
Interactions of Serotoninergic, Cholinergic, and Tachykinin-Containing Nerve Elements in the Rabbit...
11
THE ANATOMICAL RECORD 292:1548–1558 (2009) Interactions of Serotoninergic, Cholinergic, and Tachykinin-Containing Nerve Elements in the Rabbit Small Intestine VIKTORIA DE ´ NES, 1 * MARTA WILHELM, 2 AGNES NE ´ METH, 1 AND ROBERT GA ´ BRIEL 1 1 Department of Experimental Zoology and Neurobiology, University of Pe ´cs, Pe ´cs, Hungary 2 Physical Education and Sport Sciences, University of Pe ´cs, Pe ´cs, Hungary ABSTRACT This report presents novel results on the effects of serotonin (5-HT) on longitudinal muscle contractions in the rabbit ileum and the interac- tions of serotonin with some neuronal elements of the myenteric plexus. We showed previously that serotonin-triggered contractions involved two mechanisms in the rabbit ileum: neuronal excitation (via 5-HT 2 receptors in the neurons) and direct muscular stimulation (via 5-HT 4 receptors in the muscle). Here, we focus on the neuronal 5-HT 2 receptor pathway and report further pharmacological and immunocytochemical data clarifying the details of the mechanisms. We observed that antagonists for neuroki- nin (NK1 and NK2) receptors partially blocked the serotonin response, but NK3 receptor antagonists had no effect. Pretreatment by atropine (ATR) eliminated the NK1 receptor antagonist resistant contractions. In contrast, the NK1 antagonist did not depress the ATR-resistant contrac- tion when ATR was added first. 5-HT 2 receptor agonist-induced contrac- tions were partially suppressed by ATR, hexamethonium, and NK1 or NK2 receptor antagonists. In conclusion, serotonin acting through 5-HT 2 receptors could stimulate interneurons and excitatory motor neurons. Immunocytochemical staining revealed an extensive tachykinin-immuno- reactive (IR) network in the myenteric plexus. Approximately 52% of all myenteric neurons were labeled. 5-HT-IR fibers could be detected around both choline acetyltransferase- and tachykinin-IR cells, suggesting func- tional relationships between them. Consistent with our pharmacological observations, we found that immunopositive nerve elements for 5-HT 2A receptor and double-labeled immunostaining revealed a remarkable over- lap between tachykinin-IR neurons and 5-HT 2A -IR elements. Anat Rec, 292:1548–1558, 2009. V V C 2009 Wiley-Liss, Inc. Key words: ileum; 5-HT; tachykinins; 5-HT 2 receptors; rabbit One of the principal regulators of digestion in mam- mals is the enteric nervous system. Enteric neurons coordinate a variety of functions, such as the secretion of digestive enzymes, control of absorption, and the regula- tion of motility. Intestinal smooth muscle contractility is organized by polarized reflex pathways through both excitatory and inhibitory neurons (Goyal and Hirano, 1996; Kunze and Furness, 1999; Olsson and Holmgren, *Correspondence to: Vikto ´ria De ´nes, Department of Experi- mental Zoology and Neurobiology, University of Pe ´cs, H-7601 Pe ´cs, Ifju ´ sa ´g u. 6, Hungary. Fax: 36-72-501-517. E-mail: [email protected] Received 20 March 2008; Accepted 20 May 2009 DOI 10.1002/ar.20956 Published online 14 August 2009 in Wiley InterScience (www. interscience.wiley.com). V V C 2009 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Interactions of Serotoninergic, Cholinergic, and Tachykinin-Containing Nerve Elements in the Rabbit...

THE ANATOMICAL RECORD 292:1548–1558 (2009)
Interactions of Serotoninergic,Cholinergic, and Tachykinin-Containing
Nerve Elements in the Rabbit SmallIntestine
VIKTORIA DENES,1* MARTA WILHELM,2 AGNES NEMETH,1
AND ROBERT GABRIEL1
1Department of Experimental Zoology and Neurobiology, University of Pecs,Pecs, Hungary
2Physical Education and Sport Sciences, University of Pecs, Pecs, Hungary
ABSTRACTThis report presents novel results on the effects of serotonin (5-HT)
on longitudinal muscle contractions in the rabbit ileum and the interac-tions of serotonin with some neuronal elements of the myenteric plexus.We showed previously that serotonin-triggered contractions involved twomechanisms in the rabbit ileum: neuronal excitation (via 5-HT2 receptorsin the neurons) and direct muscular stimulation (via 5-HT4 receptors inthe muscle). Here, we focus on the neuronal 5-HT2 receptor pathway andreport further pharmacological and immunocytochemical data clarifyingthe details of the mechanisms. We observed that antagonists for neuroki-nin (NK1 and NK2) receptors partially blocked the serotonin response,but NK3 receptor antagonists had no effect. Pretreatment by atropine(ATR) eliminated the NK1 receptor antagonist resistant contractions. Incontrast, the NK1 antagonist did not depress the ATR-resistant contrac-tion when ATR was added first. 5-HT2 receptor agonist-induced contrac-tions were partially suppressed by ATR, hexamethonium, and NK1 orNK2 receptor antagonists. In conclusion, serotonin acting through 5-HT2
receptors could stimulate interneurons and excitatory motor neurons.Immunocytochemical staining revealed an extensive tachykinin-immuno-reactive (IR) network in the myenteric plexus. Approximately 52% of allmyenteric neurons were labeled. 5-HT-IR fibers could be detected aroundboth choline acetyltransferase- and tachykinin-IR cells, suggesting func-tional relationships between them. Consistent with our pharmacologicalobservations, we found that immunopositive nerve elements for 5-HT2A
receptor and double-labeled immunostaining revealed a remarkable over-lap between tachykinin-IR neurons and 5-HT2A-IR elements. Anat Rec,292:1548–1558, 2009. VVC 2009 Wiley-Liss, Inc.
Keywords: ileum; 5-HT; tachykinins; 5-HT2 receptors; rabbit
One of the principal regulators of digestion in mam-mals is the enteric nervous system. Enteric neuronscoordinate a variety of functions, such as the secretion ofdigestive enzymes, control of absorption, and the regula-
tion of motility. Intestinal smooth muscle contractility isorganized by polarized reflex pathways through bothexcitatory and inhibitory neurons (Goyal and Hirano,1996; Kunze and Furness, 1999; Olsson and Holmgren,
*Correspondence to: Viktoria Denes, Department of Experi-mental Zoology and Neurobiology, University of Pecs, H-7601Pecs, Ifjusag u. 6, Hungary. Fax: 36-72-501-517. E-mail:[email protected]
Received 20 March 2008; Accepted 20 May 2009
DOI 10.1002/ar.20956Published online 14 August 2009 in Wiley InterScience (www.interscience.wiley.com).
VVC 2009 WILEY-LISS, INC.

2001). Excitatory effects are mediated by serotonin (5-HT), tachykinins, acetylcholine (ACh), gastrin-releasingpeptide/bombesin, and so forth, whereas the transmit-ters that inhibit gut motility are the members of thevasoactive intestinal peptide-family, nitric oxide, c-ami-nobutyric acid, and ATP (Shuttleworth and Keef, 1995;Kunze and Furness, 1999).
5-HT has been identified as both a local hormone anda neurotransmitter that is synthesized and stored in ro-bust amounts in the gastrointestinal tract (Erspamerand Asero, 1952; Gershon et al., 1965). Numerous phar-macological studies have provided evidence that 5-HTcan act on the secretion of intestinal fluid and electro-lytes (Bjorck et al., 1988) and gastrointestinal motility(Grider et al., 1996; Wade et al., 1996; Bush et al.,2001). In guinea pig, morphological studies described arich, varicose 5-HT-immunoreactive (IR) fiber network inthe myenteric and submucous plexuses and reactivenerve cells with short, broad dendrites and a single longprocess in the myenteric plexus (Costa et al., 1982). Fur-thermore, 5-HT-IR nerve elements were also found inthe large and small intestines of many other mammals(Timmermans et al., 1990; Sang and Young, 1996; Fuji-miya et al., 1997; Toole et al., 1998). In rabbit, however,5-HT could be detected only after preloading in themyenteric plexus. Nevertheless, nerve fibers that couldaccumulate 5-HT were IR for tryptophan hydroxylase,the enzyme that synthesizes 5-HT (Denes et al., 2003).Application of exogenous 5-HT caused a concentration-dependent increase of contractions in rabbit ileum. Theresults indicated that 5-HT could act directly on smoothmuscle through 5-HT4 and on neurons through 5-HT2
receptors (Denes et al., 2003).Tachykinins are another type of widely distributed
neuroactive mediators in the mammalian ENS. Tachyki-nin family includes substance P (SP), physalaemin-likepeptide, neurokinin A (NKA), and neurokinin B (NKB)(Lazarus and Di Augustine, 1980; Kangawa et al., 1983;Nawa et al., 1984). Immunocytochemical studiesdescribed SP-IR nerve elements in all parts and layersof the gastrointestinal system (Heitz et al., 1976; Costaet al., 1980, 1981; Schultzberg et al., 1980). Applicationof tachykinins to the intestine induces various effectsincluding excitation of muscle, increase in blood flowand secretion (Holzer and Holzer-Petsche, 1997). Thediverse effects of these peptides are mediated throughthree receptors called NK1, NK2, and NK3 (Guard andWatson, 1991; Grady et al., 1996).
