
Muscarinic Receptor Subtypes in Human Iris-Ciliary Body Measured by Immunoprecipitation
9
Muscarinic Receptor Subtypes in Human Iris-Ciliary Body Measured by Immunoprecipitation Daniel W. Gil, Heather A. Krauss, Alicia M. Bogardus, and Elizabeth WoldeMussie Purpose. To determine the relative levels of the five muscarinic receptor subtypes in the anterior segment of the human eye. Methods. Antisera selective for each of the five muscarinic receptor proteins were incubated with [ a H]-QNB bound receptors solubilized from human iris sphincter, ciliary muscle, and ciliary processes. Precipitation of the radiolabeled receptor-antibody complexes and scintilla- tion counting enabled quantitation of the subtypes in the various tissues. Reverse transcrip- tion-polymerase chain reaction was performed on the tissues and cultured smooth muscle cells derived from them. Results. Approximately 60% to 75% of the muscarinic receptors in the human iris sphincter and ciliary body are the m3 subtype. Lower levels (5% to 10%) of the m2 and m4 receptors are present in these tissues. The ml receptor (7%) was detected in the ciliary processes and iris sphincter and the m5 receptor (5%), which is usually found only in the central nervous system, was present in the iris sphincter. Conclusions. The m3 subtype is the predominant muscarinic receptor in the anterior segment of the human eye. The extensive heterogeneity of muscarinic receptors makes it difficult to predict whether subtype-selective drugs will have an improved efficacy and side-effect profile. Invest Ophthalmol Vis Sci. 1997;38:1434-1442. A he clinical use of cholinergic drugs for many de- cades has revealed multiple actions mediated by mus- carinic receptors in the eye. These include reduction of pupil diameter by contraction of the iris sphincter, accommodation of the lens and enhanced aqueous humor outflow as a result of ciliary muscle contrac- tion, and increased aqueous humor secretion from the ciliary processes. The discovery of muscarinic re- ceptor subtypes that could be differentiated pharma- cologically with selective antagonists 12 and could cou- ple to different second messenger pathways 3 raised the possibility that the various effects of muscarinic drugs on the eye were elicited through different recep- tors. An M3 receptor that binds pirenzepine with inter- mediate affinity and 4-DAMP with high affinity and activates phosphoinositide (PI) hydrolysis was demon- strated in the cynomolgus monkey ciliary body, 4 hu- man ciliary muscle cells,' human nonpigmented epi- gram the Department of Biological Sciences, Alkrgan, Irvine, California. August 27, 1996: revised December 27, 1996; accepted Submitted for publ Februmy 6, 1997. l*r<tprietary interest category: E. Reprint requests: Daniel W. Gil, Biological Si: Dupont Drive, Irvine CA 92612. t (RD-2C), Allergtm, Inc., 2525 thelial cells, 6 ' 7 the rabbit iris sphincter, 8 l0 and human iris sphincter cells. 5 Tobin and Osborne" identified an M2 receptor that inhibits adenylate cyclase activity and has low affinity for pirenzepine in the rabbit cili- ary epithelium, providing the first evidence for a sec- ond muscarinic receptor in the eye. Its ability to atten- uate isoproterenol stimulation of cAMP levels sug- gested a modulatory role on aqueous humor production. 12 A population of Ml receptors that bind pirenzepine with high affinity and elicit PI hydrolysis was also identified in the rabbit ciliary processes. 13 Evidence of muscarinic receptor heterogeneity was also found in the rabbit iris sphincter 14 and in human ciliary smooth muscle, 5 but the particular subtypes in- volved could not be determined because of the limited selectivity of the available ligands. Five subtypes of muscarinic receptors have now been characterized by gene cloning, lr '~ 17 enabling a more detailed classification of the ocular muscarinic receptors. In the commonly accepted International Union of Pharmacology nomenclature, the genetically defined ml to m3 receptors correspond to the phar- macologic Ml to M3 receptors. Reverse transcription- polymerase chain reaction (RT-PCR) studies 18 and in 1434 Investigative Ophthalmology & Visual Science, June 1997, Vol. 38, No. 7 Copyright. © Association for Research in Vision and Ophthalmology
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Muscarinic Receptor Subtypes in Human Iris-Ciliary Body Measured by Immunoprecipitation

Muscarinic Receptor Subtypes in Human Iris-Ciliary BodyMeasured by Immunoprecipitation
Daniel W. Gil, Heather A. Krauss, Alicia M. Bogardus, and Elizabeth WoldeMussie
Purpose. To determine the relative levels of the five muscarinic receptor subtypes in theanterior segment of the human eye.
Methods. Antisera selective for each of the five muscarinic receptor proteins were incubatedwith [aH]-QNB bound receptors solubilized from human iris sphincter, ciliary muscle, andciliary processes. Precipitation of the radiolabeled receptor-antibody complexes and scintilla-tion counting enabled quantitation of the subtypes in the various tissues. Reverse transcrip-tion-polymerase chain reaction was performed on the tissues and cultured smooth musclecells derived from them.
Results. Approximately 60% to 75% of the muscarinic receptors in the human iris sphincterand ciliary body are the m3 subtype. Lower levels (5% to 10%) of the m2 and m4 receptorsare present in these tissues. The ml receptor (7%) was detected in the ciliary processes andiris sphincter and the m5 receptor (5%), which is usually found only in the central nervoussystem, was present in the iris sphincter.
Conclusions. The m3 subtype is the predominant muscarinic receptor in the anterior segmentof the human eye. The extensive heterogeneity of muscarinic receptors makes it difficult topredict whether subtype-selective drugs will have an improved efficacy and side-effect profile.Invest Ophthalmol Vis Sci. 1997;38:1434-1442.
