
RECOGNITION AND SELECTION OF SETTLEMENT SUBSTRATA DETERMINE POST-SETTLEMENT SURVIVAL IN CORALS
Upload
independentCategory
view
6download
0
Journal of Asian Earth Sciences 79 (2014) 345–357
Contents lists available at ScienceDirect
Journal of Asian Earth Sciences
journal homepage: www.elsevier .com/locate / jseaes
Lower Carboniferous rugose corals from the Arabian Plate: An insightfrom the Hakkari area (SE Turkey)
1367-9120/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.jseaes.2013.10.012
⇑ Corresponding author. Tel.: +32 43663258.E-mail addresses: [email protected] (J. Denayer), izzet.hosgor@viking-
intl.com (I. Hos�gör).
Julien Denayer a,⇑, Izzet Hos�gör b
a Service de Paléontologie animale et humaine, Département de Géologie, Université de Liège, Allée du Six-Août, B18, Sart Tilman, B4000 Liège, Belgiumb TransAtlantic Petroleum Ltd., Viking Int. Ltd., TR-06680, Ankara, Turkey
a r t i c l e i n f o a b s t r a c t
Article history:Received 25 February 2013Received in revised form 3 October 2013Accepted 4 October 2013Available online 18 October 2013
Keywords:Arabian PlateTurkeyHakkariRugose coralsTournaisianViséanLower CarboniferousCyathaxonia fauna
The Köprülü Formation of the Hakkari area (SE Turkey) is composed of a mixed carbonate–siliciclasticsuccession of Early Carboniferous age. The lower part of the formation yielded an abundant but poorlydiversified coral fauna composed of small non-dissepimented solitary rugose corals, namely Rotiphyllumcf. simulatum Fedorowski 2009, Zaphrentites parallela (Carruthers, 1910), cf. Gorizdronia, gen. et sp. indet.,Amplexizaphrentis sp. and Amplexizaphrentis zapense sp. nov. and specimens of Caninia cf. cornucopiaeMichelin in Gervais 1840, a dissepimented solitary coral showing a counter septum-related columella.The lower part of the Köprülü Formation is of supposed late Tournaisian age based on micropaleontolog-ical data. However, the coral assemblage indicates rather an early Viséan age. The Hakkari corals form astrongly facies-related association (‘‘Cyathaxonia fauna’’) and are compared to other areas with some dif-ficulties. The most similar, time-equivalent faunal associations is that of the Sinai Peninsula (NE Egypt).Both localities belonged, during Early Carboniferous times, to the Arabian Platform situated along thenorthern margin of Gondwana. The Gondwana-related Taurides units (Aladag), North Iran Block andAfghanistan, characterized by a dominant carbonate facies and more diversified coral faunas, formed dur-ing these times, the distal parts of the Arabian Platform.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Although Early Carboniferous (Mississippian) has been inten-sively studied worldwide, very few works concern the Lower Car-boniferous strata of the Arabian Plate. In fact, our stratigraphicalknowledge comes from subsurface survey and few scarce surfaceoutcrops of Early Carboniferous age. The scarce fauna of these stra-ta is only known from limited exposure in the Sinai Peninsula in NEEgypt. The stratigraphical and sedimentological settings of thisarea were intensively studied (Klebelsberg, 1911; Ball, 1916;Omara, 1967, 1971) but in the contrary, the Lower Carboniferousfauna are poorly known, except by the works of Omara (Omaraand Conil, 1965; Omara, 1967; Jux and Omara, 1983) and Kora(1992, 1995; Kora and Jux, 1986) dealing with rugose corals, bra-chiopods and foraminifers. Kora (1995) situated the Sinai coral fau-na in the Western Europe coral province of Sando (1990), while thenorthern part of the Arabian Plate is supposed to belong to the AsiaMinor coral province of Sando (1990) by extrapolation of what isknown from the neighboring Iran and Afghanistan zone (e.g. Flügel,1963; Von Schouppé, 1970) but few corals are known from thisarea. Moreover, the palaeogeographic conception of the
Palaeotethys has evolved since those papers. In recent models(e.g. Stampfli and Borel, 2002, 2004; Stampfli et al., 1991, 1998;Stampfli, 2000), the northern margin of Gondwana is edged by aribbon of terranes – the Cimmerian terranes – later (Late Permiantimes) rifted and drifted northward and assembled in the Alpineorogen after the Triassic. The remains of the platform edging theArabian Plate can be found in the Turkish Taurides, the North IranBlocks (e.g. Sanadaj-Sirjan terrane, Alborz terrane), Afghanistanand Pamir Blocks.
The Hakkari province (SE Turkey) provides some sectionsthrough the Paleozoic of the Arabian Plate and thus a good occa-sion to collect and analyze Lower Carboniferous corals in order toconstrain the palaeobiogeographic settings of this poorly knownarea of Middle East as a key location at the transition betweenthe eastern and western parts of the Palaeotethys Ocean.
2. Geological and stratigraphic settings
The Hakkari area belongs to the Southeast Anatolian Autochthonwhich represents the northern edge of the Arabian Plate and in-cludes more or less continuous Paleozoic successions (Fig. 1,Göncüoglu et al., 1997). The outcrops are still little known from apaleontological point of view because it has been difficult to obtainaccess to this region. The Devonian and Lower Carboniferous of the

Fig. 1. (A) Map of southeastern Turkey (Arabian Plate) with location of Palaeozoic non-metamorphic massifs. (B) Generalized stratigraphic sections of the Devonian toCarboniferous rocks in SE Turkey (modified from Gourvennec and Hos�gör (2012)).
346 J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357
Arabian Plate are represented in Southeastern Anatolia (SouthernTurkey) by sedimentary sequences observed from west to east inthe Amanos Mountains, Hazro High and Hakkari area (Fig. 1, Deanand Zhou, 1988; Yalçın and Yılmaz, 2010; Gourvennec and Hos�gör,2012). The Upper Devonian–Lower Carboniferous succession in theAmanos Mountains to the west and in the Hakkari area to the eastof the Hazro High are known as the Zap Group (Fig. 2), divided in-toYıgınlı (Middle–Late Devonian) and Köprülü formations (LateDevonian–Early Carboniferous; Perınçek et al., 1991; Gourvennecand Hos�gör, 2012). The Group is overlain by the Late Permian Hab-rol Formation (Köylüoglu and Altıner, 1989; Higgs et al., 2002).
