
Rugose corals at the Tournaisian–Viséan transition in the Central Taurides (S Turkey) –...
28
Rugose corals at the Tournaisian–Viséan transition in the Central Taurides (S Turkey) – Palaeobiogeography and palaeoceanography of the Asian Gondwana margin Julien Denayer ⇑,1 Animal and Human Palaeontology, Geology Department, University of Liège, Belgium article info Article history: Received 11 March 2014 Received in revised form 10 June 2014 Accepted 2 November 2014 Available online 22 November 2014 Keywords: Rugose corals Kueichouphyllum Palaeogeography Cimmerian Terrane Systematics abstract This paper gives the first taxonomic description of the Upper Tournaisian–Lower Viséan rugose coral fauna of the Yaricak Formation (Aladag Unit, Central Taurides, South Turkey). Fifteen species belonging to twelve genera were identified, one species is newly described: Eokoninckocarinia gemmina. The corals are stratigraphically distributed in four assemblages. The two typical assemblages of the Upper Tournai- sian are composed of widely distributed taxa (Uralinia, Caninia, Proheterelasma, Zaphrentites). The assem- blage crossing the Tournaisian–Viséan boundary is characterized by Eurasian and cosmopolitan and widely distributed taxa (Calmiussiphyllum, Siphonophyllia, Bifossularia Amygdalophyllum, Caninophyllum, Keyserlingophyllum) and Asian taxa (Kueichouphyllum). The youngest assemblage, dominated by Eokon- inckocarinia gemmina sp. nov., has yielded foraminifers Moliniacian (Lower Viséan) in age. These assem- blages form a low diversity level-bottom community which is typical of the South Palaeotethys ‘Kueichouphyllum Zone’ extending along the Asian margin of Gondwana (Cimmerian Terrane) during Lower Carboniferous times. As in the other Cimmerian blocks, all the corals are solitary and colonial taxa are virtually absent. This absence is tentatively explained by the high palaeolatitude (c. 50°S) position of the Cimmerian Terrane in the southern part of the Palaeotethys Ocean for this time slice. A cold-water palaeo-current running eastward along the Gondwana margin might also be considered as it possibly could explain the wide distribution of the Kueichouphyllum fauna, restricted east of Africa in the southern coast of the Palaeotethys. Palaeoceanography, palaeoclimate and facies issues are discussed as possible causes of the diversity gradient observed between the eastern (Australia, Malaya) and the western (North Africa) margin of the Gondwana. Ó 2014 Elsevier Ltd. All rights reserved. 1. Introduction The Late Tournaisian and Early Viséan times were a period of important change in the geobiosphere. The Devonian greenhouse climate switched to icehouse as the Late Palaeozoic glaciation started (see Fielding et al., 2008; Caputo et al., 2008 for summary); the tectonic movements of the forthcoming Variscan Orogeny modified the continental and oceanic settings around the Palaeot- ethys (e.g. Stampfli and Borel, 2002; Nance et al., 2010); the marine fauna slowly recovered after the crisis associated with the Devo- nian–Carboniferous boundary. A high relative sea level at this time drove a good communication between provinces and allows the dispersion of marine fauna, making them cosmopolitan during the uppermost Tournaisian Avins diversification event (Poty, 2007). In contrast, the earliest Viséan recorded a very low sea level and during this period, the marine basins were more isolated and consequently, the organisms – and particularly the rugose corals – became more endemic (Aretz et al., 2014). If the coral fauna of the Tournaisian–Viséan transition are well known in Western Europe (Poty, 1989, 2007; Somerville et al., 1989; Weyer, 1993), Eastern Europe (Vassiljuk, 1960; Dobroljubova et al., 1966; Poty et al., 2003) and China (Yü, 1933; Poty and Xu, 1996; Xu and Poty, 1997), those of the Middle East and Central Asia are almost unknown. With the exception of some works in Iran (Douglas, 1950; Flügel, 1963, 1991; Khaksar, 1996) and China (Fan et al., 2003), the Carboniferous fauna of the Asian margin of Gondwana is Terra Incognita. The term ‘Asian’ margin is here used instead of ‘northern’ margin as the Gondwana was centred at this time on the South Pole and its entire margin was consequently ‘northern’. This paper aims to summarize the current knowledge of these http://dx.doi.org/10.1016/j.jseaes.2014.11.008 1367-9120/Ó 2014 Elsevier Ltd. All rights reserved. ⇑ Address: Service de Paléontologie animale et humaine, Départememt de Géologie, B18, Allée du Six-Août, Sart Tilman, B-4000 Liège, Belgium. Tel.: + 32 43663248. E-mail address: [email protected] 1 Current address: School of Earth Sciences, University of Queensland, Australia. Journal of Asian Earth Sciences 98 (2015) 371–398 Contents lists available at ScienceDirect Journal of Asian Earth Sciences journal homepage: www.elsevier.com/locate/jseaes
Transcript of Rugose corals at the Tournaisian–Viséan transition in the Central Taurides (S Turkey) –...

Journal of Asian Earth Sciences 98 (2015) 371–398
Contents lists available at ScienceDirect
Journal of Asian Earth Sciences
journal homepage: www.elsevier .com/locate / jseaes
Rugose corals at the Tournaisian–Viséan transition in the CentralTaurides (S Turkey) – Palaeobiogeography and palaeoceanography of theAsian Gondwana margin
http://dx.doi.org/10.1016/j.jseaes.2014.11.0081367-9120/� 2014 Elsevier Ltd. All rights reserved.
⇑ Address: Service de Paléontologie animale et humaine, Départememt deGéologie, B18, Allée du Six-Août, Sart Tilman, B-4000 Liège, Belgium. Tel.: + 3243663248.
E-mail address: [email protected] Current address: School of Earth Sciences, University of Queensland, Australia.
Julien Denayer ⇑,1
Animal and Human Palaeontology, Geology Department, University of Liège, Belgium
a r t i c l e i n f o
Article history:Received 11 March 2014Received in revised form 10 June 2014Accepted 2 November 2014Available online 22 November 2014
Keywords:Rugose coralsKueichouphyllumPalaeogeographyCimmerian TerraneSystematics
a b s t r a c t
This paper gives the first taxonomic description of the Upper Tournaisian–Lower Viséan rugose coralfauna of the Yaricak Formation (Aladag Unit, Central Taurides, South Turkey). Fifteen species belongingto twelve genera were identified, one species is newly described: Eokoninckocarinia gemmina. The coralsare stratigraphically distributed in four assemblages. The two typical assemblages of the Upper Tournai-sian are composed of widely distributed taxa (Uralinia, Caninia, Proheterelasma, Zaphrentites). The assem-blage crossing the Tournaisian–Viséan boundary is characterized by Eurasian and cosmopolitan andwidely distributed taxa (Calmiussiphyllum, Siphonophyllia, Bifossularia Amygdalophyllum, Caninophyllum,Keyserlingophyllum) and Asian taxa (Kueichouphyllum). The youngest assemblage, dominated by Eokon-inckocarinia gemmina sp. nov., has yielded foraminifers Moliniacian (Lower Viséan) in age. These assem-blages form a low diversity level-bottom community which is typical of the South Palaeotethys‘Kueichouphyllum Zone’ extending along the Asian margin of Gondwana (Cimmerian Terrane) duringLower Carboniferous times. As in the other Cimmerian blocks, all the corals are solitary and colonial taxaare virtually absent. This absence is tentatively explained by the high palaeolatitude (c. 50�S) position ofthe Cimmerian Terrane in the southern part of the Palaeotethys Ocean for this time slice. A cold-waterpalaeo-current running eastward along the Gondwana margin might also be considered as it possiblycould explain the wide distribution of the Kueichouphyllum fauna, restricted east of Africa in the southerncoast of the Palaeotethys. Palaeoceanography, palaeoclimate and facies issues are discussed as possiblecauses of the diversity gradient observed between the eastern (Australia, Malaya) and the western (NorthAfrica) margin of the Gondwana.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
The Late Tournaisian and Early Viséan times were a period ofimportant change in the geobiosphere. The Devonian greenhouseclimate switched to icehouse as the Late Palaeozoic glaciationstarted (see Fielding et al., 2008; Caputo et al., 2008 for summary);the tectonic movements of the forthcoming Variscan Orogenymodified the continental and oceanic settings around the Palaeot-ethys (e.g. Stampfli and Borel, 2002; Nance et al., 2010); the marinefauna slowly recovered after the crisis associated with the Devo-nian–Carboniferous boundary. A high relative sea level at this timedrove a good communication between provinces and allows the
dispersion of marine fauna, making them cosmopolitan duringthe uppermost Tournaisian Avins diversification event (Poty,2007). In contrast, the earliest Viséan recorded a very low sea leveland during this period, the marine basins were more isolated andconsequently, the organisms – and particularly the rugose corals– became more endemic (Aretz et al., 2014). If the coral fauna ofthe Tournaisian–Viséan transition are well known in WesternEurope (Poty, 1989, 2007; Somerville et al., 1989; Weyer, 1993),Eastern Europe (Vassiljuk, 1960; Dobroljubova et al., 1966; Potyet al., 2003) and China (Yü, 1933; Poty and Xu, 1996; Xu andPoty, 1997), those of the Middle East and Central Asia are almostunknown. With the exception of some works in Iran (Douglas,1950; Flügel, 1963, 1991; Khaksar, 1996) and China (Fan et al.,2003), the Carboniferous fauna of the Asian margin of Gondwanais Terra Incognita. The term ‘Asian’ margin is here used instead of‘northern’ margin as the Gondwana was centred at this time onthe South Pole and its entire margin was consequently ‘northern’.This paper aims to summarize the current knowledge of these

372 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
areas and to give the first taxonomic description of the rugose cor-als of the Central Taurides for the Ivorian (Upper Tournaisian) andMoliniacian (Lower Viséan).
2. Settings
Southern Turkey (‘Anatolides and Taurides platform or block orunit’ – denomination varying from author to author) is composedof several tectonostratigraphic units (Fig. 1A). Traditionally, theAnatolides includes metamorphic Palaeozoic rocks devoid of post-Palaeozoic cover while the Taurides are mainly composed of Cam-brian–Miocene sedimentary rocks. In detail, the difference is farfrom this dual definition. The metamorphism is locally low-gradein the Anatolides (greenschist facies; Robertson and Pickett, 2000;Robertson and Ustaömer, 2009) and the sedimentary succession ispreserved. As suggested by Moix et al. (2008) the nature of the base-ment is the most convenient definition. In the Anatolides the base-ment is composed of Silurian to Carboniferous flysch witholistoliths. Viséan carbonate olistoliths were documented in theKonya (Robertson and Ustaömer, 2011) and Hadım (Denayer andAretz, 2012) in Central Turkey. The latter include reefal facies partic-ularly rich in corals which show affinities with the southern Euro-pean Province (Spain, South France, North Africa; Denayer, 2012).
In the Taurides, the basement is dominated by Precambrian–Palaeozoic sedimentary rocks and Mesozoic carbonates form thecover (Özgül, 1997). As part of the Cimmerian Terrane (S�engör,1979; Yılmaz, 1981), the Taurides recorded the rifting fromGondwana in the Permian and the Cimmerian Orogeny in the Tri-assic (S�engör and Yılmaz, 1981). The Lower Carboniferous isexposed in several tectono-stratigraphic units, namely theGeyık Dagı, Antalya and Aladag Unit. Göncüoglu et al. (2007)noticed slightly metamorphosed Tournaisian and Viséan lime-stones and siliciclastics in the Sultandag Massif of the Geyık DagıUnit. Dumont and Lys (1973) indicated the presence of Viséan sed-imentary rocks in the Gökdag near Beys�ehir in the same tectono-stratigraphic unit. Demirtas�lı et al. (1978b) described several sec-tions through the Lower Carboniferous in the Eastern Taurides nearKayseri. Tournaisian limestone and shale have been documentedby Demirtas�lı et al. (1978a) in the Korucuk section near Silifke(Antalya Unit) while Yurtsever et al. (2000) described Viséan lime-stones in Es�kibey between Silifke and Anamur. In the Aladag Unit,the Carboniferous has been documented in the eastern Tauridesnear Pinarbas�ı (Altıner and Zaninetti, 1980), Nohutluktepe (Okanand Hos�gör, 2005; Okuyucu and Vachard, 2006) and Pozantı(Frech, 1916). Moreover, the Aladag Unit is well exposed in theHadım area in the Central Taurides. Argyriadis (1978), Özgül(1984, 1997), Altıner and Özgül (2001), Turan (2001), Atakul-Özdemir et al. (2011) and Mackintosh and Robertson (2012) pro-vided litho- and bio-stratigraphic studies of the Tournaisian andViséan Yaricak Formation. The most continuous and fossiliferoussection exposing this formation is the stratotype, situated alongthe road between Hadım and the Beyreli Köyü hamlet, 9 kmsouth-west of Hadım and 10 km west of Tas�kent, (36�54046.780N32�23054.750E, Fig. 1C). The Yaricak section exposes the eponymousformation on the north and south sides of fault trending WNW–ESE, with a mean southward dip of 40�. The complementary ÇitYayla section (700 m west of the main section, Fig. 1C) providesa good outcrop of the lower part of the formation. The same suc-cession is known in the section along the Alanya road south-eastof the Tas�kent village but the Mantar Tepe Member is less fossilif-erous (Fig. 1D).
The Yaricak Formation (Fm) is divided into two members: thelower Çit Yayla Member and the upper Mantar Tepe Member.The Çit Yayla Member (Mbr) – is composed of dark-coloured,variably calcareous shale and silty shale including some bioclastichorizons with crushed brachiopods and crinoids. The macro-fauna
described by Özgül (1997) indicates a Tournaisian age of this mem-ber. Its base, not dated, rests on the Frasnian and possibly includesthe Famennian. The first limestone bed (cherty sandy limestone)occurs 170 m below the top of the formation. This horizon (Y0 inFig. 2) yields small solitary rugose corals and rare foraminifersindicating an Ivorian age (Altıner and Özgül, 2001). The topmostpart of the Çit Yayla Mbr (YR) shows the gradual appearance ofdark argillaceous bioclastic limestone alternating with shale. Thesebeds yield an abundant but poorly diversified association of Cani-nia, Uralinia and Zaphrentites. The base of the Mantar Tepe Mbr isa regional marker horizon of limestone crowded with large cylin-drical soliary rugose corals (=‘Siphonophyllum bed’ in Özgül,1997). The ten lower metres of the member (YA in Fig. 2) are dom-inated by bioclastic and sandy limestone with numerous tabulate(syringoporids) and solitary rugose corals while the upper part(YAsup) is almost devoid of macrofauna. The next unit is a 45 m-thick package of light bioclastic massive limestone, often dolomi-tized, with birds-eyes structures and evaporites pseudomorphs.Several thin oolitic beds occur in this unit. Only a few horizonsyield solitary rugose corals, mainly Kueichouphyllum and Keyser-lingophyllum (YB). This unit is topped by an argillaceous level.The overlying 25 m are dominated by dark non-fossiliferous mud-stone and wackestone (Fig. 2). The uppermost unit of the memberforms a massive rock inlier easy to recognize in the whole region. Itis made of a thick-bedded bioclastic and peloidic sandy grainstonewith abundant brachiopods, gastropods, syringoporids and solitaryrugose corals (YBsup in Fig. 2). The latter all belong to the speciesEokoninckocarinia gemmina sp. nov. This last limestone unit (40 m)is conformably overlain by sandstone and calcareous sandstonedevoid of macrofauna. The limestones have been assigned to beMiddle–Upper Viséan after Özgül (1997) and Altıner and Özgül(2001) but the foraminifers indicate an age no younger than Moli-niacian (Altıner and Özgül, 2001: zone VT3 corresponding to theMFZ11 foraminifers biozone of Poty et al., 2006). The overlyingsandstones and calcareous sandstones have been assigned to theUpper Viséan to Early Serpukhovian by Altıner and Özgül (2001).
3. Material and methods
The present paper is based on c. 75 specimens collected in theCentral Taurides in 2009 and studied in thin section (c. 150 trans-verse and longitudinal thin sections). The newly collected materialis housed in the Laboratory of Animal and Human Palaeontology,Department of Geology, University of Liège in Belgium (collectionPA.ULg). Systematic attribution follows Hill (1981) unless speci-fied. Abbreviations: TS: transverse section, LS: longitudinal section.
4. Systematic palaeontology
Subclass Rugosa Milne-Edwards and Haime 1850Order Stauriida Verrill 1865Suborder Stereolasmatina Hill 1981Family Antiphyllidae Iljina 1970Genus Proheterelasma Cotton 1973Type-species. Hadrophyllum edwardsianum(=Proheterelasma omaliusi) De Koninck 1872, Tournaisian of
Tournai.Diagnosis. See Hill (1981).Remarks. Grabau (1922) created the genus Heterelasma based on
de Koninck’s (1872) illustration of Hadrophyllum edwardsianum. Asthe generic name was preoccupied by a brachiopod, Cotton (1973)introduced the name Proheterelasma (see Vuillemin, 1990). Thegenus was often confused with Fasciculophyllum Thomson 1883after the confusion in attributing the type-species, initiallyincluded in the genus Hadrophyllum Milne-Edwards and Haime

Fig. 1. A: General structural map of Turkey (modified after Görür and Tüysüz, 2001; Moix et al., 2008; Okay, 2008). B: Geological map of the Hadım area (modified after Özgül,1984). C: Simplified locality map of the Yaricak Yayla section (Y0, YR, YA, YAsup, YB, YBsup refer to the lithological units explained in the text) and Çit Yayla section (YC)which can be correlated with the YR and YA units (after Altıner and Özgül, 2001). D: Simplified locality map of the Tas�kent–Alanya road section (ARO, AR1 and AR2 refer to thelithological units respectively equivalent to the Y0, YA and YB units in Yaricak Yayla section). Legend: CCAC: Central Anatolian Crystalline Complex, EAAC: East AnatolianAccretionary Complex, Lycian Np.: Lycian Nappes.
J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 373

Fig. 2. Composite log of the Yaricak Formation based on Yaricak Yayla section. Legend: Chrono.: chronostratigraphy, Fm.: Formation, Mbr.: Member, Litho.: lithological units(see main text), foraminiferal zones are from Altıner and Özgül (2001), *: ‘Siphonophyllum bed’ of Özgül (1997), corals: corals assemblages defined here.
374 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398

J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 375
1850. Nevertheless, Proheterelasma shows distinctive charactersknown neither in Fasciculophyllum nor in Hadrophyllum: majorsepta joined in four bundles fusing at the axis of the corallum, con-spicuous cardinal and alar fossulae and a ceratoid or trochoid hab-itus. Hadrophyllum is typically patellated and has oftencontratingent minor septa. Fasciculophyllum has a long rhopaloidcounter septum and no fossula. Rudimentary minor septa and sep-tal bundles are the main differences between Proheterelasma andSychnoelasma Lang et al. 1940. Saleelasma Weyer 1970 shows asimilar septal pattern but has carinated septa while Proheterelasmahas smooth septa. Proheterelasma can easily be distinguished fromZaphrentites Hudson 1941 and Amplexizaphrentis Vaughan 1906 byits cardinal fossula situated on the concave side of the corallumand its septa grouped in bundles joined at any stage of develop-ment (withdrawn in the adult stages of Amplexizaphrentis). Theoldest Proheterelasma is known from the base of the Ivorian (upperTournaisian) and the genus if common up to the basal Moliniacian(Boland, 2002). Occurrences in the Upper Viséan (e.g. in the Sahara;Menchikoff and Hsu, 1935) needs to be checked and verified.Hudson (1944) considered that Proheterelasma evolved into Roti-phyllum by simplification of the septal pattern and the tighteningof the axial structure. This interpretation is supported by the strongsimilarity of the juvenile stages of Rotiphyllum and the adult stagesof Proheterelasma (Weyer, 1993).
Proheterelasma omaliusi (Milne-Edwards and Haime, 1851)Fig. 3A1–A2.
*
1851 Zaphrentis omaliusi Milne-Edwards and Haime, p.377, pl. 5, fig. 3.1860
Zaphrentis omaliusi Milne-Edwards and Haime;Milne-Edwards, p. 344.1861
Zaphrentis omaliusi Milne-Edwards and Haime;de Fromentel, p. 289.?
1872 Hadrophyllum edwardsianum de Koninck, p. 52,pl. 4, figs. 2–2a.1872
Zaphrentis omaliusi Milne-Edwards and Haime;de Koninck, p. 94, pl. 9, figs. 4–4a.non
1895 Zaphrentis omaliusi Milne-Edwards and Haime;Stuckenberg, p. 36, pl. 2.p.p.
1906 Zaphrentis aff. phillipsi Milne-Edwards andHaime; Vaughan, p. 269, pl. 22, figs. 2c–e.1908
Zaphrentis omaliusi Milne-Edwards and Haime;Carruthers, p. 25, pl. 4, figs. 1–4.?
1922 Heterelasma edwardsianum (de Koninck);Grabau, pl. 1, figs. 14a and b.1931
Zaphrentis omaliusi Milne-Edwards and Haime;Fomichev, p. 19.1937
Zaphrentoides (Zaphrentoides) omaliusi (Milne-Edwards and Haime); Schindewolf, p. 448, pl. 22,figs. 2c–e.1942
Fasciculophyllum omaliusi (Milne-Edwards andHaime); Hudson, p. 260.?
1943 Fasciculophyllum thomsoni Hudson and Fox, p.105, pl. 4, figs. 1–4.?
1964 Neozaphrentis yui Wu, p. 31, pl. 2, fig. 22. 1966 Fasciculophyllum omaliusi (Milne-Edwards andHaime); Dobroljubova et al., p. 22, pl. 1, figs. 1–3.
1972 Fasciculophyllum omaliusi (Milne-Edwards andHaime); Weyer, p. 714, fig. 3.
1973 Proheterelasma edwardsianum (de Koninck);Cotton, p. 162.
1990 Proheterelasma (?) omaliusi (Milne-Edwards andHaime); Vuillemin, p. 35, pl. 1, figs. 4–5.
1991 Rotiphyllum omaliusi omaliusi (Milne-Edwardsand Haime); Flügel, p. 662, fig. 9.
1993
Rotiphyllum omaliusi (Milne-Edwards andHaime); Weyer, p. 41, pl. 2, figs. 12–13, pl. 3, figs.2, 7, pl. 4, figs. 2–5.1994
Fasciculophyllum omaliusi (Milne-Edwards andHaime); Poty and Hannay, p. 58, pl. 1, figs. 8–10.1996
Rotiphyllum? cf. omaliusi (Milne-Edwards andHaime); Khoa, p. 33, pl. 2, figs. 2–3.1999
Zaphrentites parallela (Carruthers); Heravi andKhaksar, pl. 3, fig. 10.2011
Proheterelasma omaliusi (Milne-Edwards andHaime); Denayer et al., p. 154, pl. 3, fig. O.Holotype. Specimen figured by Milne-Edwards and Haime(1851, plate 5, fig. 3), upper Tournaisian of Tournai; Museumnational d’Histoire Naturelle, Paris.
Material. Two specimens (4 TS) from Yaricak Yayla, Çit Yaylaand Tas�kent–Alanya road sections.
Diagnosis. Small Proheterelasma, maximum 13 mm in diameter.Maximum 32 major septa grouped in four bundles separated bythe cardinal and alar fossulae, and confluent with the stereo-col-umn. Septa withdrawn and rhopaloid in the counter quadrants.Minor septa rudimentary. Dissepimentarium absent. Tabulae com-plete, depressed toward the external wall. Modified from Cotton(1973).
Description. The small ceratoid coralla are about 10 mm high.The observed sections have a mean diameter of 6 mm and 19–22major septa (Fig. 4). They are long, joined in bundles of 4 or 5 septaand are confluent with the stereocolumn. The septa are concavetoward the counter sides. Seldom minor septa develop in the coun-ter quadrants (Fig. 3A2). The thickness of the septa varies between0.5 mm at the base to 0.2 mm in the middle and 0.5–0.6 mm attheir axial end. The cardinal septum is long and particularly thin.The axial stereocolumn has a mean diameter of 2–3 mm (i.e. 1/3–1/2 of the corallum diameter). The cardinal fossula is long andnarrow with convergent edges. The counter and alar fossulae areinconspicuous. The external wall is 0.4–0.5 mm thick.
Remarks. The present specimens share the characters ofP. omaliusi (Milne-Edwards and Haime, 1851) but are quite smaller(6 mm and 19–22 septa versus 11–13 mm and 30–32 septa forP. omaliusi). The observed sections showing juvenile characters(poorly conspicuous fossulae and septa densely packed in bundles),could represent the young stages of P. omaliusi. P. thomsoni Hudson,1943, figured by Hudson and Fox (1943) has similar dimensions andnumber of septa. Nevertheless, the variability of P. omaliusi coversthat of P. thomsoni and Vuillemin (1990) considers the two speciesas synonyms.
Geographic and stratigraphic range. In Belgium and England, thespecies appears at the base of the Ivorian (Vaughan, 1906; biozoneRC3a of Poty et al., 2006). The species is known in the Moliniacianin N France (Brittany: Vuillemin, 1990; Boulonnais: Poty andHannay, 1994) and in the British Isles (Hudson and Fox, 1943;Somerville and Jones, 1985). P. omaliusi is present in the Tournai-sian of Kuznetsk (Dobroljubova et al., 1966), North Iran (Flügel,1991) and Central Afghanistan (Dronov et al., 1979). In Turkey, itis known in the shaly limestone of the Çit Yayla Mbr (YaricakFm, Ivorian after Altıner and Özgül, 2001), in the Yaricak Yaylaand Tas�kent–Alanya road sections.
Family Hapsiphyllidae Grabau, 1928Genus Zaphrentites Hudson, 1941Type-species. Zaphrentis parallela Carruthers, 1910; Tournaisian
of England.Diagnosis. See Carruthers (1910).Remarks. Zaphrentites shares its simple morphology with many
zaphrentoid-like taxa. The position of the cardinal fossula – on the

