
Characterization, cloning and immunolocalization of a coronin homologue in Trichomonas vaginalis
10
Characterization, cloning and immunolocalization of a coronin homologue in Trichomonas vaginalis Genevie ` ve Bricheux 1) , Ge ´rard Coffe, Danie `le Bayle, Guy Brugerolle Laboratoire de Biologie des Protistes, Universite ´ Blaise Pascal de Clermont-Ferrand, UPRESA CNRS, Aubie ` re/France Received October 29, 1999 Received in revised version February 12, 2000 Accepted March 23, 2000 Coronin – actin cytoskeleton – immunolocalization – Trichomonas vaginalis On adhesion to host cells the flagellate Trichomonas vaginalis switches to an amoeboid form rich in actin microfilaments. We have undertaken the identification of actin-associated proteins that regulate actin dynamics. A monoclonal antibody 4C12 raised against a cytoskeletal fraction of T. vaginalis labeled a protein doublet at circa 50 kDa. These two bands were recognized by the antibody against Dictyostelium discoideum coronin. During cell extraction and actin polymerization, T. vaginalis coronin cosedimented with F-actin. By two-dimen- sional gel electrophoresis, the protein doublet was separated into two sets of isoforms covering two Ip zones around 6 and 7. By screening a T. vaginalis library with 4C12, two clones Cor 1 and Cor 2 were isolated. This gene duplicity is a particularity among unicellular organisms examined. The complete se- quence of the gene Cor 1 encodes a 435-residue protein with a calculated molecular mass of 48 kDa and Ip of 5.58. The incomplete sequence Cor 2 was very similar but with a more basic calculated Ip than Cor 1 on the same region. T. vaginalis coronin had 50% similarity with the coronin family, possessing the five WD-repeats and a leucine zipper in its C-terminal part. Double immunofluorescence labeling showed that coronin mainly colocalized with actin at the periphery of the adherent amoeboid cells. However, coronin labeling displayed patches within a reticular array. Immunogold electron microscopy confirmed the coronin labeling in the actin-rich microfilamen- tous fringe beneath the plasma membrane, with accumulation in phagocytic zones and pseudopodial extensions. In T. vaginalis, one of the first emerging lineage of eukaryotes, coronin seems to play an important role in actin dynamics and may be a downstream target of a signaling mechanism for the cytoskeleton reorganization. Introduction In most eukaryotic cells, the dynamic properties of actin are regulated by the ATP cycle of actin itself, and by numerous regulatory proteins. The ability of actin monomers to assemble into filaments and then rapidly disassemble so that new filaments can form from the monomers is essential for dynamic processes such as cell motility, cytokinesis, cell polarity and endocytosis (for reviews see Ayscough, 1998; Chen et al., 2000). When ovoid Trichomonas vaginalis cells adhere to host cells or to the substratum, they spread forming pseudopods and filopods like amoeboid cells, and microfilament inhibitors such as cytochalasins B and E prevent cell adhesion (Honig- berg and Brugerolle, 1989; Gold and Ofek, 1992; Alderete et al., 1995; Gonzalez-Robles et al., 1995; Scott et al., 1995; Brugerolle et al., 1996). The peripheral cytoplasm and the layer along the adhesion zone become microfilamentous and were found to be composed of actin (Brugerolle et al., 1996). Actin microfilaments are organized like a network and are adhering to the plasma membrane but no stress fibers have been observed (Gonzalez-Robles et al., 1995; Brugerolle et al., 1996). Cell adhesion and signal transduction are coordinated processes that may be linked through regulatory elements such as actin-associated proteins (Svitkina and Borisy, 1999). In T. vaginalis , actin dynamics are tightly regulated because filament assembly and disassembly have to occur rapidly in the parasite, at least during adhesion and lysis of host cells. To elucidate the events underlying the reorganization of the actin cytoskeleton in T. vaginalis , the identification and functional characterization of regulatory proteins of actin dynamics have been undertaken (Bricheux et al., 1998; Addis et al., 1998). Here we report on the identification of coronin proteins and genes in T. vaginalis a representative of one of the first emerging lineages of eukaryotes (Embley and Hirt, 1998). Coronin derives its name from its association with crown- shaped projections from growth-phase Dictyostelium discoi- deum (de Hostos et al., 1991). Coronin is known to interact with F-actin (for review see de Hostos, 1999) but was found to have a primary structure unlike that of any other actin-binding EJCB 413 European Journal of Cell Biology 79, 413 – 422 (2000, June) · Urban & Fischer Verlag · Jena http://www.urbanfischer.de/journals/ejcb 0171-9335/00/79/06-413 $12.00/0 1) Dr. Genevie `ve Bricheux, Laboratoire de Biologie des Protistes, Universite ´ Blaise Pascal, Les Ce ´zeaux, – 63177 Aubie `re, Cedex, France, e-mail: [email protected], Fax: 33 4 73 40 76 70.
-
Upload
univ-bpclermont -
Category
Documents
-
view
3 -
download
0
Transcript of Characterization, cloning and immunolocalization of a coronin homologue in Trichomonas vaginalis

Characterization, cloning and immunolocalizationof a coronin homologue in Trichomonas vaginalis
GenevieÁve Bricheux1), GeÂrard Coffe, DanieÁ le Bayle, Guy BrugerolleLaboratoire de Biologie des Protistes, Universite Blaise Pascal de Clermont-Ferrand, UPRESA CNRS, AubieÁre/France
Received October 29, 1999Received in revised version February 12, 2000Accepted March 23, 2000
Coronin ± actin cytoskeleton ± immunolocalization ±Trichomonas vaginalis
On adhesion to host cells the flagellate Trichomonas vaginalisswitches to an amoeboid form rich in actin microfilaments. Wehave undertaken the identification of actin-associated proteinsthat regulate actin dynamics. A monoclonal antibody 4C12raised against a cytoskeletal fraction of T. vaginalis labeled aprotein doublet at circa 50 kDa. These two bands wererecognized by the antibody against Dictyostelium discoideumcoronin. During cell extraction and actin polymerization, T.vaginalis coronin cosedimented with F-actin. By two-dimen-sional gel electrophoresis, the protein doublet was separatedinto two sets of isoforms covering two Ip zones around 6 and 7.By screening a T. vaginalis library with 4C12, two clones Cor 1and Cor 2 were isolated. This gene duplicity is a particularityamong unicellular organisms examined. The complete se-quence of the gene Cor 1 encodes a 435-residue protein with acalculated molecular mass of 48 kDa and Ip of 5.58. Theincomplete sequence Cor 2 was very similar but with a morebasic calculated Ip than Cor 1 on the same region. T. vaginaliscoronin had 50% similarity with the coronin family, possessingthe five WD-repeats and a leucine zipper in its C-terminal part.Double immunofluorescence labeling showed that coroninmainly colocalized with actin at the periphery of the adherentamoeboid cells. However, coronin labeling displayed patcheswithin a reticular array. Immunogold electron microscopyconfirmed the coronin labeling in the actin-rich microfilamen-tous fringe beneath the plasma membrane, with accumulationin phagocytic zones and pseudopodial extensions. In T.vaginalis, one of the first emerging lineage of eukaryotes,coronin seems to play an important role in actin dynamics andmay be a downstream target of a signaling mechanism for thecytoskeleton reorganization.
