
Impaired Secretory Function of the Prostate in Men With Oligo‐Asthenozoospermia
Upload
independentCategory
view
3download
0
RESEARCH Open Access
Cloning of the Zygosaccharomyces bailii GAS1homologue and effect of cell wall engineering onprotein secretory phenotypeSimone Passolunghi1dagger Luca Riboldi12dagger Laura Dato1 Danilo Porro1 Paola Branduardi1
Abstract
Background Zygosaccharomyces bailii is a diploid budding yeast still poorly characterized but widely recognisedas tolerant to several stresses most of which related to industrial processes of production Because of that it wouldbe very interesting to develop its ability as a cell factory Gas1p is a b-13-glucanosyltransglycosylase which plays animportant role in cell wall construction and in determining its permeability Cell wall defective mutants ofSaccharomyces cerevisiae and Pichia pastoris deleted in the GAS1 gene were reported as super-secretive The aimof this study was the cloning and deletion of the GAS1 homologue of Z bailii and the evaluation of its deletion onrecombinant protein secretion
Results The GAS1 homologue of Z bailii was cloned by PCR and when expressed in a S cerevisiae GAS1 nullmutant was able to restore the parental phenotype The respective Z bailii Δgas1 deleted strain was obtained bytargeted deletion of both alleles of the ZbGAS1 gene with deletion cassettes having flanking regions of ~400 bpThe morphological and physiological characterization of the Z bailii null mutant resulted very similar to that of thecorresponding S cerevisiae mutant As for S cerevisiae in the Z bailii Δgas1 the total amount of protein released inthe medium was significantly higher Moreover three different heterologous proteins were expressed and secretedin said mutant The amount of enzymatic activity found in the medium was almost doubled in the case of theCandida rugosa lipase CRL1 and of the Yarrowia lipolytica protease XPR2 while for human IL-1b secretiondisruption had no relevant effect
Conclusions The data presented confirm that the engineering of the cell wall is an effective way to improveprotein secretion in yeast They also confirmed that Z bailii is an interesting candidate despite the knowledge ofits genome and the tools for its manipulation still need to be improved However as already widely reported inliterature our data confirmed that an ldquoalways workingrdquo solution to the problems related to recombinant proteinproduction can be hardly if never found instead manipulations have to be finely tuned for each specific productandor combination of host cell and product
BackgroundThe ldquonon-conventionalrdquo yeast Zygosaccharomyces bailiihas been proposed as a possible new player in biotechprocesses arena [1] Its tolerance to high sugar concen-trations low pH environments caused by organic acidsrelatively high temperatures [2] could allow its growthunder restrictive conditions These features can be veryuseful for industrial fermentations because they can
simplify the process improving the economical value Inaddition the high specific growth rate of Z bailii andits high biomass yield render this yeast particularlyattractive for those applications were the biomass itselfor a specific protein or metabolite is the desired pro-duct In respect to recombinant protein production ithas been demonstrated that production and secretionlevels of the recombinant human interleuchin-1b (rhIL-1b) and of fungal glucoamylases in Z bailii are higherthan the levels obtained with S cerevisiae by the use ofthe same centromeric expression vector [3]
Correspondence daniloporrounimibitdagger Contributed equally1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
copy 2010 Passolunghi et al licensee BioMed Central Ltd This is an Open Access article distributed under the terms of the CreativeCommons Attribution License (httpcreativecommonsorglicensesby20) which permits unrestricted use distribution andreproduction in any medium provided the original work is properly cited
From an industrial point of view production andsecretion capabilities of the host systems currently inuse are still insufficient In particular secretion of theheterologous protein often represents one of the mainbottlenecks Different strategies can be utilized in thisrespect and among them are improvement of the het-erologous gene expression levels signal sequence opti-mization co-expression of chaperones and foldasesintroduction of mutations which improve secretion cap-abilities and reduction of the proteolytic activity insecretion vesicles [4]Since the yeast cell wall constitutes a physical barrier
to large molecules transit it represents one of the mainlimits for heterologous protein secretion Thus muta-tions which directly or indirectly alter the cell wall orga-nization can lead to better secretion capabilities of ayeast host [5]Mutations in genes involved in the construction and
in the maintenance of the cell wall such as PMR1SEC14 ERD1 MNN9 and MNN10 have in some casesbeen demonstrated to lead to supersecretive mutants inS cerevisiae and other yeasts [5] The results evidence atight correlation between glycosylation processes andprotein secretion [6] It was hypothesized that this cor-relation is not directly linked to the heterologoussecreted proteins but rather to an altered structure ofthe glycosidic residues added to the cell wall (glycol)pro-teins [5] which in turn leads to an altered permeabilityof the cell wall itselfAmong the already mentioned target also the inacti-
vation of the GAS1 gene whose product is directlyinvolved in the synthesis of the cell wall led to a hyper-secretive phenotype in S cerevisiae [7] The ScGas1p isan extracellular glycoprotein anchored to the cell mem-brane through a GPI tail which has a b-13-glucanosyl-transferase activity This enzyme plays a central role inthe formation of cell wall glucans it catalyzes the cross-linking of the glucans resulting in the b-13-glucan elon-gation [8] The identification of Gas1p homologues inother yeast species and some pathogenic fungi led tothe definition of a new glycosyl-hydrolases familyRecently the hypothetical 3D structure of the catalyticdomain of the protein was built by homology modelling[9] The GAS1 gene disruption leads to peculiar mor-phological and physiological phenotypes due to analtered cell wall structure and composition In particu-lar it was reported an altered distribution of the percen-tages of b-13- and b-16-glucans the latter being higherthan in wild type strain [10] It was also reported anincrease in the chitin and mannoproteins levels togetherwith altered linkages between the different components[11] As a direct or indirect result of these cell wallstructure modifications the gas1 mutant shows highersecretion levels if compared to the wild type both for
total and for the heterologous recombinant proteins asthe human insuline-like growth factor (rhIGF1) [7]More recently the Pichia pastoris GAS1 homologousgene was cloned and then deleted also resulting in ayeast mutant showing a supersecretive phenotype forsome heterologous proteins like a yeast lipase but notfor other like the human trypsinogen [12]Here we demonstrate that the deletion of the Z bailii
GAS1 homologue leads to an improved heterologousprotein secretion in this new promising host For reach-ing this goal the ZbGAS1 gene was firstly cloned andthen deleted thanks to the setting up of a protocol forgene deletion in this non-conventional diploid yeast Wedescribe here the morphological and physiological char-acterization of the ZbΔgas1 which in analogy to theScΔgas1 and PpΔgas1 mutants resulted to have a super-secretive phenotype
ResultsPCR cloning for Z bailii GAS1 homologueBecause of the high phylogenetic correlation between Zbailii and S cerevisiae [13] and because of the high per-centage of identity among the few known Z bailii andthe corresponding S cerevisiae gene sequences our firstattempt was to clone the gene homologous to GAS1 inZ bailii by PCR amplification using primers designedon the 3rsquo and 5rsquo ends of the ScGAS1 coding sequencebut with no successIt was therefore necessary to design internal primers
choosing the most conserved regions of the gene basedon multiple sequence alignments The Candida glabrataand the Schyzosaccharomyces pombe homologous genes(overall sequence identity to ScGAS1 71 and 62respectively) in addition to the S cerevisiae one were cho-sen for comparison Two amplification primers weredesigned on ScGAS1 in (sub)regions sharing nearly 100sequence identity with the two homologues at about 200bp and 300 bp from the ATG and the STOP codonrespectively A 1137 bp fragment was amplified using Zbailii genomic DNA as a template sharing 72 sequenceidentity with the corresponding internal region of ScGAS1Translation into the putative aminoacidic sequence led tothe identification of a region with 74 identity to theScGas1p protein fragment spanning from the catalyticdomain to the beginning of the Cys Box (WU-Blast2 align-ment results Figure 1) This confirms data reported in lit-erature which indicate the catalytic domain and the Serineand Cysteine Boxes as the most conserved regions of theGAS1 homologues in different yeast species [8]The 5rsquo and 3rsquo regions of the ZbGAS1 homologue were
subsequently amplified from a Genome Walker libraryconstructed from the Z bailii genomic DNA using theUniversal GenomeWalkerreg Kit (Clontech) Gene-specificprimers used were ZbGAS1intREV and ZbGAS1intFW
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 2 of 11
for the 5rsquo and 3rsquo ends respectively (see Methods) Thecomplete ORF of ZbGAS1 was reconstructed in thisway and additional 262 bp upstream to the ATG and177 bp after the STOP codon were also sequenced(GeneBank Accession Ndeg GU136594)Based on the new sequence it was possible to design
specific primers for the ZbGAS1 gene (see Methods)and a DNA fragment of the expected length was ampli-fied from the Z bailii genomic DNAThe complete ZbGAS1 ORF resulted to be 1710 bp
long (+ 30 bp vs ScGAS1) sharing a 67 sequence iden-tity with the S cerevisiae homologue (additional file 1)This sequence encodes for a putative aminoacidicsequence of 569 aa (+10 aminoacids vs ScGas1p) having68 identity with the ScGas1p protein (Figure 1)Sequence alignments were also performed between thenewly isolated Z bailii ORF and sequences of otherhomologous GAS genes in S cerevisiae (GAS1-5) thelower identity scores obtained for sequences other thanScGAS1 (not shown) further indicated that we clonedthe GAS1 homologue of Z bailiiHence considering the sequence similarities between
the ScGAS1 gene and their homologues in other yeaststhe GAS1 gene resulted to be highly conserved also inZ bailii The overall deduced protein structure seems tobe also conservedThe ZbGAS1 gene is one of the few Z bailii gene
sequences isolated to date hence representing a furtherlittle step towards the knowledge of this still poorlycharacterized yeast
ZbGAS1 gene disruptionTwo different ZbGAS1 deletion cassettes were con-structed based on the kanR and hphR markers Differ-ently from the case of the ZbYME2 gene deletion [14]where flanking regions of about 60 to 80 bp weredescribed to be sufficient for obtaining homologousrecombination long homologous flanking regions werenecessary to obtain gene replacement In particular weconstructed both the deletion cassettes represented inFigure 2a with flanking regions of about 400 bp whichresulted to be sufficient to disrupt the first and as wellthe second allelic copy of ZbGAS1 despite with quitelow efficiency (about only 10 of the transformantsresulted to effectively have both the deleted alleles con-firming what previously described [14]) The two cas-settes (see Methods) were therefore used to transformZ bailii cells resulting in the gas1 null mutant Theinsertion of both deletion cassettes in the homologousloci was confirmed by PCR (Figure 2b) We applied thesame strategy for the deletion of the first essential Zbailii gene leading to the first auxotrophic mutant ofthis yeast and in this case we had to further extend thelength of the homologous region for specific recombina-tion and despite that the percentage of positive eventswas even lower (manuscript in preparation) Theseresults could lead to hypothesize that for targeted genedeletion in Z bailii the length of the flanking regionscould be crucial according to the specific role that aspecific protein plays thus explaining results obtainedby other researchers [14]
Figure 1 Clustalw_aapdf Putative aminoacidic sequence of the ZbGas1p protein and alignment with the homologue ScGas1p Grey scalecoloring represents aminoacid similarity based on Blosum 62 scores Alignment was performed with ClustalW software httpwwwebiacukclustalw and graphical editing was made using Jalview software [31]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 3 of 11
Morphological and physiological characterization of thenull mutantThe GAS1 gene inactivation leads to different peculiarphenotypes in S cerevisiae that were widely character-ized [10] We therefore performed a morphological andphysiological characterization of the obtained Z bailiiΔgas1 mutant in analogy to what reported in literaturefor the model yeast First some morphological traits ofthe Sc Δgas1 mutant are visible both by light and byfluorescence microscopy after Calcofluor White stainingIt was reported for S cerevisiae (both haploid anddiploid) mutants a rounded shape and higher cell dia-meter if compared to wild type cells especially in thestationary phase [10] We performed the analysis of theZbΔgas1 and wild type strains in parallel to the S cere-visiae BY4741 Δgas1 and wt strains Microscopy imagesare shown in Figure 3 It is possible to see the typicalrounded phenotype also for the ZbΔgas1 (panels C andG) Moreover like in S cerevisiae intracellular granulesand altered vacuoles are also visible (indicated byarrows panels C and G) The altered shape is alsoreflected by cellular volumes being higher in themutants as assessed by coulter counter measurementsin cell populations growing in minimal or in rich med-ium like for the model yeast differences are more pro-nounced in stationary phase were the mutant cellvolume is 16 fold larger in respect to the wild typecells (data not shown) Another feature described forthe ScΔgas1 mutant is the appearance of large cellularaggregates still permanent after repeated sonicationcycles [10] This feature is also conserved in the Z bai-lii mutant as it is visible from the microscopy imagesof sonicated cells (Figure 3 panels C D G and H) Themorphological alterations described are likely a conse-quence of the altered structure and composition of themutant cell wall it was reported for S cerevisiae a
lowered glucans content together with a markedincrease in the chitin and mannoproteins content [15]Moreover an altered distribution of the cell wall com-ponents in the mother and daughter cell have beenreported [10] Our analyses confirmed these alterationsalso for the Z bailii mutant overall Calcofluor stainingin fact was more intense in Δgas1 than in wild typestrain (Figure 3 panels D and H) indicating a higherchitin content in the mutant The chitin seems to bedelocalized throughout the entire cell wall in themutant while in the wild type it is more concentratedat the budding ring and at the bud scars Moreover inthe mutant the cellular buds are also Calcofluor-stainedin contrast to the wild type It was proposed that theincrease in chitin levels and cross-links with b-16-glu-cosyl mannoproteins may be the effect of the activationof a compensatory mechanism in mutants characterizedby defects in the cell wall architecture like the Δgas1mutants The aim of this response would be to preventexcessive loss of mannoproteins as a consequence ofthe inability to cross-link the b-16-glucosyl mannopro-teins and the b-13-glucans a process in which Gas1pplays a central role [15] It was also hypothesized thatthe loss of Gas1p may cause a weakening of the cellwall of the emerging bud The yeast cell could inresponse to this weakening target the accumulation ofchitin at the bud surface in order to protect it fromlyses [16] Alteration of the processes directly or indir-ectly linked to budding leads to the appearance of arelative high fraction in the cell population of multi-budded cells (the so-called ldquoMickey Mouserdquo phenotype)(figure 3) A similar phenotype was also visible inZbΔgas1 mutants (Figure 3)The altered cell wall structure renders the deleted
bakersrsquo yeast strain more sensitive to sodium dodecylsulphate (SDS) and to Calcofluor White (CFW) as
Figure 2 Deletion of ZbGAS1 (a) Schematic representation of the ZbGAS1 deletion cassettes Restriction enzymes used to obtain the constructsand PCR primers used to verify recombination are indicated light grey bars represents ZbGAS1 sequence (b) Control PCR performed to verifythe deletion of both the allelic copies of ZbGAS1 Two independent mutant clones (A and B) were tested In both clones it is possible to see thepresence of bands corresponding to the two deletion cassettes and the absence of the band corresponding to the wild type gene copy
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 4 of 11
reported in literature for the W303 background [17]Here we confirm by spot assays the sensitivity to bothagents of the ScΔgas1 mutant background BY4741 (Fig-ure 3 upper right panels) and as well of the ZbΔgas1mutant (Figure 3 lower right panels)Finally to verify the functionality of the GAS1 homo-
logue of Z bailii in S cerevisiae and the similarity of itsbiological role the ZbGAS1 gene was expressed into theGAS1-deficient S cerevisiae strain resulting in wild typephenotype restoration either in respect to the morpholo-gical traits as well as for the chitin distribution (data not
shown) Moreover to verify the functionality of the Scerevisiae GAS1 in Z bailii the ScGAS1 gene wasexpressed into the GAS1-deficient Z bailii strain alsoin this case restoring the native phenotype (data notshown)Overall morphological and physiological data strongly
indicate that the ZbGAS1 gene may have a similar rolewith respect to the ScGAS1 homologous in the assess-ment of the cell wall structure and hence their inactiva-tion might have similar effects in the two yeasts inrespect to secretion abilities
Figure 3 Morphology of Z bailii and S cerevisiae wild type and gas1 null mutant strains and their sensitivity to Calcofluor White andSDS Cellular morphology From A to H dicroic and fluorescence microscope images of Z bailii and S cerevisiae wild type and gas1 null mutantstrains Cells were grown in minimal medium and harvested in stationary phase of growth Arrows indicate granules and enlarged vacuoles inmutant cells (panels C and G) Calcofluor White and SDS sensitivity of S cerevisiae (BY4741) and Z bailii (Zygo) gas1 null mutants 4 μl of culturesof the different strains (BY4741 BY4741 Δgas1 Zygo and Zygo Δgas1) were spotted starting with OD (660 nm) = 1 and then in 3 serial solution110 (234) on rich solid medium (YPD) and on the same medium added with 125 μg ml-1 of CFW or 50 μg ml-1 SDS Pictures were capturedafter 5 days of incubation at 30degC
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 5 of 11
Supersecretive phenotype of Δgas1 strainsAs already mentioned (see Introduction) the mostinteresting phenotype of Δgas1 mutants reported in lit-erature is the supersecretive property Besides theincreased secretion of heterologous proteins whichappears to be dependent on the individual protein [12]the levels of total secreted proteins in late-stationaryphase cultures of S cerevisiae mutants were two foldhigher with respect to the wild-type [7] We thereforestarted our evaluation of secretive properties of the Zbailii mutant in the perspective of a possible applica-tion in industrial processes by determining the amountof total proteins released in the growth medium duringbatch cultivation with respect to the wild type strain Zbailii and S cerevisiae wild type and Δgas1 respectivestrains were grown in shake flasks in minimal YNB-based medium containing 5 glucose Cells were har-vested at different times of cultivation and the fermen-tation broths were assayed At least triplicateexperiments were performed on three independenttransformants and Figure 4 shows the mean dataobtained reported as mg of total proteins secreted perlitre of the culture broth normalised for OD valuesThe values refers to 72 h of cultivation but it was pos-sible to observe a trend leading to this result starting atthe end of exponential phase and becoming more pro-nounced in stationary phase of growth
The data clearly underline the overall higher secretivecapacity of Z bailii if compared to S cerevisiae inaccordance to heterologous proteins secretion dataalready reported [3] Moreover they indicate a signifi-cant improvement in the secretive capability of the Zbailii mutant being nearly 80 more than the wild typein terms of total endogenous protein similarly to the Scerevisiae mutantOverexpression of CRL lipase XPR2 protease and hIL-1bin Z bailii Δgas1 strainsWith the intention to evaluate the potential effects ofcell wall engineering and to test the secretive perfor-mance in the presence of an heterologous protein threemodel proteins were tested a fungal lipase from Can-dida rugosa (CRL1) a fungal protease from Yarrowialipolytica (XPR2) and the human interleukin 1b (hIL-1b)Four clones of Z bailii Δgas1 [pZLN022XPR2] and
four clones of the parental strain Z bailii[pZLN022XPR2] together with control clones harbour-ing the respective empty plasmid were pre-cultivated for16 hours in shake flasks using YPD medium and thentransferred in shake flasks in minimal YNB-based med-ium containing 5 glucose The amount of secretedrecombinant XPR2 in culture supernatants was tested interms of enzymatic activity after 72 96 120 and 148 hwith the Azocoll method (see methods for detail) The
Figure 4 Total proteins released in the culture medium Total proteins released in the culture medium of batch cultures of the indicatedwild type and Δgas1 strains of S cerevisiae and Z bailii Yeasts were grown in shake flasks in minimal YNB-based medium with 5 glucose Cellswere inoculated at 01 OD and harvested after 72 hours of cultivation Amount of released proteins (in mg l-1 of culture broth) were normalizedfor OD values
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 6 of 11
ZbΔgas1 transformants showed a relevant increase inspecific activity levels since 72 h (+ 25 at 72 h morethan two-fold at 148 h see Figure 5a) in respect to wildtype transformantsSimilarly three clones of Z bailii Δgas1
[pZLN022LIP1] and three clones of the parental strainZ bailii [pZLN022LIP1] together with control clonesharbouring the respective empty plasmid were pre-culti-vated for 16 hours in shake flasks using YPD mediumand then transferred in shake flasks in minimal YNB-based medium containing 5 glucose The amount ofsecreted recombinant LIP1 in culture supernatants wastested in terms of enzymatic activity after 24 48 72 and120 h using the hydrolysis of p-nitrophenylestersmethod (see methods for detail) Also for this secondheterologous product the Δgas1 transformants showed arelevant increase in specific activity levels since 24 h
(+35 at 24 h more than two-fold at 120 h see Figure5b) in respect to wild type transformantsFinally three clones of Z bailii Δgas1 [pZ5KlIL-1b]
and three clones of the parental strain Z bailii[pZ5KlIL-1b] together with control clones harbouringthe respective empty plasmid were tested for secretionlevel of the human interleukin since the codingsequence was functionally linked to the signal peptide ofthe K1 killer toxin from K lactis The independenttransformants were pre-cultivated and then cultured inshake flasks containing rich medium (YPD 5 wv glu-cose) Samples were collected at the indicated times (seeFigure 5c) cells were harvested and both the superna-tants and the cell protein extracts of said samples wereloaded on a polyacrylamide gel The data obtained byWestern blot (see Figure 5c) performed on the superna-tant showed a well represented band corresponding to
Figure 5 Secretion levels of yeast protease lipase and rhIL-1b in Z bailii and ZbΔgas1 (a) Secretion levels of protease in Z bailii andZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutant transformed with controlplasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with XPR2 gene (b) Secretion levels oflipase in Z bailii and ZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutanttransformed with control plasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with LIP1 gene(c) Western blots for rhIL-1b of protein samples from supernatants and cell extracts of Z bailii and ZbΔgas1 transformants grown in YPD mediawith 5 glucose inoculated at OD660 = 01 From left to right (A) rhIL-1b (B) negative control Z bailii[pZ5] (C) (D) (E) (I) (L) (M) Z bailii[pZ5KlrhIL-1b] (F) (G) (H) (N) (O) (P) ZbΔgas1 [pZ5KlrhIL-1b] Time and sample types are indicated in the picture
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 7 of 11
the recombinant protein for both Z bailii and Z bailiiΔgas1 samples but differently from the case of recombi-nant fungal enzymes no relevant differences in favourof the deleted yeast are appreciable Remarkably the sig-nal corresponding to the protein retained intracellularlyis more intense in gas1 null mutant than the signalobtained from Z bailii parental strain indicating anincomplete secretion of this protein
DiscussionsCell wall engineering can improve the release of het-erologous proteins from yeasts [18] Generally speak-ing GAS1 which mediates the crosslinking of cell wallcomponents is directly involved in defining the perme-ability of the cell Cloning of the unknown Z bailiiGAS1 homologue by PCR was only possible startingfrom highly conserved internal regions identified uponalignment of the GAS1 homologues of three yeast spe-cies The 5rsquo and 3rsquo ends were subsequently amplifiedusing a Genome Walker library With this approachthe full-length Z bailii GAS1 homologue could beamplified and sequenced The cloned gene wasexpressed in the GAS1-deficient S cerevisiae strain andreversion of the phenotype proved that this homologueis not only structurally but also functionally similar tothe S cerevisiae gene Moreover also the ScGAS1 genecomplemented the Z bailii mutant Disruption ofGAS1 in Z bailii revealed its influence on the cell wallmorphology The GAS1 null mutants of Z bailiishowed the same phenotype reported for S cerevisiae[10] with enlarged cells often showing granular cyto-plasm an alterated chitin distribution two buds(Mickey Mouse-like appearance) and big vacuoles Thevacuole is known to be a dynamic organelle whosemorphology is highly responsive to different extracellu-lar and intracellular stimulistressors [19] In the GAS1null mutant it can be therefore speculated that thegeneration of big vacuoles is related to stressful condi-tions as already suggested by the altered chitin distri-bution being a possible defence mechanism againstlysis or related to a generalized response to sickness ofthe cells [10]Finally also the sensitivity to SDS and CFW
appeared very similar to what observed in the S cerevi-siae mutant A comparison of the different GAS1 nullphenotype mutants and the functionality of the Z bai-lii GAS1 homologue in S cerevisiae suggest a similarityof the cell wall structure of Z bailii compared with thecell wall structure of S cerevisiae Considering thesecretory pathway of a secreted protein it was specu-lated whether the protein becomes retarded in the cellwall after release from the plasma membrane The dis-ruption of GAS1 in Z bailii first of all confirmed ageneral phenomenon of protein release in respect to
the wild type strain as demonstrated by the higheraccumulation of total protein in the growing mediumof the deleted strain when compared to the parentalstrain Moreover such as compromised cell wall per-meability in the production strains for XPR2 and LIP1does lead to a clear improvement in the secretion ofthe respective proteins In the case of h IL-1b theenhancement of product secretion was not so evidentand difficult to interpret in respect to the higher accu-mulation of intracellular product It could be specu-lated that this can be related to the nature or to thesource of the protein but too few information areavailable at the moment to further comment in thisdirection What seems to be true is unfortunately whatwe very often experience for recombinant protein pro-duction which is the protein and protein-host depen-dence In fact also in P pastoris two out of threeheterologous proteins resulted more secreted in themutant as for Z bailii and when we tested the sameprotein in particular the IL-1b secretion in the S cere-visiae Δgas1 mutant (data not shown) we did not reg-ister any improvement
MethodsYeast strains media and transformationThe S cerevisiae strains used in this study were theauxotrophic haploid BY4741 (Mat a his3Δ1 leu2Δ0met5Δ0 ura3Δ0) and the relative null mutant in GAS1sequence called in this study BY4741 Δgas1 (Mat ahis3Δ1 leu2Δ0 met5Δ0 ura3Δ0 YMR307kanMX4)Both strains are commercially available from Euroscarfcollection httpwebuni-frankfurtdefb15mikroeuro-scarfcol_indexhtml The Z bailii wild type strain isATCC60483 Yeast cultures were grown in YP medium(10 g l-1 yeast extract 20 g l-1 peptone) with 2 or 5wv glucose (D) or fructose (F) as carbon sourceWhere needed NAT (80 mg l-1) or G418 (500 mg l-1
for S cerevisiae and 200 mg l-1 for Z bailii) or hygro-micine (100 mg l-1 for Z bailii) were added to themedia Alternatively yeast cultures were grown inminimal synthetic medium (067 w v-1 YNB Biolifewithout amino acids) with 2 or 5 w v-1 glucose ascarbon source When required supplements such ashistidine leucine methionine and uracil were added toa final concentration of 50 mg l-1 Media for plateswere solidified by addition of agar (Biolife Milan Italy)to 15 w v-1 Shake flask experiments were carriedout with 50 ml of medium in 250 ml shake flasks on ashaker at 30degC with 160 rpm For sensitivity tests inPetri dishes 125 μg ml-1 of CFW or 50 μg ml-1 SDSwere added to solid media Yeast transformation wasperformed using the Lithium AcetatessDNA methodas described [20] and slightly adapted for Z bailii asdescribed [3]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 8 of 11
