
STUDIES ON EFFICIENCY OF RAPD PRIMERS IN DEVELOPINGMOLECULAR PROFILES FOR GENETIC PURITY STUDIES IN...
12
___________________________ Corresponding author: Sanjay Kumar, Genetics Laboratory, Department of Zoology, Banaras Hindu University, Varanasi 225001, India (Phone: +91 7860448788; E-mail: [email protected] ) UDC 575:633 DOI: 10.2298/GENSR1403681K Original scientific paper STUDIES ON EFFICIENCY OF RAPD PRIMERS IN DEVELOPING MOLECULAR PROFILES FOR GENETIC PURITY STUDIES IN SOYBEAN (Glycine max L.) CULTIVARS Sanjay KUMAR Genetics Laboratory, Department of Zoology, Banaras Hindu University, Varanasi, India Kumar S. (2014): Studies on efficiency of RAPD primers in developing molecular profiles for genetic purity studies in soybean (Glycine max L.) cultivars. - Genetika, Vol 46, No. 3, 681- 692. Major advances have recently been made in our understanding of soybean genetics and of the application of new technologies to soybean improvement. Estimates of genetic relationships on the basis of the enzymes and molecular markers have been shown to be consistent with expectations based on origin and pedigree information. To identify efficient markers, that are to be used for genetic purity studies, polymorphism is the basic criterion. RAPD has been found to be an effective and efficient tool to evaluate and reveal genetic polymorphism in several crop species. In present study a total of 80 RAPD primers were screened, out of which 37 gave amplification and only 30 primers showed unambiguous DNA profile. Out of these 30 primers, 22 gave polymorphic banding patterns. It is evident from the result that, 30/80 primers tried (38%) provided unambiguous amplification and out of 30 primers, 22 primers (73%) were found to be polymorphic. Scorable 30 RAPD primers led to amplification of 120 fragments out of which 81 (67.5%) bands were found to be polymorphic. On an average, we got 4 bands per primer and 15 primers (50%) have been found to produce more number of bands than the average value which is encouraging. 8 primers were found to give 100% polymorphisms. Our results are indicative of the efficiency of RAPD primers towards development of molecular profiles. Key words: Glycine max, Polymorphism, PCR, RAPD, UPGMA INTRODUCTION Soybean (Glycine max (L.) Merr.) belongs to the family Leguminosae. It is an annual herbaceous leguminous crop, with a high rain protein content and easy adaptation to diverse soil-
Transcript of STUDIES ON EFFICIENCY OF RAPD PRIMERS IN DEVELOPINGMOLECULAR PROFILES FOR GENETIC PURITY STUDIES IN...

___________________________
Corresponding author: Sanjay Kumar, Genetics Laboratory, Department of Zoology, Banaras
Hindu University, Varanasi 225001, India (Phone: +91 7860448788; E-mail:
UDC 575:633
DOI: 10.2298/GENSR1403681K
Original scientific paper
STUDIES ON EFFICIENCY OF RAPD PRIMERS IN DEVELOPING MOLECULAR
PROFILES FOR GENETIC PURITY STUDIES IN SOYBEAN (Glycine max L.)
CULTIVARS
Sanjay KUMAR
Genetics Laboratory, Department of Zoology, Banaras Hindu University,
Varanasi, India
Kumar S. (2014): Studies on efficiency of RAPD primers in developing
molecular profiles for genetic purity studies in soybean (Glycine max L.) cultivars. -
Genetika, Vol 46, No. 3, 681- 692.
Major advances have recently been made in our understanding of soybean
genetics and of the application of new technologies to soybean improvement. Estimates of
genetic relationships on the basis of the enzymes and molecular markers have been shown
to be consistent with expectations based on origin and pedigree information. To identify
efficient markers, that are to be used for genetic purity studies, polymorphism is the basic
criterion. RAPD has been found to be an effective and efficient tool to evaluate and reveal
genetic polymorphism in several crop species. In present study a total of 80 RAPD
primers were screened, out of which 37 gave amplification and only 30 primers showed
unambiguous DNA profile. Out of these 30 primers, 22 gave polymorphic banding
patterns. It is evident from the result that, 30/80 primers tried (38%) provided
unambiguous amplification and out of 30 primers, 22 primers (73%) were found to be
polymorphic. Scorable 30 RAPD primers led to amplification of 120 fragments out of
which 81 (67.5%) bands were found to be polymorphic. On an average, we got 4 bands
per primer and 15 primers (50%) have been found to produce more number of bands than
the average value which is encouraging. 8 primers were found to give 100%
polymorphisms. Our results are indicative of the efficiency of RAPD primers towards
development of molecular profiles.
