
Genetic variation among seven strains of Dunaliella salina (Chlorophyta) with industrial potential,...
14
Genetic variation among seven strains of Dunaliella salina (Chlorophyta) with industrial potential, based on RAPD banding patterns and on nuclear ITS rDNA sequences Patricia I. Go ´mez * , Mariela A. Gonza ´lez Departamento de Bota ´nica, Facultad de Ciencias Naturales y Oceanogra ´ficas, Universidad de Concepcio ´n, Casilla 160-C, Concepcio ´n, Chile Received 27 October 2003; received in revised form 27 October 2003; accepted 12 November 2003 Abstract Dunaliella salina (Chlorophyta) is a halophilic microalga cultivated as a natural source of h- carotene in several countries, including Chile. Previous studies of some Chilean strains of this microalga have shown a great variability in their physiological and genetic attributes. Random amplified polymorphic DNA (RAPD) band patterns and nuclear ribosomal DNA internal transcribed spacer (ITS-1 and ITS-2) sequences were used to genotypically characterize three Chilean and four foreign (from Mexico, China, Australia and Israel) strains of D. salina with industrial potential. Unweighted pair group mean average (UPGMA) cluster analysis of RAPD data distinguished, at 70% similarity, two clusters. One cluster included all of the foreign strains, except the one from Australia, and the other grouped two of the Chilean strains (CONC-001 and CONC-007). The other Chilean strain (CONC-006) and the Australian strain appeared as single entities. Neighbor-joining (NJ), maximum parsimony (MP) and maximum likelihood (ML) analyses of the ITS sequence data yielded four clades, most of them well supported: there were two clades containing exclusively one strain, CONC-006 from Chile (86 – 94% support) and the one from Australia (98 – 100% support); the third clade grouped the strains from Mexico, China and Israel (99 – 100% support), and, the fourth clade, the strains CONC-001 and CONC-007 from Chile (100% support). Both RAPD banding pattern and ITS sequence data were consistent in resolving the same genetic relatedness among the strains analyzed. For both approaches, the strain CONC-006 from Chile and the strain from Australia were the most divergent entities within the group. Furthermore, the strain CONC-006 appeared genetically more related to the foreign strains than to the other Chilean ones. The physiological attributes exhibited by these 0044-8486/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.aquaculture.2003.11.005 * Corresponding author. Tel.: +56-41-204156; fax: +56-41-246005. E-mail address: [email protected] (P.I. Go ´mez). www.elsevier.com/locate/aqua-online Aquaculture 233 (2004) 149 – 162
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Genetic variation among seven strains of Dunaliella salina (Chlorophyta) with industrial potential,...

www.elsevier.com/locate/aqua-online
Aquaculture 233 (2004) 149–162
Genetic variation among seven strains of Dunaliella
salina (Chlorophyta) with industrial potential,
based on RAPD banding patterns and on
nuclear ITS rDNA sequences
Patricia I. Gomez*, Mariela A. Gonzalez
Departamento de Botanica, Facultad de Ciencias Naturales y Oceanograficas, Universidad de Concepcion,
Casilla 160-C, Concepcion, Chile
Received 27 October 2003; received in revised form 27 October 2003; accepted 12 November 2003
Abstract
Dunaliella salina (Chlorophyta) is a halophilic microalga cultivated as a natural source of h-carotene in several countries, including Chile. Previous studies of some Chilean strains of this
microalga have shown a great variability in their physiological and genetic attributes. Random
amplified polymorphic DNA (RAPD) band patterns and nuclear ribosomal DNA internal
transcribed spacer (ITS-1 and ITS-2) sequences were used to genotypically characterize three
Chilean and four foreign (from Mexico, China, Australia and Israel) strains of D. salina with
industrial potential. Unweighted pair group mean average (UPGMA) cluster analysis of RAPD
data distinguished, at 70% similarity, two clusters. One cluster included all of the foreign strains,
except the one from Australia, and the other grouped two of the Chilean strains (CONC-001 and
CONC-007). The other Chilean strain (CONC-006) and the Australian strain appeared as single
entities. Neighbor-joining (NJ), maximum parsimony (MP) and maximum likelihood (ML)
analyses of the ITS sequence data yielded four clades, most of them well supported: there were
two clades containing exclusively one strain, CONC-006 from Chile (86–94% support) and the
one from Australia (98–100% support); the third clade grouped the strains from Mexico, China
and Israel (99–100% support), and, the fourth clade, the strains CONC-001 and CONC-007
from Chile (100% support). Both RAPD banding pattern and ITS sequence data were consistent
in resolving the same genetic relatedness among the strains analyzed. For both approaches, the
strain CONC-006 from Chile and the strain from Australia were the most divergent entities
within the group. Furthermore, the strain CONC-006 appeared genetically more related to the
foreign strains than to the other Chilean ones. The physiological attributes exhibited by these
0044-8486/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.aquaculture.2003.11.005
* Corresponding author. Tel.: +56-41-204156; fax: +56-41-246005.
E-mail address: [email protected] (P.I. Gomez).

P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162150
strains of D. salina are mostly in agreement with the genetic relationships deduced from this
study.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Dunaliella salina strains; RAPD; Ribosomal ITS sequences; Genotypic characterization
1. Introduction
Dunaliella salina is a halotolerant green microalga widely used in mass culture for
commercial purposes. Under appropriate culture conditions, some strains of D. salina can
accumulate up to 10% carotenoids, mostly h-carotene, becoming the richest eukaryotic
source of this pigment known so far (Ben-Amotz et al., 1982; Ben-Amotz and Avron,
1983; Borowitzka and Borowitzka, 1988).