Introduction
In most eukaryotic cells, the dynamic properties of actin areregulated by the ATP cycle of actin itself, and by numerousregulatory proteins. The ability of actin monomers to assembleinto filaments and then rapidly disassemble so that newfilaments can form from the monomers is essential for dynamicprocesses such as cell motility, cytokinesis, cell polarity andendocytosis (for reviews see Ayscough, 1998; Chen et al.,2000). When ovoid Trichomonas vaginalis cells adhere to hostcells or to the substratum, they spread forming pseudopodsand filopods like amoeboid cells, and microfilament inhibitorssuch as cytochalasins B and E prevent cell adhesion (Honig-berg and Brugerolle, 1989; Gold and Ofek, 1992; Aldereteet al., 1995; Gonzalez-Robles et al., 1995; Scott et al., 1995;Brugerolle et al., 1996). The peripheral cytoplasm and thelayer along the adhesion zone become microfilamentous andwere found to be composed of actin (Brugerolle et al., 1996).Actin microfilaments are organized like a network and areadhering to the plasma membrane but no stress fibers havebeen observed (Gonzalez-Robles et al., 1995; Brugerolle et al.,1996).
Cell adhesion and signal transduction are coordinatedprocesses that may be linked through regulatory elementssuch as actin-associated proteins (Svitkina and Borisy, 1999).In T. vaginalis, actin dynamics are tightly regulated becausefilament assembly and disassembly have to occur rapidly in theparasite, at least during adhesion and lysis of host cells. Toelucidate the events underlying the reorganization of the actincytoskeleton in T. vaginalis, the identification and functionalcharacterization of regulatory proteins of actin dynamics havebeen undertaken (Bricheux et al., 1998; Addis et al., 1998).Here we report on the identification of coronin proteins andgenes in T. vaginalis a representative of one of the firstemerging lineages of eukaryotes (Embley and Hirt, 1998).Coronin derives its name from its association with crown-shaped projections from growth-phase Dictyostelium discoi-deum (de Hostos et al., 1991). Coronin is known to interactwith F-actin (for review see de Hostos, 1999) but was found tohave a primary structure unlike that of any other actin-binding
EJCB 413European Journal of Cell Biology 79, 413 ± 422 (2000, June) ´ � Urban & Fischer Verlag ´ Jenahttp://www.urbanfischer.de/journals/ejcb
0171-9335/00/79/06-413 $12.00/0
1) Dr. GenevieÁve Bricheux, Laboratoire de Biologie des Protistes,Universite Blaise Pascal, Les CeÂzeaux, ± 63177 AubieÁre, Cedex, France,e-mail: [email protected], Fax: 3347340 7670.

proteins. It belongs to the family of the WD-repeat proteinsand these repeated regions adopt a b-propeller structure,creating a stable platform that can form reversible complexeswith several proteins (Smith et al., 1999). This family of WD-repeat proteins has been implicated in many functions such assignal transduction, RNA processing, gene regulation, vesicu-lar traffic and regulation of cytoskeletal assembly (Neer et al.,1994 ). In D. discoideum coronin is found to be associated withF-actin and is enriched at the leading edge (Gerisch et al.,1995), the phagocytic cups (Maniak et al., 1995) and crownmacropinocytic structures (de Hostos et al., 1991; Hackeret al., 1997). In coronin null mutants, cell locomotion is sloweddown, cytokinesis is impaired (de Hostos et al., 1993), andphagocytosis is depressed (Maniak et al., 1995). A GFP-coronin fusion protein is recruited from cytoplasm andincorporated into the actin network of a nascent leadingedge of the cell, where it participates in the reorganization ofthe cytoskeleton (Gerisch et al., 1995; Fukui et al., 1999).Similar proteins have been identified in various organisms andcell types which are not exclusively amoeboid (de Hostos,1999). In the unicellular Plasmodium falciparum a 42-kDacoronin-like protein was found in association with actinfilaments (Tardieux et al., 1998). In vertebrates, coronin ispresent in the cytoplasm of spreading cells and concentrated atthe cell periphery in membrane ruffles and plays a role in Rac-induced lamellipodium extension and cell spreading (Mishimaand Nishida, 1999). Coronin has been shown in the set ofproteins forming the comet tail of Listeria in infectedmammalian cells (David et al., 1998). All these data showthat coronin is associated with dynamic actin filament struc-ture. A family of four coronin proteins was also described inmice and humans; three of them are ubiquitously expressed,but the expression of one is restricted to hematopoietic cells(Okumura et al., 1998). In yeast, a coronin-like protein wasevidenced in association with the actin cytoskeleton, but alsowith the microtubular network (Goode et al., 1999). Thepresent study describes the molecular characterization of acoronin-like protein from T. vaginalis associated with the actincytoskeleton and its detailed ultrastructural localization indynamic sites at the periphery of amoeboid cells.
Materials and methods
Strains and culture conditionsTrichomonas vaginalis (strain isolated by G. Brugerolle) was cultivatedaxenically on TYM medium (Diamond, 1957) at 37 8C. Cells were alsogrown on glass coverslips introduced into culture tubes to obtainamoeboid cells.
Preparation of cytoskeletal extracts and cellfractionationThe cytoskeleton extraction procedure was previously described(Brugerolle et al., 1996). Briefly, cells washed in PBS were extractedby a Tris-maleate buffer (20 mM Tris, 20 mM maleate, 10 mM EGTA,10 mM MgCl2, 0.1% NaN3, pH 7.0) containing 0.5% Triton X-100,washed and collected by low-speed centrifugation.
For cell fractionation a two-step procedure derived from a brainfodrin extraction protocol was used (Levilliers et al., 1986). Briefly,400 ml of cell suspension of T. vaginalis in late exponential phase waswashed by centrifuging three times in PBS, then once in 0.25 M sucrose.The 1 ml pellet of packed cells was lysed at 0 8C in a Tenbroeckhomogenizer at low ionic strength with 4.4 ml of 0.34 M sucrose and2 mM 2-mercaptoethanol, containing protease inhibitor cocktail
(Boehringer Mannheim, France). An average of 20 strokes wassufficient to lyse the cells (monitored by light microscopy). Aftercentrifuging for 45 min at 100000g and 4 8C, the low-ionic-strengthsupernatant was kept on ice while the pellet was re-extracted with0.3 ml of 2.1 M KCl in lysis medium (0.6 M KCl final concentration)and gentle treatment with the homogenizer. After 30 min incubation onice and centrifuging for 45 min at 100000g at 4 8C, the two low and high-ionic-strength supernatants were combined at 4 8C and re-centrifugedin the same conditions. The polymerization was triggered by addition of800 ml of 280 mM imidazole, pH 6.7, containing 0.34 M sucrose, 7 mM2-mercaptoethanol, 7 mM MgCl2, 7 mM ATP and a balanced CaCl2/EGTA of 7/10.5 mM to reach a final 1.5 mM free calcium concentration.After incubation for 1 hour at 20 8C, the extract was centrifuged for45 min at 100000g at 20 8C to sediment the F-actin. This pellet wasgently resuspended in 1 ml of 0.6 M KCl, 0.34 M sucrose, 1 mM 2-mercaptoethanol, then centrifuged for 45 min at 100000g at 4 8C.
Electrophoresis and immunoblottingCytoskeleton fractions were solubilized in boiling sample buffer andseparated on 10% SDS-PAGE (Polyacrylamide Gel Electrophoresis)mini gels according to Laemmli (Laemmli, 1970). Molecular massmarkers (Bio Rad, France) were used for molecular mass determina-tion of proteins. Gels were stained with Coomassie Brilliant Blue R-250in 40% methanol and 10% acetic acid.
Two-dimensional gel electrophoresis was performed according toO�Farell (O�Farell, 1975) using 1.6% Pharmalytes 4.0 ± 6.5 and 0.4%Pharmalytes 3.0 ± 10.0 (Pharmacia, France). Gels were either treatedfor immunoblotting or stained with Coomassie Blue after Pharmalytesextraction by two washes in 50% methanol and 10% acetic acid.
