Activation of the Ah Receptor Signaling Pathway by Prostaglandins

Prostaglandins Alter Extracellular Matrix Adjacent toHuman Ciliary Muscle Cells In Vitro
James D. Lindsey, Kenji Kashiwagi, Fumiko Kashiwagi, and Robert N. Weinreb
Purpose. This study investigates the possibility that prostaglandins (PGs) induce changes inextracellular matrix (ECM) adjacent to ciliary muscle cells.
Methods. Human ciliary smooth muscle cells were grown to confluence in monolayer culturesand were treated with PGF2a, 11-deoxy-PGEi, or PhXA85 (the nonesterified analogue ofPhXA41) for 12 to 72 hours. The amount of collagens type I, III, and IV in the cultures wasdetermined, using sandwich enzyme immunoassays. The distributions of these collagens wereassessed in the PG-treated cultures by immunocytochemistry.
Results. Twenty-four-hour treatment with 20 nM PGF2a, 11-deoxy-PGE], or PhXA85 reducedthe amount of collagen type I in extracts of the cell layer by 65 ± 10%, 56 ± 7%, and 46 ±7%, respectively, when compared with levels of those substances in vehicle-treated cultures.In similar fashion, collagen type III in cell layer extracts was reduced by 41 ± 5%, 33 ± 9%,and 3 ± 5%, respectively. When the concentration of PGs was increased to 200 nM, theamount of type III collagen in the cell layer extracts was reduced by 93 ± 7%, 99 ± 1%, and99 ± 1%, respectively. Changes in type IV collagen in cell layer extracts after treatment with20 nM PGs were not statistically significant. When the concentration of PGF2a, 11-deoxy-PGEi,or PhXA85 was increased to 200 nM, the amount of collagen type IV in the cell layer extractincreased by 101 ± 16%, 14 ± 5%, and 89 ± 11%, respectively. There were minimal changesin the staining pattern for collagen type I after 24-hour treatment with 20 nM PGs. When thePG concentration was increased to 200 nM, there were reductions in the density of collagentype I fibrils and clumping of collagen type III immunoreactive elements. The delicate lace-work of collagen type IV immunoreactivity was replaced by bundles or clumps of heavyimmunoreactive strands, separated by areas without immunoreactivity. These changes werepresent in cultures exposed to 20 nM PGs and were marked when PG concentration wasincreased to 200 nM.
Conclusion. These results indicate that PGs can induce substantial changes in the ECM aroundciliary smooth muscle cells in vitro. These data support the possibility that changes in ciliarymuscle ECM may contribute to increased uveoscleral outflow facility after topical PG adminis-tration. Invest Ophthalmol Vis Sci. 1997; 38:2214-2223.
A opical administration of certain prostaglandins(PGs) can reduce intraocular pressure (IOP) inhealthy and glaucomatous human eyes.1"4 Althoughthis effect is associated with increased uveoscleral out-flow, the precise mechanism is unknown.5"8 Reduc-
From the Glaucoma Center and Research Laboratories, University of California SanDiego, Lajolla.Presented in part at the annual meeting of the Association for Research in Visionand Ophthalmology, Ft. Lauderdale, Florida, May, 1995.Supported in part by grants from the National Eye Institute (EY 05990; RNW) andthe San Diego Eye Bank (RNW).Submitted for publication January 15, 1997; revised June 2, 1997; accepted June 2,1997.Profnietary interest category: N.Reprint requests: Robert N. Weinreb, Glaucoma Center and Research Laboratories,Shiley Eye Center, UCSD/0946, 9500 Gilman Drive, Lajolla, CA 92093-0946.
tion in the amount of extracellular matrix (ECM)among the fiber bundles of monkey ciliary muscleafter PG treatment has been reported.910 Because re-sults of tracer studies indicate that uveoscleral outflowpasses through the extracellular spaces between ciliarymuscle fibers,11'12 changes in ECM at this site mayreduce hydraulic resistance around the fibers, facilitat-ing aqueous flow through the muscle.
Matrix metalloproteinases (MMPs) are a family ofenzymes with differing specificities for peptide se-quences found in such ECM components as collagen,laminin, and fibronectin.1314 These proteases are se-creted as inactive proenzymes and converted to activeenzymes extracellularly.15"17 Recently, increased con-
2214Investigative Ophthalmology & Visual Science, October 1997, Vol. 38, No. 11Copyright © Association for Research in Vision and Ophthalmology

Prostaglandins Alter Ciliary Muscle Extracellular Matrix 2215
centrations of pro-MMP-1 and pro-MMP-3 were foundin the medium of cultured human ciliary smooth mus-cle cells treated with PGs.18 MMP-1 can degrade fibril-lar collagens, including collagen type I and collagentype III; MMP-3 can degrade collagen type IV andcollagen type III. In previous studies of human ciliarymuscle ECM, collagen type I and collagen type IIIhave been identified in the stromal matrix among themuscle bundles.19"21 Collagen type IV is found in thebasement membranes, immediately adjacent to theciliary muscle fibers.19'21 Thus, changes in ciliary mus-cle ECM after PG treatment may reflect direct alter-ations of collagens type I, III, and IV.
In the current study, this possibility was investi-gated by studying the ECM of cultured ciliary smoothmuscle cells. After exposure to PG treatment, theamount of collagen type I, III, and IV in the cultureswas evaluated using enzyme-linked immunosorbentassays (ELISAs). In addition, changes in the structuralorganization of these collagens were assessed by im-munocytochemical analysis.
MATERIALS AND METHODS
Human Ciliary Smooth Muscle Cultures
Eighteen pairs of human eyes from donors 31 to 76years old were obtained from the San Diego Eye Bankwithin 24 hours after death. Enucleation was com-pleted within 6 hours after death, and eyes were storedat 4°C for less than 24 hours before cultures weregenerated. Donors had no known history of glaucomaor other eye diseases.
