
Influence of incidence angle on detecting flooded forests using C-HH synthetic aperture radar data
Upload
independentCategory
view
0download
0
Current Biology 19, 1034–1039, June 23, 2009 ª2009 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2009.04.053
ReportThe Extracellular Domain of SmoothenedRegulates Ciliary Localization and IsRequired for High-Level Hh Signaling
Pia Aanstad,1,2,3,* Nicole Santos,1 Kevin C. Corbit,1
Paul J. Scherz,1,2 Le A. Trinh,1,6 Willi Salvenmoser,4
Jan Huisken,1,2 Jeremy F. Reiter,1,5
and Didier Y.R. Stainier1,2,5
1Department of Biochemistry & BiophysicsPrograms in Developmental Biology, Genetics,
and Human GeneticsCardiovascular Research InstituteDiabetes CenterInstitute for Regeneration Medicine2Liver CenterUniversity of California, San FranciscoSan Francisco, CA 94158USA3Institute of Molecular BiologyCenter for Molecular Biosciences Innsbruck4Institute of ZoologyUniversity of Innsbruck6020 InnsbruckAustria
Summary
Members of the Hedgehog (Hh) family of secreted proteins
function as morphogens to pattern developing tissues andcontrol cell proliferation. The seven-transmembrane domain
(7TM) protein Smoothened (Smo) is essential for the activa-
tion of all levels of Hh signaling. However, the mechanismsby which Smo differentially activates low- or high-level Hh
signaling are not known [1]. Here we show that a newly iden-tified mutation in the extracellular domain (ECD) of zebrafish
Smo attenuates Smo signaling. The Smo agonist purmorph-amine [2] induces the stabilization, ciliary translocation, and
high-level signaling of wild-type Smo. In contrast, pur-morphamine induces the stabilization but not the ciliary
translocation or high-level signaling of the Smo ECD mutantprotein. Surprisingly, a truncated form of Smo that lacks the
cysteine-rich domain of the ECD localizes to the cilium but isunable to activate high-level Hh signaling. We also present
evidence that cilia may be required for Hh signaling in earlyzebrafish embryos. These data indicate that the ECD, previ-
ously thought to be dispensable for vertebrate Smo func-tion, both regulates Smo ciliary localization and is essential
for high-level Hh signaling.
Results and Discussion
The activation of Smoothened (Smo) is correlated with itsstabilization, conformational change, and membrane localiza-tion, although the extent to which these aspects of Smo activa-tion are interrelated is not known [3–11]. In Drosophila, Smo
*Correspondence: [email protected] authors contributed equally to this work6Present address: California Institute of Technology, Pasadena, CA 91125,
USA
accumulates at the plasma membrane upon activation ofHedgehog (Hh) signaling [7], and targeting of Smo to theplasma membrane causes constitutive activation [9, 10]. Inmammals, cilia are required for Hh signaling [12]. Smo local-izes to the primary cilium in response to Hh signaling, andthis localization is required for Smo function [11]. However,the ciliary localization of Smo is not sufficient for activation,because Smo can accumulate at the cilium in the presenceof its inhibitor cyclopamine [13, 14]. The activation of Smomay thus require distinct steps [13], although the mechanismsthat control this process remain unknown.
In a screen for zebrafish mutations affecting early embryonicpatterning, we identified s294. At 24 hr postfertilization (hpf),s294 mutants (Figure 1B) show phenotypes similar to thoseseen in the hi1640 allele of smo (Figure 1C), a presumed null[15]. Mapping data showed that s294 maps close to smo,and complementation tests showed that s294 fails to comple-ment hi1640 (data not shown).
In zebrafish, Hh signaling is required for specification ofmultiple cell types within the somites [16]. Superficial slowfibers (SSFs), expressing slow myosin heavy chain (sMHC)and Prox1, depend on medium- to low-level Hh signaling,whereas muscle pioneer cells (MPs) require maximum levelsof Hh signaling and express high levels of Engrailed (Eng)[16]. In smos294 mutants, MPs were absent and the SSF popu-lation was reduced (Figures 1D–1I), indicating that high-levelHh signaling was abolished and medium- to low-level signalingdiminished. Thus, s294 is a hypomorphic allele of smo.
To identify the molecular lesion in smos294, we sequenced itsopen reading frame and found a G-to-A transition at position374, leading to a cysteine-to-tyrosine substitution at residue125 in the extracellular domain (ECD) of the protein(Figure 1J). The C125 residue is one of ten highly conservedECD cysteines in the Smo/Frizzled (Fz) family of seven-trans-membrane domain (7TM) proteins and is likely to participatein a disulfide bridge that regulates tertiary structure [17]. TheC125Y substitution in Smos294 is thus likely to alter proteinconformation through disruption of a disulfide bond.
