
THE ANTARCTIC PSYCHROPHILE, CHLAMYDOMONAS RAUDENSIS ETTL (UWO241) (CHLOROPHYCEAE, CHLOROPHYTA),...
10
THE ANTARCTIC PSYCHROPHILE, CHLAMYDOMONAS RAUDENSIS ETTL (UWO241) (CHLOROPHYCEAE, CHLOROPHYTA), EXHIBITS A LIMITED CAPACITY TO PHOTOACCLIMATE TO RED LIGHT 1 Rachael M. Morgan-Kiss 2 Department of Microbiology, University of Illinois, Urbana, Illinois 61801, USA Alexander G. Ivanov Department of Biology and The Biotron, University of Western Ontario, London, Ontario, N6A 5B7, Canada Tessa Pocock Department of Natural Science, MidSweden University, Sundsvall, Sweden MariannaKro´l Department of Biology and The Biotron, University of Western Ontario, London, Ontario, N6A 5B7, Canada Loreta Gudynaite-Savitch ECORC, Agriculture and Agri-Food Canada, Ottawa, Ontario, K1A0C6, Canada and NormanP. A. Hu ¨ner Department of Biology and The Biotron, University of Western Ontario, London, Ontario, N6A 5B7, Canada The psychrophilic Antarctic alga, Chlamydomo- nas raudensis Ettl (UWO241), grows under an ex- treme environment of low temperature and low irradiance of a limited spectral quality (blue-green). We investigated the ability of C. raudensis to accli- mate to long-term imbalances in excitation caused by light quality through adjustments in photosys- tem stoichiometry. Log-phase cultures of C. ra- udensis and C. reinhardtii grown under white light were shifted to either blue or red light for 12 h. Previously, we reported that C. raudensis lacks the ability to redistribute light energy via the short-term mechanism of state transitions. However, similar to the model of mesophilic alga, C. reinhardtii, the psychrophile retained the capacity for long-term adjustment in energy distribution between PSI and PSII by modulating the levels of PSI reaction center polypeptides, PsaA/PsaB, with minimal changes in the content of the PSII polypeptide, D1, in response to changes in light quality. The functional conse- quences of the modulation in PSI/PSII stoichiome- try in the psychrophile were distinct from those observed in C. reinhardtii. Exposure of C. raudensis to red light caused 1) an inhibition of growth and photosynthetic rates, 2) an increased reduction state of the intersystem plastoquinone pool with concom- itant increases in nonphotochemical quenching, 3) an uncoupling of the major light-harvesting com- plex from the PSII core, and 4) differential thy- lakoid protein phosphorylation profiles compared with C. reinhardtii. We conclude that the character- istic low levels of PSI relative to PSII set the limit in the capacity of C. raudensis to photoacclimate to an environment enriched in red light. Key index words: 77 K and room temperature chl fluorescence; Chlamydomonas raudensis (UWO241); light quality, PSI and PSII; photosynthetic energy balance; PQ pool; psychrophile; state transitions Abbreviations: LHCI, LHCII; light-harvesting com- plexes I and II, respectively; F o ,F m ,F v , minimal, maximal, and variable fluorescence of dark-adapted cells, respectively; F v /F m , photochemical efficiency of the dark-adapted state; F PSI , PSII , LHCII , 77 K fluo- rescence emission maxima associated with PSI, PSII, and LHCII, respectively; PQ, plastoquinone; Q A , Quinone A; q N , nonphotochemical quenching; q P , photochemical quenching; U PSII , yield of PSII electron transport Photosynthetic organisms must grow and repro- duce under highly variable light environments. The ability of an organism to modify the structure/function of the photosynthetic apparatus is paramount in main- taining maximum efficiency of light utilization while avoiding overexcitation of the photosynthetic appara- 1 Received 22 September 2004. Accepted 26 April 2005. 2 Author for correspondence: e-mail [email protected]. 791 J. Phycol. 41, 791–800 (2005) r 2005 Phycological Society of America DOI: 10.1111/j.1529-8817.2005.00094.x
Transcript of THE ANTARCTIC PSYCHROPHILE, CHLAMYDOMONAS RAUDENSIS ETTL (UWO241) (CHLOROPHYCEAE, CHLOROPHYTA),...

THE ANTARCTIC PSYCHROPHILE, CHLAMYDOMONAS RAUDENSIS ETTL (UWO241)(CHLOROPHYCEAE, CHLOROPHYTA), EXHIBITS A LIMITED CAPACITY TO
PHOTOACCLIMATE TO RED LIGHT1
Rachael M. Morgan-Kiss2
Department of Microbiology, University of Illinois, Urbana, Illinois 61801, USA
Alexander G. Ivanov
Department of Biology and The Biotron, University of Western Ontario, London, Ontario, N6A 5B7, Canada
Tessa Pocock
Department of Natural Science, MidSweden University, Sundsvall, Sweden
Marianna Krol
Department of Biology and The Biotron, University of Western Ontario, London, Ontario, N6A 5B7, Canada
Loreta Gudynaite-Savitch
ECORC, Agriculture and Agri-Food Canada, Ottawa, Ontario, K1A0C6, Canada
and
Norman P. A. Huner
Department of Biology and The Biotron, University of Western Ontario, London, Ontario, N6A 5B7, Canada
The psychrophilic Antarctic alga, Chlamydomo-nas raudensis Ettl (UWO241), grows under an ex-treme environment of low temperature and lowirradiance of a limited spectral quality (blue-green).We investigated the ability of C. raudensis to accli-mate to long-term imbalances in excitation causedby light quality through adjustments in photosys-tem stoichiometry. Log-phase cultures of C. ra-udensis and C. reinhardtii grown under white lightwere shifted to either blue or red light for 12 h.Previously, we reported that C. raudensis lacks theability to redistribute light energy via the short-termmechanism of state transitions. However, similar tothe model of mesophilic alga, C. reinhardtii, thepsychrophile retained the capacity for long-termadjustment in energy distribution between PSI andPSII by modulating the levels of PSI reaction centerpolypeptides, PsaA/PsaB, with minimal changes inthe content of the PSII polypeptide, D1, in responseto changes in light quality. The functional conse-quences of the modulation in PSI/PSII stoichiome-try in the psychrophile were distinct from thoseobserved in C. reinhardtii. Exposure of C. raudensisto red light caused 1) an inhibition of growth andphotosynthetic rates, 2) an increased reduction stateof the intersystem plastoquinone pool with concom-itant increases in nonphotochemical quenching, 3)
an uncoupling of the major light-harvesting com-plex from the PSII core, and 4) differential thy-lakoid protein phosphorylation profiles comparedwith C. reinhardtii. We conclude that the character-istic low levels of PSI relative to PSII set the limit inthe capacity of C. raudensis to photoacclimate to anenvironment enriched in red light.
Key index words: 77 K and room temperature chlfluorescence; Chlamydomonas raudensis (UWO241);light quality, PSI and PSII; photosynthetic energybalance; PQ pool; psychrophile; state transitions
Abbreviations: LHCI, LHCII; light-harvesting com-plexes I and II, respectively; Fo, Fm, Fv, minimal,maximal, and variable fluorescence of dark-adaptedcells, respectively; Fv/Fm, photochemical efficiencyof the dark-adapted state; FPSI,PSII,LHCII, 77 K fluo-rescence emission maxima associated with PSI,PSII, and LHCII, respectively; PQ, plastoquinone;QA, Quinone A; qN, nonphotochemical quenching;qP, photochemical quenching; UPSII, yield of PSIIelectron transport
Photosynthetic organisms must grow and repro-duce under highly variable light environments. Theability of an organism to modify the structure/functionof the photosynthetic apparatus is paramount in main-taining maximum efficiency of light utilization whileavoiding overexcitation of the photosynthetic appara-
1Received 22 September 2004. Accepted 26 April 2005.2Author for correspondence: e-mail [email protected].
791
J. Phycol. 41, 791–800 (2005)r 2005 Phycological Society of AmericaDOI: 10.1111/j.1529-8817.2005.00094.x

tus (Wollman 2001). This is achieved through a com-plex set of regulatory processes that are generallytermed photostasis, which in simplest terms involvesthe balance of energy absorbed and stored via photo-chemistry with energy consumed by growth andmetabolism (Oquist and Huner 2003).
