
Regulation of superoxide dismutase isoforms in resistant and susceptible strawberry cultivars...
10
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [Agriculture Canada] On: 16 September 2008 Access details: Access Details: [subscription number 793448910] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Archives Of Phytopathology And Plant Protection Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713454295 Regulation of superoxide dismutase isoforms in resistant and susceptible strawberry cultivars subjected to leaf spot disease Behrouz Ehsani-Moghaddam a ; Marie Thérèse Charles a ; Odile Carisse a ; Shahrokh Khanizadeh a a Agriculture and Agri-Food Canada, Horticultural Research Development Centre, Quebec, Canada First Published:October2008 To cite this Article Ehsani-Moghaddam, Behrouz, Charles, Marie Thérèse, Carisse, Odile and Khanizadeh, Shahrokh(2008)'Regulation of superoxide dismutase isoforms in resistant and susceptible strawberry cultivars subjected to leaf spot disease',Archives Of Phytopathology And Plant Protection,41:7,492 — 500 To link to this Article: DOI: 10.1080/03235400600833738 URL: http://dx.doi.org/10.1080/03235400600833738 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
consultant -
Category
Documents
-
view
3 -
download
0
Transcript of Regulation of superoxide dismutase isoforms in resistant and susceptible strawberry cultivars...

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Agriculture Canada]On: 16 September 2008Access details: Access Details: [subscription number 793448910]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Archives Of Phytopathology And Plant ProtectionPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713454295
Regulation of superoxide dismutase isoforms in resistant and susceptiblestrawberry cultivars subjected to leaf spot diseaseBehrouz Ehsani-Moghaddam a; Marie Thérèse Charles a; Odile Carisse a; Shahrokh Khanizadeh a
a Agriculture and Agri-Food Canada, Horticultural Research Development Centre, Quebec, Canada
First Published:October2008
To cite this Article Ehsani-Moghaddam, Behrouz, Charles, Marie Thérèse, Carisse, Odile and Khanizadeh, Shahrokh(2008)'Regulationof superoxide dismutase isoforms in resistant and susceptible strawberry cultivars subjected to leaf spot disease',Archives OfPhytopathology And Plant Protection,41:7,492 — 500
To link to this Article: DOI: 10.1080/03235400600833738
URL: http://dx.doi.org/10.1080/03235400600833738
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Regulation of superoxide dismutase isoforms in resistantand susceptible strawberry cultivars subjectedto leaf spot disease
BEHROUZ EHSANI-MOGHADDAM, MARIE THERESE CHARLES,
ODILE CARISSE, & SHAHROKH KHANIZADEH
Agriculture and Agri-Food Canada, Horticultural Research Development Centre, 430 Gouin Blvd.,
St-Jean-sur-Richelieu, Quebec, Canada J3B 3E6
(Received 19 May 2006)
AbstractMacroscopic symptoms were observed in two strawberry cultivars, with the degree of symptom intensityvarying depending on the susceptibility of the cultivars, i.e. resistant or susceptible. The symptomspresented as red spots and were observed 30 d following leaf tissue inoculation with the Mycosphaerellafragariae pathogen. A comparison of the superoxide dismutase isoform profiles obtained by gelelectrophoresis in all samples extracted from both resistant and susceptible cultivars indicated oneconstant sharp band, identified as Mn��SOD with a molecular mass of 19 kDa. The intensity of this bandwas higher in all samples derived from the resistant cultivar than in those from the susceptible cultivar.Another superoxide dismutase (SOD) isoform, identified as CuZn��SOD with a molecular mass of16 kDa, was detected in all soluble proteins derived from the resistant cultivar. This isoform was notobserved in the susceptible cultivar; however, following an incremental increase in the amount of loadedprotein, it was illuminated as a faint band in a sample collected 3 d after inoculation, indicating insufficientproduction of the CuZn��SOD isoform in the susceptible cultivar during an oxidative burst induced bythe M. fragaria pathogen. Several bands were also characterized in both cultivars containing Fe and Mn astheir co-factors (Fe, Mn��SOD). Unlike in the resistant cultivar, where the activity of Fe, Mn��SODisoforms gradually and regularly increased and reached its highest level on the third day after inoculation,the activity of the isoforms changed irregularly over 20 days of study in the susceptible cultivar.