Ciliary smooth muscle cell monolayer cultureswere prepared as previously described.22 Briefly, eacheye was bisected horizontally 3 mm posterior to thelimbus. The anterior segment was placed cornea-sidedown under a dissecting microscope, and the lens wasremoved by cutting the zonule. Strips of the outermostportion of ciliary muscle were explanted into 35-mmculture dishes (Falcon, Lincoln Park, NJ) in Dulbec-co's modified Eagle's medium and Ham's F12 nutri-ent mixture (DMEM-F12, University California, SanDiego Cell Culture Facility) supplemented with 10%fetal bovine serum (Gemini, Woodland, CA) and 1ng/ml recombinant human basic fibroblast growthfactor (bFGF; R & D Systems, Minneapolis, MN). Thecultures were incubated in a humidified atmosphereof 95% air-5% CO2. The medium was changed every3 to 4 days. Primary cells reached confluence within40 to 60 days, and subsequent passages reached con-fluence within 14 days. Confluent cultures were tryp-sinized and subcultured in the same medium at a ratioof 1:3. For the current study, we used confluent third-and fourth-passage cells. The cells were grown in 35-mm culture dishes for the ELISA studies or on acid-
cleaned glass coverslips for the immunocytochemicalstudies.
These cells express several proteins specific to cili-ary smooth muscle cells in vivo, including a-smoothmuscle actin, desmin, and calponin, an actin-bindingprotein that regulates contraction in smooth mus-cles.19'23 Also, these cells biosynthesize collagens type I,III, and IV, which assemble into a characteristic matrixadjacent to the cell monolayer.19'20
Prostaglandins
Prostaglandin ¥2a (PGF2a), 17PT-PGF2a, and 11-deoxy-PGEt were purchased from Cayman (Ann Arbor, MI),and PhXA85 (the nonesterified analogue of PhXA41)was a generous gift of J. Stjernschantz, PhD, of Phar-macia Upjohn (Uppsala, Sweden). Topically appliedPG esters (latanoprost) become hydrolyzed as theypass through the cornea and into the aqueous hu-mor.24 Thus, PhXA85, the nonesterified analogue oflatanoprost, is the appropriate choice for studies oflatanoprost action in vitro. All PGs were dissolved inethanol and diluted with DMEM-F12 to 2 mM. Thefinal concentration of ethanol in the vehicle controlwas 0.001%, the same concentration of ethanol in thehighest dose of PG treatment used in this study. Allreagents were stored at — 70°C, to minimize oxidationuntil use.
Experimental Design
Cells were maintained in a medium containing 10%serum in DMEM-F12 supplemented with 1 ng/mlbFGF for 1 week after they had become confluent. Thecultures then were switched to serum-free mediumcontaining DMEM-F12 supplemented with 1 mM in-sulin, 5 mg/ml transferrin, 100 mM ascorbic acid, 1mM /3-aminopropionitrile, and 1 ng/ml bFGF for 4days before experimental treatments were initiated.Serum-free medium was used to minimize nonspecificinduction of MMPs, because serum contains agentsknown to stimulate MMP biosynthesis.17 In the treat-ment phase, PGs or vehicle was added daily to thecultures for 1 to 3 days. Previously, 78 nM PGF2£V (28ng/ml) has been measured in aqueous humor col-lected from monkey eyes 4 hours after topical treat-ment with an IOP-lowering dose of PGF2a.
4 Also, thepeak concentration of latanoprost measured in hu-man aqueous humor samples obtained after standardtopical treatment was approximately 100 nM (33 to34 ng/ml; J. Stjernschantz, personal communication,1997). Thus, to model the range of PGs likely to beseen by ciliary smooth muscle cells in vivo after IOP-lowering topical doses, the cultures were exposed to20 or 200 nM PGs.
In vivo, molecules that can facilitate proMMP acti-vation and MMP deactivation have been detected inhuman aqueous humor.25'26 Although it is presently

2216 Investigative Ophthalmology & Visual Science, October 1997, Vol. 38, No. 11
unknown how these molecules are themselves regu-lated in the presence of PGs, their presence raises thepossibility that secreted proMMPs are converted toactive MMPs. Thus, to model the presence of proMMPactivators in aqueous humor, 100 ng/ml plasmin wasadded daily to serum-free medium along with PGs orvehicle, to convert proMMPs to active MMPs.
Immunocytochemical Analysis
Antibodies used for immunocytochemical studieswere affinity-purified rabbit polyclonal antihuman col-lagen type I (Biodesign, Kennebunkport, ME; diluted1:5), mouse monoclonal antihuman collagen type III(Biogenex, San Ramon, CA; diluted 1:15), mousemonoclonal antihuman collagen type IV (COL4;Sigma, St. Louis, MO; diluted 1:20), mouse mono-clonal antihuman calponin (hCP; Sigma; diluted1:1000), and mouse monoclonal antihuman a-smoothmuscle actin (1A4; Sigma; diluted 1:100). The speci-ficities of these antibodies have been well-character-ized.27"34 Rhodamine-conjugated goat antirabbit im-munoglobulin (Ig) G (Sigma; diluted 1:400) was usedas secondary antibody for collagen type I. Rhodamine-conjugated goatantimouse IgG (Sigma; diluted 1:400)was used demonstrate the presence of the monoclonalantibodies. In each case, the secondary antibody waspreabsorbed with homogenized human uvea to re-duce nonspecific binding.
After incubation with the experimental PGs for24 or 72 hours, the cultures were rinsed three timeswith phosphate-buffered saline (PBS), fixed with 4%formaldehyde in PBS, washed with PBS, permeatedwith 0.1% Triton X-100 in PBS for 15 minutes, washedwith PBS, blocked with 10% goat serum in PBS, andwashed again with PBS. The cells were next incubatedwith primary antibodies for 60 minutes in a moistchamber, washed with PBS, exposed to secondary anti-bodies for 30 minutes in the dark, washed with PBS,and then mounted with gelvatol. Controls for nonspe-cific secondary antibody binding were incubated withPBS instead of primary antibodies.
Enzyme Immunosorbent Assays
After administration of PGs or vehicle, media was col-lected for ELISA analysis and stored frozen at — 80°Cuntil analyzed. To isolate collagens from the cell layer,the cells were rinsed in PBS and extracted with 2 Mguanidine-HCl, 20 rnM dithiothreitol, and 0.05 MTris-HCl, (pH 7.4) for 24 hours. The extracts werecentrifuged at 10,000 rpm for 30 minutes, and thesupernatants were collected and dialyzed against PBSand 0.05% Triton X-100 for 4 hours. These sampleswere kept 1 to 3 days at 4°C until ELISA.
