
Faunal migration into the Late Permian Zechstein Basin – Evidence from bryozoan palaeobiogeography
12
Faunal migration into the Late Permian Zechstein Basin – Evidence from bryozoan palaeobiogeography Anne M. Sørensen a , Eckart Håkansson a , Lars Stemmerik a,b, ⁎ a Department of Geography and Geology, University of Copenhagen, Øster Voldgade 10, DK-1350 Copenhagen K, Denmark b Geological Survey of Denmark and Greenland, Øster Voldgade 10, DK-1350 Copenhagen K, Denmark Received 24 October 2006; received in revised form 8 March 2007; accepted 19 March 2007 Abstract Late Permian bryozoans from the Wegener Halvø, Ravnefjeld and Schuchert Formations in East Greenland have been investigated. 14 genera are recognised. Integration of the new bryozoan data from the Upper Permian of East Greenland with data on the distribution of Permian bryozoans along the northern margin of Pangea is used to test hypotheses concerning Late Palaeozoic evolution of the North Atlantic region. During the Permian, the Atlantic rift system formed a seaway between Norway and Greenland from the boreal Barents Shelf to the warm and arid Zechstein Basin. This seaway is considered to be the only marine connection to the Zechstein Basin and therefore the only possible migration route for bryozoans to enter the basin. The distribution of Permian bryozoans is largely in keeping with such a connection from the cool Barents Shelf past the East Greenland Basin to the warm Zechstein Basin and also corroborates the change in temperature through this connection. © 2007 Elsevier B.V. All rights reserved. Keywords: Bryozoans; East Greenland; Palaeobiogeography; Permian; Zechstein Basin 1. Introduction The Upper Palaeozoic successions of the Sverdrup Basin of Arctic Canada and the North Greenland– Svalbard–Norwegian Barents Sea area along the northern margin of Pangaea record overall changes in palaeoclimatic conditions from subtropical and arid in the Late Carbon- iferous to temperate in the Late Permian (Beauchamp, 1994; Beauchamp and Desrochers, 1997; Stemmerik, 1997, 2000; Beauchamp and Baud, 2003; Stemmerik and Worsley, 2005). This is in concert with global palaeogeo- graphic reconstructions indicating that Pangaea drifted more than 2500 km northward during this time period, so that the North Greenland–Norwegian Barents Sea area moved from c. 20°N latitude in the mid-Carboniferous to c. 45°N latitude in the Late Permian (Fig. 1; Golonka, 2000; Scotese, 2004). The subtropical northern Pangean shelf was connected south-eastwards to the Tethys Ocean during Late Carboniferous–Early Permian time, but collision between Laurussia and Siberia during later Permian times apparently disrupted this connection and led to develop- ment of more distinctive Late Permian faunal provinces (Fig. 1; Henderson and Mei, 2000). The Late Permian Zechstein Basin of NW Europe is a marine, partly evaporitic intracratonic basin located at c. Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198 – 209 www.elsevier.com/locate/palaeo ⁎ Corresponding author. Present address: Department of Geography and Geology, University of Copenhagen, Øster Voldgade 10, DK-1350 Copenhagen K, Denmark. E-mail address: [email protected] (L. Stemmerik). 0031-0182/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2007.03.045
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Faunal migration into the Late Permian Zechstein Basin – Evidence from bryozoan palaeobiogeography

alaeoecology 251 (2007) 198–209www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology, P
Faunal migration into the Late Permian Zechstein Basin – Evidencefrom bryozoan palaeobiogeography
Anne M. Sørensen a, Eckart Håkansson a, Lars Stemmerik a,b,⁎
a Department of Geography and Geology, University of Copenhagen, Øster Voldgade 10, DK-1350 Copenhagen K, Denmarkb Geological Survey of Denmark and Greenland, Øster Voldgade 10, DK-1350 Copenhagen K, Denmark
Received 24 October 2006; received in revised form 8 March 2007; accepted 19 March 2007
Abstract
Late Permian bryozoans from the Wegener Halvø, Ravnefjeld and Schuchert Formations in East Greenland have beeninvestigated. 14 genera are recognised.
Integration of the new bryozoan data from the Upper Permian of East Greenland with data on the distribution of Permianbryozoans along the northern margin of Pangea is used to test hypotheses concerning Late Palaeozoic evolution of the NorthAtlantic region. During the Permian, the Atlantic rift system formed a seaway between Norway and Greenland from the borealBarents Shelf to the warm and arid Zechstein Basin. This seaway is considered to be the only marine connection to the ZechsteinBasin and therefore the only possible migration route for bryozoans to enter the basin. The distribution of Permian bryozoans islargely in keeping with such a connection from the cool Barents Shelf past the East Greenland Basin to the warm Zechstein Basinand also corroborates the change in temperature through this connection.© 2007 Elsevier B.V. All rights reserved.
Keywords: Bryozoans; East Greenland; Palaeobiogeography; Permian; Zechstein Basin
1. Introduction
The Upper Palaeozoic successions of the SverdrupBasin of Arctic Canada and the North Greenland–Svalbard–Norwegian Barents Sea area along the northernmargin of Pangaea record overall changes in palaeoclimaticconditions from subtropical and arid in the Late Carbon-iferous to temperate in the Late Permian (Beauchamp,1994; Beauchamp and Desrochers, 1997; Stemmerik,1997, 2000; Beauchamp and Baud, 2003; Stemmerik and
⁎ Corresponding author. Present address: Department of Geographyand Geology, University of Copenhagen, Øster Voldgade 10, DK-1350Copenhagen K, Denmark.