The Zap Group crops out in the Zap river valley, between thetowns of Hakkari and Çukurca which are situated close to the Iranand Iraq borders. The group is dominated by brakish to marine,gray–black shale and shallow marine limestone. Parts of the Groupare absent in the Amanos and Hazro area, where they were notdeposited or, probably, removed by erosion during tectonic uplift(Cater and Tunbridge, 1992).
The thickness of the Yıgınlı Formation, in the Hakkari-Çukurcaarea, is between 200 and 300 m (Perınçek et al., 1991; Yalçın andYılmaz, 2010). Tunbridge (1988) in a sedimentological analysis,interpreted the Yıgınlı Formation as a clastic wedge and recognizedfive members within it. In ascending order the members include:(1) dolomite and red shales; (2) sandstones and gray shales; (3)red sandstones and siltstones; (4) transgressive sandstones andgray shales; and (5), dolomites and gray shales. Most fossils,including echinoderms, are well preserved in the shales and silt-stones of unit 4. The characteristic miospore taxa collected in thelast horizons include Vallatisporites hystricosus–Apiculiretusispora
verrucosa Miospore Biozone, indicating a late Famennian age(Higgs et al., 2002).
The Köprülü Formation, approximately 20 m-thick, conform-ably overlays the Yıgınlı Formation, with its base marked by thefirst limestone above the highest dolomite beds of the Yıgınlı For-mation. The Köprülü Formation represents a variety of marineenvironments ranging from agitated shallow marine to muddyshelf conditions – below fair weather wave base – and then a re-turn to more restricted shallow marine facies in the upper part(Janvier et al., 1984; Tunbridge, 1988). Hosgör et al. (2012) subdi-vided the Köprülü Formation in three informal members. The low-er member (5–7 m-thick) is made of limestone and sandylimestone, dark to grayish in color, representing the transgressivephase covering the continental deposits of the Yıgınlı Fomation.The corals described here were collected in this member. The mid-dle member (5–7 m-thick) is composed of calcareous shale andsiltstone, usually laminated and dark-colored. Several sandstonelayers are intercalated within this mudstone unit. The upper mem-ber is dominated by massive grayish limestone, often sandy ordolomitized, with some dark shale layers. This last member is 1–8 m-thick (Hosgör et al., 2012).
Janvier et al. (1984) indicated a Late Devonian age to the base ofthe Köprülü Formation on the basis of fish remains from two hori-zons (Fig. 2). A Late Devonian–Early Carboniferous age was assignedto the whole Körpülü Formation based on foraminifers, pal-ynomorphs and brachiopods by Perinçek (1990; Perınçek et al.,1991). The characteristic miospore taxa in the middle of the KörpülüFormation include the Spelaeotriletes pretiosus–Raistrickia clavataMiospore Biozone (Higgs et al., 2002), and the bivalve Posidonia

Fig. 2. (A) Schematic Cambrian to Carboniferous lithostratigraphic succession in the Zap Valley with a closer view of the Devonian–Lower Carboniferous S�ort Dere section(modified from Gourvennec and Hos�gör (2012) and Hos�gör et al. (2012)). (B) Geological map of the Köprülü zone (Hakkari area, SE Turkey).
J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357 347

348 J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357
becheri indicates a Tournaisian age (Hos�gör et al., 2012; Hos�gör andOkan, 2012).
3. Material, methods and systematic paleontology
The S�ort Dere section (Fig. 1, GPS 37�17’05.08"N 43�32’25.78"E)is situated on the southern limb of the Çukurca Anticline, croppingout along the northern flank of the valley and exposes the Yıgınlıand Köprülü formations. About 50 samples were collected nearthe base of the Köprülü Formation (approximate elevation1330 m). The fossil material comes from two bioclastic silty lime-stone levels where solitary rugose corals are numerous, togetherwith brachiopods, sponges spicules and bryozoans.
This study is based on c. 150 thin sections (transverse and lon-gitudinal) made through 51 mainly completed, rock-free small sol-itary corals. The terminology of skeletal elements is based on Hill(1981) and is summarized in Fig. 3. All the specimens, labeled withprefix HK.1 and HK.2, are housed in the collection of Paleontologieanimale et humaine of the University of Liège (Belgium). The clas-sification follows Hill (1981) with some modifications for Rotiphyl-lum and Gorizdronia (cf. Fedorowski, 2009).
Suborder Streptelasmatina Wedekind 1927Family Hapsiphyllidae Grabau 1928Genus Amplexizaphrentis Vaughan 1906Diagnose: See Hill (1981).Discussion: The septal pattern, the keyhole-shaped cardinal fos-
sula on the concave side of the corallite and well developed alarfossulae are diagnostic characters of the genus Amplexizaphrentis.The most similar genera, Zaphrentites Hudson 1941, is usuallysmaller, has no minor septa and its major septa never withdrawfrom the axis, Sychnoelasma Lang, Smith & Thomas 1940 is larger,its skeletal elements are usually thickened and has no alar fossulae.Proheterelasma Cotton 1973 is smaller and has a simpler morphol-ogy (thinner axial column, shorter cardinal fossula and less devel-oped alar fossulae). Further comparisons of Amplexizaphrentis withother zaphrentoid corals (Zaphrentites, Zaphrentoides, Zaphrufimia)are available in the revision of Fedorowski (2012).
Distribution: The genus Amplexizaphrentis is present from theKinderhookian (early Tournaisian) to the Chesterian (Serpukho-vian) in North America (Sando and Bamber, 1985) but only knownin the Viséan and Namurian in the British Isles (Vaughan, 1906;Vaughan, 1908; Smith, 1910; Garwood, 1913), in the Viséan of Iran(Heravi and Khaksar, 1999), Australia (Webb, 1990) and China (Wuand Zhao, 1989) and in the late Viséan of Morocco (Aretz, 2011).
Fig. 3. Overview of some descriptive morphological terms for the study of solitary rugos(C) cylindroid coral, and (D) external view of constriction (co) and rejuvenescence (rj) insections. Legend: sc: stereocolomun, dt: domed tabulae, ft: flat tabulae, ew: external waseptum, as: alas septum, af: alar fossula, cf: cardinal fossula. After Hill (1981).
The Amplexizaphrentis assemblage from the Sinai Peninsula (Kora,1989, 1992; Kora and Jux, 1986) in badly constraint biostratigraph-icaly (late Tournaisian to middle Viséan, see Brenckle and Mar-chant (1987) for discussion).