Variability in the quality of available light leads topreferential excitation of either PSII or PSI (Sukeniket al. 1988, Zer et al. 1999). PSI is comprised of mostlylong-wavelength absorbing chl a, whereas PSII utilizeschl b and short-wavelength absorbing chl a (Melis1998). Plants growing under a canopy are subjectedto a light environment that is depleted in blue-greenwavelengths where PSII pigments preferentially ab-sorb. Conversely, the light quality of an aquatic envi-ronment is limited in the red wavelength region whereantenna of PSI absorb (Falkowski and LaRoche 1991,Melis 1998). Earlier studies have shown that adapta-tion to different light qualities in aquatic organisms isaccompanied by changes in the thylakoid membranecomposition, ultrastructure, and molecular organiza-tion of the photosynthetic apparatus (Wilhelm et al.1985, Hoffman and Senzer 1988, Humbeck et al.1988, Sense and Senger 1990). Under blue light, thecomponents of the intersystem electron transport arelargely reduced due to preferential excitation of PSII,whereas in an environment enriched in red light ab-sorbed mainly by PSI, the intersystem electron pool islargely in an oxidized state. Preferential excitation ofeither PSII or PSI results in an imbalance of excitationbetween the photosystems, which leads to inefficientlinear electron transport.
Short-term rapid imbalances in excitation aresensed, and the signal is transduced by changes inthe redox state of the plastoquinone (PQ) pool and theCyt b6/f complex, which compensates for this imbal-ance through the regulation of state transitions (Allenand Nilsson 1997, Finazzi et al. 1999, Wollman 2001).When the rate of PSII electron transfer exceeds therate of electron flow through PSI, the thylakoid PQpool becomes reduced. The interaction of plastoquinolwith the Qo site of the Cyt b6/f complex activates a thy-lakoid protein kinase that catalyses the phosphorylat-ion of the major light-harvesting complex II (LHCII)polypeptides (Zer et al. 1999). Phosphorylation ofLHCII induces a redistribution of LCHII to PSI atthe expense of PSII. Thus, state transitions represent ashort-term redox sensing/signaling mechanism thatregulates energy distribution between PSII/PSIthrough reversible posttranslational modification ofpreexisting polypeptides.
When redistribution of LHCII via state transitions isinsufficient to maintain the poise in light energy dis-tribution between the photosystems, long-term imbal-ances are compensated through alterations in PSII/PSIstoichiometry (Fujita and Murakami 1987, Kim et al.1993, Anderson et al. 1995, Melis et al. 1996, Allenand Pfannschmidt 2000, Pfannschmidt et al. 2001).Stoichiometric modulations in PSI/PSII are primarilydependent on variability in levels of the PSI core pro-
teins, PsaA and PsaB, whereas the accumulation of thePSII reaction center polypeptide, D1, usually remainsfairly constant (Fujita et al. 1988, Aizawa and Fujita1997, Murakami et al. 1997). Recently, Pfannschmidtand coworkers (Tullberg et al. 2000, Pfannschmidtet al. 2001) showed that the redox state of the PQpool regulates the transcription of the PSI reactioncenter chloroplast-encoded genes, psaA and psaB, aswell as the nuclear-encoded PSI gene, psaF, indicatingcooperative regulation of expression of nuclear andplastid genes. When the PQ pool is in a reduced state,expression of psaAB is activated, whereas when thepool of PQ is largely oxidized, expression of psaAB isrepressed. In contrast, Kovacs and coworkers (2000)found that stoichiometric adjustments in the ratio ofPSI/PSII in the green alga, Chlamydobotrys stellata, weremainly at the transcriptional level of the psbA and psbB(genes encoding PSII core proteins), whereas only mi-nor changes in PSI expression were observed. In ad-dition to acting as a modulator of state transitions andphotosystem stoichiometry, the redox state of the PQpool also regulates the transcription of nuclear cabgenes encoding LHCII polypeptides in Dunaliella ter-tiolecta (Escoubas et al. 1995) and Dunaliella salina(Maxwell et al. 1995b) during long-term photoaccli-mation to irradiance.
In contrast with most aquatic environments, wherethe daily light regime is strongly influenced by verticalmixing of the water column (Falkowski and LaRoche1991, Talarico and Maranzana 2000), phytoplanktonthat inhabit the perennially snow- and ice-coveredlakes of the Antarctic Dry Valleys grow under an ex-tremely stable light and temperature environment as aresult of very little turbulence and continuous daylightduring the austral summer (Priscu 1991, Lizotte andPriscu 1992a). It has been suggested that the phyto-plankton populations of the Dry Valley lakes are alwayslight limited and represent some of the most shade-adapted phytoplankton in the world (Priscu et al.1987, Lizotte and Priscu 1990, 1992b). Despite theseextremely adverse conditions, the ice-free areas of theAntarctic lakes exhibit a diversity of microbial commu-nities and have been named ‘‘Antarctic oases’’ (Priscuet al. 1998). A variety of unique adaptive strategies forlife in this extreme ecosystem has been described;however, a key survival mechanism of the microbialcommunities appears to be the ability to enter the shortgrowing season of the Antarctic austral summer withan actively growing population (Laybourn-Parry2002).
Chlamydomonas raudensis Ettl (UWO241), previouslyassumed to be Chlamydomonas subcaudata (Pocock et al.2004), is a psychrophilic green alga isolated from thelowest trophic zone of Lake Bonney, Antarctica (Nealeand Priscu 1995). The light environment is character-ized by extremely low irradiance that is always lessthan 50 mmol photons �m–2 � s–1 and a spectral distri-bution that is enriched in the blue-green region (440–580 nm) (Lizotte and Priscu 1992b). Chlamydomonas ra-udensis exhibits an unusually low chl a/b ratio (1.8–2.0)
RACHAEL M. MORGAN-KISS ET AL.792
that reflects low levels of PSI relative to PSII, a reduc-tion or absence of several LHCI polypeptides, the ab-sence of the major PSI-associated low temperature(77 K) fluorescence emission maximum at 710 nm,and high rates of PSI-driven cyclic electron transportin comparison with the model mesophilic species, C.reinhardtii (Morgan et al. 1998, Morgan-Kiss et al.2002b). The Antarctic alga also possessed higher lev-els of oligomeric LHCII in conjunction with a prom-inent PSII-associated 77 K fluorescence peak (Morganet al. 1998).
Chlamydomonas raudensis is adapted to growth at lowtemperature in combination with low irradiance en-riched in the region of 440–580 nm. Although this en-vironment is considered extreme with respect totemperature, irradiance, and light quality, it is fairlyconstant over a long-term seasonal basis. As a conse-quence of adaptation to this constant environment, C.raudensis has lost its ability for short-term regulation ofenergy distribution through state transitions (Morgan-Kiss et al. 2002b). However, has this organism retainedits ability to acclimate to long-term changes in lightquality? If so, does C. raudensis possess the ability toadjust photosystem stoichiometry similar to that ob-served for the model species, C. reinhardtii? Here, weexamine the ability of C. raudensis to acclimate to long-term changes in light quality through adjustments inPSI/PSII stoichiometry relative to the model meso-philic green alga, C. reinhardtii.