Genes amplifications deletion cassettes and expressionplasmid constructionPCR amplifications were made from yeasts genomicDNA prepared as described in [21] Amplifications wereperformed by the Pwo DNA Polymerase (Roche)The amplification of the ZbGAS1 gene was as follows
the primers GAS1int_fw 5rsquo-AGT TGT TCC AGA GATATT CCA TAC CTC AA-3rsquo and GAS1int_rev 5rsquo-GCAGAA CCG CTG AAG CTA CAG T-3rsquo designed oninternal conserved region of the available sequencesfrom other yeasts were firstly used to amplify an internalregion of 1137 bp of the Z bailii gene with the follow-ing program 94degC 5 min (94degC 15 s 57degC 30 s 72degC 1min) times 25 72degC 7 min 4degC infin After that the designedprimers ZbGAS1int_fw 5rsquo-GGA CGA TAA GGT TGACGA AGA-3rsquo and ZbGAS1int_rev 5rsquo-TGC TGT GGTCCA AAG TTG TG-3rsquo were used for the amplification(program 7 cycles 94degC 2 min 72degC 3 min 32 cycles94degC 2 min 67degC 3 min 67degC 4 min 4degC infin) of theterminal ends of ZbGAS1 utilizing a Z bailii GenomeWalker library constructed using the Universal GenomeWalker kit (Clontech) following the manufacturersinstructions From the obtained and reconstructedsequence the final primers were established and used forthe ZbGAS1 1710 bp full length coding sequence ampli-fication ZbGAS1_fw 5rsquo-ACT AAT GTT ATT CCAGGC GTT TTC G-3rsquo and ZbGAS1real_rev 5rsquo-AAATCA AGC CAA AGC AAA TCC AGC A-3rsquo (program94degC 5 min (94degC 20 s 58degC 30 s 72degC 2 min) times 30 72degC 10 min 4degC infin) The DNA sequence of ScGAS1 wasPCR amplified with the following primers ScGAS1fw5rsquo-ACA ATG TTG TTT AAA TCC CTT TCA AAGTTA GCA A-3rsquo and ScGAS1rev 5rsquo-TTT TTA AACCAA AGC AAA ACC GAC ACC AG-3rsquo (program 94degC5 min (94degC 20 s 605degC 30 s 72degC 2 min) times 30 72degC10 min 4degC infin) and the DNA sequence of YlXPR2 wasPCR amplified with the following primers XPR2_fwd5rsquo-ACA ATG AAG CTC GCT ACC GCC TTT A-3rsquoand XPR2_rev 5rsquo-TGC CTA AAT GCC AAC ACCGTT GTA G-3rsquo (program 94degC 4 min (94degC 15 s 625degC 30 s 72degC 2 min) times 30 72degC 7 min 4degC infin) All theunique fragments obtained were sub-cloned in the vec-tor pSTblue-1 utilising the Perfectly Bluntreg Cloning Kit(Novagene) and sequenced The resulting respectiveplasmids utilised in this work were named as followspSTbZbGAS1int pSTbZbGAS1 pSTbScGAS1 andpSTbYlXPR2For the construction of the ZbGAS1 deletion cassettes
the KanR or the hphR cassette derived from plasmidpFA6-KanMX4 [22] and pAG26 [23] and conferringresistance to G418 and hygromycin respectively werebluntended excised and inserted in the pSTbZbGAS1intopened BstEII BglII bluntended and dephosphorylatedresulting in the plasmids pSTbZbGAS1ΔK and
pSTbZbGAS1ΔH The resistance cassettes resulted inboth cases counter-clockwise inserted The Z bailii wildtype cells were transformed with the respective deletioncassettes obtained by cutting the two described plasmidsin the AccI and SnaBI sites The effective deletion wasPCR checked by using the following couples of primersZbG1int2_fw 5rsquo- TGC AGA AGT TAC AGA CCAATG TTG T -3rsquo and and ZbG1int2_rev 5rsquo-AAG ATAGAG AAG TGC TCT TGG CA -3rsquo (see Figure 2) orZbG1int2_fw and Hph_fw 5rsquo- ATA TGA AAA AGCCTG AAC TCA CCG AC -3rsquo or Kan_fw 5rsquo- ATG GGTAAG GAA AAG ACT CAC GTT -3rsquo (not shown) inorder to verify the presence of the cassettes in thedesired locusFor complementing the yeasts GAS1 deletion pZ5
(-Nco)ScGAS1 and pYX022ZbGAS1 were constructedpZ5(-Nco)ScGAS1 was obtained from pZ5(-Nco) openedSacIEcoRV with the insertion of ScGAS1 frompSTbScGAS1 cut with SacISnaBI The pZ5(-Nco)derives from pZ3 [3] where at the KpnI site the KanR
cassette was substituted from the NATR cassetteobtained from vector pAG25 [23] by cutting PvuIISacIblunt (obtaining the pZ5 plasmid) finally this plasmidwas EcoRIBamHI cut and bluntended and reclosedpYX022ZbGAS1 was obtained with the insertion ofZbGAS1 from pSTbZbGAS1 into pYX022 (RampD Sys-tems Wiesbaden Germany) both EcoRI cutFor heterologous protein expression the plasmids
pZLN022XPR2 pZLN022LIP1 and pZ5KlhIL-1b wereconstructed The first two integrative plasmids derivesfrom pZLN022 (manuscript in preparation) obtainedfrom pYX022 opened DraIII bluntSpeI blunt (toexclude HIS3) with the insertion of ZbLEU2 cut frompSTbZbLEU2 with EcoRI blunt in KpnI blunt site theNATR cassette was inserted obtained from vectorpAG25 [23] by cutting PvuIISacIblunt ThepSTbZbLEU2 was obtained by sub-cloning ZbLEU2amplified by PCR from Z bailii genomic DNA (manu-script in preparation) in pSTBlue (Novagen) cut byEcoRV For pZLN022XPR2 construction the pZLN022was AatIINheI cut and the XPR2 from pSTbXPR2 wasinserted For pZLN022LIP1 construction the pZLN022was AatII bluntNheI blunt opened and the LIP1 wasinserted by BamHI bluntNsiI blunt excision from thepGAPsLIP [24] For rh interleukin the pZ5KlhIL-1b wasbuilt it was obtained from pZ5 opened with EcoRI-blunt with the insertion of hIL-1b in frame with Kl sig-nal sequence from vector pKSSPI3 [25] cut EcoRIbluntXbaI bluntDNA manipulations transformation and cultivation of
Escherichia coli (DH5aFrsquo (80dlacZΔM15 Δ (lacZYA-argF) U169 deo rec1 end1 sup44 l THI-1 gyrA96relA1) and Novablue Competent Cells (Novagene) wereperformed following standard protocols [26] All the
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 9 of 11
restriction and modification enzymes used were fromNew England Biolabs (Hitchin Herts UK) or fromRoche Diagnostics (Mannheim Germany)Calcofluor staining and fluorescence microscopyCells were harvested washed in PBS and resuspended in1 mg ml-1 Calcofluor White solution incubated 10-30min at room temperature washed again twice in PBSFluorescence microscope images were taken by a NikonEclipse 90i microscope using the Metamorph softwareversion 22 (Nikon) using emission filter UV-1AQuantification of total released proteinsCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were etha-nol-precipitated The amount of proteins was deter-mined by the dye-binding method of Bradford (QuickStart Bradford Dye Reagent - BIORAD) using BSA asstandard All the data were normalized by OD after ver-ification of the linear correlation between OD and dryweight as a measure of biomassQuantification of secreted proteaseCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were sepa-rated and stored at -20degC until analysis The amount ofprotease secreted in the medium was determined by theAzocoll assay (Azocoll - Calbiochem) using commercialprotease as positive control Azocoll assays were per-formed according to [27] with some modificationsReaction mixtures contained 15 mg ml-1 Azocoll 50mM TrisHCl (pH 76) 150 mM NaCl and 5 mMCaCl2 Proteolytic activities were measured by absor-bance changes at 490 nm (Shimadzu UV 1601) afterincubation for 18 h at 30degCQuantification of secreted lipaseCultures were harvested and centrifuged at maximalspeed then the supernatants were separated and storedat -20degC until analysis The amount of lipase secreted inthe medium was determined according to [28] withsome modifications Reaction mixtures contained (A)60 mg of p-nitrophenyl palmitate (Sigma Aldrich) in 20ml of isopropanol (Sigma Aldrich) (B) 1 g Triton X-100(Sigma Aldrich) and 02 g of Arabic gum (SigmaAldrich) in 200 ml TrisHCl 01 M (pH 75) To obtainthe mixture (C) add slowly 1 part of A solution to 9part of B solution under continuous agitation Add 9part of (C) solution to one part of the enzyme contain-ing solution (ie the supernatant) Lipase activities weremeasured by absorbance changes at 410 nm (ShimadzuUV 1601) and the amount of enzyme is proportional toOD according to the relation Um = 0667 times OD410 nm
(if OD410 nm lt 05)Quantification of interleukinHuman IL-1b were analysed by Western blot Indepen-dent transformants were cultured in shake flasks inminimal or rich medium and during the kinetics
samples were collected Cells were harvested (a culturevolume corresponding to 108 cells) by centrifugationOne volume of 2timesLaemmli buffer [29] was added to thesupernatants of said samples they were boiled for 3-5min and stored at -20degC until loading or loaded directlyon a polyacrylamide gel For the analysis of intracellularproduction crude extracts were prepared from 108 cellsby following the trichloroacetic acid protocol [30] andresuspending the final protein extract in 150 μl ofLaemmli buffer Samples were then loaded on standardpolyacrylamide gels (sodium dodecyl sulfate-polyacryla-mide gel electrophoresis final concentration of theseparating gel 15) after separation proteins wereblotted to nitrocellulose membranes and immunodeco-rated An anti-IL-1b rabbit polyclonal antibody (IL-1b(H-153) Santa Cruz Biotechnology USA cat no sc-7884 dilution 1200) was used for the interleukin immu-nodecoration An anti-rabbit IgG horseradish peroxi-dase-conjugated (Amersham Pharmacia Biotech UK catno NA934 dilution 110000) secondary antibody wasused For the interleukin detection as a positive controlthe human recombinant IL-1b from E coli (Roche catno 1 457 756) was always loaded in parallel on thepolyacrylamide gels The proteins were visualised usingthe ECL Western blotting system (Amersham Bios-ciences UK) according to the manufacturerrsquosinstructions
Additional file 1 Clustalw_ntd Nucleotidic sequence of the ZbGAS1gene and alignment with the homologue ScGAS1 (entry YMR307W)Grey coloring represent nucleotidic identity Alignment was performedwith ClustalW software httpwwwebiacukclustalw and graphicalediting was made using Jalview software [31]Click here for file[ httpwwwbiomedcentralcomcontentsupplementary1475-2859-9-7-S1PDF ]
AcknowledgementsSpecial thanks to Valeria Longo and Davide Zocco for technicalcontributions and to Stefania Brocca for providing the pGAPsLIP plasmidThis work was partially supported by FAR 2007 and 2008 (12-1-192) to DP
Author details1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy 2CPC Biotech Srl Naples Italy
Authorsrsquo contributionsPB and DP initiated and coordinated the project LD was responsible forZbGAS1 gene identification by PCR and its deletion LR analyzed the mutantphenotype with CFW and SDS LR and SP performed the batch cultivationanalyzed the secretory phenotype in null mutant and performed theexperiment with heterologous proteins All authors wrote the paper andapproved the final version of the manuscript
Competing interestsThe authors declare that they have no competing interests
Received 2 November 2009Accepted 26 January 2010 Published 26 January 2010
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 10 of 11
References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11

From an industrial point of view production andsecretion capabilities of the host systems currently inuse are still insufficient In particular secretion of theheterologous protein often represents one of the mainbottlenecks Different strategies can be utilized in thisrespect and among them are improvement of the het-erologous gene expression levels signal sequence opti-mization co-expression of chaperones and foldasesintroduction of mutations which improve secretion cap-abilities and reduction of the proteolytic activity insecretion vesicles [4]Since the yeast cell wall constitutes a physical barrier
to large molecules transit it represents one of the mainlimits for heterologous protein secretion Thus muta-tions which directly or indirectly alter the cell wall orga-nization can lead to better secretion capabilities of ayeast host [5]Mutations in genes involved in the construction and
in the maintenance of the cell wall such as PMR1SEC14 ERD1 MNN9 and MNN10 have in some casesbeen demonstrated to lead to supersecretive mutants inS cerevisiae and other yeasts [5] The results evidence atight correlation between glycosylation processes andprotein secretion [6] It was hypothesized that this cor-relation is not directly linked to the heterologoussecreted proteins but rather to an altered structure ofthe glycosidic residues added to the cell wall (glycol)pro-teins [5] which in turn leads to an altered permeabilityof the cell wall itselfAmong the already mentioned target also the inacti-
vation of the GAS1 gene whose product is directlyinvolved in the synthesis of the cell wall led to a hyper-secretive phenotype in S cerevisiae [7] The ScGas1p isan extracellular glycoprotein anchored to the cell mem-brane through a GPI tail which has a b-13-glucanosyl-transferase activity This enzyme plays a central role inthe formation of cell wall glucans it catalyzes the cross-linking of the glucans resulting in the b-13-glucan elon-gation [8] The identification of Gas1p homologues inother yeast species and some pathogenic fungi led tothe definition of a new glycosyl-hydrolases familyRecently the hypothetical 3D structure of the catalyticdomain of the protein was built by homology modelling[9] The GAS1 gene disruption leads to peculiar mor-phological and physiological phenotypes due to analtered cell wall structure and composition In particu-lar it was reported an altered distribution of the percen-tages of b-13- and b-16-glucans the latter being higherthan in wild type strain [10] It was also reported anincrease in the chitin and mannoproteins levels togetherwith altered linkages between the different components[11] As a direct or indirect result of these cell wallstructure modifications the gas1 mutant shows highersecretion levels if compared to the wild type both for
total and for the heterologous recombinant proteins asthe human insuline-like growth factor (rhIGF1) [7]More recently the Pichia pastoris GAS1 homologousgene was cloned and then deleted also resulting in ayeast mutant showing a supersecretive phenotype forsome heterologous proteins like a yeast lipase but notfor other like the human trypsinogen [12]Here we demonstrate that the deletion of the Z bailii
GAS1 homologue leads to an improved heterologousprotein secretion in this new promising host For reach-ing this goal the ZbGAS1 gene was firstly cloned andthen deleted thanks to the setting up of a protocol forgene deletion in this non-conventional diploid yeast Wedescribe here the morphological and physiological char-acterization of the ZbΔgas1 which in analogy to theScΔgas1 and PpΔgas1 mutants resulted to have a super-secretive phenotype
ResultsPCR cloning for Z bailii GAS1 homologueBecause of the high phylogenetic correlation between Zbailii and S cerevisiae [13] and because of the high per-centage of identity among the few known Z bailii andthe corresponding S cerevisiae gene sequences our firstattempt was to clone the gene homologous to GAS1 inZ bailii by PCR amplification using primers designedon the 3rsquo and 5rsquo ends of the ScGAS1 coding sequencebut with no successIt was therefore necessary to design internal primers
choosing the most conserved regions of the gene basedon multiple sequence alignments The Candida glabrataand the Schyzosaccharomyces pombe homologous genes(overall sequence identity to ScGAS1 71 and 62respectively) in addition to the S cerevisiae one were cho-sen for comparison Two amplification primers weredesigned on ScGAS1 in (sub)regions sharing nearly 100sequence identity with the two homologues at about 200bp and 300 bp from the ATG and the STOP codonrespectively A 1137 bp fragment was amplified using Zbailii genomic DNA as a template sharing 72 sequenceidentity with the corresponding internal region of ScGAS1Translation into the putative aminoacidic sequence led tothe identification of a region with 74 identity to theScGas1p protein fragment spanning from the catalyticdomain to the beginning of the Cys Box (WU-Blast2 align-ment results Figure 1) This confirms data reported in lit-erature which indicate the catalytic domain and the Serineand Cysteine Boxes as the most conserved regions of theGAS1 homologues in different yeast species [8]The 5rsquo and 3rsquo regions of the ZbGAS1 homologue were
subsequently amplified from a Genome Walker libraryconstructed from the Z bailii genomic DNA using theUniversal GenomeWalkerreg Kit (Clontech) Gene-specificprimers used were ZbGAS1intREV and ZbGAS1intFW
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 2 of 11
for the 5rsquo and 3rsquo ends respectively (see Methods) Thecomplete ORF of ZbGAS1 was reconstructed in thisway and additional 262 bp upstream to the ATG and177 bp after the STOP codon were also sequenced(GeneBank Accession Ndeg GU136594)Based on the new sequence it was possible to design
specific primers for the ZbGAS1 gene (see Methods)and a DNA fragment of the expected length was ampli-fied from the Z bailii genomic DNAThe complete ZbGAS1 ORF resulted to be 1710 bp
long (+ 30 bp vs ScGAS1) sharing a 67 sequence iden-tity with the S cerevisiae homologue (additional file 1)This sequence encodes for a putative aminoacidicsequence of 569 aa (+10 aminoacids vs ScGas1p) having68 identity with the ScGas1p protein (Figure 1)Sequence alignments were also performed between thenewly isolated Z bailii ORF and sequences of otherhomologous GAS genes in S cerevisiae (GAS1-5) thelower identity scores obtained for sequences other thanScGAS1 (not shown) further indicated that we clonedthe GAS1 homologue of Z bailiiHence considering the sequence similarities between
the ScGAS1 gene and their homologues in other yeaststhe GAS1 gene resulted to be highly conserved also inZ bailii The overall deduced protein structure seems tobe also conservedThe ZbGAS1 gene is one of the few Z bailii gene
sequences isolated to date hence representing a furtherlittle step towards the knowledge of this still poorlycharacterized yeast
ZbGAS1 gene disruptionTwo different ZbGAS1 deletion cassettes were con-structed based on the kanR and hphR markers Differ-ently from the case of the ZbYME2 gene deletion [14]where flanking regions of about 60 to 80 bp weredescribed to be sufficient for obtaining homologousrecombination long homologous flanking regions werenecessary to obtain gene replacement In particular weconstructed both the deletion cassettes represented inFigure 2a with flanking regions of about 400 bp whichresulted to be sufficient to disrupt the first and as wellthe second allelic copy of ZbGAS1 despite with quitelow efficiency (about only 10 of the transformantsresulted to effectively have both the deleted alleles con-firming what previously described [14]) The two cas-settes (see Methods) were therefore used to transformZ bailii cells resulting in the gas1 null mutant Theinsertion of both deletion cassettes in the homologousloci was confirmed by PCR (Figure 2b) We applied thesame strategy for the deletion of the first essential Zbailii gene leading to the first auxotrophic mutant ofthis yeast and in this case we had to further extend thelength of the homologous region for specific recombina-tion and despite that the percentage of positive eventswas even lower (manuscript in preparation) Theseresults could lead to hypothesize that for targeted genedeletion in Z bailii the length of the flanking regionscould be crucial according to the specific role that aspecific protein plays thus explaining results obtainedby other researchers [14]
Figure 1 Clustalw_aapdf Putative aminoacidic sequence of the ZbGas1p protein and alignment with the homologue ScGas1p Grey scalecoloring represents aminoacid similarity based on Blosum 62 scores Alignment was performed with ClustalW software httpwwwebiacukclustalw and graphical editing was made using Jalview software [31]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 3 of 11
Morphological and physiological characterization of thenull mutantThe GAS1 gene inactivation leads to different peculiarphenotypes in S cerevisiae that were widely character-ized [10] We therefore performed a morphological andphysiological characterization of the obtained Z bailiiΔgas1 mutant in analogy to what reported in literaturefor the model yeast First some morphological traits ofthe Sc Δgas1 mutant are visible both by light and byfluorescence microscopy after Calcofluor White stainingIt was reported for S cerevisiae (both haploid anddiploid) mutants a rounded shape and higher cell dia-meter if compared to wild type cells especially in thestationary phase [10] We performed the analysis of theZbΔgas1 and wild type strains in parallel to the S cere-visiae BY4741 Δgas1 and wt strains Microscopy imagesare shown in Figure 3 It is possible to see the typicalrounded phenotype also for the ZbΔgas1 (panels C andG) Moreover like in S cerevisiae intracellular granulesand altered vacuoles are also visible (indicated byarrows panels C and G) The altered shape is alsoreflected by cellular volumes being higher in themutants as assessed by coulter counter measurementsin cell populations growing in minimal or in rich med-ium like for the model yeast differences are more pro-nounced in stationary phase were the mutant cellvolume is 16 fold larger in respect to the wild typecells (data not shown) Another feature described forthe ScΔgas1 mutant is the appearance of large cellularaggregates still permanent after repeated sonicationcycles [10] This feature is also conserved in the Z bai-lii mutant as it is visible from the microscopy imagesof sonicated cells (Figure 3 panels C D G and H) Themorphological alterations described are likely a conse-quence of the altered structure and composition of themutant cell wall it was reported for S cerevisiae a
lowered glucans content together with a markedincrease in the chitin and mannoproteins content [15]Moreover an altered distribution of the cell wall com-ponents in the mother and daughter cell have beenreported [10] Our analyses confirmed these alterationsalso for the Z bailii mutant overall Calcofluor stainingin fact was more intense in Δgas1 than in wild typestrain (Figure 3 panels D and H) indicating a higherchitin content in the mutant The chitin seems to bedelocalized throughout the entire cell wall in themutant while in the wild type it is more concentratedat the budding ring and at the bud scars Moreover inthe mutant the cellular buds are also Calcofluor-stainedin contrast to the wild type It was proposed that theincrease in chitin levels and cross-links with b-16-glu-cosyl mannoproteins may be the effect of the activationof a compensatory mechanism in mutants characterizedby defects in the cell wall architecture like the Δgas1mutants The aim of this response would be to preventexcessive loss of mannoproteins as a consequence ofthe inability to cross-link the b-16-glucosyl mannopro-teins and the b-13-glucans a process in which Gas1pplays a central role [15] It was also hypothesized thatthe loss of Gas1p may cause a weakening of the cellwall of the emerging bud The yeast cell could inresponse to this weakening target the accumulation ofchitin at the bud surface in order to protect it fromlyses [16] Alteration of the processes directly or indir-ectly linked to budding leads to the appearance of arelative high fraction in the cell population of multi-budded cells (the so-called ldquoMickey Mouserdquo phenotype)(figure 3) A similar phenotype was also visible inZbΔgas1 mutants (Figure 3)The altered cell wall structure renders the deleted
bakersrsquo yeast strain more sensitive to sodium dodecylsulphate (SDS) and to Calcofluor White (CFW) as
Figure 2 Deletion of ZbGAS1 (a) Schematic representation of the ZbGAS1 deletion cassettes Restriction enzymes used to obtain the constructsand PCR primers used to verify recombination are indicated light grey bars represents ZbGAS1 sequence (b) Control PCR performed to verifythe deletion of both the allelic copies of ZbGAS1 Two independent mutant clones (A and B) were tested In both clones it is possible to see thepresence of bands corresponding to the two deletion cassettes and the absence of the band corresponding to the wild type gene copy
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 4 of 11
reported in literature for the W303 background [17]Here we confirm by spot assays the sensitivity to bothagents of the ScΔgas1 mutant background BY4741 (Fig-ure 3 upper right panels) and as well of the ZbΔgas1mutant (Figure 3 lower right panels)Finally to verify the functionality of the GAS1 homo-
logue of Z bailii in S cerevisiae and the similarity of itsbiological role the ZbGAS1 gene was expressed into theGAS1-deficient S cerevisiae strain resulting in wild typephenotype restoration either in respect to the morpholo-gical traits as well as for the chitin distribution (data not
shown) Moreover to verify the functionality of the Scerevisiae GAS1 in Z bailii the ScGAS1 gene wasexpressed into the GAS1-deficient Z bailii strain alsoin this case restoring the native phenotype (data notshown)Overall morphological and physiological data strongly
indicate that the ZbGAS1 gene may have a similar rolewith respect to the ScGAS1 homologous in the assess-ment of the cell wall structure and hence their inactiva-tion might have similar effects in the two yeasts inrespect to secretion abilities
Figure 3 Morphology of Z bailii and S cerevisiae wild type and gas1 null mutant strains and their sensitivity to Calcofluor White andSDS Cellular morphology From A to H dicroic and fluorescence microscope images of Z bailii and S cerevisiae wild type and gas1 null mutantstrains Cells were grown in minimal medium and harvested in stationary phase of growth Arrows indicate granules and enlarged vacuoles inmutant cells (panels C and G) Calcofluor White and SDS sensitivity of S cerevisiae (BY4741) and Z bailii (Zygo) gas1 null mutants 4 μl of culturesof the different strains (BY4741 BY4741 Δgas1 Zygo and Zygo Δgas1) were spotted starting with OD (660 nm) = 1 and then in 3 serial solution110 (234) on rich solid medium (YPD) and on the same medium added with 125 μg ml-1 of CFW or 50 μg ml-1 SDS Pictures were capturedafter 5 days of incubation at 30degC
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 5 of 11
Supersecretive phenotype of Δgas1 strainsAs already mentioned (see Introduction) the mostinteresting phenotype of Δgas1 mutants reported in lit-erature is the supersecretive property Besides theincreased secretion of heterologous proteins whichappears to be dependent on the individual protein [12]the levels of total secreted proteins in late-stationaryphase cultures of S cerevisiae mutants were two foldhigher with respect to the wild-type [7] We thereforestarted our evaluation of secretive properties of the Zbailii mutant in the perspective of a possible applica-tion in industrial processes by determining the amountof total proteins released in the growth medium duringbatch cultivation with respect to the wild type strain Zbailii and S cerevisiae wild type and Δgas1 respectivestrains were grown in shake flasks in minimal YNB-based medium containing 5 glucose Cells were har-vested at different times of cultivation and the fermen-tation broths were assayed At least triplicateexperiments were performed on three independenttransformants and Figure 4 shows the mean dataobtained reported as mg of total proteins secreted perlitre of the culture broth normalised for OD valuesThe values refers to 72 h of cultivation but it was pos-sible to observe a trend leading to this result starting atthe end of exponential phase and becoming more pro-nounced in stationary phase of growth
The data clearly underline the overall higher secretivecapacity of Z bailii if compared to S cerevisiae inaccordance to heterologous proteins secretion dataalready reported [3] Moreover they indicate a signifi-cant improvement in the secretive capability of the Zbailii mutant being nearly 80 more than the wild typein terms of total endogenous protein similarly to the Scerevisiae mutantOverexpression of CRL lipase XPR2 protease and hIL-1bin Z bailii Δgas1 strainsWith the intention to evaluate the potential effects ofcell wall engineering and to test the secretive perfor-mance in the presence of an heterologous protein threemodel proteins were tested a fungal lipase from Can-dida rugosa (CRL1) a fungal protease from Yarrowialipolytica (XPR2) and the human interleukin 1b (hIL-1b)Four clones of Z bailii Δgas1 [pZLN022XPR2] and
four clones of the parental strain Z bailii[pZLN022XPR2] together with control clones harbour-ing the respective empty plasmid were pre-cultivated for16 hours in shake flasks using YPD medium and thentransferred in shake flasks in minimal YNB-based med-ium containing 5 glucose The amount of secretedrecombinant XPR2 in culture supernatants was tested interms of enzymatic activity after 72 96 120 and 148 hwith the Azocoll method (see methods for detail) The
Figure 4 Total proteins released in the culture medium Total proteins released in the culture medium of batch cultures of the indicatedwild type and Δgas1 strains of S cerevisiae and Z bailii Yeasts were grown in shake flasks in minimal YNB-based medium with 5 glucose Cellswere inoculated at 01 OD and harvested after 72 hours of cultivation Amount of released proteins (in mg l-1 of culture broth) were normalizedfor OD values
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 6 of 11
ZbΔgas1 transformants showed a relevant increase inspecific activity levels since 72 h (+ 25 at 72 h morethan two-fold at 148 h see Figure 5a) in respect to wildtype transformantsSimilarly three clones of Z bailii Δgas1
[pZLN022LIP1] and three clones of the parental strainZ bailii [pZLN022LIP1] together with control clonesharbouring the respective empty plasmid were pre-culti-vated for 16 hours in shake flasks using YPD mediumand then transferred in shake flasks in minimal YNB-based medium containing 5 glucose The amount ofsecreted recombinant LIP1 in culture supernatants wastested in terms of enzymatic activity after 24 48 72 and120 h using the hydrolysis of p-nitrophenylestersmethod (see methods for detail) Also for this secondheterologous product the Δgas1 transformants showed arelevant increase in specific activity levels since 24 h
