
Developmental aspects of the cholinergic system
13
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of Developmental aspects of the cholinergic system
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Behavioural Brain Research 221 (2011) 367–378
Contents lists available at ScienceDirect
Behavioural Brain Research
journa l homepage: www.e lsev ier .com/ locate /bbr
Review
Developmental aspects of the cholinergic system
Yael Abreu-Villaca ∗, Cláudio C. Filgueiras, Alex C. ManhãesLaboratório de Neurofisiologia, Departamento de Ciências Fisiológicas, Instituto de Biologia Roberto Alcantara Gomes, Centro Biomédico, Universidade do Estado do Rio de Janeiro,Av. Prof. Manoel de Abreu 444, 5 andar, Vila Isabel, Rio de Janeiro, RJ 20550-170, Brazil
a r t i c l e i n f o
Article history:Received 22 December 2009Accepted 26 December 2009Available online 6 January 2010
Keywords:AcetylcholineCholine acetyltransferaseVesicular acetylcholine transporterHigh-affinity choline transporterAcetylcholinesteraseNicotinic receptorsMuscarinic receptorsDevelopment
a b s t r a c t
Beyond its importance in sustaining or modulating different aspects of the activity of the central nervoussystem (CNS), the cholinergic system plays important roles during development. In the current review,we focus on the developmental aspects associated with major components of the cholinergic system:Acetylcholine, choline acetyltransferase, vesicular acetylcholine transporter, high-affinity choline trans-porter, acetylcholinesterase, nicotinic and muscarinic receptors. We describe when and where each oneof these components is first identified in the CNS and the changes in their levels that occur during thecourse of prenatal and postnatal development. We also describe how these components are relevantto many events that occur during the development of the CNS, including progenitor cells proliferationand differentiation, neurogenesis, gliogenesis, neuronal maturation and plasticity, axonal pathfinding,regulation of gene expression and cell survival. It will be noticed that evidence regarding the develop-mental aspects of the cholinergic system comes mostly from studies that used agonists, such as nicotine,and antagonists, such as hemicholinium-3. Studies using immunohistochemistry and genetically alteredmice also provided valuable information.
© 2010 Elsevier B.V. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3672. Choline acetyltransferase, vesicular acetylcholine transporter and acetylcholine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3683. Cholinergic receptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 369
3.1. Nicotinic AChRs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3693.2. Muscarinic AChRs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 371
4. Acetylcholinesterase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3725. High-affinity choline transporter . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3736. Future directions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 374
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 374
1. Introduction
Acetylcholine (ACh) is a neurotransmitter widely distributed inthe central (and also peripheral, autonomic and enteric) nervoussystem. It is synthesized from choline and acetyl CoA by the cholineacetyltransferase (ChAT) enzyme and stored in presynaptic vesi-cles. The vesicular acetylcholine transporter (VAChT) is responsiblefor loading ACh into these vesicles. When released in the synapticcleft, ACh binds to two distinct types of receptors: Nicotinic acetyl-choline receptors (nAChR) and muscarinic acetylcholine receptors(mAChRs). Neuronal nAChR subtypes are located in pre- and post-
∗ Corresponding author. Tel.: +55 21 2587 6295; fax: +55 21 2587 6129.E-mail address: yael a [email protected] (Y. Abreu-Villaca).
synaptic membranes of neurons and also in extrasynaptic locationsthroughout the CNS. In the cleft, ACh may also be hydrolyzed byacetylcholinesterase (AChE) into choline and acetate and almost50% of the choline derived from ACh hydrolysis is recovered bya presynaptic high-affinity choline transporter (CHT) supportingcontinued ACh production and release (Fig. 1) [8].
ACh, ChAT, VAChT, nAChRs, mAChRs, AChE and CHT are com-ponents of the cholinergic system and, as described below, eachone of them follows a characteristic developmental plan, from themoment they first appear in the different regions of the CNS to thetime when adult levels are achieved. Besides, these componentsplay important roles in many events that occur during the devel-opment of the central nervous system (CNS) (Fig. 2). In the ensuingparagraphs, we also review how events such as progenitor cells pro-liferation and differentiation, neurogenesis, gliogenesis, neuronal
0166-4328/$ – see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.bbr.2009.12.049
Author's personal copy
368 Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378
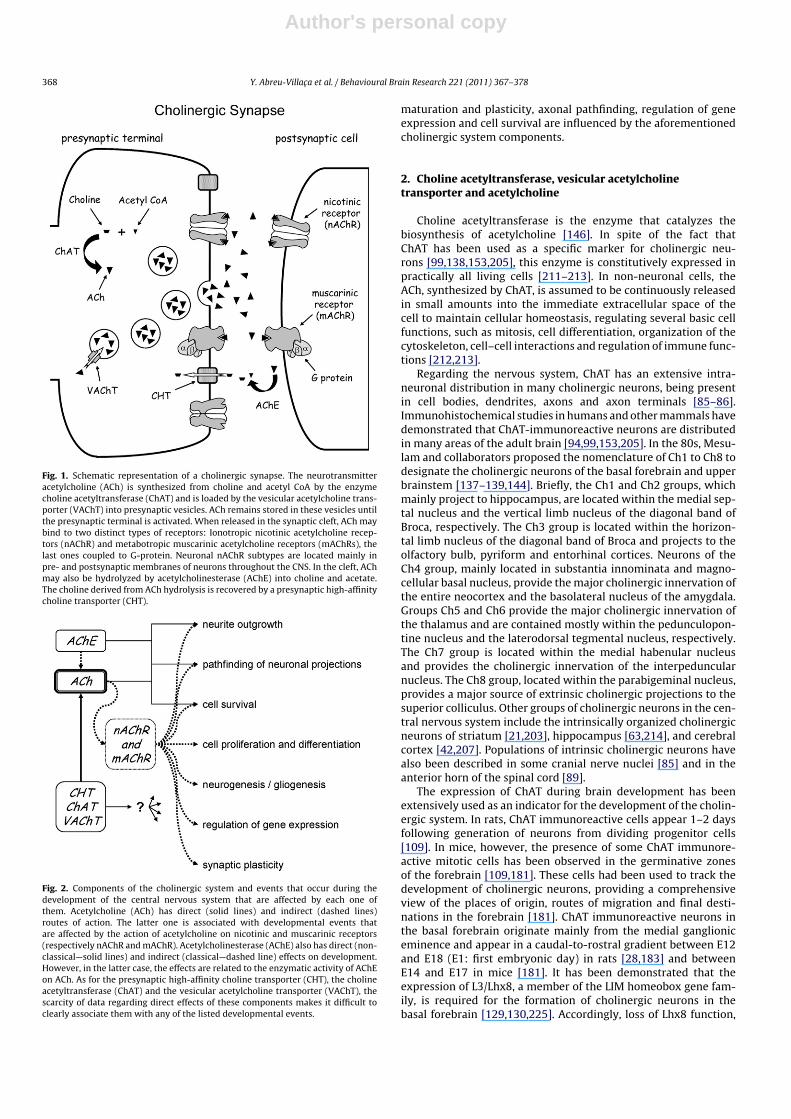
Fig. 1. Schematic representation of a cholinergic synapse. The neurotransmitteracetylcholine (ACh) is synthesized from choline and acetyl CoA by the enzymecholine acetyltransferase (ChAT) and is loaded by the vesicular acetylcholine trans-porter (VAChT) into presynaptic vesicles. ACh remains stored in these vesicles untilthe presynaptic terminal is activated. When released in the synaptic cleft, ACh maybind to two distinct types of receptors: Ionotropic nicotinic acetylcholine recep-tors (nAChR) and metabotropic muscarinic acetylcholine receptors (mAChRs), thelast ones coupled to G-protein. Neuronal nAChR subtypes are located mainly inpre- and postsynaptic membranes of neurons throughout the CNS. In the cleft, AChmay also be hydrolyzed by acetylcholinesterase (AChE) into choline and acetate.The choline derived from ACh hydrolysis is recovered by a presynaptic high-affinitycholine transporter (CHT).
Fig. 2. Components of the cholinergic system and events that occur during thedevelopment of the central nervous system that are affected by each one ofthem. Acetylcholine (ACh) has direct (solid lines) and indirect (dashed lines)routes of action. The latter one is associated with developmental events thatare affected by the action of acetylcholine on nicotinic and muscarinic receptors(respectively nAChR and mAChR). Acetylcholinesterase (AChE) also has direct (non-classical—solid lines) and indirect (classical—dashed line) effects on development.However, in the latter case, the effects are related to the enzymatic activity of AChEon ACh. As for the presynaptic high-affinity choline transporter (CHT), the cholineacetyltransferase (ChAT) and the vesicular acetylcholine transporter (VAChT), thescarcity of data regarding direct effects of these components makes it difficult toclearly associate them with any of the listed developmental events.
maturation and plasticity, axonal pathfinding, regulation of geneexpression and cell survival are influenced by the aforementionedcholinergic system components.
2. Choline acetyltransferase, vesicular acetylcholinetransporter and acetylcholine
Choline acetyltransferase is the enzyme that catalyzes thebiosynthesis of acetylcholine [146]. In spite of the fact thatChAT has been used as a specific marker for cholinergic neu-rons [99,138,153,205], this enzyme is constitutively expressed inpractically all living cells [211–213]. In non-neuronal cells, theACh, synthesized by ChAT, is assumed to be continuously releasedin small amounts into the immediate extracellular space of thecell to maintain cellular homeostasis, regulating several basic cellfunctions, such as mitosis, cell differentiation, organization of thecytoskeleton, cell–cell interactions and regulation of immune func-tions [212,213].
Regarding the nervous system, ChAT has an extensive intra-neuronal distribution in many cholinergic neurons, being presentin cell bodies, dendrites, axons and axon terminals [85–86].Immunohistochemical studies in humans and other mammals havedemonstrated that ChAT-immunoreactive neurons are distributedin many areas of the adult brain [94,99,153,205]. In the 80s, Mesu-lam and collaborators proposed the nomenclature of Ch1 to Ch8 todesignate the cholinergic neurons of the basal forebrain and upperbrainstem [137–139,144]. Briefly, the Ch1 and Ch2 groups, whichmainly project to hippocampus, are located within the medial sep-tal nucleus and the vertical limb nucleus of the diagonal band ofBroca, respectively. The Ch3 group is located within the horizon-tal limb nucleus of the diagonal band of Broca and projects to theolfactory bulb, pyriform and entorhinal cortices. Neurons of theCh4 group, mainly located in substantia innominata and magno-cellular basal nucleus, provide the major cholinergic innervation ofthe entire neocortex and the basolateral nucleus of the amygdala.Groups Ch5 and Ch6 provide the major cholinergic innervation ofthe thalamus and are contained mostly within the pedunculopon-tine nucleus and the laterodorsal tegmental nucleus, respectively.The Ch7 group is located within the medial habenular nucleusand provides the cholinergic innervation of the interpeduncularnucleus. The Ch8 group, located within the parabigeminal nucleus,provides a major source of extrinsic cholinergic projections to thesuperior colliculus. Other groups of cholinergic neurons in the cen-tral nervous system include the intrinsically organized cholinergicneurons of striatum [21,203], hippocampus [63,214], and cerebralcortex [42,207]. Populations of intrinsic cholinergic neurons havealso been described in some cranial nerve nuclei [85] and in theanterior horn of the spinal cord [89].
The expression of ChAT during brain development has beenextensively used as an indicator for the development of the cholin-ergic system. In rats, ChAT immunoreactive cells appear 1–2 daysfollowing generation of neurons from dividing progenitor cells[109]. In mice, however, the presence of some ChAT immunore-active mitotic cells has been observed in the germinative zonesof the forebrain [109,181]. These cells had been used to track thedevelopment of cholinergic neurons, providing a comprehensiveview of the places of origin, routes of migration and final desti-nations in the forebrain [181]. ChAT immunoreactive neurons inthe basal forebrain originate mainly from the medial ganglioniceminence and appear in a caudal-to-rostral gradient between E12and E18 (E1: first embryonic day) in rats [28,183] and betweenE14 and E17 in mice [181]. It has been demonstrated that theexpression of L3/Lhx8, a member of the LIM homeobox gene fam-ily, is required for the formation of cholinergic neurons in thebasal forebrain [129,130,225]. Accordingly, loss of Lhx8 function,

Author's personal copy
Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378 369
via a targeted mutation in the mouse, blocks the formation of mostcholinergic neurons in the forebrain [225]. In addition, suppressionof L3/Lhx8 expression results in a dramatic decrease in the num-ber of ChAT-positive cells obtained from cultured mouse stem cells,while overexpression of L3/Lhx8 reverts this effect [129]. The Olig2,a basic helix–loop–helix transcription factor expressed in germi-native zones associated with generation of ChAT-positive cells hasalso been implicated in the development of cholinergic neurons inthe basal forebrain [65].