Fig. 3. Solitary rugose corals of the Tournaisian–Viséan transition in Central Taurides, Turkey. A1–A2: Proheterelasma omaliusi (De Koninck, 1872), specimen AR.0.1,successive transverse sections, x5 (scale bar = 3 mm). B and C: Zaphrentites delanouei (Milne-Edwards and Haime, 1851), B: specimen YR.8.4, C: specimen YA.7.11, transversesections, x5 (scale bar = 3 mm). D1–D3: Caninia cornucopiae Michelin in Gervais, 1840, specimen YA.7.1, successive transverse sections, x5 (scale bar = 3 mm). E1–E2:Siphonophyllia sp., specimen YA.10.1, E1: longitudinal section, E2: transverse section, x3 (scale bar = 5 mm). F: Bifossularia sp., specimen YB.8.1, transverse section, x3 (scalebar = 5 mm). G1–G5: Calmiussiphyllum cf. dobroljubovae (Flügel, 1963), specimen YA.12.1, G1–4: successive transverse sections, x3 (scale bar = 5 mm), G5: close-up view ofthe dissepimentarium and contraclinant and contratingent minor septa, x6 (scale bar = 2.5 mm).
376 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398

Fig. 4. Scatter diagram showing the number of septa plotted against corallumdiameter for Zaphrentites delanouei and Proheterelasma omaliusi.
J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 377
concave side of the corallum – is a convenient criterion to distin-guish Zaphrentites because it is on the convex side in almost allthe other zaphrentoid corals. It differs from AmplexizaphrentisVaughan 1906 by larger dimensions and by its septa usually notwithdrawn from the axis in the mature stages. The septal patternnear the fossula allows the discrimination of Zaphrentites and Sych-noelasma Lang et al., 1940 while the morphology of the axial ste-reocolumn is the main distinctive feature between RotiphyllumHudson, 1942 and Zaphrentites. Based on the septal development,Fedorowski (2012) recently moved several species from Zaphren-tites to Zaphrufimia Fedorowsi, 2012.
Zaphrentites delanouei Milne-Edwards and Haime, 1851Fig. 3B and C.
*
1851 Zaphrentis delanouei Milne-Edwards and Haime,p.332, pl. 5, fig. 2.1906
Zaphrentis aff. Phillipsi Vaughan, pl. 22, figs. 2–2a. 1908 Zaphrentis delanouei Milne-Edwards and Haime;Carruthers, p. 63, pl. 5, figs. 5–7.
1910 Zaphrentis delanouei Milne-Edwards and Haime;Carruthers, p. 533, pl. 37, figs. 3a–c.
1931 Zaphrentis delanouei Milne-Edwards and Haime;Tolmachev, p. 322, pl. 18, figs. 9–11.
1931 Zaphrentis delanouei Milne-Edwards and Haime;Fomichev, p. 20, pl. 1, fig. 6.
1940 Zaphrentis delanouei Milne-Edwards and Haime;Hill, p. 144.
1941 Zaphrentis delanouei Milne-Edwards and Haime;Hudson, p. 291.
non 1981 Zaphrentoides delanouei (Milne-Edwards andHaime); Wu et al., p. 6, pl. 2, figs. 15–16.
1992 Zaphrentites delanouei (Milne-Edwards andHaime); Rodríguez and Falces, p. 171, pl. 14, figs.1–2.
1994
Zaphrentites delanouei (Milne-Edwards andHaime); Rodríguez and Falces, p. 201, fig. 5.5.1999
Zaphrentites delanouei (Milne-Edwards andHaime); Heravi and Khaksar, pl. 3, figs. 5–9.2011
Zaphrentites delanouei (Milne-Edwards andHaime); Denayer et al. p. 156, pl. 3, figs. P, R.Lectotype. Hudson (1941) choose the specimen figured byMilne-Edwards and Haime (1851, pl. 5, fig. 2).
Material. Eight specimens (10 TS) from the Çit Yayla Mbr of theYaricak Yayla and Çit Yayla sections.
Diagnosis. See Carruthers (1910).
Description. Small trochoid coral about 20 mm-high with a calyxof maximum 12 mm in diameter. Transverse sections in maturestages are on average 9 mm in diameter and have 25–30 septa(Fig. 4). The septa are long and curved. In the counter quadrants,the septa are radially disposed and join the axial top of the cardinalfossula. In the cardinal quadrants, they are curved (concavitytoward the counter quadrants, Fig. 3B) and attached to the septaforming the edges of the fossula. The septa are typically thick atthe base (up to 1 mm thick) and thin (0.3 mm) in their median part.The cardinal and cardinal lateral septa are short. The alar septa areconfluent with their counter-sided neighbours. The septa are morenumerous in the counter quadrants than in the cardinal ones. Theminor septa appear as septal crests on the external wall but canextend slightly in the counter quadrants. In the calyx, the minorsepta can be 2/3 as long as the major. The cardinal fossula is welldeveloped; its edges are parallel in juvenile stages but sag in themature stages where the fossula is typically key-hole shaped(1 mm-large near the wall, 2.5 mm in the axial part and 4–6 mm-long). The fossula is longer than the corallum radius, itstop is thus put forward to the counter side (Fig. 3B). Alar fossulaeare rarely developed. The external wall is thick (1 mm), undulatingor arched, each groove corresponding to the base of a septum.
Remarks. These specimens possess the dimensions, number ofsepta and morphology of Z. delanouei Milne-Edwards and Haime,1851. They share similar dimensions with Z. constricta (Carruthers,1910), but the latter has a narrow triangular fossula. Z. crassusHudson, 1944 is smaller (9 mm and 20 septa) and has thickened ele-ments. Z. parallela (Carruthers, 1910), is slightly smaller and charac-terized by a parallel-edged fossula and a longer counter septum.
Geographic and stratigraphic range. Zaphrentites delanouei iscommon in the Tournaisian of Western Europe. It is the guide taxafor Vaughan’s (1906) Zaphrentites delanouei Zone (i.e. Upper Tour-naisian, Mitchell and Green, 1965). It is also common in Belgium(zone RC3c of Poty et al., 2006), Brittany (Vuillemin, 1990), Poland(Poty et al., 2003), Donets Basin (Vassiljuk, 1960), Kuznetsk(Tolmachev, 1931) and Iran (Flügel, 1991; Heravi and Khaksar,1999). In Turkey, the species is known in the limestone-shale alter-nation forming the top of the Çit Yayla Mbr (Ivorian after Altınerand Özgül, 2001).
Suborder Caniniina Wang, 1950Family Cyathopsidae Dybowski, 1873Genus Caninia Michelin in Gervais, 1840Type-species. Caninia cornucopiae Michelin in Gervais, 1840;
Tournaisian of Tournai.Diagnosis. Solitary ceratoid to cylindrical corallum. Cardinal fos-
sula in the convex side of the corallum. In juvenile stages, majorsepta long and thick in cardinal quadrants. In mature stages, septawithdrawn, straight or curved toward the fossula. Counter septumusually longer, sometimes with a thickened axial end. Minor septausually short or rudimentary. Dissepimentarium narrow includinginterseptal and seldom lonsdaleoid dissepiments. Tabulae horizon-tal in the axial part of the tabularium, inclined toward the dissepi-mentarium and the cardinal fossula. Modified from Hill (1981)
Remarks. The genus Caninia was often used for any solitary cor-als devoid of axial structure and with short septa – i.e. caninimorphcorals – were classified in Caninia. Semenoff-Tian-Chansky (1974)was the first worker to give an accurate diagnosis for Caninia fromwhich he excluded taxa with a complex dissepimentarium (Cani-nophyllum Lewis, 1929), with lonsdaleoid dissepiments (Siphono-phyllia Scouler in Mc Coy, 1844), with short minor septa(Haplolasma Semenoff-Tian-Chansky, 1974) or rudimentary minorsepta (Pseudozaphrentoides Stuckenberg, 1904). Unfortunately,the type species, Caninia cornucopiae Michelin in Gervais, 1840,despite its simple morphology is extremely malleable and its inter-pretation is a sensitive issue. A revision of the abundant topotypicmaterial from the Tournaisian of Tournai would be very welcome

378 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
to help understanding the species. Specimens from the Ivorian,show commonly a long counter septum (commonly with a thick-ened axial end, unlike stated by Hill, 1981) in the mature stagesassociated with a withdrawal of the minor septa, a thickened innerrow of dissepiments, the development of lonsdaleoid dissepimentsand an underdevelopment of the cardinal fossula (Caninia cornuco-piae var. vesicularis Salée, 1910). These parallel trends produce amorphotype resembling isolated corallites of the fasciculate genusDorlodotia Salée, 1920.
Caninia cornucopiae Michelin in Gervais, 1840Fig. 3D1–D3.
*
1840 Caninia cornucopiae Michelin in Gervais, p. 485. 1842 Cyathophyllum plicatum Goldfuss; De Koninck, p.22, pl. C, figs. 4a–g.
1842 Cythophyllum mitratum (Schloth); De Koninck, p.22, pl. C, figs. 5a–f.
1847 Caninia cornucopiae Michelin; Michelin, p. 256,pl. 59, fig. 5.
1872 Campophyllum analogum De Koninck, p. 45, pl.3,fig. 6.
1872 Lophophyllum? Dumonti Milne-Edwards andHaime; De Koninck, p. 55, pl. 4, fig. 4.
1872 Amplexus ibicinus (Fischer von Waldheim); DeKoninck, p. 67, pl. 6, fig. 2.
1872 Amplexus cornu-arietis De Koninck, p. 72, pl. 6,fig. 4,
1872 Zaphrentis Edwardsiana De Koninck, p. 83, pl. 7,figs. 4a and b.
1872 Zaphrentis vermicularis De Koninck, p. 95, pl. 10,figs. 1a–c.
1872 Zaphrentis cornucopiae (Michelin); De Koninck, p.100, pl. 10, figs. 5a and b, pl. 15, fig. 2.
1872 Zaphrentis Nystiana De Koninck, p. 103, pl. 10, fig.8.
1908 Caninia cornucopiae Michelin; Carruthers, p. 159,pl. 6, figs. 1–4.
? 1910 Caninia cornucopiae Michelin; Salée, p. 20, pl. 1,figs. 1–4.
1910 Caninia cornucopiae var. vesicularis Salée, p. 20,pl. 1, figs. 5a–k.
non 1917 Caninia cornucopiae Michelin; Vaughan, p. 36, pl.4, figs. 8–9.
1939 Caninia cornucopiae Michelin; Hill, p. 106. 1950 Caninia aff. cornucopiae Michelin; Douglas, p. 21,pl. 4, figs. 5a–c.
1974 Caninia cornucopiae Michelin; S.-T.-C., p. 174, pl.44, figs. 5–7, pl. 75, fig. 2.
1979 Caninia cornucopiae Michelin; Kato, p. 140, pl. 1,figs. 8–11.
? 1981 Caninia cornucopiae Michelin; Poty, p. 52, pl. 24,fig. 5.
1989 Caninia cornucopiae Michelin; Somerville et al., p.58, fig. 5g.
1990 Caninia cornucopiae Michelin; Vuillemin, p. 49,pl. 4, fig. 1.
2011 Caninia cornucopiae Michelin; Denayer et al., p.156, pl. 3, fig. D.
Fig. 5. Scatter diagram showing the number of septa plotted against corallumdiameter for Siphonophyllia sp., Caninia cornucopiae, Calmiussiphyllum cf. dobroljubo-vae and Caninophyllum cf. patulum.
Lectotype. Type specimens from the Tournaisian of Tournai(Belgium) and Sablé (France), from the Michelin collection storedin Caen Museum, were destroyed during World War Two. Thelectotype of Lang et al. (1940) is the specimen RC331 from Tournai,figured by Carruthers (1908) and Hill (1981, fig. 222); BritishGeological Survey Collection.
Material. Two specimens (4TS) from Yaricak Yayla.Diagnosis. Caninia having thin major septa, half to one-third as
long as the corallum radius. Minor septa short or rudimentary.Counter septum long and thickened, forming a columella in thejuvenile stages, withdrawing during the growth. Simple interseptaldissepiments and occasional lonsdaleoid dissepiments. Modifiedfrom Semenoff-Tian-Chansky (1974).
Description. The solitary cylindrical corals are 5–6 cm high andup to 2 cm in diameter in the calyx. The sections are 12–18 mmin diameter (maximum 22 mm) and have a mean tabularium of16 mm (maximum 20 mm). There are 32–34 septa of both orders(maximum 36, Fig. 5). The major septa are short (no more than1/3 of the radius in length). They are thick in the juvenile stages(mostly in the cardinal quadrants) then become thinner firstly inthe counter quadrants. The cardinal septum is short. The counterseptum is up to twice as long as the other major septa (Fig. 3D2–D3). Its axial end is sometimes dilated in the juvenile stages butcan become thin in the mature stages. The minor septa are veryshort and appear as septal crests on the external wall or on theoccasional lonsdaleoid dissepiments. In the cardinal quadrants,the dissepimentarium comprises 1–2 rows of concentric intersep-tal dissepiments (Fig. 3D3). In the counter quadrants, there are 2–4rows of dissepiments, including occasional second order lonsdale-oid dissepiments. The inner row is often thickened. The externalwall is 0.6 mm-thick and regular.
Remarks. Both specimens are clear C. cornucopiae is term ofdimensions and number of septa. They share with the ‘classical’Tournaisian forms, a long counter septum in the juvenile stage thatwithdraws during growth. This last character allows distinguishingthem from C. aff. cornucopiae from the upper Tournaisian.
Geographic and stratigraphic range. Caninia cornucopiae is knownfrom the top of the Hastarian to the top of the Ivorian in Belgium(zones RC2 and RC3 of Poty et al., 2006), in the upper Tournaisianof Germany (Weyer, 2000), the British Isles (Somerville et al., 1989,1992), Brittany (Vuillemin, 1990), Kuznetsk (Dobroljubova et al.,1966), Kazakhstan (Volkova, 1941). The species is noticed in theViséan of Thailand (Fontaine et al., 1991), Siberia (Ivanowski,1967), Iran (Douglas, 1950) and Algerian Sahara (Semenoff-Tian-Chansky, 1974). In Turkey, C. cornucopiae is known in the upperTournaisian of the Eastern Taurides (Tufanbeyli area, Kato, 1979)and in the lower part of the Mantar Tepe Mbr of the Yaricak Fm,Central Taurides. Caninia aff. cornucopiae has recently been docu-mented in the Hakkari area (South-eastern Turkey, Arabian Plat-form; Denayer and Hos�gör, 2014).

J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 379
Genus Siphonophyllia Scouler in Mc Coy 1844Type species. Siphonophyllia cylindrica Scouler in Mc Coy, 1844;
Lower Viséan of Ireland.Diagnosis and remarks. See Poty and Boland (1994) and
Semenoff-Tian-Chansky (1974)Siphonophyllia sp.Fig. 3E1–E2.Material. Two specimens (6 TS, 2 LS) from Yaricak Yayla and
Tas�kent–Alanya road sections.Description. The coralla are cylindrical to scolecoid, up to 20 cm
long and up to 30 mm in diameter in the mature stages (22 mm forthe tabularium). There are 54 septa of both orders (Fig. 5). Themajor septa are long (at least half as long as the corallum radius)but do not reach the axis and leave a large free zone (8–10 mm)in the centre of the tabularium. They are straight or slightly undu-lating. They are thickened in the cardinal parts of the tabularium(Fig. 3E2) but their axial ends are sharp. The cardinal septum isshort and thick. The septa are slightly curved toward the cardinalseptum in the cardinal quadrants and toward the counter septumin the counter quadrants. The minor septa are long, entering intothe tabularium up to 1 mm, or short and reduced to septal crestson the dissepiments. The cardinal fossula is conspicuous and lar-gely opened. The dissepimentarium comprises 5 rows of first andsecond order lonsdaleoid dissepiments. The second order lonsdale-oid are more regular, concave toward the tabularium, V-shaped orherringbone between the major septa. The external wall is fes-tooned and 0.8–1 mm thick. In longitudinal section the tabulaeare complete, mesa-shaped, flat or slightly domed in the axial part.The dissepiments are 0.7–1 mm long and 1.5–4 mm high. There areabout 10 tabulae and dissepiments per centimetre.
Remarks. These specimens share with S. rivagensis Poty andBoland 1994 a dissepimentarium mainly composed of lonsdaleoiddissepiments and long minor septa. However, S. rivagensis is larger(35 mm in diameter and more than 50 septa) and has more inter-septal dissepiments. The dimensions and narrow dissepimentari-um of the Turkish specimens are similar to those observed inSiphonophyllia sp. A of Flügel (1991) and to Siphonophyllia sp. ofOgar et al. (2013).
Geographic and stratigraphic range. Siphonophyllia sp. was col-lected in the argillaceous limestone at the base of the Mantar TepeMbr (Ivorian) of the Yaricak Yaya section.
Genus Bifossularia Dobroljubova 1966Type-species. Caninia ussowi Gabunia, 1919; Tournaisian of the
Ural Mountains.Diagnosis. See Hill (1981).Discussion. The name Bifossularia is particularly badly chosen as
the counter fossula is rarely conspicuous. The genus identity ismainly based on the uniform thickening of the major septa, longminor septa and narrow dissepimentarium (sometimes no morethan one row of dissepiments). This last character allows the dis-tinction of Bifossularia from Siphonophyllia Scouler in Mc Coy,1844. Dobroljubova et al. (1966) and Hill (1981) classified Bifossu-laria in the Family Uralinidae on the base of their axially depressedtabulae but Vuillemin (1990) moved Bifossularia to the Cyathopsi-dae for its close similarity with Siphonophyllia – this second optionis adopted here. Indeed, the mature stages of Bifossularia sharewith the juvenile stages of Siphonophyllia thickened major septawith sharp ends, long minor septa and narrow dissepimentarium(e.g. Siphonophyllia sp. A in Somerville, 1994). Bifossularia possiblyevolved from Siphonophyllia by neoteny (paedomorphosis).
Bifossularia sp.Fig. 3F.Material. One single specimen (1 TS) from Yaricak Yayla section.Description. The large solitary corallum has an elliptical section
(slightly crushed) 30 � 45 mm-large. There are 80 septa of bothorders. The major septa are short and thickened in the whole
tabularium, their axial ends are sharp. The minor septa arerestricted to the dissepimentarium (eroded) or enter slightly intothe tabularium (Fig. 3F). The cardinal fossula is conspicuous, closedby the curved cardinal lateral major septa. The cardinal septum isshort. There is no conspicuous counter fossula. The dissepimentar-ium and the external wall are not preserved.
Remarks. The short major septa, thickened in the whole tabular-ium is a diagnostic character for Bifossularia. The dissepimentarium,unfortunately eroded, prevents an accurate specific attribution.Ogar et al. (2013) figured Bifossularia aghanabatiei Ogar et al., 2013from the Lower Viséan of the Alborz which has similar dimensionsbut less septa (58 major septa versus 80 in the Turkish specimen)and has minor septa long enough to enter into the tabularium.
Geographic and stratigraphic range. Bifossularia sp. was collectedin the middle part of the Mantar Tepe Mbr (Yaricak Fm) in theMoliniacian (after Altıner and Özgül, 2001). A similar speciesoccurs in the Lower Viséan of the Alborz (Ogar et al., 2013) andof the Kuznetsk Basin (Fomitchev, 1931).
Genus Uralinia Stuckenberg, 1895Type-species. Heliophyllum multiplex Ludwig, 1862; Tournaisian
of the Ural Mountains.Diagnosis. See Poty and Boland (1994).Remarks. Hill (1981) classified Uralinia among the Family Ura-
linidae Dobroljubova, 1966, together with other genera showingtabulae sagging toward the cardinal fossula and lonsdaleoid dis-sepiments (i.e. Keyserlingophyllum Stuckenberg, 1895, CystophrentisYü, 1931). However, since the phylogenetic relationship betweenSiphonophyllia Scouler in Mc Coy, 1844 and Uralinia Stuckenberg,1895 was demonstrated by Poty and Boland (1994), these authorsincluded the two genera within the Cyathopsidae Dybowski, 1873and considered the Uralinidae Dobroljubova as artificial. The othermembers of the family are most probably not related: Keyserlingo-phyllum possibly derives from Caninophyllum (see discussion ofthis genus) while Cystophrentis belongs to the Family Cystophren-tidae Yü, 1963 (see Poty and Xu, 1996). In fact the morphologicalsimilarity of these three genera might possibly be a result of home-omorphy, the three taxa developing independently a cystiphylloidtrend. Chinese palaeontologists long considered Pseudouralinia Yü,1931 as distinct from Uralinia Stuckenberg, 1895 by less inclinedtabulae and the lack of small peripheral lonsdaleoid dissepiments.Hill (1981) and Poty and Xu (1996) synonymised the two namessince both characters occur in Uralinia.
Uralinia multiplex (Ludwig, 1862)Figs. 6A1–A3, B1–B5, C and 7A.
*
1862 Heliophyllum multiplex Ludwig; p. 199. 1895 Uralinia multiplex Ludwig; Stuckenberg, p. 104, pl.8, fig. 4, pl. 11, fig. 8, pl. 20, fig. 5.
1933 Pseudouralinia gigantea Yü; p. 60, pl. 6, figs. 1a–e. 1938 Uralinia septata Gorsky; p. 25, pl. 3, fig. 1–2. 1960 Neomicroplasma dobroljubovae Rogozov; p. 49, pl. 2,fig. 1.
1960 Neomicroplasma septata Rogozov; p. 50, pl. 2, figs.4–5.
1960 Uralinia multiplex (Ludwig); Soshkina, p. 301, pl. 4,figs. 4–5.
1966 Uralinia multiplex (Ludwig); Dobroljubova et al., p.86.
1967 Uralinia multiplex (Ludwig); Ivanowski, p. 66, pl. 14,fig. 1.
? 1975 Uralinia multiplex (Ludwig); Gorsky et al., p. 78, pl.22, fig. 1.
? 1975 Uralinia septata Gorsky; Gorsky et al., p. 78, pl. 21,fig. 6.
(continued on next page)

