
Subliminal vibro-tactile based notification of CO 2 economy while driving
Upload
khangminh22Category
view
1download
0
T
Tabes Dorsalis
Definition
A late complication of neurosyphilis. It results in gaitimpairment, joint deformity, pain, lack of coordination,sensory loss, as well as autonomic dysfunction and oc-ular symptoms. Pain is felt mostly in the legs and is de-scribed as lightning and lancinating. Abdominal colickypain is also reported.� Central Nervous System Stimulation for Pain
Tachykinin
Definition
Tachykinins are a family of structurally-related pep-tides, widely scattered in vertebrate and invertebratetissues. Mammalian tachykinins are substance P (SP),neurokinin A (NKA), and neurokinin B (NKB). Allmammalian tachykinins share a common C-terminalamino acid sequence, i.e. Phe-x-Gly-Leu- MetNH,which is the minimal structural motif for the activationof tachykinin receptors (NK1, NK2 and NK3). Phar-macologically, they all cause hypotension in mammals,contraction of gut and bladder smooth muscle, andsecretion of saliva.� Neuropeptide Release in the Skin� NGF, Regulation during Inflammation� Visceral Nociception and Pain
Tachyphylaxis
� Nociceptor, Fatigue
Tactile Allodynia
Definition
Tactile allodynia refers to touch-evoked pain, i.e. paindue to a mechanical stimulus that does not normally pro-voke pain.
� OpioidsintheSpinalCordandModulationofAscend-ing Pathways (N. gracilis)
Tactile Allodynia Test
Definition
The plantar aspects of intact and neuropathic legs of ratsare probed with Von Frey hairs of different calibers orstrengths. The number of paw withdrawals per 10 trialsis counted. In general, a number of 5/10 withdrawals isobserved with hairs of strength > 20 g in rats with intactlegs, and in rats with mononeuropathy, a hair of 2 g canproduce a score of > 5/10 withdrawals.� Thalamotomy, Pain Behavior in Animals
Tactile Stimuli
Definition
Stimuli of light touch applied to the skin.� Causalgia, Assessment� Dysesthesia, Assessment
Tail Immersion
Definition
Submersion of the tail in hot water may be used as thenociceptive stimulus in the Tail-Flick Test.� Tail-Flick Test
Tail Skin Temperature Recording
Definition
Thermocouples, thermistors or infrared sensors may beused.� Tail-Flick Test

2392 Tail-Flick Latency
Tail-Flick Latency
Definition
Tail-Flick latency is the timefrom thestartof thenoxiousstimulus until the animal flicks its tail.� Tail-Flick Test
Tail-Flick Latency Correction
Definition
The tail-flick latency may be corrected for influences oftail skin temperature using a regression analysis or anal-ysis of covariance. Linearity of the data can be assumedonly for a limited range of skin temperatures. In someexperiments preheating of the tail to a certain tempera-ture may be used.� Tail-Flick Test
Tail Flick TestKJELL HOLE, ARNE TJØLSEN
University of Bergen, Bergen, [email protected], [email protected]
Definition
The tail–flick test is a test of nociception used in rats andmice. The noxious stimulus is usually � radiant heat onthe tail or � tail immersion in hot water, and the responseis a flick of the tail.
Characteristics
The tail–flick test is an extensively used test of nocicep-tion in rats and mice, and is the nociceptive test mostfrequently used in animals (Le Bars et al. 2001), firstdescribed in 1941 (D’Amour and Smith 1941). In thestandard method, radiant heat is focused on the tail, andthe time it takes until the animal flicks the tail away fromthe beam is measured. This � tail–flick latency is a mea-sure of the nociceptive sensitivity of the animal, and isprolonged by opioid analgesics, for instance. A spinaltransection above the lumbar level does not block thetail–flick response. Thus, in this test, a spinal nocicep-tive reflex ismeasured, and pain isnotmeasured directly.Still, this isconsideredaveryuseful testof“phasicpain”,both inbasicpainresearchandinpharmacological inves-tigationsofanalgesicdrugs.The relevanceof the test asameasure of pain has been discussed (LeBars et al. 2001).The test stimulus is noxious heat. In addition to the testwith radiant heat (e.g. focused light from a light bulb),the stimulus may be applied by e.g. direct contact witha heated surface, such as a Peltier element, or by sub-mersion of part of the tail in hot water. The test may be
performed in lightly anaesthetized rats or mice, as wellas in animals that are awake.Several tail–flick apparatuses are commercially avail-able, and many laboratories have made their own appa-ratus. The main requirements are stable functioning andproper focus of the light beam on the tail. The tail–flicklatency may be recorded by means of a photocell, whichisactivatedwhentheanimalflicksthe tail.Whenaphoto-cell is used, one should be aware that the reflex responsemay involve retracting the tail, without immediately re-moving the tail from the light beam. Thus the rats’ be-haviour should always be observed.The tail–flick test may be a good and useful test of noci-ception, but only if it is carefully performed and possiblesourcesoferror are taken intoaccount.One requirement,particularly in rats, is that the animals are well handled.This may require daily handling forup to a week, includ-ing adaptation to the test apparatus. Some researchersconfine the animals in a plastic tube during testing. Ifthis is used, it is necessary that the animals are so wellhandled that they freely walk in and out of the tube. Wefind it better and faster not to use a tube, but to hold thewell-adapted animal by hand.A particular problem with tests that use thermal stimu-lation is the possible confounding influence of the skintemperature. In electrophysiological studies in animals,it has been reported that changes in the temperature orblood flow of the skin (Duggan et al. 1978) alter the re-sponse to cutaneous heat stimulation. More recently, ithas been found that the tail skin temperature affects thetail–flick latency as well. This has been described usingradiant heat stimulation (Ren and Han 1979; Berge etal. 1988; Roane et al. 1998; Sawamura et al. 2002) aswell as with hot water immersion of the tail (Milne andGamble 1989). For an extensive review see Le Bars etal. (2001). However, in many laboratories the tail–flicktest is still performed without taking the tail skin temper-ature into account. This is probably a main confoundingfactor, and therefore needs special consideration in thefollowing.We have investigated the relationship between skin sur-face temperature and tail–flick latency in rats in a setupwith a radiant heat apparatus, stimulating the distal partof the tail (10–15 mm from the tip), with a stimulatedarea of 15–20 mm2 (Fig. 1). Using control latencies ofapproximately 4s, we regularly find a clear and repro-ducible relationship between tail skin temperature andtail–flick latency, with a slope of the regression equa-tion of –0.3 – 0.4s/˚C (Tjølsen et al. 1989). With similarmethodology, the same relationship has been found inmice, with a very similar slope (Eide et al. 1988).The tail is the most important thermoregulatory organ ofthe rat. The heat loss is regulated by an on–off regulationofblood flowin the tail,which leads to rapid variations inskin temperature (Milne and Gamble 1989, Tjølsen andHole 1992). The amount and duration of vasodilation ispartly determined by the relationship between the am-

T
Tail Flick Test 2393
Tail Flick Test, Figure 1 Simple test equipment for concomitant record-ing of tail skin temperatures and tail–flick latencies. A standard tail–flickapparatus can easily be modified to enable recording of tail skin temper-atures. The temperature is measured by means of a small thermocouplemounted on a plastic arm, 65 mm long, which rests on the tail with aforce corresponding to approximately 1g. For a thorough description seeTjølsen et al. (1989).
bient temperature and the acclimatization temperature.In rats at rest, the ambient temperature where vasodila-tion occurs is lower after acclimatization to cold, thanafter acclimatization to a warmer environment. Whenanimals are lightly stressed and activated due to exper-imental procedures, a considerable increase in tail skintemperature is regularly observed (Tjølsen et al. 1989).Rats restrained in tubes for a short time may show a con-siderable increase in the temperature of the tail (Tjølsenand Hole 1992), probably due to vasodilation.The relationship between skin temperatureand responselatency (Fig. 2) would be expected to vary with differentexperimentalconditions.Themost reliablevaluesfor theslope are obtained in experiments where data from re-peated measures are not pooled, but analysed separately
Tail Flick Test, Figure 2 The relationship between tail–flick latency andtail skin temperature. Data were obtained from eight measurements in eachof 12 rats. Tail skin temperature was controlled by means of a heatingblanket. Adapted from Sawamura et al. (2002), with permission.
for each time point (Tjølsen et al. 1989). In fact, if re-peated measures on the same animals are pooled in a re-gression analysis, an error in the calculated slope may beintroduced. A possible cause of error is the effect of re-peated testingonnociception itself,whetherduetostressor to local effects in the skin if the same site is stimu-lated repeatedly. The time required for heating the tissueto a critical response temperature will depend on the ini-tial skin temperature,which isdetermined by localbloodflow within the limits given by deep body and ambienttemperatures. Measuring subcutaneous tissue tempera-tures during a radiant heat stimulus, we found that therate of increase in tissue temperature was independent ofinitial skin temperature, and the time required to reach ahypothetical threshold temperature was strongly depen-dent on the initial temperature (Hole and Tjølsen 1993).Asaconsequence, the tail–flick latency isnegativelycor-related to the ambient temperature (Berge et al. 1988)and toskin temperaturewhentheheatingintensity iskeptconstant.The temperature of the tail skin of rats during an ex-periment may rise as much as 8˚C in untreated animals(Tjølsen and Hole 1992). It is reasonable to considerthis is the maximal possible difference in skin temper-ature due to changes in vasodilation. With a changein tail–flick latency of 0.3–0.4 s/˚C, it would implya potential difference in tail–flick latency of up toapproximately 3s. In a group of rats, not all animalswould show this degree of vasodilation, and hence themean difference in latency would be somewhat smaller.However, this shows that increased vasoconstrictionor inhibition of vasodilation may cause differences in

2394 Tail Flick Test
tail–flick latency that easily could be misinterpreted asanalgesia.The potential for treatment–induced vasodilation tocause reduction of the tail–flick latencies is approx-imately the same size. Under circumstances whencontrol animals are relatively vasoconstricted, vasodi-lation may lead to an increase in tail skin temperaturefrom about ambient temperature to above 30˚C. The ef-fect of vasodilation is particularly important, as smallerchanges in the tail–flick latency are required to interpretthe results as hyperalgesia than as analgesia. Even amodest increase in the mean tail skin temperature ofabout 3.5˚C, due to lesioning of descending serotoner-gic systems, leads to a reduction of the tail–flick latencyfrom 4–4.5s to about 3s (Hole and Tjølsen 1993). Ifthe change in skin temperature were not taken intoconsideration, a reduction of the tail–flick latency ofthis size would have been considered an indication of ahyperalgesic state.Many experimental treatments affect blood flow andthereby the tail skin temperature. This may by itselfinfluence the tail–flick latency, and lead to erroneousconclusions with regard to nociception. An increase intail skin temperature may shorten tail–flick latenciesand may be interpreted as hyperalgesia (Urban andSmith 1994; Roane et al. 1998; Sawamura et al. 2002).Even a reduction in tail skin temperature compared tountreated animals may occur, and may be interpreted asanalgesia. Desipramine reduced tail skin temperatureand increased tail–flick latencies at an ambient tem-perature of 24–25˚C, while no significant change wasobserved at 21–22˚C (Hole and Tjølsen 1993). Thisdifference in temperature is well within the variationin ambient temperature between laboratories, and evenwithin the range of ambient temperatures that may occurin a laboratory with insufficient control of room tem-perature. In the experiments at 24–25˚C, desipramineinhibited vasodilation so that the skin temperaturesin the drug–treated group were close to the ambienttemperature, while control animals showed higher skintemperatures and hence shorter response latencies.Stress, due to a new environment, handling or injectionprocedures, may influence peripheral blood flow and tailtemperature. In rats, stress causes motor activation, in-creased heat production, increased core temperature andan increased frequency and duration of the periods of va-sodilation and increase in skin temperature of the tail. Ithas been shown (Tjølsen et al. 1992) that immobiliza-tion may cause a considerable increase in core tempera-ture and tail vasodilation, while small doses of morphine(0.5–1mg/kg) completely abolish the vasodilation.The importance of the skin temperature for the ordinaryuse of the tail–flick test has been discussed (Roane etal. 1998). Clearly, when high doses of potent analgesicslike opioids are used, the relative influence of the skintemperature may be small. However, when the temper-ature influence is not known, this will always be an un-
predictableconfounding factor.Asdiscussed above, thishas, in several instances, lead to erroneous conclusions.
Possible Remedies
The temperature of the tail skin should always be con-sidered a possible confounding factor when performingthe tail–flick test. A minimal requirement should be thatthe tail skin temperature is measured before testing, e.g.by means of thermocouples (Fig. 1), thermistors or in-frared sensors, and the possible influence of the temper-ature evaluated.It is obviously necessary to take the effects of skintemperature into account when investigating factors,or using drugs that may influence autonomic activityand thermo – or cardiovascular regulation. Recordingthe tail skin temperature and correcting the tail–flicklatency data for changes in the temperature may reducethe problem. In some cases, a regression analysis or ananalysis of covariance may be performed for this pur-pose. Methods for tail–flick testing with measurementof skin temperature and for correction of tail–flick data(see � tail-flick latency correction)have been described(Tjølsen et al. 1989; Roane et al. 1998; Sawamura etal. 2002). However, there are some limitations whenusing this type of statistical analysis on tail–flick data.In these statistical methods, linearity of the relationshipis supposed. It seems to be a reasonable approximationto suppose linearity over a normal, limited range ofskin temperatures in untreated animals, e.g. 20–30oC.In studies where drug administration causes a large in-crease in tail–flick latency dueto changes innociception,the assumption of linearity may not be correct. Aboveall, these methods for statistical evaluation cannot ad-equately handle cut-off values for tail–flick latencies.This should be considered in each experiment, and evenwhen limitations as above are applicable, the tempera-ture of the skin of the tail should be measured and thepossible influence on the results should be evaluated.As a number of factors may possibly influence the rela-tionship between skin temperatureand response latency,it seems ideal to adjustdata from oneexperimentaccord-ing to the regression slope calculated from that experi-ment. However, this will not always be possible, in that aregression analysis requiresan adequatenumberofmea-surements to allow calculation of a reliable regressioncoefficient, and the spread of the independent variable(skin temperature) must be sufficiently large. If these re-quirements are not fulfilled, the results of the regressionanalysiswillbeinconclusive.Withanincreasingnumberof measurements in the analysis, there is an increasingprobability that a reliable regression coefficient may becalculated. In many cases, it may be a problem to ob-tain a reliable correction of tail–flick data based on thesame experiment, due to a limited number of animalsmeasured. An alternative method for correction of la-tencies is to establish the relationship between skin tem-perature and tail–flick latency in an adequate number of

T
Targeting 2395
animals under similar experimental conditions, and sub-sequently to correct tail–flick latencies according to thecalculated regression factor (Ren and Han 1979). Thismethod should be used with caution, because it mustbe assumed that the experiment is performed under thesame conditions as when the correction factor was de-termined. This is of course an approximation.Another alternative that has been used is local preheat-ing of the tail to a certain temperature before measuringthe tail–flick latency. If the temperatures of the skin andsubcutaneous tissue in the stimulated area are constantbefore the start of stimulation, this may abolish the con-founding effect of varying tissue temperatures. In elec-trophysiological experiments in anaesthetized cats, pre-heatinghasbeenusedwithaheating lampandafeedbackcontrolsystem,withathermocoupleontheareaofskintobe heated (Duggan et al. 1978). This procedure seemedto reduce the confounding effect of differences in bloodflow. For tail–flick reflex recordings, this technique maybe used in experiments in lightly anaesthetized animalsfor instance (Haws et al. 1990), or when the rat is placedin a restrainer and the tail is fixed (Carstens and Dou-glass 1995). It may probably be more difficult to use thismethod in animals that are awake when little restraint ofthe animal is required to minimize stress.When performed as described here, the tail–flick test isa reliable and useful test of nociception in rodents.
References1. Berge O-G, Garcia-Cabrera I, Hole K (1988) Response Latencies
in the Tail–Flick Test Depend on Tail Skin Temperature. NeurosciLett 86:284–288
2. Carstens E, Douglass DK (1995) Midbrain Suppression of LimbWithdrawal and Tail–Flick Reflexes in the Rat: Correlates withDescending Inhibition of Sacral Spinal Neurons. J Neurophys-iol 73:2179–2194
3. D’Amour FE, Smith DL (1941) A Method for Determining Lossof Pain Sensation. J Pharmacol Exp Ther 72:74–79
4. Duggan AW, Griersmith BT, Headley PM, Maher JB (1978) TheNeed to Control Skin Temperature when Using Radiant Heat inTests of Analgesia. Exp Neurol 61:471–478
5. Eide PK, Berge O-G, Tjølsen A, Hole K (1988) ApparentHyperalgesia in the Mouse Tail–Flick Test due to Increased TailSkin Temperature after Lesioning of Serotonergic Pathways.Acta Physiol Scand 134:413–420
6. Haws CM, Heinricher MM, Fields HL (1990) α-Adrenergic Re-ceptor Agonists, but not Antagonists, Alter the Tail–Flick La-tency when Microinjected into the Rostral Ventromedial Medullaof the Lightly Anesthetized Rat. Brain Res 533:192–195
7. Hole K, Tjølsen A (1993) The Tail–Flick and Formalin Tests inRodents: Changes in Skin Temperature as a Confounding Factor.Pain 53:247–254
8. Le Bars D, Gozariu M, Cadden SW (2001) Animal Models ofNociception. Pharmacol Rev 53:597–652
9. Milne RJ, Gamble GD (1989) Habituation to Sham Testing Pro-cedures Modifies Tail–Flick Latencies: Effects on Nociceptionrather than Vasomotor Tone. Pain 39:103–107
10. Ren MF, Han JS (1979) Rat Tail–Flick Acupuncture AnalgesiaModel. Chin Med J 92:576–582
11. Roane DS, Bounds JK, Ang C-Y, Adloo AA (1998) Quinpirole-Induced Alterations of Tail Temperature Appear as Hyperalgesiain the Radiant Heat Tail–Flick Test. Pharmacol Biochem Be-hav 59:77–82
12. Sawamura S, Tomioka T, Hanaoka K (2002) The Importance ofTail Temperature Monitoring during Tail–Flick Test in Evalu-ating the Antinociceptive Action of Volatile Anesthetics. ActaAnaesthesiol Scand 46:451–454
13. Tjølsen A, Hole K (1992) The Effect of Morphine on Core andSkin Temperature in Rats. NeuroReport 3:512–514
14. Tjølsen A, Lund A, Berge O-G, Hole K (1989) An ImprovedMethod for Tail–Flick Testing with Adjustment for Tail-SkinTemperature. J Neurosci Meth 26:259–265
15. Urban MO, Smith DJ (1994) Nuclei within the Rostral Ventro-medial Medulla Mediating Morphine Antinociception from thePeriaqueductal Gray. Brain Res 652:9–16
Talairach Coordinates
Definition
Initially developed for a specific stereotactic frame;based on one single brain; frequently used as a commoncoordinate system; X: left-right, Y: anterior-posterior,Z: superior-inferior; the reference point (0, 0, 0) is theanterior commissure (Talairach and Tournoux 1988).� Nociceptive Processing in the Secondary Somatosen-
sory Cortex
Tampa Scale for Kinesiophobia
Synonyms
TSK
Definition
The Tampa Scale for Kinesiophobia is a questionnaireaimedat theassessmentof fearof (re)injuryduetomove-ment, consisting of 17 items with 4–point likert-scales.� Disability, Fear of Movement
Tapotement
� Massage and Pain Relief Prospects
Targeting
� Trafficking and Localization of Ion Channels

2396 Tarsal Tunnel
Tarsal Tunnel
Definition
The anatomic structures that the tibial nerve passesthrough at the medial ankle are termed the tarsal tun-nel. Within this tunnel, the tibial nerve divides into themedial and lateral plantar and the calcaneal nerves,each of which has its own separate tunnel as it goesfrom the ankle to its final destination. These tunnelsrepresent sites of anatomic narrowing in which nervescan become entrapped, and which can give symptomsof chronic nerve compression in the foot. These can bepresent in the patient with a systemic neuropathy, suchas that due to diabetes.� Painful Scars� Ulceration, Prevention by Nerve Decompression
Task Force on Promotion andDissemination of PsychologicalProcedures
Definition
In 1993, Division 12 (Clinical Psychology) of the Amer-ican Psychological Association appointed a Task Force,with the goal of identifying and disseminating psycho-logical interventions that could be considered as empir-ically validated. In its 1995 report, this Task Force pub-lished criteria that allowed the classification of psycho-logical treatments as “well-established” and “probablyefficacious” In 1999, for its special issue on empiricallyvalidated treatments in pediatricpsychology, theJournalof Pediatric Psychology defined an additional categoryof “promising interventions” A treatment was consid-ered to bewell-established if therewereat least two goodbetween-group design experiments (or well-controlledsingle case studies) by at least two different investigatorsthat demonstrated the treatment’s efficacy over placebo,or at least equal efficacy as compared to an already es-tablished treatment. In addition, a treatment manual or awell defined treatment protocol needed to be available.A treatment was considered as “probably efficacious” iftherewereat least two experiments showing its superior-ity to a wait-list control, or if there was at least one study(or a small series of single-case designs) that met thewell-established treatment criteria. Finally, an interven-tion was considered “promising” if there was at least onewell-controlledandanother lesswellcontrolledstudybyseparate investigators, or a small number of single caseexperiments, or at least two well-controlled studies bythe same investigator.� Modeling, Social Learning in Pain� Psychological Treatment of Pain in Children
Task Force on Vicarious Instigation
Definition
Vicarious instigation describes the phenomenon that,possibly mediated by empathy, mere observation ofanother person’s response to a stimulus or a situation(e.g. a pain response) can induce a similar response inthe observer in the absence of any direct experiencewith the eliciting stimulus or situation. In the contextof pain, it is still a matter of debate whether observinganother person in pain can induce a pain-like vicariousresponse in the observer or whether it elicits a moregeneralized emotional response in the observer.� Modeling, Social Learning in Pain
TAUT
Definition
TAUT is a plasma membrane GABA transporter, whichtransports taurin with higher affinity than GABA.� GABA and Glycine in Spinal Nociceptive Processing
Taut Band
Definition
A taut band is a string- or cord-like structure in striatedmuscle that extends the length of the muscle fibers. Itconsists of a number of fascicles that are most palpableacross (at a right angle to) the fiber direction in the regionof fiber midpoints, where the myofascial trigger point islocated.Thetautbandis theresponsivepartof themusclein a local twitch response.� Myofascial Trigger Points
TaxonomyNIKOLAI BOGDUK
Royal Newcastle Hospital, Department of ClinicalResearch, University of Newcastle, Newcastle, NSW,[email protected]
Synonyms
Classification; Catalogue; List of Diagnoses and theirDefinitions

T
Taxonomy 2397
Definition
A � taxonomy is a catalogue that lists and classifies en-tities and provides definitions of them. It is like a dic-tionary restricted to a particular field of scholarship. Itis designed to standardize the meaning and use of par-ticular terms. In relation to pain medicine, a taxonomylists, classifies, and defines terms used to describe pain,and provides criteria for the use of diagnostic labels.
Characteristics
Two taxonomies have been produced for use in painmedicine. One, developed by the International Associa-tion for the Study of Pain (IASP), covers pain in general(Merskey and Bogduk 1994). The other, developed bythe International Headache Society, relates exclusivelyto headache (Headache Classification Subcommitteeof the International Headache Society 2004). A relatedtaxonomy – the Diagnostic and Statistical Manual ofMental Disorders (� DSM, DSM-IV, DSM-IVR), wasdeveloped by the American Psychiatric Associationand is designed to cover mental disorders, but includessome entries that potentially relate to pain (AmericanPsychiatric Association 2000).
IASP Taxonomy
The taxonomy of the IASP (Merskey and Bogduk 1994)consists of a short, introductory section devoted to thedefinition of terms used to describe pain, its differentforms, (such as � somatic pain, � Visceral Nociceptionand Pain, � referred pain, and � radicular pain), andits associated clinical features, such as � hyperalgesia,� allodynia, and � hyperpathia). The longer, moresubstantive section lists various entities that constitutepossible diagnoses for patients with � chronic pain.For each entity, criteria for making the � diagnosisare stipulated. The entities are catalogued and listedaccording to the region of the body that they affect.Conditions that affect the whole body, or which mayoccur in any region of the body, are described first,followed by conditions that affect the head, the neckand cervical spine, the upper limbs, the thoracic regionand thoracic spine, the abdomen and pelvis, the lumbarspine, and the lower limbs.The IASP Taxonomy was developed because it was rec-ognized that particular terms were being used indiscrim-inately by practitioners. Different practitioners were us-ing the same term to apply to different conditions, anddifferent terms were used to apply to the same condition.Practitioners were also applying different diagnostic la-bels to what were essentially the same patients, or wereapplying labels to patients that were not appropriate. Ineffect, the use of terms and diagnostic labels was arbi-trary. In 1979, Bonica likened the terminology for painsyndromes in use at that time to the “tower of Babel”(Bonica 1979).The first edition of the Taxonomy of the IASP (Merskey1986)listedcommonandrareconditionsassociatedwith
chronicpain,andprovideddefiningdescriptionsofeach.It allowed each condition to bedescribed along fiveaxes:The axis system, however, pertained mainly to the six-digit alphanumeric code ascribed to each condition. Theconditions themselves were classified largely accordingto Axis I, with only parenthetical mention of pathology,aetiology and other features, if these were known.The first edition of the Taxonomy was not intended tobe, or expected to be, comprehensive or fixed. Indeed,readerswere invited tosubmit revisions(Merskey1986).The second edition of the Taxonomy (Merskey andBogduk 1994) addressed many of the shortcomings ofthe first edition. Some descriptions were modernized,and involved a name change, e.g. � Reflex SympatheticDystrophy and � causalgia became � complex regionalpain syndrome Type I and Type II. Some entries weredeleted (e.g. prolapsed disc, osteophyte, spondylolysis,arachnoiditis, acute low back strain, recurrent low backstrain, and chronic mechanical low back pain) and werereplaced by more generic or alternative entries. Somenew entries were added, e.g. cervicogenic headache,xiphoidalgia, carcinoma of the lung, proctalgia fugax,piriformis syndrome, and peroneal muscular atrophy.The greatest revision pertained to entries on spinal pain.Some 96 new entries replaced all previous entries onneck pain, back pain, and other spinal pain. Moreover,the new entries were systematic and rigorous. Theywere designed to eliminate the problems of contentvalidity and of former entries.The new entries covered standard conditions such asspinal pain attributable to tumour, infection, metabolicdisease, and arthritis. Radicular pain, due to osteophyte,disc prolapse, cysts, tumours, etc, was strictly distin-guished and segregated from spinal pain on the groundsthat, although radicular pain might have a spinal aeti-ology, it was pain perceived in the limbs or trunk wallrather than in the spinal region.Perhaps the most comprehensive change was the intro-duction of the rubric – “spinal pain of unknown origin”.Users were invited, if not directed, to use this rubricwherever an alternative could not be legitimately, orhonestly, applied. Providing this rubric encouragedphysicians to avoid other poorly defined, invalid, orarbitrary rubrics, in an effort to reduce confusion andfalse labelling of patients.Nevertheless, other rubrics were offered. They cov-ered emerging entities, such as discogenic pain andzygapophysial joint pain, as well as classical entities,such as ligament strain and muscle strain, and allopathicentities such as segmental dysfunction. In providingthese entries, however, the Taxonomy stipulated strin-gent, essential diagnostic criteria, in order to avoidthe rubrics being applied on intuitive or presumptivegrounds.Thus, for “ligament strain” the ligament had to be spec-ified, and the diagnosis had to be proven with a test thatexplicitly showed that the ligament in question was the

2398 Taxonomy, Orofacial Pain
Taxonomy, Table 1 Taxonomy
Axis I Region Referred to the anatomical region in which the pain was perceived (e.g., head, abdomen, lower limb).
Axis II System Referred to the body system that ostensibly was affected by pathology to produce pain (e.g. nervous system,vascular system, musculoskeletal system
Axis II Temporal Described whether the pain was continuous, recurring, paroxysmal, etc.
Axis IV Intensity andDuration
Stated if the pain was mild, medium or severe; and lasted less than one month, between one and six months,or longer than six months.
Axis V Aetiology Stated the nature of the cause of the pain (e.g. infectious, inflammatory, and neuropathic).
source of pain. Similar criteria were applied for “musclestrain”. For “segmental dysfunction” the essential crite-ria required clinical tests of proven reliability, and estab-lished validity to implicate the specified segment as thesource of pain.Theserigorouscriteriawerestipulatedquitedeliberatelyin the full knowledge that the tests required to make thediagnosis did not (yet) exist. In effect, therefore, it wasimpossible to make the diagnosis in practice; yet it ap-peared in the Taxonomy. The purpose of this action wasto indicate to proponents of specific, but ill–defined, yetperhapspopular,diagnoses, thatresearchwasrequiredinorder for the entity to satisfy the standards of a responsi-ble Taxonomy, and for the diagnosis to be reliable, validand, therefore, respectable.
IHS Taxonomy
The taxonomy for headache catalogues the many formsof � headache according to mechanism or cause. Diag-nostic criteria are stipulated for each form of headache.These are designed to ensure that practitioners use a par-ticulardiagnostic labelonly in thosepatientswhoexhibitthe prescribed criteria.The taxonomy describes and defines those headacheswhose mechanism is not known but which have well-defined clinical features, such as � migraine, � clusterheadache, and � paroxysmal hemicrania. It contin-ues with descriptions and definitions of headachesassociated with particular circumstances (such as theheadaches of analgesic abuse, and rebound headache),headaches due to particular causes (such as raised orlowered pressure of cerebrospinal fluid, cerebral tu-mours, aneurysms, infections and granulomas), andheadaches associated with other disorders (such asdisorders of the ear, nose, and throat, or the cervicalspine) (see essay � headache).Theheadachetaxonomyiscomplementedbyatextbook,nowin its second edition (Olesen etal. 2000),witha thirdedition in preparation. The textbook follows the formatof the taxonomy, but provides descriptions, in detail, ofthe entities and their diagnosis and treatment.
References1. American Psychiatric Association (2000) DSM-IV-TR. Diagnos-
tic and Statistical Manual of Mental Disorders, 4th edn, TextRevision. American Psychiatric Association, Washington DC
2. Bonica JJ (1979) The Need for a Taxonomy. Pain 6:247–2523. Headache Classification Subcommittee of the International
Headache Society (2004) The International Classification ofHeadache Disorders, 2nd edn. Cephalalgia 24 Suppl 1:1–160
4. Merskey H (ed) (1986) Classification of Chronic Pain. Descrip-tions of Chronic Pain Syndromes and Definition of Pain Terms.Pain Suppl 3:S1–S225
5. Merskey H, Bogduk N (1994) Classification of Pain. Descriptions
of Chronic Pain Syndromes and Definitions of Pain Terms, 2nd
edn. International Association for the Study of Pain, Seattle, pp64–65
6. Olesen J, Tfelt-Hansen P, Welch KM (2000) The Headaches, 2nd
edn. Lippincott Williams & Wilkins, Philadelphia
Taxonomy, Orofacial Pain
� Orofacial Pain, Taxonomy/Classification
TCAs
� Tricyclic Antidepressants
TCD
� Thalamocortical Dysrhythmia
Team Approach
� PhysicalMedicineandRehabilitation,Team-OrientedApproach
Technique of Ultrasound Application
Definition
The most common technique is the stroking technique.� Ultrasound Therapy ofPain from theMusculoskeletal
System

T
Temporomandibular Joint 2399
Tegretol
Synonyms
Generic carbamazepine
Definition
Tegretol (Generic Carbamazepine) is an anti-epilepticdrug acting at non-voltage dependent sodium channels,which is so effective in the treatment of cranial neural-gias that lack ofa (at least transient) response to thismed-ication raises a significant question about the diagnosis.� Trigeminal, Glossopharyngeal, and Geniculate Neu-
ralgias
Telemetric
Definition
Telemetric means the transmission of data by radio orother means from a remote source.� Opioid Therapy in Cancer Pain Management, Route
of Administration
Temperament
� Personality and Pain
Temporal Arteritis
Definition
Temporal arteritis is an arterial disease with inflam-mation of the temporal arteries characterized by fever,anorexia, loss of weight, leukocytosis, and tendernessover the scalp and along the temporal vessels. The giantcell arteritis most often attacks the external arteries inthe anterior skull region – branches from the arteriacerebri externa.� Cancer Pain, Assessment in the Cognitively Impaired� Muscle Pain in Systemic Inflammation (Polymyalgia
Rheumatica, Giant Cell Arteritis, Rheumatoid Arthri-tis)
Temporal Association
Definition
Temporal association between two disorders or clinicalproblems refers to their hypothesized relationship interms of time of onset, most often inferring a causal orcontributory relationship.� Depression and Pain
Temporal Resolution
Definition
The value indicates how reliable the results are in termsof the time period. The higher, the better, and EEG andMEG are much higher than fMRI and PET.� Magnetoencephalography in Assessment of Pain in
Humans
Temporal Summation (Windup)
Definition
When synaptic potentials overlap in time, they add to-gether. In this case, repeated administration of the samestimulus, at a given interval of time, produces a progres-sively increased painful response. Temporal summationis probably the initial part of wind-up, which is the in-creased neuronal firing to a train of stimuli recorded inanimals.� Encoding of Noxious Information in the Spinal Cord� Exogenous Muscle Pain� Opioids and Muscle Pain� Opioids, Effects of Systemic Morphine on Evoked
Pain� Pain in Humans, Electrical Stimulation (Skin, Muscle
and Viscera)
Temporomandibular Disorder
Synonyms
TMD
Definition
A collective term embracing a number of clinical prob-lems that involve the masticatory musculature, the tem-poromandibular joint and associated structures, or both.Temporomandibular disorders have been identified as amajor cause of nondental pain in the orofacial region,and are considered to be a subclassification of muscu-loskeletal disorders.� Orofacial Pain, Movement Disorders� Orofacial Pain, Taxonomy/Classification� Psychological Aspects of Pain in Women
Temporomandibular Joint
Definition
The jaw joint. The joint formed between the condylarprocess of the mandible and the mandibular fossa andarticular tubercle of the temporal bone.

2400 Temporomandibular Joint and Muscle Pain Dysfunction
� Nociceptors in the Orofacial Region (Temporo-mandibular Joint and Masseter Muscle)
� Psychiatric Aspects of Pain and Dentistry� Temporomandibular Joint Disorders
Temporomandibular Joint and MusclePain Dysfunction
� Temporomandibular Joint Disorders
Temporomandibular Joint DisordersCHRISTIAN S. STOHLER
Baltimore College of Dental Surgery, University ofMaryland, Baltimore, MD, [email protected]
Synonyms
Temporomandibular joint disorders (TMJDs); Tem-poromandibular disorders (TMDs); CraniomandibularDisorders; Previously used diagnostic labels; Tem-poromandibular Joint and Muscle Pain Dysfunction;TMJD
Definition
Temporomandibular disorders (TMJDs) comprise of afamily of musculoskeletal conditions that involve deepacheorpainintheareaofthetemporomandibular joint(s)and/ or adjacent tissue structures. These conditions con-stitute a major source of non-dental pain in the cranio-facial complex.
Characteristics
Etiology and Pathogenesis
Although the etiology of these musculoskeletal paindisorders is not established and various pathogeneticconstructs have been proposed, these conditions arebelieved to develop from the combined action of manygenes, risk-conferring behaviors and environmentalfactors. The fact that pain originates in deep tissueappears to be relevant to understanding the clinicalphenomenon because, unlike superficial pain, deeppain is poorly localized and frequently associated withpronounced autonomic reactions. Genetic vulnerabilityis attributed to differences in the genetic makeup thatenhance, directly or indirectly, pro-nociceptive and/ orattenuate anti-nociceptive signalling (Fig. 1).Earlier etiological constructs have placed significantweight on the dental occlusion as a causal factor in theetio-pathogenesis of TMJDs. However, low strengths
Temporomandibular Joint Disorders, Figure 1 Etiological construct.
of association between occlusal features and TMJDs,inconsistent findings from study to study regarding therole of a given occlusal attribute, and the absence ofany gradient effect of occlusal factors put these earliertheories in question.
Case Assignment
Although research in this subject matter has been in-tensified in recent years, no biomarkers of exposure oreffect are established for valid and reliable TMJD caseascertainment. TMJD case assignment occurs on thebasis of clinical features, which consist of symptomslike pain and limited range of mandibular motion. Fa-cial pain reports focus on anatomical regions such asthe temples, cheeks, pre-auricular area, or inside theear and vary in intensity and spatial distribution, bothinter-individually and intra-individually, with time.With respect to corresponding clinical signs, allody-nia in the form of tenderness to palpation is linked topainful topographical sites. Limited range of motionis often noted and attributed to factors such as the ar-ticular disc preventing smooth gliding movement ofthe mandibular condyle along the articular eminence,constraining mandibular excursion, and/ or the recruit-ment of jaw closing muscles during their function asantagonists, limiting mandibular side-to-side excur-sions and the capacity to open the jaw fully. However,under no circumstances should observable signs beused in isolation to define a TMJD case, because ofinsufficient diagnostic validity due to high sensitivityand low specificity.
Classification Systems
Important indirectingclinical researchinthepastdecadewere efforts to produce a dual axes taxonomy for the ma-jor types of TMJDs (Dworkin and LeResche 1992). Fo-cusing on the craniofacial domain, Axis I distinguishesthree main diagnostic subsets (Fig. 2):

T
Temporomandibular Joint Disorders 2401
Temporomandibular JointDisorders, Figure 2 Overview ofthe diagnostic construct adoptedby the Research Diagnostic Criteriafor temporomandibular disorders.(For detail see Dworkin andLeResche 1992).
Temporomandibular JointDisorders, Figure 3 Overlap ofTMJDs with regional and systemicdisorders.
1. Group I: Masticatory myofascial pain2. Group II: TMJ internal derangements3. Group III: TMJ arthritides
Axis II criteriaassesspain intensity,pain-relateddisabil-ity, and the presence and severity of depressive and anx-iety symptoms. Using this classification scheme, abouthalf of all TMJD cases are identified as Group Idisorders(List and Dworkin 1996).Due to the overlap with regional myofascial pain,tension-type headache, fibromyalgia, polyarthritidesand possibly connective tissue disorders with impairedcollagen makeup, shortcomings of available TMJD tax-onomies are becoming increasingly recognized (Fig. 3).The fact that persistent TMJDs are rarely limited to asingle topographical domain underscores the need toassess these conditions in the broader context.
Phenomenology
General Characteristics
Poorly localizable ache or pain unrelated to dentalpathology, constitute the chief complaint of all major
forms of TMJDs. The sensory experience is capturedby pain descriptors, such as “aching”, “tight”, “throb-bing” and “tender” (Turp et al. 1997). Besides pain, (a)inability to freely move the jaw due to pain and/ or softor hard tissue interference, (b) sounds originating fromthe jaw joint, and (c) the disturbing perception of teethnot fitting properly constitute the other shared concerns.With respect to clinically observable signs, pressureallodynia, the experience of pain in response to definedpressure that is rarely identified as painful by subjectswithout TMJDs, represents the clinical hallmark fea-ture of this family of pain conditions. Inability to movethe jaw freely is determined by measurements of themandibular range of motion and expressed by the clini-cally observable maximum mandibular excursions in alldirections. Joint sounds are often linked to mechanicalevents between moving articular structures, such as thetemporal component of the TMJ, the articular disc andthe condyle. With respect to age, prevalence rates arelower among older subjects, and initial care-seeking inboth men and women is more likely to occur before age50 than later in life.

2402 Temporomandibular Joint Disorders
Temporomandibular JointDisorders, Figure 4 Cases(in %) reporting pain in a givendermatome. Adapted from (Turp,Kowalski, O’Leary, and Stohler,1998).
Spatial Characteristics
Not only can temporomandibular joint (TMJ) arthriditesbe part of an existing polyarthritis that affects additionaljoints other than the TMJs, those TMJDs that involvemuscle differ in the extent of their bodily involvement aswell. Distinction of local and widespread phenomena isimportant, because cases with widespread pain are morelikely in pain on follow-up examination than cases withlocalized pain (Raphael et al. 2000).In contrast to TMJD,musclepain conditions that involvethe face and adjacent head or neck regions, fibromyalgia(FMS) is understood as a clinical entity characterized bypersistentwidespreadpainandtendernessto4kilogramsof pressure at 11 of 18 anatomically defined body sites(Wolfe et al. 1990). Overlap between TMJDs and FMShas been demonstrated in a number of studies (Plesh etal. 1996; Hedenberg- Magnusson et al. 1997; Korszunet al. 1998). According to Plesh and coworkers, 75% oftheir FMS patients had TMJDs, while, on the other hand,18% of cases with TMJDs met the diagnostic criteriafor FMS. Epidemiological studies also report high as-sociations between TMJDs and the two most commontypes of headache, tension-type headache and migraineheadache (Agerberg and Carlsson 1973). In fact, per-sistent TMJD pain is associated with co-morbid pain inbody parts other than the face at much greater rates thanthe condition is limited to the face (Fig. 4) (Turp et al.1998).
Temporal Characteristics
Complaints of pain range from a local response tosimple injury to complaints of persistent widespreadbodily involvement without obvious cause. From a
phenomenological point of view, it needs to be em-phasized that the overwhelming case majority seen inthe primary care setting exhibits episodic forms, whilecases encountered in the tertiary care environment aremore likely affected by persistent conditions. The factthat the personally most devastating and clinically mostchallenging TMJD presentations occur in females ingreater numbers than males, results in up to 90% oftertiary care cases being women (Figure 5). Amongwomen, prevalence rates are higher for subjects ofreproductive age than those in postmenopausal yearswithout hormone replacement therapy (LeResche et al.1997).TMJD pain is characterized as non-progressive andfluctuating in intensity, which is often translated into“good” and “bad” days. What is applicable to a widerange of pain disorders seems also to be the case forTMJDs. As a generalization, infrequent pains of evenhigh intensity are more likely perceived as a nuisancewhen compared to persistent pain of lesser intensity.On the other hand, persistent pain disrupts the lifestyle,causing functional limitations and restrictions in dailyactivities. In this context, it is increasingly understoodthat time in pain influences the subject’s physiologicalstate and response behavior. Initial pain constitutes awarning signal, causing the subject to stop the ongoingactivity and to take actions to alleviate the pain. If painpersists, longer lasting effects on neuronal excitability,such as the up-regulation of NMDA-mediated effectsand changes in the CNS “hardwiring” occur via a seriesof events and involve alterations in intermediate andlate gene expressions. Binding of c-fos and c-jun toDNA alters the transcription of intermediate and even-

T
Temporomandibular Joint Disorders 2403
Temporomandibular JointDisorders, Figure 5 Comorbidconditions and male-to-femaleratios in different observationalsettings.
tually late effector genes, which in turn affect enzymes,growth factors, peptides, and even the phenotype.
Pain Affect
Prolonged and persistent pain can induce significantpain affect, which in itself constitutes an integral partof the TMJDs. Pain affect is captured by pain descrip-tors, such as “tiring”, “exhausting”, “frightening” and“fearful”. Great variations are observed with respectto the degree to which pain affect is expressed frompatient to patient, even within a given TMJD subset(Ohrbach and Dworkin 1998). Much of the variabilityin response to pain is believed to be of genetic origin.Consequently, intense research is beginning to iden-tify the allelic variants that underlie these responsedifferences. For example, a 3- to 4-fold reduction inthe activity due to a valine-methionine polymorphismof catechol-O-methyltransferase (COMT), an enzymethat catalyzes the O-methylation of compounds with acatechol structure, results in less or greater than normalavailability of catecholamines at the site of neurotrans-mission, which in turn significantly shapes the sensoryand affective experience of facial pain (Zubieta et al.2003).
Management
Because the causal sequence of events that leads topain and dysfunction is not known, therapeutic inter-ventions focus on symptom management rather than onthe elimination of the cause. Patients who seek care forthe first time, report symptom relief of TMJD by 65-95%. Treatments include thermal packs, non-steroidalanti-inflammatory drugs (NSAIDs) and/or muscle re-laxants, inter-occlusal appliances, physical therapy,relaxation and stress management, and acupuncture
and diet counselling to mention the most commoninterventions. There are little differences among thevarious types with respect to symptom relief. Thosepatients that do not get a satisfactory outcome, whichhappen to constitute a clear case minority in the primarycare setting, are characterized by persistent pain anddysfunction for which all current forms of treatmentfall short. Given the questionable superiority of onetype of intervention over another, the choice of careis more influenced by unwanted effects attributable tothe intervention, and/or the greater cost for care thatdoes not translate into a justifiable improvement of thetherapeutic efficacy. Consequently, case managementtends to be “conservative” and “reversible”.
References1. Agerberg G, Carlsson GE (1973) Functional Disorders of the
Masticatory System. II. Symptoms in Relation to Impaired Mo-bility of the Mandible as Judged from Investigation by Ques-tionnaire. Acta Odontol Scand 31:337–347
2. Dworkin SF, LeResche L (1992) Research Diagnostic Criteria forTemporomandibular Disorders: Review, Criteria, Examinationsand Specifications, Critique. J Craniomandib Disord 6:301–355
3. Hagberg C, Hagberg M, Kopp S (1994) MusculoskeletalSymptoms and Psychosocial Factors Among Patients withCraniomandibular Disorders. Acta Odontol Scand 52:170–177
4. Hedenberg-Magnusson B, Ernberg M, Kopp S (1997) Symptomsand Signs of Temporomandibular Disorders in Patients with Fi-bromyalgia and Local Myalgia of the Temporomandibular Sys-tem. A comparative study. Acta Odontol Scand 55:344–349
5. Korszun A, Papadopoulos E, Demitrack M, Engleberg C, Crof-ford L (1998) The Relationship Between TemporomandibularDisorders and Stress-Associated Syndromes. Oral Surg Oral MedOral Pathol Oral Radiol Endod 86:416–420
6. LeResche L, Saunders K, Von KM, Barlow W, Dworkin SF(1997) Use of Exogenous Hormones and Risk of Temporo-mandibular Disorder Pain. Pain 69:153–160
7. List T, Dworkin SF (1996) Comparing TMD Diagnoses and Clin-ical Findings at Swedish and US TMD Centers using Research

2404 Temporomandibular Pain
Diagnostic Criteria for Temporomandibular Disorders. J OrofacPain 10:240–253
8. Ohrbach R, Dworkin SF (1998) Five-Year Outcomes in TMD:Relationship of Changes in Pain to Changes in Physical and Psy-chological Variables. Pain 74:315–326
9. Plesh O, Wolfe F, Lane N (1996) The Relationship Between Fi-bromyalgia and Temporomandibular Disorders: Prevalence andSymptom Severity. J Rheumatol 23:1948–1952
10. Raphael KG, Marbach JJ, Klausner J (2000) MyofascialFace Pain. Clinical Characteristics of those with Regional vs.Widespread Pain. J Am Dent Assoc 131:161–171
11. Turp JC, Kowalski CJ, O’Leary TJ, Stohler CS (1998) Pain Mapsfrom Facial Pain Patients Indicate a Broad Pain Geography. J DentRes 77:1465–1472
12. Turp JC, Kowalski CJ, Stohler CS (1997) Pain Descriptors Char-acteristic of Persistent Facial Pain. J Orofacial Pain 11:285–290
13. Wolfe F (1997) The Relation Between Tender Points and Fi-bromyalgia Symptom Variables: Evidence that Fibromyalgia isnot a Discrete Disorder in the Clinic. Ann Rheum Dis 56:268–271
14. Wolfe F, Smythe HA, Yunus MB, Bennett RM, Bombardier C,Goldenberg DL, Tugwell P, Campbell SM, Abeles M, Clark Pet al. (1990) The American College of Rheumatology 1990 Cri-teria for the Classification of Fibromyalgia. Report of the Mul-ticenter Criteria Committee [see comments]. Arthritis Rheum33:160–172
15. Zubieta JK, Heitzeg MM, Smith YR, Bueller JA, Xu K, Xu Y,Koeppe RA, Stohler CS, Goldman D (2003) Genotype AffectsMu-Opioid Neurotransmitter Responses to a Pain Stressor. Sci-ence 299:1240–1243
Temporomandibular Pain
Definition
Chronic pain in the jaw muscles and TM joint, often as-sociated with malocclusion; also referred to as cranio-mandibular or temporo-mandibular dysfunction.� Jaw-Muscle Silent Periods (Exteroceptive Suppres-
sion)
Tender PointsMICHAEL SPAETH
Friedrich-Baur-Institute, University of Munich,Munich, [email protected]@lrz.uni-muenchen.de
Synonyms
TePsFormerly often used as synonymous: trigger points (butnowadays clearly discriminated)
Definition
If an individual reports local pain when a site is palpatedwith standardized pressure, this is considered a positive“tender point” (TP).
Characteristics
Anatomy
The anatomic TP sites do not appear to represent a sin-gle type of anatomical structure, but rather can includeligaments, tendons, skeletal muscles and bursae. TheTP hurts at the site where pressure is applied, only,whereas pain induced by pressure at a myofascial painsyndrome (MPS) “trigger point“ causes both local painand pain at a more distant area of reference (“referredpain“). Several efforts have been made to find a pri-mary origin of fibromyalgia (FM) pain at the anatomicsites themselves (Bengtsson et al. 1986; Drewes et al.1993; Henriksson et al. 1982; Yunus and Kalyan Ra-man 1989). In fact, most of these investigations studiedskeletal muscle exclusively and did not report any find-ings on other anatomical structures composing the TPregions. Morphological findings in skeletal muscle tis-sue specimens from FM patients are rather non-specificand presumably secondary to pain-related reductionof activity. Results of image analysis quantification ofsubstance P immunoreactivity in the trapezius mus-cle of patients with fibromyalgia and myofascial painsyndrome pointed to a peripheral hyperactivity of thepeptidergic nervous system in FM as well as in MPS (DeStefano et al. 2000). Recently reported ultrastructuralchanges in fibromyalgic muscle fibers may contributeto the induction and / or chronicity of nociceptive trans-mission from muscle to the central nervous system(Sprott et al. 2004). But, these alterations could not beidentified as a primary cause of hyperalgesia in FM TPareas.
Clinical Characteristics
Application of pressure on each of the TPs often inducespatient’s involuntary withdrawal. After the examinationof all 18 TPs, patients may report a persisting “deepache“, similar to that of bone pain. Some patients mayshow the symptoms with either one half (upper or lower)or one side (right or left) of the body preponderating.For standardization and for research purposes, pressuregauges are available. A dolorimeter is commonly usedand can further help to standardize the amount of pres-sure (e.g. 4 kg) applied by the examining finger of eachinvestigator.
Recent Objections
Despite the lack of information on what TPs really domeasure, research over the last few years has broughtup some interesting findings about these points. Thereis evidence from different studies, that FM patients aretenderer inbothTPandnon-TPregionsthanhealthycon-trol subjects and that these TP regions represent areas,where anyone is tenderer. TPs were revealed not to bespecific to FM (Granges and Littlejohn 1993b; Tunks etal. 1995).TPs were studied in a random sample from adults in thegeneral population. As a result, tenderness to pressure

T
Tendon Sheath Inflammation 2405
was found to occur both in people without widespreadpain and in people without any pain. The investigatorsfound that TP counts were increased in people who hadother symptoms (i.e. poor sleep and / or fatigue), even ifthey did not complain about pain at all (MacFarlaneet al.1996). Data suggest that TP counts can discriminate be-tween tender and non-tender individuals and can there-fore be considered as a clinically useful measure of ten-derness (Gracely et al. 2003). From another study, it wasconcluded that the tender point count was associated notonly with the extent of rheumatic pain, but also indepen-dently with the extent of bodily complaints (Schochatand Raspe 2003). Significant correlations were foundbetween TP count and psychological distress, evaluatedby analyzing somatic and depressive symptoms (Croftet al. 1994). Another study found that TP pain severityratings produced higher correlations with symptoms ofFM and predicted distress better than TP counts (Mc-Carberg et al. 2003).Since the TP count seems to be a composite measureof at least tenderness and psychological distress, it is oflimited value in research settings but useful in a clinicalsetting in order to recognize the tenderness-distress na-ture of FM (Gracely et al. 2003). Furthermore, the TPcount does not reflect differences in distress or pressure-pain sensitivity or provide help in subgrouping FM pa-tients (Giesecke et al. 2003). Another study replicatedprevious findings in population-based samples showingthat dolorimeter determinations are less influenced bypsychological factors than TP counts (Croft et al. 1994;Granges and Littlejohn 1993a), but there was still an im-pact of distress even on dolorimetry results (Petzke et al.2003).Most of these data-based objections were followed byrecommendations: (1) to re-consider the current defi-nition of FM, (2) to be aware of the tenderness-distressnature of both FM and TPs and (3) to re-evaluate chronicwidespread pain (CWP) in further population-basedstudies both to potentially discriminate between CWPand FM and to emphasize FM characteristics.� Muscle Pain, Fibromyalgia Syndrome (Primary, Sec-
ondary)
References1. Bengtsson A, Henriksson KG, Larsson J (1986) Muscle biopsy
in primary fibromyalgia. Light-microscopical and histochemicalfindings. Scand J Rheumatol 15:1–6
2. Croft P, Schollum J, Silman A (1994) Population study oftender point counts and pain as evidence of fibromyalgia. Bmj309:696–699
3. De Stefano R, Selvi E, Villanova M et al. (2000) Image analysisquantification of substance P immunoreactivity in the trapeziusmuscle of patients with fibromyalgia and myofascial pain syn-drome. J Rheumatol 27:2906–2910
4. Drewes AM, Andreasen A, Schroder HD et al. (1993) Pathologyof skeletal muscle in fibromyalgia: a histo-immuno-chemical andultrastructural study. Br J Rheumatol 32:479–483
5. Giesecke T, Williams DA, Harris RE et al. (2003) Subgroupingof fibromyalgia patients on the basis of pressure-pain thresholdsand psychological factors. Arthritis Rheum 48:2916–2922
6. Gracely RH, Grant MA, Giesecke T (2003) Evoked pain mea-sures infibromyalgia. BestPract ResClin Rheumatol 17:593–609
7. Granges G, Littlejohn G (1993a) Pressure pain threshold in pain-free subjects, in patients with chronic regional pain syndromes,and in patients with fibromyalgia syndrome. Arthritis Rheum36:642–646
8. Granges G, Littlejohn GO (1993b) A comparative study of clin-ical signs in fibromyalgia / fibrositis syndrome, healthy and ex-ercising subjects. J Rheumatol 20:344–351
9. Henriksson KG, Bengtsson A, Larsson J et al. (1982) Musclebiopsy findings of possible diagnostic importance in primary fi-bromyalgia (fibrositis, myofascial syndrome). Lancet 2:1395
10. MacFarlane GJ, Croft PR, Schollum J et al. (1996) Widespreadpain: is an improved classification possible? J Rheumatol23:1628–1632
11. McCarberg B, Barkin RL, Wright JA et al. (2003) Tender pointsas predictors of distress and the pharmacologic management offibromyalgia syndrome. Am J Ther 10:176–192
12. Petzke F, Gracely RH, Park KM et al. (2003) What do tenderpointsmeasure?Influence ofdistresson 4measuresof tenderness.J Rheumatol 30:567–574
13. Schochat T, Raspe H (2003) Elements of fibromyalgia in an openpopulation. Rheumatology (Oxford) 42:829–835
14. Sprott H, Salemi S, Gay RE et al. (2004) Increased DNA fragmen-tation and ultrastructural changes in fibromyalgic muscle fibres.Ann Rheum Dis 63:245–251
15. Tunks E, McCain GA, Hart LE et al. (1995) The reliability of ex-amination for tenderness in patients with myofascial pain, chronicfibromyalgia and controls. J Rheumatol 22:944–952
16. Yunus MB, Kalyan Raman UP (1989) Muscle biopsy findings inprimary fibromyalgia and other forms of nonarticular rheuma-tism. Rheum Dis Clin North Am 15:115–134
Tenderness
Definition
Tenderness describes a feeling of discomfort or paincaused by pressure that would normally be insufficientto cause such sensations.� Headache, Episodic Tension Type
Tendinitis
Definition
Tendinitus isapainful tendon,usually resultingfrom un-accustomed physical activity. Classified as a localizedSTP. Fraying and thickening of the tendon may be ob-served.� Ergonomics Essay� Muscle Pain, Fibromyalgia Syndrome (Primary, Sec-
ondary)
Tendon Sheath Inflammation
Definition
Tendon sheaths have synovial lining cells, which are in-cluded in the inflammation in rheumatoid arthritis.

2406 Tenosynovitis
� Muscle Pain in Systemic Inflammation (PolymyalgiaRheumatica, Giant Cell Arteritis, Rheumatoid Arthri-tis)
Tenosynovitis
Definition
Tenosynovitis refers to inflammation of the tendonsheaths, through which the tendons slide when themuscle length changes. Excessive fluid accumulationcan cause swelling and pain in the affected areas.� Ergonomics Essay
TENS
� Transcutaneous Electrical Nerve Stimulation
TENS, Mechanisms of ActionKATHLEEN A. SLUKA
Physical Therapy and Rehabilitation Science GraduateProgram, University of Iowa, Iowa City, IA, [email protected]
Synonyms
PES; electrical stimulation analgesia; transcutaneouselectrical nerve stimulation
Definition
Electrical stimulation applied to the skin for pain relief.
Characteristics
The mechanisms of action of � TENS primarily involvecentral mechanisms and have been extensively reviewed(see Sluka and Walsh 2003 for more details and refer-ences). There are generally two types of TENS appliedclinically, low frequency (<10 Hz) and high frequency(>50 Hz). These can be applied at either a sensory inten-sity that produces a tapping or tingling sensation or atmotor intensity that produces an additional motor con-traction. The mechanisms of action for TENS appear tobe frequency, not intensity, dependent.
High Frequency (50–100 Hz) TENS
Effects on Behavior and Dorsal Horn Neurons
Early studies utilizing acute pain tests show that highfrequency, motor intensity TENS increases the tail flicklatency to heat (i.e. analgesia) and decreases the flexionreflex response to noxious stimuli (reviewed in Slukaand Walsh 2003). Recording from spinothalamic tractcells, stimulation at an intensity activating Aβfibers (3 ×the threshold) has no effect on the spontaneous firing
rate. However, increasing the intensity so as to also ac-tivate Aδ nociceptors reduces spontaneous activity andresponses to noxiousheatorpinch(Leeetal.1985).Sim-ilarly, studies by Garrison and Foreman (1997) and bySjolund (1985) both show that increasing intensity in-creases inhibition of dorsal horn neurons and the flexionreflex response to noxious stimuli. These data suggestthat high and low frequency TENS are effective and thatincreasing intensity increases inhibition.Utilizing an animal model of joint inflammation re-veals that high frequency, sensory intensity TENS haslong-lasting effects on both primary and secondaryheat and mechanical � hyperalgesia (reviewed in Slukaand Walsh 2003) (Fig. 1). In fact, these studies showthat high frequency, sensory intensity partially reversesthe primary hyperalgesia and completely reverses thesecondary hyperalgesia associated with � carrageenaninflammation for 24 h. Importantly, modulation offrequency (4 Hz vs. 100 Hz), intensity (sensory vs.motor) or pulse duration (100 μs vs. 250 μs) showsa frequency, but not intensity or pulse duration, de-pendent effect on primary hyperalgesia to mechanicaland heat stimuli in animals with carrageenan paw in-flammation. The increased responsiveness of dorsalhorn neurons to innocuous and noxious mechanicalstimuli that occurs after inflammation is completelyreduced following high frequency, sensory intensityTENS treatment applied to the inflamed paw (Ma andSluka 2001). Utilizing a � model of neuropathic pain,Somers and Clemente (1998) demonstrated that highfrequency, sensory intensity TENS stimulation overthe paraspinal musculature reduced the heat but not themechanical hyperalgesia that normally occurs in thismodel. This inhibition of heat hyperalgesia only occursif TENS was started the first day after injury but not ifit was started 3 days after injury.
Pharmacology
In animals that were spinalized to remove descendinginhibitory pathways (Fig. 2), inhibition of the tail flickby high frequency, motor intensity TENS still occursbut is reduced by about 50% (Woolf et al. 1980). Thus,these studies suggest both spinal and � descendinginhibition are involved in the analgesia produced byhigh frequency, motor intensity TENS. Later studiesprevented the antihyperalgesia, by blockade of δ-opioidreceptors in the rostral ventral medial medulla (RVM),further supporting a role for descending inhibitorysystems in the inhibition produced by TENS.Pharmacologically, opioid peptides mediate the ef-fects of high frequency TENS. Concentrations ofbeta-endorphins increase in the bloodstream and cere-brospinal fluid and methionine-enkephalin increases inthe cerebrospinal fluid of human subjects, following ad-ministration of high frequency, sensory intensity TENS(reviewed in Sluka and Walsh 2003). High frequency,motor intensity TENS is blocked by systemic block-

T
TENS, Mechanisms of Action 2407
TENS, Mechanisms of Action, Figure 1 Effects of TENS on primary and secondary, mechanical and heat hyperalgesia induced by carrageenaninflammation. High, but not low, frequency TENS partially reverses primary hyperalgesia to heat and mechanical stimuli induced by carrageenan pawinflammation (left panels). In contrast, both high and low frequency TENS reverse secondary hyperalgesia induced by carrageenan knee joint inflammation(right panels).
TENS, Mechanisms of Action,Figure 2 Schematic drawingdemonstrating that TENS applied tothe periphery at the site of injuryactivates primary afferent fibers.This information is transmitted to thespinal cord and results in inhibitionboth locally and from descendinginhibitory pathways. Descendinginhibition from the rostral ventralmedial medulla (RVM) involves 5-HTand opioids and can be activatedby the periaqueductal gray (PAG).Previous studies show that opioidreceptors in the spinal cord and RVMand serotoninergic and muscarinicreceptors in the spinal cord mediatethe reduction in hyperalgesia byTENS.
ade of opioid receptors with naloxone and systemicdepletion of serotonin (reviewed in Sluka and Walsh2003). Blockade of δ-opioid receptors in the spinalcord or the rostral ventral medial medulla (RVM) re-verses the antihyperalgesia produced by high frequency
sensory intensity TENS in animals with carrageenanknee joint inflammation (Fig. 3) (Kalra et al. 2001;Sluka et al. 1999). Similarly, spinal δ-opioid recep-tors are implicated in the antihyperalgesic effects ofhigh frequency motor intensity TENS, since repeated

2408 TENS, Mechanisms of Action
TENS, Mechanisms of Action, Figure 3 Summary bar graph of the ef-fects of blockade of spinal receptors on the antihyperalgesia producedby low and high frequency TENS. Approximately 100% inhibition of hy-peralgesia occurs after treatment with either high or low frequency TENS(saline, purple). Blockade of μ-opioid (naloxone, dark blue) and muscarinic(atropine, red) receptors prevents the antihyperalgesia produced by highfrequency TENS. Blockade of spinal δ-opioid (naltrindole, blue), serotonin(methysergide, green) or muscarinic (atropine, red) receptors prevents theantihyperalgesia produced by low frequency TENS. Spinal blockade of α-2adrenergic (yohimbine, yellow) or nicotinic (mecamylamine, orange) recep-tors has no effect on the effects of either high or low frequency TENS.
application of high frequency, motor intensity TENSproduces tolerance (reduced effectiveness) to the an-tihyperalgesic effects of TENS and at spinal δ-opioidreceptors (Chandran and Sluka 2003).Further, blockade of muscarinic receptors (M1 and M3,but not M2) in the spinal cord also partially reverses theantihyperalgesia produced by high frequency, sensoryintensity TENS (Radhakrishnan and Sluka 2003). How-ever, blockade of serotonin or noradrenergic receptorsin the spinal cord has no effect on the reversal of hyper-algesia produced by high frequency, sensory intensityTENS (Radhakrishnan et al. 2003) (Fig. 3).
Autonomic and Peripheral Effects
TENS could have effects on autonomic function, bloodflow and peripheral afferent fibers (reviewed in Slukaand Walsh 2003). However, high frequency, sensory in-tensity TENS stimulation at intensities just above or be-low motor threshold does not affect local blood flow. Incontrast, utilizing laser Doppler imaging, increases inblood flow were observed with high frequency TENS,at an intensity “that was felt but not painful (10–15 mA).In human subjects, after application of high frequencyTENS at the threshold for discomfort (strong motor in-tensity applied to a digit), subjects report numbness andcooling justdistal to thestimulation (on thedigit).This isassociated with decreased temperature and loss of colorin the skin suggesting effects on the autonomic nervoussystem. The primary afferent neuropeptide, substanceP, is reduced in dorsal root ganglia neurons and spinalcord dorsal horn by high frequency, sensory intensityTENS in animals injected with the inflammatory irri-tant, formalin. Thus, evidence is beginning to emergethat some of the analgesic effects of TENS may be medi-ated through actions on primary afferent fibers and mod-ulation of autonomic activity.
Low Frequency (<10 Hz) TENS
Effects on Behavior and Dorsal Horn Neuron Activity
In primates without tissue injury, low rate burst TENS(3 bursts per second and 7 pulses per burst with an in-ternal frequency of 85 Hz) at an intensity that activatesAβ (presumable sensory intensity, 3 × sensory thresh-old) fibers has no effect on either the spontaneous activ-ity or responses to noxious stimuli of spinothalamic tractcells. Increasing intensity to activate Aδ fibers reducesspontaneous activity and responses to noxious stimuli ofspinothalamic tract cells (Lee et al. 1985). Similarly, lowfrequency TENS at an intensity that activates Aδ fibersreducestheventralrootreflexinresponsetoC-fiberstim-ulation (Sjolund 1985).Low frequency TENS, regardless of intensity has noeffect on the primary mechanical or heat hyperalge-sia produced by carrageenan inflammation. However,low frequency, sensory intensity TENS fully reversessecondary heat hyperalgesia and partially reverses sec-ondary mechanical hyperalgesia (reviewed in Sluka andWalsh 2003). Importantly, in these studies, increasingintensity to twice the motor threshold does not furtherreduce the secondary mechanical hyperalgesia. Theincreased responsiveness of dorsal horn neurons toinnocuous and noxious mechanical stimuli that occursafter inflammation is equally and completely reducedfollowing low frequency, sensory intensity TENS treat-ment applied to the inflamed paw (Ma and Sluka 2001).Following spinal nerve ligation, TENS reduces theresponsiveness to noxious mechanical stimulation ofdorsal horn neurons in both normal and neuropathicanimals. However, the responsiveness of spinal neurons

T
TePs 2409
to innocuous mechanical stimulation is only inhibitedby TENS in neuropathic animals (Leem et al. 1995).Behaviorally, low frequency, motor intensity TENSreduces mechanical hyperalgesia and cold allodyniainduced by nerve injury (Nam et al. 2001).
Pharmacology
Low frequency, sensory intensity, TENS antihyperalge-sia is prevented by blockade of μ-opioid receptors in thespinal cord or the RVM (Fig. 3) (Kalra et al. 2001; Slukaet al. 1999). Studies utilizing carrageenan knee joint in-flammation suggest thatμ-opioid receptorsarealso acti-vated by low frequency, motor intensity TENS since re-peatedapplicationofTENSproducestolerance(reducedeffectiveness) to the antihyperalgesic effects of TENSand of spinal μ-opioid receptors (Chandran et al. 2003).Low frequency, sensory intensity TENS is also reducedby blockade of serotonin 5-HT2A and 5-HT3 and mus-carinic M1 and M3 receptors in the spinal cord (Fig. 3)(Radhakrishnan et al. 2003; Radhakrishnan and Sluka2003).Taken together, thesestudiessuggesta roleofopi-oid, serotoninandmuscarinic receptors inthespinalcordand supraspinal opioid mechanisms in the action of lowfrequency, sensory intensity TENS.
Autonomic Effects of Low Frequency TENS
The effect of low frequency, motor intensity TENS oncold allodynia, but not mechanical hyperalgesia, is re-duced by systemic phentolamine to block α-adrenergicreceptors, suggesting activation of sympathetic nora-drenergic receptors may mediate TENS effects (Nam etal. 2001). However, phentolamine could block centralreceptors. Transient increases in blood flow with lowfrequency, burst-mode (2 Hz) TENS were observed atthe area of stimulation, if intensity was 25% above themotor threshold, but not just below (sensory intensity)or just above motor threshold (Sherry et al. 2001).
References1. Chandran P, Sluka KA (2003) Development of opioid tolerance
with repeated TENS administration. Pain 101:195–2012. Garrison DW, Foreman RD (1997) Effects of prolonged tran-
scutaneous electrical nerve stimulation (TENS) and variation ofstimulation variables on dorsal horn cell activity. Eur J Phys MedRehabil 6:87–94
3. Kalra A, Urban MO, Sluka KA (2001) Blockade of opioid re-ceptors in rostral ventral medulla prevents antihyperalgesia pro-duced by transcutaneous electrical nerve stimulation (TENS). JPharmacol Exp Ther 298:257–263
4. Lee KH, Chung JM, Willis WD (1985) Inhibition of primatespinothalamic tract cells by TENS. J Neurosurg 62:276–287
5. Leem JW, Park ES, Paik KS (1995) Electrophysiological evi-dence for the antinociceptive effect of transcutaneous electricalnerve stimulation on mechanically evoked responsiveness of dor-sal horn neurons in neuropathic rats. Neurosci Lett 192:197–200
6. Ma YT, Sluka KA (2001) Reduction in inflammation-inducedsensitization of dorsal horn neurons by transcutaneous electricalnerve stimulation in anesthetized rats. Exp Brain Res 137:94–102
7. Nam TS, Choi Y, Yeon DS et al. (2001) Differential antinoci-ceptive effect of transcutaneous electrical stimulation on painbehavior sensitive or insensitive to phentolamine in neuropathicrats. Neurosci Lett 301:17–20
8. Radhakrishnan R, Sluka KA (2003) Spinal muscarinic recep-tors are activated during low or high frequency TENS -inducedantihyperalgesia in rats. Neuropharmacology 45:1111–1119
9. Radhakrishnan R, King EW, Dickman J et al. (2003) Spinal5-HT(2) and 5-HT(3) receptors mediate low, but not high,frequency TENS-induced antihyperalgesia in rats. Pain105:205–213
10. Sherry JE, Oehrlein KM, Hegge KS et al. (2001) Effect of burst-mode transcutaneous electrical nerve stimulation on peripheralvascular resistance. Phys Ther 81:1183–1191
11. Sjolund BH (1985) Peripheral nerve stimulation suppression ofC-fiber evoked flexion reflex in rats. Part 1: Parameters of con-tinuous stimulation. J Neurosurg 63:612–616
12. Sluka KA, Walsh D (2003) Transcutaneous electrical nerve stim-ulation: Basic science mechanisms and clinical effectiveness. JPain 4:109–121
13. Sluka KA, Deacon M, Stibal A et al. (1999) Spinal blockade ofopioid receptors prevents the analgesia produced by TENS inarthritic rats. J Pharmacol Exp Ther 289:840–846
14. Somers DL Clemente FR (1998) High-frequency transcutaneouselectrical nerve stimulation alters thermal but not mechanicalallodynia following chronic constriction injury of the rat sciaticnerve. Arch Phys Med Rehabil 79:1370–1376
15. Woolf CJ, Mitchell D, Barrett GD (1980) Antinociceptive ef-fect of peripheral segmental electrical stimulation in the rat. Pain8:237–252
TENS Outcomes
� Transcutaneous Electrical Nerve Stimulation Out-comes
Tension Headache
� Headache, Episodic Tension Type
Tension Type Headache
Definition
Tension Type Headache in SLE patients is associatedwith personality changes, emotional conflicts, depres-sion, and higher disease activity scores. In some cases,tension type headache is associated with tonic contrac-tion of the cranial muscles. Many patients with tensiontype headache do not exhibit increased EMG activity inthese muscles, but have the feeling of a tight ring aroundthe head.� Headache Due to Arteritis� Headache, Episodic Tension Type� Sensitization of Muscular and Articular Nociceptors
TePs
� Tender Points

2410 Tertiary Gain
Tertiary Gain
Definition
Gains sought or obtained by others from a patient’s ill-ness.� Malingering, Primary and Secondary Gain
TES
� Transcutaneous Electrical Stimulation
Testosterone
Definition
Testosterone is normally produced in the testes in men,the ovaries in women, and in the adrenal cortex of bothmen and women. In men, testosterone is primarily re-sponsible for normal growth and development of malesex and reproductive organs, including the penis, testi-cles, scrotum, prostate, and seminal vesicles. It facili-tates thedevelopmentofsecondarymalesexcharacteris-tics such asmusculature,bonemass, fatdistribution,hairpatterns, laryngeal enlargement, and vocal chord thick-ening. In women, testosterone strengthens bone and en-sures the nipples and clitoris are sensitive to sexual plea-sure. In both men and women,normal testosterone levelsmaintain energy level, healthy mood, muscle mass, fer-tility,andsexualdesire.Decreased levelsof testosterone,induced by aging, disease, surgery, and medications (in-cluding opioids), lead to lossof libidoanddecreased sex-ual function.� Cancer Pain Management, Opioid Side Effects, En-
docrine Changes and Sexual Dysfunction
Tetrodotoxin
Synonyms
TTX
Definition
Tetrodotoxin (anhydrotetrodotoxin 4–epitetrodotoxin,tetrodonic acid, TTX) is a potent neurotoxin found inthe tissues of the puffer fish. It has a complex structure,with the active part of the molecule being a positivelycharged guanidinium moiety. The molecule can blocksodium channels from the outside (in contradistinc-tion to clinically used local anesthetic agents that mustfirst permeate the neuron and then act from the in-side). Tetrodotoxin is used by neurophysiologists tocategorize various types of sodium channel.
� Acute Pain Mechanisms� Membrane Stabilizing Drugs� Nociceptors, Action Potentials and Post-Firing Ex-
citability Changes� Tetrodotoxin
Tetrodotoxin (TTX)-Resistant SodiumChannel
Definition
A type of voltage-gated sodium channel that is not inhib-ited by thehighly potentneurotoxin, tetrodotoxin,whichis extracted from the puffer fish. Tetrodotoxin (TTX)-resistant sodium channels are found on the membraneof many DRG neurons that have nociceptive responseproperties, and contribute to the excitability of the neu-rons and generation of action potentials.� IB4-Positive Neurons, Role in Inflammatory Pain� Nociceptor Generator Potential
Thalamic Bursting Activity
� Burst Activity in Thalamus and Pain� Thalamic Bursting Activity, Chronic Pain
Thalamic Bursting Activity, Chronic PainA.TAGHVA, S.H. PATEL, A. FERNANDEZ,NIRIT WEISS, FRED A. LENZ
Department of Neurosurgery, Johns Hopkins Hospital,Baltimore, MD, [email protected]
Synonyms
Spike bursting
Definition
Spontaneous thalamic cellular activity is often catego-rized as either bursting activity (� spike bursts, burst-ing mode) or as tonic firing (tonic mode) (Steriade etal. 1990). Many studies have suggested that increased� spike bursting occurs in the thalamus of patients withchronic pain (Lenz et al. 1994; Lenz et al. 1988; Rinaldiet al. 1991; Jeanmonod et al. 1993; Lenz et al. 1998).

T
Thalamic Bursting Activity, Chronic Pain 2411
CharacteristicsThe Thalamic Region of Vc and its Importance in Pain Process-ingSeveral lines of evidence demonstrate that the ventralcaudal nucleus of the human sensory thalamus (� Vc),the human analog of monkey ventral posterior (VP)nucleus (Hirai and Jones 1989), is important in humanpain-signaling pathways. Studies of patients at autopsyfollowing lesions of the � spinothalamic tract (STT)show the most dense STT termination in the Vc regionincluding the posterior and inferior subnuclei of Vc,suprageniculate and posterior subnuclei (Mehler 1962;Walker 1943). In monkeys, STT terminations are foundin the Vmpo of Craig, which may, by immunohisto-chemistry and physiology, have a human analog (Craiget al. 1994).
Is Thalamic Functional Mode Altered in Chronic Pain States?� Spike bursting activity refers to a particular patternof � interspike intervals (ISI) between action potentials,such that a � spike burst begins after a relatively longISI and is comprised of a series of action potentials withshort ISIs (typically < 6 ms) (Lenz et al. 1989). There-after, the ISIs progressively lengthen so that the cell’sfiring decelerates throughout the spike burst. When theperiod of the bursting is completed, the cell is said to firein tonic mode when the firing rate is relatively constantand bursts do not occur.In patients with spinal transection, the highest rate ofbursting occurs in cells that do not have peripheral� receptive fields and that are located in the represen-tation of the anesthetic part of the body. These cellsalso have the lowest firing rates in the interval betweenbursts (Lenz et al. 1994). The low firing rates suggestthat these cells have decreased tonic excitatory drive andare � hyperpolarized, perhaps due to loss of excitatoryinput from the � STT (Eaton and Salt 1990; Doughertyet al. 1996). Therefore the available evidence suggeststhat affected thalamic cells in patients with spinal tran-section were dominated by spike bursting consistentwith membrane hyperpolarization ( 1990; Steriade andLlinas 1988; Lenz et al. 1998; Davis et al. 1998).Spikebursting activity ismaximal in the region posteriorand inferior to the core nucleus of Vc (Table 4 in Lenz etal. 1994). Stimulation in this area evokes the sensationof pain more frequently than does stimulation in the coreof Vc (Lenz et al. 1993; Hassler 1970). Thus, increasedspike bursting activity may be correlated with some as-pects of the abnormal sensations (e.g. � dysesthesia orpain) that thesepatientsexperience.However, inpatientswith spinal transection, the painful area and the area ofsensory loss overlap (Lenz et al. 1994). Thus, the burst-ing activity might be related to sensory loss, rather thanto pain.These findings about spike bursting activity in spinal pa-tients have been called into question by a recent study inpatients with chronic pain (Radhakrishnan et al. 1999).
It has been reported that the number of bursting cellsper trajectory in patients with movement disorders (con-trols) is not different from that in patients with chronicpain. However, there are significant differences betweenthe two studies (Lenz et al. 1994; Radhakrishnan et al.1999) in terms of patient population (spinal cord injuryvs mixed chronic pain), location of cells studied (Vc vsanterior and posterior to Vc) and analysis methods (inci-dence of bursting cells vs bursting parameters). Clearly,the increase in bursting activity demonstrated in the ear-lier study is more applicable to the region of the princi-pal somatic sensory nucleus of patients with central painfrom spinal transection (Lenz et al. 1994).Further support for increased spike bursts occurringin spinal cord transected patients is found in thalamicrecordings from monkeys with thoracic anterolateralcordotomies (Weng et al. 2000). Some of these animalsshowed increased responsiveness to electrocutaneousstimuli and thus may represent a model of central pain(Vierck 1991). The most pronounced changes in firingpattern were found in thalamic � multi-receptive cells,which respond to both cutaneous brushing and com-pressive stimuli with activity that is not graded into thenoxious range. In comparison with normal controls,multi-receptive cells in the monkeys with cordotomiesshowed significant increases in the number of burstsoccurring spontaneously or in response to brushing orcompressive stimuli. The changes in bursting behaviorwere widespread, occurring in the thalamic represen-tation of upper and lower extremities, both ipsilateraland contralateral to the cordotomy.Although there is an increase in spike burst activity inchronic pain states, there does not appear to be a directrelationship between spike burst firing and pain. Spikeburstsarealso found in the thalamic representationof themonkey upper extremity and of the representation of thearm and leg ipsilateral to the � cordotomy (Weng et al.2000). Pain is not typically experienced in these partsof the body in patients with thoracic spinal cord tran-section or cordotomy (Beric et al. 1988). Spike burstsare increased in frequency during slow wave sleep in allmammals studied (Steriade and Llinas 1988) includingman (Zirh et al. 1997). However, such bursting couldcause pain if stimulation in the vicinity of the burstingcellproduced thesensationofpain.Thisfindinghasbeenreported in two recent studies of sensations evoked bystimulation of the region of Vc in patients with chronicpain secondary to neural injury (Davis et al. 1996; Lenzet al. 1998).Thus, there is evidence from both human and animalstudies for a correlation between chronic pain statesand an altered thalamic neuronal action potential firingpattern. It appears that there is an increase in spike burstfiring in chronic pain conditions. The exact physiolog-ical relationships that link the pattern of thalamic firingto the human perception of chronic pain have yet to beelucidated.

2412 Thalamic Neurotransmitters and Neuromodulators
References1. Beric A, Dimitrijevic MR, Lindblom U (1988) Central dyses-
thesia syndrome in spinal cord injury patients. Pain 34:109–1162. Craig AD, Bushnell MC, Zhang ET et al. (1994) A thalamic
nucleus specific for pain and temperature sensation. Nature372:770–773
3. Davis KD, Kiss ZHT, Luo L et al. (1998) Phantom sensationsgenerated by thalamic microstimulation. Nature 391:385–387
4. Davis KD, Kiss ZHT, Tasker RR et al. (1996) Thalamicstimulation-evoked sensations in chronic pain patients and non-pain (movementdisorder)patients. JNeurophysiol 75:1026–1037
5. Dougherty PM, Li YJ, Lenz FA et al. (1996) Evidence that ex-citatory amino acids mediate afferent input to the primate so-matosensory thalamus. Brain Res 278:267–273
6. Eaton SA, Salt TE (1990) Thalamic NMDA receptors andnociceptive sensory synaptic transmission. Neurosci Lett110:297–302
7. Hassler R (1970) Dichotomy of facial pain conduction in the di-encephalon. In: Walker AE (ed) Trigeminal neuralgia. Saunders,Philadelphia, pp 123–138
8. Hirai T, Jones EG (1989) A new parcellation of the human tha-lamus on the basis of histochemical staining. Brain Res Rev14:1–34
9. Jeanmonod D, Magnin M, Morel A (1993) Thalamus and neuro-genic pain: physiological, anatomical and clinical data. Neurorep4:475–478
10. Lenz FA, Dostrovsky JO, Kwan HC et al. (1988) Do calciumspikes account for the bursts of action potentials which occur insensory thalamus of central pain patients? Society For Neuro-science Abstract 561
11. Lenz FA, Gracely RH, Baker FH et al. (1998) Reorganizationof sensory modalities evoked by stimulation in the regionof the principal sensory nucleus (ventral caudal - Vc) in pa-tients with pain secondary to neural injury. J Comp Neurol399:125–138
12. Lenz FA, Kwan HC, Martin R et al. (1994) Characteristics ofsomatotopic organization and spontaneous neuronal activity inthe region of the thalamic principal sensory nucleus in patientswith spinal cord transection. J Neurophysiol 72:1570–1587
13. Lenz FA, Seike M, Richardson RT et al. (1993) Thermal andpain sensations evoked by microstimulation in the area of humanventrocaudal nucleus. J Neurophysiol 70:200–212
14. Lenz FA, Zirh AT, Garonzik IM et al. (1998) Neuronal activityin the region of the principle sensory nucleus of human thalamus(ventralis caudalis) in patients with pain following amputations.Neurosci 86:1065–1081
15. Mehler WR (1962) The anatomy of the so-called “pain tract” inman: an analysis of the course and distribution of the ascendingfibers of the fasciculus anterolateralis. In: French JD, PorterRW (eds) Basic Research in Paraplegia. Thomas, Springfield,pp 26–55
16. Radhakrishnan V, Tsoukatos J, Davis KD et al. (1999) A com-parison of the burst activity of lateral thalamic neurons in chronicpain and non-pain patients. Pain 80:567–575
17. Rinaldi PC, Young RF, Albe-Fessard DG et al. (1991) Sponta-neous neuronal hyperactivity in the medial and intralaminar tha-lamic nuclei in patients with deafferentation pain. J Neurosurg74:415–421
18. Steriade M, Jones EG, Llinas RR (1990) Thalamic Oscillationsand Signaling. Wiley,John & Sons, New York
19. Steriade M, Llinas RR (1988) The functional states of thethalamus and the associated neuronal interplay. Physiol Rev68:649–742
20. Vierck CJ (1991) Can mechanisms of central pain syndromes beinvestigated in animal models? In: Casey KL (ed) Pain and centralnervous system disease: the central pain syndromes. Raven Press,New York, pp 129–141
21. Walker AE (1943) Central representation of pain. Res Publ AssocRes Nerv Ment Dis 23:63–85
22. Weng HR, Lee JI, Lenz FA, Vierck CJ, Rowland LH, DoughertyPM (2000) Functional plasticity in primate somatosensory tha-
lamus following chronic lesion of the ventral lateral spinal cord.Neuroscience 101:393-401
23. Zirh AT, Lenz FA, Reich SG et al. (1997) Patterns of burstingoccurring in thalamic cells during parkinsonian tremor. Neuro-science 83:107–121
Thalamic Neurotransmitters andNeuromodulators
KARIN N. WESTLUND, BRIDGET E. HAWKINS
Department of Anatomy and Neurosciences, Universityof Texas Medical Branch, Galveston, TX, [email protected], [email protected]
Synonyms
Signaling Molecules of Thalamic Regions; thala-mic neurotransmitters and neurochemical effectormolecules
Definition
The thalamus, a heterogeneous structure located in thedorsal diencephalic region of the brain, processes bothsensory and motor input signals prior to transmission tohigher cortical areas. � Neuromodulators are chemicaleffector molecules that determine the neural activity en-abling this process. While the microcircuitry and con-tent of neuromodulators in the thalamus can be partiallydetailed at this point in time, the precise role of trans-mitters of the sensory thalamus in the transmission andgeneration of what is perceived as “pain” is still largelyundefined. More recent observations suggest that neu-romodulation occurs throughout the excitatory and theinhibitory neurocircuitry of the thalamus. It is clear thatthechemicalneuromodulationoccurring in the thalamuspermits a significantly more important role for the tha-lamus than simply as a “relay station”. Rather, the tha-lamus is an integration site where filtering and consoli-dation of sensory information occurs through chemicalneurotransmission.
Characteristics
Neuromodulation in thesensory thalamuscanbestbede-scribed in terms of whether the neurochemicals are thecontent of input fibers, intrinsic interneurons or outputcomponents (Table 1; Fig. 1). The primary integrativesites in the thalamusfornociceptionare locatedmediallyand laterally in theposterior thalamus.Theventralposte-rior lateral thalamus is chiefly responsible for determin-ing the intensity and location of the painful stimulus be-fore relaying that information to the appropriate highercortical regions of the brain. The medial thalamus is in-volved with emotional responses to nociceptive input.
Thalamic Input
The primary sources of sensory information pro-vided to the thalamus are the incoming spinothala-mic, spinotrigeminal and medial lemniscal pathways.

T
Thalamic Neurotransmitters and Neuromodulators 2413
Thalamic Neurotransmitters and Neuromodulators, Table 1 Nociceptive Processing in the Thalamus
Thalamic Transmitters and Neuromodulators
Thalamic Input
Spinothalamic Tract and Medial Lemniscus GLU, CCK
Corticothalamic Tract GLU
Central Tegmental Field and Reticular Activation Systems ACH (ChAT), 5-HT, NA, (TH, DBH), CGRP
Input to Intralaminar and Ventral Thalamic nuclei (from hypothalamus, basal telencephalon, and lateralmidbrain)
ENK, SOM, CCK, SP, NPY
Intrinsic Interneurons and Reticular nucleus GABA, ACH, SOM
Thalamic Output
Thalamocortical Projections GLU
Thalamoamygdalar Projections CGRP
Thalamic Neurotransmitters and Neuromodulators, Figure 1 Sensory input to the thalamus is transmitted by glutamatergic (GLU) spinothalamic(STT), spinal trigeminal (SpV), dorsal column nucleus (DCN) and reticular formation (RF) neurons. The information, modulated by neuropeptides, is relayedto the medial and lateral thalamus (Thal) where it is influenced by intrinsic GABAergic (GABA) interneurons. The integrated information is routed byglutamatergic thalamic projection neurons to sites including the cerebral cortex (CTh) and amygdala. Reciprocal input back to the thalamus from thecortex is also glutamatergic. Abbreviations: ACh, acetylcholine; CCK, cholecystokinin; CRF, corticotrophin releasing factor; Enk, leu- or met-enkephalin;Glu, glutamate; 5-HT, serotonin; NA, norepinephrine; NPY neuropeptide Y; SOM, somatostatin; SP, substance P; VIP, vasoactive intestinal polypeptide.

2414 Thalamic Neurotransmitters and Neuromodulators
Thalamic Neurotransmitters and Neuromodulators, Figure 2 Schematic diagram illustrating the distribution of substance P immunoreactive fibers(dots) and cell somata (stars) in the monkey diencephalon (Jones 1988).
The corticothalamic tract, from cortical areas SI andSII, also provides a large contribution of incomingaxonal fibers to the thalamus. Glutamate, the classicalexcitatory amino acid, is believed to be the primarytransmitter of these fiber tracts, based on physiologicaland anatomical data. In other brain regions, increasedrelease of glutamate and activation of neurons by gluta-mate is typically enhanced by the modulatory effects of� neuropeptides, such as substance P (SP), CGRP andvasoactive intestinal polypeptide (VIP). Neuromodula-tors such as SP are found in abundance and probablyfunction in a similar manner in the sensory thalamus(Fig. 2). The neuropeptide content of neuronal terminalsin the thalamus is believed to be partially from thalamicinput sources (Jones 1988).
Spinothalamic Tract
Glutamate has been observed in the spinothalamicprojection neurons by electron microscopy (Westlundet al. 1992) (Fig. 1). Most spinothalamic neurons are
also believed to contain at least one neuropeptide, suchas SP, cholecystokinin (CCK), bombesin, dynorphin,enkephalin, galanin, corticotrophin releasing hormone(CRH) (Fig. 3) and / or vasoactive intestinal polypep-tide (VIP) (Coffield and Miletic 1987; Leah et al. 1988;Nahin 1988). In fact, some have been shown to con-tain multiple neuropeptides (Ju et al. 1987). LaminaX STT cells have been shown to predominantly con-tain bombesin, enkephalin, CCK, somatostatin (SOM),CRF, neuropeptide Y and SP (Battaglia et al. 1988;Leah et al. 1988). The SP seems to function as a slowintermediary for transmission of noxious stimuli aswell as a fast excitatory neurotransmitter modulator(Battaglia et al. 1988). Most of the terminals stainingfor SP are located in the medial rather than the lateralthalamus (Fig. 2). Enkephalin has also been identi-fied in spinotrigeminal neurons (Nahin 1988). Thesefindings are supported by both tract tracing / immuno-cytochemical studies and negative staining after spinalhemisection.

T
Thalamic Neurotransmitters and Neuromodulators 2415
Thalamic Neurotransmitters and Neuromodulators, Figure 3 After flu-orescent tracer is injected into the ventral thalamus, spinothalamic tractcells containing peptides such as corticotrophin releasing factor (CRF) canbe identified in the sacral spinal cord, lamina V. The panel labeled VPL illus-trates two cells retrogradely labeled after injection of fluorescent tracer intoventral thalamus. One of these cells also stained for CRF. Scale bar = 15μm.(Westlund et al., unpublished).
Medial Lemniscus
Input to the thalamus from the dorsal column nuclei isless well studied. Neuropeptide CCK content in the ven-tral posterolateral (VPL) nucleus is, however, reducedupon lesion of the contralateral dorsal column nucleus,suggesting that this modulator is involved in the relayof sensory information by the medial lemniscus (Jones1988).
Spinoreticular Pathways
Innervation by brainstem cholinergic, serotonergic andnoradrenergic neurons is also present in the thalamus(Jones 1988; Westlund et al. 1990) (Fig. 4). Sparseinnervation by � serotonin fibers from the raphe nu-cleus and adjacent periaqueductal gray region has beendemonstrated by dual labeling in monkeys. Similarly,in the same studies, norepinephrine was found sparselyinnervating the VPL. However, innervation by sero-tonergic, noradrenergic and � acetylcholine terminalsis found more prominently in the reticular and dorsallateral geniculate nuclei (as reviewed in McCormick1992). The purpose of these classical neurotransmittersin the thalamus remains unclear, though iontophoreticapplication of acetylcholine results in marked inhibition
of the activity of reticular nucleus neurons, which, if itpersists, can shift reticular nucleus cells into burstingactivity. Iontophoresis of norepinephrine or serotoninmarkedlyactivates thereticularnucleusneuronsthroughalpha1 and 5-HT2 (and possibly, 5-HT1C) receptors,due to the decrease in resting potassium conductance.The net result is facilitation of single spike activity andmarked inhibition of the rhythmic bursting activity nor-mally promoted by the slow release of norepinephrine inthis nucleus. It is assumed that these processes are partof the general arousal system throughout the neuraxisin which these transmitters participate. The highestlevels of 5-HT7 receptors are found on the intralami-nar and midline thalamic neurons. Stimulation of theraphe nuclei can alter the responses of these neurons tonociceptive input (Goaillard and Vincent 2002).
Corticothalamic InputCorticothalamic input to the thalamusisalsoglutamater-gic since there is dense staining of both glutamate andaspartate in cells in layers IV and III of SI sensory cortexand many of these cortical cells can be double labeledwith tract tracers injected into the thalamus (Guiffridaand Rustioni 1988; Rustioni et al. 1988) Only a smallpopulation ofneurons is labeled with bothglutamateandaspartate. The corticofugal fibers utilizing glutamate asthe primary neurotransmitter terminate as small endingscontaining round vesicles primarily contacting fine cal-iber (distal)dendritesof thalamicneuronsand arepartic-ularly dense in the reticular nucleus (Guiffrida and Rus-tioni 1988; McCormick 1992).
Intrinsic InterneuronsThe inhibitory neurotransmitter, gamma amino butyricacid (� GABA),hasbeenwellcharacterized in the thala-mus, particularly in the reticular nucleus of the thalamuswhere it is the predominant feature (Fig. 5). It has beendetermined that GABA is present in the interneurons ofthe ventral posterior thalamus only in carnivorous an-imals (i.e. present in primates and cats, but not in rats)(Rustioni et al. 1988). Intrinsic substance P interneuronshave also been described in the thalamus in regions re-ceiving spinothalamic tract input as demonstrated by an-terograde tracing (Battaglia et al. 1988).
Thalamic OutputInformation regarding the sensory-discriminative as-pect of pain is sent to parts of the cortex involved insomatosensory processing, such as precisely localizedareas I (SI), II (SII) and area 4, for further site specificassociation, in order that pain appropriate responseinstructions can be dispatched via the motor systems.The thalamocortical efferent pathways (Fig. 1) wereidentified as glutamate positive neurons with collateralsto the reticular nucleus (Guiffrida and Rustioni 1988;as reviewed in McCormick 1992). Thalamic output tothe cortex is activated by glutamate NMDA and non-NMDA receptors, since specific antagonists applied

2416 Thalamic Neurotransmitters and Neuromodulators
Thalamic Neurotransmitters and Neuromodulators, Figure 4 Transverse sections of the brainstem of a monkey showing retrograde labeling ofneurons (FB) after injection of Fast Blue in the ipsilateral ventral posterolateral nucleus of the thalamus, and the distribution of choline acetyltransferase(ChAT)-, tyrosine hydroxylase (TH)- and serotonin (5HT)-immunoreactive cells (Jones 1988).