The concentration of collagens type I and III inthe samples was measured using sandwich EIAs. Wellsin a polystyrene microtiter plate (Falcon; Becton Dick-
inson, Lincoln Park, NJ) were coated with donkey anti-mouse IgG (10 /ig/ml, 100 //I/well, carbonate buffer,pH 8; preabsorbed to minimize cross-reactivity to non-mouse serum proteins; Jackson ImmunoResearch,Bethesda, MD). For collagen type I assays, the plateswere rinsed and coated with monoclonal mouse anti-collagen type I antibody (2 /ig/ml, 50 /zl/well in PBSand 0.05% Tween 20; clone COL1; Sigma). The wellswere then washed, blocked with fresh nonfat milk (di-luted 1:10 in PBS), incubated with the samples orstandards (purified human collagen type I; Gibco,Grand Island, NY), washed, incubated with polyclonalrabbit anticollagen type I (Chemicon, Temecula, CA;diluted 1:100), washed, incubated with goat antirabbitIgG conjugated to horseradish peroxidase; diluted1:200), washed, and developed with 50 fi\ tetramethylbenzidine (TMB Liquid Substrate System, Sigma) for1 to 2 minutes. Color development was terminated byaddition of an equal volume of 4% sulfuric acid. Thestandard curves were linear to less than 10 ng collagentype I per well (linear regression, P > 0.97).
For collagen type III assays, the procedure was thesame, except the second antibody was mouse mono-clonal antihuman type III antibody (2 //g/ml, 50 (j,\/well in PBS-Tween 20; Chemicon) and the third anti-body was rabbit polyclonal antihuman type III antihu-man collagen type III antibody). For the standards,purified human type III collagen was obtained fromGibco. The standard curves for this assay also werelinear to less than 10 ng collagen type III per well(linear regression, P > 0.97).
Collagen type IV was measured, using a sandwichELISA with two monoclonal antibodies (Fuji Chemi-cal, Takaoka, Japan). The first antibody recognizingthe 7S domain was linked to horseradish peroxidase.The other antibody recognizing the central triple-heli-cal domain was bound to a polystyrene bead.35 Briefly,50 jA from each extract sample was incubated withthe enzyme-linked primary antibody and antibody-attached bead for 1 hour. The bead was then washedfour times with PBS and incubated for 30 minutes withtetramethylbenzidine and 0.015% hydrogen peroxidein 10 mM sodium acetate buffer (pH 5.5). 1.33 Nsulfuric acid was added to stop the reaction, and absor-bency was measured at 450 nm. For the media sam-ples, the sample size was increased to 400 ml, and theprimary incubation period was extended to two hours.Otherwise, procedures for the cell extract and mediaEIAs were the same. Standard curves were generatedfor the cell extract and media procedures.
Cell Viability After Prolonged Exposure toProstaglandins
To evaluate cell viability, cells were incubated with amixture of acetoxymethyl ester calcein (calcein AM)and ethidium homodimer (EthD-1; LIVE/DEAD

Prostaglandins Alter Ciliary Muscle Extracellular Matrix 2217
FIGURE l. Phase-contrast view of cultured ciliary muscle cellsforming confluent monolayers of spindle-shaped cells. Mag-nification, X130.
assay, Molecular Probes, Eugene, OR). Live cells,which accumulate calcein AM, are distinguished by anintense, uniform green fluorescence with fluoresceinoptics. If the cell membrane is damaged, calcein AMdoes not accumulate and EthD-1 enters the cells andbinds to the nuclear DNA. With rhodamine optics,bound EthD-1 produces a bright red fluorescence.Confluent cells grown on glass coverslips were ex-posed to 10, 60, or 200 nM PGF2o for 3 days. Positive(live) controls were cells not exposed to any treat-ment, and negative (dead) controls were culturestreated with 70% methanol for 30 minutes before eval-uation. After washing with sterile Dulbecco's phos-phate-buffered saline, the cells were incubated with0.3 mM calcein AM and 0.3 mM EthD-1 in DMEM for30 minutes. The cells were mounted on microscopeslides and immediately observed with the fluorescencemicroscope.
Statistical Analysis
Reported experiments were performed at least twice,with similar results. The data were analyzed by one-way analysis of variance and the Student-Newman-Keuls test. A P value of less than 0.05 was consideredstatistically significant.
RESULTS
Cell Culture
Cultured ciliary muscle cells formed confluent mono-layers of spindle-shaped cells (Fig. 1). These cells orga-nized into a characteristic "hill and valley" distribu-tion (i.e., areas of higher and lower cell density). Im-munocytochemical evaluation with antibodies tocalponin and a-smooth muscle actin revealed positivestaining in more than 95% and 90% of the cells, re-spectively. Prostaglandin treatment did not changethe cells' appearance when viewed under phase optics.
Collagen Type I
Enzyme immunoassay revealed that the cell layer ex-tracts of cultures treated for 24 hours with vehiclecontrol medium contained 338 ± 41 ng per dish ofcollagen type I (mean ± SE, n = 4). In parallel cul-tures treated for 24 hours with 20 nM PGF2n:, collagentype I in the cell layer extracts was reduced by 65 ±10% (Fig. 2). Similarly, in cultures treated with 20 nM11-deoxy-PGE, or PhXA85, collagen type I in the celllayer extracts was reduced by 56 ± 7% and 46 ± 7%,respectively. All of these changes were statistically sig-nificant (P< 0.05).
Results of immunocytochemical study showed thatcollagen type I in the cultures treated with vehicle for24 or 72 hours was contained in parallel fibril arrays,over and between the smooth muscle cells that werelargely aligned with the long axis of the cells (Fig. 3A).In addition, there was light cytoplasmic and nuclearstaining. Immunostaining of the cultures treated for24 hours with 10 nM PGF2a revealed stained parallelfibrils; however, the density of the fibrils and the inten-sity of the staining were reduced. This effect was moreuniformly observed throughout the cultures when thePGF2cr concentration was increased to 200 nM and thetreatment duration was extended to 72 hours (Fig,3B). Similar reductions of fibril density and stainingintensity were seen in cultures treated with 11-deoxy-PGE! or PhXA85 (Figs. 3C, 3D).
