
Ciliary Activity in the Oviduct of Cycling, Pregnant, and Muscarinic Receptor Knockout Mice
Upload
independentCategory
view
1download
0
Cerebrospinal fluid (CSF) fills the ventricles andsubarachnoid spaces enveloping the central nervous system(CNS) and has been thought to serve as a transport mediumfor disseminating global CNS signals (Nicholson, 1999).Ependymal cells form a single-layered, ciliated cellularinterface that lines the ventricular surface of vertebratebrain and the central canal of the spinal cord (Del Bigio,1995). Putative functions for these ciliated ependymal cellsinclude: acting as neural stem cells (Johansson et al. 1999),moving cellular debris in the direction of bulk CSF flow(Cathcart & Worthington, 1964), and locally mixing CSFthereby minimizing the unstirred CSF layer over theependyma and optimizing the dispersion of neuralmessengers in the CSF (Roth et al. 1985). Local mixinginduced by ependymal cilia beating can influence theexchange between CSF and brain, thereby potentiallyregulating communication within the CNS. Unlike ciliated
cells of the reproductive and respiratory epithelia, whichhave been extensively investigated, the functions andregulatory mechanisms of mammalian brain ependymalciliated cells still remain poorly understood (Del Bigio,1995). While there has been much speculation regarding theregulatory mechanisms involved in controlling the cilia beatfrequency (CBF) of ependymal cells, the specific pathwaysinvolved in such regulation have not been elucidated.
The present experiments investigated the control of ciliaryactivity in ependymal cells by serotonin (5-HT) and ATP.Here we present the first direct evidence on mechanismsthat control cilia activity in mammalian brain ependymalcells. The results show that ciliated ependymal cells havetwo unique and independent signalling pathways withopposing effects. One pathway that stimulates CBF ismediated by the activation of 5-HT receptors, resulting inIP×-mediated release of Ca¥ from intracellular Ca¥ stores.
Journal of Physiology (2001), 531.1, pp.131—140 131
Intracellular pathways regulating ciliary beating of rat brain
ependymal cells
Thien Nguyen*, Wei-Chun Chin*, Jennifer A. O’Brien†, Pedro Verdugo*‡and Albert J. Berger†
Departments of *Bioengineering, †Physiology & Biophysics and ‡Medicine, University ofWashington, Box 357962, Seattle, Washington 98195, USA
(Received 22 May 2000; accepted after revision 17 October 2000)
1. The mammalian brain ventricles are lined with ciliated ependymal cells. As yet little is knownabout the mechanisms by which neurotransmitters regulate cilia beat frequency (CBF).
2. Application of 5-HT to ependymal cells in cultured rat brainstem slices caused CBF toincrease. 5-HT had an EC50 of 30 ìÒ and at 100 ìÒ attained a near-maximal CBF increaseof 52·7 ± 4·1% (mean ± s.d.) (n = 8).
3. Bathing slices in Ca¥-free solution markedly reduced the 5-HT-mediated increase in CBF.Fluorescence measurements revealed that 5-HT caused a marked transient elevation incytosolic Ca¥ ([Ca¥]c) that then slowly decreased to a plateau level. Analysis showed thatthe [Ca¥]c transient was due to release of Ca¥ from inositol 1,4,5-trisphosphate (IP×)-sensitive stores; the plateau was probably due to extracellular Ca¥ influx through Ca¥release-activated Ca¥ (CRAC) channels.
4. Application of ATP caused a sustained decrease in CBF. ATP had an EC50 of about 50 ìÒand 100 ìÒ ATP resulted in a maximal 57·5 ± 6·5% (n = 12) decrease in CBF. The ATP-induced decrease in CBF was unaffected by lowering extracellular [Ca¥], and no changes in[Ca¥]c were observed. Exposure of ependymal cells to forskolin caused a decrease in CBF.Ciliated ependymal cells loaded with caged cAMP exhibited a 54·3 ± 7·5% (n = 9) decreasein CBF following uncaging. These results suggest that ATP reduces CBF by a Ca¥-independent cAMP-mediated pathway.
5. Application of 5-HT and adenosine-5'-O-3-thiotriphosphate (ATP-ã-S) to acutely isolatedciliated ependymal cells resulted in CBF responses similar to those of ependymal cells incultured slices suggesting that these neurotransmitters act directly on these cells.
6. The opposite response of ciliated ependymal cells to 5-HT and ATP provides a novelmechanism for their active involvement in central nervous system signalling.
11156
Keywords:

This initial elevation of cytosolic Ca¥ causes a prolongedincrease of intracellular Ca¥ due to the opening of Ca¥release-activated Ca¥ (CRAC) channels on the plasmamembrane. Ca¥ influx through CRAC channels contributesto most of the observed increase in CBF. The secondpathway that reduces CBF is activated by ATP. Theinhibitory effect is independent of intracellular Ca¥ changeand may involve the activation of purinergic receptors andis transduced by intracellular adenosine 3',5'-cyclicmonophosphate (cAMP).
METHODS
Ciliated ependymal cells of the 4th ventricle were studied either inbrain slices grown for 1—3 days in culture or in a smaller number ofexperiments as acutely isolated ependymal cells. In both casesSprague-Dawley rat pups (8—10 days old) were anaesthetized witheither a ketamine—xylazine mixture (200 mg kg¢ and 14 mg kg¢,i.m., respectively) or 30% halothane in air and then decapitated.The medulla was isolated and sliced in 300 ìm-thick sections whilebeing submerged in ice-cold Ringer solution containing (mÒ): 119NaCl, 2·5 KCl, 1·3 MgSOÚ, 1 NaHµPOÚ, 26·2 NaHCO×, 11 glucose,and 2·5 CaClµ, bubbled with 95% Oµ—5% COµ. For studies ofependymal cells in slices we transferred these acutely preparedslices to a keratinocyte serum-free medium (Life Technologies,Gaithersburg, MD, USA) with 10% fetal calf serum for 1—3 days.The University of Washington Animal Care and Use Committeeapproved all procedures.
For the preparation of isolated cells, slices were initially incubatedin Hanks’ solution containing 10 units ml¢ protease (type XIV,Sigma, MO, USA) for 20 min at 37°C. The ciliated ependymallayer was then carefully removed from the brainstem slice andrinsed with Hanks’ solution. Gentle shear force through ahypodermic needle was used to break the ependymal layer intoindividual isolated ciliated cells. Aliquots of the cell suspensionwere then allowed to attach to the floor of polylysine-coated glasschambers and mounted on the stage of a microscope for observation.
All test substances were added to a static bath solution thatcontained the cultured slices or acutely isolated cells. Measurementsof ciliary beating and cytosolic Ca¥ were carried out at 37°C,while measurements of electrical properties were performed atroom temperature, approximately 22°C.
