
Carbon–isotope stratigraphy and ammonite faunal turnover for the Middle Jurassic in the Southern...
23
Carbon–isotope stratigraphy and ammonite faunal turnover for the Middle Jurassic in the Southern Iberian palaeomargin Luis O'Dogherty a, ⁎ , José Sandoval b , Annachiara Bartolini c , Sébastien Bruchez d , Markus Bill d , Jean Guex d a Departamento Ciencias de la Tierra; CASEM, 11510, Puerto Real, Spain b Departamento Estratigrafia y Paleontologia, Universidad de Granada 18002, Granada, Spain c CNRS UMR 5143, Université Paris VI, 4 Place Jussieu, 75252, Paris, France d Institut de Géologie et Paléontologie, BFSH-2, 1015 Lausanne, Switzerland Received 19 July 2005; received in revised form 30 January 2006; accepted 31 January 2006 Abstract Variations in the stable carbon–isotope ratio of marine and continental sediments can reflect changes in sink and flux modifications of the palaeocarbon cycle. Here we report carbon–isotope compositions of Middle Jurassic marine carbonates from the Betic Cordillera (southern Spain), which represents an ideal region to link the stable carbon–isotope curves directly to ammonite zones and subzones, and thereby for the first time achieve an accurate chronostratigraphic calibration. The five sections studied represent basin and high swell deposits of the Southern Iberian palaeomargin. We find a similar δ 13 C of carbonates between different oceanic areas, suggesting a homogeneous carbon–isotope oceanic reservoir through the Middle Jurassic. The Aalenian–Bajocian transition is a critical period in ammonite evolution; hence the Early Jurassic fauna are replaced by new ammonite families which become dominant throughout the Middle and Late Jurassic. For this reason, we compared the δ 13 C values of carbonates with ammonite diversity and extinction rates at different taxonomical levels in order to explore the possible relationship between the carbon cycle and ammonite evolution. The carbon–isotope values of carbonates are not exactly linearly correlated with the extinction rate and ammonite diversity, but the main faunal turnovers follow minimum δ 13 C values, where extinct taxa are replaced by new ones. Likewise, radiation episodes are associated with increasing δ 13 C values and with transgressive sea-level rise. All these data support the idea that perturbations in the global carbon cycle reflect rapid palaeoenvironmental changes. We made detailed analyses of these faunal turnovers, using them as a proxy to identify major palaeoenvironmental crises in their ecosystems forced by modification in the carbon cycle. © 2006 Elsevier B.V. All rights reserved. Keywords: Middle Jurassic; Subbetic; Carbon–isotope; Relative sea level; Ammonite turnover 1. Introduction The Middle Jurassic of many Subbetic sections (southern Spain) is characterized by thick and contin- uous successions of pelagic marl and limestone alternations, or nodular limestones bearing abundant ammonites. A highly detailed ammonite biostratigraphy with data from many Subbetic sections has already been established at the zone and subzone levels (Sequeiros, 1979; Sandoval, 1983, 1990; Linares and Sandoval, 1993; Sandoval et al., 2001a). The established zonal Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311 – 333 www.elsevier.com/locate/palaeo ⁎ Corresponding author. E-mail address: [email protected] (L. O'Dogherty). 0031-0182/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2006.01.018
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Carbon–isotope stratigraphy and ammonite faunal turnover for the Middle Jurassic in the Southern...

laeoecology 239 (2006) 311–333www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology, Pa
Carbon–isotope stratigraphy and ammonite faunal turnover for theMiddle Jurassic in the Southern Iberian palaeomargin
Luis O'Dogherty a,⁎, José Sandoval b, Annachiara Bartolini c, Sébastien Bruchez d,Markus Bill d, Jean Guex d
a Departamento Ciencias de la Tierra; CASEM, 11510, Puerto Real, Spainb Departamento Estratigrafia y Paleontologia, Universidad de Granada 18002, Granada, Spain
c CNRS UMR 5143, Université Paris VI, 4 Place Jussieu, 75252, Paris, Franced Institut de Géologie et Paléontologie, BFSH-2, 1015 Lausanne, Switzerland
Received 19 July 2005; received in revised form 30 January 2006; accepted 31 January 2006
Abstract
Variations in the stable carbon–isotope ratio of marine and continental sediments can reflect changes in sink and fluxmodifications of the palaeocarbon cycle. Here we report carbon–isotope compositions of Middle Jurassic marine carbonates fromthe Betic Cordillera (southern Spain), which represents an ideal region to link the stable carbon–isotope curves directly toammonite zones and subzones, and thereby for the first time achieve an accurate chronostratigraphic calibration. The five sectionsstudied represent basin and high swell deposits of the Southern Iberian palaeomargin. We find a similar δ13C of carbonates betweendifferent oceanic areas, suggesting a homogeneous carbon–isotope oceanic reservoir through the Middle Jurassic.
The Aalenian–Bajocian transition is a critical period in ammonite evolution; hence the Early Jurassic fauna are replaced by newammonite families which become dominant throughout the Middle and Late Jurassic. For this reason, we compared the δ13C valuesof carbonates with ammonite diversity and extinction rates at different taxonomical levels in order to explore the possiblerelationship between the carbon cycle and ammonite evolution. The carbon–isotope values of carbonates are not exactly linearlycorrelated with the extinction rate and ammonite diversity, but the main faunal turnovers follow minimum δ13C values, whereextinct taxa are replaced by new ones. Likewise, radiation episodes are associated with increasing δ13C values and withtransgressive sea-level rise. All these data support the idea that perturbations in the global carbon cycle reflect rapidpalaeoenvironmental changes. We made detailed analyses of these faunal turnovers, using them as a proxy to identify majorpalaeoenvironmental crises in their ecosystems forced by modification in the carbon cycle.© 2006 Elsevier B.V. All rights reserved.
Keywords: Middle Jurassic; Subbetic; Carbon–isotope; Relative sea level; Ammonite turnover
1. Introduction
The Middle Jurassic of many Subbetic sections(southern Spain) is characterized by thick and contin-
⁎ Corresponding author.E-mail address: [email protected] (L. O'Dogherty).
0031-0182/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.palaeo.2006.01.018
uous successions of pelagic marl and limestonealternations, or nodular limestones bearing abundantammonites. A highly detailed ammonite biostratigraphywith data from many Subbetic sections has already beenestablished at the zone and subzone levels (Sequeiros,1979; Sandoval, 1983, 1990; Linares and Sandoval,1993; Sandoval et al., 2001a). The established zonal

312 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
scheme, synthesized by Aurell et al. (2002) is used here.Likewise, some preliminary data have been compiledfor carbon isotope stratigraphy of some Subbeticsections (Bartolini et al., 1997, 1998; Sandoval et al.,2004). Also, the connection of palaeoceanographicevents and organic crises has been frequently docu-mented (Hallam, 1987, 1989, 1990; Hallam andWignall, 1997, 1999; Wignall, 2004a,b; and otherreferences). In this context, an analysis of the diversityand faunal turnover of ammonites from Middle Jurassicof the Betic Cordillera and western Tethys in relation torelative sea-level changes has been published (Sandovalet al., 2001b, 2002b).
The aims of the present work are (a) to elaboratea composite δ13C marine isotopic curve for the LateAalenian–Early Callovian interval to be consideredas a reference for the Mediterranean regions; (b) tocalibrate the carbon–isotope curve with highlydetailed ammonite biostratigraphy; (c) to correlatethe Subbetic curve with subboreal and Submediterra-nean regions; (e) to analyse the possible connectionsbetween ammonite extinctions and radiations (includ-ing major evolutionary ammonite events), relativesea-level changes, and carbon–isotope anomalies
Fig. 1. Sketch map of the Betic Cordillera (southern Spain) showing the difsections analysed.
during the Middle Jurassic. For this purpose fivecontinuous successions with high-resolution ammo-nite biostratigraphy were sampled (bed by bed) toestablish a detailed carbon–isotope stratigraphy indifferent palaeogeographic sectors (from deep troughsto pelagic swells), which enable us to accuratelycalibrate the high-resolution carbon–isotope curveobtained for the Upper Aalenian–Lower Callovianinterval.
2. Geological setting and palaeogeography
The sections studied are located in the provinces ofCórdoba and Jáen (Andalusia, S Spain) and Murcia(SE Spain). Geologically, they correspond to theExternal Zones of the Betic Cordillera, which is abelt 600 km long by 200 km wide extending from theCádiz to the Murcia provinces, with a WSW–ENEtrend, and belonging to the western end of theEuropean Alpine Chains. From a palaeogeographicstandpoint they constituted the southern passive marginof the Iberian Plate. Within the Betic External Zonestwo main geological units have been differentiated,Prebetic and Subbetic (Fig. 1), which correspond to
ferent paleogeographic domains and position (stars on the map) of the

313L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
two palaeogeographic domains in accord with theirrelative locations with respect to the major emergedareas of the Iberian Plate. The Prebetic Domain showssedimentary rocks that were deposited closer to thecontinent and in which shallow facies, coastal, andeven continental facies dominated during Jurassic andCretaceous times. The Subbetic Domain is the area farfrom the continent in which pelagic facies weredeposited from the beginning of the main intraconti-nental phase of rifting (190 My ago) (García-Hernán-dez et al., 1989; Arias et al., 2004). The Subbetic islocated in the central part of the External Zones of theBetic Cordillera (Fig. 1), and is the only Betic domainwith more or less continuous pelagic or hemipelagicsedimentation from the middle part of the LowerJurassic (Pliensbachian) to the Upper Cretaceous; i.e.pelagic–hemipelagic deposition characterizes the strati-graphic interval analysed. The physiography of theSubbetic basin was very irregular due to severe Jurassicintracontinental rifting, which caused two high swellareas (External and Internal Subbetic) and onerelatively deep trough (Median Subbetic) in theSouthern Iberian palaeomargin.
3. Material and analytical laboratory procedure
A total of 548 limestone samples (see Appendix)covering the Upper Aalenian–Lower Callovian intervalwere analysed for their stable carbon and oxygenisotopes. All stable-isotope analyses were made onbulk carbonate samples, selecting only limestones ormarly limestones (>80% CaCO3). Rockpowder sampleswere analysed for carbon–isotope composition at theLaboratory of Isotope Geochemistry of the University ofLausanne. Except for the Casa Chimeneas section (seebelow), the δ13C and δ18O contents were measuredfollowing the standard procedure of McCrea (1950).The samples were reacted with 100% H3PO4 at 50°Cunder vacuum for 30min to produce CO2, applying afractionation factor of 1.00931 between CaCO3 andH3PO4 for oxygen isotopes (Swart et al., 1991). Theδ13C and the δ18O values were measured through a dualinlet interface in either a Finningan MAT 251 multi-collector Mass Spectrometer or in a Finningan Matdelta-S Mass Spectrometer.
The Casa Chimeneas samples were analysedfollowing the procedure describe in Spötl andVennemann (2003): reaction in 99% H3PO4 in ahelium atmosphere, separation of the CO2 produced ina Finnigan GasBench II and then sent by continuousflow interface in a Finnigan Delta Deltaplus XL MassSpectrometer.
All the isotopic composition of carbon is given in theconventional δ-notation in parts per mill (‰) relative tothe Peedee Belemnite (PDB) standard:
d13CPDB ¼13C=12Csample
13C=12Cstandard
−1
" #T1000
The CO2 reference gas (working standard) in theLausanne Laboratory was calibrated against NBS-19standard δ13C=1.95‰ and δ18O=−2.20 (Coplen et al.,1983). The standard deviation of the analyses is±0.05‰ for δ13C and ±0.1‰ for δ18O.
4. Lithology description, biostratigraphy andcarbon–isotope stratigraphy
Carbon–isotope analyses were studied in five sec-tions spanning from Upper Aalenian–Lower Callovianinterval located both on basinal area (Median Subbetic,three analysed sections) and on the swells (ExternalSubbetic, two analysed sections) (Fig. 1). In general,strata deposited on the highs are represented by lightgrey to pink biomicritic limestones, nodular limestones,and red marls, where the basinal facies have thickersequences of interbedded darker grey siliceous micriticlimestones and marls.
4.1. Median Subbetic: basinal sections
4.1.1. Casa Blanca section (JCB)This section (Figs. 1 and 2A) is located in the central
sector of the Median Subbetic, outcropping along theCasa Blanca road, approximately 3km NW of theCampillo de Arenas village, Jaén province (UTMgeographic location VG415584, Sheet 969, Valdepeñasde Jaén). Previous biostratigraphic studies have beencarried out by Olóriz et al. (1979) and by Sandoval(1983). The section shows transitional features betweenthe External and Median Subbetic such as the presenceof nodular limestones in the Upper Jurassic.
The lower part of the section (Concavum, Discites,Laeviuscula Zones and Propinquans pro part Zone),nearly 60m thick, consists of grey marls, marlylimestones and limestones (mudstones and wackes-tones of Bositra and radiolarians). Calcareous hor-izons clearly predominate over marls, and bedthickness varies from 10 to 75cm. Trace fossils, likeChondrites and, less commonly, Zoophycos occur.Occasionally, stratification is slightly altered byslumps. Ammonite-rich beds are scarce, with ammo-nite assemblages composed of diverse sonniniids

