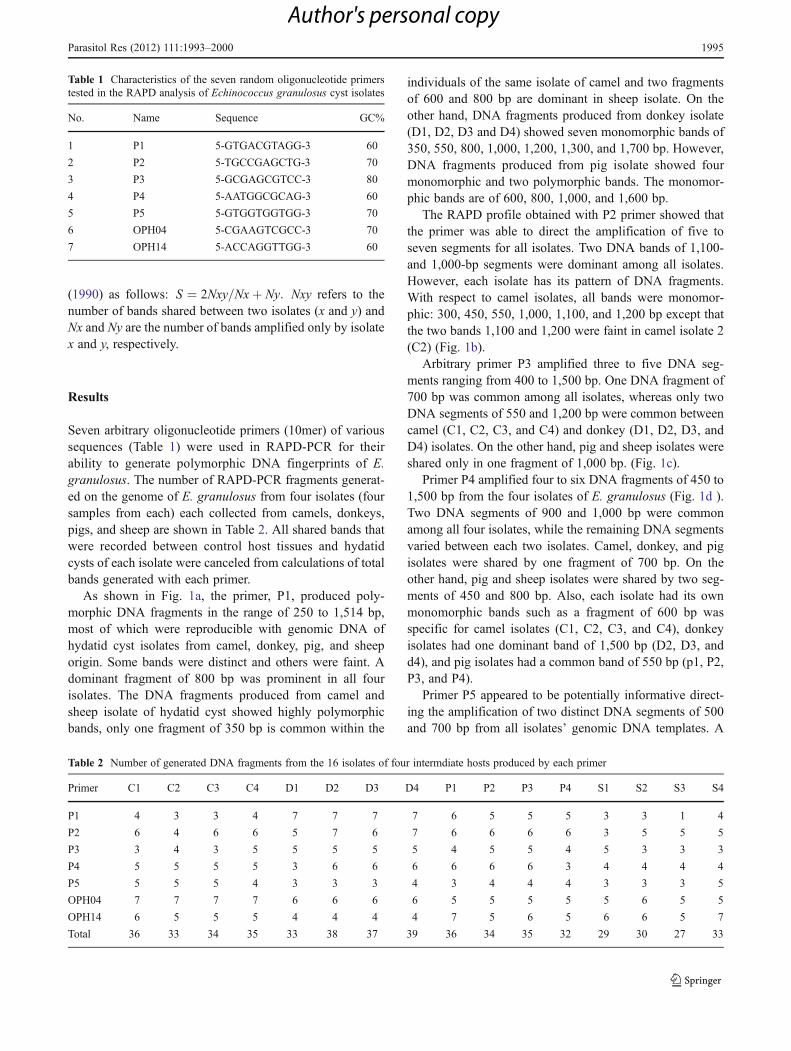
(1990) as follows: S ¼ 2Nxy Nxþ Ny= : Nxy refers to thenumber of bands shared between two isolates (x and y) andNx and Ny are the number of bands amplified only by isolatex and y, respectively.
Results
Seven arbitrary oligonucleotide primers (10mer) of varioussequences (Table 1) were used in RAPD-PCR for theirability to generate polymorphic DNA fingerprints of E.granulosus. The number of RAPD-PCR fragments generat-ed on the genome of E. granulosus from four isolates (foursamples from each) each collected from camels, donkeys,pigs, and sheep are shown in Table 2. All shared bands thatwere recorded between control host tissues and hydatidcysts of each isolate were canceled from calculations of totalbands generated with each primer.
As shown in Fig. 1a, the primer, P1, produced poly-morphic DNA fragments in the range of 250 to 1,514 bp,most of which were reproducible with genomic DNA ofhydatid cyst isolates from camel, donkey, pig, and sheeporigin. Some bands were distinct and others were faint. Adominant fragment of 800 bp was prominent in all fourisolates. The DNA fragments produced from camel andsheep isolate of hydatid cyst showed highly polymorphicbands, only one fragment of 350 bp is common within the
individuals of the same isolate of camel and two fragmentsof 600 and 800 bp are dominant in sheep isolate. On theother hand, DNA fragments produced from donkey isolate(D1, D2, D3 and D4) showed seven monomorphic bands of350, 550, 800, 1,000, 1,200, 1,300, and 1,700 bp. However,DNA fragments produced from pig isolate showed fourmonomorphic and two polymorphic bands. The monomor-phic bands are of 600, 800, 1,000, and 1,600 bp.
