
The Calcineurin A homologue from Trypanosoma cruzi lacks two important regulatory domains
Upload
independentCategory
view
2download
0
www.bba-direct.com
Biochimica et Biophysica Acta 1676 (2004) 193–202
Short sequence paper
Cloning and characterization of the promoter region of the human
prostaglandin F2a receptor gene
Dean B. Zaragozaa,*, Robyn Wilsona, Kathleen Eysterb, David M. Olsona,c,d
aPerinatal Research Centre, CIHR Group in Perinatal Health and Disease, Department of Obstetrics and Gynecology, University of Alberta,
Edmonton, AB T6G 2S2, CanadabSystems Physiology and Structural Biology Research Group, Division of Basic Biomedical Sciences, University of South Dakota School of Medicine,
Vermillion, SD, USAcDepartment of Pediatrics, University of Alberta, Edmonton, AB, CanadadDepartment of Physiology, University of Alberta, Edmonton, AB, Canada
Received 6 August 2003; received in revised form 22 October 2003; accepted 7 November 2003
Abstract
The promoter region of the human prostaglandin F2a receptor (FP) gene was isolated, sequenced, and characterized. The 5V-flanking regionhas minimal homology to the bovine, mouse, and rat FP promoters with the exception of a 100-bp region. The human promoter similarly lacks
a canonical TATA-box and a CAAT-box. Potential binding sites for SP-1, GATA-1, STAT-1, and AP-1 are present in the 5V-flanking region.
One major transcription start site was identified using 5V RLM-RACE analysis and mapped to an adenine residue 262 nucleotides upstream
from the initiator codon in exon 2. Transfection of HeLa cells with FP promoter-GFP deletion constructs indicates that the � 2437/� 1946
region contains repressor activity. DNase I footprinting analysis of this region identifies a footprint over the GATA-like site at � 2400. This
suggests repression of basal FP transcription may be mediated by a GATA binding site.
D 2003 Elsevier B.V. All rights reserved.
Keywords: Human FP promoter; Prostaglandin F2a; Transcriptional regulation
In human pregnancy, prostaglandin (PG)F2a is recognized
as one of the ‘‘triggers’’ of labor [1] because the myometrium
contracts in response to exogenous PGF2a, in vivo and in
vitro [2,3]. PGF2a acts through binding to its own plasma
membrane receptor termed the PGF2a receptor (FP). FP is a
member of a group of proteins collectively known as uterine
activation proteins (UAPs) that are involved in preparing the
uterus for the contractions of labor. As such, FP is believed to
play an important role in the maintenance and termination of
pregnancy and in women. Current data suggest that FP
expression in the human uterus is suppressed during preg-
nancy resulting in relaxation of the uterus and pregnancy
maintenance [4]. Increases in FP expression at the time of
labor are believed to contribute to uterine contraction and the
termination of pregnancy [5]. Understanding how FP is
regulated at the transcriptional level may be critical to our
understanding of how pregnancy is maintained and subse-
0167-4781/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
doi:10.1016/j.bbaexp.2003.11.004
* Corresponding author. Tel.: +1-780-492-0029; fax: +1-780-492-
1308.
E-mail address: [email protected] (D.B. Zaragoza).
quently terminated resulting in expulsion of the baby. Clues
to this regulation can be obtained from studying the FP
promoter.
FP is encoded by a single gene and is a member of the
seven-transmembrane domain receptor superfamily of G-
protein coupled receptors. The human, bovine, mouse and rat
FP gene structure has been well characterized and consists of
three exons and two introns [6–9]. Transcription initiation
sites for the FP gene have been identified for the rat, mouse,
and bovine gene. In the mouse, several sites were identified
between 222 and 274 bp upstream of the translational start
codon with the major start site 232 bp upstream of the ATG
start codon [7]. In the rat, tissue-specific transcription start
sites were identified in intron 1 as well as the typical region
f 263 bp upstream of the start ATG [9]. The bovine gene
also contains multiple initiation sites in exon 1 ranging from
98 to 265 bp upstream of ATG. Similar to the rat gene, the
bovine gene has transcription initiation sites located in intron
1 [8]. Transcription start sites have not yet been identified in
the human FP gene. In contrast to the gene, the FP promoter
is not as well characterized. Four groups have examined
sequence from the 5V flanking region of the rat [9], mouse

D.B. Zaragoza et al. / Biochimica et Biophysica Acta 1676 (2004) 193–202194
[7], bovine [8] and human [6] FP gene. A DNA fragment
containing 2590 bp of 5V flanking sequence of the rat FP
gene has been tested for regulating both basal and IL-6
mediated FP transcription. Deletion analysis identified both
positive and negative regulatory regions within this fragment
[9]. A fragment containing 1500 bp of 5V flanking region of
the mouse FP gene has been tested for promoter activity and
found to regulate transcription in a tissue dependent manner
[7], however, no specific regions or cis-acting elements were
shown to be important for regulating FP gene transcription.
Two regions of the bovine FP gene were examined for
promoter activity, 1818 bp upstream of exon 1 and 1086
bp upstream of exon 2 [8]. Again, while the region upstream
of exon 1 was found to be responsive to phorbol ester and
contain several putative transcription factor binding sites, no
specific regions or cis-acting elements were directly linked to
transcriptional regulation. While 370 bp of 5V-flankingregion of the human FP gene has been reported [6], this
region was not tested for promoter activity and no transcrip-
tion initiation start site or transcription factor binding sites
were identified. Furthermore, the authors do not suggest that
this sequence contains the entire FP promoter. It is likely that
regulatory elements could be located further upstream of the
gene and within intron 1. Crucial to the understanding of the
regulation of this molecule across human pregnancy is the
understanding of the transcriptional regulation of the gene.
Currently, very little is known about the human FP promoter.
The purpose of this study was to identify regions in the 5V-flanking sequence of the human FP gene important for
transcriptional regulation. To this end, we isolated and
characterized a larger region of 5V flanking sequence of
the human FP gene than previously reported, and examined
its ability to regulate transcription of a GFP reporter con-
struct. Upon identifying functional regions of the promoter,
we examined these regions for protein binding activity by
DNase I footprinting analysis.
The transcription initiation site of the human FP gene was
determined by Ambion’s First Choice 5V RLM-RACE
method. Fig. 1 shows the results of the analysis. A 258-bp
fragment was obtained from nested PCR reactions using the
5V RACE inner primer/hFP nested reverse primer pair (Table
1) and cDNA derived from tobacco acid pyrophosphatase
(TAP) treated myometrial RNA (Fig. 1A, lane 1). No product
was obtained when RNA was not treated with TAP (lane 2)
indicating that the 258-bp product was derived from full-
length mRNA. Lanes 3 and 4 contain nested PCR reactions
using mouse CXCR-4 primers (Table 1, 5V RACE Control
Primers) to serve as a control to validate the assay and the
expected 301-bp product was obtained in the + TAP reac-
tions (lane 3). A no-template control PCR reaction using the
hFP-RACE primers was also included in lane 5. After
cloning the 258-bp FP PCR product into pGEM-T, 20 clones
were sequenced using the hFP nested reverse primer. Of the
20 clones only five (clones 1, 2, 6, 12, and 17) contained the
adapter sequence and all of these clones mapped the 5V end
of the mRNA to an adenine 262 nucleotides upstream of the
ATG initiator codon (Fig. 1B). Eleven clones contained hFP
sequence but no adapter sequence, and for that reason we did
not believe these to contain true 5V ends. These were most
likely the result of incomplete reverse transcription. The
other four clones contained nonsense sequence containing
repeats of the hFP nested reverse primer. These clones
contained no actual FP sequence. We believe these clones
to have resulted from primer dimer artifacts of the PCR
reactions. Because our 5V RLM-RACE method gave rise to 5
clones out of 20, it is possible that we may have missed a
small population of minor transcription start sites upstream
of adenine � 265. Indeed, minor transcription start sites
have been identified upstream of the major site in the mouse,
rat, and bovine FP gene. To test for this possibility in the
human FP gene, we performed conventional RT-PCR using
two sense primers (one upstream of the adenine at � 265 and
one downstream) and one common antisense primer (hFP
nested reverse primer). Fig. 1C shows the results. Both RT-
PCR reactions using either sense primer gave rise to the
expected size PCR products. This indicates that a small
population of transcription start sites exists upstream of
adenine � 262 that was missed by our RLM 5V RACEanalysis.