Physiological variability has been reported at the intraspecific level in D. salina
(Araneda et al., 1992a,b; Cifuentes et al., 1992, 1996a,b; Markovits et al., 1993; Gomez
et al., 1999). A negative consequence of this variability is that it could lead to erroneous
assumptions about the industrial potential of this microalga, when information coming
from one strain is extrapolated to the whole species. On the other hand, this intraspecific
diversity is advantageous from a biotechnological point of view, since it permits the
application of strain selection procedures, easily improving fine chemicals production
(Lopez Alonso et al., 1992a,b; 1994).
The global demand of natural beta-carotene is around 1430 tons per annum (tpa), while
the availability is just 35 tpa produced by the companies Henkel-Cognis (Australia),
Nature Beta Technologies (a subsidiary of Nikken Sohonsha, Israel) and Dutch State
Mines (DSM; Food Specialties, Netherlands). The balance demand is met through
synthetic carotene manufactured by Roche and BASF (online business magazine do-
main-b.com, June 2002, http://www.domain-b.com).
Henkel-Cognis (previously Betatene) is the largest producer of D. salina in the world;
the company has two sites, one at Hutt Lagoon, Western Australia and the other at
Whyalla, South Australia, which are the world’s largest algal farms with a total capacity of
800 ha. Similarly, but at a lower scale, Nature Beta Technologies (NBT) which was
founded to commercialize the algal research at the Weizmann Institute of Science in
Rehovot, Israel, also cultivates D. salina as a natural carotene source. Unlike the other
companies, DSM Food Specialties produces the pigment by a fermentation process of the
fungi Blakestea trispora.
D. salina is probably the most successful microalga for mass cultivation described so
far, especially due to its high salinity requirement that minimizes the number of
competitors and predators. Furthermore, due to its lack of cell wall, dried Dunaliella
is easily and fully digestible by animals and humans (Borowitzka and Borowitzka, 1988;
Ben-Amotz and Avron, 1989). In spite of these advantages, the mass culture of D. salina
as a natural carotene source still needs to be developed to satisfy all the required
demand.
The development of mass culture conditions has required growth optimization of
growth as well as the amount of the carotene produced per unit time and culture volume.

P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162 151
This objective is complicated due to the fact that the conditions for maximum cell
growth are opposite of those for maximum carotene accumulation per cell. There is no
doubt that the selection of strains with improved growth and carotenogenic attributes
would help to make the mass culture of D. salina economically competitive with its
synthetic counterpart.
The genetic characterization of strains with industrial potential has a great relevance in
applied science because it permits partitioning of phenotypic variation into environmental
(phenotypic plasticity) and genetic components. Furthermore, genetic diversity studies
permit exotic genotypes detection, which could be the sources of genes with biotechno-
logical potential.
Many different molecular techniques can be used to reveal genetic polymorphisms. One
of them is random amplified polymorphic DNA (RAPD), a DNA-fingerprinting method
that allows the detection of multi-locus genetic variation (Welsh and Mc Clelland, 1990;
Williams et al., 1990). The RAPD technique has been successfully employed to identify
microalgae up to the strain level (Neilan, 1995; Nishihara et al., 1997; Bolch et al.,
1999a,b). By using RAPD, a recent genetic analysis of eight Chilean strains of D. salina
showed that the strain CONC-006 from an alpine salt lake (‘‘Salar de Atacama’’) was
genetically more similar to CONC-001 (42% similarity) from a coastal lagoon (‘‘La
Rinconada’’) than to almost all of the other strains collected in the ‘‘Salar de Atacama’’,
including CONC-007 (39% similarity) (Gomez and Gonzalez, 2001).
On the other hand, some non-coding rapidly evolving sequences have been used to
resolve intra- and interspecific relationships among a variety of organisms. This is the case
of the internal transcribed spacers (ITS-1 and ITS-2) of the nuclear ribosomal DNA
(Schlotterer, 2001). The ITS-1 and ITS-2 sequences have proved very useful in assessing
genetic relatedness in green algae, both unicellular and colonial (e.g. Coleman et al., 1994;
Mai and Coleman, 1997; Angeler et al., 1999).
In Dunaliella, Gonzalez et al. (1999), using ITS-RFLP analysis, did not find a
correlation between polymorphism in the ITS region (ITS1 + 5.8S rDNA+ ITS2) and
the morpho-physiological attributes used for the taxonomy within the genus. However,
after a reappraisal of the physiological attributes of nine strains of Dunaliella (excluded D.
salina) (Cifuentes et al., 2001), Gonzalez et al. (2001) recommended renaming several
strains of Dunaliella based on the phylogenetic relationships deduced from the ITS
sequences which, this time, were congruent with the morpho-physiological attributes of
the strains. Moreover, they found a much higher sequence variation among strains of D.
salina than among strains belonging to other species of the genus, stressing the need of
further work to elucidate the species status of this complex taxon.
Comparison of the physiological response within D. salina (different strains) is
rather difficult since the data come from different physiological studies, most of them
done under different experimental conditions. The only previous data that account for
physiological variability within different strains of Dunaliella is the paper published by
Cifuentes et al. (1992), which includes eight Chilean strains. Recently, an exhaustive
study on D. salina evaluated growth and carotenogenesis in three Chilean (CONC-001,
CONC-006, CONC-007) and four foreign (from Mexico, China, Australia and Israel)
strains cultivated in combined conditions of temperature and irradiance (Gomez and
Gonzalez, unpublished data). All the foreign strains, in spite of coming from

P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162152
geographically very distant locations, exhibited similar physiological responses to the
Chilean ones. In this study, our aim was to find out whether physiologically similar
strains of D. salina are also genetically related. To do this, RAPD patterns and ITS
rDNA sequence data were analyzed to quantify genetic polymorphism and to establish
phylogenetic relationships among these strains.