Peptides separated on one-dimensional or two-dimensional gelswere transferred onto Immobilon-P membranes (Amersham, France)with a semi-dry transfer device (BioRad, France). The membraneswere blocked for 1 h in 5% (w/v) skimmed milk in PBS and incubatedin the primary antibody for 2 h or overnight. After three washes in PBScontaining 1% skimmed milk, the blots were incubated for 2 h with theanti-mouse IgG alkaline phosphatase conjugate (Promega, France)diluted 1 : 7500 in PBS/milk, then washed twice for 10 min in PBS. Theactivity was revealed by Fast Red TR/Naphthol AS-MX phosphate(Sigma, France) staining.
Monoclonal antibody productionThe monoclonal antibodies (mAB) 4C12 and XG3 were produced inthe laboratory. Mice were immunized against T. vaginalis cytoskeletalfraction; cell fusion and clone isolation were performed by the generalprocedure of Galfre and Milstein (Galfre and Milstein, 1981). Antibodyscreening was done first by immunofluorescence and then by immuno-blotting.
Indirect immunofluorescence, electronmicroscopy and immunogold stainingCells were fixed in PBS containing 3.5% formaldehyde at roomtemperature for 20 min, then permeabilized in 0.5% Triton X-100 inPBS for 10 min. After treatment with 0.1 M glycine in PBS for 30 minand blocking with 1% BSA in PBS for 15 min, cells were incubated withthe primary antibody (mAB supernatant) for 2 h, then washed threetimes in PBS before incubation with a 1 : 200 dilution of anti-mouse Ig-FITC or Ig-TRITC conjugated antibody (Sigma, France). For double-labeling, cells were treated first with the anti-coronin mAB and anti-mouse Ig-FITC then with the anti-actin mAB and anti-mouse Ig-TRITC. After final washes, the slides were mounted with 1 : 1 PBS-glycerol containing 10 mg/ml 1.4-diazabicyclo (2-2-2) octane (Sigma,France). Control slides were treated with a 200-fold diluted pre-immune mouse serum as the first antibody. Microscopic examinationwas carried out using a Leica DMR HC microscope equipped with aStation Q-Fish Light.
For electron microscopy, cells grown on coverslips were fixed for 1 hin 1% (v/v) glutaraldehyde and 0.2% tannic acid in 0.1 M phosphatebuffer (pH 7.2). After two washes in phosphate buffer, cells were post-fixed in 1% osmium tetroxide in phosphate buffer. Cells were
414 G. Bricheux, G. Coffe et al. EJCB

counterstained ªen blocº in uranyl acetate-saturated 70% ethanol, thendehydrated and embedded in Epon 812. Ultrathin sections were stainedwith lead citrate and carbon coated before examination under a JEOLEX1200 transmission electron microscope.
For post-embedding immunogold staining, cells grown on coverslipswere fixed for 20 min in 3% paraformaldehyde (w/v) and 0.2%glutaraldehyde in PBS. Cells were then scraped, washed three times inPBS, treated for 20 min with 1 mg/ml sodium borohydride in PBS,centrifuged and pre-embedded in 1% agar. They were dehydrated inalcohol series and embedded in Unicryl resin (British BiolCell Intl,UK). Ultrathin sections were blocked for 15 min with 1% BSA in PBS,rinsed in PBS and labeled by the antibody at 1 : 200 dilution in PBSovernight. After three 10-min washes in PBS, they were incubated in1 : 50 diluted goat anti-mouse IgG antibody conjugated to 15-nm goldparticles (Aurion, The Netherlands) in 0.1% BSA-PBS. After severalwashes in PBS followed by water, sections were stained overnight inuranyl acetate-saturated water and then carbon coated before ob-servation.
Coronin coding sequencesA cDNA library of T. vaginalis in lgt11, constructed in the laboratory,was immunoscreened with the mAB 4C12 according to standardprotocols (Sambrook et al., 1989). After three rounds of purification,the insert of selected clones was amplified and cloned in pCR 2.1-TOPO vector (Invitrogen, The Netherlands). A genomic mini-librarywas constructed by digestion of T. vaginalis DNA with HindIII andcloning in pBluescript vector (Stratagene, California, USA) digested bythe same enzyme. The 5' extremity of clones was searched byamplification of the mini-library with one set of primer (ReverseM13/Primer 1) and a nested PCR with another set (SK/Primer 2),Primer 1 and Primer 2 being derived from the cDNA sequences.
Sequencing of double-stranded templates was performed by thedideoxynucleotide chain termination method using T7 polymerase(Pharmacia, France).
DNA isolation and Southern blottingCells were harvested in exponential phase by centrifugation andwashed with phosphate buffer. High-molecular-mass DNA was ex-tracted by the DEPC-Triton X-100 method (Riley and Krieger, 1992).DNA (10 mg) was digested with restriction endonucleases, size-fractionated in agarose gels and transferred onto Hybond-N�membranes (Amersham, France).
The membranes were hybridized at 60 8C with a random-primed 32P-labeled DNA probe in 5�SSPE, 0.5% SDS, 5�Denhardt�s solutionwith 100 mg of denatured salmon sperm DNA (Sambrook et al., 1989).
Results
Identification of a 50-kDa doublet proteinThe isolated cytoskeleton of T. vaginalis is composed of manyproteins, some of which have been previously identified astubulin (Katiyar and Edlind, 1994), actin (Bricheux andBrugerolle, 1997), a-actinin (Bricheux et al., 1998), or striatedroot proteins (Brugerolle and Viscogliosi, 1994). Using themAB 4C12 raised against a T. vaginalis cytoskeleton fractionwe detected two bands of about 50 kDa on immunoblots(Fig. 1b). These bands were weakly represented on CoomassieBlue stained cytoskeleton fraction (Fig. 1a).
An affinity-purified anti-D. discoideum coronin mAB wasused on immunoblots of a cytoskeleton fraction of T. vaginalis.This mAB presented a faint cross-reactivity with the two bandsof T. vaginalis whereas mAB 4C12 did not present any cross-reactivity with proteins from D. discoideum cytoskeleton orwith rabbit blood white cells (not shown).
Enrichment by cosedimentation with actinTo check whether this cross-reactive polypeptide behaved asan actin-associated protein, as shown for many coronins, wetried to obtain a maximal amount of this protein by a two-stepsolubilization process before its cosedimentation with actin(see Materials and methods). The gel containing the differentfractions (Fig. 2a) was transferred and treated with the anti-coronin mAB 4C12 (Fig. 2b). A low ionic strength extraction(Fig. 2, lane 1a ± 1b) optimizing the actin depolymerization wasfollowed by a high ionic strength treatment (Fig. 2, lanes 2a ±2b) to extract associated proteins. An appreciable amount ofcoronin, extracted at 0.6 M KCl, precipitated on combining thetwo supernatants (Fig. 2, lanes 3a ± 3b) owing to the reductionof the ionic strength down to 40 mM KCl. This step wasnecessary to assess the re-association of coronin with actin
415Characterization of Trichomonas vaginalis coroninEJCB
Fig. 1. Detection of a 50-kDa doublet proteinwith mAB 4C12. Coomassie Blue staining (a)and immunoblotting (b) of T. vaginalis cyto-skeleton (lanes 1 and 2, 20 and 10 mg) and ofcoronin-enriched fraction (lane 3, 5 mg). Molec-ular weight markers are shown in the left lane.