E-mail address: [email protected] (L. Stemmerik).
0031-0182/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.palaeo.2007.03.045
Worsley, 2005). This is in concert with global palaeogeo-graphic reconstructions indicating that Pangaea driftedmore than 2500 km northward during this time period, sothat the North Greenland–Norwegian Barents Sea areamoved from c. 20°N latitude in themid-Carboniferous to c.45°N latitude in the Late Permian (Fig. 1; Golonka, 2000;Scotese, 2004). The subtropical northern Pangean shelfwas connected south-eastwards to the Tethys Ocean duringLate Carboniferous–Early Permian time, but collisionbetween Laurussia and Siberia during later Permian timesapparently disrupted this connection and led to develop-ment of more distinctive Late Permian faunal provinces(Fig. 1; Henderson and Mei, 2000).
The Late Permian Zechstein Basin of NW Europe is amarine, partly evaporitic intracratonic basin located at c.

Fig. 1. Late Permian global reconstruction showing the position of the Zechstein Basin (Z), Greenland (G) and Svalbard, and conodont faunaprovinces. Based on Golonka (2000) and Henderson and Mei (2000).
199A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
20°N palaeolatitude immediately west of the TethysOcean (Fig. 1). The lower part of the depositional suc-cession is dominated by open marine platform carbo-nates and reefs with a rather endemic fauna withlittle in common with the Tethyan fauna of the time-equivalent successions in the Austrian and Italian Alps(e.g. Hollingworth and Tucker, 1987; Ernst, 2001a).This has led to the common belief that the ZechsteinBasin was semi-enclosed, separated by a land barrierfrom the Tethys Ocean and connected northwards to theArctic Boreal Ocean via the rifted seaway betweenGreenland and Norway (Fig. 1). To test and furthersubstantiate this hypothesis, we have studied the
Fig. 2. Correlation of Permian lithostratigraphic units in Spitsbergen, Bjørnøyfrom Stemmerik (2000).
bryozoan fauna of the Upper Permian carbonates ofcentral East Greenland, located at a palaeolatitude of c.32°–35°N, midway between the Boreal cool-watercarbonate platforms of North Greenland–NorwegianBarents Sea and the warm and arid areas of theZechstein basin in NW Europe (Stemmerik, 2001).The sediments and associated fauna form an importantlink between the Late Permian cool- and warm-watercarbonate realms, and the East Greenland bryozoanscombined with published bryozoan data from NorthGreenland, Svalbard and the western Tethys providebetter understanding of Late Permian biogeography andthe faunal migration into the Zechstein Basin.
a, North Greenland, East Greenland and the Zechstein Basin. Modified

200 A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
2. Permian stratigraphy and facies
Marine Permian deposits are known from outcropsand cores in NW Europe, East Greenland, North Green-land, Svalbard and the offshore areas of the BarentsShelf and the mid-Norwegian shelf (Stemmerik, 2000).Lower Permian marine strata are confined to the NorthGreenland–Norwegian Barents Sea area in the northwhereas marine Upper Permian sediments are knownfrom the entire region.
Fig. 3. Asselian–Kazanian palaeogeographic reconstructions of the westecomposition of the bryozoan fauna. For numbers, refer to Table 1. Green: lanbetween the Tethys and the Russian Platform from (A) Asselian to (B) Artinsand Norway during (C) Kungurian–(D) Kazanian time. Palaeogeography m1990), Malecki (1968, 1977), Southwood (1985, 1990), Madsen and Håka(1992), Madsen (1994) and Ernst (2000, 2001a, 2003) and our own data in Easlegend, the reader is referred to the web version of this article.)
The Permian succession in the Svalbard–NorwegianBarents Sea area belongs to three groups which reflect thechanges in palaeoclimate over time (Fig. 2; Stemmerik,1997; Larssen et al., 2005; Stemmerik and Worsley,2005). The Asselian–Sakmarian part of the GipsdalenGroup consists mostly of platform carbonates with adiverse chloro-foramol biota typical of Permian warm-water shelf areas (see Beauchamp, 1994). The carbonateplatforms surround deep halite basins thus implyingoverall warm and arid conditions during Early Permian
rn Tethys, Russian Platform, NW Europe and Greenland showingd; light blue: shelves; dark blue: oceanic. Note the closure of the straitkian time, and the stepwise opening of the seaway between Greenlandodified from Golonka (2000). Fauna data from Ross and Ross (1962,nsson (1989), Nakrem (1991, 1993, 1994a,b,c, 2004), Nakrem et al.t Greenland. (For interpretation of the references to colour in this figure

Fig. 3 (continued ).
201A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
times (Stemmerik, 2000; Stemmerik and Worsley, 2005).Dating of this part of the succession is based on fusulinids,and correlation is firmly established internally in theregion, including North Greenland, and to the Interna-tional zonation (Fig. 2; Dallmann et al., 1999; Stemmerik,2000; Larssen et al., 2005; Stemmerik andWorsley, 2005;and references therein). The Lower Permian (upperSakmarian–Artinskian (lowermostKungurian?)) Bjarme-land Group is dominated by temperate cool-watercarbonates with a bryozoan–brachiopod–echinodermdominated fauna (Stemmerik, 1997). Dating is based onlocal fusulinid assemblages, conodonts and small for-aminifers, and correlation to North Greenland andadjacent areas is generally well established (see Stem-merik, 2000).