Keywords: Antioxidant, Mycosphaerella fragariae, reactive oxygen species, superoxide dismutase,strawberry
Introduction
Leaf spot caused by Mycosphaerella fragariae is one of the most widespread foliar diseases in
strawberry crops in Canada (Fall 1951) and the United States (Fulton 1958) that can be
controlled with fungicide, owing mainly to cultivation of resistant cultivars (Maas 1984). In
previous studies, significant variation in the susceptibility of 25 cultivars to M. fragariae was
observed. According to these studies, the ‘‘Joliette’’ cultivar was considered resistant and
Correspondence: Behrouz Ehsani-Moghaddam, Agriculture and Agri-Food Canada, Horticultural Research Development Centre,
430 Gouin Blvd., St-Jean-sur-Richelieu, Quebec, Canada J3B 3E6. Tel: þ1-450-346-4494 (180). Fax: þ1-450-346-7740.
E-mail: [email protected]
Archives of Phytopathology and Plant Protection
October 2008; 41(7): 492 – 500
ISSN 0323-5408 print/ISSN 1477-2906 online ª 2008 Taylor & Francis
DOI: 10.1080/03235400600833738
Downloaded By: [Agriculture Canada] At: 14:23 16 September 2008

the Kent cultivar displayed marked susceptibility (Delhomez et al. 1995; Khanizadeh et al.
1996).
Some researchers (Hernandez et al. 1995; Foyer & Noctor 2000; Anderson 2002) reported
that interactions between host plants and fungal pathogens, and defence against invasion,
include a host resistance response, where the host plant produces reactive oxygen species
(ROS), such as superoxide- and hydroxyl-radicals and hydrogen peroxide. ROS are involved
in many aspects of a defence system because of their oxidative burst property (Mehdy 1994),
such as the oxidative cross-linking of cell walls to slow pathogen invasion (Otte & Barz 1996)
and the triggering of redox-sensitive signal cascades, which activates defence-related genes
(Vanacker et al. 1998).
An increase in ROS production by plant cells, induced by pathogen attack and other
stresses, results in an imbalance between oxidative stress and detoxification defence systems
(Kuzniak & Skłodowska 2004). There are several mechanisms by which plants protect
themselves from the toxic effects of ROS and alleviate the damage they cause, the most
effective of which is a complex antioxidant system. Among the various antioxidants,
superoxide dismutases (SODs) are the major scavengers of ROS. These antioxidants play a
crucial role in protecting cells against the toxic effects of superoxide radicals. SODs catalyze
the dismutation of O2.7 , generated in various cellular organelles (Fridovich 1986; Elstner
1991; Del Rio et al. 2002) by factors that include pathogen attack (Borden & Higgins 2002;
Barna et al. 2003), into H2O2 and O2.
SODs are ubiquitous metalloenzymes categorized, according to their metal co-factor, into
three groups: iron SOD (Fe��SOD), manganese SOD (Mn��SOD), and copper-zinc SOD
(CuZn��SOD). In a plant cell, various SOD isoforms are located in various subcellular
compartments. Typically, Fe��SOD is located in the chloroplast, Mn��SOD in the
mitochondrion and the peroxisome, and CuZn��SOD in the chloroplast, the cytosol, and
possibly the extracellular space (Alscher et al. 2002).
Evidence indicates that increased activity of SOD and its isoforms in plant tissue may be
associated with resistance to environmental stresses, such as drought and frost, and to
pathogen attack (Scandalios 1993). Conversely, evidence also indicates that susceptible
cultivars possess higher levels of superoxide production and lipid peroxidation and lower
levels of SOD activity than resistant cultivars (Wu & Tiedemann 2001). Our recent reports
indicated a positive relationship between SOD activity and the tolerance of strawberry plants
to M. fragariae attack in three kind of cultivars, i.e. susceptible, partially resistant and resistant
(Ehsani-Moghaddam et al. 2006). The present study was undertaken to better understand the
regulation of SOD isoenzymes induced in strawberry cultivars by the M. fragariae pathogen.