Our RLM 5V RACE results are in agreement with the
major transcription initiation sites mapped for the rat (270 bp
upstream of ATG), mouse (232 bp upsteam of ATG), and
bovine (265 bp upstream of ATG) FP gene [7–9]. In addition
to minor upstream start sites, these studies also report
multiple downstream transcription initiation sites which
may have resulted from incomplete reverse transcription or
degraded FP mRNA. By only examining clones that
contained the adapter sequence, we ensure that our clones
resulted from the RT reactions which fully extended to the
true 5V end of the human FP mRNA. The RLM 5V RACE
technique also controls for false-positives resulting from
degraded mRNA.
In addition to the major transcription start sites loca-
ted f 260 bp upstream of the ATG initiator codon, the rat
and bovine FP genes have additional transcription start sites
that map to intron 1 [7,9]. The authors attribute their findings
to the existence of alternate promoter regions which may be
tissue-specific. While our analysis suggests that only one
promoter is operating in human myometrial tissue, alternate
tissue-specific promoters may still exist. An analysis of
human FP transcription initiation sites in other tissues may
reveal initiation sites in intron 1 and the existence of tissue-
specific promoters for the human FP gene.
The 5V-flanking region of the human FP gene was
isolated and cloned using a Genome Walking protocol
(Clonetech, Palo Alto, CA, USA). Using this protocol, we
obtained a 4106-bp fragment containing the first 33 bp of
exon 2, a 1450-bp intron 1, a 187-bp exon 1, and 2436 bp
upstream of the transcription start site (Fig. 2). Analysis of
putative binding sites for transcription factors was carried
out using MatInspector V2.2 [10]. As shown in Fig. 2, the
sequence lacks a TATA-box and a CAAT-box, as is the case

Fig. 1. Identification of the transcription initiation start site of the human FP gene by 5V RLM-RACE. (A) Nested PCR products from 5V RLM RACE analysis
separated by agarose electrophoresis. Calf intestinal phosphatase (CIP)-treated RNA isolated from human myometrial tissue (lanes 1 and 2) or control mouse
thymus tissue (lane 3 and 4) was left untreated (lane 2 and 4) or treated with tobacco acid phosphatase (TAP) (lanes 1 and 3). Following reverse transcription
reactions and primary PCR reactions, nested PCR reactions were performed using the 5V RACE Inner primer/hFP Nested Reverse primer pair (lanes 1 and 2) or
5V RACE Inner primer/5V RACE Inner control primer. No-template reactions (NT) were performed using the 5V RACE Inner primer/hFP Nested Reverse
primer pair (lane 5). Samples were run against a 123-bp ladder (lane 6). (B) hFP 5V RLM-RACE products were cloned into pGEM-T and sequenced. Sequence
results of the five clones that were positive for the 5V RACE adapter (5V RACE Inner Primer, boxed region) are shown. All positive clones map the 5V end of
human FP mRNA to an adenine residue 262 base pairs upstream of start ATG (designated by an arrowhead). (C) Conventional RT-PCR on RNA isolated from
human myometrial tissue. RT-PCR was performed using either a sense primer located upsteam of adenine-262 (Table 1, upstream primer) or a sense primer
located downstream of adenine-262 (hFP-GFP-f1) with a common antisense primer (hFP 1rst Reverse). Water controls for each primer pair are also included.
D.B. Zaragoza et al. / Biochimica et Biophysica Acta 1676 (2004) 193–202 195
for the murine and rat FP promoters [7,9]. Of the two
potential bovine FP promoters previously identified, the
upstream promoter is also TATA-less but the promoter
localized to intron 1 did contain a TATA-like sequence
and a CAAT-like box [8]. Although the human FP promoter
contains no conventional TATA sequence or CAAT se-
quence immediately upstream of the transcription start site,
there are two SP-1/GC elements located within 100 bp of
the transcriptional start site at positions � 114 and � 86
(Fig. 2). These GC box elements may explain the lack of a
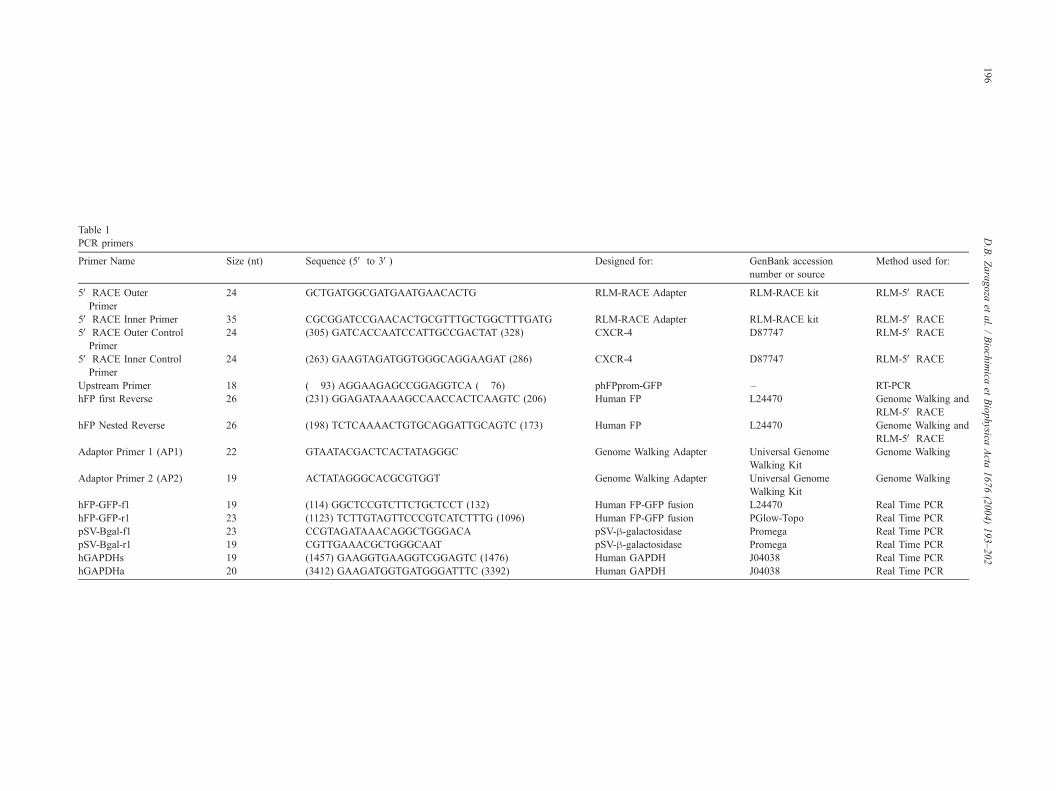
Table 1
PCR primers
Primer Name Size (nt) Sequence (5V to 3V) Designed for: GenBank accession
number or source
Method used for:
5V RACE Outer
Primer
24 GCTGATGGCGATGAATGAACACTG RLM-RACE Adapter RLM-RACE kit RLM-5V RACE
5V RACE Inner Primer 35 CGCGGATCCGAACACTGCGTTTGCTGGCTTTGATG RLM-RACE Adapter RLM-RACE kit RLM-5V RACE
5V RACE Outer Control
Primer
24 (305) GATCACCAATCCATTGCCGACTAT (328) CXCR-4 D87747 RLM-5V RACE
5V RACE Inner Control
Primer
24 (263) GAAGTAGATGGTGGGCAGGAAGAT (286) CXCR-4 D87747 RLM-5V RACE