The Lower–Upper Permian (Kungurian–Wuchiapin-gian/Changhsingian?) TempelfjordenGroup is dominatedby deeper water spiculites, shales and local sandstonesand cool-water carbonates. Dating is based on smallforaminifers, palynomorphs and conodonts, and is lesswell established than for the older Permian succession(Stemmerik, 2000; Larssen et al., 2005). In NorthGreenland, the time-equivalent upper Kim Fjelde andMidnatfjeld formations are composed of cool-water car-bonates in the platform areas and shales and spiculites inmore distal settings (Stemmerik, 1997). Correlation to themore southerly located Permian strata of central EastGreenland and the Zechstein basin is still disputed. TheWegener Halvø and Ravnefjeld formations of central EastGreenland are of Kazanian (Wuchiapingian) age based on

Fig. 3 (continued ).
202 A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
palynomorphs and conodonts and correlate, based onmicrofloral evidence with the Midnatfjeld Formation ofNorth Greenland and the uppermost TempelfjordenGroup of the Norwegian Barents Shelf, and, based onconodonts with the (?) lower part of the Zechsteinsuccession (Fig. 2; Rasmussen et al., 1990; Mangerud,1994; Utting and Piasecki, 1995; Stemmerik et al., 1996;Henderson and Mei, 2000). The overlying Schuchert DalFormation is ofChangsingian age andmay correlate to theuppermost Zechstein of NW Europe (Fig. 2).
3. Material and methods
The investigated material from East Greenland wasselected among a large collection of samples collected
during several expeditions to East Greenland and wassimply chosen with respect to density and preservationof the bryozoans. It represents both the Wegener Halvøand Schuchert Dal formations and thus spans most ofthe Late Permian. All bryozoans are embedded in matrixbut some samples have bedding planes with large andwell-preserved colonies which were used for externalinvestigations. The internal characteristics were studiedin acetate peels made from polished surfaces.
Data in Ross and Ross (1962, 1990), Malecki (1968,1977), Southwood (1985, 1990), Madsen and Håkans-son (1989), Nakrem (1991, 1993, 1994a,b,c, 2004),Nakrem et al. (1992), Madsen (1994) and Ernst (2000,2001a, 2003) on Permian bryozoans from Svalbard,North Greenland, the Russian Platform, the Zechstein
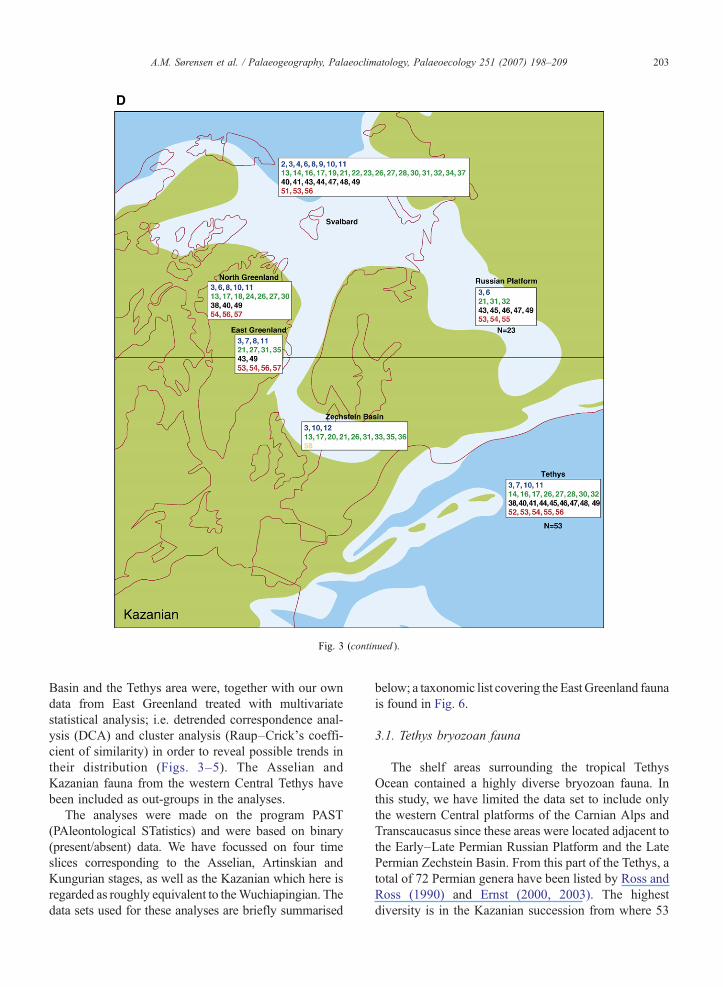
Fig. 3 (continued ).
203A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
Basin and the Tethys area were, together with our owndata from East Greenland treated with multivariatestatistical analysis; i.e. detrended correspondence anal-ysis (DCA) and cluster analysis (Raup–Crick's coeffi-cient of similarity) in order to reveal possible trends intheir distribution (Figs. 3–5). The Asselian andKazanian fauna from the western Central Tethys havebeen included as out-groups in the analyses.
