
HAT-RAPD profiling of seven fungi isolated from central Kazakhstan soils A R T I C L E I N F O
7
17 South Asian J Exp Biol; 4 (1): 17-23; 2014 ISSN: 2230-9799 Vol. 4, Issue 1, Page 17-23 hp://www.sajeb.org REGULAR ARTICLE HAT-RAPD profiling of seven fungi isolated from central Kazakhstan soils Konstann Li * , Assel Moldagulova, Eleonora Bekenova, Abzal Kazhibaev, Markhabbat Kairova, Maira Urazova, Serik Shaikhin, Kairtay Almagambetov Republic Collecon of Microorganisms (RCM) SC MES, Valikhanov st.13/1, Astana, Republic of Kazakhstan, 010000 ARTICLE INFO Arcle History: Received: 23 Feb 2014 Revised: 11 Mar 2014 Accepted: 12 Mar 2014 *Corresponding Author: Email: [email protected] Tel/FAX: 07-7172-20-07-93 Keywords: HAT-RAPD, arbitrary pri- mers, hyphomycetes, PCR, am- plicon, RAPD-profile ABSTRACT Stable HAT-RAPD (high annealing temperature) profiles were obtained for 7 micromycetes isolated from the central Kazakhstan soils with single non- specific primers. An arbitrary primer (decamer AP3), a primer derived from the intergenic spacer regions (T3B) and microsatellite primer (GTG) 5 were applied in PCR. Precise and reproducible profiles of the amplificaon prod- ucts were obtained for all seven hyphomycetes isolates in spite of increased annealing temperature applicaon. The number and size of amplicons reflect a species specificity. In some cases intraspecific variability was observed. Cluster analysis of the profiles demonstrated the phylogenec relaonships between isolates that are consistent with those of ITS-region genotyping. Obtained results completely jusfy the applicaon of the HAT-RAPD in mark- ing and rapid idenficaon of soil hyphomycetes. 1. Introducon Reliable labeling of potenally important and cul- vated with the praccal goals microscopic fungi is an urgent task for both staonary collecons as well as for temporary depositories established un- der separate research organizaons or projects. Stable properes of interest maintenance involves ensuring the stability of the microorganism as a whole, taking in account oſten pleiotropic effect of even small changes at the genec level. However, large number of morphological, biochemical and genec characteriscs permanent monitoring of the research objects is not always jusfied both scienfically and economically. Therefore, methods providing informaon about the genome stability as a whole without linking them with detectable changes in the specific properes of the organism are very useful. Currently, genec typing of microorganisms is widely used in several major fields of microbiologi- cal research. Taxonomy, research aimed at elucida- on of evoluonary dynamics or phylogenec rela- onships, populaon genecs of microorganisms, and microbial epidemiology all rely on ge- nec typing data for discriminaon between geno- types. Apart from being an essenal component of these fundamental sciences microbial typing clearly affects several areas of applied microbiological re- search. It cannot be overemphasized that limita- ons exist in both the applicability of the different techniques and their results when they are used to categorize microbial isolates. Obviously, typing da- ta should reveal differences not only between strains belonging to different species but also be-
Transcript of HAT-RAPD profiling of seven fungi isolated from central Kazakhstan soils A R T I C L E I N F O

17
South Asian J Exp Biol; 4 (1): 17-23; 2014
ISSN: 2230-9799 Vol. 4, Issue 1, Page 17-23 http://www.sajeb.org REGULAR ARTICLE
HAT-RAPD profiling of seven fungi isolated from central Kazakhstan soils Konstantin Li*, Assel Moldagulova, Eleonora Bekenova, Abzal Kazhibaev, Markhabbat Kairova, Maira Urazova, Serik Shaikhin, Kairtay Almagambetov Republic Collection of Microorganisms (RCM) SC MES, Valikhanov st.13/1, Astana, Republic of Kazakhstan, 010000
A R TI C L E I N F O Article History: Received: 23 Feb 2014 Revised: 11 Mar 2014 Accepted: 12 Mar 2014 *Corresponding Author: Email: [email protected] Tel/FAX: 07-7172-20-07-93 Keywords: HAT-RAPD, arbitrary pri-mers, hyphomycetes, PCR, am-plicon, RAPD-profile
A B S TR A C T Stable HAT-RAPD (high annealing temperature) profiles were obtained for 7 micromycetes isolated from the central Kazakhstan soils with single non-specific primers. An arbitrary primer (decamer AP3), a primer derived from the intergenic spacer regions (T3B) and microsatellite primer (GTG)5 were applied in PCR. Precise and reproducible profiles of the amplification prod-ucts were obtained for all seven hyphomycetes isolates in spite of increased annealing temperature application. The number and size of amplicons reflect a species specificity. In some cases intraspecific variability was observed. Cluster analysis of the profiles demonstrated the phylogenetic relationships between isolates that are consistent with those of ITS-region genotyping. Obtained results completely justify the application of the HAT-RAPD in mark-ing and rapid identification of soil hyphomycetes.