Collagen Type HI
The amount of collagen type III in the cell layer ex-tracts of cultures treated with vehicle control for 24hours was 228 ± 1 ng/dish (n = 4). Treatment ofparallel cultures with 20 nM PGF2ff, 11-deoxy-PGE|, or
400 i Collagen Type I20 nM PGs
Vehicle PGFact 11doPGEi PhXA85
FIGURE 2. Content of collagen type I in extracts of humanciliary muscle cell cultures after treatment with 20 nM pros-taglandins. n = 4 cultures per data point. Abbreviations arethe same as in the text.

2218 Investigative Ophthalmology & Visual Science, October 1997, Vol. 38, No. 11
FIGURE 3. Cultured ciliary muscle cultures treated for 3 dayswith vehicle or 200 nM prostaglandins and immunostainedfor collagen type I. (A) vehicle control, (B) PGF2a, (C) 11-deoxy-PGE,, and (D) PhXA85. Magnification, X400.
PhXA85 reduced the amount of collagen type III inthe cell layer extract by 41 ± 5%, 33 ± 9%, and 3 ±5%, respectively (Fig. 4A). When the treatment condi-tions were changed to 200 nM PGs for 24 hours, theamount of collagen type III in the cell layer extractsexposed to PGF2a, 11-deoxy-PGE!, or PhXA85 was re-duced by93 ± 7%, 99 ± 1%, and 99 ± 1%, respectively(n = 3, Fig. 4B). The amount of collagen type III inthe cell layer extracts from the vehicle-treated culturesin this experiment was 249 ± 37 ng/dish.
The appearance of collagen type III immunoreac-tivity in vehicle-treated control cultures included occa-sional stained fibrils above and between the cells,punctate clusters of immunoreactivity on the surfaceof the cells, and amorphous immunoreactivity distrib-uted above and between the cells (Fig. 5A). After treat-ment with 10 nM PGF2tl., there were occasional areaswhere the amount of amorphous and punctate immu-noreactivity were reduced. Also the fibrils were some-times spanned by condensed-appearing strands of im-munoreactivity. These changes were more pro-nounced and more uniformly seen in cultures treatedwith 200 nM PGF2a for 72 hours (Fig. 5B). Similarreductions in amorphous and punctate staining ac-companied by condensations were seen in culturestreated with 11-deoxy-PGE! or PhXA85 (Fig. 5C, 5D).
Collagen Type IV
The amount of collagen type IV in cell layer extractsfrom vehicle-treated control cultures was much lessthan extract of collagens type I and III. The meanamount of collagen type TV observed in vehicle-treatedcultures typically ranged from 20 to 40 ng/dish. Forthese experiments, l7-phenyltrinor-PGF2Q was addedto die list of PGs tested. In cultures treated for 24hours with 20 nM PGs, significant changes from vehi-cle controls were not observed with any of the PGs.
However, when the concentration was increased to200 nM, each of the test PGs increased the amountof collagen type IV in the cell layer extracts (Fig. 6A).Specifically, treatment with PGF2a, 17-phenyltrinor-PGF2fv, 11-deoxy-PGEi, or PhXA85 induced increasesof 101 ± 1.6%, 108 ± 17%, 14 ± 5%, and 89 ± 11%,respectively. Increases of collagen type IV in the cul-ture medium were two to four times more than wasseen in the cell layer extracts (Fig. 6B). To comparethe distribution of collagen type IV in these experi-ments, the ratio of collagen type IV in the cell layerextracts to that in the medium was calculated. Foreach of die PGs tested, this ratio decreased substan-tially (Fig. 6C).
Results of immunocytochemical analysis revealedthat collagen type IV in vehicle-treated cultures was dis-
Collagen Type III20 nM PGs
3 0 0
Vehicle PGF2a 11doPGEi PhXA85
200 nM PGs
a> 200
1 0 0 •
Vehicle PGF2a UdoPGEi PhXA85
BFIGURE 4. Content of collagen type 111 in extracts of humanciliary muscle cell cultures after treatment with prostaglan-dins. (A) Treated with 20 nM PGs; n = 4. (B) Treated with200 nM PGs; n = 3.

Prostaglandins Alter Ciliary Muscle Extracellular Matrix 2219
FIGURE 5. Cultured ciliary muscle cultures treated for 3 dayswith vehicle or 200 nM prostaglandins and immunostainedfor collagen type III. (A) vehicle control, (B) PGF2ff, (C) 11-deoxy-PGE,, and (D) PhXA85. Magnification, X400.
tributed in a delicate, even lacework between and abovethe ciliary smooth muscle cells (Fig. 7A). Structuralchanges were moderate in cultures treated with 10 nMPGs and marked when the test concentration was in-creased to 200 nM. Specifically, treatment with PGF2a
induced the condensation of collagen type IV into heavybranched beams separated by expanses with minimalimmunoreactive material (Fig. 7B). Similar changeswere observed in cultures treated with 11-deoxy-PGE] orPhXA85 (Figs. 7C, 7D). For each of die above experi-ments, similar responses were observed in repeated ex-periments using cells from different donors.
Cell Viability After Prolonged Exposure toProstaglandinsIn view of the dramatic ECM changes observed with200 nM PG treatment, the possibility that cellular tox-icity may be involved was evaluated. Treatment with200 nM PGs for 3 days did not change the appearanceof the cells observed under phase optics. To confirmwhether prolonged exposure to 200 nM PGs inducedany cellular toxicity, treated cultures were exposed to amixture of calcein AM and EthD-1. Untreated controlcells all were stained uniformly green (Fig. 8A). Allcells treated for 3 days with 10, 60, or 200 nM PGF2a
were stained uniformly green by calcein AM, and werenot stained by EthD-1 (Fig. 8B). Identical results wereobserved with l7-phenyltrinor-PGF2ff, 11-deoxy-PGEi,and PhXA85. As a positive control, cultures weretreated with methanol to kill the cells and were ex-posed to the mixture of calcein AM and EthD-1. Inthese cultures, nuclei of all cells fluoresced bright redunder rhodamine optics, and no cell contained greencytoplasmic staining under fluorescein optics (Fig.8C). No nuclear staining was observed in the PG-treated cultures under rhodamine optics (Fig. 8D).These results indicate that there was no cytotoxicity
associated with extended exposure to PGs in thisstudy.