Key words: Glycine max, Polymorphism, PCR, RAPD, UPGMA
INTRODUCTION
Soybean (Glycine max (L.) Merr.) belongs to the family Leguminosae. It is an annual
herbaceous leguminous crop, with a high rain protein content and easy adaptation to diverse soil-

682 GENETIKA, Vol. 46, No.3, 681-692, 2014
climatic conditions. Being one of the main oil crops of the world, in India it is grown in all its
major states viz. Madhya Pradesh, Uttar Pradesh, Maharashtra, Rajasthan and Gujarat. It is also
grown on a small scale in Himachal Pradesh, Punjab and Delhi. Major advances have recently
been made in our understanding of soybean genetics and of the application of new technologies to
soybean improvement. Thus it is now possible, using molecular methods, to alter the protein and
oil composition of soybean, as well as to produce other foreign proteins in the plant. Traditionally,
the study of genetic diversity has fallen within population genetics which has focussed on
measuring its extent in natural populations, in comparing levels of genetic diversity within and
among populations and in making references on the nature and intensity of evolutionary processes
from the observed patterns of genetic diversity. Hence, there is a long tradition as well as a wealth
of conceptual tools in population genetics for analyzing, measuring and partitioning genetic
diversity (KUMAR and SINGH 2014a; b).
Several thousand soybean accessions from the USDA Soybean Germplasm Collection
(Urbana, IL) have been evaluated for agronomic and seed composition traits as well as disease
resistance (NELSON et al., 1987; NELSON et al., 1989; JUVIK et al., 1989; BERNARD et al., 1989)
although a large number of G. max lines are available to soybean breeding programs, discovering
and transferring novel, favourable genes from G. soja into G. max could be a successful
approach
for enhancing the genetic variability and improving cultivated soybean (CARPENTER and FEHR,
1986). Identifying useful
diversity is a challenge. Molecular characterization of soybean
germplasm has been performed with isozymes, proteins, and DNA markers. Estimates of genetic
relationships on the basis of the enzymes and molecular markers have been shown to be consistent
with expectations based on origin and pedigree information (GRIFFIN and PALMER, 1995;
MAUGHAN et al., 1995, 1996; DOLDI et al., 1997; THOMPSON et al., 1998). KEIM et al. (1989)
surveyed 58 G. max and G. soja accessions with 17 restriction fragment length polymorphism
(RFLP) markers and found that the molecular diversity was the least among cultivated
soybeans
and greatest between species. Using simple sequence repeat (SSR) and amplified sequence length
polymorphism (ASLP) markers MAUGHAN et al. (1995) reported that five microsatellite
markers
detected a total of 79 alleles in a sample of 94 accessions of wild and cultivated soybean. Allelic
diversity for the SSR loci was greater in wild soybean than in cultivated soybean.
Overall, 43 more
SSR alleles were detected in wild than in cultivated soybean. MAUGHAN et al. (1996) evaluated 23
accessions of wild and cultivated soybean using amplified fragment length polymorphism
(AFLP).
Among the 759 AFLP fragments scored, 17% were polymorphic in G. max and 31% were
polymorphic in G. soja. Their results also indicated that AFLP phenotypic variation was greater in
