
Immunolocalization of a tachykinin-receptor-like protein in the central nervous system of Locusta...
12
Immunolocalization of a Tachykinin-Receptor-Like Protein in the Central Nervous System of Locusta migratoria migratorioides and Neobellieria bullata DIRK VEELAERT, 1 * HENDRICA B. OONK, 2 GREET VANDEN EYNDE, 1 HERBERT TORFS, 1 ROBBERT H. MELOEN, 2 LILIANE SCHOOFS, 1 MARC PARMENTIER, 3 ARNOLD DE LOOF, 1 AND JOZEF VANDEN BROECK 1 1 Laboratory for Developmental Physiology and Molecular Biology, Zoological Institute, B-3000 Leuven, Belgium 2 ID-DLO, Department of Molecular Recognition, 8200 AB Lelystad, The Netherlands 3 IRIBHN ULB, B-1070 Brussels, Belgium ABSTRACT Antisera raised against two distinct peptide regions of the Drosophila neurokinin-like receptor NKD were used to immunolocalize tachykinin-receptor-like proteins in the central nervous system of two insect species: the African migratory locust, Locusta migratoria, and the gray fleshfly, Neobellieria bullata. The resulting immunopositive staining patterns were identical for both antisera. Moreover, a very similar distribution of the immunoreactive material was observed in fleshflies and locusts. Immunoreactivity was found in nerve terminals of the retrocerebral complex, suggesting a presynaptic localization of the receptor in this part of the brain. Cell bodies were stained in the subesophageal ganglion: an anterior group of four larger cells and a posterior group of about 20 cells. These cells have axons projecting into the contralateral nervus corporis allati (NCA) II, bypassing the corpus allatum and projecting through the NCA I into the storage part of the corpus cardiacum. In the glandular part of the corpus cardiacum, the glandular adipokinetic hormone-producing cells did not show any immunopositive staining. In the locust, additional immunopositive staining was observed in internolaterally located neurons of the tritocerebrum and in important integrative parts of the neuropil such as the central body and the mushroom bodies. J. Comp. Neurol. 407:415–426, 1999. r 1999 Wiley-Liss, Inc. Indexing terms: central body; fly; insect; locust; corpus cardiacum; neuropeptide; hormone Tachykinins or neurokinins are members of a brain/gut peptide family in mammals. These peptides and their respective receptors have been implicated in a number of mammalian motor and sensory functions (Helke et al., 1990; Maggi, 1995). There is evidence for three main types of mammalian tachykinin receptors, with species variants and subtypes among each of these three receptor types (Watling et al., 1994). Compared with the high number of different peptides that have been identified in insects (Schoofs et al., 1997), very few studies on insect peptide receptors have been reported. Nevertheless, a large number of physiological effects elicited by peptides appear to be mediated by the action of G-proteins, indicating that, as in mammals, many small insect peptides act through G-protein-coupled receptors (Vanden Broeck, 1996). Complementary DNAs from Drosophila melanogaster and Stomoxys calcitrans, which code for proteins with structural and biochemical properties similar to mammalian tachykinin receptors, have been cloned (Li et al., 1991; Monnier et al., 1992; Grant sponsor: Interuniversity Poles of Attraction Programme, Belgian State, Prime Minister’s Office—Federal Office for Scientific, Technical and Cultural Affairs; Grant numbers: E.C. BIO4-CT96–0699 and IUAP/PAI-IV P4/30. *Correspondence to: Dirk Veelaert, Laboratory for Developmental Physi- ology and Molecular Biology, Zoological Institute, Naamsestraat 59, B-3000 Leuven, Belgium. E-mail: dirk.veelaert @bio.kuleuven.ac.be Received 30 July 1998; Revised 18 November 1998; Accepted 29 Decem- ber 1998 THE JOURNAL OF COMPARATIVE NEUROLOGY 407:415–426 (1999) r 1999 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Immunolocalization of a tachykinin-receptor-like protein in the central nervous system of Locusta...

Immunolocalization of aTachykinin-Receptor-Like Protein in the
Central Nervous System of Locustamigratoria migratorioides and
Neobellieria bullata
DIRK VEELAERT,1* HENDRICA B. OONK,2 GREET VANDEN EYNDE,1 HERBERT
TORFS,1 ROBBERT H. MELOEN,2 LILIANE SCHOOFS,1 MARC PARMENTIER,3
ARNOLD DE LOOF,1 AND JOZEF VANDEN BROECK1
1Laboratory for Developmental Physiology and Molecular Biology, Zoological Institute,B-3000 Leuven, Belgium
2ID-DLO, Department of Molecular Recognition, 8200 AB Lelystad, The Netherlands3IRIBHN ULB, B-1070 Brussels, Belgium
ABSTRACTAntisera raised against two distinct peptide regions of the Drosophila neurokinin-like
receptor NKD were used to immunolocalize tachykinin-receptor-like proteins in the centralnervous system of two insect species: the African migratory locust, Locusta migratoria, andthe gray fleshfly, Neobellieria bullata. The resulting immunopositive staining patterns wereidentical for both antisera. Moreover, a very similar distribution of the immunoreactivematerial was observed in fleshflies and locusts. Immunoreactivity was found in nerveterminals of the retrocerebral complex, suggesting a presynaptic localization of the receptor inthis part of the brain. Cell bodies were stained in the subesophageal ganglion: an anteriorgroup of four larger cells and a posterior group of about 20 cells. These cells have axonsprojecting into the contralateral nervus corporis allati (NCA) II, bypassing the corpus allatumand projecting through the NCA I into the storage part of the corpus cardiacum. In theglandular part of the corpus cardiacum, the glandular adipokinetic hormone-producing cellsdid not show any immunopositive staining. In the locust, additional immunopositive stainingwas observed in internolaterally located neurons of the tritocerebrum and in importantintegrative parts of the neuropil such as the central body and the mushroom bodies. J. Comp.Neurol. 407:415–426, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: central body; fly; insect; locust; corpus cardiacum; neuropeptide; hormone
Tachykinins or neurokinins are members of a brain/gutpeptide family in mammals. These peptides and theirrespective receptors have been implicated in a number ofmammalian motor and sensory functions (Helke et al.,1990; Maggi, 1995). There is evidence for three main typesof mammalian tachykinin receptors, with species variantsand subtypes among each of these three receptor types(Watling et al., 1994).
Compared with the high number of different peptidesthat have been identified in insects (Schoofs et al., 1997),very few studies on insect peptide receptors have beenreported. Nevertheless, a large number of physiologicaleffects elicited by peptides appear to be mediated by theaction of G-proteins, indicating that, as in mammals,many small insect peptides act through G-protein-coupled
receptors (Vanden Broeck, 1996). Complementary DNAsfrom Drosophila melanogaster and Stomoxys calcitrans,which code for proteins with structural and biochemicalproperties similar to mammalian tachykinin receptors,have been cloned (Li et al., 1991; Monnier et al., 1992;
Grant sponsor: Interuniversity Poles of Attraction Programme, BelgianState, Prime Minister’s Office—Federal Office for Scientific, Technical andCultural Affairs; Grant numbers: E.C. BIO4-CT96–0699 and IUAP/PAI-IVP4/30.
*Correspondence to: Dirk Veelaert, Laboratory for Developmental Physi-ology and Molecular Biology, Zoological Institute, Naamsestraat 59, B-3000Leuven, Belgium. E-mail: dirk.veelaert @bio.kuleuven.ac.be
Received 30 July 1998; Revised 18 November 1998; Accepted 29 Decem-ber 1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 407:415–426 (1999)
r 1999 WILEY-LISS, INC.