Amplexizaphrentis zapense sp. nov.Fig. 4: 1a–d, 2a–b, 3–5, 13, 16.Etymology: From the Zap River valley where the material was
collected.Holotype: Specimen HK.1.29 (5 transverse thin sections).Type locality: S�ort Dere section, Zap River valley, Hakkari area,
SE Turkey.Type horizon: Level HK.1, crinoidal silty limestone forming the
lower part of the Köprülü Formation.Diagnosis: Small Amplexizaphrentis with a mean diameter of
13 mm and 23–28 major septa. Minor septa developed only in ma-ture stages. Cardinal fossula long and closed, keyhole-shaped. Alarfossulae well marked, also keyhole-shaped. Pericounter septa con-vergent and forming, with the counter septum, a fork with theshape of the Greek ‘‘psi’’ letter.
Material: Seventeen specimens, including the holotype, fromthe type locality (46 transverse and 3 longitudinal thin sections).
Description: Small ceratoid to trochoid Amplexizaphrentites,16.6 mm high in average (maximum 22 mm), with a mean diame-ter of 13 mm (maximum 19 mm) and counting 23–28 major septa(Fig. 4:13, 16, Fig. 5). The external wall shows septal furrows andgrowth ribs. The cardinal fossula is on the concave side of the coral.In juvenile stages (diameter smaller than 4 mm), the septa are verythick, particularly the cardinal and pericardinal septa and fuse inthe center of the corallite, forming a thick axial stereocolumn(Fig. 4: 1a and b). The latter reach 1/2 of the corallite diameter.Where the corallite is around 4 mm in diameter, the column beginsto become thin. The zaphrentoid-type pattern of the septa appearsat the same time, followed by the appearance of the cardinal andalar fossulae (Fig. 4: 1b, 2a). In mature stages (around 10 mm indiameter), the septa form two groups in the cardinal quadrants,which join the edges of the cardinal fossula (Fig. 4: 1d, 5). In thecounter quadrants, two other groups of septa fuse with the axialend of the cardinal fossula (Fig. 4: 1c and d). The septal insertionis accelerated in the counter quadrants counting, in some speci-mens, 2–4 more septa than the cardinal quadrants (see septal for-mulae, Fig. 5). The cardinal and counter groups of septa areseparated from each other by two long alar fossulae (Fig. 4: 1c,5). The latter are larger in their axial part than in their peripheralpart, giving a typical keyhole shape. These fossulae are bordered,on the cardinal side, by the alar septa. At the counter side, the septa
e corals. (A–D): Outer shape: (A) external view of a ceratoid coral, (B) trochoid coral,a solitary coral. (E and F): internal characters in longitudinal (E) and transverse (F)
ll, ks: counter septum, pks: pericounter septum, cs: cardinal septa, pcs: pericardinal
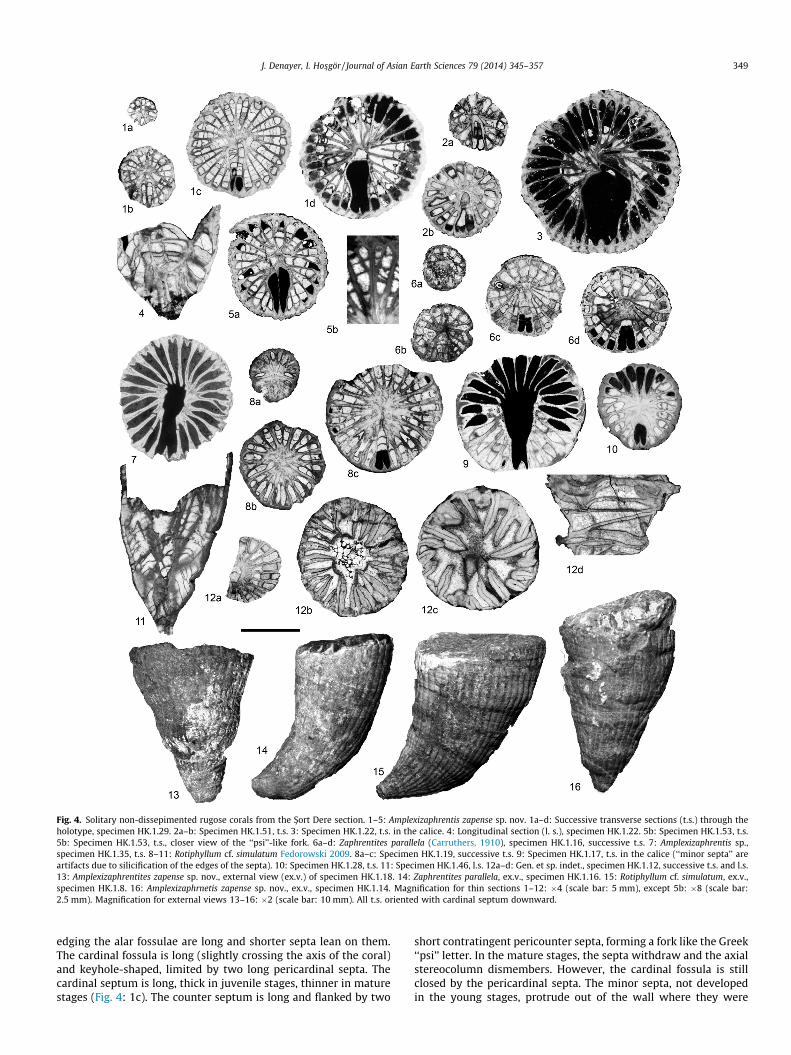
Fig. 4. Solitary non-dissepimented rugose corals from the S�ort Dere section. 1–5: Amplexizaphrentis zapense sp. nov. 1a–d: Successive transverse sections (t.s.) through theholotype, specimen HK.1.29. 2a–b: Specimen HK.1.51, t.s. 3: Specimen HK.1.22, t.s. in the calice. 4: Longitudinal section (l. s.), specimen HK.1.22. 5b: Specimen HK.1.53, t.s.5b: Specimen HK.1.53, t.s., closer view of the ‘‘psi’’-like fork. 6a–d: Zaphrentites parallela (Carruthers, 1910), specimen HK.1.16, successive t.s. 7: Amplexizaphrentis sp.,specimen HK.1.35, t.s. 8–11: Rotiphyllum cf. simulatum Fedorowski 2009. 8a–c: Specimen HK.1.19, successive t.s. 9: Specimen HK.1.17, t.s. in the calice (‘‘minor septa’’ areartifacts due to silicification of the edges of the septa). 10: Specimen HK.1.28, t.s. 11: Specimen HK.1.46, l.s. 12a–d: Gen. et sp. indet., specimen HK.1.12, successive t.s. and l.s.13: Amplexizaphrentites zapense sp. nov., external view (ex.v.) of specimen HK.1.18. 14: Zaphrentites parallela, ex.v., specimen HK.1.16. 15: Rotiphyllum cf. simulatum, ex.v.,specimen HK.1.8. 16: Amplexizaphrnetis zapense sp. nov., ex.v., specimen HK.1.14. Magnification for thin sections 1–12: �4 (scale bar: 5 mm), except 5b: �8 (scale bar:2.5 mm). Magnification for external views 13–16: �2 (scale bar: 10 mm). All t.s. oriented with cardinal septum downward.