2. Materials and methods
2.1. Strain culture and DNA extraction
In this study, unialgal cultures of seven strains of D. salina were included. All of them
are maintained in the Culture Collection of Microalgae at the Universidad de Concepcion,
Concepcion, Chile. Table 1 summarizes the geographic origin and the source of each
strain. All the strains were cultivated in 250-ml Erlenmeyer flasks with 100 ml of modified
Johnson’s medium (Borowitzka, 1988) with 15% NaCl under a continuous photon flux
density of 100 Amol m� 2 s� 1 and 25F 2 jC. Axenic culture preparation and total DNA
extraction were carried out as in Gomez and Gonzalez (2001).
2.2. RAPD reaction conditions
RAPD amplifications were done in a 25 Al volume. A total of 50 primers from the D, P,
OPA and OPD (Operon) series were used. PCR-reaction composition, amplification
program and electrophoresis conditions were performed according to previously described
methodology (Gomez and Gonzalez, 2001). Amplifications were repeated and checked for
reproducibility between reactions.
Table 1
Strains of D. salina included in this study
Strain Source and Geographic GenBank accession numberstrain number origin
ITS-1 ITS-2
D. salina
CONC-001
CONC-001 La Rinconada Lagoon,
Antofagasta, Chile
AF546091 AF546092
D. salina
CONC-006
CONC-006 Sector Burro Muerto,
Salar de Atacama, Chile
AF313424 AF313425
D. salina
CONC-007
CONC-007 Sector Puilar, Salar de
Atacama, Chile
AF313426 AF313427
D. salina
Mexican
Provided by
Ernesto Retamales,
U. Antofagasta, Chile
Yucatan, Mexico AF546093 AF546094
D. salina
Chinese
Provided by
Dr. Ralph Lewin,
Scripps Institution, USA
Tanggu, China AF546095 AF546096
D. salina
Australian
CCAP 19/18 Hutt Lagoon, Australia AF546097 AF546098
D. bardawil ATCC 30861 Bardawil Lake, Israel/Egypt AF313430 AF313431
Geographic origins and the GenBank accession number of ITS sequences are also shown for each strain.

2.3. ITS spacers sequencing
The primers used to amplify the ITS region (ITS1 + 5.8S rDNA+ ITS2) were AB28
and TW81 described in Goff et al. (1994). The amplification reaction mixtures and the
PCR program were performed as in Gonzalez et al. (1999). PCR products were analyzed
by electrophoresis of 2 Al of the reaction mixture through 2% agarose gel and visualized
by ethidium bromide staining. ITS fragments were gel purified using the kit Wizard
PCR Purification System (Promega, Madison, WI) and used directly as templates for
sequencing (without prior cloning). Sequencing was performed with an ABI Prism 377
DNA sequencer (Centro de Sıntesis y Analisis de Biomoleculas, Universidad de Chile,
Chile).
2.4. Data analysis
For RAPD-PCR data, just strong and reproducible bands were scored. Co-migrating
bands were scored as either present (score 1) or absent (score 0) to produce a binary matrix
for all the strains examined. The presence/absence matrix was used to calculate pairwise
RAPD distances between strains using the mean-distance metric of PAUP (4.0b8 version,
Swofford, 1999) and to construct a clustering dendrogram using the unweighted pair group
mean average (UPGMA) algorithm. For ITS analysis, combined ITS-1 and ITS-2 data sets
were analyzed. ITS sequences from all of the D. salina strains were aligned manually to
sequences of two reference outgroups: Dunaliella viridis (strain CONC-002; Genbank No.
AF313418, AF313419) and Dunaliella tertiolecta (strain UTEX 999; Genbank No.
AF313434, AF313435) according to Gonzalez et al. (2001). The resulting alignment
was subjected to a cladistic analysis. Both distance-based and character state-based tree-
building methods were used. Three different optimality criteria were employed: neighbor-
joining (NJ), maximum parsimony (MP) and maximum likelihood (ML). NJ analysis was
performed with the Kimura two-parameter distance model (Kimura, 1980) using the
MEGA 1.02 (Kumar et al., 1993) program. Bootstrapping the data with 500 replications
tested the robustness of each branch. In addition, sequences were analyzed with the MP
and ML algorithms of PAUP; both analyses were performed by branch and bound search
and subjected to 1000 and 500 bootstrap replicates, respectively. All sites were treated as
independent, unordered and equally weighted characters. ML analysis was based on the
Hasegawa–Kishino–Yano model (Hasegawa et al., 1985).
P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162 153
3. Results
3.1. RAPD-PCR analysis
From the total 50 primers initially tested, 12 gave clear and reproducible amplification
patterns, and consequently, were used for the comparative analysis (Table 2). The number
of bands generated with these primers was between 5 (with primers P-100, OPA-11 and
OPA-13) and 11 (with primers OPD-11 and OPD-18), with fragment size ranging between
300 and 2000 bp.