MATERIALS AND METHODS
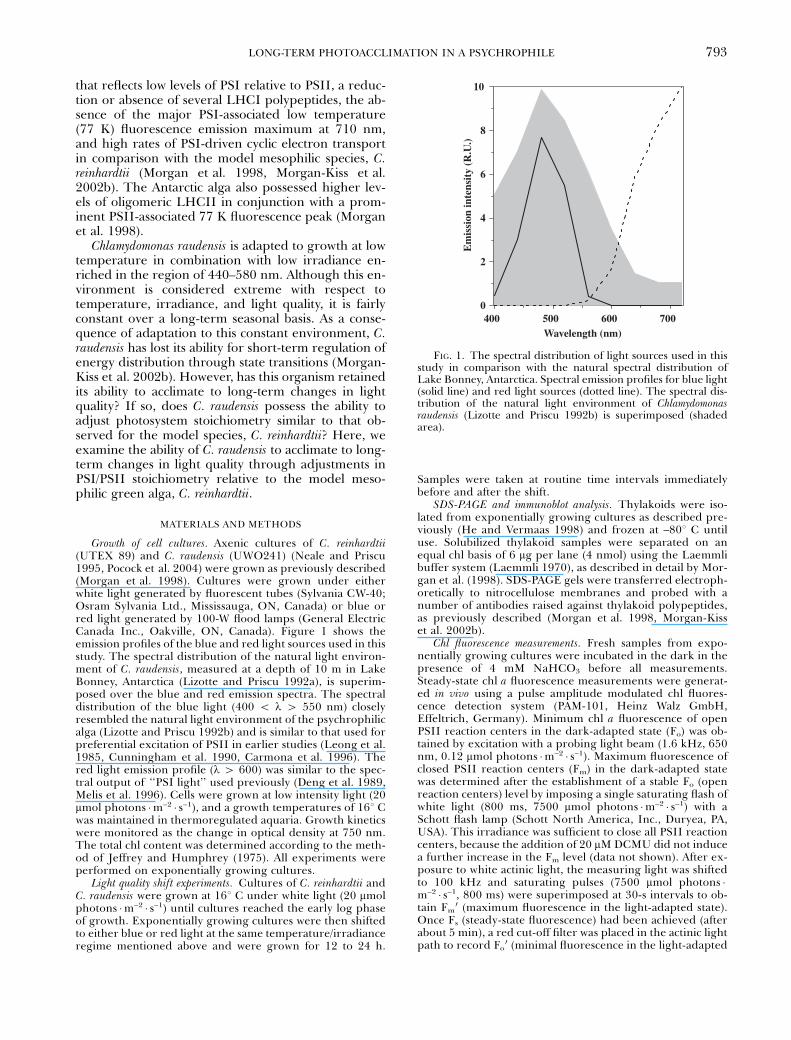
Growth of cell cultures. Axenic cultures of C. reinhardtii(UTEX 89) and C. raudensis (UWO241) (Neale and Priscu1995, Pocock et al. 2004) were grown as previously described(Morgan et al. 1998). Cultures were grown under eitherwhite light generated by fluorescent tubes (Sylvania CW-40;Osram Sylvania Ltd., Mississauga, ON, Canada) or blue orred light generated by 100-W flood lamps (General ElectricCanada Inc., Oakville, ON, Canada). Figure 1 shows theemission profiles of the blue and red light sources used in thisstudy. The spectral distribution of the natural light environ-ment of C. raudensis, measured at a depth of 10 m in LakeBonney, Antarctica (Lizotte and Priscu 1992a), is superim-posed over the blue and red emission spectra. The spectraldistribution of the blue light (400 o l 4 550 nm) closelyresembled the natural light environment of the psychrophilicalga (Lizotte and Priscu 1992b) and is similar to that used forpreferential excitation of PSII in earlier studies (Leong et al.1985, Cunningham et al. 1990, Carmona et al. 1996). Thered light emission profile (l 4 600) was similar to the spec-tral output of ‘‘PSI light’’ used previously (Deng et al. 1989,Melis et al. 1996). Cells were grown at low intensity light (20mmol photons �m–2 � s–1), and a growth temperatures of 161 Cwas maintained in thermoregulated aquaria. Growth kineticswere monitored as the change in optical density at 750 nm.The total chl content was determined according to the meth-od of Jeffrey and Humphrey (1975). All experiments wereperformed on exponentially growing cultures.
Light quality shift experiments. Cultures of C. reinhardtii andC. raudensis were grown at 161 C under white light (20 mmolphotons �m–2 � s–1) until cultures reached the early log phaseof growth. Exponentially growing cultures were then shiftedto either blue or red light at the same temperature/irradianceregime mentioned above and were grown for 12 to 24 h.
Samples were taken at routine time intervals immediatelybefore and after the shift.
SDS-PAGE and immunoblot analysis. Thylakoids were iso-lated from exponentially growing cultures as described pre-viously (He and Vermaas 1998) and frozen at –801 C untiluse. Solubilized thylakoid samples were separated on anequal chl basis of 6 mg per lane (4 nmol) using the Laemmlibuffer system (Laemmli 1970), as described in detail by Mor-gan et al. (1998). SDS-PAGE gels were transferred electroph-oretically to nitrocellulose membranes and probed with anumber of antibodies raised against thylakoid polypeptides,as previously described (Morgan et al. 1998, Morgan-Kisset al. 2002b).
Chl fluorescence measurements. Fresh samples from expo-nentially growing cultures were incubated in the dark in thepresence of 4 mM NaHCO3 before all measurements.Steady-state chl a fluorescence measurements were generat-ed in vivo using a pulse amplitude modulated chl fluores-cence detection system (PAM-101, Heinz Walz GmbH,Effeltrich, Germany). Minimum chl a fluorescence of openPSII reaction centers in the dark-adapted state (Fo) was ob-tained by excitation with a probing light beam (1.6 kHz, 650nm, 0.12 mmol photons �m–2 � s–1). Maximum fluorescence ofclosed PSII reaction centers (Fm) in the dark-adapted statewas determined after the establishment of a stable Fo (openreaction centers) level by imposing a single saturating flash ofwhite light (800 ms, 7500 mmol photons �m–2 � s–1) with aSchott flash lamp (Schott North America, Inc., Duryea, PA,USA). This irradiance was sufficient to close all PSII reactioncenters, because the addition of 20 mM DCMU did not inducea further increase in the Fm level (data not shown). After ex-posure to white actinic light, the measuring light was shiftedto 100 kHz and saturating pulses (7500 mmol photons �m–2 � s–1, 800 ms) were superimposed at 30-s intervals to ob-tain Fm
0 (maximum fluorescence in the light-adapted state).Once Fs (steady-state fluorescence) had been achieved (afterabout 5 min), a red cut-off filter was placed in the actinic lightpath to record Fo
0 (minimal fluorescence in the light-adapted
400 500 600 7000
2
4
6
8
10
Em
issi
on in
tens
ity
(R.U
.)
Wavelength (nm)
FIG. 1. The spectral distribution of light sources used in thisstudy in comparison with the natural spectral distribution ofLake Bonney, Antarctica. Spectral emission profiles for blue light(solid line) and red light sources (dotted line). The spectral dis-tribution of the natural light environment of Chlamydomonasraudensis (Lizotte and Priscu 1992b) is superimposed (shadedarea).
LONG-TERM PHOTOACCLIMATION IN A PSYCHROPHILE 793
state). Fluorescence nomenclature was applied according tovan Kooten and Snel (1990). The following parameters weregenerated from the fluorescence measurements: 1) Fv/Fm 5(Fm–Fo)/Fm, maximum photochemical efficiency of dark-adapted cells; 2) Fv
0/Fm05 (Fm
0 – Fo0)/Fm
0, maximum photo-chemical yield of light-adapted cells; 3) qp 5 (Fm
0 – Fs)/Fv0,
photochemical quenching; 4) FPSII 5 (qp � Fv0)/Fm
0, theyield of PSII electron transport; and 5) qN 5 (1 0 Fv
0)/Fv, non-photochemical quenching. All the fluorescence parameterswere calculated according to van Kooten and Snel (1990),with the exception of FPSII (Gentry et al. 1989).
Low temperature 77 K chl a fluorescence emission spectraat an excitation wavelength of 435 nm were generated fromwhole cells collected from exponentially growing cultures aswas previously described (Morgan-Kiss et al. 2002a). All emis-sion spectra were normalized to 699 nm (PSII fluorescenceemission peak) and represent the average of three independ-ent experiments.
Oxygen evolution measurements. Oxygen evolution wasmeasured in vivo in exponentially grown cultures at 161 Cand 20 mmol photons �m–2 � s–1 with a Clark type aqueousphase O2 electrode (Hansatech Ltd., King’s Lynn, UK) asdescribed by Morgan et al. (1998). Fresh samples were darkadapted as described above.