Fig. 6. Uralinia species from the Tournaisian–Viséan transition in Central Taurides, Turkey. A–D: Uralinia multiplex (Ludwig, 1862). A1–A3: specimen YA.23.3, A1: polish slabin reversed colour, transverse section, x1 (scale bar = 15 mm), A2: close-up view of the dissepimentarium, transverse section, x2 (scale bar = 7.5 mm), A3: polish slab inreverse colour, longitudinal section, x1 (scale bar = 15 mm). B1–B4: specimen YR.2. 1, successive transverse sections in juvenile stages, x3 (scale bar = 5 mm), B5: longitudinalsection, x3 (scale bar = 5 mm). C: specimen YR.8. 1, transverse section in juvenile stage, x3 (scale bar = 5 mm). D–F: Uralinia sp. D: specimen VB.0. 1, transverse section in amature stage, x3 (scale bar = 5 mm). E: specimen YC.0.1.II, transverse section in juvenile stage, x3 (scale bar = 5 mm). F: specimen YC.0.3, transverse section in juvenile stage,x6 (scale bar = 2.5 mm).
380 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
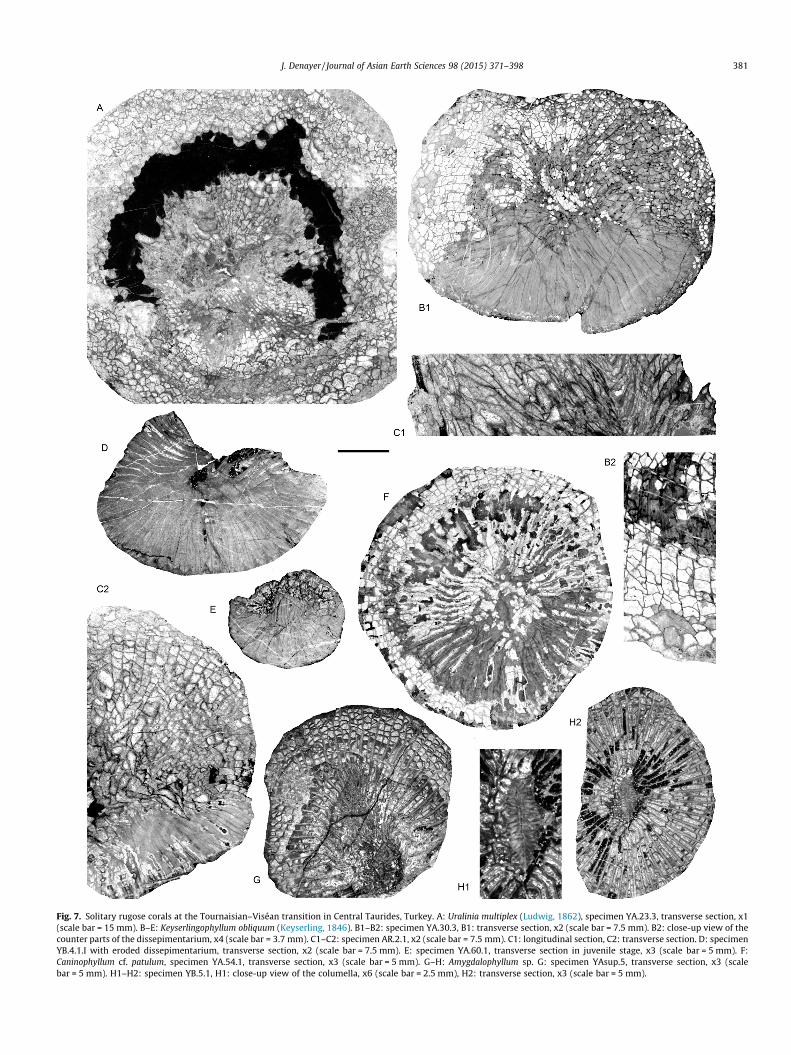
Fig. 7. Solitary rugose corals at the Tournaisian–Viséan transition in Central Taurides, Turkey. A: Uralinia multiplex (Ludwig, 1862), specimen YA.23.3, transverse section, x1(scale bar = 15 mm). B–E: Keyserlingophyllum obliquum (Keyserling, 1846). B1–B2: specimen YA.30.3, B1: transverse section, x2 (scale bar = 7.5 mm). B2: close-up view of thecounter parts of the dissepimentarium, x4 (scale bar = 3.7 mm). C1–C2: specimen AR.2.1, x2 (scale bar = 7.5 mm). C1: longitudinal section, C2: transverse section. D: specimenYB.4.1.I with eroded dissepimentarium, transverse section, x2 (scale bar = 7.5 mm). E: specimen YA.60.1, transverse section in juvenile stage, x3 (scale bar = 5 mm). F:Caninophyllum cf. patulum, specimen YA.54.1, transverse section, x3 (scale bar = 5 mm). G–H: Amygdalophyllum sp. G: specimen YAsup.5, transverse section, x3 (scalebar = 5 mm). H1–H2: specimen YB.5.1, H1: close-up view of the columella, x6 (scale bar = 2.5 mm), H2: transverse section, x3 (scale bar = 5 mm).
J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 381

382 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
?
1975 Uralinia aseptata Rogozov; Gorsky et al., p. 78, pl.21, fig. 7.1976
Uralinia multiplex (Ludwig); Onoprienko, pl. 1, figs.1–2, pl. 3, fig. 3.1976
Pseudouralinia tangpakouensis Yü; Onoprienko, pl. 1,figs. 3–4.1976
Siphonophyllia cylindrica Scouler in McCoy;Onoprienko, pl. 3, figs. 1–2.?
1976 Neomicroplasma sp.; Onoprienko, pl. 4, figs. 1–2. 1977 Pseudouralinia gigantea Yü; Jia et al., p. 181, pl. 68,fig. 1.
1979 Uralinia multiplex (Ludwig); Onoprienko, p. 20, pl. 4,figs. 3–4, pl. 5, figs. 2–3, 5.
1979 Neomicroplasma septata Rogozov; Onoprienko, p.26, pl. 2, figs. 5–6.
1979 Pseudouralinia tangpakouensis Yü; Onoprienko, p.24, pl. 3, fig. 7, pl. 4, fig. 1.
Fig. 8. Scatter diagram showing the number of septa plotted against corallum 1983 Uralinia sp.; Simakov et al., pl. 15, fig. 10. diameter for Keyserlingophyllum obliquum, Uralinia multiplex and Uralinia sp. 1984 Uralinia gigantea Yü; Poty and Onoprienko, p. 208,pl. 33, figs. 3, 5.
1987 Uralinia multiplex (Ludwig); Ivanowski, p. 2, figs. 4–5. 1989 Uralinia cf. multiplex (Ludwig); Poty, p. 270, fig. 3A. 1997 Uralinia gigantea Yü; Poty and Xu, p. 96, pl. 6, fig. 3. 2002 Uralinia gigantea Yü; Poty, p. 109, pl. 2, fig. 3. 2010 Uralinia cf. gigantea Yü; Poty, p. 391, fig. 2F. 2011 Uralinia multiplex (Ludwig); Denayer et al., p. 158,pl. 2, fig. K.
Holotype. Specimen GZG.INV.030.534, Tournaisian of the UralMountains, Collection Ludwig, Göttingen, Germany. Ludwig’s typematerial (Ludwig, 1862, pl. 24, fig. 4b), recently re-investigated byE. Poty (pers. com., June 2013) shows the typical juvenile charac-ters (short septa and a narrow dissepimentarium) of specimensdescribed in Belgium, Omolon (Onoprienko, 1979) and China(Poty and Xu, 1996) under the name U. gigantea (Yü, 1933). Conse-quently, U. gigantea must be considered as a junior synonym of U.multiplex (Ludwig, 1862). Both species are Ivorian in age. The neo-type designated by Ivanowski (1987) is a mature stage of anotherspecies (of U. lobata group) as demonstrated by its wide dissepi-mentarium. The neotype should in consequence, be rejected.
Diagnosis. Large Uralinia reaching 120 mm in diameter and pos-sessing up to 80 major septa in the cardinal quadrants. Major septatypically thickened in the cardinal parts of the tabularium andlocally in the counter parts. Dissepimentarium wide, particularlyin the counter quadrants, composed of numerous small globularlonsdaleoid dissepiments. Modified from Poty (2002).
Material. Five specimens (16 TS, 3 LS) from Yaricak Yayla.Description. The coralla are cylindrical, reaching 50 cm-long and
8 cm-wide. Sections in juvenile stages have a tabularium 20–26 mm in diameter and 35–50 septa of both orders (Fig. 8). Themajor septa are long and thick (1 mm) in the cardinal quadrantsbut thinner in the counter quadrants. The cardinal and countersepta are shorter and usually thinner than the other septa. Thecounter-lateral and cardinal-lateral septa are curved and close thecardinal and counter fossulae. The minor septa are developed onlyin the cardinal quadrants where they are intercepted by lonsdaleoiddissepiments. The tabularium is off-centred toward the cardinalside of the corallum (Fig. 6B3–B4). The external wall is thick. Inthe mature stages, the diameter reaches 80 mm while the tabular-ium is less than 30 mm large. About 100 septa are developed only inthe cardinal quadrants. They are short (less than 10 mm), thick(1 mm) and straight near the cardinal fossula but undulate in thealar sides. They are usually curved around the cardinal fossula.The dissepimentarium comprises up to 15 rows of lonsdaleoid dis-sepiments. They are small and globose in the external part and lar-
ger and more irregular in the inner parts (Fig. 6A2). In the counterquadrants, some small concentric interseptal dissepiments occur.In longitudinal section, the tabulae are incomplete, flat or domed,depressed toward the dissepimentarium where they are not easilydiscriminated from the dissepiments (Fig. 3A3 and B5). The tabulaesteeply slope at 75� toward the fossula. The height between twotabulae varies from 1 to 6 mm. The dissepiments are small (lessthan 1 mm-long and -high) in the outer dissepimentarium, larger(3 mm-long and 2 mm-high) near the tabularium.
Remarks. This species is easily differentiated from the other spe-cies of the genus by its large dimensions (U. multiplex is one of thelargest known rugose corals after Poty, 2010). Keyserlingophyllumobliquum Keyserling 1846 is similar in dimensions and morphologywith U. multiplex and the identification is sometimes difficult, par-ticularly if the counter parts of the dissepimentarium are not pre-served (see below).
Geographic and stratigraphic range. Uralinia multiplex is a guidetaxon for the Ivorian and is known in the Omolon Massif(Onoprienko, 1979), Urals Monutains (Ludwig, 1862), Novaya Zem-lya (Gorsky et al., 1975), South China (Poty and Xu, 1996) and Bel-gium (Poty, 1989; biozone RC3a of Poty et al., 2006). In Turkey, itoccurs at the top of the Çit Yayla Mbr and at the base of the MantarTepe Mbr where very large specimens are present in the ‘Siphono-phyllum bed’ in Özgül (1997), dated by foraminifers of the Ivorian(Altıner and Özgül, 2001).
Uralinia sp.Fig. 6D–F.Material. Seven specimens (8 TS) from the Çit Yayla Mbr, four
were collected in the Yaricak Yayla section and three in theAlanya–Tas�kent road section.
Description. In the juvenile stages smaller than 15 mm in diam-eter, there are about 20 septa. They are long (up to the axis) andextremely thick (Fig. 6E and F). In the mature stage (estimateddiameter: 40 mm), 38 septa are present (Fig. 8). The major septaare still thick and sinuous in the cardinal quadrants. Their axialends are sharp or abut on upturned tabulae. The cardinal septumis short and surrounded by curved cardinal lateral septa. In thecounter quadrants, the septa are longer and thinner. The dissepi-mentarium comprises 3–4 rows of interseptal dissepiments,including arched dissepiments in the fossula (Fig. 6D), and numer-ous rows of lonsdaleoid dissepiments. The size of the lonsdaleoiddissepiments increases toward the tabularium. Some of them arethickened.
Remarks. These specimens show juvenile stages very similarto those observed in U. lobata Poty and Boland 1994(‘Siphonophyllia-like stage’). The mature stages are also similar

J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 383
but have less septa. The thin long septa are another charactershared with U. lobata but this species possesses large lonsdaleoiddissepiments. The narrow dissepimentarium recalls U. uralica(Rogozov, 1960b) but the latter have commonly up to 70 septafor a similar diameter.
Geographic and stratigraphic range. All the specimens are fromthe cherty sandy limestone bed in the middle part of Çit YaylaMbr, (Ivorian after Altıner and Özgül, 2001).
Genus Keyserlingophyllum Stuckenberg, 1895Type-species. Cystiphyllum obliquum Keyserling, 1846; Tournai-
sian of the Ural Mountains.Diagnosis. See Poty and Xu (1996).Remarks. Stuckenberg (1895) created Keyserlingophyllum and
Humboldtia at the same time. After this author, the main differenceis the counter fossula developed in Humboldtia. This feature beingparticularly unstable even in the same specimen, Onoprienko(1979) and Hill (1981) considered the two genera as synonyms.Weiningophyllum Wang, 1978 and Xinningophyllum Jiang in Tanet al., 1987 were created for Keyserlingophyllum with large dissepi-mentarium but this character is more specific than generic and Xuand Poty (1997) synonymized the two names with Keyserlingophyl-lum. As stated in the discussion of Uralinia, KeyserlingophyllumStuckenberg, 1895 is morphologically similar to UraliniaStuckenberg, 1895. The distinction, although not easy in somecases, is based on: (1) longer and thicker septa in the counter quad-rants (usually poorly developed in Uralinia); (2) smaller lonsdale-oid dissepiments (compare Figs. 6A2 and 7B2); (3) interseptaldissepimentarium better developed (rarely present in Uralinia);and (4) Keyserlingophyllum shows commonly septa curved on eachsides of the cardinal and alar fossulae while they are more radiallydisposed in Uralinia. The dissepimentarium is usually eroded andthe thickened parts of the septa of both cardinal and counter quad-rants are preserved in Keyserlingophyllum while only the cardinalparts are usually preserved in Uralinia. Nevertheless, all theabove-cited characters are affected by inter- and intra-specific(even intra-individual) variation that does not help in the identifi-cation. In some stages, Keyserlingophyllum shows a regular dissepi-mentarium including several rows of interseptal dissepiments andless thickened septa extending through the whole tabularium. Thismorphology is particularly close to that of Caninophyllum patulum(Michelin, 1846). It could indicate that Keyserlingophyllum is thecystophylloid form of Caninophyllum, the same way Uralinia isthe cystiphylloid form of Siphonophyllia (Poty and Boland, 1994).A detailed study of the ontogeny of Caninophyllum would be usefulto understand the possible relationship with Keyserlingophyllum.
Keyserlingophyllum obliquum (Keyserling, 1846)Fig. 7 B1–B2, C1–C2, D, and E.
*
1846 Cystiphyllum obliquum Keyserling, p. 160, pl. 1, figs.5a–d.1895
Keyserlingophyllum obliquum (Keyserling);Stuckenberg, p. 102, pl. 5, fig. 8, pl. 6, fig. 3, pl. 20,fig. 2.1960
Keyserlingophyllum obliquum (Keyserling);Soshkina, p. 303, pl. 4, fig. 6, pl. 5, figs. 1–5.?
1971 Keyserlingophyllum concavotabulatum Katchanov inSpasky and Katchanov, p. 60, pl. 44, fig. 1.1987
Keyserlingophyllum obliquum (Keyserling);Ivanowski, p. 24, pl. 6, fig. 4.1996
Keyserlingophyllum obliquum (Keyserling); Poty andXu, p. 98, pl. 6, fig. 2.2002
Keyserlingophyllum obliquum (Keyserling); Poty, p.170, pl. 2, fig. 1.2011
Keyserlingophyllum obliquum (Keyserling); Denayeret al., p. 158, pl. 4, fig. C.Holotype. Specimen 4/46, Tournaisian, Ural Mountains; Collec-tion Keyserling.
Diagnosis. See Poty (2002).Material. Six specimens (9 TS, 2LS) from the basal part of the
Mantar Tepe Mbr: two come from the Tas�kent–Alanya road sec-tion, the other four come from Yaricak Yayla.
Description. Large cylindrical solitary corallum, more than 15 cmhigh, usually crushed and eroded. The mean diameter observed intransverse section is 63 mm (maximum 80 mm). The tabulariumdiameter varies between 30 and 40 mm. There are 65–85 septa(maximum 100, Fig. 8). The cardinal quadrants are occupied bythick (1.5–2.2 mm) and densely packed major septa (Fig. 7B1, D,and E). They form bundles on each sides of the cardinal and alarfossulae. In the counter quadrants, the septa are long, sinuous orvery tortuous. The thickening is restricted to the axial part of thesepta. In mature stages, the thickening can reduce. The cardinalfossula is well marked, the cardinal septum is shorter. In the cardi-nal quadrants, the dissepimentarium comprises 1–4 outer rows ofsmall globose lonsdaleoid and 4–6 inner rows of large and regularlonsdaleoid dissepiments. In the counter quadrants, there are 5–25rows of small regular interseptal dissepiments, concentric, angulo-concentric and V-shaped (Fig. 7B2). The external wall is thin (lessthan 0.2 mm). The tabulae are strongly divided, spaced and steeplyinclined toward the fossula. In the counter quadrants, the dissepi-ments are 1–4 mm high, 0.5–2 mm long and almost vertically dis-posed (Fig. 7C1). There are 1–4 tabulae and 3–4 dissepiments percentimetre.
Remarks. Diameter, number of septa, width of dissepimentari-um, morphology of dissepimentarium and septal pattern are thetypical characters of K. obliquum (Keyserling, 1846). It is the largestKeyserlingophyllum species. K. obliquum differs from K. ruttneri(Flügel, 1963), K. youwangense (Yü and Fan in Yü et al., 1984)and K. qinghaiense (Wang, 1981) by its septa thickened only inthe axial ends in the counter quadrants; from K. keyserlingophyllo-ides (Chu, 1933) by alar fossulae edged by the septa and from K.guangxiense (Zhang, 1986) by an inconspicuous counter fossula.
Geographic and stratigraphic range. K. obliquum is present in theIvorian of Belgium (Denayer et al., 2011), Poland (Poty et al., 2003),Ural Mountains (Ivanowski, 1967) and S China (Poty and Xu, 1996).In Turkey, the species is only known in the Mantar Tepe Mbr inYaricak Yayla and Tas�kent–Alanya road sections.
Family Bothrophyllidae Fomitchev, 1953Genus Caninophyllum Lewis, 1929Type-species. Cyathophyllum archiaci Milne-Edwards and Haime,
1852; Viséan of England.Diagnosis and remarks. See Poty (1981) and Von Schouppé
(1970).Caninophyllum cf. patulum (Michelin, 1846)Fig. 7F.Material. One single specimen from Yaricak Yayla.Description. The section is 31 mm in diameter (23 mm for the
tabularium) and shows 50 septa of both orders. The major septareach the axis of the corallum, are sinuous in the dissepimentariumand gently undulating in the tabularium. They are thickened in thecardinal parts of the tabularium but a thickening occurs somewhaton septa in the counter quadrants. The minor septa are well devel-oped but restricted to the dissepimentarium (less than 1/3 of themajor in length). The cardinal septum is shorter while the counterseptum is slightly longer than the other septa. Both cardinal andcounter fossulae are conspicuous. The dissepimentarium com-prises about ten rows of interseptal dissepiments, concentric, con-cave toward the axis or straight, rarely herringbone. The inner rowis thickened in continuity with the septa. The external wall is thinand regular.

