
TRPV4 channel is involved in the coupling of fluid viscosity changes to epithelial ciliary activity
Upload
southerndenmarkCategory
view
1download
0
Invertebrate Biology 123(2): 156-167. 0 2004 American Microscopical Society, Inc.
Ciliary feeding structures and particle capture mechanism in the freshwater bryozoan Plumatella repens (Phylactolaemata)
Hans Ulrik Riisgird, Kasper Kollerup Nielsen,* Judith Fuchs,2 Bo Fmss Rasniussen,2 Matthias Obst,2 and Peter Funch2
I Marine Biological Research Centre, University of Southern Denmark, Hindsholmvej 11, DK-5300 Kerteminde, Denmark
* Department of Zoology, Institute of Biological Sciences, University of Aarhus, DK-8000 Aarhus C, Denmark
Abstract. In contrast to marine bryozoans, the lophophore structure and the ciliary filter- feeding mechanism in freshwater bryozoans have so far been only poorly described. Specimens of the phylactolaemate bryozoan Plumatella repens were studied to clarify the tentacular ciliary structures and the particle capture mechanism. Scanning electron microscopy revealed that the tentacles of the lophophore have a frontal band of densely packed cilia, and on each side a zigzag row of laterofrontal cilia and a band of lateral cilia. Phalloidin-linked fluorescent dye showed no sign of muscular tissue within the tentacles. Video microscopy was used to describe basic characteristics of particle capture. Suspended particles in the incoming water flow, set up by the lateral ‘pump’ cilia on the tentacles, approach the tentacles with a velocity of 1-2 mm s--I. Near the tentacles, the particles are stopped by the stiff sensory laterofrontal cilia acting as a mechanical sieve, as previously seen in marine bryozoans. The particle capture mechanism suggested is based on the assumed ability of the sensory stiff laterofrontal cilia to be triggered by the deflection caused by the drag force of the through-flowing water on a captured food particle. Thus, when a particle is stopped by the laterofrontal cilia, the otherwise stiff cilia are presumably triggered to make an inward flick which brings the restrained particle back into the downward directed main current, possibly to be captured again further down in the lophophore before being carried to the mouth via the food groove. No tentacle flicks and no transport of captured particles on the frontal side of the tentacles were observed. The velocity of the me- tachronal wave of the water-pumping lateral cilia was measured to be -0.2 mm s-’, the wave- length was -7 pm, and hence the ciliary beat frequency estimated to be -30 Hz (-20 “C). The filter feeding process in P. repens reported here resembles the ciliary sieving process described for marine bryozoans in recent years, although no tentacle flicks were observed in P. repens. The phylogenetic position of the phylactolaemates is discussed in the light of these findings.
Additional key words: ciliary filter feeding, Bryozoa, Phoronida
All bryozoans, members of the phylum Bryozoa (= Ectoprocta; = Polyzoa), are colonial ciliary filter feed- ers. When feeding, the zooid extends its ciliated ten- tacle crown (lophophore) into the ambient water (Ry- land 1976, Winston 1978, Okamua & Doolan 1993, Shunatova & Ostrovsky 2001).
In the Bryozoa, three classes are commonly recog- nized, and one of these, the Phylactolaemata, lives ex- clusively in freshwater (Mukai et al. 1997, Nielsen 2001). The phylactolaemate lophophore (Figs. 1-8) is characteristically horseshoe-shaped, with two lopho-
kt Author for correspondence. E-mail: [email protected]
phore arms projecting on the dorsal side (Fig. 9). In marine bryozoans, in contrast, the lophophore is al- ways circular. The phylactolaemate horseshoe-shaped lophophore has often been regarded as a primitive character correlating with large zooid size. Thus, Mun- dy et al. (1981) suggested that phylactolaemates may be more closely related to phoronids than to marine bryozoans, and Bullivant (1968) found that the food collecting device in phylactolaemates “is an interme- diate stage” between those of phoronids and gymno- laemate bryozoans.
In recent years, the particle capture mechanism has been thoroughly described in adult marine bryozoans (Riisgird & Manriquez 1997, Nielsen & Riisgird

Ciliary feeding in Plumatella repens 157
1998, Larsen & Riisgird 2002) as well as in the adult marine phoronid Phoronis muelleri SELYS-LONCHAMPS 1903 (Riisgird 2002). The method of ciliary sieving described for P. muelleri and a number of marine bryozoans seem to be rather similar. In marine bryo- zoans, the feeding apparatus consists of a circular fun- nel-shaped lophophore, with the mouth at the centre of its base. Bands of lateral cilia on the tentacles pro- duce the feeding currents. In both the marine bryo- zoans and the phoronids, a row of stiff laterofrontal cilia is placed upstream of the water-pumping lateral cilia band, and each laterofrontal cilium is situated in a depression surrounded by a ring of microvilli, indi- cating a sensory function. When the through currents pass between the tentacles, they form a relatively strong core current directed straight down into the lophophore. Particles in the currents are retained by the stiff laterofrontal ciliary filter and either transport- ed downwards on the tentacles, by means of frontal cilia on the frontal side of the tentacles, or the particles are transferred to the core current, by means of inward tentacle flicks triggered by the arrested particles (Riis- gird & Manriquez 1997, Nielsen & Riisgbrd 1998, Larsen & Riisgird 2002). However, some differences between P. muelleri and bryozoans are also present. A strong central lophophore current is crucial for ma- rine bryozoans where tentacle flicks serve to transfer arrested particles back into the downward-directed core current. The tentacle flicks made by P. muelleri are more sinuous than seen in marine bryozoans, and this may enable the tentacles to keep an arrested par- ticle close to the frontal side of the tentacle during the whole flick cycle, which seems to indicate that pho- ronids may not be dependent on a central lophophore current.
