
Copper, zinc superoxide dismutase of Curcuma aromatica is a kinetically stable protein
Upload
independentCategory
view
1download
0
http://jhc.sagepub.com/Journal of Histochemistry & Cytochemistry
http://jhc.sagepub.com/content/45/12/1611The online version of this article can be found at:
DOI: 10.1177/002215549704501204
1997 45: 1611J Histochem CytochemSandra Moreno, Roberta Nardacci and Maria Paola Cerù
Nervous SystemRegional and Ultrastructural Immunolocalization of Copper-Zinc Superoxide Dismutase in Rat Central
Published by:
http://www.sagepublications.com
On behalf of:
Official Journal of The Histochemical Society
can be found at:Journal of Histochemistry & CytochemistryAdditional services and information for
http://jhc.sagepub.com/cgi/alertsEmail Alerts:
http://jhc.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Dec 1, 1997Version of Record >>
by guest on July 10, 2014jhc.sagepub.comDownloaded from by guest on July 10, 2014jhc.sagepub.comDownloaded from

Volume 45(12): 1611–1622, 1997The Journal of Histochemistry & Cytochemistry
© The Histochemical Society, Inc.
0022-1554/97/$3.30
1611
ARTICLE
Regional and Ultrastructural Immunolocalization of Copper–Zinc Superoxide Dismutase in Rat Central Nervous System
Sandra Moreno, Roberta Nardacci, and Maria Paola Cerù
Dipartimento di Biologia di Base ed Applicata, Università dell’Aquila, Coppito, L’Aquila, Italy (SM,MPC), and Dipartimento di Biologia Cellulare e dello Sviluppo, Università di Roma “La Sapienza,” Rome, Italy (RN)
SUMMARY
We examined the distribution of copper–zinc superoxide dismutase (CuZn-SOD) in adult rat central nervous system by light and electron microscopic immunocy-tochemistry, using an affinity-purified polyclonal antibody. The enzyme appeared to be ex-clusively localized in neurons. No immunoreactivity was seen in non-neuronal cells. Thestaining intensity was variable, depending on the brain region and, within the same re-gion, on the neuron type. Highly immunoreactive elements included cortical neuronsevenly distributed in the different layers, hippocampal interneurons, neurons of the reticu-lar thalamic nucleus, and Golgi, stellate, and basket cells of the cerebellar cortex. Otherneurons, i.e., pyramidal cells of the neocortex and hippocampus, Purkinje and granule cellsof the cerebellar cortex, and the majority of thalamic neurons, showed much weaker stain-ing. In the spinal cord, intense CuZnSOD immunoreactivity was present in many neurons,including motor neurons. Pre-embedding immunoelectron microscopy of the neocortex,hippocampus, reticular thalamic nucleus, and cerebellar cortex showed cytosolic and nucle-oplasmic labeling. Moreover, single membrane-limited immunoreactive organelles identi-fied as peroxisomes were often found, even in neurons that appeared weakly stained atthe light microscopic level. In double immunogold labeling experiments, particulate CuZn-SOD immunoreactivity co-localized with catalase, a marker enzyme for peroxisomes, thusdemonstrating that in neural tissue CuZnSOD is also present in peroxisomes.
(J Histochem Cytochem 45:1611–1622, 1997)
R
eactive oxygen species
(ROS), such as superoxideanion (O2–•), hydroxyl radical (OH•), and hydrogenperoxide (H
2
O
2
), are produced as a consequence of thephysiologic metabolism in the central nervous system(CNS). Several biochemical and physiological featuresrender the nervous tissue especially vulnerable to oxi-dative damage. These include a high rate of oxidativemetabolic activity, high concentration of molecules (i.e.,polyunsatured fatty acids) that are susceptible to ROSattack, and low levels of scavenging molecules. ROShave been implicated in brain aging processes as wellas in ischemia and neurodegenerative diseases, such asParkinson’s disease, Alzheimer’s dementia, multiple scle-rosis, and amyotrophic lateral sclerosis (for reviews see
Halliwell 1992; LeBel and Bondy 1992; Evans, 1993;Rowland 1995).
Both enzymatic and nonenzymatic systems are in-volved in the removal of ROS. Superoxide dismutases(SODs) are a family of metalloenzymes that catalyzethe dismutation of O2–• to H
2
O
2
, which, in turn, is re-duced to water by catalase or peroxidases. Mamma-lian cells possess three forms of SODs with distinct lo-calizations, i.e., extracellular SOD (EcSOD), manganeseSOD (MnSOD), and copper–zinc SOD (CuZnSOD)(Fridovich 1989).
MnSOD is located in the mitochondria, and CuZn-SOD is predominantly found in the cytosol and nucle-oplasm (Slot et al. 1986). Recent biochemical andmorphological studies have shown that CuZnSOD isalso present in peroxisomes of rat hepatocytes and fi-broblasts (Dhaunsi et al. 1992; Wanders and Denis1992) and of human fibroblasts and hepatoma cells(Keller et al. 1991). Peroxisomes are cytoplasmic or-
Correspondence to: Prof. Maria Paola Cerù, Dip. di Biologia diBase ed Applicata, Università dell’Aquila, Via Vetoio, 10 - Coppito,67010 L’Aquila, Italy.
Received for publication November 5, 1996; accepted July 10,1997 (6A4130).
KEY WORDS
free radicals
antioxidants
catalase
peroxisomes
immunohistochemistry
biotin amplification
tyramine conjugates
neurons
rat brain
by guest on July 10, 2014jhc.sagepub.comDownloaded from

1612
Moreno, Nardacci, Cerù
ganelles containing many H
2
O
2
-producing oxidases andcatalase, and are therefore primarily involved in oxy-gen metabolism. The presence of CuZnSOD in theperoxisomes, however, is still controversial (Crapo etal. 1992; Kobayashi et al. 1993) and to date has notbeen demonstrated in the nervous system.
The importance of CuZnSOD in the CNS is empha-sized by the finding that this enzyme exerts a protec-tive action against brain injury and neuronal death bothin vitro and in vivo (Imaizumi et al. 1990; Matsumiyaet al. 1991; Tagaya et al. 1992; Uyama et al. 1992; Heet al. 1993; Greenlund et al. 1995). Moreover, trans-genic mice overexpressing the human CuZnSOD genewere found to be more resistant than their nontrans-genic normal littermates to oxidative stress, includingischemia–reperfusion injury (Chan 1994). Conversely,loss of endogenous CuZnSOD anticipates the neu-ronal pathological changes due to focal or global cere-bral ischemia in rat (Liu et al. 1993,1994).
CuZnSOD has also been involved in other neurode-generative diseases, such as Parkinson’s disease (Sagguet al. 1989; Marttila et al. 1990; Nishiyama et al.1995), in which an imbalance between free radicalproduction and their scavengers has been demon-strated (Yoshikawa 1993).