384 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
Remarks. This single specimen has dimensions and number ofsepta similar to those of C. patulum (Michelin, 1846) but differsfrom the latter by major septa slightly thickened in the counterquadrants and a conspicuous counter fossula. With its relativelylong minor septa, this corallum is quite similar to Moliniacian spe-cies of Caninophyllum (described as C. patulum by Poty, 1981 andJavaux, 1994) but is smaller and has thicker septa. These charactersare, moreover, present in corals described as Caninophyllum tom-iense (Tolmachev, 1931) by Von Schouppé in Afghanistan.
Geographic and stratigraphic range. Caninophyllum cf. patulumwas collected in the lower part of the Mantar Tepe Mbr (Ivorian).
Genus Calmiussiphyllum Vassiljuk, 1959Type-species. Calmiussiphyllum calmiussi Vassiljuk, 1959; Lower
Viséan of the Donets Basin.Diagnosis. See Hill (1981).Remarks. Vassiljuk (1959) created the genus Calmiussiphyllum
for Caninophyllum-like corals having long major and minor septa.The length of the minor septa, together with a simpler dissepimen-tarium are the main features used to distinguish Calmiussiphyllumand Caninophyllum. The type-species – Calmiussiphyllum calmiussiVassiljuk, 1960 – occurs in the lower Viséan of the Donets Basin.Similar species were described in Siberia by Ivanowski (1967)under the name Caninia ussowi Gabunia, 1919 and in the Avesnois(France, Mansy et al., 1989).
Calmiussiphyllum cf. dobroljubovae (Flügel, 1963)Fig. 3G1–G5.Material. One single specimen (8 TS) from Yaricak Yayla.Description. Solitary trochoid corallum, 4.5 cm high, shows a
constriction and a rejuvenescence in the calyx. The diameter ofthe mature stages is maximum 30 mm. The tabularium has amean diameter of 14 mm (max. 24 mm). There are 44 septa ofboth orders (max. 52). The major septa are long, thick andspindle-shaped in the tabularium in the mature stages(Fig. 3G3–G4) but are also thickened in the dissepimentarium inthe juvenile stages (Fig. 3G2). In the juvenile stages, the majorsepta fuse in bundles reaching the axis but withdraw in themature stages (Fig. 3G1–G2). The central septa-free zone is about1/3 of the corallum diameter. The septa are pinnately arranged inthe cardinal quadrants but radially disposed in the counter quad-rants (Fig. 3G3). The cardinal septum is thicker and more tortuousthan the others. The minor septa are long but thinner than themajor and sometimes contratingent (Fig. 3G5). The cardinal fos-sula is deep and marked by the withdrawal of the dissepiments.The dissepimentarium is made of 3–8 rows of concentric, angu-lo-concentric and herringbone interseptal dissepiments. The innerrow is thickened in the mature stages. The external wall is thin(0.3 mm) and undulating.
Remarks. This specimen shares with Calmiussiphyllum calmiussiVassiljuk, 1960 long major septa thickened in the cardinal partsof the tabularium and long minor septa. The major differences withthis species are: smaller dimensions (up to 60 mm in diameter forC. calmiussi) and less number of septa (up to 65 in the type-spe-cies). The youngest observed stage shows a zaphrentoid septal pat-tern which has never been documented in Calmiussiphyllum but isquite common in other genera belonging to the BothrophyllidaeFamily (e.g. Caninophyllum). This corals share with Bothrophyllumdobroljubovae Flügel, 1963 from the Lower Viséan of the AlborzMountains a large dissepimentarium, septa thickened in the tabu-larium and contraclinant to contratingent minor septa. The Turkishspecimen is attributed to this species because of the similarity ofthe juvenile stages (compare Fig. 3G2 with FIügel’s fig. 3B andFig. 3G3 with Flugel’s fig. 3E). The major differences are thickersepta in the cardinal quadrants and a long counter septum in B.dobroljubovae.
Geographic and stratigraphic range. Calmiussiphyllum specieswhere described in the Lower Viséan (see above). The Turkish
specimen comes from the basal part of the Mantar Tepe Mbr, upperIvorian in age (Altıner and Özgül, 2001).
Suborder Aulophyllina Hill, 1981Family Aulophyllidae Dybowski, 1873Subfamily Amygdalophyllinae Grabau in Chi, 1935Genus Amygdalophyllum Dun and Benson, 1920Type-species. Amygdalophyllum etheridgei Dun and Benson,
1920; Viséan of New South Wales (Australia).Diagnosis and remarks. See Dun and Benson (1920), Semenoff-
Tian-Chansky (1974), Webb (1990) and Poty (2007).Amygdalophyllum sp.Fig. 7G and H1–H2.Material. Two eroded specimens (5 TS) from Yaricak Yayla.Description. Solitary corallum with a maximum diameter of
30 mm (16 mm for the tabularium) having 46–48 septa of bothorders. The major septa are long and confluent with the columella.They are spindle-shaped and maximum 0.4 mm thick. They areundulating in the dissepimentarium but straight in the tabularium,except their axial end that change in direction near the columella(Fig. 7G and H2). The minor septa are long (2/3 of the major inlength) and thin (0.2 mm) and slightly undulating. The counter sep-tum is connected to the columella (but broken in Fig. 7H1). The car-dinal septum is shortened. The cardinal fossula is shallow and open.The columella is almond-shaped in transverse section, composed ofan axial plate bearing many short septal lamellae up to 1 mm thick.The latter are arranged side by side all around the axial plate, exceptnear the cardinal end of the columella where they are lacking. Thedissepimentarium comprises about 10 rows of concentric and V-shaped interseptal dissepiments. The 2–3 inner rows are thickenedin continuity with the septa. The external wall is regular and thin.
Remarks. Amygdalophyllum with similar dimensions and num-ber of septa are: A. turbophylloides Semenoff-Tian-Chansky, 1974(25–30 mm, 50 major septa) and A. etheridgei Dun and Benson,1920 (35–40 mm, 50 major septa) but both are strongly thickenedand have a more complex dissepimentarium. A. inopinatumEtheridge, 1900 (30 mm) comprises more than 60 major septa. A.vallum Hill, 1934 is smaller (20 mm, 40–45 major septa) and hasthick minor septa. The Turkish specimens possess a similar numberof septa with that of A. kalawchense Von Schouppé, 1970 (40–44)and A. asselense Semenoff-Tian-Chansky, 1974 (45–50 major septa)but both are smaller (13–15 mm). A. praecursor (Howell, 1938) andA. sudeticum Zolinsky, 2000 are also smaller and have a roundedcolumella while the Turkish specimens show an almond-shapedone. The morphology of the columella is similar to that of A. col-umnare Pickett, 1966 but this species is smaller (20 mm, 30 majorsepta) and have short minor septa. In conclusion, the Turkish spec-imens do not fit with any Amygdalophyllum species in term ofdimensions, number of septa and morphology.
Distribution. This species was collected about 20 m above theTournaisian–Viséan boundary established by Altıner and Özgül(2001), in the lower part of Mantar Tepe Mbr in Yaricak Yayla.
Subfamily Heterocaniinae Hill, 1981Genus Kueichouphyllum Yü,1931Type-species. Kueichouphyllum sinense Yü, 1931; Upper Viséan of
Ghuizou, South China.Diagnosis. See Hill (1981).Remarks. Kueichouphyllum has been abundantly discussed (Yü,
1931, 1933, 1937; Wu and Zhao, 1989; Fontaine et al., 1991,etc.). The genus was considered as typically Viséan and its occur-rence in the Tournaisian was lately recognized (Zuo, 1986; Potyand Xu, 1996; Niikawa, 1994). Kueichouphyllum is a widely distrib-uted taxon defining the ‘Kueichophyllum Zone’ that covers South-East Asia, North-western Australia and the Middle East (see palae-obiogeographic discussion of the present paper).
Kueichouphyllum alborense Khaksar 1996Fig. 9A1–A2, B, C1–C3, D1–D3, and E.

Fig. 9. Kueichouphyllum species at the Tournaisian–Viséan transition in Central Taurides, Turkey. A–E: Kueichouphyllum alborense Khaksar, 1996. A1–A2: specimen YA.12.3,successive transverse sections, x3 (scale bar = 5 mm). B: specimen YB.9.1, transverse section in a juvenile stage, x3 (scale bar = 5 mm). C1–C3: specimen YC.1.2.II, successivetransverse sections, x3 (scale bar = 5 mm). D1–D3: specimen YC.1.4, D1: transverse section, x3 (scale bar = 5 mm), D2: longitudinal section, x3 (scale bar = 5 mm). D3: close-up view of the dissepimentarium and contratingent minor septa, x6 (scale bar = 2.5 mm). E: specimen YC.1.1, transverse section, x3 (scale bar = 5 mm). F: Kueichouphyllum cf.yabei Minato, 1943, transverse section, x3 (scale bar = 5 mm).
J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 385

Fidi
386 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
*
g. 1ame
1996
0. Scatterter for Ku
Kueichouphyllum alborense minor Khaksar, p. 101, pl.2, figs. 4–6.
1996
Kueichouphyllum alborense major Khaksar, p. 103, pl.3, figs. 1–4.1999
Kueichouphyllum alborense minor Khaksar; Heraviand Khaksar, p. 138, pl. 17, figs. 1–3, pl. 18, figs. 1–2.1999
Kueichouphyllum alborense major Khaksar; Heraviand Khaksar, p. 138, pl. 19, figs. 1–3.Holotype. Specimen GI-AA-83 (forma major), Tournaisian of theAlborz Mountains, Iran; Collection of the Geological Survey of Iran.
Material. Thirteen specimens (22 TS, 4 LS) from Yaricak Yayla.Diagnosis. Kueichouphyllum with a mean diameter of 40 mm and
having 70–90 septa of both orders. Cardinal septum slightlyshorter. Minor septa contraclinant or contratingent. Dissepimen-tarium narrow. Emended from Khaksar (1996) to include bothsub-species.
Description. The coralla are ceratoid to cylindrical 10–15cm-high. The axial structure is prominent in the calyx. The meandiameter is 27 mm (maximum 42 mm) while the tabularium is23 mm-large (maximum 40 mm). There are 65 septa of each orderon average (maximum 80, Fig. 10). The major septa are long andthick in the cardinal parts of the tabularium and thin in the counterquadrants (Fig. 8A1–C3). They are curved in the tabularium andsome group near the axis. The base of the septa is thick. The minorsepta are long (half as long as the major or more), thin and con-traclinant or contratingent (Fig. 9D3). In the juvenile stages, theyare short and strongly thickened at the base. The cardinal septumis shorter but thicker than the other septa. The cardinal fossula isnarrow and long, closed in the axial end and flanked by thicker car-dinal-lateral septa. The axial structure is half as wide as the tabu-larium diameter. It is made of numerous septal lamellae of variablelength and curvature (Fig. 8C2–C3). No lamella dominates in thisirregular structure. Some of them join the axial ends of major septaor abut on upturned axial tabellae. The density of axial tabellae ishigher in the periphery of the axial structure than in its centralpart. In juvenile stages (less than 10 mm in diameter), the axialstructure is sometimes elongated in the cardinal-counter plan(Fig. 9B). The dissepimentarium is narrow, comprises 2–7 rowsor concentric interseptal dissepiments in which the concavity isturned toward the axis or the external wall (Fig. 8D3). Occasionaloblique and V-shaped dissepiments occur. The inner row is thick-ened. In some specimens, another row situated in the periphery
diagram showing the number of septa plotted against corallumeichouphyllum alborense and Kueichouphyllum cf. yabei.
of the dissepimentarium is also thickened. The external wall is0.4 mm thick and regular. In longitudinal section, the tabulariumis complex. The axial tabellae are strongly divided, upturnedtoward the axis or domed (Fig. 8D2). The periaxial tabellae aredeclined and form a peripheral gutter. The dissepiments are glo-bose, 1–4 mm long and 1 mm high. They are gently inclined, somebeing sub-horizontal. There are about 10 tabulae and dissepimentsper centimetre.
Remarks. These specimens share with K. alborense Khaksar, 1996a similar diameter, a narrow dissepimentarium and – above all –contratingent minor septa. Khaksar (1996) distinguished twosub-species: K. alborense major (50 mm in diameter and 100 septaof both orders) and K. alborense minor (26 mm and 60 septa). Thediameter of specimens figured by Khaksar (1996) covers the rangeof both sub-species. Both of them share contratingent minor septaand a few septal lamellae connected to major septa. However, K.alborense minor shows typically juvenile characters such as apoorly developed dissepimentarium and septa thickened in bothcardinal and counter quadrants. These features suggest that K.alborense minor is the juvenile stage of K. alborense major. K. sinenseYü, 1931 and K. yabei Minato, 1943 can have similar dimensionsbut the contratingent minor septa are a diagnostic character allow-ing the easy identification of K. alborense.
Geographic and stratigraphic range. K. alborense is present in theupper Tournaisian Gerud Formation in the Alborz Mountains(Khaksar, 1996). In Turkey the species occurs in the limestone bedsforming the uppermost part of the Çit Yayla Mbr (Ivorian) and inthe Mantar Tepe Mbr (Moliniacian, VT1 zone of Altıner andÖzgül, 2001).
Kueichouphyllum cf. yabei Minato, 1943Fig. 9F.Material. Two specimens (2 TS) from Yaricak Yayla.Description. The corallum is 30 mm-large (26 mm for the tabu-
larium) and have 68–80 septa of both orders (Fig. 10). The majorsepta are long, straight in the dissepimentarium and the outer partof the tabularium but curved in the internal part of the tabulariumwhere they whirl in a vortex-like axial structure. They are thick-ened (0.7 mm) throughout the tabularium (Fig. 8F). The minorsepta are half as long as the major and enter into the dissepimen-tarium on 1 mm. They are thin and undulating in the dissepimen-tarium. The cardinal septum is short. The cardinal fossula is open,long and curved as its edges are parallel to the curved major septa.The axial structure is made by the axial ends of the major septajoined by upturned axial tabellae and some rare septal lamellaedisconnected from the septa. The dissepimentarium comprises 4–8 rows of concentric interseptal dissepiments. Their concavity iseither turned toward the axis or the external wall. The externalwall is usually eroded.
Remarks. The minor septa not contratingent allow to distinguishthese corals from K. alborense that has similar dimensions. More-over, the septa are thickened both in the cardinal and counterquadrants in this specimen while they are thickened only in thecardinal quadrants in K. alborense (compare Fig. 8C3 and F). Thevortex-like axial structure without any well-defined plate or lamel-lae is diagnostic for K. yabei Minato 1943 but the latter usuallycomprises much septa (70–120) and has shorter major septa.
Geographic and stratigraphic range. K. yabei is known in the Visé-an of S China (Wu and Zhang, 1980; Wu and Zhao, 1989), Japan(Minato, 1955), Thailand (Fontaine et al., 1991), Iran (Flügel,1963; Heravi and Khaksar, 1999) and in the Eastern Taurides (Kay-seri vicinity, Flügel and Kiratlioglu, 1956; Heritsch, 1941; Kato,1979). In Yaricak Yayla, K. cf. yabei occurs in the Mantar TepeMbr (Moliniacian, VT2 zone of Altıner and Özgül, 2001).
Kueichouphyllum sp.Fig. 11A1–A2.Material. One crushed specimen from Yaricak Yayla.

Fig. 11. Kueichouphyllum sp. and Eockoninckocarinia gemmina sp. nov. of the Lower Viséan of Central Taurides, Turkey. A1–A2: Kueichouphyllum sp. A1: specimen YB.6.1,transverse section, x2 (scale bar = 7.5 mm). A2: specimen YAsup.1, close-up view of the dissepimentarium, x3 (scale bar = 5 mm). B–E: Eokoninckocarinia gemmina sp. nov.,Moliniacian (Lower Viséan), Mantar Tepe Mbr, Yaricak Fm, Yaricak Yayla section, Central Taurides, Turkey. B1–B2: specimen YB.13.1, successive transverse sections, x3 (scalebar = 5 mm). C1–C2: specimen YB.13.3 (HOLOTYPE), successive transverse sections, x3 (scale bar = 5 mm). D: specimen YB. 13.7, transverse section, x3 (scale bar = 5 mm). E1–E5: specimen YB.13.5 (PARATYPE), E1: longitudinal section, x3 (scale bar = 5 mm), E2–3: successive transverse sections in a corallum with an offset, x3 (scale bar = 5 mm), E4:close-up view of the dissepimentarium and contratingent minor septa, x4 (scale bar = 3.7 mm), E5: close-up view of the careniform diverticulum in transverse section, x6(scale bar = 2.5 mm).
J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 387

Fig. 12. Statistical data of Eokoninckocarinia gemmina sp. nov. A: Scatter diagram showing the number of septa plotted against corallum. B: Frequency histogram of corallumdiameter. C: Frequency histogram of number of septa. A, B and C based on 28 measurements.
388 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
Description. Large solitary corallum, 20 cm high, 45 � 70 mm intransverse section, laterally crushed. There are 108 septa of bothorders. The major septa are long and reach the axial part of the tab-ularium. They are thin and slightly undulating in the peripheralpart of the dissepimentarium but thickened elsewhere. The minorsepta are short (less than ½ of the major length) and rarely enterinto the tabularium. The cardinal septum is short. The cardinal-lat-eral septa are strongly thickened and edged by arched dissepi-ments (Fig. 11A2). The axial structure is crushed and cannot bedescribed. The dissepimentarium is large and composed of 10–15rows of small concentric, angulo-concentric or rarely V-shapedinterseptal dissepiments. Several rows are thickened. The wall isnot preserved.
Remarks. The poor preservation of the specimen does not allowan accurate determination. The generic attribution is mainly basedon the morphology of the dissepimentarium which is very similarto that of Kueichouphyllum. The dimensions and number of septa ofthis specimen are sufficient to distinguish it from K. alborense pres-ent in the same beds. K. yabei Minato, 1943 and K. sinense Yü, 1931have similar dimensions but both have long minor septa.
Geographic and stratigraphic range. This specimen was collectedin the middle part of the Mantar Tepe Mbr in Yaricak Yayla. Theforaminifers associated with the corals indicate the Moliniacian(VT1 zone of Altıner and Özgül, 2001).
? Family Koninckocariniidae Dobroljubova, 1962Genus Eokoninckocarinia Wang in Zhao et al., 1984Type-species. Eokoninckocarinia xinjiangensis Wang in Zhao et al.,
1984; Middle Carboniferous of Xinjiang (China).Diagnosis. Corallum solitary or presenting a limited increase.
Major septa long extending to the axis, thin and strongly carinated.Minor septa long. Cardinal fossula conspicuous. Loose axial struc-ture. Dissepimentarium large, composed of concentric, angulo-concentric, V-shaped, herringbone and arched interseptal dissepi-ments. Seldom lonsdaleoid dissepiments. Tabulae complete ornot, horizontal or domed. Emended from Zhao et al. (1984).
Remarks. Wang in Zhao et al. (1984) established simultaneouslytwo new genera – Eokoninckocarinia and Carinokoninckophyllum –for corals with a Koninckophyllum morphology but carinated septa.Both genera are mono-specific and come from the same beds. Bothare characterized by a loose irregular axial structure, long cari-nated septa and a complex dissepimentarium. However, for thisauthor, Carinokoninckophyllum possesses rare lonsdaleoid dissepi-ments while Eokoninckocarinia has only interseptal dissepiments.The occurrence of lonsdaleoid dissepiments should be consideredas a morphological variation and does not justify the separation
of the two genera. The type species of Eokoninckocarinia xinjiangen-sis Wang in Zhao et al., 1984 is of Middle Carboniferous age but thegenus has been documented in the Lower–Middle Viséan by Xu(1996) in South China. Eokoninckocarinia is easily distinguishedfrom Kueichouphyllum by the abundant carinae and zig-zag septa.Similar structures are known in Koninckophyllum (e.g. K. magnifi-cum Thomson and Nicholson, 1876) but Eokoninckocarinia differsby longer septa and a non-persistent loose and irregular axialstructure.
Eokoninckocarinia gemmina sp. nov.Fig. 11B1–B2, C1–C2, D, and E1–E5.Etymology. From the latin gemmina for bud, in recognition of the
limited increase observed in this species.Holotype. Specimen YB.13.3 – Taurus 2009 (3 TS, 1 LS).Paratype. Specimen YB.13.5 – Taurus 2009 (2 TS) showing an
offset.Type horizon and locality. Sandy limestone in the upper part of
the Mantar Tepe Mbr (Yaricak Fm), Moliniacian, Yaricak Yayla sec-tion, Hadım vicinity, Central Taurides, S Turkey.
Material. Fifteen specimen (28 TS, 4 LS) including the type spec-imens, all from Yaricak Yayla.
Diagnosis. Small-sized Eokoninckocarinia, with a diameter of16 mm on average, having 40 septa of both orders. Septa stronglycarinated and zig-zag in the peripheral part of the dissepimentar-ium. Major septa long, axial ends confluent in a loose irregularaxial structure. Minor septa contraclinant or contratingent. Limitedlateral increase.
Description. The coralla are usually solitary but several show alimited lateral increase. They are up to 10 mm high and with a meandiameter of 16 mm (maximum 26 mm). The tabularium is 8 mm-large (maximum 11 mm). There are 40 septa of both orders on aver-age (maximum 51, Fig. 12). The major septa are thin, long and reachthe axis where they group in bundles of 3–4 septa before joining theaxial structure (Fig. 11B2). In some specimens, the septa are with-drawn and leave a free zone around the axial structure. The minorsepta are short (1/4–1/3 of the major length) and enter only a shortdistance into the tabularium. They are contraclinant or contratin-gent (Fig. 11E4). All the septa are thickened near the inner edge ofthe dissepimentarium. In the peripheral part of the dissepimentar-ium they bear numerous strong carinae, producing a tortuous rag-ged zig-zag look to the septa (Fig. 11E5, ‘careniform diverticulum’ ofSemenoff-Tian-Chansky, 1974). The cardinal septum is oftenshorter. The cardinal fossula is well marked in the juvenile stagesbut tends to become inconspicuous in mature stages. The axialstructure is irregularly developed and can occupy 1/6–1/3 of the