To test whether the SmoC125Y substitution alters proteinstability, we generated a version of mouse Smo in which theanalogous cysteine was replaced by tyrosine (SmoC151Y).When expressed in zebrafish embryos or Smo2/2 mouseembryonic fibroblasts (MEFs), SmoC151Y levels were reducedcompared to those of wild-type (WT) Smo (see Figure S1 avail-able online). Treatment with the Smo agonist purmorphamineincreased levels of both WT and mutant Smo to equal to orgreater than that of untreated WT Smo (Figure S1). However,stabilization of SmoC151Y was not sufficient to restore func-tion: whereas WT Smo was able to restore Sonic hedgehog(Shh) and purmorphamine responsiveness to Smo2/2 MEFs,SmoC151Y failed to restore full pathway activity (Figure S1).These results suggest that the ECD is an important determinantof Smo stability and that it is required for full pathway activation.
Previous studies using truncated versions of Smo proposedthat the ECD was dispensable for vertebrate Smo activity [18,19]. However, in Drosophila, three hypomorphic alleles of smohave cysteine substitutions in the ECD similar to that insmos294 (Figure 1J) [9, 20, 21], and a truncated version of

Smo ECD Is Required for High-Level Hh Signaling1035
Drosophila Smo that lacks the ECD fails to rescue Drosophilasmo mutants [9]. To further assess the signaling capabilities ofSmos294, we treated zebrafish smo mutants with purmorph-amine from 8 until 24 hpf. As expected, purmorphamine treat-ment had no effect on smohi1640 mutants at any concentrationtested (data not shown). In contrast, treatment of smos294
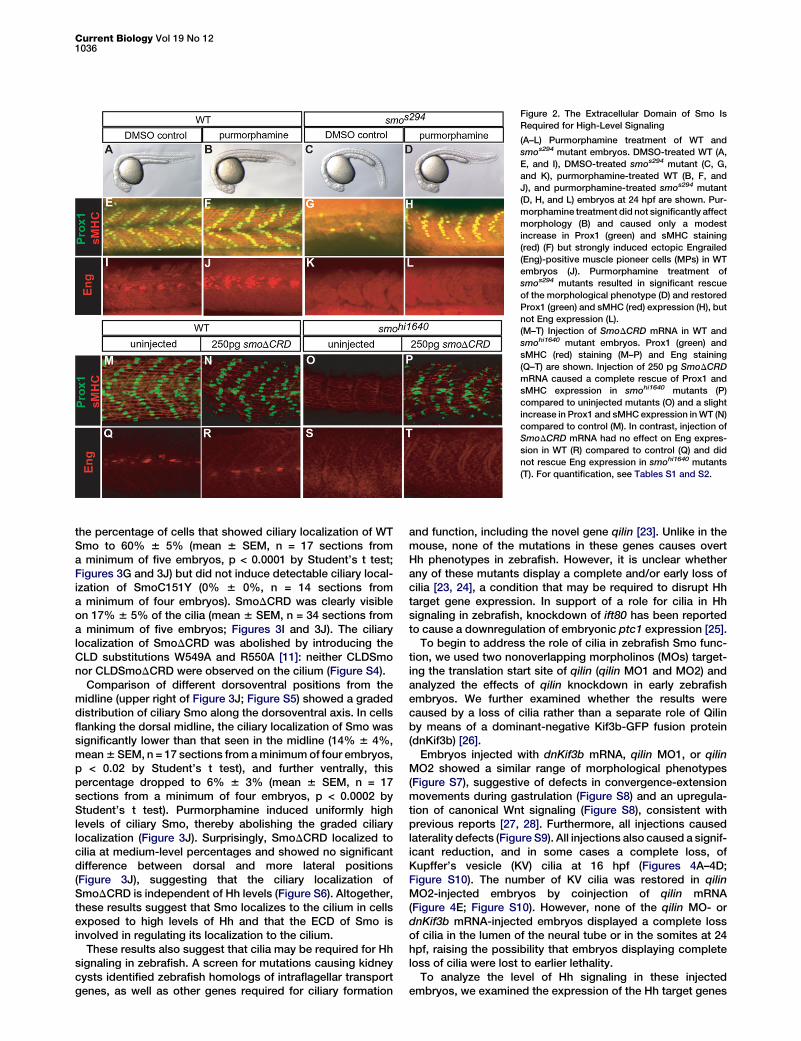
mutants with 20 or 25 mM purmorphamine resulted in a signifi-cant degree of rescue of the morphological phenotype(Figures 2A–2D). Treatment with 25 mM purmorphamineinduced complete rescue of SSFs in at least one somite in43% (10 of 23) of smos294 mutant embryos (21.6 6 0.5 SSFs,mean 6 SEM, n = 13 somites in three independent experi-ments), suggesting that medium- to low-level Hh signalingwas restored (Figures 2E–2H; Table S1). This treatment alsoinduced a small but significant increase in SSFs in WTembryos (Table S1; p < 0.02 by Student’s t test). These resultsindicate that the Smos294 protein retains the ability to signal.Although treatment with 20 mM purmorphamine significantlyincreased the number of MPs in WT embryos (p < 0.0001 byStudent’s t test; Figures 2I and 2J; Table S1), it did not restoreMP formation in smos294 mutants (Figures 2K and 2L). Higherconcentrations of purmorphamine also had no effect (TableS1). Thus, purmorphamine induced high-level Hh responsesin WT embryos but only medium- to low-level signaling insmos294 mutants.