(+35 at 24 h more than two-fold at 120 h see Figure5b) in respect to wild type transformantsFinally three clones of Z bailii Δgas1 [pZ5KlIL-1b]
and three clones of the parental strain Z bailii[pZ5KlIL-1b] together with control clones harbouringthe respective empty plasmid were tested for secretionlevel of the human interleukin since the codingsequence was functionally linked to the signal peptide ofthe K1 killer toxin from K lactis The independenttransformants were pre-cultivated and then cultured inshake flasks containing rich medium (YPD 5 wv glu-cose) Samples were collected at the indicated times (seeFigure 5c) cells were harvested and both the superna-tants and the cell protein extracts of said samples wereloaded on a polyacrylamide gel The data obtained byWestern blot (see Figure 5c) performed on the superna-tant showed a well represented band corresponding to
Figure 5 Secretion levels of yeast protease lipase and rhIL-1b in Z bailii and ZbΔgas1 (a) Secretion levels of protease in Z bailii andZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutant transformed with controlplasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with XPR2 gene (b) Secretion levels oflipase in Z bailii and ZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutanttransformed with control plasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with LIP1 gene(c) Western blots for rhIL-1b of protein samples from supernatants and cell extracts of Z bailii and ZbΔgas1 transformants grown in YPD mediawith 5 glucose inoculated at OD660 = 01 From left to right (A) rhIL-1b (B) negative control Z bailii[pZ5] (C) (D) (E) (I) (L) (M) Z bailii[pZ5KlrhIL-1b] (F) (G) (H) (N) (O) (P) ZbΔgas1 [pZ5KlrhIL-1b] Time and sample types are indicated in the picture
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 7 of 11
the recombinant protein for both Z bailii and Z bailiiΔgas1 samples but differently from the case of recombi-nant fungal enzymes no relevant differences in favourof the deleted yeast are appreciable Remarkably the sig-nal corresponding to the protein retained intracellularlyis more intense in gas1 null mutant than the signalobtained from Z bailii parental strain indicating anincomplete secretion of this protein
DiscussionsCell wall engineering can improve the release of het-erologous proteins from yeasts [18] Generally speak-ing GAS1 which mediates the crosslinking of cell wallcomponents is directly involved in defining the perme-ability of the cell Cloning of the unknown Z bailiiGAS1 homologue by PCR was only possible startingfrom highly conserved internal regions identified uponalignment of the GAS1 homologues of three yeast spe-cies The 5rsquo and 3rsquo ends were subsequently amplifiedusing a Genome Walker library With this approachthe full-length Z bailii GAS1 homologue could beamplified and sequenced The cloned gene wasexpressed in the GAS1-deficient S cerevisiae strain andreversion of the phenotype proved that this homologueis not only structurally but also functionally similar tothe S cerevisiae gene Moreover also the ScGAS1 genecomplemented the Z bailii mutant Disruption ofGAS1 in Z bailii revealed its influence on the cell wallmorphology The GAS1 null mutants of Z bailiishowed the same phenotype reported for S cerevisiae[10] with enlarged cells often showing granular cyto-plasm an alterated chitin distribution two buds(Mickey Mouse-like appearance) and big vacuoles Thevacuole is known to be a dynamic organelle whosemorphology is highly responsive to different extracellu-lar and intracellular stimulistressors [19] In the GAS1null mutant it can be therefore speculated that thegeneration of big vacuoles is related to stressful condi-tions as already suggested by the altered chitin distri-bution being a possible defence mechanism againstlysis or related to a generalized response to sickness ofthe cells [10]Finally also the sensitivity to SDS and CFW
appeared very similar to what observed in the S cerevi-siae mutant A comparison of the different GAS1 nullphenotype mutants and the functionality of the Z bai-lii GAS1 homologue in S cerevisiae suggest a similarityof the cell wall structure of Z bailii compared with thecell wall structure of S cerevisiae Considering thesecretory pathway of a secreted protein it was specu-lated whether the protein becomes retarded in the cellwall after release from the plasma membrane The dis-ruption of GAS1 in Z bailii first of all confirmed ageneral phenomenon of protein release in respect to
the wild type strain as demonstrated by the higheraccumulation of total protein in the growing mediumof the deleted strain when compared to the parentalstrain Moreover such as compromised cell wall per-meability in the production strains for XPR2 and LIP1does lead to a clear improvement in the secretion ofthe respective proteins In the case of h IL-1b theenhancement of product secretion was not so evidentand difficult to interpret in respect to the higher accu-mulation of intracellular product It could be specu-lated that this can be related to the nature or to thesource of the protein but too few information areavailable at the moment to further comment in thisdirection What seems to be true is unfortunately whatwe very often experience for recombinant protein pro-duction which is the protein and protein-host depen-dence In fact also in P pastoris two out of threeheterologous proteins resulted more secreted in themutant as for Z bailii and when we tested the sameprotein in particular the IL-1b secretion in the S cere-visiae Δgas1 mutant (data not shown) we did not reg-ister any improvement
MethodsYeast strains media and transformationThe S cerevisiae strains used in this study were theauxotrophic haploid BY4741 (Mat a his3Δ1 leu2Δ0met5Δ0 ura3Δ0) and the relative null mutant in GAS1sequence called in this study BY4741 Δgas1 (Mat ahis3Δ1 leu2Δ0 met5Δ0 ura3Δ0 YMR307kanMX4)Both strains are commercially available from Euroscarfcollection httpwebuni-frankfurtdefb15mikroeuro-scarfcol_indexhtml The Z bailii wild type strain isATCC60483 Yeast cultures were grown in YP medium(10 g l-1 yeast extract 20 g l-1 peptone) with 2 or 5wv glucose (D) or fructose (F) as carbon sourceWhere needed NAT (80 mg l-1) or G418 (500 mg l-1
for S cerevisiae and 200 mg l-1 for Z bailii) or hygro-micine (100 mg l-1 for Z bailii) were added to themedia Alternatively yeast cultures were grown inminimal synthetic medium (067 w v-1 YNB Biolifewithout amino acids) with 2 or 5 w v-1 glucose ascarbon source When required supplements such ashistidine leucine methionine and uracil were added toa final concentration of 50 mg l-1 Media for plateswere solidified by addition of agar (Biolife Milan Italy)to 15 w v-1 Shake flask experiments were carriedout with 50 ml of medium in 250 ml shake flasks on ashaker at 30degC with 160 rpm For sensitivity tests inPetri dishes 125 μg ml-1 of CFW or 50 μg ml-1 SDSwere added to solid media Yeast transformation wasperformed using the Lithium AcetatessDNA methodas described [20] and slightly adapted for Z bailii asdescribed [3]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 8 of 11
Genes amplifications deletion cassettes and expressionplasmid constructionPCR amplifications were made from yeasts genomicDNA prepared as described in [21] Amplifications wereperformed by the Pwo DNA Polymerase (Roche)The amplification of the ZbGAS1 gene was as follows
the primers GAS1int_fw 5rsquo-AGT TGT TCC AGA GATATT CCA TAC CTC AA-3rsquo and GAS1int_rev 5rsquo-GCAGAA CCG CTG AAG CTA CAG T-3rsquo designed oninternal conserved region of the available sequencesfrom other yeasts were firstly used to amplify an internalregion of 1137 bp of the Z bailii gene with the follow-ing program 94degC 5 min (94degC 15 s 57degC 30 s 72degC 1min) times 25 72degC 7 min 4degC infin After that the designedprimers ZbGAS1int_fw 5rsquo-GGA CGA TAA GGT TGACGA AGA-3rsquo and ZbGAS1int_rev 5rsquo-TGC TGT GGTCCA AAG TTG TG-3rsquo were used for the amplification(program 7 cycles 94degC 2 min 72degC 3 min 32 cycles94degC 2 min 67degC 3 min 67degC 4 min 4degC infin) of theterminal ends of ZbGAS1 utilizing a Z bailii GenomeWalker library constructed using the Universal GenomeWalker kit (Clontech) following the manufacturersinstructions From the obtained and reconstructedsequence the final primers were established and used forthe ZbGAS1 1710 bp full length coding sequence ampli-fication ZbGAS1_fw 5rsquo-ACT AAT GTT ATT CCAGGC GTT TTC G-3rsquo and ZbGAS1real_rev 5rsquo-AAATCA AGC CAA AGC AAA TCC AGC A-3rsquo (program94degC 5 min (94degC 20 s 58degC 30 s 72degC 2 min) times 30 72degC 10 min 4degC infin) The DNA sequence of ScGAS1 wasPCR amplified with the following primers ScGAS1fw5rsquo-ACA ATG TTG TTT AAA TCC CTT TCA AAGTTA GCA A-3rsquo and ScGAS1rev 5rsquo-TTT TTA AACCAA AGC AAA ACC GAC ACC AG-3rsquo (program 94degC5 min (94degC 20 s 605degC 30 s 72degC 2 min) times 30 72degC10 min 4degC infin) and the DNA sequence of YlXPR2 wasPCR amplified with the following primers XPR2_fwd5rsquo-ACA ATG AAG CTC GCT ACC GCC TTT A-3rsquoand XPR2_rev 5rsquo-TGC CTA AAT GCC AAC ACCGTT GTA G-3rsquo (program 94degC 4 min (94degC 15 s 625degC 30 s 72degC 2 min) times 30 72degC 7 min 4degC infin) All theunique fragments obtained were sub-cloned in the vec-tor pSTblue-1 utilising the Perfectly Bluntreg Cloning Kit(Novagene) and sequenced The resulting respectiveplasmids utilised in this work were named as followspSTbZbGAS1int pSTbZbGAS1 pSTbScGAS1 andpSTbYlXPR2For the construction of the ZbGAS1 deletion cassettes
the KanR or the hphR cassette derived from plasmidpFA6-KanMX4 [22] and pAG26 [23] and conferringresistance to G418 and hygromycin respectively werebluntended excised and inserted in the pSTbZbGAS1intopened BstEII BglII bluntended and dephosphorylatedresulting in the plasmids pSTbZbGAS1ΔK and
pSTbZbGAS1ΔH The resistance cassettes resulted inboth cases counter-clockwise inserted The Z bailii wildtype cells were transformed with the respective deletioncassettes obtained by cutting the two described plasmidsin the AccI and SnaBI sites The effective deletion wasPCR checked by using the following couples of primersZbG1int2_fw 5rsquo- TGC AGA AGT TAC AGA CCAATG TTG T -3rsquo and and ZbG1int2_rev 5rsquo-AAG ATAGAG AAG TGC TCT TGG CA -3rsquo (see Figure 2) orZbG1int2_fw and Hph_fw 5rsquo- ATA TGA AAA AGCCTG AAC TCA CCG AC -3rsquo or Kan_fw 5rsquo- ATG GGTAAG GAA AAG ACT CAC GTT -3rsquo (not shown) inorder to verify the presence of the cassettes in thedesired locusFor complementing the yeasts GAS1 deletion pZ5
(-Nco)ScGAS1 and pYX022ZbGAS1 were constructedpZ5(-Nco)ScGAS1 was obtained from pZ5(-Nco) openedSacIEcoRV with the insertion of ScGAS1 frompSTbScGAS1 cut with SacISnaBI The pZ5(-Nco)derives from pZ3 [3] where at the KpnI site the KanR
cassette was substituted from the NATR cassetteobtained from vector pAG25 [23] by cutting PvuIISacIblunt (obtaining the pZ5 plasmid) finally this plasmidwas EcoRIBamHI cut and bluntended and reclosedpYX022ZbGAS1 was obtained with the insertion ofZbGAS1 from pSTbZbGAS1 into pYX022 (RampD Sys-tems Wiesbaden Germany) both EcoRI cutFor heterologous protein expression the plasmids
pZLN022XPR2 pZLN022LIP1 and pZ5KlhIL-1b wereconstructed The first two integrative plasmids derivesfrom pZLN022 (manuscript in preparation) obtainedfrom pYX022 opened DraIII bluntSpeI blunt (toexclude HIS3) with the insertion of ZbLEU2 cut frompSTbZbLEU2 with EcoRI blunt in KpnI blunt site theNATR cassette was inserted obtained from vectorpAG25 [23] by cutting PvuIISacIblunt ThepSTbZbLEU2 was obtained by sub-cloning ZbLEU2amplified by PCR from Z bailii genomic DNA (manu-script in preparation) in pSTBlue (Novagen) cut byEcoRV For pZLN022XPR2 construction the pZLN022was AatIINheI cut and the XPR2 from pSTbXPR2 wasinserted For pZLN022LIP1 construction the pZLN022was AatII bluntNheI blunt opened and the LIP1 wasinserted by BamHI bluntNsiI blunt excision from thepGAPsLIP [24] For rh interleukin the pZ5KlhIL-1b wasbuilt it was obtained from pZ5 opened with EcoRI-blunt with the insertion of hIL-1b in frame with Kl sig-nal sequence from vector pKSSPI3 [25] cut EcoRIbluntXbaI bluntDNA manipulations transformation and cultivation of
Escherichia coli (DH5aFrsquo (80dlacZΔM15 Δ (lacZYA-argF) U169 deo rec1 end1 sup44 l THI-1 gyrA96relA1) and Novablue Competent Cells (Novagene) wereperformed following standard protocols [26] All the
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 9 of 11
restriction and modification enzymes used were fromNew England Biolabs (Hitchin Herts UK) or fromRoche Diagnostics (Mannheim Germany)Calcofluor staining and fluorescence microscopyCells were harvested washed in PBS and resuspended in1 mg ml-1 Calcofluor White solution incubated 10-30min at room temperature washed again twice in PBSFluorescence microscope images were taken by a NikonEclipse 90i microscope using the Metamorph softwareversion 22 (Nikon) using emission filter UV-1AQuantification of total released proteinsCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were etha-nol-precipitated The amount of proteins was deter-mined by the dye-binding method of Bradford (QuickStart Bradford Dye Reagent - BIORAD) using BSA asstandard All the data were normalized by OD after ver-ification of the linear correlation between OD and dryweight as a measure of biomassQuantification of secreted proteaseCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were sepa-rated and stored at -20degC until analysis The amount ofprotease secreted in the medium was determined by theAzocoll assay (Azocoll - Calbiochem) using commercialprotease as positive control Azocoll assays were per-formed according to [27] with some modificationsReaction mixtures contained 15 mg ml-1 Azocoll 50mM TrisHCl (pH 76) 150 mM NaCl and 5 mMCaCl2 Proteolytic activities were measured by absor-bance changes at 490 nm (Shimadzu UV 1601) afterincubation for 18 h at 30degCQuantification of secreted lipaseCultures were harvested and centrifuged at maximalspeed then the supernatants were separated and storedat -20degC until analysis The amount of lipase secreted inthe medium was determined according to [28] withsome modifications Reaction mixtures contained (A)60 mg of p-nitrophenyl palmitate (Sigma Aldrich) in 20ml of isopropanol (Sigma Aldrich) (B) 1 g Triton X-100(Sigma Aldrich) and 02 g of Arabic gum (SigmaAldrich) in 200 ml TrisHCl 01 M (pH 75) To obtainthe mixture (C) add slowly 1 part of A solution to 9part of B solution under continuous agitation Add 9part of (C) solution to one part of the enzyme contain-ing solution (ie the supernatant) Lipase activities weremeasured by absorbance changes at 410 nm (ShimadzuUV 1601) and the amount of enzyme is proportional toOD according to the relation Um = 0667 times OD410 nm
(if OD410 nm lt 05)Quantification of interleukinHuman IL-1b were analysed by Western blot Indepen-dent transformants were cultured in shake flasks inminimal or rich medium and during the kinetics
samples were collected Cells were harvested (a culturevolume corresponding to 108 cells) by centrifugationOne volume of 2timesLaemmli buffer [29] was added to thesupernatants of said samples they were boiled for 3-5min and stored at -20degC until loading or loaded directlyon a polyacrylamide gel For the analysis of intracellularproduction crude extracts were prepared from 108 cellsby following the trichloroacetic acid protocol [30] andresuspending the final protein extract in 150 μl ofLaemmli buffer Samples were then loaded on standardpolyacrylamide gels (sodium dodecyl sulfate-polyacryla-mide gel electrophoresis final concentration of theseparating gel 15) after separation proteins wereblotted to nitrocellulose membranes and immunodeco-rated An anti-IL-1b rabbit polyclonal antibody (IL-1b(H-153) Santa Cruz Biotechnology USA cat no sc-7884 dilution 1200) was used for the interleukin immu-nodecoration An anti-rabbit IgG horseradish peroxi-dase-conjugated (Amersham Pharmacia Biotech UK catno NA934 dilution 110000) secondary antibody wasused For the interleukin detection as a positive controlthe human recombinant IL-1b from E coli (Roche catno 1 457 756) was always loaded in parallel on thepolyacrylamide gels The proteins were visualised usingthe ECL Western blotting system (Amersham Bios-ciences UK) according to the manufacturerrsquosinstructions
Additional file 1 Clustalw_ntd Nucleotidic sequence of the ZbGAS1gene and alignment with the homologue ScGAS1 (entry YMR307W)Grey coloring represent nucleotidic identity Alignment was performedwith ClustalW software httpwwwebiacukclustalw and graphicalediting was made using Jalview software [31]Click here for file[ httpwwwbiomedcentralcomcontentsupplementary1475-2859-9-7-S1PDF ]
AcknowledgementsSpecial thanks to Valeria Longo and Davide Zocco for technicalcontributions and to Stefania Brocca for providing the pGAPsLIP plasmidThis work was partially supported by FAR 2007 and 2008 (12-1-192) to DP
Author details1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy 2CPC Biotech Srl Naples Italy
Authorsrsquo contributionsPB and DP initiated and coordinated the project LD was responsible forZbGAS1 gene identification by PCR and its deletion LR analyzed the mutantphenotype with CFW and SDS LR and SP performed the batch cultivationanalyzed the secretory phenotype in null mutant and performed theexperiment with heterologous proteins All authors wrote the paper andapproved the final version of the manuscript
Competing interestsThe authors declare that they have no competing interests
Received 2 November 2009Accepted 26 January 2010 Published 26 January 2010
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 10 of 11
References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11

for the 5rsquo and 3rsquo ends respectively (see Methods) Thecomplete ORF of ZbGAS1 was reconstructed in thisway and additional 262 bp upstream to the ATG and177 bp after the STOP codon were also sequenced(GeneBank Accession Ndeg GU136594)Based on the new sequence it was possible to design
specific primers for the ZbGAS1 gene (see Methods)and a DNA fragment of the expected length was ampli-fied from the Z bailii genomic DNAThe complete ZbGAS1 ORF resulted to be 1710 bp
long (+ 30 bp vs ScGAS1) sharing a 67 sequence iden-tity with the S cerevisiae homologue (additional file 1)This sequence encodes for a putative aminoacidicsequence of 569 aa (+10 aminoacids vs ScGas1p) having68 identity with the ScGas1p protein (Figure 1)Sequence alignments were also performed between thenewly isolated Z bailii ORF and sequences of otherhomologous GAS genes in S cerevisiae (GAS1-5) thelower identity scores obtained for sequences other thanScGAS1 (not shown) further indicated that we clonedthe GAS1 homologue of Z bailiiHence considering the sequence similarities between
the ScGAS1 gene and their homologues in other yeaststhe GAS1 gene resulted to be highly conserved also inZ bailii The overall deduced protein structure seems tobe also conservedThe ZbGAS1 gene is one of the few Z bailii gene
sequences isolated to date hence representing a furtherlittle step towards the knowledge of this still poorlycharacterized yeast
ZbGAS1 gene disruptionTwo different ZbGAS1 deletion cassettes were con-structed based on the kanR and hphR markers Differ-ently from the case of the ZbYME2 gene deletion [14]where flanking regions of about 60 to 80 bp weredescribed to be sufficient for obtaining homologousrecombination long homologous flanking regions werenecessary to obtain gene replacement In particular weconstructed both the deletion cassettes represented inFigure 2a with flanking regions of about 400 bp whichresulted to be sufficient to disrupt the first and as wellthe second allelic copy of ZbGAS1 despite with quitelow efficiency (about only 10 of the transformantsresulted to effectively have both the deleted alleles con-firming what previously described [14]) The two cas-settes (see Methods) were therefore used to transformZ bailii cells resulting in the gas1 null mutant Theinsertion of both deletion cassettes in the homologousloci was confirmed by PCR (Figure 2b) We applied thesame strategy for the deletion of the first essential Zbailii gene leading to the first auxotrophic mutant ofthis yeast and in this case we had to further extend thelength of the homologous region for specific recombina-tion and despite that the percentage of positive eventswas even lower (manuscript in preparation) Theseresults could lead to hypothesize that for targeted genedeletion in Z bailii the length of the flanking regionscould be crucial according to the specific role that aspecific protein plays thus explaining results obtainedby other researchers [14]
Figure 1 Clustalw_aapdf Putative aminoacidic sequence of the ZbGas1p protein and alignment with the homologue ScGas1p Grey scalecoloring represents aminoacid similarity based on Blosum 62 scores Alignment was performed with ClustalW software httpwwwebiacukclustalw and graphical editing was made using Jalview software [31]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 3 of 11
Morphological and physiological characterization of thenull mutantThe GAS1 gene inactivation leads to different peculiarphenotypes in S cerevisiae that were widely character-ized [10] We therefore performed a morphological andphysiological characterization of the obtained Z bailiiΔgas1 mutant in analogy to what reported in literaturefor the model yeast First some morphological traits ofthe Sc Δgas1 mutant are visible both by light and byfluorescence microscopy after Calcofluor White stainingIt was reported for S cerevisiae (both haploid anddiploid) mutants a rounded shape and higher cell dia-meter if compared to wild type cells especially in thestationary phase [10] We performed the analysis of theZbΔgas1 and wild type strains in parallel to the S cere-visiae BY4741 Δgas1 and wt strains Microscopy imagesare shown in Figure 3 It is possible to see the typicalrounded phenotype also for the ZbΔgas1 (panels C andG) Moreover like in S cerevisiae intracellular granulesand altered vacuoles are also visible (indicated byarrows panels C and G) The altered shape is alsoreflected by cellular volumes being higher in themutants as assessed by coulter counter measurementsin cell populations growing in minimal or in rich med-ium like for the model yeast differences are more pro-nounced in stationary phase were the mutant cellvolume is 16 fold larger in respect to the wild typecells (data not shown) Another feature described forthe ScΔgas1 mutant is the appearance of large cellularaggregates still permanent after repeated sonicationcycles [10] This feature is also conserved in the Z bai-lii mutant as it is visible from the microscopy imagesof sonicated cells (Figure 3 panels C D G and H) Themorphological alterations described are likely a conse-quence of the altered structure and composition of themutant cell wall it was reported for S cerevisiae a
lowered glucans content together with a markedincrease in the chitin and mannoproteins content [15]Moreover an altered distribution of the cell wall com-ponents in the mother and daughter cell have beenreported [10] Our analyses confirmed these alterationsalso for the Z bailii mutant overall Calcofluor stainingin fact was more intense in Δgas1 than in wild typestrain (Figure 3 panels D and H) indicating a higherchitin content in the mutant The chitin seems to bedelocalized throughout the entire cell wall in themutant while in the wild type it is more concentratedat the budding ring and at the bud scars Moreover inthe mutant the cellular buds are also Calcofluor-stainedin contrast to the wild type It was proposed that theincrease in chitin levels and cross-links with b-16-glu-cosyl mannoproteins may be the effect of the activationof a compensatory mechanism in mutants characterizedby defects in the cell wall architecture like the Δgas1mutants The aim of this response would be to preventexcessive loss of mannoproteins as a consequence ofthe inability to cross-link the b-16-glucosyl mannopro-teins and the b-13-glucans a process in which Gas1pplays a central role [15] It was also hypothesized thatthe loss of Gas1p may cause a weakening of the cellwall of the emerging bud The yeast cell could inresponse to this weakening target the accumulation ofchitin at the bud surface in order to protect it fromlyses [16] Alteration of the processes directly or indir-ectly linked to budding leads to the appearance of arelative high fraction in the cell population of multi-budded cells (the so-called ldquoMickey Mouserdquo phenotype)(figure 3) A similar phenotype was also visible inZbΔgas1 mutants (Figure 3)The altered cell wall structure renders the deleted
bakersrsquo yeast strain more sensitive to sodium dodecylsulphate (SDS) and to Calcofluor White (CFW) as
Figure 2 Deletion of ZbGAS1 (a) Schematic representation of the ZbGAS1 deletion cassettes Restriction enzymes used to obtain the constructsand PCR primers used to verify recombination are indicated light grey bars represents ZbGAS1 sequence (b) Control PCR performed to verifythe deletion of both the allelic copies of ZbGAS1 Two independent mutant clones (A and B) were tested In both clones it is possible to see thepresence of bands corresponding to the two deletion cassettes and the absence of the band corresponding to the wild type gene copy
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 4 of 11
reported in literature for the W303 background [17]Here we confirm by spot assays the sensitivity to bothagents of the ScΔgas1 mutant background BY4741 (Fig-ure 3 upper right panels) and as well of the ZbΔgas1mutant (Figure 3 lower right panels)Finally to verify the functionality of the GAS1 homo-
logue of Z bailii in S cerevisiae and the similarity of itsbiological role the ZbGAS1 gene was expressed into theGAS1-deficient S cerevisiae strain resulting in wild typephenotype restoration either in respect to the morpholo-gical traits as well as for the chitin distribution (data not
shown) Moreover to verify the functionality of the Scerevisiae GAS1 in Z bailii the ScGAS1 gene wasexpressed into the GAS1-deficient Z bailii strain alsoin this case restoring the native phenotype (data notshown)Overall morphological and physiological data strongly
indicate that the ZbGAS1 gene may have a similar rolewith respect to the ScGAS1 homologous in the assess-ment of the cell wall structure and hence their inactiva-tion might have similar effects in the two yeasts inrespect to secretion abilities
Figure 3 Morphology of Z bailii and S cerevisiae wild type and gas1 null mutant strains and their sensitivity to Calcofluor White andSDS Cellular morphology From A to H dicroic and fluorescence microscope images of Z bailii and S cerevisiae wild type and gas1 null mutantstrains Cells were grown in minimal medium and harvested in stationary phase of growth Arrows indicate granules and enlarged vacuoles inmutant cells (panels C and G) Calcofluor White and SDS sensitivity of S cerevisiae (BY4741) and Z bailii (Zygo) gas1 null mutants 4 μl of culturesof the different strains (BY4741 BY4741 Δgas1 Zygo and Zygo Δgas1) were spotted starting with OD (660 nm) = 1 and then in 3 serial solution110 (234) on rich solid medium (YPD) and on the same medium added with 125 μg ml-1 of CFW or 50 μg ml-1 SDS Pictures were capturedafter 5 days of incubation at 30degC
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 5 of 11
Supersecretive phenotype of Δgas1 strainsAs already mentioned (see Introduction) the mostinteresting phenotype of Δgas1 mutants reported in lit-erature is the supersecretive property Besides theincreased secretion of heterologous proteins whichappears to be dependent on the individual protein [12]the levels of total secreted proteins in late-stationaryphase cultures of S cerevisiae mutants were two foldhigher with respect to the wild-type [7] We thereforestarted our evaluation of secretive properties of the Zbailii mutant in the perspective of a possible applica-tion in industrial processes by determining the amountof total proteins released in the growth medium duringbatch cultivation with respect to the wild type strain Zbailii and S cerevisiae wild type and Δgas1 respectivestrains were grown in shake flasks in minimal YNB-based medium containing 5 glucose Cells were har-vested at different times of cultivation and the fermen-tation broths were assayed At least triplicateexperiments were performed on three independenttransformants and Figure 4 shows the mean dataobtained reported as mg of total proteins secreted perlitre of the culture broth normalised for OD valuesThe values refers to 72 h of cultivation but it was pos-sible to observe a trend leading to this result starting atthe end of exponential phase and becoming more pro-nounced in stationary phase of growth
The data clearly underline the overall higher secretivecapacity of Z bailii if compared to S cerevisiae inaccordance to heterologous proteins secretion dataalready reported [3] Moreover they indicate a signifi-cant improvement in the secretive capability of the Zbailii mutant being nearly 80 more than the wild typein terms of total endogenous protein similarly to the Scerevisiae mutantOverexpression of CRL lipase XPR2 protease and hIL-1bin Z bailii Δgas1 strainsWith the intention to evaluate the potential effects ofcell wall engineering and to test the secretive perfor-mance in the presence of an heterologous protein threemodel proteins were tested a fungal lipase from Can-dida rugosa (CRL1) a fungal protease from Yarrowialipolytica (XPR2) and the human interleukin 1b (hIL-1b)Four clones of Z bailii Δgas1 [pZLN022XPR2] and