The RAPD profile obtained with P2 primer showed thatthe primer was able to direct the amplification of five toseven segments for all isolates. Two DNA bands of 1,100-and 1,000-bp segments were dominant among all isolates.However, each isolate has its pattern of DNA fragments.With respect to camel isolates, all bands were monomor-phic: 300, 450, 550, 1,000, 1,100, and 1,200 bp except thatthe two bands 1,100 and 1,200 were faint in camel isolate 2(C2) (Fig. 1b).
Arbitrary primer P3 amplified three to five DNA seg-ments ranging from 400 to 1,500 bp. One DNA fragment of700 bp was common among all isolates, whereas only twoDNA segments of 550 and 1,200 bp were common betweencamel (C1, C2, C3, and C4) and donkey (D1, D2, D3, andD4) isolates. On the other hand, pig and sheep isolates wereshared only in one fragment of 1,000 bp. (Fig. 1c).
Primer P4 amplified four to six DNA fragments of 450 to1,500 bp from the four isolates of E. granulosus (Fig. 1d ).Two DNA segments of 900 and 1,000 bp were commonamong all four isolates, while the remaining DNA segmentsvaried between each two isolates. Camel, donkey, and pigisolates were shared by one fragment of 700 bp. On theother hand, pig and sheep isolates were shared by two seg-ments of 450 and 800 bp. Also, each isolate had its ownmonomorphic bands such as a fragment of 600 bp wasspecific for camel isolates (C1, C2, C3, and C4), donkeyisolates had one dominant band of 1,500 bp (D2, D3, andd4), and pig isolates had a common band of 550 bp (p1, P2,P3, and P4).
Primer P5 appeared to be potentially informative direct-ing the amplification of two distinct DNA segments of 500and 700 bp from all isolates’ genomic DNA templates. A
Table 1 Characteristics of the seven random oligonucleotide primerstested in the RAPD analysis of Echinococcus granulosus cyst isolates
No. Name Sequence GC%
1 P1 5-GTGACGTAGG-3 60
2 P2 5-TGCCGAGCTG-3 70
3 P3 5-GCGAGCGTCC-3 80
4 P4 5-AATGGCGCAG-3 60
5 P5 5-GTGGTGGTGG-3 70
6 OPH04 5-CGAAGTCGCC-3 70