1. Introduction
Reliable labeling of potentially important and culti-vated with the practical goals microscopic fungi is an urgent task for both stationary collections as well as for temporary depositories established un-der separate research organizations or projects. Stable properties of interest maintenance involves ensuring the stability of the microorganism as a whole, taking in account often pleiotropic effect of even small changes at the genetic level. However, large number of morphological, biochemical and genetic characteristics permanent monitoring of the research objects is not always justified both scientifically and economically. Therefore, methods providing information about the genome stability as a whole without linking them with detectable changes in the specific properties of the organism
are very useful.
Currently, genetic typing of microorganisms is widely used in several major fields of microbiologi-cal research. Taxonomy, research aimed at elucida-tion of evolutionary dynamics or phylogenetic rela-tionships, population genetics of microorganisms, and microbial epidemiology all rely on ge-netic typing data for discrimination between geno-types. Apart from being an essential component of these fundamental sciences microbial typing clearly affects several areas of applied microbiological re-search. It cannot be overemphasized that limita-tions exist in both the applicability of the different techniques and their results when they are used to categorize microbial isolates. Obviously, typing da-ta should reveal differences not only between strains belonging to different species but also be-

18
Li et al., South Asian J Exp Biol; 4 (1): 17-23; 2014
tween strains belonging to a same species (Van Belkum et al., 2001).
In this regard, reproducible RAPD-profiles can be convenient and informatory indicators of collection cultures stability at the genome level. They can also contribute to the rapid identification of the micro-organisms provided free access to previously estab-lished databases available online (e.g. Kazusa, SVMD, Q-bank etc.).
RAPD modification using increased primer anneal-ing temperature is of particular interest – HAT-RAPD (high annealing temperature) (Anuntalabhochai et al., 2000). More recent studies stated that at high annealing temperature (increase annealing temperature to 46–48°C), the random amplified polymorphic DNA (HAT-RAPD) technique results in greater polymorphism, higher reproduci-bility and higher resolution (Wongsawad et al., 2009).
Studies of different ecological groups of filamen-tous fungi that rapidly adapt to changing environ-mental conditions can be attributed to topical and prospective studies related to the problems of global warming and climate change on our planet.
So objects of close attention of microbiologists are the permanent inhabitants of soil layer, which may include species of such genera as Entomophthora, Isaria, Lecanicillium (Khan et al., 2012).
In addition, intensive work is carried out with rep-resentatives of two genera - Metarhizium and Beauveria (Parsal et al., 2013; Pava-Ripoll et al., 2011).
Entomopathogenic fungi naturally occur in insect hosts as infections; they can be collected in the field and grown in the laboratory for the documen-tation of the fungus. Studies on the prevalence of fungi in insects have usually been limited to species that are pests or are important non-target species such as certain predators and parasitoids. Howev-er, it is likely that almost any major insect taxon collected intensively will be found to be a natural host for B. bassiana in temperate regions (Shahid et al., 2012).
Thus hyphomycetes DNA labeling along with tradi-tional identification methods (morphology, bio-chemistry, genotyping by ribosomal RNA genes) is widely used to characterize microorganisms during their long storage, as well as their isolation from environmental conditions de novo. The latter is es-
pecially important because genetic polymorphism of fungi is a significant reserve to enhance their various biological activities, including entomopath-ogenicity.