during its polymerization. After centrifugation, some of thecoronin remained in the supernatant (Fig. 2, lanes 4a ± 4b) andthe pellet was gently resuspended in high salt buffer to releaseassociated proteins from the actin filaments. A final centrifu-gation separated the associated proteins in the supernatant(Fig. 2, lanes 6a ± 6b) from the pelletable F-actin (Fig. 2, lanes5a ± 5b). Coronin from T. vaginalis appears to be a quiteinsoluble protein at low ionic strength and needs a highconcentration of salt to dissociate from polymerized actin.
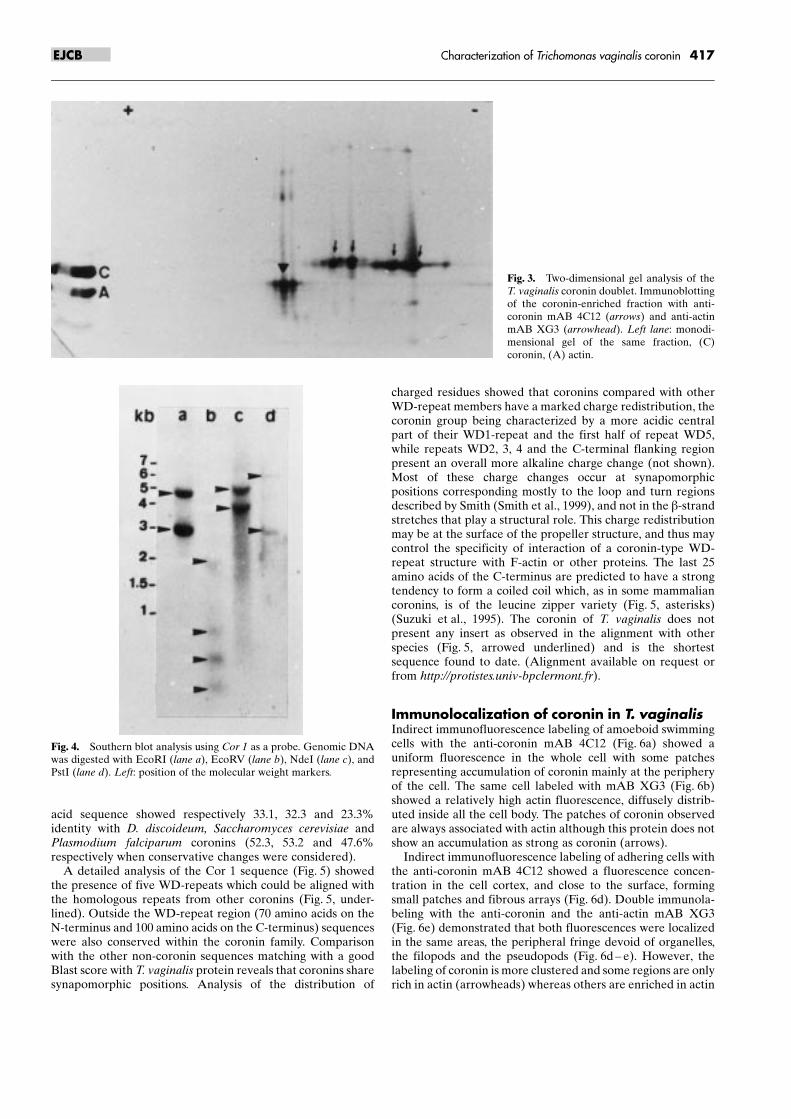
To determine the isoelectric point of the two coronin bands,the proteins present in the final fraction (Fig. 2, lane 6a) wereseparated by two-dimensional electrophoresis, transferred,and the blot treated with mAB 4C12. The two mono-dimensional bands (Fig. 3, left lane) separated into two setsof isoforms with clearly different Ip, in the range of 6 and 7,each set being represented by two major spots (Fig. 3, arrows).This corroborates the presence of two related coronins withslightly different apparent molecular masses. The same blotwas treated with the mAB XG3 to localize the different actinisoforms (Fig. 3, arrowhead).
Library screening and sequencingTo ascertain whether T. vaginalis had a protein homologous toD. discoideum coronin, as suggested by the cross-reactivityresults, we used the mAB 4C12 to screen a T. vaginalis cDNAlibrary. Four positive clones were isolated out of the 105 clonesscreened. These four clones represented two closely relatedfamilies; the longest clone of each family were called Cor 1(960 bp) and Cor 2 (1050 bp). As the two clones weretruncated at their 5' extremities, and taking advantage of thefact that T. vaginalis possesses no introns, we tried to extendtheir sequence by genomic DNA amplification of the upstreamregions with primers common to the two sequences. A band of500 bp was isolated, cloned and identified as containing theupstream region of Cor 1, an ATG starting codon plus anuntranslated region of 42 bp with a putative initiator region(TCACCTAACTTATTTTTT) (Liston and Johnson, 1998).The two clones possess a TAA termination codon, no typicalpolyadenylation signal, but a poly(A) tail. The reconstituted
clone Cor 1 was 1400 bp long with an open reading frame of1308 bp encoding a peptide of 435 residues with a calculatedmass of 48 kDa and an Ip of 5.58. To try to explain thedifference of Ip observed on two-dimensional gels, we used theprogram MWCALC (Infobiogen) to calculate the Ip of thepartial peptide Cor 2, and we compared it with the calculatedIp of Cor 1 on the same part of the sequence (aa 99 ± 435, seeFig. 5). The truncated Cor 1 had a Ip of 5.96 and Cor 2 had a Ipof 7.06. Whereas the residues differing on Cor 2 were morebasic, the two sequences were very similar (81% identity and90% homology). The difference between the calculated Ip is inclose agreement with that observed on two-dimensionalanalysis. There are a number of potentially important motifsthat are present in the coronin sequences when analysed withthe Pattern program from the AnTheProt v4.2 package(Geourjon and DeleÂage, 1993). This includes many proteinkinase C (PKC) phosphorylation sites and one tyrosine kinasephosphorylation site, which could account for the differentisoforms observed on two-dimensional immunoblots (Fig. 3).
