
Structure of a superoxide dismutase and implications for copper-ion chelation
Upload
independentCategory
view
2download
0
Rat Mitochondrial Manganese Superoxide Dismutase: Amino Acid PositionsInvolved in Covalent Modifications, Activity, and Heat Stability
Immacolata Castellano,1 Francesca Cecere,1 Alberto De Vendittis,1 Roberta Cotugno,1
Angela Chambery,2 Antimo Di Maro,2 Andzelika Michniewicz,3 Giuseppe Parlato,3
Mariorosario Masullo,1,4 Enrico Vittorio Avvedimento,5 Emmanuele De Vendittis,1
Maria Rosaria Ruocco1
1 Dipartimento di Biochimica e Biotecnologie Mediche, Universita di Napoli Federico II, Via S. Pansini 5, 80131 Napoli, Italy
2 Dipartimento di Scienze della Vita, II Universita di Napoli, Via Vivaldi 43, 81100, Caserta, Italy
3 Dipartimento di Medicina Sperimentale e Clinica ‘‘G. Salvatore’’, Universita di Catanzaro ‘‘Magna Graecia’’,
Via T. Campanella 115, 88100 Catanzaro, Italy
4 Dipartimento di Scienze Farmacobiologiche, Universita di Catanzaro ‘‘Magna Graecia’’, 88021 Roccelletta di Borgia (CZ), Italy
5 Dipartimento di Biologia e Patologia Molecolare e Cellulare, Universita di Napoli Federico II, Via S. Pansini 5, 80131 Napoli, Italy
Received 15 December 2008; revised 10 March 2009; accepted 3 April 2009
Published online 21 April 2009 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/bip.21208
This article was originally published online as an accepted
preprint. The ‘‘Published Online’’ date corresponds to the preprint
version. You can request a copy of the preprint by emailing the
Biopolymers editorial office at [email protected]
Rat Mitochondrial Manganese Superoxide Dismutase: Amino Acid PositionsInvolved in Covalent Modifications, Activity, and Heat Stability
This article is in honour of Professor Lelio Mazzarella, on theoccasion of his 70th birthday.
Correspondence to: E. De Vendittis; e-mail: [email protected] or M. R.
Ruocco; e-mail: [email protected]
ABSTRACT:
The role of three amino acid residues (Q143, Y34, S82) of
rat mitochondrial superoxide dismutase (ratSOD2) in the
enzymatic activity, thermostability, and post-translational
modification of the enzyme was investigated through site-
directed mutagenesis studies. Six recombinant forms of the
enzyme were produced, carrying the Q143 or H143 residue
with or without the Y34F or S82A replacement. All proteins
bound manganese as active cofactor and were organized as
homotetramers. The greatest effect on the activity (sixfold
reduction) was observed in ratSOD2 forms containing the
H143 variant, whereas Y34F and S82A substitutions
moderately reduced the enzymatic activity compared to the
Q143 form. Heat inactivation studies showed the high
thermo-tolerance of ratSOD2 and allowed an evaluation of
the related activation parameters of the heat inactivation
process. Compared to Q143, the H143 variant was
significantly less heat stable and displayed moderately lower
enthalpic and entropic factors; the Y34F substitution caused
a moderate reduction of heat stability, whereas the S82A
replacement slightly improved the thermo-tolerance of the
Q143 variant; both substitutions significantly increased
enthalpic and entropic factors of heat inactivation, the
greatest effect being observed with S82A substitution. All
recombinant forms of ratSOD2 were glutathionylated in
Escherichia coli, a feature pointing to the high reactivity of
ratSOD2 toward glutathione. Moreover, the S82 position of
the enzyme was phosphorylated in an in vitro system
containing human mitochondrial protein extracts as source
of protein kinases. These data highlight the role played by
some residues in ratSOD2 and suggest a fine regulation of
the enzyme occurring in vivo. # 2009 Wiley Periodicals, Inc.
Biopolymers 91: 1215–1226, 2009.
Keywords: superoxide dismutase; rat SOD2; mutagenic
analysis; post-translational modifications; thermostability
Contract grant sponsor: MIUR.
Contract grant number: PRIN 2007.�C 2009 Wiley Periodicals, Inc.
Biopolymers Volume 91 / Number 12 1215

INTRODUCTION
The cell has developed enzymatic and nonenzymatic
systems aimed at the defence against reactive oxygen
species (ROS), mainly produced during the oxygen
metabolism. Indeed, ROS are highly reactive and
toxic compounds, as they can modify many cellular
targets, such as lipids, proteins, and DNA.1 On the other
hand, ROS have also a physiological role, being messengers
in many cellular processes, such as proliferation, differentia-
tion, apoptosis, and senescence.2,3 An early cellular defence is
provided by superoxide dismutase (SOD), a key antioxidant
enzyme that eliminates the toxic superoxide anion, the first
ROS produced through the univalent reduction of oxygen.4,5
Indeed, SOD is a metal enzyme that dismutates two super-
oxide anions into hydrogen peroxide and molecular oxygen
through a catalytic mechanism conserved in taxonomically
distant organisms. However, this ubiquitous enzyme is classi-
fied in various structurally unrelated families on the basis of
the type of metal present in its active site.6,7 The two major
groups include the Cu/Zn-SOD (SOD1), present in the cyto-
sol of eukaryotic cells, and the Fe- and Mn-SOD (SOD2),
found in bacteria, archaea and also in mitochondria and
chloroplasts of eukaryotic cells. Therefore, SOD2 is widely dis-
tributed in all living kingdoms and probably displays the most
crucial defence mechanism against the oxidative stress.
Because of their intense respiratory activity, mitochondria are
the major cellular site for the production of superoxide
anions; therefore, the high efficiency of the mitochondrial
Mn-SOD is indispensable for cell viability, as confirmed by
the involvement of this enzyme in a number of diseases caused
by mitochondrial defects.8–12 The importance of SOD2 in
mammals is also highlighted by the observation that, in
contrast to SOD1, knock-out of its gene is lethal to mice.13,14
Mammalian mitochondrial Mn-SOD is encoded by the
nuclear genome as a precursor headed by a signal peptide
spanning 24 amino acid residues; this leader peptide is
removed for the entry of the enzyme in mitochondria.15 The
crystal structure of the human mitochondrial enzyme points
to its organization as a compact homotetramer, with an
interface made of two 4-helix bundles.16 Because of its com-
pactness, the human enzyme is endowed with a remarkable
thermal resistance.17 Another property shown by the human
Mn-SOD concerns its high reactivity toward peroxynitrite.18–20
Indeed, this highly toxic ROS causes the irreversible inactiva-
tion of the enzyme through the covalent modification of
Y34, a conserved residue in the channel that drives the super-
oxide anion to the active site. Other members of the Fe- and
Mn-SOD family display a similar high reactivity toward non-
physiological or physiological compounds, either on this
conserved tyrosine,21–25 or on cysteine residues.26–28
The rat mitochondrial Mn-SOD (ratSOD2), sharing 93%
amino acid identity with the human counterpart, is an
appropriate model to study the molecular and functional
properties of mammalian mitochondrial SOD2.29–31 This
investigation regards the role of some key amino acid posi-
tions of ratSOD2 regulating catalysis, reactivity, and thermal
resistance of the enzyme. The study was carried out through
the heterologous production of the mature form of ratSOD2
and mutants obtained by site-directed mutagenesis on the
corresponding gene. Among the three amino acid positions
considered in the mutagenic analysis, Q143 belongs to the
second coordination sphere of the metal ion. This residue
forms a hydrogen bond with a Mn-bound water molecule
and the hydroxyl side chain of Y34. In SODs belonging to the
same family, the position corresponding to Q143 is occupied
by glutamine or histidine.23,32 Furthermore, this position has
been hypothesized as a possible polymorphic site in rat-
SOD2, because of the occurrence of rat clones predicting the
H143 variant.29–31 The second amino acid considered in the
study is Y34, a conserved residue of the second coordination
sphere of the metal.16 This residue plays an important role in
the catalytic mechanism of superoxide dismutation and acts
as a powerful regulator of the activity of SOD2.17,21,32 Its
high reactivity toward peroxynitrite and/or phenylmethane-
sulfonyl fluoride has been proved even for bacterial and
archaeal Fe-SODs.21–25 The third residue considered is S82, a
potential target for post-translational modification of rat-
SOD2, as suggested by a phosphorylation prediction study. A
recent report, describing the phosphoproteomic screen on
proteins of the mitochondrial matrix, has identified
Mn-SOD as a target of phosphorylation.33
RESULTS
Molecular Properties of RatSOD2 Mutants
Using a heterologous expression system constituted by the
prokaryotic vector pT7-7 and the Escherichia coli host strain
BL21(DE3), a recombinant mature form of ratSOD2, called
SOD2(Q143), was produced and purified. In this recombi-
nant version, the initial Lys residue of the endogenous
mature enzyme (K25 in the premature form of ratSOD2) was
replaced by Met, representing the translation start codon in
the amplified ratSOD2 gene cloned in pT7-7. The purified
SOD2(Q143) was homogeneous when analyzed by SDS/
PAGE and showed an electrophoretic mobility corresponding
to a molecular mass of 24 kDa, slightly higher than the
expected size of the monomer subunit (22.3 kDa). The Mr of
SOD2(Q143) was also determined under nondenaturing
conditions by gel-filtration. The protein sample eluted as a
1216 Castellano et al.
Biopolymers

single symmetric peak with an apparent Mr of 90,000, a value
approaching the theoretical mass of a homotetramer.