The analyses were made on the program PAST(PAleontological STatistics) and were based on binary(present/absent) data. We have focussed on four timeslices corresponding to the Asselian, Artinskian andKungurian stages, as well as the Kazanian which here isregarded as roughly equivalent to theWuchiapingian. Thedata sets used for these analyses are briefly summarised
below; a taxonomic list covering the East Greenland faunais found in Fig. 6.
3.1. Tethys bryozoan fauna
The shelf areas surrounding the tropical TethysOcean contained a highly diverse bryozoan fauna. Inthis study, we have limited the data set to include onlythe western Central platforms of the Carnian Alps andTranscaucasus since these areas were located adjacent tothe Early–Late Permian Russian Platform and the LatePermian Zechstein Basin. From this part of the Tethys, atotal of 72 Permian genera have been listed by Ross andRoss (1990) and Ernst (2000, 2003). The highestdiversity is in the Kazanian succession from where 53

Fig. 4. Cluster analysis of the Permian bryozoan fauna in the study area based on data in Ross and Ross (1962, 1990), Malecki (1968, 1977),Southwood (1985, 1990), Madsen and Håkansson (1989), Nakrem (1991, 1993, 1994b,c, 2004), Nakrem et al. (1992), Madsen (1994) and Ernst(2000, 2001a, 2003) and our own data from East Greenland. For colour codes, see Fig. 5. (For interpretation of the references to colour in this figurelegend, the reader is referred to the web version of this article.)
204 A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
genera have been identified compared to 28 generain the Kungurian, 39 genera in the Artinskian and 29genera in the Asselian. Taxa in common with Svalbard,Greenland, the Russian Platform and the ZechsteinBasin are listed in Fig. 3; for complete fauna lists consultRoss and Ross (1990) and Ernst (2000, 2003).
The Early Permian Tethys fauna shows great simi-larities with that of the Russian Platform whereas theLate Permian fauna is related more to those found inAsia and Australia (Ernst 2000). In the multivariatestatistical analysis, we have focussed on the Asselianand the Kazanian faunas to illustrate connections to theadjacent areas (Figs. 4 and 5).
3.2. Russian platform bryozoan fauna
Data on Permian bryozoan distribution on the RussianPlatform are taken from Ross and Ross (1990) whorecorded a total of 53 Permian genera. They report 34
genera in the Asselian, 25 in the Artinskian, 17 in theKungurian and 23 in the Kazanian. Taxa in commonwith Svalbard, Greenland, the western Tethys and theZechstein Basin are listed in Fig. 3; for a complete faunalist consult Ross and Ross (1990).
3.3. Svalbard bryozoan fauna
The Permian bryozoan fauna from Svalbard was de-scribed and documented by Nakrem (1991, 1993, 1994a,b,c, 2004), Nakrem et al. (1992) and Malecki (1968,1977), subsequently revised by Nakrem (1988). Thedescribed faunas represent both the Asselian–Sakmarianwarm-water carbonate succession and the cool-watercarbonates of the Kungurian–Kazanian TempelfjodenGroup.
A total of 50 genera have been described from thePermian of Svalbard. The fauna of the Asselian suc-cession, represented by the Tyrellfjellet Member and the

Fig. 5. DCA analysis of the Permian bryozoan fauna in the study areabased on data in Ross and Ross (1962, 1990), Malecki (1968, 1977),Southwood (1985, 1990), Madsen and Håkansson (1989), Nakrem(1991, 1993, 1994a,b,c, 2004), Nakrem et al. (1992), Madsen (1994)and Ernst (2000, 2001a, 2003) and our own data from East Greenland.(For interpretation of the references to colour in this figure legend, thereader is referred to the web version of this article.)
Fig. 6. Distribution of the Kazanian bryozoan taxa in the NorthAtlantic realm. It is assumed that genera occurring both in the northernarea and in the Zechstein Basin also are present in East Greenland buthave yet to be found.
205A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
Kapp Duner Formation, comprises 21 genera with adominance of robust ramose colonies of Ascoporellaand Tabulipora together with delicate fenestrate colo-nies and fine branched Ascopora and Rhabdomesoncolonies (Fig. 3). This fauna has a general affinity tobryozoan faunas from Russia (Nakrem, 1994b). TheArtinskian fauna, represented by the upper ArtinskianVøringen Member and Hambergfjelet Formation, com-prises 27 genera and is dominated by encrusting androbust trepostomes and cystoporates and by robust fe-nestrate colonies in coarser beds. The fauna is regardedas Boreal and has affinities to Artinskian reef faunasfrom the Urals (Nakrem, 1994c). The Kungurian–Kazanian bryozoan faunas are the most diverse of thePermian faunas reported from Svalbard with 35 and 34genera, respectively (Fig. 3). The faunas are dominatedby delicate, ramose Rhombotrypella, Dyscritella andStreblascopora together with delicate fenestrate colo-nies (Table 1).
3.4. North Greenland bryozoan fauna
The bryozoan faunas described and documented byRoss and Ross (1962), Madsen and Håkansson (1989)and Madsen (1994) from the Permian of North Green-land mainly come from the upper Artinskian–Kungur-ian Kim Fjelde Formation with some additionalinformation from the Kazanian Midnatfjeld Formation.