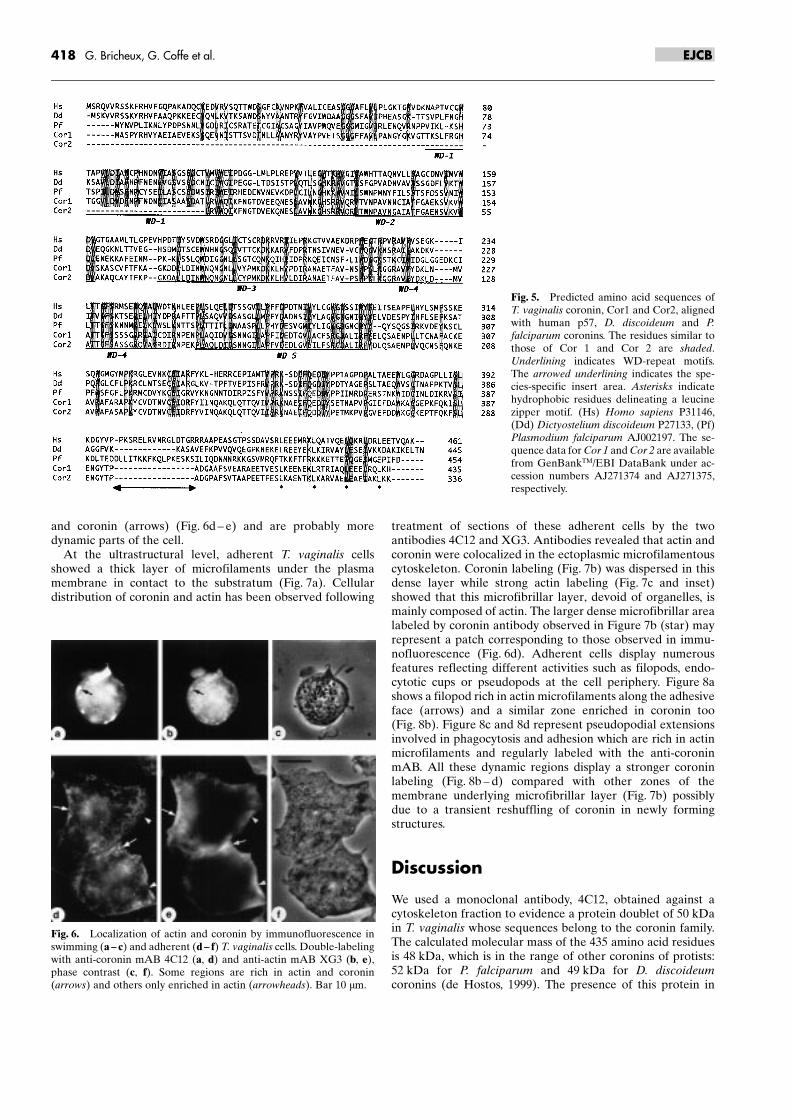
After digestion of genomic DNA by EcoRI, EcoRV, NdeIand PstI, a Southern blot analysis was performed using Cor 1 asa probe (Fig. 4). The restriction endonucleases NdeI and PstI(Fig. 4, lanes c ± d), which have no site in Cor 1 and Cor 2,produced two bands that could represent the two genes,whereas EcoRI with only one site in Cor 1 produced twobands; however, the lower one at 3 kb was more dense and mayrepresent more than one copy (Fig. 4, lane a). Digestion byEcoRV produced numerous bands below 1 kb. The samelabeling was obtained when Cor 2 was used as a probe (notshown). These results indicate that there are at least two copiesof coronin in the genome of T. vaginalis.
The deduced amino acid sequence of Cor 1 was comparedwith entries in the GenBank database with the Blast program(Altschul et al., 1997) and showed homology with coronins andother proteins belonging to the WD-repeat family. A WD-repeat comprises a 44 ± 60-residue sequence organized as asuccession of four b-strands separated by more variable loopregions (Smith et al., 1999). Cor 1 protein has a similarity withthe coronin family (Blast score in the range of 1065); the amino
416 G. Bricheux, G. Coffe et al. EJCB
Fig. 2. Cell fractionation of T. vaginalis and isolation of a coronin-enriched fraction by cosedimentation with actin. Coomassie Bluestaining (a) and immunoblotting (b) of low- and high-ionic-strengthextracts (lanes 1 and 2), insoluble material from lane 2 after lowering
ionic strength (lane 3), supernatant after actin polymerization and100000g centrifugation (lane 4), KCl-insoluble and -soluble proteinsfrom the pellet of polymerized actin (lanes 5 and 6).
acid sequence showed respectively 33.1, 32.3 and 23.3%identity with D. discoideum, Saccharomyces cerevisiae andPlasmodium falciparum coronins (52.3, 53.2 and 47.6%respectively when conservative changes were considered).
A detailed analysis of the Cor 1 sequence (Fig. 5) showedthe presence of five WD-repeats which could be aligned withthe homologous repeats from other coronins (Fig. 5, under-lined). Outside the WD-repeat region (70 amino acids on theN-terminus and 100 amino acids on the C-terminus) sequenceswere also conserved within the coronin family. Comparisonwith the other non-coronin sequences matching with a goodBlast score with T. vaginalis protein reveals that coronins sharesynapomorphic positions. Analysis of the distribution of
charged residues showed that coronins compared with otherWD-repeat members have a marked charge redistribution, thecoronin group being characterized by a more acidic centralpart of their WD1-repeat and the first half of repeat WD5,while repeats WD2, 3, 4 and the C-terminal flanking regionpresent an overall more alkaline charge change (not shown).Most of these charge changes occur at synapomorphicpositions corresponding mostly to the loop and turn regionsdescribed by Smith (Smith et al., 1999), and not in the b-strandstretches that play a structural role. This charge redistributionmay be at the surface of the propeller structure, and thus maycontrol the specificity of interaction of a coronin-type WD-repeat structure with F-actin or other proteins. The last 25amino acids of the C-terminus are predicted to have a strongtendency to form a coiled coil which, as in some mammaliancoronins, is of the leucine zipper variety (Fig. 5, asterisks)(Suzuki et al., 1995). The coronin of T. vaginalis does notpresent any insert as observed in the alignment with otherspecies (Fig. 5, arrowed underlined) and is the shortestsequence found to date. (Alignment available on request orfrom http://protistes.univ-bpclermont.fr).
Immunolocalization of coronin in T. vaginalisIndirect immunofluorescence labeling of amoeboid swimmingcells with the anti-coronin mAB 4C12 (Fig. 6a) showed auniform fluorescence in the whole cell with some patchesrepresenting accumulation of coronin mainly at the peripheryof the cell. The same cell labeled with mAB XG3 (Fig. 6b)showed a relatively high actin fluorescence, diffusely distrib-uted inside all the cell body. The patches of coronin observedare always associated with actin although this protein does notshow an accumulation as strong as coronin (arrows).
Indirect immunofluorescence labeling of adhering cells withthe anti-coronin mAB 4C12 showed a fluorescence concen-tration in the cell cortex, and close to the surface, formingsmall patches and fibrous arrays (Fig. 6d). Double immunola-beling with the anti-coronin and the anti-actin mAB XG3(Fig. 6e) demonstrated that both fluorescences were localizedin the same areas, the peripheral fringe devoid of organelles,the filopods and the pseudopods (Fig. 6d ± e). However, thelabeling of coronin is more clustered and some regions are onlyrich in actin (arrowheads) whereas others are enriched in actin
417Characterization of Trichomonas vaginalis coroninEJCB
Fig. 3. Two-dimensional gel analysis of theT. vaginalis coronin doublet. Immunoblottingof the coronin-enriched fraction with anti-coronin mAB 4C12 (arrows) and anti-actinmAB XG3 (arrowhead). Left lane: monodi-mensional gel of the same fraction, (C)coronin, (A) actin.
Fig. 4. Southern blot analysis using Cor 1 as a probe. Genomic DNAwas digested with EcoRI (lane a), EcoRV (lane b), NdeI (lane c), andPstI (lane d). Left: position of the molecular weight markers.
and coronin (arrows) (Fig. 6d ± e) and are probably moredynamic parts of the cell.