RESULTS
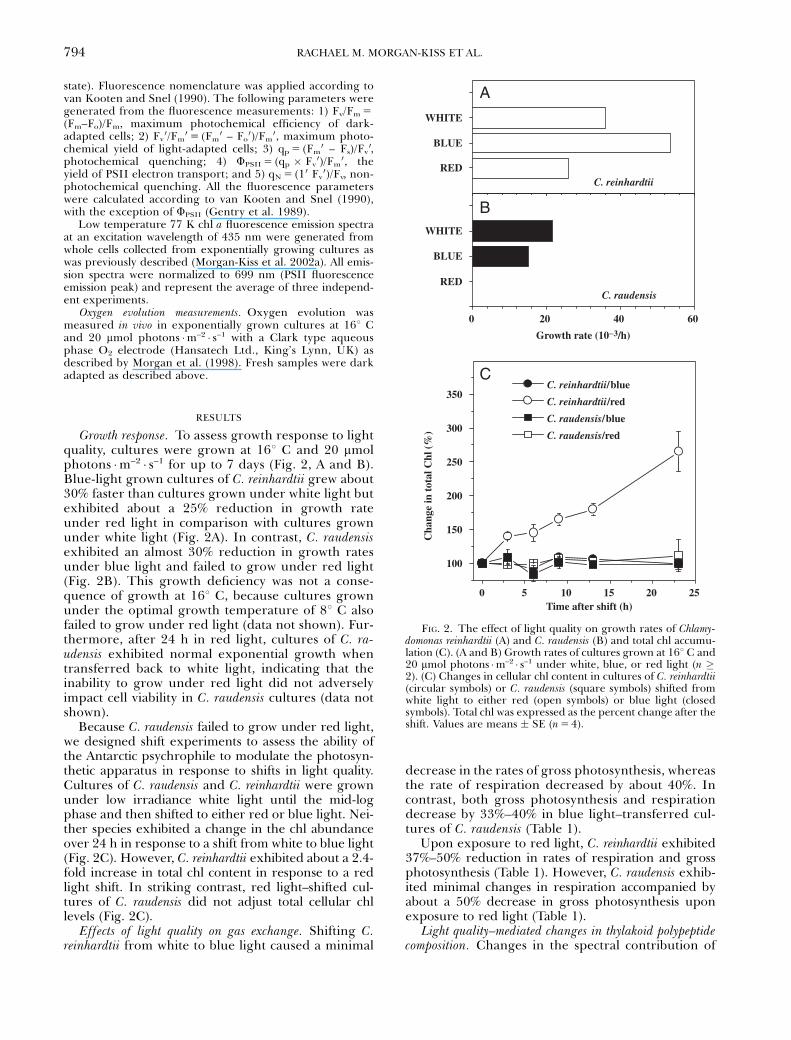
Growth response. To assess growth response to lightquality, cultures were grown at 161 C and 20 mmolphotons �m–2 � s–1 for up to 7 days (Fig. 2, A and B).Blue-light grown cultures of C. reinhardtii grew about30% faster than cultures grown under white light butexhibited about a 25% reduction in growth rateunder red light in comparison with cultures grownunder white light (Fig. 2A). In contrast, C. raudensisexhibited an almost 30% reduction in growth ratesunder blue light and failed to grow under red light(Fig. 2B). This growth deficiency was not a conse-quence of growth at 161 C, because cultures grownunder the optimal growth temperature of 81 C alsofailed to grow under red light (data not shown). Fur-thermore, after 24 h in red light, cultures of C. ra-udensis exhibited normal exponential growth whentransferred back to white light, indicating that theinability to grow under red light did not adverselyimpact cell viability in C. raudensis cultures (data notshown).
Because C. raudensis failed to grow under red light,we designed shift experiments to assess the ability ofthe Antarctic psychrophile to modulate the photosyn-thetic apparatus in response to shifts in light quality.Cultures of C. raudensis and C. reinhardtii were grownunder low irradiance white light until the mid-logphase and then shifted to either red or blue light. Nei-ther species exhibited a change in the chl abundanceover 24 h in response to a shift from white to blue light(Fig. 2C). However, C. reinhardtii exhibited about a 2.4-fold increase in total chl content in response to a redlight shift. In striking contrast, red light–shifted cul-tures of C. raudensis did not adjust total cellular chllevels (Fig. 2C).
Effects of light quality on gas exchange. Shifting C.reinhardtii from white to blue light caused a minimal
decrease in the rates of gross photosynthesis, whereasthe rate of respiration decreased by about 40%. Incontrast, both gross photosynthesis and respirationdecrease by 33%–40% in blue light–transferred cul-tures of C. raudensis (Table 1).
Upon exposure to red light, C. reinhardtii exhibited37%–50% reduction in rates of respiration and grossphotosynthesis (Table 1). However, C. raudensis exhib-ited minimal changes in respiration accompanied byabout a 50% decrease in gross photosynthesis uponexposure to red light (Table 1).
Light quality–mediated changes in thylakoid polypeptidecomposition. Changes in the spectral contribution of
RED
BLUE
WHITE
0 20 40 60
Growth rate (10−3/h)
0 5 10 15 20 25
100
150
200
250
300
350
Cha
nge
in t
otal
Chl
(%
)
Time after shift (h)
C. reinhardtii/blue
C. reinhardtii/red
C. raudensis/blue
C. raudensis/red
RED
BLUE
WHITE
C. raudensis
C. reinhardtii
B
A
C
FIG. 2. The effect of light quality on growth rates of Chlamy-domonas reinhardtii (A) and C. raudensis (B) and total chl accumu-lation (C). (A and B) Growth rates of cultures grown at 161 C and20 mmol photons �m–2 � s–1 under white, blue, or red light (n �2). (C) Changes in cellular chl content in cultures of C. reinhardtii(circular symbols) or C. raudensis (square symbols) shifted fromwhite light to either red (open symbols) or blue light (closedsymbols). Total chl was expressed as the percent change after theshift. Values are means � SE (n 5 4).
RACHAEL M. MORGAN-KISS ET AL.794

growth light are typically accompanied by adjust-ments in the stoichiometry of PSII/PSI (Kim et al.1993, Melis et al. 1996, Murakami et al. 1997, Melis1998, Pfannschmidt et al. 2001). To investigate lightquality–induced changes in the stoichiometry of ma-jor thylakoid components, relative levels of thylakoidpolypeptides were monitored over a period of 12 hafter the light shift by densitometric analysis of West-ern blots (Fig. 3). When cultures were shifted to bluelight, C. reinhardtii and C. raudensis exhibited a 1.4-and a 1.5-fold increase, respectively, in abundance ofthe major PSI reaction center proteins, PsaA/PsaB(Fig. 3A). In contrast with the blue light shift, bothChlamydomonas species exhibited a comparable75% decrease in PsaA/PsaB in response to the shiftfrom white to red light (Figs. 3A and 4). BothChlamydomonas species exhibited minimal changesin the abundance of the major PSII reactioncenter protein, D1, regardless of the light qualitytreatment (Figs. 3B and 4). These results are inagreement with previous reports in C. reinhardtiithat a red light regime induces a decrease in PSI/PSII stoichiometry (Melis et al. 1996, Murakamiet al. 1997), whereas a blue light regime induces anincrease in PSI/PSII stoichiometry (Leong et al.1985, Cunningham et al. 1990, Carmona et al.1996). Thus, it appears that adjustments in the pho-tosystem stoichiometry in response to light qualitywere mainly at the level of PSI abundance in bothChlamydomonas species.
In response to the blue light shift, both species ex-hibited a comparable increase of about 50% in LHCII,although C. reinhardtii had about a 6- to 8-h lag inLHCII accumulation (Fig. 3C). Both species also accu-mulated LHCII polypeptides under red light; howev-er, C. reinhardtii exhibited a rapid doubling in LHCIIabundance after only 3 h, whereas C. raudensis exhib-ited a slower 1.75-fold increase in LHCII over 12 h(Fig. 3C).
In the blue light–shift experiment, both speciesexhibited about a 40% increase in Cyt f abundance(Fig. 3D). In contrast, red light–shifted cultures of
C. reinhardtii exhibited a 40% increase in Cyt f,whereas C. raudensis exhibited no significant changesin the abundance of Cyt f (Fig. 3 D).
TABLE 1. Effect of light quality on maximum photochemical efficiency of dark-adapted cells (Fv/Fm), photochemical yield ofPSII (FPSII), and nonphotochemical quenching (qN) as well as rates of respiration and photosynthesis in cells of Chlamydomonasraudensis and C. reinhardtii.
Light quality treatment Fv/Fm FPSII qN Respiration Gross photosynthesis
C. reinhardtiiControl 0.69 � 0.02 0.59 � 0.02 0.17 � 0.06 –8.8 � 1.2 12.5 � 1.5Blue light (12 h) 0.65 � 0.01 0.57 � 0.07 0.15 � 0.02 –5.3 � 0.8 10.8 � 1.4Red light (12 h) 0.69 � 0.01 0.63 � 0.03 0.00 � 0.01 –5.6 � 0.3 6.8 � 0.3
C. raudensisControl 0.61 � 0.01 0.37 � 0.03 0.28 � 0.05 –9.9 � 1.2 26.2 � 0.9Blue light (12 h) 0.70 � 0.02 0.43 � 0.03 0.33 � 0.01 –5.8 � 1.4 17.5 � 2.4Red light (12 h) 0.59 � 0.03 0.31 � 0.02 0.42 � 0.02 –9.4 � 0.7 13.9 � 1.0
Cultures grown at 161 C under white light at 20 mmol photons �m–2 � s–1 (control) were transferred to either blue or red light for12 h. Fluorescence and photosynthesis parameters were measured as described in Materials and Methods at the growth regime. Rateof respiration and photosynthesis are presented as mmol O2 �mg chl–1 �h–1. Values are means � SE (n � 3).