Interest in elucidating the precise role of CuZnSODin neural tissue has furtherly increased in recent years,because point mutations in the gene encoding this en-zyme have been detected in a proportion of familieswith familial amyotrophic lateral sclerosis (FALS) (Denget al. 1993; Rosen et al. 1993; Gurney et al. 1994).
Despite the importance of CuZnSOD in neural tis-sue, light and electron microscopic studies on the lo-calization of this enzyme are few and are focused onrestricted brain areas (Thaete et al. 1986; Liu et al.1994; Pardo et al. 1995).
In this article we describe the distribution of CuZn-SOD in the rat CNS by use of a recently developed im-munocytochemical procedure that allows great signalamplification (Adams 1992; Moreno et al. 1995). Wealso performed immunoelectron microscopy in se-lected brain areas, i.e., the neocortex, the hippocam-pus, the reticular thalamic nucleus, and the cerebellarcortex, with the aim of investigating the intracellulardistribution of the enzyme, with particular regard tothe peroxisomes.
A preliminary report of our results has been pub-lished (Moreno et al. 1994).
Materials and Methods
Antibodies
Affinity-purified rabbit polyclonal anti-CuZnSOD antibodywas a generous gift of Dr. M. Giorgi, University of L’Aquila,Italy (Rossi et al. 1994). As antigen, CuZnSOD purified from
rat liver according to Yano (1990) (a generous gift of Prof.G. Rotilio, University “Tor Vergata,” Rome, Italy) was used.
Sheep polyclonal anti-catalase antibody was a generousgift from Prof. D. Fahimi (University of Heidelberg, Ger-many).
Immunoblotting
The specificity of the affinity-purified anti-CuZnSOD wastested by immunoblotting. This was performed as follows.Rat liver CuZnSOD, goat erythrocyte CuZnSOD (a gener-ous gift from Prof. L. Calabrese, University of Rome 3, Italy),rat brain, and rat liver homogenates, suspended in 60 mMTris-HCl buffer, pH 6.8, containing 2% sodium dodecyl sul-fate, 5%
b
-mercaptoethanol, 10% glycerol, and 0.001%bromophenol blue, were heated to 92C for 15 min to ensurethat all the native proteins were denatured to the monomericform. Thirty ng of the purified proteins, 60
m
g and 100
m
gof liver and brain proteins, respectively, were loaded onto a15% polyacrylamide gel. SDS-PAGE electrophoresis wasperformed according to Laemmli (1970). Western blottingwas performed according to Towbin et al. (1979). Immun-ostaining was obtained by incubating the nitrocellulose sheetswith the primary antibody (1:5000) and then with goat anti-rabbit IgG conjugated with alkaline phosphatase (1:2000)(Sigma Chemical; St Louis, MO). The immunocomplexeswere revealed according to Kincaid (1988).
Controls were performed by omission of the primary an-tibody. Because of the scarcity of available antigen, we wereunable to perform controls with the primary antibody previ-ously adsorbed with excess antigen.
Animals
Twenty adult Wistar rats of either sex, weighing about 250 gand kept on a standard diet and water ad libitum, weredeeply anesthetized with pentobarbital (35 mg/kg). Animalswere housed and handled according to approved guidelines.
Light Microscopic Immunocytochemistry
Animals were perfused through the aorta at room tempera-ture (RT) with Ringer’s variant without calcium, pH 7.3,followed by 4% freshly depolymerized paraformaldehydewith or without 0.05% glutaraldehyde in 0.12 M phosphatebuffer, pH 7.3. One hr after perfusion the brains were dis-sected and cut coronally or sagittally on a Vibratome at25
m
m. Sections were collected in 0.12 M phosphate buffer,pH 7.4, and then transferred to PBS containing 2% nonfatdry milk and 0.05% Triton X-100 for 1 hr. Sections werethen incubated with anti-CuZnSOD diluted 1:6000–1:12,000in PBS containing 1% nonfat dry milk and 0.02% TritonX-100 overnight at 4C. After rinsing with buffer, sectionswere processed applying an amplification procedure of theavidin–biotin method (Adams 1992), with the modificationspreviously described (Moreno et al. 1995). Controls per-formed by omission of the primary antibody were negative.
Biotinylated secondary antibody, normal goat serum (NGS),and avidin–biotin–horseradish peroxidase (HRP) (standardABC kit) were purchased from Vector (Burlingame, CA),NHS–biotin from Pierce Chemical (Rockford, IL), and tyra-mine and 3,3
9
-diaminobenzidine (DAB) from Sigma.
by guest on July 10, 2014jhc.sagepub.comDownloaded from

Copper–Zinc Superoxide Dismutase in Rat Brain
1613
Pre-embedding Immunoelectron Microscopy
Specimens were treated for immunocytochemistry as above.After the immunoreaction, sections were rinsed with bufferand then postfixed in OsO
4
for 1 hr, dehydrated, and flat-embedded in Epon. Selected areas were remounted on Eponblanks and sectioned on a Reichert Ultracut ultramicrotome.Ultrathin sections were briefly contrasted with uranyl ace-tate and photographed in a Philips 400 T electron micro-scope.
Post-embedding Double Immunogold Labeling
Vibratome coronal sections 100
m
m thick were collected in0.12 M phosphate buffer, pH 7.4, and then dehydrated ingraded alcohols, progressively lowering the temperature (from4C to
2
20C). Sections were infiltrated with Unicryl (BritishBioCell; Cardiff, UK), according to Scala et al. (1992), at
2
20C for 48 hr, and then at RT overnight. Polymerizationwas carried out at
2
10C under UV light.Other 100
m
m Vibratome sections were dehydrated andembedded in London Resin White (LR White) at 4C.
Ultrathin sections were sequentially incubated with (a)NGS diluted 1:10 in 10 mM PBS containing 1% bovine se-rum albumin (BSA) (Medium A) for 15 min, (b) rabbit anti-CuZnSOD diluted 1:40 in Medium A, overnight, (c) goatanti-rabbit IgG conjugated to 10-nm colloidal gold (BritishBioCell), diluted 1:30 in Medium A containing 0.01% gela-tin for 1 hr, (d) sheep anti-catalase diluted 1:10 in MediumA, overnight, and (e) donkey anti-sheep IgG conjugated to5-nm colloidal gold (British BioCell), diluted 1:30 in Me-dium A containing 0.01% gelatin for 1 hr. All incubationswere carried out at RT, (except for b and d, at 4C). Gridswere briefly contrasted with uranyl acetate and photo-graphed in a Philips 400 T electron microscope.
Results
Immunoblotting
Figure 1 shows the immunoblotting obtained by usingthe affinity-purified antibody produced against rat liverCuZnSOD. The antibody recognizes both the rat liverantigen used for immunization and goat erythrocyteCuZnSOD. In rat liver and brain homogenates, theantibody recognizes only one band at the MW corre-sponding to the CuZnSOD monomer (17 kD). Con-trols are negative.