wild soybean than in cultivated soybean. GRIFFIN and PALMER (1995) screened more than 1200 G.
max and G. soja plant introductions with eight enzymes. The numbers of alleles per locus and
average gene diversity were greater in the G. soja samples than in the
G. max samples.
MATERIALS AND METHODS
Plant material
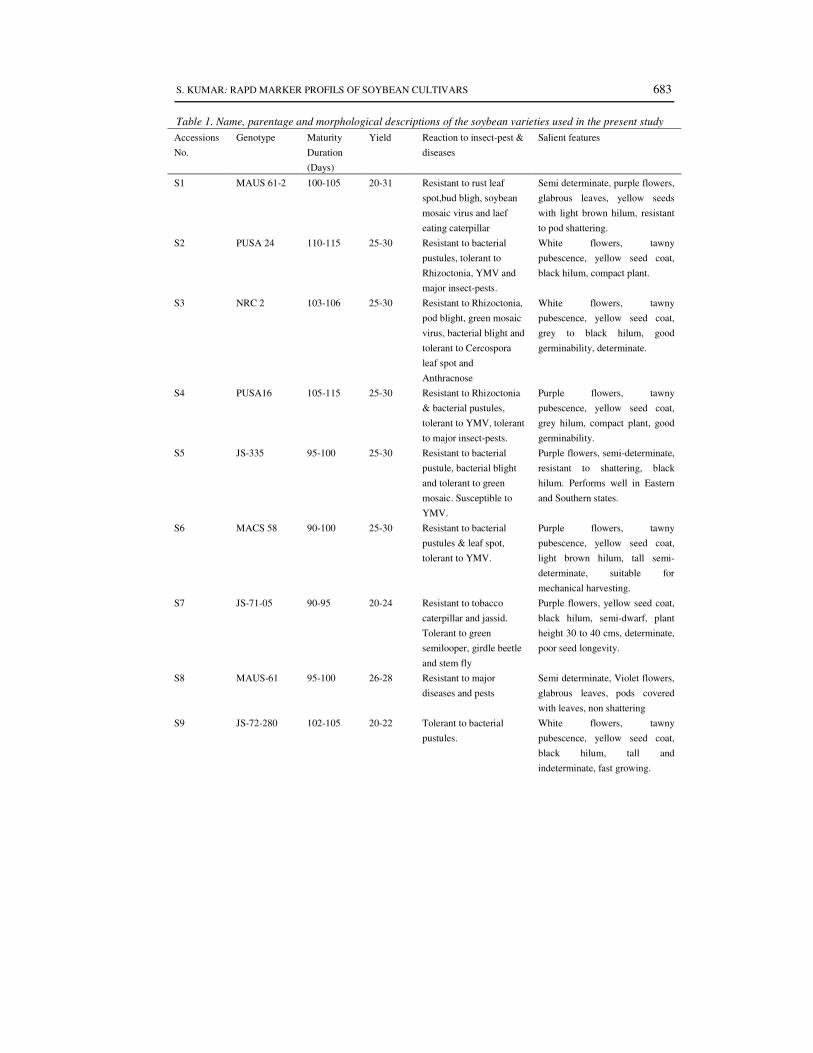
A total of 24 elite released cultivars of soybean grown across India and representing
different morphological variations, agro climatic zones and parentage were selected for molecular
polymorphism studies (Table 1). All the cultivars were collected from concerned breeder working
on soybean in SAUs or ICAR institute in India.
S. KUMAR: RAPD MARKER PROFILS OF SOYBEAN CULTIVARS 683
Table 1. Name, parentage and morphological descriptions of the soybean varieties used in the present study
Accessions
No.
Genotype Maturity
Duration
(Days)
Yield Reaction to insect-pest &
diseases
Salient features
S1 MAUS 61-2 100-105 20-31 Resistant to rust leaf
spot,bud bligh, soybean
mosaic virus and laef
eating caterpillar
Semi determinate, purple flowers,
glabrous leaves, yellow seeds
with light brown hilum, resistant
to pod shattering.
S2 PUSA 24 110-115 25-30 Resistant to bacterial
pustules, tolerant to
Rhizoctonia, YMV and
major insect-pests.
White flowers, tawny
pubescence, yellow seed coat,
black hilum, compact plant.
S3 NRC 2 103-106 25-30 Resistant to Rhizoctonia,
pod blight, green mosaic
virus, bacterial blight and
tolerant to Cercospora
leaf spot and
Anthracnose
White flowers, tawny
pubescence, yellow seed coat,
grey to black hilum, good
germinability, determinate.
S4 PUSA16 105-115 25-30 Resistant to Rhizoctonia
& bacterial pustules,
tolerant to YMV, tolerant
to major insect-pests.
Purple flowers, tawny
pubescence, yellow seed coat,
grey hilum, compact plant, good
germinability.
S5 JS-335 95-100 25-30 Resistant to bacterial
pustule, bacterial blight
and tolerant to green
mosaic. Susceptible to
YMV.
Purple flowers, semi-determinate,
resistant to shattering, black
hilum. Performs well in Eastern
and Southern states.
S6 MACS 58 90-100 25-30 Resistant to bacterial
pustules & leaf spot,
tolerant to YMV.
Purple flowers, tawny
pubescence, yellow seed coat,
light brown hilum, tall semi-
determinate, suitable for
mechanical harvesting.
S7 JS-71-05 90-95 20-24 Resistant to tobacco
caterpillar and jassid.
Tolerant to green
semilooper, girdle beetle
and stem fly
Purple flowers, yellow seed coat,
black hilum, semi-dwarf, plant
height 30 to 40 cms, determinate,
poor seed longevity.
S8 MAUS-61 95-100 26-28 Resistant to major
diseases and pests
Semi determinate, Violet flowers,
glabrous leaves, pods covered
with leaves, non shattering
S9 JS-72-280 102-105 20-22 Tolerant to bacterial
pustules.
White flowers, tawny
pubescence, yellow seed coat,
black hilum, tall and
indeterminate, fast growing.

684 GENETIKA, Vol. 46, No.3, 681-692, 2014
S10 KALITURE 120-130 18-20 Susceptible to soybean
mosaic, tolerant to
bacterial pustules
Purple flowers, tawny pubescen,
black seed coat, black hilum,
small seeded semi indeterminate
S11 PUNJAB 1 95-100 25-30 Susceptible to bacterial
pustules, less susceptible
to blue beetle.
Purple flowers, yellow pods with
brown pubescence, yellow seed
coat, small seeds, light brown
hilum, high pod-shattering, early,
good germinability, suitable for
food uses.
S12 INDIRA
SOYA 9
106 22-23 Resistant to rust.