Materials and methods
Plant material and inoculation
Two strawberry cultivars (Fragariae X ananassa Duschesne), Joliette (resistant) and Kent
(susceptible), were propagated by stolon into a 300 pot containing mineral soil, peat moss
and sand in 1:1:1 volume. Plantlets were grown in greenhouses for two months at 258C with a
16-h photoperiod until they were inoculated.
The method for production of Mycosphaerella fragariae (Tul.) Lindau (Ramularia tulasnei
Sacc.) inoculums is explained in Ehsani-Moghaddam et al. (2006). A conidial suspension
was prepared from the cultures using 10 ml of sterile and distilled water and 0.01% Tween
80 in each plate. The final spore suspension was adjusted to 1.146105 spores/ml using
a hemacytometer. Twenty plants from each cultivar were transferred to a growth chamber and
Regulation of superoxide dismutase isoforms 493
Downloaded By: [Agriculture Canada] At: 14:23 16 September 2008

the spore suspension was sprayed onto both faces of their leaves to inoculate them. The inocu-
lated plants were kept in a growth chamber at 258C and 100% humidity with a 16-h photoperiod
for 48 h. Another 20 plants from each cultivar were kept in a separate growth chamber under
the same conditions and served as non-inoculated control plants. Forty-eight hours after
inoculation, the humidity was reduced to 85% in both chambers until the end of the study.
Enzyme extraction
Samples of expanded young leaves (0.5 g) from control and inoculated plants were collected
at 1, 2, 3, 4, 10, and 20 d after inoculation. The samples were immediately frozen in liquid
nitrogen and were stored at 7808C until they were needed. All procedures for enzyme
extraction were performed at 48C. For total protein extracts, frozen leaves were homogenized
with a mortar and pestle in 3 ml of ice-cold extraction buffer (pH 7.8) containing 75 mg/l
PVPP (polyvinylpolypyrrolidone), 12.1 g/l Tris, 68 g/l sucrose, 170 mg/l EDTA, 0.031 ml/l
Tween 80, and 800 mg/l sodium thioglycolate. The homogenate was centrifuged at 12 000 g
for 20 min and the supernatant was used for SOD (EC 1.15.1.1) assay and electrophoresis.
Protein concentration in the extracts was quantified by Bradford’s method (Bradford 1976)
using BSA as the standard protein.
Detection of SOD isoforms
To separate SOD isozymes, continuous native polyacrylamide gel electrophoresis (PAGE) was
performed with a PROTEAN III vertical electrophoresis unit (Bio-Rad) according to Davis
(1964), with the following modifications: non-denaturing gel was used throughout the study, the
stacking gel was omitted, and riboflavin was used instead of ammonium persulfate for photo-
polymerization of the gel. Soluble proteins from both cultivars were collected from non-
inoculated (control) and inoculated plants (1, 2, 3, 4, 10 and 20 d after inoculation), then loaded
on the gel and run at 120 V for 58 min and 200 V for 17 min. A modified method, based on the
standard procedure (Beauchamp & Fridovich 1971), was used for gel staining. The staining
solution was 7.88 g/l Tris-HCl buffer (pH¼ 8.0), containing 0.6 mg/l riboflavin, 20 mg/l
EDTA, 100 ml TEMED, and 200 mg/l NBT (nitroblue tetrazolium). After electrophoresis, the
gel was soaked in the staining solution for 20 min, then illuminated by two 15 W fluorescent
lamps until achromatic bands containing SOD were identified on uniformly dark purple gel.