As compared to adults, ChAT levels remain relatively low untilbirth [14,46,72]. In the majority of ACh groups of the basal fore-brain, the expression of ChAT in cholinergic neurons and terminals,as well as of ChAT activity, rises progressively until adult levelsare attained approximately between the third and fourth weeksafter birth [43,51,72,84,141,151]. In the rat laterodorsal tegmen-tal nucleus for instance, ChAT activity increases strikingly duringthe second postnatal week and moderately thereafter, reaching theadult level in the fourth postnatal week [151]. This time courseof change in ChAT activity has been related to the time course ofthe development of some locomotor patterns and to decreases inthe amount of paradoxal sleep and frequency of muscle twitchesafter birth [150,151]. Of note, different patterns of developmentalchanges have been described in other cholinergic populations. Inthe striatum, ChAT activity increases slightly during the first twoweeks, rises strikingly at P15 (P1: first postnatal day), and reachesadult levels after P25 [141,198]. In the cortex and hippocampus, asharp increase in ChAT activity occurs ten days after birth, reachingclose to adult levels at three weeks of age [107].
In general, the levels of ChAT activity reflects developmentalchanges in ChAT mRNA levels, with the increase in mRNA precedingthat of enzyme activity by about 2 days [84]. Of note, the ChAT andthe vesicular acetylcholine transporter (VAChT) genes are arrangedin a common locus, referred to as “the cholinergic locus”, wherethe gene encoding the VAChT is located within the first intron ofthe gene encoding ChAT and is in the same transcriptional orienta-tion as ChAT [56,209]. The VAChT and ChAT co-expression, whichis required for the characterization of the cholinergic phenotype[34,209], has been studied during development. It was reportedthat VAChT protein levels parallels that of ChAT during develop-ment, whereas VAChT gene transcription begins much earlier indevelopment than does ChAT, with greater than half of adult VAChTmRNA levels present in the rat at birth, compared with only 20% ofadult levels of ChAT mRNA at this time [84].
Taken together, the aforementioned studies indicate that theearliest development of ACh machinery take place in regions con-taining cholinergic cell bodies, followed later by their target areas.For instance, ChAT activity in the medial septum, diagonal bandand magnocellular preoptic region preceded detectable enzymeactivity in their targets at hippocampus and cortex by some days[198]. Several studies have demonstrated that cholinergic neuronscan synthesize ACh during their migration and that these neuronscan release ACh from their growth cones prior to target contact orsynapse formation [91,196,220]. Interestingly, ACh secretion dur-ing development is not restricted to the growth cone, but has alsobeen reported along the neurite and at the soma [7,11,222]. Indeveloping Xenopus neurons, ACh secretion is mediated by vesiclesthat are recycled locally within the axon and that can be detectedthroughout the entire axonal surface [222]. In cultured Drosophilaneurons, the machinery for spontaneous quantal ACh secretionis present throughout the entire neuron, whereas ACh packets atthe soma are considerably greater than those at the growth cones[219]. It has been suggested that ACh released in the extracellu-lar medium from growing axons contributes to the developmentof neuronal networks [11,177]. Accordingly, it has been demon-strated that ACh participates in axonal pathfinding and navigationby modulating the rate and direction of axonal growth [226] and
that it affects axonal outgrowth as well as growth cone morphology[158,177].
There is a large body of literature demonstrating that the activityof both ChAT and VAChT are developmentally regulated. Partic-ularly, several extracellular factors are capable of regulating thecholinergic phenotype and of exerting trophic effects on devel-oping cholinergic neurons [6,20,123,152,199,231,232]. One of thebest characterized factors is the nerve growth factor (NGF). NGFis a member of the neurotrophin family of structurally relatedpolypeptides crucial for differentiation, survival and function ofcertain neuronal populations [17,116]. Large et al. [107] demon-strated that the developmental increase in NGF content of thebasal forebrain parallels that of the neocortex and hippocampus,and that it precedes an increase of similar magnitude in ChATactivity. NGF released from targets upregulates the expressionof the ChAT and VAChT genes in developing cholinergic neu-rons [22,120,127,161,199] interacting with two receptors: Thelow-affinity neurotrophin receptor p75NTR and the high-affinityreceptor TrkA [22,120]. In addition to NGF, other related neu-rotrophins, such as the brain-derived neurotrophic factor (BDNF),also promote the basal forebrain cholinergic phenotype [6,152].
Studies with genetically modified mice have shown that bothVAChT and ChAT play an essential role in the normal developmentof the cholinergic system. ChAT homozygous knockout mice arestillborn and present several neuromuscular alterations such asincreased number of motoneurons, muscle hyperinnervation andalterations in ACh receptor clusters in muscle [29]. Mice lackingVAChT (VAChT knockout) present neuromuscular abnormalitiessimilar to those found in ChAT knockout and also die shortly afterbirth [48]. However, mice with reduced levels of VAChT (VAChTknockdown) are viable and have been used to evaluate the role ofthis transporter [49,163]. For instance, VAChT knockdown homozy-gous mice with 65% reduction in the expression levels of VAChT aremyasthenic and have cognitive deficits in tasks involving objectand social recognition during adulthood [163]. Similarly, humanspresenting mutations in cholinergic locus present congenital myas-thenic syndrome associated with frequently fatal episodes of apnea[154]. Despite of the fact that the abnormalities observed in mutantmice and humans are likely attributable to the loss of ACh func-tion, it is not possible to discard the possibility that ChAT and/orVAChT have other nonenzymatic activities during development.The abnormal phenotype observed in knockdown and knockoutmodels may also be caused by some compensatory mechanismtriggered by the early absence of ACh. In this sense, the post-transcriptional gene knockdown through RNA interference (RNAi),which was recently developed to promptly disrupt CHAT expres-sion [178], could provide important contributions.
3. Cholinergic receptors
ACh receptors are differentially expressed throughout the CNS,where they play a role in the modulation of cell proliferation andsurvival, neuronal differentiation, regulation of gene expression,synapse formation and maturation, axonal pathfinding and neu-rotransmitter release. As described below, studies on the effectsof both nAChRs and mAChRs agonists and antagonists have beenextremely helpful to the understanding of the importance of thesereceptors to brain development (Table 1).
3.1. Nicotinic AChRs
Nicotinic receptors are pentameric ligand-gated ion channelswith different � (�2–10) and � (�2–4) subunits [71]. Multiple sub-types of neuronal nAChR can be formed from various combinationsof subunits and these subtypes can be classified into two major

Author's personal copy
370 Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378
Table 1Frequently used central acetylcholine receptors (AChRs) agonists and antagonists in experimental studies.
Nicotinic AChR Muscarinic AChR
Agonists Antagonists Agonists Antagonists
Carbachol �-bungarotoxin (�7) Carbachol AtropineCytisine (�4�2) Mecamylamine Muscarine PirenzepineDihydro-�-erythroidine D-tubocurarine Oxotremorine sesquifumarate Quinuclidinyl benzilateEpibatidine Methyllycaconitine (�7) ScopolamineNicotine5-iodo-3-(2(S)-azetidinylmethoxy)pyridine dihydrochloride (�2)
Into brackets subtype or subunit specificity.
classes containing either a heteropentameric or a homopentamericstructure. Among the heteromeric receptors, the �4�2 nAChRs arethe predominant subtype in mammalian brain [230], while amongthe homomeric class, the �7 is the one most highly expressed [39].Receptors containing �9 and/or �10 subunits are usually classifiedwith �7 [166].
Nicotinic receptors subunits are among the first membrane pro-teins to appear during CNS development. Functional nAChRs wererevealed by patch-clamp measurements in fetal mouse cerebralcortex as early as on E10 [16]. In the rat cortex, the �2 subunithas been detected on E12–E13 while the �4 subunit is initiallyexpressed on E17–19 [229]. Additionally, the �7 binding sites havebeen observed in the hippocampus and posterior hypothalamusas early as on E12 [200]. The temporal and regional expression ofindividual subunits may also vary during postnatal development.For example, in the rat cortex and hippocampus, �2 mRNA is lowon P7 and increases by P14, whereas �7 mRNA peaks on P7. Cor-tical �4 mRNA peaks on P14 and remains high on P28, whereashippocampal/septal �4 mRNA is higher on P7 and P14 when com-pared to P1 and P28 [184]. The expression of nAChRs is still modifiedlate during postnatal development so that adolescents expressgreater numbers of �4�2* and �7* nAChRs compared to the samebrain regions of adults. [53], even thought there is also evidence ofregion-dependent effects [169]. In the human fetal brain, nAChRsare detected in the first trimester, gradually increasing up to mid-gestation. Even though there is evidence that nAChR levels remainconstant or decrease throughout postnatal life depending on thebrain region and receptor composition [58,59,70,83], there is scantinformation regarding infancy and adolescence.
These changes in nAChR expression occur during critical pre-natal and postnatal periods of neuronal development, involvingmechanisms associated with neurogenesis, cell migration and dif-ferentiation, and synaptogenesis. Accordingly, the critical rolesthese receptors are believed to play in many aspects of brain devel-opment seem to depend on their regional and temporal expression.In addition, as detailed below, distinct nAChRs present particularpermeability to ions and activate distinct signaling pathways, sothat the ability of these receptors to modulate brain development isalso dependent on the pharmacology of their subunit composition.
Although the cation permeability of the subtypes is influ-enced by their subunit composition, the nAChRs are permeableto the monovalent Na+ and K+ ions, as well as to Ca2+. Ca2+
signals are pivotal in shaping nAChR-mediated effects. Two dif-ferent classes of neuronal nAChRs may be identified accordingto their Ca2+ permeability, which correlates with other pharma-cological and structural properties: (1) homopentameric nAChRscontaining subunits (�7–�9) that exhibit the highest measuredCa2+ permeability values; (2) heteropentameric nAChRs, alwayscomprising at least one � (out of �2–�6) and one � (out of �2–�4)subunits, with lower measured Ca2+ permeability [47,64]. Fol-lowing nAChR stimulation, Ca2+ can enter cells either directly,through the intrinsic ion channel, or indirectly following voltage-gated Ca2+ channel activation. Additionally, Ca2+ levels can be
amplified via Ca2+-induced Ca2+ release from intracellular stores[47].
Indeed, Ca2+ influx through nAChR channels has been shownto modulate several Ca2+-sensitive events [47,64]. Interestingly,release of Ca2+ from intracellular stores contributes to nicotine-elicited increases in intracellular Ca2+ [40]. Nicotine was shownto stimulate proliferation of neural progenitor cells derived fromthe P19 murine embryonal carcinoma, and this effect was depen-dent on Ca2+ mobilization from intracellular stores [167]. NicotinicAChRs-mediated mobilization of Ca2+ is also relevant to otherdevelopmental events. At immature synapses from neonatal rathippocampal slices, activation of presynaptic intracellular Ca2+
stores contributes to nicotine-elicited increase of glutamate release[114]. The �7 nAChR is believed to be involved in the regulationof neuronal growth, differentiation and synapse formation duringdevelopment. By modulating intracellular Ca2+ levels, this recep-tor may regulate events such as activation of second messengersystems and induction of immediate-early gene expression [5,30].The �7 nAChRs modulate neurite outgrowth. In this regard, bothstimulatory and inhibitory effects have been described. In the chickciliary ganglion from E8 embryos, where �7 nAChRs have a pre-dominantly nonsynaptic location on neurons, pulses of ACh andnicotine were shown to restrict neurite growth in cell culture andinduce retraction in a Ca2+-dependent way. This effect was blockedby the �7 nAChR antagonist �-bungarotoxin, a poisonous com-ponent present in snake venom [164]. Similar inhibitory effectson neuritic extension mediated by �7 nAChRs were described inPC12 cells [36]. Interestingly, this effect seems to be dependenton the stage of development. For instance, an opposite effect wasdescribed in cultures of dorsal root ganglia neurons obtained fromE12 chick embryos, in which treatment with the agonists muscarineand carbachol elicited longer fibers, a higher number of fibers perneuron and increased neurofilament protein expression. In addi-tion, concomitant use of agonists and antagonists, such as atropineor mecamylamine, counteracted the increase in fiber outgrowth,indicating that neurite outgrowth is mediated by both muscarinicand nicotinic receptors. In contrast, modulated expression of neu-rofilament proteins by cholinergic agonists was not evident on E18,when sensory neurons have completed their differentiation [195].Finally, in a primary culture model of neonatal rat olfactory bulbs,the agonist carbachol significantly increased neuritic length, aneffect that was abolished by co-treatment with �-bungarotoxin,which argues for a positive role for �7 nAChRs in neuritic outgrowthalso in the rat olfactory bulb [44].
Activation of nAChRs by ACh can also affect the behavior ofneuronal growth cones, regulating their morphology, motility andpathfinding. In explant cultures of thalamic neurons, ACh exposurechanged the morphology of growth cones; they became larger andextended more filopodia. A quantitative analysis of growth arearevealed that growth cones increased by almost up to 50% in sizein the presence of ACh in the culture medium [177]. In develop-ing oocytes, d-tubocurarine, a nicotinic AChR antagonist, inhibitedelectric field-induced orientation of cultured neurites toward the

Author's personal copy
Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378 371
cathode, whereas atropine, a muscarinic AChR blocker, markedlyenhanced the guidance properties of the applied field [57].