Moderately resistant to
stem tunneling and girdle
beetle and leaf folder.
Performs well under low
to moderate plant
densities
Light grey pubescence
throughout the plant parts, broad
light medium size green leaves,
yellow seeds of medium size with
black hilum and intermediate
lustre.
S13 JS-80-21 105-110 25-30 Tolerant to bacterial
pustules, viral diseases
and foliar insect-pests.
Purple flowers, tawny
pubescence, yellow seed coat,
brown/black hilum, determinate,
high seed germinability. Perform
well in eastern states.
S14 JS-2 90-95 18-20 Resistant to bacterial
pustule, tolerant to
Macrophomina
Purple flowers, tawny
pubescence, pods with dense
brown pubescene, yellow seed
coat, light brown hilum,
determinate, highly shattering
S15 MAUS 71 93-100 18-30 Not known Semi determinate, purple flowers,
glabrous leaves, yellow seed with
black hilum, resistant to pod
shattering.
S16 SHYAMA 105-110 20-25 Resistant to bacterial
pustules and seed and
seed ling rot, tolerant to
Anthracnose
Purple flowers, brown pubescen,
black seed coat, buff coloured
hilum, medium tall plants, suited
to low input conditions.
S17 MACS 124 95-105 25-30 Resistant to bud blight,
soybean mosaic and
bacterial pustules.
Purple flowers,
tawny pubescence, yellow seed
coat, dark brown hilum, semi-
determinate resistant to lodging.
S18 MACS 450 90-95 25-40 Resistant to leaf spot,
bud blight, yellow
mosaic, soybean mosaic,
bacterial pustule. Highly
resistant to stemfly and
defoliators.
Purple flowers, medium tall,
semi-determinate, tawny
pubescence, yellow seed, black
hilum.

S. KUMAR: RAPD MARKER PROFILS OF SOYBEAN CULTIVARS 685
S19 BRAGG 112-115 15-20 Resistant to bacterial
pustules, susceptible to
YMV.
White flowers, grey pubescence,
yellow seed coat, black hilum,
brown pods, determinate.
S20 PK 416 115-120 30-35 Resistant to YMV &
bacterial pustules,
tolerant to Rhizoctonia.
White flowers, tawny
pubescence, yellow seed coat,
brown hilum, semi-
determinate. Performs well in
Punjab, Haryana and also in
central zone.
S21 SL 295 120-130 20-25 Resistant to yellow
mosaic virus
White flower, tawny pubescence,
determinate, yellow seed coat,
black hilum.
S22 NRC 37 96-102 35-40 Moderately resistant to
collar rot, bacterial
pustule, pod blight and
bud blight like
syndrome. Moderately
resistant to stem fly and
leaf miner. Non lodging
under optimum plant
population, non
shattering behaviour upto
10 days after harvest
maturity
Erect, determinate plants with out
anthocyanin colouration in the
hypocotyl, white flowers, tawny
pubescence, small to medium,
spherical yellow seeds with light
to dark brown hilum.
S23 PUSA 20 105-120 25-30 Resistant to Rhizoctonia
& bacterial pustules,
tolerant to YMV and
major insect-pests.
White flowers, tawny
pubescence, yellow seed coat,
black hilum, compact
plant. Performs well in central
zone.
S24 NRC 7 90-99 25-35 Resistant to bacterial
blight, green mosaic
virus, bacterial pustules,
phyllody, soybean
mosaic, Myrothe-
cium and Cercospora
leaf spots, tolerant to
stem fly, girdle beetle,
green and grey
semilooper, leaf miner
and defoliators.
Determinate, grey pubescence,
purple flowers, yellow seed coat,
brown hilum, high oil content,
resistant to pod-shattering.
Isolation of genomic DNA
DNA was isolated followed by CTAB method (MURRAY and THOMSON, 1980). Three
grams of fresh leaf sample was ground using the extraction buffer (1.4 M NaCl; 20 mM EDTA,
pH 8.0; 100 mM Tris, pH 8.0; 2 % CTAB), incubated at 65 °C for 30 min and centrifuged at

686 GENETIKA, Vol. 46, No.3, 681-692, 2014
10,000 rpm for 10 min. The upper aqueous phase was extracted 2-3 times with fresh chloroform:
iso-amyl alcohol (24:1). The final aqueous phase was transferred to other centrifuge tubes. To
these, 0.6 volume of ice cold iso-propanol was added and mixed gently by inverting the tube. DNA
complex was spooled out with a bent pasture pipette and 20 ml of washing solution was added to
it. The pellet was gently agitated for few minutes and collected by centrifugation at 40C, dried and
an appropriate volume of TE buffer was used to dissolve the pellet. Further the RNaseA @
10µg/ml was added into the DNA which was then incubated at 370C for 30 minutes. Equal volume
of phenol: chloroform: iso-amyl alcohol (25:24:1) was added into it and centrifuged at 10000 rpm
for10 minutes. Aqueous phase was taken and equal volumes of chloroform: iso-amyl alcohol
(24:1) was added and centrifuged at 10000 rpm for 10 min. To the aqueous phase, 1/20th
volume
of sodium acetate (3M, pH-5.2) and 2.5 volume of ethanol was added. Next it was incubated at -
200C for 1h (or-70
0C for 30 min). Then the solution was centrifuged at 10000 rpm for 10 min.