To clarify if freshwater bryozoans deviate from ma- rine bryozoans, by possessing characters more in com- mon with phoronids, it is of interest to make a com- parative study of the particle capture mechanism. The present work deals with the particle capture mecha- nism in the freshwater bryozoan Plumatella repens us- ing scanning electron microscopy, muscle staining with phalloidin-linked dye visualized by epifluoresc- ence microscopy, and the same video-microscope method as applied in previous studies on marine bryo- zoans and the marine phoronid Phoronis muelleri.
Methods
selisborg, Arhus (Denmark) in early October and late November 2002. Specimens were brought to the De- partment of Zoology, Arhus, and the Marine Biologi- cal Research Centre, Kerteminde, and were kept in aerated freshwater from the collecting site. Specimens were narcotized with successive addition of droplets of dissolved MgC1, and fixed in picric acid formal- dehyde (PAF) with an osmolarity of 200-500 mOsm, according to Melone & Ricci (1995).
Zooids were dissected from fixed colonies and rinsed in droplets of 0.01 M PBS buffer on a micro- scopic slide. The buffer was replaced with 0.2% Triton X-100 in PBS for 1 h to make the specimens perme- able. Alexa-488-phalloidin was added for 40 min to visualize actin, and PBS buffer was used to rinse the specimens. The stained specimens were finally placed in Fluoromount-G and sealed with a coverslip. Epi- fluorescence images of stained muscular tissues were obtained using a Olympus BX60 light microscope with a U-MWIB2 filter cube and a ColorView I1 digital camera.
Fixed zooids, lophophores, and single tentacles of P. repens were isolated and placed in cages closed with a fine mesh (Amsellem & Clkment 1980). Some of the specimens were treated with ultrasound for 90 s, transferred to Smensen-phosphate-buffer and Vor- tex-stirred for 10 s to remove the cilia (see Rosenbaum & Carlson 1969). All cages were placed in 1% OsO, for 2 h of postfixation, transferred to distilled water, dehydrated in an ethanol series, critical point dried us- ing carbon dioxide, mounted on aluminium SEM stubs with sticky carbon pads, sputter coated with gold, and examined with a CamScan Maxim 2040 En Vac SEM.
The particle capture process in P. repens was re- corded using a video camera (Kappa CF 11/1) attached to an inverted microscope (Labovert FS), and a 50 “fields” or “half-frames’’ s- ’ video recorder (Pana- sonic NV-FS200 HQ). The animals in their natural tube were placed somewhat elevated (-2 mm) above the bottom in an observation chamber placed on the microscope. About 12-(~,mm-diameter algal cells (Tetra- selrnis sp.) were added to the water in the observation chamber in a low concentration so that the animals only encountered a particle every 1-3 s. Video frames could be copied by means of a video graphic printer (Sony UP-860 CE). Food particle movements were traced from their positions in successive frames by mounting a transparent plastic sheet onto the video screen so that the tentacle contours, as well as the po- sition of suspended particles, could be marked with a pen directly on the sheet, frame by frame.
Results The colonies of Plumatella repens comprise -750
zooids covering an area of 225 cm2 (- 1/3 of the Nym-
Colonies of the freshwater bryozoan Plumatella re- pens LINNAEUS 1758 were found growing on the un- derside of leaves and on stems of the waterlily Nym- phaea alba down to a depth of 1 m. The material was collected in a small pond in Dronningens Have, Mar-

158 Riisgiird, Nielsen, Fuchs, Rasmussen, Obst, & Funch
phea leaf). The length of an expanded zooid is -2.3 mm. The lophophore consists of 48-50 tentacles and measured - 1.2 mm in length. The bases the tentacles are connected by intei-tentacular membranes (“in”: Figs. 1, 2). The mean length of a tentacle is 780 pm, but the tentacles show different lengths depending on their position on the lophophore (Fig. 1). Short tenta- cles are situated furthest away from the mouth on the edge of the lophophore and have a length of 660 pm, whereas longer tentacles close to the mouth are -890 pm long (Fig. 9).