Fig. 2. Lithologic successions plotted against the δ13C curve in three typical Median Subbetic (deep pelagic trough) sections. Shaded boxes indicateuncertainty at subzone or zone boundaries. See location in Fig. 1.
314 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333

315L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
(Sonninia, Papilliceras, Witchelia), otoitids (Emileia,Otoites), stephanoceratids (Skirroceras), haploceratins,bradfordiins, etc., which provide an accurate biostra-tigraphy; lytoceratids and phylloceratids also appear.
The upper part of the Propinquans Zone and thelower part of the Humphriesianum Zone (Romani andHumphriesianum pro part Subzones) are nearly 69mthick. These are represented by whitish grey limestoneswith abundant chert nodules, marly limestones, andlocally thin intercalations of grey marls. Bed thicknessranges from 15cm to more than 120cm. Two conspic-uous slumps occur and trace fossils (Chondrites andZoophycos) are relatively common. Ammonites, al-though scarce, allow optimal biochronologic interpreta-tions (Sandoval, 1983).
The middle and upper parts of the HumphriesianumZone (Humphriesianum pro part and Blagdeni Sub-zones) consist of 6.5m thick of whitish marly lime-stones, overlaid by violet to red nodular siliceouslimestones (mudstones and wackestones with radiolar-ians and thin shelled pelagic bivalves) which is areference level in this succession. Bed thickness variesfrom 10 to 20cm, showing abundant Zoophycos in somelevels. The relatively abundant, but poorly preservedammonite fauna is dominated by stephanoceratids(Stephanoceras) and by phylloceratids.
The lower part of the Upper Bajocian strata consist of7.7m of decimetric beds made of slightly nodular violetor yellowish marly limestones and marls. Scarce andpoorly preserved ammonites (Holcophylloceras, Loku-ticeras, Cadomites and Leptosphinctes) identify theNiortense Zone.
Upper Bajocian and Lower Bathonian depositsconsist of approximately 36.5m of whitish marlylimestones and limestones (individual beds rangingfrom 15 to 130cm in thickness) with chert in nodules oras thin ribbons, especially in the upper part. Wack-estones with pelagic bivalves and radiolarians are thetypical microfacies. This stratigraphic interval is poor inammonites, registering only one specimen of Dimor-phinites and scarce phylloceratids. In the uppermost partof this interval a sample yielded well preservedradiolarians, this sample being assigned to the UAZ 5of Baumgartner et al. (1995) (O'Dogherty et al., 1995).
The uppermost levels sampled consist of 4m ofslightly nodular siliceous limestones with thin intercala-tions of chert. Rare and poorly preserved ammonites(Bullatimorphites, Wagnericeras) enable the identifica-tion of the Middle Bathonian (Costatus Zone, sensuSandoval, 1983).
The δ13C curve (Figs. 2 and 4A) presents a markedminimum around the Aalenian–Bajocian boundary
(1.5–2‰). The Casa Blanca section documents anincrease in δ13C values (Δ=1‰) from the Aalenian–Bajocian boundary to theDiscites–Laeviuscula Zones (upto 2.5–3.0‰). Through the Propinquans–Humphriesia-numZones carbon isotope follows the positive δ13C lowerBajocian excursion, with a restricted maximum of 3.4‰in the Propinquans–Humphriesianum Zones boundary.The values decrease to 2.0‰ in the Niortense Zone.
4.1.2. Agua Larga section (AQ1)This section, located in the central sector of the
Median Subbetic (Figs. 1 and 2C), outcrops along theAgua Larga stream, 500m NWof the Cortijo Alorí nearde villages of Campillo de Arenas and Montejicar,Province of Jaén (UTM geographic location VG514601,Sheet 969, Valdepeñas de Jaén). The section haspreviously been studied by Sandoval (1983), Linaresand Sandoval (1990, 1993, 1996) and Sandoval andChandler (2000). In this section, we report data forUpper Aalenian–Lower Bajocian interval.
The Upper Aalenian (Concavum Zone) is approxi-mately 45m thick; marly limestones dominate overmarls. Ammonite fauna is scarce but significant(Ludwidgella cornu Buckman, Malladaites, Euapteto-ceras, Graphoceras) in the lower part (ConcavumSubzone). In the upper part (Limitatum Subzone)marly deposits predominate and yield a very abundantand diversified ammonite fauna with predominance ofgraphoceratins and erycitids (Haplopleuroceras); otherammonites such as harpoceratins, hammatoceratins,sonniniids, otoitids, strigoceratids, bradfordiins, phyllo-ceratids and lytoceratids are scarcer, but some arecharacteristic biochronologic markers.
In the Lower Bajocian (Discites Zone), which isabout 17m thick, the lithology and microfacies are verysimilar to the underlying beds, with marly beds beingthicker than marly limestones. As in the previousunderlying subzone, ammonite assemblages are domi-nated by graphoceratins and Haplopleuroceras, whichare very abundant at the base and decrease progressivelyupwards to become extinct at the upper part of this zone.Sonniniids, bradfordiins, otoitids, phylloceratids andlytoceratids can also be relatively common.
The Laeviuscula Zone is nearly 10m thick. In thelower part of this zone the lithology (grey marls, marlylimestones and limestones) and the microfacies (mud-stones and packstones with radiolarians and pelagicbivalves) are similar to the underlying levels, althoughmarly limestones and locally limestones predominateover marls, locally exhibiting abundant Zoophycos. Theupper part of the Laeviuscula Zone consists of 2m ofviolet marls and nodular limestones. The ammonite

316 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
fauna is dominated by sonniniids (Sonninia, Witchellia)and bradfordiins (Bradfordia), which are the mostcharacteristic biochronological markers of this zone,together with scarce otoitids, phylloderatids andlytocetatids.
Beds belonging to the Propinquans Zone areapproximately 35m thick. The lower levels (3m) consistof violet marls and nodular limestones similar to theunderlying beds. The rest are alternating grey marls andgrey–white limestones (mudstones and packstones withradiolarians and Bositra), which episodically showabundant chert nodules. Some calcareous beds exceed1m in thickness, but decimetre beds are dominant. Thepresence of Zoophycos is a typical feature of theselevels. The ammonite assemblages are dominated byphylloceratids, sonniniids (Witchellia, Sonninia, etc.),otoitids (Emileia,Otoites) and stephanoceratids. Sphaer-oceratids (Labyrinthoceras), strigoceratids, haplocera-tins, bradfordiins and lytoceratids are also common.
The uppermost sampled levels (8.5m) are wellbedded whitish limestones with locally abundance ofchert nodules and ribbons. Scarce ammonites (Dorse-and Stephanoceras) characterize the HumphriesianumZone.
Radiolarians occur only in the lower part of thesection (Concavum Zone) and they are very abundantand have a moderate to relatively good preservation.Samples AQ1-63.0 and AQ1-71.60 yielded a well-preserved assemblage typical of the UAZones 3 ofBaumgartner et al. (1995). The most characteristicspecies are: Archaeohagiastrum longipes Baumgartner,Linaresia rifensis (El Kadiri), Palinandromeda sog-noensis Baumgartner, Parasaturnalis diplocyclis (Yao),Zartus spp., Acanthocircus suboblongus (Yao), Emilu-via splendida Carter, Parahsuum ? natorense (El Kadiri)and Mirifusus proavus Tonielli.
The Agua Larga δ13C curve is very detailed for theAalenian–Lower Bajocian interval (Figs. 2 and 4C).The curve shows a progressive tendency towards lowvalues from 2.7‰ at the Upper Aalenian (ConcavumSubzone) to reach a minimum (1.5–1.7‰) at theAalenian–Bajocian boundary, showing a difference(Δ) of 1‰. In the Lower Bajocian, the values risesharply in the Laeviuscula Zone to a main positive peakaround 2.9–3.2‰ from the base of the PropinquansZone to the Humphresianum Zone.
4.1.3. Casa Chimeneas section (SRi3)This section (Figs. 1 and 2) is located along a local
road, 1500m NW of the Cerro Mahoma in Mula,province of Murcia (UTM geographic locationsXH315202, Sheet 912, Mula). Biostratigraphic and
palaeontologic analyses were carried out by Linares andSandoval (1977), Sandoval (1983, 1990), O'Dogherty etal. (2001) and Sandoval et al. (2002a). In this section,we analysed the upper Lower Bajocian (HumpriesianumZone) to Upper Bajocian (Niortense Zone) interval.
The lower part of the analysed interval (Humphriesia-num Zone) is made up of 56m of whitish-grey marls,marly limestones, and limestones (mudstones and wack-estones of thin shelled bivalves and radiolarians). Bedthickness of marly limestones varies between 10 and60cm, and terrigenous intercalations reach 150cm thick.Downward in the section, marls predominate over marlylimestones; in the middle part (Humphriesianum Sub-zone), calcareous beds become thicker and chert nodulesare relatively common. Within the upper part of theHumphriesianum Zone (Blagdeni Subzone) chert nodulesdisappear, with bioturbation, especially Zoophycos, beingvery common. The pelagic bivalve Bositra is veryabundant throughout the section. Ammonite assemblagescontain haploceratids (Poecylomorphus cycloides (d'Or-bigny)), oppeliids (Oppelia, Oecotraustes), stephanocer-atids (Stephanoceras), sphaeroceratids (Sphaeroceras,Chondroceras), which are good biochronologic markers,strigoceratids, phylloceratids and lytoceratids.
The Niortense Zone is 47m thick is quite similar tothe underlying Humphriesianum Zone in terms oflithology and microfacies (although chert nodules arelacking), and marly intercalations clearly dominate overcalcareous sediments (limestone or marly limestonebeds vary from 5 to 30cm in thickness, while marlyintercalations are frequently thicker than 100cm).Bositra is very abundant and the ammonite assemblagesshow characteristic taxa, with prevailing oppeliids andlytoceratids in the lower part of this zone (Banksi andPolygyralis Subzones) being also common parkinso-niids, cadomitins, leptosphinctins and phylloceratids.Leptosphinctins (Leptosphinctes and Cleistosphinctes),the heteromorphous genus Spiroceras, oppeliids, stri-goceratids (Strigoceras) and cadomitins (Cadomites) areabundant and characterize the upper part of the zone(Baculata Subzone).
The δ13C values of carbonates (Figs. 2 and 4B) showmaximum values (2.5–2.7‰) in the HumphriesianumZone and then decrease (Δ=1‰) to lower values (1.5–1.8‰) in the upper Niortense Zone (Baculata Subzone).
4.2. External Subbetic: pelagic swell sections
4.2.1. Puerto Escaño section (JC4)This section (Figs. 1 and 3) is located in the Sierra de
Gaena, situated in the central sector of the ExternalSubbetic, and outcrops along the Puerto Escaño path,

Fig. 3. Lithologic succession plotted against the δ13C curve in two typical External Subbetic (high swell) sections. Shaded boxes indicate uncertainty at subzone or zone boundaries. See locationin Fig. 1.
317L.O'Dogherty
etal.
/Palaeogeography,
Palaeoclim
atology,Palaeoecology
239(2006)
311–333