The fauna thus represents a cool-water carbonate suc-cession of the same age as the Tempelfjoden Group ofSvalbard (Fig. 2). So far 22 genera have been recognizedfrom North Greenland, but the bryozoan fauna has not

Table 1Permian bryozoan genera from the study area
Trepostomata Fenestrata Cryptostomata Cystoporata Cyclostomata
1 Amphiporella 13 Acanthocladia 38 Ascopora 50 Coscinium 58 Corynotrypa2 Anisotrypella 14 Alternifenestella 39 Ascoporella 51 Cyclotrypa3 Dyscritella 15 Archimedes 40 Clausotrypa 52 Eridopora4 Dyscritellina 16 Fabifenestella 41 Gilmoropora 53 Fistulipora5 Hinaclema 17 Fenestella 42 Girtyporina 54 Goniocladia6 Neoeridotrypella 18 Fenestrellina 43 Permoheloclema 55 Meekopora7 Paralioclema 19 Flexifenestella 44 Primorella 56 Ramipora8 Rhombotrypella 20 Kalvariella 45 Rhabdomeson 57 Ramiporidra9 Stellahexaformis 21 Kingopora 46 Rhombopora10 Stenopora 22 Laxifenestella 47 Streblascopora11 Tabulipora 23 Lyrocladia 48 Streblotrypa12 Ulrichotrypa 24 Lyropora 49 Timanodictya
25 Lyroporella26 Penniretepora27 Polypora28 Polyporella29 Protoretepora30 Ptylopora31 Rectifenestella32 Reteporidra33 Ryhopora34 Septopora35 Spinofenestella36 Synocladia37 Wjatkella
For numbers, refer to Fig. 3. Fauna data from Ross and Ross (1962, 1990), Malecki (1968, 1977), Southwood (1985, 1990), Madsen and Håkansson(1989), Nakrem (1991, 1993, 1994a,b,c, 2004), Nakrem et al. (1992), Madsen (1994) and Ernst (2000, 2001a, 2003) and our own data in EastGreenland.
206 A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
been investigated in the same detail as that in Svalbardand the Zechstein Basin.
The fauna of the upper Artinskian part of the KimFjelde Formation comprises 17 genera, of which 13genera also are known from the time equivalent suc-cession in Svalbard (Fig. 3). The Kungurian fauna isslightly more diverse, including two more cryptostomegenera and one more cystoporate genus (Fig. 3). The KimFjelde Formation is dominated by Tabulipora, further-more Rhombotrypella, Dyscritella and Amphiporella arenumerous. The fauna from the Kazanian MidnatfjeldFormation includes 18 genera, of which 13 are knownfrom Svalbard and 7 are from East Greenland (Figs. 3and 6). The formation is dominated by Tabulipora,Fenestella, Polypora and Ramipora.
3.5. Zechstein bryozoan fauna
The bryozoan fauna from the Zechstein Basin hasbeen described and documented by Southwood (1985,1990) and Ernst (2001a). The bryozoan fauna includes13 genera that are primarily described from the Germanand English part of the basin. The fauna completelylacks members of the Cryptostomata and Cystoporata,
and includes the only confirmed Cyclostomata in thestudy area (Fig. 3). The bryozoans are important reefbuilders and are associated with typical warm-watercarbonates, like oolites, and evaporites.
One genus Ryhopora is endemic to the ZechsteinBasin, five genera are known from East Greenland, fivefrom North Greenland and seven genera are in commonwith Svalbard. 5 of the 13Zechstein genera are also knownfrom Kazanian-age sediments in the western TethysSea. Four genera Dyscritella, Stenopora, Fenestella andPenniretepora have a very wide geographical distributionbeing known from Svalbard, Greenland, Zechstein Basinand western Tethys (Figs. 3 and 6). Five of the Zechsteingenera have not been reported from any of the immediateadjacent areas.
3.6. East Greenland bryozoan fauna
The East Greenland bryozoan fauna occur in carbon-ates of mixed cool- and warm-water affinity midwaybetween the Late Permian Arctic Boreal Ocean of NorthGreenland, Svalbard and the Barents Sea and the lowlatitude Zechstein Basin (Stemmerik, 2001). Bryozoansare important reef builders in the Kazanian Wegener

207A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
Halvø Formation where they occur associated withtypical warm-water carbonate facies like oolites.
14 genera have been described and in addition anumber of fenestellid taxa related to the genus Fenestellahave been found but not identified. The fauna is domi-nated by the genera Tabulipora followed in frequency byRamipora. Dyscritella, Permoheloclema, Polyporaand the colonies related to Fenestella are also common.Tabulipora also dominates in the time-equivalentMidnatfjeld Formation of North Greenland where alsoRamipora, Polypora and Fenestella are common, anddominance of Tabulipora and Ramipora also charac-terises the Svalbard fauna.
13 of the East Greenland genera are also known fromthe Permian successions of Svalbard and 5 genera are incommon with the Zechstein succession.
A taxonomic description of the East Greenland fauna isunder preparation (Sørensen, Håkansson and Stemmerik).