Light Microscopic Immunocytochemistry
[Note: For all Figures the terminology follows that ofPaxinos and Watson (1986).] Immunoreactivity toCuZnSOD was seen exclusively in neurons. No stain-ing was detectable in glial or endothelial cells. The in-tensity of immunoreaction was variable, depending onthe area and on the neuronal type considered.
In the main olfactory bulb, appreciable stainingwas observed in periglomerular cells in the glomerularlayer, in a few neurons located in the external plexi-
form layer, in mitral cells, and in many elements of thegranule cell layer (Figure 2A).
The piriform cortex appeared extremely rich inCuZnSOD-positive neurons. Because of their number,size, and location in the three layers and in the subjacentendopiriform nucleus, most of the positive cells wereinterpreted as GABAergic interneurons (not shown).
In the neocortex, especially in the frontal area,many highly immunoreactive neurons were seen in allthe layers (Figure 2B). On the basis of their shape andorientation, most of these cells were identified asGABAergic interneurons (Mugnaini and Oertel 1985).Pyramidal cells showed weaker immunoreactivity.
In the telencephalon, immunostaining was detectedin many cells of the caudate putamen, including spinyneurons and presumably cholinergic interneurons (Fig-ure 2C), in the ventral pallidum, in the islands of Calleja(Figure 2D), and in some neurons of the septal nuclei(Figure 2E).
In the hippocampal formation, the immunoreactionproduct was concentrated in interneurons. These cellswere evenly distributed in all layers of Ammon’s horn,the dentate gyrus, and the subiculum (Figures 2F–2H).Hippocampal pyramidal cells and granule cells wereweakly positive.
In the diencephalon, the most remarkable degree ofstaining was seen in the reticular nucleus of the thala-mus (Figure 3A). The anterodorsal thalamic nucleusand lateral and medial geniculate nuclei also showeddistinct labeling, whereas other ventral and dorsal tha-lamic nuclei displayed little immunoreactivity.
In the mesencephalon, intense immunoreactivitywas found in neurons of all the layers of superior colli-culus (Figure 3B). A moderate positivity was also de-tected in the central gray substance and in the inferiorcolliculus (Figure 3C). The substantia nigra displayedintense immunoreactivity in both pars reticulata andpars compacta. Staining was more evident in the neu-ronal cytoplasm than in the nucleus (Figure 3D). The
Figure 1 Immunoblotting with the affinity-purified anti-CuZnSODpolyclonal antibody. (A) Lane 0, prestained molecular weight stan-dards; Lane 1, rat liver homogenate (60 mg of protein); Lane 2, ratbrain homogenate (100 mg of protein); Lane 3, rat liver CuZnSOD(30 ng of protein); Lane 4, goat erythrocyte CuZnSOD (30 ng ofprotein). B Lanes 19–49 as in (A) but incubated without the primaryantibody.
by guest on July 10, 2014jhc.sagepub.comDownloaded from

1614
Moreno, Nardacci, Cerù
by guest on July 10, 2014jhc.sagepub.comDownloaded from

Copper–Zinc Superoxide Dismutase in Rat Brain
1615
magnocellular part of the red nucleus also showedsome positivity, the staining being confined to the neu-ronal cytoplasm (Figure 3E). Many other nuclei of themesencephalon, such as dorsal and ventral nuclei ofthe lateral lemniscus, raphe nuclei, and interpedun-cular nucleus, appeared rather rich in CuZnSOD (Fig-ure 3F).
In the hindbrain, almost all the centers appeareddistinctly positive for CuZnSOD. The nuclei that ex-hibited the highest level of immunoreactivity includedthe pontine nuclei, nucleus abducens, supragenual nu-cleus, gigantocellular reticular nucleus, ventral and dor-sal cochlear nuclei, spinal trigeminal nucleus, principaltrigeminal sensory nucleus, nucleus of the trapezoidbody, facial nucleus, and lateral vestibular nucleus(Figure 3G–3J).
In the cerebellum, small neurons of deep cerebellarnuclei were moderately positive (Figure 3K). In thecerebellar cortex, remarkable immunostaining was seenin Golgi cells of the granular layer and in basket andstellate cells of the molecular layer, whereas granuleand Purkinje neurons were negative (Figure 3L).
In the spinal cord, neurons of all the Rexed’s lami-nae, comprising motor neurons and interneurons, dis-played intense immunostaining (Figures 4A and 4B).In many areas, the positive neurons showed immu-noreaction product in both their cytoplasm and nu-cleus. However, neurons of certain areas, such as mi-tral cell layer, substantia nigra, red nucleus, abducensnucleus, and reticular gigantocellular nucleus, deepcerebellar nuclei primarily showed cytoplasmic immu-noreactivity (Figures 2A, 3D, 3E, 3G, 3H,and 3K). Onthe contrary, other neurons, such as Golgi cells of thecerebellar cortex, primarily displayed nuclear positiv-ity (Figure 3L). In all the neurons, the nucleolus ap-peared negative (e.g., inset in Figure 3L).
Pre-embedding Electron Microscopic Immunocytochemistry
In all the brain regions examined at the ultrastructurallevel, CuZnSOD immunoreactivity was seen exclu-sively in neurons. The immunoreaction product wasrather homogeneously diffused in the cytosol and inthe nucleoplasm of the positive neurons. Densely stainedcytoplasmic bodies, bounded by a single membrane andmeasuring 0.2–0.3
m
m in diameter, were often observed.Endoplasmic reticulum, Golgi apparatus, vacuoles,mitochondria, and nucleolus were always unstained.
In the cerebral neocortex the immunoreaction prod-uct appeared concentrated in heterogeneous nonpyra-midal neurons, ranging from 7 to 18
m
m in mean di-ameter (Figure 4C).
In the hippocampus, intense positivity was seen insmall interneurons (not shown). Weakly immunoreac-tive pyramidal cells were occasionally observed. Inthese cases, the nucleus showed a relatively strongerlabeling than the cytoplasm. These cells sometimescontained positively labeled organelles that resembledperoxisomes (Figure 4D).
In the thalamic reticular nucleus (Figures 4E, 4F, 5A,and 5B), the majority of the neurons showed intenseimmunoreactivity. Glial cells, as in other brain re-gions, were devoid of staining (Figures 5A and 5B). Anumber of axons and dendrites, probably belonging tothe positive neurons, were immunoreactive. Other neu-rites appeared immunonegative, irrespective of thepresence or absence of a myelin sheath.