A tentacle of P. repens is triangular in cross-section. Five ciliary bands can be distinguished, extending from the tip to the base of each tentacle (Figs. 2-5). The bands are arranged in 3 functional groups, one unpaired frontal ciliary band, one pair of laterofrontal ciliary bands, and one pair of lateral ciliary bands (lc) (Fig. 4). The frontal band consists of densely packed cilia with a length of - 14 pm. The laterofrontal bands consist of two rows each, arranged in a zigzag for- mation (Fig. 5). The length of a laterofrontal cilium is -18 pm, but some are up to 25 pm long. The base of each laterofrontal cilium is surrounded by several short microvilli (Fig. 4). The distance between two cilia in one laterofrontal row is 4 pm (Fig. 4) and the distance between the two rows is 1.2 pm (Fig. 5). The lateral ciliary bands consist of 12-pm-long cilia.
Several bundles of -4 cilia are projecting from
Fig. 1. SEM. Lateral view of a zo- oid with lophophore expanded. in, intertentacular membrane; or, ori- fice; ro, rotifer; te, tentacle.
small pits on the abfrontal side of the tentacles (Figs. 2, 3). These bundles have a diameter of 1.5-2 pm, and are surrounded by a ring of microvilli. The cilia are - 18 pm long, while the microvilli are -0.5 pm long (Fig. 3). The structure is assumed to have sensory function. The distance between the bundles varies be- tween 35 pm at the base of a tentacle to 25 pm near the tip of a tentacle. The most distal bundle near the tentacle tip is larger, with a diameter -4 pm.
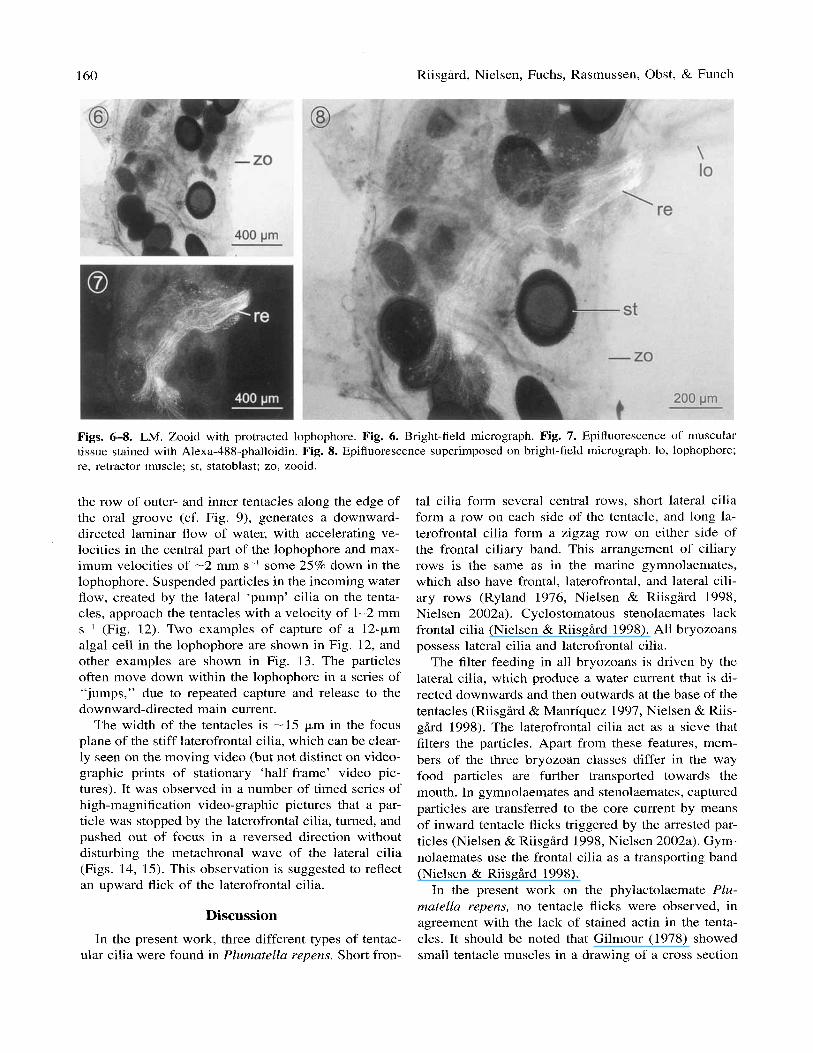
The Alexa-488-phalloidin staining shows significant binding to the actin of the lophophore retractor muscle (Figs. 6-8). Each myofibril is clearly visualized (Fig. 7). The retractor muscle consists of -50 myofibrills; it inserts in the basal part of the polypide and extends 1375 pm to the lophophore base. The staining shows no signs of actin within the lophophore. A weak fluo- rescence is observed around the polypid collar (Fig. 7).