In rabbit, we have detailed knowledge about mecha-nisms how 5-HT regulates nutrient absorption, electro-lyte transport, or chloride secretion (Donowitz et al.,1980; Sundaram et al., 1991; Salvador et al., 1997,2000). However, regarding the effect of 5-HT on intesti-nal motility in rabbit, only a few articles have been pub-lished. According to Salvador et al. (2000), 5-HTincreases contractions and tone of muscle fibers in the
duodenum, jejunum, and ileum. Based on our previouslypublished data, this study was undertaken to ascertainwhether (i) noncholinergic neuronal components takepart in 5-HT-evoked contractions and (ii) how 5-HT2
receptors are situated within the neuronal circuitry. Inaddition, (iii) we provide data regarding the morphologyand distribution of the tachykinin-containing elementsin the rabbit ileal myenteric plexus.
MATERIALS AND METHODS
Adult white rabbits of both sexes (2–2.5 kg) were usedas experimental animals. They were fed and wateredad libitum. Rabbits were killed by a single strong blowto the cervical vertebrae and pithed.
The last 10 cm of small intestine just before the ileoce-cal junction was discarded, and the rest of the ileum wasused for experiments.
Immunocytochemistry
To detect fibers taking up 5-HT, pieces of gut wereincubated in Krebs-Henseleit solution containing 10�5 M5-HT at room temperature for 25 min. To fix tissues, theileum was filled with KREBS solution (mM: NaCl 119,NaHCO3 25, KH2PO4 1.2, CaCl2 2.5, MgSO4 1.5, KCl2.5, and glucose 11) and placed into 4% paraformalde-hyde. For 5-HT and SP immunostaining, the tissueswere fixed for 6–7 hr at room temperature. For cholineacetyltransferase (ChAT) immunostaining, tissues werefixed in modified Zamboni’s fixative (2% paraformalde-hyde with saturated picric acid) overnight at 4�C. Tis-sues were washed for 6 � 10 min in phosphate buffersaline (PBS) and longitudinal muscle layer-myentericplexus whole mount preparations were made. After pre-treatment with 0.3% Triton X-100 solution (6 � 10 min)and preincubation in blocking solution (1% bovine serumalbumin, 0.1% Triton-X 100 in PBS) for 20 min, wholemount preparations were incubated in primary antiseraovernight. Primary antibodies used for immunolabelingare listed in Table 1. Whole mount preparations werewashed in PBS and placed in secondary antisera labeledwith fluorophore for 4–5 hr. The secondary antisera usedare listed in Table 2. After removing the secondary anti-sera by washing in PBS for 6 � 10 min, whole mountpreparations were mounted in Vectashield (Vector, Bur-lingame, CA). Immunostained tissues were viewed underOlympus BX 51 epifluorescence microscope connected toa digital camera (Olympus DP50), and digital photoswere recorded with the help of an image analyzer pro-gram (AnalySIS). The digital images were further proc-essed (background and contrast adjustments,assembling, and labeling of photographic tables) withthe Adobe Photoshop 7.0 program. To check the specific-ity of both primary and secondary antibodies, three con-trols were made for the immunocytochemical labeling.
TABLE 1. Primary antibodies used for immunocytochemistry
Primary antibody Host Dilution Supplier
anti-5-HT Rabbit 1:1,000 Sigma-Aldrich, St. Louis, MOanti-SP/NKA/NKB Rat 1:1,000 BD PharMingen, Heidelberg, Germanyanti-ChAT Sheep 1:1,000 Chemicon, Temecula, CAanti-5-HT2A receptor Goat 1:500 Santa Cruz Biotechnology, Heidelberg, Germany
5-HT AND TACHYKININ IN THE RABBIT ILEUM 1549

Omission of primary antibodies in the immunolabelingprocedures resulted no staining. Cross-reactivity of thenoncorresponding primary and secondary antibodies wasalso checked and ruled out. To check the specificity ofanti-5-HT2A receptor antibody, whole mount prepara-tions were prepared as described earlier. However, theywere incubated in the mixture of primary antisera (0.8lg) and blocking peptide (8 lg). As a result of presenceof blocking peptide, immunostaining could not bedetected.
To quantify the colocalization between tachykininer-gic/ChAT- and tachykininergic/5-HT2A-immunopositiveelements, three ileal flat mounts were drawn from threeanimals. Cell bodies (n ¼ 100/animal) were examinedwhether they were IR for either one of the markers orboth. The counting of cell bodies was performed directlythrough the microscope.
Counting and plotting of SP-IR neurons were per-formed on whole mounts using semiautomatic computer-assisted image analysis software (Neurolucida).
In Vitro Pharmacological Experiments
Pieces of ileum were cut and the longitudinal musclelayer was removed as described earlier by Paton andVizi (1969). The muscle strips were placed in organbaths filled with Krebs-Henseleit solution, which wasoxygenated with mixture of 5% CO2 and 95% O2 andkept at 37�C.
Drugs and their concentrations used in experimentsare listed in Table 3. They were applied directly into theorgan bath and were washed out as soon as the peakcontractions were reached. At the end of the measure-ments, 80 mM KCl was added into the baths to evoke
the maximal contraction of muscle strips. Contractionsof longitudinal muscle were measured by force-displace-ment transducers and were registered by ISOSYS pro-gram (Experimetria, Budapest, Hungary).
Data Processing
Data are presented as means � SEM (n ¼ 6 or 7), sta-tistical comparisons were made by using Wilcoxon’ssigned-rank test (two samples) and Quade-test for sev-eral related samples. A difference of P < 0.05 was con-sidered statistically significant. Any group of resultscontains at most two data from the same animal. Eachgroup of pretreated preparations was compared to a con-trol group of muscle strips taken from the same animals.
RESULTSInvolvement of Intrinsic Excitatory Pathwaysin 5-HT-Induced Contractions
Our previous data have shown that blockade of musca-rinic receptors (mAChR) does not fully abolish the 5-HT-induced contractions. At the same time, coadministra-tion of atropine (ATR) and tetrodotoxin (TTX) caused afurther decrease in the response of longitudinal muscle(Denes et al., 2003); thus, it is likely that noncholinergicnerve element(s) participate in mediating the effect of 5-HT. To test this hypothesis, we applied an NK1 receptorantagonist (N-acetyl-L-tryptophan-3,5(trifluoromethyl)-benzyl ester, 10�7 M), which decreased the 5-HT-induced(10�6 M) contractions significantly (Fig. 1a). Coapplica-tion of NK1 receptor antagonist and ATR (10�6 M) alsosignificantly reduced the contractions. Moreover, com-bined administration of NK1 receptor antagonist, ATR
TABLE 2. Secondary antibodies used for immunocytochemistry
Secondary antibody Host Dilution Supplier
Texas red-conjugated anti-rabbit IgG Donkey 1:200 Jackson, West Grove, PAFITC-conjugated anti-rabbit IgG Donkey 1:200 Jackson, West Grove, PAFITC-conjugated anti-rat IgG Donkey 1:200 Jackson, West Grove, PATexas red-conjugated anti-rat IgG Donkey 1:200 Jackson, West Grove, PATexas red-conjugated anti-sheep IgG Donkey 1:200 Jackson, West Grove, PAFITC-conjugated anti-goat IgG Rabbit 1:400 Sigma-Aldrich, St. Louis, MO
TABLE 3. Drugs used in pharmacological experiments
Drugs Target Concentration (M) Supplier
Serotonin Serotonin receptors 10�6 Sigma-Aldrich, St. Louis, MOTetrodotoxin Selective blocker of
voltage-gated Naþ channel5 � 10�8 Sigma-Aldrich, St. Louis, MO
Hexametonium Nicotinic ACh receptor(antagonist)
10�6 Sigma-Aldrich, St. Louis, MO
Atropine Muscarinic ACh receptor(antagonist)
10�6 Sigma-Aldrich, St. Louis, MO
a-Methyl-5-hydroxytryptamine 5-HT2 receptor (agonist) 10�6 Sigma-Aldrich, St. Louis, MOKetanserin 5-HT2A receptor (antagonist) 10�6 Sigma-Aldrich, St. Louis, MON-Acetyl-L-tryptophan-3,5(trifluoromethyl)benzyl ester
NK1 receptor (antagonist) 10�7 Sigma-Aldrich, St. Louis, MO
SR 48968 NK2 receptor (antagonist) 5 � 10�7 Sanofi-Synthelabo,Montpellier, France
SR 142801 NK3 receptor (antagonist) 2 � 10�7 Sanofi-Synthelabo,Montpellier, France