318 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
1200mWof the Village of Carcabuey in the province ofCórdoba (UTM geographic locations UG855447, Sheet989, Lucena). The section has been previously studiedby Delgado et al. (1980), Sandoval (1983, 1986),Steiner et al. (1987) and Sandoval (1990). In this paper,we analyse the Upper Aalenian–Middle Bathonianinterval.
The base of the section consists of marls and marlylimestones of Middle Toarcian (Upper Toarcian andAalenian correspond to a sedimentary hiatus). The restof the sampled section includes Bajocian and Bathonianbeds. Lower Bajocian (Discites, Laeviuscula andPropinquans pro part Zones) consists of nearly of 15mof light-grey micritic limestones (mudstones and wack-estones with radiolarians, crinoids, etc.); some levels areslightly nodular and chert nodules are abundant. Bedthickness varies between 20 and 40cm.
The Propinquans (pro part) to Niortense Zones (8.5mthick) consist of grey to pinkish nodular limestones(wackestones to packstones) with pelagic bivalves,echinoderms, radiolarians, benthic foraminifers, “Pro-toglobigerina”, etc.). Bed thickness fluctuates between15 and 120cm. Sedimentary hiatuses have beenidentified near the Lower–Upper Bajocian boundary(Delgado et al., 1980; Sandoval, 1983), howeverammonite assemblages provide good biostratigraphicresults. The ammonite fauna is dominated by otoitidsand stephanoceratids in the Lower Bajocian, and bystephanoceratids, leptosphinctins and spiroceratids inthe Upper Bajocian (Niortense Zone).
The Garantiana (Annulatum) and Parkinsoni Zonesare 7.5m of yellowish-red marls, nodular marly lime-stones and red nodular limestones (wackestones topackstones with abundant Bositra, benthic foraminifers,“Protoglobigerina”, echinoderms, etc.); one horizon ofcalcareous turbidites exists. Bed thickness variesbetween 10 and 50cm with a predominance of detritalsediments in the Annulatum Zone, while limestonesdominate over marls in the Parkinsoni Zone. Theanalysis of ammonite assemblages composed of oppe-liids, stephanoceratids (Cadomites), morphoceratids(Dimorphinites), leptosphinctins (Vermisphinctes), etc.,together with phylloceratids and nannolytoceratids,gives good biostratigraphic results.
The uppermost part of the studied section correspondsto the Lower andMiddle Bathonian and consists of 5m ofalternating red-violet nodular limestones and marls withmicrofacies similar to those of the underlying beds. Bedthickness varies from 20 to 30cm. Phylloceratids(Calliphylloceras and Holcophylloceras) and, especiallynannolytoceratids (Nannolytoceras tripartitum (Ras-pail)), clearly dominate the ammonite assemblages in
the Lower Bathonian, but oppeliids, morphoceratids(Morphoceras), cadomitins are also common forms. Inturn, phylloceratids together with cadomitins, tulitids(Bullatimorphites) and zigzagiceratins (Procerites, Wag-nericeras) are the dominant ammonite fauna in theMiddle Bathonian.
The δ13C values (Figs. 3 and 4D) increase from 2.2‰to a maximum (3.2–3.0‰) in the Propinquans andHumphriesianum ammonite zones (Δ=1‰) and thendecrease from the Niortense Zone to the ParkinsoniZone (2‰). From the Upper Bajocian to MiddleBathonian the values range between 2.1 and 2.5‰with a slight maximum at the Bajocian–Bathonianboundary (Δ=0.5‰).
4.2.2. La Cornicabra section (JV1)The Cornicabra section (Figs. 1 and 3) is located in
the central sector of the External Subbetic, nearly 700mNWof the Cornicabra peak and 1000mNE of the km 3.5of the local road fromValdepeñas de Jaén to Frailes in theprovince of Jaén (UTM geographic location VG272575,Sheet 969, Valdepeñas de Jaén). Linares et al. (1979) andSandoval (1983, 1990) carried out biostratigraphicstudies dealing with data from the Cornicabra section.In this section, we analysed the Upper Bajocian(Parkinsoni Zone)–Lower Callovian interval.
The lower part of the sampled section consists of14.5m of alternating grey marls and marly limestones(wackestones and packstones with abundant Bositra,radiolarians, benthic foraminifers, etc.) with chert nodules.Bed thickness varies from 25 to 125cm. The ammoniteassemblages are dominated by oppeliids (Oecotraustes),nannolytoceratids (Nannolytoceras), phylloceratids, ste-phanoceratids and morphoceratids (Dimorphinites), al-though leptosphinctins are also common forms, givingbiostratigraphic data of significant value.
Near the Bajocian–Bathonian boundary marly lime-stones progressively change from grey to violet-yellow-ish colors, and red nodular marly limestones are presentin upper levels. These levels (about 4.5m thick) representthe Lower Bathonian.Microfacies consist of wackestonesand packstones with pelagic bivalves, radiolarians,diversified benthic foraminifers, etc. Phylloceratids andespecially nannolytoceratids (N. tripartitum (Raspail),clearly dominate the ammonite assemblages. Oppeliids,morphoceratids (Morphoceras), cadomitins and zigzagi-ceratins (Procerites and Procerozigzag) are also com-mon, providing valuable biostratigraphic data.
The Middle and Upper Bathonian are represented bythe Sofanus, Costatus, and Orbis Zones (the Discusstandard Zone is not represented in the Betic Cordillera;Sandoval, 1983). This interval is made of 3.6m of red

Fig. 4. Biostratigraphic correlation of carbon–isotope curves from the Subbetic domain (Betic Cordillera).
319L.O'Dogherty
etal.
/Palaeogeography,
Palaeoclim
atology,Palaeoecology
239(2006)
311–333

320 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
nodular limestones (typical rosso ammonitico facies).Bed thickness can reach 90cm. Microfacies consist ofpackstones with pelagic bivalves, benthic foraminifers,“Protoglobigerina”, etc. Common and characteristicammonite genera are dominated by cadomitins (Cado-mites), tulitids (Bullatimorphites) and zigzagiceratins(Procerites, Wagnericeras) in the Middle Bathonian,and by oppeliids (Oxycerites), grossouvrins, and tulitidsin the Upper Bathonian.
Callovian sediments are represented by 3.6m of red,locally ferruginous, nodular limestones. Microfaciesconsist of packstones with abundant Bositra and“Protoglobigerina” (possibly Conoglobigerina); ben-thic foraminifers, ostracods and echinoderms are alsocommon faunas. Ammonite assemblages are dominat-ed by tulitids (Kheraiceras) and macrocephalitids(Dollicephalites; Macrocephalites). The Bullatus andGracilis Zones are recognized.
A relative minimum of the δ13C (about 1.7‰) occursat the base of section (Figs. 3 and 4E). The valuesincrease gradually to 2.1–2.3‰ in the upper part ofParkinsoni and Zigzag Zones. Two relative minima(1.7–1.8%) are present in the Upper Bathonian and topof the Lower Callovian, which shows a small positiveexcursion (2.6‰, Δ=0.7‰).
5. Carbon–isotope results
5.1. General aspects
If we consider the marine δ13C curve as the normalexpression for variation of the marine reservoir in theglobal carbon cycle, fluctuations of large magnitude,from light to heavy shifts and vice versa, should be ofsome palaeoenvironmental significance. The generaltrend exhibited by the δ13C curve from the UpperAalenian to Lower Callovian of the Subbetic (Figs. 5and 6) could be summarized as follows:
(a) Concavum to Limitatum Subzones (Figs. 2B, 4C,5, 6): large excursion to lighter values (Δ=0.8‰),
Fig. 5. Diversity of ammonite genera and species with indication of major(with the exception of long-ranging Phylloceratina and Lytoceratina) comrecord for the South Iberian Paleomargin (Subbetic area). Horizontal grey btext: dark-grey bands signify stronger turnover and light-grey ones (4 andfor the ammonites adapted from Sandoval et al. (2001b, 2002b) noted aperturbation on the carbon isotope. The “Middle Jurassic ammonite radconcomitant with this large increase in δ13C and the decline of the Earl(Spiroceras and Parapatoceras) are indicated and coincide with a period oisotopic excursion toward lighter values. Andean-Mixtecan (reineckeids) ana sea-level rise and increasing δ13C values. Ammonite Zones and SubzoHardenbol et al. (1998).
(b) Walkeri to Romani Subzones (Figs. 2A,B, 4C,D,5, 6): large positive excursion (Δ=1‰),
(c) Humphriesianum to Blagdeni Subzones (Figs.2A,C, 3, 4A,B,D, 5, 6): back to median values andstabilization,
(d) Banski to Baculata Subzones (Figs. 2A,C, 3, 4A,B,D, 5, 6): shift to lower values,
(e) Garantiana Zone to Daubenyi Subzone (Figs. 2A,3, 4A,D, 5, 6): stabilization,
(f) Dimorphus Subzone to Zigzag Zones (Figs. 3, 4D,E, 5, 6): weak positive shift,
(g) Aurigerus to Orbis Zones (Figs. 3, 4D,E, 5, 6):weak shift to lower values (Bathonian minimum),
(h) Orbis to Gracilis Zones (Figs. 3–6): positive shift(Δ=0.7‰).
We found a good correlation of δ13C curves amongthe different sections investigated (Fig. 4). Suchcorrelation is supported by a detailed biostratigraphiccorrelation at the ammonite subzone level.
The general trend of the δ13C curves fit well forsections belonging to a same palaeogeographic areawhere the general lithology is quite homogeneousinside. Sections from the Median Subbetic (represent-ing the carbon record in deep pelagic through) showsimilar isotopic trends but absolute values are lighterin some sections (i.e. Fig. 2, Chimeneas sectioncompared to the others). This can be explained by alithological effect as this section is the only one withdominance of marly over limestone ones (Fig. 2).The same fact is also observed in the Fig. 7, wherethe Digne section is shifted by ca. 1‰ to lowervalues but has exactly the same shape. This can beinterpreted as lower local surface productivity due tothe presence of clay minerals, which reduce thephotic zone and/or by introduction of isotopicallylight organic matter of terrestrial origin. In this sense,oxygen isotopes (available in the Appendix) wereprimary used as a rough proxy to check anydiagenetic influence in the δ13C variations as theyare provided by the methodology. No obvious
evolutionary ammonite events at the superfamily and family levelspared with sealevel changes (Haq et al., 1988) and carbon-isotopeands (1–6) correspond to major ammonite turnovers discussed in the5) indicate less marked turnovers). Genus and species diversity plotss the largest diversity periods that corresponding to the maximumiation” (undergone by Stephanoceratoidea and Haploceratoidea) isy Jurassic forms. Occurrences of uncoiled Mid-Jurassic ammonitesf maximum environmental stress, as indicated by sea-level falls andd Indo-Pacific (macrocephalitids) immigrations occurred at a time ofnes according to Sandoval in Aurell et al. (2002); time scale from

321L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333

322 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333

323L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
correlation was seen, neither within individual sec-tions nor through the detailed biostratigraphic corre-lation between them.
The δ18O values, ranging between −3.5‰ and 0‰are slightly lighter than contemporaneous belemnitevalues of the same latitude (0‰ to −2‰; Jenkyns etal., 2002 and references therein), indicating nointensive meteoric diagenesis overprint. These bulkrock carbonates values should encounter at least theeffect of burial diagenesis (lighter cements due to theburial temperature of pore fluids) and thus are difficultto interpret as a primary signal. If no small-scalecorrelation is visible, a trend can be seen from thebasinal sections (−3.5‰ to −1‰) to the high sections(−2‰ to 0‰). This could be related to their higherpaleostructural position reducing the burial diagenesisoverprint.
A similar pattern is observed (Figs. 3 and 4) in thetwo sections of the External Subbetic (representingcondensed sedimentation). Both sections exhibit similarcurves, being more or less equitable in δ13C values; inother words the curves are quite parallel, crossing onlywhen the lithology change (the ammonitico rosso faciesin one section becomes more marly with respect to theother).
The presence of a highly detailed ammonitebiostratigraphy throughout the investigated sectionsalso enables an extraordinary correlation among thecarbon signals found between deep pelagic throughsections (high sedimentation rates) and condensedswells ones (extremely low sedimentation rates).
A noteworthy aspect is that only when a highlydetailed isotopic curve is combined with a high-resolution biostratigraphy do we have a powerful toolfor stratigraphic correlations. In this sense, the well-calibrated curve obtained in the Subbetic area has to beconsidered as a reference δ13C marine isotopic curve forthe Mediterranean regions, which can facilitate isotopiccorrelations among basins, even in the absence ofprecise dating.
Fig. 6. Stratigraphic range of ammonite genera compared to general carbonissmooth curve) is made upon correlation of isotopic curves for both palaeogeodrawing has no scale, as the duration of zones and subzones are drawn after rmade by ammonite calibration at the zone or subzone levels. Note that Caslighter isotopic values than the three other ones, but it fits the general isotopicof marly interbeds throughout this section (see Section 5.1) as in Digne sectsection B). The stratigraphic ranges of ammonite genera for the Subbeticorigination of heteromorph ammonites (Spiroceras and Parapatoceras); noteenvironmental stress periods are linked to decreasing values in the carbon–morphotypes (see more discussion in the text). (For interpretation of the refeversion of this article.)
5.2. Correlation between Sub-Boreal andSub-Mediterranean records
Pelagic carbonate sections through the Aalenian–Bajocian sequence in Sub-Mediterranean province–central Italy and southern France (Bartolini et al.,1996; Bartolini and Cecca, 1999)–together with anddata from the Sub-Boreal province based on belemnitesof the Bearreraig section in the Isle of Skye (Jenkyns etal., 2002), have shown the presence of positive δ13Cexcursions in Lower Aalenian and Lower Bajocian, anda negative δ13C excursion around the Aalenian–Bajocian boundary (see Fig. 7). The Bajocian positiveexcursion has also been identified from a sequence ofredeposited oolites in the Belluno Trough, SouthernAlps (Zempolich and Erba, 1999). Carbon–isotopeanalyses of fossil wood from the Middle JurassicRavenscar Group, Yorkshire, NE England (Sub-Borealprovince), indicate a significant excursion towards lowisotopic values at about the Aalenian–Bajocian bound-ary and less marked positive excursion in the EarlyBajocian (Hesselbo et al., 2003). The similarity of theterrestrial and marine isotope curves is an indication thatthe observed isotopic signals may have recorded globalperturbation of the carbon cycle.
The Fig. 7 shows minor discrepancy concerning themajor negative excursion of the Aalenian–Bajocianboundary. Fig. 7E, for example, suggests that the majornegative excursion occurs in the Late Aalenian, whereasin other sections it is dated as Aalenian–Bajocianboundary or Early Bajocian. This suggests thatammonite based correlations are flawed in a minorway. Our Fig. 4 (sections A, B and D) clearly shows thatthe excursion towards lower δ13C is located systemat-ically near the Aalenian–Bajocian boundary, moreprecisely at the base of the Limitatum Subzone (minimavalues of 1.5‰ at the uppermost Aalenian). In thesesections no uncertainties at subzone or zone boundariesexist. Therefore the inconsistencies displayed by somesection in Fig. 7 (like Fig. 7E) are due to (a) the scarcity
otope record in the Subbetic. The general tendency of the curve (greygraphic areas (pelagic swells and troughs) of the Subbetic. Note that theelative thickness measured on the sections. The correlation of curves isa Chimeneas isotopic curve (blue triangles) is systematically showingtrend. One explanation for such a trend could be the regular dominanceion (detritic-dominated basin in the southeastern of France, see Fig. 7,are adapted from Sandoval et al. (2001b, 2002b). Red stars indicatethat these originations (as well as strong extinctions) occurring at highisotope curve. Also blue dot indicate the occurrence of serpenticonerences to colour in this figure legend, the reader is referred to the web