In the cerebellar cortex, stellate and basket cells ofthe molecular layer were highly immunoreactive (Fig-ure 5C). Parallel fibers originating from granule cellswere negative, whereas other axons and dendrites werespecifically stained (not shown). Purkinje cells showedweak positivity for CuZnSOD. In fact, only a few ofthese neurons were recognized as immunoreactive, andeven in these cases the immunoreaction product wasnon-homogeneously distributed, being more evident inthe nucleoplasm. However, clearly specific and intenseimmunostaining was rather frequently observed in sin-gle membrane-limited bodies measuring 0.2–0.4
m
min diameter, whereas other organelles were negative(Figure 5D). Golgi cells of the granular layer showedcytoplasmic and nuclear staining (Figure 5E) and alsoimmunoreactive organelles (Figure 5E, inset). Granulecells showed very weak but specific CuZnSOD immu-noreactivity in their nuclei, apparently only in corre-spondence with euchromatin (Figure 5E).
Postembedding Double Immunogold Labeling
The comparison between LR White- and Unicryl-embed-ded sections revealed that the neural tissue morphol-ogy was not satisfactorily preserved by the formerresin (not shown), whereas the latter yielded better ul-trastructural preservation. Unicryl was therefore cho-sen for postembedding experiments.
For double immunogold labeling, the cerebellar cor-tex was selected because of the presence of some cell
Figure 2
Coronal sections of rat brain processed for CuZnSOD immunocytochemistry. (
A
) Main olfactory bulb. Gl, glomerular layer; EPl, ex-ternal plexiform layer; Mi, mitral cell layer; IGr, internal granular layer. Arrows indicate positive neurons in Gl and EPl. Mitral cells showhigher immunoreactivity in their cytoplasm than the nucleus (arrowhead). (
B
) Cerebral cortex, frontal region. Many positive neurons arescattered in all the layers. Arrows indicate weakly immunoreactive pyramidal cells. (
C
) Nucleus caudate putamen (CPu). ec, external capsule.(
D
) An island of Calleja (ICj). (
E
) Septal complex. HDB and VDB, horizontal and vertical nucleus of the diagonal band of Broca. (
F–H
) Hippo-campal formation. Densely stained interneurons are seen in the entire area. (
F
) CA1 field. (
G
) Dentate gyrus. (
H
) Subiculum. Bars
5
200
m
m.
by guest on July 10, 2014jhc.sagepub.comDownloaded from

1616
Moreno, Nardacci, Cerù
by guest on July 10, 2014jhc.sagepub.comDownloaded from

Copper–Zinc Superoxide Dismutase in Rat Brain
1617
types showing peroxisome-like positivity for CuZnSODand known to contain catalase (Moreno et al. 1995),i.e., Golgi and Purkinje cells. In these cells, organellesmeasuring 0.15–0.35
m
m in diameter showed intenseimmunogold labeling for CuZnSOD, revealed by 10-nmgold particles (Figure 5F). Anti-catalase antibody (re-vealed by 5-nm gold) co-localized with anti-CuZnSODin the same organelles, thus confirming their peroxiso-mal nature (Figure 5F). Examination of 15 double la-beled organelles belonging to either Golgi or Purkinjecells gave a mean content of 5.2 gold particles of 5 nm(SD 2.17) and 26 gold particles of 10 nm (SD 4.97).
Discussion
In the present investigation we have obtained a com-plete map of the distribution of CuZnSOD in the ratCNS by means of an extremely sensitive immunocy-tochemical procedure based on the avidin–biotin sys-tem (Adams 1992). This study also describes for thefirst time the ultrastructural localization of CuZnSODin neural cells and demonstrates that the enzyme isalso present within the peroxisomes.
Regional Distribution of CuZnSOD
Our light and electron microscopic observations dem-onstrate that CuZnSOD is an exclusively neuronal en-zyme, the immunostaining being absent from any non-neuronal element, including glial and endothelial cells.This result is in agreement with other reports dealingeither with expression of CuZnSOD mRNA in the hu-man hippocampus and mesencephalon (Ceballos et al.1989; Zhang et al. 1993) or with immunolocalizationof the protein in the gerbil hippocampus (Matsuyamaet al. 1993). Furthermore, both CuZnSOD mRNA andprotein were detected only in neuronal elements in thebrains of transgenic mice carrying the human CuZn-SOD gene (Ceballos–Picot et al. 1991). In this respect,it is worth mentioning that immunologically activeCuZnSOD was detected in reactive glial cells in gerbilhippocampus after ischemic insult (Kato et al. 1995).
Although specific immunoreactivity is detectable inmost, if not all, neurons, different degrees of staining
are clearly distinguished in the various brain regions.High staining levels are found in all subdivisions ofthe CNS and particularly in the cerebral cortex, thehippocampus, the thalamus, the superior colliculus,the substantia nigra, the cerebellar cortex, and the spi-nal cord. However, within these areas, not all the neu-rons show similar immunoreactivity.
On the basis of biochemical data, a non-homoge-neous distribution of CuZnSOD in various areas ofrat brain has been suggested previously (Ciriolo et al.1991; Carrillo et al. 1992). Our data are in overallagreement with these reports because appreciable im-munoreactivity was detected in the caudate nucleus,the cerebral cortex, hippocampus, pons, and medullaoblongata. Some discrepancies between the biochemi-cal and morphological results can be easily explained,keeping in mind that the former reflect the mean val-ues of CuZnSOD biochemical activity of different celltypes within the same area, which may include highlypositive cells together with negative cells.
The morphological studies available on the distri-bution of CuZnSOD in neural tissue also point to adifferential expression of this enzyme. Immunohis-tochemical studies dealing with CuZnSOD localiza-tion recently carried out in specific CNS regions (Liuet al. 1994; Pardo et al. 1995) are in agreement withour results, because many positive neurons were ob-served in the neocortex, striatum, and spinal cord.Moreover, intense CuZnSOD immunoreactivity hasbeen described in pyramidal cells and interneurons ofthe hippocampal regions CA1 and CA3, whereas den-tate granule cells showed only moderate staining (Liuet al. 1993; Kato et al. 1995).
Our light and electron microscopic observationsare in partial agreement with these studies, because wedemonstrate that in the hippocampal formation inter-neurons are strongly immunostained for CuZnSOD,whereas pyramidal cells and granule cells show weakerimmunoreactivity.