1550 DENES ET AL.
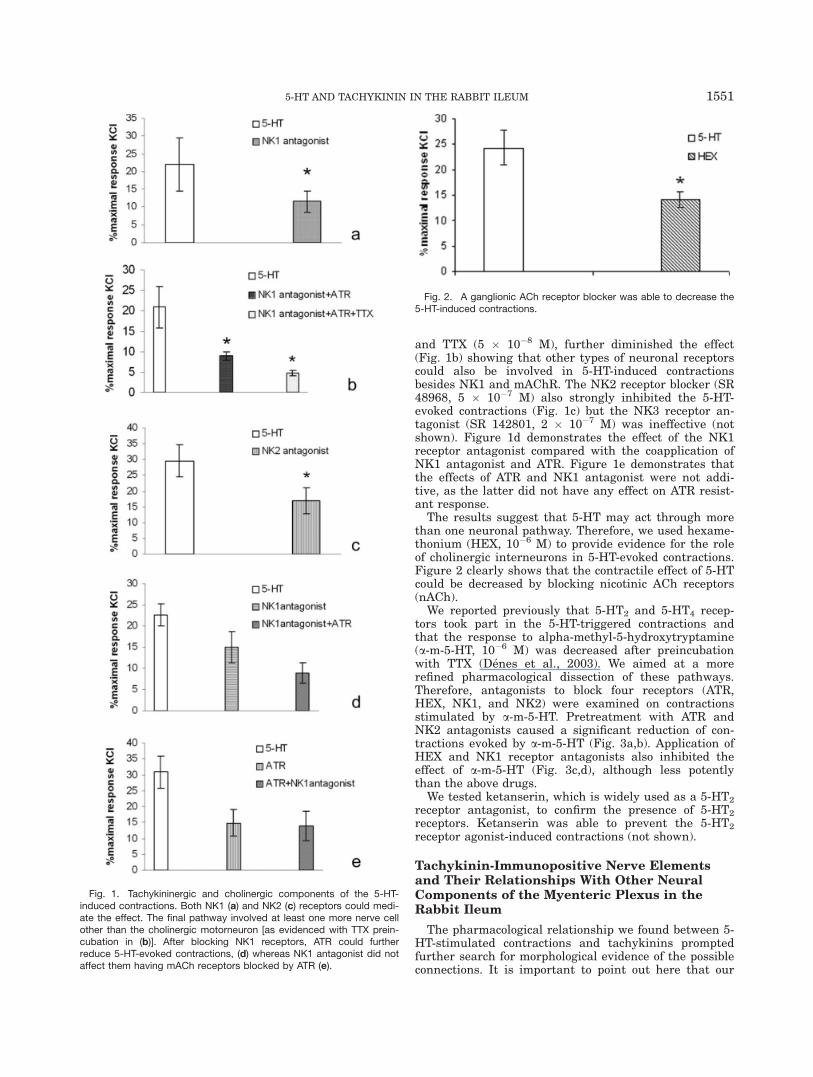
and TTX (5 � 10�8 M), further diminished the effect(Fig. 1b) showing that other types of neuronal receptorscould also be involved in 5-HT-induced contractionsbesides NK1 and mAChR. The NK2 receptor blocker (SR48968, 5 � 10�7 M) also strongly inhibited the 5-HT-evoked contractions (Fig. 1c) but the NK3 receptor an-tagonist (SR 142801, 2 � 10�7 M) was ineffective (notshown). Figure 1d demonstrates the effect of the NK1receptor antagonist compared with the coapplication ofNK1 antagonist and ATR. Figure 1e demonstrates thatthe effects of ATR and NK1 antagonist were not addi-tive, as the latter did not have any effect on ATR resist-ant response.
The results suggest that 5-HT may act through morethan one neuronal pathway. Therefore, we used hexame-thonium (HEX, 10�6 M) to provide evidence for the roleof cholinergic interneurons in 5-HT-evoked contractions.Figure 2 clearly shows that the contractile effect of 5-HTcould be decreased by blocking nicotinic ACh receptors(nACh).
We reported previously that 5-HT2 and 5-HT4 recep-tors took part in the 5-HT-triggered contractions andthat the response to alpha-methyl-5-hydroxytryptamine(a-m-5-HT, 10�6 M) was decreased after preincubationwith TTX (Denes et al., 2003). We aimed at a morerefined pharmacological dissection of these pathways.Therefore, antagonists to block four receptors (ATR,HEX, NK1, and NK2) were examined on contractionsstimulated by a-m-5-HT. Pretreatment with ATR andNK2 antagonists caused a significant reduction of con-tractions evoked by a-m-5-HT (Fig. 3a,b). Application ofHEX and NK1 receptor antagonists also inhibited theeffect of a-m-5-HT (Fig. 3c,d), although less potentlythan the above drugs.
We tested ketanserin, which is widely used as a 5-HT2
receptor antagonist, to confirm the presence of 5-HT2
receptors. Ketanserin was able to prevent the 5-HT2
receptor agonist-induced contractions (not shown).
Tachykinin-Immunopositive Nerve Elementsand Their Relationships With Other NeuralComponents of the Myenteric Plexus in theRabbit Ileum
The pharmacological relationship we found between 5-HT-stimulated contractions and tachykinins promptedfurther search for morphological evidence of the possibleconnections. It is important to point out here that our
Fig. 1. Tachykininergic and cholinergic components of the 5-HT-induced contractions. Both NK1 (a) and NK2 (c) receptors could medi-ate the effect. The final pathway involved at least one more nerve cellother than the cholinergic motorneuron [as evidenced with TTX prein-cubation in (b)]. After blocking NK1 receptors, ATR could furtherreduce 5-HT-evoked contractions, (d) whereas NK1 antagonist did notaffect them having mACh receptors blocked by ATR (e).
Fig. 2. A ganglionic ACh receptor blocker was able to decrease the5-HT-induced contractions.