Variations in the expression of particular nAChR subtypes pro-foundly influence the developmental outcome across differentbrain regions. For instance, cell survival is modulated by nAChRs,and both neurotoxic and neuroprotective effects of nAChRs activa-tion have been described. In this regard, �4�2 nAChRs have beenshown to promote survival, while �7 nAChRs seem to mediateneurotoxic effects of nicotine. Nicotine was shown to be neu-roprotective in vivo, in a rodent model of neonatal excitotoxicbrain injury, and in vitro, in primary cultures of cortical neurons.Inhibition of �4�2 nAChRs by using the antagonist dihydro-�-erythroidine (DH�E) or by using �2−/− knockout mice was shownto abolish the protective effect of nicotine both in vivo and in vitro.In contrast, activation of �7 nAChR was deleterious in neonates,whereas its blockade, either by using the �7 antagonist methyl-lycaconitine (MLA) or by using �7−/− knockout mice providedneuroprotection [108]. A genetic mutation that prolongs the dura-tion of �7 channel opening results in dramatic increases in celldeath, possibly due to increased Ca2+ influx [157]. Berger et al.[19] have shown that �7 nAChR activation by low doses of nico-tine results in apoptotic cell death of primary and immortalizedhippocampal progenitor cells.
Interestingly, neurodevelopmental actions of AChR activationmay also depend on the differentiation state of the cell. Distinctstages of differentiation are associated with differential expressionof nAChRs subunits, which, in turn, were shown to affect cell sur-vival. In HC2S2-immortalized hippocampal progenitors, nicotineis cytotoxic to undifferentiated cells, whereas it spares the samecells once differentiation has been induced. Nicotine, acting vianAChRs, inhibits proliferation in embryonal carcinoma P19 cells,but promotes it following induction of neuronal differentiation andtransition to neural progenitor cells [167]. Evidence from studiesthat used mutant mouse embryos with hypersensitive �4 subunitsshows that the continuous activation of hypersensitive nAChRson neurons in the substantia nigra and ventral tegmental area isassociated with severe reduction of nigral dopaminergic cells asmaturation progresses during the course of prenatal development[105]. In accordance with these data, nicotine was shown to exertpositive trophic effects primarily on undifferentiated pheochro-mocytoma PC12 cells, whereas with differentiation, the effectsundergo a transition to neurotoxicity [2]. Nicotine exposure wasshown to upregulate mRNA expression of growth factors in the hip-pocampus during the first postnatal week of rats, a period of intenseneurogenesis in this region [192], and this upregulation is hypoth-esized to reduce programmed cell death [90]. Finally, increasedapoptotic cell death due to nicotine exposure was identified evenduring the adolescence of rodents, resulting in reduced density ofboth neuronal and glial cell populations. These findings suggest thatnAChR-mediated modulation of cell survival persists at late stagesof brain development. [95,155].
Regulation of gene expression is an important aspect of braindevelopment. Nicotinic AChRs may directly or indirectly lead to thephosphorylation of cAMP response element binding protein (CREB),which results in gene transcription. CREB controls the transcriptionof many gene products, and it is implicated in many impor-tant developmental events [89]. Nicotinic AChR-mediated CREBphosphorylation requires both Ca2+/calmodulin-dependent pro-tein kinase (CAMK) and mitogen-activated protein (MAP) kinaseactivation as well as Ca2+ release from internal stores [37,88]. Inthe chick ciliary ganglion, this activation is opposed by L-typevoltage-gated Ca2+ channels, which induce phosphatase activitythat dephosphorylate CREB, thereby extinguishing the nAChR sig-nal [37]. In contrast, in the developing hippocampus, voltage-gatedCa2+ channels do not cause dephosphorylation of CREB and ter-mination of signal. In fact, nAChR-mediated gene transcription in
the hippocampus has both glutamate-independent and glutamate-dependent components, the latter one possibly resulting frompresynaptic facilitation of glutamate release [88]. The low levels ofsustained nicotinic stimulation required for transcriptional effectsare consistent with those achievable either by the normal septalcholinergic innervation of the hippocampus or by repeated tobaccoexposure [88].
Interestingly, both nicotine administration and exposure to anenvironment previously associated with rewarding properties ofnicotine results in an increase of CREB phosphorylation in sev-eral brain regions [31,208]. Nicotine activates a broad array ofnAChRs subtypes. Therefore, it excites different types of neurons,thus perturbing baseline neuronal communication, interferingwith synaptic properties and modulating synaptic plasticity. [131].These nicotine-elicited alterations seem to reorganize patterns ofsynaptic connectivity which, in turn, alters the operation of brainregions involved in incentive motivation and reward (such as thenucleus accumbens), judgment and inhibitory control of behavior(such as the prefrontal cortex), thus contributing to some of thepersistent sequela associated with drug use, including addiction[171].
Among the different types of neuronal plasticity, CREB has beenimplicated in those associated with learning and memory [189]. Inthis regard, long-term changes seen after in utero nicotine exposurein rodents reproduce behavioral deviations that are seen in chil-dren of smokers, including cognitive impairment [160,202]. Indeed,the period of susceptibility to nicotine extends into adolescence,since learning/memory detrimental effects of adolescent nicotineexposure were also described in mice [1].
Studies in animal models have also shown that ubiquitouslydistributed presynaptic nAChRs serve as ligand-gated ion chan-nels that promote neurotransmitter release. Thus, nAChR appearto exert a predominantly modulatory influence on brain mecha-nisms. Both Na+ and Ca2+ fluxes associated with nAChR activationcan influence neurotransmitter release [216]. In fact, the release ofclassical neurotransmitters such as dopamine, noradrenaline, ACh,glutamate and GABA are facilitated by nicotine exposure [215]. Theabove-mentioned facilitation of neurotransmitter release mediatedby nAChRs is believed to be relevant to the proper maturation ofseveral neurotransmitter systems. In this regard, during the earlypostnatal period of rats, most glutamatergic synapses in the hip-pocampus are immature and functionally silent. However, bothACh and nicotine strongly enhance neurotransmitter release andconvert presynaptically silent synapses into conductive ones viapresynaptic �7 nAChR-mediated increases in Ca2+ [128]. It is likelythat a similar facilitation of neurotransmitter release occurs inother brain regions and during distinct phases of developmentand that presynaptic nAChRs determine the maturation of distinctneurotransmitter systems. Besides, the inappropriate activationof presynaptic nAChRs may be an important mechanism throughwhich cholinergic agonists such as nicotine disturb several aspectsof brain development.
3.2. Muscarinic AChRs
Five subtypes of mAChRs, M1, M2, M3, M4 and M5 have beendescribed. The receptor protein is composed of seven transmem-brane domains. The third inner cytoplasmic mAChR loop interactswith the G-protein and this portion of the protein is believed toinfer different functional characteristics to the receptor [26,35,103].Distinct signaling pathways are activated depending on the sub-type of mAChR. The M2 and M4 subtypes were shown to inhibitadenylyl cyclase activity through activation of a pertussis toxin(PTX)-sensitive inhibitory G-protein (Gi), a mechanism that leadsto a reduction in the levels of cyclic AMP. In turn, M1, M3 andM5 subtypes are generally coupled to a PTX-insensitive G-protein

Author's personal copy
372 Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378
(Gq) which activates phospholipase C. Activation of phospholipaseC leads to the formation of inositol 1,4,5-trisphosphate (IP3) anddiacylglycerol (DAG). IP3 causes mobilization of Ca2+ from intracel-lular stores, while DAG activates protein kinase C (PKC) isozymes[45,206]. Muscarinic AChRs activation modulates several types ofion channels: For example, M1 and M2 receptors in the mouseand M1 and M4 receptors in the rat depress Ca2+ currents whilemuscarinic suppression of the M-type K+ current in the rat is alsomediated by M1 receptors [185]. MAP kinases and small GTPases,such as rho and rac, are also activated by mAChRs [45,206].
M1, M2 and M4 mAChRs are the predominate mAChR sub-types expressed in the adult brain. As examples, approximately40% of total mAChRs in the cortex of the rodent brain representM1, 37% M2 and approximately 15% M4. In the hippocampus, 36%represent M1, 33% M2 and 27% M4 [115]. The mAChR is presentin both the forebrain and the hindbrain during early develop-mental stages. Binding of [3H]-quinuclidinyl benzilate (QNB), anon-selective mAChR antagonist, has been demonstrated in humanfetal brain from early gestation [75]. In the rat, mAChR binding sitesappear in the spinal cord on E13. On E15, binding sites are evi-dent in the rhombencephalon and mesencephalic structures andon E16, in forebrain regions [182]. There is evidence of changesin receptor expression during postnatal life. As an example, in thevisual cortex of rats, the laminar distribution of the mAChRs mRNAvaries depending on the receptor subtype and on the age. In general,there are increases in mRNA expression during postnatal develop-ment and the final adult distribution is achieved during adolescence[174,175]. In the cat, both M1 and M2 subtypes of AChRs bindingsites are detected in the striatum as early as on E40 and in thesubstantia nigra on E45 (birth on E65). During fetal and perinataldevelopment, overall muscarinic binding increases in the striatumbut is stable in the substantia nigra [148,149].
When compared to nAChRs, much less is known about what theactivation of M1–M5 receptors does in the brain [162]. However,studies on the effects of mAChRs agonists and antagonists havealready demonstrated that these receptors mediate several effectsof ACh in the developing CNS. In this regard, mAChRs have beenshown to mediate effects of ACh on proliferation, differentiationand survival. In embryonic P19 carcinoma cells, a widely acceptedin vitro model for early neurogenesis, muscarine induces prolifer-ation in progenitor cells by activation of M1, M3 and M5 receptorsand mobilization of intracellular Ca2+ stores, whereas M2 receptoractivity mediates neuronal differentiation [167,168]. Neuroepithe-lial cells in the ventricular and subventricular zones of embryonicrat cortex express M2, M3 and M4 subtypes of mAChRs and acti-vation of mAChRs stimulates proliferation of not only multipotentprecursor cells, but also those cells progressing along neuronal lin-eages [125,126]. The exact intracellular mechanisms underlyingmAChR-modulated DNA synthesis and neurogenesis are not fullyunderstood. However, several pathways through PTX-sensitive G-proteins, cytosolic Ca2+ signaling, protein kinase C, MAP kinasephosphorylation, phosphatidylinositol-3 kinase (PI-3K) and c-Srchave been shown to play roles [125,126]. Li et al. [118] demon-strated that stimulation of mAChRs with the agonist carbacholactivates the extracellular signal-regulated kinase (ERK)1/2 and PI-3K. This, in turn, stimulates DNA synthesis in neural progenitorcells. These findings demonstrate that the PI-3K and MAP kinasesignaling pathways via mAChRs are involved in neural progenitorcell proliferation during early neurogenesis. [118]. The nonreceptorprotein tyrosine kinase c-src activity was also shown to be requiredfor the mAChR-mediated proliferation and neurogenesis in neuralprecursor cells via ERK1/2 and CREB [224]. Moreover, the mAChRantagonist atropine blocks the enhancement and inhibits the sub-sequent differentiation of progenitor cells [227]. The hippocampusis one of the few brain regions that maintains neurogenesis duringpostnatal life and newborn cells in the hippocampus of adult rats
express both M1 and M4 subtypes of mAChRs [142]. Lesion of septalcholinergic nuclei projecting to hippocampus suppresses prolifera-tion and short-term survival of newborn cells in the dentate gyrus,with a concurrent impairment in spatial memory [142]. In mousehippocampal slices, the activation of ERK, which is crucial for manyneural functions, including learning, memory, and synaptic plas-ticity, is mediated by the M1 mAChR subtype [20]. Donepezil, aselective AChE inhibitor, enhances survival of newborn neurons inthe hippocampal dentate gyrus and scopolamine, a mAChR blocker,suppresses it. Furthermore, donepezil enhances and scopolaminesuppresses phosphorylation of CREB, which is involved in cell sur-vival in this brain region. These results suggest that the activationof the central cholinergic transmission enhances the survival ofnewborn neurons in the dentate gyrus via CREB signaling. [102].Activation of mAChRs also plays important roles in the proliferationof rat cortical astrocytes [15,76,217] and oligodendrocyte progen-itor cells [41].
Neurite outgrowth is also modulated by mAChRs. In thalamusexplant cultures from E14 mice, application of ACh reduces thelength of axons in a dose dependent manner, an effect mediatedby both nicotinic and muscarinic AChRs, but with strongest effectsmediated by mAChR, as indicated by adding either nicotinic or mus-carinic agonists to the medium [177]. In the prenatal hippocampus,cholinergic stimulation of pyramidal neurons through the M1 sub-type of mAChR accelerates axonal growth through the inductionof Ca2+ mobilization and the activation of PKC and of ERK1/2[204]. Additionally, M3 mAChRs expressed in rat cortical astro-cytes stimulate neurite outgrowth in rat hippocampal neurons.These neurons, co-cultured with astrocytes previously exposed tothe cholinergic agonist carbachol, display longer neurites, whichdemonstrates that the release of permissive factors from astrocytesaccelerates neuronal development [77].