To further test the requirement of the ECD in Smo signaling,we generated Myc-tagged SmoDCRD, a form of mouse Smothat lacks the cysteine-rich domain (amino acids 68–182)[18]. Injection of 250 pg Smo mRNA fully rescued zebrafishsmo mutants [11], and injection of 250 pg SmoDCRD mRNAsimilarly rescued SSF development (Figures 2M–2P; Table S2).
Figure 1. s294 Is a Hypomorphic Allele of smo
with a C125Y Substitution in the Extracellular
Domain
(A–C) Comparison of wild-type (WT) (A), smos294
mutant (B), and smohi1640 mutant (C) embryos at
24 hr postfertilization (hpf) shows that s294
mutants have phenotypes similar to, although
weaker than, the smo null allele hi1640.
(D–F) Immunostaining for Prox1 (green) and slow
myosin heavy chain (sMHC) (red) assesses
medium- to low-level Hedgehog (Hh) signaling
in WT (D), smos294 mutant (E), and smohi1640
mutant (F) embryos at 24 hpf.
(G–I) Immunostaining for Engrailed (red)
assesses high-level Hh signaling in WT (G),
smos294 mutant (H), and smohi1640 mutant (I)
embryos at 24 hpf.
(J) Graphic representation of Smoothened (Smo).
Each amino acid is represented by a circle;
conserved cysteine residues in the extracellular
domain (ECD) are shown in green. The s294
mutation is shown in red; similar Drosophila
cysteine substitution alleles are shown in purple.
In contrast, SmoDCRD could not rescueMPs in smohi1640 mutants, even at higherdoses of mRNA or in the presence ofpurmorphamine (Figures 2Q–2T; TableS2), and similarly failed to restoreGli-luciferase activity in Smo2/2 MEFs(Figure S2). SmoDCRD thus behavedlike stabilized Smos294 in being able toactivate medium- to low-level but nothigh-level signaling. Together, these
results indicate that the ECD of vertebrate Smo, like that ofDrosophila Smo, is essential for full activity in vivo.
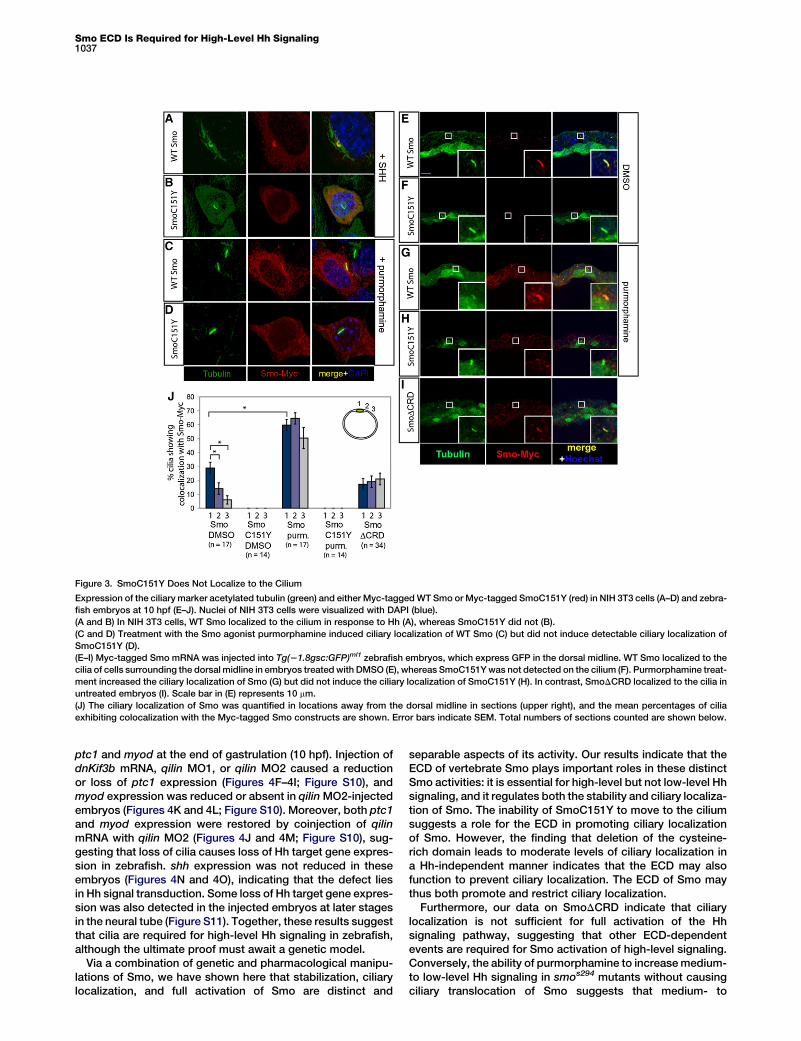
The failure of stabilization of SmoC151Y to restore high-levelsignaling suggests that the SmoC151Y mutation interfereswith other regulatory processes that control Smo activity. Totest whether the C151Y mutation alters trafficking of Smo,we examined the localization of WT Smo and SmoC151Y inNIH 3T3 cells. As shown previously, Smo localizes to the ciliumin response to Hh signaling (Figure 3A; Figure S3) [11]. Incontrast, Hh could not induce any detectable ciliary localiza-tion of SmoC151Y (Figure 3B), as is the case with ciliary local-ization-deficient Smo (CLDSmo) [11] (Figure S3). Similar to Hhexposure, treatment of NIH 3T3 cells with purmorphaminecaused the ciliary localization of WT Smo (Figure 3C) but didnot induce detectable ciliary localization of SmoC151Y orCLDSmo (Figure 3D; Figure S3).