A he clinical use of cholinergic drugs for many de-cades has revealed multiple actions mediated by mus-carinic receptors in the eye. These include reductionof pupil diameter by contraction of the iris sphincter,accommodation of the lens and enhanced aqueoushumor outflow as a result of ciliary muscle contrac-tion, and increased aqueous humor secretion fromthe ciliary processes. The discovery of muscarinic re-ceptor subtypes that could be differentiated pharma-cologically with selective antagonists12 and could cou-ple to different second messenger pathways3 raisedthe possibility that the various effects of muscarinicdrugs on the eye were elicited through different recep-tors. An M3 receptor that binds pirenzepine with inter-mediate affinity and 4-DAMP with high affinity andactivates phosphoinositide (PI) hydrolysis was demon-strated in the cynomolgus monkey ciliary body,4 hu-man ciliary muscle cells,' human nonpigmented epi-gram the Department of Biological Sciences, Alkrgan, Irvine, California.
August 27, 1996: revised December 27, 1996; acceptedSubmitted for publFebrumy 6, 1997.l*r<tprietary interest category: E.Reprint requests: Daniel W. Gil, Biological Si:Dupont Drive, Irvine CA 92612.
t (RD-2C), Allergtm, Inc., 2525
thelial cells,6'7 the rabbit iris sphincter,8 l0 and humaniris sphincter cells.5 Tobin and Osborne" identifiedan M2 receptor that inhibits adenylate cyclase activityand has low affinity for pirenzepine in the rabbit cili-ary epithelium, providing the first evidence for a sec-ond muscarinic receptor in the eye. Its ability to atten-uate isoproterenol stimulation of cAMP levels sug-gested a modulatory role on aqueous humorproduction.12 A population of Ml receptors that bindpirenzepine with high affinity and elicit PI hydrolysiswas also identified in the rabbit ciliary processes.13
Evidence of muscarinic receptor heterogeneity wasalso found in the rabbit iris sphincter14 and in humanciliary smooth muscle,5 but the particular subtypes in-volved could not be determined because of the limitedselectivity of the available ligands.
Five subtypes of muscarinic receptors have nowbeen characterized by gene cloning,lr'~17 enabling amore detailed classification of the ocular muscarinicreceptors. In the commonly accepted InternationalUnion of Pharmacology nomenclature, the geneticallydefined ml to m3 receptors correspond to the phar-macologic Ml to M3 receptors. Reverse transcription-polymerase chain reaction (RT-PCR) studies18 and in
1434Investigative Ophthalmology & Visual Science, June 1997, Vol. 38, No. 7Copyright. © Association for Research in Vision and Ophthalmology

Inununoprecipitation of mACh Receptor Subtypes in the Human Eye 1435
situ hybridization19 on human ciliary smooth muscletissue and cells identified mRNA encoding both them2 and m3 receptors. Zhang et al19 also found evi-dence for the expression of the other three subtypesin the ciliary muscle. Northern blot hybridization onRNA from the bovine iris sphincter and ciliary pro-cesses using subtype-specific oligonucleotide probes20
also demonstrated the expression of multiple musca-rinic receptor subtypes in these tissues. Surprisingly,little or no mRNA encoding the m3 receptor was de-tected, whereas transcripts encoding the m4 (re-ported20 as m3 using a different nomenclature)16 andsome m2 receptor were expressed.
Because these molecular studies were not quanti-tative and could not provide information on levels ofreceptor protein, we have immunoprecipitated musca-rinic receptors from tissues of the human anteriorchamber using receptor subtype-selective antibodies.21
The results confirm that the m3 subtype is the pre-dominant muscarinic receptor. Discrete populationsof the other subtypes, including the m5 subtype, whichis usually found only in the central nervous system,were also present.
METHODS
Tissue Preparation
Chinese hamster ovary cell lines expressing the hu-man ml to m5 muscarinic receptors were originallyconstructed by Bonner et al1517 and were obtainedfrom the National Institutes of Health. The cells weregrown in 50% F12 Ham's-50% Dulbecco's modifiedEagle's medium (Gibco, Gaithersburg, MD) supple-mented with 10% fetal calf serum and 2000 U/l peni-cillin and streptomycin. Passage 4 cells were used forall experiments.
Human ocular tissues were obtained from theUniversity of Wisconsin Eye Bank. The iris-ciliarybody tissue was cut from the eye immediately afterenucleation and frozen at — 70°C. Dissections of ciliarybody and iris sphincter were attained on ice. The cili-ary processes were carefully peeled away from the cili-ary body thus separating the muscle and the ciliaryepithelial layers. The iris sphincter was cut with finescissors at the junction of the sphincter and dilatormuscle. Iris sphincters from 73 donors (mean age of46) and ciliary bodies from 10 donors (mean age of64) were used for immunoprecipitation studies. Re-verse transcription-polymerase chain reaction studieswere done with iris sphincters from five donors (ages11, 43, 53, 60, and 64) and ciliary bodies from ninedonors (mean age, 34).
For culturing cells from ciliary or iris sphinctermuscles, eyes from a 65- and a 56-year-old man, respec-tively, enucleated 2 to 5 hours after death and kept
on ice, were obtained from the Lions-University ofCalifornia, Irvine Organ and Tissue Bank. Dissectionof tissues was performed under sterile conditions us-ing a dissection microscope. The ciliary muscle wasexposed by peeling away the ciliary processes. Themuscle was then removed and minced into smallpieces. The iris was removed from the eye, and thepigmented epithelial layer was scraped off. Thesphincter was then separated from the dilator andminced into small pieces. Tissues were placed in aPetri dish and cultured as described previously.5
Briefly, cells were allowed to migrate from the explantsfor 10 to 14 days in D-valine-containing minimal es-sential medium supplemented with 10% fetal calf se-rum, 2 mM glutamine, 100 U/ml penicillin, 100 fxg/ml streptomycin, and 25 /xg/ml Fungizone (Gibco).Cultures free of fibroblasts were maintained in me-dium 199 containing 6% glucose (Gibco). Passage 4and 5 cells were used for RNA isolation.