7 OPH14 5-ACCAGGTTGG-3 60
Table 2 Number of generated DNA fragments from the 16 isolates of four intermdiate hosts produced by each primer
Primer C1 C2 C3 C4 D1 D2 D3 D4 P1 P2 P3 P4 S1 S2 S3 S4
P1 4 3 3 4 7 7 7 7 6 5 5 5 3 3 1 4
P2 6 4 6 6 5 7 6 7 6 6 6 6 3 5 5 5
P3 3 4 3 5 5 5 5 5 4 5 5 4 5 3 3 3
P4 5 5 5 5 3 6 6 6 6 6 6 3 4 4 4 4
P5 5 5 5 4 3 3 3 4 3 4 4 4 3 3 3 5
OPH04 7 7 7 7 6 6 6 6 5 5 5 5 5 6 5 5
OPH14 6 5 5 5 4 4 4 4 7 5 6 5 6 6 5 7
Total 36 33 34 35 33 38 37 39 36 34 35 32 29 30 27 33
Parasitol Res (2012) 111:1993–2000 1995
Author's personal copy
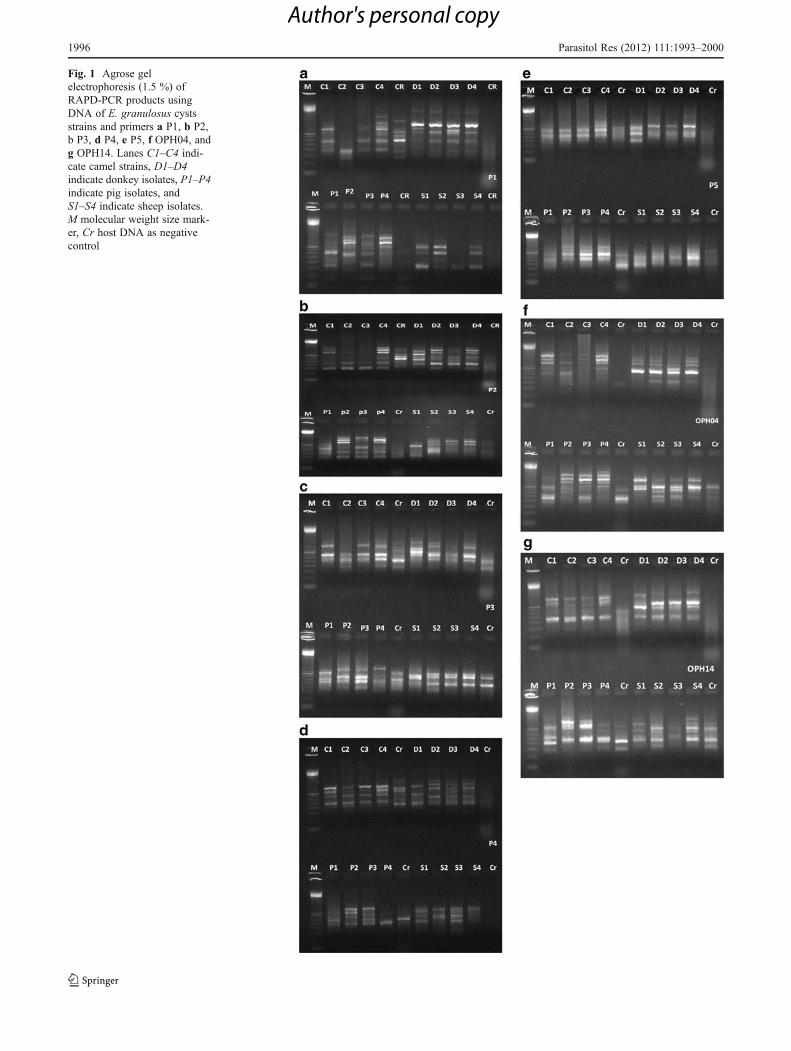
Fig. 1 Agrose gelelectrophoresis (1.5 %) ofRAPD-PCR products usingDNA of E. granulosus cystsstrains and primers a P1, b P2,b P3, d P4, e P5, f OPH04, andg OPH14. Lanes C1–C4 indi-cate camel strains, D1–D4indicate donkey isolates, P1–P4indicate pig isolates, andS1–S4 indicate sheep isolates.M molecular weight size mark-er, Cr host DNA as negativecontrol
1996 Parasitol Res (2012) 111:1993–2000
Author's personal copy

monomorphic DNA segment of 900 bp was present incamel, donkey, and pig isolates (Fig. 1e). The 300- and1,000-bp segments were only present in sheep isolate. Also,the 400-bp fragment was found only in camel isolate.
Results obtained with the OPH 04 primer showedless polymorphic DNA. This primer produced a numberof fragment ranging from five to eight DNA fragmentsof 300 to 1,400 bp (Fig. 1f). Two common DNA seg-ments of 900 and 1,000 bp were detected among allfour isolates and one dominant fragment were alsopresent (600 bp), while the remaining DNA segmentsvaried between each two and/or three isolates. Camel,donkey, and pig isolates were shared by one fragmentof 1,000 bp, while camel, donkey, and sheep isolateswere shared a 500 bp band. On the other hand, cameland donkey isolates were shared by one segment of1,100 bp. Also, each pig and sheep isolates had theirown monomorphic bands (1,300 and 1,400 bp for pigisolate and 900 bp for sheep isolate).