Upstream Primer 18 (� 93) AGGAAGAGCCGGAGGTCA (� 76) phFPprom-GFP – RT-PCR
hFP first Reverse 26 (231) GGAGATAAAAGCCAACCACTCAAGTC (206) Human FP L24470 Genome Walking and
RLM-5V RACE
hFP Nested Reverse 26 (198) TCTCAAAACTGTGCAGGATTGCAGTC (173) Human FP L24470 Genome Walking and
RLM-5V RACE
Adaptor Primer 1 (AP1) 22 GTAATACGACTCACTATAGGGC Genome Walking Adapter Universal Genome
Walking Kit
Genome Walking
Adaptor Primer 2 (AP2) 19 ACTATAGGGCACGCGTGGT Genome Walking Adapter Universal Genome
Walking Kit
Genome Walking
hFP-GFP-f1 19 (114) GGCTCCGTCTTCTGCTCCT (132) Human FP-GFP fusion L24470 Real Time PCR
hFP-GFP-r1 23 (1123) TCTTGTAGTTCCCGTCATCTTTG (1096) Human FP-GFP fusion PGlow-Topo Real Time PCR
pSV-Bgal-f1 23 CCGTAGATAAACAGGCTGGGACA pSV-h-galactosidase Promega Real Time PCR
pSV-Bgal-r1 19 CGTTGAAACGCTGGGCAAT pSV-h-galactosidase Promega Real Time PCR
hGAPDHs 19 (1457) GAAGGTGAAGGTCGGAGTC (1476) Human GAPDH J04038 Real Time PCR
hGAPDHa 20 (3412) GAAGATGGTGATGGGATTTC (3392) Human GAPDH J04038 Real Time PCR
D.B.Zaragoza
etal./Biochimica
etBiophysica
Acta
1676(2004)193–202
196

GC/SP-1 GC/SP-1
Adapter Primer 2
NF-IL-6
NFκB
NF-IL-6
NF B
AP-1
AP-1
AP-1
AP-1
AP-1 (-)
AP-1 (-)
CRE
GATA (-)
GATA
GATA (-)
GATA (-)
GATA
GATA
STAT
YGATG (-)
ACTATAGGGC ACGCGTGGTC GACGGCCCGG GCTGGTCCTC AGAAATAACA TCACACATCT ACAACCATCT GATCTTCAAC AAAACTGACA AAAACAAGCA
ATGGAGAAAG GATGTCCTAT TTAATAAATG ATGTTGGGAA AACTGGCTAG CCATATGCAA AAAACTGAAA CTGGACCCCA TTCCTTATAC CTTATACAAA
AATTAACTCA AGATGGATTA AAGACTTAAA TGTAAAACCT AAAATAATAA AAACTCTAGA AAAAAACATA GGTAATACCA TTCAGGACAT AGGCATGGGC
ATAGACTTTA TGACAAAAAC ACAAAAAACA ATGACGACAA AAACACACAA AAAAATGACA ACAAAGGCCA AAATTGACAC ATGGGGTCTA ATTAAACTAA
AGAACTTCTG TACAGCAAAA GAAACTATCA TCGGAGTGAA CAGGCAACCT AGAGAATGGG AGAAATACCT AATGTAGATG ATGGGTTGAC CACCATGGCA
CATGTATACC TATGCACATG TATCTCAGAA CTTAAAGTAT AATAATTTAA AAAAAGTTAA AATAAAATAT TAATGACTTT CAAAGAAAAA AAAAGTCAAG
AAACAATAGA TGCTGGCGAG GCTATGGAGA AATAGGAATG CTTTTACACT GTTGGTAGGA ATGTAAATTA GTTCAACCAA TATGGAAGAC GGCATGGTGA
TTCCTCAAGG ATCTAGAACC AGAAATATCA TTTGATCCAG CAATCCCATT ACTGGGTATA TACCCAAAGG AATACAAATC ATTCTATTAT GAAGACACAT
GCACACATGT GTTTACTGAA GCACTATTTA CAATAGCAAA GTCATAGAAC CAACCCAAAT GCCCATCAAT AATAGACTGG ATAAAGAAAA TGTGGTACAT
ATACACCACG GAATACTACA CAGCCATAAA AAGGAATGGG ATCATGTCCT TTGCAGGGAC ATGGATGAAC CTGGATGCTA TCATCCTCAG CAAACTAGCA
CAGGAACAGA AAACCAAACA CTGCATGTTT TCACTTATAA ATGGGAGTTC AGCAATGAGA ACACATGGAC ACAGGGAGAG GAACAACATG CACCAGGGCC
TGTTAGGGGG TGGGGGGCAG GGGGAGGGAA CTTAGAGGAC AGGTCAATAG GTGCAAAAAT TTCATGTTTT TTAAAAAAGA AAAAAATTAT ATATGCACTC
ACATATTTTA AAAAATAAAG GAAAGAATAG TACTCTGTGT CATTCATATT TTCAGTATAG CTTATCACAT AATTGCCTTT TGGATACTGG CCATTCATGT
ATAATATTTC TCTAGTAACC AGGTTATCTA ACCAAAATGT TTAATTCCCT CACCATAATT AAGTAACAGT TTAGTTTACC AGTTATGATC ATATCATTTA
AAAAAATCTC AATATCTACC ACAAGGGAAA ACTGCAATCA TTCCTTTCTA CAGTTAATGG CTAATTTATT TTATCCCGCT TTTTCATTTT TTGATCCCTT
CCAATTTCCA GGAATGTTAG AGGTCCCTTC TGGTTTTAGA ACGGAGAAAC AGGGTTCGTT TCCATGTGGT ACTGATTTTT CACATAGGTA GTGGCAACAT
TTTACTACAA TAAAAAGAGC AAAAGTCAAT AATAGATTCA CATAAGTGTT TAGCTACAAA AATGAGACAC AGCTAATGGG TGTGTGTCGA AGCAATATTA
GATAAAAGCA ATTACAGAAT AAAATGGCTC AAATCGTAAT TCCTAATGAA AAATAGATTA CTTCATCATA AATTTATCTC CACGAAATTT ATTAGACGCA
TATCTCTTGG GGTATACATA TCACTAGAAA TTCTCTCTCC AGTGGACTTC TAAAATCACT GAAATATTAC TAATCCCTTA TCTGCAGGTG TACTTCAAGA
AGTCATCTCA TCCCTTTTCC TTTTCATATT CCTTAGAAAG CTGGAAACAG TCCCAGTCCT CTGTGTGGCA GTGAACCCTG AGGCTACGGG TGTTTTCATT
TACAATACTG GGCTTGAATA CCTGTTTTTA ATTCCCTGGA GATATTTGCC ACAAAATGGC TCCCTCAAGC ACGCAACCTC TTTCTACCTG GTTTCTTTAT
GCGTTTCGAG CTGGACTTGG AATGAATTGT CTGCAAGCTA CCATCCGACA GCCTAACACA CCATCTTAGG CTTTCCTCGG CTCTCCAGAC TGCTGTCTTG
CATCCCCAGA GATGGCTTGG GGGATAAAAA TGAGTTTAAA ACCAAACTTT CTTTAGCTGG TCCAGCGGCT AAGTGCAGCT CTGATGCATC CGAGCACGGT
GCCCTCTTCT CTCCCAGTAC TGCTCTGTGC CCATGGAAAA GGGGGCGGAG GCGAGAGAGA AGAGGAAGAG GCGGAGGTCA CTCGCGCGCC CCATCCCTCC
CAGGCTGCCA GCGCAGCTAG GTGCAGAGGG ATCCCAGGAG CCGCGCGCGC CCCGCAGTTT CCGCGCTAAG GGAACGAGTG CGCGGAGGGG ACGAGCGGCT
GGACCACAGC CGGCGCCCGA TCAGGATCTC CGCGCTGGGA TCGGTGGAAC TTGAGGCAGC GGCGGCGCGG GGCGCCATGG CACACCGAGC GGCTCCGTCT
TCTGCTCCTC AGAGAGCCCG GCTGGCGGCC TGGGATGACA AGGTACCATC CCTCCAGAGG CTGATCCCAA TGCATCGGCT TCGCTTATCT AGACCCAGAT
TTAATCAACT CACCCCCTGA GACACCCAGC CACAGCTTCT GCTGGCTGAG CCTGAGCACT GAGTTTGGAC CAGATTCCCC AAGCTAGGGA GACGCATAGA
GAAAGAAGGG TACTCGAGCT AACCCATCCC CATTCTGAGC TGCTCTGCGG GGGATTGGGT AGAAAGGGTC TCAGAAGAGG GAAATCGGCA CAGATTCCGC
TTCTCGCCGT TGCGGGGTTC CCTGCGGGCG GCACTGCGAG TTCCTCACCC GGGCACGTCC TCAACTCCCC GTGCCCGGGA ATGAGGGAGA AAACAGCCCT