The flow of water around the lophophore of P. re- yens is indicated by arrows in Fig. 9. Video-graphic pictures, using different foci of the microscope, give an impression of the three-dimensional shape of the lophophore. The series of video-graphic pictures in Fig. 10 illustrate a food particle approaching the loph- ophore of P. repens and being captured. It is notable that no tentacle flicks are observed. The flow pattern in the region of the lophophore is shown in Fig. 1 1 .
The geometry of the V-shaped funnel, made up of

Ciliary feeding in Plumatella repens 159
Figs. 2-5. SEM. Lophophore with close-up of the tentacles. Arrows indicate abfrontal, putative sensory cilia; single arrowheads indicate lateral ciliary bands; double arrowheads indicate laterofrontal ciliary bands, triple arrowheads indicate the frontal ciliary band. Fig. 2. Abfrontal view of tentacle. Fig. 3. Abfrontal sensory organ. Fig. 4. Lateral view of tentacle with all ciliated bands in view except one laterofrontral band. The base of each laterofrontal cilium is surrounded by short microvilli (insert). Fig. 5. Frontal view of tentacle. The laterofrontal cilia are positioned right next to the band of dense frontal cilia. in, intertentacular membrane.
160 Riisgird, Nielsen, Fuchs, Rasmussen, Obst, & Funch
Figs. 6-8. LM. Zooid with protracted lophophore. Fig. 6. Bright-field micrograph. Fig. 7. Epifluorescence of muscular tissue stained with Alexa-488-phalloidin. Fig. 8. Epifluorescence superimposed on bright-field micrograph. lo, lophophore; re, retractor muscle; st, statoblast; zo, zooid.
the row of outer- and inner tentacles along the edge of the oral groove (cf. Fig. 9), generates a downward- directed laminar flow of water, with accelerating ve- locities in the central part of the lophophore and max- imum velocities of -2 mm s-I some 25% down in the lophophore. Suspended particles in the incoming water flow, created by the lateral ‘pump’ cilia on the tenta- cles, approach the tentacles with a velocity of 1-2 mm s- ’ (Fig. 12). Two examples of capture of a 12-pm algal cell in the lophophore are shown in Fig. 12, and other examples are shown in Fig. 13. The particles often move down within the lophophore in a series of
jumps,” due to repeated capture and release to the downward-directed main current.
The width of the tentacles is -15 pm in the focus plane of the stiff laterofrontal cilia, which can be clear- ly seen on the moving video (but not distinct on video- graphic prints of stationary ‘half-frame’ video pic- tures). It was observed in a number of timed series of high-magnification video-graphic pictures that a par- ticle was stopped by the laterofrontal cilia, turned, and pushed out of focus in a reversed direction without disturbing the metachronal wave of the lateral cilia (Figs. 14, 15). This observation is suggested to reflect an upward flick of the laterofrontal cilia.
‘‘.
Discussion In the present work, three different types of tentac-
ular cilia were found in Plumatella repens. Short fron-
tal cilia form several central rows, short lateral cilia form a row on each side of the tentacle, and long la- terofrontal cilia form a zigzag row on either side of the frontal ciliary band. This arrangement of ciliary rows is the same as in the marine gymnolaemates, which also have frontal, laterofrontal, and lateral cili- ary rows (Ryland 1976, Nielsen & Riisgird 1998, Nielsen 2002a). Cyclostomatous stenolaemates lack frontal cilia (Nielsen & Riisgird 1998). All bryozoans possess lateral cilia and laterofrontal cilia.
The filter feeding in all bryozoans is driven by the lateral cilia, which produce a water current that is di- rected downwards and then outwards at the base of the tentacles (Riisgird & Manriquez 1997, Nielsen & Riis- g k d 1998). The laterofrontal cilia act as a sieve that filters the particles. Apart from these features, mem- bers of the three bryozoan classes differ in the way food particles are further transported towards the mouth. In gymnolaemates and stenolaemates, captured particles are transferred to the core current by means of inward tentacle flicks triggered by the arrested par- ticles (Nielsen & Riisgird 1998, Nielsen 2002a). Gym- nolaemates use the frontal cilia as a transporting band (Nielsen & RiisgArd 1998).
In the present work on the phylactolaemate Plu- matella repens, no tentacle flicks were observed, in agreement with the lack of stained actin in the tenta- cles. It should be noted that Gilmour (1978) showed small tentacle muscles in a drawing of a cross section

Ciliary feeding in Plumatella repens 161
n 0.2 rnm
Fig. 9. (Above) Horseshoe-shaped tentacle crown (lophophore) with water flow indicated by arrows (modified from Gilmour 1978). (Below) Video-graphic pictures of the tentacle crown through different focal planes.