J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357 349
edging the alar fossulae are long and shorter septa lean on them.The cardinal fossula is long (slightly crossing the axis of the coral)and keyhole-shaped, limited by two long pericardinal septa. Thecardinal septum is long, thick in juvenile stages, thinner in maturestages (Fig. 4: 1c). The counter septum is long and flanked by two
short contratingent pericounter septa, forming a fork like the Greek‘‘psi’’ letter. In the mature stages, the septa withdraw and the axialstereocolumn dismembers. However, the cardinal fossula is stillclosed by the pericardinal septa. The minor septa, not developedin the young stages, protrude out of the wall where they were

Fig. 5. (A) Statistical data for Amplexizaphrentis zapense sp. nov, Zaphrentitesparallela and Rotiphyllum cf. simulatum. (B) Septal formulae for specimens ofAmplexizaphrentis zapense sp. nov.
350 J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357
confined and grow up to 1 mm long, mainly in the counter quad-rants (Fig. 4: 3). The wall is thick, strongly arched by ruguae corre-sponding to the major and minor septa (Fig. 4: 16). There are nodissepiments. In longitudinal section, the tabulae are incomplete,domed in the central part of the corallite and downturn towardthe periphery (Fig. 4: 4). They strongly dip toward the peripheryin the cardinal fossula.
Discussion: Amplexizaphrentis zapense presents dimensions andnumber of septa similar to those of A. browni Webb 1987 but con-trarily to this species, it does not show any thickening in the ma-ture stage. A. enneskileni Milne-Edwards & Haime 1851 (23 mmin diameter for 40 major septa) and A. maneri Webb and Suther-land 1993 (20 mm and 32–40 septa) are both larger than A. za-pense. A. elliptica (White, 1862) has a similar diameter butpossesses more septa and well developed minor septa in bothquadrants. A. cassa Sutherland 1958 is larger and its septa with-draw early in development. A. palmatus Easton 1944 has a verydeep alar fossulae. Moreover, all the available specimens presentthe typical ‘‘psi’’-like fork formed by the counter major septa andits shortened pericounter septa. This feature, with the smalldimensions and number of septa is sufficient to create a new spe-cies for the Hakkari fauna.
Amplexizaphrentis sp.Fig. 4: 7.Material: One specimen (4 transverse thin sections).Description: Small solitary coral, trochoid in shape, 21 mm high,
11 mm in diameter and 21 major septa. The juvenile stages showstrongly thickened elements and the septa are pinnately arrangedin four groups. The septa join in the center of the corallite, forminga wide and thick axial stereocolumn. The cardinal fossula is largeand occupied by a thick cardinal septum. The alar fossula are nar-row and not keyhole-shaped (Fig. 4: 7). The mature stages showwithdrawn thickened major septa and minor septa (up to 1 mmlong) developed in both cardinal and counter quadrants. The cardi-nal fossula is closed even in the late stages. The external wall isstrongly arched in the young stages, straighter but still thick inthe late stages.
Discussion: This single specimen is doubtlessly an Amplexiza-phrentis but clearly another species than A. zapense because themain features of the new species are not obvious in this coral. Itis quiet similar to A. palmatus Easton 1944 but the latter is larger
and has more septa. This specimen differs from the other speciesof Amplexizaphrentis by long septa (A. cassa Sutherland 1958 showsshort septa), a smaller diameter (A. enniskileni Milne-Edwards &Haime 1851 is larger) and the lack of minor septa in juvenile stages(A. elliptica (White, 1862) shows developed minor septa).
Genus Zaphrentites Hudson 1941Diagnosis: See Hill (1981).Discussion: See Carruthers (1910), Hill (1938–41) and Fedorow-
ski (2012).Distribution: Zaphrentites is a common genus through the Tour-
naisian and Viséan of Western Europe (Poty, 1981), Eastern Europe(Chwieduk, 2005), South China (Xu and Poty, 1997), the RussianPlatform (Ivanowski, 1967), the Kuznetsk Basin (Tolmachev,1931; Dobroljubova et al., 1966), Japan (Minato, 1955) and SinaiPeninsula (Kora and Jux, 1986). In Turkey, Zaphrentites is presentin the Yaricak Formation of the Aladag Unit (unpublished data ofthe first author) in the Hadım vicinity, in the Yer Köprü and Karap-unar areas in the eastern Taurides (Frech, 1916), and in the KoprülüFormation in the Hakkari area.
Zaphrentites parallela (Carruthers, 1910)Fig. 4: 6a–d, 14.