4. Discussion
The cluster analysis based on Raup-Crick's coefficientof similarity shows that the Late Permian (Kazanian)western Tethys bryozoan fauna has little in common withthe Early Permian (Asselian) western Tethys fauna(similarity index close to 0.6), and that the two Tethysfaunas deviate significantly (b0.25) from all otherinvestigated faunas (Fig. 4). The Permian bryozoanfaunas in Svalbard, North Greenland and East Greenlandshow high levels of similarity (all N0.95) and as a groupthey have a similarity index at the 0.7 level to theZechstein Basin bryozoan fauna (Fig. 4). The EarlyPermian faunas of the Russian Platform are essentiallyidentical (N0.99), whereas the Kazanian fauna differssignificantly from all other investigated faunas (Fig. 4).The combination of the results of the cluster analysis withthose of the DCA analysis indicates that the investigatedbryozoan faunas may be separated into five distinctgroups (Figs. 4 and 5). The two Tethyan faunas form afairly tight group in spite of their difference in age; thenorthern Pangaean faunas, including East Greenland,form a second distinctive group; while the Early Permianfaunas of the Russian Platform form a third, well-definedgroup. In addition two faunas are sufficiently differentfrom all others to warrant the separation of two singlefauna groups: the Zechstein Basin fauna, which has amuch higher degree of similarity to the northern Pangaeanfaunas than to the Tethyan faunas, and the Kazanian faunaof the Russian Platform, which shows the highest resem-blance to the Kazanian faunas from Greenland (Fig. 5).
The combined results of the cluster and DCA analysisindicate that the Tethyan faunas in general have little in
common with the bryozoan faunas from the RussianPlatform, northern Pangaea and the Zechstein Basin. Thehighest degree of similarity between the western Tethysand the Russian Platform is found in the Early Permian,which is in accordance with the notion that there was aconnection from the North Greenland–Svalbard areaacross the Russian Platform to the Tethys Ocean duringthis time (Fig. 3A). However, the statistical analyses alsoindicate that even during this period of open marineconnections from the Tethys to the north slope of Pangeathree rather different bryozoan faunas co-existed duringthe Early Permian in western Tethys, Russian Platformand along the northern margin of Pangea, respectively.This most likely reflects the transition from tropicalconditions in the Tethys area to warm-water, subtropicalconditions in the Barents Sea area. The almost completelack of similarity between the Tethyan fauna and RussianPlatform fauna in the Kazanian offers additional supportto a mid-Permian closure of the migration route fol-lowing the collision between Siberia and Laurussia.
The very high degree of similarity with the Sakmarianfauna of Svalbard and the younger, Artinskian–Kaza-nian bryozoan faunas of Svalbard and North Greenlandsuggests little evolution of the northern Pangean faunafollowing initial migration into the region during theLate Carboniferous–Early Permian. This is somewhatsurprising, since facies and fauna analyses indicate pro-nounced shifts in depositional conditions of the northernPangaean shelf from warm, subtropical in the Asselian–Artinskian to cool-water, temperate in the Kungurian–Kazanian (Stemmerik, 1997). The stability of thenorthern Pangaean bryozoan fauna is also in contrast tothe pronounced evolution seen both on the RussianPlatform and in the Tethyan Ocean during the Permian.
The results of the statistical analyses reveal that theEast Greenland bryozoan fauna is related more to thenorthern faunas in Svalbard and North Greenland than tothe Zechstein Basin bryozoan fauna. Thus southwardsmigration of the northern Pangaean bryozoan fauna alongthe evolving North Atlantic rifted seaway was clearlyfacilitated, at least as far south as central East Greenland(Figs. 3–5). However, the Zechstein Basin fauna,although constituting a distinctive group by itself in thestatistical analyses, has a much greater similarity to thenorthern Pangaean faunas than to any of the westernTethys faunas, thus supporting a model where migrationcontinued into the Zechstein Basin, albeit. Of the 13genera described from the Zechstein Basin four are incommon for all the studied faunas and seem to be cosmo-politan, and these four are the only genera shared betweenthe Kazanian fauna of the western Tethys platforms andthe Zechstein Basin. In contrast, the Zechstein Basin

208 A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
fauna shares three additional genera with the well-studiedKazanian fauna of Svalbard in addition to onemore genusshared with the less well-known Kazanian faunas of Eastand North Greenland (Figs. 3 and 6). This pattern ofdistribution further supports a derivation of the ZechsteinBasin fauna without any input from the western CentralTethys platforms immediately east of the Zechstein Basin.
The southward migration from the northern PangaeanBoreal faunas into the warmer settings of East Greenlandseems to have taken place without major changes infaunal composition judging from the very high degree ofsimilarity between these faunas (Figs. 3–6). However,major changes did take place as migration progressedfurther southwards into the even warmer, in part evap-oritic settings of the Zechstein Basin. Only members ofthe orders Trepostomata and Fenestrata managed suc-cessfully to get established under the somewhat adverseconditions characterising this semi-isolated basin, and itcan therefore be concluded that the main environmentalthreshold in the Atlantic rift-arm was located somewhereto the south of the East Greenland Basin.
5. Summary and conclusions
During Early Permian times, the tropical Tethys Seaand the huge shelf along the northern margin of Pangeawere connected through the seaway between Baltica andSiberia allowing the migration of bryozoans as well asother benthos. During the mid-Permian, these marineconnections were cut-off due to the beginning collisionbetween Baltica/Laurussia and Siberia, and distinctivebryozoan faunas began to develop. Rifting along theNorth Atlantic rift system formed a marine seawaybetween Norway and Greenland during the later partsof the Permian which led to flooding of first the EastGreenland Basin, and later the intracratonic ZechsteinBasin. Faunal migration into the subtropically situated,partly isolated Zechstein Basin was restricted to a routeacross the temperate Boreal shelves of northern Pangaea,which resulted in immigration of a fairly low diversityimpoverished fauna with many endemics.