Furthermore, mAChR activity is implicated in neuronal plas-ticity, including long-term potentiation (LTP). Both M1 and M2/4subtypes of mAChR are involved in LTP induction in the dentategyrus of rats [119,124], while M2/4 subtypes regulate LTP in themouse visual cortex [156]. Evidence is also available indicating thatphosphorylation of CREB, activation of MAP kinase and ERK1/2 playroles in mAChR-mediated neuronal plasticity [68,74,80,173].
4. Acetylcholinesterase
AChE catalyses the hydrolysis of ACh, thereby interruptingcholinergic activity in the synapse. Mammalian AChE appears tobe encoded by a single gene [197]. In humans, the AChE gene islocated in chromosome 7 and in mice it is located in chromo-some 5 [67,165]. In most species, the AChE gene encodes a catalyticsubunit of about 75 kDa [176]. After translation, AChE moleculesfold forming functional monomers and assemble into disulfide-bonded dimers. The dimers can be induced to tetramerize by thenon-catalytic subunit “p” peptide in the case of neurons, or the col-lagenic ColQ protein in the case of skeletal muscle. The oligomericforms are then transported through the cells and released intothe synaptic cleft where they may attach to available sites on thespecialized synaptic basal lamina [176]. Two alternatively splicedisoforms of AChE that are particularly relevant for the nervoussystem have been well documented: the tetrameric AChE-S iso-form (‘S’, synaptic) and the ‘read-through’ variant of AChE (AChE-R)[134]. Although the enzymatic activity of the two isoforms is equiv-alent, they do differ in aspects such as ontogenetic expression,multimeric assembly, cellular localization and tissue specificity.Interestingly, it seems that AChE has an additional developmentallyregulated enzymatic activity as an aryl acylamidase [27].
AChE appears quite early during embryonal development of theCNS. In fact, its mRNA is already expressed by progenitor cells in

Author's personal copy
Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378 373
the ventricular zone (VZ) and undifferentiated cells in the corti-cal plate (CP) [52]. At the beginning of neurogenesis in mice (E11),which is characterized by remarkable progenitor cell proliferation,AChE mRNA is co-localized in most VZ cells and its expression ismost intense in the apical portion, next to the ventricular lumen.At E13, clusters of adjoining cells in the basal portion of the VZpresent more pronounced cytoplasmic expression of AChE. By E15,AChE-expressing cell clusters are becoming progressively smallerso that by E17 they are limited to very small clusters or singlecells. In the intermediate zone, AChE mRNA expression, albeit lessintense than in the VZ, is observed in individual cells or in clustersof cells. These cells are radially oriented, indicating that they aremigrating from the VZ to the CP. With further development, AChEexpression in the CP appears in clusters of intensely labeled cells.These cells are more prominent in the superficial portion comparedwith the deep CP, suggesting that younger, newly arriving cells inthe CP express more AChE transcripts than cells that had alreadyundergone some differentiation. Subsequently, at E17, the intensesignals in the superficial cell layer of the CP become significantlyhigher than those of the deep CP portion [52]. In general, total AChEactivity increases about 15-fold between E9 and E19 [92]. Duringpostnatal development, up to P30, AChE activity increases in a waythat varies dramatically from one brain region to another [143,180].In humans, AChE activity also increases postnatally until it becomesfully established during adulthood [135].
In addition to this classic enzymatic role, AChE also hassome non-classical properties concerning SNC development. Forinstance, it is now accepted that AChE has functions associ-ated with adhesion, neurite growth, circuitry formation and evenapoptosis [96,113,186,188,193,223]. For instance, it was demon-strated that AChE expression correlates directly to the level ofcell–substratum adhesion of neuroblastoma cells [187]. Further-more, AChE forms a complex with amyloid precursor protein andperlecan that seems to be involved in substratum adhesion andpolarized migration of adherent cells [9]. That AChE has adhesiveproperties was also demonstrated in non-neuronal cells such asosteoblasts: blockade of some sites in the AChE molecule results ina concentration-dependent decrease in osteoblastic cell adhesion[66,93].
The adhesive properties of AChE may also help to explainanother of its non-classical roles: stimulation of neurite outgrowth.In fact, this was the first non-classical activity of AChE to beunequivocally identified [112]. As indicated above, AChE expres-sion occurs early in postmitotic neurons [111] and precedes theoutgrowth of neuritic processes [110,210]. Blocking the enzymaticactivity of AChE does not affect its role in promoting neurite growth[112]. The effects of AChE in stimulating neurite growth weredemonstrated in studies that used overexpression of this enzyme indorsal root ganglion neurons and neuroblastoma cells [24,101]. Theimportance of AChE to axon tract formation was also demonstratedin vivo: the thickness of the major longitudinal axon tract of theXenopus brain, the tract of the post-optic commissure, is reducedafter treatment with BW284C51, which blocks non-catalytic sitesin the AChE molecule [10].
In trying to explain how AChE exerts this non-classical activ-ity during development, the association between AChE and othermolecules has been a focus of study. For example, laminin, which isknown to bind to AChE [97,159], is an extracellular matrix pro-tein that participates in neuronal differentiation and adhesion.Although the exact mechanism that explains how AChE affectslaminin activity during development is yet to be described, it iswell established that laminin facilitates the migration of neuronalprecursors to their proper regions in the brain. It also elicits anddirects neurite outgrowth [73,121,170,172,190,228], possibly byinteracting with integrins, one of the most abundant family ofreceptors for molecules located in the extracellular matrix [106].
Curiously, classical and non-classical roles of AChE may associateduring development. ACh is known to arrest neurite growth afterbeing released from the growing tips. By metabolizing ACh, a clas-sical role, AChE could prevent this arresting effect, while, at thesame time, directing neurite growth toward its target through itsadhesive, non-classical, properties.
Studies using AChE knockout mice (AChE−/−) are also beginningto provide information concerning the relevance of this enzyme tothe normal development of the nervous system. In these strains,a series of adaptations allow animals to have brain and mus-cle functions that are compatible with long-term survival despitethe complete lack of AChE and abnormally high levels of ACh intheir synapses [38,54,82]. Interestingly, these animals are capableof forming central cholinergic pathways [136]. Some of the fac-tors that let animals survive postnatally are: The presence of theenzyme butyrylcholinesterase (BChE), which also metabolizes AChin the synapses; downregulation of acetylcholine (nicotinic andmuscarinic) and dopamine receptors; and morphological remod-eling of the endplate region [4,50,69,87,117]. It must be pointedout, however, that AChE−/− mice have significant behavioral andmotor deficits [55]. The analysis of the retina of AChE−/− miceindicated that AChE is fundamental to the normal development ofthis structure in a way that is unrelated to its enzymatic function,since BChE content was normal [33]. In these animals, the develop-ment of the laminar structure of the retina is disturbed during thefirst two postnatal weeks. The resulting anomalous circuits thatare assembled are not capable of sustaining photoreceptor activ-ity and long-term viability, in spite of a seemingly normal initialdevelopment of these cells. By three months, all photoreceptors,as well as cells from the inner retina, are lost due to apoptosis.In an additional study, neurons derived from dorsal root gangliaof embryonic AChE−/− and AChE+/+ mice indicated that axonalgrowth inhibition by organophosphorus pesticidesis is not relatedto the enzymatic activity of AChE [218].
5. High-affinity choline transporter
As previously mentioned, most of the ACh that is releasedinto the synaptic cleft is rapidly hydrolyzed by AChE intocholine and acetate. ACh hydrolysis interrupts cholinergic signal-ing and the resulting choline is readily transported back into thepresynaptic terminal, through the membrane, by a high-affinity,Na+-dependent, hemicholinium-3 sensitive (HC-3, a bicyclic, con-strained choline analog) mechanism. The CHT activity, supplyingthe choline that will be used by ChAT, is the rate-limiting step in thebiosynthesis of ACh. The ACh is then loaded into synaptic vesiclesby the VAChT [8,61,179]. The majority of the choline transportedby the CHT is efficiently converted to ACh [78,79,145], suggestingit is both metabolically linked and physically proximal to sites ofACh synthesis.
The CHT genes identified in mice, rats, and humans are respec-tively located on chromosomes 17, 9, and 2 [12,13]. The CHTproteins synthesized from these genes have thirteen transmem-brane domains with an extracellular N terminus and a cytoplasmicC terminus [13,201]. The development of CHT-specific antibod-ies has allowed the direct, activity-independent detection of CHTs[62,100,104,122,140]. Different pools of CHTs can be identifiedwithin cholinergic neurons [62,104,122]. At the cell body level,abundant CHT immunoreactivity is evident in association with therough endoplasmic reticulum membranes. This biosynthetic poollikely accounts for the CHT signal associated with cholinergic soma,because CHT immunoreactivity is not abundant at the cell somaplasmalemma. In contrast, CHT immunoreactivity is evident at theplasma membrane of presynaptic terminals, in line with its role insupport of ACh synthesis [32,194]. Interestingly, by far the greatest

Author's personal copy
374 Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378
density of CHT labeling is surprisingly found to be associated withvesicles located inside the terminal.
In spite of its role in cholinergic systems, only a handful ofstudies have addressed developmental aspects of CHT, particularlyin the CNS. In rats [81], on E18 and right after birth, CHT levels,as demonstrated by a quantitative autoradiography method thatused [3H]HC-3, were high only in the medial habenula. In mostbrain regions, including the striatum, nucleus accumbens, olfac-tory tubercle, cortex, and hippocampus, significant increases in CHTlevels occurred primarily during the postnatal period. Although sig-nificant variations existed between brain regions, adult levels ofCHT were reached between P 15 and P21, with 2–12-fold increasesbeing observed. Differences between brain regions regarding CHTlevels is further emphasized in another study [221]. In this study, aperiod of comparatively high levels of [3H]HC-3 binding in the cere-bral cortex and midbrain + brain stem observed before birth wasfollowed by marked reductions during the first postnatal week andsubsequent increases, faster in the midbrain + brain stem, towardadult levels by the sixth postnatal week. Regarding the hippocam-pus and the striatum, the peak levels were observed during the 5thand 6th postnatal weeks and marked reductions toward adult lev-els were observed from then on. In mice, [23], the earliest, E14, andstrongest CHT mRNA expression was found in the spinal cord. Inter-estingly, mRNA levels remained more or less constant up to P30in this structure. Conversely, in the septum and striatum, a steadyincrease, faster in the former one, in mRNA expression, from hardlydetectable on E14 to a high level on P30, was observed. In the cere-bellum, the levels of CHT mRNA expression reached the highestlevel still during gestation, on E19, a peak that was followed by arapid decline from then on. Regarding the cortex and the hippocam-pus, mRNA levels remained low, with slight variations, from E14 toP30. By P30 CHT mRNA levels were highest in the spinal cord, stria-tum and septum. Lower levels were observed for the cortex andhippocampus and even lower levels were observed for the cere-bellum. A more recent study [133], using rats and restricted to thepostnatal development of the hippocampus, indicated that duringthe first postnatal week CHT mRNA expression rapidly increasedand then stabilized from P8 to P24. This period of stability was fol-lowed by a period of marked decrease in CHT mRNA levels in whichthe lowest level was reached by P34. From then on, a slight increasefollowed by a plateau (at 50% of peak levels) was observed up to P90.As for the CHT protein [23], it was detected in the spinal cord, sep-tum, striatum, cerebellum cortex and hippocampus as early as E14.However, levels were generally small. Significant accumulation ofthe protein was detected in the spinal cord by P8, in the striatum byP15 and in the septum by P30. Taken together, the aforementionedresults in mice indicate that, in most regions, protein accumulationis delayed in relation to mRNA expression.
As demonstrated in CHT−/− mice, the absence of CHT is notlethal during gestation but results in newborn animals that arehypoxic, immobile and that die within an hour after birth, pos-sibly due to the lack of ACh in cholinergic synapses that supportrespiration [60], an observation that corroborates previous find-ings that indicate that CHT function is necessary for sustained AChproduction and release [25,98]. Interestingly, these mice are mor-phologically normal an equivalent in weight when compared toCHT+/+ mice and CHT+/−. Furthermore, no differences in mor-phology and weight were observed concerning various organs,including the brain. Nissl staining also failed to indicate defectsin brain development in the CHT−/− mice. Conversely, CHT+/−mice, which display a 50% reduction in CHT protein levels, live intoadulthood and are able to reproduce. In general, these mice do notpresent significant differences in behavior (motor co-ordination,general activity, anxiety and spatial learning and memory) whencompared to CHT+/+ ones. On the other hand, a physical challenge,such as that provided by a treadmill paradigm, is able to uncover
impairments in performance in CHT+/− mice [18]. Therefore, thefew studies that are available concerning developmental aspectsof CHT in the SNC indicate that this protein is not directly criti-cal for the normal SNC development during the gestational period.The low cholinergic activity observed during this period can bemaintained by the low-affinity, Na+ independent, HC-3-insensitivecholine transport mechanism. However, CHT is indirectly respon-sible for normal SNC development after birth, since the increaseddemand on the cholinergic systems, associated with the control ofbreathing for example, depends on a constant supply of cholineto the presynaptic terminals that can only be maintained by thistransporter system.