324 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333

325L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
of ammonites in the Presale section and (b) the existenceof another negative excursion in the Middle Aalenian(Sandoval et al., forthcoming) which is not clearlyidentified in the Bartolini and Cecca (1999).
The biostratigraphy presented by Cresta et al. (1995)is too broad for the Aalenian (Fallifax and TelegdirothiZones were identified, without more stratigraphicprecision, for the Middle to Upper Aalenian). Thiszonation was an informal subdivision and has proveddifficult to correlate to other Mediterranean areas. Forexample the Telegdirothi Zone was assigned to the LateAalenian, but in fact the zonal index (Riccardicerastelegdirothi) belongs to Discites Zone (Lower Bajocian)in well-documented Mediterranean areas (Cresta andGalácz, 1990; Linares and Sandoval, 1993, etc.)although this species does also occur in the uppermostpart of the Concavum Zone.
6. Ammonite turnovers events and sea levels
A detailed analysis of the diversity and faunal turnoverof ammonites from the Jurassic of Betic Cordillera haspreviously been published (O'Dogherty et al., 2000;Sandoval et al., 2001b, 2002b). In these works, ammonitediversity and faunal turnovers events were correlated withsea-level change curves (Haq et al., 1988; Hallam, 1989)and Tethyan eustatic cycles (Hardenbol et al., 1998).According to the extinction–radiation rates, six majorammonite turnover events in the interval studied here havebeen recorded (Sandoval et al., 2001b, 2002b) around (1)the Aalenian–Bajocian boundary (Concavum–DiscitesZones), (2) latest Early Bajocian–early Late Bajocian(Propinquans–Humphriesianum boundary; NiortenseZone respectively), (3) Bajocian–Bathonian boundary(Parkinsoni–Zigzag Zones), (4) early–Middle Bathonianboundary (Aurigerus–Sofanus Zones), (5) Middle–LateBathonian boundary (Costatus–Orbis Zones) and (6) LateBathonian–Early Callovian (Orbis to Gracilis Zones).
6.1. Turnover interval 1
One of the most important faunal turnover events inthe evolutionary history of Mesozoic ammonites tookplace within the Late Aalenian to earliest Bajocianinterval. At the subfamily level, 100% of the Mediter-ranean Ammonitina (five subfamilies) were extinct andsix new subfamilies originated before of the LaeviusculaZone (Fig. 5, turnover interval 1). This turnover gave
Fig. 7. Stratigraphic correlation of carbon isotope curves for the Middle JurUmbria basin, Italy (Bartolini et al., 1999); (D) Yorkshire, England (Hesselbo(F) Isle of Skye, Hebrides, Scotland (Jenkyns et al., 2002, and biostratigraph
rise to the origination and spreading of Stephanocer-atoidea and Haploceratoidea, which became the dom-inant ammonoids throughout the Middle Jurassic.Within the Discites Zone (Walkeri Subzone), extinctionrate was high (70% of the genera) and largely exceedednew appearances. The last Hildoceratoidea and most therepresentatives of the superfamily Hammatoceratoidea,which were the dominant Pliensbachian–Aalenianammonites, disappeared in the Walkeri Subzone. Thesubsequent radiation (important at the genus and specieslevels) expands throughout the Laeviuscula Zone. Thisturnover can be correlated with a Tethyan sequenceboundary (Bj1 of Hardenbol et al., 1998) near theAalenian–Bajocian boundary and a relative sea-levelrise in the Laeviuscula Zone (Sandoval et al., 2001b,2002b).
6.2. Turnover interval 2
During this major turnover, two phases can bedifferentiated. A minor (although important at genus orspecies level) ammonite turnover took place near theboundary between the Propinquans and Humphriesia-num Zones (Fig. 5, turnover interval 2a). The bradfor-diins, otoitids and alocolyceratids disappeared in theWestern Tethys, and oppeliids and nannolytoceratidsoriginated (Fig. 5). Extinction rates are especially highin the Hebridica Subzone. The subsequent radiation, inthe Romani Subzone, is accompanied by a strongrecovery of diversity and dominance of stephanoceratinsand oppelins (Figs. 5 and 6). Tethyan sequences(Hardenbol et al., 1998) show a regressive interval inthe Propinquans Zone, which is followed in theHumphriesianum Zone by a major transgressive se-quence that coincides with this turnover (Sandoval et al.,2001b, 2002b).
At the Early–Late Bajocian boundary a majorammonite turnover took place (Figs. 5(turnover interval2b) and 6). A relative decrease in the diversity at bothgenus and species levels in the Banksi and lower part ofthe Polygyralis Subzones followed by an increase in theupper part of Polygyralis and Baculata Subzones (Fig.5). Stephanoceratins together with the last Hammato-ceratoidea (sonniniids) became extinct, and the Peri-sphinctoidea, the most significant Middle–Late Jurassicammonites, originated. Likewise, stephanoceratids andhaploceratids underwent great changes with the extinc-tions and originations of important taxa. Five new
assic. (A) Subbetic, Spain (this paper); (B) Digne (Corbin, 1994); (C)et al., 2003); (E) Umbria basin, Italy (Bartolini and Cecca, 1999); andy according to Morton, 1990).

326 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
families or subfamilies originated (Fig. 5), representingmore than 50% of the Late Bajocian ammonites. Thisturnover correlates with a sequence boundary (Bj3 ofHardenbol et al., 1998) near the Lower–Upper Bajocianboundary and with the transgressive sequence of thePolygyralis and lower part of the Baculata Subzones(Sandoval et al., 2001b).
6.3. Turnover interval 3
The latest Bajocian has very low diversity, and manytypical Bajocian Mediterranean ammonites (sphaerocer-atins, garantianins and spiroceratins) went extinct; theleptosphinctins and strigoceratins were replaced by thezigzagiceratins and phlyctyceratins; likewise the firstmorphoceratins and hecticoceratins originated in thelatest Bajocian and later radiated in the Early Bathonian(Figs. 5(turnover interval 3) and 6). In the Mediterra-nean province a low stand (Bj4 in Hardenbol et al.,1998) is recorded in the upper part of the Garantiana andbase of the Parkinsoni Zones, where extinctionspredominate; the transgressive sequence is located inthe lower part of the Parkinsoni Zone coinciding withoccurrence of new taxa (Sandoval et al., 2001b).
6.4. Turnover interval 4
The extinction of parkinsoniids, morphoceratids,nannolytoceratids, some zigzagiceratins and manyPhylloceratina took place near the Early–Middle Bath-onian boundary, which was followed by the originationof tulitids and grossouvrins (Figs. 5(turnover interval 4)and 6). Tethyan sequences show a cycle boundary(Bat1) in the Aurigerus Zone (Postpollubrum Subzone)which cooccurs with last occurrences of many taxa(Sandoval et al., 2001b).
6.5. Turnover interval 5
Between the latest Middle Bathonian and the earliestLate Bathonian, cadomitins, diverse zigzagiceratins andmany tulitids, which were dominant in the MiddleBathonian, became extinct, while epistrenoceratins, andvarious Andean forms (eurycephalytins and neuqueni-ceratins) appeared (Figs. 5(faunal interval 5) and 6). Atthe genus and species levels, the extinctions wereespecially significant in the Suspensum Subzone, withthe consequent decrease in diversity during the LateBathonian (Fig. 5). The turnover co-occurred with athird-order cycle boundary (Bat4) in the upper part ofthe Costatus Zone and with the transgressive sequenceof the following cycle (Sandoval et al., 2001b).
6.6. Turnover interval 6
During the latest Bathonian, significant ammonitegenera went extinct and no major diversificationsoccurred, making ammonite diversity low during thistime interval (Fig. 5). Tethyan sequences show a cycleboundary (Bat5 of Hardenbol et al., 1998) coincidingwith extinctions. During the Early Callovian (Bullatusand lower part of the Gracilis Zone) the ammonites ofthe Western Tethys show a strong diversification,specially perisphinctids, tulitids and hecticoceratins;simultaneously the first records of macrocephalitids andreineckeiids took place, being dominant throughout theCallovian (Fig. 5(turnover interval6). Several Borealand Andean ammonites (kosmoceratids and eurycepha-lytins, respectively) also became diversified duringEarly Callovian. The first Callovian radiation co-occurred with a rapid relative sea-level rise in theExxon curve (Haq et al., 1988), which includes theBullatus and lower part of the Gracilis Zone (Sandovalet al., 2001b), this being equivalent to the transgressivesequence (Call2 in Hardenbol et al., 1998).
7. Major evolutionary events and the carbon-cycleperturbation
A major carbon cycle perturbation at the EarlyBajocian interval is recognized as a global phenomenonon the basis of its carbon–isotope expression in bothoceanic (Corbin, 1994; Bartolini et al., 1996; Bartoliniand Cecca, 1999; Jenkyns et al., 2002) and terrestrialreservoirs (Hesselbo et al., 2003). Large-scale and rapidpalaeoenvironmental changes are commonly associatedwith perturbations in the global carbon cycle.
The Early Bajocian positive excursion has beencorrelated in the southern margin of western Tethys witha “carbonate production crisis” concomitantly with theonset of biosiliceous sedimentation in several basins(Bartolini et al., 1996). Recently, Muttoni et al. (2005)explained the change from carbonate to chert and backto carbonate again in the western Tethys as being due toplate motions beneath a high productivity equatorialbelt. If this is the case, the increasing fertility during theMiddle Bajocian should have local causes (Equatorialupwelling). We believe that the observed link betweencarbon and silica cycles during the Middle Jurassicfavors a more global hypothesis to explain theradiolarite overspread and carbonate crises.
We remember that the “carbonate production crisis”is preceded at the Late Aalenian by a similar δ13Cexcursion toward light isotopic values (of about 2‰)in marine carbonates and terrestrial organic matter,