Thereafter, pellets were washed with 70% ethanol (10000 rpm, 5 min), air dried and dissolved in
distilled water. Yield of DNA was estimated using DNA markers with known λDNA – uncut,
through electrophoresis.
PCR amplification and gel electrophoresis
The random amplification was performed following a modified method of WILLIAMS et
al. (1990). The reaction was carried out in a thermal cycle (Mastercycler, Eppendorf). A total of 80
RAPD primers were screened, out of which 37 produced amplifications and only 30 primers (viz.
OPA 01, 03, 04, 08, 13 ; OPJ 01, 05, 09, 11, 12, 13, 16, 17, 18, 19; OPP 03, 04, 05, 06, 07, 08, 09
and OPT 01, 05, 07, 12, 13, 15, 16, 20, Operon Technologies, Canada, USA) showed
unambiguous DNA profile (Table 2). Polymerase chain reaction mixture of 25 µl contained 25 ng
of genomic DNA template, 0.6 U of Taq DNA polymerase (Bangalore Genei, Bangalore, India),
0.3 M decamer primer (Operon Technologies, Alameda, CA, USA), 2.5 µl 10 X PCR assay buffer
(50 mM KCl, 10 mM Tris–HCl, 1.5 mM MgCl2) and 0.25 µ1 pooled dNTPs (100 mM each of
dATP, dCTP, dGTP and dTTP from Promega, USA). PCR cycle conditions were as follows:
initial denaturing step at 94°C for 3 min followed by 44 cycles of 94°C for 1 min, 37°C for 1 min
and 72°C for 2 min. In the last cycle, primer extension at 72°C for 7 min was provided. PCR
products separation was done through 1.5% agarose gel electrophoresis alongside O’Gene
RulerTM 100 bp DNA Ladder Plus (Fermentas Life Sciences) as molecular mass marker. The
amplified products were documented under UV light source.
Data analysis
Clearly observed bands were scored manually for their presence (1) and absence (0)
across the genotypes and assembled in a binary data matrix table. Data on clearly resolved bands
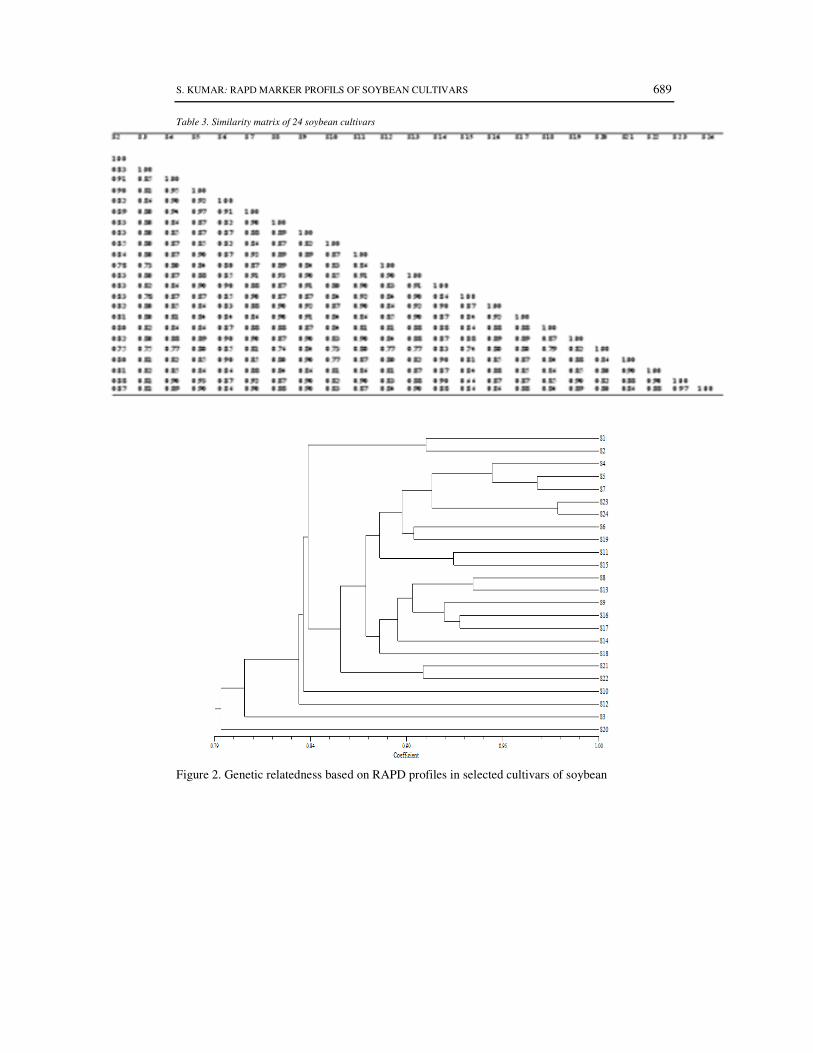
generated by 30 primers were used to estimate genetic similarity(s) among the genotypes based on
Jaccard’s coefficient. This matrix was analysed using UPGMA (Unweighted Pair Group Method
using Arithmetic averages) following the SHAN (Sequential Agglomerative Hierarchical Nested)
cluster analysis module to derive a dendrogram. All these computations were carried out using
software NTSYS-pc (ROHLF, 2000).