In the present work we studied the applicability of the HAT-RAPD to characterize the group of hypho-mycetes isolated from a soil substrate and charac-terized by morphological, cultural and genotypic levels.
2. Materials and Methods
2.1. Hyphomycetes isolation and cultivation
Filamentous fungi were isolated from the Akmola region (Central Kazakhstan) soil samples. 200 g of ground was taken from a soil surface layer in the neighborhood of Karaotkel village. Soil sample site was represented as a piece of typical steppe out of agricultural use. For the selection of filamentous fungi stable forms from natural population the method of stepwise monospore passages selection was used. The sample site of the study area was characterized by similar conditions (topography, soil structure and uniformity of vegetation, type of economic use).
Isolation of micromycetes was carried out by plating on solid nutrient medium in three stages: dilution of soil extracts, seeding on solid medium and counting of colonies. For the fungal surface cultivation Sabouraud dextrose agar, Czapek and potato agar media were used (HiMedia Laborato-ries Pvt, Ltd., India). Colonies of hyphomycetes was counted after 5-15 days of incubation depending on the speed of growth at +24-26°C in the dark.
Fungal colonies were transferred to standard iden-tification media and repeatedly plated on agar me-dia to get pure fungi cultures and saved on slop-agar tubes. Further, to identify pure isolates of fun-gi in the level of genus macro- and micromorpho-logical features analysis was performed according to conventional microbiological methods of re-search and by using of standard identification man-ual (Gilman, 1988).
2.2. Genome DNA isolation
Fungi were cultured in the Sabouraud liquid medi-um for 3-5 days at 25°C on a shaker at 100 rpm, followed by centrifugation for the mycelium har-vesting. Mycelium biomass was washed twice with sterile distilled water, lyophilized and stored at -20°C (Fegan et al., 1993). After lyophilization, the ly-

19
Li et al., South Asian J Exp Biol; 4 (1): 17-23; 2014
sate was triturated in microcentrifuge tubes in a liquid nitrogen and then further extraction of the DNA was carried out by different methods (Camberery et al., 1993; Yoon et al., 1991; Pfeifer et al., 1993; Bischoff et al., 2006; Möller et al., 1992).
2.3. ITS-region sequencing
For genetic identification of different fungal isolates the fungus-specific PCR-forward primer ITS 1 and the reverse primer ITS 4 (Sigma) were used and DNA amplification is done by using a thermal iCycler IQ5 (BioRad, USA). The sequencing of the amplified DNA fragments was carried out using of BigDye® Terminator v3.1 Ready Reaction Cycle Se-quencing Kit on the ABI PRISM 3730 DNA analyzer (Applied Biosystems Inc., USA) according to the manufacturer’s instructions. The nucleotide se-quence was determined for approximately 560 base region of the ITS and 5.8S rDNA gene from each of fungal isolates. The resulted DNA sequenc-es from all isolated genomes were aligned and ana-lyzed using Vector NTI software Version 10 (Invitrogen, USA.) and compared by the BLAST search program from the public data library of the National Center for Biotechnology Information.
2.4. Arbitrary primers RAPD-profiling
To verify the suitability of the selected primers for RAPD-profiling of fungal isolates genomic DNA sam-ples were used in amplification reactions with all the primers presented in Table 1 (Tanos et al., 1996; Muniz et al.,2010). Optimization of RAPD-PCR with primers was done to specify reaction pa-rameters settings (primer annealing temperature, number of amplification cycles, MgCl2 concentra-tion, primers and template DNA quantity).
All the arbitrary primers (as well as ITS-sequencing primers) were manufactured in NCB (National Cen-ter for Biotechnology, RK). Initial reaction condi-tions (reaction volume, DNA-matrix, primers and MgCl2 concentrations, annealing temperature, number and timing of amplification cycles) were used according to (Tanos et al., 1996; Muniz et al., 2010). Optimum values of these parameters were determined individually, depending on the particu-lar object of study.