Measurement of ciliary beating
The ciliary activity measuring system is based on a previouslydescribed method, that has been modified to measure on-lineciliary beating of single ciliated cells (Ben-Shimol et al. 1991;Sanderson & Dirksen, 1995; Nguyen, 1996). The apparatus consistsof a Nikon Diaphot inverted microscope with a ²100, 1·4 NA oilimmersion objective and a DC illumination source. Images werecaptured by a thermoelectrically cooled, low dark noise(1·3 photoelectrons s¢ pixel¢ at −36°C) digital camera with a336 ² 243 CCD matrix, a 16-bit pixel resolution, and a readout rateof 10Ç pixel s¢ (Spectra Source Model 400, Westlake Village, CA,USA). The camera was mounted in the photoport of the microscopeusing a ²20 relay lens yielding a 10 pixel ìm¢ resolution. Toincrease the sampling rate, we avoided capturing the whole image.Instead, single line scans were sampled at a rate of 256 scans s¢and an on-line fast Fourier transform of the line samples gave theCBF. Each scan sampled an area 0·2 ìm ² 30 ìm across the opticalfield of the ciliated ependymal cells. The CBF from this digitalphotography method was calibrated against the manual counting of
ciliary beating in images captured with fast cinematography (128pictures s¢). The CBF obtained by the two techniques yielded thesame beat frequency over the range of 0—28 Hz (Nguyen, 1996).
Measurement of cytosolic [Ca¥]
Intact ependymal cells in cultured slices were equilibrated for30 min in Hanks’ solution at 37°C, containing 5 ìÒ rhod-2-acetoxy-methyl ester (Molecular Probes, Eugene, OR, USA) from a stocksolution in dimethyl sulfoxide (DMSO). The final concentration ofDMSO in the Hanks’ solution was 0·1%. Rhod-2 has a high Ca¥affinity (Kd = 570 nÒ, in the absence of Mg¥), allowing themeasurement of a low range of [Ca¥]c. The Kd of rhod-2 wascalculated from direct measurements performed on cells treatedwith the Ca¥-ionophore A23187 following procedures describedelsewhere (Kao, 1994; Nguyen et al. 1998). After dye loading, thecells were washed and equilibrated in Hanks’ solution for 30 min.Ciliated cells were also loaded with 500 ng ml¢ of the fluorescentdye DiOCÜ(3) (Molecular Probes) for 5—8 min and then washed inHanks’ solution. This lipophylic, cationic dye has been usedextensively to identify intracellular organelles including the ER ofa variety of cells (Terasaki et al. 1984). Since the excitationwavelength of DiOCÜ(3) and the Ca¥ probes are different (484 and525 nm, respectively), the double-labelling strategy allows selectionof image planes and line scans without photobleaching the Ca¥probes. After a selected ciliated cell was identified and focusedusing the DiOCÜ(3) label, the filter cube was changed to excite(546 ± 10 nm) and collect the emission of rhod-2 while the cellswere stimulated with either ATP or 5-HT. The optical sectioningtechnique and its calibration have been described elsewhere (Moncket al. 1992; Nguyen et al. 1998).
Measurement of electrical properties of ependymal cells
Electrical measurements were performed on visually identified 4thventricle ependymal cells in acutely prepared brainstem slices asdescribed above from neonatal rats (8—10 days old). Immediatelyafter slicing, the slices were incubated for 1 h at 37°C. Duringwhole-cell recordings slices were perfused by a Ringer solutioncontaining (mÒ): 119 NaCl, 26·2 NaHCO×, 1 NaHµPOÚ, 2·5 KCl, 11glucose, 2·5 CaClµ, and 1·3 MgSOÚ. Using near-infrared DIC optics,ependymal cells were identified based on their location andmorphology. On most occasions cells were filled with the fixablefluorescent dye Alexa 488 (200 mÒ, Molecular Probes), allowingvisualization of ependymal cells during and after recordings.Whole-cell patch clamp recordings were performed at roomtemperature using an Axoclamp-2B amplifier and pCLAMP 7.0software (Axon Instruments). Patch electrodes (resistance 6—8 MÙ)were filled with (mÒ): 145 KCl, 10 Hepes, 1 EGTA, 2 MgClµ, 2ATP-Mg, 0·2 GTP-Tris (pH 7·2).
5-HT immunocytochemistry
Sprague-Dawley rats (7—14 days old) were anaesthetized byinjection (i.m.) of a ketamine—xylazine mixture (200 mg kg¢ and14 mg kg¢, respectively). Following decapitation, the brainstemwas removed and transverse brainstem slices (300—1000 ìm) werecut with a vibratome in a cold Ringer solution containing (mÒ): 119NaCl, 26·2 NaHCO×, 1 NaHµPOÚ, 2·5 KCl, 11 glucose, 2·5 CaClµ,and 1·3 MgSOÚ. Tissue slices were then fixed in 4%paraformaldehyde in 0·1 Ò sodium phosphate at 4°C overnight.After fixation, the tissue was washed in phosphate-buffered salinesolution (PBS) and placed in 30% sucrose in PBS overnight at 4°C.Following cryoprotection, slices were resectioned (50—70 ìm thick)using a sliding microtome.
For immunocytochemistry, free-floating tissue sections wereincubated in PBS containing 0·2% Triton X-100 and 10% donkeyserum (Vector Laboratories) for 1 h. Tissues were then incubated
T. Nguyen, W.-C. Chin, J. A. O’Brien, P. Verdugo and A. J. Berger J. Physiol. 531.1132

overnight in tryptophan hydroxylase antibody (Sigma T0678,1:400 or 1:500) in PBS containing 0·2% Triton X-100 and 10%donkey serum. After washing, tissue was then incubated in CY3donkey anti-mouse secondary antibody (Jackson ImmunoResearchInc, 1:600) for 90 min in PBS containing Triton X-100 and donkeyserum. Tissue was washed and mounted onto slides usingVectaShield (Vector Laboratories). Negative control assays wereperformed by omitting the primary antibody. Tissue was imagedusing a BioRad confocal microscope.
RESULTS
Localization of 5-HT-containing processes in the rat
brainstem ependymal cell layer
We used immunocytochemistry to demonstrate that 5-HT-containing nerve fibres and processes are present in theependymal cell layer of the 4th ventricle and central canal inrats of the same age as used in our functional studies (seebelow). To do this we used an antibody against tryptophanhydroxylase, a key enzyme in the biosynthetic pathway for5-HT, to show that the 4th ventricle ependymal cell layerhas nerve fibres and varicosities that contain this enzyme(Fig. 1A, n = 5 rats). Figure 1B shows in the same section ofthe medulla the presence of 5-HT-containing neurons in thenearby midline nucleus raphe obscurus. Since raphe neuronsare an important source of the brain’s 5-HT (Cooper et al.1996) these results serve as a positive control for our
findings in the ependymal cell layer. Further, we observedindividual 5-HT-containing axons projecting into theependymal cell layer that arose from single 5-HT-containingneurons (data not shown). Our results are in agreement withprevious studies that reported the presence of a 5-HT-containing nerve plexus extending over most of the ratbrain ependymal cell layer (Chan-Paly, 1976; Voutsinos et al.1994). While distinct classical synaptic associations between5-HT-containing nerve fibres and ependymal cells havebeen reported to be absent, it is nevertheless widely believedthat ependymal cells are the targets of this 5-HT-containingnerve plexus (Lorez & Richards, 1982; Voutsinos et al. 1994;Dinopoulos & Dori, 1995).