To investigate whether Smo localizes to cilia in zebrafish, weinjected mRNA encoding Myc-tagged Smo, SmoC151Y, andSmoDCRD into zebrafish embryos at the one-cell stage andlooked at subcellular localization in cross-sections using anti-bodies against the Myc tag and acetylated tubulin. To providespatial information, we used Tg(21.8gsc:GFP)ml1 embryos[22], which express green fluorescent protein (GFP) in the cellsof the dorsal midline, a region of shh expression. At the end ofgastrulation (10 hpf), Smo was found localized to primary ciliain 29% 6 4% (mean 6 SEM, n = 17 sections from a minimum offour embryos) of ciliated cells surrounding the GFP-positivedorsal midline (Figures 3E and 3J). In contrast, SmoC151Ywas not detected on cilia (0% 6 0%, mean 6 SEM, n = 14sections from a minimum of six embryos; Figures 3F and 3J).Treatment with 20 mM purmorphamine significantly increased
Current Biology Vol 19 No 121036
Figure 2. The Extracellular Domain of Smo Is
Required for High-Level Signaling
(A–L) Purmorphamine treatment of WT and
smos294 mutant embryos. DMSO-treated WT (A,
E, and I), DMSO-treated smos294 mutant (C, G,
and K), purmorphamine-treated WT (B, F, and
J), and purmorphamine-treated smos294 mutant
(D, H, and L) embryos at 24 hpf are shown. Pur-
morphamine treatment did not significantly affect
morphology (B) and caused only a modest
increase in Prox1 (green) and sMHC staining
(red) (F) but strongly induced ectopic Engrailed
(Eng)-positive muscle pioneer cells (MPs) in WT
embryos (J). Purmorphamine treatment of
smos294 mutants resulted in significant rescue
of the morphological phenotype (D) and restored
Prox1 (green) and sMHC (red) expression (H), but
not Eng expression (L).
(M–T) Injection of SmoDCRD mRNA in WT and
smohi1640 mutant embryos. Prox1 (green) and
sMHC (red) staining (M–P) and Eng staining
(Q–T) are shown. Injection of 250 pg SmoDCRD
mRNA caused a complete rescue of Prox1 and
sMHC expression in smohi1640 mutants (P)
compared to uninjected mutants (O) and a slight
increase in Prox1 and sMHC expression in WT (N)
compared to control (M). In contrast, injection of
SmoDCRD mRNA had no effect on Eng expres-
sion in WT (R) compared to control (Q) and did
not rescue Eng expression in smohi1640 mutants
(T). For quantification, see Tables S1 and S2.
the percentage of cells that showed ciliary localization of WTSmo to 60% 6 5% (mean 6 SEM, n = 17 sections froma minimum of five embryos, p < 0.0001 by Student’s t test;Figures 3G and 3J) but did not induce detectable ciliary local-ization of SmoC151Y (0% 6 0%, n = 14 sections froma minimum of four embryos). SmoDCRD was clearly visibleon 17% 6 5% of the cilia (mean 6 SEM, n = 34 sections froma minimum of five embryos; Figures 3I and 3J). The ciliarylocalization of SmoDCRD was abolished by introducing theCLD substitutions W549A and R550A [11]: neither CLDSmonor CLDSmoDCRD were observed on the cilium (Figure S4).