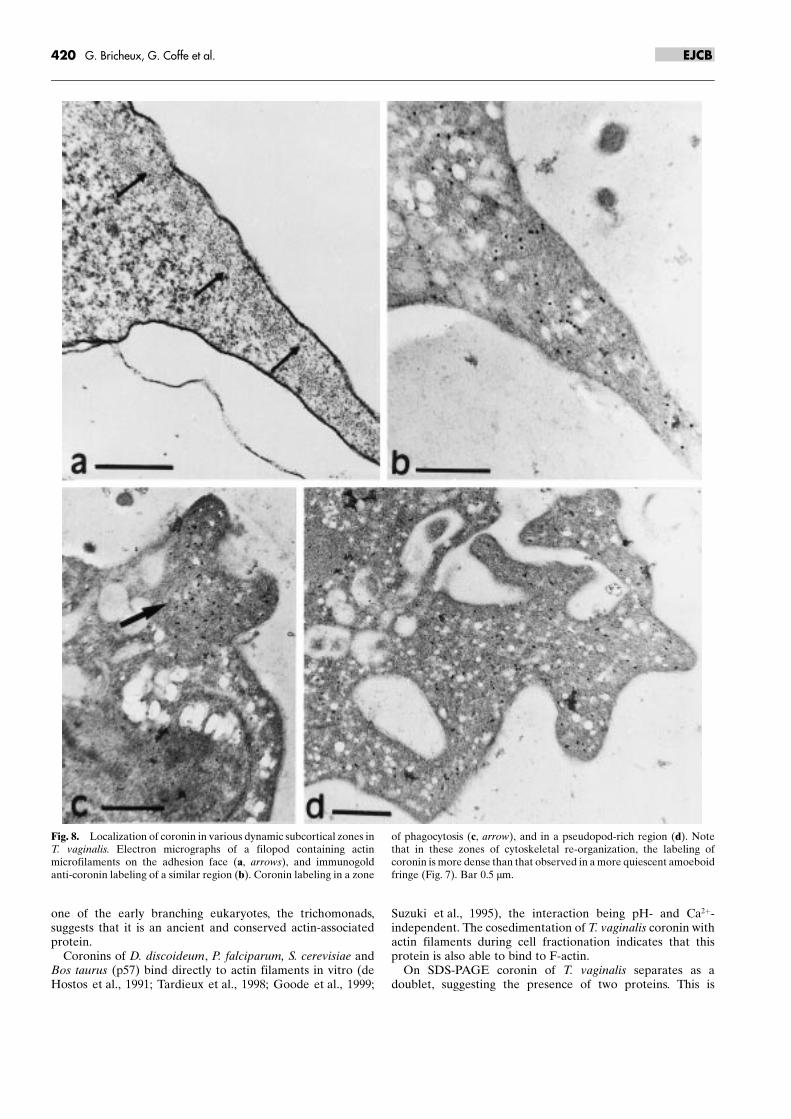
At the ultrastructural level, adherent T. vaginalis cellsshowed a thick layer of microfilaments under the plasmamembrane in contact to the substratum (Fig. 7a). Cellulardistribution of coronin and actin has been observed following
treatment of sections of these adherent cells by the twoantibodies 4C12 and XG3. Antibodies revealed that actin andcoronin were colocalized in the ectoplasmic microfilamentouscytoskeleton. Coronin labeling (Fig. 7b) was dispersed in thisdense layer while strong actin labeling (Fig. 7c and inset)showed that this microfibrillar layer, devoid of organelles, ismainly composed of actin. The larger dense microfibrillar arealabeled by coronin antibody observed in Figure 7b (star) mayrepresent a patch corresponding to those observed in immu-nofluorescence (Fig. 6d). Adherent cells display numerousfeatures reflecting different activities such as filopods, endo-cytotic cups or pseudopods at the cell periphery. Figure 8ashows a filopod rich in actin microfilaments along the adhesiveface (arrows) and a similar zone enriched in coronin too(Fig. 8b). Figure 8c and 8d represent pseudopodial extensionsinvolved in phagocytosis and adhesion which are rich in actinmicrofilaments and regularly labeled with the anti-coroninmAB. All these dynamic regions display a stronger coroninlabeling (Fig. 8b ± d) compared with other zones of themembrane underlying microfibrillar layer (Fig. 7b) possiblydue to a transient reshuffling of coronin in newly formingstructures.
Discussion
We used a monoclonal antibody, 4C12, obtained against acytoskeleton fraction to evidence a protein doublet of 50 kDain T. vaginalis whose sequences belong to the coronin family.The calculated molecular mass of the 435 amino acid residuesis 48 kDa, which is in the range of other coronins of protists:52 kDa for P. falciparum and 49 kDa for D. discoideumcoronins (de Hostos, 1999). The presence of this protein in
418 G. Bricheux, G. Coffe et al. EJCB
Fig. 5. Predicted amino acid sequences ofT. vaginalis coronin, Cor1 and Cor2, alignedwith human p57, D. discoideum and P.falciparum coronins. The residues similar tothose of Cor 1 and Cor 2 are shaded.Underlining indicates WD-repeat motifs.The arrowed underlining indicates the spe-cies-specific insert area. Asterisks indicatehydrophobic residues delineating a leucinezipper motif. (Hs) Homo sapiens P31146,(Dd) Dictyostelium discoideum P27133, (Pf)Plasmodium falciparum AJ002197. The se-quence data for Cor 1 and Cor 2 are availablefrom GenBankTM/EBI DataBank under ac-cession numbers AJ271374 and AJ271375,respectively.
Fig. 6. Localization of actin and coronin by immunofluorescence inswimming (a ± c) and adherent (d ± f) T. vaginalis cells. Double-labelingwith anti-coronin mAB 4C12 (a, d) and anti-actin mAB XG3 (b, e),phase contrast (c, f). Some regions are rich in actin and coronin(arrows) and others only enriched in actin (arrowheads). Bar 10 mm.

419Characterization of Trichomonas vaginalis coroninEJCB
Fig. 7. Immunogold localization of coronin and actin in T. vaginalis.(a) Transmission electron microscopy showing the section of anadherent cell; the cytoplasmic side of the plasma membrane is coatedwith a thick layer of microfilaments (arrowheads), nucleus (N),undulating membrane (UM). (b) Coronin labeling (mAB 4C12) is
localized in the microfilamentous fringe; a stronger coronin labeling isobserved in a zone (*) which may correspond to the patches revealed byimmunofluorescence (Fig. 6). (c and inset) actin labeling (mAB XG3)of the microfilamentous layer. Bar 1 mm (a) and 0.5 mm (b, c).
one of the early branching eukaryotes, the trichomonads,suggests that it is an ancient and conserved actin-associatedprotein.
Coronins of D. discoideum, P. falciparum, S. cerevisiae andBos taurus (p57) bind directly to actin filaments in vitro (deHostos et al., 1991; Tardieux et al., 1998; Goode et al., 1999;
Suzuki et al., 1995), the interaction being pH- and Ca2�-independent. The cosedimentation of T. vaginalis coronin withactin filaments during cell fractionation indicates that thisprotein is also able to bind to F-actin.
On SDS-PAGE coronin of T. vaginalis separates as adoublet, suggesting the presence of two proteins. This is
420 G. Bricheux, G. Coffe et al. EJCB
Fig. 8. Localization of coronin in various dynamic subcortical zones inT. vaginalis. Electron micrographs of a filopod containing actinmicrofilaments on the adhesion face (a, arrows), and immunogoldanti-coronin labeling of a similar region (b). Coronin labeling in a zone
of phagocytosis (c, arrow), and in a pseudopod-rich region (d). Notethat in these zones of cytoskeletal re-organization, the labeling ofcoronin is more dense than that observed in a more quiescent amoeboidfringe (Fig. 7). Bar 0.5 mm.

confirmed by the isolation of two different sequences byscreening a cDNA library. Moreover, Southern analysisreveals the presence of two genes for coronin (Cor 1 and Cor2). Such gene duplication was not hitherto observed in otherunicellular organisms.
The two-dimensional electrophoresis analysis shows thatcoronin is present as two sets of peptides, each one composedof several isoforms, reflecting probable post-translationalmodifications. The presence of many phosphorylation siteson Cor 1 and Cor 2 might explain the Ip heterogeneity. Aphosphorylation is likely to occur if coronin is the downstreamtarget of a PKC as suggested by phorbol ester effects on humanneutrophils (Grogan et al., 1997) and rabbit parietal cells(Parente et al., 1999).