TTGGCGTTTG AGCAATTGCC AGAAAGTTTC AGCGAGTTCG TTAGTTTGGA GGGAATGTTG CGAGGCCAAG CCGAAGGGCC GGGGGGGATA GTAACGGGCG
AAAGTCGAAC TTGTGAGTAA GACCTTGGAG AAGGCGACCG TCCTGGGGTT AGCCTAGGCA GTTCCACTCG GGCTTGGGCG CAGGAAAGGC TGGCTTCCGG
AATTCCCAGC CTCCGGGAAA GCTAGCTGCT TGGGAAACGC CCAATCGAAT TAGCTCAGAG AAGAGTGGCC GTGGCCTTGT GTATCCAGTG TCTGTGCCTC
AGGCTTCAAC CCCCTAGGGA AAGAGTGACA AGCAAATACT TCGCCTTGAA TGTTGCTACT CTGAAGCTGT AACTGGAACA GAACTTGGGC CGAGTGGAAA
ATGAACGTGG GCCCGCCCCT TCCCCCACGC GAAACAAGAA TTCTACAACA GGGGCGCTAA GACCAAAAAC CTTTCCCAGA GACTCAACAC TCTGCTTTTC
ATGGGTTCAA CGTGTGTGTG GTTTGCTGTT ATTTGATTTA TGCTTAGAAA CCATATTCAG AAAATCCGCA TTTAACATCC AATGAACTTG CTGGTCCCCT
TTTCCAAAGT TTGGAGATAG TTATGTCTGC CTGACAAGCA GGTCGTTTAA CTGAGGAATA ATCTGCAGTT TTGGGTCAAA GCCCACCAGA GGAAAGAGAA
GACACTTTAA AAGGTCTCTC TCTCTTTTTT AATAGACTTA GGATTTAAAA AATCTTATTG GGTTTAAATC TAATCTTGGT AGCGATTTTG TAAAATTTGA
AAATTTTAAG GTGTCAGTGA CGTGTCAGGG GGACATTCAG GAGGAGGAGG AGGTGCTCAT TTGCAGAGGG GCCACAGAAG TTAACCAAGG GGTCTCAATT
TCAACAGCTT CTTCACCTTG AAAATGAAAT GTGGAACCGC AGGCAGATAT GAGCTGCTTT CAGTATGATG CAACCTTGCT TATGATAGGC CAGATAAAAC
CCATCCCACC ACTGGGTGCT GGGGCACGTC AGTCATAAAC GTCAGATGAG CAGTAATGCG GTGTTCATAA TTCCTCTCCC TTTATCCTAC AGATGTCTGG
ACTGCAATCC TGCACAGTTT TGAGA
-2455
-2355
-2255
-2155
-2055
-1955
-1855
-1755
-1655
-1555
-1455
-1355
-1255
-1155
-1055
-955
-855
-755
-655
-555
-455
-355
-255
-155
-55
46
146
246
346
446
546
646
746
846
946
1046
1146
1246
1346
1446
1546
1646
GC/SP-1 GC/SP-1
Adapter Primer 2
NF-IL-6
NF B
NF-IL-6
NFκB
AP-1
AP-1
AP-1
AP-1
AP-1 (-)
AP-1 (-)
CRE
GATA (-)
GATA
GATA (-)
GATA (-)
GATA
GATA
STAT
YGATG (-)
ACTATAGGGC ACGCGTGGTC GACGGCCCGG GCTGGTCCTC AGAAATAACA TCACACATCT ACAACCATCT GATCTTCAAC AAAACTGACA AAAACAAGCA
ATGGAGAAAG GATGTCCTAT TTAATAAATG ATGTTGGGAA AACTGGCTAG CCATATGCAA AAAACTGAAA CTGGACCCCA TTCCTTATAC CTTATACAAA
AATTAACTCA AGATGGATTA AAGACTTAAA TGTAAAACCT AAAATAATAA AAACTCTAGA AAAAAACATA GGTAATACCA TTCAGGACAT AGGCATGGGC
ATAGACTTTA TGACAAAAAC ACAAAAAACA ATGACGACAA AAACACACAA AAAAATGACA ACAAAGGCCA AAATTGACAC ATGGGGTCTA ATTAAACTAA
AGAACTTCTG TACAGCAAAA GAAACTATCA TCGGAGTGAA CAGGCAACCT AGAGAATGGG AGAAATACCT AATGTAGATG ATGGGTTGAC CACCATGGCA
CATGTATACC TATGCACATG TATCTCAGAA CTTAAAGTAT AATAATTTAA AAAAAGTTAA AATAAAATAT TAATGACTTT CAAAGAAAAA AAAAGTCAAG
AAACAATAGA TGCTGGCGAG GCTATGGAGA AATAGGAATG CTTTTACACT GTTGGTAGGA ATGTAAATTA GTTCAACCAA TATGGAAGAC GGCATGGTGA
TTCCTCAAGG ATCTAGAACC AGAAATATCA TTTGATCCAG CAATCCCATT ACTGGGTATA TACCCAAAGG AATACAAATC ATTCTATTAT GAAGACACAT
GCACACATGT GTTTACTGAA GCACTATTTA CAATAGCAAA GTCATAGAAC CAACCCAAAT GCCCATCAAT AATAGACTGG ATAAAGAAAA TGTGGTACAT
ATACACCACG GAATACTACA CAGCCATAAA AAGGAATGGG ATCATGTCCT TTGCAGGGAC ATGGATGAAC CTGGATGCTA TCATCCTCAG CAAACTAGCA
CAGGAACAGA AAACCAAACA CTGCATGTTT TCACTTATAA ATGGGAGTTC AGCAATGAGA ACACATGGAC ACAGGGAGAG GAACAACATG CACCAGGGCC
TGTTAGGGGG TGGGGGGCAG GGGGAGGGAA CTTAGAGGAC AGGTCAATAG GTGCAAAAAT TTCATGTTTT TTAAAAAAGA AAAAAATTAT ATATGCACTC
ACATATTTTA AAAAATAAAG GAAAGAATAG TACTCTGTGT CATTCATATT TTCAGTATAG CTTATCACAT AATTGCCTTT TGGATACTGG CCATTCATGT
ATAATATTTC TCTAGTAACC AGGTTATCTA ACCAAAATGT TTAATTCCCT CACCATAATT AAGTAACAGT TTAGTTTACC AGTTATGATC ATATCATTTA
AAAAAATCTC AATATCTACC ACAAGGGAAA ACTGCAATCA TTCCTTTCTA CAGTTAATGG CTAATTTATT TTATCCCGCT TTTTCATTTT TTGATCCCTT
CCAATTTCCA GGAATGTTAG AGGTCCCTTC TGGTTTTAGA ACGGAGAAAC AGGGTTCGTT TCCATGTGGT ACTGATTTTT CACATAGGTA GTGGCAACAT
TTTACTACAA TAAAAAGAGC AAAAGTCAAT AATAGATTCA CATAAGTGTT TAGCTACAAA AATGAGACAC AGCTAATGGG TGTGTGTCGA AGCAATATTA
GATAAAAGCA ATTACAGAAT AAAATGGCTC AAATCGTAAT TCCTAATGAA AAATAGATTA CTTCATCATA AATTTATCTC CACGAAATTT ATTAGACGCA
TATCTCTTGG GGTATACATA TCACTAGAAA TTCTCTCTCC AGTGGACTTC TAAAATCACT GAAATATTAC TAATCCCTTA TCTGCAGGTG TACTTCAAGA
AGTCATCTCA TCCCTTTTCC TTTTCATATT CCTTAGAAAG CTGGAAACAG TCCCAGTCCT CTGTGTGGCA GTGAACCCTG AGGCTACGGG TGTTTTCATT
TACAATACTG GGCTTGAATA CCTGTTTTTA ATTCCCTGGA GATATTTGCC ACAAAATGGC TCCCTCAAGC ACGCAACCTC TTTCTACCTG GTTTCTTTAT
GCGTTTCGAG CTGGACTTGG AATGAATTGT CTGCAAGCTA CCATCCGACA GCCTAACACA CCATCTTAGG CTTTCCTCGG CTCTCCAGAC TGCTGTCTTG
CATCCCCAGA GATGGCTTGG GGGATAAAAA TGAGTTTAAA ACCAAACTTT CTTTAGCTGG TCCAGCGGCT AAGTGCAGCT CTGATGCATC CGAGCACGGT
GCCCTCTTCT CTCCCAGTAC TGCTCTGTGC CCATGGAAAA GGGGGCGGAG GCGAGAGAGA AGAGGAAGAG GCGGAGGTCA CTCGCGCGCC CCATCCCTCC
CAGGCTGCCA GCGCAGCTAG GTGCAGAGGG ATCCCAGGAG CCGCGCGCGC CCCGCAGTTT CCGCGCTAAG GGAACGAGTG CGCGGAGGGG ACGAGCGGCT
GGACCACAGC CGGCGCCCGA TCAGGATCTC CGCGCTGGGA TCGGTGGAAC TTGAGGCAGC GGCGGCGCGG GGCGCCATGG CACACCGAGC GGCTCCGTCT