*
1910 Zaphrentis parallela Carruthers, p. 533, pl. 37,Fig. 4?
1916 Zaphrentis cornu copiae Michelin; Frech, p. 241,pl. 4, Fig. 2a–f?
1916 Zaphrentis vermicularis de Koninck; Frech, p. 241,pl. 4, Fig. 1a–c?
1916 Cyathophyllum excavum M.-E. & H.; Frech, p. 242,pl. 4, Fig. 3a–c1941
Zaphrentis parallela Carruthers; Hudson, p. 292 non 1964 Zaphrentis parallela Carruthers; Wu, p. 27, pl. 1,Figs. 19 and 20
1966 Zaphrentites parallelus (Carruthers);Dobroljubova et al., p. 26, pl. 1, Figs. 4–7
non 1967 Zaphrentites parallela (Carruthers); Ivanowski, p.41, pl. 2, Fig. 2
? 1980 Zaphrentites parallelus (Carruthers); Wu & Zhang,p. 152, pl. 1, Fig. 7
1985 Zaphrentites parallelus (Carruthers); Kullmann &Liao, p. 138, pl. 1, Figs. 4 and 5
1992 Zaphrentites parallela (Carruthers); Rodríguez &Falces, p. 172, pl. 14, Fig. 3
1994 Zaphrentites parallela (Carruthers); Rodríguez &Falces, p. 201, Fig. 5.6
1999 Zaphrentites parallela (Carruthers); Heravi &Khaksar, pl. 3, Fig. 11
non 1999 Zaphrentites parallela (Carruthers); Heravi &Khaksar, pl. 3, Fig. 10
? 2003 Zaphrentites parallelus (Carruthers); Fan et al., p.281, pl. 15, Fig. 7a–b
2005 Zaphrentites parallela (Carruthers); Chwieduk, p.427, pl. 15, Fig. 6
Diagnosis: Zaphrentites with septa pinnately arranged with ax-ial ends of the septa joined to form an axial stereocolumn slightlystaggered toward the counter part of the corallite. Cardinal septumshorter, in a cardinal fossula with edges typically parallel. Alar fos-sula sometimes developed (after Hudson (1941)).
Material: Six specimens (14 transverse thin sections) from S�ortDere section (Hakkari area).
Description: Small solitary rugose coral, ceratoid or rarely tro-choid, up to 16 mm high with a mean calicular diameter of9.1 mm (maximum 11 mm, Fig. 4: 15). In transverse section, thebilateral symmetry is perfectly marked by the long cardinal

Fig. 6. Solitary dissepimented and non-dissepimented rugose corals from the S�ort Dere section. 1–4: Caninia aff. cornucopiae Michelin in Gervais 1840. 1a–d: Successive t.s.,specimen HK.1.10. 2a–d: Specimen HK.1.1, successive t.s., showing a counter septa-derived columella and some transeptal dissepiments. 3: Specimen HK.1.45. l.s. 4:Specimen HK.1.45, enlarged view of the dissepimentarium. 5–10: cf. Gorizdronia sp. 5a–c: Specimen HK.1.15, successive t.s. and l.s. showing mesa-shaped tabulae. 6:Specimen HK.1.4, t.s. 7: Specimen HK.1.21, t.s. 8: Specimen HK.1.4, ex.v. fragment of cylindroid coral showing numerous constriction-rejuvenescence cycles. 9: SpecimenHK.1.9, ex. v. with a trochoid habitus. 10: Specimen HK.1.21, ex. v., with a conical habitus showing some constriction phases. 11–12: Caninia aff. cornucopiae. 11: SpecimenHK.1.13, ex.v., with a trochoid habitus. 12a: Specimen HK.1.1, ex. v. of the calice, showing a protuberant counter septum-derived columella. 12b, specimen HK.1.1, ex.v.,largest specimen, showing a trochoid habitus. Magnification for thin sections 1–7: �4 (scale bar: 5 mm), excepted 4 (magnification: �8, scale bar: 2.5 mm). Magnification forexternal views 8–12: �2 (scale bar: 10 mm). All t.s. oriented with cardinal septum downward.
J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357 351
fossula. In thin sections, the maximum number of septa is 21 for adiameter of 8.5 mm (Fig. 5). The pinnate pattern of septa is welldeveloped. The septa are long, slightly undulating in the cardinalquadrants, joined the axial end of the fossula forming an axial ste-reocolumn which is withdrawn toward the counter quadrant of the
corallite (Fig. 4: 6). The fossula is long, closed and limited by twolong and parallel pericardinal septa (Fig. 4: 6c and d). The cardinalseptum is shorter and thinner than the other major septa. Thecounter septum has the same length and thickness than the othermajor septa. In some specimens, the alar septa are shorter and

Fig. 7. Early Carboniferous (c. 340 Ma) reconstruction of the Palaeotethys Ocean and its margins (modified form Stampfli et al. (2002)) with position of the discussed areasalong the northern margin of the Gondwana. Legend: ADH: Apulia–Dinarids–Hellenids group of terranes, Afr: Africa, Ant: Antartica, Ara: Arabian Plate, Ava: Avalonia, Bal:Baltica, EHT: group of European Hunic Terranes (including Armorica, Aquitania, Ossa-Morena, Cantabria, Adria, Anatolia, etc., see Stampfli et al., 2002 for details). IC:Indochina, Ind: India, Kaz: Kazakhstan, KT: Karakum-Turan, Nam: North America, Nch: North China Block, NT: North Tibet, Sam: South America, Sch: South China Block, Sib:Siberia, Tm: Tarim.
352 J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357
contratingent with the adjacent septa. Minor septa are not devel-oped, but visible in the stereoplasm of the wall. There are no dis-sepiments. The wall is undulating or festooned in juvenile stagesbut usually becomes straight in mature stages. Tabulae were notobserved in longitudinal section.
Discussion: These specimens clearly belong to the genusZaphrentites. The septal arrangement and axial stereocolumn aredifferent to the morphologically similar genera as Amplexizaphren-tites and Bradyphyllum Grabau 1928. The present specimens can beclassified within the species Z. parallela (Carruthers 1910) whichhas the same dimensions and shows typically a long parallel-edgedfossula with a cardinal septum withdrawn in late stages. The twoother common species of Zaphrentites, Z. delanouei (Milne-Edwards& Haime, 1851) and Z. crassus Hudson 1944 are both larger andcount more septa.
Familly Antiphyllidae Iljina 1970Genus Rotiphyllum Hudson 1942Diagnose and discussion: See Fedorowski (2009).Rotiphyllum cf. simulatum Fedorowski 2009Fig. 4: 8a–c, 9–11, 15.Material: Eleven specimens (24 transverse and 3 longitudinal
thin sections) from S�ort Dere section (Hakkari area).Description: Small solitary rugose coral, trochoid, 10–18 mm
high with a mean calicular diameter of 10.8 mm (maximum12.5 mm, Fig. 4: 14, Fig. 5). The external wall is covered with strongseptal furrows. The mean number of septa is 22 (maximum 29).The major septa are straight and thick (maximum 0.8 mm thick)
and always rhopaloid where not joined together. Their axial endsusually fuse in a large axial stereocolumn reaching 4 mm in diam-eter (Fig. 4: 8–10). The stereocolumn is circular in transverse sec-tion but incised by the cardinal fossula. The axial stereocolumn isdismembered in the very late stages but the major septa are stilljoined by their axial ends in groups of 4 to 5 (Fig. 4: 9). The cardinalseptum is usually thinner but long, slightly withdrawn toward theperiphery in the mature stages. The counter septum is slightlythinner than the others. The base of the septa are cuneiform. Minorsepta are not developed, even in the calicular part or the corallite(Fig. 4: 9). The cardinal fossula is long and elongated, borderedby the pericardinal major septa and the joined axial ends of adja-cent septa (Fig. 4: 9–10). There are no dissepiments. The wall isthick (up to 0.5 mm), festooned or arched in juvenile stages,straight afterward. In longitudinal section, the tabulae are discon-tinuous, upturned toward the axis. In the late stages of develop-ment, where the axial stereocolumn is dismembered, the tabulaebecome flat or slightly domed (Fig. 4: 11).