Incubating the gels prior to staining in 3 mM KCN and 10 mM H2O2 each for 30 min, as
inhibitors of CuZn��SOD and of CuZn��SOD and Fe��SOD activity, respectively (Santos
et al. 1999), allowed various types of SOD isoforms to be recognized. Various isoforms
were detected, depending on the sensitivity of the inhibitor: the Mn��SOD isoform is resistant
to cyanide and to H2O2, Fe��SOD is resistant to cyanide but is inhibited by H2O2, and
CuZn��SOD is rendered inactive by both inhibitors. The molecular masses of isoforms were
calculated indirectly by the separation of the proteins in a set of gels with various
polyacrylamide concentrations and by comparison of the mobility of isoforms and non-
denatured protein markers (molecular weight marker kit Sigma, USA), according to the
manufacturer’s instruction.
Results
Macroscopic symptoms presenting as red spots on leaves were observed 30 d after leaf
inoculation with the M. fragariae pathogen. The symptoms were more intense in the ‘‘Kent’’
cultivar than in the ‘‘Joliette’’ cultivar (Figure 1).
494 B. Ehsani-Moghaddam et al.
Downloaded By: [Agriculture Canada] At: 14:23 16 September 2008

Native PAGE was repeated with five extracts and similar results were obtained, therefore
data from only one experiment are reported here. During 20 d of study, M. fragariae infection
caused observably distinctive regulation of SOD isoforms in both cultivars, as follows.
Joliette (resistant cultivar)
Depending on inhibitor sensitivity, when 1.5 mg of soluble protein was loaded in each lane,
and after separation of all samples, including controls, at least two clear and sharp SOD bands
became visible. One of the bands with the fastest electrophoretic mobility was identified as the
CuZn��SOD isoform with a molecular mass of 16 kDa (Figure 2). The second most active
band was identified as an Mn��SOD isoform with a molecular mass of 19 kDa. Several
isoforms were not completely inhibited by H2O2, therefore, they were considered to be SOD
isoforms containing both metal co-factors, i.e. Fe and Mn (Fe, Mn��SOD). Such isoforms
were detected in all Joliette extracts, except in those derived 20 d after inoculation, with
several isoforms beginning with an intensive and sharp band and a molecular mass of 22 kDa
and ending with an isoform with a molecular mass of 53 kDa. The 53 kDa isoform was
observed exclusively in a sample derived 3 d after inoculation.
The number of Fe, Mn��SOD isoforms and their intensity increased from the first day of
inoculation and reached their highest level of expression on the third day after inoculation,
slowly declining thereafter (Figure 2A). While the Fe, Mn��SOD bands in the control plants
appeared as four faint bands, in samples derived from the inoculated plants 3 d after
inoculation, the bands were observed as five thick bands, three of which were condensed and
contained more than one isoform (arrow heads in Figure 2A).
Kent (susceptible cultivar)
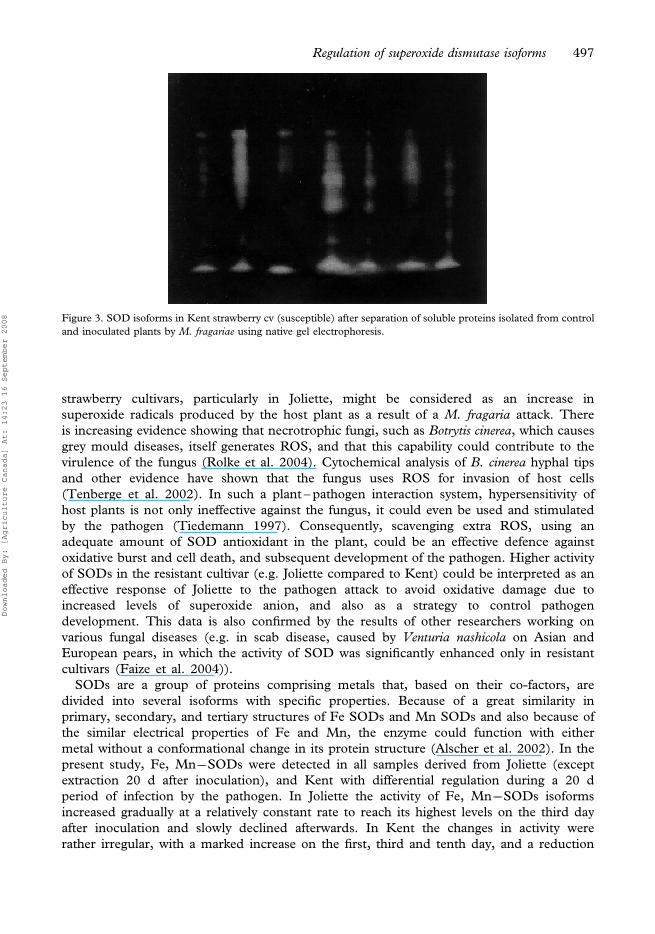
PAGE of 1.5 mg of soluble protein showed a few isoforms that were barely detectable (data
not shown). To improve detection, the amount of soluble proteins loaded on the gel was
Figure 1. Leaf spot caused by M. fragariae on two cv of strawberry plants: Kent (susceptible), and Joliette (resistant)
one month after inoculation. Inoculation of plants was accomplished in a growth chamber by spraying the spore
suspension (1.146105 spores/ml) on both faces of leaf and maintained at 258C with a 16-h photoperiod and 100%