6. Future directions
Despite the fact that there is plenty of information on severalaspects of the development of the cholinergic system, there arescant studies that aimed to investigate whether or how the devel-opment of each component of this system affects the others, i.e.to integrate information regarding all components of the system.Perhaps most importantly, as evidenced in this review (Fig. 2), thecomponents of the cholinergic system are responsible for executinginter-related events and developmental events are frequently influ-enced by more than one of these components. This suggests that theequilibrium between them is essential for their functions, there-fore, future studies are necessary to investigate how the complexnet formed by the components of the cholinergic system (whichmight go beyond the effect of each component), determines thenormal development of the CNS and how imbalances elicited forexample by environmental agents affect it.
Indeed, exposure to environmental agents that affect the cholin-ergic system may seriously compromise brain development andhave long-lasting morphologic, neurochemical, and functionalconsequences. Interestingly, the investigation of the effects ofneurotoxins is also a useful tool to identify the roles of com-ponents of this system on brain development. Until recently,most studies on developmental aspects of the cholinergic sys-tem focused on the prenatal and neonatal periods, however,it is now clear that, in several aspects, the adolescent brainis particularly vulnerable to neurotoxins. In this regard, recentstudies show that adolescents present a peculiar sensitivity tonicotine [3,191] and the fact that the maturation of central cholin-ergic system is consolidated during the periadolescent period[53,132,135,147,221] may explain, in part, this sensitivity. Accord-ingly, there has been an increasing number of studies on theeffects of neurotoxins during adolescence, which in the next fewyears may fill the gap of information regarding this period ofdevelopment.
References
[1] Abreu-Villaca Y, Medeiros AH, Lima CS, Faria FP, Filgueiras CC, Manhães AC.Combined exposure to nicotine and ethanol in adolescent mice differentiallyaffects memory and learning during exposure and withdrawal. Behav BrainRes 2007;181:136–46.
[2] Abreu-Villaca Y, Seidler FJ, Qiao D, Slotkin TA. Modeling the developmentalneurotoxicity of nicotine in vitro: cell acquisition, growth and viability in PC12cells. Dev Brain Res 2005;154(2):239–46.
[3] Abreu-Villaca Y, Seidler FJ, Tate CA, Slotkin TA. Nicotine is a neurotoxinin the adolescent brain: critical periods, patterns of exposure, regionalselectivity, and dose thresholds for macromolecular alterations. Brain Res2003;979:114–28.
[4] Adler M, Manley HA, Purcell AL, Deshpande SS, Hamilton TA, Kan RK,et al. Reduced acetylcholine receptor density, morphological remodeling,and butyrylcholinesterase activity can sustain muscle function in acetyl-cholinesterase knockout mice. Muscle Nerve 2004;30:317–27.
[5] Albuquerque E, Pereira E, Castro N, Alkondon M, Reinhardt S, Schroder H, et al.Nicotinic receptor function in the mammalian central nervous system. AnnNY Acad Sci 1995;757:48–72.

Author's personal copy
Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378 375
[6] Alderson RF, Alterman AL, Barde YA, Lindsay RM. Brain-derived neurotrophicfactor increases survival and differentiated functions of rat septal cholinergicneurons in culture. Neuron 1990;5:297–306.
[7] Allen TG, Brown DA. Detection and modulation of acetylcholine release fromneurites of rat basal forebrain cells in culture. J Physiol 1996;492:453–66.
[8] Amenta F, Tayebati SK. Pathways of acetylcholine synthesis, transport andrelease as targets for treatment of adult-onset cognitive dysfunction. CurrMed Chem 2008;15:488–98.
[9] Anderson AA, Ushakov DS, Ferenczi MA, Saffel JL. Morphoregulation by acetyl-cholinesterase in fibroblasts and astrocytes. J Cell Physiol 2008;215:82–100.
[10] Anderson RB, Key B. Role of acetylcholinesterase in the development ofaxon tracts within the embryonic vertebrate brain. Int J Dev Neurosci1999;17:787–93.
[11] Antonov I, Chang S, Zakharenko S, Popov SV. Distribution of neurotransmittersecretion in growing axons. Neuroscience 1999;90:975–84.
[12] Apparsundaram S, Ferguson SM, Blakely RD. Molecular cloning and character-ization of a murine hemicholinium-3-sensitive choline transporter. BiochemSoc Trans 2001;29(Pt 6):711–6.
[13] Apparsundaram S, Ferguson SM, George Jr AL, Blakely RD. Molecular cloning ofa human, hemicholinium-3-sensitive choline transporter. Biochem BiophysRes Commun 2000;276(3):862–7.
[14] Armstrong DM, Bruce G, Hersh LB, Gage FH. Development of choliner-gic neurons in the septal/diagonal band complex of the rat. Brain Res1987;433:249–56.
[15] Ashkenazi A, Ramachandran J, Capon DJ. Acetylcholine analogue stimulatesDNA synthesis in brain-derived cells via specific muscarinic receptor sub-types. Nature 1989;340:146–50.
[16] Atluri P, Fleck MW, Shen Q, Mah SJ, Stadfelt D, Barnes W, et al. Functionalnicotinic acetylcholine receptor expression in stem and progenitor cells ofthe early embryonic mouse cerebral cortex. Dev Biol 2001;240:143–56.
[17] Barde YA. The nerve growth factor family. Prog Growth Factor Res1990;2:237–48.
[18] Bazalakova MH, Wright J, Schneble EJ, McDonald MP, Heilman CJ, Levey AI,et al. Deficits in acetylcholine homeostasis, receptors and behaviors in cholinetransporter heterozygous mice. Genes Brain Behav 2007;6(5):411–24.
[19] Berger F, Gage FH, Vijayaraghavan S. Nicotinic receptor-induced apoptoticcell death of hippocampal progenitor cells. J Neurosci 1998;18:6871–81.
[20] Berkeley JL, Gomeza J, Wess J, Hamilton SE, Nathanson NM, Levey AI. M1 mus-carinic acetylcholine receptors activate extracellular signal-regulated kinasein CA1 pyramidal neurons in mouse hippocampal slices. Mol Cell Neurosci2001;18:512–24.
[21] Bernacer J, Prensa L, Gimenez-Amaya JM. Cholinergic interneurons are differ-entially distributed in the human striatum. PLoS One 2007;2:e1174.
[22] Berse B, Lopez-Coviella I, Blusztajn JK. Activation of TrkA by nerve growthfactor upregulates expression of the cholinergic gene locus but attenuates theresponse to ciliary neurotrophic growth factor. Biochem J 1999;342:301–8.
[23] Berse B, Szczecinska W, Lopez-Coviella I, Madziar B, Zemelko V, Kaminski R,et al. Expression of high affinity choline transporter during mouse develop-ment in vivo and its upregulation by NGF and BMP-4 in vitro. Dev Brain Res2005;157(2):132–40.
[24] Bigbee JW, Sharma KV, Chan EL, Bogler O. Evidence for the direct role of acetyl-cholinesterase in neurite outgrowth in primary dorsal root ganglion neurons.Brain Res 2000;861:354–62.
[25] Birks RI, MacIntosh FC. Acetylcholine metabolism of a sympathetic ganglion.Can J Biochem Physiol 1961;39:787–827.
[26] Bonner TI, Buckley NJ, Young AC, Brann MR. Identification of a family of mus-carinic acetylcholine receptor genes. Science 1987;237:527–32.
[27] Boopathy R, Layer PG. Aryl acylamidase activity on acetylcholinesterase ishigh during early chicken brain development. Protein J 2004;23:325–33.
[28] Brady DR, Phelps PE, Vaughn JE. Neurogenesis of basal forebrain cholinergicneurons in rat. Brain Res Dev Brain Res 1989;47:81–92.
[29] Brandon EP, Lin W, D’Amour KA, Pizzo DP, Dominguez B, Sugiura Y, et al.Aberrant patterning of neuromuscular synapses in choline acetyltransferase-deficient mice. J Neurosci 2003;23:539–49.
[30] Broide RS, Leslie FM. The a7 nicotinic acethylcholine receptor in neuronalplasticity. Mol Neurobiol 1999;20:1–16.
[31] Brunzell DH, Mineur YS, Neve RL, Picciotto MR. Nucleus accumbens CREBactivity is necessary for nicotine conditioned place preference. Neuropsy-chopharmacology 2009;34:1993–2001.
[32] Bussiere M, Vance JE, Campenot RB, Vance DE. Compartmentalization ofcholine and acetylcholine metabolism in cultured sympathetic neurons. JBiochem 2001;130:561–8.
[33] Bytyqi AH, Lockridge O, Duysen E, Wang Y, Wolfrum U, Layer PG. Impaired for-mation of the inner retina in an AChE knockout mouse results in degenerationof all photoreceptors. Eur J Neurosci 2004;20:2953–62.
[34] Castell X, Diebler MF, Tomasi M, Bigari C, De GS, Berrard S, et al. More thanone way to toy with ChAT and VAChT. J Physiol Paris 2002;96:61–72.
[35] Caulfield MP. Muscarinic receptors: characterization, coupling and function.Pharmacol Ther 1993;58:319–79.
[36] Chan J, Quik M. A role for the nicotinic a bungarotoxin receptor in neuriteoutgrowth of PC12 cells. Neuroscience 1993;56:441–51.
[37] Chang KT, Berg DK. Voltage-gated channels block nicotinic regulation of CREBphosphorylation and gene expression in neurons. Neuron 2001;32:855–65.
[38] Chatonnet F, Boudinot E, Chatonnet A, Taysse L, Daulon S, Champagnat J, et al.Respiratory survival mechanisms in acetylcholinesterase knockout mouse.Eur J Neurosci 2003;18:1419–27.
[39] Chen D, Patrick JW. The alphabungarotoxin-binding nicotinic acetylcholinereceptor from rat brain contains only the alpha7 subunit. J Biol Chem1997;272:24024–9.
[40] Chin JH, Tse FW, Harris K, Jhamandas JH. Beta-amyloid enhances intracellularcalcium rises mediated by repeated activation of intracellular calcium storesand nicotinic receptors in acutely dissociated rat basal forebrain neurons.Brain Cell Biol 2006;35:173–86.
[41] Cohen RI, Molina-Holgado E, Almazan G. Carbachol stimulates c-fos expres-sion and proliferation in oligodendrocyte progenitors. Mol Brain Res1996;43:193–201.
[42] Consonni S, Leone S, Becchetti A, Amadeo A. Developmental and neuro-chemical features of cholinergic neurons in the murine cerebral cortex. BMCNeurosci 2009;10:18.
[43] Contestabile A, Virgili M, Barnabei O. Developmental profiles of cholinergicactivity in the habenulae and interpeduncular nucleus of the rat. Int J DevNeurosci 1990;8:561–4.
[44] Coronas V, Durand M, Chabot JG, Jourdan F, Quirion R. Acetylcholine inducesneuritic outgrowth in rat primary olfactory bulb cultures. Neuroscience2000;98:213–9.
[45] Costa LG, Guizzetti M, Oberdoerster J, Yagle K, Costa-Mallen P, Tita B, et al.Modulation of DNA synthesis by muscarinic cholinergic receptors. GrowthFactors 2001;18:227–36.
[46] Coyle JT, Yamamura HI. Neurochemical aspects of the ontogenesis of cholin-ergic neurons in the rat brain. Brain Res 1976;118:429–40.
[47] Dajas-Bailador F, Wonnacott S. Nicotinic acetylcholine receptors and the reg-ulation of neuronal signalling. Trends Pharmacol Sci 2004;25:317–24.
[48] de Castro BM, De JX, Martins-Silva C, Lima RD, Amaral E, Menezes C, et al. Thevesicular acetylcholine transporter is required for neuromuscular develop-ment and function. Mol Cell Biol 2009;29:5238–50.
[49] de Castro BM, Pereira GS, Magalhaes V, Rossato JI, De JX, Martins-Silva C, et al.Reduced expression of the vesicular acetylcholine transporter causes learningdeficits in mice. Genes Brain Behav 2009;8:23–35.
[50] Decossas M, Bloch B, Bernard V. Trafficking of the muscarinic m2 autore-ceptor in cholinergic basalocortical neurons in vivo: differential regulationof plasma membrane receptor availability and intraneuronal localiza-tion in acetylcholinesterase-deficient and -inhibited mice. J Comp Neurol2003;462:302–14.
[51] Descarries L, Aznavour N, Hamel E. The acetylcholine innervation of cerebralcortex: new data on its normal development and its fate in the hAPP(SW,IND)mouse model of Alzheimer’s disease. J Neural Transm 2005;112:149–62.
[52] Dori A, Cohen J, Silverman WF, Pollack Y, Soreq H. Functional manipulationsof acetylcholinesterase splice variants highlight alternative splicing contri-butions to murine neocortical development. Cereb Cortex 2005;15:419–30.
[53] Doura MB, Gold AB, Keller AB, Perry DC. Adult and periadolescent rats differin expression of nicotinic cholinergic receptor subtypes and in the responseof these subtypes to chronic nicotine exposure. Brain Res 2008;1215:40–52.