Comparison of different dorsoventral positions from themidline (upper right of Figure 3J; Figure S5) showed a gradeddistribution of ciliary Smo along the dorsoventral axis. In cellsflanking the dorsal midline, the ciliary localization of Smo wassignificantly lower than that seen in the midline (14% 6 4%,mean 6 SEM, n = 17 sections from a minimum of four embryos,p < 0.02 by Student’s t test), and further ventrally, thispercentage dropped to 6% 6 3% (mean 6 SEM, n = 17sections from a minimum of four embryos, p < 0.0002 byStudent’s t test). Purmorphamine induced uniformly highlevels of ciliary Smo, thereby abolishing the graded ciliarylocalization (Figure 3J). Surprisingly, SmoDCRD localized tocilia at medium-level percentages and showed no significantdifference between dorsal and more lateral positions(Figure 3J), suggesting that the ciliary localization ofSmoDCRD is independent of Hh levels (Figure S6). Altogether,these results suggest that Smo localizes to the cilium in cellsexposed to high levels of Hh and that the ECD of Smo isinvolved in regulating its localization to the cilium.
These results also suggest that cilia may be required for Hhsignaling in zebrafish. A screen for mutations causing kidneycysts identified zebrafish homologs of intraflagellar transportgenes, as well as other genes required for ciliary formation
and function, including the novel gene qilin [23]. Unlike in themouse, none of the mutations in these genes causes overtHh phenotypes in zebrafish. However, it is unclear whetherany of these mutants display a complete and/or early loss ofcilia [23, 24], a condition that may be required to disrupt Hhtarget gene expression. In support of a role for cilia in Hhsignaling in zebrafish, knockdown of ift80 has been reportedto cause a downregulation of embryonic ptc1 expression [25].
To begin to address the role of cilia in zebrafish Smo func-tion, we used two nonoverlapping morpholinos (MOs) target-ing the translation start site of qilin (qilin MO1 and MO2) andanalyzed the effects of qilin knockdown in early zebrafishembryos. We further examined whether the results werecaused by a loss of cilia rather than a separate role of Qilinby means of a dominant-negative Kif3b-GFP fusion protein(dnKif3b) [26].
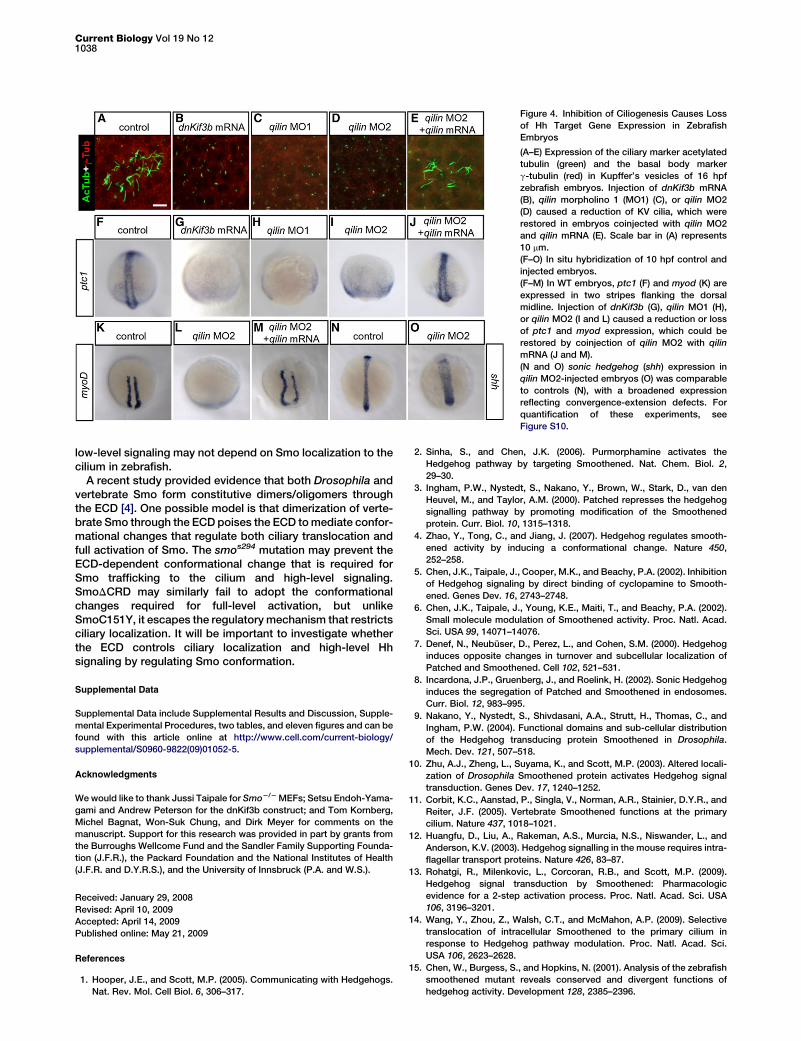
Embryos injected with dnKif3b mRNA, qilin MO1, or qilinMO2 showed a similar range of morphological phenotypes(Figure S7), suggestive of defects in convergence-extensionmovements during gastrulation (Figure S8) and an upregula-tion of canonical Wnt signaling (Figure S8), consistent withprevious reports [27, 28]. Furthermore, all injections causedlaterality defects (Figure S9). All injections also caused a signif-icant reduction, and in some cases a complete loss, ofKupffer’s vesicle (KV) cilia at 16 hpf (Figures 4A–4D;Figure S10). The number of KV cilia was restored in qilinMO2-injected embryos by coinjection of qilin mRNA(Figure 4E; Figure S10). However, none of the qilin MO- ordnKif3b mRNA-injected embryos displayed a complete lossof cilia in the lumen of the neural tube or in the somites at 24hpf, raising the possibility that embryos displaying completeloss of cilia were lost to earlier lethality.