humidity for 48 h. Lesions are small, circular with light centers and purplish margins.
Regulation of superoxide dismutase isoforms 495
Downloaded By: [Agriculture Canada] At: 14:23 16 September 2008

increased to 2.2 mg. Using this concentration, the highest number of isoforms was observed in
samples derived 3 d after inoculation. In all samples, one sharp band, identified as an
Mn��SOD isoform with a 19 kDa molecular mass (Figure 3) was observed. Several bands,
each containing more than one isoform, were also detected in all samples and were identified
as Fe, Mn��SODs. Their molecular masses ranged from 22 kDa to 53 kDa. Unlike in Joliette,
the 53 kDa isoform was observed in all samples derived from Kent. The banding patterns and
intensity of Fe, Mn��SOD isoforms were not expressed on a regular basis over the 20 d of
study. The CuZn��SOD isoform was detected only in samples derived 3 d after inoculation.
This isoform was expressed as a faint band (Figure 3).
Discussion
The increase of SOD activity (Figure 2) was concomitant with the production of germ tubes
by conidia and penetration through stomata to leaf tissue (during 12 – 96 h from inoculation)
(Carisse et al. 2000). As SODs catalyze the dismutation of superoxide radicals produced in
different cellular compartments (Fridovich 1986), the increase in SOD activity in inoculated
Figure 2. (A) SOD isoforms in strawberry (Joliette cv, resistant) after separation of soluble proteins isolated from
control and inoculated plants by M. fragariae using native gel electrophoresis, arrow heads show three condensed
Fe, Mn��SOD, each contains a group of isoforms; (B) incubation of the gel prior to staining into 3 mM KCN for
30 min for inhibition of CuZn��SOD enzyme; and (C) incubation of gel prior to staining into 10 mM H2O2 for
30 min, as inhibitor of CuZn��SOD and Fe��SOD enzymes.
496 B. Ehsani-Moghaddam et al.
Downloaded By: [Agriculture Canada] At: 14:23 16 September 2008
strawberry cultivars, particularly in Joliette, might be considered as an increase in
superoxide radicals produced by the host plant as a result of a M. fragaria attack. There
is increasing evidence showing that necrotrophic fungi, such as Botrytis cinerea, which causes
grey mould diseases, itself generates ROS, and that this capability could contribute to the
virulence of the fungus (Rolke et al. 2004). Cytochemical analysis of B. cinerea hyphal tips
and other evidence have shown that the fungus uses ROS for invasion of host cells
(Tenberge et al. 2002). In such a plant – pathogen interaction system, hypersensitivity of
host plants is not only ineffective against the fungus, it could even be used and stimulated
by the pathogen (Tiedemann 1997). Consequently, scavenging extra ROS, using an
adequate amount of SOD antioxidant in the plant, could be an effective defence against
oxidative burst and cell death, and subsequent development of the pathogen. Higher activity
of SODs in the resistant cultivar (e.g. Joliette compared to Kent) could be interpreted as an
effective response of Joliette to the pathogen attack to avoid oxidative damage due to
increased levels of superoxide anion, and also as a strategy to control pathogen
development. This data is also confirmed by the results of other researchers working on
various fungal diseases (e.g. in scab disease, caused by Venturia nashicola on Asian and
European pears, in which the activity of SOD was significantly enhanced only in resistant
cultivars (Faize et al. 2004)).