0.5
1.0
1.5
2.0
0 5 10
CytfLHCII
PSIIPSI
A
C
BC. reinhardtii/blue C. reinhardtii/redC. raudensis/blueC. raudensis/red
0 5 10
0.5
1.0
1.5
2.0
Time after shift (h)
D
Rel
ativ
e pr
otei
n ab
unda
nce
FIG. 3. Effects of blue or red light shift on the abundance ofmajor thylakoid polypeptides in cultures of Chlamydomonas rein-hardtii (circular symbols) and C. raudensis (square symbols) grownat 161 C and 20 mmol photons �m–2 � s–1. Mid-log cultures grownin white light were shifted to either blue (closed symbols) or red(open symbols) light for 12 h. Isolated thylakoid preparationswere separated via SDS-PAGE (6 mg chl per lane), and majorpolypeptides were detected via Western blotting as described inMaterials and Methods. Blots were probed with antibodiesagainst the proteins indicated. Densitometric analysis of digitizedWestern blots was performed to estimate relative abundance ofPSI reaction center proteins, PsaA/PsaB (A); PSII reaction centerprotein, D1 (B); LHCII (C); or Cyt f (D) (n � 2).
LONG-TERM PHOTOACCLIMATION IN A PSYCHROPHILE 795
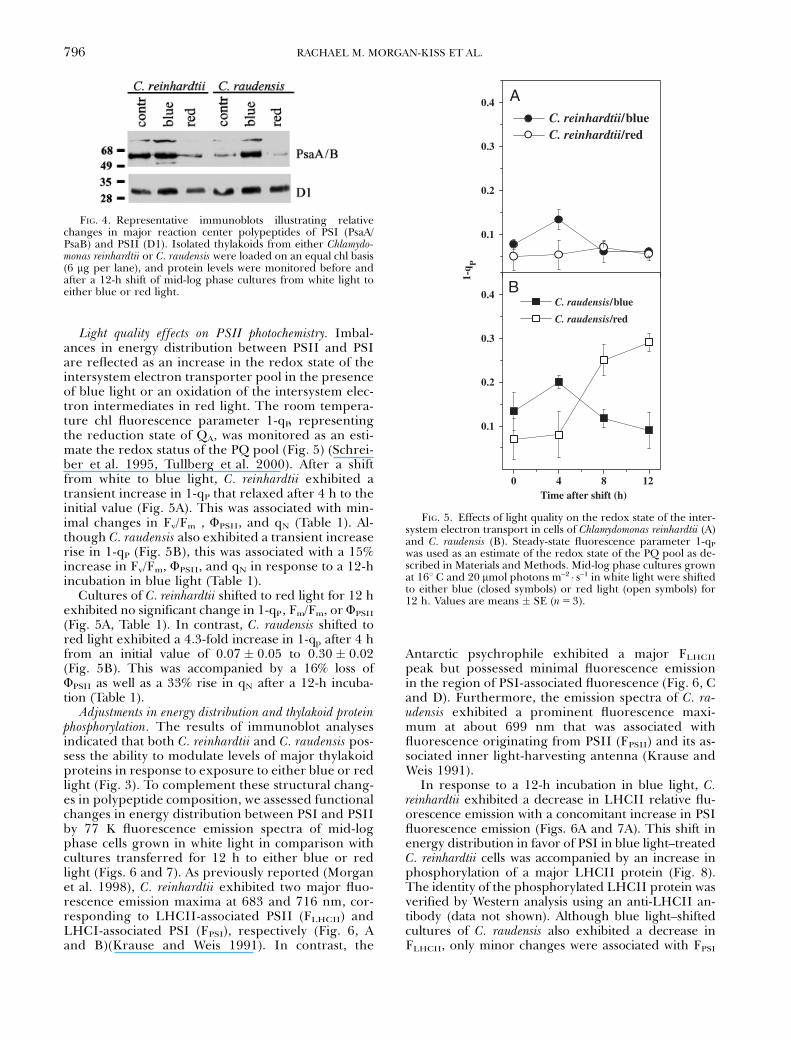
Light quality effects on PSII photochemistry. Imbal-ances in energy distribution between PSII and PSIare reflected as an increase in the redox state of theintersystem electron transporter pool in the presenceof blue light or an oxidation of the intersystem elec-tron intermediates in red light. The room tempera-ture chl fluorescence parameter 1-qP, representingthe reduction state of QA, was monitored as an esti-mate the redox status of the PQ pool (Fig. 5) (Schrei-ber et al. 1995, Tullberg et al. 2000). After a shiftfrom white to blue light, C. reinhardtii exhibited atransient increase in 1-qP that relaxed after 4 h to theinitial value (Fig. 5A). This was associated with min-imal changes in Fv/Fm , FPSII, and qN (Table 1). Al-though C. raudensis also exhibited a transient increaserise in 1-qP (Fig. 5B), this was associated with a 15%increase in Fv/Fm, FPSII, and qN in response to a 12-hincubation in blue light (Table 1).
Cultures of C. reinhardtii shifted to red light for 12 hexhibited no significant change in 1-qP, Fm/Fm, or FPSII
(Fig. 5A, Table 1). In contrast, C. raudensis shifted tored light exhibited a 4.3-fold increase in 1-qp after 4 hfrom an initial value of 0.07 � 0.05 to 0.30 � 0.02(Fig. 5B). This was accompanied by a 16% loss ofFPSII as well as a 33% rise in qN after a 12-h incuba-tion (Table 1).
Adjustments in energy distribution and thylakoid proteinphosphorylation. The results of immunoblot analysesindicated that both C. reinhardtii and C. raudensis pos-sess the ability to modulate levels of major thylakoidproteins in response to exposure to either blue or redlight (Fig. 3). To complement these structural chang-es in polypeptide composition, we assessed functionalchanges in energy distribution between PSI and PSIIby 77 K fluorescence emission spectra of mid-logphase cells grown in white light in comparison withcultures transferred for 12 h to either blue or redlight (Figs. 6 and 7). As previously reported (Morganet al. 1998), C. reinhardtii exhibited two major fluo-rescence emission maxima at 683 and 716 nm, cor-responding to LHCII-associated PSII (FLHCII) andLHCI-associated PSI (FPSI), respectively (Fig. 6, Aand B)(Krause and Weis 1991). In contrast, the
Antarctic psychrophile exhibited a major FLHCII
peak but possessed minimal fluorescence emissionin the region of PSI-associated fluorescence (Fig. 6, Cand D). Furthermore, the emission spectra of C. ra-udensis exhibited a prominent fluorescence maxi-mum at about 699 nm that was associated withfluorescence originating from PSII (FPSII) and its as-sociated inner light-harvesting antenna (Krause andWeis 1991).
In response to a 12-h incubation in blue light, C.reinhardtii exhibited a decrease in LHCII relative flu-orescence emission with a concomitant increase in PSIfluorescence emission (Figs. 6A and 7A). This shift inenergy distribution in favor of PSI in blue light–treatedC. reinhardtii cells was accompanied by an increase inphosphorylation of a major LHCII protein (Fig. 8).The identity of the phosphorylated LHCII protein wasverified by Western analysis using an anti-LHCII an-tibody (data not shown). Although blue light–shiftedcultures of C. raudensis also exhibited a decrease inFLHCII, only minor changes were associated with FPSI
FIG. 4. Representative immunoblots illustrating relativechanges in major reaction center polypeptides of PSI (PsaA/PsaB) and PSII (D1). Isolated thylakoids from either Chlamydo-monas reinhardtii or C. raudensis were loaded on an equal chl basis(6 mg per lane), and protein levels were monitored before andafter a 12-h shift of mid-log phase cultures from white light toeither blue or red light.