OPH14 primer generated from four to seven fragments,three of them (500, 700, and 800 bp) were monomorphicamong nearly all four isolates (Fig. 1g). However, pig andsheep isolates were shared by two fragments (300 and350 bp), and pig and camel were shared by a fragment of1,000 bp. On the other hand, specific fragments were en-countered for each isolate. Camel isolate had a specific bandof 400 bp, donkey isolate had a fragment of 1,100 bp, andsheep isolate had a specific fragment of 650 bp.
Collectively, a comparison of the different RAPD-PCR patterns showed that primers P1, P3, and OPH04 yielded band patterns that revealed a high degreeof divergence among the four isolates of E. granulosusthat allowed easy distinction between them. The remain-ing primers (P2, P4, P5, and OPH14) amplified DNAfragments that were common to two isolates but diver-sified in the other two or three isolates. Nevertheless,from all primers, each isolate has its own specific frag-ments as follows: camel and donkey isolates each hasfive specific fragments, pig isolate with seven specificfragments, and sheep isolate had three specific frag-ments. Also, the shared bands between each two isolateswere listed in Tables 3 and 4.
Concerning the similarity coefficients (SC) between indi-viduals of the same isolate or different isolates, data pre-sented in Tables 2 and 3 showed that the mean range of SCswithin individuals in the same isolate was 80–98 %. Thehighest SC was among isolates of sheep (mean SC092 %),followed by pig and camel isolates (mean SC087.5 %) andthen donkey isolates (mean SC087).
On the other hand, the mean range of SCs among differ-ent isolates was 33–78 %. It should be mentioned that E.granulosus isolated from camel and donkey was more close-ly related than other isolates (mean SC062 %).
Discussion
In the present work, all the collected hydatid cysts fromcamels, donkeys, pigs, and sheep were fertile in contrastwith a previous study by Abdel Aaty et al. (2012) wherethey didn’t find fertile sheep hydatid cysts during the periodof their study. This is of great epidemiological importance asthe fertile hydatid cysts are responsible for progression ofthe life cycle and therefore, acting as a reservoir for humanCE (Dyab et al. 2005).
It is well-known that six genotypes of the E. gran-ulosus complex have been identified in Africa, fromthem the sheep strains (G1 and G2 genotype), the horsestrain (G4 genotype), and the camel strain (G6 geno-type) (Anderson et al. 1997; Macpherson and Wachira1997; Sadjjadi 2006). On the other hand, Bart et al.(2006) demonstrated that three strains were identifiedbased on the mitochondrial sequences: (i) the commonsheep strain (G1) which circulates between sheep andcattle and is infective for humans, (ii) the Tasmanian sheep
Table 3 Number of shared bands (SB) among individuals from thesame isolate with calculation of similarity coefficient (SC) and geneticdistance (GD)