T
Thalamic Neurotransmitters and Neuromodulators 2417
Thalamic Neurotransmitters andNeuromodulators, Figure 5 (a)Immunocytochemical staining ofGABAergic neurons in the reticularnucleus (R) of a cat, identifiedusing antiserum to glutamicacid decarboxylase (GAD). Finedots in the dorsal thalamus areGAD-positive somata of interneurons.(b) Higher-power photomicrographof GAD-positive cells in reticularnucleus (R) and ventral posterior (VP)nucleus of a cat. Bars = 1 mm (a),250 μm (b). (Jones 1985)
to the thalamus can block transmission of thalamicoutflow. The glutamatergic input to the thalamocorticalneurons probably arises from the input sources detailedabove.There is also an affective component to pain perception,which contributes to the emotional responses generatedin response to the stimulus. This aids in memory de-velopment for future avoidance of the painful situation.These functions are relegated to the mediodorsal (ven-trocaudal) (MDvc) and intralaminar thalamic regionswhich receive input mainly from laminae I and X of thespinal cord and send projections to the cingulate cortex(in primates). The ventral posterior inferior nuclei sendsignals directly to the somatosensory area of the parietalcortex (SII) and indirectly to the insula (Millan 1999).Both may be joined by additional ipsilateral input. Athin layer of CGRP neurons is found just ventral tothe thalamus near to the meso-diencephalic junction(Kruger et al. 1988) and is associated with the thala-mus and the somatosensory pathways, as are all CGRPcomponents of the nervous system. These CGRP neu-rons have been shown to provide a thalamoamygdalarprojection which may also be important in generatingthe emotional responses to painful stimuli.Calcium calmodulin-dependent protein kinase (CaMkinase II) has been found in thalamic dendrites apposedby glutamate terminals, suggesting that this major post-synaptic activity regulator may assist in modulatingsynaptic strength, transmitter release and / or vesicle
movement in thalamic neurons (Liu and Jones 1996).The CaM kinase II terminals in the cerebral cortex arethought to be terminals of thalamocortical neurons.
References1. Battaglia G, Spreafico R, Rustioni A (1988) Substance P-
immunoreactive fibers in the thalamus from ascending so-matosensory pathways. In: Bentivoglio M, Spreafico R (eds)Cellular Thalamic Mechanisms. Elsevier Science Publishers,Amsterdam
2. Coffield JA, Miletic V (1987) Immunoreactive enkephalin is con-tained within some trigeminal and spinal neurons projecting tothe rat medial thalamus. Brain Res 425:380–383
3. Goaillard JM, Vincent P (2002) Serotonin suppresses the slowafterhyperpolarization in rat intralaminar and midline thalamicneurones by activating 5-HT(7) receptors. J Physiol 541:453–465
4. Guiffrida R, Rustioni A (1988) Glutamate and aspartate im-munoreactivity in corticothalamic neurons of rats. In: BentivoglioM, Spreafico R (eds) Cellular Thalamic Mechanisms. ElsevierScience Publishers, Amsterdam, pp 311–320
5. Jones EG (1985) Transmitters, receptors, and related compoundsin the thalamus. In: Jones EG (ed) The Thalamus. Plenum Press,New York, pp 225–256
6. Jones EG (1988) Modern views of cellular thalamic mechanisms.In: Bentivoglio M, Spreafico R, (eds) Cellular Thalamic Mech-anisms. Elsevier Science Publishers, pp 1–22
7. Ju G, Melander T, Ceccatelli S et al. (1987) Immunohisto-chemical evidence for a spinothalamic pathway co-containingcholecystokinin- and galanin-like immunoreactivities in the rat.Neuroscience 20:439–456
8. Kruger L, Sternini C, Brecha NC et al. (1988) The thalamic re-gion of calcitonin gene-related peptide (CGRP) immunoreactiv-ity and its relation to somatosensory pathways. In: BentivoglioM, Spreafico R (eds) Cellular Thalamic Mechanisms. ElsevierScience Publishers, Amsterdam, pp 375–386

2418 Thalamic Nociceptive Neurons
9. Leah J, Menetrey D, de Pommery J (1988) Neuropeptides inlong ascending spinal tract cells in the rat: evidence for parallelprocessing of ascending information. Neuroscience 24:195–207
10. Liu XB, Jones EG (1996) Localization of alpha type II calciumcalmodulin-dependent protein kinase at glutamatergic but notgamma-aminobutyric acid (GABAergic) synapses in thalamusand cerebral cortex. Proc Natl Acad Sci USA 93:7332–7336
11. McCormick DA (1992) Neurotransmitter actions in the thala-mus and cerebral cortex and their role in neuromodulation ofthalamocortical activity. Prog Neurobiol 39:337–388
12. Millan MJ (1999) The induction of pain: an integrative review.Prog Neurobiol 57:1–164
13. Nahin RL (1988) Immunocytochemical identification of long as-cending, peptidergic lumbar spinal neurons terminating in eitherthe medial or lateral thalamus in the rat. Brain Res 443:345–349
14. Rustioni A, Battaglia G, De Biasi S et al (1988) Neuromediatorsin somatosensory thalamus: an immunocytochemical overview.In: Bentivoglio M, Spreafico R (eds) Cellular Thalamic Mech-anisms. Elsevier Science Publishers, Amsterdam, pp 271–296
15. Westlund KN, Sorkin LS, Ferrington DG, Carlton SM, Willcock-son HH, Willis WD (1990) Serotonergic and noradrenergic pro-jections to the ventral posterolateral nucleus of monkey thalamus.J Comp Neurol 295:197–207
16. Westlund KN, Carlton SM, Zhang D et al. (1992) Glutamate-immunoreactive terminals synapse on primate spinothalamictract cells. J Comp Neurol 322:519–527
Thalamic Nociceptive Neurons
� Human Thalamic Nociceptive Neurons
Thalamic Nociceptive System
� Thalamic Nuclei Involved in Pain, Cat and Rat
Thalamic Nuclei
DefinitionThe thalamus, which is located in the centre of the cere-bral cortex, is comprised of many nuclei. The thalamusrelays information to and from the cerebral cortex, andalso plays a part in modulating sensory information.� Central Pain, Diagnosis
Thalamic Nuclei Involved in Pain, Cat andRat
CHRISTIANE VAHLE-HINZ
Institute of Neurophysiology and Pathophysiology,University Hospital Hamburg-Eppendorf, Hamburg,[email protected]
SynonymsThalamic Nociceptive System; Lateral and Medial Tha-lamic Nociceptive System
Definition
Nuclei of the dorsal thalamus whose neurons receiveinputs from ascending nociceptive pathways and projectto cortical nociceptive areas. The neurons respond tostimulation of � nociceptors in skin, muscles, jointsand viscera. The different thalamic regions processand transmit information subserving the � sensory-discriminative and the � motivational-affective com-ponents of pain.
Characteristics
A distinction is generally made in all species betweena lateral and a medial thalamic system, the lateral withneurons encoding stimulus quality, duration, inten-sity and location on the body and thus subserving the� sensory-discriminative component of pain, the me-dial system with neurons activated from large areasof the body and/or internal organs and involved in themotivational-affective component of pain. The differentresponse properties reflect ascending inputs dominatedbythespinothalamicandspinaltrigeminothalamictracts(STT) and projection to the SI and SII somatosensorycortices (lateral system) versus STT and spinoreticu-lothalamic tract and widespread cortical projectionsincluding limbic areas (medial system).In contrast to other sensory nuclei in the thalamus, no-ciceptive “nuclei” cannot be delineated histologically.Rather, nociceptive neurons are found within nuclei pri-marily subserving other functions like the mediodorsalor intralaminar nuclei or the ventral posterior complex(VP). Thus, “thalamic nuclei involved in pain” havebeen identified by neuroanatomical tracing of termina-tions of ascending tracts carrying nociceptive signalsor by � electrophysiological mapping of neurons re-sponsive to noxious stimuli. The latter is hinderedfurther by the problems of applying painful stimuli inawake animals or the inherent � antinociceptive effectsof general anesthetics when studies are done under� anesthesia (Vahle-Hinz and Detsch 2002; Vahle-Hinz et al. 2002). At the spinal cord level, the majorityof nociceptive somatic and visceral neurons in addi-tion have low-threshold somatic receptive fields (RFs),which may be preserved in the thalamus while their no-ciceptive components are abolished by the anesthetic.These difficulties are reflected in the heterogeneouspicture of thalamic nociception in the literature (forreview see Willis 1997) and it still awaits clarificationwhether species differences originate from princi-pally different organizations or a unifying picture mayemerge.
Lateral Thalamus
Nociceptiveneuronsarefoundinregionsofterminationsof the STT that are patchy and dispersed in and aroundthe VP and the posterior complex (PO) in the rat whilethey are confined to the PO and the margins of the VP in

T
Thalamic Nuclei Involved in Pain, Cat and Rat 2419
the cat. These regions are further characterized by smallneurons and may correspond to the small-celled matrixregionsinandaroundtheVPofmonkeys.Thus,althoughthe location of nociceptive neurons may differ betweenspecies (outside versus inside VP), this may result froma different degree of invasion of this small-celled matrixinto the VP.
Region of the Ventral Posterior Complex (VP)
The VP is characterized by input from the medial andtrigeminal lemnisci carrying a somatotopic representa-tionof low-thresholdmechanoreceptorsof thecontralat-eral half of the body surface including the furry buccalpad (rat) or a bilateral representation of intraoral struc-tures (cat) (Vahle-Hinz and Gottschaldt 1983). The car-toon in Fig. 1B shows the approximate proportions inthe cat’s VP in a frontal plane, while in fact the complexextends caudomedially to rostrolaterally with the hindlimbarearostral to theface inbothratandcat.Thesubnu-clei of the VP holding the representations of thedifferentparts of the body stand out from adjacent regions histo-logicallybydenserpackingandlargersomataofneurons(Fig. 1a). Interspersed are smaller � GABAergic cellswith local axonal arbors (interneurons, about 25% of theneurons) in the cat; these are virtually absent in the rat.The projection neurons of the VP have large round so-
Thalamic Nuclei Involved in Pain, Cat and Rat, Figure 1 Locations of nociceptive neurons in the lateral thalamus of the cat. (a, b) Subnuclei ofthe ventral posterior complex and adjacent regions as determined by histology (a), Nissl stain, coronal section, medial to the left, scale bar: 1 mm)and electrophysiological mapping of neuronal receptive fields (b). (c) Examples of neurons from the VPvp and the VP (camera lucida drawings fromGolgi-stained tissue, scale bar: 50 μm). (d) Recording sites in VPp of nociceptive neurons with cutaneous receptive fields. Nociceptive-specific (dots)and multireceptive neurons (triangles). (e) Recording sites of visceroceptive neurons in VPp and PO determined by electrical stimulation of the pelvicnerve. Eml, external medullary lamina; POl, m, medial and lateral parts of the posterior complex; R, thalamic reticular nucleus; VMb, basal ventral medialnucleus (gustatory relay); VP, ventral posterior complex; VP<Subscript>p</Subscript>, periphery of VP; VPvp, ventral periphery of VP; VPMm, l, medial(intraoral RFs) and lateral (face RFs) parts of the ventral posterior medial nucleus; VPL, ventral posterior lateral nucleus (postcranial body RFs); ZI, zonaincerta; scale bar: 1 mm. Modified from Vahle-Hinz et al. 1995 (b), Kniffki et al. 1993 (c), Kniffki and Vahle-Hinz 1987 (e), Brüggemann et al. 1994 (e).
matawith dense radialdendritic arbors (Fig.1c) (Kniffkiet al. 1993).Nociceptive neurons occur in the periphery of the VP(VPp) in the cat (Kniffki and Vahle-Hinz 1987; Vahle-Hinz et al. 1987). This peripheral region is character-ized by scattered neurons with small, spindle-shaped so-mataand fewprimary dendritesgivingrise tosparseden-dritic arbors oriented within the confines of the region,i.e. mediolaterally along its borders (Fig. 1c) (Kniffki etal. 1993). The narrow sheath of cells wraps around theVP dorsally and laterally, extending towards the exter-nalmedullary laminaonitsventralborder, thuswideningto a sizeable area ventral to the ventral posterior medialnucleus (VPM). This part resembles the ventral poste-rior inferior nucleus (VPI) of the monkey. Most of theneurons in the VPMp have somatic RFs and the major-ity are located on the head (Fig. 1d). The lateral part ofthe VPp (VPLp) is a narrow band surrounding the ven-tral posterior lateral nucleus (VPL) ventrally, laterallyand dorsally. It merges with the lateral and medial partsof the PO and its RFs are located on the fore- and hind-limbs as well as on visceral organs (Fig. 1e). Thus withrespect to somatic RFs, a coarse somatotopy is presentrunning parallel to that of the VP proper.While no nociceptive neurons are found inside the VPof the cat, they may invade the laminae between subnu-

2420 Thalamic Nuclei Involved in Pain, Human and Monkey
clei of the VP as shown in Fig. 1b. In the rat, clusteringof VP cells with similar RFs is even more pronouncedand thus laminae with nociceptive cells are more numer-ously interspersed, contributing to the appearance of amixed representation of mechanoreceptive and nocicep-tive neurons described in the literature (for review seeWillis 1997). Most of the somatic nociceptive neuronsin the rat, however, are found in the PO.
Posterior Complex (PO)
The somatosensory parts of PO adjoin VPL laterallyand dorsally and VPM dorsally (POm). The nuclearand subnuclear boundaries are not discernible histo-logically. PO receives inputs from the STT, the lateralcervical nucleus, the dorsal column, the nucleus of thesolitary tract and the parabrachial area. These affer-ents carry nociceptive somatic and visceral signals.In the cat, the VPLp and adjoining PO were found tohold visceroceptive neurons with inputs from thoracicand pelvic visceral organs (Brüggemann et al. 1993;Brüggemann et al. 1994; Horn et al. 1997; Horn etal. 1999; Vahle-Hinz et al. 1995). Also in the rat, aconcentration of visceroceptive neurons in the regionimmediately surrounding the VP was found (Berkley etal. 1993). No viscerotopic organization is discerniblebut the response properties indicate that this region maybe involved in the encoding, localization and referralof visceral pain.The somatic low-threshold RFs of PO neurons are of-ten large and bilateral and include deep structures likemuscles and joints. In the rat, somatic nociceptive neu-rons with small RFs and stimulus encoding propertiessimilar to those of the cat’s VPp are found in the POm.
Medial Thalamus
Nociceptive somatic and visceral neurons are found ina number of nuclei of the medial thalamus, includingthe central lateral and parafascicular nucleus of the in-tralaminarcomplex, themediodorsalnucleusand thenu-cleus submedius (for review see Willis 1997). The RFsare usually large, often bilateral, with convergent inputfrom skin, muscles, joints and viscera. There is no soma-totopic or viscerotopic organization; thus these regionsmaynotbeinvolvedinthespatial localizationofapainfulstimulus. In contrast, intensity is encoded in graded re-sponses, a property important for the affective compo-nent of pain. The nociceptive responses are particularlysensitive to the kind and depth of anesthesia. NS, WDRand multireceptiveneuronsoccur in themediodorsal andintralaminar nuclei. The nucleus submedius receives adense projection from � lamina I of the spinal cord incats and nociceptive-specific somatic neurons here aremore abundant than WDR neurons in both rats and cats.
References1. Berkley KJ, Guilbaud G, Benoist J-M, Gautron M (1993) Re-
sponses of neurons in and near the thalamic ventrobasal complex
of the rat to stimulation of uterus, vagina, colon, and skin. J Neu-rophysiol 69:557–568
2. Brüggemann J, Vahle-Hinz C, Kniffki K-D (1993) Representa-tion of the urinary bladder in the lateral thalamus of the cat. JNeurophysiol 70:482–491
3. Brüggemann J, Vahle-Hinz C, Kniffki K-D (1994) Projectionsfrom the pelvic nerve to the periphery of the cat’s thalamic ven-tral posterolateral nucleus and adjacent regions of the posteriorcomplex. J Neurophysiol 72:2237–2245
4. Horn AC, Vahle-Hinz C, Petersen M et al. (1997) Projectionsfrom the renal nerve to the cat’s lateral somatosensory thalamus.Brain Res 736:47–55
5. Horn AC, Vahle-Hinz C, Brüggemann J et al. (1999) Responses ofneurons in the lateral thalamus of the cat to stimulation of urinarybladder, colon, esophagus, and skin. Brain Res 851:164–174
6. Kniffki K-D, Vahle-Hinz C (1987) The periphery of the cat’sventroposteromedial nucleus (VPMp ): Nociceptive neurones. In:Besson J-M, Guilbaud G, Peschanski M (eds) Thalamus and Pain.Elsevier, Amsterdam, pp 245-257
7. Kniffki K-D, Pawlak M, Vahle-Hinz C (1993) Scaling behaviorof the dendritic branches of thalamic neurons. Fractals 1:171–178
8. Vahle-Hinz C, Brüggemann J, Kniffki K-D (1995) Thalamic pro-cessing of visceral pain. In: Bromm B, Desmedt J (eds) Pain andthe Brain. From Nociception to Cognition. Advances in Pain Re-serach and Therapy, vol 22. Raven Press, New York, pp 125–141
9. Vahle-Hinz C, Detsch O (2002) What can in vivo electrophysi-ology in animal models tell us about mechanisms of anesthesia?Br J Anaesth 89:123–142
10. Vahle-Hinz C, Freund I, Kniffki K-D (1987) Nociceptive neuronsin the ventral periphery of the cat thalamic ventroposteromedialnucleus. In: Schmidt RF, Schaible H-G, Vahle-Hinz C (eds) FineAfferent Nerve Fibers and Pain. VCH Verlagsgesellschaft, Wein-heim, pp 440–450
11. Vahle-Hinz C, Gottschaldt K-M (1983) Principal differences inthe organization of the thalamic face representation in rodentsand felids. In: Macchi G, Rustioni A, Spreafico R (eds) So-matosensory Integration in the Thalamus. Elsevier, Amsterdam,pp 125–145
12. Vahle-Hinz C, Reeker W, Detsch O et al (2002) Antinociceptiveeffects of anesthetics in vivo: Neuronal responses and cellularmechanisms. In: Urban BW, Barann M (eds) Molecular and Ba-sic Mechanisms of Anesthesia. Pabst Sci Publ, Lengerich, pp516–524
13. Willis WD (1997) Nociceptive functions of thalamic neurons.In: Steriade M, Jones EG, McCormick DA (eds) Thalamus, Vol.II, Experimental and Clinical Aspects. Elsevier, Amsterdam, pp373–424
Thalamic Nuclei Involved in Pain, Humanand Monkey
EDWARD JONES
Center for Neuroscience, University of California,Davies, CA, [email protected]
Synonyms
Nociceptive Coding in Lateral Thalamus; Lateral thala-mus encodes pain
Description
Based on anatomical considerations, afferent inputs,cortical projections and electrophysiological evidence,the core of the ventral posterior nuclei must be involvedin transmitting nociceptive information to the cortex.

T
Thalamic Nuclei Involved in Pain, Human and Monkey 2421
Characteristics
It is likely that many thalamic nuclei and therefore theirprojection areas in the cerebral cortex are “involved” inpain. However, despite many attempts to discredit it, itremains apparent that the ventral posterior nuclear com-plex is that part of the thalamus that possesses the ap-propriate input connections and the relevant cellular ma-chinery for relaying the qualitative features and periph-eral localizationofapainfulstimulus.Anecessarycorol-lary of this is that the cortical projection target of the ven-tral posterior complex, the post central gyrus, is a cen-tral element in the onward pathways to the perceptionof pain.Theessay� SpinothalamicTerminations,Coreand Ma-trix summarizes the newer connection tracing studiesthat demonstrate not only the widespread terminationsof the � central pain pathways in the primate thalamusbut also their relationships to cells within the heart oftheventralposteriornucleus, the� ventralposteriorme-dial (VPM) and � ventral posterior lateral (VPL) nuclei.Here they appear to terminate preferentially on the ma-trix cells of the two nuclei, extending these terminationsbeyond the confines of � VPM and � VPL into and sur-rounding nuclei that are also characterized by the pres-ence of � calbindin-immunoreactive matrix cells.
Thalamic Nuclei Involved inPain, Human and Monkey,Figure 1 Schematic view of thedistribution of spinothalamic tractfibers in the thalamus of the macaquemonkey and the cortical projectionsof the receiving nuclei. CL, centrallateral nucleus; Po, posterior nucleus;Pla, anterior pulvinar nucleus; SG-L,limitans-suprageniculate nucleus;VLp, posterior ventral lateral nucleus;VMb, basal ventral medial nucleus;VPI, ventral posterior inferior nucleus;VPL, ventral posterior lateral nucleus;VPM, ventral posterior medialnucleus; ZI, zona incerta.
Single unit recordings both within and around theperimeter of the ventral posterior nucleus of the tha-lamus in monkeys and humans reveal the presenceof neurons with both nociceptive-specific and � widedynamic range properties akin to those found in dorsalhorn neurons (e.g. Bushnell et al. 1993; Craig et al.1994; Lee et al. 1999; Ohara and Lenz 2003; Willisand Coggeshall 2004). Unfortunately, these have neverbeen satisfactorily identified as exclusively within thematrix, but it is clear that many of these recordingshave come not just from the matrix regions surround-ing VPM and VPL but also from within the heart ofthese nuclei themselves. From what we have describedin the essay on spino- and spinal trigeminothalamicterminations mentioned above, it is unlikely (althoughsome have considered it controversial) that noci- orthermo-specific neurons of spinal lamina I project onlyto neurons outside the confines of the ventral posteriornucleus, although their projections may be concentratedthere in the matrix domain in monkeys and humans (andin comparable regions in cats). Of special note is thepresence of neurons with nociceptive specific propertieswithin the heart of the ventral posterior nucleus. Thiscarries the implication that these neurons project tothe primary somatosensory cortex and that has indeed

2422 Thalamic Nuclei Involved in Pain, Human and Monkey
been directly demonstrated in a small number of cases(Kenshalo et al. 1980).The presence of neurons with properties appropriate forrelaying the details of the modality, location and inten-sity of a painful stimulus within the heart of the thalamicnucleus that projects to the primary somatosensory cor-tex should not be taken to imply that these neurons arefound only therein. They are indeed found within all ormost of the other sites of termination of the spinothala-mic tracts as a whole and within those sites outside theventral posterior nucleus to which neurons specificallylocated in the superficial dorsal horn project (reviewedin Willis et al. 2002). The cortical targets of these otherparts of the thalamus, which include the � ventral pos-terior inferior and � basal ventral medial nuclei of theventral posterior complex, the adjacent matrix-filled do-mains of the � posterior nucleus and � anterior pulv-inar nucleus and the caudal intralaminar nuclei, are di-verse and include the second somatosensory area, otherperi-insular areas and parts of the prefrontal and pos-sibly anterior cingulate cortex. The broad implicationis the classical one that the multi-dimensional charac-ter of that sensation that we call pain is reflected in in-puts to regions of cortex whose known or conjecturedfunctional connotations are appropriate for each of thesedimensions. But there is still no reason to rule out the be-lief that the primary somatosensory cortex is the primaryroute to the centers for perception of the quality, loca-tion and intensity of a painful stimulus. This is not theplace to consider all the data that imply the dissociationof the sensory discriminative and affective-motivationaldimensions of pain in relation to different cortical areasbut there is a growing literature (often misinterpreted ormisrepresented) in this field that is covered in other es-says in this encyclopedia.Functional imaging studies in humans support the viewof widespread thalamocortical projections in the centralpain system. Most of the areas referred to in the preced-ing paragraph show functional activation in response toapplication of painful stimuli to the skin or stimuli en-gendered in deeper tissues. Areas of the cortex report-edly activated during the appreciation of a painful stim-ulus include the first and second somatosensory areas,the insula and the anterior cingulate gyrus (e.g. Coghillet al. 1994; Davis et al. 1998; Gelnar et al. 1999). Tryingto correlate these observations with electrophysiologi-cal reports from monkeys and the effects of stimulationor lesions in humans presents a confusing picture fromwhich workers committed to any point of view about thecortical representation of pain, however eccentric, canderive sustenance.When we look at the results of recording or imaging ofthepostcentralgyrusafterpresentationofpainfulstimulito human subjects, it is difficult to escape the conclusionthat the primary somatosensory cortex is at the heart ofthe territory involved in nociception but more lateral ar-eas are likely to be involved as well (Treede et al. 2000;
Oharaetal. 2004a,2004b).These imagingresults,more-over, tend to be supported by the results of single unitstudies in monkeys (Robinson and Burton 1980; Ken-shalo et al. 2000). The role of the lateral areas has beenadduced mainly from lesion studies in humans, but thesehave been variously interpreted.My own view is that lesions involving the parietal op-erculum (which might mean involvement of the secondsomatosensory area alone but could equally undercutfibers approaching the postcentral gyrus) seem to alterthe sensory-discriminative aspects of pain apprecia-tion, while those affecting regions located in insularand postinsular regions tend to alter affective and moti-vational aspects and can result in pain asymbolia (see,for example, Greenspan et al. 1999; Ostrowsky et al.2002). This still leaves out, however, the prefrontal andcingulate areas. Perhaps it is their activation in painstates that represents the inputs from thalamic nucleisuch as the caudal intralaminar?
References1. Bushnell MC, Duncan DH, Tremblay N (1993) Thalamic VPM
nucleus in the behaving monkey. I. Multimodal and discrim-inative properties of thermosensitive neurons. J Neurophysiol69:739–752
2. Coghill RC, Talbot JD, Evans AC et al. (1994) Distributed pro-cessing of pain and vibration by the human brain. J Neurosci14:4095–4108
3. Craig AD, Bushnell MC, Zhang E-T et al. (1994) A thalamicnucleus specific for pain and temperature sensation. Nature372:770–773
4. Davis KD, Kwan CL, Crawley AP et al. (1998) Functional MRIstudy of thalamic and cortical activations evoked by cutaneousheat, cold, and tactile stimuli. J Neurophysiol 80:1533–1546
5. Gelnar PA, Krauss BR, Sheehe PR et al. (1999) A comparativefMRI study of cortical representations for thermal painful, vibro-tactile, and motor performance tasks. Neuroimage 10:460–482
6. Greenspan JD, Lee RR, Lenz FA (1999) Pain sensitivity alter-ations as a function of lesion location in the parasylvian cortex.Pain 81:273–282
7. Kenshalo DR Jr, Giesler GJ Jr, Leonard RB et al. (1980) Re-sponses of neurons in primate ventral posterior lateral nucleusto noxious stimuli. J Neurophysiol 43:1594–1614
8. Kenshalo DR, Iwata K, Sholas M, Thomas DA (2000) Re-sponse properties and organization of noiceptive neurons inarea 1 of monkey primary somatosensory cortex. J Neurophysiol84:719–729
9. Lee J-I, Dougherty PM, Antezana D et al (1999) Responses ofneurons in the region of human thalamic principal somatic sen-sory nucleus to mechanical and thermal stimuli graded into thepainful range. J Comp Neurol 410:541–555
10. Ohara S, Lenz FA (2003) Medial lateral extent of thermal andpain sensations evoked by microstimulation in somatic sensorynuclei of human thalamus. J Neurophysiol 90:2367–2377
11. Ohara S, Crone NE, Weiss N et al. (2004a) Cutaneous painfullaser stimuli evoke responses recorded directly from pri-mary somatosensory cortex in awake humans. J Neurophysiol91:2734–2746
12. Ohara S, Crone NE, Weiss N et al. (2004b) Attention to painis processed at multiple cortical sites in man. Exp Brain Res156:513–517
13. Ostrowsky K, Magnin M, Ryvlin P et al. (2002) Representationof pain and somatic sensation in the human insula: a study ofresponses to direct electrical cortical stimulation. Cereb Cortex12:376–385

T
Thalamic Plasticity and Chronic Pain 2423
14. Robinson CJ, Burton H (1980) Somatic submodality distributionwithin the second somatosensory (SII), 7b, retroinsular, postau-ditory and granular insular cortical areas of M. fascicularis. JComp Neurol 192:93–108
15. Treede RD, Apkarian AV, Bromm B et al. (2000) Cortical repre-sentation of pain: functional characterization of nociceptive areasnear the lateral sulcus. Pain 87:113–119
16. Willis WD Jr, Coggeshall RE (2004) Sensory Mechanisms ofthe Spinal Cord, 3rd edn. Plenum, New York
17. Willis WD Jr, Zhang X, Honda CN et al. (2002) A critical reviewof the role of the proposed VMpo nucleus in pain. J Pain 3:79–94
Thalamic Pain
� Central Pain, Diagnosis
Thalamic Physiology Changes Occurringin Patients with Chronic Pain
� Thalamus,ReceptiveFields,ProjectedFields,Human
Thalamic Plasticity and Chronic PainJONATHAN O. DOSTROVSKY1,DOUGLAS RASMUSSON
2
1Department of Physiology, University of Toronto,Toronto, ON, Canada2Department of Physiology and Biophysics, DalhousieUniversity, Halifax, NS, [email protected]
Synonyms
Functional Changes in Thalamus; Chronic Pain, Thala-mic Plasticity
Definition
Long term changes in processing of somatosensory in-puts in thalamus as a result of � deafferentation and/orchronic pain.
Characteristics
Most of the information regarding � plasticity in the so-matosensory system at supraspinal levels concerns theprocessing and representation of non-nociceptive tactileinputs.This ismainlybecause it isdifficult todetermineaclearmapof thebodyrepresentationofneuronsrespond-ing to nociceptive inputs. However, it is likely that thereare at least some common mechanisms between plastic-ity of innocuous information and nociceptive informa-tion processing, and thus the first section will deal with
findings regarding plasticity in processing of tactile in-formation.In contrast to the hundreds of papers that have describedreorganization of the primary somatosensory cortex af-ter deafferentation, relatively few have studied changesat the thalamic level. However, the variables that affectthalamic and cortical reorganization are the same: theage of the animal at the time of the damage; the extentand location of the damage; and the time that has expiredsince the damage. Neonatal damage, or even trimmingof whiskers from birth to decrease sensory experience,can produce dramatic changes in the somatotopic mapwithin the rodent thalamus (e.g. Nicolelis et al. 1997;Simons and Land 1994). These developmental changesare reflected in morphological changes in the structuralrepresentation of whiskers in the rodent thalamus, the“� barreloids”. In particular, the separation between af-fected barreloids is lost, suggesting a profound interfer-ence with the migration of thalamic neurons. The criticalperiod for the thalamus is intermediate between the ear-lier developing brainstem and the later developing cor-tex.In the adult animal, immediate reorganization of the� ventral posterior lateral nucleus (VPL) thalamus, byreversibly blocking afferent inputs from the skin follow-ing local anesthetic injections, has been demonstratedin several species including man (Nicolelis et al. 1993;see Dostrovsky 1999 for other references). This resultsin the appearance of new and/or enlargement of the RFsof individual neurons. Of particular interest is a studythat found that this unmasking was greatly reduced ifthe somatosensory cortex was inactivated at the time ofperipheral deafferentation, suggesting that � CorticalFeedback plays an important role in these immediatechanges (Krupa et al. 1999).Longer-term plasticity of the thalamus (requiring sev-eral months) has been demonstrated after a variety ofdifferent types of peripheral nerve damage. These in-clude transection of median and ulnar nerves in squirrelmonkeys (Garraghty and Kaas 1991), digit amputationin raccoons (Rasmusson 1996), dorsal rhizotomies inprimates(JonesandPons1998),aswellascentral lesionsaffecting the ascending branch of peripheral afferents inthedorsalcolumns(PollinandAlbe-Fessard1979).Tha-lamic reorganization also results from destruction of thedorsal column nuclei, which provide the major, directinput to VPL and to the spinothalamic tract. Lesioningthe gracile nucleus in rats results in expansion of upperlimb and shoulder representations (Parker et al. 1998;Wall and Egger 1971). In all these cases, the major effectis an expansion of intact adjacently represented regionsinto the region that has been deafferented. In addition,spinothalamic tract lesions in sub-human primates havebeen shown to increase the spontaneous firing rate andresponses to mechanical stimulation of the skin, and in-crease the degree of � bursting activity in low thresholdslowly adapting neurons (Weng et al. 2003).

2424 Thalamic Projections
Recordings and stimulation in human thalamus ofchronic pain patients suffering spinal cord damageor amputation reveal an expansion of the intact re-gions into deafferented regions, and the existence ofneurons firing spontaneously in bursts. Stimulation insuch regions usually evokes sensations referred to thedeafferented region or phantom limb. Some of thesealterations may also be involved in mediating the pa-tients’ pain (see references in Dostrovsky 1999; Wenget al. 2003).Few studies have examined the mechanisms responsiblefor thalamic plasticity. Electron microscopic evidencefor synaptogenesis, presumably resulting from sprout-ing of surviving axons, has been found in the rat afterdorsal column nuclei lesions (Wells and Tripp 1987). Ofpotential interest is the decrease in � GABAA receptorsthat follows dorsal � rhizotomy in primates (Rausellet al. 1992). In addition, other studies have revealedchanges in the morphology of � GABA terminals fol-lowing dorsal column lesions. Decreased inhibitorycontrol could contribute directly or indirectly to theplasticity mechanisms responsible for thalamic reorga-nization, and may also result in increased RF sizes andlowered thresholds of nociceptive neurons in thalamus(see references in Weng et al. 2003). There is also elec-trophysiological evidence that many cells in VPL have� subliminal receptive fields, and these may providean important substrate for the expansion of � receptivefields following deafferentation (Dostrovsky 1999).There have been several studies in animals showinggreatly increased responses, receptive field sizes, anddecreased thresholds of nociceptive neurons in me-dial and lateral (VPL) thalamus following peripheraldamage, inflammation, or central damage. Althoughit is likely that some of these alterations were due tochanges at the thalamic level, it is difficult to assessto what extent these were due to neuroplastic changesoccurring at spinal and trigeminal levels (e.g. Dostro-vsky and Gulibaud 1990; see references in Weng et al.2003).Severalstudieshavereported that followingdeafferenta-tion and � CNS damage leading to chronic pain, there isan increase in bursting activity of neurons in and near thedeafferented region in human thalamus, and have sug-gested that this may be related to the development ofcentral pain. Bursting cells have also been observed inchronic pain patients (see � Thalamic Bursting Activityand references in Weng et al. 2003).Of considerable interest are observations in humans onthe incidence of stimulation-evoked pain. In non-painpatients the incidence of evoking pain by stimulationwithin � ventral basal nucleus (VB) is very low. How-ever, in some types of pain patients, and in particularin � post-stroke central pain patients, the incidence ismuch higher, suggesting that neuroplastic changes haveoccurred in the thalamocorticalsysteminthesecases thatare causing pain to be perceived by thalamic stimuli that
normally elicit only non-painful parasthesia (see refer-ences in Dostrovsky 1999; Weng et al. 2003).� Thalamus, Dynamics of Nociception
References1. Dostrovsky JO (1999) Immediate and Long-Term Plasticity in
Human Somatosensory Thalamus and its Involvement in Phan-tom Limbs. Pain 6:37–43
2. Dostrovsky JO, Guilbaud G (1990) Nociceptive Responsesin Medial Thalamus of the Normal and Arthritic Rat. Pain40:93–104
3. Garraghty P, Kaas J (1991) Functional Reorganization in AdultMonkey Thalamus after Peripheral Nerve Injury. NeuroReport2:747–750
4. Jones EG, Pons TP (1998) Thalamic and Brainstem Contribu-tions to Large-Scale Plasticity of Primate Somatosensory Cortex.Science 282:1121–1125
5. Krupa DJ, Ghazanfar AA, Nicolelis MA (1999) Immediate Tha-lamic Sensory Plasticity Depends on Corticothalamic Feedback.Proc Natl Acad Sci USA 96:8200–8205
6. Nicolelis MA, Lin RCS, Chapin JK (1997) Neonatal WhiskerRemoval Reduces the Discrimination of Tactile Stimuli by Tha-lamic Ensembles in Adult Rats. J Neurophysiol 78:1691–1706
7. Nicolelis MAL, Lin RCS, Woodward DJ et al. (1993) Inductionof Immediate Spatiotemporal Changes in Thalamic Networks byPeripheral Block of Ascending Cutaneous Information. Nature361:533–536
8. Parker JL, Wood ML, Dostrovsky JO (1998) A Focal Zone ofThalamic Plasticity. J Neurosci 18:548–558
9. Pollin L, Albe-Fessard D (1979) Organization of Somatic Tha-lamus in Monkeys with and without Section of Dorsal SpinalTracts. Brain Res 173:431–449
10. Rasmusson DD (1996) Changes in the Organization of the Ven-troposterior Lateral Thalamic Nucleus after Digit Removal inAdult Raccoon. J Comp Neurol 364:92–103
11. Rausell E, Cusick CG, Taub E et al. (1992) Chronic Deaf-ferentation in Monkeys Differentially Affects Nociceptive andNonnociceptive Pathways Distinguished by Specific Calcium-Binding Proteins and Down-Regulates γ-Aminobutyric AcidType A Receptors at Thalamic Levels. Proc Natl Acad Sci USA89:2571–2575
12. Simons DJ, Land PW (1994) Neonatal Whisker Trimming Pro-duces Greater Effects in Nondeprived than Deprived ThalamicBarreloids. J Neurophysiol 72:1434–1447
13. Wall PD, Egger MD (1971) Formation of New Connexionsin Adult Rat Brains after Partial Deafferentation. Nature232:542–545
14. Wells J, Tripp LN (1987) Time Course of Reactive Synaptoge-nesis in the Subcortical Somatosensory System. J Comp Neurol255:466–475
15. Weng HR, Lenz FA, Vierck C et al. (2003) Physiological Changesin Primate Somatosensory Thalamus Induced by Deafferentationare Dependent on the Spinal Funiculi that are Sectioned and Timefollowing Injury. Neuroscience 116:1149–1160
Thalamic Projections
� Corticothalamic and Thalamocortical Interactions
Thalamic Reorganization
� Thalamus, Dynamics of Nociception

T
Thalamo-Amygdala Interactions and Pain 2425
Thalamic Response to Experimental Painin Humans
� Human Thalamic Response to Experimental Pain(Neuroimaging)
Thalamic Reticular Nucleus
Synonyms
TRN
Definition
A nucleus that surrounds the thalamus. It receives inputsfromthecortexandthalamusandprojectsbacktothetha-lamus with inhibitory connections. Neurones in this tha-lamic nucleus are inhibitory GABAergic neurones thatprovide inhibition onto thalamic relay neurones. TRNneurones do not project to the cerebral cortex.� Thalamocortical Loops and Information Processing� Thalamus, Metabotropic Glutamate Receptors� Thalamus, Nociceptive Neurotransmission
Thalamo-Amygdala Interactions and PainGEORGE S. BORSZCZ1, S. E. HARTE2
1Department of Psychology, Wayne State University,Detroit, MI, USA2Department of Internal Medicine, Chronic Pain andFatigue Research Center, Rheumatology University ofMichigan Health System, Ann Arbor, MI, [email protected], [email protected]
Synonyms
Affective analgesia; affective pain processing; LimbicForebrain Matrix; pain asymbolia
Definition
Processing the � affective-motivational dimension ofpain experience. Suppression of nociceptive transmis-sion through this limbic forebrain matrix producesselective inhibition of the affective reaction to pain(affective analgesia). Interactions between limbic fore-brain sites and the midbrain periaqueductal gray is onemechanism through which morphine acts to produceaffective analgesia.
Characteristics
W. H. Sweet (1980), the eminent neurosurgeon and painresearcher,observed thecriticalneed toevaluate theneu-ral mechanisms that generate and suppress the affective-motivational dimension of pain. In reviewing the extant
human neurosurgicaldata, he noted that ablation of sev-eral limbic forebrain sites proved effective in reducingthe ‘agonizing’ pain associated with advanced cancer.These areas include the anterior cingulate cortex (ACC),medial thalamic nuclei (centromedian and parafascic-ular nuclei), amygdala, and prefrontal cortex. Damageof these structures produced no loss of the � sensory-discriminative dimension of pain, but was efficacious inalleviating its affective-motivational dimension. Sweetpointed out that the mechanisms responsible for this dis-sociative analgesia were poorly understood, and calledfor the development of animal models and systematicresearch programs that could provide insights into theneurobiology of the affective dimension of pain.Subsequent research provided convergent evidencein support of the involvement of the limbic-forebrainin processing the affective-motivational dimension ofhuman pain. Additional human neurosurgical obser-vations confirmed the involvement of the � medialthalamus and ACC in pain perception. Ablations ofthese areas remain an effective treatment in alleviat-ing chronic intractable pain (Hassenbusch et al. 1990;Whittle and Jenkinson 1995). The chronic pain stateappears to be mediated by sustained abnormal neuralactivity within the limbic forebrain. High frequencyspontaneous neural activity was recorded from medialthalamic nuclei in patients with deafferentation pain(Rinaldi et al. 1991). High frequency stimulation ofthe medial thalamus produced reports of intense painin human subjects, which was suppressed by treatmentwith the μ-opiate agonist fentanyl (Velasco et al. 1998).Neuroimaging studies of the human pain, experimentaland clinical, also revealed a limbic-forebrain matrix thatunderlies the processing of the affective-motivationaldimension of pain (see review Derbyshire 2000). Thesestudies consistently report activation of the amygdala,ACC, insula, prefrontal cortex, and medial thalamus.Following the � parallel pain processing model ofMelzack and Casey (1968), the results of these neuro-surgical and neuroimaging studies havebeen interpretedas reflecting the processing of noxious stimulation bythe ‘� medial pain system’ (Vogt and Sikes 2000).The medial pain system is proposed to process nox-ious stimulation transmitted by medially projectingspinothalamic pathways (i.e. spinoreticulothalamictract). This projection system terminates in medial tha-lamic nuclei that project to limbic forebrain structuresthat underlie processing of the affective-motivationaldimension of pain. The ACC, insula, and prefrontal cor-tex are principal forebrain targets of medial thalamicprojections (Vogt and Sikes 2000). The medial thala-mus also projects to the amygdala that is reciprocallyinterconnected with the medial thalamus, ACC, andinsula. The amygdala and medial thalamus also receivenociceptive afferents from the spinal dorsal horn, re-layed via the parabrachial nucleus (Bernard and Besson1990; Bourgeais et al. 2001). The processing of noxious

2426 Thalamocortical and Corticothalamic Interactions
stimulation by this limbic-forebrain matrix generatesthe emotional reaction to pain, coordinates relevantmotor activity, and supports the development of fearconditioning and avoidance responding (Derbyshire2000).Alternately, the ‘� lateral pain system’ processes nox-ious stimulation transmitted by laterally projectingspinothalamic and trigeminothalamic pathways thatterminate in lateral thalamic nuclei (i.e. VPL andVPM). Projections of the lateral thalamic nuclei tothe primary and secondary somatosensory cortices areproposed to underlie the processing of the sensory-discriminative dimension of the pain experience. Thisdimension of pain signals the location, intensity, andphysical properties of a noxious stimulus.Systemic administration of morphine preferentiallysuppresses the affective reaction of humans and ani-mals to noxious stimulation (Borszcz et al. 1994; Priceet al. 1985). This effect is mediated by the inhibition ofpain transmission through limbic forebrain structuresthat process the affective dimension of pain (Casey etal. 2000). The ventrolateral division of the midbrain pe-riaqueductal gray (vPAG) is a major site through whichmorphine acts to suppress pain transmission. Throughits descending projections to the rostral ventromedialmedulla, the vPAG inhibits pain transmission at the levelof the spinal dorsal horn. The vPAG also contributesascending projections that suppress pain transmissiondirectly within the limbic forebrain (Borszcz 1999).The vPAG and adjacent dorsal raphé nucleus provideascending serotonergic projections to the medial tha-lamus (parafascicular nucleus) and amygdala, and themicroinjection of morphine into the vPAG increasesthe release and metabolism of serotonin in these sites(Munn and Borszcz 2002; Tao and Auerbach 1995).Moreover, the suppression of rats’ affective reaction tonoxious stimulation following injection of morphineinto vPAG is reversed by bilateral administration ofserotonin antagonists into either the parafascicularnucleus or amygdala (Borszcz 1999).The parafascicular nucleus and amygdala appear tointeract in the production of affective analgesia fol-lowing the injection of morphine into the vPAG. Theunilateral administration of serotonin antagonists intothe parafascicular nucleus or amygdala failed to alterthe antinociceptive action of vPAG-administered mor-phine. However, the combined unilateral administrationof serotonin antagonists into the parafascicular nucleusand amygdala was as effective as their bilateral admin-istration into either site in reversing the antinociceptiveaction of morphine injected into the vPAG (Borszczand Streltsov 2000). These findings suggest that a func-tional interaction exists between the medial thalamus,amygdala, and vPAG in mediating affective analgesia.Evidence also exists for the interaction between thevPAG and other limbic forebrain sites (nucleus ac-cumbens, habenula) in mediating the antinociceptive
action of vPAG-administered morphine (Ma and Han1991).
References1. Bernard JF, Besson JM (1990) The Spino(trigemino)pontoamyg-
daloid Pathway: Electrophysiological Evidence for an Involve-ment in Pain Processes. J Neurophysiol 63:473–490
2. Borszcz GS (1999) Differential Contributions of Medullary, Tha-lamic, and Amygdaloid Serotonin to the Antinociceptive Ac-tion of Morphine Administered into the Periaqueductal Gray: AModel of Morphine Analgesia. Behav Neurosci 113:612–631
3. Borszcz GS, Johnson CP, Fahey KA (1994) Comparison of MotorReflex and Vocalization Thresholds following Systemically Ad-ministered Morphine, Fentanyl, and Diazepam in the Rat: Assess-ment of Sensory and Performance Variables. Pharmacol BiochemBehav 49:827–834
4. Borszcz GS, Streltsov NG (2000) Amygdaloid-Thalamic In-teractions Mediate the Antinociceptive Action of MorphineMicroinjected into the Periaqueductal Gray. Behav Neurosci114:574–584
5. Bourgeais L, Monconduit L, Villanueva L et al. (2001)Parabrachial Internal Lateral Neurons Convey NociceptiveMessages from the Deep Laminas of the Dorsal Horn to theIntralaminar Thalamus. J Neurosci 21:2159–2165
6. Casey KL, Svensson P, Morrow TJ, Raz J, Jone C, Minoshima S(2000) Selective Opiate Modulation of Nociceptive Processingin the Human Brain. J Neurophysiol 84:525–533
7. Derbyshire SWG (2000) Exploring the Pain ‘Neuromatrix’. CurrRev Pain 4:467–477
8. Hassenbusch SJ, Pillay PK, Barnett GH (1990) Radiofre-quency Cingulotomy for Intractable Cancer Pain using Stereo-taxis Guided by Magnetic Resonance Imaging. Neurosurgery27:220–223
9. Ma QP, Han JS (1991) Neurochemical Studies on the MesolimbicCircuitry of Antinociception. Brain Res 566:95–102
10. Munn EM, Borszcz GS (2002) Increases in the Release andMetabolism of Serotonin in Nucleus Parafascicularis Thalamifollowing Systemically Administered Morphine in the Rat.Neurosci Lett 332:151–154
11. Price DD, Von der Gruen A, Miller J et al. (1985) A Psychophys-ical Analysis of Morphine Analgesia. Pain 22:261–269
12. Rinaldi PC, Young RF, Albe-Fessard D et al. (1991) SpontaneousNeuronal Hyperactivity in the Medial and Intralaminar Thala-mic Nuclei of Patients with Deafferentation Pain. J Neurosurg74:415–421
13. Sweet WH (1980) Mechanisms of Chronic Pain (Neuralgias andCertain Other Neurogenic Pain). In: Bonica JJ (ed)Pain. RavenPress, New York, pp 287–303
14. Tao R, Auerbach SB (1995) Involvement of the Dorsal Raphebut not Median Raphe Nucleus in Morphine-Induced Increasesin Serotonin Release in the Rat Forebrain. Neuroscience68:553–561
15. Velasco M, Brito F, Jimenez F et al. (1998) Effect of Fentanyl andNaloxone on a Thalamic Induced Painful Response in IntractableEpileptic Patients. Stereotact Funct Neurosurg 71:90–102
16. Vogt BA, Sikes RW (2000) The Medial Pain System, CingulateCortex, and Parallel Processing of Nociceptive Information. Progin Brain Res 122:223–235
17. Whittle IR, Jenkinson JL (1995) CT-Guided Stereotactic Antero-Medial Pulvinotomy and Centromedian-Parafascicular Thalam-otomy for Intractable Malignant Pain. Br J Neurosurg 9:195–200
Thalamocortical and CorticothalamicInteractions
� Corticothalamic and Thalamocortical Interactions

T
Thalamocortical Loops and Information Processing 2427
Thalamocortical Dysrhythmia
Synonyms
TCD
Definition
A pathophysiological chain reaction at the origin ofneurogenic pain. It consists of: 1) a reduction of ex-citatory inputs onto thalamic cells, which results incell membrane hyperpolarization, 2) the productionof low-threshold calcium spike bursts by deinactiva-tion of calcium T-channels, discharging at low (theta)frequency, 3) a progressive increase of the numberof thalamocortical modules discharging at theta fre-quency, and 4) a cortical high frequency activationthrough asymmetric corticocortical inhibition. Theseevents have been documented by thalamic and corticalrecordings in patients suffering from peripheral andcentral neurogenic pain.� Thalamotomy for Human Pain Relief� Thalamus, Dynamics of Nociception
Thalamocortical Fibers
Definition
Axons with cell bodies located in the thalamus and ter-minations in the cortex.� Corticothalamic and Thalamocortical Interactions
Thalamocortical Loops and InformationProcessing
S. MURRAY SHERMAN
Department of Neurobiology, State University of NewYork, Stony Brook, NY, [email protected]
Synonyms
Cortical Information Flow; Corticortical Pathways
Definition
Until recently, communication among related corti-cal areas (e.g., those for somatosensation and pain)was thought to involve direct connections. We (Sher-man and Guillery 2001; Sherman and Guillery 2002;Guillery and Sherman 2002a; Guillery and Sherman2002b) suggest a radically new view in which many andperhaps all corticocortical communications involve acortico-thalamo-cortical route.
Characteristics
To understand how information is processed in a tha-lamocortical system, it is important to identify and fol-low the route of information transfer. A recent sugges-tion based on thalamic circuitry is that not all pathwaysare equivalent, but instead can be divided into “drivers”which are the information bearing pathways and “mod-ulators” which serve to modulate theflowof informationrather than transmitting it. How this might apply to corti-cal processing in general and cortical processing of painmore specifically is best explained by considering howthis idea has led to important changes in our thinking ofthalamic circuitry.
Drivers and Modulators
Figure 1 shows the basic circuit of the thalamus, whichvaries only slightly among thalamic relays. As arguedpreviously, the inputs to relay cells can be divided intotwo basic types, “drivers” and “modulators” and thesediffer on a number of different morphological and func-tionalgrounds thatarebriefly summarized inTable1 (fordetails, see Sherman and Guillery 2001, 2002; Guilleryand Sherman 2002a). The first pair of listed differencesareproperties limited to thalamus,but theremainder rep-resent criteria that can be applied anywhere in the cen-tral nervous system. The drivers are the input that bringsthe information to be relayed. Examples are retinal inputto the lateral geniculate nucleus, medial lemniscal inputto the � ventral posterior nucleus and, as noted belowfor some thalamic relays, layer 5 input from cortex. Themodulatorsareeverything elseand theirmain function isto control the level and type of information relayed fromdrivers through thalamus to cortex. Examples are the lo-cal� GABAergiccells (i.e., interneuronsandcellsof thethalamic reticular nucleus), feedback from cortical layer6 and a projection from the brainstem reticular forma-tion. Drivers represent relatively few of the synaptic in-puts to relay cells (only about5–10%),but their synapsesare relatively powerful. The other 90–95% of synapsesonto relay cells are divided roughly equally among mod-ulatory inputs from local GABAergic cells, from corti-cal layer 6 and from the brainstem. The modulators re-quire thevastmajorityof inputsformanysubtle roles thataffect the relay of driver inputs (Sherman and Guillery2001, 2002; Guillery and Sherman 2002a).The main difference between thalamic relays is the ori-gin of the driver input; the modulators are basically sim-ilar throughout thalamus, although there is some varia-tion (Jones 1985; Sherman and Guillery 2001).The understanding that inputs to relay cells can be di-vided into drivers and modulators and that the formerlargely define the function of a thalamic relay has im-plications that may extend beyond thalamus (see alsobelow). Thus the � lateral geniculate nucleus is largelydefined asa relay of retinal information. It is important tounderstand thatconsiderationofanatomical information

2428 Thalamocortical Loops and Information Processing
Thalamocortical Loops and Information Processing, Figure 1 Schema of inputs to thalamic relay cells. Abbreviations: 5-HT, serotonin; ACh, acetyl-choline; BRF, brainstem reticular formation; GABA, gamma-aminobutyric acid; Glu, glutamate; LGN, lateral geniculate nucleus; NA, noradrenalin; TRN,thalamic reticular nucleus.
alone can obscure this. For the lateral geniculate nucleusfor instance, only 5-10% of synapses onto relay cells de-rive from retina and roughly one third derive from brain-stem. If we had only these anatomical data, most of uswould conclude that the lateral geniculate nucleus re-layed brainstem information and that retinal input pro-vided some obscure, minor function. In other words, wewould badly misconstrue this thalamic relay.
First and Higher Order Relays
Thus identifying the driver is a major key in determiningthe role played by a thalamic relay. For instance, we de-fine the role of the � lateral geniculate nucleus based onits relay of retinal axons and that of the ventral posteriornucleus based on its relay of � medial lemniscus axons.However, until recently, the role played by many thala-mic relays remained a mystery, because it was not clearwhat was being relayed. We used to think that the roleof the thalamus was to relay subcortical information tocortex and for large regions of thalamus, such as muchof the pulvinar, it was not clear what was the subcorticalsource being relayed.
However, therecent realization thatdriversformanytha-lamic relaysoriginate in layer5 ofcortex led to adivisionof thalamus into “first order” and “higher order” relays,and this is summarized in Fig. 2 (Sherman and Guillery2001, 2002; Guillery and Sherman 2002a). First orderrelays transmit to cortex a particular type of informa-tion (e.g. retinal) for the first time, whereas higher or-der relays are involved in further transmission of suchinformation between cortical areas. The higher order re-lay can be between a first order and higher order corticalarea (as shown in Fig. 2) or between two higher ordercortical areas (not shown).Higherorder relayshavebeenidentified for the major sensory systems, the pulvinar forvision, theposteriormedialnucleus for somatosensation(and thus for pain) and the magnocellular division of themedial geniculate nucleus for hearing. Other examplesofhigherorder relayshavealsobeen identified(seeSher-man and Guillery 2001, 2002).Several features from Fig. 2 bear further emphasis. Allthalamic relays receive a modulatory input from layer6 of cortex that is mainly feedback, whereas only thehigher order relays receive in addition a layer 5 corticalinput and this is feedforward. Note also that the driver

T
Thalamocortical Loops and Information Processing 2429
Thalamocortical Loops and Information Processing,Figure 2 First order (FO; left) and higher order (HO;right) thalamic relays. For simplicity, connections torelay cells from interneurons and brainstem are omitted.“Glomerulus” refers to a complex synaptic zone that isubiquitous to thalamus and that is often associated withdriver input.
Thalamocortical Loops and Information Processing, Figure 3 Involvement of higher order thalamic relays in corticocortical communication. Forsimplicity inputs from interneurons and cells of the thalamic reticular nucleus omitted. Abbreviations as in Fig. 1 plus: MGNv, ventral region of medialgeniculate nucleus; MGNmagno; magnocellular region of medial geniculate nucleus; POm, posterior medial nucleus; VP, ventral posterior nucleus.
afferents, both subcortical to first order relays and fromlayer 5 for higher order relays, are branches ofaxons thatalso innervate an extrathalamic target, which tends to be“motor” in nature; this is true for many and perhaps alldriver inputs (fordetails, seeGuillery and Sherman 2002a, b; Guillery 2003). For instance, many or all retinal af-
ferents to the lateral geniculate nucleus branch to alsoinnervate midbrain structures associated with control ofpupil size, eye movements, etc and many layer 5 affer-ents to higher order thalamic relays also innervate manylevels of the brainstem and may extend input to spinallevels. It isas if the information relayed to cortex through

2430 Thalamocortical Loops and Information Processing
thalamus isacorollary ofmotorcommandsandit is thesemotor commands that serve as the basis of perceptual in-formation acted upon and further elaborated by cortex(Guillery and Sherman 2002 a, b; Guillery 2003).It is also worth noting that, as sufficient information re-garding various thalamic relays develops regarding thedivision into first order and higher order, the large ma-jority of thalamus seems to be devoted to higher orderrelays.
Role of Thalamus in Corticocortical Communication
Figure 3 summarizes the major implication of this divi-sion of thalamic relays into first order and higher orderfor cortical functioning. Information of a particular sortfirst reaches cortex via a first order relay; this can applyto primary information about vision, sounds, pain, etc.Further cortical processing of this primary informationisbased on cortico-thalamo-corticalpathways involvinghigher order thalamic relays. This view of corticocorti-cal processing has the interesting feature that any newinformation reaching a cortical area, whether initiatedsubcortically or in another cortical area, benefits from a
Thalamocortical Loops andInformation Processing,Figure 4 Conventional (upper)versus alternate (lower) views ofcortical processing.
thalamic relay.Such benefitsarebeyond thescopeof thisessay to cover, but the reader can learn more of this fromother sources (Sherman and Guillery 2001; Guillery andSherman 2002a).To place this scheme in the proper perspective, it is im-portant to appreciate that most prevailing conceptionsabout functioning of cortical areas are based on directconnections between areas. For instance, the best stud-ied is visual cortex, which is divided into more than 30discreteareas inhumansandthedetailedschemeoffunc-tional organization is based almost entirely on the pat-tern of direct corticocortical connections, with no placefor thalamus (Van Essen et al. 1992; Kandel et al. 2000).A similar view dominates thinking about the organiza-tion of somatosensory cortical areas responsible for thecortical processing of pain. Understanding how corticalareas process information requires first identifying theroutes of information and, if the driver/modulator dis-tinction holds for cortical pathways as it seems to in tha-lamus, it then becomes essential to distinguish amongthe direct corticocortical pathways those that are driversfrom those that are modulators. As it happens, the cur-

T
Thalamocortical Neurones 2431
rent views of cortical organization consider only directcorticocortical connections that have been identified al-mostentirely with anatomical techniquesand an impliedassumption that needs to be made explicit is that all moreor less contribute equally, in a sort of anatomical democ-racy, to information flow. This same logic applied to thethalamus would produce the misconception that the lat-eral geniculate nucleus relayed brainstem, not retinal,inputs to cortex (see above).Given the nature of thalamocortical inputs, which havethe morphological and functional characteristics ofdrivers, it seems very likely that the cortico-thalamo-cortical pathways shown in Fig. 3 are important infor-mation routes. It follows that understanding the rela-tionships of cortical areas in various functional zones(e.g. visual, somatosensory and auditory among others)will require mapping out of all of the cortico-thalamo-cortical pathways involving higher order thalamicrelays.What, then is the function of the direct corticocorticalpathways?Ananswer to this importantquestionrequiresidentifying these pathways, one by one if necessary, forfunction as driver or modulator. One extreme possibilityis that all of these pathways are modulators. However,even if some are drivers, there is an important distinctionto bemadebetween suchputative informationroutesandthose involving higher order thalamic relays. That is, theformer involve information that remains strictly withincortex, whereas the latter involve information, perhapsinvolving motor commands, that is shared with varioussubcortical centers.
Summary and Conclusions
To understand the implications of the proposal put for-ward here for the roleof thalamus incorticocortical com-munication, it might be helpful to contrast it with theconventional view, and this is done in Fig. 4. In the con-ventional view (Fig. 4, upper), sensory information is re-layed from the periphery by thalamus to a primary sen-sory cortical area. From there, the information is pro-cessed strictly within cortex, eventually via sensorimo-tor areas to motor areas and finally this leads to a motoroutput. Note that, in this view, the only role for thala-mus is to get raw information to cortex in the first placeand that most of thalamus, which we call higher orderrelays, has no specific role to play. In the alternate view(Fig. 4, lower) offered here, information relayed to cor-tex is, from the very beginning, corollary to motor com-mands and further corticocortical processing involveshigher order thalamic relays of continuously elaboratedand updated motor commands. Thus thalamus not onlygets information to cortex in the first place but also con-tinues to play an essential role in corticocortical com-munication.This has important implications for cortical functioninggenerally and also for cortical processing of pain infor-mation more specifically. That is, the higher order tha-
lamic relays involved in pain processing could be key.The best candidate for the higher order thalamic relayof pain information would be the posterior medial nu-cleus, which lies mostly medial to the ventral posteriornucleus, most of which is the first order somatosensoryrelay.Weclearlyneedabetterunderstandingofhowpainis processed by somatosensory cortex and the purpose ofthis essay is to provide a different theoretical frameworkthat might fruitfully guide further research through thistopic.
References1. Guillery RW (2003) Branching thalamic afferents link action and
perception. J Neurophysiol 90:539–5482. Guillery RW, Sherman SM (2002a) Thalamic relay functions and
their role incorticocortical communication: Generalizations fromthe visual system. Neuron 33:1–20
3. Guillery RW, Sherman SM (2002b) Thalamocortical pathwaysas monitors of ongoing motor instructions. Philos Trans R SocLond (Biol) 357:1809–1821
4. Jones EG (1985) The Thalamus. Plenum Press, New York5. Kandel ER, Schwartz J H, Jessell TM (2000) Principles of Neural
Science. McGraw Hill, New York6. Sherman SM, Guillery RW (2001) Exploring the Thalamus. Aca-
demic Press, San Diego7. Sherman SM, Guillery RW (2002) The role of thalamus in the
flow of information to cortex. Philos Trans R Soc Lond (Biol)357:1695–1708
8. Van Essen DC, Anderson CH, Felleman DJ (1992) Informationprocessing in the primate visual system: an integrated systemsperspective. Science 255:419–423
Thalamocortical Module
Definition
Anatomofunctional entity comprising of thalamic cellsand their cortical partners, interconnected by thalamo-cortical and corticothalamic projections and sustainingperceptual, motor and cognitive hemispheric functions.The thalamocortical loop is accompanied by a shorterthalamoreticulothalamic loop. Every module may besubdivided in a specific, or content subpart, providingthe substrate for the integration of a given function,and a non-specific, or context subpart, dealing with theinteractions between functional domains.� Thalamotomy for Human Pain Relief
Thalamocortical Neurones
Definition
Neurones located within the thalamus and projecting di-rectly to the cerebral cortex.� Spinothalamic Projections in Rat

2432 Thalamotomy
Thalamotomy
Definition
A neurosurgical procedure in which a therapeutic lesionis made in a specific subnucleus of the thalamus.� Pain Treatment, Intracranial Ablative Procedures� Thalamotomy for Human Pain Relief� Thalamotomy, Pain Behavior in Animals� Thalamus,ReceptiveFields,ProjectedFields,Human
Thalamotomy for Human Pain ReliefDANIEL JEANMONOD
1, JOHANNES SARNTHEIN1,
JAIR STERN1, MICHEL MAGNIN2,CHRISTOPH AUFENBERG
1, ANNE MOREL1
1Functional Neurosurgery, Neurosurgical Clinic,University Hospital, Zürich, Switzerland2INSERM, Neurological Hospital, Lyon, [email protected]
Synonyms
Stereotactic operation with thalamic target. First ordersubtype: medial thalamotomy. Second order subtype:central lateral thalamotomy.
Definition
Neurophysiologicalstudiesat thecellular level(� singleunit activity and � local field potentials or LFP)as well as � electroencephalographic (EEG) and� magnetoencephalographic (MEG) recordings pro-vide converging evidence for a thalamocortical dys-regulation at the origin of chronic � neurogenic painof both peripheral and central origin. These data sug-gest an increase of low frequency thalamocorticalrhythmicity originating in disfacilitation of thalamicrelay neurons, followed by cortical activation due toasymmetries of corticocortical inhibition. The process,called � thalamocortical dysrhythmia (TCD) may be-come self-sustained and thus chronic, due to recurrentthalamoreticulothalamic and corticoreticulothalamicfeedback inhibition. The surgical approach presentedhere is centered on a re-establishment of a normal tha-lamocortical oscillatory activity using a strategicallyplaced medial thalamic lesion, which reduces or abol-ishes the TCD via low frequency desamplification andprovides long term therapeutic efficiency coupled withsparing of the specific � thalamocortical modules. Thisphysiopathological framework underscores the risksrun by any surgical procedure aiming at further reducingthe activation of specific thalamic relay cells and thusincreasing thalamic disfacilitation and dysrhythmicpain mechanisms. The present essay is thus focused on� medial thalamotomies, more specifically on centrallateral thalamotomy (CLT).
In 1989, Lenz and collaborators (Lenz et al. 1989)provided the first evidence for the presence of � low-threshold calcium spike (LTS) bursts in the thalamusof patients suffering from neurogenic pain. They foundthem in the somatosensory � ventral posterior (VP)complex, localized in and around the portion of VPrepresenting the deafferented and thus painful bodypart (Lenz et al. 1994). The presence of the same activi-ties was also described widely spread in and around theposterior part of the � central lateral nucleus (CL) of themedial thalamus (Jeanmonod et al. 1993; Jeanmonod etal. 1994; Jeanmonod et al. 1996). An example of suchLTS bursting activity in CL is shown in Fig. 1 a–c), withthe typical progressive increase in duration of each suc-cessive interspike interval and the inverse relationshipbetween the duration of the first interspike interval andthe number of spikes in a burst. Furthermore, it wasshown that: 1) half of the recorded neurons presentedLTS bursting activity, 2) only a minority (less than 1%) had somatosensory receptive fields, 3) LTS burstsdisplayed a theta rhythmicity, with a mean interburstdischarge rate of 4 Hz, and 4) there were no significantdifferences between recordings performed in patientssuffering from peripheral and central neurogenic pain.Arecentanalysis(Sarntheinetal.,2003)ofthalamicLFPrecordings in CL showed the presence of a high � theta(4–8 Hz) power (Fig. 1d), correlating closely with thetheta rhythmicity displayed by LTS bursts. In addition,an increase in the cortical theta power was recorded byEEG (Fig. 1d) and a high coherence between EEG andthalamic theta activities was found (Fig. 1e). This under-scores the expected high level of functional coupling be-tween thalamus and cortex. In Fig. 1f, EEG power spec-tra of patients have been averaged and compared withthose of controls, confirming the existence of a disease-related increase in theta power.Based on converging evidence from experimental andclinical data over the last 20 years (Llinás and Jahnsen1982; Steriade et al. 1990; Jeanmonod et al. 1993; Jean-monod et al. 1996; Jeanmonod et al. 2001; Steriade etal. 1997; Llinás et al. 1999; Llinás et al. 2001), a thala-mocortical concept of chronic neurogenic pain was ini-tially proposed at the thalamic level (Jeanmonod et al.1993), and extended to the cortical level, with the de-nominationof thalamocorticaldysrhythmia(Llinásetal.1999). It is characterized by the following sequential setof events.
1. Bottom-up or top-down disfacilitation by deaf-ferentation of excitatory inputs onto thalamic relaycells through a somatosensory lesion, either periph-eral or central, is at the source of the neurogenic painsyndrome. This results in cell membrane hyperpo-larization.
2. In thishyperpolarizedstate,deinactivationofcalciumT-channels causes thalamic relay neurons to fire LTSbursts at theta frequency.

T
Thalamotomy for Human Pain Relief 2433
Thalamotomy for Human PainRelief, Figure 1 (a, b, c) LTSbursts. (a) LTS bursting cell recordedin the posterior part of CL. Note theprogressive increase in duration ofeach successive interspike interval(ISI). (b) Progressive increase of theISI within the burst. (c) Logarithmicdecrease of the length of the firstISI as a function of the numberof spikes in the burst. (d, e, f)Spectral analysis of LFP and EEG.(d) LFP and EEG power spectra ofone neurogenic pain patient (powerunits: μV2/Hz). (e) Coherencebetween EEG and LFP in the samepatient. (f) Power spectra of scalpEEG recordings in 11 patients(mean age 59 +/- 13 years) and12 healthy controls (mean age56 +/- 10 years). Spectra wereaveraged over subjects.
3. These thalamic neurons impose a theta rhythmicityto the thalamocortical modules they are part of, asdemonstrated by theta power increases in both LFPand EEG recordings. The tight functional couplingbetween thalamus and cortex is confirmed by the hightheta coherence between the two. This coupling is notonly sustained by thalamocorticothalamic, but alsoby thalamoreticulothalamicandcorticoreticulothala-mic recurrent projections.
4. Divergent thalamocortical, corticothalamicandretic-ulothalamic projections provide the anatomical ba-sis for the coherent diffusion of low frequencies toan increasing number of neighboring thalamocorti-cal modules. This phenomenon may explain the fre-quently observed delay between theoccurrenceof thecausal insult and the beginning of pain.?>
5. The final step consists in the activation of high fre-quency (beta and gamma) cortical domains in thevicinity of low frequency theta areas: constrainingcorticocortical GABAergic inhibitory interneuronsto theta rhythmicity may indeed reduce lateral in-hibitory drive. This leads to disinhibition and thusactivation of neighboring thalamocortical modules(edge effect), with production of pain sensation.MEG (Llinás et al. 1999) and LFP power correlationstudies as well as LFP bicoherence data (Sarnthein etal. 2003) provide evidence for such a phenomenon,showing an increased interfrequency coherence be-tween theta and beta domains and thus indicating acoupling of low and high frequency activities.
Characteristics
Exploration of the human medial thalamus allowed theidentification of a zone located in the posterior part ofthe CL, which harbored a large majority of the recordedLTS bursts (Jeanmonod et al. 1993; Jeanmonod et al.1994). Considering in addition the evidence for lowfrequency recruitment by CL stimulation (Morison andDempsey 1942) and surgical experience in the medialthalamus, a medial thalamic target was redefined, cen-tered in the posterior part of the CL (for review, seeJeanmonod et al. 2001). This target aims at render-ing the low frequency thalamocortical power increasesubcritical by reducing theta overamplification andoversynchronization, without reducing the specificand remaining unaffected non-specific thalamocorticalloops. A magnetic resonance- and microelectrode-guided stereotactic thalamotomy in the posterior partof the CL (central lateral thalamotomy or CLT, Fig. 2a)was implemented in 96 patients (Jeanmonod et al. 2001)suffering from chronic therapy-resistant peripheral orcentral neurogenic pain (mean age 56 years; mean painduration before surgery 7.5 years). At a mean follow-upof 3 years 9 months, 53% of the patients benefited froma relief of more than 50% (Figure 2b). Patients withcontinuous pain showed only a mean relief of 20% incontrast to the 66% obtained for patients with phasicor intermittent pain manifestations. Allodynia was sup-pressed in 57% of the patients. There was only a trendfor better relief in patients with peripheral neurogenic

2434 Thalamotomy for Human Pain Relief
Thalamotomy for Human Pain Relief, Figure 2 (a) Sagittal atlas sec-tion (Morel et al., 1997) 7.2 mm distant from the ventricular border. Thehorizontal scale line is aligned to the intercommissural plane, with a crossindicating the level of the posterior commissure. Scale: 1 mm betweengraduations. The shaded area displays the position and extent of CLT, cen-tered on the penetration track (dashed line). Abbreviations: AV, anteroven-tral; CL, central lateral; CM, centre médian; LD, lateral dorsal; Li, limitans;MDpc, parvocellular part of the mediodorsal nucleus; Pf, parafascicular;PuM, medial pulvinar; VAmc, magnocellular part of the ventral anterior (VA)nucleus; VLpd and VLpv, dorsal and ventral divisions of the ventral lateralposterior nucleus; VM, ventral medial; VPMpc, parvocellular part of theventral posterior medial nucleus. ZI, zona incerta. (b) Histogram displayingthe clinical results of the CL thalamotomy for 96 patients with a meanfollow-up of 3 years and 9 months.
pain. An increase of continuous pain relief from 14 to49% was observed when CLT was performed bilater-ally instead of only contralaterally. The CLT procedureproduced no somatosensory deficits and entailed norisk of pain increase, but only the usual vascular riskrelated to the stereotactic procedure. The localizationand expanse of the posterior part of the CL, away fromneurologically eloquent areas, allow the placing ofthe therapeutic lesion with minimal risks for neuro-logical functions. Extension of bleeding into adjacentstructures, such as the centre médian-parafascicularcomplex, posterior complex or medial pulvinar, doesnot correlate with observable deficits and could evencontribute to pain relief. The medial location of the CLtarget guarantees a good distance away from the VPand lateral spinothalamic tract and its anteroposteriorcoordinate is posterior enough to avoid a significant en-croachment on the mediodorsal nucleus. At the ventrallimit of the target, 1–2 mm may be left intact in the area
of the limitans nucleus to avoid any intrusion into thepretectum below.From a review of the literature (Jeanmonod et al. 2001),relief percentages between 50 and 100% were obtainedin averages of 53, 47 and 53% after dorsal column stimu-lation, VP stimulation and motor cortex stimulation re-spectively. These results thus come close to those ob-tained after CLT. In our study, all patients selected clin-ically ashaving aneurogenicpaindiagnosisare includedin the results. To compare with stimulation studies, it isnecessary to include in the failure percentages of thesestudies those patients who received only a temporary un-satisfactory stimulation test, thus showing an immediateresistance to stimulation. In many studies, quoted suc-cess rates are based only on the patients who underwentpermanent implantation. A correction integrating all pa-tients with and without permanent implantation reducespain relief percentages very significantly.The fact that CLT does not produce clinically relevantdeficits suggests that the posterior part of the CL is nolonger normally functional, i.e. only serves as a gener-ator of low frequencies. This is supported by the unre-sponsiveness of 99% of the recorded cells (see above)and implies a redistribution of its functions to other tha-lamic nuclei. Such a transfer may be all the more com-plete in our patients who have been operated on afteryears of suffering. CLT being restricted to the medialthalamic area harboring LTS activities and exhibiting awidespread functional block leaves other medial thala-mic nuclei, all potential candidates for the redistributionof CL functions, intact. We are thus facing the paradoxi-cal and conceptually intriguing situation where a lesionnot only produces no or only clinically irrelevant deficitsbut also results in beneficial effects by the suppressionof a dysfunctional area (Welsh 1998).The therapeutic possibilities of other medial thalamictargets, particularly the posterior complex, the medialpulvinar and the centre médian-parafascicular complexseem, inourexperience, tobeinferior tothoseoftheCLT.We have, however, not explored these areas sufficientlyto make a definitive statement.
References1. Jeanmonod D, Magnin M, Morel A (1993) Thalamus and neu-
rogenic pain: physiological, anatomical and clinical data. Neu-roreport 4:475–478
2. Jeanmonod D, Magnin M, Morel A (1994) A thalamic conceptof neurogenic pain. In: Gebhart GF, Hammond DL, Jensen TS(eds) Progress in Pain Research and Management. IASP Press,Seattle, pp 767–787
3. Jeanmonod D, Magnin M, Morel A (1996) Low-thresholdcalcium spike bursts in the human thalamus. Common phys-iopathology for sensory, motor and limbic positive symptoms.Brain 119:363–375
4. Jeanmonod D, Magnin M, Morel A et al. (2001) Surgical controlof the human thalamocortical dysrhythmia: I. Central lateral tha-lamotomy in neurogenic pain. Thalamus Related Syst 1:71–79
5. Lenz FA, Kwan HC, Dostrovsky JO et al. (1989) Characteris-tics of the bursting pattern of action potentials that occurs in thethalamus of patients with central pain. Brain Res 496:357–360

T
Thalamotomy, Pain Behavior in Animals 2435
6. Lenz FA, Kwan HC, Martin R et al. (1994) Characteristics ofsomatotopic organization and spontaneous neuronal activity inthe region of the thalamic principal sensory nucleus in patientswith spinal cord transection. J Neurophysiol 72:1570–1587
7. Llinás R, Jahnsen H (1982) Electrophysiology of mammalianthalamic neurones in vitro. Nature 297:406–408
8. Llinás RR, Ribary U, Jeanmonod D et al. (1999) Thalamocorti-cal dysrhythmia: A neurological and neuropsychiatric syndromecharacterized by magnetoencephalography. Proc Natl Acad SciUSA 96:15222–15227
9. Llinás R Ribary U, Jeanmonod D et al. (2001) Thalamocorti-cal dysrhythmia I. Functional and imaging aspects. ThalamusRelated Syst 1:237–244
10. Morel A, Magnin M, Jeanmonod D (1997) Multiarchitectonicand stereotactic atlas of the human thalamus. J Comp Neurol387:588–630
11. Morison R, Dempsey E (1942) A study of thalamo-cortical re-lations. Am J Physiol 135:281–292
12. Sarnthein J, Morel A, von Stein A et al. (2003) Thalamic thetafield potentials and EEG: high thalamocortical coherence in pa-tients with neurogenic pain, epilepsy and movement disorders.Thalamus Related Syst 2:231–238
13. Steriade M, Jones EG, Llinas R (1990) Thalamic oscillations andsignalling. Wiley, New York
14. Steriade M, Jones EG, McCormick DA (1997) Thalamus: Or-ganisation and Function. Elsevier, Oxford
15. Welsh JP (1998) Systemic harmaline blocks associative and mo-tor learning by the actions of the inferior olive. Eur J Neurosci10:3307–3320
Thalamotomy, Pain Behavior in AnimalsNAYEF E. SAADÉ, SUHAYL J. JABBUR
Neuroscience Program, Faculty of Medicine, AmericanUniversity of Beirut, Beirut, [email protected]
Definition
Textbooks of neurology continue to regard the tha-lamus as an obligatory relay station for all sensorypathways (except olfaction) on their way to the cerebralcortex. Moreover, the thalamus has been considered tobe a most important brain center for the perception ofpain (Head and Holmes 1911). Over the last century,several clinical reports described disturbed sensationsand spontaneous pain produced by thalamic lesion, apathology labeled “thalamic syndrome” (Dejerine andRoussy 1906). Therefore, clinical and experimentalinvestigations have been devoted to the understandingof the mechanisms underlying sensory disturbancesfollowing thalamic lesions.
Characteristics
The work of Roussy (Roussy 1907) was among the firstattempts to reproduce the “thalamic syndrome” in an-imals by controlled thalamic lesions (thalamotomies).The stereotaxic method and the techniques used for theplacement of small or large experimental lesions in thethalamusareexplained in� Post-strokepain model, tha-lamic pain (lesion) (lesion).
Placement of a controlled lesion in the human thalamuscannot be performed for experimental purposes. There-fore, studies involving partial or total thalamotomies canonly be performed on experimental animals under con-trolled conditions and with strict adherence to ethicalguidelines for pain experimentation on animals. The ex-perimental protocol should take into consideration theeffects of lesions placed in different thalamic nucleargroups known to be involved in the processing of vari-ous aspects of pain. These areas can be classified undertwo major headings, the lateral and the medial nucleargroups.Theprotocol should also bebased on special ani-malmodelssimulatingdifferentpainconditions(includ-ing acute and chronic pain) and using appropriate teststhat allow the assessment of the different qualities of no-ciception (such as mechanonociception or thermonoci-ception etc.).Despite abundant clinical literature on the thalamic syn-drome (about 820 studies, listed in a recent Pub Medsearch), few studies have been devoted to the investiga-tion of the effects of various thalamic lesions on noci-ceptive behavior in animals. The early work by Dela-cour and Borst (Delacour and Borst 1972) showed thatlesions of centromedian and parafascicular nuclei of thethalamus did not interfere with the escape reaction tonoxious stimuli, but suppressed learning and avoidanceconditioning. Mitchell and Kaelber (Mitchell and Kael-ber1967)alsoshowedthatlesionofmorethan50%ofthesame (plus adjacent) thalamic nuclei in cats is necessaryto block the escape reaction to grid electrical shock stim-ulation. More recently, Casey and Morrow (Casey andMorrow 1983) showed that bilateral medial thalamic le-sions increased nociceptive responses in cats. This find-ing received further confirmation by the reported effectsof lesions of the nucleus submedius in rats (Roberts andDong 1994). A more recent study by Norrsell and Craig(Norrsell and Craig 1999) however, reported a mild ther-mosensory deficiency in cats subjected to � electrolyticlesions placed in the various medial nuclear groups (nu-cleus submedius, posterior medial nucleus, basal ven-tral medial nucleus). It is important to note that the elec-trolytic lesions used can involve neuronal cell bodies inaddition to passing fibers, which can have a functionalrole different from that of the lesioned centers.Recent work from our laboratory has aimed at studyingthe effects of lesions of various sizes and locations inthe thalamus on the nociceptive behavior in rats. Theselesionswereperformed by injecting either electrical cur-rent (electrolytic) or excitotoxic substances (for selec-tive lesion of cell bodies). Their effects were assessed onacute nociceptive reactions and on a rat � animal modelfor mononeuropathy (Saadé et al 1999).Chronic unilateral lesions, which were either subtotalor placed in the lateral or the medial thalamus producedsignificant and persistent decreases in the mechanicaland thermal nociceptive thresholds as assessed by the� paw pressure test (PP) and � hot plate (HP) tests, re-

2436 Thalamotomy, Pain Behavior in Animals
Thalamotomy, Pain Behavior in Animals, Figure 1 Temporal evolutionof the effects of lesions placed in the lateral thalamus of rats. Mechanical(paw pressure) and thermal (hot plate) nociceptive thresholds. The latenciesof the nociceptive tests elicit a significant decrease (hyperalgesia) after thelesion (time 0) when compared to the latencies of the same test observedin intact rats (control).
spectively (Fig. 1). No significant differences were no-ticed between the effects of electrolytic and excitotoxiclesions. Furthermore, the increased nociception was bi-lateral (Fig. 1) and more pronounced on supraspinallycoordinated nociceptive tests such as the HP. Thalamiclesions also exerted differential effects on the aversivebehavior induced by intraplantar injection of 0.05 ml offormalin 2.5%, known as the � formalin test (Dubuis-son and Dennis 1977). The most pronounced effect, ob-served as increases in the nociceptive scores, was ob-tained with subtotal thalamic lesion (Fig. 2).The effects of either lateral or medial thalamic le-sions on the neuropathic behavior observed in a rat� animal model for mononeuropathy were also ex-amined. Neuropathy was induced by selective lesionof two components (peroneal and tibial nerves) of thesciatic nerve supplying the hind leg and sparing the third
Thalamotomy, Pain Behavior in Animals, Figure 2 Summary of theeffects of different types of thalamic lesions on the formalin tests performedon three groups of rats subjected to thalamic lesions as compared to a fourthintact group (sham). (top) A composite drawing showing a microscopic viewof a transverse section of a rat brain (right) and schematic drawing (left)illustrating the placement of lesions in the thalamus. (bottom) Each barrepresents the average nociceptive score measured during a period of12 min in phase I (early phase 3–15 min), or phase II (late or tonic phase30–42 min) in the various groups of rats.
component (the sural). This model is characterized by apersistent � allodynia (nociceptive reaction induced bynon-noxious stimulus) and � hyperalgesia (increasedreactivity to a noxious stimulus). Both electrolytic orexcitotoxic lesions placed either in the lateral or medialthalamic sensory nuclei produced transient decreasesin neuropathic manifestations, which recovered theirlevels before thalamotomy within one or two weeks(Fig. 3). However, when mononeuropathy was induced1 or 2 weeks after thalamic lesions, allodynia and hyper-algesia developed without any significant differences(in intensity or temporal evolution) from those observedin animals with an intact thalamus.Although it is difficult to produce exact simulation ofclinical syndromes such as spontaneous pain by animalmodels, the batteries of tests employed appear to reflectsignificant changes in nociceptive reactivity in animals

T
Thalamus 2437
Thalamotomy, Pain Behavior in Animals, Figure 3 Transient attenuation of neuropathic manifestations by electrolytic lesions in the lateral thalamicnuclei. Each lesion was performed at day zero on a group of rats subjected to mononeuropathy. Tactile allodynia and heat hyperalgesia were decreasedduring the first 2 weeks, after which they recovered to their pre-lesion levels. Cold allodynia was not affected (Saadé et al., unpublished data).
subjected to thalamic lesions. Starting with the earliestanimal experiments by Roussy in 1907, the outcome oflesions in sensory thalamicnucleioften resulted in eithera decrease or no change in the nociceptive thresholds. Itbecomes necessary, therefore, to explain the resultingparadox as to how a function of a center can be eitherunaffected or exaggerated when that center is ablated?Possible answers to this question include the fact thata lesion to a nervous center can either affect, in vary-ing proportions, inhibitory and excitatory mechanismsor lead to plastic changes that adjust and compensate forthe insult or injury.� Lateral Thalamic Lesions, Pain Behavior in Animals
References1. Casey KL, Morrow TJ (1983) Supraspinal pain mechanisms in
the cat. In: Kitchell KL, Erickson HH (eds) Animal Pain Percep-tion and Alleviation. American Physiological Society, Bethesda,Maryland, USA, pp 63–82
2. Dejerine J, Roussy G (1906) Le syndrome thalamique. Rev Neu-rol 14:521–532
3. Delacour J, Borst A (1972) Failure to find homology in rat, cat,and monkey for functions of a subcortical structure in avoidanceconditioning. J Comp Physiol Psychol 80:458–468
4. Dubuisson D, Dennis SG (1977) The formalin test: a quantitativestudy of the analgesic effects of morphine, meperidine and brainstem stimulation in rats and cats. Pain 4:161–174
5. Head H, Holmes G (1911) Sensory disturbances from cerebrallesions. Brain 34:102–254
6. Mitchell CL, Kaelber WW (1967) Unilateral vs bilateral medialthalamic lesions and reactivity to noxious stimuli. Arch Neurol17:653–660
7. Norrsell U, Craig AD (1999) Behavioral thermosensitivity afterlesions of thalamic target areas of a thermosensory spinothalamicpathway in the cat. J Neurophysiol 82:611–625
8. Roberts VJ, Dong WK (1994) The effect of thalamic nucleussubmedius lesions on nociceptive responding in rats. Pain57:341–349
9. Roussy G (1907) Le syndrome thalamique. In: G. Steinheil: LaCouche Optique. Paris
10. Saadé NE, Kafrouni AI, Saab CY et al. (1999) Chronic thalam-otomy increases pain-related behavior in rats. Pain 83:401–409
Thalamus
Definition
ThethalamusisderivedfromtheGreekword‘Thálamos’(bedroom, chamber). It represents the biggest structure

2438 Thalamus and Cardiac Pain
of the diencephalon. Its two hemispheres are locatedin the center of the brain next to the third ventricle. Ithas reciprocal connections with the cerebral cortex andrelays sensory signals from all senses except that ofolfaction to the cerebral cortex, and is also involved inmotor, arousal and mood functions.� AnginaPectoris,NeurophysiologyandPsychophysics� Deep Brain Stimulation� Lateral Thalamic Pain-Related Cells in Humans� Pain Treatment, Intracranial Ablative Procedures� Prefrontal Cortex, Effects on Pain-Related Behavior� Spinothalamic Projections in Rat
Thalamus and Cardiac Pain
� Thalamus, Clinical Visceral Pain, Human Imaging
Thalamus and Gastrointestinal Pain
� Thalamus, Clinical Visceral Pain, Human Imaging
Thalamus and Pain
Definition
Thalamic structures (including intralaminar nuclei,principle sensory nucleus, nuclei posterior to it includ-ing supra-geniculate, posterior, ventralmedial-posteriornucleus) that have pain-related activity, as identified byanatomic and physiologic studies in primates.� Pain Treatment, Motor Cortex Stimulation
Thalamus and Visceral Pain (PositronEmission Tomography or FunctionalMagnetic Resonance Imaging)
� Thalamus, Clinical Visceral Pain, Human Imaging
Thalamus and Visceral Pain Processing(Human Imaging)
ANTHONY R. HOBSON, QASIM AZIZ
Section of GI Sciences, Hope Hospital, University ofManchester, Manchester, [email protected]
Definition
Whilst there is ample evidence from animal studiesto support the role of the thalamus in visceral pain
processing (Al-Chaer et al. 1998; Willis and Westlund1997), studies in humans have, until recently, been lim-ited to intra-operative observations from patients withimplanted deep brain stimulating electrodes (Lenz et al.1994). However, the availability of � functional brainimaging techniques (FBI) such as � positron emis-sion tomography (PET) and � functional magneticresonance imaging (fMRI), has allowed visceral painresearchers to confirm that the thalamus is not only animportant relay station for visceral pain transmission,but may also be pivotal in the generation of symptomsin several visceral pain conditions.
Characteristics
To date, information regarding the brain processingof visceral sensations and pain has been obtained fol-lowing stimulation of the esophagus, stomach, rectum,bladder and vagina, in addition to that acquired during� dobutamine induced chest pain (Aziz et al. 1997;Hobday et al. 2001; Ladabaum et al. 2001; Matsuuraet al. 2002; Rosen et al. 1996; Whipple and Komisaruk2002). In healthy subjects, thalamic activity has onlybeen reported in approximately 50% of studies, and thishas been predominantly in response to noxious ratherthan innocuous visceral stimulation.Strigo et al. have compared the processing of visceraland cutaneous pain in the human brain using fMRI(Strigo et al. 2003). In this study, esophageal distensionand contact heat stimulation of the anterior chest wallwere matched for subjective intensity and applied tohealthy subjects in a counterbalanced order. Analysisrevealed that whilst differences were seen in corticalregions such as the anterior insula, a common painneural network was activated encompassing the secondsomatosensory cortex, posterior parietal cortex, basalganglia and thalamus. The authors concluded that thesimilar activations seen within these regions implicatetheir function in the identification of a stimulus aspainful, rather than in the differentiation between thenature of painful stimuli (Strigo et al. 2003).In a PET study that examined the effect of increasing in-tensity of gastric distension on cortical and sub-corticalactivation, Ladabaum et al. reported significant thalamicactivity only at the highest distension volumes, whichproduced a noxious stimulus. Peak activations werenoted bilaterally in the ventral posteriolateral (VPL)nuclei and in the left dorsomedial nuclei (Ladabaum etal. 2001) leading the authors to conclude that noxiousgastric stimulation activates both the lateral and medialpain systems.The ability to non-invasively map the neuroanatomyof the visceral pain matrix has lead to several re-searchers embarking on clinical studies in patientswith � functional gastrointestinal disorders, predom-inantly in patients with � irritable bowel syndrome(IBS). Patients with IBS commonly report heightenedperception of rectal distension (� visceral hypersensi-

T
Thalamus and Visceral Pain Processing (Human Imaging) 2439
tivity), and the aim of these studies has been to identifyobjective neural correlates of visceral hypersensitivity.Whilst the heterogeneity of the IBS population hasmeant that results from group imaging data have beeninconsistent, several groups have shown increasedthalamic activation in response to noxious rectal dis-tension (Mertz et al. 2000; Verne et al. 2003; Yuan etal. 2003). The thalamic regions of interest identifiedin these studies encompassed the VPL and dorsome-dial nuclei. The limited spatial resolution of the fMRIimaging techniques used in these studies meant that itwas not possible to comment on activation of specificthalamic nuclei, and thus no comment could be madeon the specific contribution of the sensory and affec-tive dimensions of pain. Future clinical studies usinghigh resolution imaging of the thalamus may be ableto provide more information regarding the contributionof different thalamic regions to aberrant visceral painprocessing in IBS.Perhaps the most dramatic evidence to support the roleof the thalamus in aberrant visceral pain processingcomes from studies by Rosen et al. in patients withvarious forms of � angina pectoris (Rosen et al. 1996,2002). A previous case report from 1994 had reportedthat microstimulation of the VPL induced angina likesymptoms in a single patient, which was not coincidentwith changes in cardiovascular function, strongly im-plicating the VPL in visceral and referred pain (Lenzet al. 1994). Rosen followed this up with a PET studyin which he measured brain activity during intravenousdobutamine infusion in patients with coronary arterydisease and active angina (Rosen et al. 1994). Thesedata revealed that dobutamine induced angina wasassociated with bilateral activation of the thalamus,in addition to activation of a number of higher corti-cal structures. Following the cessation of dobutamineinfusion, symptoms ceased and no cortical activity
Thalamus and Visceral Pain Processing (Human Imaging), Figure 1This image shows bilateral activation of the thalamus following painfulesophageal balloon distension. In addition, activity is also seen in the pri-mary / secondary somatosensory cortex and insula. These four regionsare robustly activated following noxious visceral stimulation.
was seen; however, thalamic activity was still evident.The authors concluded that the thalamus acted as agateway for afferent visceral pain signals; however,cortical activity was needed to bring this to a level ofconscious perception. Further evidence to support thishas been provided by additional studies in patients withsilent ischemia and � syndrome X (Rosen et al. 1996,2002).In summary, painful visceral stimuli produce bilateralactivation of the thalamus incorporating the regions ofthe VPL and dorsomedial nuclei. Heightened activationof these regions has been associated with aberrant painprocessing in patients with gastrointestinal and cardiacdisease. As the spatial resolution of FBI techniques im-proves, itwillbepossible to study the thalamus in greaterdetail shedding further light on its role in visceral painprocessing.
References1. Al-Chaer ED, Feng Y, Willis WD (1998) A Role for the Dor-
sal Column in Nociceptive Visceral Input into the Thalamus ofPrimates. J Neurophysiol 79:3143–3150
2. Aziz Q, Andersson JL, Valind S et al. (1997) Identification of Hu-man Brain Loci Processing Esophageal Sensation using PositronEmission Tomography. Gastroenterology 113:50–59
3. Hobday DI, Aziz Q, Thacker N et al. (2001) A Study of theCortical Processing of Ano-Rectal Sensation using FunctionalMRI. Brain 124:361–368
4. Ladabaum U, Minoshima S, Hasler WL et al. (2001) GastricDistention Correlates with Activation of Multiple Cortical andSubcortical Regions. Gastroenterology 120:369–376
5. Lenz FA, Gracely RH, Hope EJ et al. (1994) The Sensation ofAngina can be Evoked by Stimulation of the Human Thalamus.Pain 59:119–125
6. Matsuura S, Kakizaki H, Mitsui T et al. (2002) Human Brain Re-gion Response to Distention or Cold Stimulation of the Bladder:A Positron Emission Tomography Study. J Urol 168:2035–2039
7. Mertz H, Morgan V, Tanner G et al. (2000) Regional CerebralActivation in Irritable Bowel Syndrome and Control Subjectswith Painful and Non-Painful Rectal Distension. Gastroenterol-ogy 118:842–848
8. Rosen SD, Paulesu E, Frith CD et al. (1994) Central NervousPathways Mediating Angina Pectoris. Lancet 344:147–150
9. Rosen SD, Paulesu E, Nihoyannopoulos P et al. (1996) SilentIschemia as a Central Problem: Regional Brain Activation Com-pared in Silent and Painful Myocardial Ischemia. Ann Intern Med124:939–949
10. Rosen SD, Paulesu E, Wise RJ et al. (2002) Central Neural Con-tribution to the Perception of Chest Pain in Cardiac SyndromeX. Heart 87:513–519
11. Strigo IA, Duncan GH, Boivin M et al. (2003) Differentiation ofVisceral and Cutaneous Pain in the Human Brain. J Neurophysiol89:3294–3303
12. Verne GN, Himes NC, Robinson ME et al. (2003) Central Rep-resentation of Visceral and Cutaneous Hypersensitivity in theIrritable Bowel Syndrome. Pain 103:99–110
13. Whipple B, Komisaruk BR (2002) Brain (PET) Responses toVaginal-Cervical Self-Stimulation in Women with CompleteSpinal Cord Injury: Preliminary Findings. J Sex Marital Ther28:79–86
14. Willis WD, Westlund KN (1997) Neuroanatomy of the Pain Sys-tem and of the Pathways that Modulate Pain. J Clin Neurophysiol14:2–31
15. Yuan YZ, Tao RJ, Xu B et al. (2003) Functional Brain Imagingin Irritable Bowel Syndrome with Rectal Balloon-Distention byusing fMRI. World J Gastroenterol 9:1356–1360

2440 Thalamus, Clinical Pain, Human Imaging
Thalamus, Clinical Pain, Human ImagingA. VANIA APKARIAN
Feinberg School of Medicine, Department ofPhysiology, Northwestern University, Chicago, IL,[email protected]
Synonyms
Chronic pain; BOLD activity; voxel-based morphome-try; Gray Matter Density
Definition
Non-invasive human brain imaging provides the oppor-tunity to examinecentralprocesses thatmay becriticallyinvolved in the induction and/or maintenance of clinicalchronic pain conditions. Such studies point to the notionthat the thalamus shows reduced signaling and reducedgray matter, suggesting that the region isat least anactiveplayer in chronic pain conditions.
Characteristics
The advent of non-invasive brain imaging technologiesaffords a unique opportunity for unraveling brain pro-cesses that may be critical in induction and/or mainte-nance of clinical chronic pain conditions. Such studieshave the potential for replacing speculations, psychoso-cial interpretations and accusations of patients by dub-bing them malingerers and other such labels, by physio-logicalparameters thatcharacterize theseconditionsandthen hopefully lead to new, more science based devel-opment of therapies. Here we briefly discuss the currentunderstanding of the role of the thalamus in clinical painstates based on human brain imaging studies.Brain activity as determined by � PET or � fMRI hasestablished a reproducible pattern of cortical activityassociated with acute or experimental painful condi-tions. A recent meta-analysis, of such studies over thelast 15 years estimated that the incidence of reporting ofthalamic activity in experimental pain conditions is 84%(16/19 studies) in PET studies and 81% (13/16 studies)in fMRI studies (Apkarian et al. 2005) (incidence hereis the ratio of the number of studies where the area wasinvestigated in contrast to the number of studies wherethe area was reported to be activated). In contrast to thisvalue, when the incidence of thalamic activity is exam-ined in brain imaging studies in clinical pain conditionsthe incidence is 59% (16/27 PET, � SPECT and fMRIstudies combined). The conditions included in the clin-ical cases are cancer pain, cluster headache, migraine,cardiac pain, irritable bowel syndrome, fibromyalgia,CRPS, phantom pain and mono- or poly-neuropathies(Apkarian et al. 2005). Contrasting the incidence ofthalamic activity in normal subjects to clinical painconditions indicates a borderline significant decreasein clinical conditions (p = 0.09, Fisher’s exact test).
The result seems paradoxical since one would naturallyassociate decreased thalamic activity with decreasedpain and not the other way. We argue below that thisobservation is consistent with the notion that chronicpain conditions are more emotional states and hence lesssensory; as a result they may involve less spinothalamicactivation and enhanced activations through pathwaysmore directly accessing emotional regions of the brain.The first clinical pain study was done in 5 patients withchronic cancer pain, where brain activity as determinedby PET was compared between them and normal sub-jects before and after high cervical cordotomies thatresulted in significant pain relief (Di Piero et al. 1991).The main outcome of the study was the observationthat thalamic activity was low in the patients duringchronic pain and normalized after the cordotomy. Atleast another 5 studies report that chronic clinicalpain conditions are associated with decreased baselineactivity or decreased stimulus related activity in thethalamus. A SPECT blood flow study (Fukumoto et al.1999) has shown a strong relationship between time ofonset of CRPS symptoms and thalamic activity. Theratio between contralateral and ipsilateral thalamic per-fusion was larger than 1.0, indicating hyperperfusion,for patients with symptoms for only 3–7 months andsmaller than 1.0, indicating hypoperfusion, for patientswith longer-term symptoms (24–36 months), with acorrelation coefficient of 0.97 (normal subjects had athalamic perfusion ratio of about 1.0). These resultsstrongly imply that the thalamus undergoes adaptivechanges in the course of CRPS. Thus, it can be assertedthat thalamic activity for pain in chronic clinical condi-tions is different from that for acute painful stimuli innormal subjects.Proving long-term reorganization of the CNS is hardwith functional imaging, since one cannot disentanglereorganization from modulation of responses due to thepresence of the condition, for example in chronic pain.On the other hand, examination of brain chemistry by� MRS indicates changes in various metabolites, mostof which are not affected by the current cognitive stateof the person. Thus, such measures document long-termchanges more readily. The limitation of the techniqueregards the specific chemicals that can be detected (thespecific functions of many of which remain unclear),and the need for regional imaging which limits thespatial extent of the measurement (Salibi and Brown1998). One such study examined thalamic metabolitesin chronic back pain patients and observed no changesin comparison to healthy subjects, although there weredecreased measures for multiple metabolites in theprefrontal cortex (Grachev et al. 2000). Another similarstudy examined thalamic metabolites (Pattany et al.2002) in patients with chronic neuropathic pain afterspinal cord injury and did see decreased metabolites inthe thalamus and observed a negative correlation be-tween pain intensity and concentration in the thalamus.