To exclude that the small difference at the N-terminus
between endogenous ratSOD2 and SOD2(Q143) caused a
partial trimming of some residues in the recombinant pro-
tein, the molecular mass of the purified SOD2(Q143) was
analyzed by ESI/Q-TOF mass spectrometry on two different
preparations of the enzyme. Indeed, the major peak had an
experimental Mr of 22294.7, in good agreement with the the-
oretical value of 22294.3 derived from the amino acid
sequence of the expected heterologous product, including
the initial Met residue (Table I). Therefore, the purified
SOD2(Q143) was produced as an intact molecule, the only
difference with the endogenous ratSOD2 being at its N-ter-
minal residue.
The described expression system was productively used
for the obtainment of mutant forms of ratSOD2. To study
the possible variance between Gln and His at position 143 of
ratSOD2, a recombinant form of the enzyme was obtained,
carrying the H143 residue and thus called SOD2(H143).
Afterward, to investigate on the role of the Y34 position, the
Y34F replacement was realized on either SOD2(Q143) or
SOD2(H143) and the corresponding mutant forms were
called SOD2(Y34F-Q) and SOD2(Y34F-H), respectively. In a
similar way, the role of the S82 position was analyzed
through the S82A substitution and the corresponding mu-
tant forms were called SOD2(S82A-Q) and SOD2(S82A-H).
Variant and mutant forms of ratSOD2 had the same behavior
of SOD2(Q143), when analyzing their electrophoretic mobil-
ity on SDS/PAGE or elution under nondenaturing conditions
by gel-filtration. Therefore, all of them conserved the homo-
tetrameric structure of the enzyme. Moreover, all the
recombinant proteins were analyzed by ESI/Q-TOF mass
spectrometry and the experimental Mr values obtained from
two different preparations are reported in Table I. For all of
them, there was a good agreement with the theoretical Mr
derived from the amino acid sequence, taking into account
the presence of the initial Met residue.
Sulfhydryl Reactivity
The ratSOD2 amino acid sequence contains two cysteine res-
idues, C140 and C196 (sequence numbering of the mature
protein). Previous investigations on the closely related
human mitochondrial Mn-SOD, containing the same cyste-
ine residues, indicated a sulfhydryl reactivity toward thiol
reagents.26,27 MS analysis of SOD2(Q143), SOD2(H143),
and their mutants indicated that the sulfhydryl groups were
reactive. Indeed, besides the presence of potassium adducts,
an additional peak corresponding to a 305 Da extra mass was
clearly evident in all mass spectra of two different prepara-
tions of the six protein samples. As an example, Figure 1
reports the mass spectra of SOD2(S82A-Q) and SOD2(S82A-
H), containing the highest (32.0%) and lowest (2.7%)
amount of this extra peak, respectively. The 305 Da extra
mass can be accounted for a glutathionylation reaction
occurring on ratSOD2, likely due to a high sulfhydryl reactiv-
ity of its cysteine residue(s). The level of the glutathionylated
adduct was variable either among different or within the
same ratSOD2 form (Table I); however, no multiple gluta-
thionylated adducts were deduced from the mass spectra,
thus indicating that only one cysteine, among C140 and
C196, was covalently modified by glutathione. These findings
are intriguing, because a glutathionylated adduct on SOD2
was recently reported even for the eubacterial Fe-SOD from
P. haloplanktis.28 Hence, our data extend the occurrence of
this glutathionylation reaction, consistently detected in the
ratSOD2 forms analyzed, also to a mitochondrial SOD. The
great variability of the glutathionylation level observed in
preparations of the same ratSOD2 form do not allow any
conclusion concerning a possible relationship between amino
acid replacements and sulfhydryl reactivity of recombinant
ratSOD2.
Table I Molecular Mass Values (Mr) Obtained by ESI-Q/TOF
Mass Spectrometry Determined on Two Different Preparations of
Recombinant Wild Type and Mutant Forms of RatSOD2 and Rel-
ative Percentages of Glutathionylated Adduct, Calculated on the
Basis of the Relative Intensities of the Deconvoluted Mass Peaks
Protein
Experimental
Mra
Delta from
Theoretical Mr
Glutathionylated
Adduct (%)b
SOD2(Q143) 22294.7 10.4 5.1
10.0
SOD2(H143) 22303.8 10.4 7.5
5.7
SOD2(Y34F–Q) 22278.9 10.6 12.9
11.8
SOD2(Y34F–H) 22287.8 10.5 6.6
22.6
SOD2(S82A–Q) 22278.7 10.4 32.0
9.7
SOD2(S82A–H) 22287.8 10.5 2.7
30.0
a Mean value of the mass peak corresponding to the unmodified protein;
the variability among the two determinations never exceeded 50 ppm.b In the calculation of the total amount of unmodified and modified
protein, the potassium adducts revealed in high amount in almost all
samples have been included, whereas other poor represented adducts
have not been considered. Each first and second figure is referred to a
protein sample purified after the incubation at 708C in the absence or in
the presence of MnCl2, respectively, as indicated in the Materials and
Methods section.
Rat Mitochondrial Key Residues in Superoxide Dismutase 1217
Biopolymers

FIGURE 1 Electrospray mass spectrum of SOD2(S82A-Q) (panel A) and SOD2(S82A-H) (panel B).
The mixed disulfide with glutathione is indicated by a solid arrow, whereas the main potassium adduct
is indicated by a dashed arrow.
Table II Manganese Content and Specific Activity of Recombinant Wild Type and Mutant Forms of RatSOD2
Protein
Manganese Contenta
(mol/mol ratSOD2 subunit)
Specific Activityb
(U/mg protein)
Specific Activity
Referred to Metal Content
(U/mg protein3 mol Mn 3 mol subunit21)
SOD2(Q143) 0.30 5350 17,800
SOD2(H143) 0.26 750 2900
SOD2(Y34F–Q) 0.22 2650 12,000
SOD2(Y34F–H) 0.16 450 2800
SOD2(S82A–Q) 0.26 3750 14,400
SOD2(S82A–H) 0.16 500 3100
a Determined by graphite furnace atomic absorption spectrometry, as indicated in Materials and Methods.b Measured through the inhibition of the cytochrome c reduction in the xanthine/xanthine oxidase system, as indicated in Materials and Methods.
1218 Castellano et al.
Biopolymers

Another evaluation of the glutathionylation reaction on
the heterologous ratSOD2 forms was obtained through
immunoblotting experiments, using anti-glutathione anti-
bodies. This experiment confirmed the presence of the gluta-
thionylated adduct in all samples examined, because of the
presence of specific immunoreactive bands (data not shown).
Metal Content and Activity
The analysis of the metal content of the purified proteins
revealed that all of them bound manganese as metal cofactor,
even though in low amounts in either wild type or mutant
forms of the enzyme. An improvement of this content was
obtained in protein samples purified after a moderate ther-
mal treatment of the protein soluble fraction in the presence
of 10 mM MnCl2 (see Materials and Methods). However, the
manganese content of the MnCl2-treated samples remained
understoichiometric, as shown in Table II. Other SODs
belonging to the same family, even when purified from their
endogenous source, displayed a similar low metal content.