The statistical analyses highlight trends in faunalmigration and evolution, within an overall history of LateCarboniferous–Early Permian open marine connectionsfrom the Tethyan Ocean to the north slope of Pangea, amid Permian closure of this sea-way, and a Late Permiandevelopment of an Atlantic rift-arm connecting thePangean north slope to the intra-cratonic ZechsteinBasin via East Greenland. During the early part of thishistory, limited evolution took place and the bryozoanfaunas of the Pangean north slope in particular remainedvery stable. In the later parts of the Permian, local
evolution was more evident, possibly as a result of morepronounced temperature and salinity gradients. Inconcord with Ernst (2001b), our analyses rule out thepossibility that any migrational connections existedbetween the Tethyan Ocean and the Zechstein Basin inthe Late Permian.
Acknowledgements
We would like to thank Hans Arne Nakrem, Oslo,for introducing us to the bryozoan fauna in Svalbard andD.A.T. Harper, Copenhagen for helping with the statis-tical analyses.
References
Beauchamp, B., 1994. Permian climatic cooling in the CanadianArctic. In: Klein, G.D. (Ed.), Pangea: Paleoclimate, Tectonics andSedimentation During Accretion, Zenith and Breakup of a Super-continent. Geological Society of America Special Paper, vol. 288,pp. 229–246.
Beauchamp, B., Baud, A., 2003. Growth and demise of Permianbiogenic chert along the northwest Pangea: evidence for end-Permian collapse of thermohaline circulation. Palaeogeography,Palaeoclimatology, Palaeoecology 184, 37–63.
Beauchamp, B., Desrochers, A., 1997. Permian warm- to very cold-water carbonates and chert in the Sverdrup Basin–Barents Seaarea, northwestern Pangea. In: James, N.P., Clarke, J.A.D. (Eds.),Cool-Water Carbonates. Society of Economic Paleontologists andMineralogists Special Publication, vol. 56, pp. 349–364.
Dallmann, W.K., Gjelberg, J.G, Harland, W.B., Johannessen, E.P.,Keilen, H.B., Lønøy, A., Nilsson, I., Worsley, D., 1999. UpperPalaeozoic lithostratigraphy. In: Dallmann, W.K. (Ed.), Lithos-tratigraphic Lexicon of Svalbard. Review and Recommendationsfor Nomenclature Use. Upper Palaeozoic to Quaternary bedrock.Norsk Polarinstitutt, Tromsø, pp. 25–126.
Ernst, A., 2000. Permian bryozoans of the NW-Tethys. FACIES 43,79–102.
Ernst, A., 2001a. Bryozoa of the Upper Permian Zechstein Formationof Germany. Senckenbergiana Lethaea 81 (1), 135–181.
Ernst, A., 2001b. Systematics and biogeography of the Permianbryozoans in Europe. In: Wyse Jackson, P.N., Buttler, C.J.,Spencer-Jones, M. (Eds.), Bryozoan Studies 2001. A.A. BalkemaPublishers, Lisse, pp. 109–112.
Ernst, A., 2003. Upper Palaeozoic bryozoans from the Carnic Alps(Austria). Freiberger Forschungshefte C499, 55–77.
Golonka, J., 2000. Cambrian–Neogene Plate Tectonic Maps. Wydaw-nictwo Uniwersyteu Jagiellonskiego, Krakow. 125 pp.
Henderson, C.M, Mei, S., 2000. Preliminary cool water Permian cono-dont zonation in north Pangea: a review. Permophiles 36, 16–23.
Hollingworth, N.T.J., Tucker, M.E., 1987. The Upper Permian(Zechstein) Tunstall Reef of North East England: palaeoecologyand early diagenesis. In: Peryt, T.M. (Ed.), The Zechstein Facies inEurope. Lecture Notes in Earth Sciences, vol. 10. Springer Verlag,pp. 5–22.
Larssen,G.B., Elvebakk,G.,Henriksen, L.B.,Kristensen, S.-E.,Nilsson, I.,Samuelsberg, T.J., Svånå, T.A., Stemmerik, L., Worsley, D., 2005.Upper Palaeozoic lithostratigraphy of the Southern Norwegian BarentsSea. Geological Survey of Norway Bulletin 444, 3–43.

209A.M. Sørensen et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 251 (2007) 198–209
Madsen, L., 1994. Bryozoans from the Upper Palaeozoic sequence inthe Wandel Sea Basin, North Greenland. In: Håkansson, E. (Ed.),Wandel Sea Basin: Basin Analysis, Scientific Report #6.Unpublished Report. University of Copenhagen, Copenhagen. 18pp., 10 plates.
Madsen, L., Håkansson, E., 1989. Upper Palaeozoic bryozoans fromthe Wandel Sea Basin, North Greenland. Rapport GrønlandsGeologiske Undersøgelse 144, 43–52.