Isolates SB SC% GD%
C1 and C2 29 87 13
C1 and C3 32 91.5 8.5
C1 and C4 32 90 10
C2 and C3 30 89.5 10.5
C2 and C4 27 80 20
C3 and C4 30 87 13
D1 and D2 29 82 18
D1 and D3 29 83 17
D1 and D4 29 80 20
D2 and D3 35 93 7
D2 and D4 36 93.5 6.5
D3 and D4 35 92 8
P1 and P2 29 86 14
P1 and P3 30 84.5 15.5
P1 and P4 25 73.5 26.5
P2 and P3 32 90 10
P2 and P4 32 96 4
P3 and P4 32 95 5
S1 and S2 26 88 12
S1 and S3 24 86 14
S1 and S4 28 90 10
S2 and S3 27 94 6
S2 and S4 31 98 2
S3 and S4 28 93 7
C camel isolate, D donkey isolate, P pig isolate, S sheep isolate
Parasitol Res (2012) 111:1993–2000 1997
Author's personal copy

Table 4 Number of sharedbands between each two isolates(SB) with calculation ofsimilarity coefficients (SC) andgenetic differences (GD)
C camel isolate, D donkeyisolate, P pig isolate, S sheepisolate
Isolate SB SC% GD% Isolate SB SC% GD%
C1 and D1 20 58 42 D1 and P1 16 46 54
C1 and D2 27 73 27 D1 and P2 17 50 50
C1 and D3 26 71 29 D1 and P3 16 47 53
C1 and D4 27 72 28 D1 and P4 14 43 57
C2 and D1 19 58 42 D2 and P1 15 40 60
C2 and D2 20 56 44 D2 and P2 19 52 48
C2 and D3 19 54 46 D2 and P3 18 49 51
C2 and D4 19 53 47 D2 and P4 14 40 60
C3 and D1 20 60 40 D3 and P1 16 43 57
C3 and D2 26 72 28 D3 and P2 18 49 51
C3 and D3 25 70 30 D3 and P3 17 47 53
C3 and D4 26 71 29 D3 and P4 13 37 63
C4 and D1 21 62 38 D4 and P1 17 45 55
C4 and D2 28 76 24 D4 and P2 19 52 48
C4 and D3 26 72 28 D4 and P3 19 51 49
C4 and D4 29 78 22 D4 and P4 14 39 61
C1 and P1 19 55 45 D1 and S1 13 41 59
C1 and P2 20 57 43 D1 and S2 15 47 53
C1 and P3 20 56 44 D1 and S3 15 50 50
C1 and P4 14 41 59 D1 and S4 18 54 46
C2 and P1 19 55 45 D2 and S1 15 45 55
C2 and P2 16 48 52 D2 and S2 15 44 56
C2 and P3 15 44 56 D2 and S3 15 46 54
C2 and P4 11 32 68 D2 and S4 18 50 50
C3 and P1 18 51 49 D3 and S1 12 36 64
C3 and P2 18 53 47 D3 and S2 16 47 53
C3 and P3 18 52 48 D3 and S3 15 44 56
C3 and P4 12 36 64 D3 and S4 18 51 49
C4 and P1 16 45 55 D4 and S1 12 35 65
C4 and P2 19 55 45 D4 and S2 15 43 57
C4 and P3 18 51 49 D4 and S3 15 45 55
C4 and P4 9 36 64 D4 and S4 18 50 50
C1 and S1 13 40 60 P1 and S1 16 49 51
C1 and S2 15 45 55 P1 and S2 21 63 37
C1 and S3 13 41 59 P1 and S3 17 54 46
C1 and S4 15 43 57 P1 and S4 21 60 40
C2 and S1 11 35 65 P2 and S1 18 57 43
C2 and S2 13 41 59 P2 and S2 22 68 32
C2 and S3 10 33 67 P2 and S3 16 52 48
C2 and S4 12 36 64 P2 and S4 22 65 35
C3 and S1 11 35 65 P3 and S1 17 53 47
C3 and S2 13 40 60 P3 and S2 21 65 35
C3 and S3 12 39 61 P3 and S3 16 51 49
C3 and S4 13 38 62 P3 and S4 22 65 35
C4 and S1 11 34 66 P4 and S1 15 50 50
C4 and S2 12 36 64 P4 and S2 18 58 42
C4 and S3 11 33 67 P4 and S3 12 40 60
C4 and S4 13 38 62 P4 and S4 19 58 42
1998 Parasitol Res (2012) 111:1993–2000
Author's personal copy

strain (G2) infecting sheep and cattle, and (iii) the pig strain(G7) predominantly found in swine.