I62 Riisgird, Nielsen, Fuchs, Rasmussen, Obst, & Funch
Fig. 10. Video sequence showing the lophophore capturing a I2-pm algal cell (indicated by arrows). Time intervals between frames a-d: 0.1s; e-i: 0.02s.
of a Plumatella sp. based on TEM observations. The tentacular muscles of P. repens seem to be weakly developed compared to other bryozoans such as Mem- hruniporu sp. (Gilmour 1978) and Electru pilosu (Mu- kai et al. 1997).
I - 0.2 mm
Fig. 11. Flow pattern in the region of the lophophore, ob- tained by means of video-recording of 12-pm algal cells. Flow lines based on a time interval of 0.02s between sub- sequent video frames. Only one particle shown is being cap- tured (two “jumps” seen), other particles are disrupted by contact to the tentacles but lost again, and finally some par- ticles follow the water without being disturbed. Only the latter particles completely reflect the flow pattern of the wa- ter.
Feeding in P. repens has previously been described by Bullivant (1 968), who observed that, between the two rows of tentacles, a central current apparently “impinges” on the oral groove and “cilia lining the trough convey particles to and into the mouth.” Par- ticles were not seen travelling down the frontal surface of the tentacles, and Bullivant suggested that the sharp deflection of the feeding current over the mouth causes particles to be thrown towards the mouth or into an eddy of comparatively still water over it. Presumably due to the high speed of the process, Bullivant was not able to observe how a particle, after the first con- tact with a tentacle, may be pushed back into the cen- tral current to be conveyed further down into the loph- ophore through a series of “jumps” (Fig. 12).
Nielsen & Riisgird (1998) found that the latero- frontal cilia of cyclostomes are mechanoreceptors with clear sensory abilities. Gilmour (1978) stated that the laterofrontal cilia in Plumatella sp. are surrounded by rings of microvilli that contain fibrils, which in turn extend into the laterofrontal cells; the cell bases make synapses with nerves running along the tentacles, in- dicating that the laterofrontal cilia have a sensory role. As presented in this work, individual of P. repens show similar morphological and distributional patterns of laterofi-ontal cilia. The laterofrontal cilia are ar- ranged in a zigzag formation with intervals of 4 pm, which has also been observed in two gymnolaemate species (Nielsen 2002a).

Ciliary feeding in Plumatella repens I63
j.0.9 (b) j o . 9
10.9
. 0.2 mm
Fig. 12. Section of the lophophore showing two examples of the capture of a 12-pm algal cell. Time interval between dots is 0.02s. Estimated velocity (mm s-’) of the particle is indicated.
The particle capture mechanism suggested in this work is based on the assumed ability of the sensory, stiff, laterofrontal cilia to be triggered by the deflection caused by the drag force of the through-flowing water on a captured food particle (cf. Larsen & Riisgbrd 2002: Fig. 7); the particle is then pushed into the cen- tral current by the flicking laterofrontal cilia. Gordon (1974) observed that the sensory (tactile) laterofrontal cilia in the marine bryozoan Cryptosula pallasiana “are not rigid, but are capable of occasional flicking movements,” using negative-field phase-contrast mi- croscopy on live specimens. This observation supports the present hypothesis that it is flicking of the later- ofrontal cilia that push a stopped particle back into the downward-directed main current of the lophophore (Figs. 14, 15). Further support comes from a study on Bowerbankia imbricata by Larsen & Riisgbrd (2002); here it was observed that MgC1,-narcotised animals no longer flicked with the tentacles but merely maintained the current driven by the lateral cilia. The laterofrontal cilia that were likewise affected by the MgCl, could no longer retain particles. This also indicates that the laterofrontal cilia interact with the particles more than simply as a mechanical barrier. Alternatively, the MgCI, could have affected the arrangement of the la- terofrontal cilia, making them unable to act as a me- chanical filter.
The method of ciliary sieving by means of stiff la- terofrontal cilia, described here for P. repens, has also been described for marine bryozoans (Riisgbrd & Manriquez 1997, Nielsen & Riisgbrd 1998, Larsen & Riisgdrd 2002) and for the adult Phoronis muelleri (Riisgbrd 2002). However, there are some differences between the freshwater bryozoan Plumatella repens, and marine bryozoans and phoronids. A strong central
_,_._._._._. -.-.-. .-. . . . . _._.-.re ....: *-.-. ... -.+./-
-\ ;
0.2 mm Fig. 13. Eight examples of particle interception by the ten- tacles in the lophophore (only the distal part of the tentacle is shown in each case). Dots represent time interval of 0.02s.
lophophore current is crucial for marine bryozoans, where tentacle flicks serve to transfer arrested particles back into the downward-directed core current. The ten- tacle flicks made by Phoronis muelleri are more sin- uous than seen in marine bryozoans. Apparently, this enables the tentacles to keep an arrested particle close to the frontal side of the tentacle during the whole flick cycle, which could indicate that phoronids may not depend on a central lophophore current to carry par- ticles toward the mouth (Riisgbrd 2002). Considering this, the lack of tentacle flicks in the particle capture process in Plumatella repens is striking. It seems pos- sible that the capture mechanism described here for P. repens may be identical to the mechanism for particle capture in the lower part of the lophophore of marine bryozoans. In the marine bryozoan Bowerbankia im- bricatu, Larsen & Riisgbrd (2002) found that when a particle is captured on the distal part of a tentacle, a tentacle flick is triggered that brings the particle into the central core and down towards the mouth. But par- ticles trapped deep down in the lophophore do not trig- ger a tentacle flick, but rather a quick displacement “possibly by cilia action,” that brings a trapped par- ticle back into the central current.