Dissected frozen tissues or cells were homoge-nized with a polytron (Brinkmann, Westbury, NY; set-ting 6: 15 to 20 seconds) in 5 to 12 ml 20 mM Tris, 1mM EDTA, pH 7.4, on ice. The homogenates werecentrifuged at 23,700g in a Beckman JA-20 rotor for15 minutes at 4°C. Pellets were resuspended in 2.5 to6 ml 10 mM Tris, 1 mM EDTA, pH 7.4 (TE) using atissuemizer (Tekmar; Cincinnati, OH; setting 60: 10to 15 seconds). Protein concentration was measuredwith the BioRad (Hercules, CA) protein assay. Typi-cally, the homogenate from one rat cortex contained4 mg protein, ciliary muscle from 2 eyes 1.9 mg pro-tein, the ciliary processes from 2 eyes 0.9 mg protein,and 20 iris sphincters 1 mg protein.
To determine receptor density, aliquots con-taining approximately 10 /xg protein were incubatedin 1 ml TE containing 0.5 nM [3H]-QNB (38.8 to44.9 Ci/mmol; Dupont-New England Nuclear, Bos-ton, MA) for 45 minutes at 32°C. Nonspecific bindingwas defined in the presence of 5 fjM atropine. Reac-tions were stopped with the addition of 3 ml ice-coldTE buffer and filtered through Whatman (Hillsboro,OR) GF/B glass fiber filters. The filters were washedtwice with 3 ml TE, dried, and bound [SH]-QNB wasmeasured by scintillation counting. The Bmax valuesdetermined for preparations of rat brain, ciliary mus-cle, ciliary processes and iris sphincter were 3, 6.6, 5.1and 2 pmol/mg protein, respectively.
Inununoprecipitation
Antisera against the five muscarinic receptor subtypeswere graciously provided by Barry Wolfe (GeorgetownUniversity, Washington, DC). Receptor labeling, solu-bilization and immunoprecipitation were carried outessentially as previously described.22 Briefly, mem-branes were incubated with 0.5 nM [3H]-QNB for 45minutes at 32° in 25 ml of TE buffer containing a

1436 Investigative Ophthalmology & Visual Science, June 1997, Vol. 38, No. 7
cocktail of protease inhibitors (TE/PIC). The labeledmembranes were pelleted and washed three times toremove unbound [3H]-QNB. All further steps weredone at 4°C to prevent dissociation of the label. Themembrane pellet was resuspended in TE containing1% digitonin and 0.2% cholic acid (1% TEDC) at afinal concentration of approximately 1 mg protein/ml, solubilized for 45 minutes, and centrifuged at80,000gfor 45 minutes. Typically, 40% to 45% of thelabeled receptors were solubilized with no apparentdifference between the subtypes.
The five muscarinic receptor subtypes and a con-trol antisera were incubated in duplicate or triplicateat a concentration of 0.6 mg/ml (0.75 mg/ml for m2)with aliquots of 100 to 200 fmole (4 to 6000 cpm) ofsolubilized labeled receptors in 0.5 ml TE/PIC for 44to 48 hours. Residual free [3H]-QNB was removedwith a 10-minute incubation in 0.75 ml of a 2.7% se-rum-treated charcoal solution in 0.1% TEDC.
Supernatant (1-15 ml) was combined with 0.2 mlof 2 X stripped Pansorbin (Calbiochem, Lajolla, CA;protein A-coated Staphylococcus aureus cells) that wasprepared the previous day. After a 30-minute incuba-tion with constant gentle rotation, the receptor-anti-body-Pansorbin complex was pelleted at 13,000g. Thesupernatant and pellet were separated, 0.2 ml of 0.1N NaOH-3% sodium deoxycholate was added, andthe volume of each fraction was brought up to 1.75ml. Ecoscint scintillation cocktail (10 ml) was addedand the counts per minute of [3H] -QNB labeled re-ceptor in each fraction determined by scintillationcounting.
Nonspecific precipitation by the control antiserarepresented 2% to 3% of total labeled receptors andwas subtracted from the pellet values. Control samplesthat had been labeled in the presence of 5 fiM atro-pine demonstrated that nonspecific [3H]-QNB bind-ing accounted for less than 5% of die total counts perminute. Percentage of precipitation by each antibodywas calculated by dividing the adjusted pellet countsper minute by the combined pellet and supernatantcounts per minute.
Reverse Transcription -Polymerase ChainReactionTo avoid artifact bands caused by contaminating geno-mic DNA, tagged subtype-specific antisense primerswith a 5' 30 bp tag were used for reverse transcription.Subsequent polymerase chain reaction (PCR) ampli-fication was carried out with a subtype-specific senseprimer and the unique tag sequence as the antisenseprimer. This "RNA template-specific PCR" has beendescribed previously by Shuldiner et al.23
Total RNA was prepared from cells and oculartissues that had been frozen no more than 5 hoursafter death by guanidinium thiocyanate- phenol -
chloroform extraction.24 Reverse transcription wascarried out on 1 to 2 /J,g total RNA using the firststrand synthesis kit from Boehringer Mannheim (Indi-anapolis, IN). A cDNA aliquot was denatured at 95°Cfor 5 minutes and combined in a 50-/̂ 1 volume with1.5 mM MgCl2, 100 /JM dNTPs, 0.625 units Ampli Taqpolymerase (Perkin Elmer, Foster City, CA), 25 //Mprimers and PCR buffer (50 mM KC1; 100 mM Tris-HC1, pH 8.3) for 40 cycles of PCR in a Perkin Elmer9600 thermal cycler. The standard cycle included a94°C, 30-second denaturation; a 60°C to 63°C, 1-mi-nute annealing, and a 72°C, 1-minute extension.There was a 6-minute extension step at the end. Prod-ucts were analyzed by agarose gel electrophoresis.