S. KUMAR: RAPD MARKER PROFILS OF SOYBEAN CULTIVARS 687
Table 2. Name, sequence and details of the polymorphic RAPD primers used in the present study
RESULTS
Out of total 80 primers (Kits OPA, OPP, OPT and OPJ) tried in this study 37 primers
showed clear and unambiguous amplification (Table 3). Scorable 30 RAPD primers led to
amplification of 120 fragments ranging from about 3,300 bp (OPT 1) to 300 bp (OPT 12), out of
which 81 (67.5%) bands were found to be polymorphic. The level of polymorphism ranged from
33.4% (OPT 16, where 1/3 bands was found to be polymorphic) to 100% (OPA1, 4, OPP 5, 7, 8,
9, 15 and OPT 13). Maximum number of 7 amplified products were obtained by primer OPT 01
and 13. Eight primers were found to be monomorphic and the primer OPA 03 amplified a
Primer Sequence (5'-3') Molecular
weight
No. of bands
amplified
No. of
polymorphic
band
%
polymorphism
OPA-01 CAGGCCCTTC 2964 6 6 100
OPA-03 AGTCAGCCAC 2997 1 0 0
OPA-04 AATCGGGCTG 3068 5 5 100
OPA-08 GTGACGTAGG 3108 2 1 50
OPA-13 CAGCACCCAC 2942 5 2 40
OPJ-01 CCCGGCATAA 2997 2 1 50
OPJ-05 CTCCATGGGG 3044 2 0 0
OPJ-09 TGAGCCTCAC 2988 1 0 0
OPJ-11 ACTCCTGCGA 2988 6 3 50
OPJ-12 GTCCCGTGGT 3035 5 4 80
OPJ-13 CCACACTACC 2917 3 1 33.3
OPJ-16 CTGCTTAGGG 3059 1 0 0
OPJ-17 ACGCCAGTTC 2988 3 0 0
OPJ-18 TGGTCGCAGA 3068 5 5 100
OPJ-19 GGACACCACT 2997 3 2 66.67
OPP-03 CTGATACGCC 2988 5 2 40
OPP-04 GTGTCTCAGG 3059 1 0 0
OPP-05 CCCCGGTAAC 2973 5 5 100
OPP-06 GTGGGCTGAC 3084 5 4 80
OPP-07 GTCCATGCCA 2988 4 4 100
OPP-08 ACATCGCCCA 2957 7 7 100
OPP-09 GTGGTCCGCA 3044 4 4 100
OPT-01 GGGCCACTCA 3013 7 6 85.7
OPT-05 GGGTTTGGCA 3099 3 0 0
OPT-07 GGCAGGCTGT 3084 5 2 40
OPT-12 GGGTGTGTAG 3139 5 4 80
OPT-13 AGGACTGCCA 3037 7 7 100
OPT-15 GGATGCCACT 3028 6 5 83.3
OPT-16 GGTGAACGCT 3068 3 1 33.3
OPT-20 GACCAATGCC 2997 3 0 0

688 GENETIKA, Vol. 46, No.3, 681-692, 2014
minimum of 1 band. On an average we obtained 4 bands per primer and 22 primers used in the
study produced polymorphic banding pattern. RAPD product (750 bp) produced by OPT 07 was
found to be specific for MAUS-61 , while OPJ 18 produced a 2350 bp band only in Indira Soya 9.
DNA amplification pattern as detected by some of the RAPD primers in the soybean cultivars has
been provided in Fig. 1.
Figure 1. RAPD profile of soybean varieties obtained with primers OPA-04, OPJ-12 and OPT-13.