In human mesencephalon, in situ hybridization re-vealed preferential expression of the CuZnSOD mRNAin dopaminergic neurons, particularly in the melanizedones, thus suggesting a role for the enzyme in the re-
Figure 3
CuZnSOD immunocytochemistry in rat brain. (
A
) Sagittal section of the thalamus. RT, reticular thalamic nucleus; VT, ventral tha-lamic nuclei; ic, internal capsule. (
B
) Coronal section of the superior colliculus. Intensely immunoreactive neurons are present in all the lay-ers. (
C
) Coronal section of inferior colliculus, external cortex. Moderate and rather uniform positivity is visible in most neurons. (
D
) Coronalsection of the substantia nigra. SNC, pars compacta; SNR, pars reticulata. Note that, at least in SNR, neurons show immunoreactivity only intheir cytoplasm (arrowhead). (
E
) Coronal section of red nucleus. Large neurons of the magnocellular portion are distinctly immunoreactivein their cytoplasm. (
F
) Coronal section of interpeduncular nucleus. (
G
) Coronal section of abducens nucleus (6) and supragenual nucleus(SGe). (
H
) Coronal section of reticular formation. Neurons of gigantocellular reticular nucleus display intense cytoplasmic immunopositivity.(
I
) Coronal section of principal trigeminal sensory nucleus. (
J
) Sagittal section of the brainstem, facial nucleus. (
K
) Coronal section of lateral(Lat) and interpositus (Int) cerebellar nuclei. (
L
) Coronal section of cerebellar cortex. ML, molecular layer; PL, Purkinje cell layer; GL, granularlayer. Stellate and basket neurons (arrowheads) of the ML and Golgi cells (arrows) in the GL are the most intensely immunoreactive ele-ments. The positivity observed in the PL appears to be external to Purkinje cells and presumably is due to basket cell terminals. (
Inset
)Higher magnification of a Golgi cell showing remarkable immunostaining in its cytoplasm, nucleus, and cell process (arrow). Note absenceof immunoreaction product from the nucleolus (arrowhead). Bars
5
200
m
m.
by guest on July 10, 2014jhc.sagepub.comDownloaded from
1618
Moreno, Nardacci, Cerù
by guest on July 10, 2014jhc.sagepub.comDownloaded from

Copper–Zinc Superoxide Dismutase in Rat Brain
1619
moval of superoxide anion generated by catechola-mine metabolism (Zhang et al. 1993). Consistently, inour specimens many aminergic areas are identified asimmunoreactive (substantia nigra, central gray sub-stance, locus ceruleus). However many other stronglypositive neurons use different neurotransmitters. Infact, interneurons of the piriform cortex, neurons ofthe reticular thalamic nucleus, principal neurons in thecorpus striatum, neurons of the dorsal nucleus of lat-eral lemniscus, and Golgi, basket, and stellate cells ofthe cerebellar cortex are GABAergic. Furthermore, thecholinergic motor neurons of the cranial and spinalnuclei are also positive.
The factors involved in the differential expressionof CuZnSOD in the various neuronal populationstherefore remain obscure, and cannot solely dependon the specific neurotransmitter used by the neuron, atleast as far as it concerns the classic mediators.
Functional Considerations
The fact that CuZnSOD is expressed exclusively inneurons may suggest that these cells require a specificprotective mechanism against the superoxide anion,although a role of CuZnSOD through its metaboliteH
2
O
2
or through a still unknown function, possiblylinked to the presence of copper and zinc in its mole-cule, cannot be ruled out.
Therefore, the issue concerning the functional im-plications of CuZnSOD abundance in certain neu-ronal subtypes is still to be addressed. Although insome instances the presence of CuZnSOD appears tofavor a greater protection against ROS-mediated neu-ronal damage (Chan 1994; Medina et al. 1996), inother cases high levels of CuZnSOD may even be re-sponsible for neuronal injury, as probably occurs inDown’s syndrome and in some strains of transgenicmice that overexpress CuZnSOD (Brooksbank andBalazs 1984; Amstad et al. 1994).
Correspondingly, some of the strongly positive neu-rons described in our specimens can be identified asoxidative stress resistant neurons (i.e., GABAergic inter-neurons of hippocampus and cerebral cortex) (Schlanderet al. 1988), whereas others clearly belong to subtypestypically vulnerable to a variety of injuries most prob-ably involving ROS (i.e., motor neurons of spinal cord,affected by ALS; neurons of the substantia nigra, af-
fected by Parkinson’s disease; and also principal neu-rons of the striatum, affected by Huntington’s disease).
This apparent contradiction is probably due to thedual aspect of the function of SOD, one as a free radi-cal scavenger (as the principal O2–• metabolizing en-zyme) and the other as a ROS generator (as an H
2
O
2
-producing enzyme).
Concerning the H
2
O
2
produced by cytosolic CuZn-SOD, one could envision that this molecule, leakedout of the neurons, is captured and eliminated by as-trocytes, which have been reported to contain glu-tathione peroxidase (Damier et al. 1993) and to pro-tect neurons against H
2
O
2
toxicity (Raps et al. 1989;Desagher et al. 1996).
CuZnSOD as a Peroxisomal Enzyme
The comparison between the distribution of CuZnSODin the rat CNS and that of catalase (McKenna et al.1976; Moreno et al. 1995) allows further consider-ations. In many instances we have found an overlap ofCuZnSOD and catalase immunostaining, e.g., in Golgicells of the cerebellar cortex, interneurons of the hip-pocampus, reticular thalamic nucleus, substantia nigrapars reticulata, interpeduncular nucleus, motor neu-rons of cranial and spinal nuclei, and neurons of sep-tal nuclei (Moreno et al. 1995).