The differences in manganese content among the protein
samples analyzed probably indicate a lower metal binding
capacity exhibited by the ratSOD2 variant containing His at
position 143. A previous analysis on several SOD2 primary
structures pointed to the relevance of position 143 for the
type of metal bound in the active site of the enzyme.34 This
position is semiconservatively occupied by Gln or His, and
this difference is usually predictive of the active metal cofac-
tor of SOD2, namely Mn or Fe with Gln or His, respectively.
To exclude that the lower Mn content of SOD2(H143) with
respect to SOD2(Q143) was due to its replacement by Fe, the
iron content of both protein samples was determined.
Interestingly, besides the Mn ion, 0.16 and 0.18 mol Fe/
mol ratSOD2 subunit were found in SOD2(Q143) and
SOD2(H143), respectively. The presence of iron bound to
ratSOD2 could be related to the heterologous production of
the enzyme and/or to the occupancy of some empty metal
binding sites, as resulting from the low content of the active
manganese. Indeed, the uptake of putatively inactive metals
was already described for another SOD of the same family.35
However, the presence of a similar Fe content in wild type
and H143 variant of ratSOD2 indicates that the Q143H
replacement did not significantly alter the specificity of
ratSOD2 for metal uptake.
The SOD activity of the six heterologous proteins
reported in Table II was measured on enzyme preparations
obtained when the thermal purification step described in
the Materials and Methods section was carried out in the
presence of MnCl2, in order to improve the manganese ion
bound to each ratSOD2 form. For a better comparison
among the different proteins, the values of specific activity
were also referred to the Mn content. All enzymes were
produced in their active state; however, significant varia-
tions among the different protein samples were found. The
highest value of specific activity was found with
SOD2(Q143), whereas a very low value was observed with
SOD2(H143), because only 16% of the activity displayed
by SOD2(Q143) was measured. Vice versa, in SOD2
(Y34F-Q) and even more in SOD2(S82A-Q), the Y34F and
S82A replacements caused a modest effect, because 67 and
81% of the activity sustained by SOD2(Q143) was deter-
mined, respectively. When the effect of the same replace-
ments was analyzed in the H143 variant of ratSOD2, the
observed decrease of activity almost exclusively reflected
the presence of His143 residue, because the activity of
SOD2(Y34F-H) and SOD2(S82A-H) was similar to that
exhibited by SOD2(H143).
Heat Stability
To evaluate the heat resistance of ratSOD2, heat inactivation
profiles of SOD2(Q143), SOD2(H143), SOD2(Y34F-Q), and
SOD2(S82A-Q) were realized, using the enzyme preparations
obtained with the thermal purification step carried out in the
presence of MnCl2. To this aim, the protein samples were
incubated for 10 min at different temperatures and after-
ward, their residual activity was measured and compared to
the corresponding untreated samples. The resulting heat
inactivation profiles indicate that all the recombinant forms
of the enzyme are endowed with a significant heat resistance,
because of the high values of half-inactivation temperatures
(T1/2), ranging between 77 and 818C (see Figure 2). An eval-
uation of the different heat stability among the proteins was
attempted. In particular, the T1/2 of 808C calculated for
SOD2(Q143) was slightly decreased to 798C in SOD2(Y34F-
Q), whereas a more pronounced decrease of heat stability
was found in SOD2(H143), whose T1/2 was 778C; on the
contrary, in SOD2(S82A-Q) the T1/2 increased to 818C. Fur-thermore, some differences were found in the shape of the
profiles; for instance, that related to SOD2(H143) had a very
broad shape.
To examine the effect of temperature on the inactivation
reaction of recombinant wild type and mutant forms of rat-
SOD2, kinetics were carried out in the temperature interval
75–838C. As an example, the time course of inactivation at
808C of the four proteins is shown in Figure 3. All reactions
followed first-order kinetics with rates depending on the rat-
SOD2 form analyzed. Indeed, compared to SOD2(Q143)
(Figure 3A), a moderately slower rate was measured with
SOD2(S82A-Q) (Figure 3C); vice versa, with SOD2(H143)
Rat Mitochondrial Key Residues in Superoxide Dismutase 1219
Biopolymers
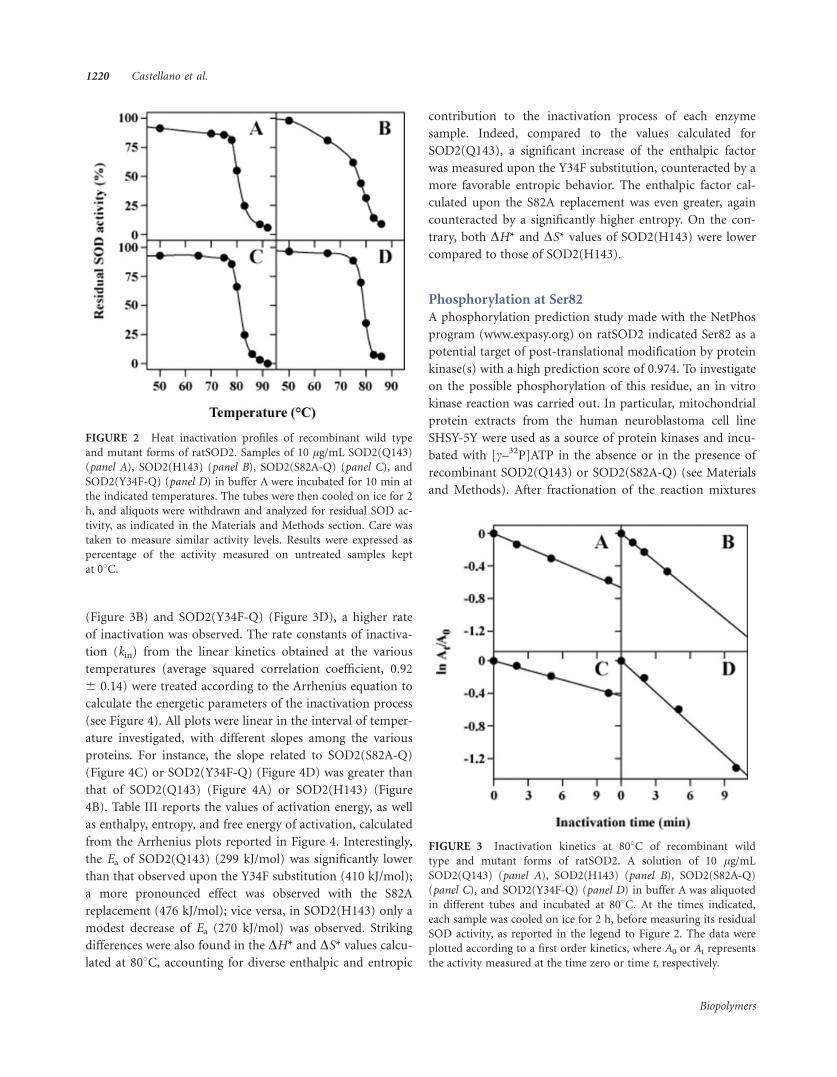
(Figure 3B) and SOD2(Y34F-Q) (Figure 3D), a higher rate
of inactivation was observed. The rate constants of inactiva-
tion (kin) from the linear kinetics obtained at the various
temperatures (average squared correlation coefficient, 0.92
6 0.14) were treated according to the Arrhenius equation to
calculate the energetic parameters of the inactivation process
(see Figure 4). All plots were linear in the interval of temper-
ature investigated, with different slopes among the various
proteins. For instance, the slope related to SOD2(S82A-Q)
(Figure 4C) or SOD2(Y34F-Q) (Figure 4D) was greater than
that of SOD2(Q143) (Figure 4A) or SOD2(H143) (Figure
4B). Table III reports the values of activation energy, as well
as enthalpy, entropy, and free energy of activation, calculated
from the Arrhenius plots reported in Figure 4. Interestingly,
the Ea of SOD2(Q143) (299 kJ/mol) was significantly lower
than that observed upon the Y34F substitution (410 kJ/mol);
a more pronounced effect was observed with the S82A
replacement (476 kJ/mol); vice versa, in SOD2(H143) only a
modest decrease of Ea (270 kJ/mol) was observed. Striking
differences were also found in the DH* and DS* values calcu-
lated at 808C, accounting for diverse enthalpic and entropic
contribution to the inactivation process of each enzyme
sample. Indeed, compared to the values calculated for
SOD2(Q143), a significant increase of the enthalpic factor
was measured upon the Y34F substitution, counteracted by a
more favorable entropic behavior. The enthalpic factor cal-
culated upon the S82A replacement was even greater, again
counteracted by a significantly higher entropy. On the con-
trary, both DH* and DS* values of SOD2(H143) were lower
compared to those of SOD2(H143).