DISCUSSIONProstaglandin F2a, 11-deoxy-PGEi, and PhXA85 eachinduce changes in amount and distribution of colla-
Collagen Type IVCell Layer, 200 nM
200
1 5 0 •
Vehicle PGF2a 17ptPGF2cc UdoPGEi PhXA85
Medium
o 100
B
1.2 -
Vehicle PGF2oc 17ptPGF2<x UdoPGEi PhXA85
Cell Layer/Medium
Vehicle PGFaa 17ptPGF2ct UdoPGEi PhXA85
FIGURE 6. Content of collagen type IV in extracts of humanciliary muscle cell cultures after treatment with 200 nM pros-taglandins. (A) Total amount in cell layer, (B) total amountin medium, (C) ratio of amount in cell layer to amount inmedium, n — 5.
2220 Investigative Ophthalmology & Visual Science, October 1997, Vol. 38, No. 11
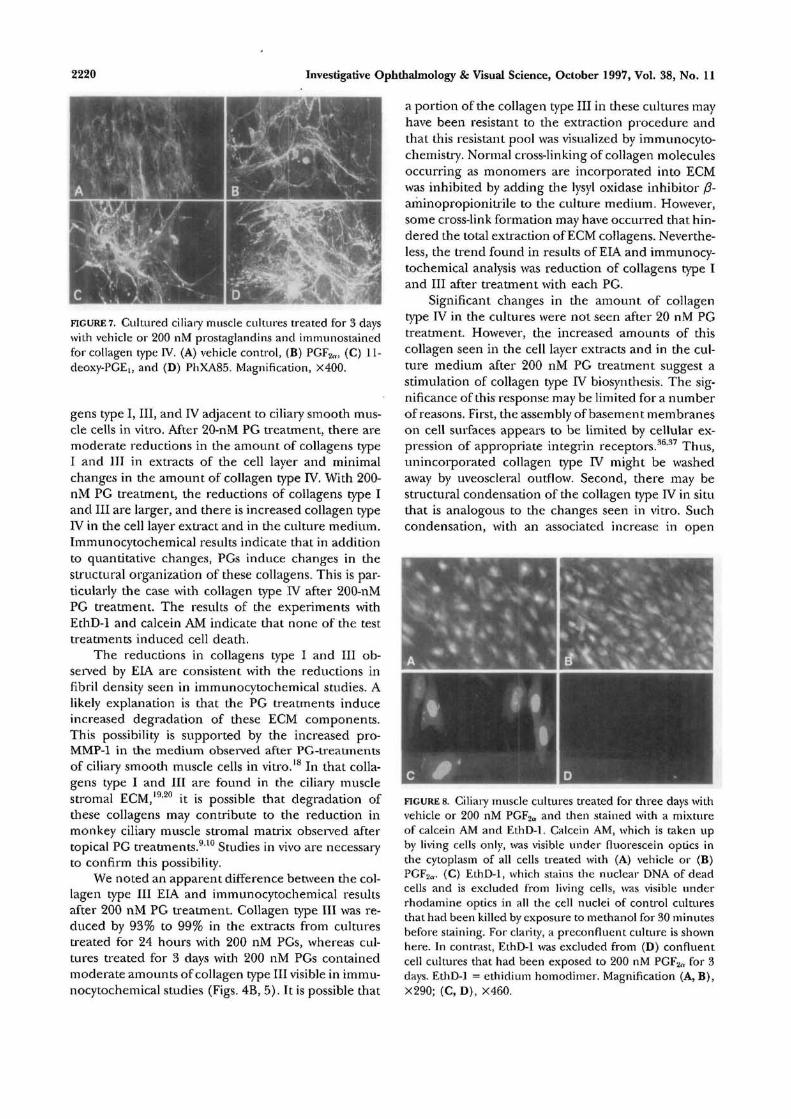
FIGURE 7. Cultured ciliary muscle cultures treated for 3 dayswith vehicle or 200 nM prostaglandins and immunostainedfor collagen type IV. (A) vehicle control, (B) PGF2n, (C) 11-deoxy-PGElt and (D) PhXA85. Magnification, X400.
gens type I, III, and IV adjacent to ciliary smooth mus-cle cells in vitro. After 20-nM PG treatment, there aremoderate reductions in the amount of collagens typeI and III in extracts of the cell layer and minimalchanges in the amount of collagen type IV. With 200-nM PG treatment, the reductions of collagens type Iand III are larger, and there is increased collagen typeIV in the cell layer extract and in the culture medium.Immunocytochemical results indicate that in additionto quantitative changes, PGs induce changes in thestructural organization of these collagens. This is par-ticularly the case with collagen type IV after 200-nMPG treatment. The results of the experiments withEthD-1 and calcein AM indicate that none of the testtreatments induced cell death.
The reductions in collagens type I and III ob-served by EIA are consistent with the reductions infibril density seen in immunocytochemical studies. Alikely explanation is that the PG treatments induceincreased degradation of these ECM components.This possibility is supported by the increased pro-MMP-1 in the medium observed after PG-treatmentsof ciliary smooth muscle cells in vitro.18 In that colla-gens type I and III are found in the ciliary musclestromal ECM,19'20 it is possible that degradation ofthese collagens may contribute to the reduction inmonkey ciliary muscle stromal matrix observed aftertopical PG treatments.910 Studies in vivo are necessaryto confirm this possibility.
We noted an apparent difference between the col-lagen type III EIA and immunocytochemical resultsafter 200 nM PG treatment. Collagen type III was re-duced by 93% to 99% in the extracts from culturestreated for 24 hours with 200 nM PGs, whereas cul-tures treated for 3 days with 200 nM PGs containedmoderate amounts of collagen type III visible in immu-nocytochemical studies (Figs. 4B, 5). It is possible that
a portion of the collagen type III in these cultures mayhave been resistant to the extraction procedure andthat this resistant pool was visualized by immunocyto-chemistry. Normal cross-linking of collagen moleculesoccurring as monomers are incorporated into ECMwas inhibited by adding the lysyl oxidase inhibitor (3-aminopropionitrile to the culture medium. However,some cross-link formation may have occurred that hin-dered the total extraction of ECM collagens. Neverthe-less, the trend found in results of EIA and immunocy-tochemical analysis was reduction of collagens type Iand III after treatment with each PG.