5-HT AND TACHYKININ IN THE RABBIT ILEUM 1551
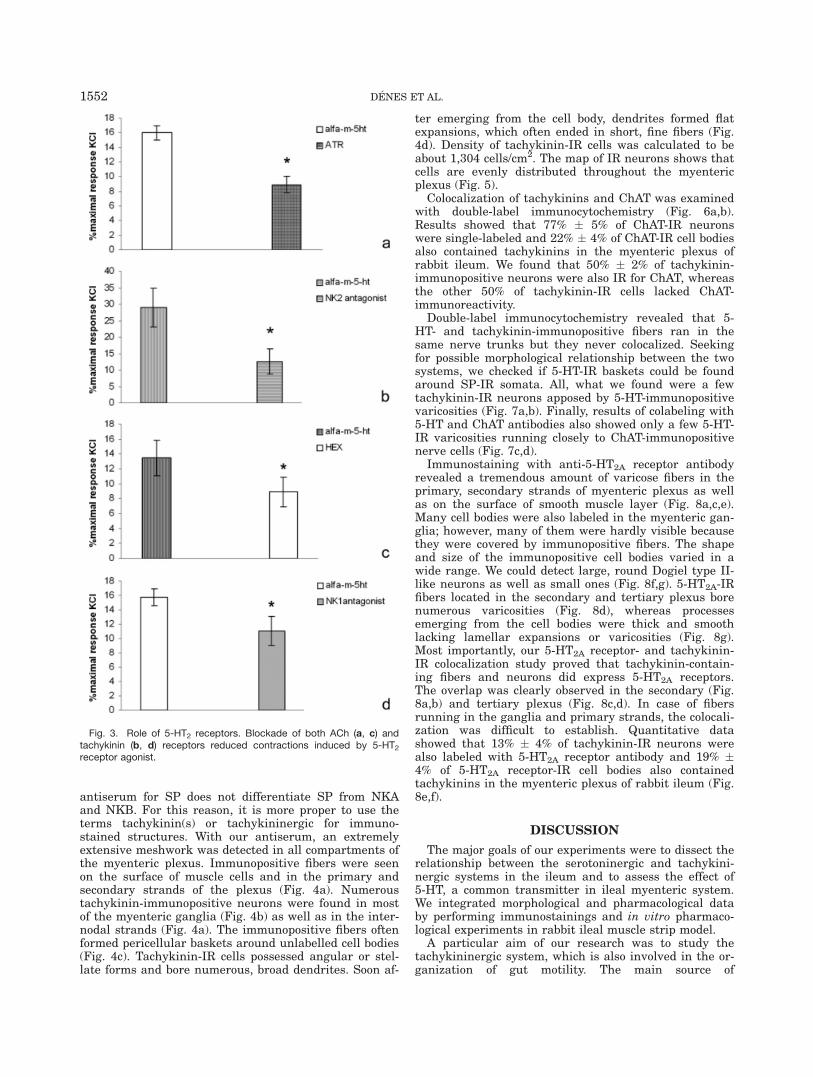
antiserum for SP does not differentiate SP from NKAand NKB. For this reason, it is more proper to use theterms tachykinin(s) or tachykininergic for immuno-stained structures. With our antiserum, an extremelyextensive meshwork was detected in all compartments ofthe myenteric plexus. Immunopositive fibers were seenon the surface of muscle cells and in the primary andsecondary strands of the plexus (Fig. 4a). Numeroustachykinin-immunopositive neurons were found in mostof the myenteric ganglia (Fig. 4b) as well as in the inter-nodal strands (Fig. 4a). The immunopositive fibers oftenformed pericellular baskets around unlabelled cell bodies(Fig. 4c). Tachykinin-IR cells possessed angular or stel-late forms and bore numerous, broad dendrites. Soon af-
ter emerging from the cell body, dendrites formed flatexpansions, which often ended in short, fine fibers (Fig.4d). Density of tachykinin-IR cells was calculated to beabout 1,304 cells/cm2. The map of IR neurons shows thatcells are evenly distributed throughout the myentericplexus (Fig. 5).
Colocalization of tachykinins and ChAT was examinedwith double-label immunocytochemistry (Fig. 6a,b).Results showed that 77% � 5% of ChAT-IR neuronswere single-labeled and 22% � 4% of ChAT-IR cell bodiesalso contained tachykinins in the myenteric plexus ofrabbit ileum. We found that 50% � 2% of tachykinin-immunopositive neurons were also IR for ChAT, whereasthe other 50% of tachykinin-IR cells lacked ChAT-immunoreactivity.
Double-label immunocytochemistry revealed that 5-HT- and tachykinin-immunopositive fibers ran in thesame nerve trunks but they never colocalized. Seekingfor possible morphological relationship between the twosystems, we checked if 5-HT-IR baskets could be foundaround SP-IR somata. All, what we found were a fewtachykinin-IR neurons apposed by 5-HT-immunopositivevaricosities (Fig. 7a,b). Finally, results of colabeling with5-HT and ChAT antibodies also showed only a few 5-HT-IR varicosities running closely to ChAT-immunopositivenerve cells (Fig. 7c,d).
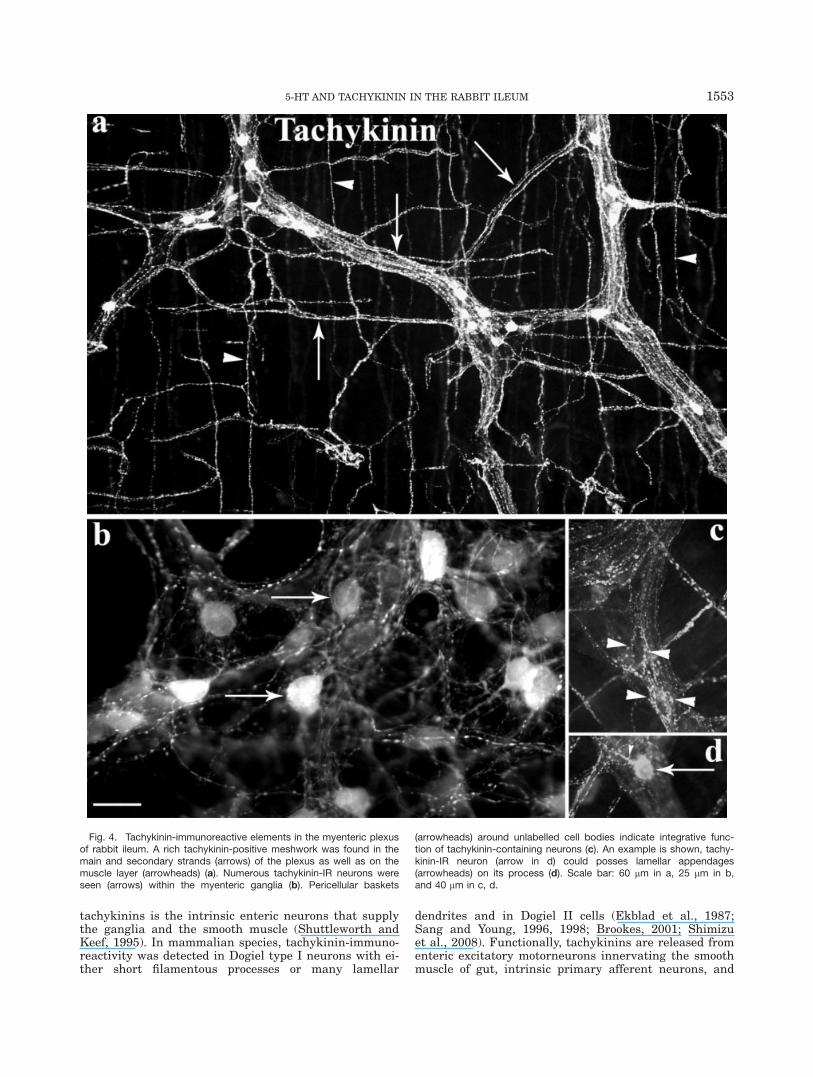
Immunostaining with anti-5-HT2A receptor antibodyrevealed a tremendous amount of varicose fibers in theprimary, secondary strands of myenteric plexus as wellas on the surface of smooth muscle layer (Fig. 8a,c,e).Many cell bodies were also labeled in the myenteric gan-glia; however, many of them were hardly visible becausethey were covered by immunopositive fibers. The shapeand size of the immunopositive cell bodies varied in awide range. We could detect large, round Dogiel type II-like neurons as well as small ones (Fig. 8f,g). 5-HT2A-IRfibers located in the secondary and tertiary plexus borenumerous varicosities (Fig. 8d), whereas processesemerging from the cell bodies were thick and smoothlacking lamellar expansions or varicosities (Fig. 8g).Most importantly, our 5-HT2A receptor- and tachykinin-IR colocalization study proved that tachykinin-contain-ing fibers and neurons did express 5-HT2A receptors.The overlap was clearly observed in the secondary (Fig.8a,b) and tertiary plexus (Fig. 8c,d). In case of fibersrunning in the ganglia and primary strands, the colocali-zation was difficult to establish. Quantitative datashowed that 13% � 4% of tachykinin-IR neurons werealso labeled with 5-HT2A receptor antibody and 19% �4% of 5-HT2A receptor-IR cell bodies also containedtachykinins in the myenteric plexus of rabbit ileum (Fig.8e,f).
DISCUSSION
The major goals of our experiments were to dissect therelationship between the serotoninergic and tachykini-nergic systems in the ileum and to assess the effect of5-HT, a common transmitter in ileal myenteric system.We integrated morphological and pharmacological databy performing immunostainings and in vitro pharmaco-logical experiments in rabbit ileal muscle strip model.