Guerrero et al., 1997). One of these putative receptorproteins, NKD, also has been shown to be activated bylocustatachykinin II (Monnier et al., 1992).
Five isoforms of locustatachykinins (Lom-TKs) havebeen isolated from brains of L. migratoria by screeningchromatographic fractions in a cockroach hindgut contrac-tion assay (Schoofs et al., 1990a,b, 1993). These neuropep-tides also have been shown to stimulate contractions of thelocust foregut and oviduct in vitro (Schoofs et al., 1993).Lom-TK I has been reported to induce the release ofadipokinetic hormone (AKH) from the glandular cells ofthe corpus cardiacum (CC) of the locust, L. migratoria, in amicromolar concentration range (Nassel et al., 1995b).
Other family members have been purified from thecockroach Leucophaea maderae (Muren and Nassel1996a,b, 1997a,b), the blowfly Calliphora vomitoria(Lundquist et al., 1994a), and the mosquito Culex sali-narius (Meola et al., 1998). To date, 19 members comprisethe insect tachykinin-related peptide family, which ischaracterized by the C-terminal pentapeptide sequenceFX1GX2Ramide (X1 most commonly is Y and X2 usually isV). Additional locustatachykinin-like peptides have beenidentified from other invertebrate species such as crusta-ceans (Christie et al., 1997; Nieto et al., 1998), molluscs(Fujisawa et al., 1994), and echiuroid worms (Ikeda et al.,1993).
Furthermore, immunocytochemical mapping of Lom-TKs in the locust and the blowfly Calliphora vomitoria hassuggested that these peptides may function as neurotrans-mitters or neuromodulators in interneurons of the centralnervous system and as regulators in the midgut (Lundquistet al., 1993, 1994b; Nassel, 1993a,b; Muren et al., 1995;Nassel et al., 1995a).
Although many neuropeptides have been immunolocal-ized in insects, the same is not true for their receptors. Inthis article, we describe the production of six antisera thatare highly specific to two different parts of the tachykinin-like receptor NKD. All antisera were used in a immunocy-tochemical study to localize this tachykinin-like receptorin the central nervous system of the African migratorylocust, Locusta migratoria, and the gray fleshfly, Neobellie-ria bullata.
MATERIALS AND METHODS
Materials
Acetonitrile (ACN) was high-performance liquid chroma-tography (HPLC) S-gradient grade; N-methylpyrrolidone(NMP), diisopropylethylamine (DIEA), dimethylformamide(DMF), trifluoroacetic acid (TFA), and piperidine werepeptide synthesis grade; and all were obtained fromBiosolve (Valkenswaard, Netherlands). N-hydroxybenzo-triazole (HOBt) and 2-(1H-benzotriazol-1-yl-1,1,3,3-tetra-methyluronium) hexafluorophosphate (HBTU) were ob-tained from Richelieu Biotechnologies Inc. (Hamon,Canada). Benzotriazol-1-yl-oxy-tris-pyrrolidino-phospho-nium hexafluorophosphate (PyBOP) was obtained fromNovabiochem (Laufelfingen, Switzerland). Thioanisole(TA), ethanedithiol (EDT), dimethylsulfoxide (DMSO), pen-tane, and dimethylaminopyridine (DMAP) were pro-analysis grade and were obtained from Merck (Darmstad,Germany). Diethylether was purified over a column ofactivated basic aluminumoxide before use. Amino acidderivatives and resins were obtained from BachemFeinchemicalien AG (Bubendorf, Switzerland).
Animals
Locusta migratoria was raised according toAshby (1972).Neobellieria bullata was reared under laboratory condi-tions as described by Huybrechts and De Loof (1981).Brains and ganglia from CO2-anesthetized adults weredissected in locust Ringer’s solution (NaCl 168 mM, KCl6.4 mM, NaHPO4 1.2 mM, NaHCO3 3.0 mM, MgCl2 3.6mM, CaCl2 2.1 mM; pH 7). Guinea pigs were housed atInstitute For Animal Science and Health [ID-DLO], TheNetherlands and kept according to the general manage-ment rules.
Multiple peptide synthesis
Synthesized peptide fragments for the NKD receptor(Monnier et al., 1992) were *CDRYIAIVDPLKRRTSR-RKV# and *CRSIGENTDRQMESMKSKRK#. Amino acidsare indicated by the single letter code. The asteriskindicates acetylation of the N-terminus, and the numbersign indicates amidation of the C-terminus. The columnswere drained automatically after each step by vacuum.The coupling cycle was based on 9-fluorenylmethyloxycar-bonyl (Fmoc)/HBTU chemistry (Rink, 1987; Fields et al.,1991) by using double coupling steps of 40 minutes. Afterthe coupling of the last amino acid, the Fmoc group wasremoved by using 30% (v:v) piperidine:NMP. The peptideswere washed, acetylated by using NMP:acetic anhydride:DIEA (10:1:0.2, v:v:v) for 30 minutes, washed again, anddried. Peptides were deprotected and cleaved in 2 hours byusing 1.5 ml of a mixture of TFA:phenol:TA:water:EDT(10:0.75:0.5:0.5:0.25, v:w:v:v:v). The cleavage mixture wasfiltered, the resin was washed with 0.5 ml TFA, and thepeptide was precipitated by adding 13 ml pentane:diethylether 1:1. After centrifugation, the precipitate wasextracted again with pentane:diethylether. The precipi-tate was dried, dissolved in ACN:water 1:1, and lyophi-lized. This procedure produces, depending on molecularweight, 25–70 mg of peptide.
Peptide analysis: HPLC, mass analysis,amino acid analysis
For analytical HPLC, an LC-MS (electrospray) systemwas used, consisting of two Waters 510 pumps, a Waters680 gradient controller, a Waters WISP 712 autoinjector,and a Waters 991 photodiode array detector (WatersAssociates, Milford, MA). Results of mass analysis wereobtained on a Micromass Quatro IIsq mass spectrometerthat was used in positive ion mode (Micromass, Wythen-shawe, UK). Mass of the largest HPLC peak was deter-mined. Products were analyzed in a linear gradient from10% ACN:water with 0.05% (v:v) TFA to 70% (v:v) ACN:water with 0.05% (v:v) TFA in 30 minutes on a WatersDelta Pak C18–100A (3.9 3 150 mm, 5:m) column at 1ml/minute. Amino acid analysis was performed with aWaters Pico-Tag system after hydrolysis in a Pico-Tagworkstation by using 6 N HCl at 150°C for 1 hour andderivatization with phenylisothiocyanate.
Antisera
Production of the locustamyotropin (Lom-MT) antise-rum has been described previously (Schoofs et al., 1992).Peptides were conjugated to keyhole limpet hemocyanin(KLH) with the aid of maleinimidobenzoyl-N-hydroxysuc-cinimide ester for the first immunization and glutardialde-hyde for subsequent immunizations (Meloen et al., 1994).