J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 389
tabularium. The axial structure is similar to that of Kueichouphyllumand made of a loose axial plate (particularly in the juvenile stages,Fig. 11E3) joined by the axial ends of the major septa. The dissepi-mentarium is composed of 2–15 rows of concentric, V-shaped andherringbone interseptal dissepiments. In the peripheral part of thedissepimentarium, the dissepiments arrangement is strongly dis-turbed by the carinae. The inner row of dissepiments is often thick-ened. The external wall, rarely preserved, is thin and regular. Thetabulae are incomplete. The axial tabellae are domed, flat or gentlyconcave. They sag toward the periphery where they pass to smallerand globose periaxial tabellae. The dissepiments are small, inclinedtoward the tabularium at 40–50�. There are 14–20 axial tabellaeand 20 dissepiments per centimetre. Several specimens show a lat-eral non-parricidal increase (Fig. 11E2–3). The offset appears in theperipheral part of the dissepimentarium and remains attached to itsparent until its diameter reaches 10 mm. The earliest stages ofdevelopment were not observed.
Remarks. These specimens show the typical carinated septa,loose axial structure and complex dissepimentarium of Eokoninck-ocarinia Wang in Zhao et al., 1984. E. xinjiangensis Wang in Zhaoet al., 1984 and E. carinata (Xu, 1981) are larger (22 mm and 20–30 mm in mean diameter) and have no contratingent minor septa.Increase was not documented in these species. The creation of thisnew species is thus justified.
Geographic and stratigraphic range. Eokoninckocarinia gemmina isonly known from the uppermost part of the Mantar Tepe Mbr inthe Yaricak Yayla section. The poor foraminiferal assemblage ofthis level indicates the Moliniacian (MFZ11 biozone of Poty et al.,2006; VT3 zone of Altıner and Özgül, 2001). Ünsalaner-Kiragli (inDemirtas�lı et al., 1978a) described from the Eastern Taurides ‘Pal-aeosmilia-like corals different from typical Palaeosmilia’. Dronovet al. (1979) noticed in Afghanistan Palaeosmilia cf. nodosa withextreme carinae. Both are possibly Eokoninckocarinia species, mis-identified at that time.
Fig. 13. Comparison between the coral zonation of Western Europe (Poty et al.,2006), South China (Tan et al., 1987, emended by Xu and Poty (1997)) and CentralTaurides. Foraminifers zones (Poty et al., 2006; Hance et al., 2011) are indicative forWestern Europe and China. Foraminifers zones of Central Taurides are from Altınerand Özgül (2001). Legend: Chrono.: chronostratigraphy.
5. Biostratigraphy
The carbonate and coral-rich parts of the Yaricak Yayla section,presented in the present paper is covered by the four lower bioz-ones defined by Altıner and Özgül (2001) in this locality. Theseare the pre-VT1, VT1 (2 subzones), VT2 (2 subzones) and VT3 zones(see below). In this interval, four successive coral assemblageswere recognized in the Yaricak Yayla and Tas�kent–Alanya road sec-tions (Figs. 2 and 13).
Proheterelasma omaliusi assemblage. The oldest corals found inthe Yaricak Yayla and Tas�kent–Alanya road sections occur in thesandy cherty limestone horizon within the thick shaly sequenceof the Çit Yayla Mbr. These are Uralinia sp. and Proheterelasma oma-liusi. These corals are not particularly useful in biostratigraphy. P.omaliusi appears in the Ivorian RC3 zone of Poty et al. (2006) whilesmall Uralinia are very rare after the Hastarian RC2 biozone. Thisfirst assemblage corresponds possibly to the transition of the bioz-ones RC2 and RC3, i.e. uppermost Hastarian to lowermost Ivorian.Foraminifers described from this interval (pre-VT1) by Altıner andÖzgül (2001) are poorly diversified and of no biostratigraphicalvalue too (small Earlandia).
Uralinia multiplex assemblage. The alternation of shale andlimestone forming the upper part of the Çit Yayla Mbr yieldsnumerous Uralinia multiplex, Caninia cornucopiae and Zaphrentitesdelanouei. This assemblage can be considered as an impoverishedequivalent of the RC3a zone of Poty et al. (2006, WesternEurope) and with the Uralinia zone of Tan et al. (1987, SouthChina) and is Ivorian in age. The foraminiferal association(pre-VT1) is dominated by earlandiids and unidentified small end-othyrids after Altıner and Özgül (2001).
Kueichouphyllum alborense assemblage. This assemblage isdefined by the first appearance of K. alborense near the base ofthe Mantar Tepe Mbr, together with the entry of Keyserlingophyl-lum obliquum. A remarkable horizon containing very large Uralinia(up to 10 cm in diameter) is a regional marker bed noticed byÖzgül (1997) as the ‘Siphonophyllum bed’. The assemblage also con-tains Siphonophyllia sp., Calmiussiphyllum cf. dobroljubovae, Canino-phyllum cf. patulum, Kueichouphyllum cf. yabei, K. sp., Bifossulariasp., and Amygdalophyllum sp. Except in the upper part, the MantarTepe Mbr is almost entirely represented by this assemblage even if

390 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
the upper 2/3 of the member is very poor in corals. The Kueichou-phyllum alborense assemblage is correlated with some difficulty(due to its poverty in guide taxa, e.g. Sychnoelasma, Rylstonia, Pala-eosmilia) with the European RC4 Zone of Poty et al. (2006) and withthe Chinese Keyserlingophyllum Zone of Tan et al. (1987) and isuppermost Ivorian to Moliniacian in age. The Kueichouphyllumalborense assemblage is covered by the VT1 and VT2 (Uralodis-cus–Glomodiscus Zone) foraminiferal assemblage of Altıner andÖzgül (2001) which can be considered as equivalent to the MFZ9and MFZ10 p.p. zones of Poty et al. (2006). The Tournaisian–Viséanboundary established by Altıner et al. (2007) based on the entry ofEoparastaffella simplex and associated microfauna, within bed 58, issituated 2 m above the base of the Mantar Tepe Mbr. None of thecorals allow the identification of this boundary since all taxa (Amy-gdalophyllum, Kueichouphyllum, Keyserlingophyllum) are known inthe Tournaisian and Viséan.
Eokoninckocarinia gemmina assemblage. The uppermost 25 m ofthe Mantar Tepe (unit YBsup) yielded the single species Eokonick-ocarinia gemmina. The genus Eokoninckocarinia is known in theLower–Middle Viséan Keyserlingophyllum–Dorlodotia Interval Zoneof Tan et al. (1987) in South China. This monospecific association isimpossible to correlate with the biozones of Poty et al. (2006). Theforaminifers identified by Altıner and Özgül (2001) in the YBsupsandy limestone define their VT3 (Endothyranopsis compressa – Lat-iendothyranopsis menneri solida Zone) which can be considered asequivalent to the MFZ10 p.p. and MFZ11 zones of Poty et al.(2006), i.e. the upper part of the Moliniacian sub-stage.
These four assemblages have a local utility for correlation buttheir use outside the studied region as yet untested due to the lackof data in the Eastern Taurides.
6. Affinity and diversity of the coral assemblages
In recent palaeogeographic models (e.g. Stampfli and Borel,2002; Stampfli et al., 2013; Fig. 14), the Cimmerian Terrane formsa ribbon edging the Asian margin of Gondwana that was later (LatePermian times) rifted and drifted northward and assembled in theAlpine orogen after the Triassic. The remains of the Gondwanamargin are now situated in the Turkish Taurides, the Iran Blocks(e.g. Sanadaj-Sirjan, Alborz, Lut-Tabas Block), Afghanistan (FarahRud, Helmland, South Pamirs) and Tibet (Lhasa, Qiantang and Bao-shan Blocks). These are included in the ‘Kueichouphyllum Zone’ or‘Kueichouphyllum Sea’ (see Niikawa, 1994) where this genus iscommon and diversified. In the southern margin of the Palaeote-thys Ocean, the ‘Kueichouphyllum Zone’ covers the Cimmerian Ter-rane but also the Bonaparte Basin (North-western Australia; Hill,1973) and possibly the Arabian Plate (Denayer and Hos�gör,2014). In the northern part of this ocean, Kueichouphyllum occursin the Chinese blocks, Japan, Tarim, Turan and possibly in thesouthern part of Kazakhstan but the specimens figured byBykova (1966) are supposed Serpukhovian in age while the taxo-nomic affinity of those cited but not figured by Shchukina (1975)needs to be re-established. Wang et al. (2003) refined the defini-tion and designed a Kueichouphyllum–Siphonophyllia–Keyserlingo-phyllum fauna which dominate the coral assemblage of the‘Kueichouphyllum Zone’. This alternative name is more correctbecause it indicates not only a geographical distribution but alsothe ecological niche shared by the three genera in the Zone duringUpper Tournaisian–Lower Viséan times. The latter correspond tothe A2 (low diversity level-bottom community) subtype in theclassification of Aretz (2010) with the particularity of thedominance of often abundant specimens of these three genera.Below are summarized the faunal composition of the tectonicblocks belonging to the Asian margin of Gondwana for the Upper
Tournaisian–Lower Viséan transition. This summary highlightsseveral observations that are discussed further.
6.1. Turkish Taurides
During Upper Tournaisian times, the coral assemblage of theTaurides included only solitary taxa belonging to the generaCaninia, Calmiussiphyllum, Caninophyllum, Keyserlingophyllum,Kueichouphyllum, Siphonophyllia, Uralinia and the undissepimentedcorals Proheterelasma and Zaphrentites. Similarly, the Lower Viséanfauna is relatively poorly diversified: Amygdalophyllum, Bifossularia,Eokoninckocarinia, Keyserlingophyllum and Kueichouphyllum. Mostof them has a wide spatial distribution throughout Eurasia andare of little interest for palaeobiogeography. Eokoninckocarinia,Calmiussiphyllum and Bifossularia are insufficiently known to proveany utility in palaeobiogeography but their presence in the Tauridesmight suggest a connection with northern Palaeotethys provinces(Western and Eastern Europe). The Upper Tournaisian entry ofKueichouphyllum alborense is remarkable, as it is one of the oldestoccurrences of this genus (as in Iran; Niikawa, 1994, 2012).
6.2. Armenia
The Transcaucasus block yielded an interesting assemblage ofUpper Tournaisian–Lower Viséan corals including Cyathoclisia,Keyserlingophyllum, Kueichouphyllum, Siphonophyllia and Uralinia(Papojan, 1969, 1977) and Calmiussiphyllum (=? Bothrophyllum inFlügel, 1963). Papojan (1977) documented Koninckophyllum andDibunophyllum in the Lower Viséan but both being typicalUpper Viséan taxa, they are most probably misidentified (=?Amygdalophyllum) and are not taken into account here.
6.3. Arabian Plate
The Upper Tournaisian–Lower Viséan coral assemblage of theArabian Plate is documented in the Um Bogma Formation of theSinai Peninsula (North-East Egypt) by Kora (1989, 1992, 1995)and Kora and Jux (1986); and in the Koprülü Formation in theHakkari area (South-East Turkey) by Denayer and Hos�gör (2014).Both faunas are dominated by undissepimented solitary rugosecorals (Amplexizaphrentis, Bradyphyllum, cf. Gorizdronia, Rotiphyl-lum, Zaphrentites and possibly Amplexocarina, see discussion inDenayer and Hos�gör, 2014) and the dissepimented genus Caninia.Their utility in palaeobiogeography is less than the Tauridesassemblage because they form a strongly facies dependant ‘Cya-thaxonia fauna’ occurring in both cases in mixed carbonate-silici-clastic facies. Hakkari and Sinai were part of the northern marginof Gondwana but were not rifted with the Cimmerian Terrane.They reached their current position during the Cenozoic AlpineOrogeny when Gondwana and Eurasia collided (S�engör andYılmaz, 1981).
6.4. Iran
Tournaisian–Viséan coral assemblages have been described fromseveral distinct tectono-stratigraphic units currently forming Iran:the Alborz Block (Northern Iran) and in the Lut-Tabas Block. The firstis the best documented by Flügel (1963, 1991), Khaksar (1996) andHeravi and Khaksar (1999). The assemblage consists of solitarycorals of the genera Amplexizaphrentis, Calmiussiphyllum (=? Bothro-phyllum in Flügel, 1963), Caninia, Caninophyllum, Cyathoclisia(=Teheranophyllum in Khaksar, 1996), Kueichouphyllum, Keyserlingo-phyllum (=Humboldtia in Flügel, 1963), Siphonophyllia, Zaphrentites,Zaphriphyllum, and Amygdalophyllum (=? Carruthersella in Heraviand Khaksar, 1999). Other taxa documented by the latter authorsare questionably misidentified but too badly figured for an accurate

Fig. 14. Early Carboniferous (c. 350 Ma, Tournaisian–Viséan transition) reconstruction of the Palaeotethys Ocean and its margins (modified from Stampfli and Borel, 2002;Stampfli et al., 2013 with some additions from Golonka, 2007; Torsvik and Cocks, 2013; Metcalfe, 2013) with position of the discussed areas along the Asian margin ofGondwana. Legend: Adr: Adria, Afr: Africa, Alp: Alpes (Helvetia, Cetic), Ana: Anatolia, Ant: Antarctica, Ara: Arabian Plate, Arm: Armorica, Aus: Australia, Bet: Betic, Bri: BritishIsles, Bru: Brunswick, Cat: Catalonia, Cib: Central Iberia, Cpt: Carpathian, Don: Donets, Eau: Eastern Australia, EMB: East Malaya Block, GC: Greater Caucasus, Hel: Hellenic,Ich: Indochina, Ind: India, IZZ: Istanbul–Zonguldak Zone, Jap: Japan, Kab: Kabylia, Kaz: Kazakhstania (not detailed), KT: KarakumTuran, Kuz: Kuznetsk, MC: Massif Central,Meg: Meguma, Mes: Meseta, MGR: Mid-German crystalline Ride, MM: Montagne Noire-Maures, Moe: Moesia, Mol: Moldanubia, Nam: North America, Nch: North China,NDB: Namur–Dinant Basin, NG: New Guinea, Noz: Novaya Zemlya, NPa: North Pamir, Nsc: Nova Scotia, OM: Ossa-Morena, Pol: Poland, Py: Pyrenees, RHN: Rheno-hercynianOcean, Rif: Moroccan Rif, Rus: Russian Platform, Sak: Sakarya Zone, Sam: South America, Sch: South China, Sib: Siberia (not detailed), Sx: Saxo-Thuringia, Tm: Tarim Basin,Ural: Ural, Wbu: Western Burma Block, WSu: Western Sumatra. Localities with Upper Tournaisian and Lower Viséan corals are indicated by *. Gondwanan localities, south ofthe Cimmerian Terrane discussed in the main text are: 1: Moroccan Tafilalt and Algerian Béchar Basin, 2: Sinai Peninsula, 3: Hakkari area (South-eastern Turkey), 4: Laristan(South-western Iran), 5: South-east Pakistan, 6: Himalayan Block, 7: Bonaparte Basin (North-western Australia).
J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 391
interpretation. Recently, Ogar et al. (2013) figured from the LowerViséan Mobarak Formation Alborzia, Bifossularia, Clisiophyllum,Kiyassarophyllum (a genus close to Arachnolasma), Rotiphyllum andSpirophyllum (=Amygdalophyllum in Ogar et al., 2011), in additionto the previously mentioned genera. This higher diversity is partlyexplained by the width of the sampling window which coversmainly the Lower Viséan but also the lower part of the Middle Viséanas testified by the presence of Hexaphyllia and Clisiophyllum gar-woodi. Poty (unpublished data) also collected Conilophyllum. Thesecorals occur within the carbonate facies of the Mobarak Formation(Brenckle et al., 2009; Khaksar, 1996). In the Lut-Tabas Block,Flügel (1991) identified a ‘Cyathaxonia fauna’ that comprisedAmplexocarinia, Claviphyllum, Cyathaxonia, Pentaphyllum, Plerophyl-lum, Pseudowannerophyllum, Rotiphyllum, Soshkineophyllum and
Ufimia in the Lower Viséan part of the Sishtu Formation, the onlydissepimented genus being Siphonophyllia. Additionally, Douglas(1950) described from Baluchistan (Lut-Tabas Block) a fauna com-posed of Allotropiophyllum, Caninia, Siphonophyllia and Zaphrentites.Siphonophyllia was also reported from Laristan (Zagros, ArabianPlate) by this author.
6.5. Afghanistan
Three distinct rugose coral faunas were documented in threedifferent tectonic blocks of Afghanistan. Dronov et al. (1979) andFontaine and Semenoff-Tian-Chansky (1977) documented in car-bonate facies of central Afghanistan (Helmand and Farah-RudBlocks) the following genera: Amplexizaphrentis, Amplexus, Caninia,