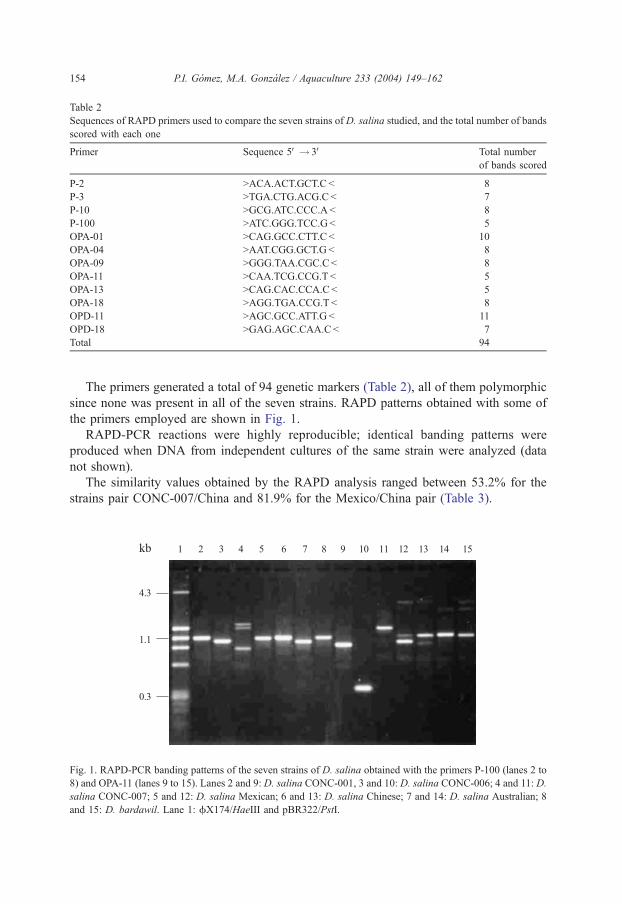
Table 2
Sequences of RAPD primers used to compare the seven strains of D. salina studied, and the total number of bands
scored with each one
Primer Sequence 5V! 3V Total number
of bands scored
P-2 >ACA.ACT.GCT.C< 8
P-3 >TGA.CTG.ACG.C< 7
P-10 >GCG.ATC.CCC.A< 8
P-100 >ATC.GGG.TCC.G< 5
OPA-01 >CAG.GCC.CTT.C< 10
OPA-04 >AAT.CGG.GCT.G< 8
OPA-09 >GGG.TAA.CGC.C< 8
OPA-11 >CAA.TCG.CCG.T < 5
OPA-13 >CAG.CAC.CCA.C< 5
OPA-18 >AGG.TGA.CCG.T < 8
OPD-11 >AGC.GCC.ATT.G < 11
OPD-18 >GAG.AGC.CAA.C< 7
Total 94
P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162154
The primers generated a total of 94 genetic markers (Table 2), all of them polymorphic
since none was present in all of the seven strains. RAPD patterns obtained with some of
the primers employed are shown in Fig. 1.
RAPD-PCR reactions were highly reproducible; identical banding patterns were
produced when DNA from independent cultures of the same strain were analyzed (data
not shown).
The similarity values obtained by the RAPD analysis ranged between 53.2% for the
strains pair CONC-007/China and 81.9% for the Mexico/China pair (Table 3).
Fig. 1. RAPD-PCR banding patterns of the seven strains of D. salina obtained with the primers P-100 (lanes 2 to
8) and OPA-11 (lanes 9 to 15). Lanes 2 and 9: D. salina CONC-001, 3 and 10: D. salina CONC-006; 4 and 11: D.
salina CONC-007; 5 and 12: D. salina Mexican; 6 and 13: D. salina Chinese; 7 and 14: D. salina Australian; 8
and 15: D. bardawil. Lane 1: fX174/HaeIII and pBR322/PstI.
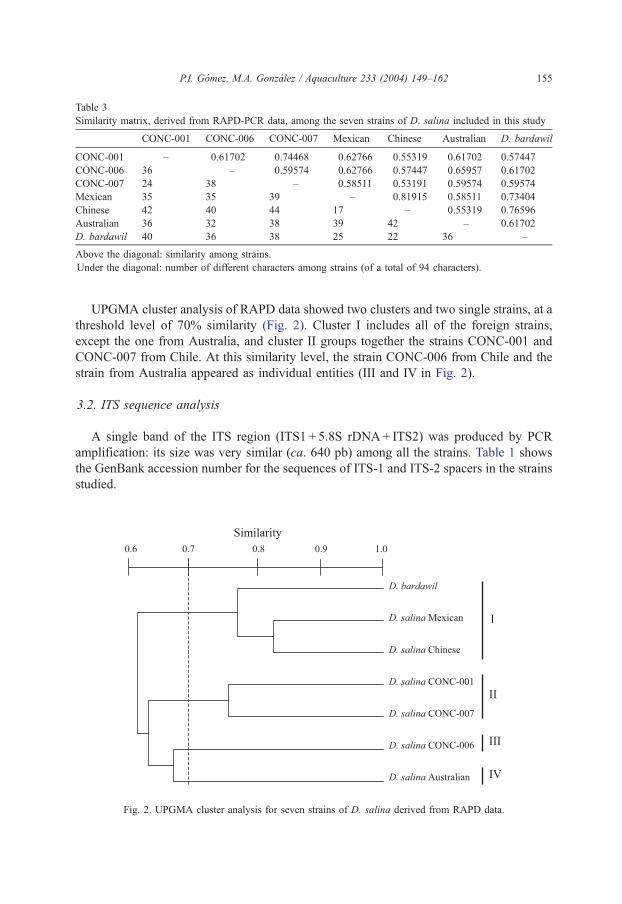
Table 3
Similarity matrix, derived from RAPD-PCR data, among the seven strains of D. salina included in this study
CONC-001 CONC-006 CONC-007 Mexican Chinese Australian D. bardawil
CONC-001 – 0.61702 0.74468 0.62766 0.55319 0.61702 0.57447
CONC-006 36 – 0.59574 0.62766 0.57447 0.65957 0.61702
CONC-007 24 38 – 0.58511 0.53191 0.59574 0.59574
Mexican 35 35 39 – 0.81915 0.58511 0.73404
Chinese 42 40 44 17 – 0.55319 0.76596
Australian 36 32 38 39 42 – 0.61702
D. bardawil 40 36 38 25 22 36 –
Above the diagonal: similarity among strains.