Phosphorylation at Ser82
A phosphorylation prediction study made with the NetPhos
program (www.expasy.org) on ratSOD2 indicated Ser82 as a
potential target of post-translational modification by protein
kinase(s) with a high prediction score of 0.974. To investigate
on the possible phosphorylation of this residue, an in vitro
kinase reaction was carried out. In particular, mitochondrial
protein extracts from the human neuroblastoma cell line
SHSY-5Y were used as a source of protein kinases and incu-
bated with [c–32P]ATP in the absence or in the presence of
recombinant SOD2(Q143) or SOD2(S82A-Q) (see Materials
and Methods). After fractionation of the reaction mixtures
FIGURE 2 Heat inactivation profiles of recombinant wild type
and mutant forms of ratSOD2. Samples of 10 lg/mL SOD2(Q143)
(panel A), SOD2(H143) (panel B), SOD2(S82A-Q) (panel C), and
SOD2(Y34F-Q) (panel D) in buffer A were incubated for 10 min at
the indicated temperatures. The tubes were then cooled on ice for 2
h, and aliquots were withdrawn and analyzed for residual SOD ac-
tivity, as indicated in the Materials and Methods section. Care was
taken to measure similar activity levels. Results were expressed as
percentage of the activity measured on untreated samples kept
at 08C.
FIGURE 3 Inactivation kinetics at 808C of recombinant wild
type and mutant forms of ratSOD2. A solution of 10 lg/mL
SOD2(Q143) (panel A), SOD2(H143) (panel B), SOD2(S82A-Q)
(panel C), and SOD2(Y34F-Q) (panel D) in buffer A was aliquoted
in different tubes and incubated at 808C. At the times indicated,
each sample was cooled on ice for 2 h, before measuring its residual
SOD activity, as reported in the legend to Figure 2. The data were
plotted according to a first order kinetics, where A0 or At represents
the activity measured at the time zero or time t, respectively.
1220 Castellano et al.
Biopolymers
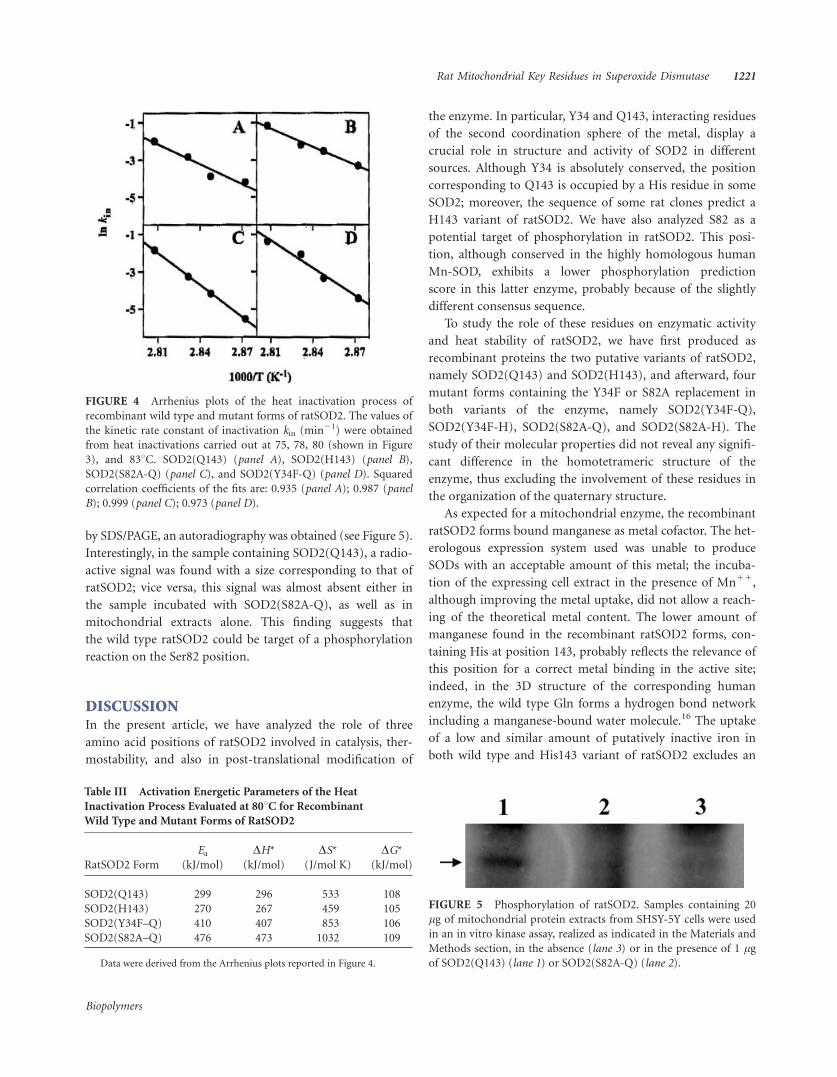
by SDS/PAGE, an autoradiography was obtained (see Figure 5).
Interestingly, in the sample containing SOD2(Q143), a radio-
active signal was found with a size corresponding to that of
ratSOD2; vice versa, this signal was almost absent either in
the sample incubated with SOD2(S82A-Q), as well as in
mitochondrial extracts alone. This finding suggests that
the wild type ratSOD2 could be target of a phosphorylation
reaction on the Ser82 position.
DISCUSSIONIn the present article, we have analyzed the role of three
amino acid positions of ratSOD2 involved in catalysis, ther-
mostability, and also in post-translational modification of
the enzyme. In particular, Y34 and Q143, interacting residues
of the second coordination sphere of the metal, display a
crucial role in structure and activity of SOD2 in different
sources. Although Y34 is absolutely conserved, the position
corresponding to Q143 is occupied by a His residue in some
SOD2; moreover, the sequence of some rat clones predict a
H143 variant of ratSOD2. We have also analyzed S82 as a
potential target of phosphorylation in ratSOD2. This posi-
tion, although conserved in the highly homologous human
Mn-SOD, exhibits a lower phosphorylation prediction
score in this latter enzyme, probably because of the slightly
different consensus sequence.
To study the role of these residues on enzymatic activity
and heat stability of ratSOD2, we have first produced as
recombinant proteins the two putative variants of ratSOD2,
namely SOD2(Q143) and SOD2(H143), and afterward, four
mutant forms containing the Y34F or S82A replacement in
both variants of the enzyme, namely SOD2(Y34F-Q),
SOD2(Y34F-H), SOD2(S82A-Q), and SOD2(S82A-H). The
study of their molecular properties did not reveal any signifi-
cant difference in the homotetrameric structure of the
enzyme, thus excluding the involvement of these residues in
the organization of the quaternary structure.
As expected for a mitochondrial enzyme, the recombinant
ratSOD2 forms bound manganese as metal cofactor. The het-
erologous expression system used was unable to produce
SODs with an acceptable amount of this metal; the incuba-
tion of the expressing cell extract in the presence of Mn11,
although improving the metal uptake, did not allow a reach-
ing of the theoretical metal content. The lower amount of
manganese found in the recombinant ratSOD2 forms, con-
taining His at position 143, probably reflects the relevance of
this position for a correct metal binding in the active site;
indeed, in the 3D structure of the corresponding human
enzyme, the wild type Gln forms a hydrogen bond network
including a manganese-bound water molecule.16 The uptake
of a low and similar amount of putatively inactive iron in
both wild type and His143 variant of ratSOD2 excludes an
FIGURE 4 Arrhenius plots of the heat inactivation process of
recombinant wild type and mutant forms of ratSOD2. The values of
the kinetic rate constant of inactivation kin (min21) were obtained
from heat inactivations carried out at 75, 78, 80 (shown in Figure
3), and 838C. SOD2(Q143) (panel A), SOD2(H143) (panel B),
SOD2(S82A-Q) (panel C), and SOD2(Y34F-Q) (panel D). Squared
correlation coefficients of the fits are: 0.935 (panel A); 0.987 (panel
B); 0.999 (panel C); 0.973 (panel D).