The tag sequence was 5' GAA CAT CGA TGA CAAGCT TAG GTA TCG ATA 3'. The subtype-specificantisense sequences to which the tag was appendedwere, ml: 3' CAT GGA GCC TTC GTC CTC T (nt837 to 855); m2: 3' AAC ACA GTT TTC AGT CAC A(nt 807 to 825); m3: 3' GGT TTC CAG CTC TTGGTT G (nt 947 to 965); m4: 3' ACG CTC TGC TTCATT AGT G (nt 743 to 761); and m5: 3' CGC AAGCAG GAT CTG AAC A (nt 746 to 764). The subtype-specific sense primers were, ml: 5' CAA CAT CACCGT CCT GGC A (nt 33 to 51); m2: 5' CCT GGCTCT TAC AAG TCC T (nt 33 to 51); m3: 5' CTT GCACAA TAA CAG TAC A (nt 6 to 24); m4: 5' CAA CTTCAC ACC TGT CAA T (nt 6 to 24); and m5: 5' TCTTAC CAC AAT GCA ACC A (nt 13 to 31). The ex-pected sizes of the PCR products were, 823 bp for ml,793 bp for m2, 960 bp for m3, 756 bp for m4 and752 bp for m5. Nucleotides are numbered from thebeginning of the coding sequence. The Genbank ac-cession numbers for the ml, m2, m3, m4, and m5receptor sequences are X15263, X15264, X15266,X15265, and M80333, respectively.
RESULTS
ImmunoprecipitationMuscarinic Receptor Expressing Chinese Hamster
Ovary Cell Lines. The specificity and sensitivity of eachantisera were tested on solubilized membranes fromCHO cell lines that stably express each of the fivemuscarinic receptors.'5'17 The ml, m2, m3, m4, andm5 receptor antisera bound 90%, 50%, 78%, 80%,and 78%, respectively, of the [3H] -QNB labeled mus-carinic receptors from the corresponding CHO celllines (Fig. 1). The ml, m2, and m4 antisera did notprecipitate muscarinic receptors from any of the othercell lines. The m3 antisera bound 2% of the musca-rinic receptors from the m l - and m2- expressingcells and the m5 antisera bound 2% of the receptorsfrom the m2 and m3 CHO cells.
To confirm the sensitivity of these antisera in a
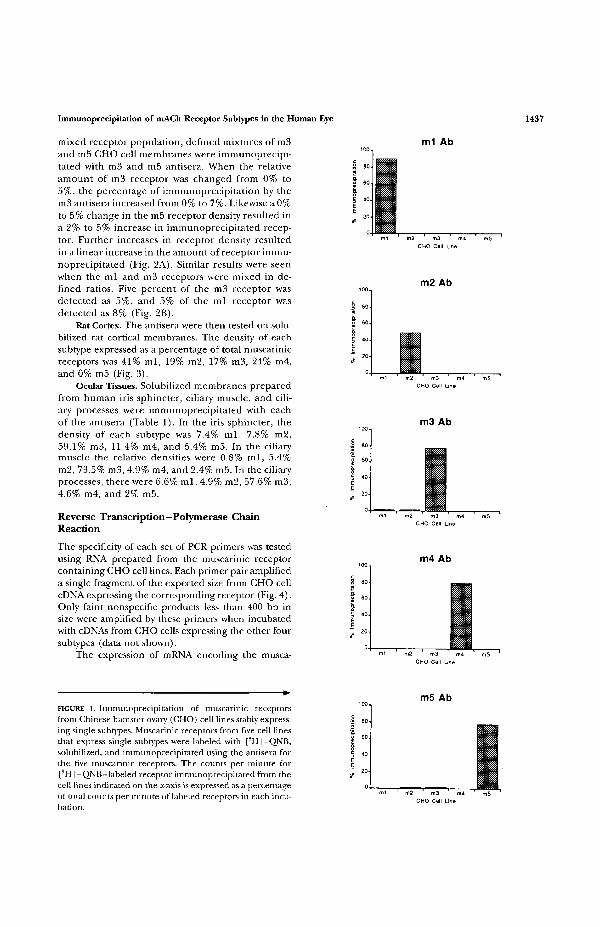
Immunoprecipitation of mACh Receptor Subtypes in the Human Eye 1437
mixed receptor population, defined mixtures of m3and m5 CHO cell membranes were immunoprecipi-tated with m3 and m5 antisera. When the relativeamount of m3 receptor was changed from 0% to5%, the percentage of immunoprecipitation by them3 antisera increased from 0% to 7%. Likewise a0%to 5% change in the m5 receptor density resulted ina 2% to 5% increase in immunoprecipitated recep-tor. Further increases in receptor density resultedin a linear increase in the amount of receptor immu-noprecipitated (Fig. 2A). Similar results were seenwhen the ml and m3 receptors were mixed in de-fined ratios. Five percent of the m3 receptor wasdetected as 5%, and 5% of the ml receptor wasdetected as 8% (Fig. 2B).
Rat Cortex. The antisera were then tested on solu-bilized rat cortical membranes. The density of eachsubtype expressed as a percentage of total muscarinicreceptors was 41% ml, 19% m2, 17% m3, 24% m4,andO% m5 (Fig. 3).
Ocular Tissues. Solubilized membranes preparedfrom human iris sphincter, ciliary muscle, and cili-ary processes were immunoprecipitated with eachof the antisera (Table 1). In the iris sphincter, thedensity of each subtype was 7.4% ml, 7.8% m2,59.1% m3, 11.4% m4, and 5.4% m5. In the ciliarymuscle the relative densities were 0.8% ml, 5.4%m2, 73.5% m3, 4.9% m4, and 2.4% m5. In the ciliaryprocesses, there were 6.6% ml, 4.9% m2, 57.6% m3,4.6% m4, and 2% m.5.