327L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
which implies a high CO2 level in the atmosphere forthis period. In such scenario the “carbonate productioncrisis” may be linked to perturbation of carbon cycle,due to important injections of CO2 associated with amajor plate tectonic reorganisation, like was theopening of the Pacific ocean, acceleration of breakupof Pangaea and major pulses of subduction-relatedmagmatism (Bartolini and Larson, 2001). High levelsof CO2 in the atmosphere could have accelerated thetransfer of nutrients from the continents to the oceans,through increasing weathering. The increasing valuesof δ13C in the Early Bajocian to a peak may be relatedto greater oceanic fertilisation, as shown by radiolarianassemblages (Bartolini et al., 1999). We cannotexclude the influence of upwelling, but in this casewe suggest an enhanced siliceous production, ratherthan passive mechanism as suggested by Muttoni et al.(2005). A major role in this scenario was played bythe opening of the Liguro-Piemontese Ocean, un-doubtedly causing a rearrangement of the oceaniccirculation patterns (Bill et al., 2001), which in turnraised eutrophication levels and induced an ecologicalcrisis of the main carbonate factories (Bartolini et al.,1996). In addition, the peculiar paleogeographicconfiguration of western Tethys as suggested by DeWever et al. (1994) together with an intensification ofgreenhouse conditions may have promoted strongerand larger upwellings zones. In this way we canconciliate the increasing fertility, high biosiliceous andcarbon burial (due to expansion of high productivityarea) with carbon isotope excursion. The Muttoni etal. (2005) interpretation is based on a pacificequatorial upwelling model, but the paleogeographicconfiguration of western Tethys had to be verydifferent from the large open Pacific, suggesting rathercoastal upwelling systems during the Middle Jurassic.
As discussed above, perturbations in the globalcarbon cycle are the final expression of large-scale andrapid palaeoenvironmental changes (Scholle and Arthur,1980; Weissert, 1989; Jenkyns et al., 2002). It is evidentthat most of organic crises occurring in the marine realmare linked with major palaeocenographic events (i.e.anoxia, changes in global circulation of oceaniccurrents, temperature changes, sea-level changes, mas-sive volcanism, etc.). Consequently, any change occur-ring in the biota must also be a proxy to help us in theinterpretation of the carbon cycle as expressed by theirstable isotopes. In fact, Margaritz (1989) was one of firstauthors to point out the relationships between δ13C andmajor extinctions in the marine biota. Later, differentauthors (Jiménez et al., 1996; Bartolini et al., 1999;Mattioli et al., 2004; Guex et al., 2004; Company et al.,
2005) have correlated different biotic crises (extinctions,faunal turnover, etc.) with anomalies in the δ13C isotopiccurve. Here we analyse the major events occurringduring the evolutionary history of Jurassic ammonites asa proxy to interpret the changes occurring in the marinebiosphere and the perturbations of the carbon cycleduring the Middle Jurassic.
7.1. The extinction of Early Jurassic ammonites and theMiddle Jurassic radiation: the Early Bajocian carboncycle perturbation
From the Upper Aalenian (upper part of theConcavum Subzone) a decreasing tendency is foundin the δ13C curve; with a minimum value of around1.5‰ at the Aalenian–Bajocian boundary (Limitatum–Walkeri Subzones). This δ13C minimum coincides withthe extinction of the last representatives of Hildocer-atoidea (one of the representative Early Jurassicammonite superfamilies) and their rapid replacementby Stephanoceratoidea and Haploceratoidea, whichrepresent the origin of the first Middle Jurassicammonite radiation, concomitant with increasing valuesin the δ13C curve. Many new genera and species occur(especially belonging to Stephanoceratoidea and Sonni-niidae) and diversity varies in parallel with the carbon–isotope values (Figs. 5 and 6). A maximum value (3–3.2‰) is reached in the Romani Subzone during thisEarly Bajocian carbon–isotope excursion.
From the upper part of the Romani to BlagdeniSubzone the δ13C curve returns to medium values andremains quite stable. In that interval the last two generadisplaying Early Jurassic characters disappear (repre-sented by the extinction of the last representatives oftypical Hammatoceratoidea). The extinction of theselatter ammonites and many Early Bajocian typicalStephanoceratoidea coincides with the origination ofthe large superfamily Perisphinctoidea. From thismoment, Stephanoceratoidea and, later on Perisphinc-toidea will be the dominant forms throughout the rest ofthe Jurassic. We can conclude that the major faunalturnover in the evolutionary history of Jurassic ammo-nites (replacement of the last Early Jurassic-type formsby Middle Jurassic ones) occurs simultaneously with theso-called “global carboncycle perturbation” whichoccurred during the Early Bajocian.
7.2. The Early Bajocian to Late Bajocian transition andthe first origination of Jurassic heteromorphs
From the topmost part of the Humphriesianum Zoneand throughout the Niortense Zone the carbon–isotope

328 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
shifts towards lighter values (from 2.6–2.9‰ to 1.4–1.8‰). Many ammonite genera, belonging mostly to theStephanoceratoidea, became extinct during this negativeexcursion (Figs. 5 and 6). This short excursion to lightervalues starts at the Blagdeni–Banski Subzones transitionwhen an important evolutionary process occurs: theorigination of extremely simple (proteromorphic) stepha-noceratids (Subcollina, Strenoceras, etc.) and includingthe origination of the heteromorphous genus Spiroceras.In our case the primitive evolute forms are represented bythe lineage formed by the genera Subcollina–Parastre-noceras–Strenoceras, giving rise to the uncoiled Spir-oceras. That lineage was probably derived from typicalStephanoceratidae such as Phaulostephanus (Sandovaland Westermann, 1986; Pavia, 2000).
The most common trends observed in ammonoidevolution during ecologically stable periods are charac-terized by an increase of shell curvature (e.g. evolute toinvolute) by the development of more complexornamentation (flexuosity of ribbing, appearance ofnodes and spines) and by a long term increase of thesuture line's fractal dimension (Guex, 1992, 2001).Major evolutionary jumps in ammonoids occur duringsevere extinction events, and are characterized by thesudden appearance of simple, primitive-looking formswhich are similar to remote ancestors of their morecomplex immediate progenitors. Such forms are oftenweakly ornamented and high evolute or even uncoiled,and we call them “proteromorphic” (Guex, 2001, 2006).The most striking example of homeomorphic groupsgenerated during such sublethal stress events isrepresented by the genera Ophiceras (earliest Triassic)and Psiloceras (earliest Jurassic). The Middle Jurassicuncoiled ammonites described as heteromorphs, such asSpiroceras and Parapatoceras, discussed above, arecompletely uncoiled. The change from coiled touncoiled morphology can be considered to be a proxyfor high environmental stress. In our case such stress canbe related to the important regressive phase and the shortδ13C excursion toward light isotopic values near theEarly–Late Bajocian boundary.
Except for a very weak trend toward decreasingvalues, no important fluctuations are observed in theδ13C curve from the Garantiana Zone to DaubenyiSubzone ((Figs. 2, 3 and 5)). From the DimorphusSubzone to Zigzag Zone the δ13C values tend to showhigher values, up to a relative maximum of 2.6‰. Ingeneral, no large isotopic fluctuations are detected andno very important changes occur in ammonite evolutionduring the interval spanning the Garantiana to ZigzagZones. The general diversity (at genus or species level)stays relatively low, although originations of new genera
dominate over extinction rates (Figs. 5 and 6). Only theprogressive domination of large and relatively involutespecies (sphaeroconid-type), together with a general sizeincreases of the ammonite shells are the most charac-teristic aspects during this interval. Such trends areconsidered characteristic of stable environmental con-ditions (Guex, 2001).
7.3. The Late Bathonian–Early Callovian immigrationsand the second origination of Jurassic heteromorphs
Through the Early–Middle Bathonian transition aprogressive dominance of Perisphinctoidea (formerlycalled Perisphinctaceae according to the new ICNZrules) over Stephanoceratoidea occurs achieved. Theextinction in the Mediterranean regions of mostStephanoceratoidea at the Zigzag and Aurigerus Zones(with only the exception of the “long-ranging” dimor-phous pair Cadomites–Polyplectites) coincides with aδ13C shift from 2.3‰ to 1.6‰ (Figs. 3 and 5). Therelative minimum δ13C value (1.6‰) is located withinthe Orbis Zone, an interval characterized by a stronglyregressive event. For the second time in the MiddleJurassic, the origination of a new uncoiled ammonitelineage (Parapatoceratidae) occurs in this zone. Theseextremely simple forms seem to have evolved rapidlyfrom the genus Cadomites via Hemigarantiana–Epis-trenoceras. Once again, these uncoiled forms rapidlyoriginate via extremely evolute forms during high-stressepisode (Guex, 1992, 2001). A second Middle Jurassicammonite radiation, defined by the appearance of thenew Perisphinctoidea, took place at the Middle–LateBathonian. From this moment and for the rest of theJurassic, this group is established as the dominant WestTethyan ammonites (Fig. 5).
The condensed character of the External Subbeticseries from the Upper Bathonian to Callovian is clearlynot accurate enough to allow a continuous isotopicstratigraphic record, but the general isotopic trend canbe traced. From the top of the Orbis Zone (probablyincluding the Discus Zone) to the middle part of theBullatus Zone the carbon isotopic ratio increases to arelative maximum (2.7‰). The most noticeable eventduring the time of this short Early Callovian positiveexcursion coincides with a relative sea-level rise and isrepresented by the immigration of the Andean-Mixtecanreineckeiid ammonites via the Hispanic Corridor. Thatgroup (Fig. 5) occurred in the Andes and Mexico beforeappearing in the Western Tethys (Cariou, 1984;Sandoval et al., 1990) and the macrocephalitids, whichhave no immediate ancestor in the Western Tethys,immigrated possibly from the East Indian Tethys to our