Under the diagonal: number of different characters among strains (of a total of 94 characters).
P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162 155
UPGMA cluster analysis of RAPD data showed two clusters and two single strains, at a
threshold level of 70% similarity (Fig. 2). Cluster I includes all of the foreign strains,
except the one from Australia, and cluster II groups together the strains CONC-001 and
CONC-007 from Chile. At this similarity level, the strain CONC-006 from Chile and the
strain from Australia appeared as individual entities (III and IV in Fig. 2).
3.2. ITS sequence analysis
A single band of the ITS region (ITS1 + 5.8S rDNA+ ITS2) was produced by PCR
amplification: its size was very similar (ca. 640 pb) among all the strains. Table 1 shows
the GenBank accession number for the sequences of ITS-1 and ITS-2 spacers in the strains
studied.
Fig. 2. UPGMA cluster analysis for seven strains of D. salina derived from RAPD data.

P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162156
In all the D. salina strains, the ITS-2 exhibited a slightly higher variability (sub-
stitutions, insertions and deletions 49/235 sites = 20.8%) than ITS-1 (32/215 sites = 14.9%)
(Fig. 3). Among substitutions, transitions were more common than transversions as
indicated by a transition/transversion ratio of 2.2 estimated via ML, over all nucleotide
positions.
Divergence values, estimated via the Kimura two-parameter model, ranged between
11.4% (CONC-006/CONC-007) and 0.2% (China–Mexico) (data not shown).
Fig. 3. Aligned sequences of ITS-1 and ITS-2 in the seven strains of D. salina studied. D. viridis (D. vir) and
D. tertiolecta (D. tert) were included as outgroups. A dash (–) indicates introduced gaps. Letters in bold and
underlined indicate mutations that characterize the Chilean strains.

P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162 157
There were DNA motifs shared by all the strains (including outgroups) and others
unique to certain D. salina strains. The three Chilean strains shared six substitutions along
the ITS spacers when compared with the foreign strains; these are located at positions 55,
56 and 120 in ITS-1 and 18, 112 and 222 in ITS-2. In addition, two short insertions in the
ITS-2 (between nucleotides 21–26 and 31–35) are found only in strains CONC-001 and
CONC-007 (Fig. 3).
The strain CONC-007 was the most divergent within the group (Fig. 4). Its farthest
conspecific was CONC-006, a strain isolated from the same biogeographic region (‘‘Salar
de Atacama’’) with which it differs by 11.4%, a value much higher than the ones observed
among the foreign strains (0.2% for the Mexico/China pair and 3.0% for the Australia/
Dunaliella bardawil pair) (data not shown).
In the parsimony analysis, 52 out of 456 characters (11.4%) were parsimony
informative and only one most parsimony tree was found at 142 steps. The data showed
low homoplasy (H.I. = 0.2289) and high consistency index (C.I. = 0.7711).
All the phylogenetic trees (distance, MP and ML) were consistent in showing five
clades: clade D includes the strains from Israel, Mexico and China; clade C consists of the
Australian strain; clade B consists of the strain CONC-006 and clade A includes the strains
CONC-001 and CONC-007. All these clades were solved with high bootstrap values (86–
100%) in the three types of phylogenetic trees. While the strain CONC-006 from Chile
Fig. 4. Phylogram based on maximum parsimony analysis of the entire ITS-1 and ITS-2 sequences of D. salina
strains. Tree shown is the only most parsimonious one at 142 steps recovered in a branch and bound search. D.
viridis and D. tertiolecta were included as outgroups. Bootstrap values are shown above (1000 replicates for
maximum parsimony-MP) and below (500 replicates for maximum likelihood-ML) branches. Consistency index
excluding uninformative characters (CI) = 0.7711; retention index (RI) = 0.7979 and rescaled consistency index
(RI) = 0.6911. Scale bar for one nucleotide change shown at the bottom.

P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162158
(clade B) and the one from Australia (clade C) each stand by itself, the strains from Israel,
Mexico and China and the two from Chile (CONC-001 and CONC-007) form two clades
(A and D), each one very well supported (bootstrap values: 99–100%), indicating a high
genetic relatedness within each group (Fig. 4).
4. Discussion
DNA-based methods have provided new possibilities for obtaining information about
genetic diversity of organisms, even at the individual level. The information provided by
this type of studies is fundamental in many fields of biology, both basic and applied (Karp
and Edwards, 1998; Weising et al., 1995).
The advantages provided by RAPD-PCR analysis have been recognized in several
papers (for reviews see Weising et al., 1995; Caetano-Anolles and Gresshoff, 1997).
However, one traditional drawback of using RAPD markers is that they represent
dominant markers, precluding discrimination between homozygous and heterozygous
diploid organisms (Van Oppen et al., 1996; Weising et al., 1995). The advantages of using
RAPD markers in the characterization of eukaryotic haploid organisms have been already
demonstrated in studies of the toxic dinoflagellate Gymnodinium catenatum (Bolch et al.,
1999b) and on strains of D. salina (Gomez and Gonzalez, 2001).
Van Oppen et al. (1994), Valatka et al. (2000), and others have stressed that RAPD
should be used only for comparisons among very closely related taxa, i.e. at the
intraspecific and/or interspecific level. In microalgae, RAPD markers have been employed
in the genetic characterization of strains of Cyanophyceae (Neilan, 1995; Nishihara et al.,
1997; Bolch et al., 1999a), Dinophyceae (Bolch et al., 1999b) and Chlorophyceae (Gomez
and Gonzalez, 2001). In the present work, the high resolution power of RAPD-markers is
demonstrated by the generation of polymorphic markers with all of the primers employed
(Fig. 1, Table 2), reinforcing previous conclusions (Gomez and Gonzalez, 2001) about the
usefulness of this technique to genetically characterize strains of D. salina with different
physiological attributes.