SODs are a group of proteins comprising metals that, based on their co-factors, are
divided into several isoforms with specific properties. Because of a great similarity in
primary, secondary, and tertiary structures of Fe SODs and Mn SODs and also because of
the similar electrical properties of Fe and Mn, the enzyme could function with either
metal without a conformational change in its protein structure (Alscher et al. 2002). In the
present study, Fe, Mn��SODs were detected in all samples derived from Joliette (except
extraction 20 d after inoculation), and Kent with differential regulation during a 20 d
period of infection by the pathogen. In Joliette the activity of Fe, Mn��SODs isoforms
increased gradually at a relatively constant rate to reach its highest levels on the third day
after inoculation and slowly declined afterwards. In Kent the changes in activity were
rather irregular, with a marked increase on the first, third and tenth day, and a reduction
Figure 3. SOD isoforms in Kent strawberry cv (susceptible) after separation of soluble proteins isolated from control
and inoculated plants by M. fragariae using native gel electrophoresis.
Regulation of superoxide dismutase isoforms 497
Downloaded By: [Agriculture Canada] At: 14:23 16 September 2008

on the second, fourth and twentieth day after inoculation. The increase of Fe, Mn��SODs
activity in both strawberry cultivars, particularly in the resistant one, might be considered
as a part of the initial activation of the antioxidant enzymatic system against the increasing
superoxide anion level generated during a time frame of pathogen establishment.
Comparison of the SOD isoform profiles in all samples derived from both resistant and
susceptible strawberry cultivars revealed a constant and sharp band identified as an
Mn��SOD with a molecular mass of 19 kDa. In all samples derived from Joliette the
intensity of bands was greater than that of Kent. Unlike in Kent, in Joliette the intensity of
the Mn��SOD band was not the same from one sample to another; it increased in the
samples derived from 2, 3, 10 and 20 d after inoculation (Figure 2A). The presence of
only one Mn��SOD band in both cultivars, rather than several bands, as observed for
Fe, Mn��SOD, may indicate either the existence of only one Mn��SOD isoform in the
defence system of the plant or the existence of several isoforms but the expression of
only one of them in response to introduction of the pathogen. It has been suggested that
differences in responses between the Mn��SODs and Fe��SODs in plants can be
correlated with their disparate subcellular locations and the action sites of various
oxidative stresses. The stresses that did not affect Mn��SOD activity may all have their
action sites in the chloroplast (Alscher et al. 2002), or outside the mitochondria, as
suggested for pathogenic interactions (Moore et al. 2002). Following this assumption, it is
possible that the action site of the M. fragaria pathogen may be located in the chloroplast,
or in a part of the invaded cell that is ignored by some Mn��SOD isoforms and not
others.