Significant changes in the amount of collagentype IV in the cultures were not seen after 20 nM PGtreatment. However, the increased amounts of thiscollagen seen in the cell layer extracts and in the cul-ture medium after 200 nM PG treatment suggest astimulation of collagen type IV biosynthesis. The sig-nificance of this response may be limited for a numberof reasons. First, the assembly of basement membraneson cell surfaces appears to be limited by cellular ex-pression of appropriate integrin receptors.36'37 Thus,unincorporated collagen type IV might be washedaway by uveoscleral outflow. Second, there may bestructural condensation of the collagen type IV in situthat is analogous to the changes seen in vitro. Suchcondensation, with an associated increase in open
FIGURE 8. Ciliary muscle cultures treated for three days withvehicle or 200 nM PGF2tt and then stained with a mixtureof calcein AM and EthD-1. Calcein AM, which is taken upby living cells only, was visible under fluorescein optics inthe cytoplasm of all cells treated with (A) vehicle or (B)PGF2tt. (C) EthD-1, which stains the nuclear DNA of deadcells and is excluded from living cells, was visible underrhodamine optics in all the cell nuclei of control culturesthat had been killed by exposure to methanol for 30 minutesbefore staining. For clarity, a preconfluent culture is shownhere. In contrast, EthD-1 was excluded from (D) confluentcell cultures that had been exposed to 200 nM PGF2a for 3days. EthD-1 = ethidium homodimer. Magnification (A, B),X290; (C, D), X460.

Prostaglandins Alter Ciliary Muscle Extracellular Matrix 2221
spaces, could reduce the hydraulic resistance contrib-uted by collagen type IV.
Another important question is the concentrationof PG seen by PG receptors on the ciliary muscle cellsurface after clinically relevant, topical PG treatments.After topical treatment of rabbit eyes with labeledPGF2a, the amount of PGF2Q. or its derivatives per milli-gram of ciliary muscle was higher than the "amountper milligram of aqueous."38 It has been suggestedthat topical PGF2o. directly passes through the sclera toenter the ciliary muscle without traversing the anteriorchamber. In contrast, when PGF2cv esters were appliedtopically to rabbit eyes, similar (within one to twofold)milligram-specific amounts of PGF2a or its derivativeswere found in the aqueous and ciliary muscle.39
Hence, with the ester PG analogues, it may be reason-able to estimate roughly the ciliary muscle concentra-tion from the aqueous concentration. In the case ofthe PG ester latanoprost, ~ 1 % of the dose given topi-cally to cynomolgus monkeys was detected in the aque-ous humor.'10 In humans, the concentration of latano-prost-free acid in the aqueous humor has been foundto peak at approximately 100 nM and to decay withhalf-life of 3.5 hours (J. Stjernschantz, personal com-munication, 1997). Thus, the appearance of ECMchanges after 20 and 200 nM PGs in the current studysupports the possibility that ECM changes may occurin vivo after IOP-lowering PG treatments.
Each of the PGs investigated in this study bindswith different affinity to ocular PG receptors and in-fluences IOP differently. For example, PGF2cv can re-duce IOP in many animals and in humans1'38'41"44 andhas heterogeneous affinity with several receptors, in-cluding strong binding to the FP receptor and moder-ate binding to EP,, EP3, and TP receptors.40'45 17PT-PGF2a, which binds with high specific affinity to theFP receptor and minimally to other PG receptors,46
significantly reduces IOP in monkeys.42 This analoguealso reduces IOP in dogs; however, its effect on IOPin cats was insignificant.41'42 Latanoprost also has highaffinity for the FP receptor and much less affinity forother PG receptors.40 Topical administration of lata-noprost substantially reduces IOP in monkeys and inglaucoma patients.3'40'47'48 In the current study, latano-prost-free acid (PhXA85) induced alterations of colla-gens type I, III, and IV adjacent to the ciliary musclecells. This suggests that FP receptor activation can me-diate lowering of IOP and alteration of ECM.
Activation of other PG receptors besides the FPreceptor can lead to IOP reduction and ECM changessimilar to specific FP receptor activation, although par-ticular details may vary. For example, 11-deoxy-PGE)does not bind to FP receptors but does have a highaffinity for EP3 receptors and moderate affinity withEP2 receptors.42'49 Topical application of this analoguereduces IOP in the monkey.42 Unlike PGF2a, however,
repeated 11-deoxy-PGEi treatments do not cause sus-tained reduction of IOP. In the current study, theminimal change in the amount of collagen type IV inthe cell layer after 11-deoxy PGEi exposure may reflectthis different receptor specificity. The structuralchanges in the collagen type IV lattice observed inimmunocytochemical analysis suggest that 11-deoxyPGE] increases degradation of this collagen. This issupported by the increased MMPs observed in themedium of ciliary smooth muscle cell cultures after11-deoxy PGEi treatment.18 In addition, the modestincrease in the amount of collagen type IV in theculture medium indicates increased collagen type IVbiosynthesis. The difference in magnitude of theseresponses from those mediated by the same concen-tration of FP receptor-selective agonists probably re-flects the different signal transduction mecha-nisms.50'51 Nevertheless, it is possible that the signaltransduction pathways activated by these various re-ceptors may converge to similarly activate ECM turn-over. It should be noted that additional physiologicalresponses (i.e., relaxation) may contribute to PG-me-diated lowering of IOP.52
Finally, an important consideration in evaluatingthe significance of these results is the presence ofmany regulators of proMMP activation, as well as spe-cific inhibitors of MMPs in aqueous humor. Theseinclude tissue-type and urokinase-type plasminogenactivator, al-protease inhibitor, a2-macroglobulin,and tissue inhibitors of matrix metalloproteinases(TIMPs).25'26'53"55 In addition, MMP-3, which is pro-duced by ciliary smooth muscle cells, also can play animportant role in the activation of several differentproMMPs.18'56 Thus, activation of proMMPs releasedin response to PGs may be highly regulated in vivo.In view of the current results, it will be important todetermine the potential role of these molecules in theocular response to topical PGs.
Key Words
extracellular matrix, glaucoma, human ciliary muscle, pros-taglandins, uveoscleral outflow
References
1. Giuffre G. The effects of prostaglandin ¥^a in the hu-man eye. Graefes Arch Clin Exp Ophthalmol. 1985;222:139-141.
2. Bito LZ, Stjernschantz J, Resul B, Miranda OC, BasuS. The ocular effects of prostaglandins and the thera-peutic potential of a new PGF2a analog, PhXA41 (lata-noprost), for glaucoma management. J Lipid Medial.1993; 6:535-543.