416 D. VEELAERT ET AL.

The loading was calculated from comparative amino acidanalysis of the conjugate and from the separate peptidesand carrier proteins and was at least 200 nmol peptide/mgKLH (approximately 0.5 peptide/mg KLH). Adjuvantswere complete Freund’s adjuvant (CFA) for the first immu-nization and incomplete Freund’s adjuvant (IFA) for subse-quent immunizations. CFA or IFA was mixed 1:1 with thepeptide solution to a stable emulsion (Meloen et al., 1994).Each guinea pig received 0.5 mg of peptide, conjugated andadjuvanted as described above. The vaccine was adminis-tered into four sites, 23 0.1 ml intramuscularly 1 23 0.1ml subcutaneously. Vaccination was repeated after 4, 8,and 12 weeks. Serum samples were taken at day 0 and at2, 5, 7, 9, and 14 weeks postvaccination by heart punction(preimmune serum) or orbita punction.
Evaluation of antisera
Peptide enzyme-linked immunosorbent assays (ELISAs)were performed in 96-well plates (Greiner, Alphen AanDen Rijn, The Netherlands; flat bottom). Each peptide wascoated in a concentration of 1 µg/well by using 50 mMNaHCO3/Na2CO3 buffer, pH 9.6. Serum dilutions weretested with rabbit anti-guinea pig coupled to horse radishperoxidase (Dako, Glostrup, Denmark) as second a anti-body and tetramethylbenzidine (Aldrich 86033–6, Aldrich,Millwaukee, WI) as a substrate. Absorbance was mea-sured at 450 nm in a Multiscan Plus ELISA reader(Titertek, Flow Laboratories, McLean, VA.
Immunocytochemistry
Dissected tissues were fixed in Bouin-Hollande’s 10%sublimate, dehydrated in an ethanol series, cleared inxylene, and embedded in Paraplast (Sherwood MedicalCo., St. Louis, MO). Serial sections were made at 4 µmthickness. Immunohistochemical staining was accom-plished by means of (1) the alkaline phosphatase–Fast-Red and (2) the peroxidase–anti-peroxidase–diaminobenzi-dine methods. Optimal dilutions were 1:400 for alltachykinin receptor antisera and 1:2,000 for Lom-MT.Specificity was controlled (1) by immunoadsorption of theserum by using the antigen (1 µl of pure serum with 1 µgsynthetic peptide) before application, (2) by preimmuneserum, (3) by anti-KLH carrier protein antiserum, and (4)by the processing of a series of immunocytochemicalstainings in which the various steps, one by one, wereomitted from the regular staining sequence.
For immunoblot analysis, brains of Locusta migratoriaand Neobellieria bullata were taken from adults. Afterextensive washing in Ringer’s solution, dissected tissueswere homogenized by using a potter, and protein concentra-tion was determined (Bradford, 1976). All steps wereperformed at 4°C. An equal volume of sodium dodecylsulfate (SDS) sample buffer was added to the samples,each containing 50 µg protein, and samples were boiled for5 minutes. Electrophoretic separation of the proteins wasperformed in a 5–15% SDS–polyacrylamide gradient gel(Laemmli, 1970). The gel was subjected to semidry blot-ting, as described by Peferoen (1988), and proteins weretransferred to nitrocellulose paper. Electroblots were ob-tained after 75 minutes under constant current conditions(1 mA/cm2). After transfer, immunostaining was per-formed as described above. The stained blot was washedwith distilled water and air dried. The part of the transfermembrane with the molecular marker proteins was stainedseparately with Amido Black.
RESULTS
Peptide synthesis
Synthetic peptides were purified by HPLC, and theiridentities were confirmed by mass analysis and amino acidanalysis.
Specificity of the antisera
The produced antisera and their antigens are shown inTable 1. ELISA was used to determine antibody formationagainst the peptides (Table 1). Titers against the first
Fig. 1. Consecutive sections through cells in the subesophagealganglion of the fly Neobellieria bullata. a: Section stained with theanti-NKD antiserum. b: Section stained with the immunosorbedantiserum (1 µl/2 µg). Scale bar 5 25 µm.
TABLE 1. Antisera Produced in Guinea Pigs Against the NKD Receptor1
Antiserum Antigen Titer
F3a *CDRYIAIVDPLKRRTSRRKV# 1.58F3b *CDRYIAIVDPLKRRTSRRKV# 2.1F3c *CDRYIAIVDPLKRRTSRRKV# 1.98G4a *CRSIGENTDRQMESMKSKRK# 4.34G4b *CRSIGENTDRQMESMKSKRK# 3.84G4c *CRSIGENTDRQMESMKSKRK# 3.83
1Peptide synthesis, vaccine preparation, immunization protocol, and enzyme-linkedimmunosorbent assays were performed as described in Materials and Methods. Titer isgiven as the logaritm of the dilution of antiserum (last bleed), producing an absorption ofthree times the background level compared with preimmune serum.
DISTRIBUTION OF INSECT TACHYKININ RECEPTOR 417
peptide *CDRYIAIVDPLKRRTSRRKV# were much lowerthan titers against the second peptide *CRSIGENT-DRQMESMKSKRK#, indicating weak immunogenicityof the former peptide. For the primary antisera, afinal dilution of 1:400 appeared to be appropriatefor an optimal immunoreaction on the tissue sections.In Locusta migratoria and Neobellieria bullata, no stainingwas observed with the preimmune sera. In immuno-adsorption controls performed on consecutive sections ofthe brains and thoracic ganglia of Locusta migratoriaand Neobellieria bullata, staining was abolished withantiserum immunoabsorbed on pure synthetic peptide(Fig. 1). Antisera raised against the carrier proteinwere immunonegative on all tissues tested.
Immunostaining in the central brain complexof Locusta migratoria
All anti-NKD antisera showed virtually identical pat-terns of immunostaining in the brains of Locusta migrato-ria and Neobellieria bullata. Between males and females,no differences were observed. A diagram representing thedistribution of the NKD-like receptor immunoreactivity inthe central nervous system of L. migratoria is shown inFigure 2.