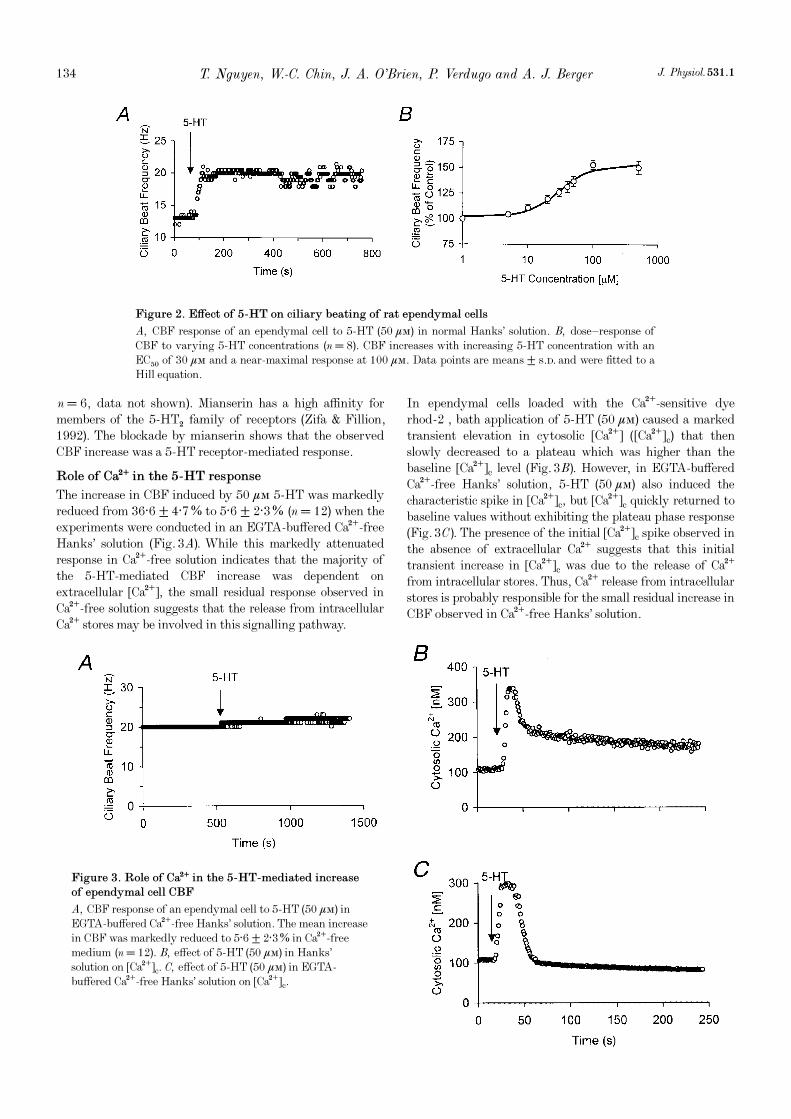
Response to 5-HT
The average basal CBF of the population of intact ciliatedependymal cells exposed to 5-HT was 19 ± 5 Hz (mean± s.d.) (n = 8). The basal CBF remained fairly stable insingle cells, with variability usually not larger than ±5%.Exposure to 5-HT caused a sustained increase of CBF(Fig. 2A). The 5-HT dose—response data were fitted to a Hillequation which gave an EC50 of 30 ìÒ and a slope factor of1·7 (Fig. 2B). At a 5-HT concentration of 100 ìÒ CBFattained a near-maximal response increase of 52·7 ± 4·1%(n = 8) in CBF (Fig. 2B). The 5-HT response was completelyblocked by the 5-HT receptor antagonist mianserin (5 ìÒ,
Regulation of brain ependymal ciliaJ. Physiol. 531.1 133
Figure 1. 5-HT immunolabelling in transverse rat brainstem slices
A, tryptophan hydroxylase-positive processes are present at the ependymal cell layer. Arrow points to thebasal side of the ependymal cell layer where there are both 5-HT-containing nerve fibres and varicosities.Tissue section is from the medulla rostral to the obex where the 4th ventricle is clearly evident. This pictureis derived from successive z_series images of the tissue section. B, tryptophan hydroxylase-positive neuronsof the nucleus raphe obscurus are present in the same tissue section as in A. This picture is derived fromsingle z-image of the tissue section. Calibration bars: 50 ìm in A and 100 ìm in B.
n = 6, data not shown). Mianserin has a high affinity formembers of the 5-HTµ family of receptors (Zifa & Fillion,1992). The blockade by mianserin shows that the observedCBF increase was a 5-HT receptor-mediated response.
Role of Ca¥ in the 5-HT response
The increase in CBF induced by 50 ìÒ 5-HT was markedlyreduced from 36·6 ± 4·7% to 5·6 ± 2·3% (n = 12) when theexperiments were conducted in an EGTA-buffered Ca¥-freeHanks’ solution (Fig. 3A). While this markedly attenuatedresponse in Ca¥-free solution indicates that the majority ofthe 5-HT-mediated CBF increase was dependent onextracellular [Ca¥], the small residual response observed inCa¥-free solution suggests that the release from intracellularCa¥ stores may be involved in this signalling pathway.
In ependymal cells loaded with the Ca¥-sensitive dyerhod_2 , bath application of 5-HT (50 ìÒ) caused a markedtransient elevation in cytosolic [Ca¥] ([Ca¥]c) that thenslowly decreased to a plateau which was higher than thebaseline [Ca¥]c level (Fig. 3B). However, in EGTA-bufferedCa¥-free Hanks’ solution, 5-HT (50 ìÒ) also induced thecharacteristic spike in [Ca¥]c, but [Ca¥]c quickly returned tobaseline values without exhibiting the plateau phase response(Fig. 3C). The presence of the initial [Ca¥]c spike observed inthe absence of extracellular Ca¥ suggests that this initialtransient increase in [Ca¥]c was due to the release of Ca¥from intracellular stores. Thus, Ca¥ release from intracellularstores is probably responsible for the small residual increase inCBF observed in Ca¥-free Hanks’ solution.
T. Nguyen, W.-C. Chin, J. A. O’Brien, P. Verdugo and A. J. Berger J. Physiol. 531.1134
Figure 2. Effect of 5-HT on ciliary beating of rat ependymal cells
A, CBF response of an ependymal cell to 5-HT (50 ìÒ) in normal Hanks’ solution. B, dose—response ofCBF to varying 5-HT concentrations (n = 8). CBF increases with increasing 5-HT concentration with anEC50 of 30 ìÒ and a near-maximal response at 100 ìÒ. Data points are means ± s.d. and were fitted to aHill equation.