3. Camras C, Aim A, Watson P, Stjernschantz J, and theLatanoprost Study Groups. Latanoprost, a prostaglan-din analog, for glaucoma therapy. Efficiency andsafety after 1 year treatment in 198 patients. Ophthal-mology. 1996; 103:1916-1924.

2222 Investigative Ophthalmology & Visual Science, October 1997, Vol. 38, No. 11
4. Camras CB, Bhuyan KC, Podos SM, Bhuyan DK, Mas-ter RW. Multiple dosing of prostaglandin F2a or epi-nephrine on cynomolgus monkey eyes. II Slit lampbiomicroscopy, aqueous humor analysis, and fluores-cein angiography. Invest Ophthalmol Vis Sci. 1987;28:921-926.
5. Gabelt BT, Kaufman PL. Prostaglandin F2a increasesuveoscleral outflow in the cynomolgus monkey. ExpEye Res. 1989;49:389-402.
6. Nilsson SF, Samuelsson M, Bill A, Stjernschantz J. In-creased uveoscleral outflow as a possible mechanismof ocular hypotension caused by prostaglandin F2a-1-isopropylester in the cynomolgus monkey. Exp Eye Res.1989; 48:707-717.
7. Crawford K, Kaufman PL. Pilocarpine antagonizesprostaglandin F2a-induced ocular hypotension inmonkeys. Evidence for enhancement of uveoscleraloutflow by prostaglandin F2a. Arch Ophthalmol. 1987;105:1112-1116.
8. Camras CB. Mechanism of the prostaglandin-inducedreduction of intraocular pressure in humans. Adv Pros-taglandin Thromboxane Leukot Res. 1995;23:519-525.
9. Tamm E, Lutjen-Drecoll E, Rohen JW. Age-relatedchanges of the ciliary muscle in comparison withchanges induced by treatment with prostaglandin F2a.An ultrastructural study in rhesus and cynomolgusmonkeys. Mech AgeingDev. 1990;51:101-120.
10. Lutjen-Drecoll E, Tamm E. Morphological study ofthe anterior segment of cynomolgus monkey eyes fol-lowing treatment with prostaglandin F2a. Exp Eye Res.1988; 47:761-69.
11. Wood RL, Koseki T, Kelly DE. Uveoscleral permeabil-ity to intracamerally infused ferritin in eyes of rabbitsand monkeys. Cell Tissue Res. 1992;270:559-567.
12. Bill A. Uveoscleral drainage of aqueous humor: Physi-ology and pharmacology. Prog Clin Biol Res. 1989;312:417-427.
13. Baramova E, Foidart JM. Matrix rnetalloproteinasefamily. Cell Biol Int. 1995; 19:239-242.
14. Woessner J Jr. The family of matrix metalloprotein-ases. Ann N Y Acad Sci. 1994;732:11-21.
15. Corcoran ML, Kleiner D Jr, Stetler-Stevenson WG.Regulation of matrix metalloproteinases during extra-cellular matrix turnover. Adv Exp Med Biol. 1995;385:151-159.
16. Matrisian LM. The matrix-degrading metalloprotein-ases. Bioessays. 1992; 14:455-463.
17. Birkedal-Hansen H, Moore WG, Bodden MK, et al.Matrix metalloproteinases: A review. Crit Rev Oral BiolMed. 1993;4:197-250.
18. LindseyJD, Kashiwagi K, Boyle D, et al. Prostaglandinsincrease proMMP-1 and proMMP-3 secretion by hu-man ciliary smooth muscle cells. Cuir Eye Res.1996; 15:869-875.
19. Weinreb RN, LindseyJ, Luo XX, Wang T-H. Extracel-lular matrix of the human ciliary muscle. / Glaucoma.1994; 3:70-78.
20. Tamm E, Baur A, Lutjen-Drecoll E. Synthesis of ex-tracellular matrix components by human ciliary mus-cle cells in culture. Curr Eye Res. 1992; 11:333-41.
21. Marshall GE, Konstas AG, Abraham S, Lee WR. Extra-
cellular matrix in aged human ciliary body: An immu-noelectron microscope study. Invest Ophthalmol Vis Sci.1992; 33:2546-2560.
22. Weinreb RN, Kim DM, Lindsey JD. Propagation ofciliary smooth muscle cells in vitro and effects of pros-taglandin F2a on calcium efflux. Invest Ophthalmol VisSci. 1992; 33:2679-2686.
23. Kashiwagi K, Lindsey JD, Kashiwagi F, Tsukahara S,Weinreb RN. Calponin distribution in human ciliarymuscle and other anterior segment tissues. Invest Oph-thalmol Vis Sci. 1997;38:349-356.
24. Basu S, Sjoquist B, Stjernschantz J, Resul B. Cornealpermeability to and ocular metabolism of phenyl sub-stituted prostaglandin esters in vitro. ProstaglandinsLeukot Essent Fatty Acids. 1994; 50:161-168.
25. Ando H, Twining SS, Yue BY, et al. MMPs and protein-ase inhibitors in the human aqueous humor. InvestOphthalmol Vis Sci. 1993;34:3541-3548.
26. Di Giroloamo N, Verma M, McCluskey P, Lloyd A,Wakefield D. Increased matrix metalloproteinases inthe aqueous humor of patients and experimental ani-mals with uveitis. Curr Eye Res. 1996; 15:1060-1068.
27. Amenta PS, Gay S, Vaheri A, Martinez-Hernandez A.The extracellular matrix is an integrated unit: Ultra-structural localization of collagen types I, III, IV, V,VI, fibronectin, and laminin in human term placenta.Coll Relat Res. 1986;6:125-152.
28. Shekhonin BV, Domogatsky SP, Muzykantov VR, Idel-son GL, Rukosuev VS. Distribution of type I, III, IVand V collagen in normal and atherosclerotic humanarterial wall: Immunomorphological characteristics.Coll Relat Res. 1985;5:355-368.
29. Rukosuev VS, Nanaev AK, Milovanov AP. Participationof collagen types I, III, IV, V, and fibronectin in theformation of villi fibrosis in human term placenta.Acta Histochem. 1990;89:ll-16.