A particular aim of our research was to study thetachykininergic system, which is also involved in the or-ganization of gut motility. The main source of
Fig. 3. Role of 5-HT2 receptors. Blockade of both ACh (a, c) andtachykinin (b, d) receptors reduced contractions induced by 5-HT2receptor agonist.
1552 DENES ET AL.
tachykinins is the intrinsic enteric neurons that supplythe ganglia and the smooth muscle (Shuttleworth andKeef, 1995). In mammalian species, tachykinin-immuno-reactivity was detected in Dogiel type I neurons with ei-ther short filamentous processes or many lamellar
dendrites and in Dogiel II cells (Ekblad et al., 1987;Sang and Young, 1996, 1998; Brookes, 2001; Shimizuet al., 2008). Functionally, tachykinins are released fromenteric excitatory motorneurons innervating the smoothmuscle of gut, intrinsic primary afferent neurons, and
Fig. 4. Tachykinin-immunoreactive elements in the myenteric plexusof rabbit ileum. A rich tachykinin-positive meshwork was found in themain and secondary strands (arrows) of the plexus as well as on themuscle layer (arrowheads) (a). Numerous tachykinin-IR neurons wereseen (arrows) within the myenteric ganglia (b). Pericellular baskets
(arrowheads) around unlabelled cell bodies indicate integrative func-tion of tachykinin-containing neurons (c). An example is shown, tachy-kinin-IR neuron (arrow in d) could posses lamellar appendages(arrowheads) on its process (d). Scale bar: 60 lm in a, 25 lm in b,and 40 lm in c, d.
5-HT AND TACHYKININ IN THE RABBIT ILEUM 1553
last but not least, orally and anally directed interneur-ons (Ekblad et al., 1987; Brookes et al., 1991, 1992,1997; Johnson et al., 1996, 1998). Tachykinins utilizedas transmitters from enteric neurons target numerousenteric elements through NK1, NK2, or NK3 receptors.NK2 receptors occur primarily on muscle, whereas NK3receptors are involved in neuroneuronal transmission(Johnson et al., 1996, 1998; Portbury et al., 1996a; Jen-kinson et al., 1999, 2000; Lecci et al., 2002). NK1 recep-tors have been shown acting on both muscle and entericneurons. More precisely, functional classes of neuronsthat expressed NK1 receptors were NOS-IR inhibitorymotorneurons, ACh/SP-IR excitatory motorneurons,secretomotor neurons, and ChAT/calbindin-IR sensoryneurons (Portbury et al., 1996b; Johnson et al., 1998;Lomax et al., 1998; Lecci et al., 2002; Thornton andBornstein, 2002).
Data that indicate functional relationship between5-HT and tachykininergic system are rare and to datenone has been obtained in rabbit. Work of Bucheit et al.(1985) on guinea pig ileum provided data that SP antag-onist reduced the effect of 5-HT; however, they did not
specify which tachykinin receptor was involved. Also,Briejer and Schuurkes (1996) found that 5-HT due to5-HT3 and 5-HT4 receptors had stimulatory effects inthe guinea pig colon and this action was mediatedthrough activation of the cholinergic and tachykininergicneurons.
A Possible Enteric Circuit UnderlyingSerotonin–Tachykinin Interaction inRabbit Ileum
In our previous article, data had been published aboutthe serotoninergic system in rabbit (Denes et al., 2003).Briefly, the 5-HT synthesizing enzyme could be detectedin the myenteric plexus but before revealing a rich vari-cose 5-HT-IR fiber meshwork, we had to incubate the il-eum with 5-HT. After preincubation with 5-HT, anextensive fiber meshwork was seen in the myentericplexus forming numerous pericellular baskets as well asrunning to the muscle layer. No 5-HT-IR neurons wereobserved in the rabbit myenteric plexus; however, 5-HTstimulated movements of the longitudinal muscle layerby direct and indirect ways in the rabbit ileum. Theindirect effect of 5-HT was partially elicited through cho-linergic pathway via 5-HT2 receptors, whereas the directaction of 5-HT was mediated by 5-HT4 receptors (Denes
Fig. 7. Double labeling of 5-HT/tachykinin and 5-HT/ACh. Oneexample of those few cases when fibers with strong 5-HT uptakecapacity (single arrowheads in (a) surrounded tachykinin-positive cell(asterisk in b). Arrows and double arrowheads point to similarlypositioned, but not identical fibers. The lower panel shows that thevaricose 5-HT-immunopositive fibers (arrowheads in c) run close toChAT-immunoreactive (asterisks in d) cell bodies. Scale bar: 20 lmrelating to all pictures.
Fig. 5. Distribution of tachykinin-positive neurons within the myen-teric plexus. Each black dot represents one tachykinin-IR neuron.Tachykinin-IR neurons were evenly distributed within the myentericplexus. Scale bar: 800 lm.
Fig. 6. Double labeling of tachykinins and ChAT. Double-labeledcell was seen in the myenteric ganglia (arrows in a, b) but ChAT cellsnot labeled for tachykinins were also found (arrowhead in a, b). Also,tachykinin-IR cells were observed lacking ChAT-immunoreactivity (as-terisk in a, b). Scale bar: 15 lm relating to all pictures.
1554 DENES ET AL.

et al., 2003). In this study, a more refined analysis of theeffect of 5-HT was performed.
Tachykinins are thought to be major excitatoric trans-mitters in enteric neurons (Costa et al., 1985; Grider,1989). Therefore, we used NK1, NK2, and NK3 receptorantagonists to analyze contractions evoked by 5-HT and5-HT2 receptor agonist. Considering that the naturalligand of NK1 receptors is SP whereas NK2 receptorsare preferred by NKA (Holzer and Holzer-Petsche,1997), we assume that 5-HT stimulates both SP andNKA release through 5-HT2 receptors. From the consec-utive and reversed administration of drugs, it is clearthat ACh has a prominent role in the residual contrac-tion after application of NK1 antagonist. At the sametime, treatment with ATR could prevent the effect of the5-HT-evoked tachykinin release. It is reasonable toassume that 5-HT could release SP from interneuronsrather than from motor neurons. Our result of withHEX treatments indicates that cholinergic interneuronscould be also involved in this circuit.
To supplement the data obtained by in vitro pharma-cological experiments, immunostainings were also per-formed. First, we found a massive tachykinin-IR systemin the myenteric plexus. We show a strong immunoposi-
tive network in the smooth muscle layer, neurons, andpericellular baskets around both unlabelled and tachyki-nin-immunopositive somata. The finding suggests thattachykinins could participate in motor as well as inte-grative functions in the rabbit ileum. By mapping SP-IRneurons, we established that the cells were distributedevenly. The proportion of tachykinin-IR neurons in rab-bit ileum is surprisingly high compared with rodents.The total neuron density of ileal myenteric plexus inrabbit was established to be �2,500 cells/cm2 (Gabriel etal., 1998). We calculated that �52% of all myenteric neu-rons synthesize tachykinins indicating that these pep-tides must have dominant roles in organizing ilealfunctions. In guinea pig, the SP-IR neuron populationamounted to 23% of all neurons of the ileal myentericplexus (Costa et al., 1996). In mice, 29% of the totalneuron population in the myenteric plexus was foundSP-immunopositive (Sang and Young, 1996).
Seeking for anatomical evidence for interconnectionsamong serotoninergic, cholinergic, and tachykinin-con-taining system, double-label studies were performed. Inmost mammalian species, both SP and NKA appeared tobe involved in neuronal transmission in the enteric nerv-ous system and they coexist with ACh in the same exci-tatory neurons (Grider, 1989; Lippi et al., 1998; Maggi,2000). The proportion of coexistence of these markers,however, varies among species. In guinea pig, all SP-IRneurons were also IR for ChAT, whereas 25–45% of allChAT-IR cells were SP-immunopositive (Steele et al.,1991). In mice, about 50% of ChAT-IR neuron populationcontained SP, and 80% of SP-IR cells contained alsoChAT. Our double-label experiments in rabbit revealed20% of ChAT-IR cells containing also tachykinins and�50% tachykinin-immunopositive cells were IR forChAT.