In general, catalase is predominantly localized toperoxisomes, whereas CuZnSOD is mainly a cytosolicenzyme. However, ultrastructural examination of manydifferent areas of the brain gives strong indication of aparticulate CuZnSOD as well. We first interpretedthese vescicles as peroxisomes on the basis of their sizeand morphology; even the dishomogeneous immuno-staining observed within these organelles is in keep-ing with their peroxisomal nature, because a subcom-partmentalization of peroxisomal enzymes has beenlargely demonstrated (for review see Van Den Munck-hof 1996). Postembedding double immunogold label-ing confirmed that the CuZnSOD-positive particlesare true peroxisomes. The low catalase/SOD rate, asevaluated by morphometric analysis in double immu-nolabeled specimens, is not in contrast to this conclu-sion. Even the absence of catalase from SOD-positiveorganelles would not contradict their peroxisomal na-ture, because catalase-negative peroxisomes have beenrecently recognized (Van Roermund et al. 1995; Wil-
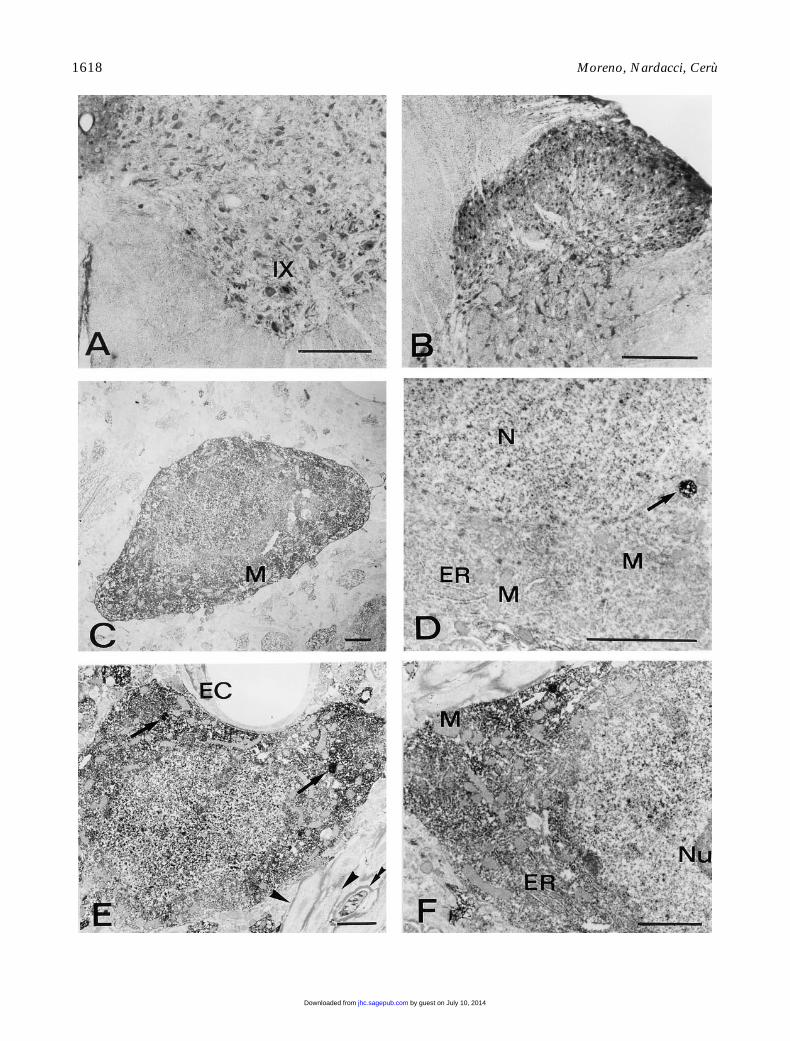
Figure 4
CuZnSOD immunocytochemistry in rat brain. (
A,B
) Coronal sections of the rat cervical cord. Bars
5
200
m
m. (
A
) Ventral horn. Mo-tor neurons of Rexed’s lamina IX, as well as many neurons belonging to the other laminae, are heavily immunostained. (
B
) Dorsal horn.Small neurons are intensely positive. (
C–F
), CuZnSOD pre-embedding immunoelectron microscopy. Bars
5
2
m
m. (
C
) Neocortex. Intenselystained nonpyramidal neuron showing similar immunoreactivity in cytoplasm and nucleus. Mitochondria (M) are unstained. (
D
) Hippocam-pus, CA3 field. Portion of a pyramidal cell containing a strongly labeled organelle, showing a dishomogeneous staining (arrow). N, nucleus;ER, endoplasmic reticulum; M, mitochondria. (
E,F
) Reticular thalamic nucleus. (
E
) Positive neuron displaying high concentration of immu-noreaction product in cytosol and nucleus. Note the presence of immunoreactive organelles (arrows) and the negative endothelial cell (EC).Positive (double arrowhead) and negative (arrowheads) myelinated axons are present in the surrounding areas. (
F
) Higher magnification ofan immunoreactive neuron, showing a densely stained organelle (arrow). ER, endoplasmic reticulum; M, mitochondria; Nu, nucleolus.
by guest on July 10, 2014jhc.sagepub.comDownloaded from

1620
Moreno, Nardacci, Cerù
by guest on July 10, 2014jhc.sagepub.comDownloaded from

Copper–Zinc Superoxide Dismutase in Rat Brain
1621
cke et al. 1995), thus confirming the high degree ofheterogeneity of these particles (Cimini et al. 1993)and the involvement of other systems in removal ofH
2
O
2
(Singh et al. 1994).The positivity for CuZnSOD in the peroxisomes
does not always correlate with a similar degree of im-munoreactivity in the cytoplasm or nucleus. In fact,neurons apparently negative or weakly stained at thelight microscopic level, such as pyramidal cells of theneocortex and hippocampus and Purkinje neurons ofthe cerebellar cortex, are often found to contain in-tensely labeled vacuoles.
Therefore, the compartimentalization of CuZnSODwithin peroxisomes strongly suggests a specific func-tion of this enzyme in the removal of intraorganellarby generated O2–•, independent of the situation in thecytosolic and nuclear compartments.
Acknowledgments
Supported by MURST 40% grant, CNR grant no.95.02196.CT04, and in part by the European Concerted Ac-tion “Peroxisomal Leukodystrophy” BMH4 CT 96-1621.
We wish to thank Dr E. Mugnaini for critical reading ofthe manuscript and for helpful discussion, Dr S. Stefanini forimmunogold analysis, and Dr A. Cimini for immunoblot-ting. We gratefully acknowledge Dr L. Cristiano, Dr B.Pruiti, and Dr P. Aimola for skillful technical assistance.
Literature Cited
Adams JC (1992) Biotin amplification of biotin and horseradishperoxidase signals in histochemical stains. J Histochem Cy-tochem 40:1457–1463
Amstad P, Moret R, Cerutti P (1994) Glutathione peroxidase com-pensates for the hypersensitivity of Cu,Zn-superoxide dismutaseoverproducers to oxidant stress. J Biol Chem 269:1606–1609
Brooksbank BW, Balazs R (1984) Superoxide dismutase, glu-tathione peroxidase and lipoperoxidation in Down’s syndromefetal brain. Dev Brain Res 16:37–44
Carrillo MC, Kanai S, Sato Y, Kitani K (1992) Age-related changesin antioxidant enzyme activities are region and organ, as well assex, selective in the rat. Mech Ageing Dev 65:187–198
Ceballos I, Javoy-Agid F, Hirsch EC, Dumas S, Kamoun PP, SinetPM, Agid Y (1989) Localization of copper-zinc superoxide dis-mutase in human hippocampus by in situ hybridization. Neuro-sci Lett 105:41–46
Ceballos–Picot I, Nicole A, Briand P, Grimber G, Delacourte A, De-fossez A, Javoy–Agid F, Lafon M, Blouin JL, Sinet PM (1991)
Neuronal-specific expression of human copper-zinc superoxidedismutase gene in transgenic mice: animal model of gene dosageeffects in Down’s syndrome. Brain Res 552:198–214
Chan PH (1994) Oxygen radicals in focal cerebral ischemia. BrainPathol 4:59–65
Cimini AM, Moreno S, Giorgi M, Serafini B, Cerù MP (1993) Puri-fication of peroxisomal fraction from rat brain. Neurochem Int23:249–260
Ciriolo MR, Fiskin K, De Martino A, Corasaniti MT, Nisticò G,Rotilio G (1991) Age-related changes in Cu,Zn superoxide dis-mutase, Se-dependent and -independent glutathione peroxidaseand calatase activities in specific areas of rat brain. Mech AgeingDev 61:287–297
Crapo JD, Oury T, Rabouille C, Slot JW, Chang L-Y (1992) Cop-per, zinc superoxide dismutase is primarily a cytosolic protein inhuman cells. Proc Natl Acad Sci USA 89:10405–10409