[54] Duysen EG, Lockridge O. Phenotype comparison of three acetylcholinesteraseknockout strains. J Mol Neurosci 2006;30:91–2.
[55] Duysen EG, Stribley JA, Fry DL, Hinrichs SH, Lockridge O. Rescue of theacetylcholinesterase knockout mouse by feeding a liquid diet; phenotypeof the adult acetylcholinesterase deficient mouse. Brain Res Dev Brain Res2002;137:43–54.
[56] Eiden LE. The cholinergic gene locus. J Neurochem 1998;70:2227–40.[57] Erskine L, McCaig CD. Growth cone neurotransmitter receptor activation
modulates electric field-guided nerve growth. Dev Biol 1995;171:330–9.[58] Falk L, Nordberg A, Seiger A, Kjaeldgaard A, Hellstrom-Lindahl E. The alpha7
nicotinic receptors in human fetal brain and spinal cord. J Neurochem2002;80:457–65.
[59] Falk L, Nordberg A, Seiger A, Kjaeldgaard A, Hellström-Lindahl E. Higherexpression of alpha7 nicotinic acetylcholine receptors in human fetal com-pared to adult brain. Dev Brain Res 2003;142:151–60.
[60] Ferguson SM, Bazalakova M, Savchenko V, Tapia JC, Wright J, Blakely RD.Lethal impairment of cholinergic neurotransmission in hemicholinium-3-sensitive choline transporter knockout mice. Proc Natl Acad Sci USA2004;101(23):8762–7.
[61] Ferguson SM, Blakely RD. The choline transporter resurfaces: new roles forsynaptic vesicles? Mol Interv 2004;4(1):22–37.
[62] Ferguson SM, Savchenko V, Apparsundaram S, Zwick M, Wright J, Heilman CJ,et al. Vesicular localization and activity-dependent trafficking of presynapticcholine transporters. J Neurosci 2003;23(30):9697–709.
[63] Frotscher M, Vida I, Bender R. Evidence for the existence of non-GABAergic, cholinergic interneurons in the rodent hippocampus. Neuro-science 2000;96:27–31.
[64] Fucile S. Ca2+ permeability of nicotinic acetylcholine receptors. Cell Calcium2004;35:1–8.
[65] Furusho M, Ono K, Takebayashi H, Masahira N, Kagawa T, Ikeda K, et al.Involvement of the Olig2 transcription factor in cholinergic neuron devel-opment of the basal forebrain. Dev Biol 2006;293:348–57.
[66] Genever PG, Birch MA, Brown E, Skerry TM. Osteoblast-derived acetyl-cholinesterase: a novel mediator of cell–matrix interactions in bone? Bone1999;24:297–303.
[67] Getman DK, Eubanks JH, Camp S, Evans GA, Taylor P. The human gene encod-ing acetylcholinesterase is located on the long arm of chromosome 7. Am JHum Genet 1992;51:170–7.
[68] Giovannini MG. The role of the extracellular signal-regulated kinase pathwayin memory encoding. Rev Neurosci 2006;17:619–34.

Author's personal copy
376 Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378
[69] Girard E, Bernard V, Minic J, Chatonnet A, Krejci E, Molgó J. Butyryl-cholinesterase and the control of synaptic responses in acetylcholinesteraseknockout mice. Life Sci 2007;80:2380–5.
[70] Gotti C, Fornasari D, Clementi F. Human neuronal nicotinic receptors. ProgNeurobiol 1997;53:199–237.
[71] Gotti C, Moretti M, Gaimarri A, Zanardi A, Clementi F, Zoli M. Heterogene-ity and complexity of native brain nicotinic receptors. Biochem Pharmacol2007;74:1102–11.
[72] Gould E, Woolf NJ, Butcher LL. Postnatal development of cholinergic neuronsin the rat: I. Forebrain. Brain Res Bull 1991;27:767–89.
[73] Greene LA, Aletta JM, Rukenstein A, Green SH. PC12 pheochromocytoma cells:culture, nerve growth factor treatment, and experimental exploitation. Meth-ods Enzymol 1987;147:207–16.
[74] Greenwood JM, Dragunow M. Muscarinic receptor-mediated phosphoryla-tion of cyclic AMP response element binding protein in human neuroblastomacells. J Neurochem 2002;82:389–97.
[75] Gremo F, Palomba M, Marchisio AM, Marcello C, Mulas ML. Heterogeneityof muscarinic cholinergic receptors in the developing human fetal brain:regional distribution and characterization. Early Hum Dev 1987;15:165–77.
[76] Guizzetti M, Costa P, Peters J, Costa LG. Acetylcholine as a mitogen: muscarinicreceptor-mediated proliferation of rat astrocytes and human astrocytomacells. Eur J Pharmacol 1996;297:265–73.
[77] Guizzetti M, Moore NH, Giordano G, Costa LG. Modulation of neuritogenesisby astrocyte muscarinic receptors. J Biol Chem 2008;283:31884–97.
[78] Guyenet P, Lefresne P, Rossier J, Beaujouan JC, Glowinski J. Inhibition byhemicholinium-3 of [14C]-acetylcholine synthesis and [3H]choline high-affinity uptake in rat striatal synaptosomes. Mol Pharmacol 1973;9:630–9.
[79] Haga T, Noda H. Choline uptake systems of rat brain synaptosomes. BiochimBiophys Acta 1973;291:564–75.
[80] Hamilton SE, Nathanson NM. The M1 receptor is required for muscarinic acti-vation of mitogen-activated protein (MAP) kinase in murine cerebral corticalneurons. J Biol Chem 2001;276:15850–3.
[81] Happe HK, Murrin LC. Development of high-affinity choline transport sites inrat forebrain: a quantitative autoradiography study with [3H]hemicholinium-3. J Comp Neurol 1992;321(4):591–611.
[82] Hartmann J, Kiewert C, Duysen EG, Lockridge O, Greig NH, Klein J. Excessivehippocampal acetylcholine levels in acetylcholinesterase-deficient mice aremoderated by butyrylcholinesterase activity. J Neurochem 2007;100:1421–9.
[83] Hellstrom-Lindahl E, Gorbounova O, Seiger A, Mousavi M, Nordberg A.Regional distribution of nicotinic receptors during prenatal development ofhuman brain and spinal cord. Dev Brain Res 1998;108:147–60.
[84] Holler T, Berse B, Cermak JM, Diebler MF, Blusztajn JK. Differences in the devel-opmental expression of the vesicular acetylcholine transporter and cholineacetyltransferase in the rat brain. Neurosci Lett 1996;212:107–10.
[85] Houser CR, Crawford GD, Barber RP, Salvaterra PM, Vaughn JE. Organizationand morphological characteristics of cholinergic neurons: an immunocyto-chemical study with a monoclonal antibody to choline acetyltransferase.Brain Res 1983;266:97–119.
[86] Houser CR. Cholinergic synapses in the central nervous system: studies ofthe immunocytochemical localization of choline acetyltransferase. J ElectronMicrosc Tech 1990;15:2–19.
[87] Hrabovska A, Farar V, Bernard V, Duysen EG, Brabec J, Lockridge O, et al. Drasticdecrease in dopamine receptor levels in the striatum of acetylcholinesteraseknock-out mouse. Chem Biol Interact 2010;183:194–201.
[88] Hu M, Liu QS, Chang KT, Berg DK. Nicotinic regulation of CREB activation inhippocampal neurons by glutamatergic and nonglutamatergic pathways. MolCell Neurosci 2002;21:616–25.
[89] Huang A, Noga BR, Carr PA, Fedirchuk B, Jordan LM. Spinal cholinergic neuronsactivated during locomotion: localization and electrophysiological character-ization. J Neurophysiol 2000;83:3537–47.
[90] Huang EJ, Reichardt LF. Neurotrophins: roles in neuronal development andfunction. Annu Rev Neurosci 2001;24:677–736.
[91] Hume RI, Role LW, Fischbach GD. Acetylcholine release from growth conesdetected with patches of acetylcholine receptor-rich membranes. Nature1983;305:632–4.
[92] Inestrosa NC, Moreno RD, Fuentes ME. Monomeric amphiphilic forms ofacetylcholinesterase appear early during brain development and may cor-respond to biosynthetic precursors of the amphiphilic G4 forms. NeurosciLett 1994;173:155–8.
[93] Inkson CA, Brabbs AC, Grewal TS, Skerry TM, Genever PG. Characterization ofacetylcholinesterase expression and secretion during osteoblast differentia-tion. Bone 2004;35:819–27.
[94] Isaacson LG, Tanaka Jr D. Cholinergic and non-cholinergic projections from thecanine pontomesencephalic tegmentum (Ch5 area) to the caudal intralaminarthalamic nuclei. Exp Brain Res 1986;62:179–88.
[95] Jang MH, Shin MC, Jung SB, Lee TH, Bahn GH, Kwon YK, et al. Alcohol andnicotine reduce cell proliferation and enhance apoptosis in dentate gyrus.Neuroreport 2002;13:1509–13.
[96] Johnson G, Moore SW. Cholinesterases modulate cell adhesion in human neu-roblastoma cells in vitro. Int J Dev Neurosci 2000;18:781–90.
[97] Johnson G, Moore SW. Human acetylcholinesterase binds to mouse laminin-1 and human collagen IV by an electrostatic mechanism at the peripheralanionic site. Neurosci Lett 2003;337:37–40.
[98] Jones SF, Kwanbunbumpen S. Some effects of nerve stimulation and hemi-cholinium on quantal transmitter release at the mammalian neuromuscularjunction. J Physiol 1970;207:51–61.
[99] Kasashima S, Muroishi Y, Futakuchi H, Nakanishi I, Oda Y. In situ mRNAhybridization study of the distribution of choline acetyltransferase in thehuman brain. Brain Res 1998;806:8–15.
[100] Kobayashi Y, Okuda T, Fujioka Y, Matsumura G, Nishimura Y, Haga T. Distri-bution of the high-affinity choline transporter in the human and macaquemonkey spinal cord. Neurosci Lett 2002;317:25–8.
[101] Koenigsberger C, Chiappa S, Brimijoin S. Neurite differentiation is modulatedin neuroblastoma cells engineered for altered acetylcholinesterase expres-sion. J Neurochem 1997;69:1389–97.
[102] Kotani S, Yamauchi T, Teramoto T, Ogura H. Donepezil, an acetyl-cholinesterase inhibitor, enhances adult hippocampal neurogenesis. ChemBiol Interact 2008;175:227–30.
[103] Kubo T, Fukuda K, Mikami A, Maeda A, Takahashi H, Mishina M, et al. Cloning,sequencing and expression of complementary DNA encoding the muscarinicacetylcholine receptor. Nature 1986;323:411–6.
[104] Kus L, Borys E, Ping Chu Y, Ferguson SM, Blakely RD, Emborg ME, et al. Distri-bution of high affinity choline transporter immunoreactivity in the primatecentral nervous system. J Comp Neurol 2003;463(3):341–57.
[105] Labarca C, Schwarz J, Deshpande P, Schwarz S, Nowak MW, Fonck C, et al. Pointmutant mice with hypersensitive alpha 4 nicotinic receptors show dopamin-ergic deficits and increased anxiety. Proc Natl Acad Sci USA 2001;98:2786–91.
[106] Lafrenie RM, Yamada KM. Integrin-dependent signal transduction. J CellBiochem 1996;61:543–53.
[107] Large TH, Bodary SC, Clegg DO, Weskamp G, Otten U, Reichardt LF.Nerve growth factor gene expression in the developing rat brain. Science1986;234:352–5.
[108] Laudenbach V, Medja F, Zoli M, Rossi FM, Evrard P, Changeux JP, et al.Selective activation of central subtypes of the nicotinic acetylcholine recep-tor has opposite effects on neonatal excitotoxic brain injuries. FASEB J2002;16:423–5.
[109] Lauder JM, Schambra UB. Morphogenetic roles of acetylcholine. EnvironHealth Perspect 1999;107(Suppl. 1):65–9.
[110] Layer PG, Alber R, Rathjen FG. Sequential activation of butyrylcholinesterasein rostral half somites and acetylcholinesterase in motoneuronesand myotomes preceding growth of motor axons. Development1988;102:387–96.
[111] Layer PG, Sporns O. Spatiotemporal relationship of embryonic cholinesteraseswith cell proliferation in chick retina and eye. Proc Natl Acad Sci USA1987;84:284–8.
[112] Layer PG, Weikert T, Alber R. Cholinesterases regulate neurite growth of chicknerve cells in vitro by means of a non-enzymatic mechanism. Cell Tissue Res1993;273:219–26.
[113] Layer PG, Willbold E. Novel functions of cholinesterases in development,physiology and disease. Prog Histochem Cytochem 1995;29:1–94.
[114] Le Magueresse C, Cherubini E. Presynaptic calcium stores contributeto nicotine-elicited potentiation of evoked synaptic transmission atCA3-CA1 connections in the neonatal rat hippocampus. Hippocampus2007;17:316–25.