PCR was performed using a thermal iCycler IQ5 (BioRad, USA). All PCR reactions of each isolate ge-nomic DNA were carried out in a total volume of 25μl containing 30-40 ng DNA template, 200 mM dNTPs, 7-15 pmol of primer, 2.5μl of 10× PCR buff-er and 1 Unit of Taq DNA polymerase (ThermoScientific, (EU) Lithuania). PCR amplifica-tion conditions were: initial denaturation at 94°C for 5 min followed by 35 cycles each of denatura-tion for 30 sec at 94°C, annealing for 30 sec at ap-propriate temperature according to primer used and extension for 1 min at 72°C and finally 5 min extension at 72°C. The reactions were repeated twice to ensure reproducibility. The amplification products were visualized on 1.5% agarose gel, de-tected by staining with ethidium bromide and ana-lyzed using the gel documentation system (BioRad, USA).
Electrophoregrams processing, cluster analysis and dendrograms were performed using GelQuest 3.1.7 and ClusterVis 1.8.1, respectively (SequentiX, Ger-many).
3. Results
3.1. Hypomycetes isolation
Soil suspension of the sample was diluted and plat-ed onto solid media. 4-15-days exposure at 28°C revealed different morphology colonies. Filamen-tous ones were selected and subcultured on Sabouraud and Czapek agar. Subsequent several passages of these isolates onto the same media confirmed purity of isolated cultures. Finally seven isolates were chosen as the objects for further study.
Microscopic analysis of isolated fungi showed that the majority of isolates belonged to the genus Peni-cillium spp. (Fig.1, A). Furthermore, other micromy-cetes were found among the isolates (Fig.1, B).
Primer Sequence (5'-3') Traits
(GTG)5 GTGGTGGTGGTGGTG microsatellite
AP3 TCACGATGCA random primer
T3B AGGTCGCGGGTTCGAATCC intergenic spacer region
Table 1: DNA fingerprinting primers used in the current study.
Figure 1: Microscopic analysis of different fungi isolates conid-iophores isolated from soil samples of Akmola region and belonging to the genus Penicillium spp. (A) and Bionectria spp. (B).
A B

20
Li et al., South Asian J Exp Biol; 4 (1): 17-23; 2014
3.2. ITS-region genotyping
The BLAST analysis of F1, F3 and F5-F7 ITS and 5.8S rDNA gene nucleotide sequences indicated the highest identities with a Penicillium roseopur-pureum, Penicillium rectrictum, Penicillium chryso-genum and Bionectria ochroleuca strains (99-100%) while sequences determined for two fungal isolates (F2, F4) was not species-specific, then their species level identification requires alternative ap-proachs application (Table 2).
3.3. Hyphomycetes HAT-RAPD profiling.
Series of experiments were performed in each of which one of the PCR parameters varied. Sets per-formed consequentially, locking in each case the test factor optimum value. Ultimately, conditions were chosen to ensure maximum yield of amplifica-tion products. Table 3 presents data on key param-eters of PCR using arbitrary primers.