four clones of the parental strain Z bailii[pZLN022XPR2] together with control clones harbour-ing the respective empty plasmid were pre-cultivated for16 hours in shake flasks using YPD medium and thentransferred in shake flasks in minimal YNB-based med-ium containing 5 glucose The amount of secretedrecombinant XPR2 in culture supernatants was tested interms of enzymatic activity after 72 96 120 and 148 hwith the Azocoll method (see methods for detail) The
Figure 4 Total proteins released in the culture medium Total proteins released in the culture medium of batch cultures of the indicatedwild type and Δgas1 strains of S cerevisiae and Z bailii Yeasts were grown in shake flasks in minimal YNB-based medium with 5 glucose Cellswere inoculated at 01 OD and harvested after 72 hours of cultivation Amount of released proteins (in mg l-1 of culture broth) were normalizedfor OD values
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 6 of 11
ZbΔgas1 transformants showed a relevant increase inspecific activity levels since 72 h (+ 25 at 72 h morethan two-fold at 148 h see Figure 5a) in respect to wildtype transformantsSimilarly three clones of Z bailii Δgas1
[pZLN022LIP1] and three clones of the parental strainZ bailii [pZLN022LIP1] together with control clonesharbouring the respective empty plasmid were pre-culti-vated for 16 hours in shake flasks using YPD mediumand then transferred in shake flasks in minimal YNB-based medium containing 5 glucose The amount ofsecreted recombinant LIP1 in culture supernatants wastested in terms of enzymatic activity after 24 48 72 and120 h using the hydrolysis of p-nitrophenylestersmethod (see methods for detail) Also for this secondheterologous product the Δgas1 transformants showed arelevant increase in specific activity levels since 24 h
(+35 at 24 h more than two-fold at 120 h see Figure5b) in respect to wild type transformantsFinally three clones of Z bailii Δgas1 [pZ5KlIL-1b]
and three clones of the parental strain Z bailii[pZ5KlIL-1b] together with control clones harbouringthe respective empty plasmid were tested for secretionlevel of the human interleukin since the codingsequence was functionally linked to the signal peptide ofthe K1 killer toxin from K lactis The independenttransformants were pre-cultivated and then cultured inshake flasks containing rich medium (YPD 5 wv glu-cose) Samples were collected at the indicated times (seeFigure 5c) cells were harvested and both the superna-tants and the cell protein extracts of said samples wereloaded on a polyacrylamide gel The data obtained byWestern blot (see Figure 5c) performed on the superna-tant showed a well represented band corresponding to
Figure 5 Secretion levels of yeast protease lipase and rhIL-1b in Z bailii and ZbΔgas1 (a) Secretion levels of protease in Z bailii andZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutant transformed with controlplasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with XPR2 gene (b) Secretion levels oflipase in Z bailii and ZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutanttransformed with control plasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with LIP1 gene(c) Western blots for rhIL-1b of protein samples from supernatants and cell extracts of Z bailii and ZbΔgas1 transformants grown in YPD mediawith 5 glucose inoculated at OD660 = 01 From left to right (A) rhIL-1b (B) negative control Z bailii[pZ5] (C) (D) (E) (I) (L) (M) Z bailii[pZ5KlrhIL-1b] (F) (G) (H) (N) (O) (P) ZbΔgas1 [pZ5KlrhIL-1b] Time and sample types are indicated in the picture
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 7 of 11
the recombinant protein for both Z bailii and Z bailiiΔgas1 samples but differently from the case of recombi-nant fungal enzymes no relevant differences in favourof the deleted yeast are appreciable Remarkably the sig-nal corresponding to the protein retained intracellularlyis more intense in gas1 null mutant than the signalobtained from Z bailii parental strain indicating anincomplete secretion of this protein
DiscussionsCell wall engineering can improve the release of het-erologous proteins from yeasts [18] Generally speak-ing GAS1 which mediates the crosslinking of cell wallcomponents is directly involved in defining the perme-ability of the cell Cloning of the unknown Z bailiiGAS1 homologue by PCR was only possible startingfrom highly conserved internal regions identified uponalignment of the GAS1 homologues of three yeast spe-cies The 5rsquo and 3rsquo ends were subsequently amplifiedusing a Genome Walker library With this approachthe full-length Z bailii GAS1 homologue could beamplified and sequenced The cloned gene wasexpressed in the GAS1-deficient S cerevisiae strain andreversion of the phenotype proved that this homologueis not only structurally but also functionally similar tothe S cerevisiae gene Moreover also the ScGAS1 genecomplemented the Z bailii mutant Disruption ofGAS1 in Z bailii revealed its influence on the cell wallmorphology The GAS1 null mutants of Z bailiishowed the same phenotype reported for S cerevisiae[10] with enlarged cells often showing granular cyto-plasm an alterated chitin distribution two buds(Mickey Mouse-like appearance) and big vacuoles Thevacuole is known to be a dynamic organelle whosemorphology is highly responsive to different extracellu-lar and intracellular stimulistressors [19] In the GAS1null mutant it can be therefore speculated that thegeneration of big vacuoles is related to stressful condi-tions as already suggested by the altered chitin distri-bution being a possible defence mechanism againstlysis or related to a generalized response to sickness ofthe cells [10]Finally also the sensitivity to SDS and CFW
appeared very similar to what observed in the S cerevi-siae mutant A comparison of the different GAS1 nullphenotype mutants and the functionality of the Z bai-lii GAS1 homologue in S cerevisiae suggest a similarityof the cell wall structure of Z bailii compared with thecell wall structure of S cerevisiae Considering thesecretory pathway of a secreted protein it was specu-lated whether the protein becomes retarded in the cellwall after release from the plasma membrane The dis-ruption of GAS1 in Z bailii first of all confirmed ageneral phenomenon of protein release in respect to
the wild type strain as demonstrated by the higheraccumulation of total protein in the growing mediumof the deleted strain when compared to the parentalstrain Moreover such as compromised cell wall per-meability in the production strains for XPR2 and LIP1does lead to a clear improvement in the secretion ofthe respective proteins In the case of h IL-1b theenhancement of product secretion was not so evidentand difficult to interpret in respect to the higher accu-mulation of intracellular product It could be specu-lated that this can be related to the nature or to thesource of the protein but too few information areavailable at the moment to further comment in thisdirection What seems to be true is unfortunately whatwe very often experience for recombinant protein pro-duction which is the protein and protein-host depen-dence In fact also in P pastoris two out of threeheterologous proteins resulted more secreted in themutant as for Z bailii and when we tested the sameprotein in particular the IL-1b secretion in the S cere-visiae Δgas1 mutant (data not shown) we did not reg-ister any improvement
MethodsYeast strains media and transformationThe S cerevisiae strains used in this study were theauxotrophic haploid BY4741 (Mat a his3Δ1 leu2Δ0met5Δ0 ura3Δ0) and the relative null mutant in GAS1sequence called in this study BY4741 Δgas1 (Mat ahis3Δ1 leu2Δ0 met5Δ0 ura3Δ0 YMR307kanMX4)Both strains are commercially available from Euroscarfcollection httpwebuni-frankfurtdefb15mikroeuro-scarfcol_indexhtml The Z bailii wild type strain isATCC60483 Yeast cultures were grown in YP medium(10 g l-1 yeast extract 20 g l-1 peptone) with 2 or 5wv glucose (D) or fructose (F) as carbon sourceWhere needed NAT (80 mg l-1) or G418 (500 mg l-1
for S cerevisiae and 200 mg l-1 for Z bailii) or hygro-micine (100 mg l-1 for Z bailii) were added to themedia Alternatively yeast cultures were grown inminimal synthetic medium (067 w v-1 YNB Biolifewithout amino acids) with 2 or 5 w v-1 glucose ascarbon source When required supplements such ashistidine leucine methionine and uracil were added toa final concentration of 50 mg l-1 Media for plateswere solidified by addition of agar (Biolife Milan Italy)to 15 w v-1 Shake flask experiments were carriedout with 50 ml of medium in 250 ml shake flasks on ashaker at 30degC with 160 rpm For sensitivity tests inPetri dishes 125 μg ml-1 of CFW or 50 μg ml-1 SDSwere added to solid media Yeast transformation wasperformed using the Lithium AcetatessDNA methodas described [20] and slightly adapted for Z bailii asdescribed [3]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 8 of 11
Genes amplifications deletion cassettes and expressionplasmid constructionPCR amplifications were made from yeasts genomicDNA prepared as described in [21] Amplifications wereperformed by the Pwo DNA Polymerase (Roche)The amplification of the ZbGAS1 gene was as follows
the primers GAS1int_fw 5rsquo-AGT TGT TCC AGA GATATT CCA TAC CTC AA-3rsquo and GAS1int_rev 5rsquo-GCAGAA CCG CTG AAG CTA CAG T-3rsquo designed oninternal conserved region of the available sequencesfrom other yeasts were firstly used to amplify an internalregion of 1137 bp of the Z bailii gene with the follow-ing program 94degC 5 min (94degC 15 s 57degC 30 s 72degC 1min) times 25 72degC 7 min 4degC infin After that the designedprimers ZbGAS1int_fw 5rsquo-GGA CGA TAA GGT TGACGA AGA-3rsquo and ZbGAS1int_rev 5rsquo-TGC TGT GGTCCA AAG TTG TG-3rsquo were used for the amplification(program 7 cycles 94degC 2 min 72degC 3 min 32 cycles94degC 2 min 67degC 3 min 67degC 4 min 4degC infin) of theterminal ends of ZbGAS1 utilizing a Z bailii GenomeWalker library constructed using the Universal GenomeWalker kit (Clontech) following the manufacturersinstructions From the obtained and reconstructedsequence the final primers were established and used forthe ZbGAS1 1710 bp full length coding sequence ampli-fication ZbGAS1_fw 5rsquo-ACT AAT GTT ATT CCAGGC GTT TTC G-3rsquo and ZbGAS1real_rev 5rsquo-AAATCA AGC CAA AGC AAA TCC AGC A-3rsquo (program94degC 5 min (94degC 20 s 58degC 30 s 72degC 2 min) times 30 72degC 10 min 4degC infin) The DNA sequence of ScGAS1 wasPCR amplified with the following primers ScGAS1fw5rsquo-ACA ATG TTG TTT AAA TCC CTT TCA AAGTTA GCA A-3rsquo and ScGAS1rev 5rsquo-TTT TTA AACCAA AGC AAA ACC GAC ACC AG-3rsquo (program 94degC5 min (94degC 20 s 605degC 30 s 72degC 2 min) times 30 72degC10 min 4degC infin) and the DNA sequence of YlXPR2 wasPCR amplified with the following primers XPR2_fwd5rsquo-ACA ATG AAG CTC GCT ACC GCC TTT A-3rsquoand XPR2_rev 5rsquo-TGC CTA AAT GCC AAC ACCGTT GTA G-3rsquo (program 94degC 4 min (94degC 15 s 625degC 30 s 72degC 2 min) times 30 72degC 7 min 4degC infin) All theunique fragments obtained were sub-cloned in the vec-tor pSTblue-1 utilising the Perfectly Bluntreg Cloning Kit(Novagene) and sequenced The resulting respectiveplasmids utilised in this work were named as followspSTbZbGAS1int pSTbZbGAS1 pSTbScGAS1 andpSTbYlXPR2For the construction of the ZbGAS1 deletion cassettes
the KanR or the hphR cassette derived from plasmidpFA6-KanMX4 [22] and pAG26 [23] and conferringresistance to G418 and hygromycin respectively werebluntended excised and inserted in the pSTbZbGAS1intopened BstEII BglII bluntended and dephosphorylatedresulting in the plasmids pSTbZbGAS1ΔK and
pSTbZbGAS1ΔH The resistance cassettes resulted inboth cases counter-clockwise inserted The Z bailii wildtype cells were transformed with the respective deletioncassettes obtained by cutting the two described plasmidsin the AccI and SnaBI sites The effective deletion wasPCR checked by using the following couples of primersZbG1int2_fw 5rsquo- TGC AGA AGT TAC AGA CCAATG TTG T -3rsquo and and ZbG1int2_rev 5rsquo-AAG ATAGAG AAG TGC TCT TGG CA -3rsquo (see Figure 2) orZbG1int2_fw and Hph_fw 5rsquo- ATA TGA AAA AGCCTG AAC TCA CCG AC -3rsquo or Kan_fw 5rsquo- ATG GGTAAG GAA AAG ACT CAC GTT -3rsquo (not shown) inorder to verify the presence of the cassettes in thedesired locusFor complementing the yeasts GAS1 deletion pZ5
(-Nco)ScGAS1 and pYX022ZbGAS1 were constructedpZ5(-Nco)ScGAS1 was obtained from pZ5(-Nco) openedSacIEcoRV with the insertion of ScGAS1 frompSTbScGAS1 cut with SacISnaBI The pZ5(-Nco)derives from pZ3 [3] where at the KpnI site the KanR
cassette was substituted from the NATR cassetteobtained from vector pAG25 [23] by cutting PvuIISacIblunt (obtaining the pZ5 plasmid) finally this plasmidwas EcoRIBamHI cut and bluntended and reclosedpYX022ZbGAS1 was obtained with the insertion ofZbGAS1 from pSTbZbGAS1 into pYX022 (RampD Sys-tems Wiesbaden Germany) both EcoRI cutFor heterologous protein expression the plasmids
pZLN022XPR2 pZLN022LIP1 and pZ5KlhIL-1b wereconstructed The first two integrative plasmids derivesfrom pZLN022 (manuscript in preparation) obtainedfrom pYX022 opened DraIII bluntSpeI blunt (toexclude HIS3) with the insertion of ZbLEU2 cut frompSTbZbLEU2 with EcoRI blunt in KpnI blunt site theNATR cassette was inserted obtained from vectorpAG25 [23] by cutting PvuIISacIblunt ThepSTbZbLEU2 was obtained by sub-cloning ZbLEU2amplified by PCR from Z bailii genomic DNA (manu-script in preparation) in pSTBlue (Novagen) cut byEcoRV For pZLN022XPR2 construction the pZLN022was AatIINheI cut and the XPR2 from pSTbXPR2 wasinserted For pZLN022LIP1 construction the pZLN022was AatII bluntNheI blunt opened and the LIP1 wasinserted by BamHI bluntNsiI blunt excision from thepGAPsLIP [24] For rh interleukin the pZ5KlhIL-1b wasbuilt it was obtained from pZ5 opened with EcoRI-blunt with the insertion of hIL-1b in frame with Kl sig-nal sequence from vector pKSSPI3 [25] cut EcoRIbluntXbaI bluntDNA manipulations transformation and cultivation of
Escherichia coli (DH5aFrsquo (80dlacZΔM15 Δ (lacZYA-argF) U169 deo rec1 end1 sup44 l THI-1 gyrA96relA1) and Novablue Competent Cells (Novagene) wereperformed following standard protocols [26] All the
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 9 of 11
restriction and modification enzymes used were fromNew England Biolabs (Hitchin Herts UK) or fromRoche Diagnostics (Mannheim Germany)Calcofluor staining and fluorescence microscopyCells were harvested washed in PBS and resuspended in1 mg ml-1 Calcofluor White solution incubated 10-30min at room temperature washed again twice in PBSFluorescence microscope images were taken by a NikonEclipse 90i microscope using the Metamorph softwareversion 22 (Nikon) using emission filter UV-1AQuantification of total released proteinsCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were etha-nol-precipitated The amount of proteins was deter-mined by the dye-binding method of Bradford (QuickStart Bradford Dye Reagent - BIORAD) using BSA asstandard All the data were normalized by OD after ver-ification of the linear correlation between OD and dryweight as a measure of biomassQuantification of secreted proteaseCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were sepa-rated and stored at -20degC until analysis The amount ofprotease secreted in the medium was determined by theAzocoll assay (Azocoll - Calbiochem) using commercialprotease as positive control Azocoll assays were per-formed according to [27] with some modificationsReaction mixtures contained 15 mg ml-1 Azocoll 50mM TrisHCl (pH 76) 150 mM NaCl and 5 mMCaCl2 Proteolytic activities were measured by absor-bance changes at 490 nm (Shimadzu UV 1601) afterincubation for 18 h at 30degCQuantification of secreted lipaseCultures were harvested and centrifuged at maximalspeed then the supernatants were separated and storedat -20degC until analysis The amount of lipase secreted inthe medium was determined according to [28] withsome modifications Reaction mixtures contained (A)60 mg of p-nitrophenyl palmitate (Sigma Aldrich) in 20ml of isopropanol (Sigma Aldrich) (B) 1 g Triton X-100(Sigma Aldrich) and 02 g of Arabic gum (SigmaAldrich) in 200 ml TrisHCl 01 M (pH 75) To obtainthe mixture (C) add slowly 1 part of A solution to 9part of B solution under continuous agitation Add 9part of (C) solution to one part of the enzyme contain-ing solution (ie the supernatant) Lipase activities weremeasured by absorbance changes at 410 nm (ShimadzuUV 1601) and the amount of enzyme is proportional toOD according to the relation Um = 0667 times OD410 nm
(if OD410 nm lt 05)Quantification of interleukinHuman IL-1b were analysed by Western blot Indepen-dent transformants were cultured in shake flasks inminimal or rich medium and during the kinetics
samples were collected Cells were harvested (a culturevolume corresponding to 108 cells) by centrifugationOne volume of 2timesLaemmli buffer [29] was added to thesupernatants of said samples they were boiled for 3-5min and stored at -20degC until loading or loaded directlyon a polyacrylamide gel For the analysis of intracellularproduction crude extracts were prepared from 108 cellsby following the trichloroacetic acid protocol [30] andresuspending the final protein extract in 150 μl ofLaemmli buffer Samples were then loaded on standardpolyacrylamide gels (sodium dodecyl sulfate-polyacryla-mide gel electrophoresis final concentration of theseparating gel 15) after separation proteins wereblotted to nitrocellulose membranes and immunodeco-rated An anti-IL-1b rabbit polyclonal antibody (IL-1b(H-153) Santa Cruz Biotechnology USA cat no sc-7884 dilution 1200) was used for the interleukin immu-nodecoration An anti-rabbit IgG horseradish peroxi-dase-conjugated (Amersham Pharmacia Biotech UK catno NA934 dilution 110000) secondary antibody wasused For the interleukin detection as a positive controlthe human recombinant IL-1b from E coli (Roche catno 1 457 756) was always loaded in parallel on thepolyacrylamide gels The proteins were visualised usingthe ECL Western blotting system (Amersham Bios-ciences UK) according to the manufacturerrsquosinstructions
Additional file 1 Clustalw_ntd Nucleotidic sequence of the ZbGAS1gene and alignment with the homologue ScGAS1 (entry YMR307W)Grey coloring represent nucleotidic identity Alignment was performedwith ClustalW software httpwwwebiacukclustalw and graphicalediting was made using Jalview software [31]Click here for file[ httpwwwbiomedcentralcomcontentsupplementary1475-2859-9-7-S1PDF ]
AcknowledgementsSpecial thanks to Valeria Longo and Davide Zocco for technicalcontributions and to Stefania Brocca for providing the pGAPsLIP plasmidThis work was partially supported by FAR 2007 and 2008 (12-1-192) to DP
Author details1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy 2CPC Biotech Srl Naples Italy
Authorsrsquo contributionsPB and DP initiated and coordinated the project LD was responsible forZbGAS1 gene identification by PCR and its deletion LR analyzed the mutantphenotype with CFW and SDS LR and SP performed the batch cultivationanalyzed the secretory phenotype in null mutant and performed theexperiment with heterologous proteins All authors wrote the paper andapproved the final version of the manuscript
Competing interestsThe authors declare that they have no competing interests
Received 2 November 2009Accepted 26 January 2010 Published 26 January 2010
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 10 of 11
References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11

Morphological and physiological characterization of thenull mutantThe GAS1 gene inactivation leads to different peculiarphenotypes in S cerevisiae that were widely character-ized [10] We therefore performed a morphological andphysiological characterization of the obtained Z bailiiΔgas1 mutant in analogy to what reported in literaturefor the model yeast First some morphological traits ofthe Sc Δgas1 mutant are visible both by light and byfluorescence microscopy after Calcofluor White stainingIt was reported for S cerevisiae (both haploid anddiploid) mutants a rounded shape and higher cell dia-meter if compared to wild type cells especially in thestationary phase [10] We performed the analysis of theZbΔgas1 and wild type strains in parallel to the S cere-visiae BY4741 Δgas1 and wt strains Microscopy imagesare shown in Figure 3 It is possible to see the typicalrounded phenotype also for the ZbΔgas1 (panels C andG) Moreover like in S cerevisiae intracellular granulesand altered vacuoles are also visible (indicated byarrows panels C and G) The altered shape is alsoreflected by cellular volumes being higher in themutants as assessed by coulter counter measurementsin cell populations growing in minimal or in rich med-ium like for the model yeast differences are more pro-nounced in stationary phase were the mutant cellvolume is 16 fold larger in respect to the wild typecells (data not shown) Another feature described forthe ScΔgas1 mutant is the appearance of large cellularaggregates still permanent after repeated sonicationcycles [10] This feature is also conserved in the Z bai-lii mutant as it is visible from the microscopy imagesof sonicated cells (Figure 3 panels C D G and H) Themorphological alterations described are likely a conse-quence of the altered structure and composition of themutant cell wall it was reported for S cerevisiae a
lowered glucans content together with a markedincrease in the chitin and mannoproteins content [15]Moreover an altered distribution of the cell wall com-ponents in the mother and daughter cell have beenreported [10] Our analyses confirmed these alterationsalso for the Z bailii mutant overall Calcofluor stainingin fact was more intense in Δgas1 than in wild typestrain (Figure 3 panels D and H) indicating a higherchitin content in the mutant The chitin seems to bedelocalized throughout the entire cell wall in themutant while in the wild type it is more concentratedat the budding ring and at the bud scars Moreover inthe mutant the cellular buds are also Calcofluor-stainedin contrast to the wild type It was proposed that theincrease in chitin levels and cross-links with b-16-glu-cosyl mannoproteins may be the effect of the activationof a compensatory mechanism in mutants characterizedby defects in the cell wall architecture like the Δgas1mutants The aim of this response would be to preventexcessive loss of mannoproteins as a consequence ofthe inability to cross-link the b-16-glucosyl mannopro-teins and the b-13-glucans a process in which Gas1pplays a central role [15] It was also hypothesized thatthe loss of Gas1p may cause a weakening of the cellwall of the emerging bud The yeast cell could inresponse to this weakening target the accumulation ofchitin at the bud surface in order to protect it fromlyses [16] Alteration of the processes directly or indir-ectly linked to budding leads to the appearance of arelative high fraction in the cell population of multi-budded cells (the so-called ldquoMickey Mouserdquo phenotype)(figure 3) A similar phenotype was also visible inZbΔgas1 mutants (Figure 3)The altered cell wall structure renders the deleted
bakersrsquo yeast strain more sensitive to sodium dodecylsulphate (SDS) and to Calcofluor White (CFW) as
Figure 2 Deletion of ZbGAS1 (a) Schematic representation of the ZbGAS1 deletion cassettes Restriction enzymes used to obtain the constructsand PCR primers used to verify recombination are indicated light grey bars represents ZbGAS1 sequence (b) Control PCR performed to verifythe deletion of both the allelic copies of ZbGAS1 Two independent mutant clones (A and B) were tested In both clones it is possible to see thepresence of bands corresponding to the two deletion cassettes and the absence of the band corresponding to the wild type gene copy
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 4 of 11
reported in literature for the W303 background [17]Here we confirm by spot assays the sensitivity to bothagents of the ScΔgas1 mutant background BY4741 (Fig-ure 3 upper right panels) and as well of the ZbΔgas1mutant (Figure 3 lower right panels)Finally to verify the functionality of the GAS1 homo-
logue of Z bailii in S cerevisiae and the similarity of itsbiological role the ZbGAS1 gene was expressed into theGAS1-deficient S cerevisiae strain resulting in wild typephenotype restoration either in respect to the morpholo-gical traits as well as for the chitin distribution (data not
shown) Moreover to verify the functionality of the Scerevisiae GAS1 in Z bailii the ScGAS1 gene wasexpressed into the GAS1-deficient Z bailii strain alsoin this case restoring the native phenotype (data notshown)Overall morphological and physiological data strongly
indicate that the ZbGAS1 gene may have a similar rolewith respect to the ScGAS1 homologous in the assess-ment of the cell wall structure and hence their inactiva-tion might have similar effects in the two yeasts inrespect to secretion abilities
Figure 3 Morphology of Z bailii and S cerevisiae wild type and gas1 null mutant strains and their sensitivity to Calcofluor White andSDS Cellular morphology From A to H dicroic and fluorescence microscope images of Z bailii and S cerevisiae wild type and gas1 null mutantstrains Cells were grown in minimal medium and harvested in stationary phase of growth Arrows indicate granules and enlarged vacuoles inmutant cells (panels C and G) Calcofluor White and SDS sensitivity of S cerevisiae (BY4741) and Z bailii (Zygo) gas1 null mutants 4 μl of culturesof the different strains (BY4741 BY4741 Δgas1 Zygo and Zygo Δgas1) were spotted starting with OD (660 nm) = 1 and then in 3 serial solution110 (234) on rich solid medium (YPD) and on the same medium added with 125 μg ml-1 of CFW or 50 μg ml-1 SDS Pictures were capturedafter 5 days of incubation at 30degC
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 5 of 11
Supersecretive phenotype of Δgas1 strainsAs already mentioned (see Introduction) the mostinteresting phenotype of Δgas1 mutants reported in lit-erature is the supersecretive property Besides theincreased secretion of heterologous proteins whichappears to be dependent on the individual protein [12]the levels of total secreted proteins in late-stationaryphase cultures of S cerevisiae mutants were two foldhigher with respect to the wild-type [7] We thereforestarted our evaluation of secretive properties of the Zbailii mutant in the perspective of a possible applica-tion in industrial processes by determining the amountof total proteins released in the growth medium duringbatch cultivation with respect to the wild type strain Zbailii and S cerevisiae wild type and Δgas1 respectivestrains were grown in shake flasks in minimal YNB-based medium containing 5 glucose Cells were har-vested at different times of cultivation and the fermen-tation broths were assayed At least triplicateexperiments were performed on three independenttransformants and Figure 4 shows the mean dataobtained reported as mg of total proteins secreted perlitre of the culture broth normalised for OD valuesThe values refers to 72 h of cultivation but it was pos-sible to observe a trend leading to this result starting atthe end of exponential phase and becoming more pro-nounced in stationary phase of growth
The data clearly underline the overall higher secretivecapacity of Z bailii if compared to S cerevisiae inaccordance to heterologous proteins secretion dataalready reported [3] Moreover they indicate a signifi-cant improvement in the secretive capability of the Zbailii mutant being nearly 80 more than the wild typein terms of total endogenous protein similarly to the Scerevisiae mutantOverexpression of CRL lipase XPR2 protease and hIL-1bin Z bailii Δgas1 strainsWith the intention to evaluate the potential effects ofcell wall engineering and to test the secretive perfor-mance in the presence of an heterologous protein threemodel proteins were tested a fungal lipase from Can-dida rugosa (CRL1) a fungal protease from Yarrowialipolytica (XPR2) and the human interleukin 1b (hIL-1b)Four clones of Z bailii Δgas1 [pZLN022XPR2] and
four clones of the parental strain Z bailii[pZLN022XPR2] together with control clones harbour-ing the respective empty plasmid were pre-cultivated for16 hours in shake flasks using YPD medium and thentransferred in shake flasks in minimal YNB-based med-ium containing 5 glucose The amount of secretedrecombinant XPR2 in culture supernatants was tested interms of enzymatic activity after 72 96 120 and 148 hwith the Azocoll method (see methods for detail) The
Figure 4 Total proteins released in the culture medium Total proteins released in the culture medium of batch cultures of the indicatedwild type and Δgas1 strains of S cerevisiae and Z bailii Yeasts were grown in shake flasks in minimal YNB-based medium with 5 glucose Cellswere inoculated at 01 OD and harvested after 72 hours of cultivation Amount of released proteins (in mg l-1 of culture broth) were normalizedfor OD values
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 6 of 11
ZbΔgas1 transformants showed a relevant increase inspecific activity levels since 72 h (+ 25 at 72 h morethan two-fold at 148 h see Figure 5a) in respect to wildtype transformantsSimilarly three clones of Z bailii Δgas1
[pZLN022LIP1] and three clones of the parental strainZ bailii [pZLN022LIP1] together with control clonesharbouring the respective empty plasmid were pre-culti-vated for 16 hours in shake flasks using YPD mediumand then transferred in shake flasks in minimal YNB-based medium containing 5 glucose The amount ofsecreted recombinant LIP1 in culture supernatants wastested in terms of enzymatic activity after 24 48 72 and120 h using the hydrolysis of p-nitrophenylestersmethod (see methods for detail) Also for this secondheterologous product the Δgas1 transformants showed arelevant increase in specific activity levels since 24 h
(+35 at 24 h more than two-fold at 120 h see Figure5b) in respect to wild type transformantsFinally three clones of Z bailii Δgas1 [pZ5KlIL-1b]