0.1
0.2
0.3
0.4
1-q P
Time after shift (h)
A
B
C. reinhardtii/blueC. reinhardtii/red
0 4 8 12
0.1
0.2
0.3
0.4C. raudensis/blue
C. raudensis/red
FIG. 5. Effects of light quality on the redox state of the inter-system electron transport in cells of Chlamydomonas reinhardtii (A)and C. raudensis (B). Steady-state fluorescence parameter 1-qP
was used as an estimate of the redox state of the PQ pool as de-scribed in Materials and Methods. Mid-log phase cultures grownat 161 C and 20 mmol photons m–2 � s–1 in white light were shiftedto either blue (closed symbols) or red light (open symbols) for12 h. Values are means � SE (n 5 3).
RACHAEL M. MORGAN-KISS ET AL.796

regardless of light treatment (Figs. 6C and 7C). UnlikeC. reinhardtii, these changes in low temperature fluo-rescence were not accompanied by phosphorylation ofLHCII polypeptides in C. raudensis but rather an in-creased accumulation of several high molecular mass(65–100 kDa) phosphorylated polypeptides (Fig. 8).
After a transfer to red light, cultures of C. reinhardtiiexhibited a shift in energy distribution in favor ofFLHCII relative to FPSI with no blue shift in the emis-sion maximum for FLHCII (Fig. 6B). Simultaneously,red light–transferred samples of C. reinhardtii exhibiteda decrease in phosphorylated LHCII polypeptides(Fig. 8). Although cultures of C. raudensis also respond-ed to a transfer to red light with an increase in FLHCII
(Figs. 6D and 7D), in contrast to C. reinhardtii, the risein FLHCII was accompanied by a blue shift in the flu-orescence maximum from 683 to 677 nm with minimalchanges in the region of PSI 77 K fluorescence emis-sion (Fig. 7D, inset). As mentioned above, no LHCIIphosphorylation was detectable in red light–trans-ferred samples of C. raudensis (Fig. 8).
DISCUSSION
Priscu and coworkers suggested that phytoplanktonpopulations living in the Dry Valley lakes of Antarcticaare always light limited and represent some of the mostshade-adapted phytoplankton in the world (Lizotte
and Priscu 1990, Lizotte and Priscu 1992a, 1992b).As a consequence of adaptation to such extreme envi-ronmental conditions, C. raudensis appears to have lostthe well-conserved short-term photoacclimatory mech-anism of state transitions (Morgan-Kiss et al. 2002b) aswell as the ability to adjust PSI photochemistry in re-sponse to thermal stress (Morgan-Kiss et al. 2002a). Incontrast, this psychrophilic alga has retained the ca-pacity for long-term adjustments in the stoichiometryof the major complexes of the thylakoid membrane inresponse to perturbations in the light quality. Underblue light, both C. raudensis and the control species, C.reinhardtii, exhibited comparable changes in PSI/PSIIstoichiometry due to the preferential accumulation ofPsaA/PsaB with minimal changes in the accumulationof the PSII reaction center polypeptide, D1. A similarresponse has been reported for a wide variety of pho-tosynthetic organisms (Fujita and Murakami 1987,Kim et al. 1993, Anderson et al. 1995, Melis et al.1996) and represents a major long-term mechanismfor maintaining balanced excitation of PSII and PSI(Falkowski and LaRoche 1991).
The accumulation of PsaA/PsaB in the presence ofblue light is a well-documented phenomenon in re-sponse to preferential absorption of light by PSII re-sulting in a limitation in rates of PSI-driven oxidationof the pool of mobile electron transporters. Such animbalance between overexcitation of PSII relative to
0.0
0.5
1.0
1.5
660 690 720 660 690 720
660 690 720 660 690 720
A PS
I
PS
IILH
CII
Rel
ativ
e fl
uo
resc
ence
em
issi
on
B
PS
I
PS
II
LH
CII
660 690 720
0.0
0.5
1.0
1.5C
PS
I
PS
II
LH
CII
Emission wavelength (nm)
660 690 720
D
PS
I
PS
II
LH
CII
FIG. 6. Low temperature (77 K)chl a fluorescence emission spectra ofwhole cells of Chlamydomonas reinhardtii(A and B) and C. raudensis (C and D)grown in white light and shifted to ei-ther blue or red light. Emission spectrawere collected from mid-log phase cul-tures grown in white light at 161 C and20 mmol photons �m–2 � s–1 (solid line)and after a 12-h shift to either blue (Aand C) or red (B and D) light (dottedlines). Fluorescence maxima of 683,699, and 716 nm corresponding tothe major chl-binding complexesLHCII, PSII, and PSI–LHCI, respec-tively, were identified previously (Mor-gan-Kiss et al. 2002a) and are indicatedat the top of the major peaks. Spectrawere normalized to PSII fluorescencemaximum (699 nm). Difference spectraare shown as an inset on the bottomright. Spectra are the average the threeindependent experiments.
LONG-TERM PHOTOACCLIMATION IN A PSYCHROPHILE 797

PSI results in a reduced PQ pool (Anderson et al.1995, Melis et al. 1996). The fluorescence parameter,1-qP, is an estimate of the redox state of QA that, inturn, reflects the redox state of the PQ pool (Allenet al. 1995, Maxwell et al. 1995a). Thus, the transientincrease in 1-qP observed after a 4-h incubation in bluelight in cultures of both C. reinhardtii and C. raudensis isconsistent with an overexcitation of PSII. After the 4-htime point, 1-qP levels returned to initial values, indi-cating that both the mesophilic and psychrophilic algaesensed the imbalance in excitation distribution be-tween the two photosystems and compensated forthe energy imbalance through the adjustment ofPSI/PSII stoichiometry.
We report here for the first time that the Antarcticpsychrophile, C. raudensis, is capable of long-term ad-justments in photosystem stoichiometry in response tolight quality similar to that observed by other me-sophilic green algae (Maxwell et al. 1995a, Durnfordand Falkowski 1997). However, unlike C. reinhardtii,acclimation to blue light in the Antarctic C. raudensisappeared to induce only minor changes in the energydistribution between PSI and PSII 77 K fluorescenceemission. This may be a consequence of reduced levels
of PSI-associated light-harvesting antenna, regardlessof light quality treatment and the reduction and/or lossof several LHCI polypeptides (Morgan et al. 1998).Furthermore, phosphorylated LHCII was undetecta-ble in C. raudensis thylakoids exposed to blue light (Fig.8). In addition, both the maximum PSII photochem-ical efficiency (Fv/Fm) as well as the yield of PSII elec-tron transport (FPSII) increased by 13%–15% in bluelight–transferred C. raudensis cells. Thus, C. raudensisappears to acclimate to blue light not only by adjustingPSI/PSII stoichiometry, but also by up-regulating PSIIphotochemistry. We suggest that this response inC. raudensis may be a consequence of adaptation to anatural low intensity light of blue-green wavelengths inthe Antarctic lake environment that would preferen-tially be absorbed by PSII.
Both Chlamydomonas species responded to a shift tored light by reducing the abundance of PsaA/PsaB. Adecrease in expression of PsaA/PsaB is well document-ed in response to red light and reflects a limitation atthe level of PSII excitation and turnover (Kim et al.1993, Melis et al. 1996). In agreement with previousfindings that red light induces a relatively oxidized PQpool (Pfannschmidt et al. 1999, 2001), in C. reinhardtii1-qp remained low in the red light–transferred cul-tures. Furthermore, in addition to structural adjust-ments to red light at the level of PSI, cultures ofC. reinhardtii also exhibited a rapid accumulation of
0.4
0.8
1.2
1.6
C. reinhardtii/redC. reinhardtii/blue
DC
A B
F
F
Rel
ativ
e fl
uore
scen
ce y
ield
0 4 8 12
0.4
0.8
1.2
1.6
C. raudensis/blue
Time after shift (h)0 4 8 12
C. raudensis/red
FIG. 7. Kinetics of modulations in major 77 K chl a fluores-cence emission maxima measured in Chlamydomonas reinhardtii (Aand B) or C. raudensis (C and D) after a shift from white light toeither blue (A and C) or red light (B and D) in whole cells ofC. reinhardtii and C. raudensis. Fluorescence emission maximawere expressed as relative fluorescence emission units of eitherLHCII fluorescence at 683 nm (squares) or PSI fluorescence at716 nm (circles) normalized to the 699-nm peak correspondingto PSII. Values are means � SE (n 5 3).