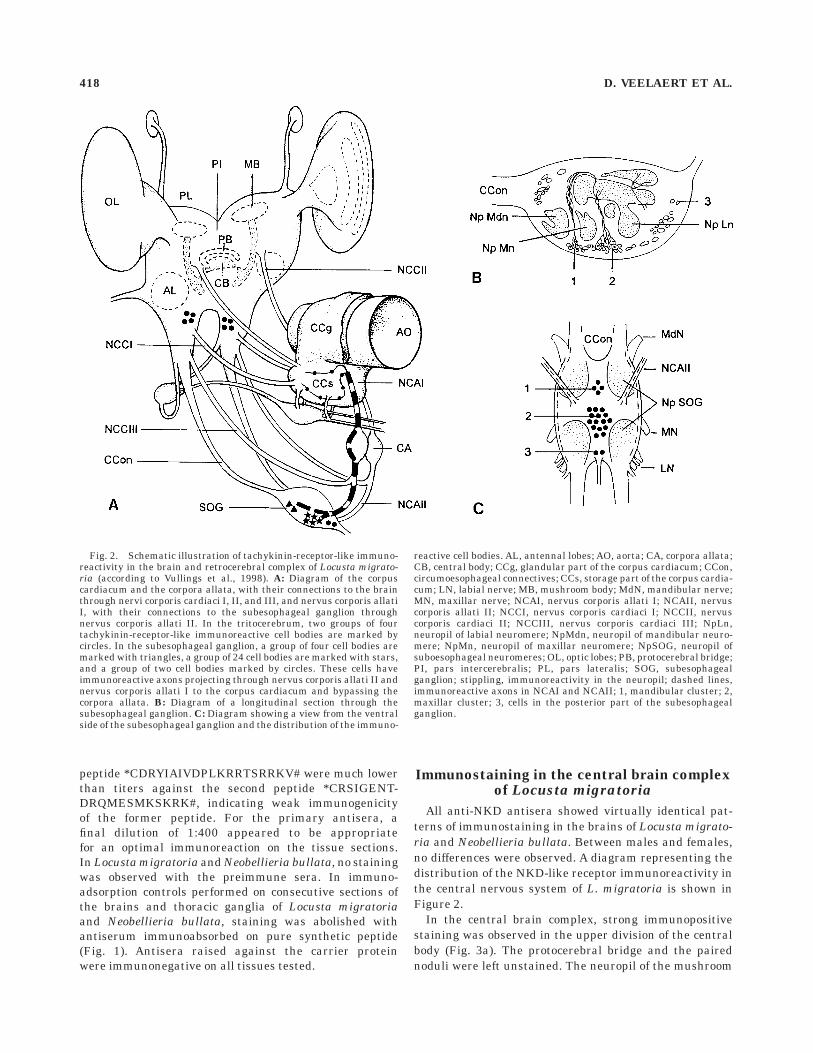
In the central brain complex, strong immunopositivestaining was observed in the upper division of the centralbody (Fig. 3a). The protocerebral bridge and the pairednoduli were left unstained. The neuropil of the mushroom
Fig. 2. Schematic illustration of tachykinin-receptor-like immuno-reactivity in the brain and retrocerebral complex of Locusta migrato-ria (according to Vullings et al., 1998). A: Diagram of the corpuscardiacum and the corpora allata, with their connections to the brainthrough nervi corporis cardiaci I, II, and III, and nervus corporis allatiI, with their connections to the subesophageal ganglion throughnervus corporis allati II. In the tritocerebrum, two groups of fourtachykinin-receptor-like immunoreactive cell bodies are marked bycircles. In the subesophageal ganglion, a group of four cell bodies aremarked with triangles, a group of 24 cell bodies are marked with stars,and a group of two cell bodies marked by circles. These cells haveimmunoreactive axons projecting through nervus corporis allati II andnervus corporis allati I to the corpus cardiacum and bypassing thecorpora allata. B: Diagram of a longitudinal section through thesubesophageal ganglion. C: Diagram showing a view from the ventralside of the subesophageal ganglion and the distribution of the immuno-
reactive cell bodies. AL, antennal lobes; AO, aorta; CA, corpora allata;CB, central body; CCg, glandular part of the corpus cardiacum; CCon,circumoesophageal connectives; CCs, storage part of the corpus cardia-cum; LN, labial nerve; MB, mushroom body; MdN, mandibular nerve;MN, maxillar nerve; NCAI, nervus corporis allati I; NCAII, nervuscorporis allati II; NCCI, nervus corporis cardiaci I; NCCII, nervuscorporis cardiaci II; NCCIII, nervus corporis cardiaci III; NpLn,neuropil of labial neuromere; NpMdn, neuropil of mandibular neuro-mere; NpMn, neuropil of maxillar neuromere; NpSOG, neuropil ofsuboesophageal neuromeres; OL, optic lobes; PB, protocerebral bridge;PI, pars intercerebralis; PL, pars lateralis; SOG, subesophagealganglion; stippling, immunoreactivity in the neuropil; dashed lines,immunoreactive axons in NCAI and NCAII; 1, mandibular cluster; 2,maxillar cluster; 3, cells in the posterior part of the subesophagealganglion.
418 D. VEELAERT ET AL.

bodies were immunopositive (Fig. 3b,c). The dispersedstaining suggests the presence of the NKD-like protein insynaptic contacts in the neuropil. No immunoreactivitywas observed in the deuterocerebrum. In the internolat-eral part of each tritocerebrum, four immunoreactive cellbodies were identified (Fig. 4a,b).
In the subesophageal ganglion, three clusters of neuro-nal cell bodies were immunoreactive. In the anterior cluster,which is situated closest to the upper brain, four cells werestained (Fig. 4c) whereas in the median cluster, about 24 cellscontained NKD-like immunoreactivity (Fig. 4e,f) and in the
posterior part two cells were immunopositive (Fig. 4d). Theanterior and median neurons have axons projecting into theNCA II. Immunoreactivity was found in presynaptic nerveterminals in the storage part of the corpus cardiacum (CCs).Extensive reaction was observed in the inner and outer lateralregions facing the glandular part of the corpus cardiacum(CCg), with the dorsal aorta on one side and the haemolymphon the other (Fig. 5a,b). Immunoreactivity was present in theNCA I, where it enters the corpus cardiacum (Fig. 5c,d). Noimmunoreactive nerve terminals were present in the CCg.ADK cells and corpora allata were left unstained.
Fig. 3. Tachykinin-receptor-like-immunoreactive material in neu-ropil regions of the central nervous system. a: Section through thecentral body, with immunoreactivity in the upper part (arrows).
b: Immunoreactivity in the a lobes of the mushroom body (arrow).c: Immunoreactivity in the b lobes of the mushroom body (arrows).Scale bars 5 50 µm.
DISTRIBUTION OF INSECT TACHYKININ RECEPTOR 419

Fig. 4. Tachykinin-receptor-like-immunoreactive cell bodies in thetritocerebrum and the subesophageal ganglion of Locusta migratoria.Punctate staining outside the intensely staining cell body and stainingin the sheath cells is background staining. a,b: Immunoreactive cellbodies at the internolateral part of the tritocerebrum. c: Two immuno-reactive cells of a group of two cells in the anterior subesophageal
ganglion, with axons projecting into the contralateral nervus corporisallati II (NCA II). d: Immunoreactive neuron in the posterior part ofthe subesophageal ganglion. e,f: Immunoreactive cell bodies of a groupof 2 3 12 cells in the central part of the subesophageal ganglion, whichhas axons projecting into the contralateral NCA II. NpMn, neuropile oflabial neuromere. Scale bars 5 25 µm.

Fig. 5. Tachykinin-receptor-like-immunoreactive nerve terminalsin the corpus cardiacum. a: Section through the corpus cardiacumwith immunoreactivity in the storage part (CCs), at the border withthe glandular part (CCg) and at the side facing the dorsal aorta (AO;arrows). No immunoreactivity is observed in the adipokinetic hormonecells (arrowheads). b: Immunoreactivity in the storage part of the
corpus cardiacum (CCs) at the side facing the hemolymph (Hae).c: Section through the storage part of the corpus cardiacum (CCs),where the nervus corporis allati (NCAI) enters the corpus cardiacum.Immunoreactivity is observed in fibers of the NCAI. d: Detail of theNCAI entering the CCs. Immunoreactive fibers in the NCAI (arrows).Scale bars 5 50 µm.
DISTRIBUTION OF INSECT TACHYKININ RECEPTOR 421

Double staining in the subesophageal ganglion with anantiserum against Lom-MT-1 showed colocalization ofFXPRLamide immunoreactivity, with the NKD-like immu-noreactivity found in the anterior and median cell clusters(Fig. 6a,b).
Immunostaining in the central brain complexof Neobellieria bullata
About 12 cell bodies of the subesophageal ganglionshowed immunoreactivity (Fig. 1). No immunoreactivitywas observed in parts of the neuropil or in other regions ofthe brain. The cells that were immunoreactive for thetachykinin receptor were also immunoreactive for theLom-MT-1 antiserum.