392 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
Caninophyllum, Cyathoclisia, Kueichouphyllum, Proheterelasma, Roti-phyllum, Siphonophyllia, Stereolasma, Ufimia, Uralinia and Zaphren-tites. Dronov et al. (1979) also reported Palaeosmilia cf. nodosawith extreme carinae which is questionably a Eokoninckocarinia,misidentified by these authors. In Badakshan (South Pamirs Block),the association comprises Amygdalophyllum, Caninophyllum, Pro-heterelasma and Zaphrentites (Von Schouppé, 1970).
6.6. Pakistan and India
Latif (1970) indicated the presence of Amplexus? from probableLower Carboniferous beds in North Pakistan (northern margin ofIndian subcontinent) but the latter is of poor interest in palaeobi-ogeography. The Yali Formation (limestone and shale) of the Hima-layan Terrane yielded an undissepimented fauna composed ofBeichuanophyllum, Commutia, Hapsiphyllum, Longiclava, Pentaphyl-lum, Ufimia, Yichunophyllum and Zaphrentites (Wang et al., 2003;Fan et al., 2003). Smith and Xu (1988) documented the dissepi-mented genera Caninia and Caninophyllum in the Lower (?) Viséanstrata of the Himalayan Terrane.
6.7. Tibet
Several blocks of Cimmerian affinity were accreted to the Chi-nese terranes during the Triassic (Wang et al., 2003 and referenceswithin). Upper Tournaisian to Lower Viséan coral associations areknown from two of them: the Lhasa and Qiantang blocks. Withinthe Lhasa Block, the carbonate Ponga Formation includes onlythe undissepimented taxa Amplexus, Barrandeophyllum, Cyathaxo-nia, Hapsiphyllum, Homalophyllites, Longiclava, Meniscophyllum,Mirusophyllum, Pentaphyllum, Stereolasma, Ufimia and Zaphrentites(Smith and Xu, 1988; Wang et al., 2003; Fan et al., 2003). Flügel(1966) also described from this area Amplexus, Caninophyllumand Siphonophyllia but these taxa might be younger. The fauna ofthe Yueyahu Formation (limestone and shale) in the QiantangBlock includes the genera Bradyphyllum, Caninia, Groenlandophyl-lum, Keyserlingophyllum (=Pseudohumboldtia in Xia and Liu, 1997,Humboldtia and Parahumboldtia in Fan et al., 2003), Siphonophyllia,Ufimia, Zaphrentites and Zaphriphyllum (=Ekvasophylloides in Fanet al., 2003).
6.8. Baoshan
The Tournaisian–Viséan Shiuhadong Formation crops out in thesouthern part of the Baoshan Block in South China (Wang et al.,2001). This carbonate formation yielded a diversified coral faunacomposed of Amplexus, Amygdalophyllum, Caninophyllum (=Siedlec-kia), Carruthersella, Conilophyllum, Cyathoclisia, Keyserlingophyllum,Kueichouphyllum, Lophophyllum, Siphonophyllia, Uralinia, Zaphren-tites and the colonial genus Heterostrotion. Moreover, Wang et al.(2001) documented a ‘Cyathaxonia fauna’ composed of Claviphyl-lum, Commutia, Cyathaxonia, Neozaphrentis?, Pentaphyllum, Ufimiaand Zaphrentites.
Fig. 15. Rugose coral diversity along the Cimmerian Terrane for the UpperTournaisian and Lower Viséan. The dotted line represents the general tendancy(linear regression of the number of occurrences of each locality). See Fig. 14 forlegend and references.
6.9. Western Australia and Sibumasu (Siam-Burma-Malaya-Sumatra)
Hill (1973) reported Kueichouphyllum in the Bonaparte Basin ofNorth-western Australia. Scrivenor (1931) noted Cyathaxonia,Siphonophyllia and a clisiophyllid in the Lower Carboniferous ofMalaya but these taxa are possibly Upper Viséan in age. The coralsdocumented by Smith (in Muir-Wood, 1948) are mainly UpperViséan, except Amygdalophyllum which could be Lower Viséan.Fontaine (1989) reported Zaphrentites from the Viséan of westernMalaya.
7. Discussion
Fig. 15 (and Table 1) summarizes the occurrence of the commongenera along the Cimmerian Terrane blocks discussed above. Asstated by Smith and Xu (1988), no endemic fauna is known fromthe Gondwanan Terrane and from the Cimmerian blocks. The dis-tribution of taxa is rather homogeneous but there is a cleardecrease of diversity from the East (Tibet, Baoshan) to the West(Armenia, Turkey, Iran). This East to West trend is obvious eitherin dissepimented corals or in the ‘Cyathaxonia fauna’, which rein-forces the interpretation of a longitudinal gradient (dotted line inFig. 15, drawn as a linear regression of the number of occurrencesof each locality). Sample biases (pseudo-absence, lack of outcrop),availability and quality of the studies, and taxonomy issues (taxo-nomical conceptions between authors, preservation of the fossils)might be evoked explaining the higher diversity of the Chineseareas, but the occurrence of a higher quantity of dissepimented sol-itary corals and – above all – colonial corals also pleads for an gen-uine East to West trend. The occurrence of the colonial genusHeterostrotion in the southern part of Baoshan Block is interpretedby Wang et al. (2001) as an influence of the closely situated Chi-nese blocks (Cathaysian continent) at this time and witness anoverlap of the two biogeographic provinces. Alternatively, it mightwitness a lower latitude location of this block. The lack of colonialrugose corals in the other blocks forming the Cimmerian Terrane isremarkable. In Western Europe, the Moliniacian yields the colonialgenera Dorlodotia, Solenodendron and Siphonodendron while inSouth China Kwangsiphyllum, Stylostrotion and Heterostrotion areabundant and diversified. What caused this diversity pattern alongthe Gondwana margin at this time? Hypothesis involving faciesdistribution, climate, currents and ecology are here tested.
7.1. Facies and palaeoclimate
Different types of facies were documented in the discussed area,from carbonate grainstone, to shale for the Tournaisian (referencesabove) but the Lower Viséan is dominated by shallow water car-bonate facies (except for ‘Cyathaxonia fauna’-bearing beds) wit-nessing a platform or a ramp edging the margin of Gondwana.No obvious trend was noticed in the facies distribution betweenthe eastern Chinese blocks and the Turkish Taurides. Oolitic faciesand evaporite pseudomorphs were documented in the Taurides
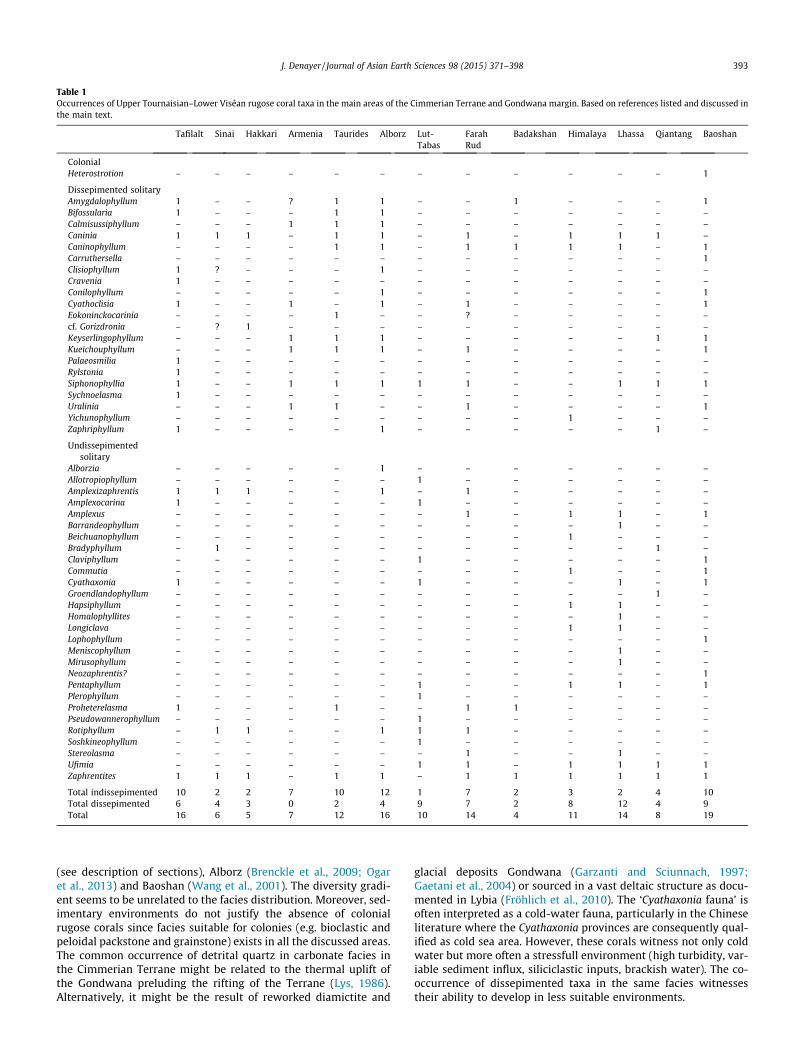
Table 1Occurrences of Upper Tournaisian–Lower Viséan rugose coral taxa in the main areas of the Cimmerian Terrane and Gondwana margin. Based on references listed and discussed inthe main text.
Tafilalt Sinai Hakkari Armenia Taurides Alborz Lut-Tabas
FarahRud
Badakshan Himalaya Lhassa Qiantang Baoshan
ColonialHeterostrotion – – – – – – – – – – – – 1
Dissepimented solitaryAmygdalophyllum 1 – – ? 1 1 – – 1 – – – 1Bifossularia 1 – – – 1 1 – – – – – – –Calmisussiphyllum – – – 1 1 1 – – – – – – –Caninia 1 1 1 – 1 1 – 1 – 1 1 1 –Caninophyllum – – – – 1 1 – 1 1 1 1 – 1Carruthersella – – – – – – – – – – – – 1Clisiophyllum 1 ? – – – 1 – – – – – – –Cravenia 1 – – – – – – – – – – – –Conilophyllum – – – – – 1 – – – – – – 1Cyathoclisia 1 – – 1 – 1 – 1 – – – – 1Eokoninckocarinia – – – – 1 – – ? – – – – –cf. Gorizdronia – ? 1 – – – – – – – – – –Keyserlingophyllum – – – 1 1 1 – – – – – 1 1Kueichouphyllum – – – 1 1 1 – 1 – – – – 1Palaeosmilia 1 – – – – – – – – – – – –Rylstonia 1 – – – – – – – – – – – –Siphonophyllia 1 – – 1 1 1 1 1 – – 1 1 1Sychnoelasma 1 – – – – – – – – – – – –Uralinia – – – 1 1 – – 1 – – – – 1Yichunophyllum – – – – – – – – – 1 – – –Zaphriphyllum 1 – – – – 1 – – – – – 1 –
Undissepimentedsolitary
Alborzia – – – – – 1 – – – – – – –Allotropiophyllum – – – – – – 1 – – – – – –Amplexizaphrentis 1 1 1 – – 1 – 1 – – – – –Amplexocarina 1 – – – – – 1 – – – – – –Amplexus – – – – – – – 1 – 1 1 – 1Barrandeophyllum – – – – – – – – – – 1 – –Beichuanophyllum – – – – – – – – – 1 – – –Bradyphyllum – 1 – – – – – – – – – 1 –Claviphyllum – – – – – – 1 – – – – – 1Commutia – – – – – – – – – 1 – – 1Cyathaxonia 1 – – – – – 1 – – – 1 – 1Groendlandophyllum – – – – – – – – – – – 1 –Hapsiphyllum – – – – – – – – – 1 1 – –Homalophyllites – – – – – – – – – – 1 – –Longiclava – – – – – – – – – 1 1 – –Lophophyllum – – – – – – – – – – – – 1Meniscophyllum – – – – – – – – – – 1 – –Mirusophyllum – – – – – – – – – – 1 – –Neozaphrentis? – – – – – – – – – – – – 1Pentaphyllum – – – – – – 1 – – 1 1 – 1Plerophyllum – – – – – – 1 – – – – – –Proheterelasma 1 – – – 1 – – 1 1 – – – –Pseudowannerophyllum – – – – – – 1 – – – – – –Rotiphyllum – 1 1 – – 1 1 1 – – – – –Soshkineophyllum – – – – – – 1 – – – – – –Stereolasma – – – – – – – 1 – – 1 – –Ufimia – – – – – – 1 1 – 1 1 1 1Zaphrentites 1 1 1 – 1 1 – 1 1 1 1 1 1
Total indissepimented 10 2 2 7 10 12 1 7 2 3 2 4 10Total dissepimented 6 4 3 0 2 4 9 7 2 8 12 4 9Total 16 6 5 7 12 16 10 14 4 11 14 8 19
J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 393
(see description of sections), Alborz (Brenckle et al., 2009; Ogaret al., 2013) and Baoshan (Wang et al., 2001). The diversity gradi-ent seems to be unrelated to the facies distribution. Moreover, sed-imentary environments do not justify the absence of colonialrugose corals since facies suitable for colonies (e.g. bioclastic andpeloidal packstone and grainstone) exists in all the discussed areas.The common occurrence of detrital quartz in carbonate facies inthe Cimmerian Terrane might be related to the thermal uplift ofthe Gondwana preluding the rifting of the Terrane (Lys, 1986).Alternatively, it might be the result of reworked diamictite and
glacial deposits Gondwana (Garzanti and Sciunnach, 1997;Gaetani et al., 2004) or sourced in a vast deltaic structure as docu-mented in Lybia (Fröhlich et al., 2010). The ‘Cyathaxonia fauna’ isoften interpreted as a cold-water fauna, particularly in the Chineseliterature where the Cyathaxonia provinces are consequently qual-ified as cold sea area. However, these corals witness not only coldwater but more often a stressfull environment (high turbidity, var-iable sediment influx, siliciclastic inputs, brackish water). The co-occurrence of dissepimented taxa in the same facies witnessestheir ability to develop in less suitable environments.

394 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
7.2. Palaeogeography and palaeoceanography
During Lower Carboniferous times, the Asian margin of Gondw-ana was situated at c. 50�S palaeolatitude after Stampfli et al.(2013), c. 40�S after Torsvik and Cocks (2013), or c. 30�S afterGolonka (2007). But in all palinspastic reconstructions, the Cimme-rian Terrane is approximately parallel to the latitude or slightlyoblique, the westernmost part lying in a lower latitude than theeastern part. The latitudinal climatic belt justifies the lower diver-sity of the Cimmerian Terrane compared to the Eurasian area (seebelow) but fails to explain satisfactory the diversity East to Westgradient. Different climatic conditions along the Gondwana marginare another hypothesis but, contrarily to the Pennsylvanian, fewstudy deals with Mississippian climate. Only a few climate modelshave been published (Mii et al., 1999; Webb, 2000; Aretz et al.,2013, 2014). In the traditional climatic models for the Late Palaeo-zoic, the Palaeotethys is run by counter-clockwise currents: a pal-aeo-equatorial westward drift along the Laurussian Palaeotethysand ending in cool or cold water currents running along the north-ern margin of Gondwana in a westward direction then mixing upwith the cool palaeo-tropical currents flowing eastward (e.g. Miiet al., 1999; Isaacson et al., 2008). In an opposite view, Ogar et al.(2013), following Brenckle et al. (2009) suggested the occurrenceof a warm current flowing westwards the Iranian coast of Gondw-ana to explain the higher diversity of this area. The geometry of thewestern part of Palaeotethys remains problematic and the width ofthe oceanic pathway between Africa and Euramerica is question-able (see Nance et al., 2010 and references within) but of greatimportance to understand the palaeo-currents.
The South-eastern Asian palaeo-archipelago (South Chinablocks) was often considered as a diversity hotspot and the cradleof many taxa. However, Poty et al. (2011) demonstrated thatalmost all the colonial genera common to Western Europe andSouth-eastern China appear earlier in Europe while the colonialgenera originated in China never reached Europe. This observationis rather contradictory with the counter-clockwise currents model.An alternative model should consider currents bypassing the Chi-nese blocks before running westwards to the Palaeotethys butagain, the geometry of the land mass and the direction of main cur-rents remain questionable. The current running eastward along theGondwana margin should have allowed the dispersion of the Wes-tern Europe taxa toward the east to the Cimmerian Terrane. But thetypical Upper Tournaisian and Lower Viséan corals of Western Eur-ope are limited to North-western Africa (Tafilalt and Béchar Basins;Aretz et al., 2013). Two hypotheses can explain the restriction. Theeastward flowing current might have been cold and have enabledthe survival of the coral larvae and thus the development of colo-nial coral along the Gondwana margin. This cold- (or at least cool-)water eastward current might also have restricted the expansion ofKueichouphyllum toward North Africa and Western Europe. Thesecond hypothesis explaining the separation of the two faunasinvolved a physical barrier. A regional siliciclastic input on theshelf might have formed such a barrier. In Lybia for example, theUpper Tournaisian and Lower Viséan deposits are represented byproximal deltaic and fluvial environments (Fröhlich et al., 2010)unsuitable for corals. This might be the barrier limitating west-wards the dispersion of the Kueichouphyllum fauna. West of thishypothetical barrier, in the Algerian Béchar Basin and MoroccanTafilalt, the corals are typical of the Western Europe Province ofSando (1990) and dominated by solitary rugose corals: Amygdalo-phyllum, Aulokoninckophyllum, Caninia, Cyathoclisia, Rylstonia,Siphonophyllia, Zaphriphyllum after Semenoff-Tian-Chansky, 1974;with Amplexocarinia, Amplexizaphrentis, Bifossularia, Cravenia, Cya-thaxonia, Merlewoodia, Palaeosmilia, Sychnoelasma and Zaphrentites(unpublished data of Aretz and Denayer) but not a single colonialgenus – since the specimen of Siphonodendron martini in Aretz
(2011) should be considered as Middle Viséan (Aretz, pers. com.,January 2014).
7.3. Palaeobiogeography
As stated above, the Gondwana margin is characterized, by anUpper Tournaisian–Lower Viséan assemblage of solitary rugosecorals dominated by Caninia, Kueichouphyllum, Keyserlingophyllum,Siphonophyllia and Uralinia, together with some less common taxa(Amygdalophyllum, Bifossularia, Calmiussiphyllum, Caninophyllum,Cyathoclisia, Eokoninckocarinia, Rotiphyllum, Zaphriphyllum) andundissepimented corals. Most of them are widely distributed taxaextending throughout the whole Palaeotethys at this time andforming the bulk component of several palaeobiogeographic prov-inces. This wide distribution is only partly explained by good mar-ine communications linked to high relative sea level (Somerville,2008). Explaining such cosmopolitanism requires a comparisonwith other invertebrates. Leighton (2005) demonstrated with Car-boniferous brachiopods and Crame (2000) with Recent bivalvesthat 70% of the genera occupy the inter-tropical belt extendingbetween 0� and 25� on both sides of the (palaeo-)Equator, whilethe remaining 30% occupy the belts extending beneath the 30� oflatitude. The first represent the tropical fauna living in the strictconditions of the inter-tropical belts while the latter are ecologi-cally less demanding taxa which are able to colonize both tropicaland temperate environments. The Kueichouphyllum fauna is typi-cally of the second category and represents the Lower Carbonifer-ous temperate rugose corals assemblage. The tropical corals arerepresented, in the investigated time slice by the colonial corals(Dorlodotia, Heterostrotion, Kwangsiphyllum, Siphonodendron, Sole-nodendron, Stylostrotion) and by the heterocaniniids, clisiophyllidsand axophyllids solitary corals diversified at the generic and spe-cific levels. The latter might have been more competitive in theinter-tropical belt and their absence in the temperate belt leftthe ecological niche available for Kueichouphyllum.
A similar latitudinal diversity gradient was documented in Mis-sissippian brachiopods by Qiao and Shen (2014) and in foramini-fers (Kalvoda, 2002). Nevertheless, no longitudinal distributionpattern was noticed by these authors.
Even if convenient and useful, the ‘Kueichouphyllum Zone’ isartificial because of a too large extension (in space and time) andby the lack of typical taxa other than the eponymous coral.Niikawa (1994) already suggested that this zone covers severalprovinces. In fact, it is easy to distinguish several different faunalassemblages in this ‘Zone’. (1) The Northern Palaeotethys includesfrom East to West the North China, Tarim, Karakum-Turan andrelated Blocks – forming the Chinese Province of Dubatolov andVassiljuk (1980) and Sando (1990). (2) The North-western Palaeot-ethys is bordered by the Hunic Terrane (Southern and Central Eur-ope) and is included in the Western European Province in which aMediterranean sub-province including South Europe and NorthAfrica is distinguished by Semenoff-Tian-Chansky (1985); it corre-sponds to the South Variscan Province sensu Korn et al. (2010). (3)The Eastern Palaeotethys is occupied by the South China, Indochinaand related blocks which are also included in the Chinese Province.Several sub-provinces are recognized within the Chinese blocks(Liao, 1990) but might simply be the result of a latitudinal gradientof diversity. (4) The southern Palaeotethys assemblage covers theCimmerian Terrane and the Asian margin of Gondwana as dis-cussed before is represented by a poorly diversified fauna of soli-tary rugose corals. This assemblage corresponds somewhat to theAsia Minor Province of Dubatolov and Vassiljuk (1980) andSando (1990) but these authors excluded from this province theTibet and Chinese Baoshan. Designating a Cimmerian Province isnot easy as its description is based on the absence of genera andnot by the occurrence of any endemic taxa (Smith and Xu, 1988),

J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 395
but its use is justified by its sub-tropical to temperate faunal char-acter and because it is a palaeobiogeographic and palaeogeograph-ic well constrained unit.
8. Conclusions
The Upper Tournaisian–Lower Viséan coral assemblage of theYaricak Yayla Formation in Central Taurides is relatively poorlydiversified, 12 genera and 15 species were identified, and one spe-cies is new. They define four assemblages easy to recognize in thefield. The Proheterelasma omaliusi assemblage and Uralinia multi-plex assemblage cover the Çit Yayla Mbr and represent the Ivorian.The Kueichouphyllum alborense assemblage crosses the Tournai-sian–Viséan boundary and covers almost entirely the Mantar TepeMember. The monospecific Eokoninckocarinia gemmina assemblagecovers the upper Molinician top of the Mantar Tepe. No post-Moli-niacian assemblage is known in the Hadım area. The Turkish Tau-rides were part, during Lower Carboniferous times, of theCimmerian Terrane during the Lower Carboniferous times. TheUpper Tournaisian–Lower Viséan coral associations of these blocksis composed of widely distributed genera representative of a sub-tropical to temperate shallow water shelf. A progressive decreaseof diversity from the East (Chinese and Tibetan blocks) to the West(Iran, Taurides, Armenia) is noticed. This longitudinal gradient ofpalaeobiodiversity along the Cimmerian Terrane, as well as thelack of any colonial genus is tentatively attributed to a cold currentrunning eastward along the Gondwana margin, maybe combinedwith the effects of a physical barrier created by a wide deltaic com-plex and/or by the competition with other rugose corals. Unfortu-nately, it must be said that all the investigated areas are as yet farfrom completely known and the results of this study deserve to bechallenged by new investigations in the Middle East and Himala-yan Lower Carboniferous successions.
Acknowledgements
D. Altıner is thanked for giving us the opportunity to study theLower Carboniferous in the Hadım vicinity. E. Poty (Liège) and M.Aretz (Toulouse) are acknowledged for their constructive remarkson taxonomy and stratigraphy. The manuscript was improved bythe review of I.D. Somerville (Dublin) and an anonymous reviewer.Field works and research were supported by a FRIA grant of theBelgian National Fund for Scientific Research (FNRS) and the Ser-vice of Paléontologie animale et humaine of the University of Liege(Belgium).
References
Altıner, D., Özgül, N., 2001. Carboniferous and Permian of the allochthonousterranes of the Central Taurides Belt, Southern Turkey. In: InternationalConference on Paleozoic Benthic Foraminifera, Guidebook, 25–29 August2001, Ankara, p. 36.
Altıner, D., Zaninetti, L., 1980. Les Archaediscidae (Foraminifères) dans leCarbonifère inférieur et moyen du Taurus oriental, Turquie. Riv. Ital.Paleontol. Stratigr. 86, 273–316.
Altıner, D., Ozkan-Altıner, S., Yılmaz, I.O., 2007. Tournaisian-Viséan boundary bedsin southern Turkey: foraminiferal biostratigraphy and sequence stratigraphy.In: Abstract of the 16th International Congress on the Carboniferous andPermian, p. 86.
Aretz, M., 2010. Habitats of colonial rugose corals: the Mississippian of westernEurope as example for a general classification. Lethaia 43, 558–572.
Aretz, M., 2011. Corals from the Carboniferous of the central Sahara (Algeria): thecollection ‘Marie Legrand-Blain’. Geodiversitas 33, 581–624.
Aretz, M., Dera, G., Lefebvre, V., Donnadieu, Y., Godderis, Y., Macouin, M., Nardin, E.,2013. The spatial and temporal distribution of Mississippian rugose corals:contribution of modelled oceanic currents and temperature data to thisproblem. In: Nardin, E., Aretz, M. (Eds.), Pre-Cenozoic Climates InternationalWorkshop, Toulouse, France. Strata 14, pp. 8–9.
Aretz, M., Nardin, E., Vachard, D., 2014. Diversity patterns andpalaeobiogeographical relationships of latest Devonian-Lower Carboniferous
foraminifers from South China: what is global, what is local? J. Palaeogeogr. 3,35–59.
Argyriadis, I., 1978. Le Permien Alpino-Méditérranéen à la charnière entrel’Hercynien et l’Alpin. Unpublished PhD Thesis, Université de Paris-Sud Orsay,p. 175.
Atakul-Özdemir, A., Altıner, D., Özkan-Altıner, S., Yılmaz, I.O., 2011. Foraminiferalbiostratigraphy and sequence stratigraphy across the mid-Carboniferousboundary in the Central Taurides, Turkey. Facies 57, 705–730.
Boland, K., 2002. Etude des Tétracoralliaires des couches de transition de la limiteTournaisien-Viséen en Europe occidentale. Unpublished PhD Thesis, Universitéde Liège, p. 390.
Brenckle, P.L., Gaetani, M., Angiolini, L., Bahrammanesh, M., 2009. Refinements inbiostratigraphy, chronostratigraphy and paleogeography of the Mississippian(Lower Carboniferous), Mobarak Formation, Alborz Mountains, Iran. GeoArabica14, 43–78.
Bykova, M.S., 1966. Carboniferous Corals of Western Kazakhstan. Akademye NaoukCCCP, Alma-Ata, p. 159.
Caputo, M.V., Melo, J.H.G., Streel, M., Isbell, J.L., 2008. Late Devonian and EarlyCarboniferous glacial records of South America. In: Fielding, C.R., Frank, T.D.,Isbell, J. (Eds.), Resolving the Late Paleozoic Ice Age in Time and Space. TheGeological Society of America, Special Paper 441, pp. 161–173.
Carruthers, R.G., 1908. A revision of some Carboniferous Corals. Geol. Mag. 5, 20–31,63–74, 158–171.
Carruthers, R.G., 1910. On the evolution of Zaphrentis delanouei in lowerCarboniferous times. Quart. J. Geol. Soc. 64, 523–538.
Chi, Y.S., 1935. Additional fossil corals from the Weiningian limestone of Hunan,Yunnan and the Kwangsi provinces, in SW China. Palaeontol. Sin. 12, 1–38.
Chu, S., 1933. The coral and brachiopod fossils from the Jinling Limestones. Bull.Inst. Geol. Acad. Sin. A2, 1–4.
Cotton, G., 1973. The Rugose Coral Genera. Elsevier, Amsterdam, p. 358.Crame, J.A., 2000. Evolution of taxonomic diversity gradients in the marine realm:
evidence from the composition of recent bivalve faunas. Paleobiology 26, 188–214.
De Koninck, L.-G., 1842. Description des animaux fossiles qui se trouvent dans leterrain carbonifère de la Belgique. Division I – Les Zoophytes. Ed. Dessein,Bruxelles, p. 32.
De Koninck, L.-G., 1872. Nouvelles recherches sur les animaux fossiles du terraincarbonifère de la Belgique. Mémoire l’Acad. Sci. Belgique 39, 1–78.
Demirtas�lı, E., Çatal, E., Dîl, N., Kiraglı, C., Salanci, A., 1978a. Carboniferous of thearea between Pınarbas�ı and Sarız field excursion on the Carboniferous of Turkeyguidebook. IUGS Subcommission on Carboniferous Stratigraphy, pp. 25–29.
Demirtas�lı, E., Çatal, E., Dîl, N., Kiraglı, C., Salanci, A., 1978b. Carboniferous of theSilifke area, Field Excursion on the Carboniferous of Turkey Guidebook. IUGSSubcommission on Carboniferous Stratigraphy, pp. 31–37.
Denayer, J., 2012. Corals of the upper Viséan microbial-sponge-bryozoan-coralbioherm and related strata of Kongul Yayla (Taurides, S Turkey). In: Denayer, J.,Aretz, M., Poty, E. (Eds.), 11th International Symposium on Fossil Cnidaria andPorifera, Liège, 2011 (Geologica Belgica 15, 317–328).
Denayer, J., Aretz, M., 2012. Discovery of a Mississippian reef in Turkey: the UpperViséan microbial-sponge-bryozoan-coral bioherm from Kongul Yayla (Taurides,S Turkey). Turk. J. Earth Sci. 21, 375–390.
Denayer, J., Hos�gör, I., 2014. Lower Carboniferous Rugose Corals from the HakkariArea (SE Turkey): a key to the understanding of the Arabian Platepalaeobiogeography in the Carboniferous. J. Asian Earth Sci. 79, 345–357.
Denayer, J., Poty, E., Aretz, M., 2011. Uppermost Devonian and Dinantian rugosecorals from Southern Belgium and surrounding areas. In: Aretz, M., Poty, E.(Eds.), 11th International Symposium on Fossil Cnidaria and Porifera, Liège,2011, Field Guide (Kölner Forum für Palaöntologie und Geologie 20, 151–201).
Dobroljubova, T.A., 1962. In: Sokolov, B.S. (Ed.), Base of Paleontology, Sponges,Archaeocyathids, Coelenterata, Worms. Osnovy Akademiye Nauk CCCP, pp.327–332.
Dobroljubova, T.A., Kabakovitsh, N.V., Sayutina, T.A., 1966. Korally nizhnegokarbona Kuznetskoj kotloviny. Trudy Academy of Sciences Ukraina USSR, p. 92.
Douglas, J.A., 1950. The Carboniferous and Permian faunas of South Iran and IranianBaluchistan. Memoirs Geol. Surv. India – Palaeontol Indica, New Ser. 22, 1–57.
Dronov, V.L., Kalimullin, S.M., Kochetkov, A.Y., Grectishnikova, I.A., Leven, E.A.,Pyzhyianov, I.V., Saltovskaya, V.D., 1979. The Carboniferous deposits of theMiddle and North-West Afghanistan. Comptes-rendus du Huitième CongrèsInternational de Stratigraphie et de Géologie du Carbonifère, Moscow, 1975, vol.2. Publishing Office Nauka, Moscow, pp. 167–174.
Dubatolov, V.N., Vassiljuk, N.P., 1980. Coral Paleozoogeography in the Devonian andCarboniferous of Eurasia. Acta Palaeontol. Pol. 25, 519–529.
Dumont, J.F., Lys, M., 1973. Description d’une série carbonifère située dans uncontexte autochtone; Göller Bölgesi Egridir. In: Congress of Earth Sciences onthe Occasion of the 50th Anniversary of the Turkish Republic, 1973, pp. 198–209.
Dun, W.S., Benson, W.N., 1920. Palaeontology. In: Benson, W.N., Dun, W., Dun, W.S.,Browne, R. (Eds.), Geology and Petrology of the Great Serpentine Belt of NewSouth Wales: The Geology, Palaeontology and Petrography of the CurrabubulaDistrict, with Notes on Adjacent Regions (Proceedings of the Linnean Society 45,337–374).
Dybowski, W.N., 1873. Monographie des Zoantharia Sclerodermata Rugosa aus derSilurformation Estlands, Nord-Livlands und der Insel Gotland. Arch. NaturkundLiv-, Ehst- Kurlands 5, 257–414.
Etheridge, R.J., 1900. Corals from the coral limestone of Lion Creek, Stanwell nearRockhampton. Queensland Geol. Surv. Bull. 12, 5–24.