Figure 3. Role of Ca¥ in the 5-HT-mediated increase
of ependymal cell CBF
A, CBF response of an ependymal cell to 5-HT (50 ìÒ) inEGTA-buffered Ca¥-free Hanks’ solution. The mean increasein CBF was markedly reduced to 5·6 ± 2·3% in Ca¥-freemedium (n = 12). B, effect of 5-HT (50 ìÒ) in Hanks’solution on [Ca¥]c. C, effect of 5-HT (50 ìÒ) in EGTA-buffered Ca¥-free Hanks’ solution on [Ca¥]c.
A similar biphasic time course of [Ca¥]c, spike followed byplateau phase, has been observed in non-excitable KNRK-PAR-2 epithelial cell lines (rat kidney epithelial cellsexpressing human proteinase-activated receptor-2)stimulated with trypsin (B�ohm et al. 1996). In that systemthe initial [Ca¥]c spike was shown to be due to release ofCa¥ from intracellular Ca¥ stores, while the subsequent[Ca¥]c plateau phase was shown to result from extracellularCa¥ influx through Ca¥ release-activated Ca¥ (CRAC)channels (B�ohm et al. 1996). Thus Ca¥ released from intra-cellular Ca¥ stores induced the opening of CRAC channelson the plasma membrane, leading to the longer-lastingplateau phase of [Ca¥]c increase.
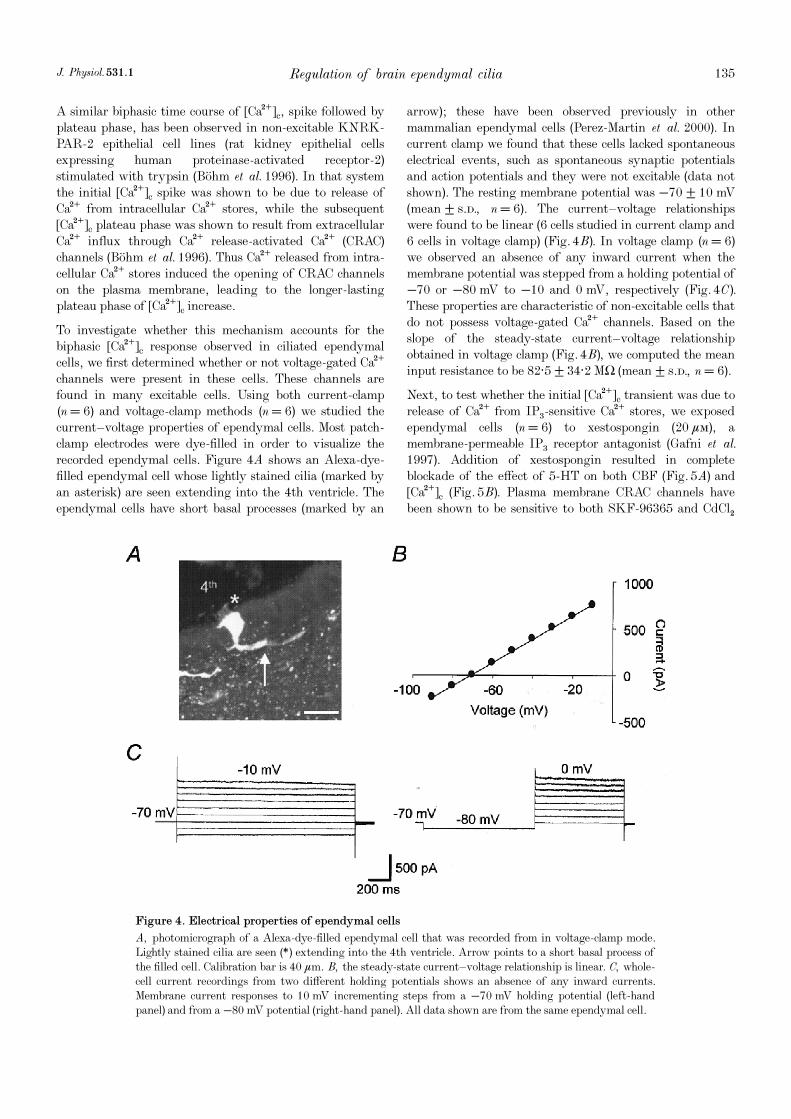
To investigate whether this mechanism accounts for thebiphasic [Ca¥]c response observed in ciliated ependymalcells, we first determined whether or not voltage-gated Ca¥channels were present in these cells. These channels arefound in many excitable cells. Using both current-clamp(n = 6) and voltage-clamp methods (n = 6) we studied thecurrent—voltage properties of ependymal cells. Most patch-clamp electrodes were dye-filled in order to visualize therecorded ependymal cells. Figure 4A shows an Alexa-dye-filled ependymal cell whose lightly stained cilia (marked byan asterisk) are seen extending into the 4th ventricle. Theependymal cells have short basal processes (marked by an
arrow); these have been observed previously in othermammalian ependymal cells (Perez-Martin et al. 2000). Incurrent clamp we found that these cells lacked spontaneouselectrical events, such as spontaneous synaptic potentialsand action potentials and they were not excitable (data notshown). The resting membrane potential was −70 ± 10 mV(mean ± s.d., n = 6). The current—voltage relationshipswere found to be linear (6 cells studied in current clamp and6 cells in voltage clamp) (Fig. 4B). In voltage clamp (n = 6)we observed an absence of any inward current when themembrane potential was stepped from a holding potential of−70 or −80 mV to −10 and 0 mV, respectively (Fig. 4C).These properties are characteristic of non-excitable cells thatdo not possess voltage-gated Ca¥ channels. Based on theslope of the steady-state current—voltage relationshipobtained in voltage clamp (Fig. 4B), we computed the meaninput resistance to be 82·5 ± 34·2 MÙ (mean ± s.d., n = 6).
Next, to test whether the initial [Ca¥]c transient was due torelease of Ca¥ from IP×-sensitive Ca¥ stores, we exposedependymal cells (n = 6) to xestospongin (20 ìÒ), amembrane-permeable IP× receptor antagonist (Gafni et al.1997). Addition of xestospongin resulted in completeblockade of the effect of 5-HT on both CBF (Fig. 5A) and[Ca¥]c (Fig. 5B). Plasma membrane CRAC channels havebeen shown to be sensitive to both SKF-96365 and CdClµ
Regulation of brain ependymal ciliaJ. Physiol. 531.1 135
Figure 4. Electrical properties of ependymal cells
A, photomicrograph of a Alexa-dye-filled ependymal cell that was recorded from in voltage-clamp mode.Lightly stained cilia are seen (*) extending into the 4th ventricle. Arrow points to a short basal process ofthe filled cell. Calibration bar is 40 ìm. B, the steady-state current—voltage relationship is linear. C, whole-cell current recordings from two different holding potentials shows an absence of any inward currents.Membrane current responses to 10 mV incrementing steps from a −70 mV holding potential (left-handpanel) and from a −80 mV potential (right-hand panel). All data shown are from the same ependymal cell.