TCTGCTCCTC AGAGAGCCCG GCTGGCGGCC TGGGATGACA AGGTACCATC CCTCCAGAGG CTGATCCCAA TGCATCGGCT TCGCTTATCT AGACCCAGAT
TTAATCAACT CACCCCCTGA GACACCCAGC CACAGCTTCT GCTGGCTGAG CCTGAGCACT GAGTTTGGAC CAGATTCCCC AAGCTAGGGA GACGCATAGA
GAAAGAAGGG TACTCGAGCT AACCCATCCC CATTCTGAGC TGCTCTGCGG GGGATTGGGT AGAAAGGGTC TCAGAAGAGG GAAATCGGCA CAGATTCCGC
TTCTCGCCGT TGCGGGGTTC CCTGCGGGCG GCACTGCGAG TTCCTCACCC GGGCACGTCC TCAACTCCCC GTGCCCGGGA ATGAGGGAGA AAACAGCCCT
TTGGCGTTTG AGCAATTGCC AGAAAGTTTC AGCGAGTTCG TTAGTTTGGA GGGAATGTTG CGAGGCCAAG CCGAAGGGCC GGGGGGGATA GTAACGGGCG
AAAGTCGAAC TTGTGAGTAA GACCTTGGAG AAGGCGACCG TCCTGGGGTT AGCCTAGGCA GTTCCACTCG GGCTTGGGCG CAGGAAAGGC TGGCTTCCGG
AATTCCCAGC CTCCGGGAAA GCTAGCTGCT TGGGAAACGC CCAATCGAAT TAGCTCAGAG AAGAGTGGCC GTGGCCTTGT GTATCCAGTG TCTGTGCCTC
AGGCTTCAAC CCCCTAGGGA AAGAGTGACA AGCAAATACT TCGCCTTGAA TGTTGCTACT CTGAAGCTGT AACTGGAACA GAACTTGGGC CGAGTGGAAA
ATGAACGTGG GCCCGCCCCT TCCCCCACGC GAAACAAGAA TTCTACAACA GGGGCGCTAA GACCAAAAAC CTTTCCCAGA GACTCAACAC TCTGCTTTTC
ATGGGTTCAA CGTGTGTGTG GTTTGCTGTT ATTTGATTTA TGCTTAGAAA CCATATTCAG AAAATCCGCA TTTAACATCC AATGAACTTG CTGGTCCCCT
TTTCCAAAGT TTGGAGATAG TTATGTCTGC CTGACAAGCA GGTCGTTTAA CTGAGGAATA ATCTGCAGTT TTGGGTCAAA GCCCACCAGA GGAAAGAGAA
GACACTTTAA AAGGTCTCTC TCTCTTTTTT AATAGACTTA GGATTTAAAA AATCTTATTG GGTTTAAATC TAATCTTGGT AGCGATTTTG TAAAATTTGA
AAATTTTAAG GTGTCAGTGA CGTGTCAGGG GGACATTCAG GAGGAGGAGG AGGTGCTCAT TTGCAGAGGG GCCACAGAAG TTAACCAAGG GGTCTCAATT
TCAACAGCTT CTTCACCTTG AAAATGAAAT GTGGAACCGC AGGCAGATAT GAGCTGCTTT CAGTATGATG CAACCTTGCT TATGATAGGC CAGATAAAAC
CCATCCCACC ACTGGGTGCT GGGGCACGTC AGTCATAAAC GTCAGATGAG CAGTAATGCG GTGTTCATAA TTCCTCTCCC TTTATCCTAC AGATGTCTGG
ACTGCAATCC TGCACAGTTT TGAGA
-2455
-2355
-2255
-2155
-2055
-1955
-1855
-1755
-1655
-1555
-1455
-1355
-1255
-1155
-1055
-955
-855
-755
-655
-555
-455
-355
-255
-155
-55
46
146
246
346
446
546
646
746
846
946
1046
1146
1246
1346
1446
1546
1646
ACTATAGGGC ACGCGTGGTC GACGGCCCGG GCTGGTCCTC AGAAATAACA TCACACATCT ACAACCATCT GATCTTCAAC AAAACTGACA AAAACAAGCA
ATGGAGAAAG GATGTCCTAT TTAATAAATG ATGTTGGGAA AACTGGCTAG CCATATGCAA AAAACTGAAA CTGGACCCCA TTCCTTATAC CTTATACAAA
AATTAACTCA AGATGGATTA AAGACTTAAA TGTAAAACCT AAAATAATAA AAACTCTAGA AAAAAACATA GGTAATACCA TTCAGGACAT AGGCATGGGC
ATAGACTTTA TGACAAAAAC ACAAAAAACA ATGACGACAA AAACACACAA AAAAATGACA ACAAAGGCCA AAATTGACAC ATGGGGTCTA ATTAAACTAA
AGAACTTCTG TACAGCAAAA GAAACTATCA TCGGAGTGAA CAGGCAACCT AGAGAATGGG AGAAATACCT AATGTAGATG ATGGGTTGAC CACCATGGCA
CATGTATACC TATGCACATG TATCTCAGAA CTTAAAGTAT AATAATTTAA AAAAAGTTAA AATAAAATAT TAATGACTTT CAAAGAAAAA AAAAGTCAAG
AAACAATAGA TGCTGGCGAG GCTATGGAGA AATAGGAATG CTTTTACACT GTTGGTAGGA ATGTAAATTA GTTCAACCAA TATGGAAGAC GGCATGGTGA
TTCCTCAAGG ATCTAGAACC AGAAATATCA TTTGATCCAG CAATCCCATT ACTGGGTATA TACCCAAAGG AATACAAATC ATTCTATTAT GAAGACACAT
GCACACATGT GTTTACTGAA GCACTATTTA CAATAGCAAA GTCATAGAAC CAACCCAAAT GCCCATCAAT AATAGACTGG ATAAAGAAAA TGTGGTACAT
ATACACCACG GAATACTACA CAGCCATAAA AAGGAATGGG ATCATGTCCT TTGCAGGGAC ATGGATGAAC CTGGATGCTA TCATCCTCAG CAAACTAGCA
CAGGAACAGA AAACCAAACA CTGCATGTTT TCACTTATAA ATGGGAGTTC AGCAATGAGA ACACATGGAC ACAGGGAGAG GAACAACATG CACCAGGGCC
TGTTAGGGGG TGGGGGGCAG GGGGAGGGAA CTTAGAGGAC AGGTCAATAG GTGCAAAAAT TTCATGTTTT TTAAAAAAGA AAAAAATTAT ATATGCACTC
ACATATTTTA AAAAATAAAG GAAAGAATAG TACTCTGTGT CATTCATATT TTCAGTATAG CTTATCACAT AATTGCCTTT TGGATACTGG CCATTCATGT
ATAATATTTC TCTAGTAACC AGGTTATCTA ACCAAAATGT TTAATTCCCT CACCATAATT AAGTAACAGT TTAGTTTACC AGTTATGATC ATATCATTTA
AAAAAATCTC AATATCTACC ACAAGGGAAA ACTGCAATCA TTCCTTTCTA CAGTTAATGG CTAATTTATT TTATCCCGCT TTTTCATTTT TTGATCCCTT
CCAATTTCCA GGAATGTTAG AGGTCCCTTC TGGTTTTAGA ACGGAGAAAC AGGGTTCGTT TCCATGTGGT ACTGATTTTT CACATAGGTA GTGGCAACAT
TTTACTACAA TAAAAAGAGC AAAAGTCAAT AATAGATTCA CATAAGTGTT TAGCTACAAA AATGAGACAC AGCTAATGGG TGTGTGTCGA AGCAATATTA
GATAAAAGCA ATTACAGAAT AAAATGGCTC AAATCGTAAT TCCTAATGAA AAATAGATTA CTTCATCATA AATTTATCTC CACGAAATTT ATTAGACGCA
TATCTCTTGG GGTATACATA TCACTAGAAA TTCTCTCTCC AGTGGACTTC TAAAATCACT GAAATATTAC TAATCCCTTA TCTGCAGGTG TACTTCAAGA
AGTCATCTCA TCCCTTTTCC TTTTCATATT CCTTAGAAAG CTGGAAACAG TCCCAGTCCT CTGTGTGGCA GTGAACCCTG AGGCTACGGG TGTTTTCATT
TACAATACTG GGCTTGAATA CCTGTTTTTA ATTCCCTGGA GATATTTGCC ACAAAATGGC TCCCTCAAGC ACGCAACCTC TTTCTACCTG GTTTCTTTAT
GCGTTTCGAG CTGGACTTGG AATGAATTGT CTGCAAGCTA CCATCCGACA GCCTAACACA CCATCTTAGG CTTTCCTCGG CTCTCCAGAC TGCTGTCTTG
CATCCCCAGA GATGGCTTGG GGGATAAAAA TGAGTTTAAA ACCAAACTTT CTTTAGCTGG TCCAGCGGCT AAGTGCAGCT CTGATGCATC CGAGCACGGT
GCCCTCTTCT CTCCCAGTAC TGCTCTGTGC CCATGGAAAA GGGGGCGGAG GCGAGAGAGA AGAGGAAGAG GCGGAGGTCA CTCGCGCGCC CCATCCCTCC