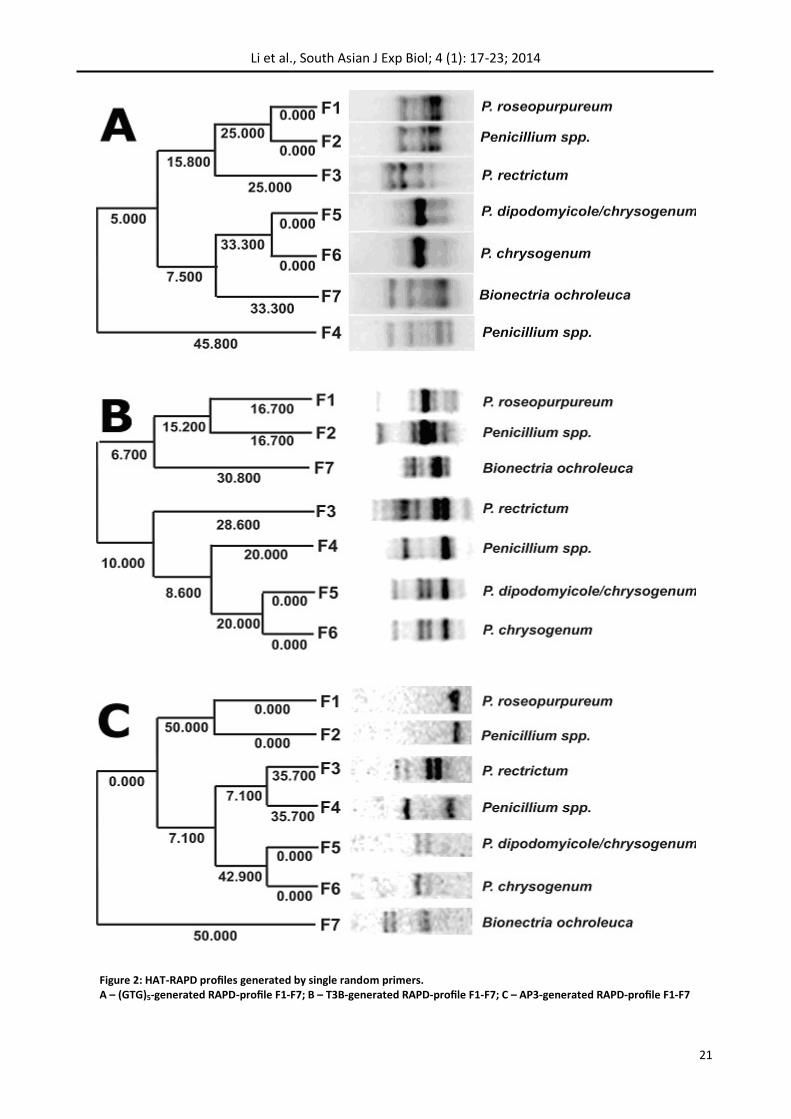
Each of three primers generated different profiles of amplification products, varying in location and intensity of the bands (Fig. 2).
In various isolates from 2 to 9 easily distinguishable and stably reproducible PCR products was formed ranging in size from 0.2 to 1.5 kb. The number and size of amplicons were characteristic for each spe-cies.
Reproducibility of RAPD-profiling is an essential problem to mark strains and species (Ramachandran et al., 2013). In the presented ex-perimental work twice reproducibility was consid-ered sufficient.
RAPD-profiles’ electrophoregrams were converted into binary matrix using the GelQuest 3.1.7 soft-ware. Formed numerical data were subjected to cluster analysis using unweighted pair-group meth-od with arithmetic averages (UPGMA). Resulting dendrograms generally reflect the regularities found on a visual level, and in addition, the evolu-tionary distance between isolates (Figure 2).
4. Discussion and Conclusions
Figure 2 illustrates the differences between patterns reflecting the polymorphism of the se-quences homologous to the used primers in the fungal genome location. The profiles obtained with the (GTG)5 and AP3 primers correspond to the iso-lates’ identification information obtained by se-quencing of ITS-region (Table 2) in spite of less dis-criminatory ability of RAPD compared with ITS-sequencing (Christianson et al., 2012).
Patterns F1 and F2 as well as F5 and F6 are very similar (but not absolutely identical) in pairs (Fig. 2 – A, C). Complete coincidence of positions of major and minor bands in 5 and 6 sample lines (Fig. 2 – A, B, C) suggests a close genetic relationship (if not identity) in F5 and F6, which was confirmed by pre-viously implemented ITS-genotyping – isolates be-long to the same species Penicillium chrysogenium. On the contrary, previously identified by ITS-genotyping as Penicillium roseopurpureum F1 iso-late demonstrates a high degree of identity with F2 (Penicillium spp.) basing on a complete coincidence of the amplicon profiles generated by the two pri-mers – (GTG)5 and AC3 (Fig. 2 – A, C) though ITS-genotyping data are not identical. It seems to be the two methods are not always precise, so it is advisable to use more than one method for the species level identification (Pál et al., 2012).
The profiles of isolates F1 and F2 generated by pri-mer AP3 represented by a single amplicon (Fig. 2. – C) are of particular interest. It seems to be justified to treat the latter as a basis for the development of species-specific identification system for Penicillium roseopurpureum as described previously (Polashock et al., 2002) because unique band (~400 bp) (Fig. 2. – C) does not correspond any band into the rest of lines and so can not be genus- or order- specific.