(B�ohm et al. 1996; Parekh & Penner, 1997). To test theinvolvement of CRAC channels, we studied 5-HT (50 ìÒ)stimulation of ependymal cells exposed to eitherSKF_96365 (50 ìÒ, Tocris) or CdClµ (500 ìÒ), althoughSKF_96365 also blocks other ion channels (Franzius et al.1994). These experiments resulted in a response verysimilar to that observed in cells stimulated with 5-HT in
Ca¥-free Hanks’ solution (Fig. 3C), i.e. their responseconsisted of a large transient increase in [Ca¥]c without thesubsequent plateau phase of [Ca¥]c increase observed incontrol experiments. Correspondingly, the CBF increaseinduced by 50 ìÒ 5-HT was also markedly reduced to5·8 ± 1·7% (n = 5) and 4·2 ± 2·1% (n = 7) in the presenceof SKF_96365 and CdClµ, respectively (data not shown).
T. Nguyen, W.-C. Chin, J. A. O’Brien, P. Verdugo and A. J. Berger J. Physiol. 531.1136
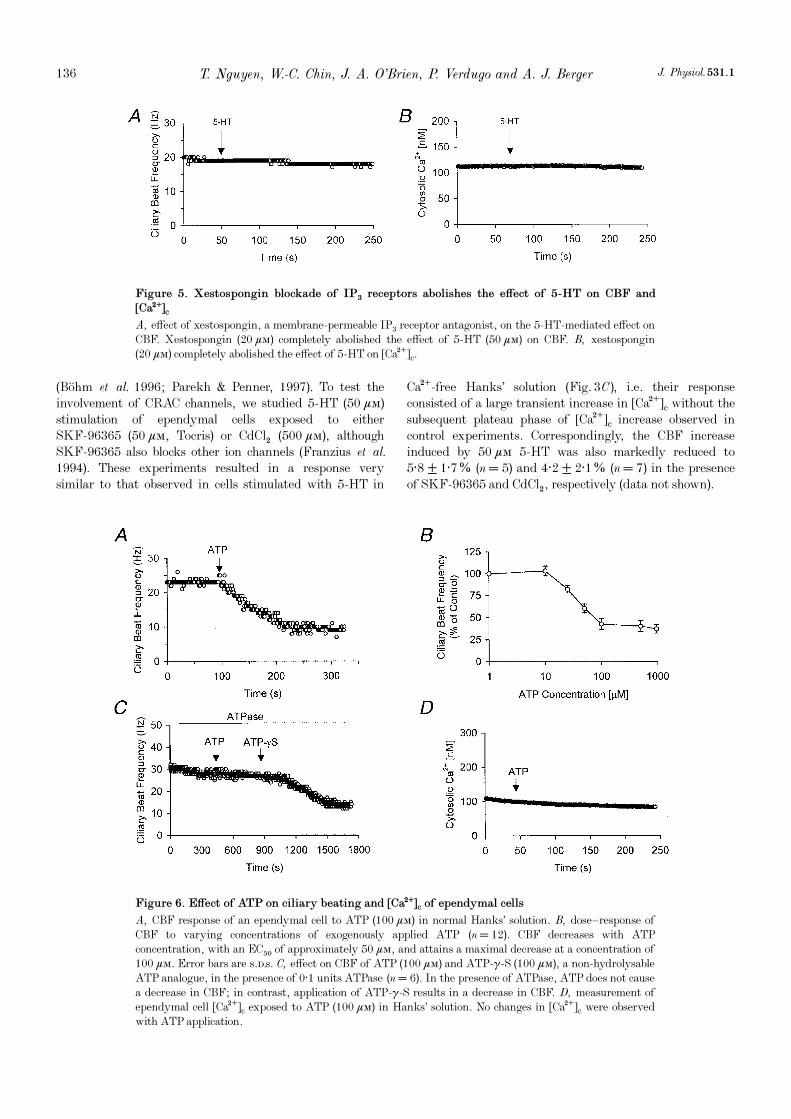
Figure 5. Xestospongin blockade of IP× receptors abolishes the effect of 5-HT on CBF and
[Ca¥]c
A, effect of xestospongin, a membrane-permeable IP× receptor antagonist, on the 5-HT-mediated effect onCBF. Xestospongin (20 ìÒ) completely abolished the effect of 5_HT (50 ìÒ) on CBF. B, xestospongin(20 ìÒ) completely abolished the effect of 5-HT on [Ca¥]c.
Figure 6. Effect of ATP on ciliary beating and [Ca¥]c of ependymal cells
A, CBF response of an ependymal cell to ATP (100 ìÒ) in normal Hanks’ solution. B, dose—response ofCBF to varying concentrations of exogenously applied ATP (n = 12). CBF decreases with ATPconcentration, with an EC50 of approximately 50 ìÒ, and attains a maximal decrease at a concentration of100 ìÒ. Error bars are s.d.s. C, effect on CBF of ATP (100 ìÒ) and ATP-ã_S (100 ìÒ), a non-hydrolysableATP analogue, in the presence of 0·1 units ATPase (n = 6). In the presence of ATPase, ATP does not causea decrease in CBF; in contrast, application of ATP-ã_S results in a decrease in CBF. D, measurement ofependymal cell [Ca¥]c exposed to ATP (100 ìÒ) in Hanks’ solution. No changes in [Ca¥]c were observedwith ATP application.
Response to ATP
ATP has been demonstrated to function as an importantneurotransmitter in the CNS. Released ATP can eitherinteract with ATP receptors present in the CNS orparticipate in extracellular ecto-protein phosphorylationthat may be involved in critical neural signalling pathways,such as long-term potentiation (LTP) (White, 1984;Edwards & Gibb, 1993; Wieraszko, 1996; Robertson &Edwards, 1998; Queiroz et al. 1999). In the mammalianrespiratory and reproductive system, and in frog oesophagi,ATP has been shown to induce an increase in CBF that ismediated by an increase in [Ca¥]c (Nelson & Wright, 1974;Villalon et al. 1989; Korngreen & Priel, 1996; Evans &Sanderson, 1999). Therefore, we investigated the effect ofATP on intact ciliated brain ependymal cells whose meanbasal CBF was 22 ± 6 Hz (n = 12). It is important to notethat the mean CBF of this population was not significantlydifferent from that of the population exposed to 5-HT (seeabove). Exposure of intact ciliated brain ependymal cells toATP in Hanks’ solution resulted in a consistent andsustained decrease in CBF (Fig. 6A). The dose—responsecurve to ATP showed an EC50 of about 50 ìÒ and amaximal response at 100 ìÒ ATP resulting in a57·5 ± 6·5% (n = 12) decrease in CBF (Fig. 6B). In thepresence of ATPase (0·1 units), ATP (100 ìÒ) was unable toinduce a decrease in CBF (n = 6) (Fig. 6C). However,ATP_ã_S (100 ìÒ), a non-hydrolysable ATP analogue, wasstill able to decrease CBF in the presence of ATPase (n = 6)(Fig. 6C). Hence, the ATP-induced inhibition of ciliarybeating does not involve the hydrolysis of ATP and mostprobably is a purinergic receptor-mediated response.