Damier P, Hirsch EC, Zhang P, Agid Y, Javoy–Agid F (1993) Glu-tathione peroxidase, glial cells and Parkinson’s disease. Neuro-science 52:1–6
Deng H-X, Hentati A, Tainer JA, Iqbal Z, Cayabyab A, Hung W-Y,Getzoff ED, Hu P, Herzfeldt B, Roos RP, Warner C, Deng G, So-riano E, Smyth C, Parge HE, Ahmed A, Roses AD, Hallewell RA,Pericak–Vance MA, Siddique T (1993) Amyotrophic lateral scle-rosis and structural defects in Cu,Zn superoxide dismutase. Sci-ence 261:1047–1051
Desagher S, Glowinski J, Premont J (1996) Astrocytes protect neu-rons from hydrogen peroxide toxicity. J Neurosci 16:2553–2562
Dhaunsi GS, Gulati S, Singh AK, Orak JK, Asayama K, Singh I(1992) Demonstration of Cu-Zn superoxide dismutase in ratliver peroxisomes. Biochemical and immunochemical evidence. JBiol Chem 267:6870–6873
Evans PH (1993) Free radicals in brain metabolism and pathology.Br Med Bull 49:577–587
Fridovich I (1989) Superoxide dismutases. An adaptation to a para-magnetic gas. J Biol Chem 264:7761–7764
Greenlund LJS, Deckwerth TL, Johnson EM (1995) Superoxide dis-mutase delays neuronal apoptosis: a role for reactive oxygen spe-cies in programmed cell death. Neuron 14:303–316
Gurney ME, Pu H, Chiu AY, Dal Canto MC, Polchow CY, Alex-ander DD, Caliendo J, Hentati A, Kwon YW, Deng H-X, ChenW, Zhai P, Sufit RL, Siddique T (1994) Motor neuron degenera-tion in mice that express a human Cu,Zn superoxide dismutasemutation. Science 264:1772–1775
Halliwell B (1992) Reactive oxygen species and the central nervoussystem. J Neurochem 59:1609–1623
He YY, Hsu CY, Ezrin AM, Miller MS (1993) Polyethylenglycol-conjugated superoxide dismutase in focal cerebral ischemia-rep-erfusion. Am J Physiol 265:H252–H256
Imaizumi S, Woolworth V, Fishman RA, Chan PH (1990) Lipo-some-entrapped superoxide dismutase reduces cerebral infarc-tion in cerebral ischemia in rats. Stroke 21:1312–1317
Kato H, Kogure K, Araki T, Liu X-H, Kato K, Itoyama Y (1995)Immunohistochemical localization of superoxide dismutase inthe hippocampus following ischemia in a gerbil model of is-chemic tolerance. J Cereb Blood Flow Metab 15:60–70
Keller GA, Warner TG, Steimer KS, Hallewell RA (1991) Cu,Zn su-
Figure 5
CuZn immunoelectron microscopy. (
A,B
) Reticular thalamic nucleus. Immunonegative non-neuronal cells. Note the presence ofnumerous positive (double arrowheads) and negative (arrowheads) myelinated axons and in
B
a strongly positive neuron with its process(arrow). Bars
5
2
mm (C–F) Cerebellar cortex. (C–E) CuZnSOD pre-embedding immunoelectron microscopy. Bars 5 0.5 mm. (F) Postembed-ding double immunogold labeling with anti-catalase and anti-CuZnSOD. (C) Stellate neuron in the molecular layer. ER, endoplasmic reticu-lum; G, Golgi apparatus; M, mitochondria; Nu, nucleolus. Arrow points to a heavily immunoreactive peroxisome. Several unstained unmye-linated processes are visible. (D) Portion of a Purkinje neuron shows weak positivity in the cytoplasm and nucleus (N), with two stronglystained peroxisomal bodies surrounded by single membrane (arrows). Note the dishomogeneous distribution of the immunoprecipitatewithin the organelles. (E) Micrograph of the granular layer showing two labeled Golgi cells and one granule cell. The latter is weakly immu-noreactive, the staining being confined to the euchromatic portion of their nucleus (N). Nu, nucleolus. (Inset) Higher magnification showinga strongly positive organelle inside a Golgi cell. (F) Portion of a Purkinje cell. Peroxisome showing co-localization of catalase (5-nm gold)and CuZnSOD (10-nm gold). Other cytoplasmic organelles, such as mitochondria (M) and endoplasmic reticulum (ER), are negative for bothantibodies.
by guest on July 10, 2014jhc.sagepub.comDownloaded from

1622 Moreno, Nardacci, Cerù
peroxide dismutase is a peroxisomal enzyme in human fibro-blasts and hepatoma cells. Proc Natl Acad Sci USA 88:7381–7385
Kincaid RL (1988) Preparation, characterization and properties ofaffinity-purified antibodies to calmodulin-dependent cyclic nu-cleotide phosphodiesterase and the protein phosphatase cal-cineurin. In Corbin JD, Johnson RA, eds. Methods in Enzymol-ogy. Vol 159. New York, Academic Press, 627–652
Kobayashi T, Saito N, Takemori N, Iizuka S, Suzuki K, TaniguchiN, Iizuka H (1993) Ultrastructural localization of superoxidedismutase in human skin. Acta Dermatol Venereol (Stockh)73:41–45
Laemmli EK (1970) Cleavage of structural proteins during the as-sembly of the head of bacteriophage T4. Nature 227:680–685
LeBel CP, Bondy SC (1992) Oxidative damage and cerebral aging.Prog Neurobiol 38:601–609
Liu X-H, Kato H, Araki T, Itoyama Y, Kato K, Kogure K (1994) Animmunohistochemical study of copper/zinc superoxide dismutaseand manganese superoxide dismutase following focal cerebral is-chemia in the rat. Brain Res 644:257–266
Liu X-H, Kato H, Nakata N, Kogure K, Kato K (1993) An immu-nohistochemical study of copper/zinc superoxide dismutase andmanganese superoxide dismutase in rat hippocampus after tran-sient cerebral ischemia. Brain Res 625:29–37
Marttila RJ, Viljanen M, Toivonen E, Lorentz H, Rinne UK (1990)Superoxide dismutase-like activity in the Parkinson’s diseasebrain. In Streifler MB, Korczyn AD, Melamed E, Youdim MBH,eds. Advances in Neurology. Vol 53. Parkinson’s Disease: Anat-omy, Pathology, and Therapy. New York, Raven Press, 141–144
Matsumiya N, Koehler RC, Kirsch JR, Traystman RJ (1991) Conju-gated superoxide dismutase reduces extent of caudate injury aftertransient focal ischemia in cats. Stroke 22:1193–1200