The presence of intraspecific variants in E. granulosushas been described from different geographical areas andintermediate hosts. These strains often differ in a number ofbiological characteristics which can influence the epidemi-ology of hydatidosis and, in turn, the measures to be takenfor diagnosis, treatment, and control of the disease (Reddy etal. 1998). The identification of such strains is considered animportant prerequisite to hydatid control and a number ofdifferent approaches are in use to realize this objective.RAPD-PCR data presented in this study revealed variationsamong the four isolates under study with several fixedalleles in each isolate in addition to the noticed variationwithin the same isolate. RAPD or AP-PCR is a DNA fin-gerprinting method used to readily produce complex andinformative genomic fingerprints (Gasser 2006) and hasproven useful for the definition of genetic markers for awide range of species and strains of protozoan and hel-minths (Gasser 1999). In the present study, RAPD analysisof 16 samples of E. granulosus recovered from four inter-mediate hosts with seven arbitrary primers indicated intra-genotype variability between different host species. In astudy from Spain, polymorphism had been found with twoof three primers in RAPD analysis of sheep, equine, and pigisolates (Daniel et al. 2004). RAPD-PCR with two primerson 21 pigs and two human patients harboring G7 genotypefrom Slovak Republic showed two discernible patterns inpig isolates and two distinct profiles in human isolates thatwere verified with nad1 sequencing and isoenzyme analysesand indicated intragenotype variation within the G7 genotype(Turcekova et al. 2003). Slight genetic variations in the cyto-chrome c oxidase subunit 1 (cox1) gene of G1 genotype (G1Ato G1E) was described in Argentina after analyzing isolates ofdifferent hosts and geographical areas (Kamenetzky et al.2002). Recently Bhattacharya et al. (2008) reported two (of26) random primers suitable for differentiating sheep, cattle,and buffalo isolates. Although they did not observe any dif-ference on ITS1-RFLP of all isolates using RsaI and Msp1restriction enzymes.
Clear differences could be observed in the number andsize of randomly amplified DNA fragments for each primeramong four isolates. A typical RAPD amplification gener-ally yielded three to seven scorable and reproducible DNAfragments in the four isolates, while the sheep isolate (gen-erated by pP1) showed only one to four fragments depend-ing upon the primer–template combination. Under identicalamplification conditions, these differences in the specificamplification of DNA fragments within sheep isolates sug-gest a possible heterogeneity of the genotype and/or thepresence of multiple genetically distinct populations. Asexplained by Reddy et al. (1998), the ability of the primersto selectively amplify one or two DNA elements more
efficiently than others in the same reaction may be due topriming within the repetitive region of the parasite genomewhich would result in more copies being produced in thePCR.
In a previous study using RAPD-PCR for characteriza-tion of human and animal hydatid cysts, it has been shownthat human and camel isolates were the most related pair.The study implies that human cases in Egypt are of thecamel/dog strain, and camels are important hosts for thetransmission of human hydatidosis (Azab et al. 2004). Inour finding, the most related isolates are camel and donkeys.This may give an impression that the transmission cycle isassociated between human and camel origins on the onehand and may be of human–donkey origin on the otherhand. Further studies are needed to investigate the relationof E. granulosus strain of donkey origin and their relation totransmission cycle to human.
References