Discussion: Rotiphyllum Hudson 1942 differs from the othernon-dissepimented coral genera with an axial structure by the latedismemberment of the axial stereocolumn. Contrarily, Bradyphyl-lum Grabau 1928 loses its axial structure rapidly during its devel-opment. Rotiphyllum differs from Monophyllum Fomichev 1953 byhaving a counter septum less protuberant and a cardinal fossulaless triangular-shaped (Fedorowski, 2009). With its strong axialstereocolumn, the present species is similar in dimensions to R.cuneatum Wu 1964 from the Hunan Province of China. However

J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357 353
the Chinese species shows thickened cardinal and peri-cardinalsepta. R. simulatum Fedorowski 2009 from the Donets Basin (Fedo-rowski, 2009) has the same dimensions and number of septa and awide axial stereocolumn but is Bashkirian while the Turkish spec-imens are late Tournaisian or early Viséan, thus it is preferable todesignate the present specimen as Rotiphyllum cf. simulatum. Theoldest representative of Rotiphyllum, R. sp. described from lower-most Tournaisian of Polish Pomerania by Chwieduk (2005) needsto be checked because the first occurrence of the genus elsewherein Europe and Asia is not older than late Tournaisian (rugose coralzone RC3 of Poty et al. (2006)). Hudson (1944) considers than Rot-iphyllum evolved from Proheterelasma Cotton 1973 by simplifica-tion of the septal pattern and the tightening of the axialstructure. Poty (personal communication) considers this lineageas correct since the oldest Rotiphyllum from Belgium (lower partof the Ivorian substage) show the typical septal pattern of Prohet-erelasma in its juvenile stages. Weyer (1993) considers Proheterel-asma as a junior synonym of Rotiphyllum after their juvenile stagesbut the two genera are clearly distinct in mature stages.
Familly Cyathopsidae Dybowski 1873Genus Caninia Michelin in Gervais 1840Diagnose: See Hill (1981).Discussion: See Semenoff-Tian-Chansky (1974).Distribution: Caninia is a frequent genus in the Tournaisian and
Viséan strata of Eurasia (Denayer et al., 2011; Mitchell, 1989;Vuillemin, 1990), Iran (Heravi and Khaksar, 1999), Siberia (Ivanow-ski, 1967), Khazakstan (Volkova, 1941), Kuznetsk (Dobroljubovaet al., 1966) and North Africa (Semenoff-Tian-Chansky, 1974). InTurkey, Caninia is present in the Taurides (Tufanbeyli area, Kato,1979), from the Aladag Unit near Hadım (unpublished data of thefirst author) and in the Zonguldak area (unpublished data). It oc-curs in the lower part of the Köprülü Formation in the S�ort Deresection.
Caninia aff. cornucopiae Michelin in Gervais 1840Fig. 6: 1a–d, 2a–d, 3–4, 11, 12a and b.Material: Eleven specimens (28 transverse and 4 longitudinal
thin sections).Description: Solitary trochoid to cylindroid coral (Fig. 5: 11–
12b). The calice is shallow, the fossula is well marked and a preem-inent counter septum extending up to the axis is visible in severalspecimens (Fig. 6: 12a). In transverse section, the mean diameter is11 mm (maximum 16 mm) for 24–30 major septa. The major septaare straight and long, thickened in the cardinal quadrants, theirbase being thick in all quadrants. In juveniles stages, all septa joinin the center of the corallite, forming a cluster incised by an opencardinal fossula (Fig. 6: 1c, 2b and c). The cardinal septum is shortand enclosed in a fossula edged by the pericardinal major septa.The counter septum is long (1–3 mm longer than the other septa),even in juvenile stages and usually thickened in its axial end, form-ing a weak columella. The minor septa appear where the diameteris larger than 10–11 mm, they never exceed 1.5 mm. The dissepi-mentarium is composed of 1–4 rows of small interseptal dissepi-ments (herringbones if more than one row, Fig. 6: 4) with one ortwo transeptal dissepiments (first and second order) where thediameter exceeds 12–13 mm (Fig. 6: 2d). In longitudinal section,the tabulae are incomplete, flat or mesa-shaped tabulae in the in-ner part of the tabularium and domed in the peripheral parts(Fig. 6: 3). The inner tabulae are upturned toward the axis wherethe counter septum is elongated. The dissepiments are large andstrongly downturned toward the tabularium. There are 16 tabulaeand 5 dissepiments along a 10 mm-long longitudinal section ofcoral.
Discussion: These specimens can easily be classified in thegenus Caninia as they share with this genus (e.g. Hill, 1981, p.F339) the same septal arrangement the same dissepimentariumand the same tabularium. As in the species C. cornucopiae Michelin
in Gervais 1840, the specimens show an elongated counter septumforming a columella in the early stages of development. Usually,this axial structure disappears rapidly but it is not rare to see spec-imen with a long counter septum in advanced growth stages. Suchphenomenon is commonly observed in Caninia aff. cornucopiae inthe Belgian Tournaisian formations (Denayer et al., 2011). How-ever, in ‘‘classical’’ C. cornucopiae, the mature stages show amplex-oid septa. The amplexoid stage is absent in our material, suggestingperhaps that the mature characters are delayed in C. aff. cornuco-piae. The oldest species of Caninia is early Tournaisian in age (Has-tarian, RC2 coral zone of Poty et al. (2006)) but specimen showing along-lasting columella are not known before the late Tournaisian(Ivorian, RC3).cf. Familly Laccophyllidae Grabau 1928cf. GenusGorizdronia Ró _zkowska 1969
cf. Gorizdronia sp.Fig. 6: 5a–c, 6–7, 8–10.Material: Four specimens (9 transverse and 5 longitudinal
sections).Description: Conical, cylindrical or scolecoid solitary coral com-
monly showing constriction–rejuvenescence cycles (Fig. 6: 8–10).The mean diameter is 11 mm and there are 22 major septa (max-imum 26). The major septa are long and sinuous but do not reachthe axis of the coral and leave a septa-free zone 4 mm-wide in thecenter of the tabularium (Fig. 6: 5a and b, 6–7). They are thickenedat the base and the thickening decreases towards the axial endwhich is sharp. One specimen shows short amplexoid septa. Inthe cardinal quadrants, the septa converge slightly towards aninconspicuous cardinal fossula (Fig. 6: 5b). The cardinal septumis slightly shorter than the others. The minor septa are restrictedto septal crests on the wall. There are no dissepiments. The wallis thin in transverse section. In longitudinal section, the tabulaeare incomplete, dome- or mesa-shaped in the central part of thetabularium. The peripheral part is depressed and counts severalsmall rounded tabulae. There are 7 tabulae (20 in the peripheralpart) in a 10 mm-long longitudinal section through the coral.