T
Thalamus, Clinical Visceral Pain, Human Imaging 2441
Since a decreased concentration of N-acetyl-aspartatehas been reported in most neurodegenerative condi-tions, it is reasonable to conclude that the observationof decreased N-acetyl-aspartate in the chronic spinalcord injury patients suggests that the condition is associ-ated with a neurodegenerative process in the thalamus.This idea was tested directly in a morphometric study.The gray matter density of chronic back pain patientswas contrasted with age and gender matched normalsubjects using high resolution anatomical MRI. Theresults indicate that the thalamus gray matter density,mainly on the right, is significantly lower in density inthe chronic back pain patients (Apkarian et al. 2004).The mechanisms inducing this atrophy remain to bedetermined, as well as the extent of its reversibility andits impact on information processing in the thalamus.However, the observation clearly implies that the thala-mus is undergoing long-term reorganization due to thepresence of the pain condition. It is also possible thatsome of this reorganization has a genetic componentthat may predispose these subjects to develop chronicpain. Thus, there are more questions raised than an-swered. Fortunately, these questions can be answeredin future studies and should provide a more accurateunderstanding of the role of the thalamus in chronicpain conditions.Decreased brain responses to painful stimuli in clinicalpain conditions have been observed in other meta-analyses of the literature (Derbyshire 1999; Peyron etal. 2000), and interpreted as evidence for a generalizeddecrease in brain activity in such patients. However,when the incidence of prefrontal cortical activity isexamined, one observes increased incidence of activat-ing this region in chronic pain patients in contrast tonormal subjects. In normal subjects, prefrontal cortex isactivated in 55% (23/42) of functional imaging studiesfor pain, while in pain patients this rate increases to81% (21/26, p =0.04, Fisher’s exact statistics). Thus,in chronic pain patients, decreased thalamic activityseems to be accompanied by increased prefrontal corti-cal activity. This implies a switch of nociceptive inputsaway from spinothalamic afferents and enhanced no-ciceptive inputs through brainstem prefrontal corticalregions. This has been suggested in animal modelsof neuropathic pain (Hunt and Mantyh 2001), basedon the response changes observed in spinal cord neu-rons following neuropathic injury. The shift suggeststhat chronic clinical pain conditions are more emo-tional/cognitive than sensory. Whether the thalamicatrophy observed in back pain patients directly con-tributes to this shift remains to be determined. It is atleast consistent with the model.Overall, multiple lines of investigations regarding theinvolvement of the thalamus in clinical pain conditionsimply that the region actively reorganizes in such con-ditions. It should be emphasized that most probablythe details of this reorganization are specific to differ-
ent types of clinical pains. The extent to which thisreorganization contributes to such conditions remainsunclear. More importantly, these observations stronglysuggest that at least part of this reorganization may be aconsequence of neuronal death and hence irreversible,pointing to the urgency for further elaborating thesedetails and for more effective therapeutic approachesfor these conditions.
References1. Apkarian AV, Sosa Y, Sonty S et al. (2004) Chronic back pain
is associated with decreased prefrontal and thalamic gray matterdensity. J Neurosci 24:10410–10415
2. Apkarian AV, Bushnell MC, Treede RD et al. (2005) Humanbrain mechanisms of pain perception and regulation in healthand disease. Eur J Pain 9:463-84
3. Derbyshire SW (1999) Meta-Analysis of Thirty-Four Indepen-dent Samples Studied Using PET Reveals a Significantly Atten-uated Central Response to Noxious Stimulation in Clinical PainPatients. Curr Rev Pain 3:265–280
4. Di Piero V, Jones AK, Iannotti F et al. (1991) Chronic pain: aPET study of the central effects of percutaneous high cervicalcordotomy. Pain 46:9–12
5. Fukumoto M, Ushida T, Zinchuk VS et al. (1999) Contralateralthalamic perfusion in patients with reflex sympathetic dystrophysyndrome. Lancet 354:1790–1791
6. Grachev ID, Fredrickson BE, Apkarian AV (2000) Abnormalbrain chemistry in chronic back pain: an in vivo proton magneticresonance spectroscopy study. Pain 89:7–18
7. Hunt SP, Mantyh PW (2001) The molecular dynamics of paincontrol. Nat Rev Neurosci 2:83–91
8. Pattany PM, Yezierski RP, Widerstrom-Noga EG et al. (2002)Proton magnetic resonance spectroscopy of the thalamus inpatients with chronic neuropathic pain after spinal cord injury.AJNR Am J Neuroradiol 23:901–905
9. Peyron R, Laurent B, Garcia-Larrea L (2000) Functional imagingof brain responses to pain. A review and meta-analysis (2000).Neurophysiol Clin 30:263–288
10. Salibi N, Brown MA (1998) Clinical MR spectroscopy. Wiley-Liss, New York
Thalamus, Clinical Visceral Pain, HumanImaging
URI LADABAUM
Division of Gastroenterology, University of California,San Francisco, CA, [email protected]
Synonyms
Thalamus and Cardiac Pain; Thalamus and Gastroin-testinal Pain; Thalamus and Visceral Pain (PositronEmission Tomography or Functional Magnetic Reso-nance Imaging)
Definition
Visceral pain arises from the internal organs, such as theheart and the gastrointestinal tract. In contrast, somaticpain arises from the skin and deeper tissues, includingmuscle. The central nervous system regions activated byvisceral pain in humans, including the thalamus, have

2442 Thalamus, Clinical Visceral Pain, Human Imaging
been studied non-invasively with � positron emissiontomography (PET) and � functional magnetic reso-nance imaging (fMRI), functional imaging techniquesthat measure increased regional cerebral blood flow asa marker of neuronal activation.
Characteristics
Thalamic activation (defined as a statistically significantincrease in regional blood flow during the condition ofinterest) has been reported in many but not all functionalcerebral imaging studies of visceral pain in humans. Thecerebral representation of visceral pain was first studiedby inducing angina in humans, and later by distendinggastrointestinal viscera to induce pain. A recent system-atic review that included inspection of published imagesby the author in addition to the results explicitly reportedby investigators found evidence for thalamic activationwith angina, noxious esophageal stimulation, gastricdistension and noxious lower gastrointestinal disten-sion in healthy volunteers and patients with � irritablebowel syndrome (IBS) (Derbyshire 2003).
Cardiac Ischemia and Angina
A pioneering � PET study reported the central nervoussystem pathways mediating � dobutamine-inducedangina in patients with coronary artery disease (Rosenet al. 1994). Compared to the resting state, regionalblood flow increased during angina in a number ofcerebral structures, including increases of 2.7% in theleft thalamus and 3.7% in the right thalamus. Afterthe resolution of angina, thalamic activity remainedsignificantly increased, whereas the cortical activityassociated with angina was no longer detected. Theauthors suggested that the thalamus receives input fromthe heart during angina and continues to receive suchinput even when angina is no longer felt, with this lessintense signal not being transmitted to the cerebralcortex and therefore not associated with consciousperception.Cerebral PET imaging was then used to gain insight intothe problem of � silent myocardial ischemia (Rosen etal. 1996). Cerebral activation patterns were comparedbetweenpatientswithstress-inducedanginaandpatientswith stress-induced myocardial ischemia but no angina.During myocardial ischemia, significant left thalamicactivation was detected in patients with angina and bi-lateral thalamic activation was detected in patients withsilent ischemia. However, much more extensive corticalactivation was seen in those with angina. Because thala-micactivationwasseen inbothgroupsofpatients, theau-thors concluded that � silent myocardial ischemia can-not be explained by peripheral nerve dysfunction. Theyproposed that abnormal central processing of afferentpain signals (possibly abnormal gating at the level ofthe thalamus), may contribute to the pathophysiology ofsilent myocardial ischemia.
Esophageal Stimulation
An early PET study of the cerebral regions involved inesophageal sensation found bilateral activations alongthe central sulcus, insulae, and frontal and parietaloperculum during non-painful esophageal distensionand more intense activations in these regions as wellas additional activation in the right anterior insularcortex and the anterior cingulate gyrus during painfuldistension (Aziz et al. 1997). Thalamic activation wasnot detected.Subsequently, a fMRI study examined the cerebralcortical response to esophageal distension or acid per-fusion (Kern et al. 1998). Acidification and distensiongenerally resulted in activations of Brodmann’s areas7, 23, 30, 32, insula, operculum and anterior cingulatecortex. Although the activated regions were similar,the temporal characteristics of the activation were dif-ferent (slower for acidification). Significant thalamicactivation was again not detected.The cortical processing of distal and proximal esopha-geal sensation has been compared using fMRI (Aziz etal. 2000). Among other differences, proximal distensionwas localized precisely to the upper chest and activatedthe left primary somatosensory cortex, whereas distaldistension was perceived diffusely over the lower chestand activated the junction of the primary and secondarysomatosensory cortices bilaterally. Significant thalamicactivation was not reported.The cerebral processing of visceral and cutaneouspains were compared using distal esophageal disten-sion and application of heat to the chest during cerebralfMRI scanning (Strigo et al. 2003). Painful esophagealdistension and painful heat stimulation both inducedstatistically significant thalamic activation on the leftand non-significant activation on the right. Overall, asimilar neural network was activated with visceral andsomatic stimulation, but notable differences were alsoapparent that probably relate to the differences in theexperience of visceral and cutaneous pain.Thedifferent resultsamong thesestudiesprobablyrelateto differences in experimental design, study populationand chance. Regional cerebral blood flow changes with“activation”aregenerallysmall (a fewpercent).Becausethe thalamus is a major relay and processing station be-tween the periphery and the cortex and basal ganglia andbecause thalamic activation has been reported in a sub-stantial proportion of all available visceral pain studiesin humans, it is likely that the lack of significant thalamicactivation in the first three studies reflects the limitationsof the available techniques.
Gastric Stimulation
Cerebral PET imaging during increasing levels ofgastric distension detected progressive increases inactivation in multiple cortical and subcortical regions,including the thalami, insulae, anterior cingulate cortex,periaqueductal gray matter and cerebellum (Ladabaum

T
Thalamus, Clinical Visceral Pain, Human Imaging 2443
et al. 2001). Statistically significant activation centeredin the right ventral posterolateral (VPL) nucleus ofthe thalamus was detected with distension producingthreshold pain as well as distension producing moder-ate pain. Statistically significant activations centeredin the VPL and dorsomedial (DM) nuclei of the leftthalamus were detected with distension producingmoderate pain. The VPL nucleus is a key componentof the lateral pain system, which is believed to subservethe sensory-discriminative component of pain and theDM nucleus is part of the medial pain system, whichis believed to mediate the affective-motivational com-ponent of pain. The identification of precise thalamicnuclei as the regions of peak activation must be inter-preted with caution given the spatial resolution limitsof PET.
Rectosigmoid Stimulation
A pioneering PET study of visceral sensation in healthyvolunteers and patients with IBS found thalamic acti-vation of borderline significance during rectal disten-sion in healthy volunteers (Silverman et al. 1997). Anearly fMRI study detected activation in numerous re-gions including the anterior cingulate, prefrontal, insu-lar and sensorimotor cortices during rectal distension inhealthy volunteers, but no thalamic activation was de-tected (Baciu et al. 1999).A larger study of healthy volunteers (16) and IBS pa-tients (18) using fMRI during rectal distension reportedincreased activity in the anterior cingulate, prefrontal,and insular cortices, as well as increased thalamic activ-ity in nearly all subjects (Mertz et al. 2000). In contrast toearlier results (Silverman et al. 1997), this study foundcomparable patterns of activation in healthy volunteersand IBS patients, but greater activation of the anteriorcingulate cortex during painful compared to non-painfulstimulation only in IBS patients.An intriguing study examined cerebral activation duringsubliminal visceral stimulation caused by rectal disten-sion (Kern and Shaker 2002). Cerebral activation duringsubliminal distension was generally bilateral in regionsincluding the sensory/motor, parieto-occipital, anteriorcingulate, prefrontal and insular cortices. Distension toliminal (threshold sensation) and supraliminal (abovethreshold sensation) levels activated regions similar tothose activated with subliminal stimulation, but the vol-ume of cortical activity increased with the stimulus in-tensity. In contrast to studies of myocardial ischemia inwhich thalamic activity was detected even after resolu-tion of angina (Rosen et al. 1994; Rosen et al. 1996),thalamic activation was not detected in this study of sub-liminal rectal sensation.Some subsequent studies involving rectal distensionhave reported significant thalamic activation (Verneet al. 2003; Yuan et al. 2003), including greater acti-vation in the thalamus and multiple other regions inIBS patients compared to healthy volunteers during
both rectal and somatic stimulation (Verne et al. 2003).However, thalamic activation has not been detected inall subsequent studies (Berman et al. 2002). As withthe esophageal stimulation studies that failed to de-tect thalamic activation, it seems likely that the lackof significant thalamic activation in some rectal dis-tension studies reflects the limitations of the availabletechnology.
Summary
The thalamus is a major relay and processing station be-tween the peripheral nervous system and higher centersin the central nervous system. It would be anticipatedthat cerebral functional imaging studies using PET orfMRI in humans would detect thalamic activation dur-ing visceral pain. Many such studies have indeed de-tected thalamic activation. The results include thalamicactivation during myocardial ischemia with or withoutangina and after the resolution of angina and thalamicactivation during distal esophageal distension, gastricdistension and rectal distension. However, not all stud-ies of functional cerebral imaging during gastrointesti-nalvisceraldistensionhavedetectedthalamicactivation.These seemingly inconsistent results are probably dueto limitations in the sensitivity of the available technol-ogy.
References1. Aziz Q, Andersson JL, Valind S et al. (1997) Identification of
human brain loci processing esophageal sensation using positronemission tomography. Gastroenterology 113:50–59
2. Aziz Q, Thompson DG, Ng VW et al. (2000) Cortical pro-cessing of human somatic and visceral sensation. J Neurosci20:2657–2663
3. Baciu MV, Bonaz BL, Papillon E et al. (1999) Central process-ing of rectal pain: a functional MR imaging study. AJNR Am JNeuroradiol 20:1920–1924
4. Berman SM, Chang L, Suyenobu B et al. (2002) Condition-specific deactivation of brain regions by 5-HT3 receptor antag-onist Alosetron. Gastroenterology 123:969–977
5. Derbyshire SW (2003) A systematic review of neuroimaging dataduring visceral stimulation. Am J Gastroenterol 98:12–20
6. Kern MK, Shaker R (2002) Cerebral cortical registration of sub-liminal visceral stimulation. Gastroenterology 122:290–298
7. Kern MK, Birn RM, Jaradeh S et al. (1998) Identification andcharacterization of cerebral cortical response to esophagealmucosal acid exposure and distention. Gastroenterology115:1353–1362
8. Ladabaum U, Minoshima S, Hasler W et al. (2000) Gastric disten-tion correlates with activatin of multiple cortical and subcorticalregions. Gastroenterology 120:369–376
9. Mertz H, Morgan V, Tanner G et al. (2000) Regional cerebralactivation in irritable bowel syndrome and control subjectswith painful and nonpainful rectal distention. Gastroenterology118:842–848
10. Rosen SD, Paulesu E, Frith CD et al. (1994) Central nervouspathways mediating angina pectoris. Lancet 344:147–150
11. Rosen SD, Paulesu E, Nihoyannopoulos P et al. (1996) Silentischemia as a central problem: regional brain activation com-pared in silent and painful myocardial ischemia. Ann Intern Med124:939–949
12. Silverman DH, Munakata JA, Ennes H et al. (1997) Regionalcerebral activity in normal and pathological perception of visceralpain. Gastroenterology 112:64–72

2444 Thalamus, Dynamics of Nociception
13. Strigo IA, Duncan GH, Boivin M et al. (2003) Differentiation ofvisceral and cutaneous pain in the human brain. J Neurophysiol89:3294–3303
14. Verne GN, Himes NC, Robinson ME et al. (2003) Central rep-resentation of visceral and cutaneous hypersensitivity in the ir-ritable bowel syndrome. Pain 103:99–110
15. Yuan YZ, Tao RJ, Xu B et al. (2003) Functional brain imaging inirritable bowel syndrome with rectal balloon-distention by usingfMRI. World J Gastroenterol 9:1356–1360
Thalamus, Dynamics of NociceptionVASCO GALHARDO
Institute of Histology and Embryology, Faculty ofMedicine of Porto, University of Porto, Porto, [email protected]
Synonyms
Thalamic plasticity; Thalamic Reorganization; sensiti-zation
Definition
Thedynamicsof the thalamic responses to noxiousstim-uli are the signature of neural mechanisms that makethe responses not rigid and immutable, since they reflectboth the nature of the incoming afferent signal and theinternal state of the neuronal populations that processnociceptive signals.
Characteristics
Somatosensory noxious information is processed by thethalamus in a complex manner that is still little under-stood. Far from being a simple relay station for painfulstimulion theirway from theperiphery to thecortex, tha-lamic neurons integrate and modulate the pain signals ina dynamic way. This implies that the modulation itselfis the result of the instantaneous status of the thalamicneuronalnetwork.By addressing the topicof thedynam-ics of thalamic responses we acknowledge the fact thatthe arriving nociceptive information will not always beprocessed by the thalamic neural networks in exactly thesame manner, since the functional properties of the pro-cessingneuralnetworkswillchangeovertime,reflectingthe history of the incoming signals.The factors that affect the status of the processing neuralnetworks may be extrinsic to the thalamus – meaningthat they arecaused by thequalitiesof thepast and imme-diate afferent information - or intrinsic – meaning thatthey are the result of the status of the thalamus, namelythe occurrence of population � oscillations.This brief essay will review four key aspects of nocicep-tive information processing dynamics within the thala-mus.
Sensitization of the Thalamus
The most basic aspect of thalamic somatosensorydynamics is the change in the response properties of
thalamic neurons induced by states of peripheral persis-tent pain. Several studies have shown that in conditionsof � hyperalgesia or � allodynia the neurons in theventrobasal complex of the lateral thalamus have en-hanced responsiveness, i.e. they have lower activationthresholds for both thermal and mechanical stimuli andlarger peripheral receptive fields and continue to dis-charge spontaneously for long periods after cessation ofthe noxious stimulation (Guilbaud et al. 1990; Shermanet al. 1997). Ultimately, it is difficult to determine towhat extent this functional plasticity corresponds to achange within the thalamic networks or if the enhancedthalamic activity is instead simply a consequence ofthe enhanced somatosensory information generated atspinal levels during chronic pain. One strong piece ofevidence for the occurrence of plasticity at the thala-mic level is that painful sensations are evoked morefrequently by microstimulation of the lateral thalamusin patients with chronic neuropathic pain than in pa-tients with non-painful movement disorders (Davis etal. 1996). Further microstimulation studies suggest thatin neuropathic pain patients, thalamic areas usuallysignaling thermal non-painful discrimination are nowevoking painful sensations when stimulated (Lenz etal. 1998).It is very tempting to assume that the enhanced sensoryactivity corresponds to a progressive change of thala-mic nociceptive-specific neurons into the wide-dynamicrange category, as was implied in some studies (Guil-baud et al. 1990). However, multielectrode recordingslasting several hours in the rat somatosensory thalamus(Brueggemann et al. 2001) showed that immediately af-ter a nerve lesion no rigid pattern of change between celltypes could be found (Fig. 1). Although the results frommultielectrode studies show immediate fluctuations inresponseproperties, they donot showaclearnet increasein nociceptive responses in the affected portion of thelateral thalamus.
Deafferentation-Induced Plasticity
Paradoxically, just as the enhanced somatosensory in-formation leads to a sensitization of thalamic networks,the decrease in somatosensory information arriving atthe thalamus due to peripheral nerve lesions also leadsto higher levels of pain sensitivity. This is probably dueto the fact that both the functional loss of afferents byperipheral lesions and the functional gain of afferentsby peripheral � hyperalgesia lead to similar net effects;some regions of the thalamus are suddenly more activethan their neighbors and this spatial imbalance causesa regional peak of activity that leads to somatosensoryhypersensitivity. � Deafferentation studies also reveala crosstalk between somatosensory modalities in whichboth the disruption of thick myelinated non-nociceptivefibers leads to altered pain perception and the disruptionof unmyelinated high-threshold � C Fiber leads to al-tered tactile perception. A recent multielectrode study

T
Thalamus, Dynamics of Nociception 2445
Thalamus, Dynamics of Nociception, Figure 1 Changes in classifica-tion of lateral thalamic neurons at the beginning (Before, before sciaticnerve ligation) and at the end of the experiments (After, after nerve liga-tion). Horizontal lines indicate no change; diagonal lines indicate changes inneuronal class. Abbreviations: Ø, unresponsive neurons; LT, Low-thresholdneurons; WDR, wide-dynamic range neurons; NS, nociceptive-specific neu-rons (Figure taken from Brueggemann et al. 2001).
in the rat lateral thalamus (Katz et al. 1999) showed thatthe silencing of � c-fibers after perioral capsaicin injec-tions changed the pattern of sensitivity to tactile deflec-tion of the whiskers, resulting in “unmasking” of newtactile responses.
Spatiotemporal Dynamics in the Thalamus
Several studies have shown novel properties in the tha-lamic integration of nociceptive signals, in both spaceand time domains. Finding spatiotemporal patterns ofcoherence in the activity of neuronal populations in thethalamus during the processing of pain will help unravelthe intrinsic organization of the processing units withinthe lateral thalamus. The use of multielectrode arrayswith precise spatial arrangements showed that the func-tional connectivity between pairs of neurons dependson the distance separating them; neighboring nocicep-tive neurons tend to be positively correlated, while mostnegative correlations in evoked spike timings occur be-tween neurons separated by more than 50 μm (Apkar-ian et al. 2000). Interestingly, the spatial maps of evokedspike coherence between pairs of neurons were differ-ent for noxious and non-noxious stimulations, with tac-tile stimuli inducing almost no inhibitory spatial effects.The results of that study also suggest that in the primatethalamus, nociceptive and non-nociceptive neurons areclustered separately in a lattice manner. This spatial ar-rangement of the lateral thalamus would have profoundimplications for the spatiotemporal dynamics of thala-mic function, since nociceptive clusters would receivespinothalamic afferents while non-nociceptive clusterswould receive mainly dorsal column afferents and both
have distinct velocity and synchrony characteristics thatwould result in complex spatiotemporal patterns, diffi-cult to interpret and predict.By the same token, it has been conclusively shown thatfor fine tactile discrimination also, the spatiotemporalmaps are complex to determine and reorganize after pe-ripheral � deafferentation and that distributed coherentactivity in the thalamus plays a key role in the encodingof somatosensory signals (Nicolelis 1997).
Thalamic Oscillations and Pain Processing
The thalamus isknown to have two modesoffiring: tonicmode, in which neurons fire single action potentialsand bursting mode, in which neurons fire in rhythmicrapid bursts of several action potentials. Traditionally,thalamic bursts were associated with non-aware states,while tonic mode was associated with alert states, al-though this simple scheme has been questioned in recentstudies (Nicolelis and Fanselow 2002). High-frequencyspike bursts are one of the most characteristic featuresobserved in the thalamus following � deafferentationby lesion of peripheral nerves, dorsal roots or the spinalcord (Albe-Fessard et al. 1985; Guilbaud et al. 1990).In the thalamus, spike bursts have been described withvery specific membrane physiology and have beenshown to function normally to synchronize activity ofneurons both within and between thalamic nuclei aswell as between the thalamus and cortex. In human tha-lamic recordings performed during surgery (Lenz et al.1989), a characteristic series of spontaneous short highfrequency spike trains (3–8 action potentials occurringat 60–160 Hz) or spike bursts was found in long-termpara- and tetraplegics. These bursts repeat themselvesin low-frequency � oscillations in pain patients and theclinical condition has been termed � thalamocorticaldysrhythmia (Llinás et al. 1999).Significance has been assigned to changes in the tem-poral patterns of activity in thalamic neurons followingdeafferentation because spike bursts in pain patients areespecially concentrated in regions of the lateral thala-mus representing the painful part of the body. In Fig. 2wepresentan exampleof such aburst activity.Thefigureis taken from Weng et al. (2000) where the authors re-port that thalamic wide-dynamic range neurons (but notthe low-threshold neurons) of monkeys partially deaf-ferentedbyspinalcord lesionsshowedenhancedactivitycompared with the same types of cells in thalamus withintact innervation; both the spontaneous and evoked dis-charges of the cells were altered, so that there was an in-creased incidenceof spike-bursts in cellsofdeafferentedthalamus.Recently, the importance of this abnormal thalamicfiring for the genesis of chronic central pain has beenchallenged by the finding of no differences betweenthe number of oscillating neurons in the thalamus ofallodynic versus non-allodynic rats with spinal cordinjury (Gerke et al. 2003), suggesting that � thalamic

2446 Thalamus Lesion
Thalamus, Dynamics ofNociception, Figure 2 Brushstimuli evoke oscillatory activityin the lateral thalamus of animalswith complete sectioning of theanterolateral spinal quadrant (Figuretaken from Weng et al. 2000).
dysrhythmia is linked to cord injury but not to the pres-ence of � allodynia. However this study was performedin anaesthetized animals, raising the question of itsapplicability to awake thalamic processing. By thesame token, Radhakrishnan et al. (1999) showed thatthe incidence of thalamic bursting was similar betweenpainful and non-painful neurological injuries. Hence,the role of the internal oscillatory states of the thalamusin pain processing is still to be clarified.
References1. Albe-Fessard D, Berkley KJ, Kruger L et al. (1985) Diencephalic
mechanisms of pain sensation. Brain Res 356:217–2962. Apkarian AV, Shi T, Brueggemann J et al. (2000) Segregation of
nociceptive and non-nociceptive networks in the squirrel monkeysomatosensory thalamus. J Neurophysiol 84:484–494
3. Brueggemann J, Galhardo V, Apkarian AV (2001) Immediatereorganization of the rat somatosensory thalamus following pe-ripheral partial nerve ligation. J Pain 2:220–228
4. Davis KD, Kiss ZHT, Tasker RR et al. (1996) Thalamicstimulation-evoked sensations in chronic pain patients andnon-pain (movement disorder) patients. J Neurophysiol75:1026–1037
5. Gerke M, Duggan A, Xu L et al. (2003) Thalamic neuronal activ-ity in rats with mechanical allodynia following contusive spinalcord injury. Neuroscience 117:715–722
6. Guilbaud G, Benoist JM, Jazat F et al. (1990) Neuronal re-sponsiveness in the ventrobasal thalamic complex of rats withan experimental peripheral mononeuropathy. J Neurophysiol64:1537–1554
7. Katz DB, Simon SA, Moody A et al. (1999) Simultaneous reorga-nization in thalamocortical ensembles evolves over several hoursafter perioral capsaicin injections. J.Neurophysiol 82:963–977
8. Lenz FA, Kwan HC, Dostrovsky JO et al. (1989) Characteris-tics of the bursting pattern of action potentials that occurs in thethalamus of patients with central pain. Brain Res 496:357–360
9. Lenz FA, Gracely RH, Baker FH et al. (1998) Reorganizationof sensory modalities evoked by microstimulation in region ofthe thalamic principal sensory nucleus in patients with pain dueto nervous system injury. J Comp Neurol 399:125–138
10. Llinas RR, Ribary U, Jeanmonod D et al. (1999) Thalamocorti-cal dysrhythmia: A neurological and neuropsychiatric syndromecharacterized by magnetoencephalography. Proc Natl Acad SciUSA 96:15222–15227
11. Nicolelis MAL (1997) Dynamic and distributed somatosensoryrepresentations as the substrate for cortical and subcortical plas-ticity. Semin Neurosci 9:24–33
12. Nicolelis MAL, Fanselow EE (2002) Thalamocortical optimiza-tion of tactile processing according to behavioral state. Nat Neu-rosci 5:517–523
13. Radhakrishnan V, Tsoukatos J, Davis KD et al. (1999) A com-parison of the burst activity of lateral thalamic neurons in chronicpain and non-pain patients. Pain 80:567–575
14. Sherman SE, Luo L, Dostrovsky JO (1997) Altered receptivefields and sensory modalities of rat VPL thalamic neuronsduring spinal strychnine-induced allodynia. J Neurophysiol78:2296–2308
15. Weng HR, Lee JI, Lenz FA et al. (2000) Functional plasticityin primate somatosensory thalamus following chronic lesion ofthe ventral lateral spinal cord. Neuroscience 101:393–401
Thalamus Lesion
� Lateral Thalamic Lesions, Pain Behavior in Animals
Thalamus, Metabotropic GlutamateReceptors
� Metabotropic Glutamate Receptors in the Thalamus
Thalamus, Nociceptive Cells in VPI, Catand Rat
CHRISTIANE VAHLE-HINZ
Institute of Neurophysiology and Pathophysiology,University Hospital Hamburg-Eppendorf, Hamburg,[email protected]
Synonyms
High-threshold neurons; nociceptive-specific neurons(NS); wide-dynamic range (WDR) neurons; multire-ceptive neurons (MR)

T
Thalamus, Nociceptive Cells in VPI, Cat and Rat 2447
Definition
Neurons of the lateral thalamus encoding stimulus qual-ity, duration, intensity and location on the body and thussubserving the � sensory-discriminative component ofpain.The neurons respond with an increase or decreaseof discharge activity to noxious stimuli exclusively(� nociceptive-specific neurons, NS) or with a higherdischarge to noxious mechanical than to innocuousmechanical stimuli (� wide-dynamic range (WDR)neurons) or to both noxious and innocuous mechanicalas well as thermal stimuli (multireceptive neurons).As defined in “Thalamic Nuclei Involved in Pain, catand rat” the region termed � ventral posterior inferiornucleus (VPI) in monkeys may correspond to the smallcelled region of the ventral periphery of the ventral pos-terior medial nucleus (VPMvp) in cats, the extensionsof which surround the ventral, lateral and dorsal periph-ery of the � ventral posterior complex (VP). In rats,nociceptive neurons are also found within and aroundthe VP, but most are concentrated in the adjoining
Thalamus, Nociceptive Cells in VPI, Cat and Rat, Figure 1 Response characteristics of nociceptive neurons of the cat’s VPp (a, b, c) and the rat’sPOm (d). (a) Spike record of a heat-evoked response. The elevated activity representing stimulus duration is followed by a lower afterdischarge for about30 s before return to prestimulus activity. (b) Multireceptive neuron responding to noxious pressure and heat stimuli in different parts of the receptivefield. (c) Wide-dynamic range neuron with graded responses to innocuous pressure and two intensities of noxious pressure. (d) The response elicitedby a radiant heat stimulus is abolished by an increase of isoflurane concentration from 0.9% to 1.2%. Spike histograms, bin width 1 s; the bar beloweach histogram represents the duration of the stimulus (modified from Vahle-Hinz et al. 1987 (a); 2002 (d); Kniffki and Vahle-Hinz 1987 (c)).
� medial part of the posterior complex (POm). Sincethese have similar � receptive field (RF) and responseproperties, it appears that these regions in the lateralthalamus of rats and cats are involved in processingand transmission of nociceptive signals in a similarway.
Characteristics
Nociceptive neurons in the VPp have a less precise so-matotopic organization than neurons of the VP proper,largely due to numerous neurons with larger and com-plex RFs consisting of discontinuous areas on the bodysurface. However, a coarse mediolateral sequence ofhead, forelimb and hind limb � RFs is present, runningparallel to that of the VP proper, thus forming a secondrepresentation of the body. Due to a lower ongoingactivity and a sparser packing of neurons, even lowimpedance electrodes hardly pick up any backgroundactivity in contrast to the noisy “hash” characteristic ofthe VP (Kniffki and Vahle-Hinz 1987; Vahle-Hinz etal. 1987).

2448 Thalamus, Nociceptive Cells in VPI, Cat and Rat
Thalamus, Nociceptive Cells in VPI, Cat and Rat, Figure 2 Excitatory responses of a PO neuron (a) and inhibitory responses of a VPp neuron (b) tonoxious distension of the bladder. Peristimulus-time histograms (bin width 1 s) and intravesical pressure (lower traces), with baseline and peak valuesindicated, are shown on the left. The low-threshold cutaneous RFs of the respective neuron (b, black area) or of the background activity (a, stippledarea) are delineated in the figurines. The locations of the recording sites are shown in the line drawings made from the histological sections. (Modifiedfrom Brüggemann et al. 1993).
In both cats and rats, the neurons respond to gradednoxious mechanical and/or heat stimuli (for reviewsee Willis 1997). Responses to noxious heat stimulicharacteristically occur after long latencies (severalseconds), which may result from receptor activationtime and transmission by C fibers. The discharge in-crease in many neurons encodes stimulus duration,often followed by a lower but still increased afterdis-charge for several tens of seconds (Fig. 1a, b, d). Athreshold temperature and a stimulus-response func-tion resembling psychophysical heat pain in humansare found. All types of nociceptive neurons are present,nociceptive-specific (Fig. 1a, d), multireceptive (Fig.1b) and WDR neurons (Fig. 1c). NS and WDR/MRneurons occur in about equal proportions in the cat,while the majority in the rat is of the NS-type. Thalamicnociceptive neurons are sensitive to anesthetics; theresponse to a noxious stimulus may be abolished bya slight increase of anesthetic dose without a markedsuppression of the neuron’s ongoing discharge activity(Fig. 1d). Different classes of anesthetic agents appearto block nociceptive signal transmission at differentsubthalamic/thalamic sites within the ascending path-ways. The low-threshold mechanoreceptive responsesin contrast are more robust (Vahle-Hinz and Detsch2002; Vahle-Hinz et al. 2002).As with noxious somatic stimuli and also with visceralstimulation, both excitation and inhibition of thalamicneuronal discharges can be elicited (Fig. 2). In contrastto the spinal cord level, where the majority of viscero-ceptive neurons have convergent somatic inputs of theWDR type, visceroceptive-specific neurons are found in
the thalamus of both rats and cats. Some of the additionalsomatic RFs of viscero-somatic convergent neurons inthe rat are nociceptive (Berkley et al. 1993), while inthe cat most are of the low-threshold type. The major-ity of these somatic low-threshold RFs are located in ar-eas including dermatomes to which pain is referred fromthe respective visceral organs (Brüggemann et al. 1993;Horn et al. 1999; Vahle-Hinz et al. 1995).The neuronal responses in the lateral thalamus indicatethat distinct response properties are segregated (e.g. so-matic and visceral nociceptive-specific), thus increasingthe diversity of response characteristics contained inthe population of nociceptive neurons. Excitatory andinhibitory interactions between inputs from differentsomatic sources (mechanoreceptive/nociceptive) orvisceral organs may help to focus the activity and hencethe sensation to a certain stimulus or organ.
References1. Brüggemann J, Vahle-Hinz C, Kniffki K-D (1993) Representa-
tion of the urinary bladder in the lateral thalamus of the cat. JNeurophysiol 70:482–491
2. Berkley KJ, Guilbaud G, Benoist J-M et al. (1993) Responses ofneurons in and near the thalamic ventrobasal complex of the ratto stimulation of uterus, vagina, colon, and skin. J Neurophysiol69:557–568
3. Horn AC, Vahle-Hinz C, Brüggemann J et al. (1999) Responses ofneurons in the lateral thalamus of the cat to stimulation of urinarybladder, colon, esophagus, and skin. Brain Res 851:164–174
4. Kniffki K-D, Vahle-Hinz C (1987) The periphery of the cat’sventroposteromedial nucleus (VPMp ): Nociceptive neurones. In:Besson J-M, Guilbaud G, Peschanski M (eds) Thalamus and Pain.Elsevier, Amsterdam, pp 245–257
5. Vahle-Hinz C, Brüggemann J, Kniffki K-D (1995) Thalamic pro-cessing of visceral pain. In: Bromm B, Desmedt J (eds) Pain and

T
Thalamus, Nociceptive Inputs in the Rat (Spinal) 2449
the Brain. From Nociception to Cognition. Advances in Pain Re-serach and Therapy, vol 22. Raven Press, New York, pp 125–141
6. Vahle-Hinz C, Detsch O (2002) What can in vivo electrophysi-ology in animal models tell us about mechanisms of anesthesia?Br J Anaesth 89:123–142
7. Vahle-Hinz C, Freund I, Kniffki K-D (1987) Nociceptive neuronsin the ventral periphery of the cat thalamic ventroposteromedialnucleus. In: Schmidt RF, Schaible H-G, Vahle-Hinz C (eds) FineAfferent Nerve Fibers and Pain. VCH Verlagsgesellschaft, Wein-heim, pp 440–450
8. Vahle-Hinz C, Reeker W, Detsch O et al (2002) Antinociceptiveeffects of anesthetics in vivo : Neuronal responses and cellularmechanisms. In: Urban BW, Barann M (eds) Molecular and Ba-sic Mechanisms of Anesthesia. Pabst Sci Publ, Lengerich, pp516–524
9. Willis WD (1997) Nociceptive functions of thalamic neurons.In: Steriade M, Jones EG, McCormick DA (eds). Thalamus, Vol.II, Experimental and Clinical Aspects. Elsevier, Amsterdam, pp373–424
Thalamus, Nociceptive Inputs in the Rat(Spinal)
JEAN-FRANÇOIS BERNARD
Institut National de la Santé et de la RechercheMédicale, INSERM U-677, Paris, [email protected]
Definition
Layers of the Spinal Cord
The gray matter of the spinal cord is divided, from dorsalto ventral, into 10 laminae on the basis of cytoarchitec-tonic criteria. The dorsal horn includes the laminae I toVI; the lamina VII is an intermediate area; the ventralhorn includes the laminae VIII and IX (motoneurons);the region around the central canal corresponds to thelamina X.
Thalamus
The caudal portion of the thalamus is an important braincenter for somatosensory and nociceptive processing.This caudal region is often divided into the lateral andthe medial thalamus. 1) The lateral thalamus is primar-ily a relay that includes chiefly, for somatosensory func-tions, the ventral posterolateral (VPL), the ventral pos-teromedial (VPM), the posterior group (Po) and the tri-angular posterior group (PoT) thalamic nuclei. 2) Themedial thalamus might be regarded as a more integra-tive region that includes intralaminar [chiefly the cen-tral lateral (CL), paracentral (PC), central medial (CM)and parafascicular (Pf) nuclei], paralaminar [notably theventral medial (VM) nucleus] and median thalamic nu-clei.
Characteristics
The Spinal Relay
Noxious messages are conveyed from the periphery(trunk and limbs) to the thalamus via a primary relay,the dorsal horn of the spinal cord. Two individualized
regions of the dorsal horn, the superficial and the deeplaminae, have key roles in the processing of nociceptivemessages.The superficial laminae (I and II) have a major role in no-ciceptive processing because this region is the main re-cipient for peripheral nociceptive inputs conveyed by Aδ
and C fibers. Whereas both the laminae I and II receivenociceptive inputs, only the lamina I neurons project tosupraspinal centers. These neurons are the main outputof this superficial region. The lamina I neurons are pri-marily nociceptive, amajority of them being nociceptivespecific (Christensen and Perl1970). In addition, a lowerproportion of lamina I neurons can encode specificallyinnocuous thermal stimuli (Light et al. 1993).The second area of the dorsal horn, which processesnociceptive messages, includes the deep laminae V, VIand the adjacent portion of the lamina VII. The involve-ment of deep laminae in nociceptive processing waschiefly demonstrated by electrophysiology; this regioncontains numerous wide dynamic range neurons thathave a great ability to encode noxious stimuli but froma clearly innocuous range (Besson and Chaouch 1987).The anatomical link of this region with peripheral Aδ
and C nociceptive fibers is less clear; deep laminaeneurons receive some collateral projections from Aδ
and C fibers, but the main nociceptive input might beconveyed indirectly via superficial laminae.The caudal thalamus is a primary brain center for painprocessing. Nociceptive information is conveyed tothe thalamus from nociceptive neurons in lamina Iand deep laminae of the spinal cord 1) directly via thespino-thalamic tract and 2) indirectly via relay nucleiin the brainstem.
Spinal Nociceptive Inputs to the Thalamus
Retrograde tracing studies show that neurons projectingto the thalamus are located in lamina I and in deep lam-inae of the spinal cord (Burstein et al. 1990). Thalamicprojecting superficial spinal neurons are clearly con-centrated in lamina I of the dorsal horn, only very feware located in lamina II. Lamina I neurons are evenlydistributed along the spinal cord, with higher concentra-tions at the levels of cervical and lumbar enlargements.On the other hand, thalamic projecting deep laminaeneurons are scattered in deep laminae of the spinal cord(III to VIII and X) although they are more numerousaround the laminae IV – VI. These thalamic projectingdeep laminae neurons are not evenly distributed in thespinal cord, since almost half of them are concentratedin the two first spinal segments (C1, 2). In the cervicalenlargement (C5 – 8), which relays nociceptive mes-sages from the forelimb, one spino-thalamic neuron inlamina I and one in the deep lamina were counted onaverage per 50 μm section (Burstein et al. 1990).Anterograde tracing studies, using a high-resolutiontracer such as the Phaseolus vulgaris leucoagglutinin(PHA-L), allowed the demonstration of separate pro-

2450 Thalamus, Nociceptive Inputs in the Rat (Spinal)
Thalamus, Nociceptive Inputs in the Rat (Spinal), Figure 1 Summary diagram of spinal projections to the thalamus. (a1) Lamina I (hatching) projectingarea in the cervical enlargement of the spinal cord. (b1) Projection in the PoT (hatching) from lamina I. (c1) Projection to VPL, Po and VPPC thalamicnuclei (hatching) from lamina I. (a2) Deep laminae (black points) and SL nucleus (gray) projecting area in the cervical enlargement. (b2) Projection toPoT thalamic nucleus (points) from the deep laminae. (c2) Projection to CL (points) and MD (gray) thalamic nuclei from deep laminae and SL nucleus,respectively. Scale bars = 1 mm. Abbreviations: I-X, laminae I-X of the spinal cord; APT, anterior pretectal nucleus; CL, central lateral thalamic nucleus;CM, central medial thalamic nucleus; DMH, dorsomedial hypothalamic nucleus; eml, external medullary lamina; f, fornix; LHp, lateral hypothalamusposterior; MD, mediodorsal nucleus; MG, medial geniculate nucleus; ml, medial lemniscus; mt, mammillothalamic tract; PAG, periaqueductal graymatter; PC, paracentral thalamic nucleus; PH, posterior hypothalamus; PIL, posterior intralaminar thalamic nucleus; Po, posterior thalamic group; PoT,posterior thalamic group, triangular part; Re, reuniens nucleus; SL, spinal lateral nucleus; SN, substantia nigra; VM, ventromedial thalamic nucleus; VMH,ventromedial hypothalamic nucleus; VPL, ventral posterolateral thalamic nucleus; VPM, ventral posteromedial thalamic nucleus; VPPC, ventral posteriorparvocellular thalamic nucleus; ZI, zona incerta.
jections from lamina I and deeplaminae of the spinalcord upon the thalamus (Gauriau and Bernard 2003).The projections from lamina I are markedly differentfrom those of deep laminae (Fig. 1).Lamina I neurons project primarily to restricted portionsof the VPL and rostral Po (Fig. 1a1–c1). In these areas,
axonal endingshave large varicosities (Fig. 2). Othersubstantial projections with small varicosities were ob-served in the PoT and the ventral posterior parvocellularnucleus(VPPC).Onlyamoderatenumberofprojectionsare observed in a few additional thalamic targets such asthe periventricular, the subparafascicular, the reuniens

T
Thalamus, Nociceptive Inputs in the Rat (Spinal) 2451
Thalamus, Nociceptive Inputs in the Rat (Spinal),Figure 2 Photomicrographs of the projection from lamina I tothe thalamus. (a) PHA-L injection site in laminae I/II of the cervicalenlargement. (b) Extensive terminal labeling with large varicosities in theVPL thalamic nucleus resulting from the injection in (a). Scale bars =500 μm in A, 100 μm in (b). Abbreviations: I-X, laminae I-X of the spinalcord; VPL, ventral posterolateral thalamic nucleus.
and the mediodorsal nuclei; no projection is observedto the submedius nucleus. Lamina I projections to thethalamus are almost exclusively contralateral. Becausethe VPL/Po system is also known to be the thalamicrelay conveying somatotopically tactile messages fromgracile and cuneate nuclei to somatosensory SI and SIIcortex, it appears likely that the lamina I – VPL/Po sys-tem is devoted to somatosensory discriminative aspectsof nociception. The role of the lamina I – PoT/VPPCsystem is lessclear, it could participate to the recognitionof the “painful” nature of nociceptive stimuli.Deep laminae neurons project chiefly to the central lat-eral intralaminar and the PoT thalamic nuclei on bothsides (Fig.1 a2–c2). Very few projections are observedto other thalamic nuclei. In fact, it appears that deep lam-inae neurons project as much to the thalamus as to ex-trathalamic targets, such as the substantia innominata,the globus pallidus, the posterior and lateral hypothala-mus and the central amygdaloid nucleus. These recentfindings clearly question the projections of deep lami-nae to the VPL, suggesting strongly that the main noci-ceptive inputs to the VPL originate primarily from thelamina I neurons, in the rat.
A last notable projection is from the spinal lateral neu-rons to the caudal portion of the mediodorsal nucleus(Fig.1 a2–c2).
Brainstem Nociceptive Inputs to the Thalamus
Thespino-thalamic tract isoften considered tobe thepri-mary pathway conveyingnociceptivemessages from thespinal cord to the thalamus. In fact, the number of spino-thalamic neurons (9500 is the higher estimation in therat;Bursteinetal.1990) issubstantialbut representsonlya very small proportion of dorsal horn neurons. Further-more,anterogradestudies inprimate(Mehleretal.1960)as well as in the rat (Gauriau and Bernard 2003) indicatethat spinal projections are clearly more extensive uponthe brainstem than to the thalamus. These data stronglysuggest that the brainstem should have an important rolein conveying nociceptive input from the spinal cord tothe thalamus and the other brain centers.The gigantocellular reticular (Gi) nucleus, located in thecenterof themedulla,was thefirst candidate to fulfill thisrole (Casey 1971; Bowsher 1976). This nucleus receivesan extensive projection from deep laminae of the spinalcord, it contains numerous nociceptive neurons and itsstimulation produces aversive reactions. However, theportion of the Gi that receives the densest spinal pro-jection projects primarily to the locus coeruleus and thespinal cord and only weakly to the thalamus.The subnucleus reticularis dorsalis (SRD), a very caudalreticular area of the medulla, located just ventral to thecuneate nucleus, now appears to be the best candidate toconvey nociceptive messages from the spinal cord to thethalamus. Indeed, the SRD receives an extensive projec-tion from deep laminae of the spinal cord (Raboisson etal. 1996). Electrophysiological studies demonstrate theinvolvement of this reticular region in nociceptive pro-cessing. Indeed, most of the SRD neurons are stronglyexcitedbynoxiousstimuli fromalowspontaneousactiv-ity and do not respond to multisensory (visual and audi-tory) stimuli. SRD neurons encode the intensity of ther-mal, mechanical and visceral noxious stimuli. They re-spond exclusively to the activation of peripheral Aδ- orAδ- and C-fibers. Such responses are depressed by in-travenous morphine in a dose-dependent and naloxone-reversible fashion (Villanuevaetal. 1996).The receptivefields of SRD neurons are very large; they often includethe whole body. The main thalamic targets of the SRDare the lateralportionsof theVMnucleus, (Fig.3) and, toa lesser extent, of the Pf nucleus (Villanueva et al. 1998).The internal lateral parabrachial (PBil) nucleus, locatedin a dorsal position at the ponto-mesencephalic junction,isanothercandidateforconveyingnociceptivemessagesfrom the spinal cord to the thalamus. The PBil shouldnot be mixed up with the lateral parabrachial area, whichreceives an extensive projection from lamina I and doesnot project substantially to the thalamus. In fact, the PBilspecifically receives a dense projection from deep lami-nae of the spinal cord (especially from the reticular por-

2452 Thalamus, Nociceptive Inputs in the Rat (Spinal)
Thalamus, NociceptiveInputs in the Rat (Spinal),Figure 3 Photomicrographs of theprojection from the SRD to thethalamus. (a) PHA-L injection site inthe SRD. (b) High density of labeledterminals in the lateral portion ofthe VM. (c) Higher magnification ofthe labeled terminals in the regiondelineated in (b). Note the numerousterminals with small varicosities.Scale bars = 500 μm in (a), (b), 100μm in (c). Abbreviations: Cu, cuneatenucleus; SRD, subnucleus reticularisdorsalis; VM, ventromedial thalamicnucleus; VPL, ventral posterolateralthalamic nucleus; VPM, ventralposteromedial thalamic nucleus.
Thalamus, NociceptiveInputs in the Rat (Spinal),Figure 4 Photomicrographs ofthe projection from the PBil to thethalamus. Note the high density oflabeled terminals concentrated in thePC (black area on both sides) with alower density of labeling in the CM(a) resulting from the PHA-L injectioncovering the PBil (b). Scale bars =500 μm. Abbreviations: 3V, thirdventricle; bc, brachium conjunctivum;CL, central lateral thalamic nucleus;CM, central medial thalamic nucleus;iml, internal medullary lamina; MD,mediodorsal thalamic nucleus; OPC,oval paracentral thalamic nucleus;PBil, internal lateral parabrachialnucleus; PC, paracentral thalamicnucleus; Rh, rhomboid thalamicnucleus.
tion of laminae IV and V) (Bernard et al. 1995). ThePBil neurons project primarily to the PC thalamic nu-cleus (Fig. 4) and, to a lesser extent, to the CM and the Pfthalamic nuclei (Bester et al. 1999). Electrophysiologi-cal studiesdemonstrate the involvementof thalamicpro-jecting PBil neurons in nociceptive processing. Indeed,most of them respond to thermal and noxious stimuli,with a maximum response in the mid-nociceptive scale(48˚C and 16 N/cm2). The PBil neurons exhibit strong“wind up” and long lasting after-discharge in responseto noxious stimuli (Bourgeais et al. 2001).With regard to the deep laminae system, the brainstemclearly has an important complementary role to the
spino-thalamic tract. Indeed, deep laminae neuronssend nociceptive messages to the medial thalamus di-rectly via the spino-thalamic tract, as well as indirectlyvia SRD and PBil neurons. Thus this system could dealwith alertness and emotional and motor aspects of painthrough a general arousal of the prefrontal and frontal(motor) cortices (the cortical targets of the medialthalamus).
Synthesis
Nociceptive inputs to the thalamus can be classified inthe two different systems summarized in Fig. 5 as fol-lows:

T
Thalamus, Nociceptive Inputs in the Rat (Spinal) 2453
Thalamus, Nociceptive Inputs in the Rat (Spinal), Figure 5 Schematicrepresentation of the main nociceptive inputs to the thalamus, in the rat.Red: Main inputs to the lateral thalamus from the lamina I of the spinalcord. Blue: Main inputs to the medial thalamus originating from the deeplaminae of the spinal cord, directly via the spino-thalamic tract, andindirectly via the SRD and the PBil nuclei. Abbreviations: I-X, laminae I-X ofthe spinal cord; APT, anterior pretectal nucleus; ar, acoustic radiation; bc,brachium conjunctivum; BL, basolateral amygdaloid nucleus; Ce, centralamygdaloid nucleus; CL, central lateral thalamic nucleus; CM, centralmedial thalamic nucleus; CPu, caudate putamen (striatum); Cu, cuneatenucleus; DMH, dorsomedial hypothalamic nucleus; Eth, ethmoid thalamicnucleus; f, fornix; Gr, gracilis nucleus; ic, internal capsule; IO, inferior olive;La, lateral amygdaloid nucleus; LC, locus coeruleus; LRt, lateral reticularnucleus; Me, medial amygdaloid nucleus; MG, medial geniculate nucleus;ml, medial lemniscus; mt, mammillothalamic tract; opt, optic tract; PBil,internal lateral parabrachial nucleus; PBl, lateral parabrachial nucleus;PBm, medial parabrachial nucleus; PC, paracentral thalamic nucleus; PIL,posterior intralaminar thalamic nucleus; Po, posterior thalamic group; PoT,posterior thalamic group, triangular part; pyx, pyramidal decussation; SG,suprageniculate thalamic nucleus; Sol, solitary tract nucleus; Sp5C, spinaltrigeminal nucleus caudal part; SPF, subparafascicular nucleus; SRD,subnucleus reticularis dorsalis; VL, ventrolateral thalamic nucleus; VM,ventromedial thalamic nucleus; VMH, ventromedial hypothalamic nucleus;VPL, ventral posterolateral thalamic nucleus; VPM, ventral posteromedialthalamic nucleus.
1. The lamina I – (lateral) thalamic system, whichcould be chiefly responsible for sensory discrimina-tion of nociceptive stimuli via thalamic projectionsto somatosensory cortices (SI, SII, Insular).
2. The deep laminae – (medial) thalamic system, whichincludes two subsystems: a direct deep laminae – me-dial thalamic pathway and an indirect deep laminae– SRD/PBil – medial thalamic pathway. This systemcouldbeinvolvedinmotorandalertness/arousalemo-tional features of pain via the medial thalamic projec-tions to the frontal motor and the prefrontal medial(cingulate) cortices.
The spinal ascending axons of all these systems arelocated around the same region of the spinal lat-
eral/ventrolateral quadrant in the rat. Thus, it appearsclear that the strikingly acute effectiveness of ventrolat-eral cordotomy is due to the interruption of both a directspino-thalamic tract and a strong spino-brainstem-thalamic pathway.An additional nociceptive pathway to the thalamus (notillustrated)wasproposed by thegroup ofWillis.Thevis-ceral (colorectal) nociceptive information, after a relayin laminae X and VII of the lumbo-sacral spinal cord,would be conveyed via the medial portion of the dorsalcolumn and the gracile nucleus to the VPL thalamic nu-cleus.Thekey point in thedemonstrationof thispathwayis the strong effect of a medial commissurotomy (in themiddle of dorsal column fasciculus) upon the responseof VPL neurons to visceral noxious stimuli and visceral

2454 Thalamus, Nociceptive Neurotransmission
pain (Willis et al. 1999). However, the existence of sucha pathway remains difficult to reconcile with previousclinical and experimental data.
References1. Bernard JF, Dallel R, Raboisson P et al. (1995) Organization of
the efferent projections from the spinal cervical enlargement tothe parabrachial area and periaqueductal gray: a PHA-L studyin the rat. J Comp Neurol 353:480–505
2. Besson JM, Chaouch A (1987) Peripheral and spinal mechanismsof nociception. Physiol Rev 67:67–186
3. Bester H, Bourgeais L, Villanueva L et al. (1999) Differentialprojections to the intralaminar and gustatory thalamus from theparabrachial area: A PHA-L study in the rat. J Comp Neurol405:421–449
4. Bourgeais L, Monconduit L, Villanueva L et al. (2001)Parabrachial internal lateral neurons convey nociceptive mes-sages from the deep laminas of the dorsal horn to the intralaminarthalamus. J Neurosci 21:2159–2165
5. Bowsher D (1976) Role of the reticular formation in responsesto noxious stimulation. Pain 2:361–378
6. Burstein R, Dado RJ, Giesler GJ Jr (1990) The cells of origin ofthe spinothalamic tract of the rat: a quantitative reexamination.Brain Res 511:329–337
7. Casey KL (1971) Somatosensory responses of bulboreticularunits in awake cat: relation to escape-producing stimuli. Science173:77–80
8. Christensen BN, Perl ER (1970) Spinal neurons specifically ex-cited by noxious or thermal stimuli: marginal zone of the dorsalhorn. J Neurophysiol 33:293–307
9. Gauriau C, Bernard JF (2003) A comparative reappraisal of pro-jections from the superficial laminae of the dorsal horn in therat: the forebrain. J Comp Neurol 468:24–56
10. Light AR, Sedivec MJ, Casale EJ et al. (1993) Physiologicaland morphological characteristics of spinal neurons projectingto theparabrachial region of the cat. SomatosensMot Res10:309–325
11. Mehler WR, Feferman ME, Nauta W (1960) Ascending axon de-generation following anterolateral cordotomy. An experimentalstudy in the monkey. Brain 83:718–751
12. Raboisson P, Dallel R, Bernard JF et al. (1996) Organizationof efferent projections from the spinal cervical enlargement tothe medullary subnucleus reticularis dorsalis and the adjacentcuneate nucleus: a PHA-L study in the rat. J Comp Neurol367:503–517
13. Villanueva L, Bouhassira D, Le Bars D (1996) The medullarysubnucleus reticularis dorsalis (SRD) as a key link in boththe transmission and modulation of pain signals. Pain 67:231–240
14. Villanueva L, Desbois C, Le Bars D et al. (1998) Organiza-tion of diencephalic projections from the medullary subnucleusreticularis dorsalis and the adjacent cuneate nucleus: A retro-grade and anterograde tracer study in the rat. J Comp Neurol390:133–160
15. Willis WD, Al-Chaer ED, Quast MJ et al. 1999 A visceral painpathway in the dorsal column of the spinal cord. Proc Natl AcadSci USA 96:7675–7679
Thalamus, NociceptiveNeurotransmission
� Nociceptive Neurotransmission in the Thalamus
Thalamus, Receptive Fields, ProjectedFields, Human
NIRIT WEISS, SHINJI OHARA, STAN ANDERSON,FRED A. LENZ
Departments of Neurosurgery, Johns HopkinsUniversity, Baltimore, MD, [email protected]
Synonyms
Thalamic Physiology Changes Occurring in Patientswith Chronic Pain
Definition
Physiologic changes in the thalamus seen in patientswith chronic pain syndromes. These changes are re-flected in alterations in the properties of neurons andassemblies of neurons in patients who suffer fromchronic pain.
Characteristics
Studies of the � plasticity of the somatosensory systemin non-human primates have focused on maps of corticalfunction determined by examining neuronal � receptivefields (RFs) (Kaas 1991). Studies in humans can exploreboth maps determined from RFs (RF maps) and maps ofthe locations and quality of sensations evoked by stimu-lation of thebrain (� projected field orPFmaps) (Lenzetal. 1994).WhileRFmapsarea reflectionof theorganiza-tion of inputs to the � central nervous system, PF mapsgive an indication of the image of the body contained inthe thalamus and cortex. There are numerous studies ofcortical plasticity secondary to injuries of the nervoussystem (Kaas 1991). Although much of the reorganiza-tion of cortical maps may reflect underlying changes inthalamic organization (Pons et al. 1991), there are rela-tively few direct studies of thalamic plasticity (Rasmus-son 1996).Medically intractable chronic pain due to nervous sys-tem injury or medically intractable movement disordersmay be surgically treated by implantation of deep brainstimulatingelectrodesintheprincipalsensorynucleusofthe thalamus (� ventral caudal – Vc) (Hosobuchi, 1986)or by � thalamotomy (Tasker et al. 1988). During suchoperations, microelectrode recordings may be used toconfirm the target predicted by the radiological studies.The physiological studies of Vc in patients with chronicpain syndromes are compared with the recordings of Vcin patients with movement disorders and thus plasticityof pain-related neuronal activity in the human thalamuscan be studied directly.The region of the principal sensory nucleus of thalamus(Vc) was explored during stereotactic surgical pro-cedures for treatment of patients with pain followingspinal cord transection and compared to results frompatients with movement disorders (Lenz et al. 1994).

T
Thalamus, Receptive Fields, Projected Fields, Human 2455
Many cells in the expected representation of the anes-thetic part of the body did not have RFs. However, in theborder zone/anesthetic zone between the anesthetic andinnervated areas of the thalamus, there was frequentlya mismatch between the location of RFs and PFs. OftenRFs were located on the chest and abdominal wall,above the level of the spinal transection, while PFswere located in the lower extremity, below the spinaltransection.Border zone/anesthetic areas of thalamus often exhib-ited increased representations of the border of the anes-thetic part of the body. Two sites were said to have a con-sistentRFif theRFofboth sites included thesamepartofthebody.Thelengthofa trajectorywithconsistentRFsisthe distance along the trajectory where each RF contin-uestoincludethesamepartof thebody(Lenzetal.1988).The maximal distance along a trajectory over which theRF or the PF stays consistent is longer for body partswith larger representations (Lenzetal. 1994).Lengthsoftrajectory with a consistent RF in a particular part of thebody were significantly longer in borderzone/anestheticzones than in controlareaswith apparentlynormal input.Neurons with RFs adjacent to the area of sensory lossin amputation patients (n=3) occupied a larger part ofthe thalamic homunculus (Lenz et al. 1998) than foundfor the same part of the body in patients with movementdisorders (Lenz et al. 1988; Lenz et al. 1994). This resultis consistent with somatotopic reorganization of affer-ent inputs from the limb. Similarly the large area overwhich PFs include the stump (Lenz et al. 1994; Lenz etal. 1993)) suggest that there has been reorganization ofthe perceptual image of the limb in the central nervoussystem (Jensen and Rasmussen 1994). It has also beenreported that phantom sensations can be evoked by stim-ulation of the region where stump RFs are located in theregion of Vc (Davis et al. 1998). Thus, in the case ofamputations and spinal cord injury, the alteration in theimage of the body in the thalamus is less than that of theinputs from that part of the body.In primates there are well-documented alterations inthalamic anatomy and physiology after peripheral nerveinjury. The distributions of thalamic regions with char-acteristic histology are altered (Rausell et al. 1992)in monkeys with a C2-T4 dorsal rhizotomy (Sweet1981). In the affected arm area of rhizotomized ani-mals, there is a reduction in the density of large cells andof � parvalbumin and � CO staining, all characteristicof the terminal zone for dorsal column inputs. There isa corresponding increase in the � calbindin staining inthe arm area, characteristic of the terminal zone for STTinputs.After cervical dorsal rhizotomy, large numbers of cellswithout RFs are encountered (Albe-Fessard and Lom-bard 1983) in the forelimb region of the monkey VP. Fol-lowing adult digit amputation, increased representationof the stump is found, with large RFs including adjacentdigits (Rasmusson 1996). The thalamic representation
of the border of the anesthetic part is increased in mon-keys with nerve sections (Garraghty and Kaas 1991).Studies have shown that stimulation of the somatic sen-sory thalamus is more likely to evoke pain in patientswith chronic pain after nervous system injury than in pa-tients without somatic sensory abnormalities (patientswith movement disorders) (Lenz et al. 1998). The re-gion of Vc was divided on the basis of projected fieldsinto areas representing the part of the body where thepatients experienced chronic pain (pain affected) or didnot experience chronic pain (pain unaffected) and intoa control area located in the thalamus of patients withmovement disorders and no experience of chronic pain.In both the core and posterior inferior regions of thethalamic sensory nucleus, the proportion of sites wherethreshold microstimulation evoked pain was larger inpain affected and unaffected areas than in control areas.Thenumberof siteswhere thermal (warm orcold) sensa-tions were evoked was correspondingly smaller, so thatthe total of pain plus thermal sites was not significantlydifferent across all areas (Lenz et al. 1998). Therefore,sites where stimulation evoked pain in patients withneuropathic pain may correspond to sites where ther-mal sensations were evoked by stimulation in patientswithout somatic sensory abnormality. In the posteriorinferior region, the number of sites where cold wasevoked by stimulation decreased significantly, whilethe number of sites where pain was evoked increasedsignificantly.These results suggest that pain is evoked in patientswith neuropathic pain by stimulation at sites wherethermal sensations would normally be evoked. There-fore, the present data suggest that the STT, or elementsto which the STT projects, signal pain rather than ther-mal sensations in patients with neuropathic pain. This isconsistent with the finding that stimulation of the STTevokes pain in patients with neuropathic pain but evokesnonpainful thermal sensations in patients who do nothave neuropathic pain (Tasker 1988). Cordotomy re-lieves pain in a much greater proportion of patients withsomatic pain than it does in patients with neuropathicpain (Sweet et al. 1994). The failure of cordotomy torelieve neuropathic pain might be anticipated from theoccurrence of central pain in patients with impairedfunction of the STT (Cassinari and Pagni 1969; Boivieet al. 1989). These results suggest that the generator forpain in patients with central pain is the terminus of theSTT.In patients with central pain, anatomic evidence ofdamage to STT is a common finding (Cassinari andPagni 1969) and loss of STT function, indicated byimpaired thermal and pain sensibility, is a uniformfinding (Boivie 1994). In patients with central pain,pain is more common than in controls, while thresholdmicrostimulation-evoked cold sensations are corre-spondingly less common. These findings suggest thatthere has been a reorganization, so that cold modalities

2456 Thalamus, Receptive Fields, Projected Fields, Human
are relabeled to signal pain in the thalamus of patientswith central pain (Ralston et al. 1996). The relationshipbetween thalamic stimulation-evoked cold and pain inpatients with central pain may explain the perception ofcold as pain (cold hyperalgesia) that can occur in thesepatients (Boivie 1994).Similar changes are observed after central nervous sys-tem injury. In patients with spinal transection, the num-bers of cellular RFs representing the border of the anes-thetic part of the body are increased (Lenz et al 1994).Following transection of the dorsal columns at the T3-T5 level, activity in simian VP (Pollin and Albe-Fessard1979) shows a decrease in the percentage of cells withhind limb RFs and an increase in the percentage of cellswith forelimb RFs. Many cells have large RFs and re-spond to high threshold inputs, consistent with inputsfrom STT. Therefore, loss of input due to peripheral orcentral nervous system injury leads to significant reor-ganization of the thalamic representation of inputs fromdifferent parts of the body.Studies of the thalamus in monkeys and humans with anervous system injury demonstrate large changes in theperceptual representation of the body in the thalamus, asrevealed by somatotopic maps of projected fields (Lenzet al. 1994; Lenz et al. 1998). Patients with amputationsshow increases in the thalamic area from which stimu-lation evokes sensations in the part of a limb that has be-come thestump (Lenzetal. 1998). Inpatientswith spinaltransections, the incidence of mismatches between neu-ronalRFsand threshold microstimulation-evokedPFsatthe same sites is much higher than in patients with move-ment disorders (Lenz et al. 1994). In patients with spinaltransection, these mismatches occur because sensationsin the anesthetic part are evoked by stimulation at siteswhere cellular RFs represent parts of the body proximalto the anesthetic part. In these patients, stimulation in thethalamic area where the representation of the anestheticpart of the body is usually located evokes sensations inthe anesthetic part (Lenz et al. 1994). These results sug-gest that the perceptual representation of the body in thethalamus reorganizes less than the representation of in-puts to the thalamus. Although the central image of thebody is relatively constant in the face of altered input,our studies show that changes in the modality organi-zation of this image can change dramatically (Lenz etal. 1998). This plasticity of modality organization maycontribute to the development of chronic pain in patientswith nervous system injury.
References1. Albe-Fessard DG, Lombard MC (1983) Use of an animal model
to evaluate the origin of and protection against deafferentationpain. Adv Pain ResTher 5:691–700
2. Boivie J (1994) Central pain. In: Wall PD, Melzack R (eds) Text-book of Pain. Churchill Livingston, Edinburgh, pp 871–902
3. Boivie J, Leijon G, Johansson I (1989) Central post-stroke pain– a study of the mechanisms through analyses of the sensoryabnormalities. Pain 37:173–185
4. Cassinari V, Pagni CA (1969) Central Pain. A neurosurgical sur-vey. Harvard University Press, Cambridge, Massachusetts
5. Davis KD, Kiss ZHT, Luo L et al. (1998) Phantom sensationsgenerated by thalamic microstimulation. Nature 391:385–387
6. Garraghty PE, Kaas JH (1991) Functional reorganization in adultmonkey thalamus after a peripheral nerve injury. Neuroreport2:747–750
7. Hosobuchi Y (1986) Subcortical electrical stimulation for controlof intractable pain in humans. J Neurosurg 64:543–553
8. Jensen TS, Rasmussen P (1994) Phantom pain and related phe-nomena after amputation. In: Wall PD, Melzack R (eds) Textbookof pain. Churchill Livingstone, New York, pp 651–665
9. Kaas JH (1991) Plasticity of sensory and motor maps in adultmammals. Annu Rev Neurosci 14:137–167
10. LenzFA, DostrovskyJO, TaskerRRetal. (1988) Single-unit anal-ysis of the human ventral thalamic nuclear group: somatosensoryresponses. J Neurophysiol 59:299–316
11. Lenz FA, Tasker RR, Kwan HC et al. (1988) Single unit analysisof the human ventral thalamic nuclear group: correlation of tha-lamic “tremor cells” with the 3-6 Hz component of parkinsoniantremor. J Neurosci 8:754–764
12. Lenz FA, Seike M, Lin YC et al. (1993) Thermal and pain sen-sations evoked by microstimulation in the area of the humanventrocaudal nucleus (Vc). J Neurophysiol 70:200–212
13. Lenz FA, Kwan HC, Martin R et al. (1994) Characteristics ofsomatotopic organization and spontaneous neuronal activityin the region of the thalamic principal sensory nucleus inpatients with spinal cord transection. J Neurophysiol 72:1570–1587
14. Lenz FA, Gracely RH, Baker FH et al. (1998) Reorganizationof sensory modalities evoked by stimulation in the region of theprincipal sensory nucleus (ventral caudal – Vc) in patients withpain secondary to neural injury. J Comp Neurol 399:125–138
15. Lenz FA, Zirh AT, Garonzik IM et al. (1998) Neuronal activityin the region of the principal sensory nucleus of human thalamus(ventralis caudalis) in patients with pain following amputations.Neurosci 86:1065–1081
16. Pollin B, Albe-Fessard DG (1979) Organization of somatic thala-mus in monkeys with and without section of dorsal spinal tracts.Brain Res 173:431–449
17. Pons TP, Garraghty PE, Ommaya AK et al. (1991) Massivecortical reorganization after sensory deafferentation in adultmacaques. Science 252:1857–1860
18. Ralston HJ, Ohara PT, Meng XW et al. (1996) Transneuronalchanges in the inhibitory circuitry of the macaque somatosensorythalamus following lesions of the dorsal column nuclei. J CompNeurol 371:325–335
19. Rasmusson DD (1996) Changes in the response properties ofneurons in the ventroposterior lateral thalamic nucleus of the rac-coon after peripheral deafferentation. J Neurophysiol 75:2441–2450
20. Rausell E, Cusick CG, Taub E et al. (1992) Chronic Deaf-ferentation in monkeys differentially affects nociceptive andnon-nociceptive pathway distinguished by specific calcium-binding protiens and down-regulates gamma-aminobutyric acidtype A receptors at thalamic levels. Proc Natl Acad Sci USA89:2571–2575
21. Sweet WH (1981) Animal models of chronic pain: their possiblevalidation from human experience with posterior rhizotomy andcongenital analgesia. Pain 10:275–295
22. Sweet WH, Poletti CE, Gybels GM (1994) Operations in thebrainstem and spinal canal with an appendix on the relationshipof open and percutaneous cordotomy. In: Wall PD, Melzack R(eds) Textbook of Pain. Churchill and Livingston, New York, pp1113–1136
23. Tasker RR (1988) Percutaneous Cordotomy: The Lateral HighCervical Technique. In: Schmidek HH, Sweet WH (eds) Oper-ative Neurosurgical Techniques Indications, Methods, and Re-sults. Saunders WB, Philadelphia, pp 1191–1205
24. Tasker RR, Doorly T, Yamashiro K (1988) Thalamotomy in Gen-eralized Dystonia. Adv Neurol 50:615–631

T
Thalamus, Visceral Representation 2457
Thalamus, Visceral RepresentationA. VANIA APKARIAN
Feinberg School of Medicine, Department ofPhysiology, Northwestern University, Chicago, IL,[email protected]
Synonyms
Interoception; Visceral Modulation; Visceral Represen-tation; referred pain
Definition
Stimulation of visceral organs distinctly activates dif-ferent populations of neurons in the thalamus. Neuronslocated in � parvocellular VP (VPpc) are described asvisceral-specific, while neurons located in other medialand lateral thalamic nuclei respond to multiple visceraand to somatic inputs convergently. Other thalamic nu-clei do not receive visceral inputs.
Characteristics
In contrast to somatic stimuli, visceral stimuli may ormay not be accompanied by perception and the per-ceptions show distinct properties. For example, baro-and chemo-receptor activation do not give rise to sen-sations/perceptions, although they are represented invisceral specific portions of the thalamus and in insularcortex. On the other hand, distension of most hollowviscera is associated with ill-defined sensations. Vis-ceral nociceptive stimuli in such organs are generallycharacterized by a sense of malaise or discomfort, withno clear ability to localize the source of the stimu-lus. Moreover, visceral pains are usually associatedwith pain referred on the skin, the location of whichis characteristic for different viscera, an associationused since antiquity to pinpoint specific viscera asthe source of injury or inflammation. Thus, the con-trast between visceral and somatic representation andassociated differences in perception provides an op-portunity for dissecting conscious perceptions fromunconscious modulation, in relationship to thalamocor-tical connectivity. Unfortunately very little effort hasbeen invested in this direction. It should also be addedthat visceral stimuli that do not evoke any conscioussensations and are represented at the level of the thala-mus and cortex, most probably play a modulatory rolein emotional responses to the environment and exertemotionally driven modulatory control over somaticand visceral responses. This notion, however, remainsmainly a speculation, since direct studies on the topicare minimal (Gebhart 1995). Here we concentrate onthalamic neuronal response properties from the view-point of coding visceral stimuli and being modulatedby visceral stimuli. All the available data are from
anesthetized preparations; thus they should be regardedas examples of responses undoubtedly stunted due toanesthesia.The earliest evidence for visceral inputs to the thala-mus and cortex used electrical stimulation of the vagus(Bailey and Bremer 1938; Dell and Olson 1951) anddemonstrated a relay from the lateral thalamus to the� insular cortex from visceral organs. More recentanatomic studies have elucidated that the visceral in-puts through vagal afferents terminate in the solitarytract, second order neurons then project ipsilaterallyto the parabrachial nucleus and third order neuronstravel contralaterally to the ventroposterior parvocel-lular nucleus of the thalamus (VPpc) and then projectto the insula. A separate projection from parabrachialregions directly accesses the insula too, see chapter byCechetto in Gebhart (1995). � Calcitonin gene relatedpeptide (CGRP) seems to label this pathway in rats aswell as in man. Using a combination of physiologicalrecordings from insular cortex and tracing techniques,it was demonstrated that the most medial portion ofthe VPpc projects to the insular region with gustatoryresponses, while a more lateral portion projects to partof the insula with gastric mechanoreceptor responsiveneurons and the most lateral portion projects to an areawith cardiopulmonary responses; parabrachial inputsto the thalamus seem to have a parallel organization aswell (Cechetto and Saper 1987; Gebhart 1995). Morerecent studies in the rat confirm the parabrachial inputsand also show inputs from spinal cord and trigemi-nal lamina I (Bester et al. 1999; Gauriau and Bernard2004). Thus, the VPpc portion of the thalamus seemsto be viscerotopically organized, where gustatory re-sponses have been shown physiologically (Ganchrowand Erickson 1972).Inputs from pelvic viscera to the cat lateral thalamushave been mapped with electrical stimulation and nat-ural stimulation of the urinary bladder (Brüggemann etal. 1993; Brüggemann et al. 1994). Identified neuronswere all found located outside the � VPL, either justin its periphery or in the adjacent � posterior nucleus(PO). All visceral responsive neurons also respondedto low-threshold stimuli applied to the skin. A simi-lar study examined renal nerve stimulus responses tomap kidney representation in and around the � VPLin the cat (Horn et al. 1997). Most responsive unitswere located in the periphery of the VPL, dorsal PO orlateral PO; none were found within the VPL. Responselatency suggested large and small myelinated afferentfibers mediating kidney inputs. The somatic receptivefields of the cells with renal nerve inputs showed mini-mal correspondence with the dermatomes of the renalnerve. A subsequent study mapped the same region ofthe thalamus in the cat for bladder, colon and esophagusinputs and again found the majority of cells localizedto either the periphery of the VPL or the � PO and noindication of segregation of neurons based on responses

2458 Thalamus, Visceral Representation
Thalamus, Visceral Representation, Figure 1 Visceral responses of aVPL neuron with low-threshold somatic response (touching digit 5, D5 onthe foot contralateral to the recording) in the anesthetized squirrel monkey.Intraluminal pressures are shown for each viscus distended. The neuronresponds by increased activity to noxious distension of the bladder, colon,and esophagus (Bruggemann, Shi and Apkarian 1994).
to any particular viscus (Ganchrow and Erickson 1972;Horn et al. 1999). Visceral responsive cells have beenstudied in the medial thalamus of the cat (1995) and
localized within the � mediodorsal (MD), � centralmedial (CM), � central lateral (CL), periphery of theVPL and � ventral posterior medial (VPM) nuclei aswell as in the � zona incerta (ZI). More than 23% re-sponded to esophagus (of 120 neurons examined), 8%to bladder and 6% to colon distension. In contrast to thelateral thalamic neurons, all medial thalamic visceralresponsive cells had nociceptive inputs from the skin.In the squirrel monkey, lateral thalamus responses todistending the urinary bladder, distal colon and loweresophagus, as well as to noxious and innocuous somaticstimuli have been mapped (Brüggemann et al. 1994).Eighty-five percent (of 106 neurons studied) respondedto at least one of the viscera. Most visceroceptive cellshad somatic low-threshold responses and convergentmultivisceral responses. Figure 1 is an example of a neu-ron located in the VPL, it responds to all three viscera byexcitation and hasasomatic tactile receptivefield onfootdigit 5. The visceral responsive cells showed increasedor decreased firing for distensions in the noxious rangeand some also coded distensions in the innocuous andnoxious ranges. Figure 2 shows visceral and somaticproperties of 20 VPL neurons identified in a singleelectrode penetration. The extent of unpredictabilitybetween adjacent neurons, regarding specific visceralinputs and their excitatory or inhibitory responses isevident. Modulatory effects of visceral stimulation havebeen examined in squirrel monkey VPL (Brüggemannet al. 1998) and indicate that noxious distensions ofurinary bladder, distal colon or lower esophagus de-crease responses to somatic stimuli by about 50%. Thehigh incidence of visceral responsive cells in the VPLimplies that this information must be transmitted to thecortex and such visceral responsive neurons have in factbeen reported in the monkey primary somatosensorycortex (Brüggemann et al. 1997).Neuronal responses to uterus, cervix, vagina, colon andskin have been examined in the rat thalamus (Berkleyet al. 1993; Berkley et al. 1995, Guilbaud et al. 1993).In the lateral thalamus, most neurons responded to mul-tiple viscera and most were located in and around theborder of ventrobasal complex (VB). Like results seenin thecatmedial thalamus,mostneurons in thecaudal in-tralaminar thalamic nuclei (IL) with visceral responsesalso responded to noxious somatic stimuli, and most hadinputs from multiple viscera (Berkley et al. 1995). Thelateral portion of ventral medial nucleus (VMl) has re-cently been established as a region important in signal-ing nociceptive information from the brainstem to thecortex (see chapter in this encyclopedia by Villanueva).Cells in this region respond to noxious mechanical andthermal stimuli applied anywhere on the body, yet sur-prisingly theseneuronsdonotrespondtovisceralstimuli(Monconduit et al. 2003).Overall, thalamic nuclei can be differentiated into threekinds: nuclei with visceral specific inputs, nuclei withvisceral convergent inputs and nuclei with no visceral

T
Thalamus, Visceral Representation 2459
Thalamus, Visceral Representation, Figure 2 Visceral and somatic re-sponses of 20 neurons identified in a single track by an electrode traversingVPL dorso-ventrally. Cell 1 (#) is located most dorsally. Somatic receptivefields are either low-threshold tactile (unmarked) or wide-dynamic rangetype marked next to the receptive fields (open circle). Visceral responsesare: 0, no response, -, inhibition, +, excitation, ±, mixed; due to distensionof the urinary bladder (B), colon (C), or lower esophagus (E) (missing sym-bols, not tested; N.A. somatic receptive field not found) (From Bruggemann,Shi and Apkarian 1994).
inputs. Surprisingly this differentiation does not followthe organization of the thalamus regarding somatic no-ciceptive and non-nociceptive responses, which was aprimary hypothesis for most of the studies in the topic.The region of the thalamus presumed to be organizedviscerotopically has not been adequately studied, espe-cially physiologically and, although it is commonly re-ferred as the visceral specific part of the thalamus, its de-tailed response properties remain unknown. However,it is possible that the observations in the cat VPL pe-riphery and PO are actually part of the same visceralspecificnucleus (VPpc)extending laterally anddorsally.There are important species differences in the visceralorganization of the thalamus. In the cat, there is goodreproducible evidence that the VPL proper, and mostlikely also the VPM proper, do not receive visceral in-puts, while the VPL and VPM in rat and monkey do.In fact, in the squirrel monkey there is good evidencethat all neurons in the VPL and VPM receive visceralinputs, non-viscerotopically. There is also good agree-ment between studies and across species for medial vis-ceroceptivecellshaving convergent somaticnociceptiveinputs, while lateral visceroceptive cells have mainly in-nocuous somatic inputs. Given the widely convergentviscero-visceral inputson somatic responsivecells in themonkey lateral thalamus, it has been proposed that thevisceral inputs through dorsal column inputs provide thesignal for poorly localized visceral perceptions and thatthe spinothalamic inputs (presumed to be more specificand more dominantly somatic) may then lead to the per-ception of referred pain on the skin; see chapter by Ap-karian in (Gebhart 1995). Undoubtedly, visceral inputsto the medial thalamus would complement this model byproviding modulation of affect. The distinct visceral or-ganizational rules of the thalamus, coupled with uniqueperceptions should provide the opportunity for studyingthe brain circuitry regarding different types of percep-tions, yet current research in this field remains limited.
References1. Bailey P, Bremer F (1938) A sensory cortical representation of the
vagus nerve with a note on the effects of the low blood pressureon the cortical electrogram. J Neurophysiol 1:405–412
2. Berkley KJ, Guilbaud G, Benoist JM et al. (1993) Responsesof neurons in and near the thalamic ventrobasal complex of therat to stimulation of uterus, cervix, vagina, colon, and skin. JNeurophysiol 69:557–568
3. Berkley KJ, Benoist JM, Gautron M et al. (1995) Responses ofneurons in the caudal intralaminar thalamic complex of the rat tostimulation of the uterus, vagina, cervix, colon and skin. BrainRes 695:92–95
4. Bester H, Bourgeais L, Villanueva L et al. (1999) Differentialprojections to the intralaminar and gustatory thalamus from theparabrachial area: a PHA-L study in the rat. J Comp Neurol405:421–449
5. Brüggemann J, Vahle-Hinz C, Kniffki KD (1993) Representa-tion of the urinary bladder in the lateral thalamus of the cat. JNeurophysiol 70:482–491
6. Brüggemann J, Shi T, Apkarian AV (1994) Squirrel monkey lat-eral thalamus. II. Viscerosomatic convergent representation ofurinary bladder, colon, and esophagus. J Neurosci 14:6796–6814

2460 Thalassotherapy
7. Brüggemann J, Vahle-Hinz C, Kniffki KD (1994) Projectionsfrom the pelvic nerve to the periphery of the cat’s thalamic ven-tral posterolateral nucleus and adjacent regions of the posteriorcomplex. J Neurophysiol 72:2237–2245
8. Brüggemann J, Shi T, Apkarian AV (1997) Viscero-somatic neu-rons in the primary somatosensory cortex (SI) of the squirrelmonkey. Brain Res 756:297–300
9. Brüggemann J, Shi T, Apkarian AV (1998) Viscerosomatic in-teractions in the thalamic ventral posterolateral nucleus (VPL)of the squirrel monkey. Brain Res 787:269–276
10. Cechetto DF, Saper CB (1987) Evidence for a viscerotopic sen-sory representation in the cortex and thalamus in the rat. J CompNeurol 262:27–45
11. Dell P, Olson R (1951) Projections thalamiques, corticales etcerebelleuses des afferences viscerales vagagles. Soc Biol (Paris)145:1084–1088
12. Ganchrow D, Erickson RP (1972) Thalamocortical relations ingustation. Brain Res 36:298–305
13. Gauriau C, Bernard JF (2004) A comparative reappraisal of pro-jections from the superficial laminae of the dorsal horn in therat: the forebrain. J Comp Neurol 468:24–56
14. Gebhart GF (1995) Visceral pain. IASP Press, Seattle15. Guilbaud G, Berkley KJ, Benoist JM et al. (1993) Responses
of neurons in thalamic ventrobasal complex of rats to gradeddistension of uterus and vagina and to uterine suprafusion withbradykinin and prostaglandin F2 alpha. Brain Res 614:285–290
16. Horn AC, Vahle-Hinz C, Petersen M et al. (1997) Projectionsfrom the renal nerve to the cat’s lateral somatosensory thalamus.Brain Res 763:47–55
17. Horn AC, Vahle-Hinz C, Bruggemann J et al. (1999) Responses ofneurons in the lateral thalamus of the cat to stimulation of urinarybladder, colon, esophagus, and skin. Brain Res 851:164–174
18. Monconduit L, Bourgeais L, Bernard JF et al. (2003) Conver-gence of cutaneous, muscular and visceral noxious inputs ontoventromedial thalamic neurons in the rat. Pain 103:83–91
Thalassotherapy
Definition
Bathing in the sea.� Spa Treatment
Tolosa-Hunt Syndrome (PainfulOphthalmoplegia)
� Headache Due to Dissection
Therapeutic Acupuncture
Definition
Therapeutic Acupuncture refers to the clinical use ofacupuncture for the more long-term relief of differentsymptoms after a course of treatments.� Acupuncture Mechanisms
Therapeutic Alliance
� Chronic Pain, Patient-Therapist Interaction
Therapeutic Cold
� Therapeutic Heat, Microwaves and Cold
Therapeutic Drug Monitoring
Definition
The use of serum drug concentrations to guide dosingof the drug in order to obtain optimum treatment effectand avoid toxicity.� Antidepressants in Neuropathic Pain
Therapeutic Exercise
Definition
Exercise as part of a treatment program designed to im-prove an individual’s ability to move, balance and co-ordination, endurance, flexibility, muscle tone, posture,and strength.� Chronic Pain in Children: Physical Medicine and Re-
habilitation
Therapeutic Gain
Definition
Therapeutic gain is the difference between the therapeu-tic response to the verum and the placebo in a random-ized controlled trial, in migraine prophylaxis, betweenthe percentage of „responders“, i.e. of patients with a50% reduction of attack frequency, to the activedrug andthe percentage of responders to placebo.� Clinical Migraine without Aura
Therapeutic Gene Transfer
� Opioids and Gene Therapy
Therapeutic Heat
� Therapeutic Heat, Microwaves and Cold

T
Therapeutic Heat, Microwaves and Cold 2461
Therapeutic Heat, Microwaves and ColdJEFFREY R. BASFORD
Department of Physical Medicine and Rehabilitation,Mayo Clinic and Mayo Foundation, Rochester, MN,[email protected]
Synonyms
Physical agent; modality; diathermy; Therapeutic Cold;Therapeutic Heat; Cold Therapy; Microwaves
Definition
The use of heat and cold to lessen pain, promote healingor obtain other therapeutic goals.
Characteristics
The body is sensitive to its environment. Temperaturesabove 42˚C or below 13˚C produce discomfort and thoseonly a few degrees higher or lower may actually injure it.Furthermore, temperaturechangeseasily obtained in theclinic alter enzymatic activity, nerve conduction and tis-sue viscosity (Oosterveld and Rasker 1994). While heatand cold may be used to alleviate the cause of pain or toproduce analgesia, the latter is far more common.Thermal agents achieve their effects as the result ofchanging the temperature of a tissue. Specifically, mostheat-based treatments attempt to warm tissues to by3–8˚C and most cold therapies attempt to reduce tis-sue temperatures by a similar amount. As a result, theoverlap between their indications and contraindica-tions is surprisingly broad (Tables 1 and 2). It shouldbe remembered that many of the contraindications ofheat and cold are relative, i.e. localized heating mightbe acceptable in a patient with cardiac disease, whilesystemic heating might place unacceptable demandson his cardiac function (Keast and Adamo 2000).
Mechanism of Action
Tissue can be heated or cooled by conduction, con-vection or the conversion of a different form of energyinto heat. Hot packs epitomize conductive heating,hydrotherapy convection and ultrasound conversion,due to its reliance on the conversion of sound into heat.
Superficial Heat
The physiological effects of the superficial heatingagents differ little and choice depends on the situationand patient / therapist preference.
Hot Packs
Hot packs are typically constructed of porous bags filledwith a hygroscopic material. They are either kept in70–80˚C water baths or warmed in a microwave beforebeing covered with an absorbent wrap and placed onthe body. Treatments may last for 30 min with the packsslowly cooling. Alternatives such as electrically heated
pads that do not cool spontaneously are convenient butmay increase the risk of burns.
Heat Lamp
Heat lamp usemay bedecliningbut is still common.Skintemperaturesarecontrolled by adjusting thedistancebe-tween the lamp and the patient. The precautions in Table1 apply to these agents but it should also be noted thatchronic use of superficial heat can produce a permanentmottling of the skin (erythema ab igne).
Hydrotherapy
Hydrotherapy uses a fluid medium to produce heating,cooling,massageanddebridement.Neitheragitationnorwater is necessary and solutes in the medium (e.g. NaClfor wound care) may be a significant factor in treatment.Hydrotherapy may also be performed with substancessuch as finely ground solid materials suspended by jetsof hot air. The benefits of this dry heat ‘fluidotherapy’approach over conventional water-based hydrotherapyare unestablished.
Whirlpool Baths
Whirlpoolbathsrangeinsizefromthose intended totreatasingleextremitytoothersinwhichtheentirebodyisim-mersed. Temperatures of 33–36˚C are usually well tol-erated, although for a healthy patient, elevations to 43˚Care possible on limited portions of the body. Treatmentwith stationary mediums may be beneficial. Sitz baths,for example, are beneficial in the treatment of anorectalpain and research demonstrates that bathing at 40–50˚Cis not only comfortable but also lessens anal tone (Pinhoet al. 1993). Agitation increases theefficiency of thermaltransfer and it should be remembered that a temperaturethat is comfortable with a motionless medium might be-come painfully hot or cold with agitation.
Contrast Bathing
Contrast bathing involves the patient shifting theirtreated extremities alternatively between a warm(38–40˚C) and a cool (13–16˚C) bath about ten times.These baths are frequently used in the treatment ofcomplex regional pain syndrome with benefits thoughtto result from reflex hyperemia and desensitization.
Paraffin Baths
Paraffin baths consist of a basin filled with a 1:7 mixtureof mineral oil and paraffin. Temperatures (45–54˚C) arehigher than those of water-based hydrotherapy and aretolerated due to the lack of agitation, the insulation pro-vided by wax as it solidifies on the treated area and thelow heat capacity of the medium. Treatment is usuallyperformed by dipping the involved extremity in the bathabout ten times, covering it and then allowing it to coolslowly in an insulated wrap. An alternative approach inwhich the treated extremity is dipped once and then keptin the bath permits more vigorous heating. 30 min treat-ments may produce increases in the intramuscular tem-

2462 Therapeutic Heat, Microwaves and Cold
Therapeutic Heat, Microwaves and Cold, Table 1 Indications and Precautions for the Use of Therapeutic Heat
Indications Precautions and Contraindications
PainAcute inflammation, trauma or hemorrhageMuscle spasmBleeding dyscrasiasContracturesInsensitivityFibromyalgiaInability to communicate or respond to painHyperemiaPoor thermal regulation (systemic heating situation)
Acceleration of metabolic processesMalignancyChronic HematomaEdemaBursitis / TenosynovitisIschemiaSuperficial thrombophlebitisAtrophic skin / scar tissueUnstable angina or blood pressureDecompensated heart failure / recent myocardial infarction
Therapeutic Heat, Microwaves and Cold, Table 2 Indications and Precautions for the Use of Therapeutic Cold
Indications Precautions and Contraindications
Acute musculoskeletal traumaIschemiaPainCold intolerance / urticariaMuscle spasmRaynaud’s phenomenon and disease
SpasticitySevere cold pressor responsesReduction of metabolic activityInability to communicate or respond to painPoor thermal regulationInsensitivity
perature of superficial muscles of about 3˚C (Abramsonet al. 1964).
Diathermy� Diathermy can be performed with short waves
(SWD), microwaves (MWD) or ultrasound (USD).USD is the most frequently used agent (Lindsay etal. 1995) but as it is discussed in another chapter willbe only summarized here. Discussion here will em-phasize SWD which is still in relatively widespreaduse and to a lesser extent, MWD whose medical useis now quite restricted.
Ultrasound
Ultrasound has both thermal and � non-thermal effects.The production and effects of heat are well understood.The benefits of non-thermal processes, which includesuch phenomena as cavitation (the production anddestruction of small bubbles), tissue micro-streamingand mechanical deformation remain to be established.Treatment usually involves stroking an USD applicatorover the treated tissue for 5–7 min. Intensities rangefrom mWs to 1.5 + W per cm2. Continuous waves areemployed when the goal is heating and pulsed treat-ments are chosen to emphasize non-thermal effects.Temperature elevations of 5˚C are easily possible andmay be particularly large at bone:soft tissue interfaces.USD phonophoresis is also used to introduce topicalmedication (e.g. lidocaine) through the skin.USD used as a treatment for conditions ranging fromcontractures, sprains, muscle strains, wounds, tendinitisand non-healing fractures to carpal tunnel syndrome.Benefits are controversial and in some situations may beno more effective than placebos or anti-inflammatories(Basford 1998). Precautions include those for heat ingeneral, as well as avoidance of fluid-filled cavities,
the gravid or mensurating uterus, the heart, brain, cer-vical ganglia, tumors, laminectomy sites, and acutelyinflamed joints.
Short Wave Diathermy
Short wave diathermy uses radio waves to heat tis-sue. Use is restricted to a limited range of frequencies(27.12 MHz, 13.56 MHz and 40.68 MHz in the U.S.).In one approach, the body acts as an antenna and theSWD machine induces eddy currents that produce heatas they flow through the body. This method deliversthe most energy to water-rich conductive tissues suchas muscles. In the second approach, the body servesas the dielectric of a capacitor that is charged by theSWD machine. In this case, the tissues are in series andheating may be most marked in water-poor, high re-sistance tissues, such as fat and ligaments. Applicatorsrange from inductive pads that are placed on the patientto the flat plates of a simple capacitor. Although nowrarely done, specialized heating can be performed bywrapping coils around a patient’s limb or with rectal orvaginal probes. Continuous waveforms are used whenthe goal is heating while pulsed waves are used whennon-thermal effects are desired. SWD can increase sub-cutaneous fat temperatures by 15˚C and 3–5 cm deepintramuscular temperatures by 4–6˚C (Draper et al.1999). A SWD device is, in effect, a radio transmitterthat is used to produce heat in two ways. As is truefor US, the benefits of SWD non-thermal phenomena(such as possible frequency dependent effects on cellfunction) remain to be established.
Microwaves
Microwaves (915 MHzand 2,456 MHz)donotpenetratetissue as deeply as SWD. In fact, their absorption is sorapid that fat overlying a site of interest will absorb a sig-

T
Therapeutic Heat, Microwaves and Cold 2463
nificantportionof thebeam.Thus,microwavediathermy(MWD) may increase subcutaneous fat temperatures by10to12˚C,while theunderlyingmuscleswillbewarmedonly a third as much (Basford 1998). Microwaves havebeen replaced in therapy by US and SWD. Today MSDuse in medicine appears to be restricted to the produc-tion of local hyperthermia and the potentiation of cancerchemotherapy and radiation treatment.SW and MW diathermy are both subject to the generalprecautions for heat outlined in Table 1. However, bothare electromagnetic in nature and metal implants / de-vices, pacemakers, stimulators, contact lenses and themenstruating or pregnant uterus should be avoided.Risks are real; diathermy treatment of the jaw resultedin severe brain damage in a man with a deep brainstimulator (Nutt et al. 2001).
Cold Therapy
Cold decreases metabolic activity, slows nerve conduc-tion, produces analgesia, lessens muscle tone, inhibitsspasticity and increases gastrointestinal motility (Bas-ford 1998; Denys 1991). The application of ice to thebody decreases skin temperature by 20˚C in about10 min. Subcutaneous temperatures decrease 3–5˚Cover the same period. If cooling is continued, forearmintramuscular temperature may decrease by 6–16˚Cand muscle blood flow by as much as 30% (Oosterveldand Rasker 1994). Although chemical and refriger-ated agents may have temperatures below 0˚C and canproduce frostbite, ice treatments of healthy people forperiods of less than 30 min do not seem to cause injury.
Technique
Ice has a high heat capacity; ice packs, massage, com-pression wraps and slushes all cool tissues rapidly.Treatments tend to last 10–20 min and a slightly damp,thin towel may be placed between the ice and the skin.Iced whirlpools provide particularly rapid cooling.However, they are poorly accepted by most people.Insulated foot coverings or fabric socks and gloves maylessen discomfort and increase acceptance. Ice massageproduces rapid cooling over a limited area. Treatmentinvolves rubbing ice (often pre-frozen in a paper cup)over the painful area. Analgesia can be achieved in7–10 min.There are a number of other cooling agents. Vapocoolantsprays can reduce skin temperature by 20˚C (Oosterveldand Rasker 1994) and are used for local skin analge-sia and the “spray and stretch” techniques. Chemical icepackscoolvia theproduction ofan endothermic reactionand while convenient tend to be expensive. Refrigeratedand pressurized water pressure cuffs are also available.Frozen juice and vegetable packages are convenient forhome use.The application of superficial cold appears to lessen hy-poxic damage, edema and compartmental pressures af-ter injury (Bert et al. 1991), but the magnitude of its ben-
efits in theultimate recovery remain debatabledespite itsability to lessen metabolic activity and blood flow (Hoet al. 1995). In practice, ice is typically used in conjunc-tion with rest, compression and elevation (RICE) in thetreatment of many musculoskeletal injuries. A commonregimen, such as for an ankle sprain, consists of usingice acutely for about 20 min every 2 h for 6–24 h. Al-though icing is almost the automatic response to acutesoft-tissue injury, it may not be a panacea in that studiesof thepostsurgicalknee (Daniel et al. 1994)andcesareansections (Amin-Hanjani et al. 1992) may not show ben-efit.After the first day or two, the choice between ice-basedtherapy and heat appears to depend on personal choice.Many find heat more comfortable and use it unlessthere is a worsening of edema or pain. Others find acombination of icing and active exercising a more ef-fective way to speed recovery. In any case, long-termrecovery depends on mobilization and exercise. Heatand cold are only important as the agents used to assistin gaining it. Patients with long-standing trocantericbursitis and lateral epicondylitis may find ice in combi-nation with friction massage extraordinarily effective.Ice massage and transcutaneous electrical nerve stim-ulation (TENS) may be equally beneficial in low backpain (Basford 1998). The precautions of Table 2 shouldbe heeded. Elevation of blood pressure as well as theeffects of cold-induced vasoconstriction in people withischemia and Raynaud’s phenomenon may be importantconsiderations.In summary, there is little evidence that heat and coldalone are of much benefit except for the temporary re-duction ofpain.Asa result, they areusuallyprescribed inconjunction with a program of education, activity mod-ification and exercise. Preference plays a role in agentchoicebut thereare someguidelines thatareappropriate.Thus, acute musculoskeletal conditions are usually ini-tially treated with ice. In addition, hydrotherapy or SWDareused to treat largeareasof thebodywhileUSDisusedfor more focal conditions. Deep heat may appear physi-ologically appealing, but at times the comforting effectsof superficial heat may prove as beneficial.
References1. Abramson DI, Tuck S, Chu LSW et al. (1964) Effect of paraffin
bath and hot fomentations on local tissue temperatures. ArchPhys Med Rehabil 45:87–94
2. Amin-Hanjani S, Corcoran J, Chatwani A (1992) Cold therapy inthe management of postoperative cesarean section pain. J ObstetGynecol 167:108–109
3. Basford JR (1998) Physical Agents. Rehabilitation Medicine:Principles and Practice, 2nd edn. Lippincott-Raven, Philadelphia,pp 483–504
4. Beenakker EA, Oparina TI, Teelken A et al. (2001) Cooling gar-ment treatment in MS: clinical improvement and decrease inleukocyte NO production. Neurology 57:892–894
5. Bert JM, Stark JG, Maschka K et al. (1991) The effect of coldtherapy on morbidity subsequent to arthroscopic lateral retinac-ular release. Orthop Rev 20:755–758

2464 Therapeutic Relationships
6. Daniel DM, Stone ML, Arendt DL (1994) The effect of cold ther-apy on pain, swelling, and range of motion after anterior cruciateligament reconstructive surgery. Arthroscopy 10:530–533
7. Denys EH (1991) AAEM minimonograph #14: the influenceof temperature in clinical neurophysiology. Muscle Nerve14:795–811
8. Draper DO, Knight K, Fujiwara T et al. (1999) Temperaturechange in human muscle during and after pulsed short-wavediathermy. J Orthop Sports Phys Ther 29:13–8; discussion 19–22
9. Ho SSW, Illgen RL, Meyer RW et al. (1995) Comparison ofvarious icing times in decreasing bone metabolism and bloodflow in the knee. Am J Sports Med 23:74–76
10. Keast ML, Adamo KB (2000) The Finnish sauna bath and itsuse in patients with cardiovascular disease. J Cardiopulm Rehabil20:225–230
11. Lindsay DM, Dearness J, McGinley CC (1995) Electrotherapyusage trends in private physiotherapy practice in Alberta. Phys-iother Can 47:30–34
12. Nutt JG, Anderson VC, Peacock JH et al. (2001) DBS anddiathermy interaction induces severe CNS damage. Neurology56:1384–1386
13. Oosterveld FG, Rasker JJ (1994) Effects of local heat and coldtreatment of surface and articular temperature of arthritic knees.Arthritis Rheum 31:1578–1582
14. Pinho M, Correa JCO, Furtado A et al. (1993) Do hot baths pro-mote anal sphincter relaxation? Dis Colon Rectum 36:273–274
Therapeutic Relationships
� Chronic Gynaecological Pain, Doctor-Patient Inter-action
Therapeutic Ultrasound
� Modalities
Therapy of Pain, HypnosisROBERT G. LARGE
The Auckland Regional Pain Service, AucklandHospital, Auckland, New [email protected]
Synonyms
Hypnotism; Mesmerism; Hypnotherapy
Definition
Hypnosis refers to the “state” of consciousness associ-ated with the phenomenon in question.Hypnotism refers to the science and art of inducing andutilising this phenomenon.This term is now less used incurrent research literature.Mesmerism is a term associated with the controversialwork ofFranzAnton Mesmer (1734–1815)and replacedhis concept of “animal magnetism”. Modern accountsoccasionally use the term to describe the non-verbal“mesmeric passes” used in some contexts.
Hypnotherapy is a term used to describe various formsof psychotherapy utilizing hypnosis as the major ingre-dient.Hypnosis has been notoriously difficult to define. TheBritish Medical Association (1955) introduced the fol-lowing operational definition:Hypnosis is a temporary condition of altered perceptionin the subject which may be induced by another per-son and in which a variety of phenomena may appearspontaneously or in response to verbal or other stimuli.These phenomena include alterations in consciousnessand memory, increased susceptibility to suggestion,and the production in the subject of responses and ideasunfamiliar to him in his normal state of mind. Furtherphenomena such as anaesthesia, paralysis and the rigid-ity of muscles, and vasomotor changes can be producedand removed in the hypnotic state.Most investigators emphasise one or more of four char-acteristics: expectations and the hypnotist-subject inter-action;� suggestibility;acognitivedimensionrelatedtorelaxation and/or � imagery; and � dissociation (Evans2001).
Characteristics
Of the wide range of phenomena associated with hyp-nosis, hypnotic analgesia is obviously the most usefulin treating pain. Clinicians have typically used one ormore of the following approaches: direct suggestion ofpain reduction or insensitivity; suggestions aimed at al-tering the experience of pain; or suggestions directingattention away from pain and its source.Social role theorists have proposed that hypnotic anal-gesia is simply a consequence of compliance with the� demand characteristics of the experimental or clin-ical situation. In other words, subjects respond in theway they expect the hypnotist wishes them to respond.However, hypnotic analgesia shows a moderately strongcorrelation with measured hypnotic susceptibility (0.5),and there is a marked difference in the subjective expe-rience of subjects simulating hypnosis compared withauthentic hypnosis (Hilgard and Hilgard 1975). Manystudies have shown that pain reduction in response tosuggestion can occur without any apparent hypnotic in-duction. Nevertheless, accumulating evidence suggeststhat a hypnotic induction at least facilitates more pro-found analgesia. Price (1999) proposes that the senseof ease, absorption of attention and lack of monitoringand censoring that are characteristic of hypnosis, laysthe foundation for an increased responsiveness to sug-gestion. Modern brain imaging techniques show differ-ences in brain activity between normal waking and hyp-notic states.Furthermore, specificanalgesia suggestionsare accompanied by specific brain changes; e.g. sugges-tions to enhance or reduce pain unpleasantness, with nochange in pain sensation, are accompanied by changesin the anterior cingulate cortex and not in the primarysensory cortex. Similarly, suggestions of sensory reduc-