We did not find 5-HT-IR pericellular baskets aroundeither ChAT or tachykinin-IR cell bodies. Those fewChAT or tachykinin-IR cells that were apposed by 5-HT-IR boutons were not prominent. Synaptology and thusthe function of serotoninergic fibers emerging from sero-toninergic descending interneurons has not been clari-fied yet since the reports that investigated the problempublished different results (Erde et al., 1985; Young andFurness, 1995; Neal and Bornstein, 2007). Erde et al.(1985) reported that mainly type II/AH neurons consid-ered sensory neurons were contacted by serotoninergicterminals. According to Young and Furness (1995),Dogiel type II neurons received a few serotonin inputsand none of them formed synapses, whereas the highestnumber of serotonin terminals were related to Dogieltype I neurons. A more recent works provided preciselyquantified data claiming that anatomical connectionbetween 5-HT-IR varicosities and the longitudinal mus-cle motor neurons was not significant. In contrast, allexcitatory circular muscle motor neurons and someascending interneurons, which are tachykininergic, wereencircled by 5-HT-IR varicosities (Brookes at al., 1997;Neal and Bornstein, 2007). However, it is important tonote that having close contacts with 5-HT-IR varicositiesdo not necessarily mean synaptic specializations (Youngand Furness, 1995). However, this is not a question incase of rabbit considering the small number of cholinergicand tachykinin-containing neurons, which were apposedby 5-HT-IR fibers. Therefore, evidence of morphologicalbasis for 5-HT/tachykinin and 5-HT/ACh interaction
Fig. 8. Expression of 5-HT2A receptors in rabbit myenteric plexus.In rabbit myenteric plexus, a varicose fiber network was immunoposi-tive for anti-5-HT2A and tachykinin antibody (arrows in a and b). Immu-noreactive processes emerging from large cell bodies or running in thestrands were smooth (arrowheads in a and g). Varicose fibers inner-vating the smooth muscle layer were immunoreactive for both tachyki-nins and 5-HT2A receptors (c and d). Some enteric neurons weredouble-labeled (arrows in e and f) but not all the tachykinin-immuno-positive neurons possessed 5-HT2A receptors (single asterisk) andneurons expressing 5-HT2A receptors lacked tachykinin-immunoreac-tivity (double asterisks). Scale bar: 25 lm relating to all pictures.
5-HT AND TACHYKININ IN THE RABBIT ILEUM 1555

could not be found in rabbit ileal myenteric plexus.Clearly, simple light microscopy used in our study is notan efficient tool to reveal synaptic inputs between twostructures. This would require ultrastructural analysis.Besides synaptic transmission, intercellular communica-tion can also occur by volume transmission, which makescommunication possible between nerve elements lackingsynaptic specializations (Zoli et al., 1999; Sykova, 2004;Vizi et al., 2004). 5-HT may be released by nonjunctionalvaricosities and can diffuse to 5-HT receptors of both neu-rons and smooth muscle cells. 5-HT might also besecreted by endocrine cells and act as a hormone orthrough activation of local neural pathways. Numerousstudies revealed that 5-HT was released in response topressure, mucosal stimuli from the intestinal mucosa andevoked peristaltic or secretory reflexes (Kirchgessner etal., 1992; Grider et al., 1996; Zhu et al., 2001; Bertrand,2007; Patel et al., 2007).
The pharmacological experiments showing indirecteffects of 5-HT through 5-HT2 receptors were also con-firmed by immunocytochemistry. We could detect numer-ous neurons as well as a varicose fiber network thatexpressed 5-HT2A receptors in the myenteric plexus. Ourcolocalization studies provided evidence that a smallpopulation of tachykininergic neurons possesses 5-HT2A
receptors that could be either excitatory motorneuronsor interneurons. In addition to this population, therewere tachykinin-IR neurons lacking 5-HT2A receptorsand many 5-HT2A receptor expressing cells, in fact, didnot show tachykinin-immunopositivity.
By comparing our observations with results obtainedin other species, we can state that the action of 5-HT onileal motility in rabbit shares both similarities and dis-similarities to other species like guinea pig, rat, andmouse that are close to rabbits in terms of diet andevolution.
Because the first description that 5-HT potentlyaffects intestinal movements (Gaddum, 1953), actions of5-HT have been investigated in a wide range of animalsand from different aspects. Most of the data have beenprovided from guinea pig small intestine. Two different5-HT receptors can mediate contractile responses named5-HT3 and 5-HT4 receptors located on neuronal compo-nents (Craig and Clarke, 1990; Craig et al., 1990; Eglenet al., 1990; Taniyama et al., 1991). 5-HT acting at5-HT3 receptors could evoke ascending as well as de-scending excitatory reflexes (Yuan et al., 1994) throughactivation of cholinergic motor neurons (Fox and Morton,1990; Monro et al., 2002). 5-HT4 receptors can also medi-ate contractile effect, increase peristaltic reflex sensitiv-ity (Craig and Clarke, 1990, 1991; Buchheit and Buhl,1991; Kilbinger and Wolf, 1992; Kilbinger et al., 1995;Galligan et al., 2003) like 5-HT3 receptors do. Besidesthe cholinergic system, evidence was reported thattachykininergic neurons also had roles in contractionsinduced by 5-HT (Buchheit et al., 1985; Ramirez et al.,1994). In mouse ileum, effect of 5-HT was mediatedthrough neuronally located 5-HT3 receptors (Tuladhar etal., 2000; Chetty et al., 2006). Data obtained in rat ileumare rather controversial. Some studies claimed that5-HT2A and 5-HT1 receptors located on the smooth mus-cle were involved in 5-HT-induced contractions, and nei-ther TTX nor atropin caused inhibition suggesting adirect effect on rat ileal muscle (Briejer et al., 1997;Yamano et al., 1997). Meanwhile, the role of 5-HT3
receptors acting through neuronal activation in rat il-eum was also proved (Chetty et al., 2006). The latterstudy also suggested that tachykinins were not media-ting 5-HT3-induced contractions.
The main conclusions of our study, therefore, were asfollows: (i) in rabbit, 5-HT could stimulate contractionson both indirect and direct ways, whereas in guinea pig,rat, and mouse 5-HT has only one way to act, eitherdirect or indirect. (ii) Cholinergic and tachykinin-con-taining motorneurons and also interneurons are partici-pating in muscle stimulation in rabbit (Fox and Morton,1990; Briejer and Schuurkes, 1996). (iii) In rabbit, move-ments of longitudinal muscle strips could be evokedpotentially via two kinds of 5-HT receptor, 5-HT2 and5-HT4, whereas 5-HT3, 5-HT4 receptors take parts inmediation of 5-HT-evoked contractions in guinea pig,5-HT2 receptors in rat and 5-HT3 receptors in mouse,respectively.
LITERATURE CITED
Bertrand PP. 2007. Real-time measurement of serotonin release andmotility in guinea pig ileum. J Physiol 577:689–704.
Bjorck S, Ahlman A, Dahlstrom A, Phillips SF, Kelly KA. 1988.Serotoninergic regulation of canine enteric motility (measured aselectrical activity) and absorption: physiologic and morphologicevidence. Acta Physiol Scand 133:247–256.
Briejer MR, Mathis C, Schuurkes JA. 1997. 5-HT receptor types inthe rat ileum longitudinal muscle: focus on 5-HT2 receptors medi-ating contraction. Neurogastroenterol Motil 9:231–237.
Briejer MR, Schuurkes JAJ. 1996. 5-HT3 and 5-HT4 receptors andcholinergic and tachykininergic neurotransmission in the guinea-pig proximal colon. Eur J Pharmacol 308:173–180.
Brookes SJ. 2001. Classes of enteric nerve cells in the guinea-pigsmall intestine. Anat Rec 262:58–70.
Brookes SJ, Meedeniya AC, Jobling P, Costa M. 1997. Orally projec-ting interneurones in the guinea-pig small intestine. J Physiol105:473–491.
Brookes SJ, Song ZM, Steele PA, Costa M. 1992. Identification ofmotor neurons to the longitudinal muscle of the guinea pig ileum.Gastroenterology 103:961–973.
Brookes SJ, Steele PA, Costa M. 1991. Identification and immuno-histochemistry of cholinergic and non-cholinergic circular musclemotor neurons in the guinea-pig small intestine. Neuroscience42:863–878.
Buchheit KH, Buhl T. 1991. Prokinetic benzamides stimulate peri-staltic activity in the isolated guinea pig ileum by activation of5-HT4 receptors. Eur J Pharmacol 205:203–208.