Gomez and Gonzalez (2001) had previously reported the high degree of genetic
variability among the Chilean strains CONC-001, CONC-006 and CONC-007 shown by
RAPD band pattern analysis. In this study a good agreement was obtained between the
phenetic and cladistic analyses of RAPD data and ITS sequences, respectively.
Different regions of the genome are exposed to different selective pressures, depending
on the genetic product for which they code (Parker et al., 1998). Even though ITS
sequences exhibit high variation rates in Volvocales, their secondary structure presents
highly conserved domains, probably because these regions affect the processing of the
rRNA primary transcript (Mai and Coleman, 1997; Coleman et al., 1998). RAPD markers,
on the other hand, are randomly generated from many loci and they can reveal genetic
differences where other markers reach their resolution limit (e.g., Van Oppen et al., 1994;
Leignel et al., 1997). Fabry et al. (1999) mentioned that RAPD data revealed substantial
differences (ca. 35%) in two African isolates of Gonium pectorale (Volvocales), which
appeared nearly identical according to intron (ypt4 and act-IX) and ITS sequences analysis
(Kohler and Fabry, unpublished results). Although in the present research both RAPD and

P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162 159
ITS analyses revealed genetic differences among the strains of D. salina studied, much
higher divergence values were obtained from RAPD data than from ITS sequences,
demonstrating the higher resolution power of RAPD.
When Gonzalez et al. (2001) sequenced the ITS spacers of the Chilean strains CONC-
006 and CONC-007, they found that the two strains did not group together as expected,
based on their geographic proximity; on the contrary, they were associated with strains of
distant geographic origins (CONC-006 with ATCC 30861 from Israel and CONC-007
with UTEX 1644 from Mexico). When more strains of D. salina are added to the analysis
(this study), the Chilean strains are again the most divergent ones, independent of the
optimality tree-building criteria used (NJ, MP and ML) (Fig. 4). On the other hand, the two
deletions in the ITS-2 reported by Gonzalez et al. (2001) as a unique character for the
strains ATCC 30861 (D. bardawil) and CONC-006, turned out to be a rule rather than an
exception in D. salina, since we detected these deletions in all of the strains analyzed,
except for CONC-001 and CONC-007 (Fig. 3, ITS-2 nucleotides 21–26 and 31–35).
When considering the physiological attributes of the analyzed strains, it should be noted
that the two strains that stand by themselves based on molecular data (this paper), namely
strains CONC-006 and Australia, exhibited contrasting physiological attributes. Thus,
CONC-006 accumulates a significantly higher amount of carotenes per cell than the
Australian strain (61.1 and 33.9 pg cell� 1, respectively); on the other hand, the Australian
strain accumulates a significantly higher amount of carotenes per culture volume than
CONC-006 (40.6 and 10 mg l� 1, respectively); this difference is due to the different
carrying capacities of both strains (Gomez and Gonzalez, unpublished data). Furthermore,
CONC-006 accumulates a-carotene (between 25% and 35%) under any culture condition
(15 or 26 jC), while the Australian strain accumulates V 10% and only at low temperature
(15 jC). The same correlation is true for all the other foreign strains (from Israel, China,
Mexico): all of them exhibited very similar physiological attributes (Gomez and Gonzalez,
unpublished data) and were grouped together based on molecular data. For comparison,
although CONC-001 and CONC-007 differed significantly in their physiological
responses (in terms of carotene content per culture volume and per cell, carotenoids/
chlorophyll ratio, and growth rate), they were grouped together based on molecular data.
The ITS data strongly support the phylogenetic closeness of CONC-001 and CONC-007,
although the RAPD data revealed a 25.5% difference between both strains. Again, RAPD
markers, as mentioned previously, can reveal genetic differences when ITS (in this case)
has reached its resolution limit.
Since intraspecific variability is a fundamental aspect in speciation processes, the
physiological diversity that exists among the strains of D. salina studied (Gomez and
Gonzalez, unpublished data) and the genetic basis that support it (this paper) might
indicate the presence of more than one species among them. Mating experiments would be
the most direct way of resolving this problem; nevertheless, due to self-mating (homo-
thalism), we have not been able to determinate interbreeding ability between the strains.
Through a series of papers, Coleman and co-workers have demonstrated that in Volvocales
(D. salina belongs to this order) there is good agreement between mating ability and ITS
sequence identity, especially in some highly conserved regions of the secondary structure
of ITS-2 (Coleman and Mai, 1997; Coleman, 1999, 2000, 2001). Future studies should be
focused on determining the secondary structure of this ribosomal spacer in the strains of D.

P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162160
salina analyzed, and looking for a qualitative type of sequence change within its highly
conserved region (hairpin loop II and III), a useful alternative in this case where the
biological species status is difficult to test by means of sexual compatibility.
Summarizing, the correlation detected between the differences in the carotenogenic
capacity of the D. salina strains and the genetic diversity among them is a clear
demonstration of the genome participation in determining these attributes. Obviously,
for mass cultivation, this is a very important result since the existence of genetically
characterized strains will minimize incorrect assumptions about the biotechnological
important traits of different strains, which could be due to phenotypic plasticity.
Acknowledgements
We thank Dr. Annette W. Coleman (Brown University, USA) for reading the
manuscript and helping us with the English. This study was supported by a grant to P.
Gomez from FONDECYT (Grant #2990047).