Sequence comparison of T. vaginalis protein with othercoronins shows a relatively weak percentage of homology(from 33 to 23%), which is in agreement with the faint cross-reactivity observed with D. discoideum mAB and the lack ofcross-reactivity of mAB 4C12 on other organisms; this is to beexpected considering the large evolutionary distance betweenthe species (Embley and Hirt, 1998). However, coroninsequences of T. vaginalis share significant structural featureswith other coronins as they are composed mainly of a WD-repeat region and a C-terminal a-helical structure. The WD-repeat protein family comprises a group of proteins involved inprotein-protein interactions at several regulatory locations inthe cell and which all contain 4 to 16 copies of a conservedmotif, the WD-repeat, all these proteins being structurallyrelated but functionally diverse (Neer et al., 1994; Smith et al.,1999). The coronin group differs from the other WD-repeatproteins by a re-distribution of the charged residues along theWD-repeat regions while not affecting the structural residuesforming b-strands. This suggests that coronins could have apropeller structure similar to that of the b-subunit of G-protein, acting as a platform used for protein-protein interac-tion, as required for a regulatory protein (Smith et al., 1999). Itis not clear which part of the coronin is responsible for actinbinding. In yeast, the N-terminus plus the WD region has beenshown to be responsible for binding to actin filaments (Goodeet al., 1999) whereas in Xenopus the WD-repeat domain plusthe N- and C-terminal domains are required for binding to F-actin (Mishima and Nishida, 1999). The C-terminal region of T.vaginalis contains a leucine zipper also present in human andbovine coronins. Such a sequence is absent in P. falciparum andS. pombe and weakly conserved in D. discoideum. A hydro-phobic cluster analysis (Callebaut et al., 1997) (not shown)showed that except for P. falciparum the coronin tails of theother organisms present an a-helical structure prone toestablish a coiled-coil interaction. In yeast this region wasshown to be essential for cross-linking F-actin in vitro (Goodeet al., 1999), probably through coronin dimerization. Yeastcoronin has been shown to interact with microtubules, inaddition to actin filaments, through a unique insert betweenthe WD-repeat region and the C-terminal region. Such aspecies-specific region, a potential carrier of a regulatory orprotein binding site, does not exist in the coronin sequence ofT. vaginalis.
Consecutive to the cell shape change from a motile flagellateto an adhering amoeboid cell, coronin labeling appears toconcentrate at the periphery in the fringe of the adhering celland in many patches. At the ultrastructural level, coroninaccumulates notably in phagocytic cups and in pseudopodswhich represent dynamic areas in T. vaginalis amoeboid cells
(Rendon-Maldonado et al., 1998). These findings are consis-tent with the observation that coronin is reversibly recruitedfrom the cytoplasm and is incorporated into the actin networkat the leading edge of lamellipodia in vertebrate cells(Mishima and Nishida, 1999), in pseudopods and phagocyticcups in D. discoideum (Maniak et al., 1995; Fukui et al., 1999)or associated with the motile actin patches in yeast (Goodeet al., 1999). Yeast coronin has been reported to promote invitro polymerization of actin, with a probable nucleating roleat the barbed end of actin filaments. Conversely, the multi-protein Arp2/3 complex promotes actin polymerization at thepointed end of actin filaments (Machesky and Gould, 1999).While the effects of both coronin and Arp2/3 complex are notclearly understood (nucleation or increased rate of elongation)the possibility of a cooperation between these two systemsremains attractive.
Topical contributions in the parasitic relation of T. vaginaliswith its host cells have been proposed for adhesin (AP 33, 51and 65) candidates (Engbring et al., 1996) and more recently inTritrichomonas foetus for Tf190 adhesin containing carbohy-drate and lipid constituents (Shaia et al., 1998). T. vaginalisundergoes a dramatic cell shape change that is probablycoupled to a major switch in its metabolism and geneexpression when it activates a still imperfectly describedmechanism of adherence through an adhesin-mediated engine.In the absence of any definitely identified membrane receptorproteins, coronin is one of the most fascinating key proteins inthe cytoskeleton regulation of this parasite in response to itsinteraction with host cells. Although the whole causal chain ofevents leading to the host cell lysis remains to be fullyelucidated, we can expect the presence of a signaling pathwayin which coronin is likely to play a major downstream role inre-organizing the cytoskeleton to adapt the parasite to anamoeboid form.
Acknowledgements. We thank Günther Gerisch for providing the anti-coronin antibody of D. discoideum and JeÂroà me Garin for providing theD. discoideum whole cell proteins. We thank Annie GueÂdon and Jean-Luc Vincenot for technical assistance. This work was supported by theCNRS (Centre National de la Recherche Scientifique).
References
Addis, M. F., Rappelli, P., Delogu, G., Carta, F., Cappuccinelli, P., Fiori,P. L. (1998): Cloning and molecular characterization of a cDNAclone coding for Trichomonas vaginalis alpha-actinin and intracel-lular localization of the protein. Infect. Immun. 66, 4924 ± 4931.
Alderete, J. F., Lehker, M. W., Arroyo, R. (1995): The mechanisms andmolecules involved in cytoadherence and pathogenesis of Tricho-monas vaginalis. Parasitol. Today 11, 70 ± 74.
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z.,Miller, W., Lipman, D. J. (1997): Gapped BLASTand PSI-BLAST: anew generation of protein database search programs. Nucleic AcidsRes. 25, 3389 ± 3402.
Ayscough, K. R. (1998): In vivo functions of actin-binding proteins.Curr. Opin. Cell Biol. 10, 102 ± 111.
Bricheux, G. , Brugerolle, G. (1997): Molecular cloning of actin genes inTrichomonas vaginalis and phylogeny inferred from actin sequences.FEMS Microbiol. Lett. 153, 205 ± 213.
Bricheux, G., Coffe, G., Pradel, N., Brugerolle, G. (1998): Evidence foran uncommon a-actinin protein in Trichomonas vaginalis. Mol.Biochem. Parasitol. 95, 241 ± 249.
421Characterization of Trichomonas vaginalis coroninEJCB

Brugerolle, G., Bricheux, G., Coffe, G. (1996): Actin cytoskeletondemonstration in Trichomonas vaginalis and in other trichomonads.Biol. Cell 88, 29 ± 36.
Brugerolle, G. , Viscogliosi, E. (1994): Organization and composition ofthe striated roots supporting the Golgi apparatus, the so-calledparabasal apparatus, in parabasalid flagellates. Biol. Cell 81, 277 ±285.
Callebaut, I., Labesse, G., Durand, P., Poupon, A., Canard, L.,Chomilier, J. (1997): Deciphering protein sequence informationthrough hydrophobic cluster analysis. Current status and perspec-tives. Cell. Mol. Life Sci. 53, 621 ± 645.
Chen, H., Bernstein, B. W., Bamburg, J. R. (2000): Regulating actin-filament dynamics in vivo. Trends Biochem. Sci. 25, 19 ± 23.
David, V., Gouin, E., Van Troys, M., Grogan, A., Segal, A. W., Ampe,C., Cossart, P. (1998): Identification of cofilin, coronin, Rac and capZin actin tails using a Listeria affinity approach. J. Cell Sci. 111, 2877 ±2884.
De Hostos, E. L. (1999): The coronin family of actin-associatedproteins. Trends Cell Biol. 9, 345 ± 350.
De Hostos, E. L., Bradtke, B., Lottspeich, F., Guggenheim, R., Gerisch,G. (1991): Coronin, an actin binding protein of Dictyosteliumdiscoideum localized to cell surface projections, has sequencesimilarities to G protein b subunits. EMBO J. 10, 4097 ± 4104.