Table III Activation Energetic Parameters of the Heat
Inactivation Process Evaluated at 808C for Recombinant
Wild Type and Mutant Forms of RatSOD2
RatSOD2 Form
Ea
(kJ/mol)
DH*
(kJ/mol)
DS*
(J/mol K)
DG*
(kJ/mol)
SOD2(Q143) 299 296 533 108
SOD2(H143) 270 267 459 105
SOD2(Y34F–Q) 410 407 853 106
SOD2(S82A–Q) 476 473 1032 109
Data were derived from the Arrhenius plots reported in Figure 4.
FIGURE 5 Phosphorylation of ratSOD2. Samples containing 20
lg of mitochondrial protein extracts from SHSY-5Y cells were used
in an in vitro kinase assay, realized as indicated in the Materials and
Methods section, in the absence (lane 3) or in the presence of 1 lgof SOD2(Q143) (lane 1) or SOD2(S82A-Q) (lane 2).
Rat Mitochondrial Key Residues in Superoxide Dismutase 1221
Biopolymers

alteration of the specificity for metal uptake upon the
Q143H replacement.
All the recombinant ratSOD2 forms were purified as
active enzymes. The different values of specific activity meas-
ured on the six protein samples reflect the specific role of the
analyzed amino acid positions in catalysis. The greatest
effects on the activity are related to the position 143. Indeed,
the lowest levels of specific activity were found with all rat-
SOD2 forms, containing His at position 143, with a sixfold
reduction of activity compared to SOD2(Q143). Also for the
human enzyme, amino acid replacements at the wild type
Q143 position cause a dramatic decrease of activity.36 At
present, we do not know the putative physiological role of
the His143 variant of ratSOD2.29–31 We can speculate that
the low activity displayed in vivo by SOD2(H143) is likely
compensated by other antioxidant mechanisms.
The effects of Y34F and S82A substitutions on the activity
are much less crucial. Indeed, the 1.5-fold reduction
observed when the activity of SOD2(Y34F-Q) was compared
with that of SOD2(Q143) is similar to that reported for the
corresponding mutation in the human Mn-SOD.17,37 It has
been proposed that the effect of this mutation on the activity
becomes more crucial when a highly efficient catalysis is
required by increased superoxide levels.17 A lower effect was
found upon the S82A replacement, because the activity of
SOD2(S82A-Q) was quite similar to that determined on
SOD2(Q143). This is not surprising, because S82 is far away
from the active site of the protein molecule. The role of this
residue, probably exposed to solvent, could be related to a
possible regulation of the SOD activity by post-translational
modifications, as suggested by the phosphorylation analysis
presented in this work. The specific activity measured on
mutants containing the Y34F or S82A replacement in the
H143 variant of ratSOD2 was very similar to that determined
on SOD2(H143). Clearly, the significant reduction of activity
only depends on the presence of His at position 143.
The study of the inactivation process of ratSOD2 has pro-
vided some important insights on the high thermo-tolerance
of this enzyme, probably reflecting a required ability for its
adaptation to different types of stress. Similarly, also the
human Mn-SOD displays a great heat resistance.17 The inac-
tivation studies on ratSOD2 led to interesting observations
on the role of the analyzed amino acid positions. Indeed, the
thermal inactivation profiles showed that SOD2(S82A-Q)
is slightly more resistant to heat inactivation than
SOD2(Q143); vice versa, a decrease in the half-inactivation
temperature is observed in SOD2(Y34F-Q) and even more in
SOD2(H143). Differential scanning calorimetry experiments
carried out on the human Mn-SOD highlighted the great im-
portance of the Q143 position for the thermal stability of the
enzyme, because of a significant reduction in the unfolding
transition temperature observed in Q143 mutants of the
enzyme.36 On the other hand, similar studies on the Y34F
substitution pointed to a significant improvement of the
thermal stability of the wild type human enzyme.17 Our inac-
tivation studies on ratSOD2 are in agreement with the pub-
lished observations on the structural role played by the
human Q143 position, thus suggesting that rat Q143 is also
important for keeping the correct structure of ratSOD2. Vice
versa, an apparent disagreement exists with the results
reported in this work for the rat Y34F mutant, possessing a
slightly reduced heat resistance; however, this feature could
be in part related to the different methodology used to mea-
sure the effect of a heat treatment on the enzyme. We have
also evaluated the possibility that the different thermal
behavior of the ratSOD2 forms could be related to a different
glutathionylation extent in the protein samples analyzed.
However, this seems unlikely, because the enzyme prepara-
tions used in this thermal inactivation studies possess a low
percentage of glutathionylation, not exceeding 11.8%. Fur-
thermore, the modification level is very similar in three out
of the four protein samples considered in the study.
We have also measured the activation parameters of the
heat inactivation process and found striking differences in
the enthalpic and entropic contribution to this reaction. The
calculated values of Ea fall in a large interval (270–476 kJ/
mol), the highest values being curiously unusual for a protein
from a mesophilic source. Among the different proteins,
SOD2(Y34F-Q) and even more SOD2(S82A-Q) exhibit a sig-
nificantly higher Ea compared to SOD2(Q143), whereas a
slight reduction of this energetic parameter is observed in
SOD2(H143). The enthalpic and entropic contribution to
the activation of the inactivation process greatly varies
among the different ratSOD2 forms. Indeed, the more favor-
able entropic contribution to the process was found with
SOD2(S82A-Q), followed by SOD2(Y34F-Q). These findings
suggest that these replacements render the active site more
disordered during the inactivation process. However, this
favorable entropy is counteracted by an unfavorable enthalpy
in these mutant forms, likely due to a greater difficulty to
reach the transition state. A different behavior was found
with SOD2(H143), because in this case both the enthalpy
and the entropy of the inactivation process were found
reduced.
To identify possible post-translational modifications in
the recombinant purified proteins, they were analyzed by
ESI/Q-TOF mass spectrometry. Surprisingly, the analysis
revealed the presence of a 305 Da extra mass, due to a
glutathionylated adduct, as confirmed by the immunoblot-
ting analysis. This modification was present in all proteins
1222 Castellano et al.
Biopolymers

produced, even though to a different extent. The gluta-
thionylation reaction observed in heterologous proteins
produced in E. coli points to the high reactivity of rat-
SOD2 toward glutathione, a physiological thiol normally
present in this host strain.38,39 Recent studies on the regu-
lation of protein function by ROS suggest that S-gluta-
thionylation is an important mechanism involved in cellu-
lar response to oxidative damage and redox signaling.40–46
An old study reported the presence of glutathionylated
forms for chicken Cu/Zn-SOD,47 but the role of this
modification was not clarified yet. Recently, we have
reported that the Fe-SOD from P. haloplanktis is gluta-
thionylated in vitro on Cys57 and that the enzyme under-
goes an in vivo S-glutathionylation when overproduced in
E. coli cells.28 Also in mammalian cells, protein glutathio-
nylation is involved both in defence against oxidative
damage and in redox signaling. In mitochondria, the high
concentration of protein thiols and glutathione has been
related to the presence of several glutathionylated pro-
teins.48,49 The human mitochondrial Mn-SOD is very sen-
sitive to reducing agents,26 and this susceptibility has been
linked to the high reactivity of Cys196 exposed on the
surface of the tetrameric enzyme. Moreover, the I58T iso-
form of the human enzyme, organized as a dimer rather
than a normal tetramer, exposes also Cys140, a residue
normally buried in the tetrameric structure.50 Interestingly,
these two cysteine residues are also conserved in the
mature ratSOD2. Our data indicate that only one cysteine
is target of glutathionylation; on the basis of the high
similarity between human and rat SOD2, the candidate
for this reaction is Cys196, because the amino acid
replacements realized on ratSOD2 did not alter its homo-
tetrameric structure and probably do not expose Cys140
to the solvent. The MS analysis has revealed great differ-
ences among the glutathionylation levels of the various
protein samples; however, these differences could not be
related to a change in the sulfhydryl reactivity of ratSOD2
caused by the amino acid replacements realized.