To analyze the level of Hh signaling in these injectedembryos, we examined the expression of the Hh target genes
Smo ECD Is Required for High-Level Hh Signaling1037
Figure 3. SmoC151Y Does Not Localize to the Cilium
Expression of the ciliary marker acetylated tubulin (green) and either Myc-tagged WT Smo or Myc-tagged SmoC151Y (red) in NIH 3T3 cells (A–D) and zebra-
fish embryos at 10 hpf (E–J). Nuclei of NIH 3T3 cells were visualized with DAPI (blue).
(A and B) In NIH 3T3 cells, WT Smo localized to the cilium in response to Hh (A), whereas SmoC151Y did not (B).
(C and D) Treatment with the Smo agonist purmorphamine induced ciliary localization of WT Smo (C) but did not induce detectable ciliary localization of
SmoC151Y (D).
(E–I) Myc-tagged Smo mRNA was injected into Tg(21.8gsc:GFP)ml1 zebrafish embryos, which express GFP in the dorsal midline. WT Smo localized to the
cilia of cells surrounding the dorsal midline in embryos treated with DMSO (E), whereas SmoC151Y was not detected on the cilium (F). Purmorphamine treat-
ment increased the ciliary localization of Smo (G) but did not induce the ciliary localization of SmoC151Y (H). In contrast, SmoDCRD localized to the cilia in
untreated embryos (I). Scale bar in (E) represents 10 mm.
(J) The ciliary localization of Smo was quantified in locations away from the dorsal midline in sections (upper right), and the mean percentages of cilia
exhibiting colocalization with the Myc-tagged Smo constructs are shown. Error bars indicate SEM. Total numbers of sections counted are shown below.
ptc1 and myod at the end of gastrulation (10 hpf). Injection ofdnKif3b mRNA, qilin MO1, or qilin MO2 caused a reductionor loss of ptc1 expression (Figures 4F–4I; Figure S10), andmyod expression was reduced or absent in qilin MO2-injectedembryos (Figures 4K and 4L; Figure S10). Moreover, both ptc1and myod expression were restored by coinjection of qilinmRNA with qilin MO2 (Figures 4J and 4M; Figure S10), sug-gesting that loss of cilia causes loss of Hh target gene expres-sion in zebrafish. shh expression was not reduced in theseembryos (Figures 4N and 4O), indicating that the defect liesin Hh signal transduction. Some loss of Hh target gene expres-sion was also detected in the injected embryos at later stagesin the neural tube (Figure S11). Together, these results suggestthat cilia are required for high-level Hh signaling in zebrafish,although the ultimate proof must await a genetic model.
Via a combination of genetic and pharmacological manipu-lations of Smo, we have shown here that stabilization, ciliarylocalization, and full activation of Smo are distinct and
separable aspects of its activity. Our results indicate that theECD of vertebrate Smo plays important roles in these distinctSmo activities: it is essential for high-level but not low-level Hhsignaling, and it regulates both the stability and ciliary localiza-tion of Smo. The inability of SmoC151Y to move to the ciliumsuggests a role for the ECD in promoting ciliary localizationof Smo. However, the finding that deletion of the cysteine-rich domain leads to moderate levels of ciliary localization ina Hh-independent manner indicates that the ECD may alsofunction to prevent ciliary localization. The ECD of Smo maythus both promote and restrict ciliary localization.
Furthermore, our data on SmoDCRD indicate that ciliarylocalization is not sufficient for full activation of the Hhsignaling pathway, suggesting that other ECD-dependentevents are required for Smo activation of high-level signaling.Conversely, the ability of purmorphamine to increase medium-to low-level Hh signaling in smos294 mutants without causingciliary translocation of Smo suggests that medium- to
Current Biology Vol 19 No 121038
low-level signaling may not depend on Smo localization to thecilium in zebrafish.