The ATP-induced decrease in CBF was unaffected bylowering extracellular [Ca¥] to below 200 nÒ in theEGTA-buffered Hanks’ solution (n = 9) (data not shown).Further, we did not observe any changes in [Ca¥]cfollowing ATP stimulation of ependymal cells (n = 9)(Fig. 6D). These findings suggest that ATP-mediatedinhibition of CBF in ependymal cells is independent ofchanges in [Ca¥]c.
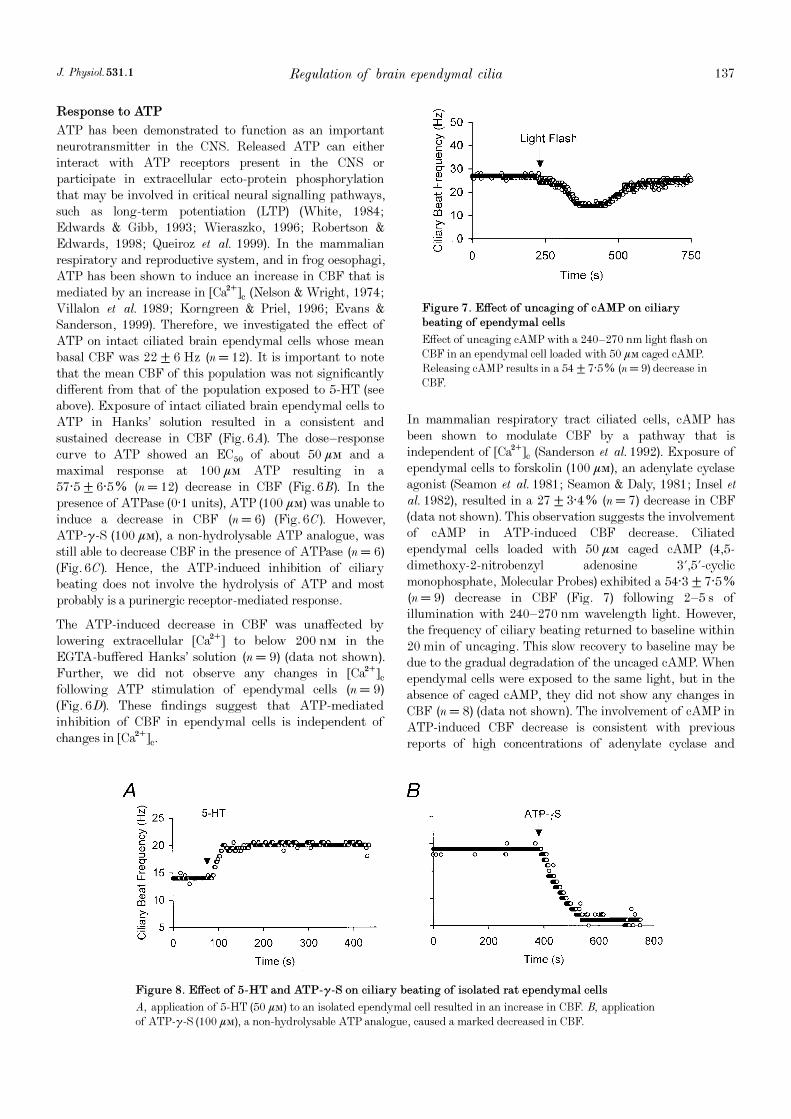
In mammalian respiratory tract ciliated cells, cAMP hasbeen shown to modulate CBF by a pathway that isindependent of [Ca¥]c (Sanderson et al. 1992). Exposure ofependymal cells to forskolin (100 ìÒ), an adenylate cyclaseagonist (Seamon et al. 1981; Seamon & Daly, 1981; Insel etal. 1982), resulted in a 27 ± 3·4% (n = 7) decrease in CBF(data not shown). This observation suggests the involvementof cAMP in ATP-induced CBF decrease. Ciliatedependymal cells loaded with 50 ìÒ caged cAMP (4,5-dimethoxy-2-nitrobenzyl adenosine 3',5'-cyclicmonophosphate, Molecular Probes) exhibited a 54·3 ± 7·5%(n = 9) decrease in CBF (Fig. 7) following 2—5 s ofillumination with 240—270 nm wavelength light. However,the frequency of ciliary beating returned to baseline within20 min of uncaging. This slow recovery to baseline may bedue to the gradual degradation of the uncaged cAMP. Whenependymal cells were exposed to the same light, but in theabsence of caged cAMP, they did not show any changes inCBF (n = 8) (data not shown). The involvement of cAMP inATP-induced CBF decrease is consistent with previousreports of high concentrations of adenylate cyclase and
Regulation of brain ependymal ciliaJ. Physiol. 531.1 137
Figure 7. Effect of uncaging of cAMP on ciliary
beating of ependymal cells
Effect of uncaging cAMP with a 240—270 nm light flash onCBF in an ependymal cell loaded with 50 ìÒ caged cAMP.Releasing cAMP results in a 54 ± 7·5% (n = 9) decrease inCBF.
Figure 8. Effect of 5-HT and ATP-ã_S on ciliary beating of isolated rat ependymal cells
A, application of 5-HT (50 ìÒ) to an isolated ependymal cell resulted in an increase in CBF. B, applicationof ATP_ã_S (100 ìÒ), a non-hydrolysable ATP analogue, caused a marked decreased in CBF.

pertussis toxin-sensitive G proteins on rat ependymal cellcilia (De Camilli et al. 1986; Shinohara et al. 1998).
We also investigated the possible interactions of the 5-HT-and ATP-stimulated pathways. At maximal concentrations,100 ìÒ ATP was able to overcome the stimulatory effect of100 ìÒ 5-HT and caused a 52·8 ± 8·2% (n = 6) decrease inCBF. Hence, cAMP decreases CBF either by a pathwayindependent of Ca¥ or by inhibition of a point downstreamof the Ca¥ regulatory cascade.
Response of isolated ciliated ependymal cells to 5-HT
and ATP
Lastly, we sought to provide evidence that 5-HT and ATPact directly on ependymal cells to increase and decreaseCBF, respectively. To do this we acutely isolated ependymalcells (see Methods) and investigated the effects of applicationof 5-HT and ATP-ã_S on CBF. Exposure of isolated ciliatedependymal cells to 5-HT (50 ìÒ) caused an increase in CBF(Fig. 8A). We observed that CBF increased, on average, by51·6 ± 14·4% (n = 3). Exposure of isolated ciliatedependymal cells to ATP-ã_S (100 ìÒ) caused a decrease inCBF (Fig. 8B). We observed that CBF decreased, onaverage, by 58·1 ± 10·4% (n = 4). Thus these resultsdemonstrate that 5-HT and ATP have a direct effect onciliated ependymal cells.