30. Godfrey M, Keene DR, Blank E, et al. Type II achon-drogenesis-hypochondrogenesis: Morphologic andimmunohistopathologic studies. Am J Hum Genet.1988; 43:894-903.
31. Gusterson BA, Warburton MJ, Mitchell D, Kraft N,Hancock WW. Invading squamous cell carcinoma canretain a basal lamina. An immunohistochemical studyusing a monoclonal antibody to type TV collagen. LabInvest. 1984; 51:82-87.
32. Hancock WW, Kraft N, Clarke F, Atkins RC. Produc-tion of monoclonal antibodies to fibronectin, type IVcollagen and other antigens of the human glomeru-lus. Pathology. 1984; 16:197-206.
33. Frid MG, Shekhonin BV, Koteliansky VE, GlukhovaMA. Phenotypic changes of human smooth musclecells during development: Late expression of heavycaldesmon and calponin. Dev Biol. 1992; 153:185-193.
34. Skalli O, Ropraz P, Trzeciak A, et al. A monoclonalantibody against alpha-smooth muscle actin: A newprobe for smooth muscle differentiation. / Cell Biol.1986; 103:2787-2796.
35. Obata K, Iwata K, Ichida T, et al. One step sandwichenzyme immunoassay for human type IV collagen us-ing monoclonal antibodies. Clin Chim Acta.1989; 181:293-303.

Prostaglandins Alter Ciliary Muscle Extracellular Matrix 2223
36. Eble J, Goblik R, Mann K, Kuhn K. The alBl integrinrecognition site of the basement membrane collagenmolecule [al(IV)]2a2(IV). EMBO J. 1993;12:4795-4802.
37. Downer C, Watt F, Speight P. Loss of a6B4 integrinsubunits coincides with loss of basement membranecomponents in oral squamous cell carcinomas. / Pa-thol. 1993;171:183-190.
38. Bito L, Baroody R. The penetration of exogenousprostaglandin and arachidonic acid into, and theirdistribution within, the mammalian eye. Curr Eye Res.1981/1982; 1:659-669.
39. Bito LZ, Baroody RA. The ocular pharmacokineticsof eicosanoids and their derivatives. 1. Comparison ofocular eicosanoid penetration and distribution follow-ing the topical application of PGF2a, PGF2a-l-methylester, and PGF2a-l-isopropyl ester. Exp Eye Res.1987;44:2l7-226.
40. Stjernschantz J, Selen G, Sjoquist B, Resul B. Preclini-cal pharmacology of latanoprost, a phenyl-substitutedPGF2a analog. Adv Prostaglandin Thromboxane LeukotRes. 1995;23:513-518.
41. Woodward DF, Burke JA, Williams LS, et al. Prosta-glandin F2o effects on intraocular pressure negativelycorrelate with FP-receptor stimulation. Invest Ophthal-mol Vis Sci. 1989; 30:1838-1842.
42. Woodward DF, Lawrence RA, Fairbairn CE, Shan T,Williams LS. Intraocular pressure effects of selectiveprostanoid receptor agonists involve different recep-tor subtypes according to radioligand binding studies./Lipid Mediat. 1993;6:545-553.
43. Woodward DF, Chan MF, Burke JA, et al. Studies onthe ocular hypotensive effects of prostaglandin F2a es-ter prodrugs and receptor selective prostaglandin ana-logs. / Ocul Pharmacol Ther. 1994; 10:177-93.
44. Woodward D, Bogardus A, Donello J, et al. Molecularcharacterization and ocular hypotensive properties ofthe prostanoid EP2 Receptor. J Ocular Pharmacol Ther.1995; 11:447-454.
45. Eglen R, Whiting R. Characterization of the prostan-oid receptor profile of enprostil and isomers in
smooth muscle and platelets in vitro. Br J Pharmacol.1989;98:1335-1343.
46. Miller WL, Weeks JR, Lauderdale JW, Kirton KT. Bio-logical activities of l7-phenyl-18,19,20-trinorprostag-landins. Prostaglandins. 1975;9:9-18.
47. Aim A, Villumsen J, Tornquist P, et al. Intraocularpressure-reducing effect of PhXA41 in patients withincreased eye pressure. A one-month study. Ophthal-mology. 1993; 100:1312-1316.
48. Racz P, Ruzsonyi MR, Nagy ZT, Gaygi Z, Bito LZ.Around-the-clock intraocular pressure reduction withonce-daily application of latanoprost by itself or incombination with timolol. Arch Ophthalmol. 1996;114:268-273.
49. Woodward D, Fairbairn C, Krauss H-P, Lawrence R,Protzman C. Radioligand binding analysis of receptorsubtypes in two FP receptor preparations that exhibitdifferent functional rank orders of potency in re-sponse to prostaglandins. / Pharmacol Exp Ther.1995;273:285-291.
50. Ashby B. Interactions among prostaglandin receptors.Receptor. 1994; 4:31-42.
51. Negishi M, Sugimoto Y, Ichikawa A. Molecular mecha-nisms of diverse actions of prostanoid receptors. Bio-chim BiophysActa. 1995; 1259:109-119.
52. Poyer J, Miller C, Kaufman P. Prostaglandin F2a effectson isolated rhesus monkey ciliary muscle. Invest Oph-thalmol Vis Sci. 1995;36:2461-2465.
53. Bernatchez SF, Tabatabay C, Belin D. Urokinase-typeplasminogen activator in human aqueous humor. In-vest Ophthalmol Vis Sci. 1992;33:2687-2692.
54. Tripathi RC, Park JK, Tripathi BJ, Millard CB. Tissueplasminogen activator in human aqueous humor andits possible therapeutic significance. Am J Ophthalmol.1988; 106:719-722.
55. Vadillo-Ortega F, Gonzalez-Avila G, Chevez P, et al.A latent collagenase in human aqueous humor. InvestOphthalmol Vis Sci. 1989;30:332-335.
56. Nagase H, Suzuki K, Morodomi T, Enghild JJ, SalvesenG. Activation mechanisms of the precursors of matrixmetalloproteinases 1, 2, and 3. Matrix Suppl. 1992;1:237-244.
Copyright © 2022 FDOKUMEN