T
Therapy of Pain, Hypnosis 2465
tion produce parallel changes in subjective ratings andin activity in the primary somatosensory cortex. There isnowsomeevidence thathypnoticanalgesiaalso involvesdescending brain-to-spinal cord inhibitory mechanismsand can inhibit spinal nociceptive reflexes. Our emerg-ing understanding of the nature of hypnotic analgesia isthat it relates to a wide range of cognitive variables, suchas placebo/nocebo, attention, distraction and emotionaltone which modulate pain through changes in neural ac-tivity in many brain structures involved in nociceptionand pain (Large et al. 2003).
Treatment for Acute Pain
Clinicians over many years have reported remarkablesuccess in using hypnosis to manage acute pain. Mes-mer’s explorations of “animal magnetism” seemedoften to precipitate pain in his patients as an indicationof the healing process (Bloch 1980). His followers, suchas John Elliotson (1791–1868), reported surgical opera-tions performed under “mesmeric sleep”. James Esdaile(1808–1859) described 345 operations performed inIndia with mesmerism as the sole anaesthetic. Modernday accounts of major surgery performed under hyp-notic analgesia continue to be collected (Hilgard andHilgard 1975). Rapid induction techniques have beendeveloped for use in managing the acute pain of traumaand the imposed pain of procedures (Barber 1982).Despite its long history and the experience of many clin-icians and patients, good outcome research has beensomewhat sparse. A recent meta-review concluded thathypnosis had been shown to be effective in controllingthe pain of procedures in children with cancer. It isalso effective in reducing the acute pain of burns andchildbirth (Hawkins 2001).
Treatment for Chronic Pain
The persistent and relapsing nature of chronic painpresents a challenge to the therapist to develop strate-gies that will endure beyond the laboratory or consul-tation room. Hypnotic analgesia in the therapy sessionmay offer some respite from pain but not a long-termcure. Clinicians have therefore turned to developingself-management techniques and the teaching of self-hypnosis (Eimer 2002). A typical clinical approach isto work with the patient in exploring hypnotic respon-siveness, emphasizing what is possible rather than whatis not possible. Techniques are explored and developedin collaboration between therapist and patient. Thesemay range from simple � relaxation, to � “specialplace” imagery, to inducing hypnotic analgesia in ahand (� glove anaesthesia) and transferring this to thesite of pain, to using � post-hypnotic suggestion withor without specific cues. The patient’s own sense ofcontrol is enhanced and encouraged.A review of clinical trials of hypnosis for chronic pain,where there has been some attempt at systemisation andcontrol, suggests that hypnosis is effective in a variety of
pain conditions. It has not been shown to be consistentlysuperior to relaxation or other psychological interven-tions, however.Hypnosis proved superior to propranalol in childrenwith migraine, but few other comparisons with drugtreatments have been made. In studies with adults,hypnosis and � autogenic training appear to be equallyefficacious for migraine.Studies in irritable bowel syndrome have shown notonly a reduction in pain and distension, but also changesin rectal sensitivity assessed by balloon manometry.This finding hints at some basic psychophysiologicalchange induced by hypnosis. However, a similar effecton tender points in fibromyalgia was looked for, but notfound. Work adherence was also improved in a hypnosisgroup treated for irritable bowel syndrome, suggestingan important shift in social and occupational function-ing. Large and James (1988) found that patients trainedin self-hypnosis changed their � self-constructs as theygained mastery over their pain. From viewing them-selves as physically ill people, they moved closer to theirconstruction of their ideal self. This and other studiessuggest that one of the major gains that can be madethrough hypnosis is the enhancement of the patient’ssense of control and � self-efficacy. Price (1999) pointsout that unlike placebo suggestion, which implies anexternal authoritative agent, hypnotic suggestion refersto a more innate, internal and self-directed capacity toalter experience.Overall, the outcome research on hypnosis is promising,but there is a difficulty in relating current clinical ap-proaches to the results of randomized controlled trials.Rigorous experimental design favours standardized ap-proachestohypnosis,withtheuseofscriptsandpreparedtape recordings. In contrast, many clinicians use sophis-ticated and individualized techniques that are difficult toreplicate across subjects. (Large et al. 2003) For exam-ple, some anecdotal reports have described using hyp-nosis as a means of returning to a presumed crucial trau-matic event, so-called � age regression. These reportssuggest that resolution of the trauma can lead to a dra-matic resolution of a chronic pain problem (e.g. Gainer1992). It is difficult to evaluate the importance of suchreports in the absence of reports of failures, which areseldom, if ever, published.As noted, � hypnotisability is correlated with hypnoticanalgesia in laboratory studies. A number of studiesusing hypnosis in chronic pain management have foundbetter results in higher hypnotizable subjects. However,it would seem that high hypnotizable subjects are alsomore likely to respond to relaxation techniques in gen-eral. This suggests that there are some commonalitiesin the various forms of psychological interventions thatutilize relaxation. It is possible that highly hypnotisablesubjects begin to access the hypnotic state in response torelaxation training in general. Hilgard’s research in thelaboratory has suggested that the simple induction of

2466 Thermal Allodynia
hypnosis without analgesic suggestions does not inducehypnotic analgesia. Specific analgesic suggestions arerequired. Many relaxation scripts, however, do includesuggestions for comfort and pain reduction, so that thereis considerable crossover between strategies labelled“hypnosis” and those not labelled “hypnosis”!
Conclusion
Hypnosis has waxed and waned in popularity since thedays of Mesmer. Systematic research in cognitive psy-chologyandphysiologicalpsychologyhasimprovedourunderstanding of the nature of hypnotic responding andhypnotic analgesia. The introduction of modern neuro-imaging techniques has begun to validate the constructof hypnosis as a phenomenon that has both subjectiveand objective reality. Clinical research continues to en-courage the development of effective strategies utilisinghypnosis. Hypnosis continues to pose important ques-tions in our understanding of pain, consciousness andcognitive influences on brain processes, as well as chal-lenging us to refine therapeutic strategies utilizing thefull potential of hypnotic analgesia.
References1. Barber J (1982) Managing Acute Pain. In: Barber J, Adrian C
(eds) Psychological Approaches to the Management of Pain.Brunner/Mazel Publishers, New York, pp 168–185
2. Bloch G (1980) Mesmerism. A Translation of the Original Scien-tific and Medical Writings of F.A. Mesmer. William Kaufmann,Inc, Los Altos
3. British Medical Association Report (1955) Medical Use of Hyp-notism. BMJ 1 Supplement 190
4. Eimer BN (2002) Hypnotize Yourself Out of Pain Now! NewHarbinger Publications, Inc, Oakland
5. Evans FJ (2001) Hypnosis in Chronic Pain Management. In:Burrows GD, Stanley RO, Bloom PB (eds) International Hand-book of Clinical Hypnosis. John Wiley and Sons, Ltd. Chichester,pp 247–260
6. Gainer MJ (1992) Hypnotherapy for Reflex Sympathetic Dys-trophy. Am J Clin Hypn 34:227–232
7. Hawkins RMF (2001) A Systemic Meta-Review of Hypnosis asan Empirically Supported Treatment for Pain. Pain Rev 8:47–73
8. Hilgard ER, Hilgard JR (1975) Hypnosis in the Relief of Pain.William Kaufmann, Inc, Los Altos
9. Large RG, James FR (1988) Personalised Evaluation of Self-Hypnosis as a Treatment of Chronic Pain: A Repertory GridAnalysis. Pain 35:155–169
10. Large RG, Price DD, Hawkins R (2003) Hypnotic Analgesiaand its Applications in Pain Management. In: Dostrovsky JO,Carr DB, Koltzenburg M (eds) Proceedings of the 10th WorldCongress on Pain. Progress in Pain Research and Management,vol 24. IASP Press, Seattle, pp 839–851
11. Price DD (1999) Mechanisms of Hypnotic Analgesia. In: PriceDD (ed) Psychological Mechanisms of Pain and Analgesia.Progress in Pain Research and Management vol 15. IASP Press,Seattle, pp 183–204
Thermal Allodynia
Definition
Allodynia evoked by a thermal stimulus.� Cognitive Behavioral Treatment of Pain
� Spinal Cord Injury Pain Model, Contusion InjuryModel
Thermal Effects of Ultrasound
Definition
The thermal effects on the target tissue result in an in-creased local metabolism, circulation, extensibility ofconnective tissue, tissue regeneration and bone growth.� Ultrasound Therapy ofPain from theMusculoskeletal
System
Thermal Hyperalgesia
Definition
Thermal Hyperalgesia is a condition of altered percep-tion of temperature. Describes heightened sensitivity tonoxious heat.� Neuropathic Pain Model, Chronic Constriction Injury� OpioidsintheSpinalCordandModulationofAscend-
ing Pathways (N. gracilis)� TRPV1, Regulation by Nerve Growth Factor� TRPV1, Regulation by Protons
Thermal Hyperalgesia Test
� Thermal Nociception Test
Thermal Neuroablation
� Radiofrequency Neurotomy, ElectrophysiologicalPrinciples
Thermal Nociception TestKENNETH M. HARGREAVES1,CHRISTOPHER M. FLORES2
1Department of Endodontics, University of TexasHealth Science Center at San Antonio, San Antonio,TX, USA2Drug Discovery, Johnson and Johnson PharmaceuticalResearch and Development, Spring House, PA, [email protected]

T
Thermal Nociception Test 2467
Synonyms
Plantar Test; Thermal Hyperalgesia Test; Radiant HeatTest; Hargreaves Test
Definition
The thermal hyperalgesia test permits highly repro-ducible evaluation of paw withdrawal thresholds ofanimals to a beam of radiant heat applied to the plantarsurface of the paw. The endpoint is detected automat-ically thereby removing an important potential sourceof observer bias. The test can be used to measure nor-mal nociceptive thresholds and to quantify hyperalgesiaand allodynia in models of inflammatory or neuropathicpain (Hargreaves et al. 1988).
Characteristics
Overview
Our understanding of the chemical and anatomicalsubstrates that underlie hyperalgesia and allodyniahas benefited tremendously from numerous importantanimal models. Consequently, we have a more pro-found appreciation for the processes that contributeto the development and maintenance of certain be-havioral responses to noxious stimulation and tissueinjury. Nonetheless, such models are only as valuableas are the methods employed to detect and quantitatethese behavioral responses. Thus, validity, reliability,sensitivity, assay reproducibility and flexibility are allcritical determinants of any paradigm by which to assessnociception and its physiological sequelae.Attempts to assess pain quantitatively in humans andanimals dates back to the first half of this century, andthere are numerous reports of “pain” measurements inanimals responding to thermal, chemical, mechanical orelectrical stimuli. However, these methods all sufferedfrom a variety of drawbacks including, but not limitedto, a lack of correlation between human clinical and ex-perimental animal studies, a lack of assay sensitivity topharmacological manipulation, especially with regardto � NSAIDs, Survey (NSAIDS), the inability to per-form within-subject controls and a virtually uniform re-liance on non-complex, reflex behaviors (often involv-ing the uniquely scaly, non-glabrous skin of the rat tail)that are difficult if not impossible to equate with the hu-man sensation of pain.The plantar test avoids many of these limitations sinceit measures complex, nociceptive behaviors follow-ing thermal stimulation of cutaneous tissues, usinga method allowing for not only the determination ofthermal nociceptive thresholds but also the quantitationof � primary hyperalgesia, � secondary hyperalgesiaand � allodynia. The test is performed in awake, freelymoving and unrestrained animals, thereby avoiding thepotential generation of stress responses. In addition,the device is flexible enough to permit the independent
testing of selected, individual receptive fields (e.g.,independent assessment of both hind paws), permittingthe use of within-subject controls, sophisticated enoughto yield a quantifiable, automated end-point, therebyremoving most of the potentially confounding observerinteraction and sensitive enough to detect relativelysmall changes in response to relevant perturbations,such as the induction of inflammation and hyperalgesiaand/or the administration of drugs. In 1988, after twoyears of design, developmentand validation, the plantartest methods paper was published. According to an ISIWeb of Science™ analysis of papers published throughMay 2005, this paper has been cited in more than 1,000publications, making it one of the most heavily citedarticles ever published in the journal Pain (Terajimaand Aneman 2003).
Procedure
The device (for detailed description, see Hargreaves etal. 1988) essentially consists of a raised glass floor be-neath which is placed a movable case containing a ra-diant heat source and photoelectric cell that are aimedthrough an aperture at the top of the case. The methodinvolves placing an animal on top of the glass floor andenclosing it in a small, clear plastic cage. Depending onthe size of the floor and the enclosures employed, severalanimals may be positioned simultaneously in individualcages. Following an acclimation period, at which pointthe animal has come to rest, the aperture is positioneddirectly beneath the plantar surface of the animal’s hindpaw. The trial is commenced by a switch that simultane-ously activates the heat source and an electronic timer.Uponwithdrawalof thepaw, theresulting interruptionofthe reflected light is detected by the photoelectric cell,thereby signaling the lamp and timer to turn off and atone to be emitted. The electronic clock circuit is wiredto a microcomputer and LED readout. The paw with-drawal latency (PWL), measured as the time to the near-est 0.1 sec between points at which the switch is turnedon and the beam of light is interrupted, is taken as a mea-sure of the thermal nociceptive threshold of the animaland is displayed on the readout. The test may then be re-peatedonthesameortheotherpaw.Owingtothefact thatthe procedure is carried out on unrestrained, freely mov-ing animals, this method also allows for the concurrentmeasurement of complex, organized behaviors in addi-tion to the PWL, including the duration and rapidity ofwithdrawal as well as various locomotor activities suchas licking, each a separate measure of hyperalgesia (de-fined as an exaggerated response to a noxious stimulus).To measure duration of paw withdrawal, a stopwatch isstarted when the device emits the tone and is stoppedwhen the paw is returned to the glass floor. Similarly,the duration of licking or other complex behavior canbe measured. Changes in latency correspond to thermalallodynia, since latency is directly correlated with cuta-neous temperature (Fig. 1). In contrast, changes in du-

2468 Thermal Nociception Test
Thermal Nociception Test, Figure 1 Effects of radiant heat on the cu-taneous temperature of rat hind paws. Reproduced with permission from:Hargreaves, et al. Pain 32:77-88, 1988, Elsevierl’.
ration of hind paw elevation following the stimulus mayreflect aspects of hyperalgesia (Fig. 2).
Advantages and Disadvantages
Many of the advantages and limitations of this radiantheatmethodcomparedwithpreviouslyexistingmethodshave been summarized (Table 1).Perhaps of greatest im-portance is that this paradigm conforms to theguidelinesfor the ethical and responsible use of animals as exper-imental subjects established by the International Asso-ciation for the Study of Pain and the Society for Neu-roscience. In this regard, it is of profound importancethat the noxious stimulus applied during testing of theanimals is readily escapable. Thus, the animal itself ter-minates the experimental session after a brief stimulusand before any thermal injury can occur, even after re-peated application of the test. In addition, the capacity toperform the test on an awake, freely moving animal thatappears ostensibly calm and comfortable during the ac-climation and testing period markedly reduces the con-cern that any stress-related neuroendocrine circuits willbe engaged. Another advantage to the freedom of move-ment afforded by this method is that a number of morecomplex behaviors related to an induced hyperalgesicstate and occurring secondary to or as a result of the test-ing can be measured. The ability to quantitate such anarray of nociceptive behaviors gives rise to a “hyperal-gesiaprofile” for agiven animal thatmay becompared toother individuals under a variety of experimental condi-tions. This set-up also readily permits the administrationof test articles or other perturbations, on a repeated basisif necessary, in between PWL measurements.As already alluded to, this method, in addition tomeasuring thermal nociceptive thresholds in normalanimals/hind paws, may also be used to quantitatehyperalgesia, operationally defined as an increased
Thermal Nociception Test, Figure 2 (a) Effects of carrageenan on thelatency for hind paw withdrawal following injection of carrageenan intoone hind paw with measurement of both the ipsilateral injected hind pawand the contralateral control hind paw. (b) Effects of carrageenan on thepost-stimulus duration of hind paw elevation above the glass floor in thesame animals as (a). Reproduced with permission from: Hargreaves et al.Pain 32:77-88, 1988, Elsevierl’.
responsiveness to noxious stimulation. It is also theo-retically possible to alter the device to deliver a sub-threshold stimulus as a means of measuring allodynia.In the context of this discussion then, it may be ap-preciated that this method is amenable to measuringhyperalgesia that develops due to inflammation or nerveinjury induced by a variety of perturbations. Thus, themethod has been applied to several models, includingthe intraplantar injection of carrageenan (Hargreaves etal. 1988), complete Freund’s adjuvant (Iadarola et al.1988), yeast (Iadarola et al. 1988), nerve growth factor(Andreev et al. 1995) or the induction of a peripheral

T
Thermal Nociception Test 2469
Thermal Nociception Test, Table 1 Methodological Papers Evaluating the Radiant Heat Divice
Modification of Parameter Evaluated Authors
Methods paper: thermal hyperalgesia in a model of inflammation Hargreaves et al. 1988
Thermal hyperalgesia in a model of peripheral neuropathy Bennett and Xie 1988
Glass surface acts as a heat sink Hirata et al. 1990
To avoid heat sink effect, only test a paw when it is in contact with the glass surface Bennett and Hargreaves 1990
A nylon mesh over the glass surface reduces the heat sink effect Murphy et al. 1991
Animals restrained during thermal testing Kerns et a1. 1991
Testing thermal thresholds in normal animals Yamamoto and Yaksh 1991
Heated glass surface using feedback control to avoid a heat sink effect Yamamoto and Yaksh 1991
Correlation of paw temperature and escape latencies are consistent with measuring nociception Yeomans et al. 1992
Thermal hyperalgesia in a model of peripheral neuropathy Kim and Chung 1992
Thermal hyperalgesia following i.t. NMDA Malmberg and Yaksh 1993
Heated glass floor, joystick control of radiant heat source, illustration of a circuit diagram Galbraith et al. 1993
Radiant heat device more sensitive than tail flick for detecting thermal hyperalgesia during tolerance Mao et al. 1994
Frogs as test subject Willenbring and Stevens 1995
Radiant heat applied to the dorsum of the hindpaw Tracey et al. 1995
Gerbils as test subject Rupniak et aI. 1995
Evaluated two intensities of radiant heat in the same subjects Abram and O’Connor 1995
neuropathy by nerve constriction (Bennett and Xie1988; Kim and Chung 1992; Bennett 1999).One of the most important advantages of this methodcompared with previous methods is the ability to havewithin-subject controls. This may be represented sta-tistically as left vs right or baseline vs post-perturbationhind paw difference scores. The utility of this transfor-mation of the data is highlighted by the demonstrationof Bennett and Xie that 140 normal rats exhibited differ-ence scores that fit a Gaussian distribution with a meanand standard deviation of 0.04 ± 0.66 sec (i.e. a right-left difference score of approximately zero) (Bennettand Xie 1988). This feature of the paradigm has beenexploited in myriad, within-animal studies evaluatingthe mechanisms of hyperalgesia and inflammation andthe effects of drugs or other therapeutic interventionsused to counteract these processes. Thus, one could de-sign a study to determine whether a given inflammatoryagent results in a localized vs systemic hyperalgesia. Inaddition, one could distinguish between peripheral vscentral mechanisms of action of antinociceptive agents(Jackson et al. 1995), in that PWL measurements can bemade in a given paw following injection of a potentiallyantinociceptive test substance directly into that paw oralternatively into the contralateral paw or some otherremote location, usually at a dose that is not sufficientto reach pharmacologically relevant systemic con-centrations. Another application of this method is the
ability to correlate somatotopically localized biochem-ical, cellular and molecular alterations with behaviorin the affected limb. Thus, several interesting studieswere performed in which carrageenan- or CFA-inducedhyperalgesia was correlated with somatotopically ap-propriate increases in opioid peptide gene expressionin the dorsal spinal cord on the side ipsilateral to theinflammation (Iadarola et al. 1988). In addition, in vivomicrodialysis has been used to measure and correlatethe time course of the local production and/or releaseof inflammatory mediators with nociceptive behaviorin parallel groups of animals (Hargreaves and Costello1990).Ultimately, the true measure of any method rests on itssensitivity, in other words, the ability to detect changesin the dependent measurement when in fact they occur.In this respect, it is significant that in the periphery, thesensitivity to detect hyperalgesia is greater for thermalvs mechanical stimuli in several tests (Handwerker etal. 1987). Consistent with this concept, the radiant heatdevice was shown to be more sensitive than the Ran-dallandSelittomechanical test indetectinghyperalgesiain response to a subcutaneous injection of carrageenan,which was manifested as an increase in signal-to-noiseratios, as indicated by improved ANOVA F ratios and in-creased carrageenan dose-response curve slopes (Harg-reaves et al. 1988). Moreover, the radiant heat device hasbeen used to evaluate several hundred compounds, rep-

2470 Thermal Receptors
resenting at least 20 distinct classes for their ability to al-ter nociception and nociceptive behaviors, highlightingits utility as a tool not only for investigating the neuro-chemistry of pain but also for the development of novelanalgesic therapeutics.The most obvious limitation of the current method isthat it does not measure mechanical nociceptive thresh-olds, allodynia or hyperalgesia. Moreover, the extentto which thermal hypersensitivity, as measured by thismethod, accurately predicts the efficacy of clinicallyuseful drugs (based on type 1 and type 2 errors) orreliably mimics one or more than one clinically rele-vant pain condition/symptom remains to be determined(although the test reliably demonstrates the efficacy of avariety of currently marketed NSAID and opioid anal-gesics with reasonably appropriate rank order potency).Notwithstanding these caveats, the combination of thethermal method with mechanical tests, such as the useof von Frey filaments, offers a powerful approach tothe study of a variety of nociception-modifying pertur-bations as well as a means for discriminating potentialsensory modality differences between them. The othermajor disadvantage of the radiant heat method is that theglass surface may act as a heat sink, leading to artifac-tually short paw withdrawal latencies and a subsequentincrease in assay variability (Bennett and Hargreaves1990; Hirata et al. 1990). This factor will vary with thespecific conductive properties of the floor, the ambienttemperature in the experimental procedure room andthe pressure exerted by the rat on the paw being tested,which last will be affected under conditions of inducedinflammation or neuropathy (Bennett and Hargreaves1990; Murphy et al. 1991). However, several sugges-tions have been made to counteract this effect, the mostimportant of these being that the skin being irradiatedshould be in contact with the glass floor (Table 1). Mod-ifications to the device itself include placing a nylonmesh screen between the rat and the glass (Murphy etal. 1991) or heating the floor to maintain it at a constanttemperature, either by implanting a heating elementwithin the glass or applying warm air to its underside(Galbraith et al. 1993). The minor inconveniences cre-ated by the rats urinating or defecating on the glass floorare easily remedied with a spray bottle of cleanser anda roll of paper towels.In conclusion, the radiant heat test described hereoffers a simple, highly reproducible and useful ap-proach to the study of thermal nociception in naïveand injured/inflamed rodents. Accordingly, it may beconsidered one of several powerful behavioral toolsavailable to pain scientists in the investigation of noci-ceptive mechanisms and in thesearch forbetter and saferanalgesic drugs. It is hoped that further improvementsin this method, including those mentioned herein, andits use in combination with complementary behavioralassays will lead to a better understanding of the neu-
rophysiolgical underpinnings of pain in humans andultimately better treatment options.
References1. Andreev NY, Dimitrieva N, Koltzenburg M et al. (1995) Periph-
eral administration of nerve growth-factor in the adult-rat pro-duces a thermal hyperalgesia that requires the presence of sym-pathetic postganglionic neurons. Pain 63:109–115
2. Bennett GJ (1999) Does a neuroimmune interaction contribute tothe genesis of painful peripheral neuropathies? Proc Natl AcadSci USA 96:7737–7738
3. Bennett GJ, Hargreaves KM (1990) A model of peripheralmononeuropathy in the rat – reply. Pain 42:255–255
4. Bennett GJ, Xie Y-K (1988) A peripheral mononeuropathy inrat that produces disorders of pain sensation like those seen inman. Pain 33:87–107
5. Galbraith JA, Mrosko BJ, Myers RR (1993) A system to measurethermal nociception. J Neurosci Methods 49:63–68
6. Handwerker HO, Anton F, Kocher L et al. (1987) Nociceptorfunctions in intact skin and in neurogenic or non-neurogenic in-flammation. Acta Physiol Hung 69:333–342
7. Hargreaves KM, Costello A (1990) Glucocorticoids suppress lev-els of immunoreactive bradykinin in inflamed tissue as evaluatedby microdialysis probes. Clin Pharmacol Ther 48:168–178
8. Hargreaves KM, Dubner R, Brown F et al. (1988) A new andsensitive method for measuring thermal nociception in cutaneoushyperalgesia. Pain 32:77–88
9. Hirata H, Pataky A, Kajander K et al. (1990) A model of pe-ripheral mononeuropathy in the rat. Pain 42:253–254
10. Iadarola MJ, Brady LS, Draisci G et al. (1988) Enhancement ofdynorphin gene-expression in spinal-cord following experimen-tal inflammation – stimulus specificity, behavioral parametersand opioid receptor-binding. Pain 35:313–326
11. Jackson DL, Graff CB, Richardson JD et al. (1995) Glutamateparticipates in the peripheral modulation of thermal hyperalgesiain rats. Eur J Pharmacol 284:321–325
12. Kim SH, Chung JM (1992) An experimental-model for peripheralneuropathy produced by segmental spinal nerve ligation in therat. Pain 50:355–363
13. Murphy LG, Alexander GM, Schwartzman RJ (1991) Improve-ment to the Hargreaves paw flick method. Pain 46:347–347
14. Terajima K, Aneman A (2003) Citation classics in anaesthesiaand pain journals: a literature review in the era of the internet.Acta Anaesth Scand 47:655–663
Thermal Receptors
Definition
Sensory receptors sensitive to thermal stimuli.� Hyperaesthesia, Assessment� Hypoesthesia, Assessment
Thermal Sensory Testing
� Quantitative Thermal Sensory Testing of InflamedSkin

T
Theta Rhythm 2471
Thermal Stimulation (Skin, Muscle,Viscera)
� Pain in Humans, Thermal Stimulation (Skin, Muscle,Viscera), Laser, Peltier, Cold (Cold Pressure), Radi-ant, Contact
Thermal Stimulation (Stimuli)
Definition
Thermal stimulation (warm and cold stimuli) consistsof transferring (adding or subtracting) calorific energybetween the skin (or mucosa, muscle, viscera) and itssurroundings.� Causalgia, Assessment� Dysesthesia, Assessment� Pain in Humans, Thermal Stimulation (Skin, Muscle,
Viscera), Laser, Peltier, Cold (Cold Pressure), Radi-ant, Contact
Thermal Therapy
� Spa Treatment
Thermal Transduction
� Polymodal Nociceptors, Heat Transduction
Thermocoagulation
Definition
Destroying tissue (e.g. nerves) by heating.� Cancer Pain Management, Anesthesiologic Interven-
tions, Neural Blockade� Radiofrequency Neurotomy, Electrophysiological
Principles
Thermode
Definition
A thermode is a device used to apply controlled temper-ature to the skin of subjects or animals. One surface ofthe thermode is placed in contact with the skin. The tem-perature at that surface is controlled with the use of heat-ing elements, Peltier elements, and/or circulating water.
The surface temperature is measured with a thermocou-ple or thermister, which is used in a feedback manner toregulate the thermode’s surface temperature.� Threshold Determination Protocols
Thermography
Definition
Detects and delineates areas of cutaneous thermalchange.� Postoperative Pain, Acute Presentation of Complex
Regional Pain Syndrome
Thermoreception
� Lateral Thalamic Pain-Related Cells in Humans
Thermoreceptor
Definition
Cutaneous, oral and visceral receptors responding onlyor preferentially to temperature within the innocuous ornonpainful range. Thermoreceptors show static and dy-namic sensitivity to temperature.� Nociceptors, Cold Thermotransduction
Thermoregulation
Definition
Rodents regulatebody temperaturemainlybyregulatingblood flow in the tail by an on-off mechanism, suddenlychanging the tail skin temperature by up to 8–10˚C.� Tail-Flick Test
Theta Rhythm
Definition
Frequency domain of oscillatory hemispheric activitybetween 4 and 8 Hz. It has been associated with differentfunctional brain states, e.g. somnolence, cognitive acti-vations, altered states of consciousness like meditation,and, relevant here, dysfunctional brain states like neuro-genic pain and tinnitus, abnormal movements, epilepsyand neuropsychiatric disorders (see thalamocorticaldysrhythmia).� Thalamotomy for Human Pain Relief

2472 Thoracic Epidural Analgesia
Thoracic Epidural Analgesia
Definition
Pain relief obtained by drugs acting directly on thespinal cord, e.g. morphine, local anesthetics, adrenaline(epinephrine), clonidine, and neostigmine.� Postoperative Pain, Acute Pain Management, Princi-
ples� Postoperative Pain, Acute Pain Team
Thoracic Epidural Anesthesia
Definition
For thoracic epidural anesthesia the catheters are in-serted dependent on the surgical procedure, betweenlevel T4–T12 by a median or paramedian approach andusually with the loss of resistance technique. Postop-eratively, the patient has perfect analgesia and can bemobilized.� Postoperative Pain, Thoracic and Cardiac Surgery
Thoracic Medial Branch Blocks andIntra-Articular Blocks
PAUL VERRILLS
Metropolitan Spinal Clinic, Prahran, VIC, [email protected]
Synonyms
Intra-Articular Blocks and Thoracic Medial BranchBlocks
Definition
Thoracicmedialbranchblocksandintra-articularblocksare both diagnostic procedures designed to test if the pa-tient’s pain arises from a thoracic zygapophysial joint.They involve anaesthetising the joint or its nerve supplywith injections of small volumes of local anaesthetic.
Characteristics
The thoraciczygapophysial jointsmay beacauseofpos-terior thoracic spinal pain. Studies in normal volunteers(Dreyfuss et al. 1994), and in patients (Fukuiet al. 1997),have shown that noxious stimulation of these joints pro-duces local and � somatic referred pain in a segmentalpattern along the posterior chest wall (Fig. 1).There are no means clinically, or by medical imaging,by which pain from these joints can be diagnosed. The
only means of determining if they are a source of painis to anaesthetise putatively symptomatic joints.The thoracic zygapophysial joints can be anaesthetisedeither by injecting local anaesthetic into the joint cav-ity, or by anaesthetising the medial branches of the dor-sal rami that innervate them. Respectively, these proce-dures are called thoracic intra-articular zygapophysialjoint blocks and thoracic medial branch blocks.
Intra-Articular Block
The thoracic zygapophysial joints are not directly evi-dent on anteroposterior views of the thoracic spine, be-cause their jointcavitiesareorientated inacoronalplane.Nevertheless, the location of each joint can be inferred.The joint lies opposite the inter-vertebral disc of the seg-ment, between the pedicles of the two vertebrae thatcon-tribute to the joint.Once the target jointhasbeen identified, apuncturepointon the skin is selected, about three-quarters of a seg-ment caudal to the joint, on a caudal extension of thesagittal bisector of the pedicle below the target joint. Aspinal needle is introduced at around 60 degrees caudalto the perpendicular, and is advanced towards the lowerquarter of the silhouette of the pedicle below the joint(Dreyfuss et al. 1994; International Spinal InterventionSociety 2004a). A contra-lateral oblique view can thenbe used to carefully advance the needle into the inferiormargin of the joint (Fig. 2a). Once the needle has en-tered the joint, an anteroposterior view is used in orderto check that the needle has not strayed medially or lat-erally (Fig. 2b).Intra-articular placement is verified by injecting 0.1 mlof contrast medium, in order to obtain an arthrogram(Dreyfuss et al. 1994; International Spinal InterventionSociety 2004a) (Fig. 3). Subsequently, the joint is anaes-thetised by injecting 0.75 ml of local anaesthetic.
Medial Branch Block
The thoracic medial branches cross the thoracic trans-verse processes obliquely, passing from the region of thesuperior lateral corner of the process to its infero-medialcorner. At segmental levels T1–4 and T9 and T10, thenerves lie on the posterior surface of the transverse pro-cess, slightly belowand justmedial to thesuperior lateralcorner of the transverse process. This point constitutesthe target point for medial branch blocks at these levels(Chua and Bogduk 1995; International Spinal Interven-tion Society 2004b).Under postero-anterior fluoroscopic screening, a spinalneedle is inserted through the skin of the posterior tho-rax, and passed towards the target point of the nerveto be blocked (Fig. 4). Once the needle has reachedbone, a test-dose of contrast medium is injected un-der continuous fluoroscopic screening to ensure thatthere is no vascular uptake, and to determine that theinjectate spreads appropriately (International SpinalIntervention Society 2004b). To block the nerve, 0.3 ml

T
Thoracic Medial Branch Blocks and Intra-Articular Blocks 2473
Thoracic Medial Branch Blocks andIntra-Articular Blocks, Figure 1 Thedistribution of referred pain from the thoraciczygapohysial joints. Based on Dreyfuss etal. 1994 and Fukui et al. 1997.
of local anaesthetic is injected. To anaesthetise a givenzygapophysial joint, both the nerves that innervate itare blocked.At the T4 to T8 levels, the medial branches do not run onbone.Fromtheintertransversespace, theyaresuspendeddorsal to the depth of the transverse process (Chua andBogduk 1995). In order to block the nerve, a needle isdelivered into the intertransverse space, slightly cranialto the tip of the transverse process (International SpinalInterventionSociety2004b).Inthis location,asmallvol-ume of contrast medium is injected, in order to confirmappropriate spread of injectate. Thereupon, local anaes-thetic can be injected to anaesthetise the nerve.The T11 and T12 medial branches assume a course sim-ilar to that of the lumbar medial branches. The targetpoints and technique for these nerves are like those for� lumbar medial branch blocks.
Evaluation
The optimal means of reducing error and securing reli-able diagnostic information is real time assessment. Theresponse to the diagnostic block is evaluated immedi-ately after the block, and for some time afterwards, atthe clinic at which the block was performed, and by anindependent observer using validated and objective in-struments or tools (Dreyfuss et al. 1994; InternationalSpinal Intervention Society 2004b).Visual analogue scores are recorded before the block,immediately afterwards, at 30 minutes and then hourly.The patient is instructed to monitor the extent and du-ration of any relief that ensues. Further, if relief occurs,the patient should carefully attempt movements and ac-tivities that are usually restricted by pain to assess theirresponse during the anaesthetic phase.

2474 Thoracic Medial Branch Blocks and Intra-Articular Blocks
Thoracic Medial Branch Blocks and Intra-Articular Blocks, Figure 2 Radiographs of a needle in position for an intra-articular injection of a thoraciczygapophysial joint. (a) Lateral view. (b) AP view. Reproduced courtesy of the International Spinal Intervention Society (2004a).
If the response to the block is negative, then zy-gapophysial joint pain can effectively be ruled outat the level tested. Adjacent levels may then be blockedbased on clinical indication.If the response to the block is positive, then a controlblock is undertaken. If the patient has a concordant re-sponse to controlled blocks, then the putative diagnosisof zygapophysial joint pain is confirmed.
Validity
Single diagnostic blocks of thoracic medial branchescarry a false-positive rate of 58% (Manchikanti etal. 2002). Control blocks are, therefore, mandatory(International Spinal Intervention Society 2004b;Manchikanti et al. 2002).Although placebo controlled blocks could be used,they require three injections, and are not readily imple-mented in clinical practice. Comparative local anaes-thetic blocks, using lignocaine 2%, as a short-actingagent, and bupivacaine 0.5% as a long-acting agent, area valid alternative (International Spinal InterventionSociety 2004b; Manchikanti et al. 2002; Barnsley etal. 1993; Lord et al. 1995).A concordant response wouldbe complete relief of pain for a shorter duration fol-lowing the short-acting agent, and a longer duration ofrelief following the long-acting agent (see � Peripheral
Nerve Blocks). Complete relief of pain on each of thetwo occasions, regardless of duration of relief, increasesthe positive yield of blocks, but increases the risk offalse-positive responses.Similar controls have not been evaluated for intra-articular blocks. Their validity has not been established.
Application
Only one study has described the application of thoracicmedial branch blocks in clinical practice (Manchikantiet al. 2002). It reported that 48% of patients had theirpain relieved with the medial branch blocks. The studythereby established that thoracic zygapophysial jointblocks have a considerable diagnostic utility in clinicalpractice. However, it did not indicate which segmentallevels were most commonly the source of pain.By analogy, with lumbar and cervical spine, one wouldexpect the therapeutic utility of thoracic medial branchblocks would be to select patients for medial branch� radiofrequency neurotomy. However, there are nopublications to describe a valid technique, or to es-tablish the efficacy of such treatment in the thoracicspine. Therefore, the therapeutic utility of thoraciczygapophysial joint blocks is only potential.Although intra-articular blocks constitute an alternativemeans of diagnosing thoracic zygapophysial joint pain,

T
Thoracic Medial Branch Blocks and Intra-Articular Blocks 2475
Thoracic Medial Branch Blocks and Intra-Articular Blocks, Figure 3A lateral radiograph of an arthrogram of a thoracic zygapophysial joint. Thearrows point to contrast medium inside the joint. Reproduced courtesy ofthe International Spinal Intervention Society (2004a).
Thoracic Medial Branch Blocks and Intra-Articular Blocks, Figure 4An AP radiograph showing a needle in place for a thoracic medial branchblock, and an injection of contrast medium to show spread of injectateover the superior lateral corner of the transverse process. The inferiorborder of the transverse process (TP) is labelled. Reproduced courtesy ofthe International Spinal Intervention Society 2004a.
no studies have described any results from their use.Some operators administer intra-articular injections ofcorticosteroidsasa treatmentfor thoraciczygapophysialjoint pain, but no studies have reported the efficacy ofthis treatment.
Patient Selection
Thoracic zygapophysial joint blocks are not indicatedin acute pain. They are relevant only for patients withpersistent thoracic spinal pain, in whom a diagnosis isrequired. A prerequisite is that serious possible causesof pain such as infection, tumours, vascular disease andmetabolic disease have been excluded by careful andthorough history and examination, laboratory tests, andmedical imaging if necessary. The working diagnosisshould be that of thoracic spinal pain of unknown origin.The minimum criterion should be that the patient haspain in a distribution that resembles that shown to em-anate from thoracic zygapophysial joint in normal vol-unteers (Dreyfuss et al. 1994; Fukui et al. 1997).
Contraindications
Absolute contra-indications include bacterial infection,either systemic or localised in the region that blocks areto be performed, bleeding diatheses or possible preg-nancy. Relative contraindications include allergy to con-trast media or local anaesthetics.
References1. Barnsley L, Lord S, Bogduk N (1993) Comparative Local Anaes-
thetic Blocks in the Diagnosis of Cervical Zygapophysial JointPain. Pain 55:99–106
2. Chua WH, Bogduk N (1995) The Surgical Anatomy of ThoracicFacet Denervation. Acta Neurochir 136:140–144
3. Dreyfuss P, Tibiletti C, Dreyer SJ (1994) Thoracic Zygapophy-seal Joint Pain Patterns. A Study in Normal Volunteers.Spine 19:807–811
4. Dreyfuss P, Tibiletti, C, Dreyer S, Sobel J (1994) ThoracicZygapophysial Joint Pain: A Review and Description of anIntra–Articular Block Technique. Pain Digest 4:46–64
5. Fukui S, Ohseto K, Shiotani M (1997) Patterns of Pain Induced byDistending the Thoracic Zygapophyseal Joints. Regional Anes-thesia 22:332–336
6. International Spinal Intervention Society (2004a). Thoracic In-tra–Articular Zygapophysial Joint Blocks. In: Bogduk N (ed).Practice Guidelines for Spinal Diagnostic and Treatment Proce-dures. International Spinal Intervention Society, San Francisco(in press)
7. International Spinal Intervention Society (2004b). ThoracicMedial Branch Blocks. In: Bogduk N (ed) Practice Guidelinesfor Spinal Diagnostic and Treatment Procedures. InternationalSpinal Intervention Society, San Francisco (in press)
8. Lord SM, Barnsley L, Bogduk N (1995) The Utility of Compara-tive Local Anaesthetic Blocks versus Placebo–Controlled Blocksfor the Diagnosis of Cervical Zygapophysial Joint Pain. Clin JPain 11:208–213
9. Manchikanti L, Singh V, Pampati V, Beyer CD, Damron KS(2002) Evaluation of the Prevalence of Facet Joint Pain in ChronicThoracic Pain. Pain Physician 5:354–359

2476 Thoracic Outlet Syndrome
Thoracic Outlet SyndromeA. LEE DELLON1, 2
1Plastic Surgery and Neurosurgery, Johns HopkinsUniversity, Baltimore, MD, USA2Plastic Surgery, University of Arizona, Tucson, AZ,[email protected]
Synonyms
Brachial Plexus Compression; Scalenus Anticus Syn-drome; Costoclavicular Syndrome
Definition
Thoracic Outlet Syndrome is a misnomer for the symp-toms caused by compression of the brachial plexus inthe thoracic inlet. The symptoms have a wide range thatcan encompass any manifestation related to the motoror sensory functions of the cervical nerve roots C5, C6,C7, C8, and T1. There can be secondary symptoms re-lated to the cervical plexus and shoulder dysfunction.The most common symptoms are aching in the shoul-der, numbness in the fingers, with the symptoms beingaggravated or initiated by elevating the arm above theshoulder level. The syndrome most commonly occursin the setting of neck or shoulder trauma, and is oftenrelated to the presence of anatomical congenital anoma-lies. Due to the inability of traditional electrodiagnostictesting to identify this syndrome, except for the isolatedproblem with the lower trunk of the brachial plexus, di-agnosis and treatment for brachial plexus compressionin the thoracic inlet remains a controversial source ofneck, shoulder and upper extremity pain.
Characteristics
The subclavian artery and subclavian vein exit the tho-rax and enter the upper extremity by a path that takesthem into the supraclavicular region, and then betweenthe clavicle and the first rib and into the axilla. The an-terior scalene muscle is usually located between thesetwo large blood vessels as it inserts into the first rib.Post-traumatic tightness or anatomic anomalies in thisregion can cause vascular symptoms such as purplishcolor and swelling due to venous obstruction, or cold-ness and digital necrosis due to arterial obstruction andemboli. These vascular forms of the “thoracic outletsyndrome” are responsible for about 5% of the patientswith symptoms in this region, and the diagnosis is madeby radiologic imaging of these vessels with the arm atrest and the arm elevated. These vessels actually leavethe thorax, and pass across the thoracic inlet to reach theaxilla (Fig. 1). Relief of obstruction is surgical, requir-ing resection of the anterior scalene muscle or excisionof the first rib. Different anatomic approaches, supra-clavicular or transaxillary, are currently in use. Thesevascular syndromes do not have neurologic symptoms,
Thoracic Outlet Syndrome, Figure 1 Brachial Plexus Neurolysis.
but can be associated with compression of the brachialplexus in this same region (Mackinnon and Dellon1988; Leffert 1992). The classic physical examinationmaneuvers related to the disappearance of the radialpulse when the hand is elevated or the head is turned,are best related to the arterial form of compression inthis region, but can be present in 33% of the normalpopulation.When the lower trunk of the brachial plexus, formedfrom the C8 nerve root and T1 nerve root, are com-pressed against the posterior border of the first rib bya tumor (Pancoast Syndrome, upper pulmonary lobebronchogenic carcinoma, or benign neural tumor likeSchwannoma or neurofibroma), trauma, or congenitalanomaly, the patient experiences symptoms of coldnessin the arm, apart from subclavian artery compression(because the sympathetic input from the stellate gan-glion enters the lower trunk). They also experiencenumbness of the little and ring finger, as well as theupper inner arm, and intrinsic muscle weakness, be-cause these are the skin territories innervated by thesenerve roots, and all the intrinsic muscles of the handare innervated by these nerve roots. Note that the com-pression site is at the thoracic inlet (the thoracic outletis the diaphragm). Therefore, this is the one form of

T
Thoracic Outlet Syndrome 2477
thoracic outlet syndrome that can be identified by elec-trodiagnostic testing (EDT) (Gilliatt et al. 1970). TheEDT will demonstrate abnormal electromyographyof both median and ulnar nerve innervated intrinsicmuscles (typically the abductor digiti minimi, the firstdorsal interosseous, and the opponens pollicis and theabductor pollicis brevis), with a decreased sensoryamplitude for the little finger while the sensory am-plitude remains normal for the index finger. This “trueneurogenic” thoracic outlet syndrome is rare, account-ing for less than 1% of patients with thoracic outletsyndrome.The vast majority of patients (94%) havecomplaints thatare often global in nature, which has led some to doubttheexistenceof thissyndrome(CheringtonandChering-ton 1992; Roos and Wilbourn 1990). Headaches, shoul-der pain, diffuse weakness in theupperextremity,numb-ness in the little and ring finger, but also in all fingers,and these symptoms being made worse by any activitythat requires elevation of the shoulder. In some womenthere is breast pain. In some patients there is upper backor scapularpain. In somepatients there ispain in the tem-poromandibular joint, requiring evaluation by a Dentistor Oral Surgeon. In some there is neck pain. It is crit-ical to appreciate that the brachial plexus does take acourse across the thoracic inlet to enter the axilla be-tween the clavicle (which is concave at this location) andthe second rib; it does not travel between the clavicleand the first rib. The upper trunk of the brachial plexusdoes travel beneath the anterior scalene muscle. Thereis a spectrum of congenital anomalies that course acrossor through or under the brachial plexus that are respon-sible for the symptoms of the nerve compression, anddepending upon the location of the compression sites,the symptoms will vary. These include:
• Cervical rib• Fibrous Bands from C7 transverse process• Extra Origins for Scalene Muscles• Pre- or Post-fixed Brachial Plexus• Intra-Plexus anomalous connections• Elevated position of Subclavian Artery• Muscle of Albinus (scalenus minimus)• Fibrous edges of Scalene Muscles• Anomalous vessels crossing Plexus• Sibson’s Fascia crossing T1 nerve root• Proximal junction of T1 to C8
Secondary compression of the cervical plexus by atight or spasmodic anterior scalene muscle is the causeof the facial pain, and this muscle pulling upon theocciput is the source of the headache (Mackinnon andDellon 1988). A winging component to the scapula isdue to compression of the long thoracic nerve as it exitsbetween the medial and posterior scalene muscles, andthis can be corrected by including a neurolysis of thisbranch of the brachial plexus in the surgical approach(Disa et al. 2001).
Diagnosisofbrachialplexuscompression in the thoracicinlet has been a challenge. Traditional EDT cannot iden-tify this compression because the site is too close to thespinal nerve roots, because the variation in thickness ofthe chest wall invalidates amplitude measurements, andbecause the inability to measure distance variable inval-idates nerve conduction measurements. The H-wave isnot reliablebecauseas the impulse travelsfromthefingerto the spinal cord and back to the hand, multiple poten-tial sitesof entrapmentareencountered at thewrist, fore-arm and elbow. (Wilbourn and Urschel 1984) Since thesymptoms are produced or aggravated when the handsare elevated above the head, provocation of the plexus insuchamannerhasbeenusedtoidentifypatientswith tho-racic outlet syndrome, a positive Roos sign (Roos 1966).Palpation over the brachial plexus in the thoracic inletwill produce a distally radiating sign when the plexus iscompressed beneath anatomicstructures in this location,positive Tinel sign or pressureprovocative test (Novak etal. 1995). Based upon the assumption that measurementof thecutaneouspressure threshold wouldchangefor theindex finger (representing the upper trunk) and the littlefinger (representing the lower trunk), when the thresh-old was compared between the at rest and the provokedmeasurement, the Pressure-Specified Sensory Device™
was introduced for diagnosis (Lee et al. 2000). This ap-proach was refined to include measurements of one andtwo-point static and moving-touch, plus pinch and gripstrength (Howard et al. 2003). A sensitivity of 100% anda specificity of 88% were obtained in the diagnosis ofsevere brachial plexus compression using this approach.Treatment of brachial plexus compression should in-clude stretching of the anterior scalene and strength-ening of the upper trapezius, rhomboids, and serratusanterior muscles (Novak et al. 1995). This approachcan relieve symptoms in 90% of patients. In those pa-tients who fail to respond to non-operative methods,an exhaustive diagnostic approach must be undertakento rule out causes that may give similar pain, prior toundertaking a surgical decompression of the plexus.Additional diagnostic testing should include chestX-ray, cervical and shoulder MRI, and evaluation ofperipheral nerve entrapments such as carpal and cubitaltunnel syndrome, since these entrapments, cervicaldisc disease and intrinsic shoulder pathology are thecommonest causes of these types of pain (Campbell etal. 1991; Levin and Dellon 1992).The surgical treatment of brachial plexus compressionmust accomplish a decompression of the brachial plexusfrom whichever structure or structures are causing thecompression. Historically, just the scalenus anteriorwas resected (Mackinnon and Dellon 1988). In 1966,the transaxillary first rib resection was introduced, withthe concept that the plexus was compressed between thefirst rib and the clavicle (Roos 1966). This is a difficultapproach, requiring wide retraction from the axilla inorder to reach the posterior border of the first rib. Com-

2478 Thoracic Surgery
plications include burning pain in the distribution of the2nd or3rd intercostobrachialnervedue to stretch/tractioninjury, and injury to the subclavian artery, pneumoth-orax, and injury to the C8 or T1 nerve roots, as theycross the posterior border of the first rib. Due to per-sistent upper trunk symptoms following transaxillaryfirst rib resection, an anterior scalene resection oftenhad to be added as a secondary procedure. This gaverise to the supra-clavicular surgical approach throughwhich the anterior (and medial) scalene muscles couldbe resected, a neurolysis of the brachial plexus couldbe done under direct vision, the major vessels could beidentified and protected, and a cervical or first rib couldstill be resected if necessary (Dellon 1993; Hempel et al.1996; Sanders 1991). The results reported by Hempelet al in 1996 are worth noting: During a 28-year period,637 patients underwent 770 supraclavicular first ribresections and scalenectomies for thoracic outlet syn-drome. Following surgery an excellent response wasachieved in 59%, good in 27%, fair in 13%, and poorin 1%. One lymphatic leakage and no brachial plexusinjuries resulted. Postoperative causalgia requiring sub-sequent sympathectomy developed in two cases. Novascular or permanent phrenic nerve injuries occurred,and 12 patients (2%) required operative interventionfor recurrence. Sanders, in 1991, reported no differencein the outcome whether he resected the first rib, ordid not resect it. The author’s preferred approach is asupraclavicular anterior scalenectomy and neurolysisof the brachial plexus, preserving the first rib. Duringfifteen years with this approach, there has not been onepneumothorax, or injury to the brachial plexus or majorvessels. There have been two patients with transientphrenic nerve palsy.
References1. Campbell JN, Naff N, Dellon AL (1991) Thoracic Outlet Syn-
drome: A Neurosurgical Perspective. Neurosurg Clin N Am2:227–234
2. Cherington M, Cherington C (1992) Thoracic Outlet Syn-drome Reimbursement Patterns and Patient Profiles. Neurology42:492–495
3. Dellon AL (1993) The Results of Supraclavicular Brachial PlexusNeurolysis (without First Rib Resection) in Management of Post-Traumatic “Thoracic Outlet Syndrome”. J Reconstr Microsurg9:11–17
4. Disa J, Wang B, Dellon AL (2001) Correction of Scapular Wing-ing by Neurolysis of the Long Thoracic Nerve. J Reconstr Mi-crosurg 17:79–84
5. Gilliatt RW, Le Quesne PM, Logue V et al. (1970) Wasting of theHandAssociatedwithaCervicalRib orBand. JNeurol NeurosurgPsychiatry 33:615–619
6. Hempel GK, Shutze WP, Anderson JF et al. (1996) 770 Consec-utive Supraclavicular First Rib Resections for Thoracic OutletSyndrome. Ann Vasc Surg 10:456–463
7. Howard M, Lee C, Dellon AL (2003) Documentation of BrachialPlexus Compression in the Thoracic Inlet Utilizing Provocationwith Neurosensory and Motor Testing. J Reconstr Microsurg19:303–312
8. Lee GW, Massry DR, Kupfer DM et al. (2000) Documentationof Brachial Plexus Compression in the Thoracic Inlet with Quan-titative Sensory Testing. J Reconstr Microsurg 16:15–20
9. Leffert RD (1992) Thoracic Outlet Syndromes. Hand Clinics8:285–291
10. Levin LS, Dellon AL (1992) Pathology of the Shoulder as itRelates to the Differential Diagnosis of Thoracic Outlet Com-pression. J Reconstr Microsurg 8:313–317
11. Mackinnon SE, Dellon AL (1988) Surgery of the PeripheralNerve, 1st edn. Thieme, New York, pp 175–191
12. Novak CB, Collins ED, Mackinnon S (1995) Outcome followingConservative Management of Thoracic Outlet Syndrome. J HandSurg 20A:542–548
13. Roos D (1966) Transaxillary Approach to the First Rib to RelieveThoracic Outlet Syndrome. Ann Surg 163:354–358
14. Roos D, Wilbourn AJ (1990) Thoracic Outlet Syndrome is Un-derrated/Overdiagnosed. Arch Neurol 47:228–230
15. Sanders RJ (1991) Thoracic Outlet Syndrome: A Common Se-quelae of Neck Injuries. JB Lippincott Co, Philadelphia
16. Wilbourn A, Urschel HC (1984) Evidence for Conduction Delayin Thoracic Outlet Syndrome is Challenged. New Eng J Med310:1052–1053
Thoracic Surgery
Definition
Thoracic surgery includes all thoracic surgical proce-dures performed with lateral thoracotomy or mediansternotomy.� Postoperative Pain, Thoracic and Cardiac Surgery
Thought Suppression
Definition
The intentional act of excluding certain thoughts fromconsciousness.� Psychology of Pain, Assessment of Cognitive Vari-
ables
Threat Appraisal
Definition
The judgment that a stressful event, such as pain, isthreatening.� Psychological Aspects of Pain in Women
Three-Way Scaling Models
Definition
Three-way scaling models provide, in addition to thegroup stimulus space, a subject weight space that pro-vides coordinates for the dimensions most salient toeach individual.� Multidimensional Scaling and Cluster Analysis Ap-
plication for Assessment of Pain

T
Threshold Determination Protocols 2479
Threshold
Definition
The threshold is the endpoint for the appearance of agiven reaction or behavior.� Randall-Selitto Paw Pressure Test� Statistical Decision Theory Application in Pain As-
sessment
Threshold Determination ProtocolsJOEL D. GREENSPAN
Department of Biomedical Sciences, University ofMaryland Dental School, Baltimore, MD, [email protected]
Synonyms
Quantitative Sensory Testing (QST). Although notstrictly synonymous, QST refers to a particular set ofthreshold determination protocols used for both painand innocuous sensory evaluation.
Definition
Threshold determination protocols are those psy-chophysical procedures used to estimate perceptualthreshold. These protocols (of which there are several)are defined by the rules by which stimuli are applied, theways that responses are measured, and how the entiresequence of events is orchestrated.
Characteristics
There are several protocols in the literature that estimatepain threshold. To a large extent, these protocols are thesame as those used to estimate the threshold of other sen-sory systems. These protocols have been developed andmodified over the last 150 years, resulting in many vari-ations (Boring 1942).One important distinction is between those protocolsthat depend upon the subject’s reaction time (RT), andthose that do not (Yarnitsky 1997). The most commonlyused RT-dependent protocol is referred to as the Methodof Limits. Within this protocol, the stimulus intensity(i.e. temperature with a thermal probe or force witha pressure � algometer) is gradually increased untilthe point at which the subject indicates he feels pain,typically by pushing a button. The stimulus level at thetime of that report is taken as the pain threshold (Fig. 1,top). One can also use this same approach and have thesubject indicate the limits of his tolerance.A major reason for its wide use is the simplicity ofthis protocol, particularly in the clinical environ-ment. However, it has some distinct disadvantages,including the obvious confound of a motor response
embedded in the measure. One must consider, for in-stance, in comparing two groups of people, whethera group difference in motor reaction time could belarge enough to produce different thresholds withthis measure. Alternatively, and perhaps more likely,is whether slower decision making in one group vs.another (i.e. aged or patient groups) could accountfor slower responses, thus leading to higher thresh-olds.One of the common variants of this protocol uses a con-tact � thermode, and runs a series of trials to estimatethe thresholdfor innocuousthermalsensation(warmandcool), and then cold pain and heat pain. In the exampleprovided in Figure 2 (Greenspan 2001), the thermodewas placed on the test site (right foot) at an adapting tem-perature of 32˚C. Then, without warning to the subject,the temperature was decreased (cool or cold pain trials)or increased (warm or heat pain trials) at a fixed rate oftemperature change (1.0˚C/s). The subject depressed abutton when she felt the particular sensation for whichshe was instructed to attend. These trials are repeatedtypically 3–4 times for each sensory modality, and anaverage value is calculated for threshold.A variant of this protocol was referred to as the� Marstock method, and used oscillating tempera-tures between warm and cool or heat and cold painthresholds (Fruhstorfer et al. 1976). While somewhatfaster than a protocol that separately evaluated warm,cool, heat pain and cold pain thresholds, the Marstockmethod relied on the subject recognizing the change intemperature direction each time, and knowing whichsensation to attend to at any point in time. This may beconfusing for a patient with � paresthesia, or those whohave completely lost the ability to feel some sensoryqualities.Almost all other pain threshold protocols use fixed dura-tion stimuli of a prescribed intensity, and do not rely onthe subject’s reaction time (see Fig 1, bottom). The mostcommon protocol of this type is a Staircase protocol, orone of its variants, including the Method of Levels. Inthis typeofmethod, a seriesof stimulus intensities ispre-scribed with a few simple rules. After a single stimulusis applied, the subject reports whether it was painful ornot. If it was not painful, the next stimulus is more in-tense. If it was painful, the next stimulus is less intense.Stimuli areapplied according to these rulesuntil aprede-fined endpoint is reached. That endpoint can be definedin terms of the number of “response reversals” (i.e. go-ing from “painful” to “non painful”, or vice versa, fromone trial to the next). Variations of this protocol includethe way in which stimulus intensity is changed from trialto trial. For instance, it is common to start the protocolwith relatively large changes (or “steps”) in stimulus in-tensity from trial to trial (such as 2–4˚C steps to deriveheat or cold pain), and reduce step size as the sessionprogresses. The initial temperature is typically chosento be well below pain threshold, so subsequent stimuli

2480 Threshold Determination Protocols
Threshold Determination Proto-cols, Figure 1 Representation ofa reaction time-dependent (top)and a reaction time independent(bottom) protocol. Top: Stimulusintensity increases until the subjectindicates (s) he feels pain (oranother specified percept), at whichtime the stimulus is returned toits baseline level. The stimulusintensity at the time of the report istaken as an estimate of threshold.This process is repeated, andthe threshold estimates averagedacross trials. Bottom: A prescribedstimulus is presented, and thesubject is instructed to provide aresponse at the end of the stimulus(i.e. “painful” or “not painful”).Depending upon the response,subsequent stimuli are of eithergreater or lesser intensity. See textfor further details.
will be of larger intensity, until the subject reports thatone of the stimuli is painful. At that point, the next stim-ulus is of a lower intensity, but the change in temperaturewill be less (for instance, half of the step size used to thatpoint). Once a stimulus intensity is reached that is notpainful, the next stimulus will be of a greater intensity,but will be yet a smaller step size. In this manner, thestimuli are “titrated” to reach the intensity closest to thepainful/non-painful level, ultimately at the smallest stepsize (resolution) desired. For the Method of Levels, thestep sizesaregradually reduced, and theprotocol iscom-pletewhentheminimallydesiredstepsize isachieved. Inthis case, the threshold is the average of the temperatureat the last tworesponsereversals.For theStaircaseproto-col, the step sizes reduce quickly to a minimal value, and
the protocol continues using that step size for several re-sponse reversals. In this case, the threshold is calculatedas an average of the last few (typically 4–6) response re-versal temperatures.A simpler variant of this type of protocol is to use a se-ries of stimuli in ascending intensity, until a stimulus isgiven that the subject reports as painful. This protocolis something like the Method of Limits (in terms of in-creasing stimulus intensity), but uses discrete stimulustrials, to make it a reaction time-independent protocol.The threshold would be estimated as the stimulus inten-sity between the first painful stimulus and the immedi-ately preceding one. This Ascending Method of Lim-its would be repeated several times, and threshold esti-matesaveraged.Aprotocolof this typehasbeensuccess-

T
Threshold Determination Protocols 2481
Threshold Determination Protocols, Figure 2 An example of a thresholddata set gathered using the Method of Limits protocol. The bars representthe temperature reached for each trial at the time the subject pressed abutton to indicate she felt a particular perception. The particular sensationfor any given set of trials is indicated at the bottom of the graph. Thethreshold for each sensory percept is calculated as the average of eachtrial, and indicated at the top of the graph (Adapted from data presentedin Greenspan 2001).
fully used for mechanical pain thresholds (Greenspanand McGillis 1991) and heat pain thresholds (Dyck etal. 1996).One problem with these protocols is the subjects’ pre-dictability of the stimulus sequence, particularly aftersome experience with it. One way to thwart such pre-dictability is to include some “out of sequence” stimulusintensities at random times, which are irrelevant to thecourse of the staircase. Another way is to use a Multi-ple Staircase protocol. For this protocol, one starts twoor more staircases, each operating by the same rules de-scribed above. However, from trial to trial, the stimu-lus may be drawn from any of the staircases, determinedrandomly. Thus, at the end of the protocol, one has twoor more estimates of pain threshold – one derived fromeach staircase – and the stimulus sequence was unpre-dictable on a trial by trail basis. Another variation of themultiple staircase protocol allows one to derive thresh-olds for more than just the pain threshold. If the subjectis instructed to report from a list of qualitative descrip-tors (i.e. “warm”, “hot”, “slight pain”, “moderate pain”,etc.), one can prescribe a staircase to titrate to any one ofthe qualitative percepts. Gracely et al. (1988) used thisprotocol to derive thresholds for “slight pain”, “moder-ate pain”, and “intense pain”, in a single test session,with heat stimuli. Taylor et al. (1993) and Greenspan etal. (1993) used a multiple staircase protocol to derivethresholds for “cool”; “slight heat pain” and “moder-ate heat pain” (see Fig. 3). However, the Greenspan etal. report revealed that mixing of cool and heat stimuliincreased the incidence of paradoxical heat sensations,rendering cool threshold estimates problematic in someinstances. Thus, it may be preferable to restrict multiplestaircases (or any other threshold protocol) to either de-tection or pain thresholds, within a single modality, forany single block of trials.
Threshold Determination Protocols, Figure 3 Data from two multiple-staircase sessions using thermal stimuli. Three different staircases wereprescribed: one for cool sensation, one for slight heat pain sensation, andone for moderate heat pain sensation. Each filled symbol represents a singletemperature presentation (a trial). After each stimulus was presented, thesubject chose a descriptor that best represented his sensation from a list ofthermal and pain terms. For each staircase, successive stimuli were moreintense until the criterion response was elicited. Then, subsequent stimuliwere progressively less intense, until a response other than the criterionresponse was elicited. Thus, the stimulus at which the subject’s responsechanged with respect to the criterion response produced a reversal of thestaircase. The trials producing such staircase reversals are marked by theopen symbols surrounding the filled symbols. Each staircase was continueduntil six reversals occurred. The dotted lines denote the calculated thresh-old for a given staircase, based on the average of its last four reversals.On any given trial, a random process selected one of the three staircasealgorithms to determine the temperature, thus preventing any prediction ofthe stimulus sequence. The adapting temperature was 34.0˚C. (Reprintedwith permission from Taylor et al. 1993; http://www.tandf.co.uk/journals).
In principle, one should be able to compare directlythreshold data gathered at different times, and by dif-ferent investigators, using the same protocol. Suchcomparisons are tenuous, due to the fact that varia-tions in protocol may systematically alter the resultingthreshold values. Differences in stimulus area, rate ofstimulus change, inter-stimulus interval, and precisebody site tested can significantly alter pain thresholds.Furthermore, situational factors such as the subject’sgeneral comfort, or the instructions to the subject, canaffect threshold values. It is for these reasons that com-parisons of threshold values are better made within agiven study, rather than across studies (Shy et al. 2003).

2482 Thromboembolism
References1. Boring EG (1942) Sensation and Perception in the History of
Experimental Psychology. Appleton-Century-Crofts, New York2. Dyck PJ, Zimmerman IR, Johnson DM et al. (1996) A Stan-
dard Test of Heat-Pain Responses using CASE IV. J.Neurol.Sci136:54–63
3. Fruhstorfer H, Lindblom U, Schmidt WG (1976) Method forQuantitative Estimation of Thermal Thresholds in Patients. JNeurol Neurosurg Psychiatry 39:1071–1075
4. Gracely RH, Lota L, Walter DJ et al. (1988) A Multiple Ran-dom Staircase Method of Psychophysical Pain Assessment. Pain32:55–63
5. Greenspan JD (2001) Quantitative Assessment of NeuropathicPain. Curr Pain Headache Rep 5:107–113
6. Greenspan JD, McGillis SLB (1991) Stimulus Features Rele-vant to the Perception of Sharpness and Mechanically EvokedCutaneous Pain. Somatosens Mot Res 8:137–147
7. Greenspan JD, Taylor DJ, McGillis SLB (1993) Body Site Vari-ation of Cool Perception Thresholds, with Observations on Para-doxical Heat. Somatosens Mot Res 10:467–474
8. Shy ME, Frohman EM, So YT et al. (2003) Quantitative SensoryTesting: Report of the Therapeutics and Technology AssessmentSubcommittee of the American Academy of Neurology. Neurol-ogy 60:898–904
9. Taylor DJ, McGillis SLB, Greenspan JD (1993) Body Site Vari-ation of Heat Pain Sensitivity. Somatosens Mot Res 10:455–466
10. Yarnitsky D (1997) Quantitative Sensory Testing. Muscle Nerve20:198–204
Thromboembolism
Definition
A thromboembolism is a blood clot (thrombus) formedin a blood vessel after surgery, which is transported asan embolus to the lungs or brain to cause an infarct.� Postoperative Pain, Acute Pain Team
Thrombosis
Definition
Thrombosis is the formation of clots inside blood ves-sels.� NSAIDs and their Indications
Thrombotic Event
Definition
Thrombotic Event refers to the formation of blood clotsin blood vessels e.g. coronary or cerebral arteries.� NSAIDs, Adverse Effects
Thromboxane
Definition
Thromboxanes are arachidonic acid derived moleculesthat are involved in platelet aggregation and blood clot-ting.� COX-1 and COX-2 in Pain� NSAIDs, Adverse Effects
Thromboxane A2
Definition
Thromboxane A2 is a prostaglandin-like material that issynthesized mainly by a COX–1–dependent process inactivatedplatelets.ThromboxaneA2promotesvasocon-striction and aggregation of platelets, thereby initiatingthe clotting of blood.� NSAIDs and their Indications
Tic and Cranial NeuralgiasZ. HARRY RAPPAPORT1, MARSHALL DEVOR2
1Department of Neurosurgery, Rabin Medical Center,Tel-Aviv University, Petah Tikva, Israel2Institute of Life Sciences and Center for Researchon Pain, Hebrew University of Jerusalem, Jerusalem,[email protected], [email protected]
Synonyms
Cranial NeuralgiasNeuralgia of Cranial Nerve V; trigeminal neuralgia; ticdouloureux; tic doloreuxNeuralgia of Cranial Nerve IX with or without CranialNerve X; glossopharyngeal neuralgia; vagoglossopha-ryngeal neuralgia; Neuralgia of Cranial Nerve VII;geniculate neuralgia
Definition
Cranial nerve neuralgia refers to a clinical pain symp-tom complex that consists of recurrent, intermittent,often paroxysmal pain felt in the head, in the distri-bution of a specific cranial nerve. By far the mostcommonly encountered cranial nerve neuralgia is thatof the trigeminal nerve and root. This is trigeminalneuralgia (tic douloureux). Neuralgias of the vago-glossopharyngeal nerves and the nervus intermediusbranch of the facial nerve are much rarer. Cranial nerveneuralgias are severely painful, but non life-threateningconditions.

T
Tic and Cranial Neuralgias 2483
Characteristics
Clinical Phenomenology
Trigeminal neuralgia is unique among cephalic-facialpain syndromes, both for its dramatic symptomatologyas for its susceptibility to both drug and interventionaltherapy. The incidence is approximately 4 new casesper 100,000 population/year. It is much more commonin the elderly than in young patients. There is no definitesex predisposition. Intense, intermittent paroxysms ofpain in the distribution of the trigeminal branches arefollowed by variable pain-free periods. The pain is uni-lateral and does not cross the midline of the face. Thereis usually a distinct trigger point, most commonly inthe perinasal or perioral area, but sometimes within theoral cavity. When the trigger point is stimulated, aftera short latency period, the painful paroxysm begins(Fig. 1). Pain may radiate from the trigger area alongthe appropriate trigeminal division, sometimes crossinginto adjacent divisions. Trigger stimuli are usually quiteweak, such as light touch, chewing, talking movementor a puff of wind. Paroxysms may also arise sponta-neously. Noxious stimulation of the trigger points isusually ineffective (Kugelberg and Lindblom 1959).The pain onset is usually sudden, and has an electricshock-like quality. A series of attacks, building to acrescendo, is common. Patients describe the pain as“shooting”, “electric”, or “cutting” in quality. Whilethe paroxysms tend to be short, in the order of secondsor occasionally minutes, they can run into each other
Tic and Cranial Neuralgias, Figure 1 A patient winces during a right-sided trigeminal neuralgia attack.
giving the impression of a prolonged attack over hours.Typically, pain attacks come in clusters separated byremissions of days to months, and sometimes evenyears (Rasmussen (1990). Following an attack the trig-ger area remains relatively refractory to further paininducing stimuli for up to 2 minutes (Kugelberg andLindblom 1959). Routine sensory examination of theface does not usually reveal any changes, but quanti-tative sensory testing may reveal elevated tactile andthermal detection thresholds in the area surrounding andincluding the trigger point (Nurmikko 1991). Trigemi-nal neuralgia is idiopathic; there is rarely any obviousprecipitating injury or disease. Prominent hypoes-thesia in the presence of typical trigeminal neuralgiasymptoms implies the presence of a paratrigeminalspace occupying lesion or multiple sclerosis. This issometimes termed atypical trigeminal neuralgia.The symptomatology of glossopharyngeal neuralgiais similar to that of trigeminal neuralgia, but pain isfelt in the distributions of the ninth (glossopharyn-geal) and/or tenth (vagus) cranial nerves (Rushton etal. 1981). � Pain paroxysms are usually triggered dur-ing swallowing, talking, or chewing, with the triggerpoint located unilaterally at the base of the tongue,tonsillar region, or in the ear. Concomitant vagal au-tonomic involvement probably accounts for episodesof syncope and bradycardia that may accompany painattacks. In more severe cases, prolonged asystole maybe life threatening. The disease is equally distributedbetween the sexes and is most frequently seen betweenthe fifth and seventh decades of life, with an incidenceof 0.8 cases/100,000/year. It is occasionally bilateral inpresentation. Pain tends to be less severe than trigeminalneuralgia, and often more steady and burning in quality(Katusic et al. 1991).Geniculate neuralgia involves the sensory distributionof the seventh cranial nerve (facial nerve) via the nervusintermedius (of Wrisberg). The painful attacks, whichare sharp and stabbing, are centered within the ear canal(Pulec 2002). A dull background pain may persist forseveral hours after an attack. Affected patients tend tobeyoungerthanintrigeminalandglossopharyngealneu-ralgia. When seen together with hemifacial spasm, theterm “ticconvulsive”hasbeen used (Yeh and Tew1984).
Pathophysiology
Medical and surgical experience in treating tic, com-bined with pathological findings and laboratory experi-ments using animal models, sheds light on the etiologyof pain paroxysms in the cranial nerve neuralgias. Dur-ing exploration of the cerebello-pontine angle on theside of the pain, vascular compression of the appropriatenerve root is usually observed (Fig. 2). Following de-compression, the patients are relieved of their pain in ahigh percentage of cases (Yeh and Tew 1984; Sindou etal. 2002; Patel et al. 2002). Ultrastructural examinationof post-mortem specimens, and cranial nerve biopsies

2484 Tic and Cranial Neuralgias
Tic and Cranial Neuralgias, Figure 2A photomicrograph of a compressedtrigeminal root (short arrow) andthe compressing blood vessel (longarrow), taken during microvasculardecompression surgery in trigeminalneuralgia. Note the discoloration of theroot at the compression site.
taken during surgical procedures, show extensive patho-logical change in the cranial nerve root and ganglion(Devor et al. 2002a; Devor et al. 2002b). Disorderingand loss of myelin is seen in the area compressed by thevessel loop, as are signs of axonal sprouting and regionsof close membrane contact between adjacent denudedaxons. The later observation is a likely anatomical sub-strate of ephaptic (electrical, non-synaptic) couplingbetween axons. These types of structural change cangive rise to ectopic pacemaker sites, and the genera-tion of abnormal impulse discharge. Moreover, theyfacilitate neuron-to-neuron spread of electrical activitythrough the injured cranial nerve root and its sensoryganglion, by means of neurophysiological mechanismssuch as � ephaptic coupling and crossed afterdischarge(Rappaport and Devor 1994). The generation of self-sustaining discharge in the root and ganglion followingperipheral triggering, and the rapid spread of this activ-ity, is thought to cause paroxysms of pain. This is the� “ignition hypothesis” of trigeminal neuralgia (Rap-paport and Devor 1994). Other properties of cranialnerve neuralgias, such as post-paroxysm refractori-ness and pain suppression by membrane stabilizinganticonvulsants, are consistent with this mechanism(Rappaport and Devor 1994). Central changes in sig-nal processing within the trigeminal brainstem havealso been proposed as causes of tic pain (Fromm andSessle–1991).
Treatment
The hallmark of cranial nerve neuralgias is their re-sponse to specific anticonvulsant medications, notablycarbamazepine and gabapentin, and to a lesser extent
phenytoin. Barbiturates are not effective, presumablybecause they act synaptically rather thanas� membranestabilizers. The effectiveness of drug therapy declinesover time in nearly one half of cases. When drug therapyno longer controls the pain paroxysms, or its side effectsimpair quality of life, interventional procedures may beconsidered (Rappaport 1996). Peripheral nerve blocksor neurectomies have only a short-term effect. Mostelderly patients undergo percutaneous partial damageto the trigeminal root and ganglion using radiofre-quency coagulation, retro-Gasserian glycerol injectionor balloon compression. The initial success rate is inthe 80 % range, with recurrence rates of up to 30 % at 1year and up to 45 % at 5 years. These procedures entailvarious degrees of facial hypoesthesia. Their efficacyis probably due to lessened triggering and reductionof trigeminal ganglion neuron populations, and hencereduction of the critical mass of electrical activity re-quired for pain ignition. Radiofrequency rhizolysis ofthe glossopharyngeal nerve is also effective for ninthnerve neuralgia; however, the procedure is technicallymore demanding.For patients who can tolerate the surgery, open exposureof the posterior fossa via a retromastoid craniotomy, and� microvascular decompression of the neural complex,optionally combined with partial rhizolysis, is the pre-ferred interventional procedure for all cranial nerve neu-ralgias.Microvasculardecompressionhastheadvantageof treating the root cause of the symptoms without de-stroying neural tissue and superimposing hypoesthesia.The low morbidity and mortality of this procedure (lessthan 4 %), and its impressive efficacy at relieving painwith only modest rates of recurrence (80 % still with sig-

T
Timolol 2485
nificant pain relief at 5 year follow-up) makes microvas-cular decompression the surgical treatment of choice, atleast in younger patients (Rappaport 1996).
Acknowledgements
We wish to thank Prof. Yair Sharav of the Hebrew Uni-versitySchoolofDentistry forprovidinguswith thepho-tograph of a patient during a trigeminal neuralgia parox-ysm used in Figure 1.
References1. Devor M, Govrin-Lippmann R, Rappaport ZH (2002a) Mech-
anism of Trigeminal Neuralgia: An Ultrastructural Analysis ofTrigeminal Root Specimens Obtained during Microvascular De-compression Surgery. J Neurosurg 96:532–543
2. Devor M, Govrin-Lippmann R, Rappaport ZH, Tasker RR,Dostrovsky JO (2002b) Cranial Root Injury in Glossopharyn-geal Neuralgia: Electron Microscopic Observations. J Neurosurg96:603–606
3. Fromm GH, Sessle BJ (1991) Trigeminal Neuralgia: CurrentConcepts Regarding Pathogenesis and Treatment. Butterworth-Heinemann, Boston, pp 1-230
4. Katusic S, Williams DB, Beard CM, Bergstralh EJ, Kurland LT(1991) Epidemiology and Clinical Features of Idiopathic Trigem-inal Neuralgia and Glossopharyngeal Neuralgia Similarities andDifferences, Rochester, Minnesota, 1945–1984. Neuroepidemi-ology 10:276–281
5. Kugelberg E, Lindblom U (1959) The Mechanism of the Pain inTrigeminal Neuralgia. J Neurol Neurosurg Psychiat 22:36–43
6. Nurmikko TJ (1991) Altered Cutaneous Sensation in TrigeminalNeuralgia. Arch Neurol 48:523–527
7. Patel A, Kassam A, Horowitz M, Chang YF (2002) Microvas-cular Decompression in the Management of GlossopharyngealNeuralgia: Analysis of 217 Cases. Neurosurgery 50:705–710
8. Pulec JL (2002) Geniculate Neuralgia: Long-Term Results ofSurgical Treatment. Ear Nose Throat J 81:30–33
9. Rappaport ZH (1996) The Choice of Therapy in Medically In-tractable Trigeminal Neuralgia. Isr J Med Sci 32:1232–1234
10. Rappaport ZH, Devor M (1994) Trigeminal Neuralgia: The Roleof Self-Sustaining Discharge in the Trigeminal Ganglion. Pain56:127–138
11. Rasmussen P (1990) Facial Pain: A Prospective Study of 1052Patients with a View of: Character of the Attacks, Onset, Course,and Character of Pain. Acta Neurochir 107:121–128
12. Rushton JG, Stevens JC, Miller RH (1981) Glossopharyngeal(Vagoglossopharyngeal) Neuralgia: A Study of 217 Cases. ArchNeurol 38:201–205
13. Sindou M, Howeidy T, Acevedo G (2002) Anatomical Obser-vations during Microvascular Decompression for IdiopathicTrigeminal Neuralgia (with Correlations between Topographyof Pain and Site of the Neurovascular Conflict). ProspectiveStudy in a Series of 579 Patients. Acta Neurochir 144:1–12
14. Tyler-Kabara EC, Kassam AB, Horowitz MH, Urgo L, Hadji-panayis C, Levy EI, Chang YF (2002) Predictors of Outcome inSurgically Managed Patients with Typical and Atypical Trigem-inal Neuralgia: Comparison of Results following MicrovascularDecompression. J Neurosurg 96:527–531
15. Yeh HS, Tew JM Jr (1984) Tic Convulsive, the Combination ofGeniculate Neuralgia and Hemifacial Spasm Relieved by Vas-cular Decompression. Neurology 34:682–68
Tic Douloureux
� Tic and Cranial Neuralgias� Trigeminal, Glossopharyngeal, and Geniculate Neu-
ralgias
� Trigeminal Neuralgia� Trigeminal Neuralgia, Diagnosis and Treatment� Trigeminal Neuralgia, Etiology, Pathogenesis and
Management
Time Constant
Definition
For a variable that is an exponential function of time
the time constant (τ) is the time after which the variable(x) has decreased by 1/e or increased by e.� Mechano-Insensitive C-Fibres, Biophysics
Time-Contingent Medication
� Time-Locked Medication
Time-Locked Medication
Synonyms
Time-Contingent Medication
Definition
Time-locked medication refers to medication intake noton an as needed (prn) schedule but at fixed points in timeto avoid maladaptive learning processes.� Operant Treatment of Chronic Pain
Timing
Definition
In the context of pre-emptive analgesia: an analgesictreatment can be initiated (or „timed“ to start) beforeor after the surgical injury.� Postoperative Pain, Pre-Emptive or Preventive Anal-
gesia
Timolol
Definition
A beta-blocker.� Migraine, Preventive Therapy

2486 Tinel Sign
Tinel SignTURO J. NURMIKKO
Department of Neurological Science and Pain ResearchInstitute, University of Liverpool, Liverpool, [email protected]
Synonyms
Tinel’s sign; Hoffman-Tinel sign; sign of formication
Definition
Tingling or sensation of pins and needles elicited by anexaminer tapping over a nerve. Tingling must be felt inthe territory supplied by the nerve.
Characteristics
The Tinel sign is a simple clinical test to identify localnerve damage or map the progress of nerve regenerationfollowing nerve injury. It was championed in the 1910sby two neuroscientists independently of each other, Dr.Jules Tinel (1879 – 1952), a French Neurologist, andPaul Hoffman (1884 – 1962), a German Neurophysi-ologist, but there are earlier descriptions of the sign inmedical literature. The phenomenon was probably wellknown to the medical community before their reports.Tinel and Hoffman are, however, credited for highlight-ing the clinical potential of the sign and giving it a mea-sure of scientific validity (Wilkins et al. 1971).The Tinel sign is a tingling perceived by a patient withnerve injury when the examiner taps over the trunk of thenerve. Tingling is felt in the skin supplied by the nerve,does not change on repetition, and outlasts the stimu-lus by no more than a few seconds. Tinel and Hoffmanconsidered this to signal the presence of regenerating� axons, which start to sprout from the proximal part ofthe nerve after injury. The further the recovery advances,the further away from the site of injury the Tinel sign canbe elicited. In this way, the successful progress of nerverecovery can be tracked.In its classical form, the test is carried out by applyingrepeated percussions by one’s extended finger over thepresumed course of the nerve, initially well below theexpected target, and continued upwardsuntil thefirst tin-gling sensation is elicited. The site where this happens isthe maximum distance over which the nerve fibers haveregenerated. In order to prevent false results from vibra-tions induced on the skin, the examiner places his freehand between the percussing finger and thesite of injury.In theearlyyearsof itspopularity, thesignwasused toes-timate the rate of recovery of the nerve following injury.Large case series were published, mostly involving pa-tients with gun shot wounds who had undergone surgeryfor their injury. At the time these were the best available
estimates of the recovery times of peripheral nerves, andranged from 1–3 mm/per day (Sunderland 1968).Not long after Tinel and Hoffman published their firstpapers, there were reports by several surgeons that thesign failed to indicateaccurately whetherornot recoveryof the injured nerve was taking place. Reports of patientswith a positive Tinel sign, who at operation were foundto have total interruption of the nerve with no chance ofrecovery, were published. It appears that the essentialrequirement for the sign to be positive – that it was pro-gressing – was overlooked by many. Nevertheless, morecriticism toward the sign was generated by observationsof good neural recovery in patients in whom the Tinelsign wasmissing (Napier1949;David and Chung 2004),and the test fell into disfavor. Later, a few investigatorspublished relatively large case series and concluded thatit is, when used critically, of moderate use as a roughguide of nerve recovery (Napier 1949; Nathan and Ren-nie 1946) There are today other more advanced meth-ods for assessment of such recovery, and the sign is tobe regarded more as a curiosity than a valid clinical tool.Despite this, it is always introduced to medical studentsand regularly appears in surgical and neurological text-books.The basis for the test lies in increased excitability of therecovering nerve fibers. Following injury, nerve fibersbegin to grow sprouts from the proximal stump. Thesesprouts are far more sensitive to mechanical stimuli thanare mature nerve fibers in intact nerves. As the sproutsgo longer, the most distant point of mechanical sensitiv-ity moves further away from the injury. � Myelin break-age and restoration, and recovery of support structuresof the nerve trunk, also have an impact on the generalexcitability of the recovering nerve. In the part of thenerve in which full or near-full recovery of fibers hastaken place, the nerve trunk loses its sensitivity to per-cussion.Tinel, and later advocates of his method, stressedthat tapping should produce tingling, not pain in thereference area. Tinel did acknowledge that at timesmechanical compression of the site of injury couldcause pain, prominent on the site of compression,with less intense pain referred to the territory of thenerve. To Tinel, the natural explanation for a stationarysign was that nerve degeneration did not take place,and that the regenerating nerves grew aimlessly intoa bundle of sprouts, forming a � neuroma. Neuromasare known for their excessive mechanical sensitivity.Inevitably, the sign was soon adopted for diagnosingneuromas, with no evidence that it provides more infor-mation than careful manual palpation or compression.It has been suggested that a neuroma can be distin-guished from other tender structures at the operationsite by using a series of light weight percussion tools(Tucker and Nancarrow 2000), but this method is notin common use. There are sophisticated electrophys-

T
TNS 2487
iological and neuroimaging methods that can be usedinstead.In the 1950s, Phalen and co-workers reported that theycould frequently elicit the Tinel sign in patients with� carpal tunnel syndrome and added the test to theirother diagnostic criteria (Phalen et al. 1950). In carpaltunnel syndrome, the median nerve is under constantcompression, imposing increasing structural damageon nerve fibers. Axonal degeneration and regenerationtake place concurrently, and with varying levels ofdemyelination and remyelination present, the nervebecomes excessively sensitive to mechanical stimuli.However, several studies have been published that showconvincingly that the Tinel sign has little to offer fordifferential diagnosis. It is too crude a test for accuratedifferentiation between patients with carpal tunnel syn-drome from those with other peripheral nerve diseases,or even healthy people (D’Arcy et al. 2000).The significance of the Tinel sign has faded with time,and these days it remains more a curiosity than a seriousclinical tool. It is still used out of its original context inpain research – more as a “neuroma sign”, rather thanthe sign of a recovering nerve than it was used for in theearly part of the 20th century.� Neuroma Pain� Peripheral Neuropathic Pain� Ulceration, Prevention by Nerve Decompression
References1. D’Arcy CA, McGee S (2000) Does this Patient have Carpal Tun-
nel Syndrome? JAMA 283:3110–31172. Dellon AL (1984) Tinel or not Tinel. J Hand Surg 9:2163. David EN, Chung KC (2004) The Tinel Sign: A Historical Per-
spective. Plast Recontr Surg 114:494–4994. Moldaver J (1978) Tinel’s Sign. J Bone Joint Surg 60-A:412–4135. Napier JR (1949) The Significance of Tinel’s Sign in Peripheral
Nerve Injuries. Brain 72:63–826. Nathan PW, Rennie AM (1946) Value of Tinel’s Sign.
Lancet I:610–6117. Phalen GS, Gardner WJ, LaLonde AA (1950) Neuropathy of
the Median Nerve Due to Compression beneath the TransverseCarpal Ligament. J Bone Joint Surg 32A:109–112
8. Sunderland S (1968) Nerves and Nerve Injuries. E&S Living-stone, Edinburgh
9. Tucker SC and Nancarrow JD (2000) Objective Assessmentof Post-Traumatic Nerve Repairs and Neuromas. Br J PlasticSurg 53:694–696
10. Wilkins RH, Brody IA (1971) Neurological Classics XXXIV.Tinel’s Sign. 24:573–575
11. Yarnitsky D, Ochoa JL (1991) The Sign of Tinel can be mediatedeither by Myelinated or Unmyelinated Primary Afferents. MuscleNerve 14: 379–380
Tinnitus
Definition
A ringing sensation in one or both ears.� NSAIDs, Adverse Effects
Tissue Fatigue
Definition
Tissue fatigue refers to a decline in the load-bearing ca-pacity of tissue in response to an applied load. This phe-nomenon is reversible provided that there is sufficienttime for recovery.� Ergonomic Counseling
TLIF
Synonyms
Transforaminal interbody fusion
Definition
Graft/cages placed between the vertebral bodies by pos-terior approach through the neural foramina without re-tracting the thecal sac.� Spinal Fusion for Chronic Back Pain
TMD
� Temporomandibular Disorder
TMS
� Transcranial Magnetic Stimulation
TMJD
� Temporomandibular Joint Disorders
TNF Alpha(α)
� Tumor Necrosis Factor Alpha(α)
TNS
� Transcutaneous Electrical Nerve Stimulation Out-comes

2488 Tolerance
Tolerance
Definition
Tolerance refers to diminishing susceptibility to the ef-fects of a drug with continued use. The phenomenon oftolerance is characteristic of opioid analgesics. The de-velopmentof tolerance to theanalgesiceffectsof theopi-oidsmeans thatwith continued opioiduse,progressivelyhigher doses may be required to maintain the same anal-gesic effect. Tolerance also develops to the nonanalgesicside effects of opioids, including respiratory depression,nausea, and sedation. The development of tolerance tothese effects is desirable as it permits dose titration.� Cancer Pain Management, Opioid Side Effects, Un-
common Side Effects� CRPS, Evidence-Based Treatment� Opiates During Development� Opioids and Inflammatory Pain� Opioids in the Periphery and Analgesia� Opioid Receptors� Opioid Responsiveness in Cancer Pain Management� Opioid Therapy in Cancer Patients with Substance
Abuse Disorders, Management� Postoperative Pain, Opioids� Purine Receptor Targets in the Treatment of Neuro-
pathic Pain� Stimulation-Produced Analgesia
Tolerance Thresholds
� Pain in Humans, Thresholds
Tolosa-Hunt Syndrome
Definition
Also called: “painful ophthalmoplegia“: a variable com-bination ofperiorbitalpain, ipsilateraloculomotornervepalsies, oculosympathetic palsy and trigeminal sensoryloss.� Headache due to Dissection
Tonic Firing Mode
Definition
A pattern of spontaneous action potential firing demon-strated by thalamic neurons, in which no clear patternof interspike intervals exists.
� Central Pain, Human Studies of Physiology� Thalamic Bursting Activity, Chronic Pain
Tonic Rebalance
Definition
Tonic rebalance is a spontaneous, physiological processby which thesystem tries to restore symmetry at the levelof the vestibular nuclei in the acute phase after a vestibu-lar crisis. The process occurs independently of the pa-tient’s activities.� Coordination Exercises in the Treatment of Cervical
Dizziness
Toothache
� Dental Pain, Etiology, Pathogenesis and Management
Top-Down Control of Pain
Definition
Neural pathways descending from higher brain struc-tures to lower brain structures modulate ascending painsignals.� Descending Modulation and Persistent Pain
Topical Drug TherapyCHARLES NG
Musculoskeletal Medicine Specialist, AustralasianFaculty of Musculoskeletal Medicine, Auckland, [email protected]
Definition
Topical drug therapy is the application of drugs to theskin over an area of pain. The drug penetrates the skinto act on the underlying painful tissues or the nerves thaninnervate them.
Characteristics
The three main groups of topical medications used intreating pain are topical NSAIDs (nonsteroidal anti-inflammatory drugs), topical local anaesthetics andtopical capsaicin. These drugs may be formulated as acream, gel, paste or patch.