A recent study provided evidence that both Drosophila andvertebrate Smo form constitutive dimers/oligomers throughthe ECD [4]. One possible model is that dimerization of verte-brate Smo through the ECD poises the ECD to mediate confor-mational changes that regulate both ciliary translocation andfull activation of Smo. The smos294 mutation may prevent theECD-dependent conformational change that is required forSmo trafficking to the cilium and high-level signaling.SmoDCRD may similarly fail to adopt the conformationalchanges required for full-level activation, but unlikeSmoC151Y, it escapes the regulatory mechanism that restrictsciliary localization. It will be important to investigate whetherthe ECD controls ciliary localization and high-level Hhsignaling by regulating Smo conformation.
Supplemental Data
Supplemental Data include Supplemental Results and Discussion, Supple-
mental Experimental Procedures, two tables, and eleven figures and can be
found with this article online at http://www.cell.com/current-biology/
supplemental/S0960-9822(09)01052-5.
Acknowledgments
We would like to thank Jussi Taipale for Smo2/2 MEFs; Setsu Endoh-Yama-
gami and Andrew Peterson for the dnKif3b construct; and Tom Kornberg,
Michel Bagnat, Won-Suk Chung, and Dirk Meyer for comments on the
manuscript. Support for this research was provided in part by grants from
the Burroughs Wellcome Fund and the Sandler Family Supporting Founda-
tion (J.F.R.), the Packard Foundation and the National Institutes of Health
(J.F.R. and D.Y.R.S.), and the University of Innsbruck (P.A. and W.S.).
Received: January 29, 2008
Revised: April 10, 2009
Accepted: April 14, 2009
Published online: May 21, 2009
References
1. Hooper, J.E., and Scott, M.P. (2005). Communicating with Hedgehogs.
Nat. Rev. Mol. Cell Biol. 6, 306–317.
2. Sinha, S., and Chen, J.K. (2006). Purmorphamine activates the
Hedgehog pathway by targeting Smoothened. Nat. Chem. Biol. 2,
29–30.
3. Ingham, P.W., Nystedt, S., Nakano, Y., Brown, W., Stark, D., van den
Heuvel, M., and Taylor, A.M. (2000). Patched represses the hedgehog
signalling pathway by promoting modification of the Smoothened
protein. Curr. Biol. 10, 1315–1318.
4. Zhao, Y., Tong, C., and Jiang, J. (2007). Hedgehog regulates smooth-
ened activity by inducing a conformational change. Nature 450,
252–258.
5. Chen, J.K., Taipale, J., Cooper, M.K., and Beachy, P.A. (2002). Inhibition
of Hedgehog signaling by direct binding of cyclopamine to Smooth-
ened. Genes Dev. 16, 2743–2748.
6. Chen, J.K., Taipale, J., Young, K.E., Maiti, T., and Beachy, P.A. (2002).
Small molecule modulation of Smoothened activity. Proc. Natl. Acad.
Sci. USA 99, 14071–14076.
7. Denef, N., Neubuser, D., Perez, L., and Cohen, S.M. (2000). Hedgehog
induces opposite changes in turnover and subcellular localization of
Patched and Smoothened. Cell 102, 521–531.
8. Incardona, J.P., Gruenberg, J., and Roelink, H. (2002). Sonic Hedgehog
induces the segregation of Patched and Smoothened in endosomes.
Curr. Biol. 12, 983–995.
9. Nakano, Y., Nystedt, S., Shivdasani, A.A., Strutt, H., Thomas, C., and
Ingham, P.W. (2004). Functional domains and sub-cellular distribution
of the Hedgehog transducing protein Smoothened in Drosophila.
Mech. Dev. 121, 507–518.
10. Zhu, A.J., Zheng, L., Suyama, K., and Scott, M.P. (2003). Altered locali-
zation of Drosophila Smoothened protein activates Hedgehog signal
transduction. Genes Dev. 17, 1240–1252.
11. Corbit, K.C., Aanstad, P., Singla, V., Norman, A.R., Stainier, D.Y.R., and
Reiter, J.F. (2005). Vertebrate Smoothened functions at the primary
cilium. Nature 437, 1018–1021.
12. Huangfu, D., Liu, A., Rakeman, A.S., Murcia, N.S., Niswander, L., and
Anderson, K.V. (2003). Hedgehog signalling in the mouse requires intra-
flagellar transport proteins. Nature 426, 83–87.
13. Rohatgi, R., Milenkovic, L., Corcoran, R.B., and Scott, M.P. (2009).
Hedgehog signal transduction by Smoothened: Pharmacologic
evidence for a 2-step activation process. Proc. Natl. Acad. Sci. USA
106, 3196–3201.
14. Wang, Y., Zhou, Z., Walsh, C.T., and McMahon, A.P. (2009). Selective
translocation of intracellular Smoothened to the primary cilium in
response to Hedgehog pathway modulation. Proc. Natl. Acad. Sci.
USA 106, 2623–2628.
15. Chen, W., Burgess, S., and Hopkins, N. (2001). Analysis of the zebrafish
smoothened mutant reveals conserved and divergent functions of
hedgehog activity. Development 128, 2385–2396.