A phosphorylation prediction study on ratSOD2 indicated
Ser82 as a potential target of post-translational modification
by protein kinase(s). Indeed, a recent article has reported
that the cytoplasmic Mn-SOD from Listeria monocytogenes, a
facultative intracellular pathogen for humans and animals, is
phosphorylated on serine and/or threonine residues and that
this post-translational modification downregulates the enzy-
matic activity during the stationary phase.51 Moreover, a
comprehensive analysis of cytoplasmic proteins phosphoryla-
tion in Campylobacter jejuni indicated Fe-SOD as one of the
major phosphoprotein.52 To date, there is no information
available on the amino acid position(s) susceptible to this
modification. Ser82 is conserved in several SOD2 from differ-
ent organisms, including human, rat, and L. monocytogenes
Mn-SOD. To investigate on the possible phosphorylation of
this residue, we have used an in vitro kinase reaction, in
which mitochondrial protein extracts from a human cell line
were considered as a source of serine/threonine protein
kinases. The experiment showed that the recombinant
SOD2(Q143) is phosphorylated by mitochondrial kinases;
vice versa, the mutant SOD2(S82A-Q) is not, suggesting that
Ser82 is the likely target of this modification. In the crystal
structure of the corresponding human enzyme, this residue
is located in a region apparently exposed to solvent.16 It
remains to be determined the function of ratSOD2 phospho-
rylation and its relation with the enzymatic activity of the rat
enzyme.
CONCLUSIONSThe produced recombinant forms of ratSOD2 allowed an
investigation on the role of three amino acid positions of the
enzyme. Among them, the most crucial role is played by
Q143, either in catalysis or structure of ratSOD2. Moreover,
our results on the possible post-translational modification of
the enzyme suggest that ratSOD2 could be finely regulated
by physiological compounds, such as glutathione, or other
modifications, such as phosphorylation by protein kinases.
We wish to stress that the resistance to peroxynitrite of
eubacterial Fe-SOD from P. haloplanktis is significantly
improved upon glutathionylation of the enzyme.28 Also rat-
SOD2, as already demonstrated for the corresponding
human enzyme, is nitrated on Y34 and inactivated by perox-
ynitrite (unpublished observation). Future research should
speculate on the in vivo glutathionylation and/or phospho-
rylation of the endogenous ratSOD2 in mammalian cells and
on the possible functional role of such post-translational
events.
MATERIALS AND METHODS
MaterialsRestriction and modifying enzymes, and [c–32P]ATP were from GE
Healthcare. Plasmid pGEM T-easy was from Promega. Vector pT7-7
and the E. coli BL21(DE3) strain were from Novagen. The Qiagen
kits (M-Medical) were used for purification of plasmids and DNA
fragments. Oligonucleotide synthesis and nucleotide sequencing
was carried out by Primm. Isopropyl-b-thiogalactopiranoside(IPTG), xanthine, xanthine oxidase, and cytochrome c were from
Sigma-Aldrich. Anti-glutathione monoclonal antibodies were from
Chemicon; horseradish peroxidase-conjugated anti-mouse antibod-
ies were from Santa Cruz. The chemiluminescent SuperSignal West
Rat Mitochondrial Key Residues in Superoxide Dismutase 1223
Biopolymers

Pico kit was from Pierce. Fuji films were used for autoradiography.
RPMI 1640 medium, fetal bovine serum (FBS), L-glutamine,
penicillin G, streptomycin, trypsin were purchased from Cambrex.
Protease inhibitor cocktail was obtained from Roche Diagnostics.
HPLC-grade solvents for mass spectrometry were obtained from
Carlo Erba. All other chemicals were of analytical grade. The follow-
ing buffers were used: buffer A, 20 mM Tris-Cl, pH 7.8; buffer B,
50 mM potassium phosphate, pH 7.2; buffer C, 20 mM Tris-Cl, pH
7.8, 50% (v/v) glycerol; buffer D, 100 mM potassium phosphate, pH
7,8, 0.1 mM Na2EDTA; buffer M, 5 mM Hepes, pH 7.4, 250 mM
mannitol, 0.5 mM EGTA, 5 mM Hepes, 0,1% BSA; PBS buffer,
10 mM sodium phosphate, pH 7.2, 150 mM NaCl; kinase buffer,
10 mM MOPS, pH 7.0, 7.7 mM MgCl2, 0.5 mM Na2EDTA, 0.5 mM
NaF, 0.5 mM Na3VO4.
MethodsSOD activity was measured at 258C in buffer D by the inhibi-
tion of cytochrome c reduction caused by superoxide anions
generated with the xanthine/xanthine oxidase method.35,53 One
unit of SOD activity was defined as the amount of enzyme
that caused 50% inhibition of cytochrome c reduction. The in
vitro kinase assay was realized by mixing mitochondrial protein
extracts with recombinant proteins in 30 lL kinase buffer sup-
plemented with 20 lM ATP and 1 lCi [c-32P]ATP. The reac-
tion mixture was incubated for 20 min at 308C and blocked by
adding the sample dilution buffer for SDS/PAGE. After frac-
tionation by SDS/PAGE, the gel was dried and radioactive signals
were visualized by autoradiography.
Protein concentration was determined by the method of Brad-
ford,54 using bovine serum albumin as standard. Purity of protein
samples was assessed by 14% SDS/PAGE according to standard pro-
tocols.55 The quaternary structure of wild type and mutant forms of
ratSOD2 was evaluated by gel-filtration on a Superdex 75 10/300
GL column (GE Healthcare). The metal content of the recombinant
protein samples was determined by graphite furnace atomic absorp-
tion spectrometry, as previously indicated.25 The presence of gluta-
thionylated adducts in the purified protein samples was revealed
through immunoblotting experiments realized with monoclonal
anti-glutathione antibodies, as previously reported.28
The energy of activation Ea and the Arrhenius constant A of the
heat inactivation process were calculated through the equation ln
kin 5 ln A 2 Ea/R 3 1/T, where kin is the heat inactivation rate con-
stant and R is the gas constant (8.314 J/mol K). The other thermo-
dynamic parameters of activation, namely enthalpy (DH*), entropy
(DS*), and free energy (DG*), were calculated at 808C according to
the equations: DH* 5 Ea 2 R 3 T; DS* 5 R 3 ln (h 3 NA 3 A/R
3 T 3 e); DG* 5 DH* 2 T 3 DS*, where h is the Planck constant
(6.624 3 10234J s), NA is the Avogadro’s number (6.023 3 1023
molecules/mol) and e is the Napier’s number, the base of natural
logarithm.
Expression Vectors and Site-Directed MutagenesisThe cDNA clone containing the ratSOD2 gene including flanking
regions has been previously described.56 Transformation of bacterial
strains, preparation of plasmids, and other details of DNA recombi-
nant technology were as previously described.57 To engineer a vector
for the heterologous production of recombinant ratSOD2 without
its leader peptide, a DNA segment was amplified by PCR, using the
rat cDNA clone as a template and the following oligonucleotides as
primers: 50d-C63TCCCGGCAT-ATG-CACAGCCT83-30 (forward
primer) and 50d-C709ACCCACCCCGGGCCTGAC691-30 (reverse
primer). Numbering in primers begins from the starting codon of
the complete ratSOD2 gene, including the segment encoding the
leader peptide; italicized letters indicate the base substitutions to
create the NdeI and XmaI cloning sites in direct and reverse primer,
respectively; the new engineered initiation codon is between two
hyphens. The 647 bp amplification product was subcloned in
pGEM T-easy vector and sequenced to confirm the identity with the
ratSOD2 gene reported in databases.29 After digestion of the
recombinant plasmid with NdeI and XmaI, the resulting 627 bp
fragment containing the ratSOD2 gene was cloned into the prokary-
otic expression vector pT7-7, previously digested with the same en-
donucleases. The vector was controlled by restriction analysis and
nucleotide sequencing, and to our surprise it contained the C499AC
codon, predicting the H143 variant of ratSOD2. Therefore, in order
to obtain the Q143 variant of ratSOD2, a site-directed mutagenesis
on the pT7-7 derivative vector was realized.58 In particular, the
following pairs of oligonucleotides were used, in which the base
mismatch introduced to create the desired amino acid replacement
is italicized: 50d-G491CTCTAATCAGGACCCACTG510-30 and 50d-
C510AGTGGGTCCTGATTAGAGC491-30 (forward and reverse
primer for the production of the Q143 variant of ratSOD2). The
PCR reaction, containing the engineered vector as template, the
specific primers and the PfuTurboTM DNA polymerase, was carried
out according to the protocol of the QuickChangeTM Site-Directed
Mutagenesis Kit (Stratagene). The correct substitution, as well as
the absence of other undesired mutations, was confirmed by
sequence analysis. In conclusion, the new engineered vectors, called
vSOD2(Q143) and vSOD2(H143), respectively, predicted the pro-
duction of Q143 and H143 variant of ratSOD2, called SOD2(Q143)
and SOD2(H143), respectively. Using these vectors, recombinant
forms of the mature ratSOD2 were produced, in which the initial
Lys residue of the endogenous mature enzyme (K25 in its premature
form) was replaced by Met; vice versa, the original stop codon of
the ratSOD2 gene was retained.