T
Topical Drug Therapy 2489
Topical NSAIDs (nonsteroidal anti-inflammatory drugs)
Mechanisms of Action
Topical NSAIDs have a peripheral action on soft tissue,inhibitingprostaglandinsynthesis, inhibitingthelipoxy-genase pathway and inhibiting excitatory amino acids.They have effects on G protein-mediated signal trans-duction (Galer 2001). They also reduce the neurogenicinflammatory response.Evidence shows that therapeutic concentrations areachieved in soft tissues (Vaile et al. 1998). These tissuescan act as a drug reservoir, and thus maintain tissueand plasma concentration after application of the drughas ceased. Systemic redistribution of NSAIDs resultsin measurable plasma levels. However, plasma, mus-cle and synovial fluid concentrations have been foundto be lower than in subcutaneous fat and soft tissue,and are possibly subtherapeutic. Consequently, topicalNSAIDs are more effective for soft tissue complaintsthan arthropathies.
Applications and Efficacy
Topical NSAIDs are most effective for soft tissuecomplaints. Evidence supports the use of ketorolac,flurbiprofen and niflumic acid in acute ankle sprain.Indomethacin and piroxicam are effective in sprainsand tendinitis. Naproxen and felbinac are effectivein a variety of soft tissue injuries (Vaile et al. 1998).Diclofenac is effective in treating lateral epicondylitis(Burnham et al. 1998).Moore et al. (1998) showed that topical NSAIDs weresignificantly more effective than placebo in the treat-ment of acute sprains and strains, with an � NNT(number needed to treat) of 3.9; and chronic arthritisand rheumatism, for which the NNT was 3.1. Keto-profen, felbinac, ibuprofen and piroxicam had similar,significant efficacy.As the natural history of soft tissue complaints tends tobeself-limiting, treatmentwith topicalNSAIDsdoesnotnecessarily alter the overall outcome.Topical NSAIDs are less effective for arthropathies, buthave been shown to be effective in treating mild to mod-erate osteoarthritis (Hosie and Bird 1994). However, ev-idence supporting their efficacy in osteoarthritis of theknee is not consistent (Vaile et al. 1998).
Side Effects
Cutaneous adverse reactions like erythema, burn-ing sensation, irritation and contact dermatitis occurin 1 to 2% of patients (Vaile et al. 1998). Asthma, renaland gastrointestinal side-effects have been reported.However, as plasma concentrations of topical NSAIDsare low compared to oral NSAIDs, gastrointestinalside-effects are less severe and less frequent. There-fore, topical NSAIDs are promoted as a safer and morecost-effective alternative.
Topical Local Anaesthetics
Mechanisms of Action
Local anaesthetics act through the blockade of sodiumchannels in sensory afferent neurons. Thereby, topicalapplication reduces ectopic discharge in superficialsomatic nerves that are stimulated in neuropathic painstates and acute injuries (Galer 2001).
Applications and Efficacy
Studies support the efficacy of topical lidocaine, ineither the Lidoderm patch or gel formulation, for re-lieving pain in chronic � postherpetic neuralgia and formechanical � allodynia (Galer 2001, Sawynok 2003).There is anecdotal evidence that topical lidocaine maybeuseful fordiabeticand otherperipheral� neuropathicpains (Galer 2001).Topical application of EMLA (eutectic mixture of lo-cal anaesthetics, 2.5% lidocaine and 2.5% prilocaine)reduces acute pain associated with invasive proceduressuch as venipuncture (Galer 2001). It also reduceschronic pain in postherpetic neuralgia and chronicpost-surgical pain (Towlerton and Rice 2003).Topical tetracaine provides effective anaesthesia forendotracheal, ocular and invasive skin procedures(Galer 2001). Topical cocaine provides effective anaes-thesia for nasal mucosa.A combination of a low dose of morphine and lidocaineresults in analgesic synergy, where the analgesic activityexceeds that from a simple additive effect of the agents(Kolesnikov et al. 2000).
Side Effects
Application of topical local anaesthetics does not resultin clinically significant plasma levels causing side ef-fects (Galer 2001; Towlerton and Rice 2003).
Topical Capsaicin
Mechanisms of Action
Capsaicin isanextractofchilipeppers. It is formulatedasa topical application, which when applied to skin causesan intensely painfulburningsensation,byselectivelyde-polarising small diameter sensory afferent neurons. Re-peated application causes a desensitising or neurotoxiceffect on nociceptors, which results in a prolonged “se-lective local analgesia” (Sawynok 2003; Towlerton andRice 2003). Desensitisation of skin results in an analge-siaeffect.Capsaicinstimulates thereleaseofsubstancePand calcitonin gene-related peptide from peripheral andcentral terminals of sensory neurons. Their release is in-hibited by the desensitisation effect (Sawynok 2003).
Applications and Efficacy
Evidence supporting the use of capsaicin in posther-petic neuralgia and painful � diabetic neuropathy isunconvincing (Galer 2001; Towlerton and Rice 2003).Evidence supports its use in osteoarthritis at a variety ofsites, including the hand and knee, and in rheumatoid

2490 Topiramate
arthritis (Galer2001;TowlertonandRice2003). It isuse-ful in non-specific neck pain (Mathias et al. 1995), clus-ter headaches and post-mastectomy pain (Galer 2001).Anecdotal evidence attests to its � effectiveness in arange of other neuropathic pain conditions and facialpain (Galer 2001).Capsaicin is often not sufficient as monotherapy fortreating chronic pain. It appears to be useful only inconjunction with other therapies.
Side Effects
Initial application of capsaicin, especially in the firstweek, causes burning pain at the site of applicationin 40–80% of patients (Towlerton and Rice 2003). This,along with a therapeutic effect time-delay of at least aweek, reduces patient compliance and continuation ofuse.
References1. Burnham R, Gregg R, Healy P, Steadward R (1998) The Effec-
tiveness of Topical Diclofenac for Lateral Epicondylitis. ClinJournal Sports Med 8:78–81
2. Galer B (2001) Topical Medications. In: Loeser JD (ed) Bonica’s
Management of Pain, 3rd edn. Lippincott Williams & Wilkins,Philadelphia, pp 1736–1742
3. Hosie G, Bird H (1994) The Topical NSAID Felbinac versus OralNSAIDs: A Critical Review. Eur J Rheumatol Inflam 14:21–28
4. Kolesnikov Y, Chereshnev I, Pasternak G (2000) Analgesic Syn-ergy between Topical Lidocaine and Topical Opioids. J Pharma-col Exp Ther 295:546–551
5. Mathias B, Dillingham T, Zeigler D, Chang A, Belandres P (1995)Topical Capsaicin for Chronic Neck Pain. Am J Phys Med Re-hab 74:39–44
6. Moore R, Tramer M, Carroll D, Wiffen P, McQuay H(1998) Quantitative Systematic Review of Topically AppliedNon–Steroidal Anti–Inflammatory Drugs. BMJ 316:333–338
7. Sawynok J (2003) Topical and Peripherally Acting Analgesics.Pharmacological Reviews 55:1–20
8. Towlerton G, Rice A (2003) Topical Analgesics for Chronic Pain.In: Jensen T, Wilson P, Rice A (eds) Clinical Pain ManagementChronic Pain. Arnold, London, 213–226
9. Vaile J, Davis P (1998) Topical NSAIDs for MusculoskeletalConditions. Adis International Limited 56:783–799
Topiramate
Definition
Anticonvulsant medication.� Migraine, Preventive Therapy
Topographical
Definition
Topographical refers to the arrangement or reference toregions of the body or of a body part, especially the re-gions of a definite and limited area of the surface.� Postsynaptic Dorsal Column Projection, Anatomical
Organization
Topography
� Magnetoencephalography in Assessment of Pain inHumans
Total Pain
Definition
Expression of suffering, particularly the expression ofpsychological distress as pain.� Cancer Pain Management, Interface between Cancer
Pain Management and Palliative Care
Touch Evoked Pain
� Allodynia (Clinical, Experimental)
Tourniquet Ischemia
� Tourniquet Test
Tourniquet Pain Ratio
Definition
It is calculated as follows: Ongoing Clinical Pain MatchTime/Tolerance Time X 100 where the Ongoing Clini-cal Pain Match time is the time point, since the begin-ning of the Tourniquet Test, at which the patient reportsa pain intensity that matches the clinical pain intensity;the Tolerance Time is the time the patient can toleratethe ischemia in the Tourniquet Test.� Tourniquet Test
Tourniquet TestGIANCARLO CARLI
Department of Physiology, University of Siena, Siena,[email protected]
Synonyms
Ischemic Test; Submaximum Effort Tourniquet Tech-nique; Forearm Ischemia Procedure; Acute IschemiaTest; Forearm Occlusion Pain; Ischemic pain; Tourni-quet Ischemia

T
Tourniquet Test 2491
Definition
Ischemic� pain iselicited by having thesubject squeezea handspring exercises 20 times after a tourniquet isinflated around his upper arm. The quality of sensa-tion is dull-aching or stinging � muscular pain, whichclosely resembles most types of � pathologic pain, butincreases progressively after cessation of squeezing.Test performance is measured in terms of elapsed timebetween cessation of squeezing and report of slight(threshold) and unbearable (tolerance) pain. Muscularpain from ischemic contractions, which is due to tran-sient stimulation of peripheral � nociceptors (LaMotteand Campbell 1978), is based on the � algogenic ac-tions of protons (Issberner et al. 1996). Noxious forearmischemia also evokes substantial elevations in arterialblood pressure (Maixner et al. 1990) and activationof endogenous opioid systems (Goldstein and Grevert1978), thus representing a � diffuse noxious inhibitorycontrols (DNIC) paradigm (Willer et al.1989).A pneumatic tourniquet cuff connected to a computercontrolled air compressor has been used recently to pro-duce pressure pain (Polianskis et al. 2001). Computer-ized cuff algometry may continuously control stimulusor adjust the pressure to maintain the programmed painlevel. This set up, which elicits an increasing pain im-mediately after tourniquet cuff application around thegastrocnemius-soleus muscle, represents a highly con-figurable tool for assessment of pain sensitivity by pres-sure, but does not involve the ischemic mechanisms asdoes the classical tourniquet test described in this chap-ter.
Characteristics
The need for a dependable experimental method to testnewanalgesicsinmanwasoneofthemainmotivationstomodify the earlier techniques of producing � IschemicPain/Test (Lewis et al. 1931, Hewer and Keele 1948),by changing from maximum to submaximum squeezingeffort.Originally, the � submaximum tourniquet test (Smith etal. 1966; Smith et al. 1968) was suggested with the fol-lowing order (see Fig. 1a, b, c).The subject, reclining on a bed, is asked to comply withthe following procedure:
1. to extend his non-dominant arm toward the ceilingand, with the arm raised, an Esmarch bandage iswrapped from the fingers to the elbow to drain thearm of venous blood (see Fig. 1a)
2. to apply a tourniquet bandage around the upper armand inflate it to obtain a pressure of 250 mm Hg, toabolish arterial supply and to render the arm hypoxic
3. to lower the arm, remove the band and, after a pauseof 60 sec, to squeeze a hand spring exerciser 20 timeswhile his arm is rested. Each squeeze is timed to last2 sec followed by a 2 sec rest, so that the whole ex-
ercise time lasts 80 sec. The schedule is presented tothe subject by means of tape-recorded signals
4. after completing thesqueezes, to remain in arecliningposition and resthisarm athis sidewith the tourniquetstill inflated, not to move his arm until the pressurehas been released (see Fig. 1c)
5. to rate, when asked at irregularly spaced intervals, theincoming sensations of pain according to a predeter-mined scale, which includes the following categoriesof experience: 0 = none; 1 = slight; 2 = moderatelydistressing; 3 = very distressing; 4 = unbearable. Ir-regular intervals should be used to minimize the cuesto the subject regarding the time he had tolerated thepain.
� Pain threshold (slight pain) occurs after about3–6 min, while unbearable pain (tolerance) variesfrom 7 to 53 min (Smith et al. 1966; Smith et al. 1968).The latency of unbearable pain is usually called � paintolerance, but formally it represents ischemia tolerance,i.e. the measurement of time elapsed since exercisetermination (beginning of ischemia) and not since painonset. When the cuff is released, pain intensity andnumbness sensations sharply decrease, but their timecourse, as not included in the formal test, have not beensystematically investigated.Several authors have modified the test, but the mainchanges that have improved the test have been pro-posed by Moore et al. (1979). According to new sug-gestions, the individual maximal grip strength of thenon-dominant arm has to be assessed using an isometricexerciser equipped with a force gauge (Fig. 1b). Afterlowering the arm, the squeeze exercise must start im-mediately after the cuff has been inflated to 200 mm Hg,instead of 250, avoiding a pause of 60 sec. as suggestedby Smith et al. (1966). The individual grip strengthshould be kept at the fixed percentage (50 %) of themaximum grip strength, in order to avoid fatigue and toreduce inter-subject differences. The measurement ofelapsed time should begin at the time of blood pressurecuff inflation, i.e. at the beginning of the ischemia, ratherthan at exercise termination. In addition, the rating mustbe in a visual analog scale from 0 (no pain) to 100 (painso severe that you would commit suicide if you had tobear it for more than a few minutes).In the same research (Moore et al. 1979), it was observedthat the amount of exercise used during the submaximaleffort tourniquet test significantly affected the level ofreported pain intensity versus time elapsed: in fact, byincreasing the duration of each contraction from 0.6 sec.to 2 sec, the threshold pain intensity was reached muchmore rapidly.
Tourniquet Pain Ratio: A Method to Evaluate Clinical Pain
Sternback et al. (1974) originally suggested that the ex-perimental pain generated by ischemic exercise couldbe used to evaluate the severity of the ongoing clinical

2492 Tourniquet Test
Tourniquet Test, Figure 1 Mainphases of the tourniquet test. (a)elevation of the bandaged forearm;(b) squeezes of isometric exerciserwhile the pneumatic tourniquetremains inflated to block forearmcirculation; (c) arm at rest while thetourniquet is still inflated.
pain. This method, called � Tourniquet Pain Ratio, re-quires thepatients to reportwhen their armpain intensitymatches the clinical pain intensity: this time point is thenused to measure the subjective severity of patient’s clin-ical pain and is calculated as follows: (Ongoing ClinicalPain Match Time / Tolerance Time) X 100. This methodhas several limitations, mainly because the TourniquetPain Matching Score is significantly lower than the pa-tient’s own pain estimate (Moore et al. 1979). As for therelationship between pain intensity and elapsed time, thedata indicate that thepain ratingsproducedbythe test arenot a linear function of elapsed time but rather a sigmoidshaped curve. Nonetheless, the Touriquet Pain Ratio isstill used to provide an additional index of clinical painmodulation (Sigurdsson and Maixner 1994).
Tourniquet Test in the Diagnosis of Altered Nociceptive Trans-mission
Both threshold and tolerance to ischemic pain has beenrepetitively tested in clinical syndromes such as local-ized musculoscheletal pain, � fibromyalgia, peripheralneuralgias, bulimia, hypertension and other conditionsin which a dysfunction in the excitability of the nocicep-tive system is expected.
Tourniquet Test and DNIC
The efficacy of DNIC, the neurophysiological mecha-nism that underlines the long established clinical phe-nomenon of � counterirritation (Wand-Tetley 1956),has been successfully tested in healthy subjects follow-ing the procedure of the submaximum tourniquet test,
in electrical threshold to evoke muscle pain (Pantaleo etal. 1988), cutaneous warming pain threshold (Pertovaraet al. 1982) and in other sensory modalities. In patientswith ongoing pain, it has been established that pain byclinical origin does not stimulate DNIC on ischemicor thermal pain perception (Ekbloom and Hansson1987; Hansson et al.1988). On the contrary, forearmischemia elicited by the procedure of the submaximumtourniquet test elicits a generalized non-segmental in-hibition of tooth pain, resulting from acute irreversiblepulpitis (Sigurdsson and Maixner 1994). The lattereffects remain for at least 5 minutes after removal ofthe tourniquet, while the arm is free from pain. It hasto be underlined that the mechanisms of DNIC are notunivocal since, for instance, the tourniquet test elicitsinhibition of static mechano-allodynia sensations, trig-gered by pressure stimuli, but has no effect on dynamicmechano-allodynia sensations, elicited by brushing(Bouhassira et al. 2003). In fibromyalgia and painfulosteoarthritispatients theprocedureof thesubmaximumtourniquet test does not elicit modulation of pressurepain, as opposed to controls (Kosek et al. 1966), whilethe DNIC is still effective in patients suffering fromlong-term trapezius myalgia.
Touniquet Test and Drug Screening
It has been repeatedly established that placebo and sug-gestions of analgesia, both in subjects who are awakeand during hypnosis, can reduce pain elicited by thetourniquet test. Similarly, small doses of opiates (mor-phine, dipipanone, codeine) and the NMDA-receptor

T
Toxic Neuropathies 2493
antagonist ketamine display analgesic effects. On thecontrary, the NMDA-receptor antagonist dextromethor-phan, diazepan and anti-inflammatory drugs such asaspirin or indomethacin do not affect ischemic pain.Finally, adenosine, which mediates ischemic pain inhumans, at low doses exerts analgesic effects on thetourniquet pain by membrane-bound peripheral adeno-sine receptors (Eriksson et al. 2000).
Concluding Remarks
In conclusion, the tourniquet test can provide usefulindications about individual reactivity and toleranceto deep ischemic stimuli, both in healthy subjects andin patients. There are, however, some limitations to beunderlined. First of all, in patients suffering from deeppressure pain or other deep pains in the forearms, un-bearable pain can occur following the application and/orinflation of the bandage. Moreover, the tourniquet pro-cedure has shown a considerable between subjects andbetween session variability (Sigurdsson and Maixner1994), and the sensitivity displayed may be inadequateto assess the analgesic effects of some pharmacologicalagents (Sternbach et al. 1977).
References1. Bouhassira D, Danziger N, Atta N et al. (2003) Comparison of the
Pain Suppressive Effects of Clinical and Experimental PainfulConditioning Stimuli. Brain 126:1068–1078
2. ErikssonBE, SadigB, SvedenhagJ et al. (2000) Analgesic Effectsof Adenosine in Syndrome X are Counteracted by Theophylline:A Double-Blind Placebo-Controlled Study. Clin Sci 98:15–20
3. Ekblom A, Hansson P (1987) Thermal Sensitivity is not Changedby Acute Pain or Afferent Stimulation. J Neurol Neurosurg Psy-chiat 50:1216–1220
4. Goldstein A, Grevert P (1978) Placebo Analgesia, Endorphinsand Naloxone. Lancet 2:1385
5. Hansson P, Ekbloom A, Lindblom U et al. (1988) Does AcuteIntraoral Pain Alter Cutaneous Sensibility? J Neurol NeurosurgPsychiat 51:10032–1036
6. Hewer AJ, Keele CA (1948) A Method of Testing Analgesicsin Man. Lancet 2:683–688
7. Issberner U, Rehe PW, Steen KH (1996) Pain due to Tissue Aci-dosis: A Mechanism for Inflammatory and Ischemic Myalgia?Neurosci Lett 208:191–194
8. Kosek E, Ekholm J, Hansson P (1966) Sensory Dysfunction inFibromyalgia Patients with Implications for Pathogenic Mech-anisms. Pain 68:375–383
9. LaMotte RH, Campbell JN (1978) Comparison of Responses ofWarm and Nociceptive C-Fiber Afferents in Monkey with HumanJudgements of Thermal Pain. J Neurophysiol 41:509–528
10. Lewis D, Pikering GW, Rothschild P (1931) Observations uponMuscular Pain in Intermittent Claudication. Heart 15:359–383
11. Maixner W, Gracely RH, Zuniga JR et al. (1990) Cardiovascularand Sensory Responses to Forearm Ischemia and Dynamic HandExercise. Am J Physiol 259:R1156–R1163
12. Moore PA, Duncan GH, Scott DS et al. (1979) The SubmaximalEffort Tourniquet Test: Its use in Evaluating Experimental andChronic Pain. Pain 6:375–382
13. Pantaleo T, Duranti R, Bellini F (1988) Effects of HeterotopicIschemic Pain on Muscular Pain Threshold and Blink Reflex inHumans. Neurosci Letter 85:56–60
14. Pertovara A, Kemppainen P, Johannson G et al. (1982) IschemicPain Nonsegmentally Produces a Predominant Reduction of Painand Thermal Sensitivity in Man: A Selective Role for Endoge-nous Opioids. Brain Res 251:82–93
15. Polianskis R, Graven Nielsen T, Arendt-Nielsen (2002) Spatialand Temporal Aspects of Deep Tissue Pain Assessed by CuffAlgometry. Pain 100:19–26
16. Sigurdsson A, Maixner W (1994) Effects of Experimental andClinical Noxious Counterirritants on Pain Perception. Pain57:265–275
17. Smith GM, Egbert LD, Markowitz RA et al. (1966) An Exper-imental Pain Method Sensitive to Morphine in Man: The Sub-maximum Effort Tourniquet Technique. J Pharmacol Exp Ther154:324–332
18. Smith GM, Lowenstein E, Hubbard JH et al. (1968) Experi-mental Pain Produced by the Submaximum Effort TourniquetTechnique: Further Evidence and Validity. J Pharmacol Exp Ther163:468–474
19. Sternbach RA, Deems LM, Timmermans G et al. (1977) On theSensitivity of the Tourniquet Pain Test. Pain 3:105–110
20. Wand-Tetley JI (1956) Historical Methods of Counter-Irritation.Ann Phys Med 3:90–98
21. Willer JC, DeBroucker T, LeBars D (1989) Encoding of Noci-ceptive Thermal Stimuli by Diffuse Noxious Inhibitory Controlsin Humans. J Neurophysiol 62:1028–1038
22. Williams MW (1959) Ischemic Arm Pain and Non-NarcoticAnalgesics. Toxicol Appl Pharmacol 1:590–597
Toxic NeuropathiesSUNG-TSANG HSIEH
Department of Anatomy and Cell Biology andDepartment of Neurology, National Taiwan University,Taipei, [email protected]
Definition
Toxic neuropathies are peripheral nerve disorders due toacute or chronic adverse effects of chemicals or medi-cations which occur in individuals or as an outbreak ina certain population after occupational exposure or en-vironmental contamination.
Characteristics
Neurons, Axons and Neuropathy� Neuropathies are diseases of the peripheral nervoussystem from various etiologies, including metabolic dis-orders, such as � diabetic neuropathy due to � diabetesmellitus (Vinik AI et al. 2000), and toxic effects ofchemotherapy for cancer. In the peripheral nervoussystem, the length of � axons is much greater thanthe diameter of the neuronal cell body. This results ina much larger cytoplasmic volume in the entire axoncompared to the volume of the neuronal cell body, oftenby several orders of magnitude. As an example, takea motor neuron of an adult human. The diameter of amotor neuron in the lumbar spinal cord is up to 100 μm;in contrast, the length of a motor axon from this motorneuron innervating the foot muscles is up to 1 m inlength. This greatly differs from other cells in the body,such as fibroblasts, which have a cellular extension ofonly a limited length.To maintain such unusual structural requirements, ax-ons possess a rather complicated cytoskeletal system

2494 Toxic Neuropathies
(Griffin et al. 1995). Components of the � cytoskeletoninclude microtubules (> 24 nm), intermediate filaments(10 nm), and microfilaments (6–8 nm) according to theirdiameters and composition. These cytoskeletal proteinsinteract with each other through associated proteins,such as microtubule-associated proteins, to form athree-dimensional interlacing structure. In addition, themetabolic demands of axons heavily depend on the neu-ronal cell body, and an � axoplasmic transport systemhas developed to transport materials both to and fromneuronal cell bodies. Some cytoskeleton-associatedproteins, such as the kinesin and dynein superfamily,are also responsible for transporting organelles andneurotransmitter containing vesicles (Hirokawa 1998).Peripheral nerves, particularly those in nerve terminalregions, are distributed throughout the entire body, andare thus the most vulnerable part of the nervous systemduring toxin exposure and metabolic derangement.Consequences of these insults, including dysregulatedcytoskeletal maintenance or blockade of � axonaltransport, may impair the functioning of axons, andeventually result in � axonal degeneration. Alterna-tively, neurons are vulnerable to toxins via systemicabsorption. Toxins directly act on neuronal cell bodies,resulting in neuronal cell death and subsequent axonaldegeneration.
Structural Organization of Peripheral Nerves and Pathology ofToxic Neuropathies
The structural organization of peripheral nerves in-cludes two major cellular components: neurons withcytoplasmic extensions and axons, and ensheathingglia (� Schwann cells) with myelin sheaths, modifiedmembranous insulating materials. Nerve injury in toxicneuropathies occurs at different levels along the neuralaxis through different mechanisms. Toxins can act onneuronal components (neuronal cell bodies, axons, andnerve terminals) and glial components (Schwann cellsand myelin sheaths). Neuronal degeneration results insubsequent nerve fiber degeneration. Most frequently,the major target site is at axons, either proximally or dis-tally along the nerve fibers, while neuronal cell bodieslargely remain intact. The common outcome is axonaldegeneration, or Wallerian-like degeneration, and sec-ondary demyelination. The pathology of most toxicneuropathies is axonal degeneration, particularly of thedistal axons, such as with � acrylamide (Schaumburget al. 1974). Only rare experimental toxins, such asβ,β’-iminodipropionitrile, act on proximal axons (Grif-fin et al. 1983). If a toxin selectively damages Schwanncells or impairs myelin organization, neuropathies ofprimary demyelination will develop. Because of con-duction failure in primary � demyelination, the majorelectrophysiological abnormality of � demyelinatingneuropathy is the marked slowing of nerve conduction.This type of toxic neuropathy is relatively rarecomparedto neuropathies of the primarily axonal degeneration
type. An example of a primarily demyelinating neu-ropathy is tellurium-induced toxic neuropathy (Bouldinet al. 1988).
Clinical Applications
Peripheral neuropathies are common neurological dis-orders of the community with an annual incidenceof 100–200 per 100,000 people. Toxic neuropathiesaccount for a minor proportion of etiology (–5%)depending on economical and medical factors and ge-ographic distribution. In the 1950-1970’s, most toxicneuropathies have been related to outbreaks of indus-trial chemicals, such as acrylamide and organic solvents(hexacarbons). The list of these etiologies depends onthe industrial development of different countries. Overthe past decades, major outbreaks of toxic neuropathyhave decreased to a great extent due to stricter regulationof chemicals used in industry.In recent years, most reported toxic neuropathies havebeen due to side effects of medications or from acciden-tal exposure. Common chemotherapeutic agents caus-ing toxic neuropathies include cis-platinum, taxol, andthalidomide. Thus, toxic neuropathies are relatively un-common compared to other etiologies, such as diabetesmellitus, genetic neuropathies and inflammatory disor-ders. Nevertheless, toxic neuropathies have great impacton environmental and industrial regulation, and serve asimportantmodels for investigating mechanismsofnerveinjury and the correlation between pathology and clin-ical manifestations.
Clinical Manifestations of Toxic Neuropathies
Functionally, the peripheral nervous system consistsof motor, sensory, and autonomic nerve fibers (Fig. 1).Motor nerves and proprioceptive nerves, which conveysensory information from bones, muscles, and joints,are large � myelinated nerves (large-diameter nervesor � large fibers). Nociceptive, thermal, and autonomicnerves belong to small myelinated nerves or unmyeli-nated nerves (small-diameter nerves or � small fibers).All of these nerve fibers have different cytoskeletalorganizations, origins, and terminals (Chen et al. 1999).The severity of neurological deficits depends on severalfactors, including the vulnerability of each nerve fibertype, the duration of toxin exposure, the total dose oftoxins, and the extent of nerve degeneration. A majorfactor in determining clinical presentations is the differ-ential susceptibility of neurons to toxins or pathologicprocesses, a characteristic of neurological diseases: forexample, motor neurons in amyotrophic lateral sclerosis(Lou Gehrig’s disease), cortical neurons in Alzheimer’sdisease, and substantia nigra neurons in Parkinson’sdisease. Each disease has a distinct pattern of clinicalpresentation because certain parts of the nervous systemare selectively damaged.Clinical manifestations of neuropathies, therefore, de-pend on the damage to different fiber types (Thomas

T
Toxic Neuropathies 2495
Toxic Neuropathies, Figure 1 Structural organization of peripheral nerves. Peripheral nerves consist of: (1) motor nerves from ventral horn motorneurons of the spinal cord (black), (2) proprioceptive nerves from large-diameter sensory neurons of dorsal root ganglia which terminate in skeletalmuscles, tendons, and joints (olive), and (3) nociceptive and thermal nerves from small-diameter sensory neurons of dorsal root ganglia which terminatein the skin (pink) and autonomic nerves from autonomic ganglia innervating smooth muscles and glands (blue).
and Ochoa 1993). When motor nerve fibers are in-jured, neurological deficits range from mild weaknessof distal limb muscles to marked paralysis of all fourlimbs. When sensory nerves responsible for joint move-ments are injured, unsteadiness during walking maydevelop. This type of neuropathy is sometimes called� Large Fiber Neuropathy. On the other hand, in pa-tients with damaged sensory nerves, neuropathic pain,such as tingling or electric-shock sensations can beexperienced. Various autonomic complaints, such astachycardia, constipation, or diarrhea can be presentingsymptoms in patients with damaged autonomic nerves.Sensory neuropathy with painful features and loss ofnociceptive functions and autonomic neuropathy aretermed as � small-fiber neuropathy. Often, neurolog-ical presentations are combined effects due to deficitsof different fibers types: for example, a patient withmotor, sensory, and autonomic neuropathy may havemanifestations of weakness, sensory disturbances, anddysautonomia. Various etiologies of toxic neuropathieswith their pathologic mechanisms and manifestationsare summarized below. This list is not intended to bea comprehensive catalogue, and readers can refer tovarious sources for detailed information (Spencer andSchaumburg 2000).
Classification of Toxic Neuropathies:Sources
• Environmental contaminants: lead• Occupational outbreaks: n-hexane• Adverse effects of medications: cisplatin• Accidental exposure or ingestion: lead• Experimentaluses:e.g.hexacarbons,2,5-hexanediol,
acrylamide, capsaicin
Functional components
• Motor neuropathy: lead• Sensory neuropathies
– Proprioceptive type:cis-platinum,vincristine, iso-niazid
– Nociceptive type: capsaicin
• Autonomic neuropathy: acrylamide, alcoholic, ar-senic (inorganic), diphtheritic toxin
Clinical presentations
• Weakness• Sensory disturbances: neuropathic pain, hypesthesia• Dysautonomia

2496 Toxic Neuropathies
Structural components
• Large myelinated nerves
– Motor nerves: acrylamide– Sensory nerves (proprioception): n-hexane
• Small myelinated nerves
– Sensory nerves (thermal sense, nociception): cap-saicin
– Autonomic nerves: acrylamide, alcoholic, arsenic(inorganic), diphtheritic toxin
• Unmyelinated nerves
– Sensory nerves (thermal senses, nociception):capsaicin
– Autonomic nerves: acrylamide, alcoholic, arsenic(inorganic), diphtheritic toxin
Sites of actions and mechanisms
• Proximal axons: filamentous swelling
– β,β’-iminodipropionitrile
• Terminal portion: swelling and degeneration
– Neuromuscular junctions: 2,5-hexanediol, acry-lamide
– Pacinian corpuscles: 2,5-hexanediol– Epidermalnervesof theskin:acrylamide,cisplatin
Pathology
• Primarily axonal degeneration: hexacarbon• Primarily demyelination: tellurium
The terminals of the longest nerves are the most sus-ceptible part during toxin exposure, and clinical symp-toms usually develop in the corresponding innervatedareas of these damaged nerves in the early phase of toxicneuropathy. In addition, clinical symptoms and signs areusually more severe in regions with early-onset symp-toms than in regions with late-onset symptoms. For ex-ample, numbness and neuropathic pain may appear inthe toes and feet earlier than in the legs and thighs. Sim-ilarly, symptoms in the fingers and hands are earlier thanthose in the forearms. Neurological deficits are usuallymore severe in the lower limbs than in the upper limbs.This “glove-stocking” type of distribution is character-istic of length-dependent neuropathies, particularly forthe majority of toxic neuropathies, although exceptionsmay exist in certain types of toxic neuropathies.
Therapeutic Consequences
Evolution of Pathology in Toxic Neuropathies
The typical pathology of toxic neuropathies of large-diameter motor and sensory nerves has been demon-
strated in both humans and experimental animals overthe past few decades: for example, neuropathies dueto exposure to hexacarbons, particularly the activecomponents, 2,5–hexanediol, acrylamide, and carbondisulfide. The longest and largest nerves are affectedearlier, with the major pathology in the terminal partsof axons, i.e. � distal axonopathy. Intoxication resultsfrom absorption of organic solvents in poorly venti-lated environments. Fig. 2 illustrates the scenario ofpathology in neuromuscular junctions after hexacar-bon and acrylamide intoxication (Ko et al. 1999). Inanimals intoxicated with such compounds as, for ex-ample, acrylamide, there is no visible weakness orataxia during the early phase. At that stage, however,motor nerve terminals have already begun to swell up.Obvious weakness and ataxia of hind limbs graduallydevelops. The weakness and unsteadiness progress atvariable speeds, and eventually the forelimbs are af-fected, resulting in quadriparesis at the late stage. Asintoxication proceeds, axonal swelling extends fromjunctional folds into the intramuscular nerves, whichresults in Wallerian-like degeneration of motor nervesand denervation of neuromuscular junctions (Fig. 2a).Similar changes also develop in the central terminals oflong axons, such as terminals of the posterior column atthe brainstem, the central axons of dorsal root ganglia,thus the term, � central-peripheral distal axonopathy,or � dying-back neuropathy (LoPachin 2000).
Nociceptive Nerves in Toxic Neuropathies
Nociceptive nerves subserving thermal sensations areAδ or C fibers based on physiological classification,with diameters in the range of 1–5 μm. These axons,with � free nerve endings, are peripheral processes ofsmall neurons in � dorsal root ganglia, and terminatein the most-superficial layer of the skin, the epidermis(Fig. 1). Their central processes end in the dorsal hornand synapse with spinothalamic and other nociceptiveneurons. Due to their small size, these nerve fibershave traditionally been studied with high-resolution� electron microscopy. The application of sensitive� immunohistochemistry with various neuronal mark-ers has enabled the evaluation of skin innervation ata global scale (Hsieh et al. 2000). Among these neu-ronal proteins, protein gene product 9.5, an ubiquitinC-terminal hydrolase, is particularly useful for demon-strating the rich innervation of the epidermis (Fig. 2b).Based on technical improvements, it is now possibleto address the issue of neuropathy of small-diameternerves due to toxin exposure.� Capsaicin is an active compound from hot peppers,which is known to activate the vallinoid receptor. A ma-jor use of capsaicin ointment is to treat neuropathic pain;its efficacy probably occurs through damage to sensorynerve endings. Systemic injection of capsaicin in neona-tal rats abolishes primary afferent terminals in the dorsalhorn of the spinal cord (Wall et al. 1982). In human stud-

T
Toxic Neuropathies 2497
Toxic Neuropathies, Figure 2 Diagram of the progression of toxic neuropathy. The diagram illustrates degeneration of nerve terminals in neuromuscularjunctions (a) and in the epidermis of the skin (b) during the evolution of toxic neuropathy with acrylamide-induced neuropathy as an example. Axons andnerve terminals were immunostained with protein gene product 9.5 (dark brown), and neuromuscular junctions with substrates of cholinesterase (blue).(a) In normal muscles, motor nerve terminals travel into every fold of the neuromuscular junction (NMJ) with quite-uniform thickness. In the early phaseof degeneration, focal swelling of nerve terminals in the neuromuscular junctions develops, and axonal swelling extends into the motor nerve trunks.Finally in the late stage, neuromuscular junctions are denervated, and motor nerves become degenerated with a segmented appearance. (b) Bundles ofnerve fibers form subepidermal nerve plexuses (SNP) in the dermis (DERM) below the epidermis (EPI) in normal skin. Epidermal nerves arise from thesubepidermal nerve plexuses, and ascend vertically in the epidermis with a varicose appearance. In the early phase of degeneration, epidermal nervesbecome swollen, and branching increases. During the late phase, epidermal nerves disappear, and the epidermis becomes denervated. Subepidermalnerve plexuses have a fragmented appearance, indicating that they are undergoing degeneration.
ies, capsaicin causes degeneration of nerve terminals inthe epidermis after local application, with correspond-ing loss of thermal sensations in the application area (Si-mone et al. 1998). A certain degree of epidermal nerveregeneration can later be observed, suggesting that themajor target of capsaicin-induced nerve damage by lo-cal treatment is at the terminal portion, instead of at theneuronal cell body.Traditionally, acrylamide and cisplatin have been con-sidered to cause neuropathy of large-diameter nerves.The development of staining for cutaneous nerve ter-minals allowed the influence of acrylamide on small-diameter sensory nerves in the skin to be assessed (Fig.2b). At the initial stage of intoxication, � epidermalnerves show two major changes: terminal swelling andincreased branching (Ko et al. 2002). There is a progres-sive reduction in epidermal nerve density thereafter. Atthe late stage, there is significant dermal nerve degen-eration with ultrastructural demonstration of vacuolarchanges. These findings have established the patho-logical consequences of acrylamide neurotoxicity incutaneous sensory nerves for studying the “dying-back”pathology of nociceptive nerves. These phenomena arequite similar to the pathology of cutaneous nervesin human skin, including epidermal nerve swelling,increased branching points of epidermal nerves, andfragmentation of dermal nerve fibers.Neuropathic pain is an important manifestation of sometoxicneuropathies, such ascis-platinum and vincristine-induced neuropathies. The exact mechanisms remainobscure. Responses to nerve injury include neuronaland glial responses at the levels of transcription andpost-translation, re-organization of neuronal and ax-
onal structures and changes in synaptic plasticity. Forexample, different subtypes of sodium channels areup-regulated or down-regulated in animals with neu-ropathic pain. In damaged nerves, sodium channelsare re-distributed along the entire axons or accumulatein the neuroma, instead of clustering around the nodeof Ranvier in normal axons. Many of these changesmay contribute to the generation and maintenance ofneuropathic pain.Our knowledge of toxic neuropathies has broadlyexpanded over the past decade because many newinvestigative techniques have been developed, particu-larly toxic neuropathies of nociceptive nerves, whosedysfunctions cause diverse manifestations of neuro-pathic pain. The identification of environmental toxinsas etiologies of neuropathies provides new insights intomechanisms of nerve degeneration and its consequentneuropathicpain. Agood example is2,5-hexanediol, theactive metabolic product of organic solvents containinghexacarbons. The recognition of hexacarbon-inducedcentral-peripheral distal axonopathy has provided anew model and opened a new field to understand nervedegeneration. The causes of a considerable proportion(10–20%) of peripheral neuropathies have not yet beenidentified. Among these neuropathies, toxin-exposurehas rarely been explored. Detailed investigation of ex-posure history is mandatory, and may offer a new lookat some neuropathies of unknown etiology.
References1. Bouldin TW, Samsa G, Earnhardt TS, Krigman MR (1988)
Schwann Cell Vulnerability to Demyelination is Associatedwith Internodal Length in Tellurium Neuropathy. J NeuropatholExp Neurol 47:41-47

2498 Traction
2. Chen WP, Chang YC, Hsieh ST (1999) Trophic Interactions be-tween Sensory Nerves and the Targets. J Biomed Sci 6:79-85
3. Griffin JW, Fahnestock KE, Price DL, Cork LC (1983) Cy-toskeletal Disorganization Induced by Local Application ofbeta, beta’–iminodipropionitrile and 2,5–hexanedione. AnnNeurol 14:55-61
4. Griffin JW, George EB, Hsieh ST, Glass JD (1995) Axonal De-generation and Disorders of the Axonal Cytoskeleton. In: Wax-man SG, Kocsis JD, Stys PK (eds) The Axon, Oxford UniversityPress, New York, pp 375-390
5. Hirokawa N (1998) Kinesin and Dynein Superfamily Proteinsand the Mechanism of Organelle Transport. Science 279:519-526
6. Hsieh ST, Chiang HY, Lin WM (2000) Pathology of Nerve Termi-nal Degeneration in the Skin. J Neuropathol Exp Neurol 59:297-307
7. Ko MH, Chen WP, Hsieh ST (2002) Neuropathology of SkinDenervation in Acrylamide-Induced Neuropathy. Neurobiol Dis11:155-165
8. Ko MH, Chen WP, Lin-Shiau SY, Hsieh ST (1999) Age-Dependent Acrylamide Neurotoxicity in Mice: Morphology,Physiology, and Function. Exp Neurol 158:37-46
9. LoPachin RM (2000) Redefining Toxic Distal Axonopathies.Toxicol Lett 112-113:23-33
10. Schaumburg HH, Wisniewski HM, Spencer PS (1974) Ultra-structural Studies of the Dying-Back Process. I. Peripheral NerveTerminal and Axon Degeneration in Systemic Acrylamide In-toxication. J Neuropathol Exp Neurol 33:260-284
11. Simone DA, Nolano M, Johnson T, Wendelschafer-Crabb G,Kennedy WR (1998) Intradermal Injection of Capsaicin in Hu-mans Produces Degeneration and Subsequent Reinnervation ofEpidermal Nerve Fibers: Correlation with Sensory Function. JNeurosci 18:8947-8959
12. Spencer PS, Schaumburg HH (2000) Experimental and ClinicalNeurotoxicology. Oxford University Press, New York
13. Thomas PK, Ochoa J (1993) Clinical Features and DifferentialDiagnosis. In: Dyck PJ, Thomas PK, Griffin JW, Low PA, PodusloJF (eds) Peripheral Neuropathy, W.B.Saunders, Philadelphia, pp749-774
14. Vinik AI, Park TS, Stansberry KB, Pittenger GL (2000) DiabeticNeuropathies. Diabetologia 43:957-973
15. Wall PD, Fitzgerald M, Nussbaumer JC, Van der Loos H, DevorM (1982) Somatotopic Maps are Disorganized in Adult RodentsTreated Neonatally with Capsaicin. Nature 295:691-693
TractionSUSAN MERCER
School of Biomedical Sciences, University ofQueensland, Brisbane, QLD, [email protected]
Synonyms
Intermittent; progressive adjustive; manual or continu-ous traction; autotraction; gravity-assisted traction; selftraction; unloading
Definition
Traction is a treatment for spinal pain. It involves ap-plying a pulling force to the lower limbs or to the head,in order to separate the vertebrae of the spine and / orto stretch the surrounding muscles and ligaments.The traction force may be delivered manually, or viaweights, pulleys or mechanical devices as a continuous,sustained, intermittent or intermittent pulsed force.
Characteristics
Mechanism
No mechanism has been established whereby tractionmight relieve pain. Nevertheless, proponents of tractionbelieve that it works, and have speculated on its mech-anism of effect (Krause et al. 2000).There is clear evidence that spinal traction does sepa-ratevertebralbodies.However, in the lumbarspinemuchof the separation observed arises from flattening of thelumbar lordosis (Twomey 1985). In the cervical spine,30 pounds of traction achieves only fractions of a mil-limetre separation between vertebral bodies, amountingto 2 mm total elongation anteriorly and 6 mm posteri-orly between C2 and T1 (Colachis and Strohm 1969).The purported benefit of this separation is, however, am-biguous.When traction is used to treat radicular pain, the im-plicitmechanism isdecompression of theaffected spinalnerve. However, separation of vertebrae increases thelongitudinal dimension of the intervertebral foram-ina but longitudinal compression of spinal nerves isan uncommon phenomenon. Most commonly, spinalnerves are affected in the sagittal dimension, anteriorlyby disc herniations or osteophytes or posteriorly byosteophytes of the zygapophysial joints. Longitudinalseparation does not relieve encroachment in the sagittaldimension. Moreover, upon the patient resuming theupright posture, any benefit of traction is immediatelylost, as gravity restores the compression load on thespine. Indeed, its has been shown that, without rising,after simply resting on the traction table for 20 min, theeffects of cervical traction are all but lost (Colachis andStrohm 1969).Another conjecture is that traction reduces disc her-niations. The available data, however, are limited andconflicting. In one small study, although traction didreduce disc herniations in two of three patients, theprotrusions reappeared within 14 min after release ofthe force (Matthews 1968).In the absence of firm evidence for a mechanical effectof traction, some authorities have pursued alternativerationales, such as silencing ectopic impulse genera-tors and normalisation of conduction in spinal nerves(Krause et al. 2000). These speculations, however, nev-ertheless presuppose that traction reverses compressionof the spinal nerve by separating the vertebral bodies.They also require that relatively brief traction somehowachieves lasting reversal of the pathophysiology thatcauses pain.There iseven lessofaphysiological rationale for tractionwhen it is applied for spinal pain, as opposed to radicu-lar pain. In the first instance, nerve root irritation causespain in the limbs, not in the back or in the neck. The ratio-nalefor tractionforspinalpain, therefore,cannot involvedecompression of spinal nerves. Instead, it has been pro-posed that perhaps spinal pain might be relieved by “in-

T
Traditional Pharmacological Pain Relief 2499
creasing non-nociceptive input and recruitment of de-scending inhibition” (Krauseetal. 2000).Whileperhapsattractive as a conjecture, such a statement falls shortof actually constituting evidence of how traction mightrelieve spinal pain. Another proposition is that tractionserves to stretch spinal tissues (Krause et al. 2000),but inthat event it is questionable whether elaborate and pas-sive traction offers any advantage over simple stretch-ing exercises that the patients can undertake themselves.The proposition that traction reduces intervertebral discpressures is confounded not only by the lack of experi-mental data that demonstrate this effect, but also by thelack of a cogent theory as to how raised disc pressurecauses back pain and why that pain should stay relievedonce the traction is released and the patient resumes anupright posture.
Applications
Traction has been used to treat neck pain, cervical radic-ular pain and radiculopathy, back pain and lumbar radic-ular pain and radiculopathy. Some practitioners use it totreat thoracic spinal pain, but there is no literature on itsefficacy for this condition.Traction can be applied manually by the therapist (man-ual traction), by amotorizedpulley(motorised lumbarorcervical traction), by the patients themselves providingthe pulling force (autotraction) or by suspension from adevice (gravitational traction) (Twomey 1985).
Efficacy
Systematic reviews have been confounded by the irregu-lar and inconsistentdiagnosticcriteriaused in thestudiesreviewed. It has not always been evident if the patientshadsomaticpain, somatic referredpainor radicularpain.Although some earlier reviews offered open or encour-aging conclusions, these have been supplanted by sub-sequent studies and reviews.No randomized controlled trials have shown if tractionis effective for neck pain (Aker et al. 1996; Harms-Ringdahl and Nachemson 2000). The available studiesindicated that it is not effective for acute neck pain(Bogduk and McGuirk 2006). For chronic neck pain,traction has not been subjected to scientific studies. Forcervical radicular pain, studies have shown that tractionis no more effective than sham traction or placebo treat-ment (British Association of Physical Medicine 1966;Goldie and Landquist 1970; Klaber et al. 1990). Thesame conclusions apply for the treatment of lumbarradicular pain (Coxhead et al. 1981; Matthews andHickling 1975; Pal et al. 1986). For non-specific lowback pain, traction is no more effective than a placebotreatment (Beursken et al. 1997).
Indications
In the face of the available scientific evidence, there areno legitimate indications for traction in themodernera. It
isatreatment,steepedintraditionbutdevoidofevidence.� Lumbar traction
References1. Aker PD, Gross AR, Goldsmith CH et al. (1996) Conservative
management of mechanical neck pain: systematic overview andmeta-analysis. BMJ 313:1291–1296
2. Beurskens AJ, de Vet HC, Koke AJ et al. (1997) Efficacy oftraction for non-specific low back pain: 12-week and 6-monthresults of a randomized clinical trial. Spine 22:2756–2762
3. Bogduk N, McGuirk B (2006) Medical Management of Acuteand Chronic Neck Pain. An Evidence-Based Approach. Elsevier,Amsterdam (in press)
4. British Association of Physical Medicine (1966) Pain in the neckand arm: a multicentre trial of the effects of physiotherapy. BMJ1:253–258
5. Colachis SC, Strohm BR (1969) Effects of intermittent trac-tion on separation of lumbar vertebrae. Arch Phys Med Rehabil44:251–258
6. Coxhead CE, Inskip H, Meade TW et al. (1981) Multicentre trialof physiotherapy in the management of sciatic symptoms. Lancet1:1065–1068
7. Goldie I, Landquist A (1970) Evaluation of the effects of differentforms of physiotherapy in cervical pain. Scand J Rehab Med2–3:117–121
8. Harms-Ringdahl K, Nachemson A (2000) Acute and subacuteneck pain: nonsurgical treatment. In: Nachemson A, Jonsson E(eds) Neck and Back Pain: The Scientific Evidence of Causes,Diagnosis, and Treatment. Lippincott Williams & Wilkins,Philadelphia, pp 327–338
9. Harte AA, Baxter GD, Gracey JH (2203) The efficacy of tractionfor back pain: a systematic review of randomized controlled trials.Arch Phys Med Rehabil 84:1542–1553
10. Klaber Moffett JA, Hughes GI, Griffiths P (1990) An investiga-tion of the effects of cervical traction. Part 1: clinical effective-ness. Clin Rehab 4:205-211
11. Krause M, Refshauge KM, Dessen M et al. (2000) Lumbar spinetraction: evaluation of effects and recommended application fortreatment. Manual Therapy 5:72–81
12. Matthews JA (1968).Dynamic discography: a study of lumbartraction. Ann Phys Med 9:275–279
13. Matthews JA, Hickling J (1975) Lumbar traction: a double-blindcontrolled study for sciatica. Rheumatol Rehabil 14:222–225
14. Pal B, Mangion P, Hossain MA et al. (1986) A controlled trialof continuous lumbar traction in the treatment of back pain andsciatica. Brit J Rheumatol 25:181–183
15. Twomey LT (1985) Sustained lumbar traction. An experimentalstudy of long spine segments. Spine 10:146–149
Tractus Trigeminothalamicus
� Trigeminothalamic Tract Projections
Traditional Pharmacological Pain Relief
Definition
Pain relieved by oral and subcutaneous injections ofanalgesic drugs when the patient demands pain relief.Often ineffective in that doses are too low and dosingintervals are too long.� Postoperative Pain, Acute Pain Team

2500 Trafficking
Trafficking
Definition
Trafficking refers to synthesis and targeting of proteinsto specific locations within the cell.� Opioid Receptor Trafficking in Pain States� Trafficking and Localization of Ion Channels
Trafficking and Localization of IonChannels
PETER SHRAGER1, MATTHEW N. RASBAND2
1Deptartment of Neurobiology and Anatomy,University of Rochester Medical Center, Rochester,NY, USA2University of Connecticut Health Center, Farmington,CT, [email protected], [email protected]
Synonyms
Voltage-Dependent Pores; Clustering; Targeting; IonChannel Trafficking
Definition� Ion channels are pore-forming proteins in the surface
membrane through which electrical charges moveto establish and alter the membrane potential. Thesechannels must be synthesized and inserted into themembrane in the proper locations within the neuronin order for signaling to occur normally.
Characteristics
Neurons are highly polarized cells, and during devel-opment individual neurites become either dendritesor axons. An essential element in this polarization isthe proper � trafficking of proteins to these processes,and, in particular, the targeting of ion channels to spe-cific locations. In dendrites, ligand-gated channels mustaccumulate at postsynaptic regions. In axons, Na+ chan-nels are clustered at the initial segment, which is thesite of integration of postsynaptic potentials and of ini-tiation of the action potential. Na+ and � K+ channelsare distributed at low density throughout the remainderof the fiber and, if the axon is myelinated, they arealso clustered within specific zones in the region of thenode of Ranvier. Ectopic localization of these channelscan lead to repetitive or spontaneous firing, and canthus contribute to pain. Localization may be controlledby domains within the protein that direct transport orinsertion to specific sites, or by differential rates ofendocytosis. For example, a protein may be axonalbecause its endocytotic removal rate is much higher indendrites. There has been much progress recently inuncovering these mechanisms.
Multiple targeting domains have been discovered thatdirect K+ channels to different sites within the neuron.The differential targeting of Kv2.1 and Kv2.2 to alter-native dendritic regions is governed, at least in part,by a 26 amino acid � motif on the cytoplasmic tail ofKv2.1. Manganas et al. (2001) have found that specificresidues within the pore region of Kv1.× control sur-face expression of those channels. Voltage-dependention channels are typically heteromultimers consist-ing of a pore-forming alpha subunit and one or morebeta subunits that modulate gating. Kv channel betasubunits are bound at the cytoplasmic surface of thechannel. It was recently found that beta subunits alsoparticipate in localization. Axonal targeting of Kv1.2 inhippocampal neurons is driven by the tetramerizationdomain on the N-terminus, at a site that binds Kvbeta2(Gu et al. 2003). Kvbeta subunits have a binding pocketfor NADP+, and mutation of this site eliminates theiraxonal targeting capability (Campomanes et al. 2002).Nodal regions have 3 major zones: the nodal gap; para-nodes in which axoglial junctions link the paranodalloops to the axolemma; and the juxtaparanodes, zonesthat flank the paranodes, begin the internode, and haveno special morphological characteristics. At nodal re-gions KCNQ2 and KCNQ3, slowly activating channelsthat can be modulated by neurotransmitters, are foundwithin the nodal gap (Devaux et al. 2004). Kv1.1 andKv1.2 are clustered in the juxtaparanodes in both thePNS and CNS. The localization of Kv1.× is dependenton the integrity of the axoglial paranodal junctions.When the latter are disrupted, these K+ channels arefound in the paranodes and diffusely throughout theinternode.At both axon initial segments and � nodes of Ranvier,voltage-dependent � Na + � channels are linked tothe spectrin-actin cytoskeleton by the adapter proteinankyrinG. AnkyrinG also serves to link Na+ chan-nels in a large molecular complex that includes theL1–family proteins NrCAM and neurofascin. Thebeta subunits of Na+ channels are membrane proteinsand they, along with NrCAM, neurofascin, and con-tactin (a GPI-anchored protein) are all members ofthe immunoglobulin superfamily, implicating them inintercellular (perhaps neuron-glial) signaling. The linkto ankyrinG also appears to be important in targeting.In spinal motor neurons in culture, Na+ channels arenormally synthesized in the soma, but are inserted inthe surface membrane only at the axon initial segment(Alessandri-Haber et al. 1999). Nav1.2 has a C-terminalmotif that is involved in axonal targeting. It has alsorecently been demonstrated that the intracellular linkerbetween domains II and III servesasa localization signalto the initial segment (Garrido et al. 2003). Of particularinterest, Lemaillet et al. (2003) showed that this latterloop contains an ankyrinG binding motif. Thus, estab-lishment of these regions of high Na+ channel densityappears to involve targeting to the axon and insertion in

T
Trafficking of Proteins 2501
the surface membrane, followed by cytoskeletal immo-bilization via ankyrinG. In the case of nodes of Ranvier,there is an additional requirement for glial modelingof the axonal surface. Myelinating Schwann cells inthe PNS direct the clustering of Na+ channels, per-haps by reorganizing the low density of these channelspresent throughout the axon prior to myelin formation(Dugandzija-Novakovic et al. 1995). The neuron-glialcommunication involved in this trafficking may involvethe other members of the Na+ channel complex men-tioned above (Custer et al. 2003). In the CNS, someearly aspects of clustering may involve soluble factorsreleased by oligodendrocytes, but final node formationseems to be contact dependent, as in the PNS (Kaplan etal. 2001). Finally, there is a developmentally regulatedprogression in Na+ channel subtype in axons. Nav1.2 isexpressed early, and is later replaced by Nav1.6 (Boikoet al. 2001). The physiological consequences of thisshift are not known, but Nav1.6 has a unique resurgentcurrent (Raman and Bean 1997), which could renderaxons more susceptible to re-entry excitation that maycontribute to pain, if other regulatory zones of nodesare compromised. For example, loss of juxtaparanodalKv1.× can lead to instabilities.Turnover rates of ion channels in myelinated axonshave not been measured directly, but some data areavailable. Nodal Na+ channels are particularly stable,and clusters can be detected up to 9 days after myelindisruption (Custer et al. 2003). This is undoubtedlydue to the cytoskeletal link via ankyrinG. The stabi-lization of these channels seems to occur graduallyafter their initial clustering, and may involve the forma-tion of large complexes through the multiple bindingsites of ankyrinG (Custer et al. 2003). JuxtaparanodalKv1.× channel clusters are significantly less stable, andbreak up within 1–2 days after initiating demyelination(Rasband et al. 1998).There is considerable evidence that altered ion channeltrafficking, localization, and turnover are involved inpainful neuropathies, although the precise mechanismsremain to be determined. Painful neuromas developwithin peripheral nerves after axonal injury, and thespontaneous discharge at these sites is thought to re-sult from an accumulation of Na+ channels (Devor etal.1989).Further, thereisevidencethatspecificsubtypesof Na+ channels are involved, including PN1 and PN3.The Na+ channels accumulating at neuromas are co-localized with ankyrinG (Kretschmer et al. 2002), andthe increased levels of both proteins, may reflect dys-regulation of trafficking and/or turnover rates. As moreinformation becomes available on the neurobiologicalmechanisms regulating ion channel biosynthesis, traf-ficking, and targeting, it will be interesting to apply theseprinciples to models of neuropathic pain. For example,if some forms of neuropathic pain are a consequenceof altered Na+ channel trafficking, then therapeuticstrategies designed to disrupt or perturb the trafficking
of these proteins may prove useful in the treatment ofthese disorders.
References1. Alessandri-Haber N, Paillart C, Arsac C, Gola M, Couraud F,
Crest M (1999) Specific Distribution of Sodium Channels in Ax-ons of Rat Embryo Spinal Motoneurones. J Physiol 518:203–214
2. Boiko T, Rasband MN, Levinson SR, Caldwell JH, Mandel G,Trimmer JS, Matthews G (2001) Compact Myelin Dictates theDifferential Targeting of Two Sodium Channel Isoforms in theSame Axon. Neuron 30:91–104
3. Campomanes CR, Carroll KI, Manganas LN, Hershberger ME,Gong B, Antonucci DE, Rhodes KJ, Trimmer JS (2002) Kv BetaSubunit Oxidoreductase Activity and Kv1 Potassium ChannelTrafficking. J Biol Chem 277:8298–8305
4. Custer AW, Kazarinova-Noyes K, Sakurai T, Xu X, SimonW, Grumet M, Shrager P (2003) The Role of the Ankyrin-Binding Protein NrCAM in Node of Ranvier Formation. JNeurosci 23:10032–10039
5. Devaux JJ, Kleopa KA, Cooper EC, Scherer SS (2004) KCNQ2is a Nodal K+ Channel. J Neurosci 24:1236–1244
6. Devor M, Keller CH, Deerinck TJ, Levinson SR, Ellisman MH(1989) Na+ Channel Accumulation on Axolemma of AfferentEndings in Nerve End Neuromas in Apteronotus. NeurosciLett 102:149–154
7. Dugandzija-Novakovic S, Koszowski AG, Levinson SR, ShragerP (1995) Clustering of Na Channels and Node of Ranvier For-mation in Remyelinating Axons. J Neurosci 15:492–502
8. Garrido JJ, Giraud P, Carlier E, Fernandes F, Moussif A, FacheMP, Debanne D, Dargent B (2003) A Targeting Motif Involvedin Sodium Channel Clustering at the Axonal Initial Segment.Science 300:2091–2094
9. Gu C, Jan YN, Jan LY (2003) A Conserved Domain in AxonalTargeting of Kv1 (Shaker) Voltage-Gated Potassium Channels.Science 301:646–649
10. Kaplan MR, Cho MH, Ullian EM, Isom LL, Levinson SR, Bar-res BA (2001) Differential Control of Clustering of the SodiumChannels Na(v)1.2 and Na(v)1.6 at Developing CNS Nodes ofRanvier. Neuron 30:105–119
11. Kretschmer T, England JD, Happel LT, Liu ZP, Thouron CL,Nguyen DH, Beuerman RW, Kline DG (2002b) Ankyrin Gand Voltage Gated Sodium Channels Co-Localize in HumanNeuroma-Key Proteins of Membrane Remodeling after AxonalInjury. Neurosci Lett 323:151–155
12. Lemaillet G, Walker B, Lambert S (2003) Identification of a Con-served Ankyrin-Binding Motif in the Family of Sodium ChannelAlpha Subunits. J Biol Chem 278:27333–27339
13. Manganas LN, Wang Q, Scannevin RH, Antonucci DE, RhodesKJ, Trimmer JS (2001) Identification of a Trafficking Determi-nant Localized to the Kv1 Potassium Channel Pore. Proc NatlAcad Sci USA 98:14055–14059
14. Raman IM, Bean BP (1997) Resurgent Sodium Current and Ac-tion Potential Formation in Dissociated Cerebellar Purkinje Neu-rons. J Neurosci 17:4517–4526
15. Rasband MN, Trimmer JS, Schwarz TL, Levinson SR, EllismanMH, Schachner M, Shrager P (1998) Potassium Channel Distri-bution, Clustering, and Function in Remyelinating Rat Axons.J Neurosci 18:36–47
Trafficking of Proteins
Definition
Proteins are synthesized on ribosomes in the cell body,packaged into vesicles in the Golgi apparatus, and thentransported along cytoskeletal structures to their final lo-

2502 Training by Quotas
cation in the cell. The process of targeted transport istrafficking.� Peripheral Neuropathic Pain� Trafficking and Localization of Ion Channels
Training by QuotasWILBERT E. FORDYCE
Department of Rehabilitation, University ofWashington School of Medicine, Seattle, WA, [email protected]
Synonyms
Exercise; quotas; contingency management
Definition
Training by quota describes methods for increasing ex-ercise and activity level by use of the quota system, aform of contingency management based on behavioralscience. The methods make use of sensitivity to our en-vironments to improve our state. Review of the theoret-ical and conceptual background can be found in texts byFordyce (1976, 1990). “How” to do it will be described.PINPOINT the � behavior to be changed (i.e. specifyin � movement cycles, walking-steps, exercise repeti-tions), RECORD the pre-intervention rate of the behav-ior and CONSEQUATE, i.e. spell out the rate of rein-forcement and by whom, always with the patient’s in-formed consent.
Characteristics
The term chronic pain encompasses many conditions.Increasing exercise and / or activity level is not indicatedfor all of them. Evaluation of a problem of chronic pain(ideally a multi-disciplinary, multi-modal process) in-dicating increased exercise or activity is clearly to thepatient’s benefit points to using these methods. Quotamethods help patients increase exercise or activity lev-els. They are not methods for treating pain. Increases inactivity level help restore patient access to activities usu-ally engaged in.When target levelsofactivity orexerciseare reached, the program can be faded to maintenancelevels or stopped altogether. Re-entry into sustaining ac-tivities should be achieved beforehalting or reducing ex-ercise levels.� Reinforcers are usually simple to define and ap-
ply in the practical case, using the Premack Principle(Premack 1959). This states that high strength behav-iors reinforce low strength behaviors; paraphrased as,what a person does a lot of can be used to reinforceactivities or movements targeted to be increased.Observing the consequences of what the person doesa lot can usually readily identify effective reinforcers.To a restless person frequently on the move, activity,
a high strength behavior, is likely to be an effective re-inforcer. If programmed or scheduled carefully, it canserve to enhance or reinforce exercise. To an inactiveperson, one who moves relatively little, activity isunlikely to be reinforcing but rest or “time out” fromactivity is. In the context of pain management, restor “time out” from activity and attention or encour-agement of important others around the person (e.g.the therapist) usually suffice as reinforcers, whenprogrammed appropriately.
Baseline
Baseline is the starting point or rate (i.e. number of rep-etitions) at which the target behavior is occurring, e.g.number of steps walked beforeweakness, pain or fatiguecauses stopping. If the target behavior is not in the per-son’s present repertoire, establishing it by teaching or“shaping” is indicated (Fordyce 1976).Walking, performing selected exercises, engaging in ac-tivities intended to diminish reclining time and increas-ing activity are common targets for behavior change.Talking about pain, grimacing, asking for and / or takinganalgesics are less frequently targets for change andshould become so only with careful consideration ofappropriateness to the patient’s condition and with fullinformed consent.
Quota
Once baseline performance is identified, set quotas to beapplied foreach trial.The initialquota should beapprox-imately 2
3 or ¿ of baseline, e.g. laps walked = 7, initialquota = 4 or 5, baseline of an exercise repetition = 5,initial quota = 2 or 3. Initial quota level should be deter-mined by confidence that it is an amount well within thepatient’s current repertoire, ensuring successes at earlytrials. It is better to have too low an initial quota and besure of success than too high and a risk of failure. Quotasare then incremented at a rate driven mainly by an as-cending performance curve confidently expected to pro-vide several early succeeding trials before initial base-line is reached. In short, “success breeds success.”
Quota Increment Rates
Most commonly, incrementing by 1 repetition per trialor session isappropriate. Incasesofmarginallyadequateability to increment, a slower rate (e.g. 1 × each 2 or 3trials) may suffice. An amount easily performed may in-dicate increments of 2 or more with each trial, but dothat only if confident that several successful sessions areachieved prior to reaching baseline. In all cases, full in-formed consent prior to performance is indicated. Quotaincrement rates should be spelled out to the patient fol-lowing their determination, including the expected end-point.
Quota Endpoints
Within physiological limits, quota endpoints or targetsare defined mainly by practical considerations. Target

T
Trajectory 2503
levels must consider time to permit achieving currentquota levels, as well as overall fatigue considerationsif there is an array of exercises to be performed. Quotaincrements and endpoints are also moderated by patientaccess to monitoring. For example, for a patient forwhom 3 weeks of daily trials is the logistical limit,quota endpoints may be limited by the time available toreach them using the quota increments selected, unlesssurrogate monitoring is available, e.g. family.
Examples
Walking
Pinpoint
Lay out the course to be walked (e.g. a 25 m lap in a hos-pital corridor or distance in meters from the front doorof home to a selected landmark).
Baseline
The initial baseline should be within expected patienttolerance. The patient is asked to walk laps (start-end-return to start) until “pain,weakness or fatigue cause youto want to stop. You decide when to stop.” If the patient isunable to do at least one lap, shorten the lap distance un-til two or more repetitions are within starting tolerance.Theoretically, baseline is defined by a stabilizing perfor-mance curve. Usually, however, in the exercise context,2–3 baseline trials suffice. Each trial is spaced to provideample rest between trials.
Record
Record laps walked each trial. A graph on the wall atthe bedside or in a folder carried by the patient is ade-quate. By making recorded results “visible”, an elementof social support or reinforcement is also provided.
Quota
One lap increments per trial usually suffices.
Consequate
Reinforcement from rest (i.e. time out from the activ-ity) and social approval of contextual personnel usuallysuffice. Resort to some form of tangible reinforcementis rarely needed. See Fordyce (1976, 1990) for more onthis.
Exercise: (e.g. Hip Abduction, Deep Knee Bends, Arm Exten-sion)
Pinpoint
The patient is asked to perform the assigned exercise (al-ways with full informed consent) and instructed as tohow to do it.
Baseline
The patient is asked to perform the assigned exercise“until pain, weakness or fatigue cause you to want tostop. You decide when to stop.” Amounts performedacross 2–3 trials, spaced appropriately, are recorded.
Quota
Usually one repetition per trial suffices. Increment ratesof >1 repetition per trial could be designated if there isample confidence that performance will succeed, partic-ularly if access to monitoring is limited by the calendar.
Consequate
Rest or time out from the exercise, plus therapist atten-tion and regard suffice.
Quota Failures (see also Fordyce 1976, 1990)
The options re failure divide into (a) problems of patientstatus and access and (b) performance.
Status
If opportunity to perform was not the problem, ask thepatient why failure? If change in health or status ofpain is the problem, clarify with the attending physi-cian whether to proceed with quotas, change exercises,etc. Scheduling problems or access to equipment withwhich to perform can be dealt with directly.
Performance
Your value as a source of reinforcement should not beexpended carelessly. Admonitions or urging better per-formance lets your attention become contingent on fail-ure. Instead, convey that you hope and expect he / shewill be able to resume meeting quotas and leave it at that.If on subsequent trials success is achieved, be quick topraise.If the patient is not invested in getting better, the patientshould choose whether to proceed or to halt the program.Discussions with significant others to the patient mayalso help.
References1. Fordyce WE (1976) Behavioral methods in chronic pain and ill-
ness. C.V. Mosby, St. Louis, pp 2362. Fordyce WE (1990) Contingency Management. In: Bonica JJ
Management of Pain, vol 2, 2nd edn. Lea & Febiger, Philadel-phia, pp 1702–1710 (also in: Bonica JJ (2001) Management ofPain, 3rd edn. Lippincott, Williams & Wilkins, Philadelphia,pp 1745–1750)
3. Premack D (1959) Toward Empirical Behavior Laws: I PositiveReinforcement. Psychol Rev 66:219–233
Trait
� Personality and Pain
Trajectory
Definition
The path of pain expression over time.� Pain Assessment in Neonates

2504 Tramadol
TramadolDefinitionTramadol is a synthetic, centrally acting analgesic agentthat is structurally related to codeine and morphine; it isa racemic mixture, and the 2 enantiomers function in acomplementary manner to enhance analgesic actions. Italso acts as a weak opioid agonist and inhibits the sero-tonin release and reuptake of norepinephrine and sero-tonin. The analgesic effects of oral tramadol (100mg)peak at 1–2 hrs and last for 3–6 hours after drug admin-istration.� Acute Pain in Children, Post-Operative� Drugs with Mixed Action and Combinations, Empha-
sis on Tramadol� Post-Operative Pain, Tramadol� Tramadol Hydrochloride
Tramadol HydrochlorideJAYANTILAL GOVIND
Department of Anaesthesia, Pain Clinic, University ofNew South Wales, Liverpool Hospital, Sydney, NSW,[email protected]
SynonymsTramadol; Tramal®; Zydol
DefinitionAn analogue of codeine, tramadol hydrochloride is acentrally acting analgesic. It is not derived from naturalsources, and structurally is not related to opioids; butdoes exhibit certain opioid characteristics.
CharacteristicsMechanismAnimal and in-vitro studies suggest that in addition toits mu-opioid effect, tramadol synergistically inhibitsthe reuptake of norepinephrine (NE) and serotonin, andsimultaneously stimulates the pre-synaptic release ofserotonin (Raffa and Fridericks 1996; Bamigbade et al.1997). It has a weak affinity for opioid receptors andis less potent than morphine. The analgesic effect isapportioned between the opioid and monoaminergiccomponents (Desmoules et al. 1996). The response isdose-dependent but the relationship between analgesiceffect and serum concentration varies considerablybetween individuals.
PharmacokineticsAfter oral administration, tramadol is rapidly and almostcompletely absorbed with a mean bio-availability of68%–72%. The drug is widely distributed and 20%is bound to plasma proteins. The peak serum level isreached within two hours (range 1–3 hours) (Tramal2004).
Tramadol is metabolized, via the CY2D6 isoenzyme ofcytochrome P450, to eleven metabolites of which onlyO-desmethyltramadol (M1) is active. M1 has a greateraffinity for opioid receptors and exerts greater analgesiathan tramadol. About 7%–8% of Caucasians lack thisisoenzyme. Tramadol has a half-life of about 6–7 hoursand is principally excreted via the kidneys. At least 30%remains unchanged (Tramal 2004).
Routes of Administration
Tramadol can be administered orally, rectally and par-enterally.For formulationsanddosing schedules readersshould consult the product information pertinent to theirrespective jurisdictions.
Applications
The agent is indicated for the treatment of mild to mod-erate pain. It is less effective than morphine for severepain (Kaye 2004; Pang et al. 1999; Osipova et al. 1991).
Efficacy
When administered to post-surgical and post-traumaticpatients, 100 mg injectable tramadol is equivalent to5–10mg of morphine. As an oral dose, 100 mg tra-madol is as effective as 1000 mg paracetamol (Mooreand McQuay 1997). For achieving 50% reduction ofpain, tramadol has an NNT of 4.6 (Moore and McQuay1997). For 150 mg the NNT is 2.9 (Drugdex DrugEvaluations. Micromedex® Healthcare series).Tramadol has been shown to be effective for the relief ofneuropathic pain (Duhmke 2004). Patients with chronicpainful neuropathy reported relief of their pain, paraes-thesia and touch evoked allodynia by 2 median points ona10-pointscale (Sindrup1999).Toachievea50%reduc-tion in pain level, the � NNT is between 3 and 6, using adaily dose of 200 mg – 400 mg. Studies investigating theefficacy of tramadol for the treatment of pain of diabetesneuropathy (Harati 1998) and � post-herpetic neuralgiahave been inconclusive (Wareham DW 2004).The safety and efficacy of tramadol in patients under theage of 16 years has not been established (Drugdex DrugEvaluations. Micromedex® Healthcare series).
Contraindications
Tramadol is contraindicated in patients with: por-phyria; known hypersensitivity to tramadol, opioids orany excipients; acute intoxication with alcohol, opioids,hypnotics, analgesics, or psychotropic drugs; who usemonoamine oxidase inhibitors, or have used them inthe last 14 days. Tramadol must not be used for opi-oid dependency, addiction or for opioid withdrawaltreatment (Kaye 2004; Pang et al. 1999; Drugdex DrugEvaluations. Micromedex® Healthcare series).
Side effects and complications
Adverse reactions are common and patients must begiven guidance about appropriate action. The potentialfor serious side effects including anaphylaxis should

T
Tramadol Hydrochloride 2505
not be underestimated. Either the monoaminergic oropioid effects may predominate. The adverse effectsof tramadol may be difficult to distinguish or recog-nize in patients taking multiple medications. Even withtherapeutic doses opioid side-effects can occur, exceptrespiratory depression and inhibition of smooth mus-cle, which are usually less pronounced (Kaye 2004;Drugdex Drug Evaluations. Micromedex® Healthcareseries).Central effects include seizures, hallucinations, eupho-ria, ataxia, and suicidal ideation. Co-administrationwith alcohol, general anaesthetics, or other respiratorydepressants can precipitate profound respiratory de-compensation. Dizziness can cause at-risk patients tofall.Gastrointestinal side effects are very common, includ-ing nausea, vomiting, and constipation. Dyspepsia, diar-rhoea and increased flatulence have been reported (Kaye2004).Although uncommon, cardiovascular side-effects suchas hypertension, palpitations, tachycardia, bradycardia,other ECG abnormalities, and orthostatic dysregulationdo occur and can be serious.Additional reported side effects include urinary reten-tion, erythema, hypertonia, hyponatraemia, itch, rhab-domyolysis and diaphoresis.
Drug interactions
Tramadolisknowntointeractwithat least72otherdrugs,some with serious consequences (Kaye 2004; DrugdexDrug Evaluations. Micromedex® Healthcare series).Carbamazepine reduces tramadol’s analgesic effect byincreasing its metabolism. Drugs that inhibit CY2D6activity (e.g. Selective Serotonin Reuptake Inhibitors[SSRI], or quinidine) will prevent conversion to itsactive metabolite, M1.Serotonin syndrome is a potentially serious toxicstate caused mainly by excess serotonin within thecentral nervous system (Hall and Buckley 2003). Itmanifests as a dose-related range of toxic symptomsdue to a variety of mental, autonomic and neuro-muscular changes. The clinical features are highlyvariable and the onset could be insidious or dra-matic. It is nearly always caused by a drug inter-action involving two or more ‘serotonergic’ drugsincluding SSRIs, MAOI, pethidine, tricyclic antide-pressants, St John’s wort and lithium. Severe hyper-thermia, rhabdomyolysis, disseminated intravascularcoagulation and adult respiratory distress syndromeare potentially life-threatening (Hall and Buckley2003).
Reproduction
Tramadol is embryotoxic and fetotoxic but not tetrato-genic in animal studies (Drugdex Drug Evaluations.Micromedex® Healthcare series).
Precautions
Tramadol is best avoided in patients with epilepsy, andin patients who are at risk of developing respiratorydepression. Because of miosis and its central effects,it is inadvisable to prescribe tramadol for patients witha head injury or raised intracranial pressure. Elderlypatients and patients with renal or hepatic decompen-sation, myxoedema, hypothyroidism, hypoadrenalismare particularly vulnerable to severe side effects andcomplications. Its administration may complicate theclinical assessment of patients with acute abdominalconditions (Kaye 2004; Drugdex Drug Evaluations.Micromedex® Healthcare series).
Pregnancy and lactation
Tramadol has been detected in breast milk. It is bestavoided during pregnancy and whilst breast-feeding(Kaye 2004).
Overdose
Treatment is symptomatic. The effects are not com-pletely reversed by naloxone (Kaye 2004), and thelatter may increase the risk of seizure (Drugdex DrugEvaluations. Micromedex® Healthcare series).
References1. Raffa RB, Fridericks E (1996) The basic science aspect of tra-
madol hydrochloride. Pain Rev 3:249–2712. Bamigbade TA, Davidson C, Langdord RM, Stamford JA (1997)
Actions of tramadol: its enantiomers and principle metabolite,O-desmethyltramadol on serotonin (5-HT) efflux and uptake inthe rat dorsal raphe nucleus. Br J Anaesth 79:352–356
3. Desmoules JB, Piquet R, Collart L, Dayer P (1996) Combinationof monoaminergic modulation to the analgesic effect of tramadolBr J Clin Pharmacol 41:7–12
4. Tramal (2004) Product information. CSL Limited. Parkville,VIC, Australia
5. Kaye K (2004) Trouble with tramadol (editorial) Australian Pre-scriber 27:26–27
6. Pang, WW, Mok MS, Lin CH, et al. (1999) Comparison of patientcontrolled analgesia (PCA) with tramadol or morphine. Can JAnesth 46:1030–1035
7. Osipova NA, Novikov GA, Beresnev VA et al. (1991) Analgesiceffect of tramadol in cancer patients with chronic pain: a com-parison with prolonged action morphine sulfate. Curr Ther Res50:812–821
8. Moore RA, McQuay HJ (1997) Single-patient data meta-analysisof 3453 postoperative patients: oral tramadol versus placebo,codeine and combination analgesics. Pain 69:287–294
9. Drugdex Drug Evaluations. Micromedex® Healthcare se-ries. � http://micromedex.hen.net.au/mde-3261/display.exeCTL=apache/products/micromede. Accessed 13.10.04
10. Duhmke RM, Cornblath DD, Hollingshead JR (2004) Tramadolfor neuropathic pain. Cochrane Database Syst Rev: CD003726
11. Sindrup SH, Andersen G, Madsen C, Smith T, Brosen K, JensenTS (1999) Tramadol relieves pain and allodynia in polyneuropa-thy: a randomised double blind controlled trial. Pain 83:85–90
12. Harati Y, Gooch C, Swenson M et al. (1998) Double blind ran-domized trial of tramadol for the treatment of the pain of diabeticneuropathy. Neurology 50:1842–1846
13. Wareham DW (2004) Postherpetic Neuralgia. In: Godlee F (ed)Clinical Evidence. edn
14. Hall M, Buckley N (2003) Serotonin Syndrome. Australian Pre-scriber 26:62–63

2506 Tramal®
Tramal®
� Postoperative Pain, Tramadol� Tramadol Hydrochloride
Transcranial Magnetic Stimulation
Synonyms
TMS
Definition
Transcranial Magnetic Stimulation refers to stimulationof the cerebral cortex by an electromagnetic field, gener-ated by a magnetic stimulator placed over the scalp with-out the need for surgery or external electrodes. Magneticstimuli can also activate the peripheral nervous system.� Clinical Migraine without Aura� Motor Cortex, Effect on Pain-Related Behavior� Stimulation Treatments of Central Pain
Transcription Factor
Definition
A protein, product of gene transcription and translation,which enters the nucleus where it binds to a nucleotidesequence in the regulatory regions of responsive genes,and has the effect of either enhancing or repressing theexpression ofoneormoregenesdownstreamof thebind-ing site. Expression of one gene is normally controlledby several transcription factors.� Central Changes after Peripheral Nerve Injury� COX-1 and COX-2 in Pain� Cytokines, Effects on Nociceptors� NSAIDs and Cancer
Transcutaneous Electrical NerveStimulation
JAMES WATT
North Shore Hospital, Auckland, New [email protected]
Synonyms
TENS; AL-TENS (Acupuncture like TENS); ElectricalTherapy.
Definition
Transcutaneous electrical nerve stimulation (TENS) is ameans of relieving pain. It entails delivering an electricalstimulus through electrodes to the skin overlying or nearthe region in which pain is perceived. The stimulus isdelivered from a battery-driven generator.
Characteristics
TENS is a commonly used non-invasive modality thatprovides an alternative to medication for pain relief. Ithasbeenusedformore than30years,but itseffectivenessremains controversial.
Mechanism
TENS was developed on the basis of the gate controltheory of pain proposed by Melzack and Wall (Melzackand Wall1965).This theory predicted that stimulation oflarge-diameter primary afferent fibres (A fibres) wouldhave an inhibitory effect on transmission from thesmall-diameter, unmyelinated afferent fibres (C fibres). Ac-cordingly, pain should be relieved if cutaneous afferentsfrom a region of pain could be artificially stimulated us-ing an electrical current.A battery-operated, transistorised unit generates theelectrical stimulus. There are usually three separatecontrols. The first varies the amplitude or intensity,and allows for administration of a range from low(2–4Hz) to high (100–250 Hz) frequency stimulation.A second varies pulse width, usually from 0.04-0.1 ms,and a third controls the mode to select continuous orpulsed ± ramped, ± random stimulus. The stimulus isdelivered through pads attached to the skin surface. Theelectrical field generated has to be of sufficient magni-tude to excite the adjacent afferent nerve fibres withoutdamaging the local skin, producing dysaesthesia orproducing painful muscle contractions.The mechanism of action of TENS has not been explic-itly or directly demonstrated. However, it is believedthat low frequency stimulation releases mu opioids (β-endorphins), while high frequency stimulation releasesdelta opioids (met-enkephalin and leu-enkephalin) inthe central nervous system. The mu receptor antago-nist – naloxone, inhibits the effects of low frequencystimulation, but not those of high frequency stimulation(Sjolund and Eriksson 1979; Freeman et a. 1983). Ratsdevelop opioid tolerance after 4 days when treated withTENS for 20 min. a day (Chandran and Sluka 2003).Due to the development of tolerance, treatment is moreeffective when TENS is used intermittently. Daily ad-ministration lessens the analgesic effect. The short du-rationofbenefit(260minutes) (Cheingetal.2003),how-ever, encourages frequent application in chronic condi-tions; but this in turn reduces pain relief.
Applications
TENS is used for the control of pain of various types invarious regions of the body. The portability, safety and

T
Transcutaneous Electrical Nerve Stimulation 2507
low cost of the transistorised impulse generator have allcontributed to its wide and varied use in the field of painmedicine.A number of factors need to beconsidered when offeringTENS as a treatment option. The aim is to produce theoptimal tolerable stimulus. Application is critical to thesuccess of the treatment. The site of stimulation shouldbe chosen to produce maximal input in the segmentwhere the pain originates, and should be proximal to it.It is often not possible to predict the optimal placementor type of electrodes, or the stimulus that will producemaximal pain relief. Consequently, with guidance, thepatient should be encouraged to experiment with am-plitude, frequency and pulse width, as well as electrodeplacement and duration of stimulus, in order to maxi-mize their relief of pain. Most choose frequencies from40–70 Hz; with a pulse-width of 0.1–0.5 ms. Stimu-lation at lower frequencies requires higher intensityof stimulus, which tends to produce painful musclecontractions.Maximal comfortable stimulus is more effective in re-lieving pain than stimulation that is barely detectable.However, the theoretical argument of providing a suffi-ciently powerful stimulus to activate both small myeli-nated A-delta fibres and large A-beta fibres is negatedby the difficulty patients have in tolerating continuoushigh-intensity stimulation.
Efficacy
Two Cochrane reviews, one of chronic pain (Carroll etal.2002),and theotherofchroniclowbackpain(Milneetal. 2002), demonstrate lack of effectiveness. There werewide variations in the parameters of type, site and fre-quency of application, treatment duration, intensity andfrequency of the electrical impulse in both reviews.Many studies show lack of evidence of benefit. Therewas no evidence that TENS was any more effective thanother conservative treatments for acute and chronic non-specific low back pain (van Tulder et al. 1997) or forchronic pain (McQuay e al. 1997). TENS was foundnot to be effective in relieving labour pain (Carroll etal. 1997).In contrast, a Cochrane Review by Proctor (Proctor etal. 2002) showed high frequencyTENSto bemoreeffec-tive than placebo in relieving primary dysmenorrhoea,while low frequency TENS showed no difference. Areview by Osiri (Osiri et al. 2001) on knee osteoarthritisshowed TENS provided significantly better pain reliefand improved movement, compared with placebo. Theeffect had previously been shown in three randomisedcontrolled trials reviewed in a paper by Puett and Grif-fin (Puett and Griffin 1994). However, Osiri’s reviewshowed that TENS only differed from placebo whentreatment continued for more than 4 weeks in duration.The efficacy of TENS in acute situations is attested ina meta-analysis of studies of reduction of analgesic usein post-operative patients. Bjordal (Bjordal et al. 2003)
reviewed 21 randomised placebo-controlled trials witha total of 1350 patients, and demonstrated a mean reduc-tion in analgesic consumption after TENS/AL-TENSof 26.5% (range –6 to +51%). Eleven of these trials, cov-ering 964 patients, reported that a strong sub-noxiouselectrical impulse of adequate stimulus frequency wasadministered. These trials demonstrated a reductionin analgesic consumption of 35.5% (range 1–51%). Incomparison, the nine trials that did not confirm sufficientcurrent intensity or adequate frequency, demonstrateda reduction of only 4.1% (range –10 to +29%).The short-term analgesic efficacy of TENS was demon-strated in a trial using distension arthrography (a moder-ately painful procedure) for frozen shoulder. There wasa 50% reduction using high intensity TENS, and 38% re-duction using low intensity TENS, compared to controls(Morgan et al. 1996).Thus, the evidence is mixed. This is due, in part, to thepoor quality of studies available, and the lack of cleardefinition of the treatment given. The effect has beenshowntodifferdependingonanumberofvariables in thetreatmentapplication.Thesearefrequency, intensityandwaveform of the stimulus, site and duration of its appli-cation, and conduction medium. All need to be recordedclearly for every subject ina trialof theefficacyofTENS.
Side-Effects
Opioid tolerance can develop with repeated daily use.There are few other side effects from the administrationof TENS, probably due to the application generallybeing under the patient’s control. There is the theo-retical risk that too high a stimulus could damage theskin by electrical burn, but it is unusual for patients toachieve such damage. The range of machine settings,and the noxious stimulus produced by painful musclecontraction at the higher stimulus range, affords pro-tection. Side effects really only occur in the presenceof excessive zeal.� Acupuncture Mechanisms� Chronic Pain in Children, Physical Medicine and Re-
habilitation� Complex Chronic Pain in Children, Interdisciplinary
Treatment� McGill Pain Questionnaire� Pain in Humans, Electrical Stimulation (Skin, Muscle
and Viscera)� Postoperative Pain, Appropriate Management� Transcutaneous Electrical Nerve Stimulation Out-
comes� TranscutaneousElectricalNerveStimulation (TENS)
in Treatment of Muscle Pain
References1. Bjordal JM, Johnson MI, Ljunggreen AE (2003) Transcutaneous
Electrical Nerve Stimulation (TENS) can Reduce PostoperativeAnalgesic Consumption. A Meta-Analysis with Assessment ofOptimal Treatment Parameters for Postoperative Pain. Eur JPain 7:181–188

2508 Transcutaneous Electrical Nerve Stimulation (TENS) in Treatment of Muscle Pain
2. Carroll D, Moore RA, McQuay HJ, Fairman F, Tramer M, LeijonG (2002) Transcutaneous Electrical Nerve Stimulation (TENS)for Chronic Pain (Cochrane Review). The Cochrane Library, Is-sue 2
3. Carroll D, Moore RA, Tramer MR, McQuay HJ (1997) Transcu-taneous Electrical Nerve Stimulation Does Not Relieve LabourPain: Updated Systematic Review. Contemporary Reviews in Ob-stetrics and Gynaecology: 195–205
4. Chandran P, Sluka KA (2003) Development of Opioid Tolerancewith Repeated Transcutaneous Electrical Nerve Stimulation Ad-ministration. Pain 102(1–2):195–201
5. Cheing GL, Tsui AY, Lo SK, Hui-Chan CW (2003) Optimal Stim-ulation Duration of TENS in the Management of OsteoarthriticKnee Pain. J Rehabil Med 35(2):62–68
6. Freeman TB, Campbell JN, Long DM (1983) Naloxone DoesNot Affect Pain Relief Induced by Electrical Stimulation in Man.Pain 17:189–195
7. McQuay HJ, Moore RA, Eccleston C, Morley S, DeC WilliamsAC (1997) Systematic Review of Outpatient Services for ChronicPain Control. Health Technology Assessment 1(6):1–137
8. Melzack R, Wall PD (1965) Pain Mechanisms: A New Theory.Science 15:971–979
9. Milne S, Welch V, Brosseau L, Saginur M, Shea B, TugwellP, Wells G (2002) Transcutaneous Electrical Nerve Stimulation(TENS) for Chronic Low Back Pain (Cochrane Review). TheCochrane Library, Issue 1
10. Morgan B, Jones AR, Mulcahy KA, Finlay DB, Collett B(1996) Transcutaneous Electrical Nerve Stimulation (TENS)during Distension Shoulder Arthrography: A Controlled Trial.Pain 64:265–267
11. Osiri M, Welch V, Brosseau L, Shea B, McGowan J, Tugwell P,Wells G (2001) Transcutaneous Electrical Nerve Stimulation forKnee Osteoarthritis (Cochrane Review). The Cochrane Library,Issue 1
12. Proctor ML, Smith CA, Farquhar CM, Stones RW (2002) Tran-scutaneous Electrical Nerve Stimulation and Acupuncture forPrimary Dysmenorrhoea (Cochrane Review). The Cochrane Li-brary, Issue 1
13. Puett DW, Griffin MR (1994) Published Trials of Non-Medicinaland Non-Invasive Therapies for Hip and Knee Osteoarthritis. AnnInt Med 121:133–140
14. Sjolund BH, Eriksson MBE (1979) The Influence of Naloxoneon Analgesia Produced by Peripheral Conditioning Stimulation.Brain Res 173:295–301
15. van Tulder MW, Koes BW, Bouter LM (1997) ConservativeTreatment of Acute and Chronic Non-Specific Low Back Pain:A Systematic Review of Randomised Controlled Trials of theMost Common Interventions. Spine 22:2128–2156
Transcutaneous Electrical NerveStimulation (TENS) in Treatment ofMuscle Pain
MARK I. JOHNSON
Faculty of Health, Leeds Metropolitan University,Leeds, [email protected]
Synonyms
Transcutaneous Nerve Stimulation (TNS); Transcuta-neous Electrical Stimulation; (TES) Electrical Stimu-lation Therapy (EST); Percutaneous Electrical NerveStimulation; Electroanalgesia
Transcutaneous Electrical Nerve Stimulation (TENS) in Treatment ofMuscle Pain, Figure 1 An electrical pulse generator delivers currents viaconducting electrodes attached to the intact surface of the skin. Tradition-ally, carbon rubber electrodes smeared with conducting gel and attachedto the skin using self-adhesive tape were used to deliver the electricalcurrents. Nowadays, self-adhesive electrodes are used.
Definition� Transcutaneous electrical nerve stimulation (TENS)is a non-invasive analgesic intervention, which deliverselectrical currents across the intact surface of the skinto stimulate the underlying nerves (see Fig. 1). TENS isused extensively for the symptomatic relief of all typesof pain, including pains of musculoskeletal origin.The purpose of TENS is to activate selectively thosepopulations of nerve fibres that are concerned with� segmental and � extrasegmental anti-nociceptivemechanisms. The two main TENS techniques are� conventional TENS and � acupuncture-like TENS(AL-TENS).
Characteristics
TENS is popular because patients can administer TENSthemselves and can titrate the dosage of treatment asrequired. TENS effects are often rapid in onset andthere are few side effects and no potential for toxicity oroverdose. The technical specifications of TENS devicesvary according to manufacturer, although most utilisebiphasic pulsed currents that may be either symmetricalor non-symmetrical square waves. Pulse durations liebetween 50 μs–1000 μs, pulse frequencies between1–200 pulses per second (pps) and pulse amplitudes be-tween 1–60 mA. Most devices offer continuous, burst(used for AL-TENS) and modulated pulse patterns.Factors that influence the success of TENS include thepatient, the condition and the appropriateness of theTENS technique employed (see Fig. 2).TENS is commonly used to treat chronic pains, includ-ing those of musculoskeletal origin. Systematic reviewson TENS and chronic pain have been inconclusive dueto the low methodological quality of RCTs (Reeve et al.1996; McQuay and Moore 1998; Carroll et al. 2003).However, meta-analyses have demonstrated that TENSis beneficial for knee osteoarthritis (Osiri et al. 2000)and rheumatoid arthritis in the hand (Brosseau et al.2003). Meta-analysis on TENS for low back pain are

T
Transcutaneous Electrical Nerve Stimulation (TENS) in Treatment of Muscle Pain 2509
Transcutaneous Electrical NerveStimulation (TENS) in Treatmentof Muscle Pain, Figure 2 Theoutput characteristics of a typicalTENS device (topographic view).The amplitude, frequency, durationand pattern of electrical pulsescan be controlled by the enduser (I, intensity; F, frequency; D,duration; C, continuous; B, burst;M, modulation; pps, pulses persecond).
conflicting (Flowerdew and Gadsby 1997; Brosseauet al. 2002). Under dosing of TENS and the use ofinappropriate TENS techniques in some RCTs haveinfluenced the findings of systematic reviews (Bjordalet al. 2003).There have been no systematic reviews on TENS andmuscle pain because of a lack of randomised controlledclinical trials. TENS is used extensively for reducing lo-calised muscle pain, especially in the neck and shoulder,arising from muscle tension. TENS is also used to treatacute traumatic muscle pain that results from physicalinjury from sport and minor accidents, and to treat post-exertional muscle pain. Reports suggest that TENS ishelpful for myofascial pain syndrome (Graff-Radford etal.1989;Houetal.2002)andlocalisedpaininfibromyal-gia (Offenbacher and Stucki 2000), although in practice,TENS reduces pain in some of these patients and aggra-vates it in others. It is unlikely that TENS will be helpfulin widespread pain in fibromyalgia, because it is difficultto direct TENS currents into the painful area.In clinical practice, conventional TENS is used in thefirst instance for most pains, including those of muscu-loskeletal origin. The purpose of conventional TENS isto activate selectively large diameter non-nociceptivecutaneous afferents (Aβ) without concurrently activat-ing small diameter nociceptive afferents (Aδ and C),which would cause pain, or muscle efferents, whichwould cause muscle contractions. TENS-induced Aβ
activity has been shown to inhibit ongoing activity insecond order nociceptive neurones in the dorsal horn ofthe spinal cord (Garrison and Foreman 1994). In prac-tice, large diameter non-nociceptive cutaneous afferentactivity is recognised by a ‘strong but comfortable’ non-painful electrical paraesthesia beneath the electrodes,and patients are trained to titrate current amplitude toachieve this outcome (see Fig. 3).
Transcutaneous Electrical Nerve Stimulation (TENS) in Treatment ofMuscle Pain, Figure 3 The purpose of conventional TENS is to activatenon-nociceptive cutaneous afferents (Aβ) without concurrent activation no-ciceptive (Aδ/C) or muscle afferents. Arrows indicate impulses travellingtowards the central nervous system.
During conventional TENS electrodes are applied tohealthy skin, at the site of pain, to stimulate large diame-ter non-nociceptive cutaneous afferents which enter thesame spinal segment as the nociceptive fibres associatedwith the origin of the pain. When it is not possible toapply electrodes at the site of pain, for example whenthe skin is damaged and/or sensitive to touch, electrodescan be applied proximally over the main nerve trunkthat innervates the skin at the site of pain. Alternatively,electrodes can be placed over the spinal cord at a levelsegmentally related to the site of origin of the pain, orat a site which is contralateral (mirror image) to the siteof pain. Dual channel devices using 4 electrodes can beused for pains covering large areas such as in a glutealregion or lower limb, and for multiple pains such as lowback pain and sciatica (see Fig. 4).For conventional TENS, maximum pain relief isachieved when the TENS device is switched on. There-
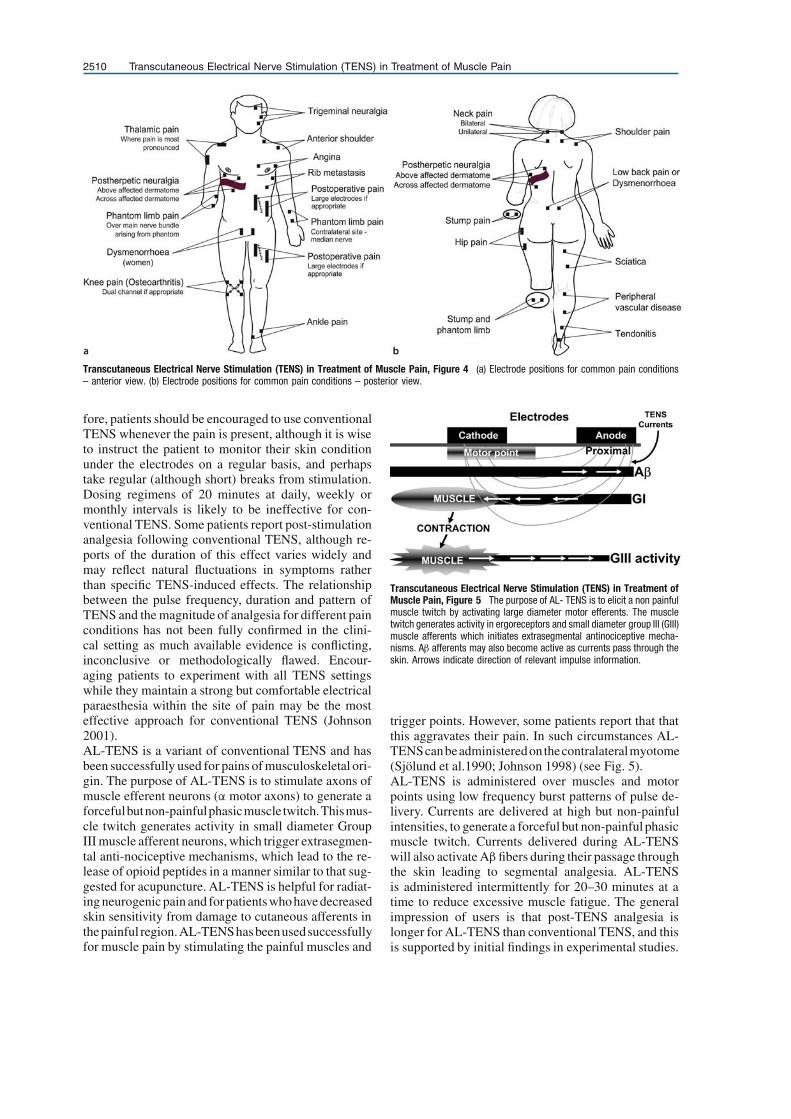
2510 Transcutaneous Electrical Nerve Stimulation (TENS) in Treatment of Muscle Pain
Transcutaneous Electrical Nerve Stimulation (TENS) in Treatment of Muscle Pain, Figure 4 (a) Electrode positions for common pain conditions– anterior view. (b) Electrode positions for common pain conditions – posterior view.
fore, patients should be encouraged to use conventionalTENS whenever the pain is present, although it is wiseto instruct the patient to monitor their skin conditionunder the electrodes on a regular basis, and perhapstake regular (although short) breaks from stimulation.Dosing regimens of 20 minutes at daily, weekly ormonthly intervals is likely to be ineffective for con-ventional TENS. Some patients report post-stimulationanalgesia following conventional TENS, although re-ports of the duration of this effect varies widely andmay reflect natural fluctuations in symptoms ratherthan specific TENS-induced effects. The relationshipbetween the pulse frequency, duration and pattern ofTENS and the magnitude of analgesia for different painconditions has not been fully confirmed in the clini-cal setting as much available evidence is conflicting,inconclusive or methodologically flawed. Encour-aging patients to experiment with all TENS settingswhile they maintain a strong but comfortable electricalparaesthesia within the site of pain may be the mosteffective approach for conventional TENS (Johnson2001).AL-TENS is a variant of conventional TENS and hasbeen successfully used for pains of musculoskeletal ori-gin. The purpose of AL-TENS is to stimulate axons ofmuscle efferent neurons (α motor axons) to generate aforcefulbutnon-painfulphasicmuscle twitch.Thismus-cle twitch generates activity in small diameter GroupIII muscle afferent neurons, which trigger extrasegmen-tal anti-nociceptive mechanisms, which lead to the re-lease of opioid peptides in a manner similar to that sug-gested for acupuncture. AL-TENS is helpful for radiat-ing neurogenic pain and forpatients who have decreasedskin sensitivity from damage to cutaneous afferents inthepainful region.AL-TENShasbeenusedsuccessfullyfor muscle pain by stimulating the painful muscles and
Transcutaneous Electrical Nerve Stimulation (TENS) in Treatment ofMuscle Pain, Figure 5 The purpose of AL- TENS is to elicit a non painfulmuscle twitch by activating large diameter motor efferents. The muscletwitch generates activity in ergoreceptors and small diameter group III (GIII)muscle afferents which initiates extrasegmental antinociceptive mecha-nisms. Aβ afferents may also become active as currents pass through theskin. Arrows indicate direction of relevant impulse information.
trigger points. However, some patients report that thatthis aggravates their pain. In such circumstances AL-TENScanbeadministeredonthecontralateralmyotome(Sjölund et al.1990; Johnson 1998) (see Fig. 5).AL-TENS is administered over muscles and motorpoints using low frequency burst patterns of pulse de-livery. Currents are delivered at high but non-painfulintensities, to generate a forceful but non-painful phasicmuscle twitch. Currents delivered during AL-TENSwill also activate Aβ fibers during their passage throughthe skin leading to segmental analgesia. AL-TENSis administered intermittently for 20–30 minutes at atime to reduce excessive muscle fatigue. The generalimpression of users is that post-TENS analgesia islonger for AL-TENS than conventional TENS, and thisis supported by initial findings in experimental studies.