[115] Levey AI, Kitt CA, Simonds WF, Price DL, Brann MR. Identification andlocalization of muscarinic acetylcholine receptor proteins in brain withsubtype-specific antibodies. J Neurosci 1991;11:3218–26.
[116] Levi-Montalcini R. The nerve growth factor: thirty-five years later. EMBO J1987;6:1145–54.
[117] Li B, Stribley JA, Ticu A, Xie W, Schopfer LM, Hammond P, et al. Abundant tissuebutyrylcholinesterase and its possible function in the acetylcholinesteraseknockout mouse. J Neurochem 2000;75:1320–31.
[118] Li BS, Ma W, Zhang L, Barker JL, Stenger DA, Pant HC. Activation ofphosphatidylinositol-3 kinase (PI-3K) and extracellular regulated kinases(Erk1/2) is involved in muscarinic receptor-mediated DNA synthesis in neuralprogenitor cells. J Neurosci 2001;21:1569–79.
[119] Li S, Cullen WK, Anwyl R, Rowan MJ. Muscarinic acetylcholine receptor-dependent induction of persistent synaptic enhancement in rat hippocampusin vivo. Neuroscience 2007;144(2):754–61.
[120] Li Y, Holtzman DM, Kromer LF, Kaplan DR, Chua-Couzens J, Clary DO, et al.Regulation of TrkA and ChAT expression in developing rat basal forebrain:evidence that both exogenous and endogenous NGF regulate differentiationof cholinergic neurons. J Neurosci 1995;15:2888–905.
[121] Liesi P. Neuronal migration on laminin involves neuronal contact forma-tion followed by nuclear movement inside a preformed process. Exp Neurol1992;117:103–13.
[122] Lips KS, Pfeil U, Haberberger RV, Kummer W. Localisation of the high-affinity choline transporter-1 in the rat skeletal motor unit. Cell Tissue Res2002;307:275–80.
[123] Lopez-Coviella I, Blusztajn JK. Bone morphogenetic proteins and cholinergicneurons in the central nervous system. Rev Neurol 2001;33:1054–60.
[124] Luo L, Chen WH, Wang M, Zhu DM, She JQ, Ruan DY. Modulation of long-termpotentiation by individual subtypes of muscarinic acetylcholine receptor inthe rat dentate gyrus. Hippocampus 2008;18(10):989–95.
[125] Ma W, Li BS, Zhang L, Pant HC. Signaling cascades implicated in muscarinicregulation of proliferation of neural stem and progenitor cells. Drug NewsPerspect 2004;17:258–66.
[126] Ma W, Maric D, Li BS, Hu Q, Andreadis JD, Grant GM, et al. Acetylcholinestimulates cortical precursor cell proliferation in vitro via muscarinic receptoractivation and MAP kinase phosphorylation. Eur J Neurosci 2000;12:1227–40.
[127] Madziar B, Shah S, Brock M, Burke R, Lopez-Coviella I, Nickel AC, et al. Nervegrowth factor regulates the expression of the cholinergic locus and the high-

Author's personal copy
Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378 377
affinity choline transporter via the Akt/PKB signaling pathway. J Neurochem2008;107:1284–93.
[128] Maggi L, Le Magueresse C, Changeux JP, Cherubini E. Nicotine acti vates imma-ture “silent” connections in the developing hippocampus. Proc Natl Acad SciUSA 2003;100:2059–64.
[129] Manabe T, Tatsumi K, Inoue M, Makinodan M, Yamauchi T, Makinodan E, et al.L3/Lhx8 is a pivotal factor for cholinergic differentiation of murine embryonicstem cells. Cell Death Differ 2007;14:1080–5.
[130] Manabe T, Tatsumi K, Inoue M, Matsuyoshi H, Makinodan M, Yokoyama S,et al. L3/Lhx8 is involved in the determination of cholinergic or GABAergiccell fate. J Neurochem 2005;94:723–30.
[131] Mansvelder HD, Mertz M, Role LW. Nicotinic modulation of synaptictransmission and plasticity in cortico-limbic circuits. Semin Cell Dev Biol2009;20:432–40.
[132] Matthews DA, Nadler JV, Lynch GS, Cotman CW. Development of cholin-ergic innervation in the hippocampal formation of the rat. Dev Biol1974;36:130–41.
[133] Mellott TJ, Kowall NW, Lopez-Coviella I, Blusztajn JK. Prenatal cholinedeficiency increases choline transporter expression in the septum and hip-pocampus during postnatal development and in adulthood in rats. Brain Res2007;1151:1–11.
[134] Meshorer E, Erb C, Gazit R, Pavlovsky L, Kaufer D, Friedman A, et al. Alter-native splicing and neuritic mRNA translocation under long-term neuronalhypersensitivity. Science 2002;295:508–12.
[135] Mesulam MM, Geula C. Acetylcholinesterase-rich neurons of the humancerebral cortex: cytoarchitectonic and ontogenetic patterns of distribution.J Comp Neurol 1991;306:193–220.
[136] Mesulam MM, Guillozet A, Shaw P, Levey A, Duysen EG, Lockridge O.Acetylcholinesterase knockouts establish central cholinergic pathways andcan use butyrylcholinesterase to hydrolyze acetylcholine. Neuroscience2002;110:627–39.
[137] Mesulam MM, Mufson EJ, Levey AI, Wainer BH. Cholinergic innervation ofcortex by the basal forebrain: cytochemistry and cortical connections of theseptal area, diagonal band nuclei, nucleus basalis (substantia innominata),and hypothalamus in the rhesus monkey. J Comp Neurol 1983;214:170–97.
[138] Mesulam MM, Mufson EJ, Wainer BH, Levey AI. Central cholinergic pathwaysin the rat: an overview based on an alternative nomenclature (Ch1-Ch6).Neuroscience 1983;10:1185–201.
[139] Mesulam MM, Mufson EJ. Neural inputs into the nucleus basalis of the sub-stantia innominata (Ch4) in the rhesus monkey. Brain 1984;107(Pt 1):253–74.
[140] Misawa H, Nakata K, Matsuura J, Nagao M, Okuda T, Haga T. Distribution ofthe high-affinity choline transporter in the central nervous system of the rat.Neuroscience 2001;105:87–98.
[141] Mobley WC, Woo JE, Edwards RH, Riopelle RJ, Longo FM, Weskamp G, et al.Developmental regulation of nerve growth factor and its receptor in the ratcaudate-putamen. Neuron 1989;3:655–64.
[142] Mohapel P, Leanza G, Kokaia M, Lindvall O. Forebrain acetylcholine reg-ulates adult hippocampal neurogenesis and learning. Neurobiol Aging2005;26:939–46.
[143] Moreno RD, Campos FO, Dajas F, Inestrosa NC. Developmental regula-tion of mouse brain monomeric acetylcholinesterase. Int J Dev Neurosci1998;16:123–34.
[144] Mufson EJ, Martin TL, Mash DC, Wainer BH, Mesulam MM. Cholinergic projec-tions from the parabigeminal nucleus (Ch8) to the superior colliculus in themouse: a combined analysis of horseradish peroxidase transport and cholineacetyltransferase immunohistochemistry. Brain Res 1986;370:144–8.
[145] Mulder AH, Yamamura HI, Kuhar MJ, Snyder SH. Release of acetylcholine fromhippocampal slices by potassium depolarization: dependence on high affinitycholine uptake. Brain Res 1974;70:372–6.
[146] Nachmansohn D, Machado AL. The formation of acetylcholine. A new enzyme“choline acetylase”. J Neurophysiol 1943;6:397–404.
[147] Nadler JV, Matthews DA, Cotman CW, Lynch GS. Development of cholin-ergic innervation in the hippocampal formation of the rat. II. Quantitativechanges in choline acetyltransferase and acetylcholinesterase activities. DevBiol 1974;36:142–54.
[148] Nastuk MA, Graybiel AM. Ontogeny of M1 and M2 muscarinic binding sitesin the striatum of the cat: relationships to one another and to striatal com-partmentalization. Neuroscience 1989;33:125–47.
[149] Nastuk MA, Graybiel AM. Pharmacologically defined M1 and M2 muscariniccholinergic binding sites in the cat’s substantia nigra: development and matu-rity. Dev Brain Res 1991;61:1–10.
[150] Ninomiya Y, Kayama Y, Koyama Y. Postnatal development of cholinergic neu-rons in the mesopontine tegmentum revealed by histochemistry. Int J DevNeurosci 2005;23:711–21.
[151] Ninomiya Y, Koyama Y, Kayama Y. Postnatal development of choline acetyl-transferase activity in the rat laterodorsal tegmental nucleus. Neurosci Lett2001;308:138–40.
[152] Nonomura T, Nishio C, Lindsay RM, Hatanaka H. Cultured basal fore-brain cholinergic neurons from postnatal rats show both overlapping andnon-overlapping responses to the neurotrophins. Brain Res 1995;683:129–39.
[153] Oda Y. Choline acetyltransferase: the structure, distribution and pathologicchanges in the central nervous system. Pathol Int 1999;49:921–37.
[154] Ohno K, Tsujino A, Brengman JM, Harper CM, Bajzer Z, Udd B, et al. Cholineacetyltransferase mutations cause myasthenic syndrome associated withepisodic apnea in humans. Proc Natl Acad Sci USA 2001;98:2017–22.
[155] Oliveira-da-Silva A, Vieira FB, Cristina-Rodrigues F, Filgueiras CC, Manhães AC,Abreu-Villaca Y. Increased apoptosis and reduced neuronal and glial densitiesin the hippocampus due to nicotine and ethanol exposure in adolescent mice.Int J Dev Neurosci 2009;27:539–48.
[156] Origlia N, Kuczewski N, Aztiria E, Gautam D, Wess J, Domenici L. Mus-carinic acetylcholine receptor knockout mice show distinct synaptic plasticityimpairments in the visual cortex. J Physiol 2006;577:829–40.
[157] Orr-Urtreger A, Broide RS, Kasten MR, Dang H, Dani JA, Beaudet AL, et al. Micehomozygous for the L250T mutation in the alpha7 nicotinic acetylcholinereceptor show increased neuronal apoptosis and die within 1 day of birth. JNeurochem 2000;74:2154–66.
[158] Owen A, Bird M. Acetylcholine as a regulator of neurite outgrowth and motil-ity in cultured embryonic mouse spinal cord. Neuroreport 1995;6:2269–72.
[159] Paraoanu LE, Layer PG. Mouse acetylcholinesterase interacts in yeast with theextracellular matrix component laminin-1beta. FEBS Lett 2004;576:161–4.
[160] Pauly JR, Slotkin TA. Maternal tobacco smoking, nicotine replacement andneurobehavioural development. Acta Paediatr 2008;97:1331–7.
[161] Pongrac JL, Rylett RJ. NGF-induction of the expression of ChAT mRNA in PC12cells and primary cultures of embryonic rat basal forebrain. Brain Res MolBrain Res 1998;62:25–34.
[162] Potter LT, Flynn DD, Liang JS, McCollum MH. Studies of muscarinic neuro-transmission with antimuscarinic toxins. Prog Brain Res 2004;145:121–8.
[163] Prado VF, Martins-Silva C, de Castro BM, Lima RF, Barros DM, Amaral E, et al.Mice deficient for the vesicular acetylcholine transporter are myasthenic andhave deficits in object and social recognition. Neuron 2006;51:601–12.
[164] Pugh CP, Berg DK. Neuronal acetylcholine receptors that bind �-bungarotoxinmediate neurite retraction in a calcium-dependent manner. J Neurosci1994;14:889–96.
[165] Rachinsky TL, Camp S, Li Y, Ekström TJ, Newton M, Taylor P. Molecular cloningof mouse acetylcholinesterase: tissue distribution of alternatively splicedmRNA species. Neuron 1990;5:317–27.
[166] Rahman S, López-Hernández GY, Corrigall WA, Papke RL. Neuronal nicotinicreceptors as brain targets for pharmacotherapy of drug addiction. CNS NeurolDisord Drug Targets 2008;7:422–41.
[167] Resende RR, Alves AS, Britto LR, Ulrich H. Role of acetylcholine receptors inproliferation and differentiation of P19 embryonal carcinoma cells. Exp CellRes 2008;314:1429–43.
[168] Resende RR, Gomes KN, Adhikari A, Britto LR, Ulrich H. Mechanism ofacetylcholine-induced calcium signaling during neuronal differentiation ofP19 embryonal carcinoma cells in vitro. Cell Calcium 2008;43:107–21.
[169] Ribeiro-Carvalho A, Lima CS, Medeiros AH, Siqueira NR, Filgueiras CC, Man-hães AC, et al. Combined exposure to nicotine and ethanol in adolescentmice: effects on the central cholinergic systems during short and long termwithdrawal. Neuroscience 2009;162:1174–86.
[170] Rivas RJ, Burmeister DW, Goldberg DJ. Rapid effects of laminin on the growthcone. Neuron 1992;8:107–15.
[171] Robinson TE, Kolb B. Structural plasticity associated with exposure to drugsof abuse. Neuropharmacology 2004;47:33–46.