Malecki, J., 1968. Permian bryozoans from the Trokrossøya beds,Sørkkappland, Vestspitsbergen. Studia Geologica Polonica 21,7–32.
Malecki, J., 1977. Permian bryozoans from southern Spitsbergen andBjørnøya (Svalbard). Studia Geologica Polonica 51, 75–87.
Mangerud, G., 1994. Palynostratigraphy of the Permian and lowermostTriassic succession, Finnmark Platform, Barents Sea. Review ofPalaeobotany and Palynology 82, 317–349.
Nakrem, H.A., 1988. Permian bryozoans from southern Spitsbergenand Bjørnøya. A review of bryozoans described by J. Malecki(1968, 1977). Polar Research 6, 113–121.
Nakrem, H.A., 1991. Distribution of bryozoans in the Permiansuccession of Svalbard (Preliminary data). In: Bigley, F.P. (Ed.),Bryozoaires Actuels et Fossiles: Bryozoa Living and Fossil.Bullentin de la societe des Sciences Naturelles de l'Quest de laFrance, Memoire HS 1, Nantes, France, pp. 291–298.
Nakrem, H.A., 1993. Upper Palaeozoic–Lower Triassic Bryozoa fromSvalbard: Taxonomy, Stratigraphy and Palaeoenvironments.Doctor ScientiarumUniversity of Oslo, Oslo. 338 pp.
Nakrem, H.A., 1994a. Environmental distribution of bryozoans in thePermian of Spitsbergen. In: Hayward, P.J., Ryland, J.S., Taylor, P.D.(Eds.), Biology and Palaeobiology of Bryozoans. Olsen & Olsen,Fredensborg, pp. 133–137.
Nakrem, H.A., 1994b. Middle Carboniferous–Lower Permianbryozoans from Spitsbergen. Acta Palaeontologica Polonica 39(1), 45–116.
Nakrem, H.A., 1994c. Bryozoans from the Lower Permian VøringenMember (Kapp Starostin Formation), Spitsbergen (Svalbard).Norsk Polarinstitutt Skrifter, vol. 196. 92 pp.
Nakrem, H.A., 2004. Natural History Museum, University of Oslo,Palaeontological Type Collection. http://natmus.uio.no/svalex/svalbard_fossils/index.html.
Nakrem, H.A., Nilsson, I., Mangerud, G., 1992. Permian biostratig-raphy of Svalbard (Arctic Norway) – a review. InternationalGeology Review 34, 933–959.
Rasmussen, J.A., Piasecki, S., Stemmerik, L., Stouge, S., 1990. LatePermian conodonts from central East Greenland. Neues Jahrbuchfür Geologie und Paläontologie, Abhandlungen 178, 309–324.
Ross, J.R.P., Ross, C.A., 1962. Faunas and correlation of the LatePalaeozoic rocks of Northeast Greenland: Part 4. Bryozoa.Meddelelser om Grønland 167 (7) 65 pp.
Ross, J.R.P., Ross, C.A., 1990. Late Palaeozoic Bryozoan biogeog-raphy. In: McKerrow, W.S., Scotese, C.R. (Eds.), PalaeozoicPalaeogeography and Biogeography. Memoir Geological Society,London, pp. 353–362.
Scotese, C.R., 2004. A continental drift flipbook. The Journal ofGeology 112, 729–741.
Southwood, D. A., 1985. The taxonomy and paleoecology of Bryozoafrom the Upper Permian Zechstein reef of N.E. England.Unpublished PhD Thesis, University of Durham. 407 pp.
Southwood, D.A., 1990. New bryozoan taxa from the Upper PermianZechstein reef in N.E. England. Proceedings of the YorkshireGeological Society 48 (1), 33–40.
Stemmerik, L., 1997. Permian (Artinskian–Kazanian) cool-watercarbonates in North Greenland, Svalbard and the western BarentsSea. In: James, N.P., Clarke, J.A.D. (Eds.), Cool-Water Carbo-nates. Society of Economic Paleontologists and MineralogistsSpecial Publication, vol. 56, pp. 349–364.
Stemmerik, L., 2000. Late Palaeozoic evolution of the North Atlanticmargin of Pangea. Palaeogeography, Palaeoclimatology, Palaeoe-cology 161, 95–126.
Stemmerik, L., 2001. Stratigraphy of the Upper Permian WegenerHalvø Formation, Karstryggen area, East Greenland – a lowproductivity carbonate platform. Sedimentology 48, 79–97.
Stemmerik, L., Worsley, D., 2005. 30 years on: Arctic UpperPalaeozoic stratigraphy, depositional evolution and hydrocarbonprospectivity. Norwegian Journal of Geology 85, 151–168.
Stemmerik, L., Håkansson, E., Madsen, L., Nilsson, I., Piasecki, S.,Pinard, S., Rasmussen, J.A., 1996. Stratigraphy and depositionalevolution of the Upper Palaeozoic sedimentary succession ineastern Peary Land, North Greenland. Bulletin GrønlandsGeologiske Undersøgelse 171, 45–71.
Utting, J., Piasecki, S., 1995. Palynology of the Permian of northerncontinents: a review. In: Scholle, P.A., Peryt, T.M., Ulmer-Scholle,D.S. (Eds.), The Permian of Northern Pangea. Springer-Verlag,Berlin, Heidelberg, pp. 236–261.




