Immunostaining in the thoracic andabdominal ganglia of Locusta migratoria
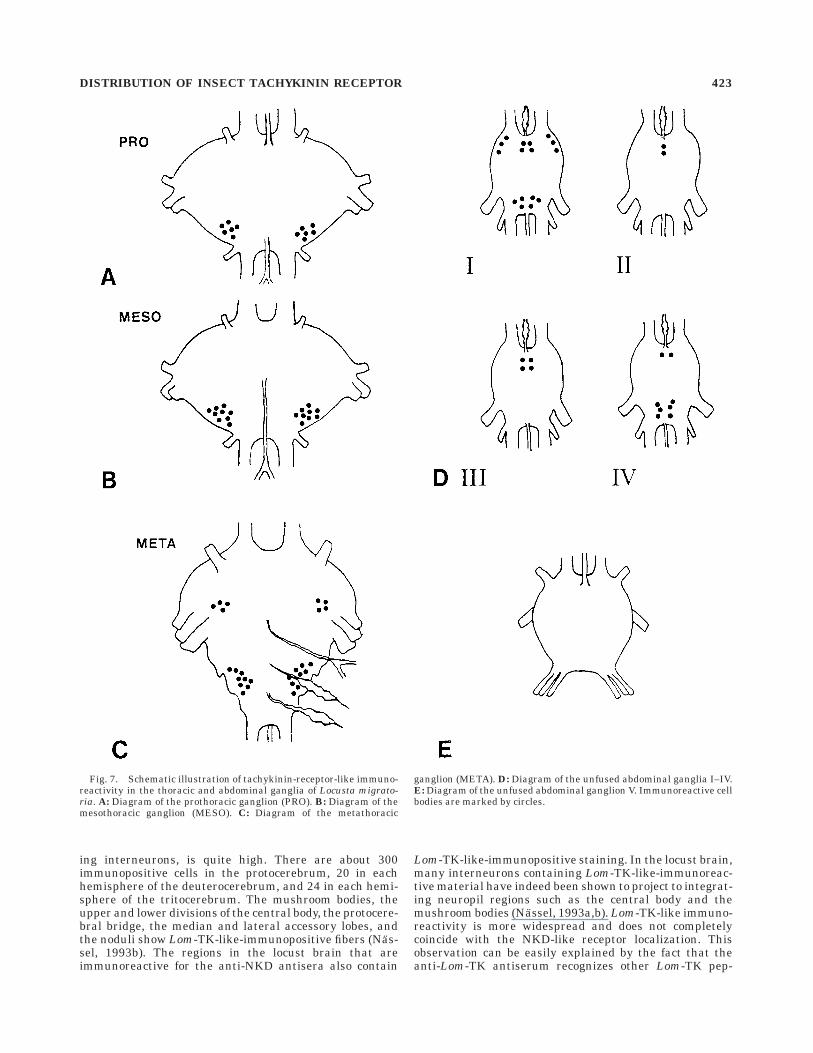
Many cell bodies were stained in the thoracic andabdominal ganglia. Probably due to the individual physi-
ological differences between the animals, it was quitedifficult to reproduce the entire staining pattern in allthese ganglia. As an example, the distribution of immu-nopositive staining obtained with the anti-NKD antiserais shown for one animal (Fig. 7). In the prothoracicganglion, two symmetrical groups of about six cell bodieswere observed in the posterior part. In the mesothoracicganglion, two symmetrical groups of about nine neuronswere observed. In the metathoracic ganglion, two symmetri-cal groups of four and two of nine cells were stained, thelatter ones in the fused abdominal ganglia. In most of theabdominal ganglia, additional symmetrical groups of im-munopositive cells were observed. The fifth unfused ab-dominal ganglion was left unstained. No immunoreactiv-ity was observed in neuropil regions of the thoracic andabdominal ganglia.
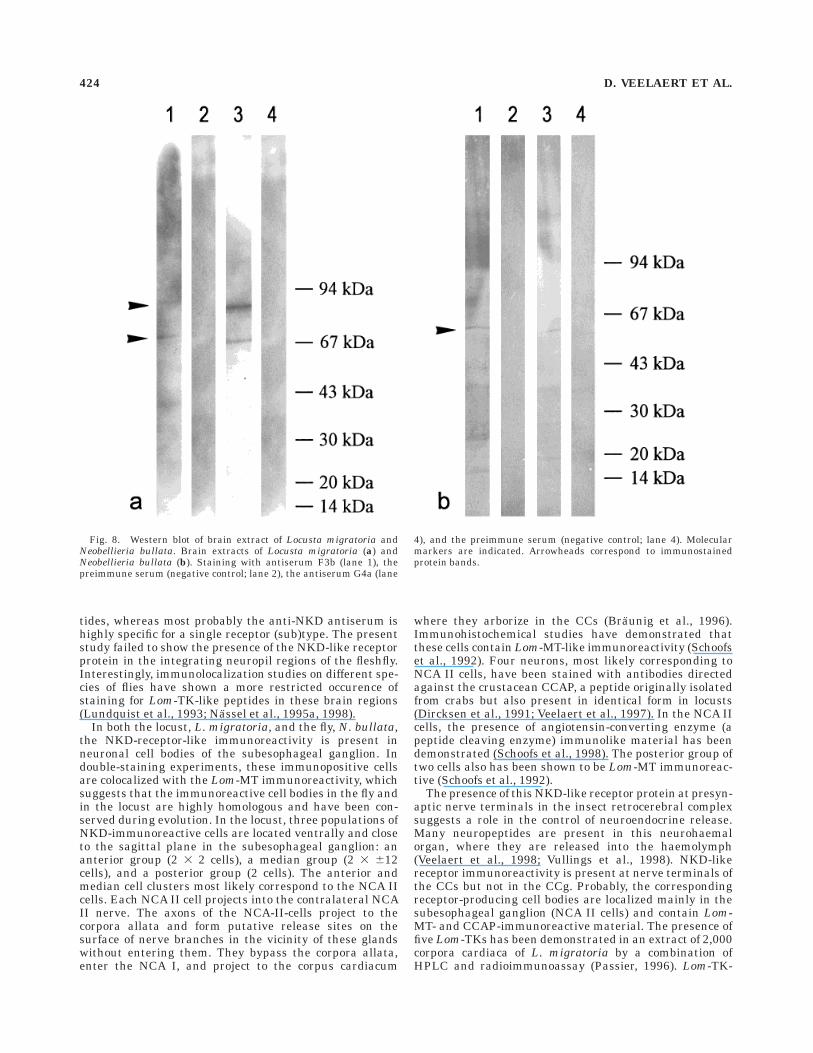
Western blot analysis of brain extracts
To investigate whether all antibodies recognize the sameprotein(s), we performed an immunoblotting with brainextracts of Locusta migratoria and Neobellieria bullata. InLocusta migratoria, we observed two protein bands (ap-proximately 67 kDa and 87 kDa) that reacted with boththe F3 and G4 antibodies. The preimmune controls werenegative. In Neobellieria bullata, only one protein of about57 kDa was immunopositive, and the preimmune serashowed no reaction (Fig. 8).
DISCUSSION
Very specific antisera were raised against two syntheticpeptide fragments corresponding to selected immunogenicparts of the NKD receptor. Staining in the central braincomplex was reproducable with all antisera producedagainst these two peptide fragments, despite the fact thatone of the peptides was much less immunogenic in guineapigs than the other. Apparently, however, the affinity ofboth peptide antisera to the NKD receptor was approxi-mately equal. No reaction was observed with the preim-mune sera, anti-KLH (carrier protein) antisera, and theimmunoadsorption controls. Western blot analysis showedthe presence of two protein bands in Locusta migratoriaand a single protein in Neobellieria bullata brain extracts.These bands were immunoreactive for all antisera tested.The estimated molecular weights of these bands are in therange expected for G-protein-coupled receptors. The pres-ence of two immunopositive protein bands in locust brainextracts suggests that locust brain probably contains twoNKD-like isoforms or related receptors.