Reverse Transcription-Polymerase ChainReaction
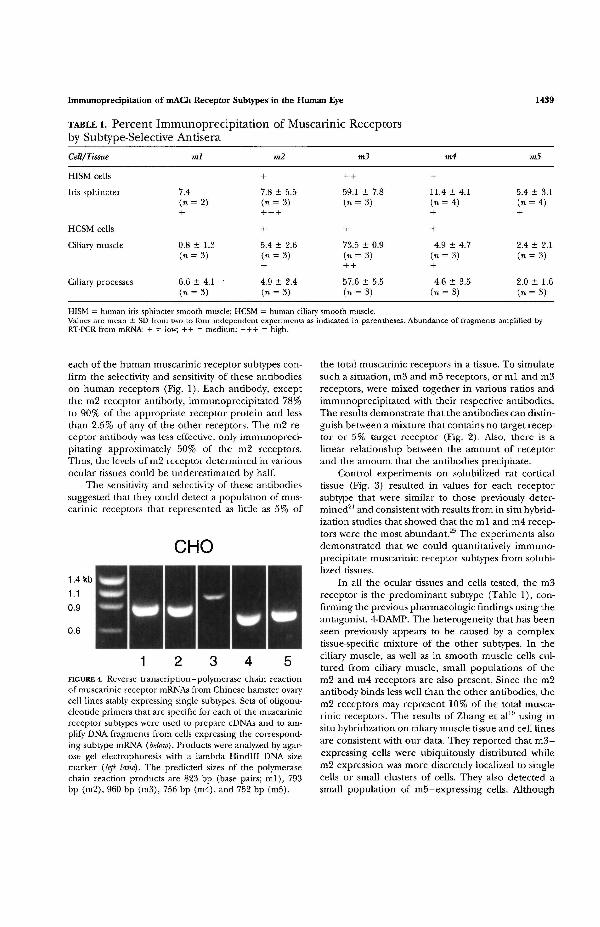
The specificity of each set of PCR primers was testedusing RNA prepared from the muscarinic receptorcontaining CHO cell lines. Each primer pair amplifieda single fragment of the expected size from CHO cellcDNA expressing the corresponding receptor (Fig. 4).Only faint nonspecific products less than 400 bp insize were amplified by these primers when incubatedwith cDNAs from CHO cells expressing the other foursubtypes (data not shown).
The expression of mRNA encoding the musca-
m1 Ab
% 4 0 .
S
o 60.
§ 40.
CHO Cell Line
m2 Ab
CHO Cell Line
m3 Ab
CHO Cell Line
m4 Ab
CHO Cell Line
FIGURE 1. Immunoprecipitation of muscarinic receptorsfrom Chinese hamster ovary (CHO) cell lines stably express-ing single subtypes. Muscarinic receptors from five cell linesthat express single subtypes were labeled with [SH]-QNB,solubilized, and immunoprecipitated using the antisera forthe five muscarinic receptors. The counts per minute for[3H] -QNB-labeled receptor immunoprecipitated from thecell lines indicated on the x-axis is expressed as a percentageof total counts per minute of labeled receptors in each incu-bation.
m5 Ab

1438 Investigative Ophthalmology & Visual Science, June 1997, Vol. 38, No. 7
rinic receptor subtypes in the human ocular tissueswas determined by RT-PCR (Fig. 5). In addition tothe tissues used for immunoprecipitation studies, cul-tures of smooth muscle cells isolated from the humaniris sphincter and human ciliary muscle were studied.In the iris sphincter sample (Fig. 5A), primers specificfor the ml, m2, m4 and m5 receptors amplified PCRproducts, with the m2 particularly abundant. In sam-ples from iris smooth muscle cells (Fig. 5B), only prod-ucts from reactions with m2, m3, and m4 primers wereapparent. Products of PCR were amplified by primersselective for the m2, m3, and m4 subtypes from bothciliary muscle tissue (Fig. 5C) and ciliary muscle cellsamples (Fig. 5D).
100
o 80
- 20 -
e 5ooo,
20 40 60 80% m3 Receptor
B100
•5 20 -
40 60 80% m3 Receptor
100
FIGURE 2. Immunoprecipitation of defined mixtures of Chi-nese hamster ovary cell membranes. Homogenized cellmembrane preparations of either m3 and m5 cells (A) orml and m3 cells (B) were mixed in various ratios based ontheir receptor densities. The mixtures were labeled, solubi-lized, and immunoprecipitated. The actual percentage ofprecipitation by the m3 ( • ) , m5 (•) and ml (A) antiserais plotted versus the relative m3 receptor density in the mix-ture. The data are the mean of at least two separate determi-nations.
Rat Cortex19% 17% 24% 0%
m5
FIGURE 3. Quantitative immunoprecipitation of muscarinicreceptor subtypes from rat cortex. Rat cortex homogenatewas labeled with [3H]-QNB, solubilized, and immunopre-cipitated using antisera selective for the subtypes indicatedon the x-axis. The counts per minute of [3H] -QNB-labeledreceptor immunoprecipitated by each antisera are indicatedby the solid bar and the counts per minute of [3H]-QNBlabeled receptor not recognized by each antisera, represent-ing the other four subtypes, are indicated by the hatchedpart of the bars. For each antisera, the total counts perminute were approximately 4250. The percentage of immu-noprecipitation calculated from the data is indicated abovethe bars. The data are representative of two separate experi-ments.
DISCUSSION
100 Data collected in previous pharmacologic and molecu-lar biologic studies on muscarinic cholinergic recep-tors in anterior ocular tissues have led to the conclu-sion that there is subtype heterogeneity. The m3 andm2 subtypes appear to be the most prevalent, butthere is also evidence for the expression of the otherthree subtypes (ml, m4, and m5). The lack of highlyselective pharmacologic agents and the qualitative na-ture of the studies on gene expression results in asomewhat confusing picture to date.