The mutant forms of ratSOD2, containing the replacements
Y34F and S82A were obtained through a site-directed mutagenesis
of the ratSOD2 gene present in vSOD2(Q143) and vSOD2(H143).
The following pairs of oligonucleotides were used: 50d-A164CGCGACCTTCGTGAACAAT183-3
0 and 50d-A183TTGTTCAC-
GAAGGTCGCGT164-30 (forward and reverse primer for the Y34F
replacement); 5d0-A307CAAACCTGGCCCCTAAGGGT327-30 and
50d-A327CCCTTAGGGGCCAGGTTTGT307-30 (forward and reverse
primer for S82A replacement). Therefore, four new vectors were
obtained, called vSOD2(Y34F-Q), vSOD2(Y34F-H), vSOD2(S82A-
Q) and vSOD2(S82A-H).
Expression and Purification of Recombinant ProteinsThe E. coli BL21(DE3) strain was transformed with the prokary-
otic expression vector vSOD2(Q143) or its mutant forms and
grown at 378C in Luria-Bertani medium containing 0.1 mg/mL
ampicillin in the absence or in the presence of 1 mg/L MnSO4.
When OD600 reached 0.6, 0.4 mM IPTG was added for induction,
and cultures were grown up for additional 3 h. Bacterial cells
were harvested by centrifugation at 5000 rpm for 15 min and pel-
1224 Castellano et al.
Biopolymers

lets were resuspended in buffer A. Cell lysis was obtained by a
cell disruption system (Constant System, UK) at 1.5 kbar; after-
ward, the cell homogenate was incubated at 378C for 30 min in the
presence of 40 U/mL DNAasi A to remove nucleic acids that some-
times form complexes with proteins. Cell homogenate was then
centrifuged at 100,000g for 2 h and supernatant was incubated at
708C for 30 min to denature most of the E. coli host proteins,
removed by centrifugation at 30,000g for 30 min. This purification
strategy, suggested by the high thermal stability of the closely
related human SOD2,17 was validated by the heat stability experi-
ments of ratSOD2 shown in this work. To improve the metal uptake
of all heterologous ratSOD2 forms, the incubation at 708C was car-
ried out in the presence of 10 mM MnCl2. The supernatant was
then extensively dialyzed against buffer B and loaded onto an
Econo-Pac cartridge (5 mL) hydroxyapatite column (Bio-Rad),
connected to a FPLC system (Pharmacia), equilibrated at room
temperature with buffer B. Protein elution was achieved by a linear
50–500 mM potassium phosphate gradient and the identification of
the recombinant product was made by the SOD assay and/or elec-
trophoretic mobility on SDS/PAGE. The recombinant protein was
eluted as a single symmetrical peak at around 200 mM potassium
phosphate; however, an additional purification step was necessary,
and the active fractions were concentrated by Aquacide IIA (Calbio-
chem) and loaded on a Superdex 75 HR10/30 column to remove
minor protein contaminants. Sometimes, during this gel-filtration
experiment we observed an additional peak with an absorbance
maximum at 260 nm; this material was identified as nucleic acids
copurifying with the heterologous product on the hydroxyapatite
column. Fractions containing single protein bands were pooled
together and concentrated. Care was taken to avoid precipitation of
the purified protein, when its concentration was higher than 1 mg/
mL. The concentrated material was then dialyzed against buffer C
and stored at 2208C. Under these conditions, the final yield was
nearly 1 mg from 1 L culture. No significant differences emerged in
the purification procedure among wild type or mutant forms of rat-
SOD2. The presence of MnSO4 during growth of the bacterial cul-
ture did not improve the metal uptake of ratSOD2. Other details of
the purification procedure were as previously described.24
Mass Spectrometry AnalysisThe relative molecular mass (Mr) of wild type and mutant forms of
ratSOD2 was determined by a Q-TOF Micro mass spectrometer
(Waters, Milford, MA), fitted with a Z-spray electrospray ion
source. The capillary source voltage and the cone voltage were set at
3000 and 35 V, respectively. The source temperature was kept at
808C and nitrogen was used as a drying gas (flow rate about 50 L/
h). Before ESI/MS analysis, proteins were desalted by RP-HPLC, as
previously reported.28 Using a mixture of acetonitrile/0.1% formic
acid in water (50/50, v/v), the sample was brought to a concentra-
tion of 1 pmol/lL and infused into the system at a flow rate of
5 lL/min. The acquisition and deconvolution of data were per-
formed by MassLynx software.
Mitochondrial ExtractsThe human neuroblastoma cell line SHSY-5Y was obtained from
American Type Culture Collection. SHSY-5Y cells were grown in
RPMI 1640 medium supplemented with 10% FBS, 2 mM L-gluta-
mine, 100 IU/mL penicillin G and 100 lg/mL streptomicyn in
humidified incubator at 378C under 5% CO2 atmosphere. Cells
were split and seeded in plates (75 cm2) every 3 days and used for
assays during exponential phase of growth.
For the preparation of mitochondrial protein extracts, SHSY-5Y
cells were plated at a density of 2 3 106 cells/plate (75 cm2) and
grown to subconfluency. Cells were then harvested, washed in PBS
buffer and then suspended in buffer M supplemented with a prote-
ase inhibitors cocktail, and homogenized. The homogenate was cen-
trifuged at 800g for 10 min at 48C and the supernatant was than
centrifuged at 12,000g for 30 min at 48C. The resulting pellet (mito-
chondrial fraction) was suspended in buffer M, whereas the final
supernatant represented the cytosolic fraction.
REFERENCES1. Finkel, T. Curr Opin Cell Biol 2003, 15, 247–254.
2. Adler, V.; Yin, Z.; Tew, K. D.; Ronai, Z. Oncogene 1999, 18,
6104–6111.
3. Sauer, H.; Wartenberg, M.; Hescheler, J. Cell Physiol Biochem
2001, 11, 173–186.
4. McCord, J. M.; Boyle, J. A.; Day, E. D., Jr.; Rizzo, L. J.; Salin, M.
L. In Superoxide and Superoxide dismutases; Michelson, A. M.;
McCord, J. M.; Fridovich, I., Eds.; Academic Press: London,
1977, pp 129–138.
5. Fridovich, I. J Biol Chem 1989, 264, 7761–7764.
6. Bannister, J. V.; Bannister, W. H.; Rotilio, G. CRC Crit Rev
Biochem 1987, 22, 111–180.
7. Zelko, I. N.; Mariani, T. J.; Folz, R. J. Free Radic Biol Med 2002,
33, 337–349.
8. Wispe, J. R.; Warner, B. B.; Clark, J. C.; Dey, C. R.; Neuman, J.;
Glasser, S. W.; Crapo, J. D.; Chang, L. Y.; Whitsett, J. A. J Biol
Chem 1992, 267, 23937–23941.
9. Hirose, K.; Longo, D. L.; Oppenheim, J. J.; Matsushima, K.
FASEB J 1993, 7, 361–368.
10. Li, Y.; Huang, T. T.; Carlson, E. J.; Melov, S.; Ursell, P. C.; Olson,
J. L.; Noble, L. J.; Yoshimura, M. P.; Berger, C.; Chan, P. H.;
Wallace, D. C.; Epstein, C. J. Nat Genet 1995, 11, 376–381.
11. MacMillan-Crow, L. A.; Cruthirds, D. L. Free Radic Res 2001,
34, 325–336.
12. Vincent, A. M.; Russell, J. W.; Sullivan, K. A.; Backus, C.; Hayes,
J. M.; McLean, L. L.; Feldman, E. L. Exp Neurol 2007, 208, 216–
227.
13. Lebovitz, R. M.; Zhang, H.; Vogel, H.; Cartwright, J., Jr.;
Dionne, L.; Lu, N.; Huang, S.; Matzuk, M. M. Proc Natl Acad
Sci USA 1996, 93, 9782–9787.