CAGGCTGCCA GCGCAGCTAG GTGCAGAGGG ATCCCAGGAG CCGCGCGCGC CCCGCAGTTT CCGCGCTAAG GGAACGAGTG CGCGGAGGGG ACGAGCGGCT
GGACCACAGC CGGCGCCCGA TCAGGATCTC CGCGCTGGGA TCGGTGGAAC TTGAGGCAGC GGCGGCGCGG GGCGCCATGG CACACCGAGC GGCTCCGTCT
TCTGCTCCTC AGAGAGCCCG GCTGGCGGCC TGGGATGACA AGGTACCATC CCTCCAGAGG CTGATCCCAA TGCATCGGCT TCGCTTATCT AGACCCAGAT
TTAATCAACT CACCCCCTGA GACACCCAGC CACAGCTTCT GCTGGCTGAG CCTGAGCACT GAGTTTGGAC CAGATTCCCC AAGCTAGGGA GACGCATAGA
GAAAGAAGGG TACTCGAGCT AACCCATCCC CATTCTGAGC TGCTCTGCGG GGGATTGGGT AGAAAGGGTC TCAGAAGAGG GAAATCGGCA CAGATTCCGC
TTCTCGCCGT TGCGGGGTTC CCTGCGGGCG GCACTGCGAG TTCCTCACCC GGGCACGTCC TCAACTCCCC GTGCCCGGGA ATGAGGGAGA AAACAGCCCT
TTGGCGTTTG AGCAATTGCC AGAAAGTTTC AGCGAGTTCG TTAGTTTGGA GGGAATGTTG CGAGGCCAAG CCGAAGGGCC GGGGGGGATA GTAACGGGCG
AAAGTCGAAC TTGTGAGTAA GACCTTGGAG AAGGCGACCG TCCTGGGGTT AGCCTAGGCA GTTCCACTCG GGCTTGGGCG CAGGAAAGGC TGGCTTCCGG
AATTCCCAGC CTCCGGGAAA GCTAGCTGCT TGGGAAACGC CCAATCGAAT TAGCTCAGAG AAGAGTGGCC GTGGCCTTGT GTATCCAGTG TCTGTGCCTC
AGGCTTCAAC CCCCTAGGGA AAGAGTGACA AGCAAATACT TCGCCTTGAA TGTTGCTACT CTGAAGCTGT AACTGGAACA GAACTTGGGC CGAGTGGAAA
ATGAACGTGG GCCCGCCCCT TCCCCCACGC GAAACAAGAA TTCTACAACA GGGGCGCTAA GACCAAAAAC CTTTCCCAGA GACTCAACAC TCTGCTTTTC
ATGGGTTCAA CGTGTGTGTG GTTTGCTGTT ATTTGATTTA TGCTTAGAAA CCATATTCAG AAAATCCGCA TTTAACATCC AATGAACTTG CTGGTCCCCT
TTTCCAAAGT TTGGAGATAG TTATGTCTGC CTGACAAGCA GGTCGTTTAA CTGAGGAATA ATCTGCAGTT TTGGGTCAAA GCCCACCAGA GGAAAGAGAA
GACACTTTAA AAGGTCTCTC TCTCTTTTTT AATAGACTTA GGATTTAAAA AATCTTATTG GGTTTAAATC TAATCTTGGT AGCGATTTTG TAAAATTTGA
AAATTTTAAG GTGTCAGTGA CGTGTCAGGG GGACATTCAG GAGGAGGAGG AGGTGCTCAT TTGCAGAGGG GCCACAGAAG TTAACCAAGG GGTCTCAATT
TCAACAGCTT CTTCACCTTG AAAATGAAAT GTGGAACCGC AGGCAGATAT GAGCTGCTTT CAGTATGATG CAACCTTGCT TATGATAGGC CAGATAAAAC
CCATCCCACC ACTGGGTGCT GGGGCACGTC AGTCATAAAC GTCAGATGAG CAGTAATGCG GTGTTCATAA TTCCTCTCCC TTTATCCTAC AGATGTCTGG
ACTGCAATCC TGCACAGTTT TGAGA
-2455
-2355
-2255
-2155
-2055
-1955
-1855
-1755
-1655
-1555
-1455
-1355
-1255
-1155
-1055
-955
-855
-755
-655
-555
-455
-355
-255
-155
-55
46
146
246
346
446
546
646
746
846
946
1046
1146
1246
1346
1446
1546
1646
Fig. 2. Sequence of the 4106-bp hFP promoter DNA fragment that contains � 2436 bp of 5V-flanking sequence. Shown in grey and underlined is the Adapter
Primer used for the genome walking protocol. Bases are numbered relative to the transcription start site determined from 5V RLM-RACE analysis (arrowhead).
The fragment also contains all of exon 1 and the first 33 bp of exon 2 (shaded regions) as well as intron 1. Potential transcription factor binding sites are
underlined. Shown in bold italics is the homologous region found in the mouse, rat, and bovine FP promoter.
D.B. Zaragoza et al. / Biochimica et Biophysica Acta 1676 (2004) 193–202 197

D.B. Zaragoza et al. / Biochimica et Biophysica Acta 1676 (2004) 193–202198
TATA box as they are found in many TATA-less promoters.
It is believed that SP-1 can function as a tethering moiety to
recruit the general transcription machinery to TATA-less
promoters [11–14]. This is also supported by the existence
of SP-1/GC box elements immediately upstream of the
transcription initiation sites for the TATA-less mouse, rat,
and bovine FP genes [7–9].
The human FP promoter bears no homology to the mouse,
rat, or bovine FP promoters with the exception of a region
Fig. 3. Ability of the hFP promoter fragment (4106 bp) to promote gene transcrip
phFPprom-GFP. (B) PCR primers (hFP-GFP f1/hFP-GFP r1) were designed acro
These primers will yield a 435-bp product if intron 1 is spliced out. (C) Results of R
with phFPprom-GFP. A minus reverse transcriptase reaction (�RT) was performe
PCR product was sequenced to verify that it was a FP-GFP fusion and that intro
between � 1013 and � 920 (Fig. 2, shown in bold italics).
This ~100-bp region is found at position � 989 of the mouse
promoter [7], � 1154 of the rat promoter [9], and � 1484 of
the bovine promoter [8], and therefore may play some
common regulatory role in transcription (see below). How-
ever, the overall lack of homology among promoters sug-
gests that regulation of transcription of the FP gene differs
among species. Nevertheless, similar transcription binding
sites are found in all of these promoters. Several GATA and
tion. (A) The hFP fragment was cloned into a GFP reporter vector to make
ss intron 1 in order to detect FP-GFP fusion mRNA in RT-PCR reactions.
T-PCR reactions from RNA isolated from HeLa cells transiently transfected
d in parallel to ensure that the 435-bp product was derived from RNA. The
n 1 was spliced out.