Figure 4. Inhibition of Ciliogenesis Causes Loss
of Hh Target Gene Expression in Zebrafish
Embryos
(A–E) Expression of the ciliary marker acetylated
tubulin (green) and the basal body marker
g-tubulin (red) in Kupffer’s vesicles of 16 hpf
zebrafish embryos. Injection of dnKif3b mRNA
(B), qilin morpholino 1 (MO1) (C), or qilin MO2
(D) caused a reduction of KV cilia, which were
restored in embryos coinjected with qilin MO2
and qilin mRNA (E). Scale bar in (A) represents
10 mm.
(F–O) In situ hybridization of 10 hpf control and
injected embryos.
(F–M) In WT embryos, ptc1 (F) and myod (K) are
expressed in two stripes flanking the dorsal
midline. Injection of dnKif3b (G), qilin MO1 (H),
or qilin MO2 (I and L) caused a reduction or loss
of ptc1 and myod expression, which could be
restored by coinjection of qilin MO2 with qilin
mRNA (J and M).
(N and O) sonic hedgehog (shh) expression in
qilin MO2-injected embryos (O) was comparable
to controls (N), with a broadened expression
reflecting convergence-extension defects. For
quantification of these experiments, see
Figure S10.

Smo ECD Is Required for High-Level Hh Signaling1039
16. Wolff, C., Roy, S., and Ingham, P.W. (2003). Multiple muscle cell identi-
ties induced by distinct levels and timing of hedgehog activity in the
zebrafish embryo. Curr. Biol. 13, 1169–1181.
17. Dann, C.E., Hsieh, J.C., Rattner, A., Sharma, D., Nathans, J., and Leahy,
D.J. (2001). Insights into Wnt binding and signalling from the structures
of two Frizzled cysteine-rich domains. Nature 412, 86–90.
18. Taipale, J., Cooper, M.K., Maiti, T., and Beachy, P.A. (2002). Patched
acts catalytically to suppress the activity of Smoothened. Nature 418,
892–897.
19. Murone, M., Rosenthal, A., and de Sauvage, F.J. (1999). Sonic hedgehog
signaling by the Patched-Smoothened receptor complex. Curr. Biol.
9, 76–84.
20. Chen, Y., and Struhl, G. (1998). In vivo evidence that Patched and
Smoothened constitute distinct binding and transducing components
of a Hedgehog receptor complex. Development 125, 4943–4948.
21. Alcedo, J., Zou, Y., and Noll, M. (2000). Posttranscriptional regulation of
Smoothened is part of a self-correcting mechanism in the Hedgehog
signaling system. Mol. Cell 6, 457–465.
22. Doitsidou, M., Reichman-Fried, M., Stebler, J., Koprunner,M., Dorries, J.,
Meyer, D., Esquerra, C.V., Leung, T., and Raz, E. (2002). Guidance of
primordial germ cell migration by the chemokine SDF-1. Cell 111,
647–659.
23. Sun, Z., Amsterdam, A., Pazour, G.J., Cole, D.G., Miller, M.S., and
Hopkins, N. (2004). A genetic screen in zebrafish identifies cilia genes
as a principal cause of cystic kidney. Development 131, 4085–4093.
24. Tsujikawa, M., and Malicki, J. (2004). Intraflagellar transport genes are
essential for differentiation and survival of vertebrate sensory neurons.
Neuron 42, 703–716.
25. Beales, P.L.,Bland, E.,Tobin, J.L., Bacchelli, C.,Tuysuz,B.,Hill, J., Rix, S.,
Pearson, C.G., Kai, M., Hartley, J., et al. (2007). IFT80, which encodes
a conserved intraflagellar transport protein, is mutated in Jeune asphyx-
iating thoracic dystrophy. Nat. Genet. 39, 727–729.
26. Haraguchi, K., Hayashi, T., Jimbo, T., Yamamoto, T., and Akiyama, T.
(2006). Role of the kinesin-2 family protein, KIF3, during mitosis.
J. Biol. Chem. 281, 4094–4099.
27. Gerdes, J.M., Liu, Y., Zaghloul, N.A., Leitch, C.C., Lawson, S.S., Kato, M.,
Beachy, P.A., Beales, P.L., DeMartino, G.N., Fisher, S., et al. (2007).
Disruption of the basal body compromises proteasomal function and
perturbs intracellular Wnt response. Nat. Genet. 39, 1350–1360.
28. Corbit, K.C., Shyer, A.E.,Dowdle,W.E., Gaulden, J., Singla, V., Chen, M.H.,
Chuang, P.T., and Reiter, J.F. (2008). Kif3a constrains beta-catenin-
dependent signalling through dual ciliary and non-ciliary mechanisms.
Nat. Cell Biol. 10, 70–76.
Copyright © 2022 FDOKUMEN




