T
Transcutaneous Electrical Nerve Stimulation Outcomes 2511
For this reason, AL-TENS is useful for patients whoobtain brief after-effects from conventional TENS, orwho are resistant to conventional TENS.TENS should not be administered to patients fitted withcardiac pacemakers or women in the first trimester ofpregnancy. TENS should not be used while operatingvehicles or potentially hazardous equipment, and elec-trodes should not be positioned over the anterior partof the neck, over areas of broken skin or directly overa pregnant uterus (although it is safe when applied tothe lower back to treat labour pains). Patients should betested fornormal skin sensation prior tousingTENS.Pa-tients should be warned not to use TENS in the showeror bath and to keep TENS appliances out of the reach ofchildren.
References1. Bjordal JM, Johnson MI, Ljunggreen AE (2003) Transcutaneous
Electrical Nerve Stimulation (TENS) can Reduce PostoperativeAnalgesic Consumption. A Meta-Analysis with Assessment ofOptimal Treatment Parameters for Postoperative Pain. Eur J Pain7:181–188
2. Brosseau L, Milne S, Robinson V et al. (2002) Efficacy of theTranscutaneous Electrical Nerve Stimulation for the Treatmentof Chronic Low Back Pain: A Meta-Analysis. Spine 27:596–603
3. Brosseau L, Yonge KA, Robinson V et al. (2003) Transcuta-neous Electrical Nerve Stimulation (TENS) for the Treatmentof Rheumatoid Arthritis in the Hand (Cochrane Review). TheCochrane Library Oxford: Update Software Issue 3:1–19
4. Carroll D, Moore RA, McQuay HJ et al. (2003) TranscutaneousElectrical Nerve Stimulation (TENS) for Chronic Pain. In: TheCochrane Library, Issue 3. Update Software, Oxford
5. Flowerdew M, Gadsby G (1997) A Review of the Treatmentof Chronic Low Back Pain with Acupuncture-Like Transcuta-neous Electrical Nerve Stimulation and Transcutaneous Electri-cal Nerve Stimulation. Complement Ther Med 5:193–201
6. Garrison DW, Foreman RD (1994) Decreased Activity of Sponta-neous and Noxiously Evoked Dorsal Horn Cells during Transcu-taneous Electrical Nerve Stimulation (TENS). Pain 58:309–315
7. Graff-Radford SB, Reeves JL, Baker RL et al. (1989) Effectsof Transcutaneous Electrical Nerve Stimulation on MyofascialPain and Trigger Point Sensitivity. Pain 37:1–5
8. Hou CR, Tsai LC, Cheng KF et al. (2002) Immediate Effects ofVarious Physical Therapeutic Modalities on Cervical Myofas-cial Pain and Trigger-Point Sensitivity. Arch Phys Med Rehabil83:1406–1414
9. Johnson M (1998) The Analgesic Effects and Clinical Use ofAcupuncture-Like TENS (AL-TENS). Phys Ther Rev 3:73–93
10. Johnson MI (2001) Transcutaneous Electrical Nerve Stimula-tion. In: Kitchen S (ed) Electrotherapy: Evidence-Based Practice.Churchill Livingstone, Edinburgh, pp 259–286
11. McQuay H, Moore A (1998) TENS in Chronic Pain. In: McQuayH, Moore A (eds) An Evidence-Based Resource for Pain Relief.Oxford University Press, Oxford, pp 207–211
12. Offenbacher M, Stucki G (2000) Physical Therapy in the Treat-ment of Fibromyalgia. Scand J Rheumatol 113:78–85
13. Osiri M, Welch V, Brosseau L et al. (2003) TranscutaneousElectrical Nerve Stimulation for Knee Osteoarthritis. In: TheCochrane Library, Issue 3. Update Software, Oxford
14. Reeve J, Menon D, Corabian P (1996) Transcutaneous Electri-cal Nerve Stimulation (TENS): A Technology Assessment. IntJ Technol Assess Health Care 12:299–324
15. Sjölund B, Eriksson M, Loeser J (1990) Transcutaneous and Im-planted Electric Stimulation of Peripheral Nerves. In: Bonica JJ(ed) The Management of Pain, vol 2. Lea & Febiger, Philadel-phia, pp 1852–1861
Transcutaneous Electrical NerveStimulation Outcomes
MARK I. JOHNSON
School of Health and Human Sciences, Faculty ofHealth, Leeds Metropolitan University, Leeds, [email protected]
Synonyms
TENS Outcomes; transcutaneous nerve stimulation;TNS; transcutaneous electrical stimulation (TES);electrical stimulation therapy (EST); PercutaneousElectrical Nerve Stimulation; Electroanalgesia
Definition� Transcutaneous electrical nerve stimulation (TENS)is a non-invasive analgesic intervention that deliv-ers electrical currents across the intact surface ofthe skin to stimulate the underlying nerves. TENSis used extensively for the symptomatic relief of alltypes of pain including pains of nociceptive, neuro-pathic and musculoskeletal origin. The purpose ofTENS is to activate selectively those populations ofnerve fibres that are concerned with segmental andextrasegmental anti-nociceptive mechanisms. The twomain TENS techniques are � conventional TENS and� acupuncture-like TENS (AL-TENS).
Characteristics
Clinical experience supports a role for TENS in themanagement of acute and chronic pain. Clinical re-search on TENS also reports beneficial effects for awide range of chronic pain conditions including backpain, neck pain, headache, osteoarthritis, rib fracture,orofacial pain, post-herpetic neuralgia, trigeminal neu-ralgia, post-stroke pain, phantom limb and stump pain,brachial plexus avulsion, causalgia, angina pectoris,myalgia, postoperative pain, labour pain, dental painand cancer pain (Hansson and Lundeberg 1999). How-ever, many clinical trials lack appropriate controlsand / or randomisation leading to overestimation oftreatment effects. In recent years, systematic reviewsand meta-analyses have challenged the effectiveness ofTENS.Many clinical trials on TENS and chronic pain suffermethodological inadequacies and fail to meet the inclu-sion criteria for systematic reviews. Systematic review-ers report that the lack of good quality RCTs makes itdifficult to estimate the effectiveness of TENS. Reeveet al. reported that TENS was more effective than sham(dummy) TENS or no treatment in 9 / 20 RCTs on avariety of chronic pain conditions (Reeve et al. 1996).McQuay et al. reported that TENS was better than shamTENS, placebo pills or inappropriate electrode place-ments in 10 / 24 RCTs on chronic pain (McQuay andMoore 1998). A follow-up review identified 107 reports

2512 Transcutaneous Electrical Nerve Stimulation Outcomes
Transcutaneous Electrical Nerve Stimulation Outcomes, Table 1 Physical Medicine, TENS outcomes
Ref. Patients Results Reviewers’Conclusion
Reeve et al. (1996) Mixed conditions (low back, pancreatitis,arthritis, angina)
TENS >control in 9 / 20 RCTs Evidenceinconclusive
McQuay and Moore(1998)
Mixed conditions (low back, pancreatitis,osteoarthritis, dysmenorrhoea)
TENS >control in 10 / 24 RCTs Evidenceinconclusive
Carroll et al. (2003) Mixed conditions (low back, pancreatitis,osteoarthritis, dysmenorrhoea)
TENS >control in 10 / 15 RCTs Evidenceinconclusive
Flowerdue andGadsby (1997)
Low back pain 288 patients (6 RCTs)TENS >sham for pain relief (OR = 2.11)
Evidence of effect
Brosseau et al.(2002)
Low back pain 421 patients (5 RCTs)TENS = sham for pain relief (SMD = -0.207)
Evidence of noeffect
Price and Pandyan(2001)
Post-stroke shoulder pain 170 patients (4 RCTs). Any surface electricalstimulation (ES)ES = sham / no treatment control for pain relief(WMD = 0.13)ES >sham / no treatment control for range ofmovement (WMD = 9.17)
Evidenceinconclusive
Proctor et al. (2002) Primary dysmenorrhoea 213 patients (8 RCTs)HF TENS >sham for pain relief (OR = 7.2)LF TENS = sham for pain relief (OR = 1.3)
Evidence ofeffect – HF TENSonly
Brosseau et al.(2003)
Rheumatoid arthritis of the hand 78 patients (3 RCTs)AL-TENS reduced pain at rest (67% relative benefitversus placebo)
Evidence of effect
Osiri et al. (2003) Knee osteoarthritis 294 patients (7 RCTs)TENS >sham for pain relief (SMD = -0.448 – althoughonly 2 / 7 RCTs +ve)
Evidence of effect
Abbreviations: RCT, randomised controlled clinical trial; OR, odds ratio; SMD, standardised mean difference; WMD, weighted mean difference; +ve, positiveoutcome; HF, high frequency; LF, low frequency
on TENS and chronic pain but only 19 met the inclusioncriteria (Carroll et al. 2003). TENS provided better painrelief than sham orno treatmentcontrols in 10 /15RCTs.Evidence for TENS effectiveness in specific chronicpain conditions is also inconclusive or discordant. Ameta-analysis of 288 low back pain (LBP) patients (6RCTs) found that TENS reduced pain and improvedthe range of motion (Flowerdew and Gadsby 1997). Incontrast, a meta-analysis of 321 LBP patients (5 RCTs)found no statistically significant differences betweenactive and sham TENS for pain relief (Brosseau et al.2002). A meta-analysis of 294 patients (7 RCTs) withknee osteoarthritis found that TENS produced statis-tically significantly more pain relief and reductionsin knee stiffness than placebo (Osiri et al. 2003). Ameta-analysis of 78 patients (3 RCTs) with rheuma-toid arthritis of the hand found that AL-TENS reducedpain intensity whereas conventional TENS did not,although conventional TENS improved the patient’sassessment of their disease state (Brosseau et al. 2003).The beneficial effects of AL-TENS were achieved usingparticularly low dosage regimens of one 15 min ses-
sion per week for 3 weeks. A meta-analysis of TENSfor primary dysmenorrhoea found that high frequencybut not low frequency TENS was more effective forpain relief than sham TENS (Proctor et al. 2003). Ameta-analysis of any form of surface electrical stimu-lation (ES) on 170 patients with post-stroke shoulderpain found no significant change in pain incidence orpain intensity after ES compared with control (Priceand Pandyan 2001). ES was reported to improve thepain-free range of passive humeral lateral rotation andreduce the severity of glenohumeral subluxation. Insummary, the effectiveness of TENS on chronic painremains uncertain.The effectiveness of TENS in reducing labour pain hasbeen challenged by systematic reviews. TENS did notimprove pain relief when compared to sham TENS or ano treatment control in 3 / 8 RCTs (Reeve et al. 1996)and 10 / 10 RCTs (Carroll et al. 1997). These systematicreviews seem to conflict with clinical experience wheremidwives and patients report satisfaction with TENS ef-fects. Interestingly, one RCT found significantly morewomen and midwives favoured active rather than sham
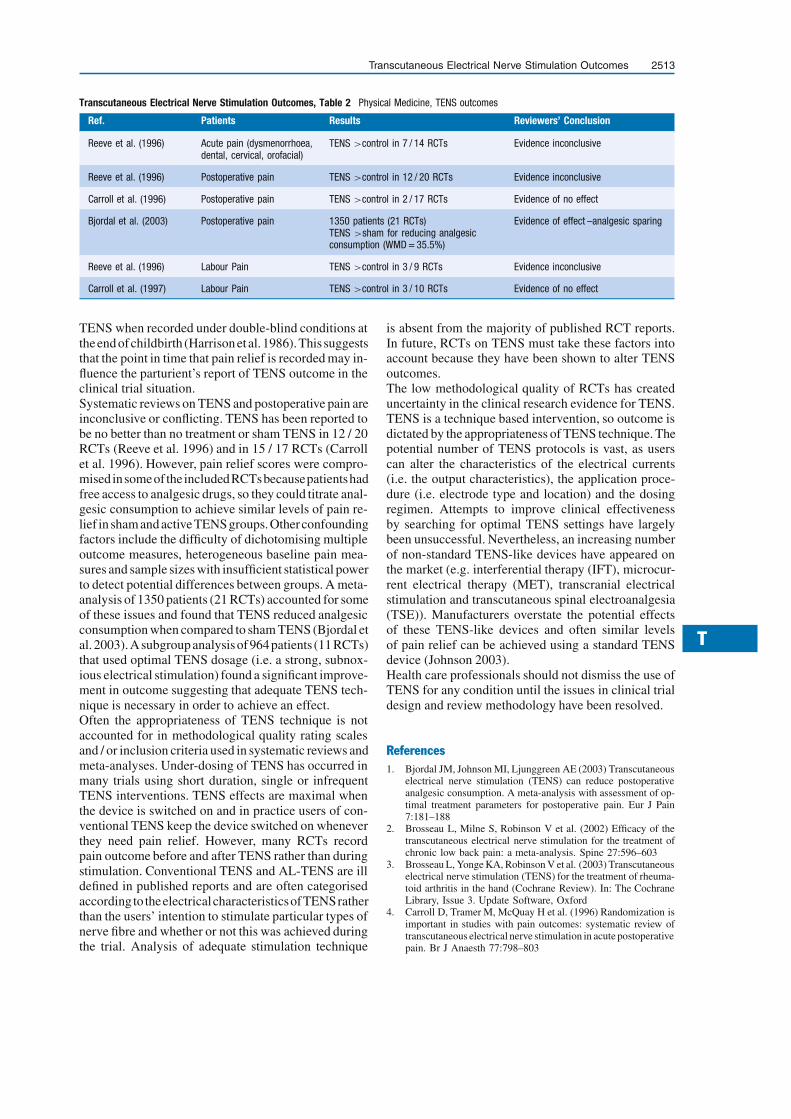
T
Transcutaneous Electrical Nerve Stimulation Outcomes 2513
Transcutaneous Electrical Nerve Stimulation Outcomes, Table 2 Physical Medicine, TENS outcomes
Ref. Patients Results Reviewers’ Conclusion
Reeve et al. (1996) Acute pain (dysmenorrhoea,dental, cervical, orofacial)
TENS >control in 7 / 14 RCTs Evidence inconclusive
Reeve et al. (1996) Postoperative pain TENS >control in 12 / 20 RCTs Evidence inconclusive
Carroll et al. (1996) Postoperative pain TENS >control in 2 / 17 RCTs Evidence of no effect
Bjordal et al. (2003) Postoperative pain 1350 patients (21 RCTs)TENS >sham for reducing analgesicconsumption (WMD = 35.5%)
Evidence of effect –analgesic sparing
Reeve et al. (1996) Labour Pain TENS >control in 3 / 9 RCTs Evidence inconclusive
Carroll et al. (1997) Labour Pain TENS >control in 3 / 10 RCTs Evidence of no effect
TENS when recorded under double-blind conditions attheend ofchildbirth (Harrison etal. 1986).This suggeststhat the point in time that pain relief is recorded may in-fluence the parturient’s report of TENS outcome in theclinical trial situation.Systematic reviews on TENS and postoperative pain areinconclusive or conflicting. TENS has been reported tobe no better than no treatment or sham TENS in 12 / 20RCTs (Reeve et al. 1996) and in 15 / 17 RCTs (Carrollet al. 1996). However, pain relief scores were compro-mised insomeof the includedRCTsbecausepatientshadfree access to analgesic drugs, so they could titrate anal-gesic consumption to achieve similar levels of pain re-lief in sham and activeTENSgroups.Otherconfoundingfactors include the difficulty of dichotomising multipleoutcome measures, heterogeneous baseline pain mea-sures and sample sizes with insufficient statistical powerto detect potential differences between groups. A meta-analysis of 1350 patients (21 RCTs) accounted for someof these issues and found that TENS reduced analgesicconsumption when compared to sham TENS (Bjordal etal. 2003).Asubgroup analysisof964patients (11 RCTs)that used optimal TENS dosage (i.e. a strong, subnox-ious electrical stimulation) found a significant improve-ment in outcome suggesting that adequate TENS tech-nique is necessary in order to achieve an effect.Often the appropriateness of TENS technique is notaccounted for in methodological quality rating scalesand / or inclusion criteria used in systematic reviews andmeta-analyses. Under-dosing of TENS has occurred inmany trials using short duration, single or infrequentTENS interventions. TENS effects are maximal whenthe device is switched on and in practice users of con-ventional TENS keep the device switched on wheneverthey need pain relief. However, many RCTs recordpain outcome before and after TENS rather than duringstimulation. Conventional TENS and AL-TENS are illdefined in published reports and are often categorisedaccording to theelectricalcharacteristicsofTENSratherthan the users’ intention to stimulate particular types ofnerve fibre and whether or not this was achieved duringthe trial. Analysis of adequate stimulation technique
is absent from the majority of published RCT reports.In future, RCTs on TENS must take these factors intoaccount because they have been shown to alter TENSoutcomes.The low methodological quality of RCTs has createduncertainty in the clinical research evidence for TENS.TENS is a technique based intervention, so outcome isdictated by the appropriateness of TENS technique. Thepotential number of TENS protocols is vast, as userscan alter the characteristics of the electrical currents(i.e. the output characteristics), the application proce-dure (i.e. electrode type and location) and the dosingregimen. Attempts to improve clinical effectivenessby searching for optimal TENS settings have largelybeen unsuccessful. Nevertheless, an increasing numberof non-standard TENS-like devices have appeared onthe market (e.g. interferential therapy (IFT), microcur-rent electrical therapy (MET), transcranial electricalstimulation and transcutaneous spinal electroanalgesia(TSE)). Manufacturers overstate the potential effectsof these TENS-like devices and often similar levelsof pain relief can be achieved using a standard TENSdevice (Johnson 2003).Health care professionals should not dismiss the use ofTENS for any condition until the issues in clinical trialdesign and review methodology have been resolved.
References1. Bjordal JM, Johnson MI, Ljunggreen AE (2003) Transcutaneous
electrical nerve stimulation (TENS) can reduce postoperativeanalgesic consumption. A meta-analysis with assessment of op-timal treatment parameters for postoperative pain. Eur J Pain7:181–188
2. Brosseau L, Milne S, Robinson V et al. (2002) Efficacy of thetranscutaneous electrical nerve stimulation for the treatment ofchronic low back pain: a meta-analysis. Spine 27:596–603
3. Brosseau L, Yonge KA, Robinson V et al. (2003) Transcutaneouselectrical nerve stimulation (TENS) for the treatment of rheuma-toid arthritis in the hand (Cochrane Review). In: The CochraneLibrary, Issue 3. Update Software, Oxford
4. Carroll D, Tramer M, McQuay H et al. (1996) Randomization isimportant in studies with pain outcomes: systematic review oftranscutaneous electrical nerve stimulation in acute postoperativepain. Br J Anaesth 77:798–803

2514 Transcutaneous Electrical Stimulation
5. Carroll D, Moore A, Tramer M et al. (1997) Transcutaneouselectrical nerve stimulation does not relieve in labour pain: up-dated systematic review. Contemporary Reviews in Obstetricsand Gynecology September 1997:195–205
6. Carroll D, Moore RA, McQuay HJ et al. (2003) Transcutaneouselectrical nerve stimulation (TENS) for chronic pain. In: TheCochrane Library, Issue 3. Update Software, Oxford
7. Flowerdew M, Gadsby G (1997) A review of the treatment ofchronic low back pain with acupuncture-like transcutaneous elec-trical nerve stimulation and transcutaneous electrical nerve stim-ulation. Complem Ther Med 5:193–201
8. Hansson P, Lundeberg T (1999) Transcutaneous electrical nervestimulation, vibration and acupuncture as pain-relieving mea-sures. In: PD Wall, R Melzack (eds) Textbook of Pain. ChurchillLivingstone, Edinburgh, pp 1341–1351
9. Harrison R, Woods T, Shore M et al. (1986) Pain relief in labourusing transcutaneous electrical nerve stimulation (TENS). ATENS / TENS placebo controlled study in two parity groups.Br J Obstet Gynaecol 93:739–746
10. Johnson M (2003) Transcutaneous Electrical Nerve Stimulation(TENS) and TENS-like devices. Do they provide pain relief?Pain Rev 8:121–128
11. McQuay H, Moore A (1998) TENS in chronic pain. In: McQuayH, Moore A (eds) An evidence-based resource for pain relief.Oxford University Press, Oxford, pp 207–211
12. Osiri M, Welch V, Brosseau L et al. (2003) Transcutaneous elec-trical nerve stimulation for knee osteoarthritis. In: The CochraneLibrary, Issue 3. Update Software, Oxford
13. Price CI, Pandyan AD (2001) Electrical stimulation for prevent-ing and treating post-stroke shoulder pain: a systematic Cochranereview. Clin Rehab 15:5–19
14. Proctor ML, Smith CA, Farquhar CM et al. (2003) Transcuta-neous electrical nerve stimulation and acupuncture for primarydysmenorrhoea. (Cochrane Review). The Cochrane Library, Is-sue 3. Update Software, Oxford
15. Reeve J, Menon D, Corabian P (1996) Transcutaneous electri-cal nerve stimulation (TENS): a technology assessment. Int JTechnol Assess Health Care 12:299–324
Transcutaneous Electrical Stimulation
� Transcutaneous Electrical Nerve Stimulation Out-comes
� TranscutaneousElectricalNerveStimulation (TENS)in Treatment of Muscle Pain
Transdermal
Definition
The route of analgesic action is through transdermal ap-plication for systemic effect.� Analgesic Guidelines for Infants and Children
Transduction
Definition
Transduction is the conversion of one form of signal toanother.Forexample, in sensoryendings, theconversion
of the stimulus (e.g. pressure or cold) into an electricalsignal.� Somatic Pain� Species Differences in Skin Nociception� Visceral Nociception and Pain
Transduction Channel
Definition
Ionic channel gated (i.e. open or closed) by a physicalor chemical stimulus applied to the membrane of a pe-ripheral sensory cell.� Nociceptor Generator Potential
Transduction Sites
Definition
Membrane patches of sensory receptor terminals wheresensory transduction takes place.� Nociceptor Generator Potential
Transfected Cells
Definition
Transfection refers to the introduction of exogenousgenetic material encoding a gene of interest into cells.Transfected cells are used as model systems to studyfunctional and structural properties of the transfectedprotein.� Purine Receptor Targets in the Treatment of Neuro-
pathic Pain
Transfer and Generalization
Definition
Transfer refers to the maintenance of learned behaviorchange in the patient’s environment outside the treat-ment context; generalization refers to the extension oflearned behaviors to similar problems.� Operant Treatment of Chronic Pain
Transforaminal Injection of Steroids
� Epidural Steroid Injections

T
Transduction and Encoding of Noxious Stimuli 2515
Transduction and Encoding of NoxiousStimuli
CARLOS BELMONTE, FÉLIX VIANA
Instituto de Neurociencias de Alicante, UniversidadMiguel Hernández-CSIC, San Juan de Alicante,[email protected]
Sensory Transduction
Sensory transduction is the process by which externalphysical changes are transformed into internal bio-chemical and/or electrical signals that are propagatedand processed through different levels of the centralnervous system to elicit a sensation. Despite the spe-cialized design of sensory receptors for the differentsensory modalities, the cellular and molecular mech-anisms involved in sensory transduction have certaincommon basic principles that can be outlined in a uni-fied scheme (Fig. 1) (Block 1992; Belmonte 1996).This process involves sequential detection, amplifica-tion and filtering of the incoming signal. In nociceptorneurons, as in other types of receptor neurons, thetransformation of physical and chemical stimuli intoelectrical signals normally takes place at the periph-eral nerve terminals, where the transduction machin-ery is located. This machinery is formed by a variety
Transduction and Encoding of Noxious Stimuli, Figure 1 Schematicdiagram for sensory transduction steps. Adapted from Belmonte (1996)and Block (1992).
of specialized proteins called receptors. Activation ofthese receptors by the different stimuli leads to a con-formational change in the protein, forming pores in themembrane that allow the flow of ions (ion channels).
Stimulating Energy
Nociceptors were defined by Charles Sherrington assensory receptors activated by stimuli that potentiallylead to tissue injury (noxious stimuli). Nociceptors, asin the case of other sensory receptors, are directly ac-tivated by a limited number of the various forms of en-ergy that are continuously impinging upon the exter-nal surface and internal organs of the body and onlywithin a narrow range of intensities (noxious stimuli).Effective stimuli include mechanical forces as well astemperature and a relatively large number of chemi-cal substances. The largest part of the electromagneticspectrum goes undetected by nociceptors. Only whenthe action of these otherwise unnoticed forces leads tocell damage may indirect stimulation of nociceptorsfollow, due to release of chemical mediators by injuredcells located nearby. This is the case, for instance, in ul-traviolet light or nuclear radiation exposure. What dis-tinguishes nociceptors from low threshold sensory re-ceptors isnot thephysicalnatureoftheadequatestimulibut the required threshold intensity for their activation(Belmonte 1996).
Perireceptor Elements
Nociceptorsarenaked(or free)nerveterminalsembed-ded in an intercellularmatrix thatcontainscollagen fib-rils and proteoglycansand devoid of specialized struc-tures (perireceptor elements) that in other receptors actas filters for the transmission of forces to the transduc-tion sites. Receptive areas in the nociceptor membraneappear to be discontinuous patches of bare axolemma,covered only by the basal lamina of the nerve fiber andthus exposed to the direct action of stimuli. However,macromolecules present in the intercellularmatrix sur-rounding nociceptive nerve endings may play a role inthe filtering of noxious stimuli reaching the receptivemembrane.
Transduction Molecules
The diversity of forces that selectively activate noci-ceptor endings suggests that nociceptive neurons areequipped with separate transduction mechanisms foreach of the stimulating energies. Nevertheless, someof the transducer molecules are multimodal and can beactivated by more than one class of stimulus.The detection of stimuli by nociceptor neurons isbased on membrane signaling molecules that con-vert the stimulus energy into an allosteric molecularchange, leading ultimately to the gating of membraneion channels and depolarization of the nerve terminal.

2516 Transduction and Encoding of Noxious Stimuli
Transduction and Encoding of Noxious Stimuli, Figure 2 Summary of transduction channels, modulatory molecules and voltage gated channelsinvolved in noxious stimulus transduction and modulation and encoding of propagated nerve impulses by the various classes of nociceptors.
In nociceptors, most transduction molecules are ionchannelsdirectly gated by thestimulus. Several classesof ion channels have been associated with the transduc-tion of the various forms of energy and the productionof generator potentials at nociceptor nerve terminals(Fig.2).The listof channels locatedatnociceptornerveterminals continues to grow and their functional roleis under intense scrutiny. In addition, stimuli can in-fluence other ion channels in the terminal and receptormolecules that are part of G protein signaling cascadesthatwillexertamodulatoryroleinthegenesisandprop-agation of impulse discharges.
Transduction Channels Directly Gated by the Stimulus
The � TRP superfamily of cation channels is com-posed of six subfamilies (TRPC, TRPV, TRPM, TRPP,TRPML and TRPA). Many TRPs have emerged as im-portant cellular sensors in a variety of sensory modali-ties including taste, olfaction, phototransduction, os-
mosensation, touch, hearing, thermal sensations andpain (Clapham 2003). Members of the TRPC, TRPVand TRPM families and TRPA1 (the single memberof the TRPA subfamily) are expressed in mammalianprimary sensory neurons and participate in the trans-duction of heat, cold, chemical stimuli and mechanicalforces.The degenerin/epithelium sodium channel superfam-ily (DEG/ENaC) includes five major subfamilies ofchannels (Kellenberger and Schild 2002). Mutationsin some of the members, named degenerins, makeconstitutively active channels that lead to neurode-generation. DEG/ENaC channels are gated by gen-tle mechanical forces and mediate touch in nematodesand flies. In vertebrates, the homologs of degener-ins include various subunits of Na+ selective, � acid-sensing ion channels that are blocked by amilorideand were called ASICs (Waldmann and Lazdunski1998). These subunits are encoded by 4 separate

T
Transduction and Encoding of Noxious Stimuli 2517
genes and are processed in different splice variants.Some subunits, such as ASIC1b and ASIC3, are ex-pressed almost exclusively in the peripheral nervoussystem. Others, like ASIC2a, are expressed in special-ized mechanosensory structures. These channels havetwotransmembranedomainsandare thought toensem-ble into homomeric and heteromeric combinations of4 subunits (i.e. tetramers). They are gated by protonsand give rise to transient inward currents with fast acti-vating and variable desensitizing kinetics. In additionto activation by low pH, some of these channels areactivated by mechanical forces and temperature andcould play important roles in nociception (Waldmannand Lazdunski 1998).The � two-pore domain superfamily of K+ channels(K2p) is a class of voltage insensitive, K+ selectivechannels that are open at rest. From this property theyare also known as background or leak K+ channels.They control neuronal excitability by changing theresting membrane potential and membrane resistance(Talley et al. 2003). Activity of these channels is reg-ulated by a variety of signaling molecules, includingprotons, O2 tension polyunsaturated fatty acids andphospholipids (Patel et al. 2001). They are also modu-lated by physical variables such as mechanical stretchand temperature, making them candidates for sensorytransduction channels. Various K2p subunits have beenidentified in primary sensory neurons. However, verylittle information is available regarding the expressionpattern in specific subpopulations of somatosensoryafferents. K2p2.1 (TREK-1), K2p10.1 (TREK-2) andK2p4.1 (TRAAK) may play a role in mechanosensoryand thermal transduction (Patel et al. 2001; Kang et al.2005). Some of these channels, such as members of theTASK subfamily, are closed by extracellular acidic so-lutions leading to depolarization and could be involvedin pH sensing by nociceptors (Talley et al. 2003).Several types of Ligand-gated ion channels are alsopresent in nociceptor neurons and may play a rolein transduction of stimuli. These channels open inresponse to a variety of chemical substances, someof which are released locally following tissue injury.A prominent example is offered by the family ofionotropic � purinergic receptors (P2X). These areATP gated, cation selective ion channels. Of the sevenP2X channels identified so far, all except P2X7 are ex-pressed in sensory neurons (Burnstock 2000). Onlythe P2X3 subunit is selectively expressed in nocicep-tors (Chen et al. 1995). Also, some members of theCys-loop family of transmitter-gated ion channels arepresent in nociceptive terminals. These include thecation permeable nicotinic ACh receptors and the5-HT3 serotonin receptors, which are non-selectivecation channels, gated respectively by acetylcholineand serotonin, substances that are released in inflamed
tissues. Recently, ionotropic receptors for excitatoryamino acids have been reported in nociceptive termi-nals, associated with their excitation during injury.
Remote Sensors and Modulators of the Stimulus
Activation of metabotropic membrane receptor pro-teins by endogenous substances released locally in theenvironment of nociceptive nerve endings plays an im-portant role in modulating their activity and respon-siveness following injury. These substances includepeptides, kinins, purines and excitatory amino acids. Inthiscase, theeffectsonnociceptormembraneexcitabil-ity are not exerted directly on an ion channel. Rather,the presence of an agonist substance causes the activa-tion of second messenger cascades that will finally leadto the opening or closure of ion channels. The effectscan be immediate or be the consequence of long-termchanges in gene expression that include variations inthe number of ion channels and then sustained modifi-cations of neuronal excitability.Receptors of this typeinclude thesuperfamily ofG-proteincoupled receptors(GPCR); a large number of G-protein coupled recep-tors have been shown to be present in sensory nerveendings and up- and down-modulate their excitabilityfollowing application of the specific ligand. Very of-ten sensitization is specific to a certain type of stimu-lus (mechanical versus thermal). The list ofpro-algesicsubstances includes acetylcholine, bradykinin (BK),noradrenaline, histamine, serotonin, PGE2 and nervegrowth factor (NGF).Among the GPCRs capable ofacutely sensitizing nociceptors, we count B1 and B2BK receptors and alpha-1, alpha-2 and beta-2 adren-ergic receptors. Histamine H1 receptors also sensi-tize nociceptors. The sensitization of nociceptors pro-duced by bradykinin through the activation of B2 andB1 receptors involves the phospholipase C and pro-tein kinase C signaling pathways that lead ultimately tothe modulation of the TRPV1 (capsaicin) ion channel(Chuang et al. 2001; Vellani et al. 2001).In contrast, M2 muscarinic receptors participate in thedepression of nociceptive responsiveness to heat andmechanical stimulation produced by the acetylcholineliberated during injury by skin keratinocytes and otherlocal cells. Activation of somatostatin sst2 receptors,present in about 10% of cutaneous afferent terminals,also has antinociceptive effects. Intradermal injectionof metabotropic glutamate receptor type 5 antagonistsproduces full reversal of thermal hyperalgesia in an-imal models of neuropathic pain. Opiates also exerta peripheral antinociceptive activity and these uncon-ventional effects of opioids appear to be mediated byopening of K+-ATP channels following activation ofthe arginine/NO/cGMP/ pathway by protein kinase G.Tyrosine kinase receptors, another superfamily ofmembrane receptor proteins, also participate in the

2518 Transduction and Encoding of Noxious Stimuli
modulation of nociceptor excitation. This group in-cludes trkA receptors that are activated by nervegrowth factor (NGF) released by injury in the en-vironment of nociceptive terminals. The effect ofNGF appears to be mediated in part by the samemechanism that is activated by BK, namely PLCstimulation with subsequent hydrolysis of membranephosphatidylinositol-4,5-bisphosphate (PIP2) yield-ing inositol 1,4,5-trisphosphate (IP3) and diacylglyc-erol.Onefinal targetof thiscascade is theTRPV1chan-nel, which is modulated both by PKC dependent andindependent mechanisms (Vellani et al. 2001; Chuanget al. 2001).� Proteinase-activated receptors (PARs) are alsomembers of the superfamily of G-protein coupled re-ceptors.They initiate intracellular signaling by thepro-teolytic activity of extracellular serine proteases thatcleave the N-terminus of the receptor. PAR-1 and PAR-2 are expressed in many peptidergic sensory neuronsand their activation induces neurogenic inflammationand mediates peripheral sensitization of nociceptors(Vergnolle et al. 2003).Membrane lipids are emerging as a new class of ionchannelmodulators,with important implicationsinno-ciceptor activation. A growing list of ion channels aremodulated directly by interactions with phosphatidyli-nositol 4,5-bisphosphate (PIP2) (Suh and Hille 2005).As already mentioned, a number of agonists exert theiractions by their ability to activate the phospholipase C(PLC) signaling pathway, which stimulates phospho-inositide hydrolysis. The end result is a reduction intheconcentrationsofplasmamembranephosphoinosi-tides to produce IP3. In the case of TRPV1, a reduc-tion in membrane PIP2 levels, as occurs when PLC isstimulated by NGF or BK, leads to channel activation(Chuang et al. 2001). The opposite appears to occur forTRPM8 channels, inhibited by low PIP2 levels.
Transduction Mechanisms for the Different Stimuli
Ion channels and receptor proteins of the different su-perfamilies exhibit a variable sensitivity to mechanicalforces, chemical substances, heat or cold. The charac-teristic expression patterns of these proteins in the var-ious subtypes of nociceptor neurons confer on themtheir specific transduction capabilities for the differentforms of stimulating energy.
Mechanotransduction
Propagated impulse responses to mechanical forcesare prominent in the peripheral endings of noci-ceptor neurons exclusively activated by mechanicalstimuli (� mechano-nociceptors); they also appear in� polymodal nociceptor neurons that are additionallyexcited by chemical and thermal stimuli. Mechani-cally evoked nerve impulse discharges are absent in
� ‘silent’ nociceptor nerve fibers but they are obtainedfollowing local tissue inflammation. Ion channels di-rectly gated by mechanical stimuli were first recog-nized in the 1980s but the identification of the molecu-lar entities involved in the transduction of mechanicalforces remained elusive until recently.Mechanical distortion, like stretch or pressure, pro-duces the opening of mechanically sensitive channels(mechanosensitive channels, MSCs). MSCs are a het-erogeneous population of channels with differences insensitivity, type of response, pharmacology and bio-physical properties like ionic selectivity conductanceand adaptation. They are present in a great variety ofcell types and, in addition to stimulus detection, par-ticipate in a variety of other cell functions, such asvolume regulation, cell movement, cell division, os-mosensation and contraction. The effect of mechan-ical force on a channel can be direct, leading to gat-ing by tension exerted directly on the channel proteinsor indirect, involving second messengers controlledby mechanosensitive enzymes. However, the precisemechanism coupling the supply of energy provided bythe mechanical stimulus to the gating of the channelsis still unresolved in nociceptors.The molecular identity of ion channels involved in thetransduction of lowintensity mechanical forcesbyspe-cific mechanosensory cells has been partly elucidatedin ciliated mechanosensory cells of invertebrates andin the hair cells of the auditory, vestibular and lateralline organs of vertebrates (reviewed by Corey 2003).With the application of genetic screens, Deg/ENaCchannels and TRP channels have been identified asessential for mechanosensation in different types ofmechanosensors infliesandworms. Inmostcaseshow-ever, it remains uncertain whether they are the trans-duction channels themselves and what are the specificmechanisms of activation. The fact that mechanosen-sation requires the concerted function of several pro-teins acting in an ensemble (� transduction apparatus)make these studies especially difficult.In the nematode C. elegans, gentle touch mechanosen-sation depends on members of the DEG/ENaC fam-ily. In particular, mutations in the channel subunitsMEC-4 and MEC-10 led to loss of responses to touchand abolished mechanotransduction currents. OSM-9 and OCR-2, two related TRPV channels are alsorequired for touch sensitivity in the nose region inC. elegans. In flies, TRPN1 (previously known asNompC) is essential for mechanotransduction in sen-sory bristles. This channel is also at the core ofmechanotransduction in zebra fish hair cells. Sur-prisingly, no TRPN-like genes could be found inhigher vertebrates. Also in flies, a TRPV-like chan-nel protein, NAN, is expressed selectively in chordo-tonal neurons and is essential for hearing. No ortho-

T
Transduction and Encoding of Noxious Stimuli 2519
logues of nanchung havebeen identified in vertebrates,suggesting that, in this case, hearing is mediated bydifferent transducers. Genetic screens in Drosophilalarvae for mutations that alter responses to noxiousmechanical and thermal stimuli identified Painless, aTRPAchannel (TraceyJretal.2003).Recently,TRPA1channels have been located on apical hair bundles (i.e.the mechanotransduction site) of vertebrate hair cells.Furthermore, inhibition of TRPA1 expression inhibitstransduction currents generated by the movement ofthe cilia (Corey et al. 2004).In contrast to this knowledge, the nature of the chan-nels directly involved in the transduction of mechan-ical stimuli by mammalian somatosensory endings islargely ignored. Moreover, the differences in thresholdand adaptation characteristics found between low- andhigh-threshold mechanoreceptor neurons are still un-explained at the cellular level. These differences maysimply lie in the density of mechanosensory channelsin the transducing areas, be due to the presence of dif-ferent types of mechanosensory transduction channelsor to other factors like variations in the arrangementor composition of the mechanotransduction appara-tus, cytoskeleton organization and/or second messen-ger pathways.Several channels with apparent mechanosensitivityhave been identified in primary sensory neurons ofmammals but evidence for their direct mechanicalactivation is slim in all cases. Candidate transducermolecules for mechanotransduction include TRPchannels and members of the acid-sensing ion channel(ASIC) family. Modulatory roles have been assignedto other channels like K2P channels and purinergicreceptors.Within the TRP superfamily of ion channels, TRPC1and TRPV4 appear to besensitive tomembranestretch.TRPC1 is highly expressed in frog oocyte membranesandopensin response to tensionexertedwithin the lipidbilayer (Maroto et al. 2005). TRPC1 is also found invisceral sensory neurons, including fine nerve termi-nals within the carotid body. TRPV4 opens in responseto osmoticcell swelling when expressed inmammaliancultured cells. TRPV4 is a multifunctional channel thatis also activated by warm temperatures and lipoxy-genase metabolites (Patapoutian et al. 2003). Percep-tion of noxious pressure is reduced in trpv4-/- mice,while gentle touch detection is unimpaired. However,TRPV4 doesn’t appear to be directly mechanosensi-tive; rather, its activation appears to depend on a sec-ond messenger cascade. Finally, TRPA1 channels arepresent in a large proportion of mammalian DRG neu-rons and may be activated by mechanical forces, as oc-curs in hair cells.Among channels of the DEG/ENaCs superfamily,ASIC1, ASIC2 and ASIC3 subunits are expressed
in somas and peripheral terminals of primary sen-sory neurons (reviewed by Kellenberger and Schild2002). Nonetheless, assigning a functional role to spe-cific subunits is complicated by the fact that, in mostcases, functional channels in vivo are heteromulti-meric. Somewhat predictably, knockout of individualASIC subunits in mice only leads to modest deficits intouch sensitivity. The existence of numerous touch re-ceptors with overlapping mechanical sensitivities andintermixed receptive fields limits the possibility ofmarked functional deficits in the case of single genedisruptions. In addition, properties of human subunitsare not identical to those displayed by homologs inother species. So far, there is no definitive evidencefor a major role of a particular ASIC subunit in so-matosensory mechanotransduction.Early studies sug-gested that loss of ASIC2a reduced the sensitivity oflow threshold rapidly adapting Aβ mechanoreceptorsto touch. Deletion of ASIC3, also know as DRASIC,increased the sensitivity of mechanoreceptors detect-ing light touch, but reduced the sensitivity of Aδ noci-ceptors responding to noxious pinch. However, otherstudies indicate that lack of ASIC1, ASIC2 or ASIC3does not impair cutaneous or visceral mechanosensa-tion nor does it reduce mechanogated currents in thesoma of DRG neurons (Roza et al. 2004).The roleof the lipid sensitive,mechano-gatedK+chan-nelsTREK-1,TREK-2andTRAAKoftheK2Pchannelfamily in nociceptor mechanotransduction is still opento discussion (Patel et al. 2001).P2 purinergic receptors also seem to participate in-directly in mechanotransduction, possibly modu-lating the opening probability of other classes ofmechanosensory channels. Expression of P2Y1 recep-tors in oocytes determines the development of ionicresponses to the application of light mechanical stim-uli. These stimuli appear to release intracellular ATPthat in turn activates P2Y1 receptors (Nakamura andStrittmatter1996).Similarly, releaseofATPfromdam-aged cells following mechanical injury may stimulateP2X receptors present in small unmyelinated fibersand may be involved in mechanical sensitivity of no-ciceptors (Cook and McCleskey 2002).Finally, mechanical stretching modifies the activ-ity of voltage dependent Ca2+ channels but not ofNa+ and K+ channels or K+ leakage channels. ThemechanosensitivityofCa2+ channelsmaycontribute tomodulating the neuron response to mechanical stimulithrough changes in intracellular Ca2+.Extracellular matrix attachments have been proposedas the general mechanism involved in transmittingexternal mechanical forces to the neuron surfaceand subsequently to MSCs. In turn, MSCs are teth-ered to the internal cytoskeleton. Relative displace-ment of these structures would transmit tension to
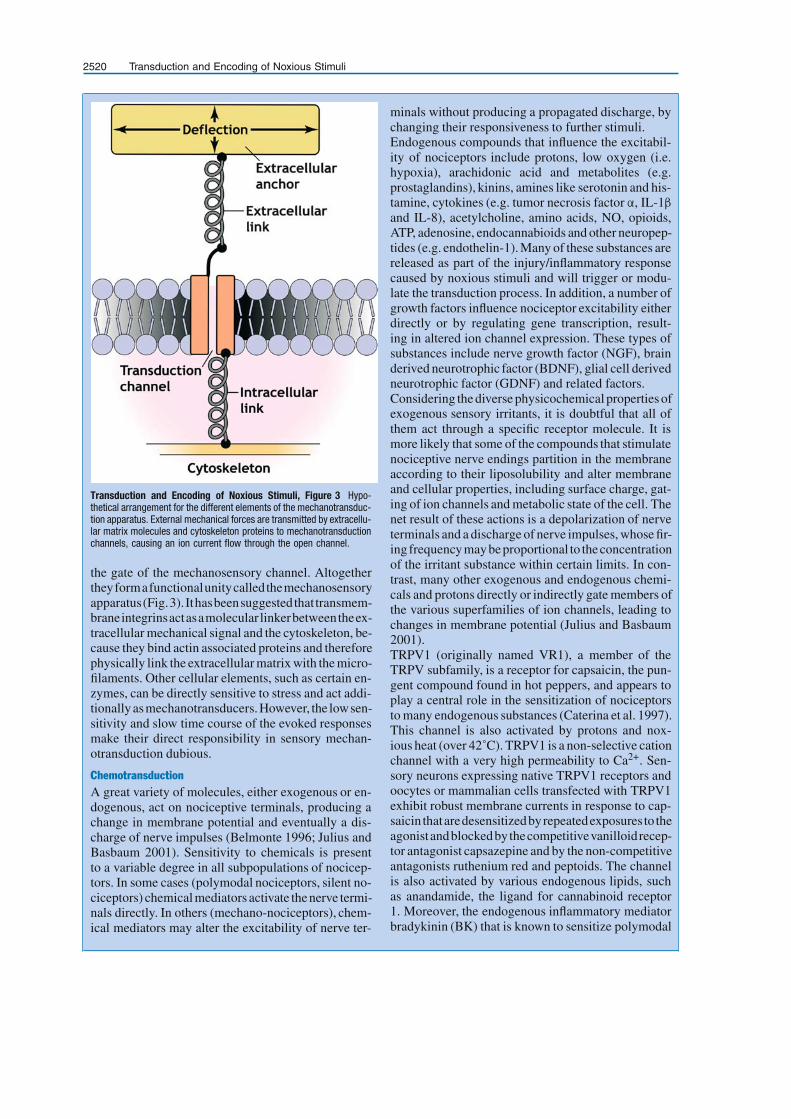
2520 Transduction and Encoding of Noxious Stimuli
Transduction and Encoding of Noxious Stimuli, Figure 3 Hypo-thetical arrangement for the different elements of the mechanotransduc-tion apparatus. External mechanical forces are transmitted by extracellu-lar matrix molecules and cytoskeleton proteins to mechanotransductionchannels, causing an ion current flow through the open channel.
the gate of the mechanosensory channel. Altogethertheyformafunctionalunitycalledthemechanosensoryapparatus(Fig.3). Ithasbeensuggestedthat transmem-braneintegrinsactasamolecular linkerbetweentheex-tracellular mechanical signal and the cytoskeleton, be-cause they bind actin associated proteins and thereforephysically link the extracellular matrix with the micro-filaments. Other cellular elements, such as certain en-zymes, can be directly sensitive to stress and act addi-tionally asmechanotransducers.However, the lowsen-sitivity and slow time course of the evoked responsesmake their direct responsibility in sensory mechan-otransduction dubious.
ChemotransductionA great variety of molecules, either exogenous or en-dogenous, act on nociceptive terminals, producing achange in membrane potential and eventually a dis-charge of nerve impulses (Belmonte 1996; Julius andBasbaum 2001). Sensitivity to chemicals is presentto a variable degree in all subpopulations of nocicep-tors. In some cases (polymodal nociceptors, silent no-ciceptors) chemical mediators activate thenerve termi-nals directly. In others (mechano-nociceptors), chem-ical mediators may alter the excitability of nerve ter-
minals without producing a propagated discharge, bychanging their responsiveness to further stimuli.Endogenous compounds that influence the excitabil-ity of nociceptors include protons, low oxygen (i.e.hypoxia), arachidonic acid and metabolites (e.g.prostaglandins), kinins, amines like serotonin and his-tamine, cytokines (e.g. tumor necrosis factor α, IL-1β
and IL-8), acetylcholine, amino acids, NO, opioids,ATP, adenosine, endocannabioids and other neuropep-tides (e.g. endothelin-1). Many of these substances arereleased as part of the injury/inflammatory responsecaused by noxious stimuli and will trigger or modu-late the transduction process. In addition, a number ofgrowth factors influence nociceptor excitability eitherdirectly or by regulating gene transcription, result-ing in altered ion channel expression. These types ofsubstances include nerve growth factor (NGF), brainderived neurotrophic factor (BDNF), glial cell derivedneurotrophic factor (GDNF) and related factors.Considering the diverse physicochemical properties ofexogenous sensory irritants, it is doubtful that all ofthem act through a specific receptor molecule. It ismore likely that some of the compounds that stimulatenociceptive nerve endings partition in the membraneaccording to their liposolubility and alter membraneand cellular properties, including surface charge, gat-ing of ion channels and metabolic state of the cell. Thenet result of these actions is a depolarization of nerveterminals and a discharge of nerve impulses, whose fir-ing frequency may beproportional to theconcentrationof the irritant substance within certain limits. In con-trast, many other exogenous and endogenous chemi-cals and protons directly or indirectly gate members ofthe various superfamilies of ion channels, leading tochanges in membrane potential (Julius and Basbaum2001).TRPV1 (originally named VR1), a member of theTRPV subfamily, is a receptor for capsaicin, the pun-gent compound found in hot peppers, and appears toplay a central role in the sensitization of nociceptorsto many endogenous substances (Caterina et al. 1997).This channel is also activated by protons and nox-ious heat (over 42˚C). TRPV1 is a non-selective cationchannel with a very high permeability to Ca2+. Sen-sory neurons expressing native TRPV1 receptors andoocytes or mammalian cells transfected with TRPV1exhibit robust membrane currents in response to cap-saicin thataredesensitizedbyrepeatedexposures to theagonistandblockedbythecompetitivevanilloidrecep-tor antagonist capsazepine and by the non-competitiveantagonists ruthenium red and peptoids. The channelis also activated by various endogenous lipids, suchas anandamide, the ligand for cannabinoid receptor1. Moreover, the endogenous inflammatory mediatorbradykinin (BK) that is known to sensitize polymodal

T
Transduction and Encoding of Noxious Stimuli 2521
nociceptor endings through activation of the G-protein-coupled receptor BK2 also enhances capsaicincurrents. A similar effect is produced by nerve growthfactor (NGF), which also sensitizes polymodal nerveendings to noxious stimuli by activation of the tyrosinekinase receptor TrkA. In addition, GDNF up-regulatesB1 receptor expression in small non-peptidergic noci-ceptive neurons. Furthermore, activation of B1 recep-tor causes a marked increase in the amplitude of theheat-activated current in these neurons. BK2 and TrkAreceptors act through the stimulation of phospholipaseC (PLC), which enhances both basal TRPV1 activityand capsaicin-evoked responses. PLC catalyzes thehydrolysis of membrane PIP2 to form IP3 and diacyl-glycerol. PIP2 inhibits TRPV1 channels directly andits hydrolysis results in TRPV1 potentiation (Chuanget al. 2001). In contrast, potentiation of TRPV1 by B2and P2Y2 receptors appears to be mediated by proteinkinase C (PKC) activation.It has been claimed that ATP released by injuredcells excites nearby nociceptive terminals. Impulse re-sponses to ATP have been evoked in the soma and neu-rites of tooth pulp cultured nociceptive neurons labeledretrogradely (Cook et al., 1997). This effect is medi-atedbyP2X3ionotropicchannels.Also, inmammaliancells expressing cloned TRPV1 channels and in DRGneurons, ATP enhances the responses to capsaicin, toheat and to protons (Moriyama et al. 2003). This effectseems to besecondary to activation of themetabotropicpurinergic receptor P2Y2, that activates phospholipaseC through a G–protein (Gq/11), leading to the produc-tion of IP3 and diacylglycerol.Among 2-pore domain K+ channels, TREK-1, TREK-2 and TRAAK are gated by different biolipids likearachidonic acid and lysophosphatidic acid (Patel et al.2001).
Acid Sensing
Intradermal injection of acidic solutions induces pain.Furthermore, tissue acidosis is a common occurrencefollowing inflammation, ischemia and tissue injury.This acidosis, to pH values as low as 5, is known toactivate and sensitize polymodal nerve terminals (Bel-monte et al. 1991). In fact, most acid sensing neuronshavesmalldiameters, typical fornociceptors.Theexci-tatory effects of protons could be mediated by a varietyof ion channels including TRPV1, ASICs and TASKchannels that are sensitive to pH changes. These find-ings are of great significance in explaining the pain andprimary hyperalgesia that appear during inflammationand ischemia.In TRPV1-transfected cells, protons evoke distinct in-ward currents and enhance the response to capsaicinup to five times over control values (Caterina et al.1997). Single channel activity in outside-out mem-
brane patches of TRPV1-transfected cells also in-creaseswhen theexternalpHis reduced. Theacid sens-ing ion channels (ASIC), members of the DEG/ENaCsuperfamily, are also activated by extracellular protonsand are over-expressed during inflammation. The con-tribution of different DEG/ENaC channel subunits topH sensitive currents in DRG neurons has been investi-gated with targeted disruption of the various subunits.Deletion of any one subunit did not abolish proton-gated currents, suggesting that two or more ASIC sub-units coassemble as heteromultimers. As mentionedabove, ASIC3 is present in presumed primary nocicep-tive neurons of the mouse and possibly participates inthedetectionofstrong localpHreductionsaccompany-ing ischemiaor inflammation.Thischannel isprobablyinvolved in the sensation of cardiac pain (Sutherland etal. 2001). The ASIC3 knockout mouse exhibits a low-ered sensitivity to intramuscular injection of acid. Fur-thermore, DRG neurons respond less to low pH solu-tion. Psychophysical and pharmacological data in hu-mans suggest that ASIC channels have a more impor-tant role than TRPV1 channels in the sensation of mod-erate cutaneous acid induced pain.Among the K2P channels, TASK subunits (TASK-1,TASK-2, TASK-3) represent background outward rec-tifiers that are constitutively active at all voltages andare inhibited by extracellular acidic pH values. Closureof these channels depolarizes the membrane potential.TASK channels have been found in some sensory neu-rons with nociceptive properties but their role in pH-induced pain has not been firmly established yet.
Thermal Transduction
Extremely low or high temperatures evoke distinctpain sensations that are mediated by activation of sub-populations of sensory afferents. Heat activates poly-modal nociceptor fibers, which respond to tempera-tures greater than 41˚–42˚C. For the sensation of painevoked by cold, it was suggested that a specific groupof cold-activated sensory fibers might exist. However,there is also evidence favoring the view that the sen-sation of cold pain rather results from the concomitantactivation by lowtemperaturesofnon-nociceptivecoldthermoreceptors and a fraction of polymodal nocicep-tors (Campero et al. 1996). The transduction mecha-nisms for heat and cold in primary sensory neurons ap-pear to be different at the cellular and molecular levels(Fig. 4). Several receptor molecules have been iden-tified recently that seem to be involved in the detec-tion of temperature changes greater than normal values(Patapoutian et al. 2003). In contrast, the cellular andmolecular mechanisms involved in the detection of in-nocuous versus noxious temperature decreases are stillincompletely understood.

2522 Transduction and Encoding of Noxious Stimuli
Transduction and Encoding of Noxious Stimuli, Figure 4 Diagram of ion channels hypothetically involved in transduction of innocuous andnoxious temperatures by peripheral sensory receptor terminals.
Heat
Temperature elevations influence ionic pumps andconductances inallcell types includingnociceptive ter-minals. However, polymodal nociceptor nerve fibers,unlike thermal sensory fibers that detect innocuouswarming, begin to discharge nerve impulses when thetissue temperature increasesover40˚C.Moreover, theybecome sensitized by repeated thermal stimulation inthe noxious range. Based on threshold, latency andpeak discharges to controlled heat pulses, two types ofimpulse responses were identified in separate groupsof Aδ nociceptors. One had a high threshold (over53˚C), a slow build-up and latencies measured in sec-onds; the second type was characterized by a lowerthreshold (46˚C), latencies measured in millisecondsand rapid peak discharges. In turn, C nociceptors be-gin to respond at around 41˚C, giving a peak dis-charge near the stimulus onset. Overall, these obser-vations suggest that various transduction mechanismsmay contribute to the final activation of nociceptorsby heat. Changes in responsiveness of polymodal no-ciceptive terminals to heat often go in parallel with amodified chemosensitivity. Thus, thermo- and chemo-sensitivities were simultaneously altered by capsaicinand exhibited cross-sensitization (Belmonte et al.1991).Theseobservationssuggested that in polymodal
neurons the specific transduction mechanisms forheat were closely associated with chemosensitivity tocapsaicin.Cesare and McNaughton (Cesare and McNaughton1996) first described an inward current in a subpop-ulation of small dorsal root ganglion neurons that wasactivated by noxiousheat, sensitizedbybradykinin andpresumably mediated by a non-selective cation chan-nel. Identification and cloning of the ‘capsaicin recep-tor’ TRPV1 (Caterina et al. 1997) proved that heat is astimulus for this channel as well as for other channelsof the TRPV family. Heat induced currents in mem-brane patches of TRPV1 transfected cells showed thesame outwardly rectifying current-voltage relations asthose evoked with capsaicin. Both were blocked bycapsazepine and displayed analogous ionic selectivi-ties. However, in transgenic mice where the TRPV1gene was disrupted, sensitivity to capsaicin was lost,while responses to noxious heat still persisted. Also, afraction of cultured primary sensory neurons respondto heat but not to capsaicin. A candidate molecule forthe additional sensitivity to noxious heat is anothervanilloid receptor-like channel named TRPV2 that is50% identical to TRPV1, responds to heat over 52˚Cand is insensitive to capsaicin, being inhibited by thenon-competitive antagonist ruthenium red (Caterina etal. 1999).

T
Transduction and Encoding of Noxious Stimuli 2523
Finally, sensitivity to rising temperatures has beenshown in two more members of the TRPV family.TRPV3 is structurally analogous to TRPV1 and isexpressed in the skin keratinocytes, the brain andother tissues. When transfected into mammalian cells,this channel responds to temperature with a thresh-old around 33˚C but not to capsaicin or pH changes,showing an activation profile closely similar to that ofTRPV1 (reviewed by Patapoutian et al. 2003). TRPV3is also activated by camphor. Although no immunore-activity to this channel can be found in the neuronsof peripheral sensory ganglia, TRPV3 null mice havestrong deficits in responses to innocuous and noxiousheat, indicating that it participates in thermosensation(Moqrichetal.2005).TRPV4,acationchannel thathasbeen implicated in hypo-osmolality sensing by neu-rons of the anterior hypothalamus, is also tempera-ture sensitive when expressed in oocytes and HEK 293cells,with an activation threshold of25˚Cand maximalresponses around 40˚C (Guler et al. 2002). Thus, trans-duction of heat by primary sensory neurons appears tobe mediated by several ion channel proteins that coverdifferent temperature ranges and may explain the dif-ferences in heat threshold among nociceptors. Forma-tion of heteromeric channel assemblies by the mem-bers of the TRPV subfamily in nociceptive neurons isan additional possibility for explaining the presence ofa range of receptors with a wide spectrum of responsesto temperature.
Cold
The identification of the cellular mechanisms involvedin the transduction of cold stimuli by low- and high-threshold cold thermal receptors has been quite elu-sive.Low-threshold(innocuous)coldsensoryfibers in-crease their firing frequency with changes of skin tem-perature as small as 0.01˚C (Gallar et al. 2003). Onthe other hand, high-threshold cold fibers respondingspecifically to cutaneous cold below 27˚C have beenreported (reviewed by Reid 2005). Finally, a fractionofpolymodalnociceptorafferentsdischargewhentheyare exposed to very low temperatures (Campero et al.1996; Acosta et al. 2001).Theunifying hypothesisoriginallyproposed to explainthe sensitivity of primary sensory neurons to low tem-peratures was that the electrogenic Na+-K+ pump washighly temperature dependent, so that temperature de-creases cause a reduction in pump activity, leading todepolarization. The differential temperature sensibil-ity of sodium and potassium ion channels would ad-ditionally contribute to a depolarized membrane po-tential during cooling. However, blockers of the Na+-K+ pump do not eliminate the sensitivity of cold sen-sory fibers to temperature decreases. More recently,it has been proposed that cold selectively activates an
ion channel of the TRP family, TRPM8 (originallynamed CMR1 because of its cold and menthol sensi-tivity, McKemy et al. 2002; Peier et al. 2002) that isfound in about 15% ofprimary sensory neurons. In het-erologous expression systems, TRPM8 channels acti-vateat temperaturesbelow25˚C,a temperaturenotablylower than the activity threshold of cold thermorecep-tors (de la Pena et al. 2005). However, this mechanismis possibly not sufficient to explain the firing charac-teristics of cold receptors and the differences in sen-sitivity to temperature of the various types of thermalreceptors. Cold also closes a background K+ current inthe soma of cultured cold sensitive neurons (Viana etal. 2002; Reid 2005), producing a net inward current(Icold) that leads to depolarization and impulse firing.This effect is partly counteracted by the prominent in-wardly rectifying Ih current present in these cells thattends to depolarize the neuron at the basal membranepotential but decreases progressively with depolariza-tion as well as as a consequence of the direct action ofcold. Depolarization by cold occurs in other types ofprimary sensory neuron but they are less sensitive thancold specificneurons,because theyexpressa slowly in-activating, transient, outward current named IKD thatreduces their excitability during cooling and preventsthe impulse discharge. Low concentrations of 4-AP se-lectively block this current and render cold insensitiveneurons responsive to cold (Viana et al. 2002).
Noxious Cold
The reduction of cutaneous surface temperature below15˚C elicits a sensation of cold pain. The discovery ofTRPA1, a TRP channel that is expressed in a fractionof primary sensory neurons, some of which also har-bor TRPV1 channels, and the fact that TRPA1 is acti-vated by temperatures below 18˚C made it a good can-didate for the transduction of noxious cold (Story et al.2003). However, TRPA1 is also gated by a number ofpungent compounds like cinnamaldehyde, mustard oiland allicin that, when applied to the skin, evoke a burn-ing rather than a cold sensation (Namer et al. 2005).These findings and the disputed activation of TRPA1by low temperature leave the specific role of TRPA1in the transduction of noxious cold temperatures un-settled. In neuropathic pain models, the time course ofcold hyperalgesia matches the expansion of a popu-lation of TRPA1 expressing neurons, but only in theuninjured ganglion (Obata et al. 2005). Cultured pri-mary sensory neurons responding to cold have a widerange of temperature thresholds from 32˚C to 18˚C,butexhibithomogeneousmembranepropertiesand fir-ing characteristics and the great majority are sensi-tive to the TRPM8 activator, menthol. About 50% ofthem are also activated by capsaicin (Viana etal. 2002).Therefore, the possibility that noxious cold is signaled

2524 Transduction and Encoding of Noxious Stimuli
by a fraction of cold sensitive neurons biophysicallyanalogous to those responding to innocuous cold butwith different central connections cannot be excluded.In that respect, it is interesting that topical applicationof menthol, the specific agonist of TRPM8 channels,canalsoinducesensationsofirritationandpain(Acostaet al. 2001; Wasner et al. 2004). Nonetheless, there isexperimental evidence that noxious cooling also acti-vates identified polymodal nociceptor fibers in animalsand humans. Thus, it is also possible that noxious coldsensations are evoked through the parallel excitationof polymodal nociceptor and innocuous cold receptorfibers by very low temperatures.
Generation and Encoding of Nerve Impulses
In nociceptors, the change in local membrane conduc-tance caused by physical and chemical stimuli is ex-pected to produce a generator potential (see genera-tor current), whose magnitude and duration will reflectthe intensity, duration and time course of the stimu-lus. These generator potentials associated with gatingof transduction channels in nociceptive terminals havenever been recorded, due to technical limitations im-posed by the small size of nociceptive endings. Thegenerator potential will in turn propagate electroton-ically and initiate, at the closest point of the axonalmembrane endowed with regenerative properties, adischarge of propagated nerve impulses, whose fre-quency of discharge is also proportional to the ampli-tude of the stimulus. The various ion channels asso-ciated with the stimulus transduction and the genera-tion of electrical activity in primary nociceptive neu-rons appear to be distributed unevenly among the pe-ripheral terminal arborizations, the parent axon and thecellbody, thusconferringdifferentelectricalpropertieson the different portions of the neuron.
Voltage-gated Channels and the Encoding Noxious Stimuli
Primary sensory neuronsalso express ion channels thatgate in response to changes in membrane potential andwhose primary role in terms of stimulus detection isnot transduction but propagation and modulation ofthe impulse discharge. They belong to the superfamilyof � voltage-gated channels that includes a large va-riety of K+, Na+ and Ca2+ channels, Cl- channels andnon-selective cation channels like the HCN channels(hyperpolarization activated cyclic nucleotide gatedK+ channels) (Hille 2001). Nociceptors have charac-teristic electrophysiological properties (Koerber andMendell 1992) and many different voltage gated ionchannels contribute to their cellular excitability.Abnormal excitability of injured neurons has beenlinked to alterations in the expression and functionalcharacteristicsofmanytypesofvoltagegated ionchan-nels (Cummins et al. 2000). Given the diversity of ion
channels in nerve terminals and their opposite effectson cell firing, the net change in excitability may not al-ways correspond to that predicted by disturbances ina single class of voltage gated ion channel.
Na+ Channels
Pharmacologically, � voltage-gated Na+ channelshave been broadly separated into two groups; one classis blocked by nanomolar concentrations of the natu-ral marine toxin tetrodotoxin (tetrodotoxin-sensitive,TTXs) while the other group of Na+ channels is resis-tant to tetrodotoxin (� tetrodotoxin resistant, TTXr).The results of various molecular approaches indicatethat nine of the ten alpha subtypes of sodium channelsare present in sensory neurons. Some of these chan-nels are found in virtually all sensory neurons, whileothers, specially the TTXr subtypes, show a more re-stricted expression pattern. The expression profile alsochanges during development.Propagation of action potentials along all sensory ax-ons is mediated by TTXs channels. In contrast, some ofthe TTXr Na+ channels subtypes (Nav1.8 and Nav1.9)are selectively found in small diameter, primarily noci-ceptive neurons (Akopian et al. 1996; McCleskey andGold 1999). Nav1.8 channels are particularly abun-dant in polymodal nociceptive terminals, where theycan sustain propagated action potentials that may con-tribute to local release of � neuropeptides (Brock et al.1998).Expression of Na+ channels is up- or down-regulatedfollowing local inflammation and injury of peripheralsensory axons (Lai et al. 2004). Functional propertiesare also affected by injury. These factors possibly con-tribute to the changes in sensitivity and the ectopic ac-tivity found in damaged sensory nerves that may giverise to � neuropathic pain. The role of Na+ channels inhyperalgesia is further substantiated by the effective-ness of use-dependent sodium channel blockers for thetreatment of various types of chronic pain. It must benoted that abnormal activity in uninjured primary af-ferents may also be critical for the observed hypersen-sitivity to sensory input in animal pain models (Lai etal. 2004).
K+ Channels
Potassium channels form a diverse superfamily of ionchannels. On the basis of their structure and func-tional properties they can be separated into voltagegated (Kv), calcium activated (KCa), inward recti-fier (Kir) and two-pore domain K2P channels. Theyare involved in a variety of neuronal functions in-cluding maintenance of membrane potential, actionpotential repolarization and regulation of firing fre-quency and arecritical regulatorsofneuronalexcitabil-ity (Hille 2001). Because K+ ions have a negative equi-

T
Transduction and Encoding of Noxious Stimuli 2525
librium potential across the plasma membrane, activa-tion of these channels tends to dampen excitation.
Voltage Gated K+ Channels (Kv)
A large number of Kv channels are expressed in noci-ceptors (McCleskey and Gold 1999). A variable com-bination of rapidly inactivating A-type (IA) and slowlyor non-inactivating (I K) K+ currents are observed inthe various types of nociceptor neurons. These func-tional currents can be further subdivided into sev-eral types with distinct kinetic and pharmacologicalcomponents. Pharmacological, functional and geneticstudies confirm that K+ channels play a role in nocicep-tor excitability. Furthermore, severalKv channel genesare down-regulated in sensory neurons following axo-tomy or chronic constriction of peripheral nerves, sug-gesting that, together with Nav channels, they partici-pate in the alteration of axonal excitability that accom-panies peripheral nerve injury (Rasband et al. 2001).Substances like PGE2 can sensitize nociceptive sen-sory neurons by reducing activity in Kv channels.Kv1.4, an A-type K+ channel, is expressed in sen-sory neurons of small size, that co-express the cap-saicin channel TRPV1 and the TTX-R Na+ channelNav1.8 and arepresumably polymodalnociceptorneu-rons (Rasband et al. 2001). Kv1.1 and Kv1.2 chan-nels are also expressed in small DRG neurons and ge-netic studies have linked the lack of Kv1.1 channelswith thermal hyperalgesia. Kv7 (also named KCNQ)channels, responsible for M-type K+ currents, are alsopresent in nociceptive sensory neurons and pharmaco-logical activation of these channels inhibits responsesto algesic substances in animal models of pain (Pass-more et al. 2003). Finally, activation of small conduc-tance calcium activated K+ channels and ATP sensi-tive K+ channels has been also implicated in antinoci-ception. Kir3.2 knockout and Kir3.3 knockout micedisplay hyperalgesia to elevated temperatures (>50˚C)suggesting implication of G-protein-gated K+ chan-nels in thermal nociception. These modulatory effectsappear to be occurring at the level of the spinal cordrather than at the periphery.
Two-pore Domain K+ Channels (KCNK)
In addition to their role as cellular sensors, these chan-nels, responsible for leak K+ currents, are key elementsin the regulation of background excitability. As such,they are thought to play an important role in triggeringectopic discharges associated with chronic pain con-ditions.
Ca2+ Channels
Voltage activated Ca2+ channels are channels withsteeply voltage dependent gates that open in response
tomembranedepolarization.Theyhavethespecial roleof translating electrical signals into chemical signalsthrough their control of the flow of Ca2+ ions into thecytoplasm, thereby regulating a variety of Ca2+ de-pendent intracellular events (Hille 2001). Functionaland pharmacological studies followed by cloning andgenetic analysis have led to the identification of alarge number of voltage gated Ca2+ channels. Sub-populations of primary sensory neurons exhibit dif-ferences in their functional type of Ca2+ channels andsome subunits are selectively expressed in nociceptors(Bell et al. 2004). The properties of voltage-gated Ca2+
channels are modulated directly and indirectly by anumber of endogenous mediators released during in-jury, thereby changing the excitability and responsive-ness of nociceptive neurons. The expression of voltagegated Ca2+ channel subunits is also altered in condi-tions such as peripheral nerve injury, contributing toabnormal nerve activity and neuropathic pain. In ad-dition,Ca2+ channelsalsoplayakeyrole inthefunctionof central neurons involved in nociceptive processing.The end result of these findings is that Cav2.1 (P/Q-type), Cav2.2 (N-type), and Cav3.x (T-type) calciumchannels have been validated as useful targets for thetreatment of pain (reviewed by Bourinet and Zamponi2005).
HCN Channels
The currents carried by hyperpolarization activated,cyclic nucleotide gated channels (HCN) have beentermed If, Ih or Iq and are poorly selective K+ currentsactivatedby membranehyperpolarization.Thecurrentreverses at membrane potentials of about -25 mV andis inwardly directing at rest. Ih currents contribute tothe resting membrane potential, input conductanceandsubthreshold membrane oscillations in many types ofneurons (Robinson and Siegelbaum 2003).HCN chan-nels, particularly HCN1 and HCN2, are abundantlyexpressed in sensory neurons. Inflammatory media-tors, such as serotonin, raise intracellular cAMP lev-els, which, in turn, increase I h current by binding tothe HCN channel, shifting its voltage dependent acti-vation to less negative potentials (reviewed by Robin-son and Siegelbaum 2003). Likewise, the amplitude ofI h currents in DRG neurons increases markedly fol-lowing spinal nerve ligation or chronic compression ofthe ganglion (Yao et al. 2003). Furthermore, pharma-cological blockade of HCN activity with the specificinhibitor ZD7288 reduces the spontaneous ectopic ac-tivity secondary to nerve injury and reverses abnormalhypersensitivity to light touch. These results suggestthat abnormal expression of HCN channels may be animportant factor in the hyperexcitability secondary toperipheral nerve injury.

2526 Transduction and Encoding of Noxious Stimuli
Transduction and Encoding of Noxious Stimuli, Figure 5 (A) Propa-gation of nerve impulses at the peripheral branches of nociceptor fibers.The stimulus extends to various branches where it generates propa-gated action potentials that travel centripetally. These action potentialsmay collide at the branching points and this determines the final firingfrequency in the parent axon (Modified from Weidner et al. 2003). (B,C) Nerve terminal impulses recorded from nociceptors endings in thecornea, before (B) and after (C) local application of lidocaine. The localanesthetic blocks the regenerative nerve impulse that normally occursin the terminal (transducing) region of the branch (b) and only impulsespassively propagated from neighbor endings are recorded (c).
Electrical Activity in Peripheral Terminal Arborizations
Parentsensoryaxonsofnociceptorsbranchextensivelyin the peripheral territory. The distribution of the vari-ous transduction and voltage gated channels is presum-ably non-homogeneous among the terminal branchesof the various subclasses of nociceptor fibers and per-haps even between branches of a single neuron. It isconceivable that in nociceptors, each terminal branchof an axonal arborization acts as an independent sitefor transduction and generation of action potentials.Action potentials of nociceptor branches travel cen-tripetally at a conduction velocity that is below that ofthe parent axon (Weidner et al. 2003). When arrivingat a branching point, impulses of the fastest brancheswillusually travelantidromicallyand invadetheslowerbranch, transiently changing its excitability or even oc-cluding the generation of action potentials (Fig. 5). Inpolymodal nociceptors, antidromic action potentialsinvade the terminals due to the high density of TTXrNa+ channels present in their membrane (Brock et al.1998).
Impulse Firing in Parent Axons and Soma
Impulses generated at the endings progress centrallythrough terminal branches of different length, conduc-tion velocity and possibly duration of their refractoryand supernormal periods, becoming finally integrated
in the parent axon. This integration will determine thefrequency and firing pattern of the discharge of prop-agated nerve impulses traveling to the CNS along thecomplete axonal path of the sensory ganglion neuron.The differences in membrane properties between sub-classes of nociceptive neurons are reflected in theconduction velocity and the shape of propagated ac-tion potentials at the parent axon and soma, far awayfrom the peripheral endings. C nociceptive axons havelonger duration APs and AP undershoot durations thanlow threshold mechanosensory axons (Koerber andMendell 1992). Further differences in axonal spikeduration and post-spike excitability are noticed be-tween classes of nociceptors (Lawson 2002), possi-bly influencing the pattern and frequency of their im-pulse discharge. The soma of the various classes ofnociceptive neurons also possesses a number of spe-cific membrane characteristics. Neurons tentativelyidentified as polymodal have slow somatic action po-tentials with a hump in the falling phase and do notpresent inward rectification in response to hyperpo-larizing pulses, while low-threshold mechanosensoryneurons produce short-lasting action potentials andhave a prominent inward rectification (Lawson 2002).These data further suggest that nociceptive neuronspossess a specific set of voltage activated Na+, K+ andCa2+ channels that confer distinct electrical propertieson them and ultimately determine their excitability andpattern of impulse firing in response to peripheral stim-uli.It is worth noticing that the electrical behavior of no-ciceptor neurons is profoundly influenced by previ-ous history. Repeated stimulation can transiently mod-ify their excitability, through changes in some of theionic mechanisms associated with the codification andpropagation of nerve impulses, contributing to fatigueand/orsensitization(Serraetal.1999).Similarchangeshave been observed after nerve damage. This plasticityis important in understanding the altered excitability ofnociceptor neurons following injury.
Conclusions
Electrophysiological, pharmacological and molecu-lar evidence suggests that nociceptor neurons pos-sess multiple mechanisms for detection, amplificationand encoding of input signals. These mechanisms willmediate transduction of the various forms of stim-ulating energy and also modulate the input signalat the successive steps of the transduction and en-coding processes. Different types of stimuli may in-teract at threshold or subthreshold levels to producepropagated responses. Short- and long-term modu-latory mechanisms will further modify the final re-sponse characterized by a discharge of nerve impulsesof a given frequency, time course and firing pattern.

T
Transduction and Encoding of Noxious Stimuli 2527
The interplay of transduction and modulatory mecha-nisms also defines other characteristics of the impulseresponse such as ongoing activity, post-discharge, sen-sitization, fatigue and inactivation.From the perspective that several molecular mecha-nismsmayco-exist innociceptiveendingsfor the trans-duction and amplification of various forms of stimulat-ing energy, a sharp categorization of nociceptors basedonly on the dominant presence of a given transduc-tionmoleculeappearstobesomewhatsimplistic.Manyof the ‘specific’ transduction molecules for a particu-lar form of energy are also present in sensory neuronsthat respondpreferentially toothermodalitiesofstimu-lus, as is the case of TRPV1 channels in cold receptorneurons. Nonetheless, it is also incomplete to distin-guish nociceptors according only to the type of stim-ulus that evokes a propagated impulse response. No-ciceptor functional subtypes categorized by this crite-rion may in fact represent cases where the transducermechanism for a given form of energy is prevalent overthe others, so that under normal circumstances onlythis form of stimulus will elicit a propagated response.When changes in excitability and/or summation of theeffects of other subthreshold stimuli take place, as isprobably the case following inflammation and axonalinjury, propagated responses evoked by other forms ofenergy may become apparent, as occurs for examplewith ‘silent’ nociceptors. In spite of their biophysicalspecialization for certain stimuli, most nociceptor neu-rons appear to be potentially polymodal and endowedwith ahigh degreeofplasticity tomodify their responseaccording to the characteristics of the stimulus and itstemporal course.
References1. Acosta MC, Belmonte C, Gallar J (2001) Sensory experiences
in humans and single-unit activity in cats evoked by polymodalstimulation of the cornea. J Physiol 534:511–525
2. Akopian AN, Sivilotti L, Wood JN (1996) A tetrodotoxin-resistant voltage-gated sodium channel expressed by sensoryneurons. Nature 379:257–262
3. Bell TJ, Thaler C, Castiglioni AJ et al. (2004) Cell-specificalternative splicing increases calcium channel current densityin the pain pathway. Neuron 41:127–138
4. Belmonte C (1996) Signal transduction in nociceptors: generalprinciples. In: Belmonte C, Cervero F (eds) Neurobiology ofNociceptors. Oxford University Press, New York, pp 243–257
5. Belmonte C, Gallar J, Pozo MA et al. (1991) Excitation by ir-ritant chemical substances of sensory afferent units in the cat’scornea. J Physiol 437:709–725
6. Block S (1992) Biophysical principles of sensory transduction.In: Corey D, Roper SD (eds) Sensory Transduction. The Rock-efeller University Press, New York, pp 1–17
7. Bourinet E, Zamponi GW (2005) Voltage gated calcium chan-nels as targets for analgesics. Curr Top Med Chem 5:539–546
8. Brock JA, McLachlan EM, Belmonte C (1998) Tetrodotoxin-resistant impulses in single nociceptor nerve terminals inguinea-pig cornea. J Physiol 512:211–217
9. Burnstock G (2000) P2X receptors in sensory neurones. Br JAnaesth 84:476–488
10. Campero M, Serra J, Ochoa JL (1996) C-polymodal nocicep-tors activated by noxious low temperature in human skin. JPhysiol 497:565–572
11. Caterina MJ, Schumacher MA, Tominaga M et al. (1997) Thecapsaicin receptor: a heat-activated ion channel in the pain path-way. Nature 389:816–824
12. Caterina MJ, Rosen TA, Tominaga M et al. (1999) A capsaicin-receptor homologue with a high threshold for noxious heat.Nature 398:436–441
13. Cesare P, McNaughton P (1996) A novel heat-activated cur-rent in nociceptive neurons and its sensitization by bradykinin.Proc Natl Acad Sci USA 93:15435–15439
14. Chen CC, Akopian AN, Sivilotti L et al. (1995) A P2Xpurinoceptor expressed by a subset of sensory neurons. Na-ture 377:428–431
15. Chuang HH, Prescott ED, Kong H et al. (2001) Bradykininand nerve growth factor release the capsaicin receptor fromPtdIns(4,5)P2-mediated inhibition. Nature 411:957–962
16. Clapham DE (2003) TRP channels as cellular sensors. Nature426:517–524
17. Cook SP, McCleskey EW (2002) Cell damage excites nocicep-tors through release of cytosolic ATP. Pain 95:41–47
18. Cook SP, Vulchanova L, Hargreaves KM et al. (1997) DistinctATP receptors on pain-sensing and stretch-sensing neurons.Nature 387:505–508
19. Corey DP (2003) New TRP channels in hearing andmechanosensation. Neuron 39:585–588
20. Corey DP, Garcia-Anoveros J, Holt JR et al. (2004) TRPA1 isa candidate for the mechanosensitive transduction channel ofvertebrate hair cells. Nature 432:723–730
21. Cummins TR, Dib-Hajj SD, Black JA et al. (2000) Sodiumchannels and the molecular pathophysiology of pain. ProgBrain Res 129:3–19
22. de la Pena E, Malkia A, Cabedo H et al. (2005) The contributionof TRPM8 channels to cold sensing in mammalian neurones.J Physiol 567:415–426
23. Gallar J, Acosta MC, Belmonte C (2003) Activation of scleralcold thermoreceptors by temperature and blood flow changes.Invest Ophthalmol Vis Sci 44:697–705
24. Guler AD, Lee H, Iida T et al. (2002) Heat-evoked activationof the ion channel, TRPV4. J Neurosci 22:6408–6414
25. Hille B (2001) Ion Channels of Excitable Membranes. SinauerAssociates, Sunderlad, MA
26. Julius D, Basbaum AI (2001) Molecular mechanisms of noci-ception. Nature 413:203–210
27. Kang D, Choe C, Kim D (2005) Thermosensitivity of the two-pore domain K+ channels TREK-2 and TRAAK. J Physiol564:103–116
28. Kellenberger S, Schild L (2002) Epithelial sodium chan-nel/degenerin family of ion channels: a variety of functionsfor a shared structure. Physiol Rev 82:735–767
29. Koerber HR, Mendell LM (1992) Functional heterogeneity ofdorsal root ganglion cells. In: Scott SA (ed) Sensory Neu-rons Diversity, Development, and Plasticity. Oxford UniversityPress, New York, pp 77–96
30. Lai J, Porreca F, Hunter JC et al. (2004) Voltage-gated sodiumchannels and hyperalgesia. Annu Rev Pharmacol Toxicol44:371–397
31. Lawson SN (2002) Phenotype and function of somatic primaryafferent nociceptive neurones with C-, Adelta- or Aalpha/beta-fibres. Exp Physiol 87:239–244
32. Maroto R, Raso A, Wood TG et al. (2005) TRPC1 forms thestretch-activated cation channel in vertebrate cells. Nat CellBiol 7:179–185
33. McCleskey EW, Gold MS (1999) Ion channels of nociception.Annu Rev Physiol 61:835–856
34. McKemy DD, Neuhausser WM, Julius D (2002) Identificationof a cold receptor reveals a general role for TRP channels inthermosensation. Nature 416: 52–58

2528 Transforaminal Interbody Fusion
35. Moqrich A, Hwang SW, Earley TJ et al. (2005) Impaired ther-mosensation inmice lacking TRPV3, a heat andcamphorsensorin the skin. Science 307:1468–1472
36. Moriyama T, Iida T, Kobayashi K et al. (2003) Possible involve-ment of P2Y2 metabotropic receptors in ATP-induced transientreceptor potential vanilloid receptor 1-mediated thermal hyper-sensitivity. J Neurosci 23:6058–6062
37. Nakamura F, Strittmatter SM (1996) P2Y1 purinergic recep-tors in sensory neurons: contribution to touch-induced impulsegeneration. Proc Natl Acad Sci USA 93:10465–10470
38. Namer B, Seifert F, Handwerker HO et al. (2005) TRPA1 andTRPM8 activation in humans: effects of cinnamaldehyde andmenthol. Neuroreport 16:955–959
39. Obata K, Katsura H, Mizushima T et al. (2005) TRPA1 inducedin sensory neurons contributes to cold hyperalgesia after in-flammation and nerve injury. J Clin Invest 115:2393–2401
40. Passmore GM, Selyanko AA, Mistry M et al. (2003) KCNQ/Mcurrents in sensory neurons: significance for pain therapy. JNeurosci 23:7227–7236
41. Patapoutian A, Peier AM, Story GM et al. (2003) ThermoTRPchannels and beyond: mechanisms of temperature sensation.Nat Rev Neurosci 4:529–539
42. Patel AJ, Lazdunski M, Honore E (2001) Lipid and mechano-gated 2P domain K(+) channels. Curr Opin Cell Biol13:422–428
43. Peier AM, Moqrich A, Hergarden AC et al. (2002) A TRP chan-nel that senses cold stimuli and menthol. Cell 108:705–715
44. Rasband MN, Park EW, Vanderah TW et al. (2001) Distinctpotassium channels on pain-sensing neurons. Proc Natl AcadSci USA 98:13373–13378.
45. Reid G (2005) ThermoTRP channels and cold sensing: whatare they really up to? Pflugers Arch 451:250–263
46. Robinson RB, Siegelbaum SA (2003) Hyperpolarization-Activated Cation Currents: From Molecules to PhysiologicalFunction. Ann Rev Physiol 65:453–480
47. Roza C, Puel JL, Kress M et al. (2004) Knockout of the ASIC2channel in mice does not impair cutaneous mechanosensation,visceral mechanonociception and hearing. J Physiol (Lond)558:659–669
48. Serra J, Campero M, Ochoa J et al. (1999) Activity-dependentslowing of conduction differentiates functional subtypes of Cfibres innervating human skin. J Physiol 515:799–811
49. Story GM, Peier AM, Reeve AJ et al. (2003) ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated bycold temperatures. Cell 112:819–829
50. Suh BC, Hille B (2005) Regulation of ion channels byphosphatidylinositol 4,5-bisphosphate. Curr Opin Neurobiol15:370–378
51. Sutherland SP, Benson CJ, Adelman JP et al. (2001) Acid-sensing ion channel 3 matches the acid-gated current in car-diac ischemia-sensing neurons. Proc Natl Acad Sci USA98:711–716
52. Talley EM, Sirois JE, Lei Q et al. (2003) Two-pore-Domain(KCNK) potassium channels: dynamic roles in neuronal func-tion. Neuroscientist 9:46–56
53. Tracey WD, Jr., Wilson RI, Laurent G et al. (2003) Painless, aDrosophila gene essential for nociception. Cell 113:261–273
54. Vellani V, Mapplebeck S, Moriondo A et al. (2001) Proteinkinase C activation potentiates gating of the vanilloid receptorVR1 by capsaicin, protons, heat and anandamide. J Physiol534:813–825
55. Vergnolle N, Ferazzini M, D’Andrea MR et al. (2003)Proteinase-activated receptors: novel signals for peripheralnerves. Trends Neurosci 26:496–500
56. Viana F, de la Pena E, Belmonte C (2002) Specificity of coldthermotransduction is determined by differential ionic channelexpression. Nat Neurosci 5: 254–260
57. Waldmann R, Lazdunski M (1998) H(+)-gated cation channels:neuronal acid sensors in the NaC/DEG family of ion channels.Curr Opin Neurobiol 8:418–424
58. Wasner G, Schattschneider J, Binder A et al. (2004) Topicalmenthol–a human model for cold pain by activation and sen-sitization of C nociceptors. Brain 127:1159–1171
59. Weidner C, Schmidt R, Schmelz M et al. (2003) Action po-tential conduction in the terminal arborisation of nociceptiveC-fibre afferents. J Physiol (Lond) 547:931–940
60. Yao H, Donnelly DF, Ma C et al. (2003) Upregulation of thehyperpolarization-activated cation current after chronic com-pression of the dorsal root ganglion. J Neurosci 23:2069–2074
Transforaminal Interbody Fusion� TLIF
Transforaminal Steroids� Lumbar Transforaminal Injection of Steroids
Transformed Migraine� Chronic Daily Headache in Children� Transformed Migraine Headache
Transformed Migraine Headache
Definition
Classification proposed by Silberstein et al (1994),describing a subtype of chronic daily or near daily
headache (> 15 days per month) that developed gradu-allyover timefromapre-existing,well-definedmigraineheadache. Headache is longer than 4 hours per day, caninclude a mixture of autonomic and tension-type symp-toms, and symptoms have progressed with increasingfrequency and decreasing severity over at least threemonths.� Chronic Daily Headache in Children
Transganglionic Transport
Definition
Oneof the tract tracingmethods,e.g.aneural tracer takenup by a peripheral axon or branches of primary afferentsis transportedcentripetallyandthencentrifugally totheircentral terminals.� Trigeminal Brainstem Nuclear Complex, Anatomy

T
Transient Headache and CSF Lymphocytosis 2529
Transgene
Definition
Any gene inserted into a vector that is expressed in cellsin vitro or animals in vivo following vector-mediatedtransduction.� Opioids and Gene Therapy
Transgenic Knockout Mice
Definition
Genetically engineered null mutant mice. Via blastocystmicroinjection of transgenic (i.e. possessing an alteredDNA sequence) embryonic stem cells, mice can be cre-ated that lack all expression of a particular gene. Thesemice can be evaluated for their altered pain sensitivitycompared to „wild-type“ mice, and the function of themissing gene/protein inferred.� Heritability of Inflammatory Nociception� NerveGrowthFactorOverexpressingMiceasModels
of Inflammatory Pain
Transgenic Mice
� NerveGrowthFactorOverexpressingMiceasModelsof Inflammatory Pain
Transient Headache and CSFLymphocytosis
ZAZA KATSARAVA, HANS-CHRISTOPH DIENER
Department of Neurology, University Hospital ofEssen, Essen, [email protected]
Synonyms
Migraine With Pleocytosis, Pseudomigraine With Lym-phocyctic Pleocytosis
Definition
The International Headache Society proposed the fol-lowing diagnostic criteria for HaNDL (IHS 2003):
a) Moderate or severe � headacheb) Cerebrospinal fluid pleocytosis with lymphocytic
predominance (greater than 15 cells/μl) and nor-mal neuroimaging, CSF culture and other tests foretiology
c) Headache occurs at time of CSF pleocytosisd) Episodes of headache and neurological deficits last
< 3 months.
Characteristics
HaNDL is a transient syndrome characterized by severeheadache, focal neurological symptoms and CSF lym-phocytosis (IHS 2003; for review see Pascual and Valle2003).The HaNDL syndrome was first described by Bartlesonet al. (Bartleson et al. 1981). Until now about 100 casesof HaNDL have been published. Fifty patients have beenreported by Gomez-Aranda et al. (Gomez-Aranda et al.1997).BergandWilliamspresentedaseriesof7patients,as well as a systemic review of the literature (Berg andWilliams 1995).Age of onset of HaNDL ranges between 7 and 50 years,withameanof27years.HaNDLismorefrequent inmen,with a male to female ratio of 3:1 (Berg and Williams1995; Gomez-Aranda et al. 1997).Approximately 25–30% of patients report a viral-likeillness 2–3 weeks before the onset of HaNDL. Typi-cal symptoms are cough, rhinitis, diarrhea and fatigue(Gomez-Aranda et al. 1997).The timing between the headache and neurologicdeficit is variable. In some patients the initial symptomis headache, in others neurologic deficits occur first. Insome patients neurologic deficits with CSF lymphocy-tosis can occur without a severe headache (Oldani et al.1998).Headache is moderate to severe, pulsating or throbbingand mostly bilateral. In some patients the headache isunilateral and localized contralateral to the neurologicsymptoms.Durationofheadacherangesbetween1hand1week,onaverage19h(Gomez-Arandaetal.1997).Ac-companying symptoms are nausea and vomiting, photo-and phonophobia.Neurologic signs usually consist of hemiparesis,hemisensory symptoms or aphasia. Sensory symp-toms are more frequent than motor signs. Pure motoraphasia is the most frequent speech disorder followedby global aphasia and pure sensory aphasia (Gomez-Aranda et al. 1997). Visual symptoms are rare. The focalneurologic symptoms usually evolve progressively, insome cases, however, suddenly. The deficits are tran-sient, lasting from 5 minutes to hours and at most 3days. Atypical cases with longer duration up to a weekhave been described. The recovery is always complete.Routine blood and immunological tests are usuallynormal. Two exceptional cases associated with a cy-tomegalovirus infection have been reported (Ferrariet al. 1983; Richert et al. 1987). The examination ofcerebrospinal fluid reveals a lymphocytic pleocyto-sis (range 10–760/μl) and increased total protein. In50% of the cases an elevated CSF pressure has beenmeasured (Berg and Williams 1995; Gomez-Aranda etal. 1997). Immunological, bacterial and viral studiesproduce normal results.Electroencephalographyreveals a unilateral focal slow-ing during the acute HaNDL phase in up to 70% of the

2530 Transient Receptor Potential
patients (Berg and Williams 1995; Gomez-Aranda et al.1997). In 10% � EEG slowing was observed bilaterally(Gomez-Aranda et al. 1997). The changes normalizedagain after the symptomatic period.Cranial computed tomography, as well as cranialmagnetic resonance imaging, is usually normal. Ina few patients nonspecific small areas of high signalin T2-weighted images have been observed (Berg andWilliams 1995; Gomez-Aranda et al. 1997). Very re-cently, in one patient, a diffusion weighted MRI couldbe performed during acute HaNDL, which did notreveal diffusion changes (Gekeler et al. 2002).Single photon emission computed tomography hasbeen suggested to be the most informative neuroimag-ing technique. A focus of decreased tracer uptake hasbeen detected during the acute phase, which becomesnormal several days after recovery (Caminero et al.1997; Fuentes et al. 1998).Cerebral angiography is usually not informative. In thevast majority of patients studied no changes could befound. In some patients inflammation like processes inthe wall of the opercular arteries have been detected, andin some patients new episodes of HaNDL were triggeredby the angiography, therefore it has to be avoided.The pathophysiology of HaNDL is still unclear. Thereare some obvious similarities to � migraine with aura.However, only few patients with HaNDL report ahistory of migraine. The duration of the focal neuro-logical symptoms in HaNDL is longer than in migrainewith aura. The most important difference between theHaNDL and migraine is the CSF pleocytosis. HaNDLhas also been separated from other diseases such asfamilial hemiplegic migraine or progressive cerebellarataxia.There are several infectious diseases that can causeheadache, focal neurologic symptoms and CSF pleocy-tosis.Mollaret’smeningitis ischaracterized by recurrentepisodes of aseptic meningitis with headache and symp-toms of meningeal irritation. However, focal neurologi-cal symptoms are not typical for Mollaret’s meningitis.Furthermore, the pleocytosis in Mollaret’s meningitisis a mixture of lymphocytes, polymorphonuclear- andlarge ‘Mollaret’s’ cells.HaNDL has to be separated from further infectiousconditions such as Lyme disease (Pal et al. 1987), neu-rosyphilis (Berg and Williams 1995), neurobrucellosis(Roldan-Montaud et al. 1991), mycoplasma infection(Dalton and Newton 1991), granulomatous meningitis(Mayer et al. 1993), and secondary cytomegalovirusencephalitis associated with human immunodeficiencyvirus (HIV) infection (Richert et al. 1987).HaNDL is always a self-limiting disorder. Therefore,the therapy is restricted to symptomatic treatment withanalgesics and antiemetics. No preventive treatment isneeded.
References1. Bartleson JD, Swanson JW, Whisnant JP (1981) A Migrainous
Syndrome with Cerebrospinal Fluid Pleocytosis. Neurology31:1257–1262
2. Berg M, Williams L (1995) The Transient Syndrome of Headachewith Neurologic Deficits and CFS Lymphocytosis. Neurology45:1648–1954
3. Caminero AB, Pareja JA, Arpa J et al. (1997) MigrainousSyndrome with CSF Pleocytosis. SPECT Findings. Headache37:511–515
4. Dalton M, Newton RW (1991) Aseptic Meningitis. Dev MedChild Neurol 33:446–451
5. Ferrari MD, Buruma OJ, van Laar-Ramaker M et al. (1983) AMigrainous Syndrome with Pleocytosis. Neurology 33:813
6. Fuentes B, Diez Tejedor E, Pascual J et al. (1998) Cerebral BloodFlow Changes in Pseudomigraine with Pleocytosis Analyzed bySingle Photon Emission Computed Tomography. A SpreadingDepression Mechanism? Cephalalgia 18:570–573, discussion531
7. Gekeler F, Holtmannspotter M, Straube A et al. (2002) Diffusion-Weighted Magnetic Resonance Imaging during the Aura of Pseu-domigraine with Temporary Neurologic Symptoms and Lympho-cytic Pleocytosis. Headache 42:294–296
8. Gomez-Aranda F, Canadillas F, Marti-Masso JF et al. (1997)Pseudomigraine with Temporary Neurological Symptomsand Lymphocytic Pleocytosis. A Report of 50 Cases. Brain120:1105–1113
9. IHS (2003) Headache Classification Committee of the Interna-tional Headache Society. Classification and Diagnostic Crite-ria for Headache Disorders, Cranial Neuralgias and Facial Pain.Cephalalgia (in press)
10. Mayer SA, Yim GK, Onesti ST et al. (1993) Biopsy-Proven Iso-lated Sarcoid Meningitis. Case Report. J Neurosurg 78:994–996
11. Oldani A, Marcone A, Zamboni M et al. (1998) The TransientSyndrome of Headache with Neurologic Deficits and CSF Lym-phocytosis. Report of a Case without Severe Headache. Headache38:135–137
12. Pal GS, Baker JT, Humphrey PR (1987) Lyme Disease Presentingas Recurrent Acute Meningitis. Br Med J (Clin Res Ed) 295:367
13. Pascual J, Valle N (2003) Pseudomigraine with LymphocyticPleocytosis. Curr Pain Headache Rep 7:224–228
14. Richert JR, Potolicchio S Jr, Garagusi VF et al. (1987) Cy-tomegalovirusEncephalitisAssociated withEpisodic NeurologicDeficits and OKT-8+ Pleocytosis. Neurology 37:149–152
15. Roldan-Montaud A, Jimenez-Jimenez FJ, Zancada F et al. (1991)Neurobrucellosis Mimicking Migraine. Eur Neurol 31:30–32
Transient Receptor Potential
Definition
Transient change in electrical potential across the cellmembrane, for example, induced by activation of a TRPreceptor.� TRPV1 Receptor, Species Variability
Transient Receptor Potential Family ofIon Channels
Definition
The mammalian transient receptor potential (TRP) ionchannels are named after the role of these channels inDrosophila phototransduction. They are encoded by

T
Transition from Acute to Chronic Pain 2531
at least 21 different channel genes. The TRP channelprimary structures predict six transmembrane domains,with a pore domain between the fifth and sixth segments,and both the C- and N-termini are located intracellularly.The mammalian TRP channel family is comprised ofthree subfamilies, including TRPC, TRPV and TRPM.The family members are at least 25% homologouswithin their amino acid sequences. Most of the chan-nels are non-selective to cations, allowing sodium andcalcium to flow in and depolarize neurons. The mostwell-characterized TRP channels in DRG neurons arethe vanilloid family of TRPV1–4 channels, which areactivated by a range of heat and/or warm temperatures.TRPV1 is the prototype vanilloid channel and is acti-vated by noxious heat, acidic pH and the alkaloid irritantcapsaicin. Additional TRP channels in DRG neuronsare TRPM8, which is activated by cool temperaturesand menthol, and ANKTM1, which is activated bymustard oil derivatives and may be activated by cold.� Immunocytochemistry of Nociceptors� TRPV1 Receptor, Species Variability
Transient Receptor Potential Vanilloid 1(TRPV1 or VR1) Receptor
� TRPV1
Transition from Acute to Chronic PainSARAH J. HARPER, STEPHAN A. SCHUG
Royal Perth Hospital and University of WesternAustralia, Perth, WA, [email protected]
Synonyms
Pain Progression; Pain Chronification; Central sensiti-sation
Definition
It is now well established that chronic pain can developas the consequence of repeated or severe episodes ofacute pain, e.g. in the context of trauma, surgery or acutepainful illness. This progression is most likely to be theconsequence of central nervous processes, commonlycalled central � neuroplasticity or sensitisation.
Characteristics
It is well recognised that chronic pain states often followan acutely painful stimulus such as surgery or trauma.A large study of over 5000 patients referred to chronicpain clinics in the UK, revealed that 22.5% of these pa-tients had developed their pain after surgery, and 18.7%after trauma (Crombie et al. 1998). Similar observations
have been made when following patients after surgery ortrauma; and severe early pain after thoracotomy (Katz etal. 1996) and orthopaedic trauma (Gehling et al. 1999)predicts development of chronic pain states.
Risk Factors for Transition to Chronic Pain States
In 2000,Perkinsand Kehletpublisheda� meta-analysisof the predictive factors of chronic pain after surgery(Perkins and Kehlet 2000). They identified a number ofsignificant risk factors for this transition, which includetype of surgery, preoperative, intraoperative and postop-erative factors:
Type of Surgery
After lower limb amputation phantom limb pain occursin 30–81% and stump pain in >50%. Post-thoracotomypain syndrome (PTPS) occurs in >50% of patients.Breast surgery can give rise to chest wall, scar, breastor shoulder pain in 11–57% of patients, and phantombreast pain in 13–24%; post-mastectomy pain syndrome(PMPS) has an overall incidence of 50% at one year(Kwekkeboom 1996). Gallbladder surgery carries a riskbetween 3 and 56% of post-cholecystectomy syndrome(PCS).
Preoperative Factors
Intense preoperative pain increases the incidence ofphantom limb pain (from 33% to 72%) (Nikolajsenet al. 1997; Krane and Heller 1995). Preoperative riskfactors for PCS include ‘psychologic vulnerability’, fe-male gender and long-standing preoperativesymptoms.Other factors are repeat surgery and issues of compen-sation. Preoperative epidural pain control may decreasethe risk of chronic pain after amputation (Schug 2004,Bach et al. 1988).
Intraoperative Factors
Technical issuesof thesurgery, including thesurgicalap-proach and the risk of nerve injury, are important here.Video-assisted-thoracoscopic lung surgery reduces therisk of PTPS compared to open lung resection. Positionof the incision for thoracotomy may influence the inci-dence of PTPS. Intraoperative epidural analgesia seemsto reduce the risk of PTPS. The incidence of PMPS maybe increased by: breast-conserving surgery, immediateinsertion of implants, extent of axillary dissection anddamage to the intercostobrachial nerve.
Early Postoperative Factors
Stump pain at one week post amputation correlates withthe risk ofphantom pain. Intensityofacutepostoperativepain is an independent predictor for PTPS and PMPS.Intercostal nerve dysfunction (loss of the superficial ab-dominal reflex) is associated with more acute, subacuteand chronic pain. Adjuvant postoperative radiotherapyto the breast increases the risk of development of PMPS.

2532 Transition from Acute to Chronic Pain
Late Postoperative Factors
Long-term stump pain predicts long-term phantom limbpain (Nikolajsen et al. 1997). More severe or prolongedacute pain in the postoperative period as well as postop-erative complications, commonly leading to increased� nociception, significantly predict the development ofchronicpain after surgery(Katzetal.1996,Gehlingetal.1999).Otherrelevantfactorsareradiotherapyandneuro-toxic chemotherapy, but also psychological factors suchas anxiety, depression, neuroticism and psychologicalvulnerability.
Mechanisms for the Transition from Acute to ChronicPain
The development of chronic pain is based on the phe-nomenon of central neuroplasticity. Disruption of thenormalspecialisationof thesomatosensorysystemleadsto increasing mismatch between stimulus and response.The underlying mechanisms are not fully elucidatedyet, however, the physiological principles have beenreviewed in detail (Pockett 1995). The implications forthe development of chronic pain as a separate diseaseentity have been summarised recently by Siddall andCousins (Siddall and Cousins 2004). Some of thesemechanisms are presented in the following:
Wind Up
A progressive increase in the number of action poten-tials elicited per stimulus occurs in dorsal and ventralhorn neurons when the stimulus exceeds 0.5 Hz. Abovethis frequency, the post-synaptic depolarising responsessummate to produce a cumulative � depolarisation, re-sulting in a burst of action potentials, instead of a singleaction potential, in response to each stimulus. It is medi-ated via N-methyl-D-aspartic acid (NMDA) glutamatereceptors and therefore blocked and reversed by NMDAantagonists. � Wind-up lasts as long as ventral horn celldepolarisation, i.e. about 60 seconds.
Long Term Potentiation
Repeated episodes of wind-up may trigger � long-termpotentiation (LTP). It was first studied in the hippocam-pus and is now known to occur in visual, sensorimotorand pre-frontal cortex, as well as in the spinal cord. Itsmechanism is complex, but in essence high-frequencypre-synaptic activity causes a pre-synaptic glutamaterelease, which activates α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptors. AMPAreceptor activation opens ion channels allowing post-synaptic depolarisation. If the depolarisation reachesa certain threshold, a magnesium-dependent block ofNMDA receptors is released, and these then open theirassociated ion channels. There is an overall influx ofcalcium ions which triggers additional calcium releasefrom intracellular calcium stores. The intracellular cal-cium rise triggers a complex chain of events, whichincludes the release of one or more retrograde factors
by the post-synaptic cell. By diffusing back to thepre-synaptic membrane, these cause an increased trans-mitter release in response to each pre-synaptic actionpotential. Subsequently, calcium-dependent enzymesare activated, such as protein kinases A and C (PKA,PKC) and calcium/calmodulin kinase, leading to phos-phorylation of membrane proteins including receptorsand ion channels. This makes the post-synaptic cellmore excitable, and upregulation of AMPA receptorsand growth of dendrites/spines on the post-synaptic celloccurs.The overall result is LTP, which can last from one hourto several months. It can be slowed or prevented fromoccurring in vitro by NMDA antagonists, early coolingor PKC inhibitors, but in contrast to wind up cannot bereversed.
Recruitment
Chronic inflammation and nerve injury have an ef-fect on the presence and distribution of voltage-gatedsodium channels, which can become concentrated inareas of injury and produce ectopic discharges. Studieshave shown that neurone-specific sodium channelsbecome concentrated in neurones proximal to a siteof nerve injury, and play a role in the � hyperalgesiaand � allodynia of chronic pain states. Not all sensoryneurons are active all the time, and this � peripheralsensitisation will ‘recruit’ dormant nociceptors, thusincreasing the receptive fields of dorsal horn neuronsand increasing the intensity and area of pain (Mannionand Woolf 2000).
Immediate Early Gene Expression
Immediate early genes are a family of genes (e.g. c-fos,c-jun) that share thecharacteristicofhaving theirexpres-sion rapidly and transiently induced upon stimulationof neuronal and non-neuronal cells (Caputto and Guido2000). Damaged sensory neurones may undergo alteredgeneexpression, such that they releaseadifferent typeofneurotransmitter. The release of neurotransmitters usu-ally associated with noxious stimuli, such as substanceP, may contribute to � central sensitisation.A change in gene expression can also lead to up or down-regulation of ion channels leading to changes in cellularexcitability.
Excitotoxicity� Excitotoxicity is a phenomenon that was first de-scribed by Olney in the seventies (Olney et al. 1972).It involves the activation of glutamate receptors in thecentral nervous system (CNS). Glutamate, an excita-tory amino acid, activates different types of ion channelforming receptors to develop their essential role in thefunctional activity of the brain. However, high concen-trations of glutamate or neurotoxins acting at the samereceptors, cause cell death by apoptosis through theexcessive activation of these receptors.

T
Transsynaptic Changes after Peripheral Nerve Injury 2533
The physiological role of the NMDA receptor seemsto be related to synaptic plasticity and learning. Inaddition, working together with G-protein coupledglutamate receptors, it ensures the establishment of thelong-term potentiation phenomenon (LTP) describedabove. Research into the phenomenon has focused onfinding clinically useful NMDA receptor antagonists,for use in both chronic pain conditions and neurodegen-erative disorders in which excitotoxicity plays a part,such as � Parkinson’s disease and Alzheimer’s disease(Sureda 2000).
Conclusion
These mechanisms imply that chronic pathologicalpain may persist long after the initial noxious insulthas ceased and tissue damage has healed. The processof synaptic plasticity and learning begins early andis difficult to reverse. It seems that untreated acutepain persisting for long periods of time can imprintmemory-like processes into the central nervous system(Schug 2004). Currently the recommendation is thatan extended, balanced, multi-modal approach to painmanagement should begin in the preoperative periodand continue postoperatively.
References1. Bach S, Noreng MF, Tjellden NU (1988) Phantom Limb Pain
in Amputees during the First 12 Months following Limb Am-putation, After Preoperative Lumbar Epidural Blockade. Pain33:297–301
2. Caputto BL, Guido ME (2000) Immediate early gene expres-sion within the visual system: light and circadian regulationin the retina and the suprachiasmatic nucleus. Neurochem Res25:153–162
3. Crombie IK, Davies HT, Macrae WA (1998) Cut and Thrust:Antecedent Surgery and Trauma amongst Patients Attending aChronic Pain Clinic. Pain 76:167–171
4. Gehling M, Scheidt C-E, Neibergall H et al. (1999) PersistentPain after Elective Trauma Surgery. Acute Pain 2:110–114
5. Katz J, Jackson M, Kavanaugh B, Sandler A (1996) Acute Painafter Thoracic Surgery Predicts Long-Term Post-ThoracotomyPain. Clin J Pain 12:50–55
6. Krane EJ, Heller LB (1995) The Prevalence of Phantom LimbSensation and Pain in Pediatric Amputees. J Pain Symptom Man-agement 10:21–29
7. Kwekkeboom K (1996) Postmastectomy Pain Syndromes. Can-cer Nurs 19:37–43
8. Mannion RJ, Woolf CJ (2000) Pain mechanisms and manage-ment: A central perspective. Clin J Pain 16(3 suppl):S144–156
9. Nikolajsen L, Ilkjaer S, Kroner K et al. (1997) The Influenceof Preamputation Pain on Postamputation Stump and PhantomPain. Pain 72:393–405
10. Olney JW, Sharpe LG, Feigin RD (1972) Glutamate-inducedbrain damage in infant primates. J Neuropathol Exp Neurol31:464–488
11. Perkins FM, Kehlet H (2000) Chronic Pain as an Outcomeof Surgery – A Review of Predictive Factors. Anesthesiology93:1123–1133
12. Pockett S (1995) Spinal Cord Synaptic Plasticity and ChronicPain. Anesth Analg 80:173–179
13. Schug SA (2004) Acute Pain Management – Its Role in thePrevention of Chronic Pain States. ASEAN J Anaesthesiol5:166–169
14. Siddall PJ, Cousins MJ (2004) Persistent pain as a disease entity:implications for clinical management. Anesth Analg 99:510–520
15. Sureda FX (2000) Excitotoxicity and the NMDA receptor.From EUROSIVA meeting Vienna. www.eurosiva.org/Archive/Vienna/abstracts/Speakers
Transition from Parenteral to OralAnalgesic Drugs
� Postoperative Pain, Transition from Parenteral to Oral
Translaminar Epidural Steroid Injection
� Epidural Steroid Injections for Chronic Back Pain
Transmitters in the Descending Circuitry
� Descending Circuitry, Transmitters and Receptors
Transmucosal
Definition
Absorption across the buccal mucosa.� Pain Control in Children with Burns
Transmucosal Fentanyl
Definition
Transmucosal fentanyl is also available as a lollipoppreparation (transmucosal preparation) that has beensuccessfully used in children.� Postoperative Pain, Fentanyl
Transmural
Definition
Passing through a wall, as of the body or of a cyst or anyhollow structure.� Animal Models of Inflammatory Bowel Disease
Transsynaptic Changes after PeripheralNerve Injury
� Central Changes after Peripheral Nerve Injury

2534 Traumatic Angiospasm
Traumatic Angiospasm
� Complex Regional Pain Syndromes, General Aspects
Traumatic Nerve Endbulb Pain
� Neuroma Pain
Treating Pediatric Burns
� Pain Control in Children with Burns
Treatment Adherence
Definition
The extent to which patients or clients follow treatmentrecommendations made by the clinician, or treatmentgoals negotiated with the clinician.� Chronic Pain, Patient-Therapist Interaction
Treatment Alliance
Definition
The extent to which patients and clinicians agree ontreatment goals and hold each other in positive regard.� Chronic Pain, Patient-Therapist Interaction
Treatment Matching
Definition
Prescribing treatments based on specific features ofpatients that are believed to be important to outcomes.Treatments may be matched on the basis of physi-cal, behavioral, or any unique individual differencesassociated with pain and disability.� Psychological Assessment of Pain
Treatment of Neuropathic Pain
� Phantom Limb Pain, Treatment
Treatment of Phantom Pain
� Phantom Limb Pain, Treatment
Treatment Outcome
� Psychological Treatment of Chronic Pain, Predictionof Outcome
Treatment Outcome Research
� Psychology of Pain, Efficacy
Treatment-Related Neutropenia
Definition
Low white blood counts that develops after chemother-apy or radiation therapy.� Cancer Pain Management, Orthopedic Surgery
Tremor
� Orofacial Pain, Movement Disorders
Tricyclic AntidepressantsMARK JOHNSTON
Musculoskeletal Physician, Hibiscus Coast Highway,Orewa, New [email protected]
Synonyms
TCAs; tricyclics
Definition
Tricylic antidepressants are a group of drugs developedand prescribed primarily as antidepressants. It was dis-covered in the 1960s that they had an analgesic effect,which was separate from their antidepressant effect.They have since been used increasingly as an analgesicin managing chronic pain.TCAs consist of a 3 ring central core (Fig. 1). Theirproperties are related to the degree of saturationof the terminal amine. Tertiary amines available atpresent include; amitriptyline, clomipramine, dox-epine, imipramine and trimipramine. Some of thesecompounds are metabolised to active secondary aminetricylic compounds e.g. imipramine to desipramine andnortriptyline from amitriptyline.