D.B. Zaragoza et al. / Biochimica et Biophysica Acta 1676 (2004) 193–202 199
AP-1 sites are present throughout the human FP promoter
(Fig. 2). GATA sites were also found in the rat promoter [9]
and in the bovine FP promoter although they were not
reported [8]. AP-1 sites were also found in rat FP promoter
[9]. In addition, we identified a STAT site at position � 950
and a cAMP response element (CRE) was found at position
� 2124 (Fig. 2). A CRE site was also present in the rat
promoter [9]. Interestingly, this STAT site at � 950 and the
GATA site at � 984 are located in the region homologous to
the mouse, rat, and bovine promoter. Whether these sites
have any functional significance remains to be determined.
Intron 1 of the human FP gene was also examined for
potential transcription factor binding sites and found to
contain two NFnB sites at + 746 and + 777, and two NF-
IL-6 sites at + 593 and + 1102 (Fig. 2). The NFnB sites may
be important for the IL-1h induction of human FP mRNA in
human granulosa-luteal cells [15]. We have observed a
similar effect of IL-1h in human myometrial cells (unpub-
lished data). The NF-IL-6 sites may also be important given
recent evidence showing IL-6 regulation of FP transcription
in the rat [9]. Mutational analysis of these sites under
conditions of IL-1h or IL-6 treatment will provide more
information on their functional relevance.
The basal transcriptional activity of the 4106-bp hFP
DNA fragment was assessed by transfecting GFP reporter
constructs into HeLa cells and measuring mRNA levels of
hFP-GFP fusion mRNA. HeLa cells were used because of
the high transfection efficiency achieved, and RT-PCR
analysis of RNA isolated from HeLa cells indicated that
these cells do express FP (data not shown). The full-length
construct contains 2436 bp of sequence upstream of the
Fig. 4. Transcriptional activity of the 5V-flanking region of the human FP gene. He
5V deletions of the hFP promoter. Cells (1�105) were plated in each well of si
antimycotic (Invitrogen) for 24 h (f 60% confluent). Cells were co-transfected wi
(Promega, Canada) (to normalize for transfection efficiency) using Effectene Trans
with Qiagen’s EC buffer (Qiagen proprietary compound mixture) to 100 Al. Twelveto each sample and incubated 5 min at room temperature. Fifteen microliters of
incubation at room temperature. After addition of 600 Al of growth media (D-MEM
to the HeLa cells and incubated overnight. Following a 19-h incubation, cells were
determined by real-time RT-PCR. Both FP-GFP mRNA levels and h-galactosidasewere then corrected for transfection efficiency by dividing this value by GAPDH
independent experiments. Values greater than two standard deviations from the m
Student’s t tests were performed on consecutive paired samples.
transcription initiation site as well as 1450 bp of intron 1
(Fig. 3A). RT-PCR reaction primers (Table 1, hFP-GFP f1/
hFP-GFP r1) spanned intron 1 and were designed to detect a
FP-GFP fusion mRNA of 435 bp if intron 1 was properly
spliced out of the fusion mRNA (Fig. 3B). We were able to
detect a single RT-PCR product, f 435 bp, from RNA
isolated from HeLa cells transfected with the full-length
phFPprom-GFP construct (Fig. 3C). Sequencing confirmed
that this product was a FP-GFP fusion that was missing
intron 1. This result indicates that the 4106-bp hFP fragment
is able to promote basal gene transcription.
In order to identify basal transcription regulatory regions
in the 5V-flanking sequence of the hFP gene, we assessed the
ability of constructs containing successive 5V deletions of
hFP promoter to direct transcription of the FP-GFP fusion
mRNA by real-time PCR. Because we are detecting actual
fusion FP-GFP transcripts (sense primer-FP; antisense prim-
er-GFP), an empty control vector is not shown since no FP-
GFP would be detected in this case. Deletion of the � 2437-
bp to � 1946-bp region resulted in a 2.6-fold increase in the
levels of FP-GFP mRNA, suggesting the presence of a
repressor element (Fig. 4). Successive deletions to � 1118
bp resulted in no significant changes in the levels of FP-GFP
mRNA; however, deleting to � 582 bp resulted in a fivefold
decrease, suggesting the presence of an enhancer element in
the � 1118/� 582 region (Fig. 4).
The existence of repressor and enhancer regions in the
FP promoter has important implications for the regulation
of pregnancy. The repressor regions may be important for
the down-regulation of FP expression observed during
pregnancy resulting in relaxation of the myometrium [4].
La cells were transfected with GFP reporter constructs containing successive
x-well culture plates and grown in D-MEM containing 10% FBS and 1�th the appropriate hFP-GFP construct and pSV-hgalactosidase control vectorfection Reagent (Qiagen Inc., Canada). Each plasmid (0.75 Ag) was dilutedmicroliters of enhancer (Qiagen proprietary compound mixture) was added
effectene was added (1:10 DNA to effectene ratio) followed by a 10-min
, 10% FBS, and 1� antimycotic), DNA–effectene complexes were added
harvested and hFP-GFP, hGAPDH, and h-galactosidase mRNA levels were
levels were first normalized to GAPDH. GAPDH normalized FP-GFP levels
normalized h-galactosidase values. Values are meansF S.E.M. from six
ean were discarded from the analysis resulting in N = 5 for some groups.

D.B. Zaragoza et al. / Biochimica et Biophysica Acta 1676 (2004) 193–202200
Further, the enhancer region may be important for up-
regulating FP expression at the time of birth, resulting in
the contraction of the uterus and termination of pregnancy
[5].
There are several candidate repressors that may bind this
repressor region (� 2436/� 1946). Within this region, there
are three AP-1 sites (� 2371, � 2145, and � 1969), one
CRE site (� 2124), and two GATA-like sites (� 2400,
� 2030). The presence of a CRE site with upstream AP-1
sites may be relevant for transcriptional repression. AP-1
sites in this configuration, through binding CBP/AP-1
complexes, may sequester CBP away from transcription
preinitiation complexes formed on CRE sites [16]. Several
Fig. 5. DNase I footprinting analysis of the � 2436/� 1946 repressor region. phF
Labeled linearized plasmid was then digested with NcoI. The labeled SpeI/Nc
chromatography. Binding reactions were performed for 30 min on ice and containe
(pH 8.0), 20% glycerol, 1 mM DTT, 0.02 Ag/Al poly (dI–dC), 10000 cpm probe
reactions, DNase I treatments were carried out for each reaction in a staggered fas
CaCl2, 10 mM MgCl2), reactions were incubated at room temperature for 1.5 mi
temperature for 1.5 min. Samples were phenol/chloroform-extracted, ethanol-prec
Samples were run against a Maxam and Gilbert G +A sequencing reaction, and a
Full-length undigested probe was also included in the run. Four identical reaction
regions in the + nuclear extract lanes are shown with a bracket ( ] ).
factors have also been shown to bind to GATA-like
sequences (GATA/G) and mediate transcriptional repression
[17–19].
In an effort to determine whether the � 2436/� 1946
contains functional DNA binding sites, DNase I footprinting
was performed on this region (Fig. 5). Incubating a 32P-end-
labeled probe (corresponding to the � 2436/� 1946 region)
with nuclear extracts resulted in a DNase I protected region
at f� 2400 as determined by a G+A sequencing reaction
and the fX174RF marker (Fig. 5, + nuclear extract lanes).
This region corresponds to a GATA-like motif. This site may
also be capable of mediating suppression of gene transcrip-
tion through binding of the GAP-12 repressor, Ash1, or
Pprom-GFP was digested with SpeI and end-labeled with Klenow fragment.
oI fragement was purified by agarose electrophoresis and reverse-phase
d 50 mM Tris HCl (pH 8.0), 100 mM KCl, 12.5 mM MgCl2, 1 mM EDTA
and either 40 Ag of nuclear extract or extraction buffer. Following binding
hion, 30 s apart. Following addition of 50 Al of Ca2 +/Mg2 + solution (5 mM
n. Reactions were either treated with water or 0.6 U RQ DNase I at room
ipitated and loaded onto a 6% PAGE gel and analyzed by autoradiography.
n fX174 RF DNA/HaeIII ladder in order to identify the protected regions.
s are shown comparing � nuclear extact with + nuclear extract. Protected

D.B. Zaragoza et al. / Biochimica et Biophysica Acta 1676 (2004) 193–202 201
TRPS1 [17–20]. These proteins have all been demonstrated
to repress transcription through binding to GATA-like ele-
ments. While the site found at � 2400 lacks a conventional
GATA core sequence, it does contain the GATA-like motif
YTGAT (-strand) which has been shown to bind the Ash1
repressor [19]. Mutational analysis of this site will reveal
whether it has any functional role in the repressive activity of
this region of the hFP promoter. In addition, gel shift and
supershift assays will identify which DNA binding proteins
bind to the GATA-like site at position � 2400 of the FP
promoter.