Serial number of the varieties corresponds to Table 1. M = Standard DNA marker, 100 bp
DNA Ladder Plus
OPA-04
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
OPJ-12
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
OPT-13
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
S. KUMAR: RAPD MARKER PROFILS OF SOYBEAN CULTIVARS 689
Table 3. Similarity matrix of 24 soybean cultivars
Figure 2. Genetic relatedness based on RAPD profiles in selected cultivars of soybean

690 GENETIKA, Vol. 46, No.3, 681-692, 2014
Thus, it is evident that, 30/80 primers tried (38%) provided unambiguous amplification
and out of 30 primers, 22 primers (73%) were found to be polymorphic.
DISSCUSSION
RAPD has been found to be an effective and efficient tool to evaluate and reveal genetic
polymorphism in several crop species like rice (VIRK et al., 1995; RAY-CHOUDHURY et al., 2001),
wheat (CAO et al., 2000), maize (PEJIC et al., 1998), barley (YU et al., 2002) and soybean
(BARANEK et al., 2002; BARAKAT, 2004). In our present study 67.5% fragments were found to be
polymorphic as compared to 46% obtained by BARANEK et al. (2002) in 19 Czech National
Collection of soybean genotypes. Further, THOMPSON et al. (1998) were able to identify only 34%
polymorphism using RAPD. Very high degree of polymorphism detected in our present study
indicated an accurate selection of the polymorphic RAPD primers. The 30 RAPD primers used in
the present study has been selected after screening of 80 primers, 22 out of 30 were found to be
polymorphic. Thus, quite a high percentage of primers (73%, as mentioned earlier). Two
genotypes, were able to generate specific unique band. However, validation of the identified
primers and reproducibility of those bands are required for proper identification of the genotypes.
Moreover, the unique bands could be converted into SCAR (Sequence Characterized Amplified
Region) markers for specificity. In this regard, few more RAPD primers could be tried for
identification of unique bands in newer varieties. On an average, we got 4 bands per primer.
Further, 8 primers were found to give 100% polymorphism among the genotypes studied. 15
primers (50%) have been found to develop more number of bands than the average value which is
encouraging (Table 2) and indicates the efficiency of RAPD primers towards development of
molecular profiles.
ACKNOWLEDGEMENTS
Author is very grateful to Directorate of Seed Research, Kushmaur, Mau. India, to provide
facilities for this work. Special thanks are due to Dr. P. RAY-CHOUDHURY and Mr. S.P. TRIPATHI,
for his guidance during this work.
Received December 16th, 2013
Accepted September 05th, 2014
REFERENCES
BARAKAT, H. (2004): Genetic fingerprinting and relationships of six soybeans (Glycine max L.) cultivars based on protein
and DNA polymorphism. Int. J. Agri. and Biol., 6: 877-883.
BARANEK, M., M. KADLEC, J. RADDOVA, M. VACHUN and M. PIDRA (2002): Evaluation of genetic diversity in 19 Glycine max
(L.) Merr. accessions included in the Czech national collection of soybean genotypes. Czech. J. Genet. Plant
Breed, 38: 69–74.
BERNARD, R.L., G.A. JUVIK and R.L. NELSON (1989): USDA Soybean Germplasm Collection Inventory. Int. Agri. Pub.,
Urbana, IL.
CAO, W., G. SCOLES, P. HUCL and R.N. CHIBBAR (2000): Phylogenetic relationships of five morphological groups of
hexaploid wheat (Triticum aestivum L.) based on RAPD analysis. Genome, 43: 724–727.
CARPENTER, J.A. and W.R. FEHR (1986): Genetic variability for desirable agronomic traits in populations containing Glycine
soja germplasm. Crop Sci., 26: 681–686.
DOLDI, M.L., J. VOLLMANN and T. LELLEY (1997): Genetic diversity in soybean as determined by RAPD and microsatellite
analysis, Plant Breed., 116: 331-335.

S. KUMAR: RAPD MARKER PROFILS OF SOYBEAN CULTIVARS 691
GRIFFIN, J.D. and R.G. PALMER (1995): Variability of thirteen isozyme loci in the USDA soybean germplasm collections.
Crop Sci., 35: 897–904.
KUMAR, S. and A.K. SINGH (2014a): Complete absence of linkage disequilibrium between enzyme loci in natural
populations of Drosophila ananassae. Genetika 46: 227–234.
KUMAR, S. and A.K. SINGH (2014b): Allozyme polymorphism in Drosophila. Proc. Zool. Soc. 67: (in press) DOI
10.1007/s12595-014-0126-3
JUVIK, G., R.L. BERNARD, R.Z. CHANG and J.F. CAVINS (1989): Evaluation of the USDA Wild Soybean Germplasm
Collection: Maturity Group 000 to IV (PI 65.549 to PI 483.464). U.S. Department of Agriculture. Tech Bull
1761, U.S. Govt Print Office, Washington, DC.
KEIM, P., R.C. SHOEMAKER and R.G. PALMER (1989): Restriction fragment length polymorphism diversity in soybean. Theor.
Appl. Genet., 77: 786–797.