DISCUSSION
To our knowledge, this study is the first report to documentthe effects of 5-HT and ATP on ciliary beating ofmammalian brain ependymal cells and to identify twodistinct, independent pathways with opposing effects onCBF. Figure 9 presents a working model for these pathways.In the first pathway 5-HT increases ciliary beating byactivating a 5-HT receptor, probably a member of the5_HTµ receptor family located on the ependymal cell plasmamembrane, that triggers the generation of cytosolic IP×. Thebinding of IP× to its receptor on IP×-sensitive Ca¥ storesresults in release of Ca¥ to the cytosol. This initial rise in[Ca¥]c opens CRAC channels on the plasma membrane,leading to a more sustained Ca¥ influx from theextracellular environment. The sustained elevation in[Ca¥]c is responsible for most of the 5-HT-stimulatedincrease in CBF. In contrast in the second pathway, ATPbinding, probably to a purinergic receptor, possiblyactivates adenylate cyclase that generates cAMP that theninhibits ciliary beating. cAMP works by either inhibiting apoint downstream of the Ca¥ regulatory cascade or apathway independent of Ca¥. However, the exact nature bywhich cAMP and [Ca¥]c ultimately change ciliarymovement remains unclear.
T. Nguyen, W.-C. Chin, J. A. O’Brien, P. Verdugo and A. J. Berger J. Physiol. 531.1138
Figure 9. A model of the signalling pathways involved in the effect of 5-HT and ATP on ciliary
beating of ependymal cells
Exposure to 5-HT results in the intracellular production of IP×, which releases Ca¥ from intracellular Ca¥stores. The released Ca¥ mediates the opening of CRAC channels that results in the influx of Ca¥ from theextracellular environment. The increase in [Ca¥]c ultimately increases CBF by an as yet unidentifiedmechanism. In contrast, ATP activates the production of intracellular cAMP, which inhibits CBF by a stillto be determined mechanism.

While it is conceivable that the results observed in our sliceculture experiments may involve not only ependymal cellsbut other cell types, we think this is unlikely. The fact thatwe observed the same effects of 5-HT and ATP on CBF inacutely isolated ciliated ependymal cells supports the modelproposed in Fig. 9.
The 5-HT-mediated increase in CBF may have importantfunctional implications. 5-HT is released into the CSF fromthe ependymal surface (Ternaux et al. 1977). Stimulation ofthe brainstem raphe nuclei, the location of the cell bodiesthat are the source of the 5-HT-containing nerve plexus ofthe ependymal layer (Chan-Paly, 1976), causes increasedrelease of 5-HT into the CSF (Sheard & Zolovick, 1971).Thus the 5-HT-mediated increase in CBF may provide amechanism for the wider distribution of this importantneurotransmitter, whereby the CSF may be suited as acorridor for distributing neuronal messengers. This isanalogous in the endocrine system to utilizing blood todistribute hormones. Finally, it is of interest that patientswith immotile cilia syndrome (primary ciliary dyskinesia)have a number of neurological deficits including headache,depression and schizophrenia (Afzelius & Mossberg, 1995),all of which have been in part associated with alterations inthe brain’s serotonergic system (Johnson et al. 1998;Peroutka, 1998).
Although the specific roles of ciliated ependymal cells in theCNS remain to be elucidated (Cathcart & Worthington,1964; Roth et al. 1985; Del Bigio, 1995), here we haveshown that these ciliated ependymal cells responded to 5-HT and ATP with opposing effects. By increasing theirCBF, ciliated ependymal cells can preferentially disperse ortransmit certain neural messengers into the bulk CSF andsubstantially spread these molecules to other regions ofCNS, thereby initiating a more widely distributed signal.Conversely, by reducing the CBF, ciliated ependymal cellscan also selectively retain or localize the dispersion ofcertain messengers in CSF, generating a short-rangeinfluence. These results provide compelling evidence thatependymal cells can play an active role in neural messengerexchange at the CSF—brain interface.
Afzelius, B. A. & Mossberg, B. (1995). Immotile-cilia syndrome(primary ciliary dyskinesia), including Kartagener syndrome. InThe Metabolic and Molecular Bases of Inherited Disease,ed. Scriver, C. R., Beaudet, A. L., Sly, W. S. & Valle, D.,pp. 3943—3954. McGraw-Hill, New York.
Ben-Shimol, Y., Dinstein, I., Meisels, A. & Priel, Z. (1991).Ciliary motion features from digitized video photography. Journalof Computer-Assisted Microscopy 3, 103—116.
B�ohm, S. K., Khitin, L. M., Grady, E. F., Aponte, G., Payan, D. G.& Bunnett, N. W. (1996). Mechanisms of desensitization andresensitization of proteinase-activated receptor-2. Journal ofBiological Chemistry 271, 22003—22016.
Cathcart, R. S. & Worthington, W. C. (1964). Ciliary movement inthe rat cerebral ventricles: clearing action and directions of currents.Journal of Neuropathology and Experimental Neurology 23,609—618.
Chan-Paly, V. (1976). Serotonin axons in the supra- andsubependymal plexuses and in the leptomeninges; their roles inlocal alterations of cerebrospinal fluid and vasomotor activity. BrainResearch 102, 103—130.
Cooper, J. R., Bloom, F. E. & Roth, R. H. (1996). The BiochemicalBasis of Neuropharmacology, 7th edn. Oxford University Press,New York.
De Camilli, P., Moretti, M., Donini, S. D., Walter, U. &Lohmann, S. M. (1986). Heterogeneous distribution of the cAMPreceptor protein RII in the nervous system: evidence for its intra-cellular accumulation on microtubules, microtubule-organizingcenters, and in the area of the Golgi complex. Journal of CellBiology 103, 189—203.
Del Bigio, M. R. (1995). The ependyma: a protective barrier betweenbrain and cerebrospinal fluid. Glia 14, 1—13.
Dinopoulos, A. & Dori, I. (1995). The development of theserotonergic fiber network of the lateral ventricles of the rat brain: alight and electron microscopic immunocytochemical analysis.Experimental Neurology 133, 73—84.
Edwards, F. A. & Gibb, A. J. (1993). ATP — a fast neurotransmitter.Federation of European Biochemical Societies 325, 86—89.
Evans, J. H. & Sanderson, M. J. (1999). Intracellular calciumoscillations regulate ciliary beat frequency of airway epithelial cells.Cell Calcium 26, 103—110.
Franzius, D., Hoth, M. & Penner, R. (1994). Non-specific effects ofcalcium entry antagonists in mast cells. Pfl�ugers Archiv 428,433—438.
Gafni, J., Munsch, J. A., Lam, T. H., Catlin, M. C., Costa, L. G.,Molinski, T. F. & Pessah, I. N. (1997). Xestospongins: potentmembrane permeable blockers of the inositol 1,4,5-trisphosphatereceptor. Neuron 19, 723—733.
Insel, P. A., Stengel, D., Ferry, N. & Hanoune, J. (1982).Regulation of adenylate cyclase of human platelet membranes byforskolin. Journal of Biological Chemistry 257, 7485—7490.