14. Melov, S.; Coskun, P.; Patel, M.; Tuinstra, R.; Cottrell, B.; Jun,
A. S.; Zastawny, T. H.; Dizdaroglu, M.; Goodman, S. I.; Huang,
T. T.; Miziorko, H.; Epstein, C. J.; Wallace, D. C. Proc Natl Acad
Sci USA 1999, 96, 846–851.
15. Wispe, J. R.; Clark, J. C.; Burhans, M. S.; Kropp, K. E.;
Korfhagen, T. R.; Whitsett, J. A. Biochim Biophys Acta 1989,
994, 30–36.
16. Borgstahl, G. E.; Parge, H. E.; Hickey, M. J.; Beyer, W. F., Jr.;
Hallewell, R. A.; Tainer, J. A. Cell 1992, 71, 107–118.
17. Guan, Y.; Hickey, M. J.; Borgstahl, G. E.; Hallewell, R. A.;
Lepock, J. R.; O’Connor, D.; Hsieh, Y.; Nick, H. S.; Silverman,
D. N.; Tainer, J. A. Biochemistry 1998, 37, 4722–4730.
Rat Mitochondrial Key Residues in Superoxide Dismutase 1225
Biopolymers

18. MacMillan-Crow, L. A.; Crow, J. P.; Kerby, J. D.; Beckman, J. S.;
Thompson, J. A. Proc Natl Acad Sci USA 1996, 93, 11853–
11858.
19. MacMillan-Crow, L. A.; Crow, J. P.; Thompson, J. A. Biochemis-
try 1998, 37, 1613–1622.
20. Yamakura, F.; Taka, H.; Fujimura, T.; Murayama, K. J Biol
Chem 1998, 273, 14085–14089.
21. Sorkin, D. L.; Duong, D. K.; Miller, A. F. Biochemistry 1997, 36,
8202–8208.
22. Soulere, L.; Claparols, C.; Perie, J.; Hoffmann, P. Biochem J
2001, 360, 563–567.
23. Ursby, T.; Adinolfi, B. S.; Al-Karadaghi, S.; De Vendittis, E.; Boc-
chini, V. J Mol Biol 1999, 286, 189–205.
24. De Vendittis, E.; Ursby, T.; Rullo, R.; Gogliettino, M. A.;
Masullo, M.; Bocchini, V. Eur J Biochem 2001, 268, 1794–1801.
25. Castellano, I.; Di Maro, A.; Ruocco, M. R.; Chambery, A.;
Parente, A.; Di Martino, M. T.; Parlato, G.; Masullo, M.; De
Vendittis, E. Biochimie 2006, 88, 1377–1389.
26. Matsuda, Y.; Higashiyama, S.; Kijima, Y.; Suzuki, K.; Kawano,
K.; Akiyama, M.; Kawata, S.; Tarui, S.; Deutsch, H. F.;
Taniguchi, N. Eur J Biochem 1990, 194, 713–720.
27. Hernandez-Saavedra, D.; McCord, J. M. Cancer Res 2003, 63,
159–163.
28. Castellano, I.; Ruocco, M. R.; Cecere, F.; Di Maro, A.;
Chambery, A.; Michniewicz, A.; Parlato, G.; Masullo, M.; De
Vendittis, E. Biochim Biophys Acta 2008, 1784, 816–826.
29. Ho, Y. S.; Crapo, D. J. Nucleic Acids Res 1987, 15, 10070.
30. Ho, Y. S.; Howard, A. J.; Crapo, D. J. Am J Respir Cell Mol Biol
1991, 4, 278–286.
31. Hurt, J.; Hsu, J. L.; Dougal, W. C.; Visner, G. A.; Burr, I. M.;
Nick, H. S. Nucleic Acids Res 1992, 20, 2985–2990.
32. Gogliettino, M. A.; Tanfani, F.; Scire, A.; Ursby, T.; Adinolfi, B.
S.; Cacciamani, T.; De Vendittis, E. Biochemistry 2004, 43,
2199–2208.
33. Hopper, R. K.; Carroll, S.; Aponte, A. M.; Johnson, D. T.;
French, S.; Shen, R. F.; Witzmann, F. A.; Harris, R. A.; Balaban,
R. S. Biochemistry 2006, 45, 2524–2536.
34. Wintjens, R.; Gilis, D.; Rooman, M. Proteins 2008, 70, 1564–
1577.
35. Dello Russo, A.; Rullo, R.; Nitti, G.; Masullo, M.; Bocchini, V.
Biochim Biophys Acta 1997, 1343, 23–30.
36. Leveque, V. J. P.; Stroupe, M. E.; Lepock, J. R.; Cabelli, D. E.;
Tainer, J. A.; Nick, H. S.; Silverman, D. N. Biochemistry 2000,
39, 7131–7137.
37. MacMillan-Crow, L. A.; Thompson, J. A. Arch Biochem Bio-
phys 1999, 366, 82–88.
38. Masip, L.; Veeravalli, K.; Georgiou, G. Antioxid Redox Signal
2006, 8, 753–762.
39. Hondorp, E. R.; Matthews, R. G. PLoS Biol 2004, 2, 1738–1753.
40. Rahman, I.; Biswas, S. K.; Jimenez, L. A.; Torres, M.; Forman,
H. J. Antioxid Redox Signal 2005, 7, 42–59.
41. Biswas, S.; Chida, A. S.; Rahman, I. Biochem Pharmacol 2006,
71, 551–564.
42. Wang, J.; Tekle, E.; Oubrahim, H.; Mieyal, J. J.; Stadtman, E. R.;
Chock, P. B. Proc Natl Acad Sci USA 2003, 100, 5103–5106.
43. Pan, S.; Berk, B. C. Circ Res 2007, 100, 213–219.
44. Adachi, T.; Weisbrod, R. M.; Pimentel, D. R.; Ying, J.; Sharov, V.
S.; Schoneich, C.; Cohen, R. A. Nat Med 2004, 10, 1200–1207.
45. Chen, C. L.; Zhang, L.; Yeh, A.; Chen, C. A.; Green-Church, K.
B.; Zweier, J. L.; Chen, Y. R. Biochemistry 2007, 46, 5754–5765.
46. Chen, Y. R.; Chen, C. L.; Pfeiffer, D. R.; Zweier, J. L. J Biol Chem
2007, 282, 32640–32654.
47. Schinina, M. E.; Carlini, P.; Polticelli, F.; Zappacosta, F.; Bossa,
F.; Calabrese, L. Eur J Biochem 1996, 237, 433–439.
48. Beer, S. M.; Taylor, E. R.; Brown, S. E.; Dahm, C. C.; Costa, N.
J.; Runswick, M. J.; Murphy, M. P. J Biol Chem 2004, 279,
47939–47951.
49. Hurd, T. R.; Costa, N. J.; Dahm, C. C.; Beer, S. M.; Brown, S. E.;
Filipovska, A.; Murphy, M. P. Antioxid Redox Signal 2005, 7,
999–1010.
50. Borgstahl, G. E.; Parge, H. E.; Hickey, M. J.; Johnson, M. J.;
Boissinot, M.; Hallewell, R. A.; Lepock, J. R.; Cabelli, D. E.;
Tainer, J. A. Biochemistry 1996, 35, 4287–4297.
51. Archambaud, C.; Nahori, M. A.; Pizarro-Cerda, J.; Cossart, P.;
Dussurget, O. J Biol Chem 2006, 281, 31812–31822.
52. Voisin, S.; Watson, D. C.; Tessier, L.; Ding, W.; Foote, S.; Bhatia,
S.; Kelly, J. F.; Young, N. M. Proteomics 2007, 7, 4338–4348.
53. McCord, J. M.; Fridovich, I. J Biol Chem 1969, 244, 6049–6055.
54. Bradford, M. Anal Biochem 1976, 72, 248–254.
55. Laemmli, U. K. Nature 1970, 227, 680–685.
56. Ginsberg, M. D.; Feliciello, A.; Jones, J. K.; Avvedimento, V. A.;
Gottesman, M. E. J Mol Biol 2003, 327, 885–897.
57. Maniatis, T.; Fritsch, E. F.; Sambrook, J. Molecular Cloning;
Cold Spring Harbor: New York, 1982.
58. Braman, J.; Papworth, C.; Greener, A. Methods Mol Biol 1996,
57, 31–44.
Reviewing Editor: Laurence Nafie
1226 Castellano et al.
Biopolymers
Copyright © 2022 FDOKUMEN




