and three clones of the parental strain Z bailii[pZ5KlIL-1b] together with control clones harbouringthe respective empty plasmid were tested for secretionlevel of the human interleukin since the codingsequence was functionally linked to the signal peptide ofthe K1 killer toxin from K lactis The independenttransformants were pre-cultivated and then cultured inshake flasks containing rich medium (YPD 5 wv glu-cose) Samples were collected at the indicated times (seeFigure 5c) cells were harvested and both the superna-tants and the cell protein extracts of said samples wereloaded on a polyacrylamide gel The data obtained byWestern blot (see Figure 5c) performed on the superna-tant showed a well represented band corresponding to
Figure 5 Secretion levels of yeast protease lipase and rhIL-1b in Z bailii and ZbΔgas1 (a) Secretion levels of protease in Z bailii andZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutant transformed with controlplasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with XPR2 gene (b) Secretion levels oflipase in Z bailii and ZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutanttransformed with control plasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with LIP1 gene(c) Western blots for rhIL-1b of protein samples from supernatants and cell extracts of Z bailii and ZbΔgas1 transformants grown in YPD mediawith 5 glucose inoculated at OD660 = 01 From left to right (A) rhIL-1b (B) negative control Z bailii[pZ5] (C) (D) (E) (I) (L) (M) Z bailii[pZ5KlrhIL-1b] (F) (G) (H) (N) (O) (P) ZbΔgas1 [pZ5KlrhIL-1b] Time and sample types are indicated in the picture
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 7 of 11
the recombinant protein for both Z bailii and Z bailiiΔgas1 samples but differently from the case of recombi-nant fungal enzymes no relevant differences in favourof the deleted yeast are appreciable Remarkably the sig-nal corresponding to the protein retained intracellularlyis more intense in gas1 null mutant than the signalobtained from Z bailii parental strain indicating anincomplete secretion of this protein
DiscussionsCell wall engineering can improve the release of het-erologous proteins from yeasts [18] Generally speak-ing GAS1 which mediates the crosslinking of cell wallcomponents is directly involved in defining the perme-ability of the cell Cloning of the unknown Z bailiiGAS1 homologue by PCR was only possible startingfrom highly conserved internal regions identified uponalignment of the GAS1 homologues of three yeast spe-cies The 5rsquo and 3rsquo ends were subsequently amplifiedusing a Genome Walker library With this approachthe full-length Z bailii GAS1 homologue could beamplified and sequenced The cloned gene wasexpressed in the GAS1-deficient S cerevisiae strain andreversion of the phenotype proved that this homologueis not only structurally but also functionally similar tothe S cerevisiae gene Moreover also the ScGAS1 genecomplemented the Z bailii mutant Disruption ofGAS1 in Z bailii revealed its influence on the cell wallmorphology The GAS1 null mutants of Z bailiishowed the same phenotype reported for S cerevisiae[10] with enlarged cells often showing granular cyto-plasm an alterated chitin distribution two buds(Mickey Mouse-like appearance) and big vacuoles Thevacuole is known to be a dynamic organelle whosemorphology is highly responsive to different extracellu-lar and intracellular stimulistressors [19] In the GAS1null mutant it can be therefore speculated that thegeneration of big vacuoles is related to stressful condi-tions as already suggested by the altered chitin distri-bution being a possible defence mechanism againstlysis or related to a generalized response to sickness ofthe cells [10]Finally also the sensitivity to SDS and CFW
appeared very similar to what observed in the S cerevi-siae mutant A comparison of the different GAS1 nullphenotype mutants and the functionality of the Z bai-lii GAS1 homologue in S cerevisiae suggest a similarityof the cell wall structure of Z bailii compared with thecell wall structure of S cerevisiae Considering thesecretory pathway of a secreted protein it was specu-lated whether the protein becomes retarded in the cellwall after release from the plasma membrane The dis-ruption of GAS1 in Z bailii first of all confirmed ageneral phenomenon of protein release in respect to
the wild type strain as demonstrated by the higheraccumulation of total protein in the growing mediumof the deleted strain when compared to the parentalstrain Moreover such as compromised cell wall per-meability in the production strains for XPR2 and LIP1does lead to a clear improvement in the secretion ofthe respective proteins In the case of h IL-1b theenhancement of product secretion was not so evidentand difficult to interpret in respect to the higher accu-mulation of intracellular product It could be specu-lated that this can be related to the nature or to thesource of the protein but too few information areavailable at the moment to further comment in thisdirection What seems to be true is unfortunately whatwe very often experience for recombinant protein pro-duction which is the protein and protein-host depen-dence In fact also in P pastoris two out of threeheterologous proteins resulted more secreted in themutant as for Z bailii and when we tested the sameprotein in particular the IL-1b secretion in the S cere-visiae Δgas1 mutant (data not shown) we did not reg-ister any improvement
MethodsYeast strains media and transformationThe S cerevisiae strains used in this study were theauxotrophic haploid BY4741 (Mat a his3Δ1 leu2Δ0met5Δ0 ura3Δ0) and the relative null mutant in GAS1sequence called in this study BY4741 Δgas1 (Mat ahis3Δ1 leu2Δ0 met5Δ0 ura3Δ0 YMR307kanMX4)Both strains are commercially available from Euroscarfcollection httpwebuni-frankfurtdefb15mikroeuro-scarfcol_indexhtml The Z bailii wild type strain isATCC60483 Yeast cultures were grown in YP medium(10 g l-1 yeast extract 20 g l-1 peptone) with 2 or 5wv glucose (D) or fructose (F) as carbon sourceWhere needed NAT (80 mg l-1) or G418 (500 mg l-1
for S cerevisiae and 200 mg l-1 for Z bailii) or hygro-micine (100 mg l-1 for Z bailii) were added to themedia Alternatively yeast cultures were grown inminimal synthetic medium (067 w v-1 YNB Biolifewithout amino acids) with 2 or 5 w v-1 glucose ascarbon source When required supplements such ashistidine leucine methionine and uracil were added toa final concentration of 50 mg l-1 Media for plateswere solidified by addition of agar (Biolife Milan Italy)to 15 w v-1 Shake flask experiments were carriedout with 50 ml of medium in 250 ml shake flasks on ashaker at 30degC with 160 rpm For sensitivity tests inPetri dishes 125 μg ml-1 of CFW or 50 μg ml-1 SDSwere added to solid media Yeast transformation wasperformed using the Lithium AcetatessDNA methodas described [20] and slightly adapted for Z bailii asdescribed [3]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 8 of 11
Genes amplifications deletion cassettes and expressionplasmid constructionPCR amplifications were made from yeasts genomicDNA prepared as described in [21] Amplifications wereperformed by the Pwo DNA Polymerase (Roche)The amplification of the ZbGAS1 gene was as follows
the primers GAS1int_fw 5rsquo-AGT TGT TCC AGA GATATT CCA TAC CTC AA-3rsquo and GAS1int_rev 5rsquo-GCAGAA CCG CTG AAG CTA CAG T-3rsquo designed oninternal conserved region of the available sequencesfrom other yeasts were firstly used to amplify an internalregion of 1137 bp of the Z bailii gene with the follow-ing program 94degC 5 min (94degC 15 s 57degC 30 s 72degC 1min) times 25 72degC 7 min 4degC infin After that the designedprimers ZbGAS1int_fw 5rsquo-GGA CGA TAA GGT TGACGA AGA-3rsquo and ZbGAS1int_rev 5rsquo-TGC TGT GGTCCA AAG TTG TG-3rsquo were used for the amplification(program 7 cycles 94degC 2 min 72degC 3 min 32 cycles94degC 2 min 67degC 3 min 67degC 4 min 4degC infin) of theterminal ends of ZbGAS1 utilizing a Z bailii GenomeWalker library constructed using the Universal GenomeWalker kit (Clontech) following the manufacturersinstructions From the obtained and reconstructedsequence the final primers were established and used forthe ZbGAS1 1710 bp full length coding sequence ampli-fication ZbGAS1_fw 5rsquo-ACT AAT GTT ATT CCAGGC GTT TTC G-3rsquo and ZbGAS1real_rev 5rsquo-AAATCA AGC CAA AGC AAA TCC AGC A-3rsquo (program94degC 5 min (94degC 20 s 58degC 30 s 72degC 2 min) times 30 72degC 10 min 4degC infin) The DNA sequence of ScGAS1 wasPCR amplified with the following primers ScGAS1fw5rsquo-ACA ATG TTG TTT AAA TCC CTT TCA AAGTTA GCA A-3rsquo and ScGAS1rev 5rsquo-TTT TTA AACCAA AGC AAA ACC GAC ACC AG-3rsquo (program 94degC5 min (94degC 20 s 605degC 30 s 72degC 2 min) times 30 72degC10 min 4degC infin) and the DNA sequence of YlXPR2 wasPCR amplified with the following primers XPR2_fwd5rsquo-ACA ATG AAG CTC GCT ACC GCC TTT A-3rsquoand XPR2_rev 5rsquo-TGC CTA AAT GCC AAC ACCGTT GTA G-3rsquo (program 94degC 4 min (94degC 15 s 625degC 30 s 72degC 2 min) times 30 72degC 7 min 4degC infin) All theunique fragments obtained were sub-cloned in the vec-tor pSTblue-1 utilising the Perfectly Bluntreg Cloning Kit(Novagene) and sequenced The resulting respectiveplasmids utilised in this work were named as followspSTbZbGAS1int pSTbZbGAS1 pSTbScGAS1 andpSTbYlXPR2For the construction of the ZbGAS1 deletion cassettes
the KanR or the hphR cassette derived from plasmidpFA6-KanMX4 [22] and pAG26 [23] and conferringresistance to G418 and hygromycin respectively werebluntended excised and inserted in the pSTbZbGAS1intopened BstEII BglII bluntended and dephosphorylatedresulting in the plasmids pSTbZbGAS1ΔK and
pSTbZbGAS1ΔH The resistance cassettes resulted inboth cases counter-clockwise inserted The Z bailii wildtype cells were transformed with the respective deletioncassettes obtained by cutting the two described plasmidsin the AccI and SnaBI sites The effective deletion wasPCR checked by using the following couples of primersZbG1int2_fw 5rsquo- TGC AGA AGT TAC AGA CCAATG TTG T -3rsquo and and ZbG1int2_rev 5rsquo-AAG ATAGAG AAG TGC TCT TGG CA -3rsquo (see Figure 2) orZbG1int2_fw and Hph_fw 5rsquo- ATA TGA AAA AGCCTG AAC TCA CCG AC -3rsquo or Kan_fw 5rsquo- ATG GGTAAG GAA AAG ACT CAC GTT -3rsquo (not shown) inorder to verify the presence of the cassettes in thedesired locusFor complementing the yeasts GAS1 deletion pZ5
(-Nco)ScGAS1 and pYX022ZbGAS1 were constructedpZ5(-Nco)ScGAS1 was obtained from pZ5(-Nco) openedSacIEcoRV with the insertion of ScGAS1 frompSTbScGAS1 cut with SacISnaBI The pZ5(-Nco)derives from pZ3 [3] where at the KpnI site the KanR
cassette was substituted from the NATR cassetteobtained from vector pAG25 [23] by cutting PvuIISacIblunt (obtaining the pZ5 plasmid) finally this plasmidwas EcoRIBamHI cut and bluntended and reclosedpYX022ZbGAS1 was obtained with the insertion ofZbGAS1 from pSTbZbGAS1 into pYX022 (RampD Sys-tems Wiesbaden Germany) both EcoRI cutFor heterologous protein expression the plasmids
pZLN022XPR2 pZLN022LIP1 and pZ5KlhIL-1b wereconstructed The first two integrative plasmids derivesfrom pZLN022 (manuscript in preparation) obtainedfrom pYX022 opened DraIII bluntSpeI blunt (toexclude HIS3) with the insertion of ZbLEU2 cut frompSTbZbLEU2 with EcoRI blunt in KpnI blunt site theNATR cassette was inserted obtained from vectorpAG25 [23] by cutting PvuIISacIblunt ThepSTbZbLEU2 was obtained by sub-cloning ZbLEU2amplified by PCR from Z bailii genomic DNA (manu-script in preparation) in pSTBlue (Novagen) cut byEcoRV For pZLN022XPR2 construction the pZLN022was AatIINheI cut and the XPR2 from pSTbXPR2 wasinserted For pZLN022LIP1 construction the pZLN022was AatII bluntNheI blunt opened and the LIP1 wasinserted by BamHI bluntNsiI blunt excision from thepGAPsLIP [24] For rh interleukin the pZ5KlhIL-1b wasbuilt it was obtained from pZ5 opened with EcoRI-blunt with the insertion of hIL-1b in frame with Kl sig-nal sequence from vector pKSSPI3 [25] cut EcoRIbluntXbaI bluntDNA manipulations transformation and cultivation of
Escherichia coli (DH5aFrsquo (80dlacZΔM15 Δ (lacZYA-argF) U169 deo rec1 end1 sup44 l THI-1 gyrA96relA1) and Novablue Competent Cells (Novagene) wereperformed following standard protocols [26] All the
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 9 of 11
restriction and modification enzymes used were fromNew England Biolabs (Hitchin Herts UK) or fromRoche Diagnostics (Mannheim Germany)Calcofluor staining and fluorescence microscopyCells were harvested washed in PBS and resuspended in1 mg ml-1 Calcofluor White solution incubated 10-30min at room temperature washed again twice in PBSFluorescence microscope images were taken by a NikonEclipse 90i microscope using the Metamorph softwareversion 22 (Nikon) using emission filter UV-1AQuantification of total released proteinsCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were etha-nol-precipitated The amount of proteins was deter-mined by the dye-binding method of Bradford (QuickStart Bradford Dye Reagent - BIORAD) using BSA asstandard All the data were normalized by OD after ver-ification of the linear correlation between OD and dryweight as a measure of biomassQuantification of secreted proteaseCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were sepa-rated and stored at -20degC until analysis The amount ofprotease secreted in the medium was determined by theAzocoll assay (Azocoll - Calbiochem) using commercialprotease as positive control Azocoll assays were per-formed according to [27] with some modificationsReaction mixtures contained 15 mg ml-1 Azocoll 50mM TrisHCl (pH 76) 150 mM NaCl and 5 mMCaCl2 Proteolytic activities were measured by absor-bance changes at 490 nm (Shimadzu UV 1601) afterincubation for 18 h at 30degCQuantification of secreted lipaseCultures were harvested and centrifuged at maximalspeed then the supernatants were separated and storedat -20degC until analysis The amount of lipase secreted inthe medium was determined according to [28] withsome modifications Reaction mixtures contained (A)60 mg of p-nitrophenyl palmitate (Sigma Aldrich) in 20ml of isopropanol (Sigma Aldrich) (B) 1 g Triton X-100(Sigma Aldrich) and 02 g of Arabic gum (SigmaAldrich) in 200 ml TrisHCl 01 M (pH 75) To obtainthe mixture (C) add slowly 1 part of A solution to 9part of B solution under continuous agitation Add 9part of (C) solution to one part of the enzyme contain-ing solution (ie the supernatant) Lipase activities weremeasured by absorbance changes at 410 nm (ShimadzuUV 1601) and the amount of enzyme is proportional toOD according to the relation Um = 0667 times OD410 nm
(if OD410 nm lt 05)Quantification of interleukinHuman IL-1b were analysed by Western blot Indepen-dent transformants were cultured in shake flasks inminimal or rich medium and during the kinetics
samples were collected Cells were harvested (a culturevolume corresponding to 108 cells) by centrifugationOne volume of 2timesLaemmli buffer [29] was added to thesupernatants of said samples they were boiled for 3-5min and stored at -20degC until loading or loaded directlyon a polyacrylamide gel For the analysis of intracellularproduction crude extracts were prepared from 108 cellsby following the trichloroacetic acid protocol [30] andresuspending the final protein extract in 150 μl ofLaemmli buffer Samples were then loaded on standardpolyacrylamide gels (sodium dodecyl sulfate-polyacryla-mide gel electrophoresis final concentration of theseparating gel 15) after separation proteins wereblotted to nitrocellulose membranes and immunodeco-rated An anti-IL-1b rabbit polyclonal antibody (IL-1b(H-153) Santa Cruz Biotechnology USA cat no sc-7884 dilution 1200) was used for the interleukin immu-nodecoration An anti-rabbit IgG horseradish peroxi-dase-conjugated (Amersham Pharmacia Biotech UK catno NA934 dilution 110000) secondary antibody wasused For the interleukin detection as a positive controlthe human recombinant IL-1b from E coli (Roche catno 1 457 756) was always loaded in parallel on thepolyacrylamide gels The proteins were visualised usingthe ECL Western blotting system (Amersham Bios-ciences UK) according to the manufacturerrsquosinstructions
Additional file 1 Clustalw_ntd Nucleotidic sequence of the ZbGAS1gene and alignment with the homologue ScGAS1 (entry YMR307W)Grey coloring represent nucleotidic identity Alignment was performedwith ClustalW software httpwwwebiacukclustalw and graphicalediting was made using Jalview software [31]Click here for file[ httpwwwbiomedcentralcomcontentsupplementary1475-2859-9-7-S1PDF ]
AcknowledgementsSpecial thanks to Valeria Longo and Davide Zocco for technicalcontributions and to Stefania Brocca for providing the pGAPsLIP plasmidThis work was partially supported by FAR 2007 and 2008 (12-1-192) to DP
Author details1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy 2CPC Biotech Srl Naples Italy
Authorsrsquo contributionsPB and DP initiated and coordinated the project LD was responsible forZbGAS1 gene identification by PCR and its deletion LR analyzed the mutantphenotype with CFW and SDS LR and SP performed the batch cultivationanalyzed the secretory phenotype in null mutant and performed theexperiment with heterologous proteins All authors wrote the paper andapproved the final version of the manuscript
Competing interestsThe authors declare that they have no competing interests
Received 2 November 2009Accepted 26 January 2010 Published 26 January 2010
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 10 of 11
References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11
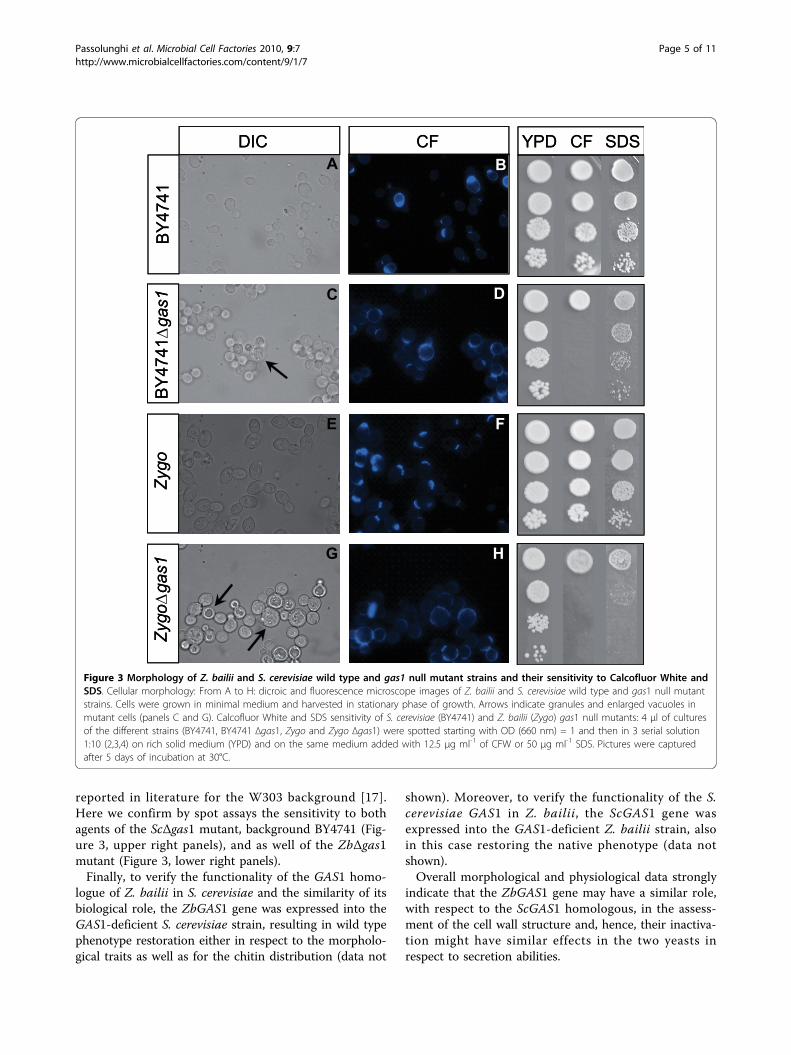
reported in literature for the W303 background [17]Here we confirm by spot assays the sensitivity to bothagents of the ScΔgas1 mutant background BY4741 (Fig-ure 3 upper right panels) and as well of the ZbΔgas1mutant (Figure 3 lower right panels)Finally to verify the functionality of the GAS1 homo-
logue of Z bailii in S cerevisiae and the similarity of itsbiological role the ZbGAS1 gene was expressed into theGAS1-deficient S cerevisiae strain resulting in wild typephenotype restoration either in respect to the morpholo-gical traits as well as for the chitin distribution (data not
shown) Moreover to verify the functionality of the Scerevisiae GAS1 in Z bailii the ScGAS1 gene wasexpressed into the GAS1-deficient Z bailii strain alsoin this case restoring the native phenotype (data notshown)Overall morphological and physiological data strongly
indicate that the ZbGAS1 gene may have a similar rolewith respect to the ScGAS1 homologous in the assess-ment of the cell wall structure and hence their inactiva-tion might have similar effects in the two yeasts inrespect to secretion abilities
Figure 3 Morphology of Z bailii and S cerevisiae wild type and gas1 null mutant strains and their sensitivity to Calcofluor White andSDS Cellular morphology From A to H dicroic and fluorescence microscope images of Z bailii and S cerevisiae wild type and gas1 null mutantstrains Cells were grown in minimal medium and harvested in stationary phase of growth Arrows indicate granules and enlarged vacuoles inmutant cells (panels C and G) Calcofluor White and SDS sensitivity of S cerevisiae (BY4741) and Z bailii (Zygo) gas1 null mutants 4 μl of culturesof the different strains (BY4741 BY4741 Δgas1 Zygo and Zygo Δgas1) were spotted starting with OD (660 nm) = 1 and then in 3 serial solution110 (234) on rich solid medium (YPD) and on the same medium added with 125 μg ml-1 of CFW or 50 μg ml-1 SDS Pictures were capturedafter 5 days of incubation at 30degC
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 5 of 11
Supersecretive phenotype of Δgas1 strainsAs already mentioned (see Introduction) the mostinteresting phenotype of Δgas1 mutants reported in lit-erature is the supersecretive property Besides theincreased secretion of heterologous proteins whichappears to be dependent on the individual protein [12]the levels of total secreted proteins in late-stationaryphase cultures of S cerevisiae mutants were two foldhigher with respect to the wild-type [7] We thereforestarted our evaluation of secretive properties of the Zbailii mutant in the perspective of a possible applica-tion in industrial processes by determining the amountof total proteins released in the growth medium duringbatch cultivation with respect to the wild type strain Zbailii and S cerevisiae wild type and Δgas1 respectivestrains were grown in shake flasks in minimal YNB-based medium containing 5 glucose Cells were har-vested at different times of cultivation and the fermen-tation broths were assayed At least triplicateexperiments were performed on three independenttransformants and Figure 4 shows the mean dataobtained reported as mg of total proteins secreted perlitre of the culture broth normalised for OD valuesThe values refers to 72 h of cultivation but it was pos-sible to observe a trend leading to this result starting atthe end of exponential phase and becoming more pro-nounced in stationary phase of growth
The data clearly underline the overall higher secretivecapacity of Z bailii if compared to S cerevisiae inaccordance to heterologous proteins secretion dataalready reported [3] Moreover they indicate a signifi-cant improvement in the secretive capability of the Zbailii mutant being nearly 80 more than the wild typein terms of total endogenous protein similarly to the Scerevisiae mutantOverexpression of CRL lipase XPR2 protease and hIL-1bin Z bailii Δgas1 strainsWith the intention to evaluate the potential effects ofcell wall engineering and to test the secretive perfor-mance in the presence of an heterologous protein threemodel proteins were tested a fungal lipase from Can-dida rugosa (CRL1) a fungal protease from Yarrowialipolytica (XPR2) and the human interleukin 1b (hIL-1b)Four clones of Z bailii Δgas1 [pZLN022XPR2] and
four clones of the parental strain Z bailii[pZLN022XPR2] together with control clones harbour-ing the respective empty plasmid were pre-cultivated for16 hours in shake flasks using YPD medium and thentransferred in shake flasks in minimal YNB-based med-ium containing 5 glucose The amount of secretedrecombinant XPR2 in culture supernatants was tested interms of enzymatic activity after 72 96 120 and 148 hwith the Azocoll method (see methods for detail) The
Figure 4 Total proteins released in the culture medium Total proteins released in the culture medium of batch cultures of the indicatedwild type and Δgas1 strains of S cerevisiae and Z bailii Yeasts were grown in shake flasks in minimal YNB-based medium with 5 glucose Cellswere inoculated at 01 OD and harvested after 72 hours of cultivation Amount of released proteins (in mg l-1 of culture broth) were normalizedfor OD values
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 6 of 11
ZbΔgas1 transformants showed a relevant increase inspecific activity levels since 72 h (+ 25 at 72 h morethan two-fold at 148 h see Figure 5a) in respect to wildtype transformantsSimilarly three clones of Z bailii Δgas1
[pZLN022LIP1] and three clones of the parental strainZ bailii [pZLN022LIP1] together with control clonesharbouring the respective empty plasmid were pre-culti-vated for 16 hours in shake flasks using YPD mediumand then transferred in shake flasks in minimal YNB-based medium containing 5 glucose The amount ofsecreted recombinant LIP1 in culture supernatants wastested in terms of enzymatic activity after 24 48 72 and120 h using the hydrolysis of p-nitrophenylestersmethod (see methods for detail) Also for this secondheterologous product the Δgas1 transformants showed arelevant increase in specific activity levels since 24 h
(+35 at 24 h more than two-fold at 120 h see Figure5b) in respect to wild type transformantsFinally three clones of Z bailii Δgas1 [pZ5KlIL-1b]
and three clones of the parental strain Z bailii[pZ5KlIL-1b] together with control clones harbouringthe respective empty plasmid were tested for secretionlevel of the human interleukin since the codingsequence was functionally linked to the signal peptide ofthe K1 killer toxin from K lactis The independenttransformants were pre-cultivated and then cultured inshake flasks containing rich medium (YPD 5 wv glu-cose) Samples were collected at the indicated times (seeFigure 5c) cells were harvested and both the superna-tants and the cell protein extracts of said samples wereloaded on a polyacrylamide gel The data obtained byWestern blot (see Figure 5c) performed on the superna-tant showed a well represented band corresponding to
Figure 5 Secretion levels of yeast protease lipase and rhIL-1b in Z bailii and ZbΔgas1 (a) Secretion levels of protease in Z bailii andZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutant transformed with controlplasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with XPR2 gene (b) Secretion levels oflipase in Z bailii and ZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutanttransformed with control plasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with LIP1 gene(c) Western blots for rhIL-1b of protein samples from supernatants and cell extracts of Z bailii and ZbΔgas1 transformants grown in YPD mediawith 5 glucose inoculated at OD660 = 01 From left to right (A) rhIL-1b (B) negative control Z bailii[pZ5] (C) (D) (E) (I) (L) (M) Z bailii[pZ5KlrhIL-1b] (F) (G) (H) (N) (O) (P) ZbΔgas1 [pZ5KlrhIL-1b] Time and sample types are indicated in the picture
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 7 of 11
the recombinant protein for both Z bailii and Z bailiiΔgas1 samples but differently from the case of recombi-nant fungal enzymes no relevant differences in favourof the deleted yeast are appreciable Remarkably the sig-nal corresponding to the protein retained intracellularlyis more intense in gas1 null mutant than the signalobtained from Z bailii parental strain indicating anincomplete secretion of this protein
DiscussionsCell wall engineering can improve the release of het-erologous proteins from yeasts [18] Generally speak-ing GAS1 which mediates the crosslinking of cell wallcomponents is directly involved in defining the perme-ability of the cell Cloning of the unknown Z bailiiGAS1 homologue by PCR was only possible startingfrom highly conserved internal regions identified uponalignment of the GAS1 homologues of three yeast spe-cies The 5rsquo and 3rsquo ends were subsequently amplifiedusing a Genome Walker library With this approachthe full-length Z bailii GAS1 homologue could beamplified and sequenced The cloned gene wasexpressed in the GAS1-deficient S cerevisiae strain andreversion of the phenotype proved that this homologueis not only structurally but also functionally similar tothe S cerevisiae gene Moreover also the ScGAS1 genecomplemented the Z bailii mutant Disruption ofGAS1 in Z bailii revealed its influence on the cell wallmorphology The GAS1 null mutants of Z bailiishowed the same phenotype reported for S cerevisiae[10] with enlarged cells often showing granular cyto-plasm an alterated chitin distribution two buds(Mickey Mouse-like appearance) and big vacuoles Thevacuole is known to be a dynamic organelle whosemorphology is highly responsive to different extracellu-lar and intracellular stimulistressors [19] In the GAS1null mutant it can be therefore speculated that thegeneration of big vacuoles is related to stressful condi-tions as already suggested by the altered chitin distri-bution being a possible defence mechanism againstlysis or related to a generalized response to sickness ofthe cells [10]Finally also the sensitivity to SDS and CFW
appeared very similar to what observed in the S cerevi-siae mutant A comparison of the different GAS1 nullphenotype mutants and the functionality of the Z bai-lii GAS1 homologue in S cerevisiae suggest a similarityof the cell wall structure of Z bailii compared with thecell wall structure of S cerevisiae Considering thesecretory pathway of a secreted protein it was specu-lated whether the protein becomes retarded in the cellwall after release from the plasma membrane The dis-ruption of GAS1 in Z bailii first of all confirmed ageneral phenomenon of protein release in respect to
the wild type strain as demonstrated by the higheraccumulation of total protein in the growing mediumof the deleted strain when compared to the parentalstrain Moreover such as compromised cell wall per-meability in the production strains for XPR2 and LIP1does lead to a clear improvement in the secretion ofthe respective proteins In the case of h IL-1b theenhancement of product secretion was not so evidentand difficult to interpret in respect to the higher accu-mulation of intracellular product It could be specu-lated that this can be related to the nature or to thesource of the protein but too few information areavailable at the moment to further comment in thisdirection What seems to be true is unfortunately whatwe very often experience for recombinant protein pro-duction which is the protein and protein-host depen-dence In fact also in P pastoris two out of threeheterologous proteins resulted more secreted in themutant as for Z bailii and when we tested the sameprotein in particular the IL-1b secretion in the S cere-visiae Δgas1 mutant (data not shown) we did not reg-ister any improvement
MethodsYeast strains media and transformationThe S cerevisiae strains used in this study were theauxotrophic haploid BY4741 (Mat a his3Δ1 leu2Δ0met5Δ0 ura3Δ0) and the relative null mutant in GAS1sequence called in this study BY4741 Δgas1 (Mat ahis3Δ1 leu2Δ0 met5Δ0 ura3Δ0 YMR307kanMX4)Both strains are commercially available from Euroscarfcollection httpwebuni-frankfurtdefb15mikroeuro-scarfcol_indexhtml The Z bailii wild type strain isATCC60483 Yeast cultures were grown in YP medium(10 g l-1 yeast extract 20 g l-1 peptone) with 2 or 5wv glucose (D) or fructose (F) as carbon sourceWhere needed NAT (80 mg l-1) or G418 (500 mg l-1