Although most movement of captured particles in marine bryozoans seems to depend on flicking-assisted transport, particles may also be conveyed downwards on the frontal surface of the tentacle (e.g., Larsen & Riisgzrd 2002: Fig. 3). No downward transport of par- ticles was seen on the frontal surface of the tentacles in Plumatella repens, but more studies on other fresh- water bryozoans are needed to evaluate if any gener-

164 RiisgArd, Nielsen, Fuchs, Rasmussen, Obst, & Funch
* 16 8
3 I
&
I I --..-- 2 *. 50 pm 1
Fig. 14. (Above) Example of particle interception by the tentacles in the lophophore (only the bases of three tentacles are seen in the series of video-graphic pictures). (Below) Drawing of the path of the particle in the above photos. #1 = incoming particle. The time interval between the following outlines of the particle is 0.02s, and the numbers correspond with those in the upper panel. The direction of the laeoplectic metachronal wave (indicated by arrow) shows that the lophophore is being observed from the outside. The particle approaches the tentacle (#1-5; out of focus indicated by dashed outline), comes into focus (#7, 9; indicated by unbroken outline) to be subsequently turned and pushed inwards into the downward- directed main current of the lophophore, which brings the particle further down towards the mouth (#11-16).
alization about similarities between freshwater and marine bryozoans can be made.
Strathmann (1973, 1982) observed that particle cap- ture on tentacles of two adult, marine bryozoans was correlated with a change in the metachronal pattern of the lateral ciliary bands. But Strathmann did not ob- serve the laterofrontal cilia, and concluded that the particles were captured by the lateral cilia due to local reversal of beat induced by individual particles. Based on this interpretation, Strathmann & McEdward (1986) concluded that ciliary sieving by means of stiff later- ofrontal cilia, observed in the cyphonautes larvae of a gymnolaemate bryozoan, “breaks a biological rule.” Based on a study of the conspicuous morphological similarities between adult and larvae bryozoan filter- ing structures, Nielsen (2002a) suggested that the basic particle retention mechanisms are similar, “both strain particles by a sieve formed by the lateral row of la- terofrontal cilia,” but the method of transferring cap-
tured particles further towards the mouth may differ. The suggested ability of the laterofrontal cilia to flick when stimulated by a captured particle may perhaps be the method of transferring particles to the frontal ciliary band on the immobile cyphonautes ciliary ridge. Such mechanism does not exclude the possibil- ity that local disruption of the metachronal wave of the lateral cilia, possibly triggered simultaneously with a tentacle- or laterofrontal ciliary flick, may play a crucial role in the transfer of the captured particle from the filter as discussed by RiisgArd (2002).
In conclusion, the observations presented here (in- cluding the absence of disruptions of the metachroni- cal waves of the lateral cilia) suggest that the particle capture mechanism in Plumatella repens is based on the ability of the sensory, stiff, laterofrontal cilia to flick when triggered by the drag force on a captured food particle, Thus, when a particle is stopped by the laterofrontal cilia, the otherwise stiff cilia are triggered

Ciliary feeding in Plumatella repens 165
I I
Fig. 15. Example of particle interception by the tentacles in the lower part of the lophophore. (Above) #1 = incoming particle; #3 = same particle seen 2 video frames later, etc. (i.e., time interval between successive particle outlines is 2/50=0.04s). The direction of the laeoplectic metachronal wave (indicated by arrow) shows that the lophophore is being observed from the outside. The particle approaches the tentacle (#l; out of focus indicated by dashed outline), comes into focus (#3) (indicated by unbroken outline) to be subsequently turned and pushed inwards (#5-9) into the downward-directed main current of the lophophore, which brings the particle further down towards the mouth (#lo-15). (Below) Series of video-graphic pictures where numbers correspond with those indicated in the upper panel. Scale, 50 km.
to make an inward flick, which brings the restrained particle back into the downward-directed main current, possibly to be captured again further down in the loph- ophore before being carried to the mouth via the food groove. However, video-microscope recordings with sufficient resolution are needed to conclusively reveal how an arrested particle that does not trigger a tentacle flick (as in the present work) may be transferred from the laterofrontal filter and back into the downward- directed currents in the lophophore, and if this process is coincident with a brief local disruption in the me- tachronal wave of the lateral cilia pumping water be- low the arrested particle. Further, the distances that particles are suggested to be “pushed” out into the downward-directed main current by laterofrontal-cili- ary flicks should be measured and compared to the length of the laterofrontal cilia in order to evaluate the laterofrontal-ciliary flick hypothesis.