T
Tricyclic Antidepressants 2535
Tricyclic Antidepressants, Figure 1 The core chemical structure ofthe tricyclic antidepressants, showing the three-ring structure. Individualagents differ in their substitution of carbon or nitrogen in the central ring,and in the radicle on the amine chain.
Characteristics
Mechanisms
Whereas their antidepressant action takes about 2–3weeks to develop, the analgesic effect of TCAs oc-curs in 3–7 days. For many years, the most appealinghypothesis was that the analgesic and antidepressanteffects are related to the block of reuptake of nore-pinephrine and serotonin (5–HT) at spinal dorsal hornsynapses. Presynaptically, TCAs inhibit the reuptakeof serotonin, norepinephrine, and to a lesser degree,dopamine. Postsynaptic activity is variable. Amitripty-line blocks cholinergic, histamine, alpha adrenergic,muscarinic, N-methyl-D-aspartate (NMDA), substanceP, and various subsets of serotonergic receptors.Recent evidence from animal studies has shown thatother mechanisms may also be having antinociceptiveeffects. These include: modulation of the sympatheticnervous system, blockade of sodium channels, someanti-inflammatory effects, and mechanisms involvingGABA or opioid receptors (Cohen and Abdi 2001).
Routes of Administration
TCAs are only administered orally. However, intra-venous preparations can be obtained.
Applications
TCAs have been recommended and used in many painconditions, but this is not supported by the evidence. Ev-idence supports their use only for:
• diabetic neuropathy• postherpetic neuralgia• tension-type headache• prevention of migraine and tension-type headache
There is no evidence, or no more than dubious evidence,for the use of TCA in: acute or chronic low back pain,‘rheumatologic pain’ (including fibromyalgia), andchronic facial pain.Combining TCAswith other analgesicshasbeen recom-mended in patients with chronic pain, but there are nogood studies to support this practice.
Side Effects
Cardiovascular
The most serious side effects of TCAs are on the car-diovascular system, where they can cause heart block,arrhythmias, and postural hypotension.TCAs slow conduction through the heart. The effectis more marked in patients with pre-existing cardiacconduction disease, particularly bundle branch block.Ischemic heart disease is frequently associated withconduction defects. Some 18% of depressed patientswith ischemic heart disease, treated with nortriptyline,developed potentially dangerous sinus tachycardia orcomplex arrythmias, compared with 2% in those treatedwith paroxitene (Roose et al. 1998). Treating the elderlyis a problem, as ischemic heart disease is common, andas many episodes may be silent it may not be obvi-ous from the history that the patient is suffering fromischemic heart disease.Postural hypotension is a result of α-adrenergic receptorblockade. Syncope can occur at any age, but it is moreserious in the elderly. The elderly are more prone to syn-cope, as part of the normal aging process. Other diseases(congestive heart failure etc.) and medication (vasodila-tors, diuretics etc.) may intensify the normal posturalblood pressure drop. The consequences of falls are alsomore serious in the elderly.
Sedation
Sedation can be a useful side effect if the patient has in-somnia. Sedation is caused by blockade of H1 receptors.Different TCAs produce different degrees of sedation.The sedative effects may diminish after several weeks.
Sexual Dysfunction
TCAs have all been reported to delay or prevent orgasmin both sexes.
Psychiatric
Manic episodes may be triggered in patients with bipo-lar disease. A greater concern is that of a possible sui-cide attempt using TCAs, in someone who has severedepression. In large doses the cardiovascular effects canbe lethal.
Withdrawal Syndromes
Sudden or gradual discontinuation of TCAs may cause anumber of symptoms. These include: nausea, vomiting,headache, malaise, sleep disturbance, akathisia, or para-doxical behavioural activation resulting in hypomanicsymptoms. These effects may start within 24 to 48 hours

2536 Tricyclic-Type Antidepressants
of the last dose and last up to 1 month. To reduce with-drawal symptoms, the dose should be tapered gradually;if symptoms are upsetting, the dose may even be in-creased a little.
Serotonin Syndrome
Combining TCAs with selective serotonin re-uptakeinhibitors or monoamine reuptake inhibitors may pre-cipitate a “serotonin overload syndrome”, characterisedby myoclonus, hyperreflexia, tremor, increased muscletone, fever, shivering, sweating, diarrhoea, delirium,coma or death (Bodner et al. 1995). The condition isusually reversible if the drugs involved are stopped.
Efficacy
For the treatment of chronic low back pain, a systematicreview (van Tulder et al. 1997) found moderate evidencethat TCAs are not effective for chronic LBP. There is noevidence of efficacy in acute low back pain.For � neuropathic pain, TCAs are considered ‘adjuvantanalgesics’. A systematic review (McQuay et al. 1996)concluded that TCA are effective for the treatment ofneuropathic pain: “of 100 patients . . . 30 will obtainmore than50%painrelief”Evidence isstrongest forpainrelief with amitriptyline and desipramine.For � fibromyalgia, TCAs provide pain relief and im-provesleep (O’Malley etal. 2000).The� NNTwas4 forthe outcome ‘to obtain significant benefit’.A meta-analysis (Tomkins et al. 2001) of the outcomesfrom TCAs for the treatment of chronic headacheshowed an NNT of 3.2, for ‘improvement in headaches’.The effect was the same for TCA and serotonin antago-nists, and forboth tension-likeheadachesand migraines.
Selection
When deciding which TCA to use, the broader spectrumones, such as amitriptyline, imipramine and nortripty-line, have greater efficacy than the selective re-uptakeblockers. Nortriptyline may be the best as it has less se-dation, less postural hypotension and less anticholiner-gic effects.� Antidepressants in Neuropathic Pain� Drugs Targeting Voltage-Gated Sodium and Calcium
Channels� Drugs with Mixed Action and Combinations, Empha-
sis on Tramadol� Postoperative Pain, Postamputation Pain, Treatment
and Prevention
References1. Bodner RA, Lynch T, Lewis L, Kahn D (1995) Serotonin Syn-
drome. Neurology 45:219–2232. Cohen SP, Abdi S (2001) New Developments in the Use of Tri-
cyclic Antidepressants for the Management of Pain. Curr OpinAnaesthesiol 14:505–511
3. McQuay HJ, Tramer M, Nye BA, Carroll D, Wiffen J, Moore RA(1996) A Systematic Review of Antidepressants in NeuropathicPain. Pain 68:217–227
4. O’Malley PG, Balden E, Tompkins G, Santoro J, Kroenke K,Jackson JL (2000) Treatment of Fibromyalgia with Antidepres-sants: A Meta-Analysis. J Gen Inter Med 15:659–666
5. Roose SP, Laghrissi-Thode F, Kennedy JS, Nelson JC, Big-ger JT, Pollock BG, Gaffney A, Narayan M, Finkel MS,McCafferty J, Gergel I (1998) Comparison of Paroxitene andNortriptyline in Depressed Patients with Ischemic Heart Disease.JAMA 279:287–291
6. Tomkins GE, Jackson JL, O’Malley PG, Balden E, Santoro JE(2001) Treatment of Chronic Headache with Antidepressants: AMeta-Analysis. Am J Med 111:54–63
7. van Tulder MW, Koes BW, Bouter LM (1997) ConservativeTreatment of Acute and Chronic Non-Specific Low Back Pain.Spine 22:2128–2156
Tricyclic-Type Antidepressants
� Antidepressant Analgesics in Pain Management
Tricyclics
� Tricyclic Antidepressants
Trigeminal Brainstem Nuclear Complex,Anatomy
YOSHIO SHIGENAGA, ATSUSHI YOSHIDA
Department of Oral Anatomy and Neurobiology,Osaka University, Osaka, [email protected],[email protected]
Definition
The trigeminal brainstem nuclear complex in mam-mals is generally recognized to include the principalnucleus (Vp), the spinal nuclear complex (Vsp; some-times also termed the spinal tract nucleus), and the� mesencephalic nucleus (Vmes). It extends throughthe pons and medulla from the C2 spinal segment andupward through the mesencephalon. In a caudoros-tral direction, these nuclei are the Vsp, Vp, and Vmes(Fig. 1). Three further divisions of the Vsp are differ-entiated (Olszewski 1950): the subnuclei caudalis (Vc)(Fig. 1e), interpolaris (Vi) (Fig. 1d), and oralis (Vo)(Fig. 1b, c). The Vmes differs from the other nuclei inthat it contains cell bodies of primary afferents inner-vating jaw muscle spindles or periodontal ligaments.
Characteristics
The Vp and Vsp integrate the trigeminal afferent systemthat is organized to serve the � exteroceptive, interocep-tive, and proprioceptivesensoryfunctionsof theoral andcraniofacial structures, but also receive projections from

T
Trigeminal Brainstem Nuclear Complex, Anatomy 2537
Trigeminal BrainstemNuclear Complex, Anatomy,Figure 1 Camera lucida drawingsillustrating the distribution ofhorseradish peroxidase (HRP)reactions produced at differentlevels of the trigeminal brainstemnuclear complex after injections ofHRP into the lower tooth pulps. (a) to(e) are arranged rostrocaudally. BC,brachum conjunctivum; S, nucleusof solitary tract; SO, superior olivarynucleus; Vtr, spinal trigeminal tract;VII, facial nucleus; 5, trigeminalmotor nerve; 7, facial nerve. D-L,dorsal-lateral. Scale bar = 0.5 mm.(Modified from Fig. 1 in Shigenagaet al., 1986c).
the facial, glossopharyngeal and vagus nerves and uppercervical nerves.The trigeminal spinal tract (Vtr) is somatotopically or-ganized, with fibers of the ophthalmic, maxillary, andmandibular divisions lying successively more dorsally.This dorsoventral arrangement is also applicable to thefacial region of theVp and Vsp,while the intraoral repre-sentation is organized in a complex manner (see below)(Shigenaga et al. 1986a; Shigenaga et al. 1986b; Shige-naga et al. 1989b).Primary afferent projections are not identical at eachlevel, as the density of terminals varies along differ-ent nuclei, or in different parts of the same nucleus(Tsuru et al. 1989). The individual nuclei are connectedby ascending and descending internuclear pathways(Nasution and Shigenaga 1987).
Nucleus Principalis (Vp)
This is divided into a subnucleus dorsomedialis (Vpd)and a subnucleus ventrolateralis (Vpv; Fig. 1a). TheVpv extends further caudally than the Vpd, to the rostralpole of the facial nucleus. The caudal limit of Vpd corre-sponds to thecaudalpoleof the trigeminalmotornucleus(Vmo). The neurons are densely packed and have a uni-form appearance, with small and medium-sized, roundor oval cell bodies. The Vpd is distinguished by thecompact aggregation of its cells, although caudally itbecomes loosely arranged (Shigenaga et al. 1986b).The central projections of primary afferents have beenexamined using the techniques of � transganglionictransport and intraaxonal labeling of horseradish per-oxidase (HRP). The Vpd receives a projection from

2538 Trigeminal Brainstem Nuclear Complex, Anatomy
trigeminal primary afferents innervating intraoral struc-tures, but both the intraoral and facial afferents projectto the Vpv (Shigenaga et al. 1986b). Both the subnucleireceive mesencephalic afferent projections (Shigenagaet al. 1988; Shigenaga et al. 1989a). Tooth pulp afferentsterminate in the Vpd in the cat (Shigenaga et al. 1989b;Westrum et al. 1981), and in the Vpd and Vpv in the rat(Marfurt and Turner 1984).Intraaxonal labeling shows that stained axons (centralprocesses) of primary afferents in the Vtr ascend anddescend (bifurcating fibers), or descend without bifur-cation (descending non-bifurcating fibers) (Tsuru et al.1989; Hayashi 1985). The ascending fiber and the de-scending fiber give off axon collaterals mainly in the Vpand the Vsp, respectively.At the electron microscopic level (Bae et al. 2003),all pulpal afferent boutons in rats contain clear, roundvesicles (S-type), and make synaptic contact withnon-primary dendrites with � asymmetric junctions.Approximately one-third of the boutons show charac-teristic glomeruli, in which synaptic contact is madewith small dendrites as well as with other axon ter-minals. The � synaptic glomerulus has an importantrole to activate multiple second-order neurons at thesame time. The presynaptic axon terminals contain� pleomorphic vesicles and form symmetric contactswith the postsynaptic membrane. The presynapticboutons are immunoreactive for GABA. These ultra-structural features are also common in axon terminalsfrom low-threshold mechanoreceptive afferents.The distribution of cell bodies of trigeminothalamicneurons has been mapped in cats with the � retrogradelabeling technique (Shigenaga et al. 1983). It was foundthat most Vpv neurons project of the contralateral tha-lamus to the nucleus ventralis posteromedialis (VPM)via the crossed ventral tract, while most Vpd neuronsproject to the ipsilateral VPM via the uncrossed dorsaltract. Thus sensory information from the intraoral struc-tures is mediated through both crosswed (via Vpv) anduncrossed (via Vpd) pathways, whereas facial sensoryinformation is mediated via Vpv by a single crossedpathway, although in the rat most Vpd neurons projectinto the crossed pathway. The axons of the crossedpathway give off collaterals in the central lateral nu-cleus (CL) of the intralaminar complex (Shigenagaet al. 1983). An � intracellular labeling study in cats(Yoshida et al. 1998) has shown that the Vp also contains� local-circuit cells whose axon collaterals terminatein the jaw-closing region of the Vmo and the lateralreticular formation.
Subnucleus Oralis (Vo)
This extends from the caudal tip of the facial nucleus tothe level a little posterior to the caudal end of Vmo andthus, the rostral half of the Vo (termed Vo.r) is situated inthe region dorsomedial to the Vpv (Fig. 1b). Its caudallimit corresponds to the rostral pole of the facial nucleus.
The Vo.r is characterized by large � multipolar cells andmerges medially with the lateral reticular formation.More caudally, two regions represent subgroupings, aventrolateral region (Vo.c) and a dorsomedial region(Vo.dm, Fig. 1c). The Vo.dm is composed mainly ofsmall, compactly arranged cells and merges with theventrolateral border of the solitary nucleus. In the cat,this subdivision is present between the levels corre-sponding to the facial nucleus, whereas in the rat itcontinues without change of structure caudally into theVi. The Vo.c is composed of oval- or spindle-shapedsmall cells, triangular or fusiform medium-sized cells,and large multipolar cells. The large cells are sparselyscattered throughout this subdivision (Shigenaga et al.1986b). The Vo.r and Vo.dm receive projections fromprimary afferents innervating intraoral structures aswell as from mesencephalic primary afferents (Shige-naga et al. 1989a), while facial afferents project to theVo.c. A dorsoventral organization is not apparent in theVo.r and Vo.dm. In cats, pulpal afferents terminate inthe Vo.r and Vo.dm, where the upper and lower teethare represented in a mediolateral sequence, and projec-tions from the anterior to posterior teeth are organizedin a ventrolateral to dorsomedial sequence, with anextensive overlap in projections from adjoining teeth(Shigenaga et al. 1989b).At the electron microscopic level (Bae et al. 2003),synaptic organization of rat pulpal afferent boutons inthe Vo differs from that in Vp, in that the number ofpostsynaptic elements per bouton and the frequencyof axoaxonic contacts are lower in the Vo, indicatingless frequent synaptic glomeruli. These ultrastruc-tural features are common to those of low-thresholdmechanoreceptive afferent terminals in the Vp and Voin cats.Vo neurons, especially in Vo.r and Vo.dm, send few ax-ons to the VPM in cats (Shigenaga et al. 1983), while aconsiderable number of Vo.r neurons project to the CLof the intralaminar complex.There are, however, species differences, and a signif-icant projection to VPM might exist in the rat (see� Trigeminal Brainstem Nuclear Complex, Physiol-ogy). In addition, the Vo.r and Vo.dm contain pre-motoneurons projecting to either jaw-closing or jaw-opening regions of the motor nucleus (Yoshida etal.1994). Although many neurons respond to light me-chanical stimulation of intraoral structures, some areactivated by noxious stimuli. These nociceptive neuronssend their axon collaterals to the jaw-closing region ofthe V motor nucleus, suggesting that they may be in-volved in a reflex circuit that modulates jaw-closingalpha-motoneurons.
Subnucleus Interpolaris (Vi)
This lies between the Vo and Vc and ends a little cau-dal to the obex, and is composed of three neural popula-tions with small, medium, and large cells. Its dorsome-
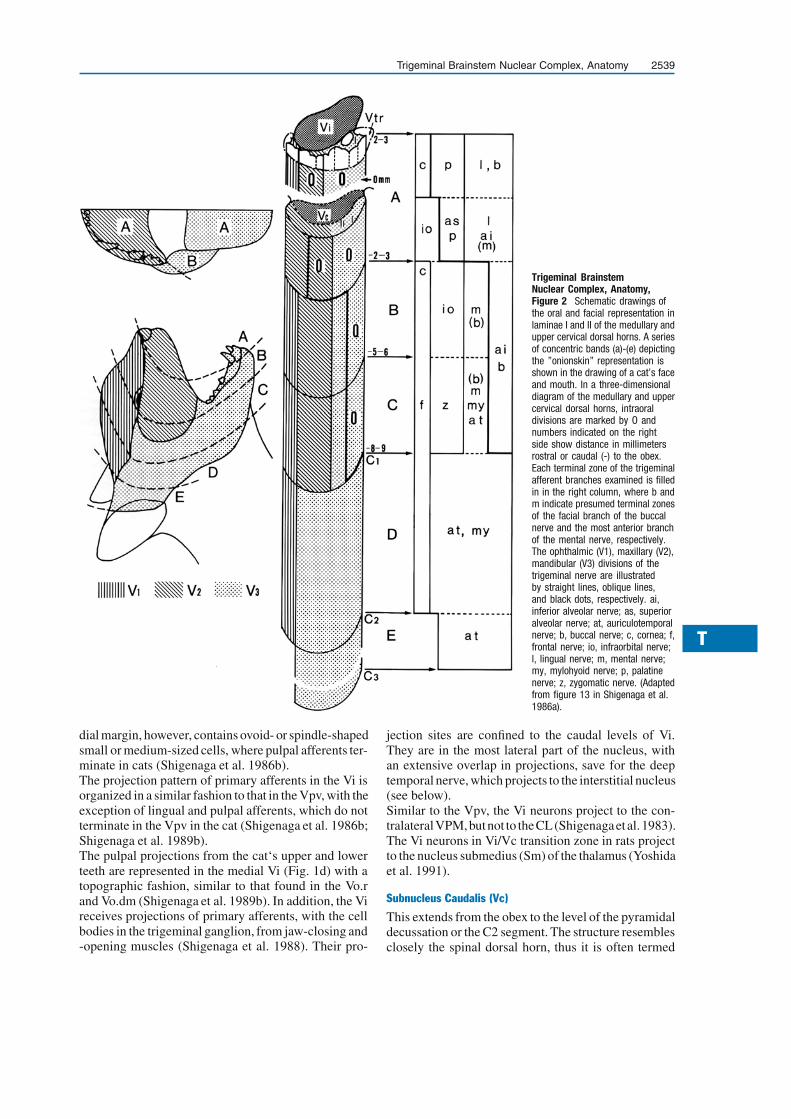
T
Trigeminal Brainstem Nuclear Complex, Anatomy 2539
Trigeminal BrainstemNuclear Complex, Anatomy,Figure 2 Schematic drawings ofthe oral and facial representation inlaminae I and II of the medullary andupper cervical dorsal horns. A seriesof concentric bands (a)-(e) depictingthe ”onionskin” representation isshown in the drawing of a cat’s faceand mouth. In a three-dimensionaldiagram of the medullary and uppercervical dorsal horns, intraoraldivisions are marked by O andnumbers indicated on the rightside show distance in millimetersrostral or caudal (-) to the obex.Each terminal zone of the trigeminalafferent branches examined is filledin in the right column, where b andm indicate presumed terminal zonesof the facial branch of the buccalnerve and the most anterior branchof the mental nerve, respectively.The ophthalmic (V1), maxillary (V2),mandibular (V3) divisions of thetrigeminal nerve are illustratedby straight lines, oblique lines,and black dots, respectively. ai,inferior alveolar nerve; as, superioralveolar nerve; at, auriculotemporalnerve; b, buccal nerve; c, cornea; f,frontal nerve; io, infraorbital nerve;l, lingual nerve; m, mental nerve;my, mylohyoid nerve; p, palatinenerve; z, zygomatic nerve. (Adaptedfrom figure 13 in Shigenaga et al.1986a).
dial margin, however, contains ovoid- or spindle-shapedsmall or medium-sized cells, where pulpal afferents ter-minate in cats (Shigenaga et al. 1986b).The projection pattern of primary afferents in the Vi isorganized in a similar fashion to that in the Vpv, with theexception of lingual and pulpal afferents, which do notterminate in the Vpv in the cat (Shigenaga et al. 1986b;Shigenaga et al. 1989b).The pulpal projections from the cat‘s upper and lowerteeth are represented in the medial Vi (Fig. 1d) with atopographic fashion, similar to that found in the Vo.rand Vo.dm (Shigenaga et al. 1989b). In addition, the Vireceives projections of primary afferents, with the cellbodies in the trigeminal ganglion, from jaw-closing and-opening muscles (Shigenaga et al. 1988). Their pro-
jection sites are confined to the caudal levels of Vi.They are in the most lateral part of the nucleus, withan extensive overlap in projections, save for the deeptemporal nerve, which projects to the interstitial nucleus(see below).Similar to the Vpv, the Vi neurons project to the con-tralateralVPM,butnot to theCL(Shigenagaetal. 1983).The Vi neurons in Vi/Vc transition zone in rats projectto the nucleus submedius (Sm) of the thalamus (Yoshidaet al. 1991).
Subnucleus Caudalis (Vc)
This extends from the obex to the level of the pyramidaldecussation or the C2 segment. The structure resemblesclosely the spinal dorsal horn, thus it is often termed

2540 Trigeminal Brainstem Nuclear Complex, Anatomy
the medullary dorsal horn. Olszewski (1950) dividedthe Vc into three laminar zones: marginalis, gelati-nosus, and magnocellularis. The Rexed laminationscheme of the spinal dorsal horn is also applicable tothe Vc, and the marginalis, gelatinosus, and magnocel-lularis correspond to lamina I, lamina II, and laminaeIII/IV, respectively (Fig. 1e). In the most rostral part,cell islands of lamina I (termed interstitial nucleus)are seen intercalated in the dorsal, lateral and ventralparts of the Vtr at the caudal levels of Vi. The detailedmorphology of cells in the different laminae has beenreported by several studies (Gobel et al. 1981; Rene-han et al. 1986). In cats, laminae I, outer II and V (thelateral parts of medullary reticular formation) receiveinputs from small A-delta fibers and c-fibers, whichconvey information about pain or thermal sensation.In contrast, low-threshold mechanoreceptive afferents(A-beta) terminate throughout inner lamina II to laminaV. However, rat laminae III/IV also receive input fromnociceptive primary afferents (Jacquin et al. 1988).Pulpal afferent boutons in the superficial layers in ratscan be classified into those with clear, round (S-type)vesicles and dense-cored vesicles, in contrast to pulpalboutons in the rostral nuclei (Bae et al. 2003). Un-myelinated nociceptive primary afferents have beenfound to bear terminal varicosities that contain a largenumber of dense-cored vesicles as well as clear, roundvesicles (Bae et al. 2003). In addition, the occurrence ofsynaptic glomeruli and of axoaxonic contacts is muchless frequent in the Vc than in the Vp.The projection patterns of primary afferents in the Vchave also been examined in cats using the techniqueof transganglionic transport of HRP (Shigenaga et al.1986a; Shigenaga et al. 1988; Shigenaga et al. 1989b).All three trigeminal divisions terminate throughoutlaminae I-V, with the exception of tooth pulp, jaw mus-cle and corneal afferents, which terminate in laminaeI, outer II, and V. However, the mediolateral arrange-ments and caudal extensions differ between the differentnerves or branches.Theintraoralandfacialstructuresarearrangedasaseriesof concentric semicircular rings that are centered aroundthe midline of the most anterior face and mouth, and arerepresented, especially in laminae I/II, in a consecutiveorder. In this way, the midline of the mouth and the mostanterior face are represented most rostrally, while morelateral or posterior structures are represented at succes-sively more caudal levels in themedullary and uppercer-vical dorsal horns (Fig. 2). The fact that neurons in lam-inae I, outer II and V receive inputs from nociceptiveor thermoreceptive afferents, and that lamina I neuronsproject to the VPM (Shigenaga etal. 1983),Sm (Yoshidaet al. 1991) and in monkey, the posterior ventral medialnucleus (VMpo) (Beggsetal. 2003), support theconceptthat the onionskin-like organization of pain and thermalsensations is defined by the arrangement of the sensoryprojections to lamina I and outer lamina II. An anatom-
ical substrate for referred pain phenomena may be pro-vided by the extensive mediolateral overlap in projec-tions from different nerve branches. Neurons in laminaeIII and IV project to the rostral nuclei (Nasution and Shi-genaga 1987) and medullary reticular formation.
References1. Bae YC, Kim JP, Choi BJ et al. (2003) Synaptic Organization
of Tooth Pulp Afferent Terminals in the Rat Trigeminal SensoryNuclei. J Comp Neurol 463:13–24
2. Beggs J, Jordan S, Ericson AC et al. (2003) Synaptology ofTrigemino- and Spinothalamic Lamina I Terminations in the Pos-terior Ventral Medial Nucleus of the Macaque. J Comp Neurol459:334–354
3. Gobel S, Hockfield S, Ruda MA (1981) An Anatomical Analysisof the Similarities between Medullary and Spinal Dorsal Horns.In: Kawamura Y and Dubner R (eds) Oral-Facial Sensory andMotor Functions. Quintessence, Tokyo, pp 211–223
4. Hayashi H (1985) Morphology of Terminations of Small andLarge Myelinated Trigeminal Primary Afferent Fibers in the Cat.J Comp Neurol 240:71–89
5. Jacquin MF, Stennett RA, Renehan WE et al. (1988) Structure-Function Relationships in the Rat Brainstem Subnucleus Interpo-laris: II. Low and High Threshold Trigeminal Primary Afferents.J Comp Neurol 267:107–130
6. Marfurt CF, Turner DF (1984) The Central Projections of ToothPulp Afferent Neurons in the Rat as Demonstrated by the Trans-ganglionic Transport of Horseradish Peroxidase. J Comp Neurol223:535–547
7. Nasution ID, Shigenaga Y (1987) Ascending and DescendingInternuclear Projections within the Trigeminal Sensory NuclearComplex. Brain Res 425:234–247
8. Olszewski J (1950) On the Anatomical and Functional Or-ganization of the Spinal Trigeminal Nucleus. J Comp Neurol92:402–413
9. Renehan WE, Jacquin MF, Mooney RD, Rhoades RW (1986)Structure-Function Relationships in Rat Medullary and CervicalDorsal Horns. II. Medullary Dorsal Horn Cells. J Neurophysiol55:1187–1201
10. Shigenaga Y, Chen IC, Suemune S et al. (1986a) Oral and FacialRepresentation within the Medullary and Upper Cervical DorsalHorns in the Cat. J Comp Neurol 243:388–408
11. Shigenaga Y, Doe K, Suemune S et al. (1989a) Physiologicaland Morphological Characteristics of Periodontal MesencephalicTrigeminal Neurons – Intra-Axonal Staining with HRP. Brain Res505:91–110
12. Shigenaga Y, Nakatani Z, Nishimori T et al. (1983) The Cellsof Origin of Cat Trigeminothalamic Projections: Especially inthe Caudal Medulla. Brain Res 277:201–222
13. Shigenaga Y, Nishimura M, Suemune S et al. (1989b) Somato-topic Organization of Tooth Pulp Primary Afferent Neurons inthe Cat. Brain Res 477:66–89
14. Shigenaga Y, Okamoto T, Nishimori T et al. (1986b) Oral andFacial Representation in the Trigeminal Principal and RostralSpinal Nuclei of the Cat. J Comp Neurol 244:1–18
15. Shigenaga Y, Sera M, Nishimori T et al. (1988) The Central Pro-jection of Masticatory Afferent Fibers to the Trigeminal SensoryNuclear Complex and Upper Cervical Spinal Cord. J Comp Neu-rol 268:489–507
16. Shigenaga Y, Suemune S, Nishimura T et al. (1986c) TopographicRepresentation of Lower and Upper Teeth Within the Trigemi-nal Sensory Nuclei od Adult Cat as Demonstrated by the Trans-ganglionic Transport of Horseradish Peroxidase. J Comp Neurol251:299–316
17. Tsuru K, Otani K, Kajiyama K et al. (1989) Central Terminationsof Periodontal Mechanoreceptive and Tooth Pulp Afferents inthe Trigeminal Principal and Oral Nuclei of the Cat. Brain Res485:29–61

T
Trigeminal Brainstem Nuclear Complex, Immunohistochemistry and Neurochemistry 2541
18. Westrum LE, Canfield RC, O’Conner TA (1981) Each CanineTooth Projects to all Brain Stem Trigeminal Nuclei in Cat. ExpNeurol 74:787–799
19. Yoshida A, Dostrovsky JO, Sessle BJ et al. (1991) TrigeminalProjections to the Nucleus Submedius of the Thalamus in theRat. J Comp Neurol 307:609–625
20. Yoshida A, Hiraga T, Moritani Met al. (1998) Morphologic Char-acteristics of Physiologically Defined Neurons in the Cat Trigem-inal Nucleus Principalis. J Comp Neurol 401:308–328
21. Yoshida A, Yasuda K, Dostrovsky JO et al. (1994) Two MajorTypes of Premotoneurons in the Feline Trigeminal Nucleus Oralisas Demonstrated by Intracellular Staining with Horseradish Per-oxidase. J Comp Neurol 347:495–514
Trigeminal Brainstem Nuclear Complex,Immunohistochemistry andNeurochemistry
DAVID A. BEREITER
Department of Surgery, Brown Medical School,Providence, RI, USA
Definition
The trigeminal brainstem nuclear complex (TBNC) iscomprised of the principal or main sensory nucleus andspinal trigeminal nucleus.Thespinaltrigeminalnucleusisfurthersubdivided,fromrostral to caudal, into subnucleus oralis (Vo), subnucleusinterpolaris (Vi) and subnucleus caudalis (Vc), which is
Trigeminal Brainstem Nuclear Complex, Immunohistochemistry andNeurochemistry, Figure 1 Trigeminal brainstem nuclear complex of therat. Abbreviations: I-II, laminae I-II; III-V, laminae III-V; m, trigeminal motorn.; nts, n. tractus solitarii; Pa5, paratrigeminal islands; Vc, subnucleuscaudalis; Vi, subnucleus interpolaris; Vi/Vc, interpolaris/caudalis transitionregion; Vo, subnucleus oralis; Vp, principal sensory nucleus; Vtr, spinaltrigeminal tract. Numbers below each outline indicate approximate distancein mm caudal to bregma, a skull surface landmark.
often referred to as the medullary dorsal horn since it dis-plays several features, such as a laminated organization,similar to the spinal dorsal horn.
Characteristics
The role of the different subnuclei of the TBNC in me-diating the various aspects of pain remains controver-sial, since injurious stimuli can excite neurons through-out the TBNC (Sessle 2000), and orofacial tissues arerepresented somatotopically at multiple, but intercon-nected, levels of the TBNC (Bereiter et al. 2000). Theneurochemical organization of the TBNC has shed ad-ditional light on possible contributions of different por-tions of the TBNC to pain processing.Immunohistochemical approaches have identified twobroad classes of small-diameter myelinated or unmyeli-nated nerve fibers: i) fibers stained positive for the neu-ropeptides substance P, calcitonin gene-related peptideand � neurotrophins and ii) fibers stained positive forthe cell-surface marker, � isolectin B4, and negative forneuropeptides. As summarized in Table 1, both classesof small-diameter fibers terminate densely in laminae I-II of Vc and the paratrigeminal islands and weaker, butsignificant, labeling in deep laminae of Vc and dorso-medial portions of rostral regions of the TBNC (Sugi-moto et al. 1997a; Sugimoto et al. 1997b). Immunoreac-tivity for NK1, the substance P receptor, and TrkA, thehighaffinity-receptor fornervegrowthfactor,areconsis-tent generally with the distribution of substance P- andnerve growth factor-positive fibers, respectively, withinthe TBNC (Krol et al. 2001; Nakaya et al. 1994).Markers associated with opioid analgesia such the� endogenous opioid peptide, endomorphin 2 (Martin-Schild et al. 1999), and the mu opioid receptor subtype,MOR1 (Ding et al. 1996), display moderate to denseimmunoreactivity in laminae I-II of Vc, paratrigeminalislands and at the Vi/Vc transition region, while deeperlaminae of Vc and more rostral regions of the TBNCdisplay weak or no staining.Physiological studies revealthat cornea-responsive neurons at the Vi/Vc transitionregion, i.e., the most rostral pole of Vc, are often en-hanced by increasing doses of morphine, suggesting arole for this region in recruitment of endogenous paincontrols (Hirata et al. 2000).Receptors for monoamine transmitters (catecholaminesand serotonin) display diffuse staining throughout mostrostral regions of the TBNC; however, the superfi-cial laminae of Vc display moderate to dense levelsof immunoreactivity (Day et al. 1997; Wright et al.1995). Staining for choline acetyltransferase (ChAT),the biosynthesis enzyme for acetylcholine, is weak inmost rostral regions of TBNC, while the superficiallaminae of Vc display moderate levels, consistent withthe distribution of nicotinic receptors (Wada et al. 1989).Neurons that contain the inhibitory amino acid trans-mitter, GABA, are found in all regions of the TBNC,with the highest density in laminae I-II of Vc (Ginestal

2542 Trigeminal Brainstem Nuclear Complex, Immunohistochemistry and Neurochemistry
Trigeminal Brainstem Nuclear Complex, Immunohistochemistry and Neurochemistry, Table 1 Summary of the distribution of immunohistochemicalmarkers associated with nociceptive processing in different portions of the trigeminal brainstem nuclear complex
Vp Vo Vi Vi/Vc Pa5 Vc
dm vl dm vl I-II III-V
IB4 + - + - + - ++ +++ -
SP + - + - + ++ ++ +++ +
CGRP ++ - ++ - + ++ ++ +++ +
TrkA + - + - + + + +++ -
ChAT + - + - + + - ++ +
NR1 ++ ++ ++ ++ ++ ++ +++ +++ +
GABA ++ + ++ + + ++ + +++ ++
AR α/β + - + - + + + ++ +
5HT1/2 + - + - + + + ++ +
Endo2 + - + - + + ++ +++ -
MOR1 - - - - - + ++ +++ -
EP3 - - - - - - + +++ -
P2X2 + + + + + + ++ ++ +
ER α - - - - - - - +++ -
Symbols and abbreviations:- = very weak or no staining; +,++,+++ = weak, moderate and dense staining; AR α/β, adrenergic receptor subtypes;CGRP, calcitonin gene-related peptide; ChAT, choline acetyltransferase; dm, dorsomedial; Endo2, endomorphin 2; EP3, prostaglandin receptor; ER α,estrogen receptor alpha subtype; GABA, gamma aminobutyric acid; 5HT1/2, serotonin receptor subtypes; IB4, isolectin B4; MOR1, mu opioid receptor;NR1, NMDA receptor subunit; P2X2, ATP receptor; Pa5, paratrigeminal islands; SP, substance P; TrkA, tyrosine kinase A receptor subtype; Vc, subnucleuscaudalis; Vi, subnucleus interpolaris; Vi/Vc, interpolaris/caudalis transition region; Vo, subnucleus oralis; Vp, principal sensory nucleus; vl, ventrolateral
and Matute 1993) in agreement with the distribution ofsubunits for the GABAA receptor (Fritschy and Mohler1995).Immunoreactivity for EP3, a subtype of the prostaglan-din receptor, and well associated with inflammatorypain, occurs only in laminae I-II of Vc and the para-trigeminal islands (Nakamura et al. 2000). By contrast,the purinergic receptor, P2X2, an ATP-gated ion chan-nel, (Kanjhan et al. 1999) and NR1, a subunit of theNMDA receptor (Petralia et al. 1994), receptors that areupregulated during inflammation and associated with� neuroplasticity, are widely distributed throughoutthe TBNC, although they have a higher density in Vc.Several orofacial pain conditions (e.g., migraineheadache, temporomandibular disorders) display asignificant sex-related prevalence. The basis for thesesex differences is not certain. However, the distributionof ERα, a major subtype of estrogen receptor, occursmainly in lamina II of Vc and not in more rostral regionsof the TBNC (Bereiter et al. 2005).The neurochemical organization of the TBNC is con-sistent with the notion that, laminae I-II of Vc and theparatrigeminal islands receive direct input from small-diameter nerve fibers, and play an essential role in theprocessing and modulation of trigeminal pain. Althoughrostral regions of the TNBC receive only sparse direct
input from small-diameter nerve fibers, the appearanceof selected markers for nociceptors and neuroplasticityin the dorsomedial portions of the principal sensory nu-cleus and Vo, suggest that these regions also contributeto the integration of specific forms of trigeminal pain.
References1. Bereiter DA, Cioffi IL, Bereiter DF (2005) Oestrogen receptor-
immunoreactive neurons in the trigeminal sensory system of maleand cycling female rats. Arch Oral Biol 50:971–9
2. Bereiter DA, Hirata H, Hu JW (2000) Trigeminal SubnucleusCaudalis: Beyond Homologies with the Spinal Dorsal Horn. Pain88:221–224
3. Day HE, Campeau S, Watson SJ, Akil H (1997) Distribution ofα1a, α1b, and α1d-Adrenergic Receptor mRNA in the Rat Brainand Spinal Cord. J Chem Neuroanat 13:115–139
4. Ding Y-Q, Kaneko T, Nomura S, Mizuno N (1996) Immunohis-tochemical Localization of μ-opioid Receptors in the CentralNervous System of the Rat. J Comp Neurol 367:375–402
5. Fritschy J-M, Mohler H (1995) GABAA-Receptor Heterogeneityin the Adult Rat Brain: Differential Regional and Cellular Dis-tribution of Seven Major Subunits. J Comp Neurol 359:154–194
6. Ginestal E, Matute C (1993) Gamma-Aminobutyric Acid-Immunoreactive Neurons in the Rat Trigeminal Nuclei. Histo-chemistry 99:49–55
7. Hirata H, Takeshita S, Hu JW, Bereiter DA (2000) Cornea–Responsive Medullary Dorsal Horn Neurons: Modulationby Local Opioid Agonists and Projections to Thalamus andBrainstem. J Neurophysiol 84:1050–1061
8. Kanjhan R, Housley GD, Burton LD, Christie DL, KippenbergerA, Thorne PR, L.Luo, Ryan AF (1999) Distribution of the P2X2

T
Trigeminal Brainstem Nuclear Complex, Physiology 2543
Receptor Subunit of the ATP-gated Ion Channels in the Rat Cen-tral Nervous System. J Comp Neurol 407:11–32
9. Krol KM, Stein EJ, Elliot J, Kawaja MD (2001) TrkA-ExpressingTrigeminal Sensory Neurons Display both Neurochemical andStructural Plasticity Despite a Loss of p75 NTR Function: Re-sponses to Normal and Elevated Levels of Nerve Growth Factor.Eur J Neurosci 13:35–47
10. Martin-Schild S, Gerall AA, Kastin AJ, Zadina JE (1999) Dif-
ferential Distribution of Endomorphin 1-and Endomorphin2-likeImmunoreactivies in the CNS of the Rodent. J Comp Neurol405:450–471
11. Nakamura K, Kaneko T, Yamashita Y, Hasegawa H, KatohH, Negishi M (2000) Immunohistochemical Localization ofProstaglandin EP3 Receptor in the Rat Nervous System. J CompNeurol 421:543–569
12. Nakaya Y, Kaneko T, Shigemoto R, Nakanishi S, Mizuno N(1994) Immunohistochemical Localization of Substance P Re-ceptor in the Central Nervous System of the Adult Rat. J CompNeurol 347:249–274
13. Petralia RS, Yokotani N, Wenthold RJ (1994) Light and ElectronMicroscope Distribution of the NMDA Receptor Subunit NM-DAR1 in the Rat Nervous System using a Selective Anti-PeptideAntibody. J Neurosci 14:667–696
14. Sessle BJ (2000) Acute and Chronic Craniofacial Pain: BrainstemMechanisms of Nociceptive Transmission and Neuroplasticity,and their Clinical Correlates. Crit Rev Oral Biol Med 11:57–91
15. Sugimoto T, Fujiyoshi Y, He Y-F, Xiao C, Ichikawa H (1997a)Trigeminal Primary Projection to the Rat Brain Stem Sen-sory Trigeminal Nuclear Complex and Surrounding StructuresRevealed by Anterograde Transport of Cholera Toxin BSubunit-Conjugated and Bandeiraea Simplicifolia Isolectin B4-Conjugated Horseradish Peroxidase. Neurosci Res 28:361–371
16. Sugimoto T, Fujiyoshi Y, Xiao C, He Y-F, Ichikawa H (1997b)Central Projection of Calcitonin Gene-Related Peptide (CGRP)-and Substance P (SP)-Immunoreactive Trigeminal Primary Neu-rons in the Rat. J Comp Neurol 378:425–442
17. Wada E, Wada K, Boulter J, Deneris E, Heinemann S, PatrickJ, Swanson LW (1989) Distribution of alpha2, alpha3, aplha4,and beta2 Neuronal Nicotinic Receptor Subunit mRNAs in theCentral Nervous System: A Hybridization Histochemical Studyin the Rat. J Comp Neurol 284:314–335
18. Wright DE, Seroogy KB, Lundgren KH, Davis BM, JennesL (1995) Comparative Localization of Serotonin 1A, 1C and2 Receptor Subtype mRNAs in Rat Brain. J Comp Neurol351:357–373
Trigeminal Brainstem Nuclear Complex,Physiology
JAMES W. HU1, ALAIN WODA2
1Faculty of Dentistry, University of Toronto, Toronto,ON, Canada2Faculté de Chirurgie Dentaire, University Clermont-Ferrand, Clermont-Ferrand, [email protected], [email protected]
Definition
Trigeminal (V) Brainstem Nuclear Complex (VBNC,Fig. 1) is the site of the first synapse of most V sen-sory primary afferents and of considerable sensoryprocessing and modulation. Its anatomy and physiol-ogy, although generally similar to the spinal sensorysystem, display several differences. These differences,and the existence of specialized organs and tissues, are
important features that have prompted the study of Vnociception, and may be manifested clinically in theunique trigeminal pain conditions, such as toothache,migraine, V neuralgia, temporomandibular disordersand other idiopathic orofacial pains.
Characteristics
Trigeminal Brainstem Nuclear Complex (VBNC) Organization
Most V primary afferents have their cell bodies in theV ganglion and project centrally to the VBNC, wherethey ascend to the main (or principal, Vp) sensory nu-cleus and/or descend to the spinal tract nucleus, whichincludes subnuclei oralis (Vo), interpolaris (Vi) and cau-dalis (Vc); see essay by Shigenaga and Yoshida (Fig. 1).The most caudal, Vc, is a laminated structure that ex-tends caudally into the dorsal horn of the cervical spinalcord with which it is homologous. For this reason, it isalso called the medullary dorsal horn (Gobel et al. 1981).Almost all nociceptive � C Fiber afferents from the Vnerve have their central terminals distributed in the Vcand Vi/Vc transition zone. Terminals of the A-δ fiberafferents are also found in Vc; however some, mostlyfrom the oral and perioral area, are found rostral to Vc;in particular, A-δ fibers from the dental pulp have beendescribed to be in the rostral divisions, Vp, Vo, Vi, (seeessay by Shigenaga and Yoshida). The somatotopic or-ganization of VBNC is also described in the essay byShigenaga and Yoshida.
General Features of Nociceptive Processing in SubnucleusCaudalis
In 1938, Sjöqvist reported that � trigeminal tractotomy,which interrupts the inputs to the Vc by severing theV spinal tract at the upper level of Vc, was an efficienttreatment for � trigeminal neuralgia. The effect ofVc deafferentation by tractotomy has repeatedly beenconfirmed by neurosurgeons and animal researchers,and it has therefore long been considered that Vc isthe crucial brainstem center for orofacial nociceptiveprocessing. However, this procedure has little effecton tactile sensation and intraoral pain. These resultsindicate the involvement of the rostral nuclei, such asthe Vo, in toothache or other pain sensations from theoral area (Young 1982). Thus neurons in V0, as well asVc, are involved in ascending pathways contributing toorofacial pain (also see below). In the case of autonomicresponses to noxious stimuli, such as cardiovascularand secretory reflexes (e.g. lacrimation and salivation),the trigeminal brainstem regions that mediate theseevoked responses include the Vi/Vc transition zone andmedial regions of the VBNC that border the reticularformation.The fact that Vc is critical for V nociception is supportedby 7 pieces of evidence:
1. Vc receives A-δ and C nociceptive afferents that ter-minate in laminae I and II

2544 Trigeminal Brainstem Nuclear Complex, Physiology
Trigeminal BrainstemNuclear Complex, Physiology,Figure 1 Schema of nociceptivesomatosensory organisation of theorofacial area. The trigeminal nerveis made up of three divisions,ophthalmic (V1), maxillary (V2) andmandibular (V3), which supplya wide variety of tissues; notethat V2 and V3 as well as V1also supply part of the meninges.The primary afferent neuronsmediating nociceptive messages(A-delta and C) project to thespinal tract nucleus. Ascendingpathways arise from spinal tractnucleus to reach suprasegmentallevels such as the thalamus andcerebral cortex. There are alsoimportant pathways descendingfrom the suprasegmentalcenters to modulate theafferent messages, such as thecerebral cortex. Modified withpermission from Dallel et al. 2003Medecine/sciences 19: 567–574.
2. Vc is a laminated structure with a substantia gelati-nosa (lamina II) region, which is known to be asso-ciated with pain processing and a high concentrationof neuropeptide markers for nociception
3. Vc contains nociceptive neurons, either � wide-dynamic-range (WDR) or nociceptive-specific (NS).These nociceptive neurons predominate in the lam-inae I, V and VI, and display positive stimulus-response functions to various afferent inputs, andcan therefore code the intensity of different types ofnoxious stimuli, including those arising from spe-cialized orofacial structures (Price et al. 1976; Sessle2000). In addition, these Vc nociciceptive neuronsalso receive a wide range of inputs (see Table 1)
4. Some Vc nociceptive neurons send their axons intoascending nociceptive pathways that reach the thala-mus (Price et al. 1976)
5. Some Vc neurons serve as reflex interneurons in re-flex responses to noxiousorofacial stimuli (seeSessle2000)
6. Vc nociceptive neurons are subject to several affer-ent and descending antinociceptive modulatory in-fluences, as well as pharmacological antinociceptivemodulations (Sessle 2000; Ren and Dubner 2002)
7. In certain conditions (e.g. inflammation or nerveinjury), these nociceptive neurons manifest someneuroplastic changes, such as increase in sponta-neous activity, reduction of activation threshold,increase response magnitude to noxious stimuli andreceptive field expansion (Hu et al. 1992). Inductionof these neuroplastic changes is an NMDA receptor-dependent process (Chiang et al. 1998) (Fig. 2).These neuroplastic changes in the nociceptive neu-
Trigeminal Brainstem Nuclear Complex, Physiology, Table 1 Conv-ergence of Nociceptive Afferent Inputs to Nociceptive Neurons in TrigeminalSubnucleus Caudalis of Cats
Facial Skin/Oral Mucosa 100%
Tooth pulp 66%
Muscle (jaw or tongue) 55%
Temporomandibular joint (TMJ) 35%
Laryngeal mucosa (Viscera) 55%
Upper cervical nerves 50%
Based upon a sample of approximately 100 WDR and NS neurons withcutaneous or intraoral mechanoreceptive fields; from Sessle et al. 1986
rons correlate well with the clinical observation ofallodynia and hyperalgesia associated with inflam-matory (Chiang et al. 1998) or nerve injury-inducedneuropathic (Iwata et al. 2001) pain conditions.
Special Features of Trigeminal Brainstem Nociceptive Pro-cessing
1. Nociceptive processing in VBNC is not restricted toVc: Nociceptive neurons activated from the orofacialarea are also observed rostrally in Vo and Vi (Dallel etal. 1990; Sessle 2000). Vo convergent neurons share allthe properties of other neurons of the deep spinal dorsalhorn, or of deep layers of Vc, (Dallel et al. 1990; Hu et al.1992) including plasticity (Sessle 2000). These neuronsmay play a role in reflex responses to noxious orofacialstimulation (see Shigenaga and Yoshida essay), butresults of V tractotomy (see above) indicate that oral

T
Trigeminal Brainstem Nuclear Complex, Physiology 2545
Trigeminal Brainstem Nuclear Complex, Physiology, Figure 2 Rat caudalis nociceptive-specific (NS) neuron showing neuroplastic changes in neuronalproperties after small-fiber excitant, and inflammatory irritant mustard oil (MO) application to the maxillary right molar pulp. In (a), histologically retrievedrecording site and the arrow indicates the neuronal recording site in caudalis and in (b), mechanoreceptive field (RF) sizes before (0 min.) and 10 min.after MO application (i.e. solid area represents pinch RF and ‘Red’ area, with a thin red arrow, represents a newly defined tactile RF that also respondedto pinch stimulus after MO application). In (b), blackened area indicates the RF location and size, which only noxious pinch stimulation activated, beforeMO application. The RF expansion 10 minutes after MO application and a small stripped area that appeared only after the application of MO, indicatedby a thin arrow, that a brush (low-threshold mechanical) stimulus can also activate this NS neuron that previously was unable to respond to this brushstimulus. In (c), MO-induced a brief burst of discharges followed by higher firing rate than the baseline level (0 min). This response represents an inputfrom the maxillary tooth pulp. In (d), neuronal responses to mechanical and thermal stimuli applied to the cutaneous RF. Top trace: marker of brush (Br),pressure (Pr), pinch (Pi) and radiant heat (RH). Middle trace: neuronal responses in control conditions (i.e. Pre-MO before MO application). Bottom trace:neuronal responses to same stimuli 20 min after MO application (i.e. Post-MO after MO application). In (e), neuronal responses to graded mechanicalstimuli (50, 100, and 200 g). Each stimulus lasts for 3 s, and the y -axis scale (omitted) is the same for middle and bottom traces in a and b. Binwidth is1 s. Note that after MO application this NS neuron became responsive to light brushing (thin arrow) and radiant heating (thick arrow) of the cutaneousRF, and strongly responsive to graded pinch stimuli. Modified with permission from Chiang et al. (1998).
and perioral pain sensation also depends on Vo. Anotherindication of a sensory role for Vo as well as Vc is thepresence of a similar density of anatomical projectionsfrom these two subnuclei to higher levels of the brain.In addition, nociceptive thalamic neurons can still berecorded after stimulation of oral and perioral areas, inspite of the deafferentation of Vc by trigeminal tracto-tomy (Raboisson et al. 1989). In Vo, the C-fiber primaryafferent endings are scarce and a substantia gelatinosais lacking. However, C-fiber evoked activities recordedin Vo relay in the substantia gelatinosa of Vc (Dallel etal. 1998). The influence on the properties of Vo neuronsof the substantia gelatinosa of Vc constitutes a uniquefeature, which can be used as an experimental model(Fig. 3), since it offers the possibility of recording fromdeep layer nociceptive neurons, i.e. in Vo, while in-jecting neurochemicals in the Vc substantia gelatinosa.Indeed, it has been shown that plastic changes observed
in nociceptive neurons of Vo can be modulated, eitherfacilitated or inhibited, by neurochemicals injected intothe Vc substantia gelatinosa (Dallel et al. 1998; Wodaet al. 2001; Hu et al. 2002).2. Some specialized orofacial tissues receive innerva-tion patterns unique to the trigeminal system. Unlike theskin, specialized V structures, such as the tooth pulp,meninges and cornea are innervated mainly by A-δ andC nociceptive afferents, since very few or no A-β affer-ents can be found. Single unit-recording studies haveshown that the VBNC nociceptive neurons activatedfrom these specialized tissues often receive convergentinputs from a wide range of structures including facialskin. The distribution of V neurons that encode nocicep-tive information from these specialized orofacial tissuesdisplay a complex pattern, which is characterized bytheir occurrence at different rostrocaudal levels of theVBNC.

2546 Trigeminal Ganglion
Trigeminal Brainstem Nuclear Complex, Physiology, Figure 3 Schem-atic dorsal view of the spinal tract nucleus of VBNC that illustrates the sub-nucleus oralis-subnucleus caudalis experimental model. The subnucleusoralis is activated only indirectly by primary afferent C-fibers. A relay in thesubstantia gelatinosa in subnucleus caudalis is mandatory, since there is nodirect C fiber projection to Vo. This allows for recordings from nociceptiveneurons while injecting chemicals into the substantia gelatinosa.
3. The VBNC also offers the possibility of extendingthe knowledge base relevant to spinal neuronal proper-ties from anaesthetized to awake conditions. Taking ad-vantage of the unique location of the medullary dorsalhorn (Vc), Dubner and his colleagues recorded Vc sin-gle neurons in conscious monkeys while performing be-havioral tasks relevant to nociception (Hoffman et al.1981). While the properties of WDR and NS neurons inVc observed in anaesthetized animals during recordingswere confirmed in the conscious monkeys, these experi-ments also showed that many of these Vc neurons couldbe modulated through attentional and motivational fac-tors.
References1. Bereiter DA, Hirata H, Hu JW (2000) Trigeminal Subnucleus
Caudalis: Beyond Homologies with the Spinal Dorsal Horn. Pain88:221–224
2. Chiang CY, Park SJ, Kwan CL, Hu JW, Sessle BJ (1998) NMDAReceptor Mechanisms Contribute to the Trigeminal NociceptiveNeuronal Plasticity Induced by Mustard Oil Application to theRat Molar Tooth Pulp. J Neurophysiol 80.2621–2631
3. DallelD, DualeC, Molat JL (1998)Morphine Administered in theSubstantia Gelatinosa of the Spinal Trigeminal Nucleus CaudalisInhibits Nociceptive Activities in the Spinal Trigeminal NucleusOralis. J Neurosci 18:3529–3536
4. Dallel R, Raboisson P, Woda A, Sessle BJ (1990). Properties ofNociceptive and Non-Nociceptive Neurons in Trigeminal Sub-nucleus Oralis of the Rat. Brain Res 521:95–106
5. Gobel S, Hockfield S, Ruda MA (1981) Anatomical Similari-ties between Medullary and Spinal Dorsal Horns. In: KawamuraY, Dubner R (eds) Oral-Facial Sensory and Motor Functions.Quintessence Publishing Co, Tokyo, Berlin, pp 211–223
6. Hoffman DS, Dubner R, Hayes RL, Medlin TP (1981) NeuronalActivity in Medullary Dorsal Horn of Awake Monkeys Trainedin a Thermal Discrimination Task. I. Responses to Innocuousand Noxious Thermal Stimuli. J Neurophysiol 46:409–427
7. Hu B, Chiang CY, Hu JW, Dostrovsky JO, Sessle BJ (2002) P2XReceptors in Trigeminal Subnucleus Caudalis Modulate CentralSensitization in Trigeminal Subnucleus Oralis. J Neurophysiol88:1614–1624
8. Hu JW, Sessle BJ, Raboisson P, Dallel R, Woda A (1992). Stimu-lation of Craniofacial Muscle Afferents Induces Prolonged Facil-itatory Effects in Trigeminal Nociceptive Brain-Stem Neurones.Pain 48:53–60
9. Iwata K, Imai T, Tsuboi Y, Tashiro A, Ogawa A, Morimoto T,Masuda Y, Tachibana Y, Hu J (2001) Alteration of Medullary Dor-sal Horn Neuronal Activity Following Inferior Alveolar NerveTransection in Rats. J Neurophysiol 86:2868–2877
10. Price DD, Dubner R, Hu JW (1976) Trigeminothalamic Neu-rons in Nucleus Caudalis Responsive to Tactile, Thermal, andNociceptive Stimulation of Monkey’s Face. J Neurophysiol39:936–953
11. Raboisson P, Dallel R, Woda A (1989) Responses of Neurones inthe Ventrobasal Complex of the Thalamus to Orofacial NoxiousStimulation after Large Trigeminal Tractotomy. Exp Brain Res77:569–576
12. Ren K, Dubner R (2002) Descending Modulation in PersistentPain: An Update. Pain 100:1–6
13. Sessle BJ (2000) Acute and Chronic Craniofacial Pain: BrainstemMechanisms of Nociceptive Transmission and Neuroplasticity,and their Clinical Correlates. Crit Rev Oral Biol Med 11:57–91
14. Woda A, Molat JL, Luccarini P (2001) Low Doses of N-methyl-D-aspartate Antagonists in Superficial Laminae of MedullaOblongata Facilitate Wind-Up of Convergent Neurones. Neu-roscience 107:317–327
15. Young RF (1982) Effect of Trigeminal Tractotomy on DentalSensation in Humans. J Neurosurg 56:812–818
Trigeminal Ganglion
Definition
A nerve ganglion where the cell bodies of the afferentneurones innervating the orofacial area are located.� Nociceptors in the Dental Pulp
Trigeminal Lemniscus
Definition
Trigeminal cutaneous projection to the thalamus, in con-trast to the spinal lemniscus.� Parafascicular Nucleus, Pain Modulation
Trigeminal Motor Nucleus
Definition
Also called masticatory nucleus, it is located in the dor-sal mid-pons, close to the trigeminal principal sensorynucleus, and contains motoneurons for both jaw-closingand jaw-opening muscles.� Jaw-Muscle Silent Periods (Exteroceptive Suppres-
sion)

T
Trigeminal Neuralgia, Diagnosis and Treatment 2547
Trigeminal NerveDefinition
The trigeminal nerve is the largest cranial nerve and in-nervatesmuchof thecutaneouscraniofacial regions.Thetrigeminal nerve also innervates the muscles of mastica-tion. The three major divisions of the trigeminal nerveare ophthalmic, maxillary and mandibular nerves. Anextremely painful condition, tic douloureux, occurs inthe territory of the trigeminal nerve and likely involvesprimary afferent fibers of the trigeminal nerve.� Amygdala, Pain Processing and Behavior in Animals� Nociception in Nose and Oral Mucosa� Trigeminothalamic Tract Projections
Trigeminal NeuralgiaSynonyms
Tic douloureux
Definition
Trigeminal neuralgia is an idiopathic, episodic severepain condition of the orofacial region often described aselectric-shock-like sensations that can be triggered byinnocuous stimuli, associated with injury or dysfunctionof the fifth cranial nerve (trigeminal nerve) or its gan-glion and felt in the distribution of this nerve. It is con-sidered as a prototype of neuropathic pain. Trigeminalneuralgia (tic douloureux) may have no apparent causeor be associated with neurovascular conflict between thetrigeminal root and an anomalous vascular loop (classi-cal trigeminal neuralgia), or be secondary to benign tu-mors of the cerebellopontine angle or multiple sclerosis.� Central Pain in Multiple Sclerosis� Demyelination� Jaw-Muscle Silent Periods (Exteroceptive Suppres-
sion)� Neuralgia, Assessment� Neuralgia, Diagnosis� Pain Paroxysms� Paroxysmal Hemicrania� Primary Stabbing Headache� Tic and Cranial Neuralgias� Trigeminal Brainstem Nuclear Complex, Physiology� Trigeminal Neuralgia, Etiology, Pathogenesis and
Management
Trigeminal Neuralgia, Aims of SurgicalManagement
Definition
Surgical management of trigeminal neuralgia can pro-vide complete pain relief for periods ranging from a yearto 10 years.
� Trigeminal Neuralgia, Etiology, Pathogenesis andManagement
Trigeminal Neuralgia, Diagnosis andTreatment
DONALD R. NIXDORF
Department of Diagnostic and Biological Sciences,School of Dentistry, University of Minnesota,Minneapolis, MN, [email protected]
Synonyms
Tic douloureux
Definition
Trigeminal neuralgia is an excruciating pain felt in thedistribution of the trigeminal or fifth cranial nerve and isclassically described as a brief sharp shooting pain (Kittet al. 2000). Trigeminal neuralgia has been defined bythe International Headache Society’s classification sub-committee as (IHS 2004):
A) � Paroxysmal attacks of pain lasting from a fractionof a second to 2 minutes, affecting one or more divi-sions of the trigeminal nerve and fulfilling criteria Band C.
B) Pain has at least one of the following characteristics:
1. intense, sharp, superficial or stabbing2. precipitated from trigger areas or by trigger factors
C) Attacks are stereotyped in the individual patientD) There is no clinically evident neurological deficitE) Not attributed to another disorder
Although this definition has never been tested for reli-ability and proven to be valid, either it, or a derivation,are widely accepted and utilized extensively for both re-search and patient care purposes.Note: the terms classical trigeminal neuralgia and idio-pathic trigeminal neuralgia are used when a secondarycause for this pain is not readily identifiable. Symp-tomatic trigeminal neuralgia is used when a secondarycause for this pain has been identified, such as ananeurysm, tumor or stroke.
Characteristics
Trigeminal neuralgia occurs in approximately 4 per100,000 people and has an onset later in life, with themedian age of diagnosis being 67 years old. It occursabout twice as often in women as in men (Katusic et al.1990), but this may be from sampling error, since goodprevalence data do not exist (Zakrzewska and Hamlyn1999). This severe pain is described by patients as hav-ing an electric or lightening bolt quality and being briefin duration, lasting less than 30 seconds. Sometimes

2548 Trigeminal Neuralgia, Diagnosis and Treatment
patients describe a dull, burning or throbbing pain thatoccurs between attacks of intense pain. Trigeminalneuralgia pain is most commonly felt in the maxillarydivision and to a lesser extent in the mandibulardivision.Pain is infrequently felt only in the ophthalmic divisionof the trigeminal nerve. It is almost always unilateral,especially on initial presentation and does not cross themidline. There may be predilection for right-sided pain(Zakrzewska and Hamlyn 1999), which would suggestan anatomical correlation. Paroxysms of pain may bespontaneous, occurring without a known reason ortriggered by non-painful stimuli such as light touch orwind on the face, eating and grooming. The triggeringarea may be the area where the pain is felt or different,but it is always on the same side as the pain and is oftenperioral. Local anesthetic applied to the triggering areawill abate the ability to trigger the pain. Once pain istriggered it is self-sustaining and frequent triggeringoften results in a decrease in pain intensity in the laterbouts of pain. Interestingly, even though this pain isone of the worst pains imaginable, it is not triggerableduring unconsciousness and rarely wakes people fromsleep. Also, trigeminal neuralgia is characterized ashaving periods of spontaneous remission, which maylast days, months or even years.
Diagnostic Process
The diagnosis of trigeminal neuralgia is based strictlyon clinical data, i.e. a history and physical examination,since there are no laboratory tests or imaging studiesthat can either confirm or refute its presence (Merskeyand Bogduk 1994; IHS 2004). Brain imaging is fre-quently ordered to ensure that trigeminal neuralgia isnot caused by an intracranial space-occupying lesionor � demyelinating process. Trigeminal neuralgia is20 times more common in people with multiple scle-rosis (Katusic et al. 1990), so when it occurs in youngpatients, the appropriate evaluative measures should beperformed.Patients sometimes present to their dentist first, becausethis pain is frequently felt in the jaws and teeth. Oftentheir description is of a continuous dull ache, similar totoothache. There is controversy about this pain becauseitmay actually beaprodrome to theclassically describedtrigeminal neuralgia, occurring days to years before themore recognizable symptoms occur. This prodromepain has been termed pre-trigeminal neuralgia and cor-responds to the initialpresentation featuresof trigeminalneuralgia. This concept is supported by patient reportingand by clinical experience that this prodrome pain re-sponds to pharmacotherapy like classic trigeminal neu-ralgia does (Fromm et al. 1990). The opposing opinionis that the continuous dull ache pain is really of odonto-genic origin. Dental interventions, like root canals andtooth extractions, employed to address pathology resultin peripheral nerve injury and this cumulative injuryleads, in part, to the formation of trigeminal neuralgia.
Etiology and Pathophysiology
The etiology of trigeminal neuralgia is not establishedand little is truly understood about the underlying painmechanisms, since a valid animal model does not exist.Trigeminal nerve root compression, at the entry zoneinto the � pons, has been observed in patients withtrigeminal neuralgia (Dandy 1934). The anatomicalarrangement of blood vessel impingement on the nerveroot has also been described in patients after surgicalexploration (Jannetta 1967), but it is not present in allpatients with trigeminal neuralgia and sometimes it isnoted in imaging of non-trigeminal neuralgia patients(Majoie et al. 1997). It has been hypothesized that thepresence of � ectopic action potentials arising fromperipheral nerves and the failure of central inhibitionwithin the trigeminal system occur simultaneously toproduce paroxysmal pain. This explains why certainmedications are beneficial and others are not in thetreatment of trigeminal neuralgia (Fromm et al. 1984).Currently, the phenomenon of ephaptic cross talk isused to explain how non-painful stimuli are translatedinto painful ones and crossed � afterdischarge has beencited as the mechanism implicated in further pain am-plification and prolongation. These two assumptionshave been collectively termed the ‘ignition hypothe-sis’, which defines trigeminal neuralgia as a peripheralnerve disorder (Devor et al. 2002). This hypothesis ex-plains why people with demyelinating disorders, suchas multiple sclerosis, frequently experience trigeminalneuralgia. It may also account for the clinical observa-tion that trigeminal neuralgia seems to start after recentdental interventions. The continuous dull burning back-ground pain sometimes felt by patients would best beexplained by the development of central sensitizationsecondary to a lack of inhibition, akin to deafferentationpain and other continuous neuropathic pains.
Treatment
Overview
Whenever possible, treatment should be directed atthe etiological factors or tailored to the mechanismsinvolved. Since these are largely unknown, currenttreatments have developed from empiric practice. Dataexist regarding pharmacological and surgical treatmentoptions, but little or nothing is published about psy-chological interventions. In my opinion, as with otherchronic pains, some trigeminal neuralgia patients maybenefit from a cognitive behavioral approach to care thatalso includes stress reduction and coping strategies. Thefollowing is a summation of the best available researchpublished.
Pharmacological Treatment Options
Pharmacotherapy, also known as medication or drugtherapy, is considered the first-line treatment for trigem-inal neuralgia (Kitt et al. 2000). Anticonvulsant medi-cations or drugs that have membrane-stabilizing prop-

T
Trigeminal Neuralgia, Diagnosis and Treatment 2549
erties, meaning that they reduce spontaneous nerve ac-tivity, are typically used for the treatment of trigeminalneuralgia.Thesemedicationshavebeenshowninanimalexperiments to suppress the response of mechanorecep-tors and augment presynaptic afferent inhibition withinthe maxillary nerve; this has been correlated with clin-ical drug efficacy (Fromm et al. 1981). Therefore, thegoal of pharmacotherapy is to augment inhibitory path-ways and reduce ectopic and � ephaptic neuronal firing.Randomized placebo controlled trials for the treatmentof trigeminal neuralgia are rare. The low prevalencein the population and the occurrence of spontaneoussymptom remission make it difficult to study.Carbamazepine is an older anticonvulsant medicationthat has been studied the most and clinically utilizedoften. Three randomized controlled studies have eval-uated carbamazepine for the treatment of trigeminalneuralgia and have found that it reduces pain severity,frequency and trigger sensitivity. Up to 75% of patientstaking carbamazepine find it effective for long-termpain reduction (Sindrup and Jensen 2002). Sodiumchannel blockade is the accepted mechanism of actionfor carbamazepine; this results in a reduction of nerveexcitation and axonal firing. Dosing is by titration tothe clinical effect of pain reduction, initiating at 200 mga day, increasing at a maximum rate of 200 mg per dayto a maximum dosage of 1200 mg per day in divideddoses. Most patients respond between 400–800 mg perday, which corresponds to serum levels between 20 and40 μmol/L (Sindrup and Jensen 2002). Side effects ofcarbamazepine include hepatic induction and toxicity,hematopoietic suppression, electrolyte imbalances,multiple drug interactions and cognitive impairment.These side effects can be serious; therefore, laboratorymonitoring to ensure early detection of potential prob-lems is required. Side effects are often very limiting forpatients and can cause them to discontinue use, evenwhen profound pain reduction is achieved.Phenytoin and valproic acid are two other older anticon-vulsant medications that have not been systematicallystudied. Both phenytoin and valproic acid block sodiumchannels, while valproic acid also increases gamma-aminobutyric acid (GABA) and enhances glutamatedecarboxylase. Case series information suggests thatdoses of 300 mg per day of phenytoin and 1200 mgper day of valproic acid are effective for pain reduction(Sindrup and Jensen 2002). Both drugs are becomingused less often because they are hampered by similarside effects and monitoring issues as carbamazepine.Also, clinical experience suggests that they do notproduce the same robust reduction in pain symptoms.Clonazepam, a GABAA agonist, and baclofen, aGABAB agonist, are labeled as muscle relaxants and arebeing used for the treatmentof trigeminalneuralgia.Onesmall, uncontrolled trial suggested that clonazepam, atdoses of 6–8 mg per day, may be helpful. In a smallcontrolled study of baclofen, as an add-on therapy
to carbamazepine, pain frequency was reduced withdosages of up to 80 mg per day (Sindrup and Jensen2002).Newer anticonvulsant medications have shown somepotential for successful future use based primarily ontheir lower side effect profile, as well as on some limitedresearch data. Lamotrigine has been studied in one smallcontrolled trial as an add-on medication. Its mechanismof action has not been totally elucidated, but it has beenreported to block sodium channels and inhibit presy-naptic glutamate release. At doses of 400 mg per dayit has been shown to reduce trigeminal neuralgia pain(Zakrzewska et al. 1997). Topiramate has been studiedin a very small controlled trial as an add-on medication.The exact mechanisms of action are also not known, butit is thought to block sodium channels, inhibit glutamatebinding and act as a GABAA agonist. At doses of upto 600 mg per day, it reduced trigeminal neuralgia painintensity, duration and frequency (Gilron et al. 2001).Gabapentin, which is thought to cause calcium channelblockade, has been reported to be helpful in case reportsin dosages of up to 2700 mg per day (Sist et al. 1997).Its side effect profile tends to be the most favorable ofall anticonvulsant medications but dizziness, fatigue,ataxia and weight gain can occur. Oxcarbazepine wasevaluated in a small, uncontrolled study of patients withrefractory trigeminal neuralgia; patients reported painreliefon dosesaround 1200mgperday (ZakrzewskaandPatsalos 2002). Oxcarbazepine is chemically relatedto carbamazepine and also blocks sodium channels. Itis reported to have far fewer side effects and does notrequire monitoring with laboratory testing, but dosedependent � hyponatremia has been reported.Theutility ofothermedications typically used in chronicpain, such as tricyclic antidepressants and opioid anal-gesics for the treatment of trigeminal neuralgia has notbeen systematically researched. Conceptually, mostclinicians agree that these medications have a role inthe management of trigeminal neuralgia when it is re-fractory to previous treatment or when it has an atypicalpresentation, such as concurrent continuous pain.
Surgical Treatment Options
Surgical treatment is usually reserved for patients whofail first line drug therapy due to uncontrollable pain orintolerable side effects. The target for surgery is the pe-ripheral nerve, with either nerve ablation or decompres-sion the goal. There are no randomized controlled tri-als that investigate the effects of surgical treatment op-tions; thus the advantages and disadvantages of the var-ious procedures are based on personal opinion (Kitt etal. 2000). Historically, peripheral � neurectomy was thepreferred procedure for trigeminal neuralgia, using ei-ther mechanical or chemical means. This procedure hasbeen replaced by other surgical options that result in re-duced sensory deficits and higher success. Dental treat-ments, which result in peripheral nerve ablation, usually

2550 Trigeminal Neuralgia, Diagnosis and Treatment
result in a short-term remission, but pain usually reoc-curs in 3 to 6 weeks.Gasserian or trigeminal ganglion � rhizotomy is aneuroablative procedure performed by inserting a nee-dle within the trigeminal cistern. Several neurolyticmethods have been developed and are routinely em-ployed, each of them having relative advantages anddisadvantages. They all provide immediate pain reliefin more than 90% of patients. Radiofrequency ther-mocoagulation can be directed to a specific division ofthe trigeminal nerve and therefore has the advantage ofproviding a limited and well-controlled lesion. Correctpositioning of the needle tip can be verified by electricalstimulation of the awakened patient prior to lesioning.Disadvantages of this technique include a potentiallyhigher risk for anesthesia dolorosa and corneal anes-thesia. Also, other techniques may be superior andmore practical when large areas of the trigeminal nerveare involved, since multiple lesions will be required.Injection of glycerol as a neurolytic agent is anotherpossible approach. Glycerol is thought to affect un-myelinated nerve fibers selectively, thereby sparing� proprioception, touch and motor functions of thetrigeminal nerve. Anatomic variations in nerve ablationare achieved by altering the patient’s head position andadjusting the volume of glycerol injected. The majordisadvantages are the relatively imprecise technique,with the potential of glycerol spreading into the brain,as well as the moderate risk for anesthesia dolorosa andcorneal anesthesia. Balloon compression is the newesttechnique and has the lowest reported complicationsof anesthesia dolorosa and corneal anesthesia (Tekkökand Brown 1996). This procedure has the advantage ofnot requiring patient participation, but it has the highestincidence of muscle weakness and bradycardia requir-ing pacing has been reported during balloon inflation.Overall, all three techniques provide comparable painrelief and differ mostly in their potential advantagesand disadvantages (Slavin and Burchiel 2002).Stereotactic radiosurgery or gamma knife is a procedureperformed using radiation to cause nerve injury. The tar-get of treatment is along the trigeminal nerve about 2 to4 mm after it exits the brainstem at the level of the pons.Advantages of this technique are that pain relief percent-agesarecomparable to rhizotomy andithasa lowrisk foranesthesia dolorosa and corneal anesthesia. Disadvan-tages include a potential for delayed onset of pain reliefand high rates of pain recurrence (Slavin and Burchiel2002).Microvascular decompression, also known by theacronym MVD, is an open surgical procedure of thebrain. The goal is the alleviation of existing vascularpressure on the trigeminal nerve root. Touted as beingpotentially curative, this procedure has initial successrates in pain elimination comparable to those of rhizo-tomy, but it has less pain recurrence and a much lowerchance of anesthesia dolorosa, corneal anesthesia and
muscle weakness. The potential disadvantage is a lessthan 1% mortality rate. Other possible complicationsinclude stroke, meningitis, cerebrospinal fluid leakand various ipsilateral cranial nerve deficits, such ashearing loss (Slavin and Burchiel 2002). As surgicalmicrovascular decompression techniques become lessinvasive, such as with endoscopic assistance, it is likelythat it will become more commonly practised and withfavorable research published, it may even become afirst-line treatment option.Due to the varied presentation of trigeminal neuralgiaand the reported relapse of positive treatment effects,long-termfollow-upandmanagement isessential inpro-viding the best possible patient care.
References1. Dandy WE (1934) Concerning the cause of trigeminal neuralgia.
Am J Surg 242. Devor M, Amir R, Rappaport ZH (2002) Pathophysiology of
Trigeminal Neuralgia: The Ignition Hypothesis. Clin J Pain18:4–13
3. Fromm GH, Chattha AS, Terrence CF et al. (1981) Role ofinhibitory mechanisms in trigeminal neuralgia. Neurology31:683–687
4. Fromm GH, Terrence CF, Maroon JC (1984) Trigeminal neural-gia: current concepts regarding etiology and pathogenesis. ArchNeurol 41:1204–1207
5. Fromm GH, Graff-Radford SB, Terrence CF et al. (1990) Pre-trigeminal neuralgia. Neurology 40:1493–1495
6. Gilron I, Booher SL, Rowan JS et al. (2001) Topiramate intrigeminal neuralgia: a randomized, placebo-controlled multiplecrossover pilot study. Clin Neuropharmacol 24:109–112
7. International Headache Society Classification of Headache Dis-orders (2004) Cranial neuralgia, central and primary facial pain,and other headache disorders. Cephalalgia Suppl 1:125–135
8. Jannetta PJ (1967) Arterial compression of the trigeminal nerveat the pons in patients with trigeminal neuralgia. J Neurosurg26:159–162
9. Katusic S, Beard CM, Bergstrath E et al. (1990) Incidence andclinical features of trigeminal neuralgia, Rochester, Minnesota,1945–1984. Ann Neurol 27:80–95
10. Kitt CA, Gruber K, Davis M et al. (2000) Trigeminal neuralgia:opportunities for research and treatment. Pain 85:3–7
11. Majoie CB, Hulsmans FJ, Verbeeten B Jr et al. (1997) Trigemi-nal neuralgia: comparison of two MR imaging techniques in thedemonstration of neurovascular contact. Radiology 204:455–460
12. Merskey H, Bogduk N (1994) Relatively localized syndromes ofthe head and neck. In: Merskey H, Bogduk N (eds) Classificationof chronic pain. IASP Press, Seattle, pp 59–92
13. Sindrup SH, Jensen TS (2002) Pharmacotherapy of trigeminalneuralgia. Clin J Pain 18:22–27
14. Sist T, Filadora V, Miner M et al. (1997) Gabapentin for idiopathictrigeminal neuralgia: report of two cases. Neurology 48:1467
15. Slavin KV, Burchiel KJ (2002) Surgical options for facial pain.In: Burchiel KJ (ed) Surgical management of pain. Thieme, NewYork, pp 849–864
16. Tekkök IH, Brown JA (1996) Trigeminal neuralgia. NeurosurgQuart 6:267–288
17. Zakrzewska JM, Hamlyn PJ (1999) Facial pain. In: Crombie IK,Croft PR, Linton SJ et al (eds) Epidemiology of pain. IASP Press,Seattle, pp 171–202
18. Zakrzewka JM, Patsalos PN (2002) Long-term cohort comparingmedical (oxcarbazepine) and surgical management of intractabletrigeminal neuralgia. Pain 95:259–266
19. Zakrzewka JM, Chaudhry Z, Nurmikko TJ et al. (1997) Lamot-rigine (lamictal) in refractory trigeminal neuralgia: results from adouble-blind placebo controlled crossover trial. Pain 73:223–230

T
Trigeminal Neuralgia, Etiology, Pathogenesis and Management 2551
Trigeminal Neuralgia, Diagnostic Method
Definition
Diagnosis of trigeminal neuralgia is made principally onhistory as there are no diagnostic tests to validate the di-agnosis.� Trigeminal Neuralgia, Etiology, Pathogenesis and
Management
Trigeminal Neuralgia, Etiology,Pathogenesis and Management
JOANNA M. ZAKRZEWSKA
Clinical Diagnostic and Oral Sciences, Institute ofDentistry, University of London, Barts and the LondonQueen Mary’s School of Medicine & Dentistry,London, [email protected]
Synonyms
Trigeminal neuralgia; tic douloureux; SecondaryTrigeminal Neuralgia; Symptomatic Trigeminal Neu-ralgia; Atypical Trigeminal Neuralgia
Definition
The International Association for the Study of Pain(IASP) defines trigeminal neuralgia as “A sudden, usu-ally unilateral, severe, brief, stabbing, recurrent painin the distribution of one or more branches of the fifthcranial nerve” (Merskey and Bogduk 1994).
Characteristics
Epidemiology
The incidence of � trigeminal neuralgia is 4.3/100,000(2.96 male, 3.47 female/100,000 based on data from theUS). The point prevalence is 0.1%. The peak incidenceis in the age group 60–69, and it is rare in patients un-der the age of 40. There is a strong link between multi-ple sclerosis and trigeminal neuralgia, and hypertensionmay also be a risk factor. There are little data on the natu-ral history and prognostic features (� trigeminal neural-gia, features), but data from the US suggests that it doesnotaffect survival, although attacksgetmoreseverewithtime (Zakrzewska and Hamlyn 1999).
Etiology and Pathogenesis
As there are no satisfactory animal models of trigeminalneuralgia, it still remains difficult to elucidate fully theaetiology and pathogenesis of trigeminal neuralgia. Theignition hypothesis by Devor et al. (2002) (� trigeminalneuralgia, ignition theory) isacceptedbymany,althoughdirect support of it from trigeminal electrophysiologicalstudies is very limited. According to this theory, chronic
irritation of the trigeminal nerve leads to focal demyeli-nation, which results in the generation of ectopic actionpotentials and impaired segmental inhibition. This leadsto hyper-excitability of the afferents that give rise to painparoxysms as a result of synchronised after-dischargeactivity. This theory is supported by clinical observa-tions thatpatientswith trigeminalneuralgia in themajor-ity of cases are found to have blood vessels compressingthe trigeminal nerve, either at the nerve root entry zoneor less commonly the brain stem. Electron microscopicexamination of nerve roots taken from patients withsuch compressions has revealed focal demyelination inthe region of the compression, with close apposition ofdemyelinated axons and an absence of intervening glialprocesses. A process of re-myelination does occur, andthis could be responsible for the spontaneous remissionof the neuralgia. The most effective drugs are anti-convulsants, and they probably work by suppressing ec-topic hyper-excitability in the nerve or central neurons.
Clinical History
The following list provides the diagnostic criteria assuggested by the International Headache Society (Anon1988) (� trigeminal neuralgia, diagnostic method):
1. Paroxysmal attacks of facial or frontal pain last a fewseconds to less than two minutes.
2. Pain has at least 4 of the following characteristics:
– Distributionalongoneormoredivisionsof trigem-inal nerve
– Sudden, intense, sharp, superficial, stabbing orburning in quality
– Pain intensity severe– Precipitation from trigger areas, or by certain daily
activities such as eating, talking, washing the faceor cleaning the teeth
– Between paroxysms entirely asymptomatic
3. No neurological deficit.4. Attacks are stereotyped in individual patients.5. Exclusion of other causes of facial pain.
It is essential to take a very careful history, as this is theonly reliable method of making the diagnosis. It is es-pecially important to elucidate the sharpness and parox-ysmal quality of this pain, which differentiates it frommost other facial pains. A particular feature of trigem-inal neuralgia is that it is usually precipitated from oneor more trigger areas (especially in the second or thirdtrigeminal divisions), upon tactile stimulation or dailyactivities such as eating, talking, washing or shaving theface or cleaning the teeth. Each bout of pain is very quick(seconds), but patients may get many of these in quicksuccession,andso thepainmayseemtobelastinglonger.The nerve eventually becomes refractory and there is aperiod when the patient is pain free. Classically, thereare also periods of complete pain remission which lastfor weeks or months. These pain remissions gradually

2552 Trigeminal Neuralgia, Etiology, Pathogenesis and Management
Trigeminal Neuralgia, Etiology, Pathogenesis and Management, Table 1 Medical management of trigeminal neuralgia, most drugs need to beescalated and withdrawn slowly
Drug Daily dosagerange
Outcome Numberneeded to treat NNT(95% CI)
Side effects, Number needed to harmNNH(95% CI)
Comments
Baclofen 50–80 mg NNT 1.4 (1–2.6) only10 patients, possiblyeffective
Ataxia, lethargy, fatigue, nausea Useful as add on therapy
Carbamazepine 300–1200 mg NNT 2.6 ( 2–4) ,effective
Ataxia, dizziness, diplopia, lethargy NNH 3.4(2.5–5.2) for side effects, NNH for withdrawal24 (13–110)
Reduced white cell count,hyponatraemia higher doses
Lamotrigine 200–400 mg NNT 2.1 ( 1.3–6.1) asadd on medication
dizziness, drowsiness, constipation, ataxia,diplopia, irritability
Rapid dose escalationincreases incidence ofrashes
Pimozide 4–12 mg NNT 2 (2–3) Extrapyramidal e.g. tremor, rigidity NNH 2.9(2–4)
Side effects too severe torecommend routine use
Proparcaine 2 drops of0.5% solution
Not effective Toxic keratopathy in long term use Short lasting even if givenrepeatedly
Tizanidine 6–18 mg May be effective Nil reported Effect is short lasting
Tocainide 60 mg/kg Not effective Nausea, parasthesia, rash Risk of aplastic anaemiaprecludes its routine use
Drugs evaluated in case reports only none over 25 patients
Capsaicin topicalon skin
mg for 21–28days
Little benefit Burning sensation Temporary relief ,avoidcontact with the eye
Clonazepam 2–8 mg May be effective Lethargy in 60%, fatigue, dizziness,personality change
Thromocytopenia can occur
Gabapentin 1200–3600 mg
May be effective Ataxia, dizziness, drowsiness, nausea,headache
Better tolerated thancarbamazepine
Oxcarbazepine 300–1200 mg Effective Ataxia, dizziness, diplopia, lethargy whichmay be related to hyponatraemia dosedependent
Better tolerated than withcarbamazepine
Phenytoin 200 – 300 mg Effective Ataxia, lethargy, nausea, headache,behavioural changes , folate deficiency inprolonged use, gingival hypertrophy
Small margin for doseescalation
Valproic acid 600 – 2000 mg May be effective Irritability, restlessness, tremor, confusion,nausea, rash, weight gain
get shorter and shorter. The most common divisions tobe affected are the second and third, and it is rare for thefirst division alone to be affected. In about 3% ofpatientsthepain becomesbilateral, but it isunusual forboth sidesto be active at the same time.Other features that need to be assessed include the qual-ity of life and the level of anxiety and depression. Mostpatients with severe trigeminal neuralgia find it impos-sible to socialise because of fear of developing an at-tack of pain while eating. These patients will often loseweight and become depressed. Ideally, patients shouldbe evaluated with standard assessment measures suchas the McGill Pain Questionnaire, some form of anx-iety or depression scale such as the Hospital Anxietyand Depression Scale, and a quality of life assessmentsuch as the Brief Pain Inventory or SF36. Some patients
will have, what are termed, atypical features and will re-port a constant, dull, aching background pain, exhibit noparoxysmal features, or have no pain free periods. Someof these symptoms may be related to stress and anxiety.Although 70% of patients will gain relief of pain fromthe use of carbamazepine, this is not always diagnostic.
Examination
In most patients, neurological examination will be nega-tive but some subtle sensory changes may be present. Insome cases, this could indicate that there is a secondarycause for trigeminalneuralgia. It is important to examinethe oral cavity to exclude any dental causes for pain, es-pecially if thepatient reports intra-oralpain.Assessmentof hearing may be important if patients wish to undergoan operation that carries a risk of causing hearing loss.

T
Trigeminal Neuralgia, Etiology, Pathogenesis and Management 2553
Investigation
There are no diagnostic tests for trigeminal neuralgia. ACT scan can be used to look for evidence of secondarycauses such as benign or malignant tumours or cysts.MRIscanswill also showwhether there isacompressionof the nerve by blood vessels. However, their specificityand sensitivity in being able to predict operative findingsstill remain under investigation.
Management
Trigeminal neuralgia is a rare condition, and althoughit can be treated in its initial phases in primary care,most of these patients benefit from being referred to thesecondary care sector, to clinicians who specialise inthe management of this condition. Patients, themselves,should be well-informed about the condition since com-pliance and satisfaction have been shown to be improvedin those patients who have a better understanding oftheir condition, and are given an opportunity to expresstheir wishes in terms of treatment. The � expert patientis also able to give truly informed consent to treatment.Most patients will initially be managed medically andthen surgery will be offered as a secondary procedure.There remains considerable debate as to the best timingof surgery.
Medical management (trigeminal neuralgia, medicalmanagement)
The mainstay of treatment for this condition is the useof anti-convulsants, and carbamazepine is the only drugthat has been the subject of three randomised, con-trolled trials (McQuay et al. 1995; Wiffen et al. 2005;Zakrzewska and Lopez 2005). It therefore remains thegold standard against which other drugs are evaluated.
Trigeminal Neuralgia, Etiology, Pathogenesis and Management, Table 2 Surgical management of trigeminal neuralgia ( � trigeminal neuralgia,aims of surgical management ), data on 5 years not available for some procedures, sensory loss common in all Gasserian ganglion* procedures
Procedure % Probability of beingpain free
Mortality Morbidity
Peripheral neurectomy, cryother-apy, alcohol, injection, acupunc-ture
Two years: 22 Nil Low, sensory loss, transient haematoma, oedema
Radiofrequency thermorhizotomy(RFT) *
Two years: 68Five years: 48
Low 28% complications mainly relating to trigeminal nerve,dyseasthesia, anesthesia dolorosa, eye problems, massetericproblems
Percutaneous glycerol rhizotomy* Two years: 63Five years: 45
Low 25% complications as for RFT
Balloon microcompression* Two years: 79 Low 10% complications as for RFT
Microvascular decompression Two years: 81Five years: 76Ten years: 71
0.5% Overall 75% no complications, 14% peri-operative complications,
5% transient cranial nerve 4th,6th,8th dysfunction, 2% permanentdeafness
Gamma Knife surgery Two years: 58 Nil Late onset of relief, may only be partial, 8% sensory loss up totwo years post treatment
Table 1 gives an indication of the drugs that are in use.Patients should be encouraged to keep pain diaries sothey learn to take control of their medication. Once thepatient has become pain free for a month, the drugsshould be slowly tapered off in the hope that the patienthas gone into a natural remission period. Patients, how-ever, need to be warned that they should re-start theirmedication as soon as they develop new paroxysms ofpain.
Surgery
Major indications for referring patients for surgeryinclude inability to control the pain, poor quality oflife, and side effects from medication. Surgery can becarried out at three levels, at the peripheral level wheretreatments are aimed at the trigger points, at the levelof the Gasserian ganglion which involves ablative pro-cedures, and at the level of the posterior fossa whereone procedure is ablative whereas the other, that ofmicrovascular decompression (� trigeminal neuralgia,microvascular decompression), is non-ablative. All ab-lative surgery (� trigeminal neuralgia, types of ablativesurgery) is likely to result in sensory loss in the area ofthe trigeminal nerve as tactile, thermal and pain fibresmay be destroyed. There are no randomised controlledtrials of surgical treatments, and the quality of data re-porting outcomes after surgical treatments is relativelypoor (Zakrzewska and Lopez 2003). However, there isgeneral consensus that the more central the procedure,the more likely patients are to gain long-term pain reliefand high satisfaction rates (Zakrzewska 2002). Table 2provides data on different surgical procedures (Burchiel1999; Maesawa et al. 2001; Nurmikko and Eldridge2001).