Buchheit KH, Engel G, Mutschler E, Richardson B. 1985. Study ofthe contractile effect of 5-hydroxytryptamine (5-HT) in the iso-lated longitudinal muscle strip from guinea-pig ileum. Evidencefor two distinct release mechanisms. Naunyn Schmiedebergs ArchPharmacol 329:36–41.
Bush TG, Spencer NJ, Watters N, Sanders KM, Smith TK. 2001.Effects of alosetron on spontaneous migrating motor complexes inmurine small and large bowel in vitro. Am J Physiol GastrointestLiver Physiol 281:G974–G983.
Chetty N, Irving HR, Coupar IM. 2006. Activation of 5-HT3 recep-tors in the rat and mouse intestinal tract: a comparative study.Br J Pharmacol 148:1012–1021.
Costa M, Brookes SJH, Steele PA, Gibbins I, Burcher E, KandiahCJ. 1996. Neurochemical classification of myenteric neurons inthe guinea-pig ileum. Neuroscience 75:949–967.
Costa M, Cuello AC, Furness JB, Franco R. 1980. Distribution ofenteric neurons showing immunoreactivity for substance P in theguinea-pig ileum. Neuroscience 5:323–331.
Costa M, Furness JB, Cuello AC, Verhofstad AAJ, SteinbuschHWJ, Elde RP. 1982. Neurons with 5-hydroxytryptamine-likeimmunoreactivity in the enteric nervous system: their
1556 DENES ET AL.

visualization and reactions to drug treatment. Neuroscience7:351–363.
Costa M, Furness JB, Llewellyn-Smith IJ, Cuello AC. 1981. Projec-tions of substance P-containing neurons within the guinea-pigsmall intestine. Neuroscience 6:411–424.
Costa M, Furness JB, Llewellyn-Smith IJ, Murphy R, Bornstein JC,Keast JR. 1985. Functional roles for substance P-containing neu-rons in the gastrointestinal tract. In: Jordan CC,Ohme P, editors.Substance P metabolism and biological actions. London: Taylorand Francis. p 99–119.
Craig DA, Clarke DE. 1990. Pharmacological characterization of aneuronal receptor for 5-hydroxytryptamine in guinea pig ileumwith properties similar to the 5-hydroxytryptamine receptor.J Pharmacol Exp Ther 252:1378–1386.
Craig DA, Clarke DE. 1991. Peristalsis evoked by 5-HT andrenzapride: evidence for putative 5-HT4 receptor activation. Br JPharmacol 102:563–564.
Craig DA, Eglen RM, Walsh LK, Perkins LA, Whiting RL, ClarkeDE. 1990. 5-Methoxytryptamine and 2-methyl-5-hydroxytrypta-mine-induced desensitization as a discriminative tool for the5-HT3 and putative 5-HT4 receptors in guinea pig ileum. NaunynSchmiedebergs Arch Pharmacol 342:9–16.
Denes V, Lazar ZS, Bartho L, Gabriel R. 2003. Serotonin in the rab-bit ileum: localization, uptake and effect on motility. Anat Rec271:368–376.
Donowitz M, Tai YH, Asarkof N. 1980. Effect of serotonin on activeelectrolyte transport in rabbit ileum, gallbladder and colon. Am JPhysiol 239:G463–G472.
Eglen RM, Swank SR, Walsh LKM, Whiting RL. 1990. Character-isation of 5-HT3 and atypical 5-HT receptors mediating guinea-pig ileal contractions in vitro. Br J Pharmacol 101:513–520.
Ekblad E, Winther C, Ekman R, Hakanson R, Sundler F. 1987.Projections of peptide-containing neurons in rat small intestine.Neuroscience 20:169–188.
Erde SM, Sherman D, Gershon MD. 1985. Morphology and seroto-nergic innervation of physiologically identified cells of the guineapig’s myenteric plexus. J Neurosci 5:617–633.
Erspamer V, Asero B. 1952. Identification of enteramine, the spe-cific hormone of the enterochromaffin cell system as 5-hydroxy-tryptamine. Nature 169:800–801.
Fox AJ, Morton IKM. 1990. An examination of the 5-HT3 receptormediating contraction and evoked [3H]-ACh release in the guinea-pig ileum. Br J Pharmacol 101:553–556.
Fujimiya M, Okumiya K, Yamane T, Maeda T. 1997. Distribution ofserotonin-immunoreactive nerve cells and fibers in rat gastroin-testinal tract. Histochem Cell Biol 107:105–114.
Gabriel R, Pasztor I, Denes V, Wilhelm M. 1998. Some neurohisto-chemical properties of nerve elements in myenteric plexus of rab-bit ileum: similarities and dissimilarities to the rodent pattern.Cell Tissue Res 292:283–291.
Gaddum JH. 1953. Tryptamine receptors. J Physiol 119:363–368.Galligan JJ, Pan H, Messori E. 2003. Signalling mechanism coupledto 5-hydroxytryptamine4 receptor-mediated facilitation of fastsynaptic transmission in the guinea-pig ileum myenteric plexus.Neurogastroenterol Motil 15:523–529.
Gershon MD, Drakontides AB, Ross LL. 1965. Serotonin: synthesisand release from the myenteric plexus of the mouse intestine. Sci-ence 149:197–199.
Goyal RK, Hirano I. 1996. Mechanisms of disease: the enteric nerv-ous system. New Engl J Med 334:1106–1115.
Grady EF, Baluk P, Bohm S, Gamp PD, Wong H, Payan DG, AnselJ, Portbury AL, Furness JB, McDonald DM, Bunett NW. 1996.Characterization of antisera specific to NK1, NK2 and NK3 neu-rokinin receptors and their utilization to localize receptors in therat gastrointestinal tract. J Neurosci 16:6975–6986.
Grider JR. 1989. Tachykinins as transmitters of ascending contract-ile component of the peristaltic reflex. Am J Physiol 257:G709–G714.
Grider JR, Kuemmerle JF, Jin JG. 1996. 5-HT released by mucosalstimuli initiates peristalsis by activating 5-HT4/5-HT1p receptorson sensory CGRP neurons. Am J Physiol 270:G778–G782.
Guard S, Watson SP. 1991. Tachykinin receptors types: classifica-tion and membrane signalling mechanisms. Neurochem Int18:149–165.
Heitz P, Polak JM, Timson CM, Pearse AGE. 1976. Enterochromaf-fin cells as endocrine source of gastrointestinal substance P. His-tochemistry 49:343–347.
Holzer P, Holzer-Petsche U. 1997. Tachykinins in the gut. II. Rolesin neural excitation, secretion and inflammation. Pharmacol Ther73:219–263.
Jenkinson KM, Mann PT, Southwell BR, Furness JB. 2000. Inde-pendent endocytosis of the NK(1) and NK(3) tachykinin receptorsin neurons of the rat myenteric plexus. Neuroscience 100:191–199.
Jenkinson KM, Morgan JM, Furness JB, Southwell BR. 1999. Neu-rons bearing NK(3) tachykinin receptors in the guinea-pig ileumrevealed by specific binding of fluorescently labelled agonists. His-tochem Cell Biol 112:233–246.
Johnson PJ, Bornstein JC, Burcher E. 1998. Roles of neuronalNK1 and NK3 receptors in synaptic transmission during motilityreflexes in the guinea-pig ileum. Br J Pharmacol 124:1375–1384.
Johnson PJ, Bornstein JC, Yuan SY, Furness JB. 1996. Analysis ofcontributions of acetylcholine and tachykinins to neuro-neuronaltransmission in motility reflexes in guinea-pig ileum. Br J Phar-macol 118:973–983.
Kangawa K, Minamino N, Fukuda A, Matsuo H. 1983. NeuromedinK: a novel mammalian tachykinin identified in porcine spinalcord. Biochem Biophys Res Commun 114:533–540.
Kilbinger H, Gebauer A, Haas J, Ladinsky H, Rizzi CA. 1995. Ben-zimidazolones and renzapride facilitate acetylcholine release fromguinea-pig myenteric plexus via 5-HT4 receptors. NaunynSchmiedebergs Arch Pharmacol 351:229–236.
Kilbinger H, Wolf D. 1992. Effects of 5-HT4 receptor stimulation onbasal and electrically evoked release of acetylcholine from guinea-pig myenteric plexus. Naunyn Schmiedebergs Arch Pharmacol345:270–275.