In the locust central body, which is implicated in visualprocessing because it links pathways from the compoundeyes and ocelli (Bausenwein et al., 1994; Burrows, 1996)and in the control of motor output such as walking andflight (Homberg, 1994; Ilius et al., 1994), strong immu-nopositive staining was observed. This integrative neuro-pil region contains several monoamine neurotransmitterssuch as octopamine, dopamine, and serotonin (Homberg etal., 1989; Homberg, 1991; Wendt and Homberg, 1992;Burrows 1996), but some neuropeptides were also demon-strated to be present in this part of the protocerebrum,e.g., allatostatins (Veelaert et al., 1995; Vitzthum et al.,1996), crustacean cardioactive peptide (CCAP; Dircksenand Homberg, 1995), and Lom-TK (Nassel, 1993a; Vitz-thum and Homberg, 1998). In the locust brain, the totalnumber of Lom-TK-immunoreactive neurons, mostly be-
Fig. 6. Colocalization of locustamyotropin-like and tachykinin-receptor-like immunoreactivity in cell bodies on consecutive sectionsthrough the subesophageal ganglion. The NKD staining is not asuniform and is more restricted than locustamyotropin-1 because ofdifferences in the concentration of the antigen and the low titer of theNKD antibodies. a: Section stained with the tachykinin-receptorantiserum. b: Section stained with locustamyotropin antiserum. Scalebars 5 50 µm.
422 D. VEELAERT ET AL.
ing interneurons, is quite high. There are about 300immunopositive cells in the protocerebrum, 20 in eachhemisphere of the deuterocerebrum, and 24 in each hemi-sphere of the tritocerebrum. The mushroom bodies, theupper and lower divisions of the central body, the protocere-bral bridge, the median and lateral accessory lobes, andthe noduli show Lom-TK-like-immunopositive fibers (Nas-sel, 1993b). The regions in the locust brain that areimmunoreactive for the anti-NKD antisera also contain
Lom-TK-like-immunopositive staining. In the locust brain,many interneurons containing Lom-TK-like-immunoreac-tive material have indeed been shown to project to integrat-ing neuropil regions such as the central body and themushroom bodies (Nassel, 1993a,b). Lom-TK-like immuno-reactivity is more widespread and does not completelycoincide with the NKD-like receptor localization. Thisobservation can be easily explained by the fact that theanti-Lom-TK antiserum recognizes other Lom-TK pep-
Fig. 7. Schematic illustration of tachykinin-receptor-like immuno-reactivity in the thoracic and abdominal ganglia of Locusta migrato-ria. A: Diagram of the prothoracic ganglion (PRO). B: Diagram of themesothoracic ganglion (MESO). C: Diagram of the metathoracic
ganglion (META). D: Diagram of the unfused abdominal ganglia I–IV.E: Diagram of the unfused abdominal ganglion V. Immunoreactive cellbodies are marked by circles.
DISTRIBUTION OF INSECT TACHYKININ RECEPTOR 423
tides, whereas most probably the anti-NKD antiserum ishighly specific for a single receptor (sub)type. The presentstudy failed to show the presence of the NKD-like receptorprotein in the integrating neuropil regions of the fleshfly.Interestingly, immunolocalization studies on different spe-cies of flies have shown a more restricted occurence ofstaining for Lom-TK-like peptides in these brain regions(Lundquist et al., 1993; Nassel et al., 1995a, 1998).
In both the locust, L. migratoria, and the fly, N. bullata,the NKD-receptor-like immunoreactivity is present inneuronal cell bodies of the subesophageal ganglion. Indouble-staining experiments, these immunopositive cellsare colocalized with the Lom-MT immunoreactivity, whichsuggests that the immunoreactive cell bodies in the fly andin the locust are highly homologous and have been con-served during evolution. In the locust, three populations ofNKD-immunoreactive cells are located ventrally and closeto the sagittal plane in the subesophageal ganglion: ananterior group (2 3 2 cells), a median group (2 3 612cells), and a posterior group (2 cells). The anterior andmedian cell clusters most likely correspond to the NCA IIcells. Each NCA II cell projects into the contralateral NCAII nerve. The axons of the NCA-II-cells project to thecorpora allata and form putative release sites on thesurface of nerve branches in the vicinity of these glandswithout entering them. They bypass the corpora allata,enter the NCA I, and project to the corpus cardiacum
where they arborize in the CCs (Braunig et al., 1996).Immunohistochemical studies have demonstrated thatthese cells contain Lom-MT-like immunoreactivity (Schoofset al., 1992). Four neurons, most likely corresponding toNCA II cells, have been stained with antibodies directedagainst the crustacean CCAP, a peptide originally isolatedfrom crabs but also present in identical form in locusts(Dircksen et al., 1991; Veelaert et al., 1997). In the NCA IIcells, the presence of angiotensin-converting enzyme (apeptide cleaving enzyme) immunolike material has beendemonstrated (Schoofs et al., 1998). The posterior group oftwo cells also has been shown to be Lom-MT immunoreac-tive (Schoofs et al., 1992).
The presence of this NKD-like receptor protein at presyn-aptic nerve terminals in the insect retrocerebral complexsuggests a role in the control of neuroendocrine release.Many neuropeptides are present in this neurohaemalorgan, where they are released into the haemolymph(Veelaert et al., 1998; Vullings et al., 1998). NKD-likereceptor immunoreactivity is present at nerve terminals ofthe CCs but not in the CCg. Probably, the correspondingreceptor-producing cell bodies are localized mainly in thesubesophageal ganglion (NCA II cells) and contain Lom-MT- and CCAP-immunoreactive material. The presence offive Lom-TKs has been demonstrated in an extract of 2,000corpora cardiaca of L. migratoria by a combination ofHPLC and radioimmunoassay (Passier, 1996). Lom-TK-
Fig. 8. Western blot of brain extract of Locusta migratoria andNeobellieria bullata. Brain extracts of Locusta migratoria (a) andNeobellieria bullata (b). Staining with antiserum F3b (lane 1), thepreimmune serum (negative control; lane 2), the antiserum G4a (lane
4), and the preimmune serum (negative control; lane 4). Molecularmarkers are indicated. Arrowheads correspond to immunostainedprotein bands.
424 D. VEELAERT ET AL.

like fibers were localized in the NCA II nerves to the corpuscardiacum and a sparse distribution of varicose fibers wasdetected within the CCg and CCs. Electron microscopicimmunocytochemistry has shown that Lom-TK-like axonterminals containing granular vesicles make contact withthe glandular cells (Nassel et al., 1995b).