2554 Trigeminal Neuralgia, Ignition Theory
Patient InformationandSupport (trigeminal neuralgia, patientinformation)
There is now a considerable amount of literature ontrigeminal neuralgia for patients in the English lan-guage. The information is in the form of printed articlesas well as a book (Weigel and Casey 2000), and there isa considerable body of data on the internet produced bypatient support groups (www.tna-uk.org.uk; www.tna-support.org). Patients need to be aware that continuingcare may be necessary and not everyone can be cured.
References1. Anonymous (1988) Classification and Diagnostic Crite-
ria for Headache Disorders, Cranial Neuralgias and FacialPain. Headache Classification Committee of the InternationalHeadache Society. Cephalalgia 8:1–96
2. Devor M, Amir R, Rappaport ZH (2002) Pathophysiology ofTrigeminal Neuralgia: the Ignition Hypothesis. Clin J Pain18:4–13
3. Burchiel KJ (1999) (ed) Trigeminal Neuralgia in Techniques inNeurosurgery 5:200–266 Series of Articles by variety of authors
4. Maesawa S, Salame C, Flickinger JC et al. (2001) Clinical Out-comes after Stereotactic Radiosurgery for Idiopathic TrigeminalNeuralgia. J Neurosurg 94:14–20
5. McQuay H, Carroll D, Jadad AR et al. (1995) AnticonvulsantDrugs for Management of Pain: A Systematic Review. BMJ311:1047–1052
6. Merskey H, Bogduk N (1994) Classification of Chronic Pain.Descriptors of Chronic Pain Syndromes and Definitions of PainTerms. IASP Press, Seattle
7. Nurmikko TJ, Eldridge PR (2001) Trigeminal Neuralgia – Patho-physiology, Diagnosis and Current Treatment. Br J Anaesth87:117–132
8. Weigel G, Casey KF (2004) Striking Back – The Trigeminal Neu-ralgia and Face Pain Handbook. Whitehall Printing Company,Naples, Florida
9. Wiffen PJ, McQuay HJ, Moore RA (2005) Carbamazepine foracute and chronic pain. The Cochrane Database of SystematicReviews: Reviews 2005. Issue 3 John Wiley & Sons, Ltd. Chich-ester, UK DOI.:10.1002/14851858.CD005451
10. Zakrzewska JM (2002) Trigeminal Neuralgia. In: ZakrzewskaJM, Harrison SD (eds) Assessment and Management of OrofacialPain. Elsevier Sciences, Amsterdam, pp 267–370
11. Zakrzewska JM, Hamlyn PJ (1999) Facial Pain. In: CrombieIKCPR, Linton SJ, LeResche L et al. (eds) Epidemiology of Pain.IASP Press, Seattle, pp 171–202
12. Zakrzewska JM, Lopez BC (2003) Quality of Reporting in Eval-uations of Surgical Treatments of Trigeminal Neuralgia: Recom-mendations for Future Reports. Neurosurgery 53:110–122
13. Zakrzewska JM, Lopez BC (2005) Trigeminal Neuralgia. ClinEvid 14:1669–1677
Trigeminal Neuralgia, Ignition Theory
Definition
Trigeminal neuralgia is thought to be caused by chronicirritation of the trigeminal nerve, which leads to ectopichyperexcitability,and this is the basis of the ignition hy-pothesis.� Trigeminal Neuralgia, Etiology, Pathogenesis and
Management
Trigeminal Neuralgia, MedicalManagement
Definition
Medical management of trigeminal neuralgia is princi-pally with the use of anticonvulsant drugs, few of whichhaveundergoneevaluationunderrandomizedcontrolledtrial conditions.� Trigeminal Neuralgia, Etiology, Pathogenesis and
Management
Trigeminal Neuralgia, MicrovascularDecompression
Definition
Microvascular decompression is a major neurosurgicalprocedure that involves gaining entry into the posteriorfossaof theskull, and identifying anddecompressing thetrigeminal nerve in order to provide pain relief withoutsensory loss.� Trigeminal Neuralgia, Etiology, Pathogenesis and
Management
Trigeminal Neuralgia, Patient Information
Definition
There is a considerable amount of information, bothwritten and electronic, on trigeminal neuralgia for bothhealthcare workers and patients, which would aid inmanagement.� Trigeminal Neuralgia, Etiology, Pathogenesis and
Management
Trigeminal Neuralgia, Types of AblativeSurgery
Definition
Ablative surgery for trigeminal neuralgia involves se-lective destruction of the trigeminal nerve, and many ofthese procedures can be done in elderly medically un-fit patients, but most result in facial sensory loss. Theseprocedures include radiofrequency thermocoagulation(electrically heating the Gasserian ganglion), percuta-neous glycerol injection (injecting glycerol round theGasserian ganglion) or balloon microcompression (ap-plying pressure on the Gasserian ganglion for a few sec-onds).� Trigeminal Neuralgia, Etiology, Pathogenesis and
Management

T
Trigeminal, Glossopharyngeal, and Geniculate Neuralgias 2555
Trigeminal Nucleus Caudalis
Definition
Part of the trigeminal nuclei that relays intracranial no-ciceptive information to higher integrative centers.� Brainstem Subnucleus Reticularis Dorsalis Neuron
Trigeminal Subnucleus Caudalis
Definition
Trigeminal subnucleus caudalis is the most caudal com-ponentof thetrigeminalsystem,continuouswiththedor-sal horn of the spinal cord.� Dental Pain, Etiology, Pathogenesis and Management
Trigeminal Tractotomy
Definition
A surgical procedure to sever the V spinal tract. The le-sion also involves some lateral part of Vc. When per-formed in humans, the purpose is to deprive Vc of theinputs from the primary afferents, and therefore to pro-vide relief to patients from the excruciating pain of Vneuralgia.� Trigeminal Brainstem Nuclear Complex, Physiology
Trigeminal Transition Zone
Definition
A region at the obex level where the subnuclei interpo-laris (Vi) and caudalis (Vc) of the spinal trigeminal nu-cleus converge. The ventral portion of the laminated Vcis pushed dorsomedially by Vi. The substantia gelati-nosa is still identifiable but often interrupted. Neuronsin the trigeminal transition zone project to the thalamus,and are involved in processing of nociceptive informa-tion from the orofacial regions.� Trigeminothalamic Tract Projections
Trigeminal, Glossopharyngeal, andGeniculate Neuralgias
WILLIAM S. ANDERSON, CAROL JAMES,BENJAMIN S. CARSON SR.Department of Neurological Surgery, Johns HopkinsHospital, Baltimore, MD, [email protected]
Synonyms
Trigeminal neuralgia: tic douloureux, Fothergill’s dis-ease, epileptiform neuralgiaGlossopharyngeal neuralgia: vagoglossopharyngealneuralgiaNervus Intermedius, Primary Otalgia
Definitions
Cranial neuralgias are idiopathic chronic pain con-ditions characterized by sudden, often short-lived,episodes of pain that may result from non-painfulcutaneous stimulation, and that involve the distribu-tions of the involved trigeminal, glossopharyngeal, orgeniculate nerve branches.
Characteristics
Epidemiology is challenged by the lack of consistentlyapplieddiagnosticcriteria, theabsenceofobjectivemea-sures, and the variety of facial pain syndromes encoun-tered by the clinician (Kitt 2000). Trigeminal neuralgia(TN) is the most common cranial neuralgia. Studies inMinnesota report a TN incidence rate of approximately5/100,000 population (Katusic et al. 1991). Rates arehigher in women, 5.9 vs 3.4. The ratio of glossopharyn-geal neuralgia (GN) has been reported to be from 5 to200-fold less than TN. Geniculate neuralgia (GenN) isconsidered rare, with experienced neurosurgeonsreportseeing 10–15 GenN cases among several thousand casesof cranial neuralgias.Cranial neuralgias are usually diseases of seniors (IHS1997). The median age of diagnosis in the Minnesotaseries was 67 (Katusic et al. 1991), although the rangeis wide and younger patients are common, especially inmultiple sclerosis (see � Central Pain in Multiple Scle-rosis). A few series and case reports have described fa-milial occurrences of TN, although no specific geneticdefects have been identified, and no linkage studies ofthese families have been performed. An aggregation ofthe case reports shows that family members tend to beafflicted on the same side of the face, have a tendency todevelop bilateral TN, have a younger age of onset (44.4years), and tend to demonstrate an autosomal dominantmode of transmission with reduced penetrance (Fleet-wood 2001).The known causes of TN include demyelinating dis-eases such as multiple sclerosis, infiltration of the root orGasserian ganglion by tumor or amyloid deposition, orsmall pontine or medullary infarcts (Love and Coakham2001). About 4% of TN patients have multiple sclero-sis, and 1–5% of multiple sclerosis patients developsTN (Nurmikko and Eldridge 2001). The majority ofcases are believed to be due to nerve compression ef-fects, usually by vascular structures (80–90%) (Loveand Coakham 2001). A recent operative series of mi-crovascular decompression procedures showed that

2556 Trigeminal, Glossopharyngeal, and Geniculate Neuralgias
90% of first operations revealed compression of thenerve root by either an artery or vein, while in the restof the cases compression of the root was caused byarachnoid thickening, or angulation or torsion of theroot axis (Ishikawa et al. 2002). Approximately 70%of this series had arterial compression, of which 82%were caused by the superior cerebellar artery (SCA).More than 9% had demonstrable venous compression.Herpes zoster is commonly associated with outbreaks.The causes of GN and GenN may be similar to that ofTN because effective therapies are similar, but direct ev-idence is lacking. Several groups have studied the patho-logic consequences of trigeminal nerve root compres-sion. Devor et al. describe areas of axonal loss and de-myelination close to the compressing vessel with an in-crease in number of astrocytic processes, a froth of li-posomes (the residual myelin sheaths) as well as largeand small diameter denuded axons (Devor et al. 2002).Clumps of pure collagen are found in the demyelinationzone, as well as regions of dysmyelination in the rootadjacent to the offending vessel. The length of a given� cranial nerve central nervous system (CNS) portionmay be associated with the incidence of neuralgia af-fecting that cranial nerve (De Ridder et al. 2002). For in-stance, TN has a higher incidence than hemifacial spasm(CN VII), which has a higher incidence than GN, co-varying with the central segment of the involved nerveidentified by oligodendroglial cell derived myelin. TheCNS portion of the nerve may be subject to injury bycompression because it lacks the strong fascicular struc-ture of the peripheral nerve system portion (De Ridderet al. 2002). The relief many patients feel immediatelyafter decompressive surgery is too short for remyelina-tion to be a mechanism. More likely, relief may relateto a decrease in ectopic pulse generation, to an increasein fiber separation brought about by the decompression,and to the rapid reversal of conduction blockade in theremaining myelinated fibers (Love and Coakham 2001).
Clinical Features
TN is characterized by a painful sensation in the face,characterized by paroxysms of electric shock like painwithin one or more of the trigeminal nerve divisions.The pain can occur spontaneously, or be elicited by non-noxious stimuli such as touch, talking, eating, or wind.The pain is acute in onset and termination, and mayshow periods of remission (IHS 1997). Pain is usuallyunilateral, but bilateral cases are observed. Neurologicdeficits are generally absent. Case reports of atypicalTN are common.Severe paroxysmal pain in the sensory domain of theglossopharyngeal nerve is similar to TN, except for theposition within the distribution of the auricular and pha-ryngeal branches of the vagus nerve and that of the glos-sopharyngealnerve: theposteriorpartof the tongue, ton-sillar fossa, pharynx, beneath the angle of the lower jaw,or in the ear (Loeser 1990). GN may be mistaken for
mandibular division TN. The trigger can be swallowing,yawning, clearing the throat, or talking. GN, like TN, isgenerally unilateral although there are reports of bilat-eral GN, GN with TN, and atypical cases.GenN is a very rare condition, which is characterized bydeep pain affecting the distribution of the nervus inter-medius: inner ear with radiation to parts of the face withthe pinna of the ear being the most common spot. Un-like trigeminal neuralgia, the pain can last for hours at atime, but like trigeminal neuralgia spontaneous remis-sions can occur. Pain can be triggered by non-noxiousstimuli of the ear canal, swallowing, or talking (Loeser1990).
Pharmacological Treatments
Patients who present with TN are initially treatedwith medication, and the first line drug therapy iscarbamazepine, with typical maintenance doses of1500–2000 mg/day (Sindrup and Jensen 2002). Carba-mazepine was shown to be effective at reducing painseverity, number of paroxysms, and number of triggersin approximately 75% of patients. Side effects includesedation, rash, hyponatremia, and rarely agranulocy-tosis. Many clinicians have switched their patients tooxcarbamazepine, a keto-derivative of carbamazepinewith a lower side-effect profile, and less chance ofhematologic problems.Phenytoin is often used as a second-line agent, but fewcontrolled trialshavebeenperformedwiththisdrug; typ-ical maintenance doses are 300–400 mg/day (Sindrupand Jensen 2002). Baclofen has also been used for paincontrol in these syndromes. Newer anti-epileptic drugshave also been investigated. Lamotrigine, as an add-ondrug to either carbamazepine or Phenytoin, was shownto be effective.The relative roles of pharmacotherapy and surgery (seebelow) for TN are in a state of flux, although certainlymedically refractory patients tend to progress to surgerymore quickly. A few authors have argued that surgeryshould be utilized earlier, before the typical TN featuresevolve into the atypical features of constant backgroundpainandsensorydisturbances,whileothersquestion thistheory ofdiseaseprogression. Manyreportsobserve thatthe pharmacotherapy of GN and the rare GenN are thesame as TN.
Surgical Treatment
Surgical therapy of TN includes an assortment of treat-ments, ranging from relatively low invasive therapiessuch as stereotactic radiosurgery, peripheral nervebranch procedures including peripheral neurectomy,foraminal neurectomy and occlusion, or cryotherapy,percutaneous ganglion level procedures including RFthermocoagulation, glycerolysis, balloon compression,and � microvascular decompression (MVD) proce-dures. These latter procedures involve a suboccipitalcraniectomy to reverse vascular compressive of the

T
Trigeminocervical Complex 2557
nerve. Jannetta and coworkers described the largest se-ries of cranial neuralgias treated with MVD, includingTN and GN (McLaughlin et al. 1997).Reviews of the various surgical treatments have gener-ally found that ganglion level procedures tended to bemoreeffective than peripheralprocedures;however,nei-ther group produced long-term pain relief. For instance,RF thermocoagulation by itself provided initial pain re-lief rates of 91–99% with subsequent recurrence rates of10–25% over the various study times (25% at 14 years),while peripheral neurectomy by itself had a success rateof only 64% at one year, which fell to 26% by 4 years(Peters and Nurmikko 2002).An emerging treatment for TN is stereotactic radio-surgery. Pollock et al. published a series of 117 patientswith an average age of 67.8 years, 58% of whom hada previous surgical treatment. An excellent outcomeoverall (complete pain relief without medication) wasachieved of 57% at 1 year, and 55% at 3 years. Factorsassociated with a good outcome were normal preoper-ative facial sensation, increased radiation dosage, noprior surgery, and trigeminal dysfunction/numbness.Negative factors included multiple sclerosis and atypi-cal pain features. These authors stress that since the longterm effects of radiation therapy close to the brainstem isunknown, first line treatment for young healthy patientsshould continue to address the vascular compressivelesion via an MVD procedure (Pollock et al. 2002).Patel et al. (2002) reviewed the experience of 217 pa-tients who underwent MVD for GN neuralgia between1973 and 2000. In this group, immediate relief (no pain,and no medication) was obtained by 67% and partialrelief by 25%, while the long-term results showed com-plete relief 58% and partial relief 18%. Complicationsof the procedure were few and included intracranialhematoma, brainstem infarction, cranial nerve palsy,CSF leak, operative death (none since 1987), and dys-phagia, and seemed to decrease over time (Patel et al.2002).Surgery forGenNisbased on theconcept thatpain isme-diated through the nervous intermedius and the genic-ulate ganglion. In general, rare conditions such as thisshould be referred to individuals who have had signifi-cant experience with them, since historically morbidityassociated with surgical treatment of this condition isrelatively high. The surgical treatment involves dissect-ing out the nervous intermedius from the seventh cra-nial nerve in the cerebellar pontine angle and section-ing it. Variable results have been reported with MVD.One of the major problems with GenN is distinguishingit from GN or even from TN, which affects the third divi-sion of the nerve. This confusion has led some to advo-cate not only cutting the nervous intermedius, but alsothe glossopharyngeal nerve and the upper third of thevagus nerve. Unfortunately, there are no specific teststhat are guaranteed to predict surgical success, althoughsome otolaryngologists feel that it is useful to do pha-
ryngealcocaineblocksprior toattemptingnervesection-ing. Long-term results for surgical treatment of GenN in64 patients have been reviewed by Pulec (Pulec 2002).
References1. De Ridder D, Møller A, Verlooy J, Cornelissen M, De Ridder L
(2002) Is the Root Entry/Exit Zone Important in MicrovascularCompression Syndromes? Neurosurgery 51:427–434
2. Devor M, Govrin-Lippmann R, Rappaport ZH (2002a) Mech-anism of Trigeminal Neuralgia: An Ultrastructural Analysis ofTrigeminal Root Specimens Obtained During Microvascular De-compression Surgery. J Neurosurg 96:532–543
3. Fleetwood IG, Innes AM, Hansen SR, Steinberg GK (2001) Fa-milial Trigeminal Neuralgia: Case Report and Review of the Lit-erature. J Neurosurg 95:513–517
4. International Headache Society (IHS) (1997) Member’s Hand-book 1997/98: Classification and Diagnostic Criteria forHeadache Disorders, Cranial Neuralgias, and Facial Pain.Scandinavian University Press, Oslo, pp 102–103
5. Ishikawa M, Nishi S, Aoki T, Takase T, Wada E, Ohwaki H, Kat-suki T, Fukuda H (2002) Operative Findings in Cases of Trigemi-nal Neuralgia Without Vascular Compression: Proposal of a Dif-ferent Mechanism. J Clin Neurosci 9:200–204
6. Kastusic S, Williams DB, Beard CM, Bergstralh EJ, Kurland LT(1991) Epidemiology and Clinical Features of Idiopathic Trigem-inal Neuralgia and Glossopharyngeal Neuralgia: Similarities andDifferences, Rochester, Minnesota, 1945–1984. Neuroepidemi-ology 10:276–281
7. Loeser JD (1990) In: The Management of Pain, vol 1, 2nd edn.Cranial Neuralgias, Lea & Febiger, Philadelphia
8. Love S, Coakham HB (2001) Trigeminal Neuralgia: Pathologyand Pathogenesis. Brain 124:2347–2360
9. McLaughlin MR, Jannetta PJ, Clyde BL, Subach BR, Comey CH,Resnick DK (1997) Microvascular Decompression of CranialNerves: Lessons Learned after 4400 Operations. J Neurosurg90:1–8
10. Nurmikko TJ, Eldridge PR (2001) Trigeminal Neuralgia:Pathophysiology, Diagnosis and Current Treatment. Br J Anaes87:117–132
11. Patel A, Kassam A, Horowitz M, Chang Y-F (2002) Microvas-cular Decompression in the Management of GlossopharyngealNeuralgia: Analysis of 217 Cases. Neurosurgery 50:705–711
12. Peters G, Nurmikko TJ (2002) Peripheral and GasserianGanglion-Level Procedures for the Treatment of TrigeminalNeuralgia. Clin J of Pain 18:28–34
13. Pollock BE, Phuong LK, Gorman DA, Foote RL, Stafford SL(2002) Stereotactic Radiosurgery for Idiopathic Trigeminal Neu-ralgia. J Neurosurg 97:347–353
14. Pulec JL (2002) Geniculate Neuralgia: Long-Term Results ofSurgical Treatment. Ear Nose Throat J 81:30–33
15. Sindrup SH, Jensen TS (2002) Pharmacotherapy of TrigeminalNeuralgia. Clin J of Pain 18:22–27
Trigeminocervical Complex
Definition
Neurons in the trigeminal nucleus caudalis and dorsalhorn of spinal cord segments C1 and C2, which act to-gether as a functional relay for pain input from intracra-nial structures, and overlap with input from the front andback of the head.� Migraine, Pathophysiology

2558 Trigeminohypothalamic Tract
Trigeminohypothalamic TractXIJING J. ZHANG
Department of Neuroscience, University of Minnesota,Minneapolis, MN, [email protected]
Definition
The trigeminohypothalamic tract is a bundle of nervefibersoriginating from allof thesubnucleiof the trigemi-nalbrainstem nuclear complexand thegraymatterofup-percervical spinalcord segments (C1–2), and projectingto or through the hypothalamus. The trigeminohypotha-lamic tract is responsible for conveying sensory infor-mation, especially nociceptive information, from facialskin, cornea, oral mucosa, and intracranial dura to thehypothalamus, a brain area that regulates homeostasisand other hormonal responses required for survival ofthe organism.
Characteristics
Anatomical retrograde labeling and physiologicalantidromic activation techniques have identified thelocations of the trigeminohypothalamic neurons in thetrigeminal brainstem nuclear complex and the gray mat-ter of upper cervical spinal cord segments (Malick andBuratein 1998; Malick et al. 2000). After the retrogradetracer Fluoro-Gold injection into the hypothalamus,trigeminohypothalamic neurons are found throughoutall of the subnuclei of the trigeminal brainstem nuclearcomplex (56%) and upper cervical spinal cord seg-ment (44%) (Malick and Buratein 1998). Within thetrigeminal brainstem nuclear complex, over 75% of theneurons were distributed caudal to the obex; most of theneurons were located in the nucleus caudalis. Over 90%of the neurons in the nucleus caudalis were distributedin laminae I, II and V. The nucleus caudalis is the subnu-cleus in the trigeminal brainstem nuclear complex thatprocesses nociceptive information arising in oral andfacial organs (Sessle 1987; Jacquin et al. 1986). In cer-vical cord segments 1 and 2, approximately 85% of theneurons were found in laminae I, II and V. These areasreceive direct input from trigeminal primary afferentfibers and contain neurons that respond to mechanicalor thermal stimulation of the cornea, oral mucosa, tem-poromandibular joint, facial skin, and intracranial dura(Lu et al. 1993; Broton et al. 1988, Burstein et al. 1998).Trigeminohypothalamic neurons were also recordedin laminae I, II and IV, V of the nucleus caudalis, andupper cervical (C1) spinal cord by physiological an-tidromic activation studies (Malick et al. 2000). Most ofthe trigeminohypothalamic neurons that were recordedresponded maximally or exclusively to noxious me-chanical or thermal stimulation to the head and orofacialreceptive fields innervated by the trigeminal nerve.
The trajectory and termination of the trigeminohypotha-lamic tract have been examined using antidromic acti-vation (Malick et al. 2000). The trigeminohypothalamicaxons cross the midline and ascend on the contralateralside of the brainstem to the level of the contralateralthalamus. Within the thalamus, trigeminohypothalamicaxons shift through the optic tract, internal capsule,and supraoptic decussation and reach the rostral ven-tral hypothalamus. In the rostral ventral hypothalamusmore than half of the axons cross the midline again atthe posterior optic chiasm to the ipsilateral hypotha-lamus, turn caudally and descend along the identicalpath in which they ascended in the contralateral hy-pothalamus (Malick et al. 2000). The axons of thetrigeminohypothalamic neurons and their collateralbranches terminate bilaterally in many nuclei withinhypothalamus, such as the lateral, perifornical, dorso-medial, suprachiasmatic, and supraoptic hypothalamicnuclei (Malick et al. 2000). Through this complexprojection, the axons of the trigeminohypothalamicneurons are capable of carrying nociceptive informa-tion bilaterally to many nuclei in the hypothalamus thatare involved in the production of various responses tonoxious stimuli.Responses of the trigeminohypothalamic neurons tonoxiousstimulihavebeendescribed(Malicketal.2000).The overwhelming majority of the trigeminohypothala-mic neurons responded strongly to noxious mechanicaland thermal stimuli applied to the skin. The receptivefields of the trigeminohypothalamic neurons includedthe skin of the head and neck, as well as orofacialorgans such as the oral mucosa, tongue, lips, cornea,and intracranial dura. Laminae I-II neurons gener-ally exhibited small to medium receptive fields, whilethose located in deeper layers had medium to largereceptive fields. Over 80% of the recorded trigemi-nohypothalamic neurons were nociceptive. Amongthem, 42% were wide dynamic range, and 38% werehigh-threshold neurons. The remaining 20% were low-threshold neurons (Malick et al. 2000). Figure 1 showsthat a high-threshold trigeminohypothalamic neuronresponded to noxious mechanical stimulus applied toits receptive field. The neuron was located in dorsome-dial lamina V of the nucleus caudalis (b), and its axonprojected to the contralateral hypothalamus (a). Theneuron had a small cutaneous receptive field locatedaround the ipsilateral side of the mouth (c), and onlyresponded to noxious mechanical stimuli applied to theskin (d).The majority of nociceptive neurons responded primar-ily to noxious intensities of thermal stimulation. In con-trast, all low-threshold neurons responded to both heatand cold stimuli at innocuous and noxious intensities, aphenomenon that has not been reported for other pop-ulations of spinal or trigeminal low-threshold neurons(Malick et al. 2000). Thus, trigeminohypothalamic neu-rons can transmit innocuous and primarily noxious me-

T
Trigeminohypothalamic Tract 2559
TrigeminohypothalamicTract, Figure 1 Responseproperty of a high-thresholdtrigeminohypothalamic neuron. (a)Location of low-threshold pointfor antidromic activation in thehypothalamus. (b) Recording site indorsomedial lamina V. (c) Receptivefield. (d) Peristimulus histogramand record of window discriminatoroutput (below histogram) illustratingthe response to mechanical stimuliapplied to the receptive field.Numbers in parentheses depictmean response (spikes/sec) to eachstimulus. (From Malick et al. 2000).Hyp, hypothalamus; IC, internalcapsule; VBC, ventrobasal complex.
chanical and thermal information from facial skin di-rectly to the hypothalamus.Many trigeminohypothalamic neurons were also pow-erfully activated by electrical and mechanical stimuli ofthe oral mucosa, tongue, lips, cornea, and intracranialdura mater (Malick et al. 2000). Most (85%) oral-sensitive trigeminohypothalamic neurons encoded theintensity of the noxious mechanical and thermal stim-uli. Since pain is the only sensation that can be evokedby stimulating the cornea and intracranial dura andsinuses, regardless of whether the stimulus is electrical,mechanical, or chemical (Lele and Weddell 1959, Rayand Wolff 1940), the cornea- and dura-sensitive neuronswere considered nociceptive. Therefore, trigeminohy-pothalamic neurons can transmit nociceptive infor-mation from these specific organs in the head to thehypothalamus.The hypothalamus plays an important role in regulatingbody temperature, food and water intake, sleep andcircadian rhythms, endocrine adjustments, and a widerange of behavior (Swanson 1987). Through the termi-nations and collateral branches in the hypothalamus, thetrigeminohypothalamic tract is likely to bring noxiousand innocuous sensory signals from orofacial skin andorgans to the hypothalamus, and is involved in pain-
related autonomic responses, endocrine adjustments,or emotion reactions.
References1. Broton JG, Hu JW, Sessle BJ (1988) Effects of Temporomandibu-
lar Joint Stimulation on Nociceptive and Non-Nociceptive Neu-rons of the Cat’s Trigeminal Subnucleus Caudalis (MedullaryDorsal Horn). J Neurophysiol 59:1575–1589
2. Burstein R, Yamamura H, Malick A, Strassman AM (1998)Chemical Stimulation of the Intracranial Dura Induces EnhancedResponses to Facial Stimulation in Brainstem Trigeminal Neu-rons. J Neurophysiol 79:964–982
3. Jacquin MF, Renehan WE, Mooney RD, Rhoades RW (1986)Structure-Function Relationships in Rat Medullary and CervicalDorsal Horns. I. Trigeminal Primary Afferents. J Neurophysiol55:1153–1186
4. Lele PP, Weddell G (1959) Sensory Nerves of the Cornea andCutaneous Sensibility. Exp Neurol 1:334–359
5. Lu J, Hathaway CB, Bereiter DA (1993) Adrenalectomy En-hances Fos-Like Immunoreactivity within the Spinal TrigeminalNucleus Induced by Noxious Thermal Stimulation of the Cornea.Neuroscience 54:809–818
6. Malick A, Buratein R (1998) Cells of Origin of the Trigemino-hypothalamic Tract in the Rat. J Comp Neurol 400:125–144
7. Malick A, Strassman AM, Burstein R (2000) Trigeminohypotha-lamic and Reticulohypothalamic Tract Neurons in the Upper Cer-vical Spinal Cord and Caudal Medulla of the Rat. J Neurophysiol84:2078–2112

2560 Trigeminothalamic Tract Projections
8. Ray BS, Wolff HG (1940) Experimental Studies on Headache.Pain-Sensitive Structures of the Head and their Significance inHeadache. Arch Surg 41:813–856
9. Sessle BJ (1987) The Neurobiology of Facial and Dental Pain:Present Knowledge, Future Directions. J Dent Res 66:617–626
10. Swanson LW (1987) The Hypothalamus. In: Hökfelt T and Swan-son LW (eds) Handbook of Chemical Neuroanatomy, vol 5, Inte-grated Systems of the CNS, Part I. Elsevier, Amsterdam, pp 1–124
Trigeminothalamic Tract ProjectionsKE REN
Department of Biomedical Sciences, University ofMaryland, Baltimore, MD, [email protected]
Synonyms
Tractus Trigeminothalamicus; Lemnicus Trigeminalis
Definition
The axon bundles of the secondary sensory neurons inthe spinal trigeminal nucleus and principal sensory nu-cleus of the � trigeminal nerve, which carry somatosen-sory information from the head and face and terminate intheventralposteriorpartof theopposite thalamus.As thecounterpart of the spinothalamic tract, the trigeminotha-lamic axons from the spinal trigeminal nucleus, mainlythe subnucleus caudalis, convey nociceptive input. Thetrigeminothalamic axons from the principal sensory nu-cleus, equivalent to the medial � lemniscal fibers, trans-mit discriminative tactile as well as proprioceptive im-pulses.
Characteristics
The somesthetic information from orofacial regions isconveyed via thefifthcranialnerve, the trigeminalnerve,to the two major trigeminal sensory nuclei, the spinaltrigeminal and the principal sensory nuclei. Populationsof neurons in the trigeminal sensory nuclei then sendtheir axons to the thalamus via trigeminothalamic pro-jections.Thepattern and functionsof trigeminothalamicprojections are specifically related to the individual sub-nuclei of the spinal trigeminal nucleus and the thalamus(FukushimaandKerr1979;RausellandJones1991).Forthe purpose of completeness, the following discussionwill go beyond the projection to the ventrobasal thala-mus, to also include trigeminal projections to other nu-clei of the thalamus that are related to pain processing.
Spinal Trigeminal Nucleus
The spinal trigeminal nucleus is further divided rostro-caudally into the subnuclei oralis, interpolaris and cau-dalis. Cytoarchitecturally, the general classification ofthe subnuclei for humans is applicable to lower animalssuch as the rat.The subnucleus caudalis appears at approximately thelevel of the obex and continues caudally to merge with
the upper cervical spinal dorsal horn. The caudal portionof the subnucleus caudalis is analogous to the spinaldorsal horn both histologically and physiologically. TheRexed classification of the dorsal horn can be appliedto its outer layers. Thus, the subnucleus caudalis isregarded as a laminated structure, and is sometimes re-ferred to as the � medullary dorsal horn. After enteringthe central nervous system at the middle of the pons,small-diameter primary afferent (nociceptive) fibersin the trigeminal nerve turn caudally, then descend inthe spinal trigeminal tract, and finally terminate in thesubnucleus caudalis. Nociceptive neurons, both noci-ceptive specific and wide dynamic range types, can berecorded from the subnucleus caudalis. The thalamic-projecting axons of subnucleus caudalis neurons carrypain and temperature information to the thalamus.The subnucleus interpolaris appears gradually froma transition region at the level of the obex, where thesubnuclei caudalis and interpolaris coexist in the sametransverse plane for a small distance (the � trigeminalVi/Vc transition zone). The subnucleus interpolarisneurons have a well-defined and extensive projection tothe medial ventroposterior thalamic nucleus (VPM, themedial part of the ventrobasal complex) (Phelan andFalls 1991). Nociceptive thalamic projecting neuronshave been identified in subnucleus interpolaris. How-ever, most thalamic projecting subnucleus interpolarisneurons respond to deflection of mystacial vibrissaeand are low-threshold mechanoreceptive (Jacquin etal. 1986; Hayashi et al. 1984). Two types of thalamicascending fibers have been identified in the subnucleusinterpolaris: fast-conducting thick fibers terminate in theposterior nucleus of the thalamus, and slow-conductingthin fibers project to VPM (Veinante et al. 2000).The subnucleus oralis is the most rostral subnucleusof the spinal trigeminal nucleus. Fukushima and Kerr(1979) found that the subnucleus oralis had no projec-tions to the thalamus in the rat. Using fluorescent dyetracers, however, subnucleus oralis neurons have beenshown to project to the thalamus in rodents (Bruce etal. 1987). The neurons of the subnucleus oralis mayproject to the contralateral thalamus via a relay in thejuxtatrigeminal nucleus and principal sensory nucleus(Zhang and Yang 1999).
Principal Sensory Nucleus of the Trigeminal Nerve, TrigeminalNucleus Principalis
The principal sensory nucleus (main sensory nucleus)can be traced caudally to the level of the rostral tip ofthe facial nucleus and contiguous obliquely with sub-nucleus oralis. The principal sensory nucleus mainly re-ceives large-diameter primary afferents and contributesto discriminative sensations. In the rat, the principal sen-sory nucleus thalamicprojectingaxonsarise fromsmall-and medium-sized neurons; large neurons (20–42 μm)of the principal sensory nucleus do not project to the tha-lamus (Fukushima and Kerr 1979). The ascending fibers

T
Trigeminothalamic Tract Projections 2561
from the ventral and dorsal principal sensory nucleusproject to the contralateral and ipsilateral VPM, respec-tively.
Subnuclei of the Thalamus
The ascending trigeminal projections terminate in sev-eral thalamic subnuclei including the VPM, posteriorthalamic nucleus, nucleus submedius and intralaminarnuclei centralis medialis and lateralis and parafas-cicular nucleus. The principal sensory nucleus andspinal trigeminal nucleus have complementary pro-jection foci in the thalamus (Rausell and Jones 1991,Williams et al. 1994). The principal sensory nucleusinput enters the barreloid portion of VPM that contains� parvalbumin-immunoreactive cells. The ascend-ing fibers from spinal trigeminal nucleus terminate innon-barreloid VPM, which contains another calcium-binding protein, � calbindin-D28k. Calbindin D-28kand parvalbumin belong to calcium binding proteinfamilies and can be used as selective neural markers ofcentral neurons. The trigeminal inputs to the posteriorthalamic nucleus are mainly from the principal sensorynucleus and subnucleus interpolaris (Waite and Tracey1995). The parafascicular nucleus of the thalamus re-ceives input from subnuclei caudalis and interpolaris,but not from principal sensory nucleus and subnucleusoralis (Krout et al. 2002). The nucleus submedius re-ceives a major input from the Vi/Vc transition zone,as well as from the subnucleus caudalis (Yoshida et al.1991, Craig and Dostrovsky 2001).Most trigeminothalamic projections are contralateral,except that the subnucleus caudalis projects to bilateral� intralaminar thalamic nuclei (Peschanski 1984), andthe principal sensory nucleus has an ipsilateral pro-jection to VPM (Fukushima and Kerr 1979) (Fig. 1).Although the major trigeminothalamic projections arethe inputs to VPM and posterior thalamic nucleus fromthe principal sensory nucleus and subnucleus interpo-laris (Waite and Tracey 1995), they are generally notconcerned with nociceptive transmission, or at leastdo not play a major role. Most VPM neurons relayprecisely organized tactile information from the faceand mouth. Neurons in the posterior nucleus and medialthalamus including nucleus submedius, intralaminarnuclei and parafascicular nucleus are less somatotopi-cally organized in relaying sensory information than isthe VPM nucleus.
Trigeminothalamic Projection and Pain
Trigeminothalamic neurons are involved in transmit-ting pain information from the craniofacial region. Thetrigeminothalamic projection involved in pain is mainlyfrom the subnucleus caudalis, where small-diametertrigeminal primary afferents terminate. Specifically,neurons in the marginal layer (lamina I) of the subnu-cleus caudalis project to VPM and posterior thalamicnucleus (Shigenaga et al. 1979). Neurons in the sub-
Trigeminothalamic Tract Projections, Figure 1 Major thalamic projec-tions from the trigeminal sensory nuclei. Sensory inputs from the head andface are relayed in the trigeminal brain stem nuclear complex, includingthe principal sensory nucleus (PrV) and spinal trigeminal nucleus (SpV).The spinal trigeminal nucleus is further divided into subnuclei oralis (Vo),interpolaris (Vi) and caudalis (Vc). The most caudal subnucleus interpo-laris and rostral caudalis merge to form a transition zone (Vi/Vc) at theobex level. Large-diameter myelinated primary afferents mainly synapsein the principal sensory nucleus, and small-diameter A-delta and C-fiberstravel caudally to terminate in the subnucleus caudalis. Neurons in thespinal trigeminal nucleus, primarily the subnucleus caudalis, project tocontralateral medial ventroposterior nucleus of the thalamus (VPM), pos-terior thalamic nucleus (not shown), as well as bilateral intralaminar nuclei(ILN) and nucleus submedius (Sm). The trigeminal projection to the sub-medius is predominantly contralateral. The principal sensory nucleus andthe subnucleus caudalis project to the barreloid and non-barreloid regions(open circles in VPM) of the VPM, respectively. Note that discriminative andaffective components of pain may be relayed by distinct pathways. Dashedlines indicate transmission of discriminative tactile information from thehead and face.
nuclei interpolaris and oralis are also involved in theorofacial pain process, although limited informationis available on their ascending thalamic projectionsrelated to pain. One characteristic of the nociceptivetrigeminothalamic projection is that a relatively smallnumber of nociceptive neurons in the subnucleus cau-dalis projects to the VPM. This is in sharp contrast withthe principal sensory nucleus, where a vast majorityof cells are connected to VPM. However, nociceptivetrigeminothalamic neurons also project to the posteriorthalamic nucleus, nucleus submedius and intralaminarthalamic nuclei.

2562 Trigeminovascular System
Most lamina I trigeminothalamic nociceptive neu-rons in the subnucleus caudalis project to nucleussubmedius, but not VPM. In contrast, most thermore-ceptive trigeminothalamic cells project to VPM (Craigand Dostrovsky 2001). Subnucleus caudalis nocicep-tive neurons are antidromically activated by stimulationof the posterior nucleus of the thalamus (Hirata et al.1999).Using combined retrograde tracing and nuclear Fos pro-tein expression techniques, it has been shown that a pop-ulation of nucleus submedius-projecting neurons in theVi/Vc transition zone is activated by inflammation of themasseter muscle (Ikeda et al. 2003). In contrast, cornealresponsive units in the Vi/Vc transition zone do not ap-pear to project to the nucleus submedius (Hirata et al.1999).Thus, theVi/Vc-submediusprojectionmay bese-lectively activated by injury of deep orofacial structures.The trigeminal nociceptive input to the VPM is furtherrelayed to the somatosensory cerebral cortex and relatedto discriminative information of pain. In contrast, infor-mation about affective and emotional aspects of painfrom the head and face is likely to be related throughthe thalamic nucleus submedius and intralaminar nuclei(Fig. 1).
Trigeminothalamic Projection and Discriminative Sensations
Information related to discriminative sensations fromthe head and face is relayed in the VPM throughtrigeminothalamic projections. Neurons in the principalsensory nucleus receive synaptic contact from large-diameter primary afferent terminals of the trigeminalnerve and send axons ascending to VPM. This pathwaycorresponds to the spinal dorsal column/medial lem-niscal system, and is highly somatotopically organized.In rodents, distinct clusters of VPM cells receive inputsfrom designated whiskers, mediated by correspondingaggregates of neurons in the principal sensory nucleus.Neurons in the principal sensory nucleus, VPM andthe somatosensory cortex are grouped into rows (rods,cellular cylinders) to give topographical representa-tion of the whisker pad. This specific array of cells isnamed “barrels” in cortex, “barreloids” in VPM and“barrelets” in the principal sensory nucleus (Waite andTracey 1995; Hendry and Hsiao 2003). Informationrelated to discriminative pain from the head and faceis relayed in the non-barreloid thalamic regions of theVPM, complementary to those mediating tactile sensa-tions, or the barreloid areas (Rausell and Jones 1991;Williams et al. 1994).
References1. Bruce LL, McHaffie JG, Stein BE (1987) The Organization of
Trigeminotectal and Trigeminothalamic Neurons in Rodents: ADouble-Labeling Study with Fluorescent Dyes. J Comp Neurol262: 315–330
2. Craig AD, Dostrovsky JO (2001) Differential Projections ofThermoreceptive and Nociceptive Lamina I Trigeminothalamicand Spinothalamic Neurons in the Cat. J Neurophysiol 86:856–870
3. Fukushima T, Kerr FW (1979) Organization of Trigeminothala-mic Tracts and Other Thalamic Afferent Systems of the Brain-stem in the Rat: Presence of Gelatinosa Neurons with ThalamicConnections. J Comp Neurol 183: 169–184
4. Hayashi H, Sumino R, Sessle BJ (1984) Functional Organizationof Trigeminal Subnucleus Interpolaris: Nociceptive and Innocu-ous Afferent Inputs, Projections to Thalamus, Cerebellum, andSpinal Cord, and Descending Modulation from PeriaqueductalGray. J Neurophysiol 51:890–905
5. Hendry SH, Hsiao SS (2003) The Somatosensory System, In:Squire LR, Bloom FE, McConnell SK, Roberts JL, Spitzer NC,Zigmond MJ (eds) Fundamental neuroscience. 2nd ed. AcademicPress, London, pp 667–697
6. Hirata H, Hu JW, Bereiter DA (1999) Responses of MedullaryDorsal Horn Neurons to Corneal Stimulation by CO2 Pulses inthe Rat. J Neurophysiol 82:2092–2107
7. Ikeda T, Terayama R, Jue SS et al. (2003) Differential RostralProjections of Caudal Brainstem Neurons receiving TrigeminalInput after Masseter Inflammation. J Comp Neurol 465:220–233
8. Jacquin MF, Mooney RD, Rhoades RW (1986) Morphology, Re-sponse Properties, and Collateral Projections of Trigeminotha-lamic Neurons in Brainstem Subnucleus Interpolaris of Rat. ExpBrain Res 61:457–468
9. Krout KE, Belzer RE, Loewy AD (2002) Brainstem Projectionsto Midline and Intralaminar Thalamic Nuclei of the Rat. J CompNeurol 448:53–101
10. Peschanski M (1984) Trigeminal Afferents to the Diencephalonin the Rat. Neuroscience 12:465–487
11. Phelan KD, Falls WM (1991) A Comparison of the Distributionand Morphology of Thalamic, Cerebellar and Spinal ProjectionNeurons in Rat Trigeminal Nucleus Interpolaris. Neuroscience40:497–511
12. Rausell E, Jones EG (1991) Chemically Distinct Compartmentsof the Thalamic VPM Nucleus in Monkeys Relay Principaland Spinal Trigeminal Pathways to Different Layers of theSomatosensory Cortex. J Neurosci 11:226–237
13. Shigenaga Y, Takabatake M, Sugimoto T et al. (1979) Neuronsin Marginal Layer of Trigeminal Nucleus Caudalis Projecting toVentrobasal Complex (VB) and Posterior Nuclear Group (PO)Demonstrated by Retrograde Labeling with Horseradish Perox-idase. Brain Res 166:391–396
14. Veinante P, Jacquin MF, Deschenes M (2000) Thalamic Projec-tions from the Whisker-Sensitive Regions of the Spinal Trigem-inal Complex in the Rat. J Comp Neurol 420: 233–243
15. Waite Phil ME, Tracey DJ (1995) Trigeminal Sensory System.In: Paxinos G (ed) The rat nervous system, 2nd edn. AcademicPress, San Diego, pp 705–724
16. Williams MN, Zahm DS, Jacquin MF (1994) Differential Fociand Synaptic Organization of the Principal and Spinal TrigeminalProjections to the Thalamus in the Rat. Eur J Neurosci 6:429–453
17. Yoshida AK, Dostrovsky JO, Sessle BJ et al. (1991) TrigeminalProjections to the Nucleus Submedius of the Thalamus in theRat. J Comp Neurol 307:609–625
18. Zhang JD, Yang XL (1999) Projections from Subnucleus Oralisof the Spinal Trigeminal Nucleus to Contralateral Thalamus viathe Relay of Juxtatrigeminal Nucleus and Dorsomedial Part ofthe Principal Sensory Trigeminal Nucleus in the Rat. J Hirnforsch39:301–310
Trigeminovascular System
Definition
The trigeminovascular system designates the visceralterminals of the 1st division of the trigeminal nervesurrounding vessels in the meninges. It is the majorpain-signaling structure of the visceral organ brain.� Clinical Migraine without Aura� Migraine, Pathophysiology

T
TrkA Receptor(s) 2563
Trigger Point
Definition
A trigger point is a spot of localized tenderness in a pal-pable taut band of muscle. Pressure stimulation or pal-pation on trigger points evoke a characteristic pattern ofreferred pain and typically a local twitchresponse.Thereare also scar, skin, and connective tissue sensitive spotsthat, when stimulated, mechanically refer pain and aresometimes called trigger points.� Chronic Pelvic Pain, Musculoskeletal Syndromes� Muscle Pain, Fibromyalgia Syndrome (Primary, Sec-
ondary)� Myofascial Trigger Points� Psychophysiological Assessment of Pain� Referred Muscle Pain, Assessment� Stretching
Trigger Point Pain
� Myofascial Pain
Trigger Point Pressure Release
Definition
Trigger point pressure release is a simple, often effec-tive, manual technique for the treatment of myofascialtriggerpoints.Slowly increasing,gentle,digitalpressureis applied to the trigger point until a barrier of tissue re-sistance is encountered. Constant pressure is maintaineduntil the resistance of the barrier tension decreases. Thenpressure is slowly increased to reach a new barrier. Theterm ischemic compression has previously been used todescribe this treatment approach, but a different conceptof the treatment mechanism was attributed to it, and theprocedure was unnecessarily painful.� Myofascial Trigger Points
Triple Response
Definition
The triple response consists of three components. Inresponse to an intradermal histamine injection, thereis a local erythema (reddening reaction) occurringwithin 10 seconds due to a direct histamine inducedcapillary dilatation at the injection site. Then, there isa local swelling, a „wheal“, due to locally increasedpermeability of capillaries and postcapillary venuleswith consecutive fluid extravasation. Finally, there is a„flare“, i.e. an erythema occurring within 30 seconds
in neighboring skin areas due to axon reflex activation.Histamine activates nociceptive, unmyelinated C-fibersat the injection site. Their impulses travel orthodromi-cally. As soon as they have reached axon branchingpoints, impulses also propagate antidromically downother neighboring branches of the sensory nerve fibers.The antidromic impulses reach axon terminals andinduce release of vasodilating and itching neuropep-tides, such as calcitonine gene related peptide (CGRP)or substance P, which also causes fluid extravasation(Ganong WF (1997) Review of Medical Physiology.Appleton & Lange; Stamford, CT p581-582).� Congenital Insensitivity to Pain with Anhidrosis
Triptans
Definition
Serotonin, 5–HT1B/1D, receptor agonist, i.e. compoundsthat activate or turn on both these receptor sub-types.These compounds are highly effective in the treatmentof acute migraine.� Clinical Migraine without Aura� Migraine, Pathophysiology
trkA
� Tyrosine Kinase A
TrkA Receptor(s)
Definition
TrkA is a member of the trk family of tyrosine kinase re-ceptors and is the high affinity receptor for nerve growthfactor found on NGF-dependent sensory neurons. WhentrkA binds NGF, the receptor autophosphorylateson ty-rosine residues, leading to activation of multiple down-stream effector proteins, including the Ras-MAP kinasesignaling cascade and PI3–kinase activation.� Congenital Insensitivity to Pain with Anhidrosis� IB4-Positive Neurons, Role in Inflammatory Pain� Immunocytochemistry of Nociceptors� Nerve Growth Factor, Sensitizing Action on Nocicep-
tors� TRPV1, Regulation by Nerve Growth Factor

2564 TrkA-IgG
TrkA-IgG
Definition
NGF sequestering molecule.� Spinal Cord Nociception, Neurotrophins
TRN
� Thalamic Reticular Nucleus
Trochanteric Bursitis
Definition
Inflammation of the bursa near the greater trochanter ofthe hip. This condition can produce pain in the thigh.� Sciatica
Trophic Factors
Definition
Molecules that cause growth/regeneration of variousparts of a cell/neuron; more recently some trophicfactors have also been shown to have transmitter-likeactions.� Retrograde Cellular Changes after Nerve Injury
Tropism
Definition
Movement or growth of an organism in response to anexternal stimulus.� Hansen’s Disease
TRP Channels
Definition
Denominates ion channels of theTransient ReceptorPo-tential (TRP) family. The first member of this ion chan-nel family was identified by localization of a gene thatcaused deficiencies in signaling in the visual system ofthemodelorganismDrosophilamelanogaster (fruitfly).Mammals have more than 14 TRP channel genes. TRPchannels play important roles in the regulation of neuralexcitability and in sensory systems, such as vision, ol-faction, and pheromone sensation. TRP channels serve
as receptors for temperature and irritant chemicals (cap-saicin, mustard oil) in the pain pathway, and play impor-tant functional roles in neurogenic inflammation and hy-peralgesia.� Nociceptors, Cold Thermotransduction� TRPV1, Regulation by Nerve Growth Factor� TRPV1, Regulation by Protons
TRPA1
Definition
Temperature-sensitive, non-selective cation channel ac-tivated near 17˚C. The channel is insensitive to mentholbut is activated by natural pungent compounds like cin-namon and mustard oil. The transcripts are expressedin a small percentage (4%) of primary sensory neurons.Found exclusively in neurons expressing TRPV1 andpeptidergic markers of nociceptors, such as calcitoningene-related peptide (CGRP). Suggested as the molec-ular transducer of noxious cold temperatures.� Nociceptors, Cold Thermotransduction
TRPM8 Channel
Definition
Calcium permeable, voltage-gated, non-selective cationchannel that isactivatedbytemperature(threshold25˚C)and natural cooling compounds like menthol and euca-lyptol.Expressed inaround10%ofprimarysensoryneu-ronsofsmalldiameter.Acrucialelement in the transduc-tion of temperature signals by low-threshold peripheraland visceral thermoreceptors.� Nociceptors, Cold Thermotransduction
TRPV1
Synonyms
Transient Receptor Potential Vanilloid Type 1
Definition
The TRPV1 receptor is a member of the transient recep-tor vanilloid subfamily. It is sensitive to capsaicin andhas a well defined temperature threshold of about 43˚C,similar to the threshold for thermal nociception. The re-sponse of this receptor can be sensitized to NGF andother proinflammatory mediators providing a basis forthermal hyperalgesia. Other TRP subfamily membersare not sensitive to capsaicin, and have different temper-ature thresholds (e.g. TRPV2 – 52˚C; TRPV3 – 35˚C).Still other members of this subfamily appear to respondto other stimuli, e.g. TRPV4 – osmoreceptors. TRPV1

T
TRPV1 Modulation by p2Y Receptors 2565
is the charter member of a family of TRP channels iden-tified in mammalian sensory neurons that are activatedby temperatures ranging from cold to warm to intenseheat.� Capsaicin Receptor� ERK Regulation in Sensory Neurons during Inflam-
mation� IB4-Positive Neurons, Role in Inflammatory Pain� Muscle Pain Model, Ischemia-Induced and Hyper-
tonic Saline-Induced� Nerve Growth Factor, Sensitizing Action on Nocicep-
tors� NGF, Regulation during Inflammation� Nociceptor, Categorization� Satellite Cells and Inflammatory Pain� Species Differences in Skin Nociception� TRPV1 Modulation by p2Y Receptors� TRPV1 Receptor, Species Variability� TRPV1, Regulation by Nerve Growth Factor� TRPV1, Regulation by Protons� Visceral Pain Model, Esophageal Pain
TRPV1 Modulation by p2Y ReceptorsMAKOTO TOMINAGA
Department of Cellular and Molecular Physiology,Mie University School of Medicine, Tsu Mie, [email protected]
Synonyms
TRPV1 Receptor, Modulation by P2Y Receptors
Definition� Capsaicin receptor � TRPV1 is a non-selectivecation channel expressed in a subset of sensory neurons,� nociceptors. � P2Y receptor activation potentiates orsensitizes TRPV1 activity through a � PKC-dependentpathway. In the presence of � ATP released in thecontext of tissue damage, the temperature thresholdfor TRPV1 activation is reduced to less than 35˚C, sothat body temperature is capable of activating TRPV1,leading to the sensation of pain.
Characteristics
Pain is initiated when noxious thermal, mechanical, orchemical stimuli excite the peripheral terminals of spe-cialized primary afferent neurons called nociceptors.Many different kinds of ionotropic and metabotropicreceptors are known to be involved in this process(McCleskey and Gold 1999). � Vanilloid receptors arenociceptor-specific cation channels that serve as themolecular target of capsaicin, the pungent ingredient inhot chili peppers (Szallasi and Blumberg 1999). A geneencoding a capsaicin receptor was isolated using an
expression-cloning method and the receptor proteinwas found to be an ion channel with six transmembranedomains having high Ca2+ permeability (Caterina et al.1997). When expressed in heterologous systems, thecloned capsaicin receptor TRPV1 can also be activatedby noxious heat (with a thermal threshold > 43˚C) orprotons (acidification), both of which cause pain invivo (Caterina et al. 1997; Tominaga et al. 1998; Cate-rina and Julius 2001). Furthermore, analyses of micelacking TRPV1 have shown that TRPV1 is essentialfor selective modalities of pain sensation and for tissueinjury-induced thermal hyperalgesia (Caterina et al.2000; Davis et al. 2000).Tissue damage associated with infection, inflammationor ischemia produces an array of chemical mediatorsthat activate or sensitize nociceptor terminals to elicitpain at the site of injury. One important componentof this pro-algesic response is adenosine triphosphate(ATP) released from different cell types (North andBarnard 1997; Burnstock and Williams 2000). Extra-cellular ATP excites the nociceptive endings of nearbysensory nerves, evoking a sensation of pain. In theseneurons, the most widely studied targets of extracellu-lar ATP have been ionotropic ATP � (P2X) receptors.Indeed, several P2X receptor subtypes have been iden-tified in sensory neurons, including one (P2X3) whoseexpression is largely confined to these cells (North andBarnard 1997). The importance of widely distributedmetabotropic ATP (P2Y) receptors in nociception hasbeen recently reported (Molliver et al. 2002; Zimmer-mannetal.2002). Inparticular, thefunctionalinteractionbetween P2Y receptors and TRPV1 has received muchattention because TRPV1 is one of the key moleculesdetecting nociceptive stimuli and because the signalingpathway downstream of P2Y receptor activation canbe applicable for other Gq-protein coupled receptoractivation.Pretreatment with 100 μM extracellular ATP (an at-tainable concentration in the context of tissue damage)causes more than 6 times the potentiation of capsaicin(low doses)-evoked current responses in HEK293 cellsheterologously expressing TRPV1 channels (Fig. 1a)(Tominaga et al. 2001). A similar potentiating effect ofextracellular ATP is observed on proton-evoked activa-tion of TRPV1. The dose-response curves for capsaicinin the presence or absence of ATP demonstrate thatATP enhances capsaicin and proton action on TRPV1by lowering � EC50 values without altering maximalresponses (Fig. 1b). Extracellular ATP also lowers thethreshold temperature for TRPV1 activation signifi-cantly (from about 43˚C to about 35˚C) (Fig. 1c). Thus,in the presence of ATP, normally non-painful thermalstimuli (even body temperature) are capable of acti-vating TRPV1. These data show that TRPV1 currentsevoked by any of three different stimuli (capsaicin,proton or heat) are potentiated or sensitized by extra-cellular ATP. Activation of protein kinase C (PKC) by

2566 TRPV1 Modulation by p2Y Receptors
TRPV1 Modulation by p2Y Receptors, Figure 1 (a) Extracellular ATP (100 μM) potentiates capsaicin (CAP)-activated currents in HEK293 cells expressingTRPV1. Whole-cell patch-clamp recordings were carried out with holding potential of -60 mV. Cells were perfused for 2 min with solution containingATP before re-exposure to capsaicin. (b) Capsaicin dose-response curves for TRPV1 in the absence (λ) and presence (μ) of 100 μM extracellular ATP.Currents were normalized to the currents maximally activated by 1 μM capsaicin in the absence of ATP. Figure shows averaged data fitted with theHill equation. EC50 = 114.7 nM and Hill coefficient = 1.48 in the absence of ATP. EC50 = 49.3 nM and Hill coefficient = 1.56 in the presence of ATP.Values represent the mean ± SEM. (c) Reduction of the threshold temperature for TRPV1 activation by extracellular ATP. Representative temperature-response profiles in the absence and presence of 100 μM extracellular ATP. Dashed lines show the threshold temperature for heat activation of VR1.Temperature threshold for activation of TRPV1 in the presence of ATP (35.3 ± 0.7˚C) was significantly lower than that in the absence of ATP (41.7 ± 1.1˚C).*, p < 0.001 (Tominaga et al. 2001).
diacylglycerol (DAG) downstream of P2Y1 receptorsis found to be a mechanism for the ATP-induced po-tentiation from various pharmacological analyses inHEK293 cells (Fig. 2a), consistent with the reportthat PKC-ε is specifically involved in sensitization ofheat-activated channels by � bradykinin in dorsal rootganglion (DRG) neurons.The interaction between ATP and TRPV1 in the con-text of ATP-induced hyperalgesia in vivo is confirmedby a behavioral analysis using wild type mice andTRPV1-deficient mice (Moriyama et al. 2003). A sig-nificant reduction in paw withdrawal latency to radiantpaw heating is observed for 5 to 30 min followingATP injection in wild type mice. On the other hand,TRPV1-deficient mice develop no such thermal hyper-sensitivity in response to ATP injection, suggesting afunctional interaction between ATP and TRPV1 (Fig.2b). Mice lacking P2Y1 receptors, a subtype provedto be involved in ATP-induced potentiation of TRPV1currents in HEK293 cells, showed similar thermalhyperalgesia to wild type mice, indicating that other
P2Y subtypes are involved in ATP-induced thermalhyperalgesia in mice.Electrophysiological and pharmacological analysesusing DRG neurons of mice revealed that P2Y2 isa subtype involved in ATP-induced potentiation ofTRPV1 currents and ATP-induced thermal hyperal-gesia in mice (Moriyama et al. 2003). UTP, a potentP2Y2 receptor agonist, potentiates the capsaicin-evokedcurrent responses and causes thermal hyperalgesia to asimilar extent as ATP in mice, further confirming theinvolvement of P2Y2 receptors. P2Y2 receptor mRNAbut not P2Y1 mRNA is found to be co-expressed withTRPV1 mRNA in the rat lumbar DRG using doublein situ hybridization, suggesting that P2Y2, not P2Y1receptors, can functionally interact with TRPV1 in
DRG neuronsThe data described above suggest that direct phosphory-lation of TRPV1 or a closely associated protein by PKCchanges the agonist sensitivity of this ion channel. Thein vivo phosphorylation of TRPV1 by PKC is confirmedin HEK293 cells expressing TRPV1 (Numazaki et al.

T
TRPV1 Modulation by p2Y Receptors 2567
TRPV1 Modulation by p2YReceptors, Figure 2 (a)Regulation mechanisms of TRPV1by P2Y receptors. Gq-coupledP2Y receptor activation leadsto production of IP3 and DAGthrough PLCβ. PKC activationby DAG causes phosphorylationof TRPV1, leading to functionalpotentiation. PLCβ, phospholipaseCβ; DAG, diacylglycerol; IP3,inositol 1,4,5-trisphosphate; P,phosphorylation; o and I, outsideand inside of cell, respectively.(b) TRPV1 is essential for thedevelopment of ATP-inducedthermal hypersensitivity in vivo.Wild type (γ) or TRPV1 -/- mice (γ)were injected intraplantarly withATP (100 nmol), and the responselatency to radiant heating of thehind paw was measured at varioustime points after injection. * p <
0.05 and ** p < 0.01 vs. wild typemice. (Moriyama et al. 2003).
2002).Furthermore, two serine residuesassubstrates forPKC-dependent phosphorylation, S502 in the first intra-cellular loop and S800 in the C-terminus, were identifiedusing an in vitro kinase assay (Numazaki et al. 2002). Amutant lacking substrates for PKC-dependent phospho-rylation (S502A/S800A) exhibited almost no potentia-tion effects of capsaicin- or proton-evoked current re-sponses by PMA (a direct activator of PKC). Further-more, the double mutant showed no reduction in tem-perature thresholdforTRPV1activation, suggesting thatthe two serine residues are the major substrates for PKC-dependent phosphorylation.Inflammatory pain is initiated by tissue damage/inflam-mation and is characterized by hypersensitivity both atthe site of damage and in adjacent tissue. One mech-anism underlying these phenomena is the modulation(sensitization) of ion channels, such as TRPV1, that de-tect noxious stimuli at the nociceptor terminal. Sensi-tization is triggered by extracellular inflammatory me-diators that are released in vivo from surrounding dam-aged or inflamed tissue and from nociceptive neuronsthemselves.Amongthemediators,ATPnotonlypotenti-atescapsaicin-orproton-evokedcurrentsbutalso lowersthe temperature threshold for heat activation of TRPV1,such that normally non-painful thermal stimuli (i.e. nor-malbody temperature) arecapableofactivating TRPV1,
making ATP act like a direct activator of TRPV1. Thisrepresents a novel mechanism through which extracel-lular ATP may cause inflammatory pain. Most attentionin the pain field has focused on the role of ionotropicATP receptors in ATP-evoked nociception. The abovefindings suggest that P2Y2 is also involved in this pro-cess and may represent a fruitful target for the develop-ment of drugs that blunt nociceptive signaling throughcapsaicin receptors. P2Y2 receptors confer responsive-ness to uridine triphosphate (UTP) and ATP to a simi-lar extent, suggesting a possible role for UTP as an im-portant component of pro-algesic response in the con-text of tissue injury. UTP was indeed found to potentiatecapsaicin-activated currents and cause thermal hyperal-gesia in mice (Moriyama et al. 2003).These results suggest that activation of similar PKC-dependent events might underlie certain nociceptiveeffects of other Gq-coupled metabotropic receptors.Indeed, bradykinin is found to potentiate or sensitizeTRPV1 in a similar PKC-dependent pathway throughthe activation of B2 receptors (Sugiura et al. 2002).
References1. Burnstock G, Williams M (2000) P2 purinergic receptors: mod-
ulation of cell function and therapeutic potential. J PharmacolExp Ther 295:862–869

2568 TRPV1 Modulation by PKC
2. Caterina MJ, Schumacher MA, Tominaga M et al. (1997) Thecapsaicin receptor: a heat-activated ion channel in the pain path-way. Nature 389:816–824
3. Caterina MJ, Leffler A, Malmberg AB et al. (2000) Impairednociception and pain sensation in mice lacking the capsaicin re-ceptor. Science 288:306–313
4. Caterina MJ, Julius D (2001) The vanilloid receptor: a moleculargateway to the pain pathway. Annu Rev Neurosci 24:487–517
5. Davis, J.B., Gray, J., Gunthorpe, M.J et al. (2000) Vanilloidreceptor-1 is essential for inflammatory thermal hyperalgesia.Nature 405:183–187
6. McCleskey EW, Gold MS (1999) Ion channels of nociception.Annu Rev Physiol 61:835–856
7. Molliver DC, Cook SP, Carlsten JA et al. (2002) ATP and UTP ex-cite sensory neurons and induce CREB phosphorylation throughthe metabotropic receptor, P2Y2. Eur J Neurosci 16:1850–1860
8. Moriyama T, Iida T, Kobayashi K et al. (2003) Possible involve-ment of P2Y2 metabotropic receptors in ATP-induced transientreceptor potential vanilloid receptor 1-mediated thermal hyper-sensitivity. J Neurosci 23:6058–6062
9. North AN, Barnard EA (1997) Nucleotide receptors. Curr OpinNeurobiol 7:346–357
10. Numazaki M, Tominaga T, Toyooka H et al. (2002) Direct phos-phorylation of capsaicin receptor VR1 by PKCε and identificationof two target serine residues. J Biol Chem 277:13375–13378
11. Sugiura T, Tominaga M, Katsuya H et al. (2002) Bradykininlowers the threshold temperature for heat activation of vanilloidreceptor 1. J Neurophysiol 88:544–548
12. Szallasi A, Blumberg PM (1999) Vanilloid (capsaicin) receptorsand mechanisms. Pharmacol Rev 51:159–211
13. Tominaga M, Caterina MJ, Malmberg AB et al. (1998) The clonedcapsaicin receptor integrates multiple pain-producing stimuli.Neuron 21:531–543
14. Tominaga M, Wada M, Masu M (2001) Potentiation of capsaicinreceptor activity by metabotropic ATP receptors as a possiblemechanism for ATP-evoked pain and hyperalgesia. Proc NatlAcad Sci USA 98:6951–6956
15. Zimmermann K, Reeh PW, Averbeck B (2002) ATP can en-hance the proton-induced CGRP release through P2Y receptorsand secondary PGE(2) release in isolated rat dura mater. Pain97:259–265
TRPV1 Modulation by PKCPETER A. MCNAUGHTON
Department of Pharmacology, University ofCambridge, Cambridge, [email protected]
Synonyms
VR1; PKC; protein kinase C; TRPV1 Modulation byPKC
Definition
TRPV1 is an ion channel located in the surface mem-brane of pain-sensitive nerve terminals or nocicep-tors, which is activated by capsaicin, heat and protons.TRPV1 when open conducts electric charge, in the formof cations (mainly Na+ and Ca2+) and therefore makesthe interior of the nociceptor more positive, leadingto the generation of action potentials. The temperaturethreshold of TRPV1 is lowered when inflammatory me-diators such as bradykinin or ATP interact with surfacemembrane receptors on nociceptive nerve terminals and
so the nerve terminal becomes hyperalgesic (hypersen-sitive to heat pain). Many inflammatory mediators actby stimulating an intracellular enzyme known as pro-tein kinase C (a member of the large kinase family ofenzymes), which attach phosphate groups to specificlocations on the portion of the TRPV1 protein exposedto the intracellular milieu of the nociceptor terminal.
Characteristics
A role for PKC in nociceptor sensitization was firstsuspected in studies of the potent sensitizing agentbradykinin, a pro-inflammatory nonapeptide which isreleased from a larger precursor protein by proteolyticcleavage following tissue damage (Dray and Perkins1993). Bradykinin is one of the most potent pain-producing substances known, and as well as causingpain directly, it acts as a sensitising agent which lowersthe temperature threshold for the activation of heat pain
TRPV1 Modulation by PKC, Figure 1 Action potentials and generatorcurrents elicited in a nociceptive neuron by a noxious heat stimulus. (a)Application of a 49˚C heat stimulus depolarizes a nociceptive neuron tothreshold and elicits a train of action potentials. This temperature did notdamage the neuron as repeated application of the stimulus gave similarresults. (b) Dependence of the membrane current on temperature in a heat-sensitive neuron before and after activation of PKC by phorbol myristateacetate (PMA), a PKC-specific activator. After activation of PKC the activationthreshold shifts to lower temperatures and the magnitude of the currentincreases.

T
TRPV1 Modulation by PKC 2569
TRPV1 Modulation by PKC,Figure 2 Pathways leading to heathyperalgesia. Binding of bradykininto its receptor activates PLCβ, whichin releases DAG from PIP2, leadingto activation of PKCε, which in turnphosphorylates TRPV1 at two serineresidues, leading to the lowering ofheat threshold shown in Fig. 1b.
in vivo (Mizumura and Kumazawa 1996). In isolatednociceptive neurons action potentials are elicited byheat stimuli (Fig. 1a), just as they are in vivo, and theinward membrane current responsible can be recordedusing the whole-cell voltage clamp technique (Fig. 1b).The properties of the heat-gated ion current, such asits threshold and dependence on temperature, closelycorrespond to those of heat pain in vivo (Cesare andMcNaughton 1996). Bradykinin is a potent agonist atthe G-protein coupled B2 receptor, leading to activa-tion of Gq and phospholipase C β (PLCβ), followedby metabolism of phosphatidylinositol bisphosphate(PIP2) and release of diacylglycerol (DAG) and inositoltrisphosphate (IP3). These two products have differentactions: DAG activates protein kinase C (PKC) whileIP3 releases calcium from intracellular organelles. Thecritical member of this cascade which is responsible forcausing sensitization of the heat-gated membrane cur-rent isPKC,because thesensitising effectsofbradykininare mimicked by direct PKC activation (Fig. 1b), areantagonized by PKC inhibitors such as staurosporineand are promoted by phosphatase inhibitors such ascalyculin A (Cesare and McNaughton 1996; Cesareand McNaughton 1997).TRPV1 is activated by heat stimuli, and is the moleculeresponsible for the heat hyperalgesia caused by inflam-mation, because no heat hyperalgesia is seen in animalsfrom which TRPV1 has been genetically deleted (Daviset al. 2000; Caterina et al. 2000). Two main models havebeen proposed for the molecular basis of this hyperal-gesia. In experiments on isolated neurons (see above)PKC has been identified as a critical mediator of heathyperalgesia. PKC acts by phosphorylating serine orthreonine residues on its target proteins and phosphory-lation sites relevant for the process of sensitisation havebeen identified by the use of site-directed mutagenesis
of individual serine or threonine residues, followedby expression of the mutant TRPV1 in a heterologousexpression system (Numazaki et al. 2002). Mutationof two serines to alanine, which cannot be phosphory-lated, abolished sensitization following PKC activation(Fig. 2). The isoform of PKC responsible has also beenidentified (Cesare et al. 1999). Of the eleven knownisoforms of PKC only five, namely PKCβI, βII, δ, ε
and ζ, are expressed to any significant extent in sen-sory neurons and of these only PKCε is translocatedto the membrane following exposure to bradykinin,suggesting that it is this isoform which is responsiblefor sensitization of TRPV1. A central role for PKCε insensitization was confirmed by showing that constitu-tively active PKCε incorporated into the cell was indeedcapable of sensitising the heat-gated current, and thatthe incorporation of a specific PKCε inhibitor into thecell largely abolished sensitisation (Cesare et al. 1999).A specific role for PKCε in nociceptor sensitization isalso suggested by the reduced heat hyperalgesia seen instudies using PKCε knockout mice (Khasar et al. 1999).The model of specific phosphorylation of TRPV1 byPKCε is shown in outline in Fig. 2.In an alternative model (Chuang et al. 2001) TRPV1 isproposed tobetonically inhibitedbyPIP2in theneuronalcell membrane. Removal of PIP2, when bradykinin ac-tivates PLCβ, is then proposed to release TRPV1 frominhibition. This model is supported by experimentsin which application of PLCβ to isolated membranepatches in order to break down PIP2, or removal ofPIP2 with an antibody, potentiated gating of TRPV1(Chuang et al. 2001). The proposal also draws on workcarried out on other ion channels, particularly on in-ward rectifier potassium channels, where a modulationof ion channel function by PIP2 is well established(Hilgemann 2003). It has yet to be clearly established

2570 TRPV1 Modulation by PKC
whether this mechanism plays an important role inTRPV1 sensitization, but the observations cited abovesuggest that it plays at most a minor role by comparisonwith the PKCε pathway summarized in Fig. 2.
References1. Caterina MJ, Leffler A, Malmberget AB et al. (2000) Impaired
nociception and pain sensation in mice lacking the capsaicin re-ceptor. Science 288:306–313
2. Cesare P, McNaughton PA (1996) A novel heat-activated currentin nociceptive neurons, and its sensitization by bradykinin. ProcNatl Acad Sci USA 93:15435–15439
3. Cesare P, McNaughton PA (1997) Peripheral pain mechanisms.Curr Opin Neurobiol 7:493–499
4. Cesare P, Dekker LV, Sardini A et al. (1999) Specific involve-ment of PKC-epsilon in sensitization of the neuronal responseto painful heat. Neuron 23:617–624
5. Chuang HH, Prescott ED, Kong H et al. (2001) Bradykininand nerve growth factor release the capsaicin receptor fromPtdIns(4,5)P2-mediated inhibition. Nature 411:957–962
6. Davis JB, Gray J, Gunthorpe MJ et al. (2000) Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature405:183–187
7. Dray A, Perkins M (1993) Bradykinin and inflammatory pain.Trends Neurosci 16:99–104
8. Hilgemann DW (2003) Getting ready for the decade of the lipids.Annu Rev Physiol 65:697–700
9. Khasar SG, Lin Y-H, Martin A et al. (1999) A novel nociceptorsignaling pathway revealed in protein kinase c epsilon mutantmice. Neuron 24:253–260
10. Mizumura K, Kumazawa T (1996) Modification of nociceptorresponses by inflammatory mediators and second messengersimplicated in their action – A study in canine testicular polymodalreceptors. Prog Brain Res 113:115–141
11. Numazaki M, Tominaga T, Toyooka H et al. (2002) Direct phos-phorylation of capsaicin receptor VR1 by protein kinase Cep-silon and identification of two target serine residues. J Biol Chem277:13375–13378
TRPV1 Modulation by PKC
� TRPV1 Modulation by PKC
TRPV1-Null Mice
Definition
Mice that have a disruption of the TRPV1 gene. Thesemice lack expression of the TRPV1 protein and have noresponse to capsaicin.� IB4-Positive Neurons, Role in Inflammatory Pain
TRPV1 Receptor
� TRPV1
TRPV1 Receptor, Modulation by P2YReceptors
� TRPV1 Modulation by p2Y Receptors
TRPV1 Receptor, Species VariabilityPETER MCINTYRE
Department of Pharmacology, University ofMelbourne, Melbourne, VIC, [email protected]
Synonyms
Transient receptor potential cation channel, subfamilyV, member 1; TRPV1; Capsaicin receptor; Vanilloid Re-ceptor Subtype 1; VR1
Definition
TRPV1isanon-specificcationchannelof the� transientreceptor potential protein family, which is expressedin � polymodal sensory neurons in the peripheral sen-sory nervous system and is activated by noxious heat(>43˚C), pH below 6.5, capsaicin, the pungent ingre-dient of chili peppers, resiniferatoxin and a numberof other vanilloid, phorbol-related compounds as wellas the endogenous activators anandamide and someproducts of lipoxygenase action on arachidonic acid.
Characteristics
Relatives of TRPV1
TRPV1 is a non-selective cation channel with high cal-cium permeability. It has a long cytoplasmic N-terminaltail with 3 ankyrin repeats and 6 hydrophobic domains(termed S1–S6 here) which may be transmembrane do-mains and a putative pore-forming region between S5and S6. It is a member of the superfamily of ion channelswith six hydrophobic transmembrane domains and a pu-tativepore-forming loop betweenthe last two hydropho-bic domains. This superfamily includes the voltage acti-vated potassium channels, hyperpolarization and cyclicnucleotide gated channels (HCN) and the transient re-ceptor potential (TRP) ion channels, which are involvedin many sensory processes.
Splice Variants
TRPV1 was first isolated by expression cloning fromrat sensory neurons (Caterina et al. 1997) and has sub-sequently been isolated from human (McIntyre et al.2001; Cortright et al. 2001; Hayes et al. 2000), guineapig (Savidge et al. 2002), mouse (Accession numberAJ620495), rabbit (Accession number AY487342) andchicken (Jordt and Julius 2002) sources and partial se-

T
TRPV1 Receptor, Species Variability 2571
quences are available from gene predictions from dogand zebrafish. Cloning and sequencing of the humanand mouse genes has shown that the trpv1 gene has 16exons in mouse and human predicted by Ensembl with15 coding exons and that there are four splice variants ofmRNA. The TRPV1 gene is found at position 17p13.2in the human genome and in a highly � syntenic regionon chromosome 10 in the rat genome (Xue et al. 2001)and chromosome 11 in the mouse genome (Caterinaet al. 2000). The predicted protein size is 839 aminoacids in human, guinea pig and mouse (~95 kD), 838amino acids in rat, 842 in the rabbit and 843 (96.5 kD)in chicken. The rat protein can be glycosylated at as-paragine 604, migrates with a higher apparent molecularmass in its glycosylated form and can form tetramersthat are likely to be the functional form of the channel(Kedei et al. 2001).An N-terminal splice variant, with most of the ankyrindomains and the cytoplasmic tail missing, has beenobserved (Schumacher et al. 2000b). The transcripttermed VR.5’sv is not functional but the protein is ex-pressed in rat kidney and this splice variant is expressedat low levels in dorsal root ganglia compared to TRPV1(Sanchez et al. 2001). The importance of this transcript,if any, is still not understood but it appears that an intactN-terminus is essential for TRPV1 to be functional.An earlier report of a TRPV1 transcript termed stretch-inhibited channel (SIC) with differences at both C-and N-termini (Schumacher et al. 2000a) seems to bederived from two independent genes and is unlikely tobe a functional TRPV1 transcript (Xue et al. 2001).
Single Nucleotide Polymorphism
One non-synonymous � single nucleotide polymor-phism resulting in the amino acid substitution fromvaline to isoleucine at position 585 has been reported inthe human TRPV1 sequence (Hayes et al. 2000). Geno-typing of DNA from 123 randomly selected, mixedrace individuals, showed that 51% of the populationare heterozygous, the homozygous valine-encodingallele occurs in 15% and the homozygous isoleucine-encoding allele occurs in 34% of the population. These
TRPV1 Receptor, Species Variability, Table 1 TRPV1 percent amino acid identity (top triangle) and divergence (lower triangle) calculated usingClustal W
rat mouse rabbit human guinea pig chicken
rat *** 94.9 86.9 85.7 88.0 65.8
mouse 5.1 *** 88.0 86.3 88.5 66.0
rabbit 14.2 13.0 *** 88.0 87.5 65.2
human 15.4 14.5 13.0 *** 86.5 64.0
guinea pig 12.6 12.3 13.6 14.3 *** 66.0
chicken 41.0 40.9 41.3 43.1 40.7 ***
TRPV1 alleles both give rise to functional channelswith no reported differences in pharmacology (Hayeset al. 2000).
Protein Sequence Alignment
Alignment of the predicted proteins using Clustal Wshows that the highest variation occurs at the N- and C-terminal regions of the protein and within the putativepore region. Overall homology scores (Table 1) showthat rat and mousesequenceshavemost identity (94.9%)and that apart from these closely related sequences, themammalian proteins determined so-far share between85.7% and 88.5% identity. The mammalian proteinshave between 64% and 66% identity to the chickenprotein.
Pharmacological Differences
Agonists
The most extreme example of variation in pharmaco-logical characteristics of TRPV1 is between the rat andchicken � orthologues. Primary sensory neurons fromchicks are responsive to heat (~45˚C) and low pH (pH 4)but not capsaicin (Marin-Burgin et al. 2000). The clonedchicken TRPV1 has these properties when expressed inXenopus oocytes or mammalian cells (Jordt and Julius2002). Jordt and Julius showed, using chimeric recep-tors, that the molecular determinants for this differencelay insequencewithin theS3andS4regions;specificallyresidues responsible for capsaicin sensitivity are tyro-sine511andserine512,whereasarginine491wasshownto modulate the ratio of capsaicin-evoked current to pH-evoked current in electrophysiological experiments.Differences inagonistsensitivityhavebeenfoundwithinTRPV1 from mammalian species. The rabbit TRPV1 isunresponsive to capsaicin and substitution of threonine553 for isoleucine is largely responsible for this phe-notype (Gavva et al. 2003). Rat TRPV1 responds wellto the agonist phorbol 12-phenylacetate 13-acetate 20-homovanillate (PPAHV) but the human (McIntyre et al.2001) and guinea pig (Savidge et al. 2002) channels donot. This difference has been mapped to a single con-servative amino acid change. Mutation of leucine 457

2572 TRPV1 Receptor, Species Variability
TRPV1 Receptor, SpeciesVariability, Figure 1 Alignment ofthe available predicted full-lengthproteins using Clustal W (DNAstar).Predicted ankyrin repeat regionsare shaded grey and hydrophobictransmembrane domains areunderlined and marked S1−S6.The most conserved regions arethe hydrophobic domains whichare thought to be buried in themembrane and the least conservedregions are the N-terminal andC-terminal regions and the proximalpart of the putative pore regionbetween S5 and S6.
to methionine enables the human TRPV1 to respond toPPAHV (Phillips et al. 2004).
Antagonists
There is a pharmacological difference in the ability ofcapsazepine to antagonize the low pH and heat activa-tion of the rat TRPV1 (McIntyre et al. 2001). Whereasit blocks these modalities well in human and guinea pigTRPV1, it has little or no effect on these modalities in rat
TRPV1. The residues responsible for this change weremutated to the human equivalents and restored the phe-notype and thus the difference was mapped to aminoacids isoleucine 514, valine 518 and methionine 547 inthe rat TRPV1.In summary, several groups have shown that residues be-tween hydrophobicdomains S2 and S4 are important inrecognition of agonists and antagonists and that smallvariations in sequence can have significant effects on thepharmacology of this ion channel.

T
TRPV1 Receptor, Species Variability 2573
TRPV1 Receptor, SpeciesVariability, Figure 1 (continued)
References1. Caterina MJ, Schumacher MA, Tominaga M et al. (1997) The
capsaicin receptor: a heat-activated ion channel in the pain path-way. Nature 389:816–824
2. Caterina MJ, Leffler A, Malmberg AB et al. (2000) Impairednociception and pain sensation in mice lacking the capsaicin re-ceptor. Science 288:306–313
3. Cortright DN, Crandall M, Sanchez JF et al. (2001) The tis-sue distribution and functional characterization of human VR1.Biochem Biophys Res Commun 281:1183–1189
4. Gavva NR, Klionsky L, Qu Y et al. (2003) Molecular deter-minants of capsaicin sensitivity in rabbit TRPV1. Society forNeuroscience Abstracts 811.12
5. Hayes P, Meadows HJ, Gunthorpe MJ et al. (2000) Cloning andfunctional expression of a human orthologue of rat vanilloidreceptor-1. Pain 88:205–215
6. Jordt SE, Julius D (2002) Molecular basis for species-specificsensitivity to “hot” chili peppers. Cell 108:421–430
7. Kedei N, Szabo T, Lile JD et al. (2001) Analysis of the na-tive quaternary structure of vanilloid receptor 1. J Biol Chem276:28613–28619

2574 TRPV1, Regulation by Acid
8. Marin-Burgin A, Reppenhagen S, Klusch A et al. (2000) Low-threshold heat response antagonized by capsazepine in chick sen-sory neurons, which are capsaicin-insensitive. Eur J Neurosci12:3560–3566
9. McIntyre P, McLatchie LM, Chambers A et al. (2001) Pharmaco-logical differences between the human and rat vanilloid receptor1 (VR1). Br J Pharmacol 132:1084–1094
10. Phillips E, Reeve A, Bevan S et al. (2004) Identification ofspecies-specific determinants of the action of the antagonistcapsazepine and the agonist PPAHV on TRPV1. J Biol Chem279:17165–17172
11. Sanchez JF, Krause JE, Cortright DN (2001) The distribution andregulation of vanilloid receptor VR1 and VR1 5’ splice variantRNA expression in rat. Neuroscience 107:373–381
12. Savidge J, Davis C, Shah K et al. (2002) Cloning and func-tional characterization of the guinea pig vanilloid receptor 1.Neuropharmacology 43:450–456
13. Schumacher MA, Jong BE, FreySL et al. (2000a) The stretch-inactivated channel, a vanilloid receptor variant, is expressedin small-diameter sensory neurons in the rat. Neurosci Lett287:215–218
14. Schumacher MA, Moff I, Sudanagunta SP et al. (2000b) Molecu-lar cloning of an N-terminal splice variant of the capsaicin recep-tor. Loss of N-terminal domain suggests functional divergenceamong capsaicin receptor subtypes. J Biol Chem 275:2756–2762
15. Xue Q, Yu Y, Trilk SL et al. (2001) The genomic organization ofthe gene encoding the vanilloid receptor: evidence for multiplesplice variants. Genomics 76:14–20
TRPV1, Regulation by Acid
� TRPV1, Regulation by Protons
TRPV1, Regulation by Nerve GrowthFactor
SVEN-ERIC JORDT
Department of Pharmacology, Yale University Schoolof Medicine, New Haven, CT, [email protected]
Synonyms
TRPV1, regulation by NGF; Capsaicin receptor, regu-lation by NGF; VR1, Regulation by NGF; Vanilloid Re-ceptor, Regulation by NGF
Definition� TRPV1, the receptor for � capsaicin, the pungentingredient in chili peppers, is a polymodal receptor forphysical (heat) and chemical painful stimuli in sen-sory neurons. The neuropeptide � nerve growth factor(NGF), which is generated during injury or inflam-mation, causes � thermal hyperalgesia, an increasedsensitivity to thermal stimuli. NGF binds to its receptor,TrkA, which activates intracellular signaling pathwaysthat shift the thermal dependence of TRPV1 activationto lower temperatures.
Characteristics
Tissue damage produces a variety of mediators that ac-tivate or sensitize nociceptor terminals and elicit pain.These mediators include peptides such as bradykinin ornerve growth factor (NGF) that bind to their receptorson the nociceptor membrane and activate intracellularsignaling pathways that lead to neural activation or sen-sitization.NGF is the prototypical member of the neurotrophinfamily. During mammalian development NGF is essen-tial for the survival of sensory neurons and contributesto the maintenance of their phenotypes within the firsttwo weeks after birth. In the adult NGF is not requiredfor cell survival.The NGF peptide consists of three subunits, alpha, betaand gamma. The active neurotrophic peptide is the betasubunit that is processed by proteases and contains 118amino acids. NGF binds to a specific high-affinity recep-tor, � TrkA, on the cell membrane (Kaplan and Miller2000). TrkAbelongs to the receptor tyrosinekinase fam-ily of membrane receptors. This peptide receptor proteinfamily has > 50 members and includes the receptors forother neurotrophins and epidermal growth factor (EGF),platelet-derived growth factor (PDGF) and insulin. TheTrkA protein consists of an extracellular ligand bindingdomain, a single transmembrane domain that transmitsthe extracellular signal and an intracellular domain thatis responsible for intracellular signaling and for the for-mation of a signaling complex with other proteins.While the role of NGF in the developing nervous sys-tem is firmly established, the concept of NGF as a no-ciceptive signaling molecule is relatively new. NGF hasbeen shown to contribute to inflammationandinflamma-tory pain in numerous studies. During injury and inflam-mation the concentration of NGF in the affected tissueis increased (Constantinou et al. 1994; Donnerer et al.1992; Woolf et al. 1994). For example, inflammatory in-terleukins that are produced by macrophages and mastcells have been found to trigger the release of NGF fromkeratinocytes in the skin. NGF release is also activatedbytumornecrosisfactor (TNFalpha),apotent inflamma-tory peptide. When NGF is injected into rats the animalsdevelopreducedpawwithdrawal thresholdsformechan-ical and thermal stimuli (Lewin et al. 1993). After injec-tion of NGF, mechanical and thermal hyperalgesia de-velop with different time courses. Thermal hyperalge-sia can be observed within minutes and is thought to bemediated through a peripheral mechanism, whereas me-chanical hyperalgesia has a longer onset and is believedto be caused by � central sensitization on the level of thespinal cord (Lewin et al. 1993; Lewin et al. 1994).The capsaicin receptor, TRPV1, is an ion channel that isactivated by heat and by painful chemical stimuli suchas acid (Caterina et al. 1997). Activation of TRPV1by capsaicin does not only induce acute pain but italso causes thermal hyperalgesia. Heat hyperalgesia

T
TRPV1, Regulation by Protons 2575
is absent in mice deficient in TRPV1 (Caterina et al.2000; Davis et al. 2000). In addition, injection of NGFdoes not result in thermal hyperalgesia in these mice,indicating that TRPV1 might be the downstream targetof NGF signaling in nociceptors. Indeed, when TrkAand TRPV1 were co-expressed in a heterologous ex-pression system, activation of TrkA by NGF resulted ina pronounced sensitization of TRPV1 currents towardstemperature, with channels already active below bodytemperature (Chuang et al. 2001).Recent studies revealed the signaling mechanismsthat are involved in the sensitization of TRPV1 byNGF. The initial signaling event after NGF binding isthe dimerization of TrkA, followed by autocatalyticphosphorylation of tyrosine residues within the intra-cellular domain of the receptor (Kaplan and Miller2000). This results in the activation of phospholipaseC (PLC) that catalyses the hydrolysis of the mem-brane phospholipid phosphoinositol-4,5-bisphosphate(PIP2), yielding inositol-1,4,5-trisphosphate (IP3) anddiacylglycerol (DAG). PIP2 has been shown to inhibitTRPV1 through interaction with a C-terminal domainin the protein (Chuang et al. 2001; Prescott and Julius2003). Hydrolysis of PIP2 relieves this inhibition andleads to sensitization of TRPV1 (Chuang et al. 2001).Biochemical studies revealed that TrkA, TRPV1 andPLC proteins interact in vitro and may form a signalingcomplex. If this interaction occurs in vivo remains tobe determined (Chuang et al. 2001).In addition to PIP2 hydrolysis by PLC, TrkA activatesintracellular kinase pathways that play important rolesin the sensitization of TRPV1 in sensory neurons. Oneof the kinases involved is phosphoinositide-3-kinase(PI3K), a kinase that phosphorylates membrane phos-pholipids. PI3K contributes to TRPV1 sensitizationby reducing PIP2 levels in the membrane. In addition,PI3K activates downstream kinases such as � ERKthat may induce long-term sensitization of the neuron(Bonnington and McNaughton 2003; Zhuang et al.2004).
References1. Bonnington JK, McNaughton PA (2003) Signalling pathways
involved in the sensitisation of mouse nociceptive neurones bynerve growth factor. J Physiol 551:433–446
2. Caterina MJ, Schumacher MA, Tominaga M et al. (1997) Thecapsaicin receptor: a heat-activated ion channel in the pain path-way. Nature 389:816–824
3. Caterina MJ, Leffler A, Malmberg AB et al. (2000) Impairednociception and pain sensation in mice lacking the capsaicin re-ceptor. Science 288:306–313
4. Chuang, HH, Prescott ED, Kong H et al. (2001) Bradykininand nerve growth factor release the capsaicin receptor fromPtdIns(4,5)P2-mediated inhibition. Nature 411:957–962
5. Constantinou J, Reynolds ML, Woolf CJ et al. (1994) Nervegrowth factor levels in developing rat skin: upregulation follow-ing skin wounding. Neuroreport 5:2281–2284
6. Davis JB, Gray J, Gunthorpe MJ et al. (2000) Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature405:183–187
7. Donnerer J, Schuligoi R, Stein C (1992) Increased content andtransport of substance P and calcitonin gene-related peptide insensory nerves innervating inflamed tissue: evidence for a reg-ulatory function of nerve growth factor in vivo. Neuroscience49:693–698
8. Kaplan DR, Miller FD (2000) Neurotrophin signal transductionin the nervous system. Curr Opin Neurobiol 10:381–391
9. Lewin GR, Ritter AM, Mendell LM (1993) Nerve growth factor-induced hyperalgesia in the neonatal and adult rat. J Neurosci13:2136–2148
10. Lewin GR, Rueff A, Mendell LM (1994) Peripheral and cen-tral mechanisms of NGF-induced hyperalgesia. Eur J Neurosci6:1903–1912
11. Prescott ED, Julius D (2003) A modular PIP2 binding siteas a determinant of capsaicin receptor sensitivity. Science300:1284–1288
12. Woolf CJ, Safieh-Garabedian B, Ma QP et al. (1994) Nervegrowth factor contributes to the generation of inflammatorysensory hypersensitivity. Neuroscience 62:327–331
13. Zhuang ZY, Xu H, Clapham DE et al. (2004) Phosphatidylinositol3-kinase activates ERK in primary sensory neurons and mediatesinflammatory heat hyperalgesia through TRPV1 sensitization. JNeurosci 24:8300–8309
TRPV1, Regulation by ProtonsSVEN-ERIC JORDT
Department of Pharmacology, Yale University Schoolof Medicine, New Haven, CT, [email protected]
Synonyms
TRPV1,RegulationbyAcid;Capsaicin receptor, regula-tion by protons; VR1, Regulation by Protons; VanilloidReceptor, Regulation by Protons
Definition� TRPV1, the receptor for � capsaicin, the pungent in-
gredient in chili peppers, is a polymodal receptor forphysical (heat) and chemical painful stimuli in sen-sory neurons. Extracellular acidification, caused byinflammation or tissue injury, increases TRPV1 ac-tivation through interaction with glutamate residuesin the channel protein.
Characteristics
Tissue damage produces a variety of chemical medi-ators that activate or sensitize nociceptor terminals toelicit pain. An important component of this proalgesicresponse is local � acidosis, namely, a reduction in ex-tracellular pH to levels below the physiological norm of~7.4 (Reeh and Steen 1996). Tissue acidosis has beenobserved in many painful clinical disorders, whichinclude inflammation, skeletal muscle and cardiac� ischemia, � arthritis, hematoma and bone cancer.In chronic cough and asthma, acidification is thoughtto contribute to the induction of cough through sen-sitization of sensory neurons. Tissue acidification isperceived as painful by humans. Test subjects report asignificant correlation of the perceived intensity of pain

2576 TRPV1, Regulation by Protons
TRPV1, Regulation by Nerve Growth Factor, Figure 1 Sensitization of TRPV1 by nerve growth factor – activated signaling pathways. The capsaicinreceptor, TRPV1, is coexpressed with the receptor for nerve growth factor (NGF), TrkA, in the plasma membrane of many sensory neurons. Binding ofNGF to TrkA leads to TrkA dimerization and subsequent activation of intracellular signaling. TrkA activates phospholipase C (PLC) and phosphoinositide-3-kinase (PI3K). Both enzymes catalyze reactions that reduce the concentration of the phospholipid phosphoinositol-(4,5)-bisphosphate (PIP2) in the plasmamembrane. Whereas PLC hydrolyses PIP2, yielding inositol-triphosphate (IP3) and diacylglycerol (DAG), PI3K phosphorylates PIP2, thereby generatingPIP3. PIP2 inhibits TRPV1 currents by direct interaction with the receptor protein. PIP2 removal leads to sensitization of TRPV1, resulting in thermalhyperalgesia.
with decreasing pH when acidic solution is perfusedinto the forearm muscle (Issberner et al. 1996).Protons cause excitation or sensitization of sensoryneurons by activating ionic currents across the neu-ral membrane. Two different proton-activated inwardcationic currents have been described (Bevan and Yeats1991). One current is sustained and non-desensitizing.The other current is a transient, rapidly activating anddesensitizing current. In concert with these currents, aproton-dependent reduction in background potassiumconductances has been observed in some neurons thatmay contribute to an increase in neural excitability.Protons are capable of modulating the activity of anumber of cloned receptors and ion channels expressedby primary afferent nociceptors. Pharmacological,electrophysiological and genetic evidence suggestthat the capsaicin receptor, TRPV1, is underlying thesustained proton-activated current, whereas differentcombinations of � ASICs (acid sensitive ion channelsof the degenerin family) may give rise to transientacid-sensitive currents (Krishtal 2003). In addition,ATP-gated channels (P2X-receptors) and backgroundpotassium channels (TASK-channels) have been shownto be modulated by protons.The capsaicin receptor, TRPV1, is a nociceptor-specificcation channel that serves as the molecular target forcapsaicin, the main pungent ingredient in “hot” chilipeppers (Caterina et al. 1997). TRPV1 is a member ofthe transient receptor potential (TRP) ion channel genefamily that includes several other channels involved insensory transduction, such as � TRPM8, the receptorfor cold temperature and � menthol, and � TRPV2, achannelactivatedathightemperatures(Jordtetal.2003).
In addition to being sensitive to capsaicin and protons,TRPV1 is also activated by noxious heat and is essentialfor � bradykinin- or � nerve growth factor-dependent� thermal hyperalgesia (Caterina and Julius 2001).TRPV1 is predominantly expressed in small diameterunmyelinated neurons. In these neurons, measure-ments of capsaicin-activated currents correlate with thepresence of the sustained proton-activated current (Be-van and Geppetti 1994; Petersen and LaMotte 1993).The sustained current can be reduced by the TRPV1antagonists capsazepine (Liu and Simon 1994) andiodo-resiniferatoxin, indicating that capsaicin and pro-tons use the same target to activate neural excitation.The effectiveness of TRPV1 antagonists, especially� capsazepine, is strongly dependent on the speciesused in the individual studies. Whereas proton-inducedfiber responses in guinea pigs are strongly reduced bycapsazepine, responses in rodents are less affected.However, new high-affinity TRPV1 antagonists such asBCTC (N-(4-tertiarybutylphenyl)-4-(3-chloropyridin-2-yl)-tetrahydropyrazine-1(2H)-carboxamide) are veryeffective at blocking proton-induced fiber responses inrats at nanomolar concentrations. Capsaicin-activatedcurrents in sensory neurons show sensitivity to ex-tracellular acidification. For example, a decrease inextracellular pH from 7.3 to 6.3 leads to a seven-foldpotentiation of the capsaicin-activated current (300 nM)in dissociated rat sensory neurons (Petersen and LaM-otte 1993). Single ion channel recordings from sensoryneurons show that acidification leads to a dramaticincrease in the open probability of capsaicin-activatedchannels, whereas the single channel conductance isslightly diminished (Baumann and Martenson 2000).

T
TRPV1, Regulation by Protons 2577
TRPV1, Regulation by Protons, Figure 1 Activation and modulation or TRPV1 by protons (left): TRPV1 currents recorded from a TRPV1−expressingXenopus oocyte are shown, recorded at a membrane voltage of -40 mV. TRPV1 channels were first activated for 20 s by acidic solution (pH 4.0) atroom temperature, inducing a rapidly activating inward current. To record heat-activated currents, the bath temperature was then elevated from roomtemperature to 47˚C within 10 s (red bar). This procedure was repeated seven times in 2 min. intervals. During the fifth heat application, the bath pHwas decreased from 7.6 to 6.3. Heat activated currents are potentiated >2-fold by the drop in pH.Localization of proton-sensing residues in the TRPV1channel (right): TRPV1 has a transmembrane moiety with six putative transmembrane domains and a P-loop structure between transmembrane domains5 and 6 that contributes to the ion permeation pathway. The N- and C-termini are localized intracellularly. Two glutamate residues (E) are essential forregulation by protons (E600) and activation by protons (E648). Both residues are in or near the putative pore domain of the channel protein.
The analysis of mice with a targeted deletion in the geneencoding for TRPV1 provided further evidence for animportant role of TRPV1 in proton-dependent signalingin sensory neurons. In these mice, proton-activated sus-tained currents in DRG neurons were absent and proton-activated Ca2+-uptake was greatly diminished (Caterinaet al. 2000). In addition, recordings in the skin-nerve-preparation showed a dramatic reduction in the preva-lence of proton-sensitive C-fibers.In heterologous systems, TRPV1 channels are acti-vated by extracellular protons in the absence of otheractivating stimuli, such as capsaicin or heat. When ex-pressed in cultured mammalian cells, protons activateTRPV1 starting at pH around 6.5, with a half maximalactivation at pH 5.4 (Tominaga et al. 1998). Thesecurrents are inhibited by TRPV1 antagonists. Like thesituation in the native neuron, the degree of inhibitiondepends on the individual antagonist and differs be-tween TRPV1 species homologs. For example, protonactivated human and guinea pig TRPV1 channels aremore strongly inhibited by capsazepine than rat TRPV1channels.The potentiation of capsaicin-activated currents by pro-tons in sensory neurons can be recapitulated with clonedTRPV1 channels. More importantly, it was found thatprotons also potentiate heat-activated TRPV1 currents,effectively lowering the temperature activation thresh-old of TRPV1 channels during persistent activation andincreasing current amplitudes (Tominaga et al. 1998).This indicates that at body temperature, and especiallyin inflammatory situations with elevated concentrations
of other sensitizing factors (e.g. bradykinin or � nervegrowth factor), even moderate acidification can lead tostrong activation of TRPV1.The capsaicin receptor is a non-selective cation chan-nel with a central ion pore. The channel is formedby four homomeric protein subunits. Each monomerhas a transmembrane moiety with six predicted trans-membrane domains and extended cytosolic N-terminaland C-terminal domains. The transmembrane moietycontains a region, the P-loop between the fifth andsixth transmembrane domains, that contributes to theion conduction pathway of the central pore. Extensivesite-directed mutagenesis studies pinpointed putativeproton interaction sites to regions near or within thepore domain of the channel (Jordt et al. 2000; Welch etal. 2000). In particular, two glutamate residues, at posi-tions E600 and E648 in the rat protein, were identifiedas essential for the regulation and activation of TRPV1by protons. Negatively charged glutamate residues canserve as acceptors for protons. In addition, glutamateresidues may interact with permeating cations near orin the conduction pathway. These mutagenesis studiesalso provided further evidence that protons play a dualrole in the activation of TRPV1. Mutations introducedat position E600 dramatically shifted or eliminated thepH-dependenceofheat-andcapsaicin-activatedTRPV1currents. On the other hand, mutations at position E648led to a significant reduction in proton-activated cur-rents, whereas heat- and capsaicin-activated currentsmaintained their pH-dependence. These results indi-cated that the modulation of TRPV1 currents by protons

2578 TSK
and direct channel gating by protons can be separatedat the structural level.
References1. Baumann TK, Martenson ME (2000) Extracellular protons both
increase the activity and reduce the conductance of capsaicin-gated channels. J Neurosci 20:RC80
2. Bevan S, Geppetti P (1994) Protons: small stimulants ofcapsaicin-sensitive sensory nerves. Trends Neurosci 17:509–512
3. Bevan S, Yeats J (1991) Protons activate a cation conductance ina sub-population of rat dorsal root ganglion neurones. J Physiol(Lond) 433:145–161
4. Caterina MJ, Julius D (2001) The vanilloid receptor: a moleculargateway to the pain pathway. Annu Rev Neurosci 24:487–517
5. Caterina MJ, Schumacher MA, Tominaga M et al. (1997) Thecapsaicin receptor: a heat-activated ion channel in the pain path-way. Nature 389:816–824
6. Caterina MJ, Leffler A, Malmberg AB et al. (2000) Impairednociception and pain sensation in mice lacking the capsaicin re-ceptor. Science 288:306–313
7. Issberner U, Reeh PW, Steen KH (1996) Pain due to tissue acido-sis: a mechanism for inflammatory and ischemic myalgia? Neu-rosci Lett 208:191–194
8. Jordt SE, Tominaga M, Julius D (2000) Acid potentiation of thecapsaicin receptor determined by a key extracellular site. ProcNatl Acad Sci USA 97:8134–8139
9. Jordt SE, McKemy DD, Julius D (2003) Lessons from peppersand peppermint: the molecular logic of thermosensation. CurrOpin Neurobiol 13:487–492
10. Krishtal O (2003) The ASICs: signaling molecules? Modulators?Trends Neurosci 26:477–483
11. Liu L, Simon SA (1994) A rapid capsaicin-activated currentin rat trigeminal ganglion neurons. Proc Natl Acad Sci USA91:738–741
12. Petersen M, LaMotte RH (1993) Effect of protons on the inwardcurrent evoked by capsaicin in isolated dorsal root ganglion cells.Pain 54:37–42
13. Reeh PW, Steen KH (1996) Tissue acidosis in nociception andpain. Prog Brain Res 113:143–151
14. Tominaga M, Caterina MJ, Malmberg AB et al. (1998) The clonedcapsaicin receptor integrates multiple pain-producing stimuli.Neuron 21:531–543
15. Welch JM, Simon SA, Reinhart PH (2000) The activation mech-anism of rat vanilloid receptor 1 by capsaicin involves the poredomain and differs from the activation by either acid or heat.Proc Natl Acad Sci USA 97:13889–13894
TSK
� Tampa Scale for Kinesiophobia
TTX
� Tetrodotoxin
Tuberculoid Leprosy
Definition
The relatively benign and least infectious type of lep-rosy, which is characterized by early severe damage to
the nerves and by the presence of one to a few sharplydefined, anesthetic, hypopigmented skin lesions.� Hansen’s Disease
Tubo-Ovarian Complex
� Chronic Pelvic Pain, Pelvic Inflammatory Diseaseand Adhesions
Tumor (National Cancer InstituteTerminology – Neoplasm)
Definition
An abnormalmassof tissue that resultswhencellsdividemore than they should or do not die when they should.Tumors may be benign (not cancerous), or malignant(cancerous).� Cancer Pain Management, Treatment of Neuropathic
Components
Tumor Necrosis Factor Alpha(α)
Synonyms
TNF Alpha(α)
Definition
TNFα is a pro-inflammatory cytokine and member ofthe ’TNF-superfamily’, with algesic actions. It partici-pates in inflammation, wound healing and remodelingof tissue.� Cytokines as Targets in the Treatment of Neuropathic
Pain� Inflammatory Neuritis� Wallerian Degeneration
Twin Studies
Definition
The comparison of traits among pairs of monozygotic(MZ; identical) versus dizygotic (DZ; fraternal) twins.MZ twins are clones, sharing 100% of their DNA se-quence,whereasDZtwinsshareonly 50%(nomore thanany other siblings). If a painful pathology occurs moreoften in both individuals in a MZ twin pair than a DZtwin pair, it can be said to be heritable. A caveat is thatthis analysis is dependent on the assumption of equalenvironments of MZ and DZ twins, which may not berealistic.� Heritability of Inflammatory Nociception� Heritable

T
Tyrosine Kinase A 2579
Twitch-Obtaining IntramuscularStimulation
Definition
This is a technique of dry needling developed by Chu,using an EMG needle to perform dry needling.� Dry Needling
Two Pore Domain K+ Channels
Definition
A large, structurally related, class of K+ channels thatprovide a large fraction of the background conductancein many neurons. They are modulated by a variety ofphysical (temperature, mechanical) and chemical stim-uli (pH, anesthetics, lipids).� Nociceptors, Cold Thermotransduction
Two-Way Scaling Models
Definition
Two-way scaling models yield only the group stimulusspace; individual differences are lost.� Multidimensional Scaling and Cluster Analysis Ap-
plication for Assessment of Pain
Type-1 Reaction (Leprosy)
Synonyms
Reversal reaction
Definition
A leprosy reaction usually occurring during chemother-apy in borderline leprosy, representing a delayed hyper-sensitivity reaction with upgrading of cell-mediated im-munity to Mycobacterium leprae. It is characterized byerythema, edema, and tenderness of preexisting skin le-sions, neuritis with nerve damage, and fever.� Hansen’s Disease
Type-2 Reaction (Leprosy)
Synonyms
Erythema Nodosum Leprosum
Definition
A leprosy reaction resembling an Arthus reaction andrepresenting humoral hypersensitivity. It is due to anantigen-antibody reaction with the formationof immunecomplexes at the site of antigen deposits in various tis-sues and gives rise to acute inflammatory foci. It ischaracterized by painful red nodules or plaques, anddistributes especially on the face and limbs. It may beassociated with severe systemic and visceral symptoms.� Hansen’s Disease
Type-II Receptors
Definition
Type II receptors are G-protein coupled receptors with7 transmembrane domains.� Opioid Modulation of Nociceptive Afferents In Vivo
Tyrosine Kinase A
Synonyms
trkA
Definition
The biological actions of neurotrophins are mediated byspecific neurotrophin receptor tyrosine kinases (trks).The proto-oncogene trkA is now recognized as theprimary high affinity receptor for nerve growth fac-tor (NGF), while two related tyrosine kinase receptors,trkB and trkC, mediate the biological functions of brain-derived neurotrophic factor (BDNF)andneurotrophin-3(NT-3), respectively.� ERK Regulation in Sensory Neurons during Inflam-
mation

Copyright © 2022 FDOKUMEN




