Abdel Aaty HE, Abdel-Hameed DM, Alam-Eldin YH, El-ShennawySF, Aminou HA, Makled SS, Darweesh SK (2012) Moleculargenotyping of Echinococcus granulosus in animal and humanisolates from Egypt. Acta Trop 121(2):125–128
Ammann R, Eckert R (1995) Clinical diagnosis and treatment ofechinococcosis in humans, Echinococcus and hydatid disease.CAB International, Wallingford
Anderson FL, Ouhelli H, Kachemi M (1997) Compendium on cysticechinococcosis in Africa and Middle Eastern countries with spe-cial reference to Morocco. Brigham Young University Print Serv-ices, Provo
Azab ME, Bishara SA, Helmy H, Oteifa NM, El-Hoseiny LM, RamzyRM, Ahmed MA (2004) Molecular characterization of Egyptianhuman and animal Echinococcus granulosus isolates by RAPD-PCR technique. J Egypt Soc Parasitol 34:83–96
Bart JM, Morariu S, Knapp J, Ilie MS, Pitulescu M, Anghel A,Cosoroaba I, Piarroux R (2006) Genetic typing of Echinococcusgranulosus in Romania. Parasitol Res 98(2):130–137
Bhattacharya D, Bera AK, Bera BC, Pan D, Das SK (2008) Molecularappraisal of Indian animal isolates of Echinococcus granulosus.Indian J Med Res 127:383–387
Bowles J, McManus DP (1993a) Rapid discrimination of Echinococ-cus species and strains using a polymerase chain reaction-basedRFLP method. Mol Biochem Parasitol 57:231–240
Bowles J, McManus DP (1993b) Molecular variation in echinococcus.Acta Trop 53:291–305
Bowles J, McManus DP (1993c) NADH dehydrogenase 1 genesequences compared for species and strains of the genus Echino-coccus. Int J Parasitol 23:969–972
Bowles J, Blair D, Mc Manus DP (1992) Genetic variants within thegenus Echinococcus identified by mitochondrial DNA sequenc-ing. Mol Biochem Parasitol 54:165–174
Bowles J, Blair D, McManus DP (1994) Molecular genetic character-ization of the cervid strain (northern form) of Echinococcusgranulosus. Parasitology 109:215–221
Bowles J, Blair D, McManus DP (1995) A molecular phylogeny of thegenus Echinococcus. Parasitology 110:317–328
Budke CM, Deplazes P, Torgerson PR (2006) Global socioeconomicimpact of cystic echinococcosis. Emerg Infect Dis 12:296–303
Parasitol Res (2012) 111:1993–2000 1999
Author's personal copy

Daniel MK, Ponce-Gordo F, Cuesta-Bandera C (2004) Genetic identi-fication and host range of the Spanish strains of Echinococcusgranulosus. Acta Trop 91:87–93
Dyab KA, Hassanein R, Hussein AA, Metwally SE, Gaad HM (2005)Hydatidosis among man and animals in Assiut and Aswan gov-ernorates. J Egypt Soc Parasitol 35(1):157–166, 258
Eckert J, Thompson RCA (1997) Intraspecific variation of Echinococ-cus granulosus and related species with emphasis on their infec-tivity to humans. Acta Trop 64:19–34
Gasser RB (1999) PCR-based technology in veterinary parasitology.Vet Parasitol 84:229–258
Gasser RB (2006) Molecular tools advances, opportunities and pros-pects. Vet Parasitol 136:69–89
Gasser RB, Chilton NB (1995) Characterization of taeniid cestodespecies by PCR-RFPL of IST2 ribosomal DNA. Acta Trop59:31–40
Hobbs RP, Lymbery AJ, Thompson RCA (1990) Rostellar hook mor-phometry of Echinococcus granulosus (Batsch, 1786) from natu-ral and experimental Australian hosts, and its implication.Parasitology 101:273–281
Hope M, Bowles J, McManus DP (1991) Reconsideration of theEchinococcus granulosus strain situation in Australia followingRFLP analysis of cystic material. Int J Parasitol 21:471–475
Kamenetzky L, Gutierrez AM, Canova SG, Haag KL, Guarnera EA,Parra A, Garcia GE, Rosenzvit MC (2002) Several strains ofEchinococcus granulosus infect livestock and humans in Argen-tina. Infect Genet Evol 2:129–136
Lymbery AJ, Thompson RCA (1996) Species of Echinococcus: patternand process. Parasitol Today 12:486–491
Lynch M (1990) The similarity index and DNA fingerprinting. MolBiol Evol 5:584–599