Kirchgessner AL, Tamir H, Gershon MD. 1992. Identification andstimulation by serotonin of intrinsic sensory neurons of the sub-mucosal plexus of the guinea pig gut: activity-induced expressionof Fos immunoreactivity. J Neurosci 12:235–248.
Kunze WAA, Furness JB. 1999. The enteric nervous system andregulation of intestinal motility. Annu Rev Physiol 61:117–142.
Lazarus LH, Di Augustine RP. 1980. Radioimmunoassay for thetachykinin peptide physalaemin: detection of physalaemin-likesubstance in rabbit stomach. Anal Biochem 107:350–357.
Lecci A, Santicioli P, Maggi CA. 2002. Pharmacology of transmis-sion to gastrointestinal muscle. Curr Opin Pharmacol 2:630–641.
Lippi A, Santicioli P, Criscuoli M, Maggi CA. 1998. Depolarizationevoked co-release of tachykinins from enteric nerves in theguinea-pig proximal colon. Naunyn Schmiedebergs Arch Pharma-col 357:245–251.
Lomax AEG, Bertrand PP, Furness JB. 1998. Identification of thepopulations of enteric neurons that have NK1 tachykinin recep-tors in the guinea-pig small intestine. Cell Tissue Res 294:27–33.
Maggi CA. 2000. Principles of tachykininergic co-transmission inthe peripheral and enteric nervous system. Regul Pept 93:53–64.
Monro RL, Bertrand PP, Bornstein JC. 2002. ATP and 5-HT are theprincipal neurotransmitters in the descending excitatory reflexpathway of the guinea-pig ileum. Neurogastroenterol Motil14:255–264.
Nawa H, Doteuchi M, Igano K, Inouye K, Nakanishi S. 1984. Sub-stance K: a novel mammalian tachykinin that differs from sub-stance P in its pharmacological profile. Life Sci 34:1153–1160.
Neal KB, Bornstein JC. 2007. Mapping 5-HT inputs to entericneurons of the guinea-pig small intestine. Neuroscience 145:556–567.
Olsson C, Holmgren S. 2001. The control of gut motility. Comp Bio-chem Physiol 128:481–503.
Patel BA, Bian X, Quaiserova-Mocko V, Galligan JJ, Swain GM.2007. In vitro continuous amperometric monitoring of 5-hydroxy-tryptamine release from enterochromaffin cells of the guinea pigileum. Analyst 132:41–47.
5-HT AND TACHYKININ IN THE RABBIT ILEUM 1557

Paton WDM, Vizi ES. 1969. The inhibitory action of noradrenalineon acetylcholine output by guinea-pig ileum longitudinal musclestrip. Br J Pharmacol 35:10–18.
Portbury AL, Furness JB, Southwell BR, Wong H, Walsh JH, Bun-nett NW. 1996b. Distribution of neurokinin-2 receptors in theguinea-pig gastrointestinal tract. Cell Tissue Res 286:281–292.
Portbury AL, Furness JB, Young HM, Southwell BR, Vigna SR.1996a. Localisation of NK1 receptor immunoreactivity to neuronsand interstitial cells of the guinea-pig gastrointestinal tract.J Comp Neurol 367:342–351.
Ramırez MJ, Cenarruzabeitia E, Del Rıo J, Lasheras B. 1994.Involvement of neurokinins in the non-cholinergic response toactivation of 5-HT3 and 5-HT4 receptors in guinea-pig ileum. BrJ Pharmacol 111:419–424.
Salvador MT, Murillo MD, Rodriguez-Yoldi MC, Alcalde AI, Meso-nero JE, Rodriguez-Yoldi MJ. 2000. Effects of serotonin on thephysiology of the rabbit small intestine. Can J Physiol Pharmacol78:359–366.
Salvador MT, Rodriguez-Yoldi MC, Alcalde AI, Rodriguez-Yoldi MJ.1997. 5-HT receptor subtypes involved in the serotonin-induced inhi-bition of L-leucin absorption in rabbit jejunum. Life Sci 61:309–318.
Sang Q, Young HM. 1996. Chemical coding of neurons in myentericplexus and external muscle of the small and large intestine of themouse. Cell Tissue Res 284:39–53.
Sang Q, Young HM. 1998. The identification and chemical coding ofcholinergic neurons in the small and large intestine of the mouse.Anat Rec 251:185–199.
Schultzberg M, Hokfelt T, Nilsson G, Terenius L, Rehfeld JF, BrownM, Elde R, Goldstein M, Said S. 1980. Distribution of peptide-and catecholamine-containing neurons in the gastro-intestinaltract of rat and guinea-pig: immunohistochemical studies withantisera to substance P, vasoactive intestinal polypeptide, enke-phalins, somatostatin, gastrin/cholecystokinin, neurotensin anddopamine beta-hydroxylase. Neuroscience 5:689–744.
Shimizu Y, Matsuyama H, Shiina T, Takewaki T, Furness JB. 2008.Tachykinins and their functions in the gastrointestinal tract. CellMol Life Sci 65:295–311.
Shuttleworth CWR, Keef KD. 1995. Roles of peptides in enteric neu-romuscular transmission. Regul Pept 56:101–120.
Steele PA, Brookes SJ, Costa M. 1991. Immunohistochemical identi-fication of cholinergic neurons in the myenteric plexus of guinea-pig small intestine. Neuroscience 45:227–239.
Sundaram U, Knickelbein RG, Dobbins JW. 1991. Mechanism ofintestinal secretion. Effect of serotonin on rabbit ileal crypt andvillus cells. J Clin Invest 87:743–746.
Sykova E. 2004. Extrasynaptic volume transmission and diffu-sion parameters of the extracellular space. Neuroscience129:861–876.
Taniyama K, Nakayama S, Takeda K, Matsuyama S, Shirakawa J,Sano I, Tanaka C. 1991. Cisapride stimulates motility of theintestine via the 5-hydroxytryptamine receptors. J PharmacolExp Ther 258:1098–1104.
Thornton PD, Bornstein JC. 2002. Slow excitatory synaptic poten-tials evoked by distension in myenteric descending interneuronesof guinea-pig ileum. J Physiol 539:589–602.
Timmermanns JP, Scheuermann DW, Stach W, Adriaensen D, DeGroodt-Lasseel MHA. 1990. Distinct distribution of CGRP-, en-kephalin-, galanin-, neuromedin U-, neuropeptide Y-, somatosta-tin-, substance P-, VIP- and serotonin-containing neurons in twosubmucosal ganglionic neural networks of the porcine small intes-tine. Cell Tissue Res 260:367–379.
Toole L, Belai A, Burnstock G. 1998. A neurochemical characterisa-tion of the golden hamster myenteric plexus. Cell Tissue Res291:385–394.
Tuladhar BR, Womack MD, Naylor RJ. 2000. Pharmacological char-acterization of the 5-HT receptor-mediated contraction in themouse isolated ileum. Br J Pharmacol 131:1716–1722.
Vizi ES, Kiss JP, Lendvai B. 2004. Nonsynaptic communication inthe central nervous system. Neurochem Int 45:443–451.
Wade PR, Chen J, Jaffe B, Kassem IS, Blakely RD, Gershon MD.1996. Localization and function of a 5-HT transporter incrypt epithelia of the gastrointestinal tract. J Neurosci16:2352–2364.
Yamano M, Ito H, Miyata K. 1997. Species differences in the 5-hy-droxytryptamine-induced contraction in the isolated distal ileum.Jpn J Pharmacol 74:267–274.
Young HM, Furness JB. 1995. Ultrastructural examination of sero-tonin-immunoreactive descending interneurons in guinea pigsmall intestine. J Comp Neurol 356:101–114.
Yuan SY, Bornstein JC, Furness JB. 1994. Investigation of the roleof 5-HT3 and 5-HT4 receptors in ascending and descendingreflexes to the circular muscle of guinea-pig small intestine. Br JPharmacol 112:1095–1100.
Zhu JX, Wu XY, Owyang C, Li Y. 2001. Intestinal serotonin acts asa paracrine substance to mediate vagal signal transmissionevoked by luminal factors in the rat. Am J Physiol GastrointestLiver Physiol 281:G916–G923.
Zoli M, Jansson A, Sykova E, Agnati LF, Fuxe K. 1999. Volumetransmission in the CNS and its relevance for neuropsychophar-macology. Trends Pharmacol Sci 20:142–150.
1558 DENES ET AL.




