396 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
Fan, Y., Yu, X., He, Y., Pan, Y., Li, X., Wang, F., Tang, D., Chen, S., Zhao, P., Liu, J., 2003.The Late Palaeozoic Rugose Corals of Xizang (Tibet) and Adjacent Regions andTheir Palaeobiogeography. Hunan Science and Technology Press, p. 679.
Fedorowsi, J., 2012. The new Upper Serpukhovian genus Zaphrufimia andhomeomorphism in some rugose corals. Palaeontogr. Abteilung 296, 109–161.
Fielding, C.R., Frank, T.D., Isbell, J., 2008. The Late Paleozoic ice Age – a review ofcurrent understanding and synthesis of global climate patters. In: Fielding, C.R.,Frank, T.D., Isbell, J. (Eds.), Resolving the Late Paleozoic Ice Age in Time andSpace. The Geological Society of America, Special Paper 441, pp. 343–353.
Flügel, H., 1963. Korallen aus der oberen Visé-Stufe (Kueichouphyllum-zone) Nord-Irans. Jahrbuch Geol. Bundesanstalt 106, 365–404.
Flügel, H.W., 1966. Palaeozoische korallen aus der Tibetischen zone von Dolpo(Nepal). Jahrbuch Geol. Bundesanstalt 12, 109–139.
Flügel, H., 1991. Rugosa aus dem Karbon der Ozbak-Kuh- Gruppe Ost-Iran. JahrbuchGeol. Bundesanstalt 134, 657–688.
Flügel, H., Kiratlioglu, E., 1956. Zur Paläontologie des anatolischen Paläozoikums. VI.Visékorallen aus dem Antitaurus. Neues Jahrbuch Geol. Paläontol. Monatshefte11, 512–520.
Fomitchev, V.D., 1931. New data on Lower Carboniferous corals of the KuznetskBasin. Trans. Geol. Prospect. Serv. USSR 49, 1–49.
Fomitchev, V.D., 1953. Corally rugosa i stratigraphia Sredene i Verkhenekameno iPermski otdojenii Donetsskogho basseyna. Moscow, p. 622.
Fontaine, H., 1989. Peculiarities of the Permian of peninsular Thailand. CCOP Newsl.14, 15–20.
Fontaine, H., Semenoff-Tian-Chansky, P., 1977. Aperçu sur les coraux du Carbonifèrede l’Hazarjat et des autres régions de l’Afghanistan. CR Soc. Géol. France 1977,235–237.
Fontaine, H., Suteethorn, V., Jongkanjanasoontorn, Y., 1991. Carboniferous corals ofThailand. Committee for co-ordination of joint prospecting for mineralresources in Asian offshore areas 22, 1–82.
Frech, F., 1916. Geologie Kleinasiens im Bereich der Bagdadbahn, Ergebnisse eigenerReisen, vergleichender Studien und paläontologischer Untersuchungen. Sonder-Abdruck aus Zeitschrift Deutschen Geol. Gesellschaft 68, 1–322.
Fröhlich, S., Petitpierre, L., Redfern, J., Grech, P., Bodin, S., Lang, S., 2010.Sedimentological and sequence stratigraphic analysis of Carboniferousdeposits in western Lybia: recording the sedimentary response of thenorthern Gondwana margin to climate sea-level changes. J. Afr. Earth Sci. 57,279–296.
Fromentel de, E., 1861. Introduction a l’étude des polypiers fossiles. Ed. Savy, Paris,p. 357.
Gabunia, K.E., 1919. Materialy k izucheniyu fauny korallov iznizhnekamennougolnykh otlozheniy okolo drevni Royki na reke Tomi.Sibirskogo Nauchno-Issledovatelskogo Instituta Geologii, Geofiziki iMineralnogo Syrya 1, 1–48.
Gaetani, M., Zanchi, A., Angiolini, L., Olivini, O., Sciunnach, D., Brunton, H., Nicora, A.,Mawson, R., 2004. The Carboniferous of the Western Karakoram (Pakistan). J.Asian Earth Sci. 23, 275–305.
Garzanti, E., Sciunnach, D., 1997. Early Carboniferous onset of Gondwanaglaciation and Neo-tethyan rifting in South Tibet. Earth Planet. Sci. Lett. 148,359–365.
Gervais, P., 1840. Astrées, Astrea, Dictionnaire des Sciences naturelles, supplément1, Paris, pp. 481–487.
Golonka, J., 2007. Phanerozoic paleoenvironment and paleolithofacies maps, LatePaleozoic. Geologia 33, 145–209.
Göncüoglu, M.C., Çapkinoglu, S�., Gürsu, S., Noble, P., Turhan, N., Tekin, U.K.,Okuyucu, C., Göncüoglu, Y., 2007. The Mississippian in the Central and EasternTaurides (Turkey): constraints on the tectonic setting of the Tauride-AnatolidePlatform. Geol. Carpath. 58, 427–442.
Gorsky, I., 1938. In: Nalivkin, D.V. (Ed.), Carboniferous Corals from Novaya Zemlya.Paleontology of the Soviet Arctic, Part II. Chief Administration of the NorthernSea route Arctic Institute of the USSR, Leningrad, p. 21.
Gorsky, I., Degtyarev, D.D., Katchanov, E.I., Rakchtschn, P., Stchakova, M.A., 1975.Tetracoralla (Rugosa). In: Stephanov, D.L., Krilova, A.K., Gorozpilova, L.P.,Pozner, V.M., Soultanaev, A.A. (Eds.), Paleontologitcheskii AtlasKamennougol’nykh Otlojenii Urala. Niedra, Leningrad, pp. 73–89.
Görür, N., Tüysüz, O., 2001. Cretaceous to Miocene palaeogeographic evolution ofTurkey: implications for hydrocarbon potential. J. Pet. Geol. 24, 119–146.
Grabau, A.W., 1922. Palaeozoic corals of China: part 1, Tetraseptata. Palaeontol. Sin.2, 1–76.
Grabau, A.W., 1928. Palaeozoic corals of China: part 1, Tetraseptata, secondcontribution. Acta Palaeontol. Sin. 2, 1–175.
Hance, L., Hou, H.-F., Vachard, D., 2011. Upper Famennian to Viséan Foraminifersand Some Microproblematica from South China. Geological Publishing House,Beijing, p. 359.
Heravi, M.A., Khaksar, K., 1999. An Assemblage of Corals of Iran. Geological Surveyof Iran, Mahshad, p. 357.
Heritsch, F., 1941. Unterkarbonische Korallen aus Anatolien. Zentralblatt Mineral.,Geol. Paläontol., Abteilung, Geol. Paläontol. 6, 188–190.
Hill, D., 1934. The Lower Carboniferous corals from Australia. Proc. R. Soc.Queensland 45, 63–115.
Hill, D., 1938. A Monograph on the Carboniferous Rugose Corals of Scotland.Palaeontographical Society, London, pp. 1–78 (1938), 79–144 (1939), 115–204(1940), 205–213 (1941).
Hill, D., 1973. Lower Carboniferous corals. In: Hallam, A. (Ed.), Atlas ofPalaeobiogeography. Elsevier Scientific Publishing Company, pp. 133–142.
Hill, D., 1981. Coelenterata; supplement 1, Rugosa and Tabulata. In: Teichert, C.(Ed.), Treatise on Invertebrate Paleontology. The Geological Society of Americaand the University of Kansas, Boulder, Colorado, Lawrence, Kansas, p. 762.
Howell, E.J., 1938. Rugose corals from the mid-Avonian limestone of Glamorgan.Ann. Mag. Nat. Hist. 1, 1–22.
Hudson, R.G.S., 1941. On the Carboniferous corals: Zaphrentis carruthersi sp. nov.from the Mirk Fell Beds and its relation to the Z. delanouei species-group. Proc.Yorkshire Geol. Soc. 24, 290–311.
Hudson, R.G.S., 1942. Fasciculophyllum Thomson and other genera of the ‘Zaphrentis’omaliusi group of Carboniferous corals. Geol. Mag. 79, 257–263.
Hudson, R.G.S., 1944. Lower Carboniferous corals of the genera Rotiphyllum andPermia. J. Paleontol. 18, 355–362.
Hudson, R.G.S., Fox, T., 1943. An Upper Viséan Zaphrentoid fauna from the YoredaleBeds of North-West Yorkshire. Yorkshire Geol. Soc. Bull. 25, 101–126.
Iljina, T.G., 1970. Nekotorye novye permskie rugozy yugovstochnogo Pamira. In:Astrova, G.C., Chudinova, I.I. (Eds.), Novye vidy paleozoyskikh mshanok ikorllov, Moscow, pp. 146–151.
Isaacson, P.E., Diaz-Martinez, E., Grader, G.W., Kalvoda, J., Babek, O., Devuyst, F.-X.,2008. Late Devonian–earliest Mississippian glaciation in Gondwanaland and itsbiogeographic consequences. Palaeogeogr. Palaeoclimatol. Palaeoecol. 268,126–142.
Ivanowski, A.B., 1967. Etyudy o rannekamennougolnykh rugozakh. Nauka, Moscow,p. 97.
Ivanowski, A.B., 1987. Rugosie opisannie A. A. Stukenberg’om (1888–1905). Akad.Naouk CCCP, 45.
Javaux, E., 1994. Palaeoecology of rugose corals in the Neffe Formation (MiddleViséan of Belgium. Courrier Forschunginstitut Senckenberg 172, 127–139.
Jia, H., Xu, S., Kuang, G., Zhang, B., Zhuo, Z., Wu, J., 1977. Class Anthozoa. In:Palaeontological Atlas of Central South China, Part I, Late Paleozoic. GeologicalPublishing House, Beijing, pp. 109–272.
Kalvoda, J., 2002. Late Devonian-Early Carboniferous Foraminiferal Fauna:Zonations, Evolutionary Trends, Paleobiogeography and Tectonic Implications.Masaryk University, Brno, Czech Republic, p. 213.
Kato, M., 1979. Some Upper Palaeozoic corals from Turkey. J. Facul. Sci., HokkaidoUniv. 19, 137–148.
Keyserling, A., 1846. Geognostihe Beobachtungen. Wissenschaftliche Beobachtungenauf einer Reise in das Petschora-land im Jahre 1843, Saint-Petersbourg,pp. 149–406.
Khaksar, K., 1996. New subfamily, genus and species of Lower Carboniferous Rugosecorals from central Alborz (N Iran). J. Sci. IR Iran 7, 97–112.
Khoa, N.D., 1996. The Upper Famennian and Tournaisian Rugosa and stratigraphy ofViet Nam. Geologos 1, 19–68.
Kora, M., 1989. Lower Carboniferous (Viséan) fauna from wadi Budra, west-centralSinai, Egypt. Neues Jahrbuch für Geol. Paläontol. Monatshefte 9, 523–538.
Kora, M., 1992. Carboniferous Macrofauna from Wadi Khaboba, West-Central Sinai(Egypt). Geol. Palaeontol. 26, 13–27.
Kora, M., 1995. Carboniferous macrofauna from Sinai, Egypt: biostratigraphy andpalaeogeography. J. Afr. Earth Sci. 20, 37–51.
Kora, M., Jux, U., 1986. On the early Carboniferous macrofauna from the Um BogmaFormation, Sinai. Neues Jb. Geol. Paläontol. Monat. 2, 85–98.
Korn, D., Jovanovic, D., Novak, M., Sudar, M.N., 2010. Early late Viséan ammonoidfaunas from the Jadar Block (NW Serbia). Geol. Carpath. 61, 355–364.
Lang, W.D., Smith, S., Thomas, H.D., 1940. Index of Palaeozoic Coral Genera. BritishMuseum of Natural History, London, p. 231.
Latif, M.A., 1970. Lower Carboniferous rocks near Nowshera, West Pakistan. Geol.Soc. Am. Bull. 81, 1585–1588.
Leighton, L.R., 2005. The latitudinal diversity gradient through deep time: testingthe ‘Age of the Tropics’ hypothesis using Carboniferous productidinebrachiopods. Evol. Ecol. 19, 563–581.
Lewis, H.P., 1929. On the avonian Coral Caninophyllum gen. nov. and Caninophyllumarchiaci Edw. et H. Ann. Mag. Nat. Hist. 3, 546–568.
Liao, W.-H., 1990. The biogeographic affinities of East Asian corals. In: McKerrow,W.S., Scotese, C.R. (Eds.), Palaeozoic Palaeogeography and Biogeography.Geological Society Memoir 81, pp. 175–179.
Ludwig, R., 1862. Paleontologie des Ural’s: Actinozoen und Bryozoen aus demCarbon-Kalkstein im Gouvernement Perm. Palaeontographica 10, 179–226.
Lys, M., 1986. Biostratigraphie du Carbonifère et du Permien en Mésogée (Espagne,Afrique du Nord, Régions égéennes, Proche-Orient). Etudemicropaléontologique (Foraminifères), paléobiogéographie. Unpublished PhdThesis, Université de Paris-Sud Orsay, France, p. 239.
Mackintosh, P.W., Robertson, A., 2012. Late Devonian–Late Triassic sedimentarydevelopment of the central Taurides, S. Turkey: implications for the northernmargin of Gondwana. Gondwana Res. 21, 1089–1114.
Mansy, J.-L., Conil, R., Meilliez, F., Khatir, A., Delcambre, B., Groessens, E., Lys, M.,Poty, E., Swennen, R., Trentesaux, A., Weyant, M., 1989. Nouvelles donnéesstratigraphiques et structurales sur le Dinantien de l’Avesnois. Ann. Soc. Géol.du Nord 108, 125–142.
Mc Coy, F., 1844. A Synopsis of the Characters of the Carboniferous LimestoneFossils of Ireland. University Press, Dublin, p. 207.
Menchikoff, N., Hsu, T.-H., 1935. Les polypiers carbonifères du Sahara occidental.Bull. Soc. Géol. France 5, 229–261.
Metcalfe, I., 2013. Tectonic evolution of the Malay Peninsula. J. Asian Earth Sci. 76,195–213.
Michelin, J.L.H., 1841–1848. Iconographie zoophytologique, description parlocalités et terrains des polypiers fossiles de Frances et pays environnants. Ed.