Although Lom-TK-1 has been shown to induce therelease of AKH by the glandular cells (Nassel et al.,1995b), no NKD-receptor-like immunoreactivity was pres-ent at these AKH cells. There are three possible explana-tions for this observation: (1) a NKD-like receptor ispresent in very low, undetectable amounts; (2) anotherLom-TK receptor subtype is involved in the release ofAKH; or (3) Lom-TKs have an indirect effect on the releaseof AKH. The absence of NKD immunoreactivity on theAKH cells probably explains the need for unexpectedlyhigh concentrations of Lom-TK-1 (ED50 6 50 µM) to inducethe release of AKHs (Nassel et al., 1995b). This explana-tion is in contrast with the activity of CCAP, which inducesthe release of AKHs at 1,000-fold lower concentrations(ED50 6 10 nM; Veelaert et al., 1997). Immunoreactivityfor CCAP has been observed in some NCA II cells and inthe CCs, but not in the CCg (Dircksen et al., 1991). Thepresent study suggests that the NKD-like receptor ispresent at presynaptic termini of NCA II cells, and some ofthese cells have been reported to contain CCAP, a potentAKH-releasing neurohormone.
The presence of NKD receptor immunoreactivity in thethoracic and abdominal ganglia suggests the existence ofadditional roles that remain to be determined. Lo-custatachykinins also play an important role in the diges-tive tract. Six tachykinin-related peptides have recentlybeen isolated from the midgut of the cockroach Leucophaeamaderae (Muren et al., 1995; Muren and Nassel, 1996a,b,1997a,b). These peptides are stimulators of hindgut con-tractility in this cockroach species (Schoofs et al., 1993;Winther et al., 1998). Moreover, the NCA II cells in thesubesophageal ganglion, which have been shown to beNKD immunopositive, also possess ramifications towardthe esophagus (Braunig et al., 1996).
This is the first report to describe the localization of acognate neuropeptide receptor protein in insect nervoustissues. The study couples the existing immunolocaliza-tion data for insect tachykinin, myotropin, and CCAPpeptides to those of NKD, an insect tachykinin receptorshown to be activated by Lom-TK-like peptides.
ACKNOWLEDGMENTS
We thank M. Christiaens, J. Gijbels, R. Jonkers, S.Kotanen, G.A. Posthuma-Trumpie, and J. Puttemans fortechnical assistance. Dirk Veelaert is a postdoctoral stu-dent and Jozef Vanden Broeck is a Senior ResearchAssociate of the FWO-Flanders.
LITERATURE CITED
Ashby G. 1972. Locusts In: UFAW, editor. The UFAW handbook on the careand management of laboratory animals. Edinburgh: Churchill Living-stone. p 582–587.
Bausenwein B, Muller RM, Heisenberg M. 1994. Behaviour dependentactivity labeling in the central complex of Drosphila melanogasterduring controlled visual stimuli. J Comp Neurol 340:255–268.
Bradford MM. 1976. A rapid and sensitive method for quantification ofmicrogram quantities of protein utilizing the principe of protein dyebinding. Anal Biochem 72:248–254.
Braunig P, Bohme C, Staufer B. 1996. Morphology of locust neurosecretorycells projecting into the nervus corporis allati II of the suboesophagealganglion. Microsc Res Tech 35:230–241.
Burrows M. 1996. The neurobiology of an insect brain. Oxford: OxfordUniversity Press. p 699.
Christie AE, Lundquist CT, Nassel DR, Nusbaum MP. 1997. Two noveltachykinin-related peptides from the nervous system of the crab Cancerborealis. J Exp Biol 200:2279–2294.
Dircksen H, Homberg U. 1995. Crustacean cardioactive peptide-immunore-active neurons innervating brain neuropils, retrocerebral complex andstomatogastric nervous system of the locust Locusta migratoria. CellTissue Res 279:495–515.
Dircksen H, Muller A, Keller R. 1991. Crustacean cardioactive peptide inthe nervous system of the locust, Locusta migratoria: an immunocyto-chemical study on the ventral nerve cord and peripheral innervation.Cell Tissue Res 263:439–457.
Fields CG, Lloyd DH, Macdonald RL, Otteson KM, Noble RL. 1991. HBTUactivation for automated Fmoc solid-phase peptide synthesis. PeptideRes 4:95–101.
Fujisawa J, Muneoka Y, Takashashi T, Takao T, Shimonishi Y, Kubota I,Ikeda T, Minakata H, Nomoto K, Kiss T, Hiripi L. 1994. An invertebrate-type tachykinin isolated from the freshwater bivalve mollusc, Anodontacygnea. In: Okoda Y, editor. Peptide chemistry. Osaka: Protein ResearchFoundation. p 161–164.
Guerrero FD, Clottens FL, Holman GM, Vanden Broeck J, De Loof A. 1997.Cloning and sequencing of a Stomoxys calcitrans cDNA which encodes aputative tachykinin peptide receptor. 7TM J 6:1–4.
Helke CJ, Krause JE, Mantyh PW, Couture R, Bannon MJ. 1990. Diversityin mammalian tachykinin peptidergic neurons: multiple peptides,receptors, and regulatory mechanisms. FASEB J 4:1606–1615.
Homberg U. 1991. Neuroarchitecture of the central complex in the brain ofthe locust Schistocerca gregaria and S. americana as revealed byserotonin immunocytochemistry. J Comp Physiol 303A:245–254.
Homberg U. 1994. Flight-correlated activity changes in neurons of thelateral accessory lobes in the brain of the locust Schistocerca gregaria. JComp Physiol 175A:597–610.
Homberg U, Binkle U, Lehmann HK, Vullings HGB, Eckert M, Rapus J,Hildebrand JG. 1989. Octopamine-immunoreactivity in the brain of twoinsect species. In: Elsner N, Richter DW, editors. Rhythmogenesis inneurons and networks. Stuttgart: Thieme. p 477.
Huybrechts R, De Loof A. 1981. Effect of ecdysteron on vitellogeninconcentration in haemolymph of male and female Sarcophaga. Int JInvert Reprod 3:91–98.
Ikeda T, Minakata H, Nomoto K, Kubota I, Muneoka Y. 1993. Two noveltachykinin-related neuropeptides in the echiuroid worm, Urechisunicinctus. Biochem Biophys Res Commun 192:1–6.
Ilius M, Wolf R, Heisenberg M. 1994. The central complex of Drosophilamelanogaster is involved in flight control: studies on mutants andmosaics of the gene ellipsoid body open. J Neurogenet 9:189–206.
Laemmli UK. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227:680–685.
Leary JJ, Brigati DJ, Ward DC. 1983. Rapid and sensitive colorimetricmethod for visualizing biotin-labeled DNA probes hybridized to DNA orRNA immobilized on nitrocellulose. Proc Natl Acad Sci USA 80:4045–4049.
Li XJ, Wolfgang W, Wu YN, North RA, Forte M. 1991. Cloning, heterologousexpression and developmental regulation of a Drosophila receptor fortachykinin-like peptides. EMBO J 10:3221–3229.
Lundquist CT, Brodin E, Muren JE, Nassel DR. 1993. Tachykinin- andleucokinin-related peptides in the nervous system of the blowfly:immunocytochemical and chromatographical diversity. Peptides 14:651–663.
Lundquist CT, Clottens FL, Holman GM, Nichols R, Nachman RJ, NasselDR. 1994a. Callitachykinin I and II, two novel myotropic peptidesisolated from the blowfly, Calliphora vomitoria, that have resemblancesto tachykinins. Peptides 15:761–768.
Lundquist CT, Clottens FL, Holman GM, Riehm JP, Bonkale W, Nassel DR.1994b. Locustatachykinin immunoreactivity in the blowfly centralnervous system and intestine. J Comp Neurol 341:225–240.
Maggi CA. 1995. The mammalian tachykinin receptors. Gen Pharmacol26:911–944.