FIG. 8. Western blot showing phosphorylation patterns ofmajor thylakoid proteins in response to either a red or a bluelight quality shift. Thylakoid membranes were isolated from cellsof Chlamydomonas reinhardtii and C. raudensis according to He andVermaas (1998), separated via SDS-PAGE, and transferred tonitrocellulose as described in Materials and Methods. Blots wereprobed with antibodies raised against phosphothreonine (BioradLaboratories, Mississauga, ON, Canada). Cultures were grownuntil log phase in white light and then transferred to either blueor red light. Samples were loaded on an equal chl basis (6 mg perlane).
RACHAEL M. MORGAN-KISS ET AL.798

total chl and LHCII polypeptides, which has been re-ported in other green algae under conditions wherethe PQ pool is oxidized (Escoubas et al. 1995, Maxwellet al. 1995b). Changes in the functional organization ofthe photosynthetic apparatus of C. reinhardtii were alsoaccompanied by a substantial decrease in both respi-ration and photosynthetic rates in the red light–trans-ferred cultures. This is probably a consequence ofgrowth at a light quality of low intensity and low en-ergy wavelengths and probably accounts for the de-crease in growth rates under red light in cultures ofC. reinhardtii.
In striking contrast to C. reinhardtii, incubation ofC. raudensis cultures under red light caused a 4-foldincrease in 1-qP, indicative of an increase in the reduc-tion state of the PQ pool. How can we account for anapparent reduced PQ pool under red light in thepsychrophile? It is clear from our previous studiesthat an unusual characteristic of the photosyntheticapparatus of C. raudensis is its inherently low content ofPSI relative to PSII even when grown under whitelight (Morgan et al. 1998, Morgan-Kiss et al. 2002a,b).Therefore, upon exposure to red light, it is possiblethat the further loss of PSI via the decrease in abun-dance of PsaA/PsaB reaction center proteins resulted ina photochemical apparatus that was severely limited atthe level PSI. This is consistent with the observed in-hibition of growth and photosynthesis in C. raudensisexposed to red light.
To protect the photosynthetic apparatus of C. ra-udensis from over-reduction, the increase in 77 K flu-orescence emission at 677 nm indicates that LHCII isfunctionally detached from PSII, which decreasesthe efficiency of energy transfer from LHCII to thePSII core complex (Krause and Weis 1991). Thesedata are consistent with the fact that FPSII and ratesof gross photosynthesis declined by 16% and 70%,respectively, after a 12-h exposure to red light inC. raudensis with a concomitant 1.5-fold increase inqN. Thus, although C. raudensis can adjust PSI/PSIIstoichiometry, these results together with the inhibitionof growth under red light indicate that the psych-rophile is much more limited than C. reinhardtii inits capacity to maintain growth and photosynthesiswhen PSI is preferentially excited. We suggest thatthis is probably due to the inherently low levels of PSIrelative to PSII in C. raudensis (Fig. 4) (Morgan et al.1998).
In conclusion, the Antarctic psychrophile, C. ra-udensis (UWO241), has retained the capacity for long-term adjustment in energy distribution between PSIand PSII by modulating photosystem stoichiometry inresponse to light quality. The loss of short-term acc-limatory responses (such as state transitions; Morgan-Kiss et al. 2002b) in the psychrophile most likely re-flects the fact that although Lake Bonney of the Ant-arctic Dry Valleys is an extremely harsh aquaticecosystem, C. raudensis has adapted to an environmentthat is relatively constant with respect to light qualityand temperature. Finally, the functional consequences
of the modulation in PSI/PSII stoichiometry in thepsychrophile are distinct from those observed in themesophilic alga, C. reinhardtii, based growth rates, 77 Kfluorescence emission, the redox state of the PQ pool,and thylakoid protein phosphorylation. We suggestthat this reflects the inherently low levels of PSI rela-tive to PSII, which sets the limit for the capacity ofC. raudensis to photoacclimate to an environmentenriched in red light.
We thank Dr. J. Priscu for his generous gift of an axenic cul-ture of Chlamydomonas raudensis. This work was supported bythe National Science and Engineering Council of Canada.
Aizawa, K. & Fujita, Y. 1997. Regulation of synthesis of PSI in thecyanophytes Synechocystis PCC6714 and Plectonema boryanumduring the acclimation of the photosystem stoichiometry to thelight quality. Plant Cell Physiol. 38:319–26.
Allen, J. F., Alexciev, K. & Kakansson, G. 1995. Regulation by re-dox signalling. Curr. Biol. 5:869–71.
Allen, J. F. & Nilsson, A. 1997. Redox signalling and the structuralbasis of regulation of photosynthesis by protein phosphorylat-ion. Physiol. Plant. 100:863–8.
Allen, J. F. & Pfannschmidt, T. 2000. Balancing the two photosys-tems: photosynthetic electron transfer governs transcription ofreaction centre genes in chloroplasts. Philos. Trans. R. Soc.Lond. B Biol. Sci. 355:1351–9.
Anderson, J. M., Chow, W. S. & Park, Y. 1995. The grand design ofphotosynthesis: acclimation of the photosynthetic apparatus toenvironmental cues. Photosynth. Res. 46:129–39.
Carmona, R., Vergara, J. J., Perez-Llorens, J. L., Figueroa, F. L. &Niell, F. X. 1996. Photosynthetic acclimation and biochemicalresponses of Gelidium sesquipedale cultured in chemostats underdifferent qualities of light. Mar. Biol. 127:25–34.
Cunningham, F. X. J., Dennenberg, R. J., Jusinic, P. A. & Grant, E.1990. Growth under red light enhances photosystem II rela-tive to photosystem I and phycobilisomes in red algaPorphyridium cruetum. Plant Physiol. 93:88–895.
Deng, X.-W., Tonkyn, J. C., Peter, G. F., Thornber, J. P. & Gruissem,W. 1989. Post-transcriptional control of plastid mRNA accu-mulation during adaptation of chloroplasts to different lightquality environments. Plant Cell 1:645–54.
Durnford, D. G. & Falkowski, P. 1997. Chloroplast redox regula-tion of nuclear gene transcription during photoacclimation.Photosynth. Res. 266:229–41.
Escoubas, J., Lomas, M., LaRoche, J. & Falkowski, P. G. 1995. Lightintensity regulation of cab gene transcription is signalled by theredox state of the plastoquinone pool. Proc. Natl. Acad. Sci.USA 92:10237–41.
Falkowski, P. G. & LaRoche, J. 1991. Acclimation to spectral irra-diance in algae. J. Phycol. 27:8–14.
Finazzi, G., Furia, A., Barbagallo, R. P. & Forti, G. 1999. State tran-sitions, cyclic and linear electron transport and photo-phosphorylation in Chlamydomonas reinhardtii. Biochim.Biophys. Acta 1413:117–29.
Fujita, Y. & Murakami, A. 1987. Regulation of electron transportcomposition in cyanobacterial photosynthetic system: stoic-hiometry among photosystem I and II complexes and theirlight-harvesting antennae and cytochrome b6/f complex. PlantCell Physiol. 28:1547–53.
Fujita, Y., Murakami, A., Ohki, K. & Hagiwara, N. 1988. Regula-tion of photosystem composition in cyanobacterial photosyn-thetic system: evidence indicating that photosystem Iformation is controlled in response to the electron transportstate. Plant Cell Physiol. 29:557–64.
Gentry, B., Briantais, J. & Baker, N. R. 1989. The relationship be-tween the quantum yield of photosynthetic electron transportand quenching of chlorophyll fluorescence. Biochim. Biophys.Acta 990:87–92.
LONG-TERM PHOTOACCLIMATION IN A PSYCHROPHILE 799

He, Q. & Vermaas, W. 1998. Chlorophyll a availability affects psbAtranslation and D1 precursor processing in vivo in Synechocystissp. PCC 6803. Proc. Natl. Acad. Sci. USA 95:5830–5.
Hoffman, B. & Senzer, H. 1988. Kinetics of photosynthetic appa-ratus adaptation in Scenedesmuus obliquus to changes in irradi-ance and light quality. Photochem. Photobiol. 47:737–9.
Humbeck, K., Hoffman, B. & Senzer, H. 1988. Influence ofenergy flux and quality of light on the molecular organizationof the photosynthetic apparatus in Scenedesmus. Planta 173:205–12.
Jeffrey, S. W. & Humphrey, G. F. 1975. New spectrophotometricequations for determining chlorophyll a, b, c1, c2 in higherplants, algae and natural phytoplankton. Biochem. Physiol.Pflanz 167:191–4.