J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398 397
Bertrand, Paris, pp. 1–40 (1841), 42–72 (1842), 73–104 (1843), 105–144 (1844),145–184 (1845), 185–248 (1846), 249–328 (1847), 329–348 (1848).
Mii, H.-J., Grossman, F.L., Yancey, T.E., 1999. Carboniferous isotopes stratigraphies ofNorth America: implication for Carboniferous paleoceanography andMississippian glaciation. Geol. Soc. Am. Bull. 111, 960–973.
Milne-Edwards, H., Haime, J., 1850–1855. A Monograph of the British Fossil Corals.Palaeontographic Society Monograph, London, pp. 1–71 (1850), 147–210(1852), 211–244 (1853), 245–299 (1855).
Milne-Edwards, H., Haime, J., 1851. Monographie des Polypiérites fossiles desterrains paléozoïques, précédée d’un tableau général de la classification desPolypes, p. 502.
Minato, M., 1943. Note on some Lower Carboniferous fossils from Kirin Formationin Mincheng, Panshih-Hsien Province, Manchoukuo. J. Facul. Sci. HokkaidoImperial Univ. Ser. IV 5, 59–66.
Minato, M., 1955. Japanese Carboniferous and Permian corals. J. Facul. Sci.,Hokkaido Univ. Ser. IV: Geol. Mineral. 9, 1–202.
Mitchell, W.I., Green, G.W., 1965. Table 1: the faunal succession in theCarboniferous limestone of Burrington Combe. In: Green, G.W., Welch, F.B.A.(Eds.), Geology of the Country along Wells and Cheddar. Memoirs of theGeological Survey of Great Britain 280, pp. 177–197.
Moix, P., Beccaletto, L., Kozur, H.W., Hochard, C., Rosselet, F., Stampfli, G.M., 2008. Anew classification of the Turkish terranes and sutures and its implication for thepaleotectonic history of the region. Tectonophysics 451, 7–39.
Muir-Wood, H., 1948. Malayan Lower Carboniferous Fossils and Their Bearing andthe Viséan Palaeogeography of Asia. British Museum of Natural History, London,p. 118.
Nance, R.D., Guti Rrez-Alonso, G., Keppie, J.D., Linnemann, U., Murphy, J.B., Quesada,C., Strachan, R.A., Woodcock, N.H., 2010. Evolution of the Rheic Ocean.Gondwana Res. 17, 194–222.
Niikawa, I., 1994. The palaeobiogeography of Kueichouphyllum. CourierForschungsinstitut Senckenberg 172, 43–50.
Niikawa, I., 2012. The Upper Paleozoic coral fauna from Southern Turkey andadjacent area, and paleobiogeography of the rugose coral Kueichouphyllum. In:Yalçın, M.N., Çorbacıoglu, H., Aksu, Ö., Bozdogan, N. (Eds.), Paleozoic of NorthernGondwana and Its Petroleum Potential A Field Workshop. Turkish Association ofPetroleum Geologists Special Publication, Kayseri, Turquie, pp. 32–34.
Ogar, V., Falahatgar, M., Mosaddegh, H., 2011. Viséan corals of the Kiyasar area(North of Iran). In: Aretz, M., Delculée, S., Denayer, J., Poty, E. (Eds.), 11thInternational Symposium on Fossil Cnidaria and Porifera, Liège 2011, Abstract(Kölner Forum für Geologie und Paläontologie 19, 123–124).
Ogar, V., Falahatgar, M., Mosaddegh, H., 2013. Viséan corals of the Kiyasar area,South of Sari, Northern Iran. Rev. Bras. Paleontol. 16, 375–396.
Okan, Y., Hos�gör, I., 2005. Nohutluk Tepe (Aladag Bölgesi, Dogu Toroslar) istifindebulunan Bas�kirien (erken Geç Karbonifer) yas�lı Bivalvia türü Astartellaconcentrica (Conrad)’nın tanımı ve paleocografik dagılımı. J. Earth Sci. Appl.Res. Center Hacettepe Univ. 26, 13–23.
Okay, A.I., 2008. Geology of Turkey: a synopsis. Anschnitt 21, 19–42.Okuyucu, C., Vachard, D., 2006. Late Viséan foraminifers and algae from the
Cataloturan Nappe, Aladag Mountains, eastern Taurides, southern Turkey.Geobios 39, 535–554.
Onoprienko, Y., 1976. Early Carboniferous colonial rugose corals from the North-East of USSR. In: Gramm, M.N. (Ed.), Morphology and Systematics of Soviet Far-East Fossil Invertebrates. Institute of Biology and Pedology Far-East ScienceCentre, Academy of Sciences of the USSR, Vladivostok, pp. 5–34.
Onoprienko, Y., 1979. Some Early Carboniferous solitary Rugosa from the OmolonMassif. In: Gramm, M.N. (Ed.), Iskopcemye Bespozvonotchiye Dal’nego Vostoka,Vladivostok, pp. 1–32.
Özgül, N., 1984. Stratigraphy and tectonic evolution of the Central Taurides. In:Tekeli, O., Göncuöglü, M.C. (Eds.), Geology of the Taurus Belt. Mineral Researchand Exploration Institute of Turkey Publication, pp. 77–90.
Özgül, N., 1997. Bozkır-Hadim-Tas�kent (Orta Toroslar’in kuzey kesimi) dolayindayer alan tektono-stratigrafik birliklerin stratigrafisi. Bull. Mineral Res. Explor.Inst. Turkey (MTA Bull.) 119, 113–174.
Papojan, A.S., 1969. O nekotorykh rannekamennougol’nykh vidakh rodaKueichouphyllum v Armenii. Paleontol. Z. 1, 19–30.
Papojan, A.S., 1977. The coral complexes of Early Carboniferous in the SouthernTranscaucasus and its relation with some biogeographical provinces. In: SecondInternational Symposium on Corals and Fossil Corals Reefs. Mémoire du Bureaude Recherches Géologiques et Minières (BRGM), Paris 1975, pp. 197–202.
Pickett, J., 1966. Lower Carboniferous coral faunas from the New England District ofNew South Wales. Memoirs Geol. Surv. New South Wales, Palaeontol. 15, 1–38.
Poty, E., 1981. Recherches sur les Tétracoralliaires et les Hétérocoralliaires duViséen de la Belgique. Meded. Rijks Geol. Dienst 35, 1–161.
Poty, E., 1989. Distribution and palaeogeographic affinities of Belgian Tournaisianrugose corals. Memoirs Assoc. Aust. Palaeontol. 8, 267–273.
Poty, E., 2002. Interstitial tabellae: a new type of tabellae developed between highlythickened septa in Rugosa. Coral Res. Bull. 7, 167–173.
Poty, E., 2007. The Avins event: a remarkable worldwide spread of corals at the endof the Tournaisian (Lower Carboniferous). In: Hubmann, B., Piller, W.E. (Eds.),Fossil Corals and Sponges. Proceedings of the 9th International Symposium onFossil Cnidaria and Porifera, Schriftenreihe der ErdwissenschaftlichenKommissionen, vol. 2003. Oesterreichische Akademie der Wissenschaften,Graz, pp. 231–249.
Poty, E., 2010. Morphological limitation to the diversification of the rugose andtabulate corals. In: Kossovaya, O., Somerville, I.D. (Eds.), 10th International
Symposium on Fossil Cnidaria and Porifera, Saint-Petersburg, 2007(Palaeoworld 19, 389–400).
Poty, E., Boland, K., 1994. Révision des Tétracoralliaires caninomorphes del’Hastarien (Tournaisien) belge. Ann. Soc. Géol. Belgique 117, 201–225.
Poty, E., Hannay, D., 1994. Stratigraphy of rugose corals in the Dinantian of theBoulonnais (France). Mémoires l’Inst. Géol. l’Univ. Catholique Louvain 35, 51–82.
Poty, E., Onoprienko, Y., 1984. Rugose corals. In: Shilo, N.A., Bouckaert, J., Afanasieva,M.J.M., Bless, M.J.M., Conil, R., Erlanger, O.A., Gagiev, M.H., Lazarev, S.S.,Onoprienko, Y., Poty, E., Razina, T.P., Simakov, K.V., Smirnova, L.V., Streel, M.,Swennen, R. (Eds.), Sedimentological and Palaeontological Atlas of the LateFamennian and Tournaisian Deposits in the Omolon Region (NE USSR) (Annalesde la Société géologique de Belgique 107, 200–213).
Poty, E., Xu, S., 1996. Rugosa from the Devonian-Carboniferous transition in Hunan,China. Mémoires l’Inst. Géol. l’Univ. Louvain 36, 89–139.
Poty, E., Xu, S., 1997. Systematic position of some Strunian and Lower CarboniferousHeterocoral-like colonial corals. Bolletíno del Real Soc. Española Hist. Nat.(seccion Geología) 91, 99–106.
Poty, E., Berkowski, B., Chevalier, E., Hance, L., 2003. Biostratigraphic and sequencestratigraphic correlations between the Dinantian deposits of Belgium andSouthern Poland (Krakow area). In: Wong, T.E. (Ed.), XVth InternationalCongress on Carboniferous and Permian Stratigraphy. Royal NetherlandsAcademy of Arts and Sciences, Utrecht, The Netherlands, pp. 97–106.
Poty, E., Devuyst, F.-X., Hance, L., 2006. Upper Devonian and Mississippianforaminiferal and rugose coral zonations of Belgium and northern France: atool for Eurasian correlations. Geol. Mag. 143, 829–857.
Poty, E., Aretz, M., Xu, S., 2011. A comparison of Mississippian colonial rugose coralsfrom Western Europe and South China. In: Aretz, M., Delculée, S., Denayer, J.,Poty, E. (Eds.), 11th International Symposium on Fossil Cnidaria and Porifera,Liège, 2011, Abstracts (Kölner Forum für Geologie und Paläontologie 19, 135–136).
Qiao, L., Shen, S.-Z., 2014. Global paleobiogeography of brachiopods during theMississippian – response to the global tectonic reconfiguration, oceancirculation, and climate changes. Gondwana Res. 26, 1173–1185.
Robertson, A.H.F., Pickett, E.A., 2000. Palaeozoic–Early Tertiary Tethyian evolutionof mélanges, rift and passive margin units in Karaburun Peninsula (WesternTurkey) and Chios Island (Greece). In: Bozkurt, E., Piper, J.D.A., Winchester, J.A.(Eds.), Tectonics and Magmatism in Turkey and the Surrounding Area Tectonicsand Magmatism in Turkey and the Surrounding Areas. Geological Society,London, Special Publication 173, pp. 43–82.
Robertson, A., Ustaömer, T., 2009. Formation of the Late Palaeozoic Konya Complexand comparable units in southern Turkey by subduction – accretion processes:implications for the tectonic development of Tethys in the EasternMediterranean region. Tectonophysics 473, 113–148.
Robertson, A.H., Ustaömer, T., 2011. Role of tectonic-sedimentary melange andPermian-Triassic cover units, central southern Turkey in tethyan continentalmargin evolution. J. Asian Earth Sci. 40, 98–120.
Rodríguez, S., Falces, S., 1992. Corales Rugosos. In: Rodríguez, S., Arribas, M.E.,Comas-Rengifo, M., de la Peña, J., Falces, S., Kullmann, J., Gegundez, P., Legrand-Blain, M., Martinez-Chacon, M.L., Moreno-Eiris, Y.E., Perejon, A., Sanchez, J.L.,Sanchez-Chico, F., Sarmiento, G. (Eds.), Análisis paleontológico ysedimentológico de la cuenca carbonífera de Los Santos de Maimona(Bajadoz) (Coloquios de Paleontología 44, 49–90).
Rodríguez, S., Falces, S., 1994. Coral distribution patterns in the Los Santos deMaimona Lower Carboniferous Basin (Badajoz, SW Spain). CourrierForschunginstitut Seckenberg 172, 193–202.
Rogozov, Y.G., 1960a. Novye Turnejiskij rod Neomicroplasma (Rugosa).Paleontologicheski Z. 1960 (3), 48–51.
Rogozov, Y.G., 1960b. Pseudouralinia i Cystophrentis iz verjnego turne PripolyarnogoUrala. Paleontologicheski Z. 1960 (2), 40–43.
Salée, A., 1910. Contribution à l’étude des polypiers du Calcaire carbonifère de laBelgique, le genre Caninia. Nouveaux Mémoires Soc. belge Géol. 3, 1–62.
Salée, A., 1920. Sur un genre nouveau de Tétracoralliaire (Dorlodotia) et sur la valeurstratigraphique des Lithostrotion. Ann. Soc. Sci. Bruxelles Sér. B 39, 145–154.
Sando, W.J., 1990. Global Mississippian coral zonation. Courier ForschungsinstitutSenckenberg 130, 173–187.
Schindewolf, O.H., 1937. Zur stratigraphie und paläontologie der Wocklumerschichten (Oberdevon). Vertrieb bei der Preussischen Geol. landesanstalt 178,1–132.
Scrivenor, J.B., 1931. The Geology of Malaya. McMillian and Co., London, p. 217.Semenoff-Tian-Chansky, P., 1974. Recherches sur les Tétracoralliaires du
Carbonifère du Sahara Occidental. Mémoires du Mus. Natl. d’Hist. Nat. Sér. C:Sci. Terre 30, 1–316.
Semenoff-Tian-Chansky, P., 1985. Corals. In: Wagner, R.H., Winkler-Prins, C.F.,Granados, L.F. (Eds.), The Carboniferous of the World, II: Australia, IndianSubcontinent, South Africa, V. Instituto Geologico Y Minero de Espana Empresanacional adaro de investigaciones mineras, pp. 374–381.
S�engör, A.M.C., 1979. Mid-Mesozoic closure of Permo-Triassic Tethys and itsimplications. Nature 279, 590–593.
S�engör, A.M.C., Yılmaz, Y., 1981. Tethyan evolution of Turkey: a plate tectonicapproach. Tectonophysics 75, 181–241.
Shchukina, V., 1975. Rugose corals and biostratigraphy of northern Tian Shan, LowerCarboniferous. In: Sokolov, B.S. (Ed.), Ancient Cnidaria II, pp. 180–185.
Simakov, K.V., Bless, M.J.M., Conil, R., Gaviev, M.H., Kolesov, Y.V., Onoprienko, Y.I.,Poty, E., Razina, T.P., Shilo, N.A., Smirnova, L.V., Streel, M., Swennen, R., 1983.

398 J. Denayer / Journal of Asian Earth Sciences 98 (2015) 371–398
Upper Famennian and Tournaisian deposits of the Omolon region (NE-USSR).Ann. Soc. Géol. Belgique 106, 335–399.
Smith, A.B., Xu, J., 1988. Palaeontology of the 1985 Tibet Geotraverse, Lhasa toGolmud. Philos. Trans. R. Soc. Lond. Ser. A: Math. Phys. Sci. 327 (1594), 53–105.
Somerville, I.D., 1994. Early Carboniferous rugose coral assemblages from theDublin Basin, Ireland: possible bathymetric and palaeoecologic indicators.Courier Forschungsinstitut Senckenberg 172, 223–229.
Somerville, I.D., 2008. Biostratigraphic zonation and correlation of Mississippianrocks in Western Europe: some case studies in the late Viséan/Serpukhovian.Geol. J. 43, 209–240.
Somerville, I.D., Jones, G.L., 1985. The Courceyan stratigraphy of Pallaskenryborehole, County Limerick, Ireland. Geol. J. 20, 377–400.
Somerville, I.D., Strank, A.R.E., Welsh, A., 1989. Chadian Faunas and Flora fromDyserth: depositional environments and palaeogeographic settings of Viséanstrata in northeast Wales. Geol. J. 24, 49–66.
Somerville, I.D., Strogen, P., Jones, G.L., 1992. Biostratigraphy of Dinantian limestoneand associated volcanic rocks in the Limerick Syncline, Ireland. Geol. J. 27, 201–220.
Soshkina, E.D., 1960. Turneyskie korally Rugosa i ikh vzaimootnosheniya sdevonskimi. Akademiya Nauk SSSR Sbornik trudov po geologii i paleontologiiSyktyvkar, pp. 273–339.
Spassky, Y., Kachanov, E.I., 1971. Novye primitivnye rannekamennougolnye korallyAltay i Urala, vol. 59. Leningrad Gornyi Institut Zapiski, pp. 48–64.
Stampfli, G., Borel, G.D., 2002. A plate tectonic model for the Palaeozoic andMesozoic constrained by dynamic plate boundaries and restored syntheticocean isochrons. Earth Planet. Sci. Lett. 169, 17–33.
Stampfli, G.M., Hochard, C., Vérard, C., Wilhem, C., von Raumer, J., 2013. Theformation of Pangea. Tectonophysics 593, 1–19.
Stuckenberg, A., 1895. Korallen und Bryozoen der Steinkohlen ablagerungen desUral und des Timan. Mémoires Comité Géol. Saint-Petersbourg 10, 1–244.
Stuckenberg, A.A., 1904. Korally i mshanki nizhnego otdela srednerusskogokamennougolnogo izvestnyaka. Geol. Komiteta, Trudy 14, 1–362.
Tan, Z., Dong, Z., Jin, Y., Li, S., Yang, Y., Jiang, S., 1987. The Late Devonian and EarlyCarboniferous Strata and Palaeobiocoenosis of Hunan. Geological PublishingHouse, Beijing, p. 200.
Thomson, J., 1883. On the development and generic relation of the corals of theCarboniferous System of Scotland. Proc. R. Philos. S. Glasgow 14, 296–502.
Thomson, J., Nicholson, H.A., 1875–1876. Contributions to the Study of the chiefgeneric types of the Palaeozoic corals. Ann. Mag. Nat. Hist. Mus. Ser. IV 16, 305–309, 424–429 (1875); 17, 60–70, 123–158, 290–305, 451–461 (1876); 18, 68–73 (1876).
Tolmachev, I.P., 1931. Faune du Calcaire du Bassin du Kuznetsk. Comité Géol.Leningrad, Matér. pour Géol. Gén. Appl. 25, 1–663.
Torsvik, T.H., Cocks, L.R., 2013. Gondwana from top to base in space and time.Gondwana Res. 24, 999–1030.
Turan, A., 2001. Karaköy-(Gümdogmus�-Antalya)-Tas�kent-Hadim (Konya) arasınınmorfotektonik özllikleri. Cumhuriyet Üniv. Mühendislik Fakültesi Dergisi Ser. A18, 113–122.
Vassiljuk, N.P., 1959. Noviye Tetrakorally iz Nijnyego Karbona DonetskogoBassyeina. Paleontol. J. Akad. Naouk CCCP 4, 85–89.
Vassiljuk, N.P., 1960. Lower Carboniferous corals of the Donets Basin. Trudy Acad.Sci. Ukraina USSR, Stratigr. Palaeontol. 13, 179.
Vaughan, A., 1906. The Carboniferous Limestone series (Avonian) of the Avon Gorge.Proc. Bristol Nat. Soc. 4th Serie 1, 73–166.
Vaughan, A., 1917. Correlation of Dinantian and Avonian. Quart. J. Geol. Soc. 521, 1–52.
Verrill, A.E., 1865. Classification of Polyps. Ann. Mag. Nat. Hist. 16, 191–197.Volkova, M.S., 1941. Nizhnekamennougol’nye korally Tsentral’nogo Kazakhstana.
Mater Geol. Polez i iskop vost Kazakhsatana 11, 1–119.Von Schouppé, A., 1970. Lower Carboniferous corals from Badakhshan (North-East
Afghanistan). In: Deslo, A. (Ed.), Italian Expedition to the Karakorum and Hindu-Kush. Scientific Reports – IV Palaeontology, Zoology, Botany: Fossils of theNorth-East Afghanistan, vol. 2, pp. 1–23.
Vuillemin, C., 1990. Les Tétracoralliaires (Rugosa) du Carbonifère inférieur duMassif Armoricain (France). Editions du CNRS, p. 167.
Wang, H.C., 1950. A revision of Zoantharia Rugosa in the light of their minuteskeletal structures. Philos. Trans. R. Soc. Lond. 611, 175–246.
Wang, H., 1978. Subclass Rugosa. The Palaeontological Atlas of West South China,Guizhou, vol. 2. Geological Publishing House, Beijing, pp. 106–188.
Wang, Z., 1981. Rugose corals from the early Lower Carboniferous ChuashangouFormation in Amunike Mountain, Qinghai Province. Acta Geol. Sin. 3, 170–178.
Wang, X.-D., Ueno, K., Mizuno, Y., Sugiyama, T., 2001. Late Paleozoic faunal, climaticand geographic changes in the Baoshan blocks as a Gondwana-derived
continental fragment in southwest China. Palaeogeogr. Palaeoclimatol.Palaeoecol. 170, 197–218.
Wang, X.-D., Shen, S.Z., Sugiyama, T., West, R.R., 2003. Late Palaeozoic corals of Tibet(Xizang) and West Yunnan, Southwest China: successions andpalaeobiogeography. Palaeogeogr. Palaeoclimatol. Palaeoecol. 191, 385–397.
Webb, G.E., 1990. Lower Carboniferous coral fauna of the Rockhampton Group,East-Central Queensland. Memoirs Assoc. Aust. Palaeontol. 10, 1–167.
Webb, G.E., 2000. The Palaeobiogeography of Eastern Australian LowerCarboniferous Corals. Hist. Biol. 15, 91–119.
Weyer, D., 1970. The middle Tournaisian Rugose Coral Zaphrentites delepineiVaughan 1915. Bull. Soc. belge Géol. 79, 55–84.
Weyer, D., 1972. Zur Morphologie der Rugosa (Pterocorallia). Geol. Jahrbrug 21,710–737.
Weyer, D., 1993. Korallen aus dem Obertournai und Untervisé des Inseln Hiddenseeund Rügen. Abhandlungen Berichte Naturkunde 16, 31–69.
Weyer, D., 2000. Korallen im Unterkarbon Deutschlands. Abhandlungen BerichteNaturkunde 23, 57–91.
Wu, W., 1964. Lower Carboniferous Corals in Central Hunan, vol. 3. Memoirs ofInstitute of Geology and Palaeontology, pp. 1–100.
Wu, W.-S., Zhang, Y.S., 1980. Carboniferous rugose corals from eastern Xizang andwestern Sichuan. In: Regional Geological Survey of Sichuan Province andNanjing Institute of Geology and Palaeontology, Academia Sinica Stratigraphyand Palaeontology in Western Sichuan and Eastern Xizang, China, vol. 4, pp.103–162.
Wu, W., Zhao, J., 1989. Carboniferous and early Permian Rugosa from westernGuizhou and eastern Yunnan, SW China. Palaeontol. Sin. 177, 1–224.
Wu, W., Zhao, J., Jiang, S., 1981. Corals from the Shaodong Formation (Etroeungt) ofSouth China. Acta Palaeontol. Sin. 20, 1–14.
Xia, D.X., Liu, S.K., 1997. Stratigraphy (Lithostratigraphy) of Xizang AutonomousRegion, China. University Geological Press, Wuhan, p. 302.
Xu, S., 1981. On new Taxa of Rugose Corals from the Lower Carboniferous ofGuangdong Province, vol. 1. Bulletin of the Yichang Institute of Geology andMineral Research, pp. 42–48.
Xu, S., 1996. Strunian and Dinantian Rugose Corals from Hunan and Guangxi (SouthChina). Unpublished PhD Thesis, Université de Liège, Belgium, p. 267.
Xu, S., Poty, E., 1997. Rugose corals near the Tournaisian-Viséan boundary in SouthChina. Bolletinó Real Soc. Española Hist. Nat. (seccion Geología) 92, 349–363.
Yılmaz, I., 1981. Geology of the Antalya Complex, SW Turkey. University of Kansas,Austin, p. 193.
Yü, C.C., 1931. The correlation of the Fengninian System, the Lower Carboniferous asbased on Corals zones. Bull. Geol. Soc. China 10, 1–4.
Yü, C.C., 1933. Lower Carboniferous corals of China. Palaeontol. Sin. 12, 1–211.Yü, C.C., 1937. The Fenginian corals of South China. Memoirs Natl. Res. Inst. Geol. 16,
1–111.Yü, C.C., 1963. Discussion on relationship between Cystophrentis and Hexacoralla,
and establish Order Mesocorallia (ord. nov.) and Family Cystophrentidae (fam.nov.). Acta Palaeontol. Sin. 11, 307–316.
Yü, C.C., Lin, Y., Fan, Y., 1984. New material of Mesocorallia from the Lowercarboniferous of China, Neuvième Congrès international de Stratigraphie et deGéologie du Carbonifère. Southern Illinois University Press, p. 193.
Yurtsever, T.S., Gûrçay, H., Demirel, I.H., 2000. Aladag birligi nin Üst Devoniyen-altTriyas istifinin Çökelme ortamları ve hidrokarbon kaynak kayadegerlendirmesi: Orta Toroslar, Türkiye. Geol. Bull. Turkey 43, 33–57.
Zhang, J., 1986. New species of Lower Carboniferous Humboldtia found from Xigan,Guangxi. Acta Palaeontol. Sin. 25, 194–199.
Zhao, Z., Han, J., Wang, Z., 1984. The Carboniferous Strata and its Fauna from theSouthwestern Margin of Tarim Basin in Xinjiang. Geological Publishing House,Beijing, p. 187.
Zolinsky, L., 2000. Amygdalophyllum sudeticum sp. nov. (Rugosa) from a LowerViséan gneissic conglomerate, Bardzkie Mts., Sudetes (Poland). Acta Geol. Pol.50, 335–342.
Zuo, Z., 1986. Discovery of Kueichouphyllum from Yanguan stage of LowerCarboniferous. Acta Palaeontol. Sin. 25, 296–300.
Further Reading
Denayer, J., in press. Taxonomy, Biostratigraphy and Palaeobiogeography of the LateTournaisian rugose corals of North-Western Turkey. Paläont. Z.
Minato, M., Kato, M., 1977. Tethys sea corals in the Upper Palaeozoic. MémoiresBureau Recherches Géol. minières (BRGM) 89, 228–233.