for S cerevisiae and 200 mg l-1 for Z bailii) or hygro-micine (100 mg l-1 for Z bailii) were added to themedia Alternatively yeast cultures were grown inminimal synthetic medium (067 w v-1 YNB Biolifewithout amino acids) with 2 or 5 w v-1 glucose ascarbon source When required supplements such ashistidine leucine methionine and uracil were added toa final concentration of 50 mg l-1 Media for plateswere solidified by addition of agar (Biolife Milan Italy)to 15 w v-1 Shake flask experiments were carriedout with 50 ml of medium in 250 ml shake flasks on ashaker at 30degC with 160 rpm For sensitivity tests inPetri dishes 125 μg ml-1 of CFW or 50 μg ml-1 SDSwere added to solid media Yeast transformation wasperformed using the Lithium AcetatessDNA methodas described [20] and slightly adapted for Z bailii asdescribed [3]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 8 of 11
Genes amplifications deletion cassettes and expressionplasmid constructionPCR amplifications were made from yeasts genomicDNA prepared as described in [21] Amplifications wereperformed by the Pwo DNA Polymerase (Roche)The amplification of the ZbGAS1 gene was as follows
the primers GAS1int_fw 5rsquo-AGT TGT TCC AGA GATATT CCA TAC CTC AA-3rsquo and GAS1int_rev 5rsquo-GCAGAA CCG CTG AAG CTA CAG T-3rsquo designed oninternal conserved region of the available sequencesfrom other yeasts were firstly used to amplify an internalregion of 1137 bp of the Z bailii gene with the follow-ing program 94degC 5 min (94degC 15 s 57degC 30 s 72degC 1min) times 25 72degC 7 min 4degC infin After that the designedprimers ZbGAS1int_fw 5rsquo-GGA CGA TAA GGT TGACGA AGA-3rsquo and ZbGAS1int_rev 5rsquo-TGC TGT GGTCCA AAG TTG TG-3rsquo were used for the amplification(program 7 cycles 94degC 2 min 72degC 3 min 32 cycles94degC 2 min 67degC 3 min 67degC 4 min 4degC infin) of theterminal ends of ZbGAS1 utilizing a Z bailii GenomeWalker library constructed using the Universal GenomeWalker kit (Clontech) following the manufacturersinstructions From the obtained and reconstructedsequence the final primers were established and used forthe ZbGAS1 1710 bp full length coding sequence ampli-fication ZbGAS1_fw 5rsquo-ACT AAT GTT ATT CCAGGC GTT TTC G-3rsquo and ZbGAS1real_rev 5rsquo-AAATCA AGC CAA AGC AAA TCC AGC A-3rsquo (program94degC 5 min (94degC 20 s 58degC 30 s 72degC 2 min) times 30 72degC 10 min 4degC infin) The DNA sequence of ScGAS1 wasPCR amplified with the following primers ScGAS1fw5rsquo-ACA ATG TTG TTT AAA TCC CTT TCA AAGTTA GCA A-3rsquo and ScGAS1rev 5rsquo-TTT TTA AACCAA AGC AAA ACC GAC ACC AG-3rsquo (program 94degC5 min (94degC 20 s 605degC 30 s 72degC 2 min) times 30 72degC10 min 4degC infin) and the DNA sequence of YlXPR2 wasPCR amplified with the following primers XPR2_fwd5rsquo-ACA ATG AAG CTC GCT ACC GCC TTT A-3rsquoand XPR2_rev 5rsquo-TGC CTA AAT GCC AAC ACCGTT GTA G-3rsquo (program 94degC 4 min (94degC 15 s 625degC 30 s 72degC 2 min) times 30 72degC 7 min 4degC infin) All theunique fragments obtained were sub-cloned in the vec-tor pSTblue-1 utilising the Perfectly Bluntreg Cloning Kit(Novagene) and sequenced The resulting respectiveplasmids utilised in this work were named as followspSTbZbGAS1int pSTbZbGAS1 pSTbScGAS1 andpSTbYlXPR2For the construction of the ZbGAS1 deletion cassettes
the KanR or the hphR cassette derived from plasmidpFA6-KanMX4 [22] and pAG26 [23] and conferringresistance to G418 and hygromycin respectively werebluntended excised and inserted in the pSTbZbGAS1intopened BstEII BglII bluntended and dephosphorylatedresulting in the plasmids pSTbZbGAS1ΔK and
pSTbZbGAS1ΔH The resistance cassettes resulted inboth cases counter-clockwise inserted The Z bailii wildtype cells were transformed with the respective deletioncassettes obtained by cutting the two described plasmidsin the AccI and SnaBI sites The effective deletion wasPCR checked by using the following couples of primersZbG1int2_fw 5rsquo- TGC AGA AGT TAC AGA CCAATG TTG T -3rsquo and and ZbG1int2_rev 5rsquo-AAG ATAGAG AAG TGC TCT TGG CA -3rsquo (see Figure 2) orZbG1int2_fw and Hph_fw 5rsquo- ATA TGA AAA AGCCTG AAC TCA CCG AC -3rsquo or Kan_fw 5rsquo- ATG GGTAAG GAA AAG ACT CAC GTT -3rsquo (not shown) inorder to verify the presence of the cassettes in thedesired locusFor complementing the yeasts GAS1 deletion pZ5
(-Nco)ScGAS1 and pYX022ZbGAS1 were constructedpZ5(-Nco)ScGAS1 was obtained from pZ5(-Nco) openedSacIEcoRV with the insertion of ScGAS1 frompSTbScGAS1 cut with SacISnaBI The pZ5(-Nco)derives from pZ3 [3] where at the KpnI site the KanR
cassette was substituted from the NATR cassetteobtained from vector pAG25 [23] by cutting PvuIISacIblunt (obtaining the pZ5 plasmid) finally this plasmidwas EcoRIBamHI cut and bluntended and reclosedpYX022ZbGAS1 was obtained with the insertion ofZbGAS1 from pSTbZbGAS1 into pYX022 (RampD Sys-tems Wiesbaden Germany) both EcoRI cutFor heterologous protein expression the plasmids
pZLN022XPR2 pZLN022LIP1 and pZ5KlhIL-1b wereconstructed The first two integrative plasmids derivesfrom pZLN022 (manuscript in preparation) obtainedfrom pYX022 opened DraIII bluntSpeI blunt (toexclude HIS3) with the insertion of ZbLEU2 cut frompSTbZbLEU2 with EcoRI blunt in KpnI blunt site theNATR cassette was inserted obtained from vectorpAG25 [23] by cutting PvuIISacIblunt ThepSTbZbLEU2 was obtained by sub-cloning ZbLEU2amplified by PCR from Z bailii genomic DNA (manu-script in preparation) in pSTBlue (Novagen) cut byEcoRV For pZLN022XPR2 construction the pZLN022was AatIINheI cut and the XPR2 from pSTbXPR2 wasinserted For pZLN022LIP1 construction the pZLN022was AatII bluntNheI blunt opened and the LIP1 wasinserted by BamHI bluntNsiI blunt excision from thepGAPsLIP [24] For rh interleukin the pZ5KlhIL-1b wasbuilt it was obtained from pZ5 opened with EcoRI-blunt with the insertion of hIL-1b in frame with Kl sig-nal sequence from vector pKSSPI3 [25] cut EcoRIbluntXbaI bluntDNA manipulations transformation and cultivation of
Escherichia coli (DH5aFrsquo (80dlacZΔM15 Δ (lacZYA-argF) U169 deo rec1 end1 sup44 l THI-1 gyrA96relA1) and Novablue Competent Cells (Novagene) wereperformed following standard protocols [26] All the
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 9 of 11
restriction and modification enzymes used were fromNew England Biolabs (Hitchin Herts UK) or fromRoche Diagnostics (Mannheim Germany)Calcofluor staining and fluorescence microscopyCells were harvested washed in PBS and resuspended in1 mg ml-1 Calcofluor White solution incubated 10-30min at room temperature washed again twice in PBSFluorescence microscope images were taken by a NikonEclipse 90i microscope using the Metamorph softwareversion 22 (Nikon) using emission filter UV-1AQuantification of total released proteinsCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were etha-nol-precipitated The amount of proteins was deter-mined by the dye-binding method of Bradford (QuickStart Bradford Dye Reagent - BIORAD) using BSA asstandard All the data were normalized by OD after ver-ification of the linear correlation between OD and dryweight as a measure of biomassQuantification of secreted proteaseCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were sepa-rated and stored at -20degC until analysis The amount ofprotease secreted in the medium was determined by theAzocoll assay (Azocoll - Calbiochem) using commercialprotease as positive control Azocoll assays were per-formed according to [27] with some modificationsReaction mixtures contained 15 mg ml-1 Azocoll 50mM TrisHCl (pH 76) 150 mM NaCl and 5 mMCaCl2 Proteolytic activities were measured by absor-bance changes at 490 nm (Shimadzu UV 1601) afterincubation for 18 h at 30degCQuantification of secreted lipaseCultures were harvested and centrifuged at maximalspeed then the supernatants were separated and storedat -20degC until analysis The amount of lipase secreted inthe medium was determined according to [28] withsome modifications Reaction mixtures contained (A)60 mg of p-nitrophenyl palmitate (Sigma Aldrich) in 20ml of isopropanol (Sigma Aldrich) (B) 1 g Triton X-100(Sigma Aldrich) and 02 g of Arabic gum (SigmaAldrich) in 200 ml TrisHCl 01 M (pH 75) To obtainthe mixture (C) add slowly 1 part of A solution to 9part of B solution under continuous agitation Add 9part of (C) solution to one part of the enzyme contain-ing solution (ie the supernatant) Lipase activities weremeasured by absorbance changes at 410 nm (ShimadzuUV 1601) and the amount of enzyme is proportional toOD according to the relation Um = 0667 times OD410 nm
(if OD410 nm lt 05)Quantification of interleukinHuman IL-1b were analysed by Western blot Indepen-dent transformants were cultured in shake flasks inminimal or rich medium and during the kinetics
samples were collected Cells were harvested (a culturevolume corresponding to 108 cells) by centrifugationOne volume of 2timesLaemmli buffer [29] was added to thesupernatants of said samples they were boiled for 3-5min and stored at -20degC until loading or loaded directlyon a polyacrylamide gel For the analysis of intracellularproduction crude extracts were prepared from 108 cellsby following the trichloroacetic acid protocol [30] andresuspending the final protein extract in 150 μl ofLaemmli buffer Samples were then loaded on standardpolyacrylamide gels (sodium dodecyl sulfate-polyacryla-mide gel electrophoresis final concentration of theseparating gel 15) after separation proteins wereblotted to nitrocellulose membranes and immunodeco-rated An anti-IL-1b rabbit polyclonal antibody (IL-1b(H-153) Santa Cruz Biotechnology USA cat no sc-7884 dilution 1200) was used for the interleukin immu-nodecoration An anti-rabbit IgG horseradish peroxi-dase-conjugated (Amersham Pharmacia Biotech UK catno NA934 dilution 110000) secondary antibody wasused For the interleukin detection as a positive controlthe human recombinant IL-1b from E coli (Roche catno 1 457 756) was always loaded in parallel on thepolyacrylamide gels The proteins were visualised usingthe ECL Western blotting system (Amersham Bios-ciences UK) according to the manufacturerrsquosinstructions
Additional file 1 Clustalw_ntd Nucleotidic sequence of the ZbGAS1gene and alignment with the homologue ScGAS1 (entry YMR307W)Grey coloring represent nucleotidic identity Alignment was performedwith ClustalW software httpwwwebiacukclustalw and graphicalediting was made using Jalview software [31]Click here for file[ httpwwwbiomedcentralcomcontentsupplementary1475-2859-9-7-S1PDF ]
AcknowledgementsSpecial thanks to Valeria Longo and Davide Zocco for technicalcontributions and to Stefania Brocca for providing the pGAPsLIP plasmidThis work was partially supported by FAR 2007 and 2008 (12-1-192) to DP
Author details1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy 2CPC Biotech Srl Naples Italy
Authorsrsquo contributionsPB and DP initiated and coordinated the project LD was responsible forZbGAS1 gene identification by PCR and its deletion LR analyzed the mutantphenotype with CFW and SDS LR and SP performed the batch cultivationanalyzed the secretory phenotype in null mutant and performed theexperiment with heterologous proteins All authors wrote the paper andapproved the final version of the manuscript
Competing interestsThe authors declare that they have no competing interests
Received 2 November 2009Accepted 26 January 2010 Published 26 January 2010
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 10 of 11
References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11
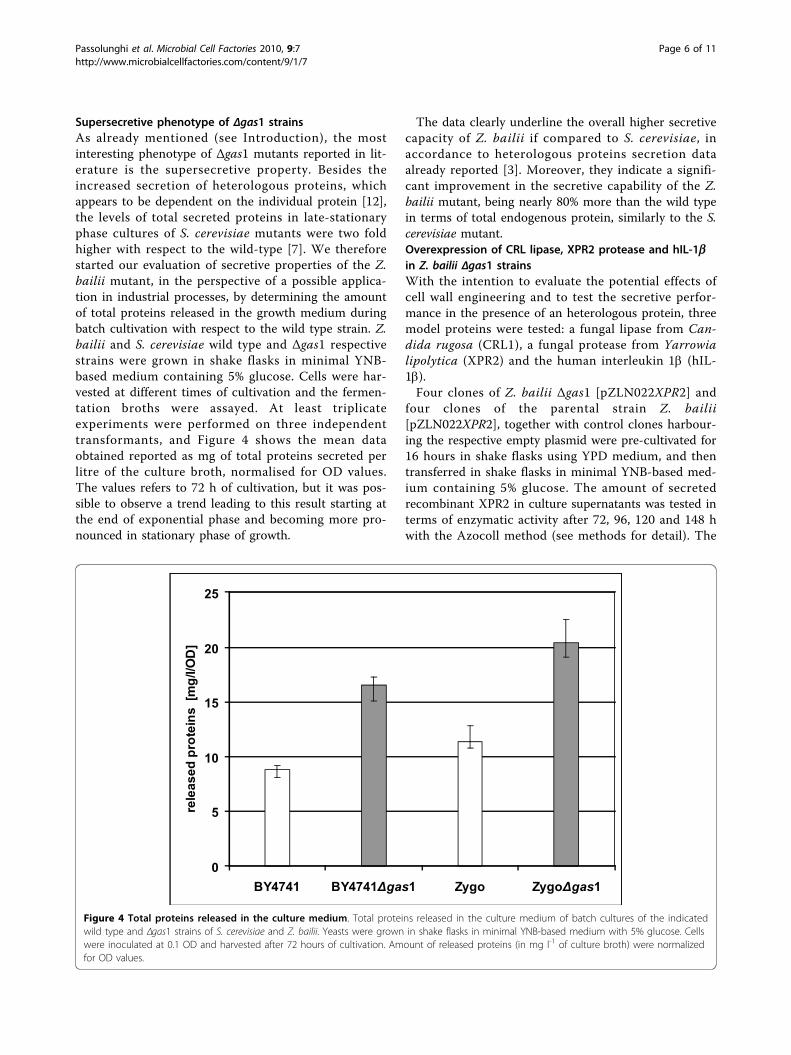
Supersecretive phenotype of Δgas1 strainsAs already mentioned (see Introduction) the mostinteresting phenotype of Δgas1 mutants reported in lit-erature is the supersecretive property Besides theincreased secretion of heterologous proteins whichappears to be dependent on the individual protein [12]the levels of total secreted proteins in late-stationaryphase cultures of S cerevisiae mutants were two foldhigher with respect to the wild-type [7] We thereforestarted our evaluation of secretive properties of the Zbailii mutant in the perspective of a possible applica-tion in industrial processes by determining the amountof total proteins released in the growth medium duringbatch cultivation with respect to the wild type strain Zbailii and S cerevisiae wild type and Δgas1 respectivestrains were grown in shake flasks in minimal YNB-based medium containing 5 glucose Cells were har-vested at different times of cultivation and the fermen-tation broths were assayed At least triplicateexperiments were performed on three independenttransformants and Figure 4 shows the mean dataobtained reported as mg of total proteins secreted perlitre of the culture broth normalised for OD valuesThe values refers to 72 h of cultivation but it was pos-sible to observe a trend leading to this result starting atthe end of exponential phase and becoming more pro-nounced in stationary phase of growth
The data clearly underline the overall higher secretivecapacity of Z bailii if compared to S cerevisiae inaccordance to heterologous proteins secretion dataalready reported [3] Moreover they indicate a signifi-cant improvement in the secretive capability of the Zbailii mutant being nearly 80 more than the wild typein terms of total endogenous protein similarly to the Scerevisiae mutantOverexpression of CRL lipase XPR2 protease and hIL-1bin Z bailii Δgas1 strainsWith the intention to evaluate the potential effects ofcell wall engineering and to test the secretive perfor-mance in the presence of an heterologous protein threemodel proteins were tested a fungal lipase from Can-dida rugosa (CRL1) a fungal protease from Yarrowialipolytica (XPR2) and the human interleukin 1b (hIL-1b)Four clones of Z bailii Δgas1 [pZLN022XPR2] and
four clones of the parental strain Z bailii[pZLN022XPR2] together with control clones harbour-ing the respective empty plasmid were pre-cultivated for16 hours in shake flasks using YPD medium and thentransferred in shake flasks in minimal YNB-based med-ium containing 5 glucose The amount of secretedrecombinant XPR2 in culture supernatants was tested interms of enzymatic activity after 72 96 120 and 148 hwith the Azocoll method (see methods for detail) The
Figure 4 Total proteins released in the culture medium Total proteins released in the culture medium of batch cultures of the indicatedwild type and Δgas1 strains of S cerevisiae and Z bailii Yeasts were grown in shake flasks in minimal YNB-based medium with 5 glucose Cellswere inoculated at 01 OD and harvested after 72 hours of cultivation Amount of released proteins (in mg l-1 of culture broth) were normalizedfor OD values
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 6 of 11
ZbΔgas1 transformants showed a relevant increase inspecific activity levels since 72 h (+ 25 at 72 h morethan two-fold at 148 h see Figure 5a) in respect to wildtype transformantsSimilarly three clones of Z bailii Δgas1
[pZLN022LIP1] and three clones of the parental strainZ bailii [pZLN022LIP1] together with control clonesharbouring the respective empty plasmid were pre-culti-vated for 16 hours in shake flasks using YPD mediumand then transferred in shake flasks in minimal YNB-based medium containing 5 glucose The amount ofsecreted recombinant LIP1 in culture supernatants wastested in terms of enzymatic activity after 24 48 72 and120 h using the hydrolysis of p-nitrophenylestersmethod (see methods for detail) Also for this secondheterologous product the Δgas1 transformants showed arelevant increase in specific activity levels since 24 h
(+35 at 24 h more than two-fold at 120 h see Figure5b) in respect to wild type transformantsFinally three clones of Z bailii Δgas1 [pZ5KlIL-1b]
and three clones of the parental strain Z bailii[pZ5KlIL-1b] together with control clones harbouringthe respective empty plasmid were tested for secretionlevel of the human interleukin since the codingsequence was functionally linked to the signal peptide ofthe K1 killer toxin from K lactis The independenttransformants were pre-cultivated and then cultured inshake flasks containing rich medium (YPD 5 wv glu-cose) Samples were collected at the indicated times (seeFigure 5c) cells were harvested and both the superna-tants and the cell protein extracts of said samples wereloaded on a polyacrylamide gel The data obtained byWestern blot (see Figure 5c) performed on the superna-tant showed a well represented band corresponding to
Figure 5 Secretion levels of yeast protease lipase and rhIL-1b in Z bailii and ZbΔgas1 (a) Secretion levels of protease in Z bailii andZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutant transformed with controlplasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with XPR2 gene (b) Secretion levels oflipase in Z bailii and ZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutanttransformed with control plasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with LIP1 gene(c) Western blots for rhIL-1b of protein samples from supernatants and cell extracts of Z bailii and ZbΔgas1 transformants grown in YPD mediawith 5 glucose inoculated at OD660 = 01 From left to right (A) rhIL-1b (B) negative control Z bailii[pZ5] (C) (D) (E) (I) (L) (M) Z bailii[pZ5KlrhIL-1b] (F) (G) (H) (N) (O) (P) ZbΔgas1 [pZ5KlrhIL-1b] Time and sample types are indicated in the picture
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 7 of 11
the recombinant protein for both Z bailii and Z bailiiΔgas1 samples but differently from the case of recombi-nant fungal enzymes no relevant differences in favourof the deleted yeast are appreciable Remarkably the sig-nal corresponding to the protein retained intracellularlyis more intense in gas1 null mutant than the signalobtained from Z bailii parental strain indicating anincomplete secretion of this protein
DiscussionsCell wall engineering can improve the release of het-erologous proteins from yeasts [18] Generally speak-ing GAS1 which mediates the crosslinking of cell wallcomponents is directly involved in defining the perme-ability of the cell Cloning of the unknown Z bailiiGAS1 homologue by PCR was only possible startingfrom highly conserved internal regions identified uponalignment of the GAS1 homologues of three yeast spe-cies The 5rsquo and 3rsquo ends were subsequently amplifiedusing a Genome Walker library With this approachthe full-length Z bailii GAS1 homologue could beamplified and sequenced The cloned gene wasexpressed in the GAS1-deficient S cerevisiae strain andreversion of the phenotype proved that this homologueis not only structurally but also functionally similar tothe S cerevisiae gene Moreover also the ScGAS1 genecomplemented the Z bailii mutant Disruption ofGAS1 in Z bailii revealed its influence on the cell wallmorphology The GAS1 null mutants of Z bailiishowed the same phenotype reported for S cerevisiae[10] with enlarged cells often showing granular cyto-plasm an alterated chitin distribution two buds(Mickey Mouse-like appearance) and big vacuoles Thevacuole is known to be a dynamic organelle whosemorphology is highly responsive to different extracellu-lar and intracellular stimulistressors [19] In the GAS1null mutant it can be therefore speculated that thegeneration of big vacuoles is related to stressful condi-tions as already suggested by the altered chitin distri-bution being a possible defence mechanism againstlysis or related to a generalized response to sickness ofthe cells [10]Finally also the sensitivity to SDS and CFW
appeared very similar to what observed in the S cerevi-siae mutant A comparison of the different GAS1 nullphenotype mutants and the functionality of the Z bai-lii GAS1 homologue in S cerevisiae suggest a similarityof the cell wall structure of Z bailii compared with thecell wall structure of S cerevisiae Considering thesecretory pathway of a secreted protein it was specu-lated whether the protein becomes retarded in the cellwall after release from the plasma membrane The dis-ruption of GAS1 in Z bailii first of all confirmed ageneral phenomenon of protein release in respect to
the wild type strain as demonstrated by the higheraccumulation of total protein in the growing mediumof the deleted strain when compared to the parentalstrain Moreover such as compromised cell wall per-meability in the production strains for XPR2 and LIP1does lead to a clear improvement in the secretion ofthe respective proteins In the case of h IL-1b theenhancement of product secretion was not so evidentand difficult to interpret in respect to the higher accu-mulation of intracellular product It could be specu-lated that this can be related to the nature or to thesource of the protein but too few information areavailable at the moment to further comment in thisdirection What seems to be true is unfortunately whatwe very often experience for recombinant protein pro-duction which is the protein and protein-host depen-dence In fact also in P pastoris two out of threeheterologous proteins resulted more secreted in themutant as for Z bailii and when we tested the sameprotein in particular the IL-1b secretion in the S cere-visiae Δgas1 mutant (data not shown) we did not reg-ister any improvement
MethodsYeast strains media and transformationThe S cerevisiae strains used in this study were theauxotrophic haploid BY4741 (Mat a his3Δ1 leu2Δ0met5Δ0 ura3Δ0) and the relative null mutant in GAS1sequence called in this study BY4741 Δgas1 (Mat ahis3Δ1 leu2Δ0 met5Δ0 ura3Δ0 YMR307kanMX4)Both strains are commercially available from Euroscarfcollection httpwebuni-frankfurtdefb15mikroeuro-scarfcol_indexhtml The Z bailii wild type strain isATCC60483 Yeast cultures were grown in YP medium(10 g l-1 yeast extract 20 g l-1 peptone) with 2 or 5wv glucose (D) or fructose (F) as carbon sourceWhere needed NAT (80 mg l-1) or G418 (500 mg l-1
for S cerevisiae and 200 mg l-1 for Z bailii) or hygro-micine (100 mg l-1 for Z bailii) were added to themedia Alternatively yeast cultures were grown inminimal synthetic medium (067 w v-1 YNB Biolifewithout amino acids) with 2 or 5 w v-1 glucose ascarbon source When required supplements such ashistidine leucine methionine and uracil were added toa final concentration of 50 mg l-1 Media for plateswere solidified by addition of agar (Biolife Milan Italy)to 15 w v-1 Shake flask experiments were carriedout with 50 ml of medium in 250 ml shake flasks on ashaker at 30degC with 160 rpm For sensitivity tests inPetri dishes 125 μg ml-1 of CFW or 50 μg ml-1 SDSwere added to solid media Yeast transformation wasperformed using the Lithium AcetatessDNA methodas described [20] and slightly adapted for Z bailii asdescribed [3]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 8 of 11
Genes amplifications deletion cassettes and expressionplasmid constructionPCR amplifications were made from yeasts genomicDNA prepared as described in [21] Amplifications wereperformed by the Pwo DNA Polymerase (Roche)The amplification of the ZbGAS1 gene was as follows
the primers GAS1int_fw 5rsquo-AGT TGT TCC AGA GATATT CCA TAC CTC AA-3rsquo and GAS1int_rev 5rsquo-GCAGAA CCG CTG AAG CTA CAG T-3rsquo designed oninternal conserved region of the available sequencesfrom other yeasts were firstly used to amplify an internalregion of 1137 bp of the Z bailii gene with the follow-ing program 94degC 5 min (94degC 15 s 57degC 30 s 72degC 1min) times 25 72degC 7 min 4degC infin After that the designedprimers ZbGAS1int_fw 5rsquo-GGA CGA TAA GGT TGACGA AGA-3rsquo and ZbGAS1int_rev 5rsquo-TGC TGT GGTCCA AAG TTG TG-3rsquo were used for the amplification(program 7 cycles 94degC 2 min 72degC 3 min 32 cycles94degC 2 min 67degC 3 min 67degC 4 min 4degC infin) of theterminal ends of ZbGAS1 utilizing a Z bailii GenomeWalker library constructed using the Universal GenomeWalker kit (Clontech) following the manufacturersinstructions From the obtained and reconstructedsequence the final primers were established and used forthe ZbGAS1 1710 bp full length coding sequence ampli-fication ZbGAS1_fw 5rsquo-ACT AAT GTT ATT CCAGGC GTT TTC G-3rsquo and ZbGAS1real_rev 5rsquo-AAATCA AGC CAA AGC AAA TCC AGC A-3rsquo (program94degC 5 min (94degC 20 s 58degC 30 s 72degC 2 min) times 30 72degC 10 min 4degC infin) The DNA sequence of ScGAS1 wasPCR amplified with the following primers ScGAS1fw5rsquo-ACA ATG TTG TTT AAA TCC CTT TCA AAGTTA GCA A-3rsquo and ScGAS1rev 5rsquo-TTT TTA AACCAA AGC AAA ACC GAC ACC AG-3rsquo (program 94degC5 min (94degC 20 s 605degC 30 s 72degC 2 min) times 30 72degC10 min 4degC infin) and the DNA sequence of YlXPR2 wasPCR amplified with the following primers XPR2_fwd5rsquo-ACA ATG AAG CTC GCT ACC GCC TTT A-3rsquoand XPR2_rev 5rsquo-TGC CTA AAT GCC AAC ACCGTT GTA G-3rsquo (program 94degC 4 min (94degC 15 s 625degC 30 s 72degC 2 min) times 30 72degC 7 min 4degC infin) All theunique fragments obtained were sub-cloned in the vec-tor pSTblue-1 utilising the Perfectly Bluntreg Cloning Kit(Novagene) and sequenced The resulting respectiveplasmids utilised in this work were named as followspSTbZbGAS1int pSTbZbGAS1 pSTbScGAS1 andpSTbYlXPR2For the construction of the ZbGAS1 deletion cassettes
the KanR or the hphR cassette derived from plasmidpFA6-KanMX4 [22] and pAG26 [23] and conferringresistance to G418 and hygromycin respectively werebluntended excised and inserted in the pSTbZbGAS1intopened BstEII BglII bluntended and dephosphorylatedresulting in the plasmids pSTbZbGAS1ΔK and
pSTbZbGAS1ΔH The resistance cassettes resulted inboth cases counter-clockwise inserted The Z bailii wildtype cells were transformed with the respective deletioncassettes obtained by cutting the two described plasmidsin the AccI and SnaBI sites The effective deletion wasPCR checked by using the following couples of primersZbG1int2_fw 5rsquo- TGC AGA AGT TAC AGA CCAATG TTG T -3rsquo and and ZbG1int2_rev 5rsquo-AAG ATAGAG AAG TGC TCT TGG CA -3rsquo (see Figure 2) orZbG1int2_fw and Hph_fw 5rsquo- ATA TGA AAA AGCCTG AAC TCA CCG AC -3rsquo or Kan_fw 5rsquo- ATG GGTAAG GAA AAG ACT CAC GTT -3rsquo (not shown) inorder to verify the presence of the cassettes in thedesired locusFor complementing the yeasts GAS1 deletion pZ5
(-Nco)ScGAS1 and pYX022ZbGAS1 were constructedpZ5(-Nco)ScGAS1 was obtained from pZ5(-Nco) openedSacIEcoRV with the insertion of ScGAS1 frompSTbScGAS1 cut with SacISnaBI The pZ5(-Nco)derives from pZ3 [3] where at the KpnI site the KanR
cassette was substituted from the NATR cassetteobtained from vector pAG25 [23] by cutting PvuIISacIblunt (obtaining the pZ5 plasmid) finally this plasmidwas EcoRIBamHI cut and bluntended and reclosedpYX022ZbGAS1 was obtained with the insertion ofZbGAS1 from pSTbZbGAS1 into pYX022 (RampD Sys-tems Wiesbaden Germany) both EcoRI cutFor heterologous protein expression the plasmids
pZLN022XPR2 pZLN022LIP1 and pZ5KlhIL-1b wereconstructed The first two integrative plasmids derivesfrom pZLN022 (manuscript in preparation) obtainedfrom pYX022 opened DraIII bluntSpeI blunt (toexclude HIS3) with the insertion of ZbLEU2 cut frompSTbZbLEU2 with EcoRI blunt in KpnI blunt site theNATR cassette was inserted obtained from vectorpAG25 [23] by cutting PvuIISacIblunt ThepSTbZbLEU2 was obtained by sub-cloning ZbLEU2amplified by PCR from Z bailii genomic DNA (manu-script in preparation) in pSTBlue (Novagen) cut byEcoRV For pZLN022XPR2 construction the pZLN022was AatIINheI cut and the XPR2 from pSTbXPR2 wasinserted For pZLN022LIP1 construction the pZLN022was AatII bluntNheI blunt opened and the LIP1 wasinserted by BamHI bluntNsiI blunt excision from thepGAPsLIP [24] For rh interleukin the pZ5KlhIL-1b wasbuilt it was obtained from pZ5 opened with EcoRI-blunt with the insertion of hIL-1b in frame with Kl sig-nal sequence from vector pKSSPI3 [25] cut EcoRIbluntXbaI bluntDNA manipulations transformation and cultivation of
Escherichia coli (DH5aFrsquo (80dlacZΔM15 Δ (lacZYA-argF) U169 deo rec1 end1 sup44 l THI-1 gyrA96relA1) and Novablue Competent Cells (Novagene) wereperformed following standard protocols [26] All the
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 9 of 11
restriction and modification enzymes used were fromNew England Biolabs (Hitchin Herts UK) or fromRoche Diagnostics (Mannheim Germany)Calcofluor staining and fluorescence microscopyCells were harvested washed in PBS and resuspended in1 mg ml-1 Calcofluor White solution incubated 10-30min at room temperature washed again twice in PBSFluorescence microscope images were taken by a NikonEclipse 90i microscope using the Metamorph softwareversion 22 (Nikon) using emission filter UV-1AQuantification of total released proteinsCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were etha-nol-precipitated The amount of proteins was deter-mined by the dye-binding method of Bradford (QuickStart Bradford Dye Reagent - BIORAD) using BSA asstandard All the data were normalized by OD after ver-ification of the linear correlation between OD and dryweight as a measure of biomassQuantification of secreted proteaseCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were sepa-rated and stored at -20degC until analysis The amount ofprotease secreted in the medium was determined by theAzocoll assay (Azocoll - Calbiochem) using commercialprotease as positive control Azocoll assays were per-formed according to [27] with some modificationsReaction mixtures contained 15 mg ml-1 Azocoll 50mM TrisHCl (pH 76) 150 mM NaCl and 5 mMCaCl2 Proteolytic activities were measured by absor-bance changes at 490 nm (Shimadzu UV 1601) afterincubation for 18 h at 30degCQuantification of secreted lipaseCultures were harvested and centrifuged at maximalspeed then the supernatants were separated and storedat -20degC until analysis The amount of lipase secreted inthe medium was determined according to [28] withsome modifications Reaction mixtures contained (A)60 mg of p-nitrophenyl palmitate (Sigma Aldrich) in 20ml of isopropanol (Sigma Aldrich) (B) 1 g Triton X-100(Sigma Aldrich) and 02 g of Arabic gum (SigmaAldrich) in 200 ml TrisHCl 01 M (pH 75) To obtainthe mixture (C) add slowly 1 part of A solution to 9part of B solution under continuous agitation Add 9part of (C) solution to one part of the enzyme contain-ing solution (ie the supernatant) Lipase activities weremeasured by absorbance changes at 410 nm (ShimadzuUV 1601) and the amount of enzyme is proportional toOD according to the relation Um = 0667 times OD410 nm