329L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
Mediterranean regions (Westermann and Callomon,1988).
8. Conclusions
• The well calibrated carbon–isotope curve drawn in theBetic Cordillera correlates well with those curvespreviously established in pelagic sequences of central-northern Italy and southern France, or with those basedon isotope data measured on belemnites from Scotlandand on analyses of fossil wood from NE England. Allthese curves indicate a significant negative excursionat the Aalenian–Bajocian, which is followed by apronounced positive excursion during the EarlyBajocian. Such wide correlation and similaritybetween terrestrial and marine carbon–isotope curvesis an indication that the observed isotopic signals mayhave recorded a global perturbation of the carbon cycleas stated by Hesselbo et al. (2003).
• An evident link is observed at the latest Aalenian–Early Bajocian times among (i) the similar δ13Cpositive excursion in marine carbonates and terres-trial organic matter, (ii) the “carbonate productivitycrisis”, (iii) the onset of biosiliceous sedimentationand (iv) the major faunal turnover occurring in theJurassic–Early Cretaceous Ammonoidea. All thisevidence strongly supports the idea that large-scaleand rapid palaeoenvironmental changes are com-monly associated with perturbations in the globalcarbon cycle.
• The two originations of the Middle Jurassic uncoiledammonites (Spiroceras in the middle part of thePolygiralis Subzone and Parapatoceras in the OrbisZone) were considered by Guex (1992, 2001) to beindicative of high environmental stress. In our study,we show that these originations coincide with rapidshifts to lighter values in the δ13C curve and that theyare associated with important ammonite extinctions.
• We found a good correlation of extinctions with δ13Cminima, and radiations with δ13C increasing. Suchcorrelation supports the idea that perturbations in theglobal carbon cycle reflect rapid palaeoenvironmentalchanges. Hence, our study reveals how major faunalturnover in the marine realm, as shown by ammoniteevolution, can be used as a proxy to identify majorpalaeoenvironmental crises of their ecosystems.
Acknowledgments
We would like to express our gratitude to Mr. D.Nesbbit for reviewing the English text. We thankFinn Surlyk, Stephen Hesselbo and an anonymous
reviewer for their comments on this manuscript. Thisstudy forms part of the results obtained in ResearchProjects: BTE 2001-3020, financed by the DGICT(Dirección General de Investigación Científica yTécnica, Spain), by the EMMI Research Group(Junta de Andalucía, Spain) and by the Project no.2000-052555 financed by the Swiss National ScienceFoundation.
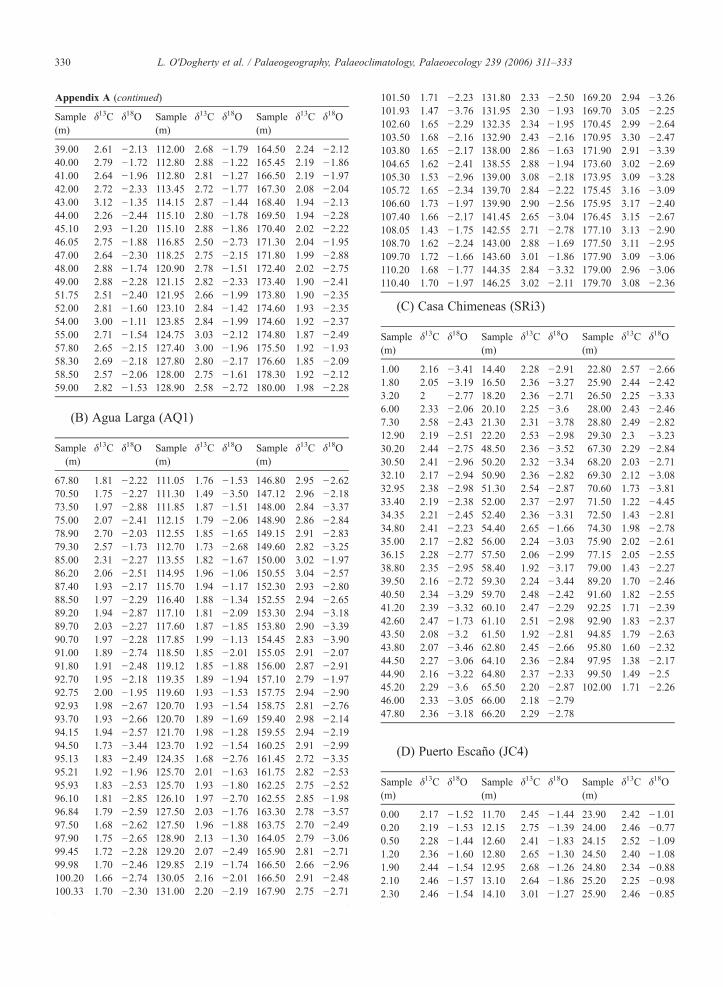
Appendix A. Carbon and oxygen isotopes of UpperAalenian–Lower Callovian bulk carbonate samplesfrom the Southern Iberian Palaeomargin
A. Casa Blanca (JCB)
Sample(m)
δ13C
δ18O Sample(m)δ13C
δ18O Sample(m)δ13C
δ18O0
2.03 −2.65 60.00 2.71 −1.71 129.30 2.81 −0.92 1.00 1.57 −3.29 60.80 2.37 −2.52 129.50 2.70 −1.49 2.20 2.09 −2.86 62.30 2.66 −2.02 130.00 2.77 −1.82 4.90 1.80 −2.95 63.00 2.51 −3.04 130.45 2.77 −1.63 5.85 2.02 −3.10 65.20 2.75 −1.74 130.80 2.65 −1.31 6.38 1.52 −3.56 66.05 2.74 −1.93 131.05 2.76 −1.45 7.00 1.77 −3.50 67.20 2.68 −2.30 131.85 2.64 −1.69 7.57 1.77 −3.08 68.10 2.95 −1.00 132.15 2.78 −1.51 7.60 1.80 −3.25 69.20 2.82 −1.19 132.50 2.59 −1.27 9.15 1.99 −3.22 70.00 2.64 −2.38 133.10 2.62 −1.17 11.20 2.19 −2.47 74.30 2.76 −1.69 133.50 2.78 −1.32 12.45 2.08 −3.47 75.35 2.78 −2.31 134.00 2.79 −1.09 13.15 2.44 −2.52 76.15 2.77 −2.83 135.00 2.94 −1.11 14.00 2.28 −2.94 77.15 2.95 −2.27 136.00 2.67 −1.32 15.60 2.58 −2.12 78.05 2.88 −1.96 136.50 2.76 −1.09 16.15 2.21 −2.85 79.10 3.07 −1.40 138.00 2.38 −1.29 16.90 2.39 −2.36 80.20 2.99 −1.65 139.00 2.25 −0.65 16.90 2.40 −2.47 81.10 2.75 −2.18 140.80 1.97 −2.05 18.00 2.31 −3.28 82.00 2.87 −2.17 142.45 1.86 −2.28 19.00 2.46 −2.94 83.00 2.86 −2.09 143.60 1.88 −2.32 20.20 2.58 −2.52 83.95 3.14 −1.17 144.40 2.08 −1.78 20.45 2.60 −2.53 85.80 3.21 −1.58 145.60 2.06 −1.81 21.00 2.44 −2.81 86.85 3.20 −2.01 146.45 2.18 −1.52 21.90 2.41 −2.45 87.00 3.29 −1.84 147.30 2.13 −1.79 23.40 2.36 −2.95 87.80 3.28 −2.25 148.40 2.18 −2.35 25.00 2.38 −2.85 89.85 3.35 −1.46 149.35 2.10 −2.32 27.42 2.49 −2.22 90.80 3.30 −1.57 150.40 2.18 −2.15 27.85 2.23 −2.50 91.55 3.17 −1.83 151.35 2.11 −2.02 29.20 2.44 −2.34 91.55 3.28 −1.90 152.50 2.08 −2.22 29.20 2.20 −2.61 92.75 3.25 −1.69 153.60 2.16 −1.86 30.00 2.44 −2.63 93.52 2.95 −1.83 154.30 2.15 −2.32 30.35 2.21 −2.59 94.00 2.90 −1.59 155.20 2.05 −1.97 31.10 2.70 −2.52 95.30 2.83 −1.92 156.40 2.11 −2.85 32.00 2.32 −2.62 100.00 2.75 −2.06 157.60 2.13 −1.96 33.00 2.22 −2.36 101.10 2.66 −1.87 158.30 2.11 −2.13 33.50 2.82 −1.96 101.80 2.67 −2.89 159.30 2.10 −2.32 34.80 2.72 −2.25 102.80 2.72 −1.84 160.40 2.13 −2.52 36.00 2.91 −1.39 102.80 2.69 −1.82 161.20 2.11 −2.20 37.10 2.55 −1.98 103.20 2.84 −1.82 162.40 2.18 −2.39 38.00 2.66 −2.29 107.00 2.44 −2.92 163.20 2.27 −2.19(continued on next page)
330 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
Appendix A (continued)
Sample(m)
δ13C
δ18O Sample(m)δ13C
δ18O Sample(m)δ13C
δ18O39.00
2.61 −2.13 112.00 2.68 −1.79 164.50 2.24 −2.12 40.00 2.79 −1.72 112.80 2.88 −1.22 165.45 2.19 −1.86 41.00 2.64 −1.96 112.80 2.81 −1.27 166.50 2.19 −1.97 42.00 2.72 −2.33 113.45 2.72 −1.77 167.30 2.08 −2.04 43.00 3.12 −1.35 114.15 2.87 −1.44 168.40 1.94 −2.13 44.00 2.26 −2.44 115.10 2.80 −1.78 169.50 1.94 −2.28 45.10 2.93 −1.20 115.10 2.88 −1.86 170.40 2.02 −2.22 46.05 2.75 −1.88 116.85 2.50 −2.73 171.30 2.04 −1.95 47.00 2.64 −2.30 118.25 2.75 −2.15 171.80 1.99 −2.88 48.00 2.88 −1.74 120.90 2.78 −1.51 172.40 2.02 −2.75 49.00 2.88 −2.28 121.15 2.82 −2.33 173.40 1.90 −2.41 51.75 2.51 −2.40 121.95 2.66 −1.99 173.80 1.90 −2.35 52.00 2.81 −1.60 123.10 2.84 −1.42 174.60 1.93 −2.35 54.00 3.00 −1.11 123.85 2.84 −1.99 174.60 1.92 −2.37 55.00 2.71 −1.54 124.75 3.03 −2.12 174.80 1.87 −2.49 57.80 2.65 −2.15 127.40 3.00 −1.96 175.50 1.92 −1.93 58.30 2.69 −2.18 127.80 2.80 −2.17 176.60 1.85 −2.09 58.50 2.57 −2.06 128.00 2.75 −1.61 178.30 1.92 −2.12 59.00 2.82 −1.53 128.90 2.58 −2.72 180.00 1.98 −2.28(B) Agua Larga (AQ1)
Sample(m)
δ13C
δ18O Sample(m)δ13C
δ18O Sample(m)δ13C
δ18O67.80
1.81 −2.22 111.05 1.76 −1.53 146.80 2.95 −2.62 70.50 1.75 −2.27 111.30 1.49 −3.50 147.12 2.96 −2.18 73.50 1.97 −2.88 111.85 1.87 −1.51 148.00 2.84 −3.37 75.00 2.07 −2.41 112.15 1.79 −2.06 148.90 2.86 −2.84 78.90 2.70 −2.03 112.55 1.85 −1.65 149.15 2.91 −2.83 79.30 2.57 −1.73 112.70 1.73 −2.68 149.60 2.82 −3.25 85.00 2.31 −2.27 113.55 1.82 −1.67 150.00 3.02 −1.97 86.20 2.06 −2.51 114.95 1.96 −1.06 150.55 3.04 −2.57 87.40 1.93 −2.17 115.70 1.94 −1.17 152.30 2.93 −2.80 88.50 1.97 −2.29 116.40 1.88 −1.34 152.55 2.94 −2.65 89.20 1.94 −2.87 117.10 1.81 −2.09 153.30 2.94 −3.18 89.70 2.03 −2.27 117.60 1.87 −1.85 153.80 2.90 −3.39 90.70 1.97 −2.28 117.85 1.99 −1.13 154.45 2.83 −3.90 91.00 1.89 −2.74 118.50 1.85 −2.01 155.05 2.91 −2.07 91.80 1.91 −2.48 119.12 1.85 −1.88 156.00 2.87 −2.91 92.70 1.95 −2.18 119.35 1.89 −1.94 157.10 2.79 −1.97 92.75 2.00 −1.95 119.60 1.93 −1.53 157.75 2.94 −2.90 92.93 1.98 −2.67 120.70 1.93 −1.54 158.75 2.81 −2.76 93.70 1.93 −2.66 120.70 1.89 −1.69 159.40 2.98 −2.14 94.15 1.94 −2.57 121.70 1.98 −1.28 159.55 2.94 −2.19 94.50 1.73 −3.44 123.70 1.92 −1.54 160.25 2.91 −2.99 95.13 1.83 −2.49 124.35 1.68 −2.76 161.45 2.72 −3.35 95.21 1.92 −1.96 125.70 2.01 −1.63 161.75 2.82 −2.53 95.93 1.83 −2.53 125.70 1.93 −1.80 162.25 2.75 −2.52 96.10 1.81 −2.85 126.10 1.97 −2.70 162.55 2.85 −1.98 96.84 1.79 −2.59 127.50 2.03 −1.76 163.30 2.78 −3.57 97.50 1.68 −2.62 127.50 1.96 −1.88 163.75 2.70 −2.49 97.90 1.75 −2.65 128.90 2.13 −1.30 164.05 2.79 −3.06 99.45 1.72 −2.28 129.20 2.07 −2.49 165.90 2.81 −2.71 99.98 1.70 −2.46 129.85 2.19 −1.74 166.50 2.66 −2.96 100.20 1.66 −2.74 130.05 2.16 −2.01 166.50 2.91 −2.48 100.33 1.70 −2.30 131.00 2.20 −2.19 167.90 2.75 −2.71 100.53 1.72 −2.03 131.45 2.22 −2.30 168.55 2.89 −3.16101.50
1.71 −2.23 131.80 2.33 −2.50 169.20 2.94 −3.26 101.93 1.47 −3.76 131.95 2.30 −1.93 169.70 3.05 −2.25 102.60 1.65 −2.29 132.35 2.34 −1.95 170.45 2.99 −2.64 103.50 1.68 −2.16 132.90 2.43 −2.16 170.95 3.30 −2.47 103.80 1.65 −2.17 138.00 2.86 −1.63 171.90 2.91 −3.39 104.65 1.62 −2.41 138.55 2.88 −1.94 173.60 3.02 −2.69 105.30 1.53 −2.96 139.00 3.08 −2.18 173.95 3.09 −3.28 105.72 1.65 −2.34 139.70 2.84 −2.22 175.45 3.16 −3.09 106.60 1.73 −1.97 139.90 2.90 −2.56 175.95 3.17 −2.40 107.40 1.66 −2.17 141.45 2.65 −3.04 176.45 3.15 −2.67 108.05 1.43 −1.75 142.55 2.71 −2.78 177.10 3.13 −2.90 108.70 1.62 −2.24 143.00 2.88 −1.69 177.50 3.11 −2.95 109.70 1.72 −1.66 143.60 3.01 −1.86 177.90 3.09 −3.06 110.20 1.68 −1.77 144.35 2.84 −3.32 179.00 2.96 −3.06 110.40 1.70 −1.97 146.25 3.02 −2.11 179.70 3.08 −2.36(C) Casa Chimeneas (SRi3)
Sample(m)
δ13C
δ18O Sample(m)δ13C
δ18O Sample(m)δ13C
δ18O1.00
2.16 −3.41 14.40 2.28 −2.91 22.80 2.57 −2.66 1.80 2.05 −3.19 16.50 2.36 −3.27 25.90 2.44 −2.42 3.20 2 −2.77 18.20 2.36 −2.71 26.50 2.25 −3.33 6.00 2.33 −2.06 20.10 2.25 −3.6 28.00 2.43 −2.46 7.30 2.58 −2.43 21.30 2.31 −3.78 28.80 2.49 −2.82 12.90 2.19 −2.51 22.20 2.53 −2.98 29.30 2.3 −3.23 30.20 2.44 −2.75 48.50 2.36 −3.52 67.30 2.29 −2.84 30.50 2.41 −2.96 50.20 2.32 −3.34 68.20 2.03 −2.71 32.10 2.17 −2.94 50.90 2.36 −2.82 69.30 2.12 −3.08 32.95 2.38 −2.98 51.30 2.54 −2.87 70.60 1.73 −3.81 33.40 2.19 −2.38 52.00 2.37 −2.97 71.50 1.22 −4.45 34.35 2.21 −2.45 52.40 2.36 −3.31 72.50 1.43 −2.81 34.80 2.41 −2.23 54.40 2.65 −1.66 74.30 1.98 −2.78 35.00 2.17 −2.82 56.00 2.24 −3.03 75.90 2.02 −2.61 36.15 2.28 −2.77 57.50 2.06 −2.99 77.15 2.05 −2.55 38.80 2.35 −2.95 58.40 1.92 −3.17 79.00 1.43 −2.27 39.50 2.16 −2.72 59.30 2.24 −3.44 89.20 1.70 −2.46 40.50 2.34 −3.29 59.70 2.48 −2.42 91.60 1.82 −2.55 41.20 2.39 −3.32 60.10 2.47 −2.29 92.25 1.71 −2.39 42.60 2.47 −1.73 61.10 2.51 −2.98 92.90 1.83 −2.37 43.50 2.08 −3.2 61.50 1.92 −2.81 94.85 1.79 −2.63 43.80 2.07 −3.46 62.80 2.45 −2.66 95.80 1.60 −2.32 44.50 2.27 −3.06 64.10 2.36 −2.84 97.95 1.38 −2.17 44.90 2.16 −3.22 64.80 2.37 −2.33 99.50 1.49 −2.5 45.20 2.29 −3.6 65.50 2.20 −2.87 102.00 1.71 −2.26 46.00 2.33 −3.05 66.00 2.18 −2.79 47.80 2.36 −3.18 66.20 2.29 −2.78(D) Puerto Escaño (JC4)
Sample(m)
δ13C
δ18O Sample(m)δ13C
δ18O Sample(m)δ13C
δ18O0.00
2.17 −1.52 11.70 2.45 −1.44 23.90 2.42 −1.01 0.20 2.19 −1.53 12.15 2.75 −1.39 24.00 2.46 −0.77 0.50 2.28 −1.44 12.60 2.41 −1.83 24.15 2.52 −1.09 1.20 2.36 −1.60 12.80 2.65 −1.30 24.50 2.40 −1.08 1.90 2.44 −1.54 12.95 2.68 −1.26 24.80 2.34 −0.88 2.10 2.46 −1.57 13.10 2.64 −1.86 25.20 2.25 −0.98 2.30 2.46 −1.54 14.10 3.01 −1.27 25.90 2.46 −0.85
331L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
2.85
2.44 −2.05 14.50 3.03 −1.61 26.00 2.34 −0.92 2.85 2.40 −1.60 14.95 3.05 −1.62 26.40 2.24 −1.24 2.95 2.52 −1.40 15.10 3.10 −1.65 26.80 2.25 −0.63 3.45 2.28 −1.67 15.80 3.08 −1.48 27.70 2.17 −1.03 3.75 2.45 −1.32 16.45 3.13 −1.50 28.40 2.09 −1.64 3.80 2.44 −1.42 16.62 3.06 −1.41 28.70 2.10 −1.84 4.25 2.55 −1.33 16.80 3.19 −1.21 28.90 2.23 −0.37 4.50 2.58 −1.43 17.45 3.13 −1.23 29.40 2.22 −1.32 4.90 2.53 −1.44 17.50 3.08 −1.17 29.80 2.40 −1.06 5.05 2.64 −1.21 18.00 3.17 −0.92 30.00 2.29 −0.89 5.38 2.58 −1.41 18.15 3.03 −1.12 30.30 2.41 −1.88 5.65 2.42 −1.78 18.35 2.91 −1.37 30.60 2.50 −1.60 5.86 2.53 −1.35 18.80 2.90 −1.34 30.75 2.38 −1.53 6.00 2.53 −1.31 19.00 2.43 −1.57 31.00 2.53 −0.73 6.60 2.46 −1.36 19.45 2.84 −0.70 31.75 2.44 −0.57 7.15 2.48 −1.45 19.55 2.95 −0.86 32.00 2.47 −1.25 7.50 2.46 −1.57 20.15 3.01 −0.90 32.40 2.35 −1.14 7.95 2.58 −1.39 20.60 2.96 −0.89 33.30 2.42 −1.38 8.40 2.57 −1.88 21.40 2.61 −1.18 33.50 2.34 −1.00 8.90 2.55 −1.43 21.60 2.47 −1.29 33.75 2.27 −1.24 9.45 2.95 −0.23 22.10 2.36 −1.53 34.00 2.40 −1.02 9.50 2.66 −1.90 22.20 1.90 −2.14 34.40 2.38 −1.08 9.80 2.62 −1.72 22.30 2.19 −1.37 34.45 2.33 −1.15 10.55 2.73 −1.80 22.60 2.43 −1.14 35.65 2.41 −0.68 10.95 2.71 −1.82 22.75 2.39 −1.44 35.80 2.20 −0.07 11.35 2.40 −1.42 23.90 2.39 −0.46(E) La Cornicabra (JV1)
Sample(m)
δ13C
δ18O Sample(m)δ13C
δ18O Sample(m)δ13C
δ18O0.20
1.71 −0.28 9.90 2.16 −0.40 18.90 2.14 0.21 0.60 1.83 −0.51 10.50 2.22 −0.25 19.25 2.08 0.09 1.20 1.85 −0.59 11.30 2.24 −0.36 19.60 1.99 0.02 1.70 1.88 −0.33 12.10 2.27 −0.31 20.00 2.00 0.10 1.90 1.90 −0.65 12.60 2.25 −0.28 20.45 1.87 −0.05 2.10 1.95 −0.81 13.20 2.29 −0.29 20.80 1.87 −0.09 2.30 1.96 −0.48 14.30 2.25 0.01 21.15 1.84 0.17 3.25 1.96 −1.27 15.50 2.16 0.03 21.80 1.81 0.10 3.85 1.85 −1.96 16.00 2.26 0.14 22.25 1.92 −0.02 4.85 2.04 −0.20 16.30 2.27 0.15 22.50 2.11 −0.08 5.25 1.92 −0.46 16.60 2.30 0.00 23.00 2.30 0.12 5.80 2.13 −0.19 16.90 2.28 0.22 23.40 2.63 0.42 6.40 2.07 −0.35 17.30 2.32 0.13 24.20 2.34 0.38 6.70 2.04 −0.52 17.90 2.29 0.00 24.70 2.35 0.20 7.60 2.06 −0.38 18.30 2.20 −0.12 24.95 1.77 0.20 8.60 2.10 −0.20 18.40 2.21 0.04 26.25 2.24 0.27 9.20 2.18 −0.16 18.60 2.18 −0.10References
Arias, C., Castro, J.M., Chacón, B., Company, M., Diaz deFederico, A., Estévez, A., Fernández, J., García-Hernández,M., de Gea, G.A., López-Garrido, A.C., Martín-Algarra, A.,Martín-Chivelet, J., Molina, J.M., Morata, D., Nieto, L.M.,O'Dogherty, L., Pérez-López, A., Puga, E., Rey, J., Rivas, P.,Ruiz-Ortiz, P., Sandoval, J., Sanz de Galdeano, C., Vera, J.A.,Vilas, L., 2004. Las Zonas Externas Béticas. In: Vera, J.A. (Ed.),Geología de España. Sociedad Geológica de España, Ministerio
de Educación y Ciencia, Instituto Geológico y Minero deEspaña, Madrid, pp. 354–395.
Aurell, M., Meléndez, G., Olóriz, F., Bádenas, B., Caracuel, J., García-Ramos, J.C., Goy, Linares, A., Quesada, S., Robles, S., Rodríguez-Tovar, F.J., Rosales, I., Sandoval, J., Suarez de Centi, C., Tavera, J.M., Valenzuela, M., 2002. Jurassic. In: Gibbsons, W., Moreno, T.(Eds.), The Geology of Spain. The Geological Society, London,pp. 213–253.
Bartolini, A., Cecca, F., 1999. 20 My hiatus in the Jurassic of Umbria-Marche Apennines (Italy): carbonate crisis due to eutrophication.Comptes Rendus de l'Académie des Sciences de Paris. Sciences dela Terre et des Planétes 329, 587–595.
Bartolini, A., Larson, R., 2001. The Pacific microplate and the Pangeasupercontinent in the Early-Middle Jurassic. Geology 29,735–738.
Bartolini, A., Baumgartner, P.O., Hunziker, J.C., 1996. Middle andLate Jurassic carbon stable-isotope stratigraphy and radiolaritesedimentation of the Umbria-Marche Basin (Central Italy).Eclogae Geologicae Helvetiae 89, 811–844.
Bartolini, A., Morettini, E., O'Dogherty, L., Sandoval, J., Baumgart-ner, P.O., Hunziker, J.C., 1997. Carixian-Bajocian carbon stablestratigraphy of the Umbria-Marche (central Italy) and Subbetic(Spain) areas. EUG 9, Abstr. Suppl. 1. Terra Nova, vol. 9, p. 403.
Bartolini, A., Sandoval, J., O'Dogherty, L., Morettini, E., Baumgart-ner, P.O., Hunziker, J.C., 1998. Middle Jurassic carbon isotopestratigraphy of the Subbetic Domain correlated with ammonite andradiolarian zones. 5th International Symposium on the JurassicSystem. Vancouver, Canada, pp. 7–8.
Bartolini, A., Baumgartner, P.O., Guex, J., 1999. Middle and Lateradiolarian palaeoecology versus carbo-isotope stratigraphy.Palaeogeography, Palaeoclimatology, Palaeoecology 145, 43–60.
Baumgartner, P.O., O'Dogherty, L., Gorican, S., Urquhart, E.,Pillevuit, A., De Wever, P. (Eds.), 1995. Middle Jurassic toLower Cretaceous Radiolaria of Tethys: Occurrences, Systematics,Biochronology. Mémoires de Géologie, Lausanne, vol. 23, i–xxix,pp. 1–1172.
Bill, M., O'Dogherty, L., Guex, J., Baumgartner, P.O., Masson, H.,2001. Radiolarite ages in Alpine-Mediterranean ophiolites: con-straints of the continental breakup and the Tethys-Atlanticconnection. Bulletin of the Geological Society of America 113(1), 129–143.
Cariou, E., 1984. Structure, origine et paleobiogéographie de la familleReineckeiidae, Ammonitina, du Jurassique moyen. Comptesrendus de l'Academie des Sciences de Paris 298 (II), 245–248.
Company, M., Aguado, R., Jiménez de Cisneros, C., Sandoval, J.,Tavera, J.M., Vera, J.A., 2005. Biotic changes linked to a minoranoxic event (Faraoni level, Latest Hauterivian, Early Cretaceous).Palaeogeography, Palaeoclimatology, Palaeoecology 224,186–199.
Coplen, T.B., Kendall, C., Hopple, J., 1983. Comparison of stableisotope reference samples. Nature 302, 236–238.
Corbin, J.C., 1994. Evolution géochimique du Jurassique du Sud-estde la France: influence des variations du niveau marin et de latectonique. PhD thesis, Paris VI Univ., France.
Cresta, S., Galácz, A., 1990. Mediterranean basal Bajocian ammonitefaunas. Examples from Hungary and Italy. Memorie Descrittivedella Carta Geologica d'Italia 40, 165–197.
Cresta, S., Basilici, G., Bussoletti, A., Comas, M.J., Elmi, S.,Fernández-López, S., Ferretti, A., Goy, A., Henriques, M.H.,Kalin, O., Jiménez, A., Martínez, G., Sandoval, J., Ureta, S.,Venturi, F., 1995. Relazione sul Workshop “Paleontologia eBiostratigrafia dei sedimenti Toarciano-Bajociani in successioni di