We also analyzed the enhancer region (� 1118/� 582)
for nuclear extract binding activity; however, to date, we
have not identified any DNase I protected regions (data not
shown). It is possible that protein binding to these sites
may require additional downstream sequences that are not
present in the 32P-labeled probe. A thorough mutational
analysis may be necessary to identify specific sites respon-
sible for the enhanced transcription observed in our experi-
ments. Oddly, this enhancer region of the hFP promoter
(Fig. 4, � 1118/� 582) also contains a GATA element
(� 984) and an AP-1 site (� 833), in addition to a
STAT-1 site (� 950). Whether these sites are required for
basal transcription of the hFP gene remains to be deter-
mined. This region also contains the portion of the hFP
promoter that is found in the mouse, rat, and bovine FP
promoters (� 1013 and � 920). This finding suggests the
existence of a common regulatory element of FP gene
transcription among different species. In fact, 5V deletions
of the rat FP promoter from � 1418 to � 608 which
contain this common region (starting at � 1154 of the rat
FP promoter) resulted in a f 5-fold decrease in luciferase
reporter activity [9]. To date, no analysis of the homolo-
gous region has been performed on the mouse or bovine FP
promoters. Nevertheless, our data and the rat data indicate
an important role for this homologous region in regulating
FP gene transcription. A more detailed analysis within this
100-bp region will be crucial to understanding the regula-
tion of FP gene transcription.
To date, the transcription regulatory mechanisms of the
human FP receptor are largely unknown. In this study we
have taken the initial steps to understanding how this
important molecule is regulated at the transcriptional level.
The characterization of the promoter presented here will be
important for future studies analyzing the transcriptional
regulation of FP and how it relates to the maintenance and
termination of human pregnancy. The identification of both
negative and positive cis-acting regions important for basal
transcription may provide clues to how this gene regulates
uterine contractions during pregnancy. Further, the identifi-
cation of a GATA-like site able to bind nuclear extracts in
the repressor region should prove beneficial to more in-
depth analyses of the control of transcription of the human
FP gene. Ultimately such studies will provide important
information on how this gene is regulated in the context
parturition.
Acknowledgements
The authors would like to thank Dr. Jacob T. Ross for
technical assistance. This work was supported by the Ca-
nadian Institutes of Health Research (CIHR), the Alberta
Heritage Foundation for Medical Research, the Faculty of
Medicine and Dentistry, University of Alberta, the University
of Alberta Perinatal Research Centre and the National
Science Foundation POWRE Program.
References
[1] J.R. Challis, D.M. Olson, in: E. Knobil, J. Neill (Eds.), The Physiol-
ogy of Reproduction, Raven Press, New York, 1988, pp. 2177–2216.
[2] J. Senior, K. Marshall, R. Sangha, J.K. Clayton, In vitro characte-
rization of prostanoid receptors on human myometrium at term preg-
nancy, Br. J. Pharmacol. 108 (1993) 501–506.
[3] J. Senior, R. Sangha, G.S. Baxter, K. Marshall, J.K. Clayton, In vitro
characterization of prostanoid FP-, DP-, IP- and TP-receptors on the
non-pregnant human myometrium, Br. J. Pharmacol. 107 (1992)
215–221.
[4] T. Matsumoto, N. Sagawa, M. Yoshida, T. Mori, I. Tanaka, M. Mu-
koyama, M. Kotani, K. Nakao, The prostaglandin E2 and F2 alpha
receptor genes are expressed in human myometrium and are down-
regulated during pregnancy, Biochem. Biophys. Res. Commun. 238
(1997) 838–841.
[5] J. Brodt-Eppley, L. Myatt, Prostaglandin receptors in lower segment
myometrium during gestation and labor, Obstet. Gynecol. 93 (1999)
89–93.
[6] R. Betz, J. Lagercrantz, D. Kedra, J.P. Dumanski, A. Nordenskjold,
Genomic structure, 5V flanking sequences, and precise localization in
1P31.1 of the human prostaglandin F receptor gene, Biochem. Bio-
phys. Res. Commun. 254 (1999) 413–416.
[7] K. Hasumoto, Y. Sugimoto, M. Gotoh, E. Segi, A. Yamasaki, M.
Yamaguchi, H. Honda, H. Hirai, M. Negishi, A. Kakizuka, A. Ichi-
kawa, Characterization of the mouse prostaglandin F receptor gene: a
transgenic mouse study of a regulatory region that controls its ex-
pression in the stomach and kidney but not in the ovary, Genes Cells 2
(1997) 571–580.
[8] T. Ezashi, K. Sakamoto, K. Miwa, E. Okuda-Ashitaka, S. Ito, O.
Hayaishi, Genomic organization and characterization of the gene en-
coding bovine prostaglandin F2alpha receptor, Gene 190 (1997)
271–278.
[9] F. Neuschafer-Rube, U. Moller, G.P. Puschel, Structure of the 5V-flanking region of the rat prostaglandin F(2alpha) receptor gene and
its transcriptional control functions in hepatocytes, Biochem. Bio-
phys. Res. Commun. 278 (2000) 278–285.
[10] K. Quandt, K. Frech, H. Karas, E. Wingender, T. Werner, MatInd and
MatInspector: new fast and versatile tools for detection of consensus
matches in nucleotide sequence data, Nucleic Acids Res. 23 (1995)
4878–4884.
[11] P.W. Rigby, Three in one and one in three: it all depends on TBP, Cell
72 (1993) 7–10.
[12] B.F. Pugh, R. Tjian, Diverse transcriptional functions of the multi-
subunit eukaryotic TFIID complex, J. Biol. Chem. 267 (1992)
679–682.
[13] B.F. Pugh, R. Tjian, Mechanism of transcriptional activation by Sp1:
evidence for coactivators, Cell 61 (1990) 1187–1197.
[14] W.S. Dynan, S. Sazer, R. Tjian, R.T. Schimke, Transcription factor
Sp1 recognizes a DNA sequence in the mouse dihydrofolate reductase
promoter, Nature 319 (1986) 246–248.
[15] K. Narko, O. Ritvos, A. Ristimaki, Induction of cyclooxygenase-2
and prostaglandin F2alpha receptor expression by interleukin-1beta in

D.B. Zaragoza et al. / Biochimica et Biophysica Acta 1676 (2004) 193–202202
cultured human granulosa-luteal cells, Endocrinology 138 (1997)
3638–3644.
[16] A.S. Guberman, M.E. Scassa, L.E. Giono, C.L. Varone, E.T. Canepa,
Inhibitory effect of AP-1 complex on 5-aminolevulinate synthase
gene expression through sequestration of cAMP-response element
protein (CRE)-binding protein (CBP) coactivator, J. Biol. Chem.
278 (2003) 2317–2326.
[17] T.H. Malik, S.A. Shoichet, P. Latham, T.G. Kroll, L.L. Peters, R.A.
Shivdasani, Transcriptional repression and developmental functions
of the atypical vertebrate GATA protein TRPS1, EMBO J. 20
(2001) 1715–1725.
[18] F.J. Kaiser, K. Tavassoli, G.J. Van den Bemd, G.T. Chang, B. Hors-
themke, T. Moroy, H.J. Ludecke, Nuclear interaction of the dynein
light chain LC8a with the TRPS1 transcription factor suppresses the
transcriptional repression activity of TRPS1, Hum. Mol. Genet. 12
(2003) 1349–1358.
[19] M.E. Maxon, I. Herskowitz, Ash1p is a site-specific DNA-binding
protein that actively represses transcription, Proc. Natl. Acad. Sci.
U. S. A. 98 (2001) 1495–1500.
[20] C. Becker, S. Wirtz, X. Ma, M. Blessing, P.R. Galle, M.F. Neurath,
Regulation of IL-12 p40 promoter activity in primary human mono-
cytes: roles of NF-kappaB, CCAAT/enhancer-binding protein beta,
and PU.1 and identification of a novel repressor element (GA-12)
that responds to IL-4 and prostaglandin E(2), J. Immunol. 167
(2001) 2608–2618.
Copyright © 2022 FDOKUMEN




