Discussion: These specimens share several characters with thegenus Gorizdronia: (1) the septa are simple and sinuous; (2) thecardinal septum is short; (3) the minor septa are almost not devel-oped; (4) there is no dissepimentarium; and (5) the tabulae are flatand mesa-shaped in the central part and more globular in theperiphery. The main difference is the relative development of thecardinal fossula, usually inconspicuous in Gorizdronia. Moreover,this genus is characterized by an aulos in the youngest stages ofdevelopment (‘‘axial tube’’ of Ró _zkowska (1969)) that disappearsrapidly, contrarily to the genera Nalivkinella and Amplexocarinia.The remains of this aulos is questionably observed in the almostfused axial ends of septa in our smallest sections (Fig. 6: 5a). Mor-phology and dimensions are similar to G. profunda profunda figuredby (Ró _zkowska, 1969) but this species is Famennian while thepresent specimens are upper Tournaisian or lower Viséan. Kora(1992) figured a large Amplexocarinia sp. from the Viséan of Sinaiwhich is morphologically similar to our cf. Gorizdronia and is ques-tionably the same taxa. Since Gorizdronia is a Devonian genus, theauthors prefer to keep these specimens in open nomenclature (seebelow). Several Gorizdronia species are known in the Famennian ofPoland (Berkowski, 2002; Ró _zkowska, 1969), North China (Guo,1990; Liao and Cai, 1987), Siberia (Poty & Onoprienko, 1984), SouthFrance (Semenoff-Tian-Chansky, 1988) and New Mexico (Sorauf,1992) but the latter is questionably a distinct genus. Gorizdroniais not supposed to cross the Devonian–Carboniferous boundary(Berkowski, 2002). Except the coral figured by Kora (1992) asAmplexocarinia, Gorizdronia was never signaled in the Early Car-boniferous. Note that our material is also morphologically similarto n. gen. n. sp. aff. Kabakovitchiella described from the uppermostDevonian of the Rhenish Massif (Germany, Weyer, 1993) alsoshowing a simple morphology, withdrawn septa and flat mesa-

354 J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357
shapes tabulae. The main differences with the German taxa is alower ratio number of septa/diameter and straighter septa.
Gen. et sp. indet.Fig. 2: 12a–d.Material: One single specimen (3 transverse and 2 longitudinal
thin sections).Description: The single specimen is a fragment (20 mm long) of
a scolecoid coral with a diameter of 12 mm, showing two constric-tion/rejuvenescence cycles. The smallest section has a diameter of7.5 mm and shows 22 septa, pinnately arranged (Fig. 4: 12a). Thesepta are long and thick, they reach the coral axis and fuse in athick axial stereocolumn incised by a long cardinal fossula(Fig. 4: 12a). In the mature stages (11 mm, 22 septa) the septaare irregular in thickness and length (0.8–4 mm long), their axialends are sharp, rounded or rhopaloid (Fig. 4: 12c). There are nominor septa. The cardinal (?) fossula is poorly marked and the car-dinal and counter septa are not recognizable. The wall is straightand up to 1 mm-thick. The tabulae are flat, horizontal or oblique,complete or not (Fig. 4: 12d).
Discussion: The pinnately arranged (zaphrentoid-type) septaobserved in the early stage of development, associated with septaof different length in the mature stages are two characters of Ufi-mia Stuckenberg 1895 and Zaphrufimia Fedorowski 2012 (see dis-cussion of these genera in Fedorowski (2012)). However, thecounter, cardinal and alar septa are shorter while pericounterand perialar septa are longer in these genera. In the Turkish spec-imen, the septa are very irregular in length and shape (sharp,rounded, rhopaloid). The generic attribution of this specimen isnot easy and the authors do not exclude the idea of consideringit as a pathologic form of any zaphrentoid coral or even of a caninidone. Note that the specimen figured by Kora (1995) as Soshkineo-phyllum sp. also possesses a irregular septal pattern but the coun-ter septum is particularly long in this specimen.
4. Discussion
4.1. Biostratigraphy
None of the corals in the Köprülü Formation has a precise bio-stratigraphical value, all being long-ranging taxa. Caninia aff. cor-nucopiae indicates an age at least late Tournaisian as columellatespecimen of this species are not known in Europe before the Ivori-an (Denayer et al., 2011). In the same manner, Amplexizaphrentis isnot known before the Moliniacian (lower Viséan) in Eurasia(Vaughan, 1906). Nonetheless it is present in the Kinderhookian(lower Tournaisian) in North America (Sando and Bamber, 1985).Rotiphyllum is also more common in the Viséan than in the Tour-naisian in Europe and China (Xu and Poty, 1997) but already pres-ent in the Tournaisian in North America (Sando and Bamber, 1985).In conclusion, an early Viséan age is suspected for the S�ort Derecoral assemblage. Palynomorphs from the same horizons give alate Tournaisian age (pretiosus-clavata-PC Miosopore zone; Higgset al., 2002). The poor foraminifers assemblage preliminary de-scribed by Perinçek et al. (1991) are compatible with a late Tour-naisian or early Viséan age.