References
Angeler, D.G., Schagerl, M., Coleman, A.W., 1999. Phylogenetic relationships among isolates of Eudorina
species (Volvocales, Chlorophyta) inferred from molecular and biochemical data. J. Phycol. 35, 815–823.
Araneda, P., Jimenez, C., Gomez-Silva, B., 1992a. Microalgae from Northern Chile: III. Growth and h-carotenecontent of three isolates of Dunaliella salina from the Atacama Desert. Rev. Biol. Mar. Valpso. 27, 157–162.
Araneda, P., Tapia, I., Gomez-Silva, B., Tapia, L., 1992b. Microalgae from Northern Chile: II. Cultivo en medios
de bajo costo de dos cepas de Dunaliella salina (Teodoresco 1905) nativas del desierto de Atacama. Estud.
Oceanol. 11, 53–59.
Ben-Amotz, A., Avron, M., 1983. On the factors which determine massive h-carotene accumulation in the
halotolerant alga Dunaliella bardawil. Plant Physiol. 72, 593–597.
Ben-Amotz, A., Avron, M., 1989. The biotechnology of mass culturing Dunaliella for products of commercial
interest. In: Cresswell, R.C., Rees, T.A.V., Shah, N. (Eds.), Algal and Cyanobacterial Biotechnology. Long-
man Scientific and Technical Press, New York, pp. 91–114.
Ben-Amotz, A., Katz, A., Avron, M., 1982. Accumulation of h-carotene in halotolerant algae: purification and
characterization of h-carotene rich globules from Dunaliella bardawil (Chlorophyceae). J. Phycol. 18,
529–537.
Bolch, C.J.S., Orr, P., Jones, G.J., Blackburn, S.J., 1999a. Genetic, morphological and toxicological variation
among globally distributed strains of Nodularia (Cyanobacteria). J. Phycol. 35, 339–355.
Bolch, C.J.S., Blackburn, S.J., Hallegraeff, G.M., Vaillancourt, R.E., 1999b. Genetic variation among strains of
the toxic dinoflagellate Gymnodinium catenatum (Dinophyceae). J. Phycol. 35, 356–367.
Borowitzka, M., 1988. Algal growth media and sources of algal cultures. In: Borowitzka, M., Borowitzka, L.
(Eds.), Microalgal Biotechnology. Cambridge Univ. Press, London, pp. 460–461.
Borowitzka, M., Borowitzka, L., 1988. Dunaliella. In: Borowitzka, M., Borowitzka, L. (Eds.), Microalgal
Biotechnology. Cambridge Univ. Press, London, pp. 27–58, 460–461.
Caetano-Anolles, G., Gresshoff, P.M., 1997. DNA Markers: Protocols, Applications and Overviews, 1st ed.
Wiley, New York.
Cifuentes, A.S., Gonzalez, M., Conejeros, M., Dellarossa, V., Parra, O., 1992. Growth and carotenogenesis in
eight strains of Dunaliella salina Teodoresco from Chile. J. Appl. Phycol. 4, 111–118.
Cifuentes, A.S., Gonzalez, M., Parra, O., 1996a. The effect of salinity on the growth and carotenogenesis in two
Chilean strains of Dunaliella salina Teodoresco. Biol. Res. 29, 227–236.

P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162 161
Cifuentes, A.S., Gonzalez, M., Parra, O., Zuniga, M., 1996b. Cultivo de cepas de Dunaliella salina (Teodoresco
1905) en diferentes medios bajo condiciones de laboratorio. Rev. Chil. Hist. Nat. 69, 105–112.
Cifuentes, A.S., Gonzalez, M., Inostroza, I., Aguilera, A., 2001. Reappraisal of physiological attributes of nine
strains of Dunaliella (Chlorophyceae): growth and pigment content across a salinity gradient. J. Phycol. 37,
334–344.
Coleman, A.W., 1999. Phylogenetic analysis of ‘‘Volvocacae’’ for comparative genetic studies. Proc. Natl. Acad.
Sci. U. S. A. 96, 13892–13897.
Coleman, A.W., 2000. The significance of a coincidence between evolutionary landmarks found in mating
affinity and DNA sequence. Protist 151, 1–9.
Coleman, A.W., 2001. Biogeography and speciation in Pandorina/Volvulina (Chlorophyta) superclade. J. Phycol.
37, 836–851.
Coleman, A.W., Mai, J.C., 1997. Ribosomal DNA ITS-1 and ITS-2 sequence comparisons as a tool for predicting
genetic relatedness. J. Mol. Evol. 45, 168–177.
Coleman, A.W., Suarez, A., Goff, L.J., 1994. Molecular delineation of species and syngens in volvocacean green
algae (Chlorophyta). J. Phycol. 30, 80–90.
Coleman, A.W., Preparata, R.M., Mehrotra, B., Mai, J.C., 1998. Derivation of the secondary structure of the ITS-1
transcript in Volvocales and its taxonomic correlations. Protist 149, 135–146.
Fabry, S., Kohler, A., Coleman, A., 1999. Intraspecies analysis: comparison of ITS sequence data and gene intron
sequence data with breeding data for a world-wide collection of Gonium pectorale. J. Mol. Evol. 48, 94–101.
Goff, L.J., Moon, D.A., Coleman, A.W., 1994. Molecular delineation of species and species relationships in the
red algal agarophytes Gracilariopsis and Gracilaria (Gracilariales). J. Phycol. 30, 521–537.
Gomez, P., Gonzalez, M., 2001. Genetic polymorphism in eight Chilean strains of the carotenogenic microalga
Dunaliella salina Teodoresco (Chlorophyta). Biol. Res. 34, 23–30.