(if OD410 nm lt 05)Quantification of interleukinHuman IL-1b were analysed by Western blot Indepen-dent transformants were cultured in shake flasks inminimal or rich medium and during the kinetics
samples were collected Cells were harvested (a culturevolume corresponding to 108 cells) by centrifugationOne volume of 2timesLaemmli buffer [29] was added to thesupernatants of said samples they were boiled for 3-5min and stored at -20degC until loading or loaded directlyon a polyacrylamide gel For the analysis of intracellularproduction crude extracts were prepared from 108 cellsby following the trichloroacetic acid protocol [30] andresuspending the final protein extract in 150 μl ofLaemmli buffer Samples were then loaded on standardpolyacrylamide gels (sodium dodecyl sulfate-polyacryla-mide gel electrophoresis final concentration of theseparating gel 15) after separation proteins wereblotted to nitrocellulose membranes and immunodeco-rated An anti-IL-1b rabbit polyclonal antibody (IL-1b(H-153) Santa Cruz Biotechnology USA cat no sc-7884 dilution 1200) was used for the interleukin immu-nodecoration An anti-rabbit IgG horseradish peroxi-dase-conjugated (Amersham Pharmacia Biotech UK catno NA934 dilution 110000) secondary antibody wasused For the interleukin detection as a positive controlthe human recombinant IL-1b from E coli (Roche catno 1 457 756) was always loaded in parallel on thepolyacrylamide gels The proteins were visualised usingthe ECL Western blotting system (Amersham Bios-ciences UK) according to the manufacturerrsquosinstructions
Additional file 1 Clustalw_ntd Nucleotidic sequence of the ZbGAS1gene and alignment with the homologue ScGAS1 (entry YMR307W)Grey coloring represent nucleotidic identity Alignment was performedwith ClustalW software httpwwwebiacukclustalw and graphicalediting was made using Jalview software [31]Click here for file[ httpwwwbiomedcentralcomcontentsupplementary1475-2859-9-7-S1PDF ]
AcknowledgementsSpecial thanks to Valeria Longo and Davide Zocco for technicalcontributions and to Stefania Brocca for providing the pGAPsLIP plasmidThis work was partially supported by FAR 2007 and 2008 (12-1-192) to DP
Author details1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy 2CPC Biotech Srl Naples Italy
Authorsrsquo contributionsPB and DP initiated and coordinated the project LD was responsible forZbGAS1 gene identification by PCR and its deletion LR analyzed the mutantphenotype with CFW and SDS LR and SP performed the batch cultivationanalyzed the secretory phenotype in null mutant and performed theexperiment with heterologous proteins All authors wrote the paper andapproved the final version of the manuscript
Competing interestsThe authors declare that they have no competing interests
Received 2 November 2009Accepted 26 January 2010 Published 26 January 2010
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 10 of 11
References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11
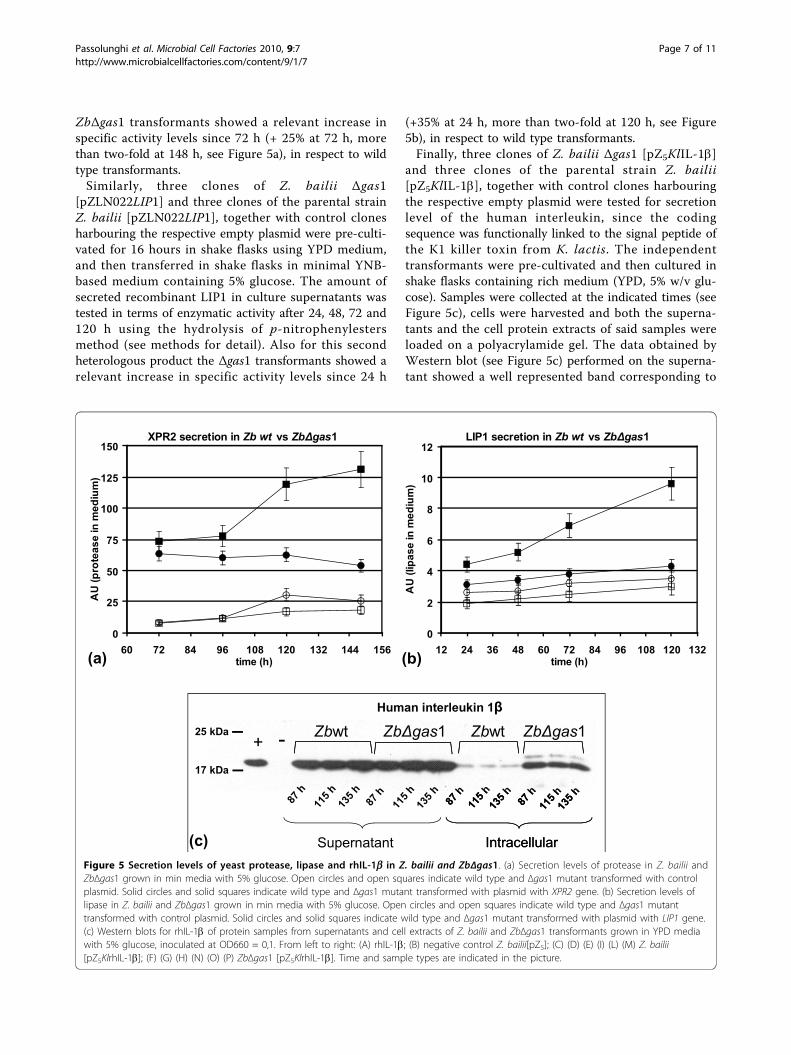
ZbΔgas1 transformants showed a relevant increase inspecific activity levels since 72 h (+ 25 at 72 h morethan two-fold at 148 h see Figure 5a) in respect to wildtype transformantsSimilarly three clones of Z bailii Δgas1
[pZLN022LIP1] and three clones of the parental strainZ bailii [pZLN022LIP1] together with control clonesharbouring the respective empty plasmid were pre-culti-vated for 16 hours in shake flasks using YPD mediumand then transferred in shake flasks in minimal YNB-based medium containing 5 glucose The amount ofsecreted recombinant LIP1 in culture supernatants wastested in terms of enzymatic activity after 24 48 72 and120 h using the hydrolysis of p-nitrophenylestersmethod (see methods for detail) Also for this secondheterologous product the Δgas1 transformants showed arelevant increase in specific activity levels since 24 h
(+35 at 24 h more than two-fold at 120 h see Figure5b) in respect to wild type transformantsFinally three clones of Z bailii Δgas1 [pZ5KlIL-1b]
and three clones of the parental strain Z bailii[pZ5KlIL-1b] together with control clones harbouringthe respective empty plasmid were tested for secretionlevel of the human interleukin since the codingsequence was functionally linked to the signal peptide ofthe K1 killer toxin from K lactis The independenttransformants were pre-cultivated and then cultured inshake flasks containing rich medium (YPD 5 wv glu-cose) Samples were collected at the indicated times (seeFigure 5c) cells were harvested and both the superna-tants and the cell protein extracts of said samples wereloaded on a polyacrylamide gel The data obtained byWestern blot (see Figure 5c) performed on the superna-tant showed a well represented band corresponding to
Figure 5 Secretion levels of yeast protease lipase and rhIL-1b in Z bailii and ZbΔgas1 (a) Secretion levels of protease in Z bailii andZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutant transformed with controlplasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with XPR2 gene (b) Secretion levels oflipase in Z bailii and ZbΔgas1 grown in min media with 5 glucose Open circles and open squares indicate wild type and Δgas1 mutanttransformed with control plasmid Solid circles and solid squares indicate wild type and Δgas1 mutant transformed with plasmid with LIP1 gene(c) Western blots for rhIL-1b of protein samples from supernatants and cell extracts of Z bailii and ZbΔgas1 transformants grown in YPD mediawith 5 glucose inoculated at OD660 = 01 From left to right (A) rhIL-1b (B) negative control Z bailii[pZ5] (C) (D) (E) (I) (L) (M) Z bailii[pZ5KlrhIL-1b] (F) (G) (H) (N) (O) (P) ZbΔgas1 [pZ5KlrhIL-1b] Time and sample types are indicated in the picture
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 7 of 11
the recombinant protein for both Z bailii and Z bailiiΔgas1 samples but differently from the case of recombi-nant fungal enzymes no relevant differences in favourof the deleted yeast are appreciable Remarkably the sig-nal corresponding to the protein retained intracellularlyis more intense in gas1 null mutant than the signalobtained from Z bailii parental strain indicating anincomplete secretion of this protein
DiscussionsCell wall engineering can improve the release of het-erologous proteins from yeasts [18] Generally speak-ing GAS1 which mediates the crosslinking of cell wallcomponents is directly involved in defining the perme-ability of the cell Cloning of the unknown Z bailiiGAS1 homologue by PCR was only possible startingfrom highly conserved internal regions identified uponalignment of the GAS1 homologues of three yeast spe-cies The 5rsquo and 3rsquo ends were subsequently amplifiedusing a Genome Walker library With this approachthe full-length Z bailii GAS1 homologue could beamplified and sequenced The cloned gene wasexpressed in the GAS1-deficient S cerevisiae strain andreversion of the phenotype proved that this homologueis not only structurally but also functionally similar tothe S cerevisiae gene Moreover also the ScGAS1 genecomplemented the Z bailii mutant Disruption ofGAS1 in Z bailii revealed its influence on the cell wallmorphology The GAS1 null mutants of Z bailiishowed the same phenotype reported for S cerevisiae[10] with enlarged cells often showing granular cyto-plasm an alterated chitin distribution two buds(Mickey Mouse-like appearance) and big vacuoles Thevacuole is known to be a dynamic organelle whosemorphology is highly responsive to different extracellu-lar and intracellular stimulistressors [19] In the GAS1null mutant it can be therefore speculated that thegeneration of big vacuoles is related to stressful condi-tions as already suggested by the altered chitin distri-bution being a possible defence mechanism againstlysis or related to a generalized response to sickness ofthe cells [10]Finally also the sensitivity to SDS and CFW
appeared very similar to what observed in the S cerevi-siae mutant A comparison of the different GAS1 nullphenotype mutants and the functionality of the Z bai-lii GAS1 homologue in S cerevisiae suggest a similarityof the cell wall structure of Z bailii compared with thecell wall structure of S cerevisiae Considering thesecretory pathway of a secreted protein it was specu-lated whether the protein becomes retarded in the cellwall after release from the plasma membrane The dis-ruption of GAS1 in Z bailii first of all confirmed ageneral phenomenon of protein release in respect to
the wild type strain as demonstrated by the higheraccumulation of total protein in the growing mediumof the deleted strain when compared to the parentalstrain Moreover such as compromised cell wall per-meability in the production strains for XPR2 and LIP1does lead to a clear improvement in the secretion ofthe respective proteins In the case of h IL-1b theenhancement of product secretion was not so evidentand difficult to interpret in respect to the higher accu-mulation of intracellular product It could be specu-lated that this can be related to the nature or to thesource of the protein but too few information areavailable at the moment to further comment in thisdirection What seems to be true is unfortunately whatwe very often experience for recombinant protein pro-duction which is the protein and protein-host depen-dence In fact also in P pastoris two out of threeheterologous proteins resulted more secreted in themutant as for Z bailii and when we tested the sameprotein in particular the IL-1b secretion in the S cere-visiae Δgas1 mutant (data not shown) we did not reg-ister any improvement
MethodsYeast strains media and transformationThe S cerevisiae strains used in this study were theauxotrophic haploid BY4741 (Mat a his3Δ1 leu2Δ0met5Δ0 ura3Δ0) and the relative null mutant in GAS1sequence called in this study BY4741 Δgas1 (Mat ahis3Δ1 leu2Δ0 met5Δ0 ura3Δ0 YMR307kanMX4)Both strains are commercially available from Euroscarfcollection httpwebuni-frankfurtdefb15mikroeuro-scarfcol_indexhtml The Z bailii wild type strain isATCC60483 Yeast cultures were grown in YP medium(10 g l-1 yeast extract 20 g l-1 peptone) with 2 or 5wv glucose (D) or fructose (F) as carbon sourceWhere needed NAT (80 mg l-1) or G418 (500 mg l-1
for S cerevisiae and 200 mg l-1 for Z bailii) or hygro-micine (100 mg l-1 for Z bailii) were added to themedia Alternatively yeast cultures were grown inminimal synthetic medium (067 w v-1 YNB Biolifewithout amino acids) with 2 or 5 w v-1 glucose ascarbon source When required supplements such ashistidine leucine methionine and uracil were added toa final concentration of 50 mg l-1 Media for plateswere solidified by addition of agar (Biolife Milan Italy)to 15 w v-1 Shake flask experiments were carriedout with 50 ml of medium in 250 ml shake flasks on ashaker at 30degC with 160 rpm For sensitivity tests inPetri dishes 125 μg ml-1 of CFW or 50 μg ml-1 SDSwere added to solid media Yeast transformation wasperformed using the Lithium AcetatessDNA methodas described [20] and slightly adapted for Z bailii asdescribed [3]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 8 of 11
Genes amplifications deletion cassettes and expressionplasmid constructionPCR amplifications were made from yeasts genomicDNA prepared as described in [21] Amplifications wereperformed by the Pwo DNA Polymerase (Roche)The amplification of the ZbGAS1 gene was as follows
the primers GAS1int_fw 5rsquo-AGT TGT TCC AGA GATATT CCA TAC CTC AA-3rsquo and GAS1int_rev 5rsquo-GCAGAA CCG CTG AAG CTA CAG T-3rsquo designed oninternal conserved region of the available sequencesfrom other yeasts were firstly used to amplify an internalregion of 1137 bp of the Z bailii gene with the follow-ing program 94degC 5 min (94degC 15 s 57degC 30 s 72degC 1min) times 25 72degC 7 min 4degC infin After that the designedprimers ZbGAS1int_fw 5rsquo-GGA CGA TAA GGT TGACGA AGA-3rsquo and ZbGAS1int_rev 5rsquo-TGC TGT GGTCCA AAG TTG TG-3rsquo were used for the amplification(program 7 cycles 94degC 2 min 72degC 3 min 32 cycles94degC 2 min 67degC 3 min 67degC 4 min 4degC infin) of theterminal ends of ZbGAS1 utilizing a Z bailii GenomeWalker library constructed using the Universal GenomeWalker kit (Clontech) following the manufacturersinstructions From the obtained and reconstructedsequence the final primers were established and used forthe ZbGAS1 1710 bp full length coding sequence ampli-fication ZbGAS1_fw 5rsquo-ACT AAT GTT ATT CCAGGC GTT TTC G-3rsquo and ZbGAS1real_rev 5rsquo-AAATCA AGC CAA AGC AAA TCC AGC A-3rsquo (program94degC 5 min (94degC 20 s 58degC 30 s 72degC 2 min) times 30 72degC 10 min 4degC infin) The DNA sequence of ScGAS1 wasPCR amplified with the following primers ScGAS1fw5rsquo-ACA ATG TTG TTT AAA TCC CTT TCA AAGTTA GCA A-3rsquo and ScGAS1rev 5rsquo-TTT TTA AACCAA AGC AAA ACC GAC ACC AG-3rsquo (program 94degC5 min (94degC 20 s 605degC 30 s 72degC 2 min) times 30 72degC10 min 4degC infin) and the DNA sequence of YlXPR2 wasPCR amplified with the following primers XPR2_fwd5rsquo-ACA ATG AAG CTC GCT ACC GCC TTT A-3rsquoand XPR2_rev 5rsquo-TGC CTA AAT GCC AAC ACCGTT GTA G-3rsquo (program 94degC 4 min (94degC 15 s 625degC 30 s 72degC 2 min) times 30 72degC 7 min 4degC infin) All theunique fragments obtained were sub-cloned in the vec-tor pSTblue-1 utilising the Perfectly Bluntreg Cloning Kit(Novagene) and sequenced The resulting respectiveplasmids utilised in this work were named as followspSTbZbGAS1int pSTbZbGAS1 pSTbScGAS1 andpSTbYlXPR2For the construction of the ZbGAS1 deletion cassettes
the KanR or the hphR cassette derived from plasmidpFA6-KanMX4 [22] and pAG26 [23] and conferringresistance to G418 and hygromycin respectively werebluntended excised and inserted in the pSTbZbGAS1intopened BstEII BglII bluntended and dephosphorylatedresulting in the plasmids pSTbZbGAS1ΔK and
pSTbZbGAS1ΔH The resistance cassettes resulted inboth cases counter-clockwise inserted The Z bailii wildtype cells were transformed with the respective deletioncassettes obtained by cutting the two described plasmidsin the AccI and SnaBI sites The effective deletion wasPCR checked by using the following couples of primersZbG1int2_fw 5rsquo- TGC AGA AGT TAC AGA CCAATG TTG T -3rsquo and and ZbG1int2_rev 5rsquo-AAG ATAGAG AAG TGC TCT TGG CA -3rsquo (see Figure 2) orZbG1int2_fw and Hph_fw 5rsquo- ATA TGA AAA AGCCTG AAC TCA CCG AC -3rsquo or Kan_fw 5rsquo- ATG GGTAAG GAA AAG ACT CAC GTT -3rsquo (not shown) inorder to verify the presence of the cassettes in thedesired locusFor complementing the yeasts GAS1 deletion pZ5
(-Nco)ScGAS1 and pYX022ZbGAS1 were constructedpZ5(-Nco)ScGAS1 was obtained from pZ5(-Nco) openedSacIEcoRV with the insertion of ScGAS1 frompSTbScGAS1 cut with SacISnaBI The pZ5(-Nco)derives from pZ3 [3] where at the KpnI site the KanR
cassette was substituted from the NATR cassetteobtained from vector pAG25 [23] by cutting PvuIISacIblunt (obtaining the pZ5 plasmid) finally this plasmidwas EcoRIBamHI cut and bluntended and reclosedpYX022ZbGAS1 was obtained with the insertion ofZbGAS1 from pSTbZbGAS1 into pYX022 (RampD Sys-tems Wiesbaden Germany) both EcoRI cutFor heterologous protein expression the plasmids
pZLN022XPR2 pZLN022LIP1 and pZ5KlhIL-1b wereconstructed The first two integrative plasmids derivesfrom pZLN022 (manuscript in preparation) obtainedfrom pYX022 opened DraIII bluntSpeI blunt (toexclude HIS3) with the insertion of ZbLEU2 cut frompSTbZbLEU2 with EcoRI blunt in KpnI blunt site theNATR cassette was inserted obtained from vectorpAG25 [23] by cutting PvuIISacIblunt ThepSTbZbLEU2 was obtained by sub-cloning ZbLEU2amplified by PCR from Z bailii genomic DNA (manu-script in preparation) in pSTBlue (Novagen) cut byEcoRV For pZLN022XPR2 construction the pZLN022was AatIINheI cut and the XPR2 from pSTbXPR2 wasinserted For pZLN022LIP1 construction the pZLN022was AatII bluntNheI blunt opened and the LIP1 wasinserted by BamHI bluntNsiI blunt excision from thepGAPsLIP [24] For rh interleukin the pZ5KlhIL-1b wasbuilt it was obtained from pZ5 opened with EcoRI-blunt with the insertion of hIL-1b in frame with Kl sig-nal sequence from vector pKSSPI3 [25] cut EcoRIbluntXbaI bluntDNA manipulations transformation and cultivation of
Escherichia coli (DH5aFrsquo (80dlacZΔM15 Δ (lacZYA-argF) U169 deo rec1 end1 sup44 l THI-1 gyrA96relA1) and Novablue Competent Cells (Novagene) wereperformed following standard protocols [26] All the
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 9 of 11
restriction and modification enzymes used were fromNew England Biolabs (Hitchin Herts UK) or fromRoche Diagnostics (Mannheim Germany)Calcofluor staining and fluorescence microscopyCells were harvested washed in PBS and resuspended in1 mg ml-1 Calcofluor White solution incubated 10-30min at room temperature washed again twice in PBSFluorescence microscope images were taken by a NikonEclipse 90i microscope using the Metamorph softwareversion 22 (Nikon) using emission filter UV-1AQuantification of total released proteinsCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were etha-nol-precipitated The amount of proteins was deter-mined by the dye-binding method of Bradford (QuickStart Bradford Dye Reagent - BIORAD) using BSA asstandard All the data were normalized by OD after ver-ification of the linear correlation between OD and dryweight as a measure of biomassQuantification of secreted proteaseCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were sepa-rated and stored at -20degC until analysis The amount ofprotease secreted in the medium was determined by theAzocoll assay (Azocoll - Calbiochem) using commercialprotease as positive control Azocoll assays were per-formed according to [27] with some modificationsReaction mixtures contained 15 mg ml-1 Azocoll 50mM TrisHCl (pH 76) 150 mM NaCl and 5 mMCaCl2 Proteolytic activities were measured by absor-bance changes at 490 nm (Shimadzu UV 1601) afterincubation for 18 h at 30degCQuantification of secreted lipaseCultures were harvested and centrifuged at maximalspeed then the supernatants were separated and storedat -20degC until analysis The amount of lipase secreted inthe medium was determined according to [28] withsome modifications Reaction mixtures contained (A)60 mg of p-nitrophenyl palmitate (Sigma Aldrich) in 20ml of isopropanol (Sigma Aldrich) (B) 1 g Triton X-100(Sigma Aldrich) and 02 g of Arabic gum (SigmaAldrich) in 200 ml TrisHCl 01 M (pH 75) To obtainthe mixture (C) add slowly 1 part of A solution to 9part of B solution under continuous agitation Add 9part of (C) solution to one part of the enzyme contain-ing solution (ie the supernatant) Lipase activities weremeasured by absorbance changes at 410 nm (ShimadzuUV 1601) and the amount of enzyme is proportional toOD according to the relation Um = 0667 times OD410 nm
(if OD410 nm lt 05)Quantification of interleukinHuman IL-1b were analysed by Western blot Indepen-dent transformants were cultured in shake flasks inminimal or rich medium and during the kinetics
samples were collected Cells were harvested (a culturevolume corresponding to 108 cells) by centrifugationOne volume of 2timesLaemmli buffer [29] was added to thesupernatants of said samples they were boiled for 3-5min and stored at -20degC until loading or loaded directlyon a polyacrylamide gel For the analysis of intracellularproduction crude extracts were prepared from 108 cellsby following the trichloroacetic acid protocol [30] andresuspending the final protein extract in 150 μl ofLaemmli buffer Samples were then loaded on standardpolyacrylamide gels (sodium dodecyl sulfate-polyacryla-mide gel electrophoresis final concentration of theseparating gel 15) after separation proteins wereblotted to nitrocellulose membranes and immunodeco-rated An anti-IL-1b rabbit polyclonal antibody (IL-1b(H-153) Santa Cruz Biotechnology USA cat no sc-7884 dilution 1200) was used for the interleukin immu-nodecoration An anti-rabbit IgG horseradish peroxi-dase-conjugated (Amersham Pharmacia Biotech UK catno NA934 dilution 110000) secondary antibody wasused For the interleukin detection as a positive controlthe human recombinant IL-1b from E coli (Roche catno 1 457 756) was always loaded in parallel on thepolyacrylamide gels The proteins were visualised usingthe ECL Western blotting system (Amersham Bios-ciences UK) according to the manufacturerrsquosinstructions
Additional file 1 Clustalw_ntd Nucleotidic sequence of the ZbGAS1gene and alignment with the homologue ScGAS1 (entry YMR307W)Grey coloring represent nucleotidic identity Alignment was performedwith ClustalW software httpwwwebiacukclustalw and graphicalediting was made using Jalview software [31]Click here for file[ httpwwwbiomedcentralcomcontentsupplementary1475-2859-9-7-S1PDF ]
AcknowledgementsSpecial thanks to Valeria Longo and Davide Zocco for technicalcontributions and to Stefania Brocca for providing the pGAPsLIP plasmidThis work was partially supported by FAR 2007 and 2008 (12-1-192) to DP
Author details1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy 2CPC Biotech Srl Naples Italy
Authorsrsquo contributionsPB and DP initiated and coordinated the project LD was responsible forZbGAS1 gene identification by PCR and its deletion LR analyzed the mutantphenotype with CFW and SDS LR and SP performed the batch cultivationanalyzed the secretory phenotype in null mutant and performed theexperiment with heterologous proteins All authors wrote the paper andapproved the final version of the manuscript
Competing interestsThe authors declare that they have no competing interests
Received 2 November 2009Accepted 26 January 2010 Published 26 January 2010
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 10 of 11
References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11

the recombinant protein for both Z bailii and Z bailiiΔgas1 samples but differently from the case of recombi-nant fungal enzymes no relevant differences in favourof the deleted yeast are appreciable Remarkably the sig-nal corresponding to the protein retained intracellularlyis more intense in gas1 null mutant than the signalobtained from Z bailii parental strain indicating anincomplete secretion of this protein
DiscussionsCell wall engineering can improve the release of het-erologous proteins from yeasts [18] Generally speak-ing GAS1 which mediates the crosslinking of cell wallcomponents is directly involved in defining the perme-ability of the cell Cloning of the unknown Z bailiiGAS1 homologue by PCR was only possible startingfrom highly conserved internal regions identified uponalignment of the GAS1 homologues of three yeast spe-cies The 5rsquo and 3rsquo ends were subsequently amplifiedusing a Genome Walker library With this approachthe full-length Z bailii GAS1 homologue could beamplified and sequenced The cloned gene wasexpressed in the GAS1-deficient S cerevisiae strain andreversion of the phenotype proved that this homologueis not only structurally but also functionally similar tothe S cerevisiae gene Moreover also the ScGAS1 genecomplemented the Z bailii mutant Disruption ofGAS1 in Z bailii revealed its influence on the cell wallmorphology The GAS1 null mutants of Z bailiishowed the same phenotype reported for S cerevisiae[10] with enlarged cells often showing granular cyto-plasm an alterated chitin distribution two buds(Mickey Mouse-like appearance) and big vacuoles Thevacuole is known to be a dynamic organelle whosemorphology is highly responsive to different extracellu-lar and intracellular stimulistressors [19] In the GAS1null mutant it can be therefore speculated that thegeneration of big vacuoles is related to stressful condi-tions as already suggested by the altered chitin distri-bution being a possible defence mechanism againstlysis or related to a generalized response to sickness ofthe cells [10]Finally also the sensitivity to SDS and CFW
appeared very similar to what observed in the S cerevi-siae mutant A comparison of the different GAS1 nullphenotype mutants and the functionality of the Z bai-lii GAS1 homologue in S cerevisiae suggest a similarityof the cell wall structure of Z bailii compared with thecell wall structure of S cerevisiae Considering thesecretory pathway of a secreted protein it was specu-lated whether the protein becomes retarded in the cellwall after release from the plasma membrane The dis-ruption of GAS1 in Z bailii first of all confirmed ageneral phenomenon of protein release in respect to
the wild type strain as demonstrated by the higheraccumulation of total protein in the growing mediumof the deleted strain when compared to the parentalstrain Moreover such as compromised cell wall per-meability in the production strains for XPR2 and LIP1does lead to a clear improvement in the secretion ofthe respective proteins In the case of h IL-1b theenhancement of product secretion was not so evidentand difficult to interpret in respect to the higher accu-mulation of intracellular product It could be specu-lated that this can be related to the nature or to thesource of the protein but too few information areavailable at the moment to further comment in thisdirection What seems to be true is unfortunately whatwe very often experience for recombinant protein pro-duction which is the protein and protein-host depen-dence In fact also in P pastoris two out of threeheterologous proteins resulted more secreted in themutant as for Z bailii and when we tested the sameprotein in particular the IL-1b secretion in the S cere-visiae Δgas1 mutant (data not shown) we did not reg-ister any improvement
MethodsYeast strains media and transformationThe S cerevisiae strains used in this study were theauxotrophic haploid BY4741 (Mat a his3Δ1 leu2Δ0met5Δ0 ura3Δ0) and the relative null mutant in GAS1sequence called in this study BY4741 Δgas1 (Mat ahis3Δ1 leu2Δ0 met5Δ0 ura3Δ0 YMR307kanMX4)Both strains are commercially available from Euroscarfcollection httpwebuni-frankfurtdefb15mikroeuro-scarfcol_indexhtml The Z bailii wild type strain isATCC60483 Yeast cultures were grown in YP medium(10 g l-1 yeast extract 20 g l-1 peptone) with 2 or 5wv glucose (D) or fructose (F) as carbon sourceWhere needed NAT (80 mg l-1) or G418 (500 mg l-1
for S cerevisiae and 200 mg l-1 for Z bailii) or hygro-micine (100 mg l-1 for Z bailii) were added to themedia Alternatively yeast cultures were grown inminimal synthetic medium (067 w v-1 YNB Biolifewithout amino acids) with 2 or 5 w v-1 glucose ascarbon source When required supplements such ashistidine leucine methionine and uracil were added toa final concentration of 50 mg l-1 Media for plateswere solidified by addition of agar (Biolife Milan Italy)to 15 w v-1 Shake flask experiments were carriedout with 50 ml of medium in 250 ml shake flasks on ashaker at 30degC with 160 rpm For sensitivity tests inPetri dishes 125 μg ml-1 of CFW or 50 μg ml-1 SDSwere added to solid media Yeast transformation wasperformed using the Lithium AcetatessDNA methodas described [20] and slightly adapted for Z bailii asdescribed [3]
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 8 of 11
Genes amplifications deletion cassettes and expressionplasmid constructionPCR amplifications were made from yeasts genomicDNA prepared as described in [21] Amplifications wereperformed by the Pwo DNA Polymerase (Roche)The amplification of the ZbGAS1 gene was as follows
the primers GAS1int_fw 5rsquo-AGT TGT TCC AGA GATATT CCA TAC CTC AA-3rsquo and GAS1int_rev 5rsquo-GCAGAA CCG CTG AAG CTA CAG T-3rsquo designed oninternal conserved region of the available sequencesfrom other yeasts were firstly used to amplify an internalregion of 1137 bp of the Z bailii gene with the follow-ing program 94degC 5 min (94degC 15 s 57degC 30 s 72degC 1min) times 25 72degC 7 min 4degC infin After that the designedprimers ZbGAS1int_fw 5rsquo-GGA CGA TAA GGT TGACGA AGA-3rsquo and ZbGAS1int_rev 5rsquo-TGC TGT GGTCCA AAG TTG TG-3rsquo were used for the amplification(program 7 cycles 94degC 2 min 72degC 3 min 32 cycles94degC 2 min 67degC 3 min 67degC 4 min 4degC infin) of theterminal ends of ZbGAS1 utilizing a Z bailii GenomeWalker library constructed using the Universal GenomeWalker kit (Clontech) following the manufacturersinstructions From the obtained and reconstructedsequence the final primers were established and used forthe ZbGAS1 1710 bp full length coding sequence ampli-fication ZbGAS1_fw 5rsquo-ACT AAT GTT ATT CCAGGC GTT TTC G-3rsquo and ZbGAS1real_rev 5rsquo-AAATCA AGC CAA AGC AAA TCC AGC A-3rsquo (program94degC 5 min (94degC 20 s 58degC 30 s 72degC 2 min) times 30 72degC 10 min 4degC infin) The DNA sequence of ScGAS1 wasPCR amplified with the following primers ScGAS1fw5rsquo-ACA ATG TTG TTT AAA TCC CTT TCA AAGTTA GCA A-3rsquo and ScGAS1rev 5rsquo-TTT TTA AACCAA AGC AAA ACC GAC ACC AG-3rsquo (program 94degC5 min (94degC 20 s 605degC 30 s 72degC 2 min) times 30 72degC10 min 4degC infin) and the DNA sequence of YlXPR2 wasPCR amplified with the following primers XPR2_fwd5rsquo-ACA ATG AAG CTC GCT ACC GCC TTT A-3rsquoand XPR2_rev 5rsquo-TGC CTA AAT GCC AAC ACCGTT GTA G-3rsquo (program 94degC 4 min (94degC 15 s 625degC 30 s 72degC 2 min) times 30 72degC 7 min 4degC infin) All theunique fragments obtained were sub-cloned in the vec-tor pSTblue-1 utilising the Perfectly Bluntreg Cloning Kit(Novagene) and sequenced The resulting respectiveplasmids utilised in this work were named as followspSTbZbGAS1int pSTbZbGAS1 pSTbScGAS1 andpSTbYlXPR2For the construction of the ZbGAS1 deletion cassettes
the KanR or the hphR cassette derived from plasmidpFA6-KanMX4 [22] and pAG26 [23] and conferringresistance to G418 and hygromycin respectively werebluntended excised and inserted in the pSTbZbGAS1intopened BstEII BglII bluntended and dephosphorylatedresulting in the plasmids pSTbZbGAS1ΔK and