To determine which are the significant muscarinicreceptor subtypes in the iris and ciliary body that mayserve as targets of selective drug action, we have quan-tified receptor protein levels using subtype-selectiveantibodies.21'22 These antibodies were generatedagainst fusion proteins containing fragments of thethird intracellular loop regions of these receptors, ex-cept for the m3 antisera that is directed against a car-boxyl terminal synthetic peptide. These domains havethe least sequence similarity between the subtypes. Re-sults of previous studies have shown that these anti-bodies could quantitatively immunoprecipitate rat-brain muscarinic receptors that have been solubilizedand radiolabeled with the nonselective ligand, [3]H-QNB.21
Control experiments carried out on solubilizedmembranes from CHO cell lines that stably express
Immunoprecipitation of mACh Receptor Subtypes in the Human Eye 1439
TABLE l. Percent Immunoprecipitation of Muscarinic Receptorsby Subtype-Selective AntiseraCell/Tissue
HISM cells
Iris sphincter
HCSM cells
Ciliary muscle
Ciliary processes
7.4(n
0.8(n
6.6(n
ml
= 2)
± 1.3
± 4.1 •
= 3)
m2
+
7.8 ± 5.5( n = 3 )
+
5.4 ± 2.6( « = 3 )
4.9 ± 2.4
+ +
59.1 ± 7.8( n = 3 )
+
73.5 ± 0.9( n = 3 )
57.6 ± 5.5( n = 3 )
+
11.4 ± 4.1(n = 4)
+
4.9 ± 4.7( n = 3 )
4.6 ± 3.5( n = 3 )
m5
5.4 ± 3.1(n = 4)
2.4 ± 2.1( n = 3 )
2.0 ± 1.6( n = 3 )
HISM = human iris sphincter smooth muscle; HCSM = human ciliary smooth muscle.Values are mean ± SD from two to four independent experiments as indicated in parentheses. Abundance of fragments amplified byRT-PCR from mRNA: + = low; ++ = medium; + + + = high.
each of the human muscarinic receptor subtypes con-firm the selectivity and sensitivity of these antibodieson human receptors (Fig. 1). Each antibody, exceptthe m2 receptor antibody, immunoprecipitated 78%to 90% of the appropriate receptor protein and lessthan 2.5% of any of the other receptors. The m2 re-ceptor antibody was less effective, only immunopreci-pitating approximately 50% of the m2 receptors.Thus, the levels of m2 receptor determined in variousocular tissues could be underestimated by half.
The sensitivity and selectivity of these antibodiessuggested that they could detect a population of mus-carinic receptors that represented as little as 5% of
CHO
0.6
FIGURE 4. Reverse transcription-polymerase chain reactionof muscarinic receptor mRNAs from Chinese hamster ovarycell lines stably expressing single subtypes. Sets of oligonu-cleotide primers that are specific for each of the muscarinicreceptor subtypes were used to prepare cDNAs and to am-plify DNA fragments from cells expressing the correspond-ing subtype mRNA (below). Products were analyzed by agar-ose gel electrophoresis with a lambda Hindlll DNA sizemarker (left lane). The predicted sizes of the polymerasechain reaction products are 823 bp (base pairs; ml) , 793bp (m2), 960 bp (m3), 756 bp (m4), and 752 bp (m5).
the total muscarinic receptors in a tissue. To simulatesuch a situation, m3 and m5 receptors, or ml and m3receptors, were mixed together in various ratios andimmunoprecipitated with their respective antibodies.The results demonstrate that the antibodies can distin-guish between a mixture that contains no target recep-tor or 5% target receptor (Fig. 2). Also, there is alinear relationship between the amount of receptorand the amount that the antibodies precipitate.
Control experiments on solubilized rat corticaltissue (Fig. 3) resulted in values for each receptorsubtype diat were similar to those previously deter-mined21 and consistent widi results from in situ hybrid-ization studies that showed that the ml and m4 recep-tors were the most abundant.25 The experiments alsodemonstrated fhat we could quantitatively immuno-precipitate muscarinic receptor subtypes from solubi-lized tissues.
In all the ocular tissues and cells tested, the m3receptor is the predominant subtype (Table 1), con-firming the previous pharmacologic findings using theantagonist, 4-DAMP. The heterogeneity that has beenseen previously appears to be caused by a complextissue-specific mixture of the other subtypes. In theciliary muscle, as well as in smooth muscle cells cul-tured from ciliary muscle, small populations of them2 and m4 receptors are also present. Since the m2antibody binds less well than the other antibodies, them2 receptors may represent 10% of the total musca-rinic receptors. The results of Zhang et al19 using insitu hybridization on ciliary muscle tissue and cell linesare consistent with our data. They reported that m 3 -expressing cells were ubiquitously distributed whilem2 expression was more discretely localized to singlecells or small clusters of cells. They also detected asmall population of m5-expressing cells. Although

1440
A. Iris Sphincter
Investigative Ophthalmology & Visual Science, June 1997, Vol. 38, No. 7
B. HISM C. Ciliary Muscle D. HCSM
1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 2 3 4 5
FIGURE 5. Reverse transcription-polymerase chain reaction analysis of muscarinic receptorsubtype expression in human iris sphincter (A), human iris smooth muscle cells (B), humanciliary muscle (C), and human ciliary smooth muscle cells (D). Total RNAs isolated fromeach of the tissues and cells were aliquoted into five reactions using the primer sets specificfor the five muscarinic receptor subtypes (below each panel). Products of polymerase chainreaction were analyzed by agarose gel electrophoresis. The lambda Hindlll DNA size markeris shown in the right lane of panel D. The predicted sizes of the products are 823 bp (basepairs; ml), 793 bp (m2), 960 bp (m3), 756 bp (m4), and 752 bp (m5).
there was some receptor immunoprecipitated by them5 antisera in the current data, the value (2.4%) istoo small to be considered conclusive.