Apart from the sensory laterofrontal cells, a second type of presumed sensory structure occurs on the abfrontal side of the tentacles of P. repens. We regard them as sensory due to the uniform pattern in which they occur and their morphology, both similar to the abfrontal row of sensory cells that has been docu- mented in several marine species (Shunatova & Niel- sen 2002). The distal-most structure in the abfrontal sensory cell row is enlarged, and might correspond to the terminal multiciliated sensory cells that were found in all studied gymnolaemates by Shunatova & Nielsen (2002). These structures may serve the purpose of de- tecting elements like food particles and potential pred- ators as Shunatova & Nielsen (2002) suggested for several gymnolaemate bryozoans.
The position of phylactolaemates within the bryo- zoans is unclear and has not yet been addressed in a thorough phylogenetic analysis (Nielsen 2001, Nielsen

166 Riisgkd, Nielsen, Fuchs, Kasmussen, Obst, & Funch
2002b). In most molecular phylogenetic studies, a sin- gle bryozoan (Halanych et al. 1995, Mackey et al. 1996, Morris et al. 1996) or only one class of bryo- zoans (Cohen et al. 1998, Cohen 2000) has been in- cluded. In a combined morphological and molecular analysis of tribloblastic relationships, including data from three different bryozoan classes, Giribet et al. (2000) did find bryozoan monophyly supported in their preferred total evidence tree, but not in their 18s rDNA trees.
Morphological evidence, drawn mainly from fossils, considers Phylactolaemata as a sister taxon to all other bryozoans (Todd 2000). However, Mundy et al. (1981) suggested that phylactolaemates may be more closely related to phoronids than to marine bryozoans. Phy- lactolaemates commonly have larger zooids compared to other bryozoans, and possess a horseshoe-shaped lophophore instead of circular one. It has been argued that the shape of the lophophore may be an adaptation to a larger size of zooid, a hypothesis that explains similar observations in phoronids and some sabellid polychaetes (Nielsen 2001).
The filter feeding apparatus in P. repens, as de- scribed in this study, is very similar to that of marine bryozoans, such as the gymnolaemate Flustrellidra hispida (Atkins 1933, Nielsen 2002a). It is also similar to that of the phoronid Phoronis muelleri (Riisgbrd 2002). In Plumatella repens, the lophophore only has weak muscles and does not flick the tentacles, but the arrangement of the two zigzag rows of stiff laterofron- tal cilia surrounded by microvilli, located in between the bands of water-pumping lateral cilia and the band of frontal cilia, are similar in all these groups. Fur- thermore, the presence of presumed sensory structures, consisting of abfrontal bundles of cilia that are sur- rounded by microvilli and regularly spaced along the abfrontal side of the tentacles, are described so far only for phylactolaemates (here) and marine bryozoans (Shunatova & Nielsen 2002). The construction of a ciliary feeding apparatus that may depend on a strong central lophophore current is a common feature of phylactolaemates and marine bryozoans.
Acknowledgements. This study has been supported by two grants from the Danish Natural Science Research Council (Peter Funch grant no. 51-00-0278; Hans Ulrik RiisgHrd grant no. 5 1-00-0314). Furthermore, we acknowledge the technical assistance from Else B. Rasmussen and Per G. Henriksen. Thanks are due to Dr. Claw Nielsen and two anonymous referees for constructive comments and criticism of the manuscript.
References Amsellem J & ClCment P 1980. A simplified method for
preparation of rotifers for transmission and scanning elec- tron microscopy. Hydrobiologia 73: 119-122.
Atkins D 1933. The ciliary feeding mechanism of the en- toproct Polyzoa, and a comparison with that of the ecto- proct Polyzoa. Q. J. Microsc. Sci. 75: 393-423.
Bullivant JS 1968. The methods of feeding of lophophorates (Bryozoa, Phoronida, Brachiopoda). New Zeal. J. Mar. Fresh. 2: 135-146.
Cohen BL 2000. Monophyly of brachiopods and phoronids: reconciliation of molecular evidence with Linnaean clas- sification (the subphylum Phoroniformea nov.). Phil. Trans. R. SOC. Lond. B 267: 225-231.
Cohen BL, Gawthrop A, & Cavalier-Smith T 1998. Molec- ular phylogeny of brachiopods and phoronoids based on nuclear-encoded small subunit ribosomal RNA gene se- quences. Phil. Trans. R. SOC. Lond. B 353: 2039-2061.
Gilmour THJ 1978. Ciliation and function of the food-col- lecting and waste-rejecting organs of lophophorates. Can. J. Zool. 56: 2142-2155.