pSTbZbGAS1ΔH The resistance cassettes resulted inboth cases counter-clockwise inserted The Z bailii wildtype cells were transformed with the respective deletioncassettes obtained by cutting the two described plasmidsin the AccI and SnaBI sites The effective deletion wasPCR checked by using the following couples of primersZbG1int2_fw 5rsquo- TGC AGA AGT TAC AGA CCAATG TTG T -3rsquo and and ZbG1int2_rev 5rsquo-AAG ATAGAG AAG TGC TCT TGG CA -3rsquo (see Figure 2) orZbG1int2_fw and Hph_fw 5rsquo- ATA TGA AAA AGCCTG AAC TCA CCG AC -3rsquo or Kan_fw 5rsquo- ATG GGTAAG GAA AAG ACT CAC GTT -3rsquo (not shown) inorder to verify the presence of the cassettes in thedesired locusFor complementing the yeasts GAS1 deletion pZ5
(-Nco)ScGAS1 and pYX022ZbGAS1 were constructedpZ5(-Nco)ScGAS1 was obtained from pZ5(-Nco) openedSacIEcoRV with the insertion of ScGAS1 frompSTbScGAS1 cut with SacISnaBI The pZ5(-Nco)derives from pZ3 [3] where at the KpnI site the KanR
cassette was substituted from the NATR cassetteobtained from vector pAG25 [23] by cutting PvuIISacIblunt (obtaining the pZ5 plasmid) finally this plasmidwas EcoRIBamHI cut and bluntended and reclosedpYX022ZbGAS1 was obtained with the insertion ofZbGAS1 from pSTbZbGAS1 into pYX022 (RampD Sys-tems Wiesbaden Germany) both EcoRI cutFor heterologous protein expression the plasmids
pZLN022XPR2 pZLN022LIP1 and pZ5KlhIL-1b wereconstructed The first two integrative plasmids derivesfrom pZLN022 (manuscript in preparation) obtainedfrom pYX022 opened DraIII bluntSpeI blunt (toexclude HIS3) with the insertion of ZbLEU2 cut frompSTbZbLEU2 with EcoRI blunt in KpnI blunt site theNATR cassette was inserted obtained from vectorpAG25 [23] by cutting PvuIISacIblunt ThepSTbZbLEU2 was obtained by sub-cloning ZbLEU2amplified by PCR from Z bailii genomic DNA (manu-script in preparation) in pSTBlue (Novagen) cut byEcoRV For pZLN022XPR2 construction the pZLN022was AatIINheI cut and the XPR2 from pSTbXPR2 wasinserted For pZLN022LIP1 construction the pZLN022was AatII bluntNheI blunt opened and the LIP1 wasinserted by BamHI bluntNsiI blunt excision from thepGAPsLIP [24] For rh interleukin the pZ5KlhIL-1b wasbuilt it was obtained from pZ5 opened with EcoRI-blunt with the insertion of hIL-1b in frame with Kl sig-nal sequence from vector pKSSPI3 [25] cut EcoRIbluntXbaI bluntDNA manipulations transformation and cultivation of
Escherichia coli (DH5aFrsquo (80dlacZΔM15 Δ (lacZYA-argF) U169 deo rec1 end1 sup44 l THI-1 gyrA96relA1) and Novablue Competent Cells (Novagene) wereperformed following standard protocols [26] All the
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 9 of 11
restriction and modification enzymes used were fromNew England Biolabs (Hitchin Herts UK) or fromRoche Diagnostics (Mannheim Germany)Calcofluor staining and fluorescence microscopyCells were harvested washed in PBS and resuspended in1 mg ml-1 Calcofluor White solution incubated 10-30min at room temperature washed again twice in PBSFluorescence microscope images were taken by a NikonEclipse 90i microscope using the Metamorph softwareversion 22 (Nikon) using emission filter UV-1AQuantification of total released proteinsCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were etha-nol-precipitated The amount of proteins was deter-mined by the dye-binding method of Bradford (QuickStart Bradford Dye Reagent - BIORAD) using BSA asstandard All the data were normalized by OD after ver-ification of the linear correlation between OD and dryweight as a measure of biomassQuantification of secreted proteaseCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were sepa-rated and stored at -20degC until analysis The amount ofprotease secreted in the medium was determined by theAzocoll assay (Azocoll - Calbiochem) using commercialprotease as positive control Azocoll assays were per-formed according to [27] with some modificationsReaction mixtures contained 15 mg ml-1 Azocoll 50mM TrisHCl (pH 76) 150 mM NaCl and 5 mMCaCl2 Proteolytic activities were measured by absor-bance changes at 490 nm (Shimadzu UV 1601) afterincubation for 18 h at 30degCQuantification of secreted lipaseCultures were harvested and centrifuged at maximalspeed then the supernatants were separated and storedat -20degC until analysis The amount of lipase secreted inthe medium was determined according to [28] withsome modifications Reaction mixtures contained (A)60 mg of p-nitrophenyl palmitate (Sigma Aldrich) in 20ml of isopropanol (Sigma Aldrich) (B) 1 g Triton X-100(Sigma Aldrich) and 02 g of Arabic gum (SigmaAldrich) in 200 ml TrisHCl 01 M (pH 75) To obtainthe mixture (C) add slowly 1 part of A solution to 9part of B solution under continuous agitation Add 9part of (C) solution to one part of the enzyme contain-ing solution (ie the supernatant) Lipase activities weremeasured by absorbance changes at 410 nm (ShimadzuUV 1601) and the amount of enzyme is proportional toOD according to the relation Um = 0667 times OD410 nm
(if OD410 nm lt 05)Quantification of interleukinHuman IL-1b were analysed by Western blot Indepen-dent transformants were cultured in shake flasks inminimal or rich medium and during the kinetics
samples were collected Cells were harvested (a culturevolume corresponding to 108 cells) by centrifugationOne volume of 2timesLaemmli buffer [29] was added to thesupernatants of said samples they were boiled for 3-5min and stored at -20degC until loading or loaded directlyon a polyacrylamide gel For the analysis of intracellularproduction crude extracts were prepared from 108 cellsby following the trichloroacetic acid protocol [30] andresuspending the final protein extract in 150 μl ofLaemmli buffer Samples were then loaded on standardpolyacrylamide gels (sodium dodecyl sulfate-polyacryla-mide gel electrophoresis final concentration of theseparating gel 15) after separation proteins wereblotted to nitrocellulose membranes and immunodeco-rated An anti-IL-1b rabbit polyclonal antibody (IL-1b(H-153) Santa Cruz Biotechnology USA cat no sc-7884 dilution 1200) was used for the interleukin immu-nodecoration An anti-rabbit IgG horseradish peroxi-dase-conjugated (Amersham Pharmacia Biotech UK catno NA934 dilution 110000) secondary antibody wasused For the interleukin detection as a positive controlthe human recombinant IL-1b from E coli (Roche catno 1 457 756) was always loaded in parallel on thepolyacrylamide gels The proteins were visualised usingthe ECL Western blotting system (Amersham Bios-ciences UK) according to the manufacturerrsquosinstructions
Additional file 1 Clustalw_ntd Nucleotidic sequence of the ZbGAS1gene and alignment with the homologue ScGAS1 (entry YMR307W)Grey coloring represent nucleotidic identity Alignment was performedwith ClustalW software httpwwwebiacukclustalw and graphicalediting was made using Jalview software [31]Click here for file[ httpwwwbiomedcentralcomcontentsupplementary1475-2859-9-7-S1PDF ]
AcknowledgementsSpecial thanks to Valeria Longo and Davide Zocco for technicalcontributions and to Stefania Brocca for providing the pGAPsLIP plasmidThis work was partially supported by FAR 2007 and 2008 (12-1-192) to DP
Author details1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy 2CPC Biotech Srl Naples Italy
Authorsrsquo contributionsPB and DP initiated and coordinated the project LD was responsible forZbGAS1 gene identification by PCR and its deletion LR analyzed the mutantphenotype with CFW and SDS LR and SP performed the batch cultivationanalyzed the secretory phenotype in null mutant and performed theexperiment with heterologous proteins All authors wrote the paper andapproved the final version of the manuscript
Competing interestsThe authors declare that they have no competing interests
Received 2 November 2009Accepted 26 January 2010 Published 26 January 2010
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 10 of 11
References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11

Genes amplifications deletion cassettes and expressionplasmid constructionPCR amplifications were made from yeasts genomicDNA prepared as described in [21] Amplifications wereperformed by the Pwo DNA Polymerase (Roche)The amplification of the ZbGAS1 gene was as follows
the primers GAS1int_fw 5rsquo-AGT TGT TCC AGA GATATT CCA TAC CTC AA-3rsquo and GAS1int_rev 5rsquo-GCAGAA CCG CTG AAG CTA CAG T-3rsquo designed oninternal conserved region of the available sequencesfrom other yeasts were firstly used to amplify an internalregion of 1137 bp of the Z bailii gene with the follow-ing program 94degC 5 min (94degC 15 s 57degC 30 s 72degC 1min) times 25 72degC 7 min 4degC infin After that the designedprimers ZbGAS1int_fw 5rsquo-GGA CGA TAA GGT TGACGA AGA-3rsquo and ZbGAS1int_rev 5rsquo-TGC TGT GGTCCA AAG TTG TG-3rsquo were used for the amplification(program 7 cycles 94degC 2 min 72degC 3 min 32 cycles94degC 2 min 67degC 3 min 67degC 4 min 4degC infin) of theterminal ends of ZbGAS1 utilizing a Z bailii GenomeWalker library constructed using the Universal GenomeWalker kit (Clontech) following the manufacturersinstructions From the obtained and reconstructedsequence the final primers were established and used forthe ZbGAS1 1710 bp full length coding sequence ampli-fication ZbGAS1_fw 5rsquo-ACT AAT GTT ATT CCAGGC GTT TTC G-3rsquo and ZbGAS1real_rev 5rsquo-AAATCA AGC CAA AGC AAA TCC AGC A-3rsquo (program94degC 5 min (94degC 20 s 58degC 30 s 72degC 2 min) times 30 72degC 10 min 4degC infin) The DNA sequence of ScGAS1 wasPCR amplified with the following primers ScGAS1fw5rsquo-ACA ATG TTG TTT AAA TCC CTT TCA AAGTTA GCA A-3rsquo and ScGAS1rev 5rsquo-TTT TTA AACCAA AGC AAA ACC GAC ACC AG-3rsquo (program 94degC5 min (94degC 20 s 605degC 30 s 72degC 2 min) times 30 72degC10 min 4degC infin) and the DNA sequence of YlXPR2 wasPCR amplified with the following primers XPR2_fwd5rsquo-ACA ATG AAG CTC GCT ACC GCC TTT A-3rsquoand XPR2_rev 5rsquo-TGC CTA AAT GCC AAC ACCGTT GTA G-3rsquo (program 94degC 4 min (94degC 15 s 625degC 30 s 72degC 2 min) times 30 72degC 7 min 4degC infin) All theunique fragments obtained were sub-cloned in the vec-tor pSTblue-1 utilising the Perfectly Bluntreg Cloning Kit(Novagene) and sequenced The resulting respectiveplasmids utilised in this work were named as followspSTbZbGAS1int pSTbZbGAS1 pSTbScGAS1 andpSTbYlXPR2For the construction of the ZbGAS1 deletion cassettes
the KanR or the hphR cassette derived from plasmidpFA6-KanMX4 [22] and pAG26 [23] and conferringresistance to G418 and hygromycin respectively werebluntended excised and inserted in the pSTbZbGAS1intopened BstEII BglII bluntended and dephosphorylatedresulting in the plasmids pSTbZbGAS1ΔK and
pSTbZbGAS1ΔH The resistance cassettes resulted inboth cases counter-clockwise inserted The Z bailii wildtype cells were transformed with the respective deletioncassettes obtained by cutting the two described plasmidsin the AccI and SnaBI sites The effective deletion wasPCR checked by using the following couples of primersZbG1int2_fw 5rsquo- TGC AGA AGT TAC AGA CCAATG TTG T -3rsquo and and ZbG1int2_rev 5rsquo-AAG ATAGAG AAG TGC TCT TGG CA -3rsquo (see Figure 2) orZbG1int2_fw and Hph_fw 5rsquo- ATA TGA AAA AGCCTG AAC TCA CCG AC -3rsquo or Kan_fw 5rsquo- ATG GGTAAG GAA AAG ACT CAC GTT -3rsquo (not shown) inorder to verify the presence of the cassettes in thedesired locusFor complementing the yeasts GAS1 deletion pZ5
(-Nco)ScGAS1 and pYX022ZbGAS1 were constructedpZ5(-Nco)ScGAS1 was obtained from pZ5(-Nco) openedSacIEcoRV with the insertion of ScGAS1 frompSTbScGAS1 cut with SacISnaBI The pZ5(-Nco)derives from pZ3 [3] where at the KpnI site the KanR
cassette was substituted from the NATR cassetteobtained from vector pAG25 [23] by cutting PvuIISacIblunt (obtaining the pZ5 plasmid) finally this plasmidwas EcoRIBamHI cut and bluntended and reclosedpYX022ZbGAS1 was obtained with the insertion ofZbGAS1 from pSTbZbGAS1 into pYX022 (RampD Sys-tems Wiesbaden Germany) both EcoRI cutFor heterologous protein expression the plasmids
pZLN022XPR2 pZLN022LIP1 and pZ5KlhIL-1b wereconstructed The first two integrative plasmids derivesfrom pZLN022 (manuscript in preparation) obtainedfrom pYX022 opened DraIII bluntSpeI blunt (toexclude HIS3) with the insertion of ZbLEU2 cut frompSTbZbLEU2 with EcoRI blunt in KpnI blunt site theNATR cassette was inserted obtained from vectorpAG25 [23] by cutting PvuIISacIblunt ThepSTbZbLEU2 was obtained by sub-cloning ZbLEU2amplified by PCR from Z bailii genomic DNA (manu-script in preparation) in pSTBlue (Novagen) cut byEcoRV For pZLN022XPR2 construction the pZLN022was AatIINheI cut and the XPR2 from pSTbXPR2 wasinserted For pZLN022LIP1 construction the pZLN022was AatII bluntNheI blunt opened and the LIP1 wasinserted by BamHI bluntNsiI blunt excision from thepGAPsLIP [24] For rh interleukin the pZ5KlhIL-1b wasbuilt it was obtained from pZ5 opened with EcoRI-blunt with the insertion of hIL-1b in frame with Kl sig-nal sequence from vector pKSSPI3 [25] cut EcoRIbluntXbaI bluntDNA manipulations transformation and cultivation of
Escherichia coli (DH5aFrsquo (80dlacZΔM15 Δ (lacZYA-argF) U169 deo rec1 end1 sup44 l THI-1 gyrA96relA1) and Novablue Competent Cells (Novagene) wereperformed following standard protocols [26] All the
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 9 of 11
restriction and modification enzymes used were fromNew England Biolabs (Hitchin Herts UK) or fromRoche Diagnostics (Mannheim Germany)Calcofluor staining and fluorescence microscopyCells were harvested washed in PBS and resuspended in1 mg ml-1 Calcofluor White solution incubated 10-30min at room temperature washed again twice in PBSFluorescence microscope images were taken by a NikonEclipse 90i microscope using the Metamorph softwareversion 22 (Nikon) using emission filter UV-1AQuantification of total released proteinsCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were etha-nol-precipitated The amount of proteins was deter-mined by the dye-binding method of Bradford (QuickStart Bradford Dye Reagent - BIORAD) using BSA asstandard All the data were normalized by OD after ver-ification of the linear correlation between OD and dryweight as a measure of biomassQuantification of secreted proteaseCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were sepa-rated and stored at -20degC until analysis The amount ofprotease secreted in the medium was determined by theAzocoll assay (Azocoll - Calbiochem) using commercialprotease as positive control Azocoll assays were per-formed according to [27] with some modificationsReaction mixtures contained 15 mg ml-1 Azocoll 50mM TrisHCl (pH 76) 150 mM NaCl and 5 mMCaCl2 Proteolytic activities were measured by absor-bance changes at 490 nm (Shimadzu UV 1601) afterincubation for 18 h at 30degCQuantification of secreted lipaseCultures were harvested and centrifuged at maximalspeed then the supernatants were separated and storedat -20degC until analysis The amount of lipase secreted inthe medium was determined according to [28] withsome modifications Reaction mixtures contained (A)60 mg of p-nitrophenyl palmitate (Sigma Aldrich) in 20ml of isopropanol (Sigma Aldrich) (B) 1 g Triton X-100(Sigma Aldrich) and 02 g of Arabic gum (SigmaAldrich) in 200 ml TrisHCl 01 M (pH 75) To obtainthe mixture (C) add slowly 1 part of A solution to 9part of B solution under continuous agitation Add 9part of (C) solution to one part of the enzyme contain-ing solution (ie the supernatant) Lipase activities weremeasured by absorbance changes at 410 nm (ShimadzuUV 1601) and the amount of enzyme is proportional toOD according to the relation Um = 0667 times OD410 nm
(if OD410 nm lt 05)Quantification of interleukinHuman IL-1b were analysed by Western blot Indepen-dent transformants were cultured in shake flasks inminimal or rich medium and during the kinetics
samples were collected Cells were harvested (a culturevolume corresponding to 108 cells) by centrifugationOne volume of 2timesLaemmli buffer [29] was added to thesupernatants of said samples they were boiled for 3-5min and stored at -20degC until loading or loaded directlyon a polyacrylamide gel For the analysis of intracellularproduction crude extracts were prepared from 108 cellsby following the trichloroacetic acid protocol [30] andresuspending the final protein extract in 150 μl ofLaemmli buffer Samples were then loaded on standardpolyacrylamide gels (sodium dodecyl sulfate-polyacryla-mide gel electrophoresis final concentration of theseparating gel 15) after separation proteins wereblotted to nitrocellulose membranes and immunodeco-rated An anti-IL-1b rabbit polyclonal antibody (IL-1b(H-153) Santa Cruz Biotechnology USA cat no sc-7884 dilution 1200) was used for the interleukin immu-nodecoration An anti-rabbit IgG horseradish peroxi-dase-conjugated (Amersham Pharmacia Biotech UK catno NA934 dilution 110000) secondary antibody wasused For the interleukin detection as a positive controlthe human recombinant IL-1b from E coli (Roche catno 1 457 756) was always loaded in parallel on thepolyacrylamide gels The proteins were visualised usingthe ECL Western blotting system (Amersham Bios-ciences UK) according to the manufacturerrsquosinstructions
Additional file 1 Clustalw_ntd Nucleotidic sequence of the ZbGAS1gene and alignment with the homologue ScGAS1 (entry YMR307W)Grey coloring represent nucleotidic identity Alignment was performedwith ClustalW software httpwwwebiacukclustalw and graphicalediting was made using Jalview software [31]Click here for file[ httpwwwbiomedcentralcomcontentsupplementary1475-2859-9-7-S1PDF ]
AcknowledgementsSpecial thanks to Valeria Longo and Davide Zocco for technicalcontributions and to Stefania Brocca for providing the pGAPsLIP plasmidThis work was partially supported by FAR 2007 and 2008 (12-1-192) to DP
Author details1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy 2CPC Biotech Srl Naples Italy
Authorsrsquo contributionsPB and DP initiated and coordinated the project LD was responsible forZbGAS1 gene identification by PCR and its deletion LR analyzed the mutantphenotype with CFW and SDS LR and SP performed the batch cultivationanalyzed the secretory phenotype in null mutant and performed theexperiment with heterologous proteins All authors wrote the paper andapproved the final version of the manuscript
Competing interestsThe authors declare that they have no competing interests
Received 2 November 2009Accepted 26 January 2010 Published 26 January 2010
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 10 of 11
References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11

restriction and modification enzymes used were fromNew England Biolabs (Hitchin Herts UK) or fromRoche Diagnostics (Mannheim Germany)Calcofluor staining and fluorescence microscopyCells were harvested washed in PBS and resuspended in1 mg ml-1 Calcofluor White solution incubated 10-30min at room temperature washed again twice in PBSFluorescence microscope images were taken by a NikonEclipse 90i microscope using the Metamorph softwareversion 22 (Nikon) using emission filter UV-1AQuantification of total released proteinsCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were etha-nol-precipitated The amount of proteins was deter-mined by the dye-binding method of Bradford (QuickStart Bradford Dye Reagent - BIORAD) using BSA asstandard All the data were normalized by OD after ver-ification of the linear correlation between OD and dryweight as a measure of biomassQuantification of secreted proteaseCultures were harvested and centrifuged at maximalspeed then total proteins in the supernatants were sepa-rated and stored at -20degC until analysis The amount ofprotease secreted in the medium was determined by theAzocoll assay (Azocoll - Calbiochem) using commercialprotease as positive control Azocoll assays were per-formed according to [27] with some modificationsReaction mixtures contained 15 mg ml-1 Azocoll 50mM TrisHCl (pH 76) 150 mM NaCl and 5 mMCaCl2 Proteolytic activities were measured by absor-bance changes at 490 nm (Shimadzu UV 1601) afterincubation for 18 h at 30degCQuantification of secreted lipaseCultures were harvested and centrifuged at maximalspeed then the supernatants were separated and storedat -20degC until analysis The amount of lipase secreted inthe medium was determined according to [28] withsome modifications Reaction mixtures contained (A)60 mg of p-nitrophenyl palmitate (Sigma Aldrich) in 20ml of isopropanol (Sigma Aldrich) (B) 1 g Triton X-100(Sigma Aldrich) and 02 g of Arabic gum (SigmaAldrich) in 200 ml TrisHCl 01 M (pH 75) To obtainthe mixture (C) add slowly 1 part of A solution to 9part of B solution under continuous agitation Add 9part of (C) solution to one part of the enzyme contain-ing solution (ie the supernatant) Lipase activities weremeasured by absorbance changes at 410 nm (ShimadzuUV 1601) and the amount of enzyme is proportional toOD according to the relation Um = 0667 times OD410 nm
(if OD410 nm lt 05)Quantification of interleukinHuman IL-1b were analysed by Western blot Indepen-dent transformants were cultured in shake flasks inminimal or rich medium and during the kinetics
samples were collected Cells were harvested (a culturevolume corresponding to 108 cells) by centrifugationOne volume of 2timesLaemmli buffer [29] was added to thesupernatants of said samples they were boiled for 3-5min and stored at -20degC until loading or loaded directlyon a polyacrylamide gel For the analysis of intracellularproduction crude extracts were prepared from 108 cellsby following the trichloroacetic acid protocol [30] andresuspending the final protein extract in 150 μl ofLaemmli buffer Samples were then loaded on standardpolyacrylamide gels (sodium dodecyl sulfate-polyacryla-mide gel electrophoresis final concentration of theseparating gel 15) after separation proteins wereblotted to nitrocellulose membranes and immunodeco-rated An anti-IL-1b rabbit polyclonal antibody (IL-1b(H-153) Santa Cruz Biotechnology USA cat no sc-7884 dilution 1200) was used for the interleukin immu-nodecoration An anti-rabbit IgG horseradish peroxi-dase-conjugated (Amersham Pharmacia Biotech UK catno NA934 dilution 110000) secondary antibody wasused For the interleukin detection as a positive controlthe human recombinant IL-1b from E coli (Roche catno 1 457 756) was always loaded in parallel on thepolyacrylamide gels The proteins were visualised usingthe ECL Western blotting system (Amersham Bios-ciences UK) according to the manufacturerrsquosinstructions
Additional file 1 Clustalw_ntd Nucleotidic sequence of the ZbGAS1gene and alignment with the homologue ScGAS1 (entry YMR307W)Grey coloring represent nucleotidic identity Alignment was performedwith ClustalW software httpwwwebiacukclustalw and graphicalediting was made using Jalview software [31]Click here for file[ httpwwwbiomedcentralcomcontentsupplementary1475-2859-9-7-S1PDF ]
AcknowledgementsSpecial thanks to Valeria Longo and Davide Zocco for technicalcontributions and to Stefania Brocca for providing the pGAPsLIP plasmidThis work was partially supported by FAR 2007 and 2008 (12-1-192) to DP
Author details1University of Milano-Bicocca Department of Biotechnology and BioscienceMilan Italy 2CPC Biotech Srl Naples Italy
Authorsrsquo contributionsPB and DP initiated and coordinated the project LD was responsible forZbGAS1 gene identification by PCR and its deletion LR analyzed the mutantphenotype with CFW and SDS LR and SP performed the batch cultivationanalyzed the secretory phenotype in null mutant and performed theexperiment with heterologous proteins All authors wrote the paper andapproved the final version of the manuscript
Competing interestsThe authors declare that they have no competing interests
Received 2 November 2009Accepted 26 January 2010 Published 26 January 2010
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 10 of 11
References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11

References1 Branduardi P Molecular cloning and sequence analysis of the
Zygosaccharomyces bailii HIS3 gene encoding the imidazoleglycerolphosphate dehydratase Yeast 2002 19(13)1165-1170
2 Thomas DS Davenport RR Zygosaccharomyces bailii - a profile ofcharacteristics and spoilage activities Food Microbiology 19852(2)157-169
3 Branduardi P Valli M Brambilla L Sauer M Alberghina L Porro D The yeastZygosaccharomyces bailii a new host for heterologous proteinproduction secretion and for metabolic engineering applications FEMSYeast Res 2004 4(4-5)493-504
4 Chow T Ash J Dignard D Thomas D Screening and identification of agene PSE-1 that affects protein secretion in Saccharomyces cerevisiae JCell Sci 1992 101(Pt 3)709-719
5 Bartkeviciūte D Sasnauskas K Disruption of the MNN10 gene enhancesprotein secretion in Kluyveromyces lactis and Saccharomyces cerevisiaeFEMS Yeast Res 2004 4(8)833-840
6 Bankaitis V Aitken J Cleves A Dowhan W An essential role for aphospholipid transfer protein in yeast Golgi function Nature 1990347(6293)561-562
7 Vai M Brambilla L Orlandi I Rota N Ranzi B Alberghina L Porro DImproved secretion of native human insulin-like growth factor 1 fromgas1 mutant Saccharomyces cerevisiae cells Appl Environ Microbiol 200066(12)5477-5479
8 Popolo L Vai M The Gas1 glycoprotein a putative wall polymer cross-linker Biochim Biophys Acta 1999 1426(2)385-400
9 Papaleo E Fantucci P Vai M De Gioia L Three-dimensional structure ofthe catalytic domain of the yeast beta-(13)-glucan transferase Gas1 amolecular modeling investigation J Mol Model 2006 12(2)237-248
10 Popolo L Vai M Gatti E Porello S Bonfante P Balestrini R Alberghina LPhysiological analysis of mutants indicates involvement of theSaccharomyces cerevisiae GPI-anchored protein gp115 in morphogenesisand cell separation J Bacteriol 1993 175(7)1879-1885
11 Kapteyn J Ram A Groos E Kollar R Montijn R Ende Van Den H Llobell ACabib E Klis F Altered extent of cross-linking of beta16-glucosylatedmannoproteins to chitin in Saccharomyces cerevisiae mutants withreduced cell wall beta13-glucan content J Bacteriol 1997179(20)6279-6284
12 Marx H Sauer M Resina D Vai M Porro D Valero F Ferrer PMattanovich D Cloning disruption and protein secretory phenotype ofthe GAS1 homologue of Pichia pastoris FEMS Microbiol Lett 2006264(1)40-47
13 James S Collins M Roberts I Genetic interrelationship among species ofthe genus Zygosaccharomyces as revealed by small-subunit rRNA genesequences Yeast 1994 10(7)871-881
14 Mollapour M Piper P Targeted gene deletion in Zygosaccharomyces bailiiYeast 2001 18(2)173-186
15 Popolo L Gilardelli D Bonfante P Vai M Increase in chitin as an essentialresponse to defects in assembly of cell wall polymers in the ggp1deltamutant of Saccharomyces cerevisiae J Bacteriol 1997 179(2)463-469
16 Carotti C Ferrario L Roncero C Valdivieso M Duran A Popolo LMaintenance of cell integrity in the gas1 mutant of Saccharomycescerevisiae requires the Chs3p-targeting and activation pathway andinvolves an unusual Chs3p localization Yeast 2002 19(13)1113-1124
17 Turchini A Ferrario L Popolo L Increase of external osmolarity reducesmorphogenetic defects and accumulation of chitin in a gas1 mutant ofSaccharomyces cerevisiae J Bacteriol 2000 182(4)1167-1171
18 Ferrer P Diers I Asenjo J Andrews B Yeast cell permeabilizing beta-13-glucanases A tool for the integration of downstream processes andmetabolic engineering applications to yeast Biotechnol Bioeng 1998 58(2-3)321-324
19 Li S Kane P The yeast lysosome-like vacuole endpoint and crossroadsBiochim Biophys Acta 2009 1793(4)650-663
20 Gietz R Woods R Transformation of yeast by lithium acetatesingle-stranded carrier DNApolyethylene glycol method Methods Enzymol2002 35087-96
21 Hoffman C Winston F A ten-minute DNA preparation from yeastefficiently releases autonomous plasmids for transformation ofEscherichia coli Gene 1987 57(2-3)267-272
22 Wach A Brachat A Poumlhlmann R Philippsen P New heterologous modulesfor classical or PCR-based gene disruptions in Saccharomyces cerevisiaeYeast 1994 10(13)1793-1808
23 Goldstein A McCusker J Three new dominant drug resistance cassettesfor gene disruption in Saccharomyces cerevisiae Yeast 199915(14)1541-1553
24 Brocca S Schmidt-Dannert C Lotti M Alberghina L Schmid R Design totalsynthesis and functional overexpression of the Candida rugosa lip1gene coding for a major industrial lipase Protein Sci 1998 7(6)1415-1422
25 Camattari A Bianchi M Branduardi P Porro D Brambilla L Induction byhypoxia of heterologous-protein production with the KlPDC1 promoterin yeasts Appl Environ Microbiol 2007 73(3)922-929
26 Sambrook J Maniatis T Fritsch EF Molecular cloning a laboratory manualCold Spring Harbor NY Cold Spring Harbor Laboratory 2 1989
27 Chavira RJ Burnett T Hageman J Assaying proteinases with azocoll AnalBiochem 1984 136(2)446-450
28 Brocca S Persson M Wehtje E Adlercreutz P Alberghina L Lotti MMutants provide evidence of the importance of glycosydic chains in theactivation of lipase 1 from Candida rugosa Protein Sci 2000 9(5)985-990
29 Laemmli U Cleavage of structural proteins during the assembly of thehead of bacteriophage T4 Nature 1970 227(5259)680-685
30 Surana U Amon A Dowzer C McGrew J Byers B Nasmyth K Destructionof the CDC28CLB mitotic kinase is not required for the metaphase toanaphase transition in budding yeast EMBO J 1993 12(5)1969-1978
31 Waterhouse A Procter J Martin D Clamp M Barton G Jalview Version 2ndashamultiple sequence alignment editor and analysis workbenchBioinformatics 2009 25(9)1189-1191
doi1011861475-2859-9-7Cite this article as Passolunghi et al Cloning of the Zygosaccharomycesbailii GAS1 homologue and effect of cell wall engineering on proteinsecretory phenotype Microbial Cell Factories 2010 97
Submit your next manuscript to BioMed Centraland take full advantage of
bull Convenient online submission
bull Thorough peer review
bull No space constraints or color figure charges
bull Immediate publication on acceptance
bull Inclusion in PubMed CAS Scopus and Google Scholar
bull Research which is freely available for redistribution
Submit your manuscript at wwwbiomedcentralcomsubmit
Passolunghi et al Microbial Cell Factories 2010 97httpwwwmicrobialcellfactoriescomcontent917
Page 11 of 11
Copyright © 2022 FDOKUMEN




