Assessment of the SOD isoform profiles also revealed a constant band as a CuZn��SOD
with a molecular mass of 16 kDa that was detected in all soluble proteins extracted
from Joliette. This isoform was not detectable in Kent; however, when the amount of
loaded protein was increased from 1.5 to 2.2 mg, the isoform was illuminated as a
faint band in samples derived 3 d after inoculation, indicating insufficient production of
the CuZn��SOD isoform in the Kent cultivar during the oxidative burst induced by
the pathogen. Unlike Fe��SODs and Mn��SODs, CuZn��SODs have been found mostly
in eukaryotes and throughout the plant cell (Alscher et al. 2002). The importance
of CuZn��SODs in plants and their protective actions against various oxidative
stresses have been documented by several researchers. The transfer of tomato chloroplastic
and cytosolic CuZn��SOD genes into potato plants improved tolerance levels against
the oxidative stress caused by methyl viologen (Perl et al. 1993). The data presented
here are, to the best of our knowledge, the first concerning the involvement of
a CuZn��SOD isoform as a distinctive protein that could be linked to resistance against
a fungal disease in plants. The presence of this isoform in Joliette, as a resistant
cultivar, in the non-inoculated plant and during infection by the pathogen, as well as its
absence in Kent, as a susceptible cultivar, in the non-inoculated plant and with
insubstantial activity during infection, indicates the importance of this SOD in the
defence system of strawberry plants against oxidative bursts induced by the M. fragaria
pathogen.
Our data suggest that the resistance of the Joliette cultivar to red spot disease infection may
be related to the suppression of oxidative bursts and cell death induced by the M. fragaria
pathogen in a timely manner and in a such a way that several Mn, Fe��SODs and one
Mn��SOD are regulated to reach maximum activity when most needed. The involvement of a
CuZn��SOD in the defence system of the resistant cultivar, which was not detected in the
interaction between the susceptible cultivar and the pathogen, deserves further consideration.
In the interaction between Kent as a susceptible cultivar and M. fragaria, rapid necrosis and
498 B. Ehsani-Moghaddam et al.
Downloaded By: [Agriculture Canada] At: 14:23 16 September 2008

cell death leading to the expression of severe symptoms are perhaps the consequence of
unsuccessful regulation of SOD isoforms to adequately scavenge superoxide anions generated
during pathogen attack.
References
Alscher RG, Erturk N, Heath LS. 2002. Role of superoxide dismutases (SODs) in controlling oxidative stress in
plants. J Exp Bot 372:1331 – 41.
Anderson JA. 2002. Catalase activity, hydrogen peroxide content and thermotolerance of pepper leaves. Scientia Hor
95:277 – 84.
Barna B, Fodor J, Pogany M, Kiraly Z. 2003. Role of reactive oxygen species and antioxidants in plant disease
resistance. Pest Manag Sci 59:459 – 64.
Beauchamp C, Fridovich I. 1971. Superoxide dismutase: improved assays and an assay applicable to acrylamide gels.
Anal Biochem 44:276 – 87.
Borden S, Higgins VJ. 2002. Hydrogen peroxide plays a critical role in the defence response of tomato to Cladosporium
fulvum. Physiol Mol Plant P 61:227 – 36.
Bradford M. 1976. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the
principle of dye binding. Anal Biochem 72:248 – 54.
Carisse O, Bourgeois G, Duthie JA. 2000. Influence of temperature and leaf wetness duration on infection of
strawberry leaves by Mycosphaerella fragariae. Phytopathology 90:1120 – 5.
Davis BJ. 1964. Disc electrophoresis. Method and application to human serum proteins. Ann NY Acad Sci 121:
404 – 27.
Delhomez N, Carisse O, Lareau M, Khanizadeh S. 1995. Susceptibility of strawberry cv. and advanced selection to
leaf spot caused by Mycosphaerella fragariae. HortScience 30:592 – 5.
Del Rio LA, Corpas FJ, Sandalio LM, Palma JM, Gomez M, Barroso JB. 2002. Reactive oxygen species, antioxidant
systems and nitric oxide in peroxisomes. Exp Bot 53:1 – 31.
Ehsani-Moghaddam B, Charles MT, Carisse O, Khanizadeh S. 2006. Superoxide dismutase responses of strawberry
cultivars to infection by Mycosphaerella fragariae. J Plant Physiol 163: 147 – 53.
Elstner EF. 1991. Mechanisms of oxygen activation in different compartments of plant cells. In: Pell EJ, Steffen KL,
editors. Active oxygen species, oxidative stress and plant metabolism. Rockville, MD: The American Society of
Plant Physiologists. pp 13 – 25.