332 L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
alto morfostructturale dell'Appennino Umbro-Marchigiano: l'areadi Monte Nerone. Correlazioni con altre regioni della TetideOccidentale”. (Piobbico, 2-14 agosto 1993). Bollettino delServizio Geologico d'Italia 112, 129–139.
Delgado, F., Linares, A., Sandoval, J., Vera, J.A., 1980. Contribution al'étude de l'Ammonitico Rosso du Dogger dans la ZoneSubbétique. In: Farinacci, A., Elmi, S. (Eds.), Rosso AmmoniticoSymposium Proceedings. Tecnoscienza, Roma, pp. 181–197.
De Wever, P., Azema, J., Fourcade, E., 1994. Radiolarians andradiolarite: primary production, diagenesis and paleogeography.Bulletin des Centres de Recherche et Exploration-Productiond'Elf-Aquitaine 18, 315–379.
García-Hernández, M., López-Garrido, A.C., Martín-Algarra, A.,Molina, J.M., Ruiz-Ortiz, P.A., Vera, J.A., 1989. Las discontinui-dades mayores del Jurásico de las Zonas Externas de lasCordilleras Béticas: Análisis e interpretación de los ciclossedimentarios. Cuadernos de Geología Ibérica 13, 35–52.
Guex, J., 1992. Origine des sauts évolutives chez les ammonites.Bulletin de Géologie Lausanne 316, 117–144.
Guex, J., 2001. Environmental stress and atavism in ammonoidevolution. Eclogae Geologicae Helvetiae 94, 321–328.
Guex, J., 2006. Reinitialization of evolutionary clocks during sublethalenvironmental stress in some invertebrates. Earth and PlanetaryScience Letters 242, 240–253.
Guex, J., Bartolini, A., Atudorei, V., Taylor, D., 2004. High-resolutionammonite and carbon isotope stratigraphy across the Triassic-Jurassic boundary at New Cork Canyon (Nevada). Earth andPlanetary Science Letters 225, 29–41.
Hallam, A., 1987. Radiations and extinctions in relation toenvironmental change in the marine Lower Jurassic of northwestEurope. Paleobiology 13, 152–168.
Hallam, A., 1989. The case for sea-level change as a dominantcausal factor in mass extinction of marine invertebrates.Philosophical Transactions of the Royal Society of London. B325, 437–455.
Hallam, A., 1990. Biotic and abiotic factors in the evolutions of earlymesozoic marine mollusks. In: Ross, R.M., Allmon, W.D. (Eds.),Cause of Evolution. A Paleontological Perspective. UniversityChicago Press, Chicago, pp. 249–260.
Hallam, A., Wignall, P.B., 1997. Mass Extinctions and theirAftermath. Oxford University Press, Oxford.
Hallam, A., Wignall, P.B., 1999. Mass Extinctions and sea-levelchanges. Earth-Science Reviews 48, 217–250.
Haq, B.U., Hardenbol, J., Vail, P.R., 1988. Mesozoic and Cenozoicchronostratigraphy and cycles of sea-level change. In: Wilgus,C.K., et al. (Eds.), Sea-level Changes: An Integrated Approach.S.E.P.M. Special Publication, vol. 42, pp. 71–108.
Hardenbol, J., Thierry, J., Farley, M.B., Jacquin, T., De Graciansky, P.C., Vail, P.R., 1998. Mesozoic and Cenozoic sequence chronos-tratigraphic framework of European basins. Jurassic sequencechronostratigraphy. In: De Graciansky, P.C., Hardenbol, J.,Jacquin, J., Vail, P.R. (Eds.), Mesozoic and Cenozoic sequencestratigraphy of European Basins. S.E.P.M. Special Publication, vol.60. chart 6.
Hesselbo, S.P., Morgans-Bell, H.S., McElwain, J.C., Rees, P.M.,Robinson, S.A., Ross, C.E., 2003. Carbon-Cycle Perturbation inthe Middle Jurassic and accompanying changes in the terrestrialpaleoenvironment. Journal of Geology 111, 259–276.
Jenkyns, H.C., Jones, C.E., Gröcke, D.R., Hesselbo, S.P., Parkinson,D.N., 2002. Chemostratigraphy of the Jurassic System: applica-tions, limitations and implications for palaeoceanography. Journalof the Geological Society (London) 159, 351–378.
Jiménez, A.P., Jiménez de Cisneros, C., Rivas, P., Vera, J.A., 1996.The Early Toarcian anoxic event in the westernmost Tethys(Subbetic): paleogeographic and paleobiogeographic significance.Journal of Geology 10, 399–416.
Linares, A., Sandoval, J., 1977. Contribución al estudio del Bajocensede la Sierra de Ricote (Murcia). Cuadernos de Geología-Universidad de Granada 8, 277–312.
Linares, A., Sandoval, J., 1990. The lower boundary of the Bajocian inthe “Barranco de Agua Larga” section, (Subbetic Domain,Southern Spain). Memorie Descrittive della Carta Geologicad'Italia 40, 13–22.
Linares, A., Sandoval, J., 1993. El Aaleniense de la Cordillera Bética(Sur de España). Análisis bioestratigráfico y caracterizaciónpaleobiogeográfica. Revista de la Sociedad geológica de España6, 177–206.
Linares, A., Sandoval, J., 1996. The genus Haplopleuroceras(Erycitidae, Ammonitina) in the Betic Cordillera, SouthernSpain. Geobios 29, 287–305.
Linares, A., Olóriz, F., Sandoval, J., Tavera, J.M., 1979. El Bajocensesuperior y el Bathonense en la serie de la Cornicabra (Prov. deJaén, sector central de las Cordilleras Béticas). Cuadernos deGeología-Universidad de Granada 10, 253–266.
Margaritz, M., 1989. Delta13 C minima follow extinction events; aclue to faunal radiation. Geology 17, 337–340.
Mattioli, E., Pittet, B., Bucefalo Palliani, R., Rohl, H.J., Schmid-Rohl,A., Morettini, E., 2004. Phytoplankton evidence for the timing andcorrelation of palaeoceanographical changes during the earlyToarcian oceanic anoxic event (Early Jurassic). Journal of theGeological Society (London) 161, 685–693.
McCrea, J.M., 1950. On isotopic chemistry of carbonates andpaleotemperature scale. Journal of Chemical Physics 18, 849–857.
Morton, N., 1990. Bearreraig (Isle of Skye, NW Scotland) as boundarystratotype for the base of the Bajocian Stage. Memorie Descrittivedella Carta Geologica d'Italia 40, 23–48.
Muttoni, G., Erba, E., Kent, D.V., Bachtadse, V., 2005. Mesozoicalpine facies deposition as a result of past latitudinal plate motion.Nature 434, 59–63.
O'Dogherty, L., Baumgartner, P.O., Sandoval, J., Martín-Algarra, A.,Pillevuit, A., 1995. Middle and Upper Jurassic radiolarianassemblages co-occurring with ammonites from the SubbeticRealm (Southern Spain). In: Baumgartner, P.O., O'Dogherty, L., etal. (Eds.), Middle Jurassic to Lower Cretaceous Radiolaria ofTethys: Occurrences, Systematics, Biochronology. Mémoires deGéologie, Lausanne, vol. 23, pp. 717–724.
O'Dogherty, L., Sandoval, J., Vera, J.A., 2000. Ammonite faunalturnover tracing sea level changes during the Jurassic (BeticCordillera, southern Spain). Journal of the Geological Society(London) 157, 723–736.
O'Dogherty, L., Nieto, L.M., Sandoval, J., 2001. Depósitos de brechasintraformacionales y slumps en el Jurásico Medio del SubbéticoMedio oriental (Sierra de Ricote, provincia de Murcia). Geotemas3, 249–253.
Olóriz, F., Rivas, P., Tavera, J.M., 1979. La secuencia Jurásica de CasaBlanca: atención especial al Dogger-Malm. Cuadernos deGeología-Universidad de Granada 10, 605–616.
Pavia, G., 2000. New Subcollina (Ammonitida) from the TopmostLower Bajocian: their Phylogenetic and Paleogeographic Signif-icance. GeoResearch Forum 6, 397–406.
Sandoval, J., 1983. Bioestratigrafía y Paleontología (Stephanoceratoi-dea y Perisphinctoidea) del Bajocense y Bathonense de lasCordilleras Béticas. PhD thesis, Servicio de Publicaciones, Univ.Granada, Granada, Spain.