Johansson, C. B., Momma, S., Clarke, D. L., Risling, M.,Lendahl, U. & Fris�en, J. (1999). Identification of a neural stemcell in the adult mammalian central nervous system. Cell 96, 25—34.
Johnson, K. W., Phebus, L. A. & Cohen, M. L. (1998). Serotonin inmigraine: theories, animal models and emerging therapies. Progressin Drug Research 51, 219—244.
Kao, J. P. Y. (1994). Practical aspects of measuring [Ca¥] withfluorescent indicators. In Methods in Cell Biology, ed. Nuccitelli,R., pp. 155—181. Academic Press, San Diego.
Korngreen, A. & Priel, Z. (1996). Purinergic stimulation of rabbitciliated airway epithelia: control by multiple calcium sources.Journal of Physiology 497, 53—66.
Lorez, H. P. & Richards, J. G. (1982). Supra-ependymalserotoninergic nerves in mammalian brain: morphological,pharmacological and functional studies. Brain Research Bulletin 9,727—741.
Monck, J. R., Oberhauser, A. F., Keating, T. J. & Fernandez,J. M. (1992). Thin-section ratiometric Ca¥ images obtained byoptical sectioning of fura_2 loaded mast cells. Journal of CellBiology 116, 745—759.
Regulation of brain ependymal ciliaJ. Physiol. 531.1 139

Nelson, D. J. & Wright, E. M. (1974). The distribution, activity, andfunction of the cilia in the frog brain. Journal of Physiology 243,63—78.
Nguyen, T. (1996). Dynamics of cytosolic and compartmentalizedCa¥ during signal transduction. PhD Thesis, University ofWashington, Seattle, USA.
Nguyen, T., Chin, W.-C. & Verdugo, P. (1998). Role of Ca¥ÏK¤ ionexchange in intracellular storage and release of Ca¥. Nature 395,908—912.
Nicholson, C. (1999). Signals that go with the flow. Trends inNeurosciences 22, 143—145.
Parekh, A. B. & Penner, R. (1997). Store depletion and calciuminflux. Physiological Reviews 77, 901—930.
Perez-Martin, M., Grondona, J. M., Cifuentes, M., Perez-Figares, J. M., Jimenez, J. A. & Fernandez-Llebrez, P. (2000).Ependymal explants from the lateral ventricle of the adult bovinebrain: a model system for morphological and functional studies ofthe ependyma. Cell Tissue Research 300, 11—19.
Peroutka, S. J. (1998). Serotonin receptor variants in disease: newtherapeutic opportunities? Annals of the New York Academy ofSciences 861, 16—25.
Queiroz, G., Meyer, D. K., Meyer, A., Starke, K. & vonK�ugelgen, I. (1999). A study of the mechanism of the release ofATP from rat cortical astroglial cells evoked by activation ofglutamate receptors. Neuroscience 91, 1171—1181.
Robertson, S. J. & Edwards, F. A. (1998). ATP and glutamate arereleased from separate neurones in the rat medial bavenula nucleus:frequency dependence and adenosine-mediated inhibition of release.Journal of Physiology 508, 691—701.
Roth, Y., Kimhi, Y., Edery, H., Aharonson, E. & Priel, Z. (1985).Ciliary motility in brain ventricular system and trachea ofhamsters. Brain Research 330, 291—297.
Sanderson, M. J. & Dirksen, E. R. (1995). Quantification of ciliarybeat frequency and metachrony by high-speed digital video. Methodsin Cell Biology 47, 289—297.
Sanderson, M. J., Lansley, A. B. & Dirksen, E. R. (1992).Regulation of ciliary beat frequency in respiratory tract cells. Chest101, 69S—71S.
Seamon, K. B. & Daly, J. W. (1981). Activation of adenylate cyclaseby the diterpene forskolin does not require the guanine nucleotideregulatory protein. Journal of Biological Chemistry 256,9799—9801.
Seamon, K. B., Padgett, W. & Daly, J. W. (1981). Forskolin: uniquediterpene activator of adenylate cyclase in membranes and in intactcells. Proceedings of the National Academy of Sciences of the USA78, 3363—3367.
Sheard, M. H. & Zolovick, A. J. (1971). Serotonin: release in catbrain and cerebrospinal fluid on stimulation of midbrain raph�e.Brain Research 26, 455—458.
Shinohara, H., Asano, T., Kato, K., Kameshima, T. & Semba, R.(1998). Localization of a G protein Gi2 in the cilia of rat ependyma,oviduct and trachea. European Journal of Neuroscience 10,699—707.
Terasaki, M., Song, J., Wong, J. R., Weiss, M. J. & Chen, L. B.(1984). Localization of endoplasmic reticulum in living andglutaraldehyde-fixed cells with fluorescent dyes. Cell 38, 101—108.
Ternaux, J. P., Hery, F., Hamon, M., Bourgoin, S. & Glowinski, J.(1977). 5-HT release from ependymal surface of the caudate nucleusin ‘enc�ephale isol�e’ cats. Brain Research 132, 575—579.
Villalon, M., Hinds, T. R. & Verdugo, P. (1989). Stimulus-responsecoupling in mammalian ciliated cells. Demonstration of twomechanisms of control for cytosolic Ca¥. Biophysical Journal 56,1255—1258.
Voutsinos, B., Chouaf, L., Mertens, P., Ruiz-Flandes, P.,Joubert, Y., Belin, M. F. & Didier-Bazes, M. (1994). Tropism ofserotonergic neurons towards glial targets in the rat ependyma.Neuroscience 59, 663—672.
White, T. D. (1984). Characteristics of neuronal release of ATP.Progress in Neuro-Psychopharmacology and Biological Psychiatry 8,487—493.
Wieraszko, A. (1996). Extracellular ATP as a neurotransmitter: itsrole in synaptic plasticity in the hippocampus. Acta NeurobiologiaeExperimentalis 56, 637—648.
Zifa, E. & Fillion, G. (1992). 5-Hydroxytryptamine receptors.Pharmacological Reviews 44, 401—458.
Acknowledgements
We thank Dr D. A. Bayliss, University of Virginia, for his adviceon the 5-HT immunocytochemistry, and J. Steed and P. Huynh fortechnical assistance. This work was supported by grants from theNSF, NIH, DOE and Mellon Foundation. T. Nguyen wassupported by training grants from the MSTP program and InternalMedicine residency program at University of Washington School ofMedicine. J. A. O’Brien was supported by an NIH training grant.W.-C. Chin was supported by a fellowship from the Centre forNanotechnology at the University of Washington.
Corresponding author
A. J. Berger: Department of Physiology & Biophysics, School ofMedicine, University of Washington, Box 357290, Seattle,WA 98195-7290, USA.
Email: [email protected]
T. Nguyen, W.-C. Chin, J. A. O’Brien, P. Verdugo and A. J. Berger J. Physiol. 531.1140
Copyright © 2022 FDOKUMEN




