Gomez, P., Gonzalez, M., Becerra, J., 1999. Quantity and quality of h-carotene produced by two strains of
Dunaliella salina (Teodoresco 1905) from the North of Chile. Bol. Soc. Chil. Quım. 44, 463–468.
Gonzalez, M.A., Gomez, P., Montoya, R., 1999. Comparison of PCR-RFLP analysis of the ITS region with
morphological criteria of various strains of Dunaliella. J. Appl. Phycol. 10, 573–580.
Gonzalez, M., Coleman, A.W., Gomez, P., Montoya, R., 2001. Phylogenetic relationship among various strains of
Dunaliella (Chlorophyceae) based on nuclear ITS rDNA sequences. J. Phycol. 37, 604–611.
Hasegawa, M., Kishino, H., Yano, T., 1985. Dating the human–ape split by a molecular clock mitochondrial
DNA. J. Mol. Evol. 22, 160–174.
Karp, A., Edwards, K.J., 1998. DNA markers: a global overview. In: Caetano-Anolles, G., Gresshoff, P.M. (Eds.),
DNA Markers: Protocols, Applications and Overviews. Wiley, New York, pp. 1–14.
Kimura, M., 1980. A simple method for estimating evolutionary rates of base substitutions through comparative
studies of nucleotide sequences. J. Mol. Evol. 16, 111–120.
Kumar, S., Tamura, K., Nei, M., 1993. MEGA: Molecular Evolutionary Genetics Analysis. Version 1.01 Penn-
sylvania State University, University Park, USA, 130 pp.
Leignel, V., Humbert, J.F., Elard, L., 1997. Study by ribosomal DNA ITS-2 sequencing and RAPD analysis on
the systematics of four Metastrongylus species (Nematoda: Metastrongyloidea). J. Parasitol. 83, 606–611.
Lopez Alonso, D., Molina Grima, E., Sanchez Perez, J.A., Garcıa Sanchez, J.L., Garcıa Camacho, F., 1992a.
Fatty acid variation among different isolates of a single strain of Isochrysis galbana. Phytochemistry 31,
3901–3904.
Lopez Alonso, D., Molina Grima, E., Sanchez Perez, J.A., Garcıa Sanchez, J.L., Garcıa Camacho, F., 1992b.
Isolation of clones of Isochrysis galbana rich in eicosapentaenoic acid. Aquaculture 102, 363–371.
Lopez-Alonso, D., Segura del Castillo, C.I., Garcıa Sanchez, J.L., Sanchez Perez, J.A., Camacho, F.G., 1994.
Quantitative genetics of fatty acid variation in Isochrysis galvana (Prymnesiophyceae) and Phaeodactylum
tricornutum (Bacillariophyceae). J. Phycol. 30, 553–558.
Mai, J.C., Coleman, A.W., 1997. The internal transcribed spacer 2 exhibits a common secondary structure in
green algae and flowering plants. J. Mol. Evol. 44, 258–271.
Markovits, A., Gianelli, M.P., Conejeros, R., Erazo, S., 1993. Strain selection for h-carotene production by
Dunaliella. World J. Microbiol. Biotechnol. 9, 534–537.
Neilan, B.A., 1995. Identification and phylogenetic analysis of toxigenic cyanobacteria by multiplex randomly
amplified polymorphic DNA PCR. Appl. Environ. Microbiol. 61, 2286–2291.

P.I. Gomez, M.A. Gonzalez / Aquaculture 233 (2004) 149–162162
Nishihara, H., Mina, H., Watanabe, M., Nagashima, M., Yagi, O., Takamura, Y., 1997. Random amplified
polymorphic DNA (RAPD) analysis for discriminating genotypes of Microcystis cyanobacteria. Biosci. Bio-
technol. Biochem. 61, 1067–1072.
Parker, P.G., Snow, A.A., Schug, M.D., Booton, G.C., Fuerst, P.A., 1998. What molecules can tell us about
populations: choosing and using a molecular marker. Ecology 79, 361–382.
Schlotterer, C., 2001. Ribosomal DNA probes and primers. In: Karp, A., Isaac, P.G., Ingram, D.S. (Eds.),
Molecular Tools for Screening Biodiversity. Kluwer Academic Publishers, Netherlands, pp. 267–276.
Swofford, D.L., 1999. PAUP: Phylogenetic Analysis Using Parsimony, Version 4.0b4a. Smithsonian Institution,
Washington, DC, USA.
Valatka, S., Marinen, A., Yli-Mattila, A., 2000. Analysis of genetic diversity of Furcellaria lumbricalis (Giga-
rtinales, Rhodophyta) in the Baltic Sea by RAPD-PCR technique. Phycologia 39, 109–117.
Van Oppen, M.J.H., Diekman, O.E., Wiencke, C., Stam, W.T., Olsen, J.L., 1994. Tracking dispersal routes:
phylogeography of the arctic–antarctic disjunct seaweed Acrosiphonia arcta (Chlorophyta). J. Phycol. 30,
67–80.
Van Oppen, M.J., Klerk, H., De Graaf, M., Stam, W.T., Olsen, J.L., 1996. Assessing the limits of random
amplified polymorphic DNAs (RAPDs) in seaweed biogeography. J. Phycol. 32, 433–444.
Weising, K., Nybom, H., Wolff, K., Meyer, W., 1995. DNA Fingerprinting in Plants and Fungi. CRC Press, Boca
Raton, FL.
Welsh, J., Mc Clelland, M., 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res.
18, 7213–7218.
Williams, J.G.K., Kubelik, A.R., Livak, K.J., Rafalski, J.A., Tingey, S., 1990. DNA polymorphisms amplified by
arbitrary primers are useful as genetic markers. Nucleic Acids Res. 18, 6531–6535.




