Results for T3B profiles, along with a high percent-age of coincidence in pairs F1-F2 and F5-F6 show a certain polymorphism in fragments length and their composition (Fig. 2 – B). This makes it possible to
Isolates Systematic name by ITS-genotyping
F1 Penicillium roseopurpureum
F2 Penicillium spp.
F3 Penicillium rectrictum
F4 Penicillium spp.
F5 Penicillium chrysogenum
F6 Penicillium chrysogenum
F7 Bionectria ochroleuca
Table 2: Hyphomycetes isolated from the soil samples of Akmola region of Kazakhstan.
Primer Annealing t° pmol/25 µl
AP3 40 15
T3B 52 15
(GTG)5 60 7
Table 3: Optimized settings for hyphomycetes HAT-RAPD.
21
Li et al., South Asian J Exp Biol; 4 (1): 17-23; 2014
Figure 2: HAT-RAPD profiles generated by single random primers. A – (GTG)5-generated RAPD-profile F1-F7; B – Т3В-generated RAPD-profile F1-F7; C – АР3-generated RAPD-profile F1-F7

22
Li et al., South Asian J Exp Biol; 4 (1): 17-23; 2014
analyze, in future, intra-specific differences in hy-phomycetes. In this case, T3B could be a primer of choice, though there are some data concerning successful species-specific identification by use of T3B-primer in Ascomycetes (de Oliveira et al., 2012).
Despite of limited isolates sample size cluster anal-ysis distinguished them between two subgroups based around the identical pairs: F1-F2 and F5-F6, regardless of the specific primer generated the DNA profiles. That is consistent with the results of genotyping by ITS-region (Table 2).
The specific nature of the manifestation of the temperature discontinuity at the annealing step during HAT-RAPD is of particular note.
In accordance with (Anuntalabhochai et al., 2000; Wongsavad et al., 2009) successful annealing tem-perature RAPD-PCR using unique primers differ from calculated values at 10-12°C. And that is what allows you to get stable reproducible profiles.
Number of produced bands is reduced due to the elimination of non-specific amplicons initiated by the primer annealing in the matrix sites with partial homology and therefore makes problematic the reproducibility of the profiles obtained.
We do not consider worthy the data showing few amount of amplicons in low annealing temperature RAPD-PCR (Ranga et al., 2011) because of bulky smear presenting in the electrophoretic patterns.
Thus, the suitability of HAT-RAPD as a means for a stable marking on the genomic level was shown for a group of hyphomycetes isolated from the Akmola region soils. It is suitable also in establishing phy-logeny and quick species identification. The latter is possible via sufficient databases use. Application of single primers HAT-RAPD discriminate interspecific and intraspecific differences in the soil hyphomy-cetes DNA-profiles.
Acknowledgements
The authors would like to extend their sincere ap-preciation to the Science committee of the Ministry of Science RK for its funding of this research through the Research Project no. 0112RK00457.