[172] Rogers SL, Letourneau PC, Palm SL, McCarthy J, Furcht LT. Neurite extensionby peripheral and central nervous system neurons in response to substratum-bound fibronectin and laminin. Dev Biol 1983;98:212–20.
[173] Rosenblum K, Futter M, Jones M, Hulme EC, Bliss TV. ERKI/II regulation by themuscarinic acetylcholine receptors in neurons. J Neurosci 2000;20:977–85.
[174] Rossner S, Kues W, Witzemann V, Schliebs R. Laminar expression of m1-, m3-and m4-muscarinic cholinergic receptor genes in the developing rat visualcortex using in situ hybridization histochemistry. Effect of monocular visualdeprivation. Int J Dev Neurosci 1993;11:369–78.
[175] Rossner S, Kumar A, Witzemann V, Schliebs R. Development of laminarexpression of the m2 muscarinic cholinergic receptor gene in rat visual cortexand the effect of monocular visual deprivation. Dev Brain Res 1994;77:55–61.
[176] Rotundo RL. Expression and localization of acetylcholinesterase at the neu-romuscular junction. J Neurocytol 2003;32:743–66.
[177] Rüdiger T, Bolz J. Acetylcholine influences growth cone motility and morphol-ogy of developing thalamic axons. Cell Adhes Migr 2008;2:30–7.
[178] Santamaria J, Khalfallah O, Sauty C, Brunet I, Sibieude M, Mallet J, et al.Silencing of choline acetyltransferase expression by lentivirus-mediated RNAinterference in cultured cells and in the adult rodent brain. J Neurosci Res2009;87:532–44.
[179] Sarter M, Parikh V. Choline transporters, cholinergic transmission and cogni-tion. Nat Rev Neurosci 2005;6(1):48–56.
[180] Sawyer TW, Weiss MT. Parallel development of acetylcholinesterasein vivo and in primary neuron surface culture. Brain Res Dev Brain Res1993;71:147–9.
[181] Schambra UB, Sulik KK, Petrusz P, Lauder JM. Ontogeny of cholinergic neuronsin the mouse forebrain. J Comp Neurol 1989;288:101–22.
[182] Schlumpf M, Palacios JM, Cortes R, Lichtensteiger W. Regional developmentof muscarinic cholinergic binding sites in the prenatal rat brain. Neuroscience1991;45:347–57.
[183] Semba K, Fibiger HC. Time of origin of cholinergic neurons in the rat basalforebrain. J Comp Neurol 1988;269:87–95.
[184] Shacka JJ, Robinson SE. Postnatal developmental regulation of neuronal nico-tinic receptor subunit alpha 7 and multiple alpha 4 and beta 2 mRNA speciesin the rat. Dev Brain Res 1998;109:67–75.
[185] Shapiro MS, Gomeza J, Hamilton SE, Hille B, Loose MD, Nathanson NM, et al.Identification of subtypes of muscarinic receptors that regulate Ca2+ and K+channel activity in sympathetic neurons. Life Sci 2001;68:2481–7.

Author's personal copy
378 Y. Abreu-Villaca et al. / Behavioural Brain Research 221 (2011) 367–378
[186] Sharma KV, Bigbee JW. Acetylcholinesterase antibody treatment results inneurite detachment and reduced outgrowth from cultured neurons: furtherevidence for a cell adhesive role for neuronal acetylcholinesterase. J NeurosciRes 1998;53:454–64.
[187] Sharma KV, Koenigsberger C, Brimijoin S, Bigbee JW. Direct evidence for anadhesive function in the noncholinergic role of acetylcholinesterase in neuriteoutgrowth. J Neurosci Res 2001;63:165–75.
[188] Silman I, Sussman JL. Acetylcholinesterase: ‘classical’ and ‘non-classical’ func-tions and pharmacology. Curr Opin Pharmacol 2005;5:293–302.
[189] Silva AJ, Kogan JH, Frankland PW, Kida S. CREB and memory. Annu Rev Neu-rosci 1998;21:127–48.
[190] Slotkin TA, Ryde IT, Wrench N, Card JA, Seidler FJ. Nonenzymatic role of acetyl-cholinesterase in neuritic sprouting: regional changes in acetylcholinesteraseand choline acetyltransferase after neonatal 6-hydroxydopamine lesions.Neurotoxicol Teratol 2009;31:183–6.
[191] Slotkin TA. If nicotine is a developmental neurotoxicant in animal studies,dare we recommend nicotine replacement therapy in pregnant women andadolescents? Neurotoxicol Teratol 2008;30:1–19.
[192] Son JH, Winzer-Serhan UH. Chronic neonatal nicotine exposure increasesmRNA expression of neurotrophic factors in the postnatal rat hippocampus.Brain Res 2009;1278:1–14.
[193] Soreq H, Seidman S. Acetylcholinesterase—new roles for an old actor. Nat RevNeurosci 2001;2:294–302.
[194] Suszkiw JB, Pilar G. Selective localization of a high-affinity choline uptake sys-tem and its role in ACh formation in cholinergic nerve terminals. J Neurochem1976;26:1133–8.
[195] Tata AM, Cursi S, Biagioni S, Augusti-Tocco G. Cholinergic modulation of neu-rofilament expression and neurite outgrowth in chick sensory neurons. JNeurosci Res 2003;73:227–34.
[196] Tatsumi H, Tsuji S, Anglade P, Motelica-Heino I, Soeda H, Katayama Y. Syn-thesis, storage and release of acetylcholine at and from growth cones of ratcentral cholinergic neurons in culture. Neurosci Lett 1995;202:25–8.
[197] Taylor P, Radic Z. The cholinesterases: from genes to proteins. Annu RevPharmacol Toxicol 1994;34:281–320.
[198] Thal LJ, Gilbertson E, Armstrong DM, Gage FH. Development of the basal fore-brain cholinergic system: phenotype expression prior to target innervation.Neurobiol Aging 1992;13:67–72.
[199] Tian X, Sun X, Suszkiw JB. Developmental age-dependent upregulationof choline acetyltransferase and vesicular acetylcholine transporter mRNAexpression in neonatal rat septum by nerve growth factor. Neurosci Lett1996;209:134–6.
[200] Tribollet E, Bertrand D, Marguerat A, Raggenbass M. Comparative distributionof nicotinic receptor subtypes during development, adulthood and aging: anautoradiographic study in the rat brain. Neuroscience 2004;124:405–20.
[201] Turk E, Wright EM. Membrane topology motifs in the SGLT cotransporterfamily. J Membr Biol 1997;159:1–20.
[202] Vaglenova J, Birru S, Pandiella NM, Breese CR. An assessment of the long-termdevelopmental and behavioral teratogenicity of prenatal nicotine exposure.Behav Brain Res 2004;150:159–70.
[203] van Vulpen EH, Van Der KD. Differential maturation of cholinergic interneu-rons in the striatal patch versus matrix compartments. J Comp Neurol1996;365:683–91.
[204] VanDeMark KL, Guizzetti M, Giordano G, Costa LG. The activation of M1muscarinic receptor signaling induces neuronal differentiation in pyramidalhippocampal neurons. J Pharmacol Exp Ther 2009;329:532–42.
[205] Varga C, Hartig W, Grosche J, Keijser J, Luiten PG, Seeger J, et al. Rabbitforebrain cholinergic system: morphological characterization of nuclei anddistribution of cholinergic terminals in the cerebral cortex and hippocampus.J Comp Neurol 2003;460:597–611.
[206] Volpicelli LA, Levey AI. Muscarinic acetylcholine receptor subtypes in cerebralcortex and hippocampus. Prog Brain Res 2004;145:59–66.
[207] von Engelhardt J, Eliava M, Meyer AH, Rozov A, Monyer H. Functional char-acterization of intrinsic cholinergic interneurons in the cortex. J Neurosci2007;27:5633–42.
[208] Walters CL, Cleck JN, Kuo YC, Blendy JÁ. Mu-opioid receptor and CREB activa-tion are required for nicotine reward. Neuron 2005;46:933–43.
[209] Weihe E, Schafer MK, Schutz B, Anlauf M, Depboylu C, Brett C, et al.From the cholinergic gene locus to the cholinergic neuron. J Physiol Paris1998;92:385–8.
[210] Weikert T, Rathjen FG, Layer PG. Developmental maps of acetyl-cholinesterase and G4-antigen of the early chicken brain: long-distancetracts originate from AChE-producing cell bodies. J Neurobiol 1990;21:482–98.
[211] Wessler I, Kilbinger H, Bittinger F, Kirkpatrick CJ. The biological role of non-neuronal acetylcholine in plants and humans. Jpn J Pharmacol 2001;85:2–10.
[212] Wessler I, Kirkpatrick CJ, Racke K. The cholinergic ‘pitfall’: acetylcholine, auniversal cell molecule in biological systems, including humans. Clin ExpPharmacol Physiol 1999;26:198–205.
[213] Wessler I, Kirkpatrick CJ. Acetylcholine beyond neurons: the non-neuronalcholinergic system in humans. Br J Pharmacol 2008;154:1558–71.
[214] Widmer H, Ferrigan L, Davies CH, Cobb SR. Evoked slow muscarinicacetylcholinergic synaptic potentials in rat hippocampal interneurons. Hip-pocampus 2006;16:617–28.
[215] Wonnacott S. Presynaptic nicotinic ACh receptors. Trends Neurosci1997;20:92–8.
[216] Wonnacott S, Barik J, Dickinson J, Jones IW. Nicotinic receptors modulatetransmitter cross talk in the CNS: nicotinic modulation of transmitters. J MolNeurosci 2006;30:137–40.
[217] Yagle K, Lu H, Guizzetti M, Moller T, Costa LG. Activation of mitogen-activatedprotein kinase by muscarinic receptors in astroglial cells: role in DNA synthe-sis and effect of ethanol. Glia 2001;35:111–20.
[218] Yang D, Howard A, Bruun D, Ajua-Alemanj M, Pickart C, Lein PJ. Chlor-pyrifos and chlorpyrifos-oxon inhibit axonal growth by interfering withthe morphogenic activity of acetylcholinesterase. Toxicol Appl Pharmacol2008;228:32–41.
[219] Yao WD, Rusch J, Poo M, Wu CF. Spontaneous acetylcholine secretion fromdeveloping growth cones of Drosophila central neurons in culture: effects ofcAMP-pathway mutations. J Neurosci 2000;20:2626–37.
[220] Young SH, Poo MM. Spontaneous release of transmitter from growth conesof embryonic neurones. Nature 1983;305:634–7.
[221] Zahalka EA, Seidler FJ, Lappi SE, Yanai J, Slotkin TA. Differential developmentof cholinergic nerve terminal markers in rat brain regions: implications fornerve terminal density, impulse activity and specific gene expression. BrainRes 1993;601:221–9.
[222] Zakharenko S, Chang S, O’Donoghue M, Popov SV. Neurotransmitter secretionalong growing nerve processes: comparison with synaptic vesicle exocytosis.J Cell Biol 1999;144:507–18.
[223] Zhang XJ, Yang L, Zhao Q, Caen JP, He HY, Jin QH, et al. Induction of acetyl-cholinesterase expression during apoptosis in various cell types. Cell DeathDiffer 2002;9:790–800.
[224] Zhao WQ, Alkon DL, Ma W. c-Src protein tyrosine kinase activity is required formuscarinic receptor-mediated DNA synthesis and neurogenesis via ERK1/2and c-AMP-responsive element-binding protein signaling in neural precursorcells. J Neurosci Res 2003;72:334–42.
[225] Zhao Y, Marin O, Hermesz E, Powell A, Flames N, Palkovits M, et al. TheLIM-homeobox gene Lhx8 is required for the development of many cholin-ergic neurons in the mouse forebrain. Proc Natl Acad Sci USA 2003;100:9005–10.
[226] Zheng JQ, Felder M, Connor JA, Poo MM. Turning of nerve growth conesinduced by neurotransmitters. Nature 1994;368:140–4.
[227] Zhou C, Wen ZX, Shi DM, Xie ZP. Muscarinic acetylcholine receptors involvedin the regulation of neural stem cell proliferation and differentiation in vitro.Cell Biol Int 2004;28:63–7.
[228] Zhou FC, Azmitia EC. Laminin facilitates and guides fiber growth of trans-planted neurons in adult brain. J Chem Neuroanat 1988;1:133–46.
[229] Zoli M, Le Novere N, Hill Jr JA, Changeux JP. Developmental regulation of nico-tinic ACh receptor subunit mRNAs in the rat central and peripheral nervoussystems. J Neurosci 1995;15:1912–39.
[230] Zoli M, Léna C, Picciotto MR, Changeux JP. Identification of four classesof brain nicotinic receptors using beta2 mutant mice. J Neurosci 1998;18:4461–72.
[231] Zurn A. Fibroblast growth factor differentially modulates the neuro-transmitter phenotype of cultured sympathetic neurons. J Neurosci1992;12:4195–201.
[232] Zurn AD, Werren F. Development of CNS cholinergic neurons in vitro: selec-tive effects of CNTF and LIF on neurons from mesencephalic cranial motornuclei. Dev Biol 1994;163:309–15.




