Macpherson NL, Wachira TWM (1997) Cystic echinococcosis inAfrica south of the Sahara. In: Anderson FL, Ouhelli H, KachaniM (eds) Compendium on cystic echinococcosis in Africa and inMiddle Eastern countries with special reference to Morocco.Brigham Young University, Print Services, Provo, pp 245–277
McManus DP (2002) The molecular epidemiology of Echinococcusgranulosus and cystic hydatid disease. Trans R Soc Trop MedHyg 96:151–157
McManus DP, Bowles J (1996) Molecular genetic approaches to par-asite identification: their value in diagnostic parasitology andsystematics. Int J Parasitol 26:687–704
McManus DP, Rishi AK (1989) Genetic heterogeneity within Echino-coccus granulosus: isolate from different hosts and geographicalareas characterized with DNA probes. Parasitology 99:17–29
McManus DP, Smyth JD (1986) Hydatidosis: changing concepts inepidemiology and speciation. Parasitol Today 2:163–168
Mostafa OMS, Taha HA, Ramadan G (2003) Diagnosis of Fasciolagigantica in snail using the polymerase chain reaction (PCR)assay. J Egypt Soc Parasitol 33(3):733–742
Mullis KB, Faloona F, Scharf S, Saiki R, Horn G, Erlich H (1986)Specific enzymatic amplification of DNA in vitro: the polymerase
chain reaction. Cold Spring Harb Symp Quant Biol 51:263–273
Nei M, Li WS (1979) Mathematical model for studying genetic vari-ation in terms of restriction endonucleases. Proc Natl Acad SciUSA 76:5269–5273
Reddy YA, Rao JR, Butchaiahb GB, Sharma B (1998) Random am-plified polymorphic DNA for the specific detection of bubalineEchinococcus granulosus by hybridization assay. Vet Parasitol79:315–323
Sadjjadi S (2006) Present situation of echinococcosis in the MiddleEast and Arabic North Africa. Parasitol Int 55(Suppl):S197–S202
Scott JC, Stefaniak J, Pawlowski ZS, McManus DP (1997) Moleculargenetic analysis of human cystic group (G9) of Echinococcusgranulosus. Parasitology 114:37–43
Siles-lucas M, Cuesta-Bandera C, Cesar-Bento M (1993) Randomamplified polymorphic DNA technique for speciation studies ofEchinococcus granulosus. Parasitol Res 79:343–345
Šnábel V, Altintas N, D’Amelio S, Nakao M, Romig T, Yolasigmaz A,Gunes K, Turk M, Busi M, Hüttner M et al (2009) Cystic echi-nococcosis in Turkey: genetic variability and first record of thepig strain (G7) in the country. Parasitol Res 105(1):145–154
Thompson RCA (1986) Biology and systematics of Echinococcus. In:Thompson RCA (ed) The biology of Echinococcus and hydatiddisease. G. Allen, London, pp 5–43
Thompson RCA (1988) Intraspecific variation and epidemiology. In:Mehlhorn H (ed) Parasitology in focus. Springer, Berlin, pp 391–411
Thompson RCA, Lymbery AJ (1988) The nature, extent and signifi-cance of variation within the genus Echinococcus. Adv Parasitol27:209–258
Thompson RCA, Lymbery AJ (1991) The epidemiological signifi-cance of biological variation in Echinococcus. Arch Hydatid30:195–200
Thompson RCA, McManus DP (2002) Towards a taxonomic revisionof the genus Echinococcus. Trends Parasitol 18:452–457
Thompson RCA, Lymbery AJ, Constantine CC (1995) Variation inEchinococcus: towards a taxonomic revision of the genus. AdvParasitol 35:145–176
Turcekova L, Snabel V, D’Amelio S, Busi M, Dubinsky P (2003)Morphological and genetic characterization of Echinococcusgranulosus in the Slovak Republic. Acta Trop 85:223–229
Varcasia A, Canu S, Kogkos A, Pipia AP, Scala A, Garippa G, SeimenisA (2007) Molecular characterization of Echinococcus granulosus insheep and goats of Peloponnesus, Greece. Parasitol Res 101(4):1135–1139
Welsh J, McClelland M (1990) Fingerprinting genomes using PCRwith arbitrary primers. Nucleic Acid Res 18:7213–7218
Williams JGK, Kubelik AR, Livak KJ, Rafalski JA, Tingey SV (1990)DNA polymorphisms amplified by arbitrary primers are useful asgenetic markers. Nucleic Acid Res 18(22):6531–6535
Yap KW, Thompson RCA (1987) CTAB precipitation of cestode DNA.Parasitol Today 3(7):220–222
2000 Parasitol Res (2012) 111:1993–2000
Author's personal copy




