Meloen RH, Turkstra JA, Lankhof H, Puijk WC, Schaaper WMM, DijkstraG, Wensing CJG, Oonk RB. 1994. Efficient immunocastration of malepiglets by immunoneutralization of GnRH, using a new GnRH-likepeptide. Vaccine 12:741–746.
DISTRIBUTION OF INSECT TACHYKININ RECEPTOR 425

Meola SM, Clottens FL, Holman GM, Nachman RJ, Nichols R, Schoofs L,Wright MS, Olson JK, Hayes TK, Pendleton MW. 1998. Isolation andimmunocytochemical characterization of three tachykinin related pep-tides from the mosquito, Culex salinarius. Neurochem Res 23:189–202.
Monnier D, Colas JF, Rosay P, Hen R, Borrelli E, Maroteaux L. 1992. NKD,a developmentally regulated tachykinin receptor in Drosophila. J BiolChem 267:1298–1302.
Muren JE, Nassel DR. 1996a. Isolation of five tachykinin-related peptidesfrom the midgut of the cockroach Leucophaea maderae: existence ofN-terminally extended isoforms. Regul Peptides 65:185–196.
Muren JE, Nassel DR. 1996b. Radioimmunoassay determination of tachy-kinin-related peptide in different portions of the central nervous systemand intestine of the cockroach Leucophaea maderae. Brain Res 739:314–321.
Muren JE, Nassel DR. 1997a. Novel tachykinin-related peptides in thecockroach nervous system and intestine. Structure, distribution, andactions. Ann NY Acad Sci 814:312–314.
Muren JE, Nassel DR. 1997b. Seven tachykinin-related peptides isolatedfrom the brain of the Madeira cockroach: evidence for tissue-specificexpression of isoforms. Peptides 18:7–15.
Muren JE, Lundquist CT, Nassel DR. 1995. Abundant distribution oflocustatachykinin-like peptide in the nervous system and intestine ofthe cockroach Leucophaea maderae. Phil Trans R Soc Lond B Biol Sci348:423–444.
Nassel DR. 1993a. Insect myotropic peptides: differential distribution oflocustatachykinin- and leucokinin-like immunoreactive neurons in thelocust brain. Cell Tissue Res 274:27–40.
Nassel DR. 1993b. Neuropeptides in the insect brain: a review. Cell TissueRes 273:1–29.
Nassel DR, Kim MY, Lundquist CT. 1995a. Several forms of callitachyki-nins are distributed in the central nervous system and intestine of theblowfly Calliphora vomitoria. J Exp Biol 198:2527–2536.
Nassel DR, Passier PCCM, Elekes K, Dircksen H, Vullings HGB, CanteraR. 1995b. Evidence that locustatachykinin is involved in release ofadipokinetic hormone from the locust corpora cardiaca. Regul Peptides57:297–310.
Nassel DR, Lundquist CT, Muren JE, Winther AME. 1998. An insectpeptide family in search of functions: the tachykinin-related peptides.In: Coast GM, Webster SG, editors. Recent advances in arthropedandocrinology. Cambridge: Cambridge University Press. p 248–277.
Nieto J, Veelaert D, Derua R, Cerstiaens A, Waelkens E, Coast G, CalderonJ, De Loof A, Schoofs L. 1998. Identification of one tachykinin and twokinin-related petpides in the brain of the white shrimp, Penaeusvannamei. Biochem Biophys Res Comm 248:406–411.
Passier PCCM. 1996. The multifactorial control of the release of adipoki-netic hormones from the locust corpus cardiacum [PhD thesis]. Univer-sity of Utrecht.
Peferoen M. 1988. Blotting with plate electrodes. In: Walker J, editor.Methods in molecular biology. Volume 3. London: Humana Press. p 222.
Rink H. 1987. Solid-phase synthesis of protected peptide fragments using atrialkoxy-diphenyl-methylester resin. Tetrahedron Lett 28:3787–3790.
Schoofs L, Holman GM, Hayes TK, Kochansky JP, Nachman RJ, De Loof A.1990a. Locustatachykinin III and IV: two additional insect neuropep-tides with homology to peptides of the vertebrate tachykinin family.Regul Peptides 31:199–212.
Schoofs L, Holman GM, Hayes TK, Nachman RJ, De Loof A. 1990b.Locustatachykinin I and II, two novel insect neuropeptides withhomology to peptides of the vertebrate tachykinin family. FEBS Lett261:397–401.
Schoofs L, Tips A, Holman GM, Nachman RJ, De Loof A. 1992. Distributionof locustamyotropin-like immunoreactivity in the nervous system ofLocusta migratoria. Regul Peptides 37:237–254.
Schoofs L, Vanden Broeck J, De Loof A. 1993. The myotropic peptides ofLocusta migratoria: structures, distribution, functions and receptors.Insect Biochem Mol Biol 23:859–881.
Schoofs L, Veelaert D, Vanden Broeck J, De Loof A. 1997. Peptides in thelocusts, Locusta migratoria and Schistocerca gregaria. Peptides 18:145–156.
Schoofs L, Veelaert D, De Loof A, Huybrechts R, Isaac E. 1998. Immunocy-tochemical distribution of angiotensin I-converting enzyme-like immu-noreactivity in the brain and testis of insects. Brain Res 785:215–227.
Vanden Broeck J. 1996. G-protein-coupled receptors in insect cells. Int RevCytol 164:189–268.
Veelaert D, Schoofs L, Tobe SS, Yu CG, Vullings HG, Couillaud F, De Loof A.1995. Immunological evidence for an allatostatin-like neuropeptide inthe central nervous system of Schistocerca gregaria, Locusta migratoriaand Neobellieria bullata. Cell Tissue Res 279:601–611.
Veelaert D, Passier P, Devreese B, Vanden Broeck J, Van Beeumen J,Vullings HGB, Diederen JHB, Schoofs L, De Loof A. 1997. Isolation andcharacterization of an adipokinetic hormone release-inducing factor inlocusts: the crustacean cardioactive peptide. Endocrinology 138:138–142.
Veelaert D, Schoofs L, De Loof A. 1998. Peptidergic control of the corpuscardiacum-corpora allata complex of locusts. Int Rev Cytol 182:249–302.
Vitzthum H, Homberg U. 1998. Immunocytochemical demonstration oflocustatachykinin-related peptides in the central complex of the locustbrain. J Comp Neurol 390:455–469.
Vitzthum H, Homberg U, Agricola H. 1996. Distribution of Dip-allatostatinI-like immunoreactivity in the brain of the locust Schistocerca gregariawith detailed analysis of immunostaining in the central complex. JComp Neurol 369:419–437.
Vullings HGB, Diederen JHG, Veelaert D. 1999. The multifactorial controlof the release of hormones from the locust retrocerebral complex.Microsc Res Tech, in press.
Watling KJ, Guard S, Boyle SJ, McKnight AT, Woodruff GN. 1994. Speciesvariants of tachykinin receptor types. Biochem Soc Trans 22:118–122.
Wendt B, Homberg U. 1992. Immunocytochemistry of dopamine in thebrain of the locust Schistocerca gregaria. J Comp Neurol 321:387–403.
Winther AM, Muren JE, Lundquist CT, Osborne RH, Nassel DR. 1998.Characterization of actions of Leucophaea tachykinin-related peptides(LemTRPs) and proctolin on cockroach hindgut contractions. Peptides19:445–458.
426 D. VEELAERT ET AL.




