Giribet G, Distel DL, Polz M, Sterrer W, & Wheeler WC 2000. Tribloblastic relationships with emphasis on the acoelomates and the position of Gnathostomulida, Cyclio- phora, Plathelminthes, and Chaetognatha: a combined ap- proach of 18s rDNA sequences and morphology. Syst. Biol. 49: 539-562.
Gordon DP 1974. Microarchitecture and function of the lophophore in the bryozoan Cryptosula pallasiana. Mar. Biol. 27: 147-163.
Halanych KM, Bacheller JD, Aguinaldo AMA, Liva SM, Hillis DM, & Lake JA 1995. Evidence from 18s ribosom- al DNA that the lophophorates are protostome animals. Science 267: 1641-1642.
Larsen PS & RiisgHd HU 2002. On ciliary sieving and pumping in bryozoans. J. Sea Res. 48: 181-195.
Mackey LY, Winnepenninckx, Wachter R De, Backeljau T, Emschermann P, & Garey JR 1996. 18s rRNA suggests that Entoprocta are protostomes, unrelated to Ectoprocta. J Mol Evol 42: 552-559.
Melone G & Ricci C 1995. Rotatory apparatus in bdelloids. Hydrobiologia 313/314: 91-98.
Morris SC, Cohen BL, Gawthrop AB, Cavalier-Smith T, & Winnepenninckx B 1996. Lophophorate phylogeny. Sci- ence 272: 282.
Mukai H, Terakado K, & Reed CG 1997. Bryozoa. In: Mi- croscopic Anatomy of Invertebrates, vol. 13. Lophophor- ates, Entoprocta, and Cycliophora. Harrison FW & Wool- lacott RM, eds., pp. 45-206. Wiley-Liss, New York.
Mundy SP, Taylor PD, & Thorpe JP 1981. A reinterpretation of phylacyolaemate phylogeny. In: Recent and Fossil Bry- ozoa. Larwood GP & Nielsen C, eds., pp 185-190. Olsen & Olsen, Fredensborg, Denmark.
Nielsen C 2001. Animal Evolution: Interrelationships of the Living Phyla. 2nd edition. Oxford University Press, Ox- ford. 563 pp.
Nielsen C 2002a. Ciliary filter-feeding structures in adult and larval gymnolaemate bryozoans. Invertebr. Biol. 121 : 255-261.
Nielsen C 2002b. The phylogenetic position of Entoprocta, Ectoprocta, Phoronida, and Brachiopoda. Integ. Comp. Biol. 42: 685-691.

Ciliary feeding in Plumatella repens 167
Nielsen C & Riisgkd HU 1998. Tentacle structure and filter- feeding in Crisia eburnea and other cyclostomatous bryo- zoans, with a review of upstream-collecting mechanisms. Mar Ecol Prog-Ser 168: 163-186.
Okamua B & Doolan LA 1993. Patterns of suspension feed- ing in the freshwater bryozoan Plumatella repens. Biol
Riisghd HU 2002. Methods of ciliary filter feeding in adult Phoronis muelleri (phylum Phoronida) and its free-swim- ming actinotroch larva. Mar Biol 141: 75-87.
RiisgHrd HU & Manriquez P 1997. Filter-feeding in fifteen marine ectoprocts (Bryozoa): particle capture and water pumping. Mar Ecol Prog-Ser 154: 223-239.
Rosenbaum JL & Carlson K 1969. Cilia regeneration in Tet- rahymena and its inhibition by colchicine. J Cell Biol 40:
Ryland JS 1976. Physiology and ecology of marine bryo-
Shunatova N & Nielsen C 2002. Putative sensory structures
Bull 198: 52-56.
415-425.
zoans. Adv Mar Biol 14: 285-443.
in marine bryozoans. Invertebr. Biol. 121: 262-270.
Shunatova NN & Ostrovsky AN 2001. Individual autozooi- dal behaviour and feeding in marine bryozoans. Sarsia 86:
Strathmann R 1973. Function of lateral cilia in suspension feeding of lophophorates (Brachiopoda, Phoronida, Ecto- procta). Mar. Biol. 23: 129-136.
Strathmann RR 1982. Cinefilms of particle capture by an induced local change of beat of lateral cilia of a bryozoan. J. Exp. Mar. Biol. Ecol. 62: 225-236.
Strathmann RR & McEdward LR 1986. Cyphonautes’ cili- ary sieve breaks a biological rule of inference. Biol. Bull.
Todd JA 2000. The central role of ctenostomes in bryozoan phylogeny. In: Proceedings of the 1 1 th International Bry- ozoology Association Conference. Herrera Cubilla A & Jackson JBC, eds., pp 104-135. Smithsonian Tropical Re- search Institute, Balboa, Panama.
Winston JE 1978. Polypide morphology and feeding behav- ior in marine ectoprocts. Bull. Mar. Sci. 28: 1-31.
113-142.
17 1 : 694-700.
Copyright © 2022 FDOKUMEN




