Matsuyama T, Michishita H, Nakamura H, Tsuchiyama M,Shimizu S, Watanabe K, Sugita M (1993) Induction of copper-zinc superoxide dismutase in gerbil hippocampus after ischemia.J Cereb Blood Flow Metab 13:135–144
McKenna O, Arnold G, Holtzman E (1976) Microperoxisome dis-tribution in the central nervous system of the rat. Brain Res117:181–194
Medina L, Figueredo–Cardenas G, Reiner A (1996) Differentialabundance of superoxide dismutase in interneurons versus pro-jection neurons and in the matrix versus striosome neurons inmonkey striatum. Brain Res 708:59–70
Moreno S, Cimini A, Giorgi M, Cerù MP (1994) Immunocytochem-ical localization of Cu,Zn-superoxide dismutase in rat brain. CellBiol Int 18:520
Moreno S, Mugnaini E, Cerù MP (1995) Immunocytochemical lo-calization of catalase in the central nervous system of the rat. JHistochem Cytochem 43:1253–1267
Mugnaini E, Oertel WH (1985) An atlas of the distributionGABAergic neurons and terminals in the rat CNS as revealed byGAD immunohistochemistry. In Bjorklund A, Hökfelt T, eds.Handbook of Chemical Neuroanatomy. Vol 4. Part I. New York,Elsevier 436–608
Nishiyama K, Murayama S, Shimizu J, Ohya Y, Kwak S, AsayamaK, Kanazawa I (1995) Cu/Zn superoxide dismutase-like immu-noreactivity is present in Lewy bodies from Parkinson disease: alight and electron microscopic immunocytochemical study. ActaNeuropathol (Berl) 89:471–474
Pardo CA, Xu Z, Borchelt DR, Price DL, Sisodia SS, Cleveland DW(1995) Superoxide dismutase is an abundant component in cellbodies, dendrites, and axons of motor neurons and in a subset ofother neurons. Proc Natl Acad Sci USA 92:954–958
Paxinos G, Watson C (1986) The Rat Brain in Stereotaxic Coordi-nates. 2nd ed. San Diego, CA, Academic Press
Raps SP, Lai JCK, Hertz L, Cooper JL (1989) Glutathione is presentin high concentrations in cultured astrocytes but not in culturedneurons. Brain Res 493:398–401
Rosen DR, Siddique T, Patterson D, Figlewicz DA, Sapp P, Hentati
A, Donaldson D, Goto J, O’Regan JP, Deng H-X, Rahmani Z,Krizus A, McKenna–Yasek D, Cayabyab A, Gaston SM, BergerR, Tanzi RE, Halperin JJ, Herzfeldt B, Van den Bergh R, HungW-Y, Bird T, Deng G, Mulder DW, Smyth C, Laing NG, SorianoE, Pericak–Vance MA, Haines J, Rouleau GA, Gusella JS, Hor-vitz HR, Brown RH (1993) Mutations in Cu/Zn superoxide dis-mutase gene are associated with familial amyotrophic lateralsclerosis. Nature 362:59–62
Rossi L, Ciriolo MR, Marchese E, De Martino A, Giorgi M, RotilioG (1994) Differential decrease of copper content and of copperbinding to superoxide dismutase in liver, heart and brain of cop-per-deficient rats. Biochem Biophys Res Commun 203:1028–1034
Rowland LP (1995) Amyotrophic lateral sclerosis: human challengefor neuroscience. Proc Natl Acad Sci USA 92:1251–1253
Saggu H, Cooksey J, Dexter D, Wells FR, Lees A, Jenner P, Mars-den CD (1989) A selective increase in particulate superoxide dis-mutase activity in parkinsonian substantia nigra. J Neurochem53:692–697
Scala C, Cenacchi G, Ferrari C, Pasquinelli G, Preda P, Manara GC(1992) A new acrylic resin formulation: a useful tool for histo-logical, ultrastructural and immunocytochemical investigations. JHistochem Cytochem 40:1799–1804
Schlander M, Hoyer S, Frotscher M (1988) Glutamate decarboxy-lase-immunoreactive neurons in the aging rat hippocampus aremore resistant to ischemia than CA1 pyramidal cells. NeurosciLett 91:241–246
Singh AK, Dhaunsi GS, Gupta MP, Asayama K, Singh I (1994)Demonstration of glutathione peroxidase in rat liver peroxi-somes and its intraorganellar distribution. Arch Biochem Bio-phys 315:331–338
Slot JW, Geuze HJ, Freeman BA, Crapo JD (1986) Intracellular lo-calization of the copper-zinc and manganese superoxide dismu-tases in rat liver parenchymal cells. Lab Invest 55:363–371
Tagaya M, Matsumoto M, Kitagawa K, Niinobe M, Ohtsuki T,Hata R, Ogawa S, Handa N, Mikoshiba K, Kamada T (1992)Recombinant human superoxide dismutase can attenuate is-chemic neuronal damage in gerbils. Life Sci 51:253–259
Thaete LG, Crouch RK, Nakagawa F, Spicer SS (1986) The immu-nocytochemical demonstration of copper-zinc superoxide dismu-tase in the brain. J Neurocytol 15:337–343
Towbin H, Staehelin T, Gordon J (1979) Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: proce-dure and some applications. Proc Natl Acad Sci USA 76:4350–4354
Uyama O, Matsuyama T, Michishita H, Nakamura H, Sugita M(1992) Protective effects of human recombinant superoxide dis-mutase on transient ischemic injury of CA1 neurons in gerbils.Stroke 23:75–81
Van Den Munckhof RJM (1996) In situ heterogeneity of peroxiso-mal oxidase activities: an update. Histochem J 28:401–429
Van Roermund CWT, Van Den Berg M, Wanders RJA (1995) Lo-calization of peroxisomal 3–oxoacyl–CoA thiolase in particles ofvaried density in rat liver: implications for peroxisome biogene-sis. Biochim Biophys Acta 1245:348–358
Wanders RJA, Denis S (1992) Identification of superoxide dismu-tase in rat liver peroxisomes. Biochim Biophys Acta 1115:259–262
Wilcke M, Hultenby K, Alexson SEH (1995) Novel peroxisomalpopulations in subcellular fractions from rat liver. J Biol Chem270:6949–6958
Yano S (1990) Multiple isoelectric variants of copper, zinc-superox-ide dismutase from rat liver. Arch Biochem Biophys 279:60–69
Yoshikawa T (1993) Free radicals and their scavengers in Parkin-son’s disease. Eur Neurol 33:60–68
Zhang P, Damier P, Hirsch EC, Agid Y, Ceballos–Picot I, Sinet PM,Nicole A, Laurent M, Javoy–Agid F (1993) Preferential expres-sion of superoxide dismutase messenger RNA in melanized neu-rons in human mesencephalon. Neuroscience 55:167–175
by guest on July 10, 2014jhc.sagepub.comDownloaded from
Copyright © 2022 FDOKUMEN




