MAUGHAN, P.J., M.M.A. SAGHAI and G.R. BUSS (1996): Molecular-marker analysis of seed-weight: Genomic locations, gene
action, and evidence for orthologous evolution among three legume species. Theor. Appl. Genet., 93: 574–579.
MAUGHAN, P.J., M.M.A. SAGHAI and G.R. BUSS (1995): Microsatellite and amplified sequence length polymorphisms in
cultivated and wild soybean. Genome, 38: 715-723.
MURRAY, M.G. and W.F. THOMSON (1980): Rapid isolation of high molecular weight plant DNA. Nucl. Acid Res., 8: 4321-
4325.
NELSON, A.I., W.B. WIJERATNE, S.W. YEN, T.M. WEI and L.S. WEI (1987): Dry extrusion as an aid to mechanical expelling of
oil from soybeans. J. Am. Oil. Chem. Soc., 64: 1341–1347.
NELSON, R.L., C.D. NICKELL, J.H. ORF, H. TACHIBANA, E.T. GRITTON, C.R. GRAU and B.W. KENNEDY (1989): Evaluating
Soybean Germ Plasm for Brown Stem Rot Resistance. Plant Disease, 73: 110-114.
PEJIC, I., P. AJMONE-MARSON, M. MORGANTE, V. KOZUMPLICK, P. CASTIGLIONI, G. TARAMINO and M. MOTT (1998):
Comparative analysis of genetic similarity among maize inbred lines detected by RFLPs, RAPDs, SSRs and
AFLPs. Theor. Appl. Genet., 97: 1248–1255.
RAY-CHOUDHURY, P., S. KOHLI, K. SRINIVASAN, T. MOHAPATRA and R.P. SHARMA (2001): Identification and classification
of aromatic rice based on DNA fingerprinting. Euphytica, 118: 243–251.
ROHLF, F.J. (2000): NTSYS-pc version 1.70 numerical taxonomy and multivariate analysis system. - Applied Biostatistics
Inc., Exeter Software. Setauket - New York.
THOMPSON, J.A., R.L. ELSON and L.O. VODKIN (1998): Identification of diverse soybean germplasm using RAPD markers.
Crop Sci., 38: 1348–1355.
VIRK, P.S., B.V. FORD-LLOYD, M.T. JACKSON and H.J. NEWBURY (1995): Use of RAPD for the study of diversity within plant
germplasm collections. Heredity, 4: 170–179.
WILLIAMS, J.G.K., A.R. KUBELIK, K.J. LIVAK, J.A. RAFALSKI and S.V. TINJOY (1990): DNA polymorphism amplified by
arbitrary primers are useful as genetic markers. Nucl. Acid Res., 18: 6531-6535.
YU, Z., L. LI-QIONG, L. HUAN, B. JIE, Y. MAN-YE, M. CHEN, C. YING-FAN, Q. XIAO-LIN and C. FANG (2002): RAPD markers in
diversity detection and variety identification of Tibetan hulless barley. Plant Mol. Bio. Report, 20: 369–377.

692 GENETIKA, Vol. 46, No.3, 681-692, 2014
ISPITIVANJE EFIKASNOSTI RAPD MARKERA U RAZVOJU MOLEKULARNOG
PROFILA ZA ISPITIVANJE GENETIČKE ČISTOĆE (Glycine max L.)
Sanjay KUMAR
Genetička laboratorija, Odelenje za zoologiju Banaras Hindu Univerzitet, Varanasi, Indija
Izvod Glavni napredak u istraživanjima soje je u razumevanju genetičkih osobina i primena
novih tehnologija u njenom poboljšanju. Utvrđivanje genetičkog odnosa aktivnosti enzima i
molekularnih markera je pokazalo da je konzistentno sa očekivanjima zasnovanim na poreklu i
informacijama pedigrea. Da bi se identifikovali efikasni markeri koji će biti korišćeni u
ispitivanjima genetičke čistoće polimorfizam je osnovni kriterijum. U ovom radu vršena je
evaluacije 80 RAPD markera od kojih je moglo da se amplifikuje 37 a samo 30 prajmera je dalo
nedvosmislene profile. Od tih 30, 22 je imalo polimorfnu sliku fragmenata DNK (banding).
Analiziranih 30 RAPD markera je dovelo do umnožavanja 120 fragmenata od kojih je 81 (67.5 %)
fragmenata bilo polimorfno. U proseku dobijena su 4 fragmenta po prajmeru i 115 prajmera (50
%) je dalo veći broj fragmenata od prosečnog broja što je ohrabrujuće. 8 prajmera je imalo 100%
polimorfizam. Rezultati su indikativni za efikasnost RAPD prajmera u razvoju molekularnih
profila.
. Primljeno 16. XII. 2013.
Odobreno 05. IX. 2014.




