The ciliary processes express the ml, m2, and m4subtypes in addition to the m3 receptor. Both the mland m2 receptors have been detected previously insecond-messenger studies conducted on rabbit ciliaryprocesses,1113 which suggests that these subtype popu-lations, though small, are functional. Since ml mRNAhas not been detected by RT-PCR in cultured NPEcells,26 it is possible that ml receptors are expressedin the pigmented epithelium. Their distribution andtheir function in aqueous humor secretion remain tobe determined. The location and role of the m4 sub-type is also unknown. Transcripts encoding m4 werepreviously detected in bovine ciliary processes byNorthern blot analysis.20
All five subtypes were detected in the iris sphinc-ter. Because of the small size of this tissue, however,it is possible that contamination with receptors fromother cell types associated with the tissue is more sig-nificant. The ml and m5 receptor mRNAs were notdetected in the cultured smooth muscle cells from theiris sphincter (Fig. 5B), but this may be the resultof culturing the cells. Alternatively, the ml and m5receptors may only be expressed on a subset of irissphincter cells, as described for m2 and m5 receptorsin the ciliary muscle,19 or on an infiltrating or adjacentcell type—vascular cells or the iris epithelium, for ex-ample. The ml subtype was not detected in previousNorthern blot hybridizations on RNA from bovine irissphincter.20
The presence of the m5 receptor is a surprise. Ithas previously been seen in small amounts in a popula-tion of dopaminergic neurons in the substantia nigra25
and in the midbrain27 of rats. Outside the central ner-vous system, m5 mRNA has been detected in humanmacrophages28 and in the human ciliary muscle.19 Thepresence of m5 protein in a small number of humancells outside the central nervous system is demon-strated for the first time in these results.
The same subtypes were detected by RT-PCR and
immunoprecipitation with one surprising exception.Even though the m3 subtype is the predominant mus-carinic receptor at the protein level in iris sphincter,no m3 transcripts were detected (Fig. 5A). Controlexperiments on m3—expressing CHO cells (Fig. 4)indicate that this primer set is not as sensitive as theother primers, but it did detect m3 transcripts in theciliary muscle and iris sphincter cell lines (Fig. 5).Interestingly, in the Northern blot studies of Honka-nen et al,20 they also did not detect any m3 mRNA(m4 in their nomenclature)16 in the iris sphincter.Perhaps the level of m3 transcripts in the iris sphincteris low and not easily detected. One explanation for thisresult is the recent finding that m3 receptor protein isdegraded very slowly.29 There is no need, therefore,for cells to express much m3 mRNA. Such disparitiesbetween mRNA and protein levels demonstrate thevalue of measuring levels of receptor protein, as wehave done.
The coexistence of multiple subtypes, some thatactivate the same second messenger pathways, in suchapparently homogeneous tissues as smooth muscle ispuzzling. The current data and the in situ hybridiza-tion study on ciliary muscle of Zhang et al19 do notdetermine whether receptor subtypes coexist in a sin-gle cell. But the in situ results clearly demonstrate thatcells within the ciliary muscle have unique patterns ofmuscarinic receptor expression. This tissue and thecell lines derived from it should be useful models fordetermining the physiological significance of the vari-ous receptor subtypes. The longitudinal muscle of therat ileum has both the m2 and m3 receptor subtypes,and it has been suggested that inhibition of adenylatecyclase by the m2 subtype antagonizes /3-adrenergic-mediated relaxation, thereby enhancing m3-inducedmuscle contraction.30 It will be interesting to seewhether the m2-expressing cells in the ciliary musclealso express /3-adrenergic receptors.
In conclusion, the m3 muscarinic receptor is the pre-dominant receptor in the iris-ciliary body of the humaneye. The m2 and m4 subtypes are also present at lowerlevels in all the tissues. The ml and m5 subtypes appear

Immunoprecipitation of mACh Receptor Subtypes in the Human Eye 1441
to be expressed in discrete cell populations. The extensiveheterogeneity of muscarinic receptor subtypes makes itlikely that subtype-selective drugs will have different activi-ties than current muscarinic drugs. Whedier such drugswill have an improved efficacy and side-effect profile, how-ever, is difficult to predict.
Key Words
ciliary muscle, ciliary processes, immunoprecipitation, irissphincter, muscarinic receptor
Acknowledgments
The authors thank Kathy Crawford at the University of Wis-consin for her efforts in obtaining, dissecting, and freezinghuman eye tissue from the University of Wisconsin eye bank;Barry Wolfe and members of his laboratory at GeorgetownUniversity for providing the antibodies and helpful advicein setting up the immunoprecipitation studies; Barbara Feld-mann at Allergan for her assistance in dissecting tissues andculturing cells; and Bernadette Cumming for her secretarialassistance.
References
1. Hammer R, Berrie CP, Birdsall NJM, Burgen ASV,Hulme EC. Pirenzepine distinguishes between differ-ent subclasses of muscarinic receptors. Nature. 1980;283:90-92.
2. Hammer R, Giachetti A. Muscarinic receptor sub-types: M, and M2 biochemical and functional charac-terization. LifeSci. 1983; 31:2991-2998.
3. Gil DW, Wolfe BB. Pirenzepine distinguishes betweenmuscarinic receptor-mediated phosphoinositidebreakdown and inhibition of adenylate cyclase. ]Phar-macol Exp Ther. 1985;232:608-616.
4. Barany E, Berrie CP, Birdsall NJM, Burgen ASV,Hulme EC. The binding properties of the muscarinicreceptors of the cynomolgus monkey ciliary body andthe response to the induction of agonist subsensitivity.Br JPharmacol. 1982; 77:731-739.