References
Anuntalabhochai S, Chiangda J, Chandet R, Apawat P (2000) Ge-netic diversity within lychee (Litchi chinensis Soonn.) based on
RAPD analysis. International Symposium on Tropical and Subtropi-cal Fruit. 26th Nov–1st Dec, Cairhs, Australia, p. 45
Bischoff JF, Rehner SA, Humber RA (2006) Metarhizium frigidum sp. nov.: a cryptic species of M. anisopliae and a member of the M. flavoviride complex. Mycologia 98: 737–745
Cambereri EB, Kinsey JA (1993) An ultra-fast method of DNA ex-traction from Neurospora. Fungal Geneics Newsletters 40: 22–23
Christianson S, Wolfe J, Soualhine H, Sharma MK (2012) Compari-son of repetitive-sequence-based polymerase chain reaction with random amplified polymorphic DNA analysis for rapid geno-typing of nontuberculosis mycobacteria. Canadian Journal of Mi-crobiology 58(8): 953-964
De Oliveira MM, Sampaio P, Almeida-Paes R, Pais C, Gutierrez-Galhardo MC, Zancope-Oliveira RM.J (2012) Rapid identification of Sporothrix species by T3B fingerprinting. Journal of Clinical Micro-biology 50(6): 2159-2162
Fegan M, Manners JM, Maclean DJ, Irwin JA, Samuels KD, Holdom DG, Li DP (1993) Random amplified polymorphic DNA markers reveal a high degree of genetic diversity in the entomopathogenic fungus Metarhizium artisopliae var. anisopliae. Journal of General Microbiology 139: 2075-208
Gilman JC (1998) A Manual Of Soil Fungi. Daya Publishing House, Delhi
Inglis GD, Duke GM, Goettel MS, Kabaluk JTJ (2008) Genetic diver-sity of Metarhizium anisopliae var. anisopliae in southwest-ern British Columbia. Journal of Invertebrate Pathology 98(1): 101-113
Khan S, Guo L, Maimaiti Y, Mijit M, Qiu D (2012) Entomopathogen-ic Fungi as Microbial Biocontrol Agent. Molecular Plant Breeding 3(7): 63-79
Möller EM, Bahnweg G, Sandermann H, Geiger HH (1992) A simple and efficient protocol for the isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tis-sues. Nucleic Acids Research 18: 1657
Muniz MM, Tavares PMS, Meyer W, Nosanchuk JD, Zancope-Oliveira RM (2010) Comparison of different DNA-based methods for molecular typing of Histoplasma capsulatum. Applied and Environmental Microbiology 76(13): 4438–4447
Pál K, Szén O, Kiss A, Naár Z (2012) Comparison and evaluation of molecular methods used for identification and discrimination of lactic acid bacteria. Journal of the Science of Food and Agriculture 92(9): 1931-1936
Parsal S, Ortiz V, Vega FE (2013) Establishing Fungal Entomopatho-gens as Endophytes: Towards Endophytic Biological Control. Jour-nal of Visualized Experiments http://www.jove.com/video/50360 accessed on 20 February 2014
Pava-Ripoll M, Angelini C, Fang W, Wang S, Posada FJ, St. Leger R (2011) The rhizosphere-competent entomopathogen Metarhizium anisopliae expresses a specific subset of genes in plant root exu-date. Microbiology (157): 47–55
Pfeifer TA, Khachatourians GG (1993) Isolation of DNA from ento-mopathogenic fungi grown in liquid cultures. Journal of Inverte-brate Pathology 61: 113-116
Polashock JJ, Vorsa N (2002) Development of SCAR markers for DNA fingerprinting and germplasm analysis of american cranberry. Journal of American Society of Horticultures Sciences 127(4): 677-684

23
Li et al., South Asian J Exp Biol; 4 (1): 17-23; 2014
Ramachandran P, Lacher DW, Pfeiler EA, Elkins CA (2013) Develop-ment of a tiered multilocus sequence typing scheme for members of the Lactobacillus acidophilus complex. Applied and Environmen-tal Microbiology 79(23): 7220-7228
Ranga V, Saini GK (2011) Biochemical and molecular characteriza-tion of wild-type and fused protoplasts of Beauveria bassiana and Metarhizium anisopliae. Folia Microbiologia 56: 289–295
Shahid AA, Rao AQ, Bakhsh A, Husnain T (2012) Enthomopatho-genic fungi as biological controllers: new insights into their viru-lence and pathogenicty. Archives of Biological Sciences 64(1): 21-42
Thanos M, Schonian G, Meyer W, Schweinoch C, Graser Y, Mitchell
TG,. Presber W, Tietz H-J (1996) Rapid identification of Candida species by DNA fingerprinting with PCR. Journal of Clinical Micro-biology 34(3): 615–621
Van Belkum A, Struelens M, De Visser A, Verbrugh H, Tibayrenc M (2001) role of genomic typing in taxonomy, evolutionary genetics, and microbial epidemiology. Clinical Microbiology Reviews 14(3): 547-60
Wongsawad C, Wongsawad P, Chai JY, Anuntalabhochai S (2009) Haplorchis taichui, Witenberg, 1930: Development of a HAT-RAPD marker for the detection of minute intestinal fluke infection. Experimental Parasitology 123(2): 158-161




