De Hostos, E. L., Rehfuess, C., Bradtke, B., Waddell, D. R., Albrecht,R., Murphy, J., Gerisch, G. (1993): Dictyostelium mutants lacking thecytoskeletal protein coronin are defective in cytokinesis and cellmotility. J. Cell Biol. 120, 163 ± 173.
Diamond, L. S. (1957): The establishment of various trichomonads ofanimals and man in axenic cultures. J. Parasitol. 43, 488 ± 490.
Embley, T. M., Hirt, R. P. (1998): Early branching eukaryotes? Curr.Opin. Genet. Dev. 8, 624 ± 629.
Engbring, J. A., O�Brien, J. L., Alderete, J. F. (1996): Trichomonasvaginalis adhesin proteins display molecular mimicry to metabolicenzymes. Adv. Exp. Med. Biol. 408, 207 ± 233.
Fukui, Y., Engler, S., InoueÂ, S., de Hostos, E. L. (1999): Architecturaldynamics and gene replacement of coronin suggest its role incytokinesis. Cell Motil. Cytoskeleton 42, 204 ± 217.
GalfreÂ, G., Milstein, C. (1981): Preparation of monoclonal antibodies.Strategies and procedures. Methods Enzymol. 73, 3 ± 47.
Geourjon, C., DeleÂage, G. (1993): Interactive and graphic couplingbetween multiple alignments, secondary structure predictions andmotif/pattern scanning into proteins. Comp. Appl. Biosc. 9, 87 ± 91.
Gerisch, G., Albrecht, R., Heizer, C., Hodgkinson, S., Maniak, M.(1995): Chemoattractant-controlled accumulation of coronin at theleading edge of Dictyostelium cells monitored using a greenfluorescent protein-coronin fusion protein. Curr. Biol. 5, 1280 ± 1285.
Gold, D., Ofek, I. (1992): Adhesion of Trichomonas vaginalis to plasticsurfaces: requirement for energy and serum constituents. Parasitol-ogy 105, 55 ± 62.
Gonzalez-Robles, A., Lazaro-Haller, A., Espinosa-Cantellano, M.,Anaya-Velazquez, F., Martinez-Palomo, A. (1995): Trichomonasvaginalis: ultrastructure bases of the cytopathic effect. J. Euk.Microbiol. 42, 5641 ± 5651.
Goode, B. L., Wong, J. J., Butty, A. C., Peter, M., McCormack, A. L.,Yates, J. R., Drubin, D. G., Barnes, G. (1999): Coronin promotes therapid assembly and cross-linking of actin filaments and may link theactin and microtubule cytoskeletons in yeast. J. Cell Biol. 144, 83 ±98.
Grogan, A., Reeves, E., Keep, N., Wientjes, F., Totty, N. F., Burlingame,A. L., Hsuan, J. J., Segal, A. W. (1997): Cytosolic phox proteinsinteract with and regulate the assembly of coronin in neutrophils. J.Cell Sci. 110, 3071 ± 3081.
Hacker, U., Albrecht, R., Maniak, M. (1997): Fluid-phase uptake bymacropinocytosis in Dictyostelium. J. Cell Sci. 110, 105 ± 112.
Honigberg, B. M., Brugerolle, G. (1989): Structure. In B. M. Honigberg(ed.): Trichomonads Parasitic in Humans. Springer-Verlag, NewYork, pp. 5 ± 35.
Katiyar, S. K. , Edlind, T. D. (1994): Beta-tubulin genes of Trichomonasvaginalis. Mol. Biochem. Parasitol. 64, 33 ± 42.
Laemmli, U. K. (1970): Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680 ± 685.
Levilliers, N., PeÂron-Renner, M., Coffe, G., Pudles, J. (1986): Gelationand fodrin purification from rat brain extracts. Biochem. Biophys.Acta 882, 113 ± 126.
Liston, D. R., Johnson, P. J. (1998): Gene transcription in Trichomonasvaginalis. Parasitol. Today 14, 261 ± 265.
Machesky, L. M., Gould, K. L. (1999): The Arp2/3 complex: a multi-functional actin organiser. Curr. Opin. Cell Biol. 11, 117 ± 121.
Maniak, M., Rauchenberger, R., Albrecht, R., Murphy, J., Gerisch, G.(1995): Coronin involved in phagocytosis: Dynamics of particle-induced relocalization visualized by a green fluorescent protein tag.Cell 83, 915 ± 924.
Mishima, M., Nishida, E. (1999): Coronin localizes to leading edges andis involved in cell spreading and lamellipodium extension invertebrate cells. J. Cell Sci. 112, 2833 ± 2842.
Neer, E. J., Schmidt, C. J., Nambudripad, R., Smith, T. F. (1994): Theancient regulatory-protein family of WD-repeat proteins. Nature371, 297 ± 300.
O�Farell, P. H. (1975): High resolution two-dimensional electrophoresisof protein. J. Biol. Chem. 250, 4007 ± 4021.
Okumura, M., Kung C., Wong, S., Rodgers, M., Thomas, M. L. (1998):Definition of family of coronin-related proteins conserved betweenhumans and mice: close genetic linkage between coronin-2 andCD45-associated protein. DNA Cell Biol. 17, 779 ± 787.
Parente, J. A., Chen, X., Zhou, C., Petropoulos, A. C., Chew, C. S.(1999): Isolation, cloning, and characterization of a new mammaliancoronin family member, coronin(se), which is regulated within theprotein kinase C signaling pathway. J. Biol. Chem. 274, 3017 ± 3025.
Rendon-Maldonado, J. G., Espinosa-Cantellano, M., Gonzales-Robles,A., Martinez-Palomo, A. (1998): Trichomonas vaginalis: In vitrophagocytosis of lactobacilli, vaginal epithelial cells, leukocytes anderythrocytes. Exp. Parasitol. 89, 241 ± 250.
Riley, D. E., Krieger, J. N. (1992): Rapid and practical DNA isolationfrom Trichomonas vaginalis and other nuclease-rich protozoa. Mol.Biochem. Parasitol. 51, 161 ± 164.
Sambrook, J., Fritsch, E. F., Maniatis, T. (1989): Molecular Cloning: ALaboratory Manual. Cold Spring Harbor Laboratory Press, NewYork.
Scott, D. A., North, M. J., Coombs, G. H. (1995): Trichomonas vagina-lis: amoeboid and flagellated forms synthesize similar proteinases.Exp. Parasitol. 80, 345 ± 348.
Shaia, C. I., Voyich, J., Gillis, S. J., Singh, B. N., Burgess, D. E. (1998):Purification and expression of the Tf190 adhesin in Trichomonasvaginalis. Infect. Immun. 66, 1100 ± 1105.
Smith, T. F., Gaitatzes, C., Saxena, K., Neer, E. J. (1999): The WDrepeat: a common architecture for diverse functions. TrendsBiochem. Sci. 24, 181 ± 185.
Suzuki, K., Nishihata, J., Arai, Y., Honma, N., Yamamoto, K., Irimura,T., Toyoshimata, S. (1995): Molecular cloning of a novel actin-binding protein, p57, with a WD repeat and a leucine zipper motif.FEBS Lett. 364, 283 ± 288.
Svitkina, T. M., Borisy, G. G. (1999): Progress in protrusion: the tell-talescar. Trends Biochem. Sci. 24, 432 ± 436.
Tardieux, I., Liu, X., Poupel, O., Parzi, D., Dehoux, P., Langsley, G.(1998): A Plasmodium falciparum novel gene encoding a coronin-like protein which associates with actin filaments. FEBS Lett. 441,251 ± 256.
422 G. Bricheux, G. Coffe et al. EJCB




