333L. O'Dogherty et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 239 (2006) 311–333
Sandoval, J., 1986. Middle Jurassic Haploceratidae (Ammonitina)from the Subbetic Zone (South Spain). Geobios 19, 435–463.
Sandoval, J., 1990. A revision of the Bajocian divisions in the SubbeticDomain (Southern Spain). Memorie Descrittive della CartaGeologica d'Italia 40, 141–162.
Sandoval, J., Chandler, R.B., 2000. The Sonninniid ammoniteEuhoploceras from the Middle Jurassic of south-west Englandand southern Spain. Palaeontology 43, 495–533.
Sandoval, J., Westermann, G.E.G., 1986. The Bajocian (Jurassic)ammonite fauna of Oaxaca, Mexico. Journal of Paleontology 60,1220–1271.
Sandoval, J., Westermann, G.E.G., Marshall, M.C., 1990. Ammonitefauna, stratigraphy and ecology of the Bathonian–Callovian(Jurassic) Tecocoyunca group, South Mexico. PalaeontographicaAbteilung A 210, 93–149.
Sandoval, J., Henriques, M.H., Ureta, M.S., Goy, A., Rivas, P., 2001a.The Lias/Dogger boundary in Iberia: Betic and Iberian cordillerasand Lusitanian basin. Bulletin de la Société Géologique de France172, 18–21.
Sandoval, J., O'Dogherty, L., Guex, J., 2001b. Evolutionary rates ofJurassic ammonites in relation to sea-level fluctuations. Palaios 16,330–363.
Sandoval, J., Aguado, R., O`Dogherty, L., Bartolimi, A., 2002a.Biostratigraphic events at the Lower–Upper Bajocian boundary inthe Subbetic (Southern Spain). 6th International Symposyum onthe Jurassic System. Mondello, Italy, p. 159.
Sandoval, J., O'Dogherty, L., Vera, J.A., Guex, J., 2002b. Sea levelchanges and ammonite faunal turnover during the Lias/Doggertransition in the western Tethys. Bulletin de la Société Géologiquede France 173, 57–66.
Sandoval, J., Bill, M., O'Dogherty, L., 2004. Carbon isotopestratigraphy and ammonite faunal turnover for the Middle Jurassicin the South Iberian Margin. 24rd IAS Meeting, Coimbra,Portugal, p. 240.
Scholle, P.A., Arthur, M.A., 1980. Carbon isotope fluctuations inCretaceous pelagic limestones; potential stratigraphic and petro-leum exploration tool. AAPG Bulletin 64 (1), 67–87.
Sequeiros, L., 1979. El Calloviense en la Zona Subbética. Cuadernosde Geología-Universidad de Granada 10, 452–462.
Spötl, C.V., Vennemann, T.W., 2003. Continuous-flow isotope ratiomass spectrometric analysis of carbonate minerals. Rapid Com-munications in Mass Spectrometry 17, 1004–1006.
Steiner, M., Ogg, J., Sandoval, J., 1987. Jurassic magnetostratigraphy,3. Bathonian-Bajocian of Carcabuey, Sierra Harana and Campillode Arenas (Subbetic Cordillera, southern Spain). Earth andPlanetary Science Letters 82, 357–372.
Swart, P.J., Burns, S.K., Leder, S.J., 1991. Fractionation of the stableisotopes of oxygen and carbon dioxide during the reaction ofcalcite with phosphoric acid as a function of temperature andtechnique. Chemical Geology 86, 89–96.
Weissert, H., 1989. C-isotope stratigraphy, a monitor of paleoenviron-mental change; a case study from the Early Cretaceous. Surveys inGeophysics 10, 1–61.
Westermann, G.E.G., Callomon, J.H., 1988. The macrocephalitinaeand associated Bathonian and early Callovian (Jurassic) Ammo-noids of the Sula Islands and New Guinea. PalaeontographicaAbteilung A 140, 1–116.
Wignall, P.B., 2004a. Causes of mass extinctions. In: Taylor, P.D.(Ed.), Extinction in the History of Life. Cambridge UniversityPress, Cambridge, pp. 119–150.
Wignall, P.B., 2004b. Biotic Recovery After Mass Extinction.McGraw-Hill 2004, Yearbook of Science and Technology.McGraw-Hill Professional, New York.
Zempolich, W.G., Erba, E., 1999. Sedimentologic and chemostrati-graphic recognition of third-order sequences in resedimentedcarbonates: the Middle Jurassic Vajont Limestone. In: Harris, P.M.,Seller, A.H., Simo, J.A. (Eds.), Advances in Carbonate SequencesStratigraphy: Application to Reservoirs, Outcrops and Models. S.E.P.M. Special Publication, vol. 63, pp. 355–370.




