5. WoldeMussie E, Feldmann BJ, ChenJ. Characteriza-tion of muscarinic receptors in cultured human irissphincter and ciliary smooth muscle cells. Exp Eye Res.1993; 56:385-392.
6. Wax MB, Coca-Prados M. Receptor-mediated phos-phoinositide hydrolysis in human ocular ciliary epithe-lial cells. Invest Ophthalmol Vis Sci. 1989;30:1675-1679.
7. Crook RB, PolanskyJR. Neurotransmitters and neuro-peptides stimulate inositol phosphates and intracellu-lar calcium in cultured human nonpigmented ciliaryepithelium. Invest Ophthalmol Vis Sci. 1992;33:1706-1716.
8. Akhtar RA, Honkanen RE, Howe PH, Abdel-Latif AA.M2 muscarinic receptor subtype is associated with ino-sitol triphosphate accumulation, myosin light chainphosphotylation and contraction in sphincter smoothmuscle of rabbit iris. / Pharmacol Exp Ther. 1987;243:624.
9. Honkanen RE, Abdel-Latif AA. Characterization of
cholinergic muscarinic receptors in the rabbit iris. Bio-chem Pharmacol. 1988;37:2575-2583.
10. Bognar IT, Baumann B, Dammann F, et al. M2 musca-rinic receptors on the iris sphincter muscle differ fromthose on iris noradrenergic nerves. Eur j Pharmacol.1989; 163:263-274.
11. Tobin AB, Osborne NN. Evidence for the presence ofcholinergic muscarinic receptors negatively linked toadenylate cyclase in the iris-ciliary body. NeurochemInt. 1988; 13:517-523.
12. Jumblatt JE, North GT, Hackmiller RC. Muscariniccholinergic inhibition of adenylate cyclase in the rab-bit iris-ciliary body and ciliary epithelium. Invest Oph-thalmol Vis Sci. 1990;31:1103-1108.
13. Mallorga P, Babilon RW, Buisson S, Sugrue MF. Mus-carinic receptors of the albino rabbit ciliary process.Exp Eye Res. 1989;48:509-522.
14. Bognar IT, Altes U, Beinhauer C, Kessler I, Fuder H.A muscarinic receptor different from the Mi, M2, M5,and M4 subtypes mediates the contraction of the rab-bit iris sphincter. Naunyn-Schmiedeberg's Arch Pharmacol.1992;345:611-618.
15. Bonner TI, Buckley NJ, Young AC, Brann MR. Identi-fication of a family of muscarinic acetylcholine recep-tor genes. Science. 1987; 237:527-532.
16. Peralta EG, Ashkenazi A, Winslow JW, Smith DH, Ra-machandranj, Capon DJ. Distinct primary structures,ligand-binding properties and tissue-specific expres-sion of four human muscarinic acetylcholine recep-tors. EMBOJ. 1987; 6:3923-3929.
17. Bonner TI, Young AC, Brann MR, Buckley NJ. Clon-ing and expression of the human and rat m5 musca-rinic acetylcholine receptor genes. Neuron. 1988;1:403-410.
18. Bogardus AM, Feldmann BJ, WoldeMussie E, Gil DW.Muscarinic receptor subtypes in human eye (abstract).SocNeurosd. 1991; 17:587.
19. Zhang X, Hernandez MR, Yang H, Erickson K. Expres-sion of muscarinic receptor subtype mRNA in the hu-man ciliary muscle. Invest Ophthalmol Vis Sci. 1995;36:1645-1657.
20. Honkanen RE, Howard EF, Abdel-Latif AA. M3-mus-carinic receptor subtype predominates in the bovineiris sphincter smooth muscle and ciliary processes. In-vest Ophthalmol Vis Sci. 1990;31:590-593.
21. Wall SJ, Yasuda RP, Li M, Ciesla W, Wolfe BB. Differ-ential regulation of subtypes m l - m 5 of muscarinicreceptors in forebrain by chronic atropine administra-tion. J Pharmacol Exp Ther. 1992; 262:584-588.
22. Wall SJ, Yasuda RP, Hory F, et al. Production of anti-sera selective for ml muscarinic receptors using fusionproteins: Distribution of ml receptors in rat brain.Mol Pharmacol. 1991; 39:643-649.
23. Shuldiner AR, Tanner K, Moore CA, Roth J. RNAtemplate-specific PCR: An improved method that dra-matically reduces false positives in RT-PCR. Biotech-niques. 1991; 11:760-763.
24. Chomczynski P, Sacchi N. Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chlorofonn extraction. Analyl Biochem. 1987; 162:156-159.

1442 Investigative Ophthalmology & Visual Science, June 1997, Vol. 38, No. 7
25. Weiner DM, Levey AI, Brann MR. Expression of mus-carinic acetylcholine and dopamine receptor mRNAsin rat basal ganglia. Proc Natl Acad Sci USA. 1990;87:7050-7054.
26. Gil DW, Bogardus AM, WoldeMussie E, Coca-Prados M. Identification of muscarinic receptorsubtypes in human eye by polymerase chain reac-tion. ARVO Abstracts. Invest Ophthalmol Vis Sci.1991;32:256.
27. Wolfe BB, Yasuda RP, Wall SJ, Li M, Ciesla W. Develop-ment and utilization of a panel of antisera selective
for each of the subtypes of muscarinic cholinergicreceptor. Life Sci. 1993; 52:584. Abstract.
28. Ferrari-Dileo G, Flynn DD. Characterization of mus-carinic receptors on cultured microglial cells. Life Sci.1995;56:1037. Abstract.
29. Fraser CM, Lee NH. Regulation of muscarinic recep-tor expression by changes in mRNA stability. Life Sci.1995; 56:899-906.
30. Thomas EA, Baker SA, Ehlert FJ. Selective blockadeof M3 receptor mediated responses by 4-DAMP mus-tard. Life Sci. 1993; 52:573. Abstract.




