Faize M, Faize L, Ishizaka M, Ishii H. 2004. Expression of potential defense responses of Asian and European pears
to infection with Venturia nashicola. Physiol Mol Plant P 64:319 – 30.
Fall J. 1951. Studies on fungus parasites of strawberry leaves in ontario. Can J Bot 29:301 – 15.
Foyer CH, Noctor G. 2000. Oxygen processing in photosynthesis: regulation and signaling. New Phytol 146:
359 – 88.
Fridovich I. 1986. Superoxide dismutases. Adv Enzymol Relat Areas Mol Biol 58:61 – 97.
Fulton RH. 1958. Studies on strawberry leaf spot in Michigan. Michigan Expt Sta Quart Bull 40:581 – 8.
Hernandez JA, Olmos E, Corpas FJ, Sevilla F, del Rio LA. 1995. Salt induced oxidative stress in chloroplast of pea
plants. Plant Sci 105:151 – 67.
Khanizadeh S, Buszard D, Carrise O, Thibodeau PO. 1996. ‘Joliette’ strawberry. HortScience 31:1036 – 7.
Kuzniak E, Skłodowska M. 2004. The effect of Botrytis cinerea infection on the antioxidant profile of mitochondria
from tomato leaves. J Exp Bot 55:605 – 12.
Maas JL. 1984. Compendium of strawberry diseases. St. Paul, MN: APS Press.
Mehdy MC. 1994. Active oxygen species in plant defence against pathogens. Plant Physiol 105:467 – 72.
Moore S, de Vries OMH, Tudzynski P. 2002. The major Cu, Zn SOD of the phytopathogen Claviceps purpurea is not
essential for pathogenicity. Mol Plant Pathol 3:9 – 22.
Otte O, Barz W. 1996. The elicitor-induced oxidative burst in cultured chickpea cells drives the rapid insolubilization
of two cell wall structural proteins. Planta 200:238 – 46.
Perl A, Perl-Treves R, Galili S, Aviv D, Shalgi E, Malkin S, Galun E. 1993. Enhanced oxidative stress defence in
transgenic potato expressing tomato Cu,Zn superoxide dismutases. Theor Appl Genet 85:568 – 76.
Rolke Y, Liu S, Quidde T, Williamson B, Schouten A, Weltring KM, Siewers V, Tenberge KB, Tudzynski B,
Tudzynski P. 2004. Functional analysis of H2O2-generating systems in Botrytis cinerea: the major Cu-Zn-
superoxide dismutase (BCSOD1) contributes to virulence on French bean, whereas a glucose oxidase
(BCGOD1) is dispensable. Mol Plant Pathol 5:17 – 27.
Santos I, Almedia J, Salema R. 1999. The influence of UV-B radiation on the superoxide dismutase of maize, potato,
sorghum, and wheat leaves. Can J Bot 77:70 – 6.
Regulation of superoxide dismutase isoforms 499
Downloaded By: [Agriculture Canada] At: 14:23 16 September 2008

Scandalios JG. 1993. Oxygen stress and superoxide dismutase. Plant Physiol 101:7 – 12.
Tenberge KB, Beckedorf M, Hoppe B, Schouten A, Solf M, von den Driesch M. 2002. In situ localization of AOS in
host-pathogen interactions. Microsc Microanal 8:250 – 1.
Tiedemann AV. 1997. Evidence for a primary role of active oxygen species in induction of host cell death during
infection of bean leaves with Botrytis cinerea. Physiol Mol Plant P 50:151 – 66.
Vanacker H, Carver TLW, Foyer C. 1998. Pathogen-induced changes in the antioxidant status of the apoplast in
barely leaves. Plant Phsiol 117:1103 – 14.
Wu YX, Tiedemann A. 2001. Evidence for oxidative stress involved in physiological leaf spot formation in winter and
spring barley. Phytopathology 92:145 – 55.
500 B. Ehsani-Moghaddam et al.
Downloaded By: [Agriculture Canada] At: 14:23 16 September 2008




