Kim, J. H., Glick, R. E. & Melis, A. 1993. Dynamics of photosystemstoichiometry adjustment by light quality in chloroplasts. PlantPhysiol. 102:181–90.
Kovacs, L., Wiessner, W., Kis, M., Naagy, F., Mende, D. & Demeter,S. 2000. Short- and long-term redox regulation of photosyn-thetic light energy distribution and photosystem stoichiometryby acetate metabolism in the green alga, Chlamydobotrys stellata.Photosynth. Res. 65:231–47.
Krause, G. H. & Weis, E. 1991. Chlorophyll fluorescence and pho-tosynthesis: the basics. Annu. Rev. Physiol. Plant Mol. Biol. 42:313–49.
Laemmli, U. K. 1970. Cleavage of structural proteins duringthe assembly of the head of bacteriophage T4. Nature227:680–5.
Laybourn-Parry, J. 2002. Survival mechanisms in Antarctic lakes.Philos. Trans. R. Soc. Lond. B Biol. Sci. 357:863–9.
Leong, T.-Y., Goodchild, D. J. & Anderson, J. M. 1985. Effect oflight quality on the composition, function, and structure ofphotosynthetic thylakoid membranes of Asplenium austral-asicum (Sm.) hook. Plant Physiol. 78:561–7.
Lizotte, M. P. & Priscu, J. C. 1990. Photosynthesis-irradiance rela-tionships in phytoplankton from Lake Bonney. Antarc. J. U.S.25:223–4.
Lizotte, M. P. & Priscu, J. C. 1992a. Photosynthesis-irradiance re-lationships in phytoplankton from the physically stable watercolumn of a perennially ice-covered lake (Lake Bonney, Ant-arctica). J. Phycol. 28:179–85.
Lizotte, M. P. & Priscu, J. C. 1992b. Spectral irradiance and bio-optical properties in perennially ice-covered lakes of the dryvalleys (McMurdo Sound, Antarctica). Antarc. Res. Ser. 57:1–14.
Maxwell, D. P., Falk, S. & Huner, N. P. A. 1995a. Photosystem IIexcitation pressure and development of resistance to photo-inhibition. Plant Physiol. 107:687–94.
Maxwell, D. P., Laudenbach, D. E. & Huner, N. P. A. 1995b. Redoxregulation of light-harvesting complex II and cab mRNAabundance in Dunaliella salina. Plant Physiol. 109:787–95.
Melis, A. 1998. Photostasis in plants: mechanisms and regulation.In Thistle, W. [Ed.] Photostasis and Related Phenomena. LenumPress, New York, pp. 207–21.
Melis, A., Murakami, A., Nemson, J. A., Aizawa, K., Ohki, K. &Fujita, Y. 1996. Chromatic regulation in Chlamydomonas rein-hardtii alters photosystem stoichiometry and improves thequantum efficiency of photosynthesis. Photosynth. Res.47:253–65.
Morgan, R. M., Ivanov, A. G., Priscu, J. C., Maxwell, D. P. & Huner,N. P. A. 1998. Structure and composition of the photochemicalapparatus of the Antarctic green alga, Chlamydomonas subcau-data. Photosynth. Res. 56:303–14.
Morgan-Kiss, R., Ivanov, A. G., Williams, J., Mobashsher, K. &Huner, N. P. 2002a. Differential thermal effects on the energydistribution between photosystem II and photosystem I inthylakoid membranes of a psychrophilic and a mesophilic alga.Biochim. Biophys. Acta 1561:251–65.
Morgan-Kiss, R. M., Ivanov, A. G. & Huner, N. P. 2002b. The Ant-arctic psychrophile, Chlamydomonas subcaudata, is deficient instate I-state II transitions. Planta 214:435–45.
Murakami, A., Fujita, Y., Nemson, J. A. & Melis, A. 1997. Chro-matic regulation in Chlamydomonas reinhardtii: time course of
photosystem stoichiometry adjustment following a shift ingrowth light quality. Plant Cell Physiol. 38:188–93.
Neale, P. J. & Priscu, J. C. 1990. Structure and function of thephotochemical apparatus in the phytoplankton of ice-coveredLake Bonney. Antarc. J. U.S. 25:225–34.
Neale, P. J. & Priscu, J. C. 1995. The photosynthetic apparatus ofphytoplankton from a perennially ice-covered Antarctic lake:acclimation to an extreme shade environment. Plant CellPhysiol. 36:253–63.
Oquist, G. & Huner, N. P. 2003. Photosynthesis of overwinteringevergreen plants. Annu. Rev. Plant Physiol. Plant Mol. Biol.54:329–55.
Pfannschmidt, T., Nilsson, A., Tullberg, A., Link, G. & Allen, J. F.1999. Direct transcriptional control of the chloroplast genespsbA and psaAB adjusts photosynthesis to light energy distri-bution in plants. IUBMB Life 48:271–6.
Pfannschmidt, T., Schutze, K., Brost, M. & Oelmuller, R. 2001. Anovel mechanism of nuclear photosynthesis gene regulationby redox signals from the chloroplast during photosystem sto-ichiometry adjustment. J. Biol. Chem. 276:36125–33.
Pocock, T., Lachance, M.-A., Proschold, T., Priscu, J. C., Kim, S. &Huner, N. P. A. 2004. Identification of a psychrophilicgreen alga from Lake Bonney Antarctica: Chlamydomonasraudensis Ettl. (UWO 241) (Chlorophyceae). J. Phycol.40:1138–48.
Priscu, J. C. 1991. Variation in light attenuation by the permanentice cap of Lake Bonney during spring and summer. Antarc. J.U.S. 26:223–4.
Priscu, J. C., Fritsen, C. H., Adams, E. E., Giovannoni, S. J., Paerl,H. W., McKay, C. P., Doran, P. T., Gordon, D. A., Lanoil, B. D.& Pinckney, J. L. 1998. Perennial Antarctic lake ice: an oasisfor life in a polar desert. Science 280:2095–8.
Priscu, J. C., Priscu, L. R., Vincent, W. F. & Howard-Williams, C.1987. Photosynthate distribution by microplankton in perma-nently ice-covered Antarctic desert lakes. Limnol. Oceanogr.32:260–70.
Schreiber, U., Hormann, H., Neubauer, C. & Klughammer, C.1995. Assessment of photosystem II photochemical quantumyield by chlorophyll fluorescence quenching analysis. Aust. J.Plant Physiol. 22:209–20.
Sense, M. & Senger, H. 1990. Functional changes in the photosyn-thetic apparatus during light adaptation of the green algaChlorella fusca. J. Photochem. Photobiol. B Biol. 8:63–71.
Sukenik, A., Bennett, J. & Falkowski, P. 1988. Changes in theabundance of individual apoproteins of light-harvesting chlo-rophyll a/b-protein complexes of photosystem I and II withgrowth irradiance in the marine chlorophyte Dunaliella ter-tiolecta. Biochim. Biophys. Acta 932:206–15.
Talarico, L. & Maranzana, G. 2000. Light and adaptive responsesin red macroalgae: an overview. J. Photochem. Photobiol. B 56:1–11.
Tullberg, A., Alexciev, K., Pfannschmidt, T. & Allen, J. F. 2000.Photosynthetic electron flow regulates transcription of thepsaB gene in pea (Pisum sativum L.) chloroplasts through theredox state of the plastoquinone pool. Plant Cell Physiol.41:1045–54.
van Kooten, O. & Snel, J. F. H. 1990. The use of chlorophyll flu-orescence nomenclature in plant stress physiology. Photosynth.Res. 25:147–50.
Wilhelm, C., Kramer, P. & Wild, A. 1985. Effects of different lightqualities on the ultrastructure, thylakoid membrane composi-tion and assimilation metabolism of Chlorella fusa. Physiol.Plant. 64:359–64.
Wollman, F. A. 2001. State transitions reveal the dynamics andflexibility of the photosynthetic apparatus. EMBO J. 20:3623–30.
Zer, H., Vink, M., Keren, N., Dilly-Hartwig, H. G., Paulsen, H.,Herrmann, R. G., Andersson, B. & Ohad, I. I. 1999. Regula-tion of thylakoid protein phosphorylation at the substratelevel: reversible light-induced conformational changes exposethe phosphorylation site of the light-harvesting complex II.Proc. Natl. Acad. Sci. USA 96:8277–82.
RACHAEL M. MORGAN-KISS ET AL.800




