4.2. Faunal assemblage
This coral assemblage is a typical example of the so-called ‘‘Cya-thaxonia Fauna’’ sensu Hill (1938–41) also named laccophyllid fau-na sensu Oliver (1992). Taxonomically the corals are cosmopolitanand long-ranging genera often with a simple skeletal organization.They also constitute a pioneer coral fauna, rapidly recovering theextinction of ‘‘complex’’ taxa after a crisis. In the original descrip-tion of Hill (1938–41) and Oliver (1992), this coral assemblage is
supposed to witness a deep environment but is common in mar-ginal, stressed or unfavorable environments where other coralwere not able to live. Such associations usually indicate dysphotic,dysaerobic, cold-water, hyperhaline (?) and turbid environments(Scrutton, 1998). Small solitary horn-shaped non-dissepimentedcorals are common in mixed carbonate–siliciclastic facies, inde-pendent of the age or palaeogeographical position. Table 1 summa-rize the diversity of such assemblage in some well studiedlocalities. Usually, the number of taxa is low and the ratio numberof species/number of genera is close to 1, indicating a weak dispar-ity. The presence of columellate and/or dissepimented taxa is occa-sional, witnessing the ability of some ‘‘complex’’ coral (e.g. Caniniaaff. cornucopiae) to colonize less favorable environments. In theclassification of Aretz (2010), this small coral fauna belongs tothe A2 type (low diversity level-bottom community).
4.3. Palaeobiogeography
A stated in the introductive part, the Hakkari area forms thenorthern part of the Arabian Plate which belongs to the northernmargin of the Gondwana continent during Early Carboniferoustimes (Fig. 7). The Hakkari area was situated in a more proximal(southward) position than the Taurides, North Iran Block and Al-borz also forming the northern edge of Gondwana (e.g. reconstruc-tions by Stampfli and Borel (2002), von Raumer et al. (2003) andFig. 8) and was obviously submitted to a detrital influx witnessedby the mixed carbonate–siliciclastic sedimentation. The sedimen-tary record is anyway more complete in the latter zone than inthe Arabian Plate s. s. (Husseini, 1992). The fossil components isalso poorer in that zone as shown by weak diversity of coral faunadescribed here. The comparison can be made with several Gondw-anian-related areas yielding Lower Carboniferous coral fauna.
(1) The Sinai Peninsula (NE Egypt). Kora (1989; 1992) and Koraand Jux (1986) described form the Um Bogma Formation(lower? to middle Viséan) a poorly diversified fauna com-posed mainly of small non-dissepimented solitary rugosecorals, namely Amplexizaphrentis, Zaphrentites, Bradyphyllumand, secondarily, Amplexocarinia (possibly corresponding toour cf. Gorizdronia), Soshkineophyllum (our gen. et sp. indet.?)and Clisiophyllum (questionably a Canina after Kora’s (1995)Fig. 4g, j–k). This fauna (6 genera belonging to 5 families) is atypical assemblage of coral taxa found in mixed carbonated–siliciclastic sediments in the Carboniferous (Sobhy andEzaki, 2006). There is an important facies bias that enablethe attribution of this assemblage to any palaeobiogeo-graphic province, even if Kora (1995) did it in consideringits fauna as typical of the Western Europe Province of Sando(1990).
(2) The Aladag Unit in the Turkish Taurides. Data from Frech(1916) and unpublished material collected by the firstauthor in the Hadım vicinity is composed – for the late Tour-naisian–early Viséan – of small non-dissepimented solitarycorals (Zaphrentites and Proheterelasma) in shale and lime-stone alternation, but also of large dissepimented taxa inpurer carbonate facies (Caninia, Bifossularia, Siphonophyllia,Uralinia, Keyserlingophyllum, Caninophyllum, Kueichouphyl-lum, Calmiussiphyllum, Amygdalophyllum). The assemblageis more diversify (10 genera belonging to 6 families), includ-ing columellate and dissepimented taxa.
(3) North Iran Block (Sanadaj-Sirjan and Alborz). The compila-tions of Flügel (1963) and Heravi and Khaksar (1999) pres-ent, for the Tournaisian, at least 10 genera from 5 families(Amplexizaphrentis, Zaphrentites, Keyserlingophyllum Caninia,Siphonophyllia, Caninophyllum, Kueichouphyllum, Zaphriphyl-lum, and questionably Cyathoclisia and Carruthersella). As

Table 1Age, location and diversity of some well studied Upper Palaeozoic ‘‘Cyathaxonia fauna’’.
Age Locality Facies Nb. sp./gen.
Dispt.taxa
References
Emsian Anti-Atlas (Morocco) Deep-water mound 19/12 2 Berkowski (2008); Berkowski (2004)Late Emsian–early Eifelian Istanbul area (NW Turkey) Deep-water silicicalstic 5/5 0 Kullmann (1973)Givetian Anti-Atlas (Morocco) Deep-water mound 4/4 0 Berkowski (2006)Early Famennian Harz (Germany) Deep water carbonate 4/4 0 Weyer (1991)Early Famennian Thuringia (Germany) Deep water mixed carbonate–
siliciclastic5/5 0 Weyer (2004)
early Famennian Sauerland (Germany) Deep water mixed carbonate–siliciclastic
8/6 0 Weyer (2004)
Early Famennian Namur-Dinant Basin(Belgium)
Shallow water siliciclastic 4/4 2 Denayer et al. (2012)
Late Famennian Sinkiang (NE China) Deep water carbonate 11/9 2 Soto and Liao (2002)Latest Famennian–early
TournaisianRhenish Massif (Germany) Deep-water silicicalstic and
carbonate17/13 1 Weyer (1994), Korn and Weyer (2003)
Early Viséan Hakkari (SE Turkey) Mixed carbonate–siliciclastic 7/6 1 This paperEarly-mid. Viséan Sinai (NE Egypt) Mixed carbonate–siliciclastic 9/6 1 Kora (1989, 1992, 1995) and Kora and Jux
(1986)Late Tournaisian–early Viséan Baltic coast (NE Germany) Mixed carbonate–siliciclastic 9/7 1 Weyer (1993)Late Tournaisian–early Viséan Montagne Noire (S France) Deep-water carbonate 9/7 1 Semenoff-Tian-Chansky (1988)Late Serpukhovian Arkansas (USA) Reefal (thrombolite) limestone 9/8 1 Webb (1987)Late Serpukhovian Oklahoma and Arkansas
(USA)Siliciclastics 11/5 0 Webb and Sutherland (1993)
Lower Pennsylvanian Texas (USA) Carbonate and siliciclastic 24/10 3 Moore and Jeffords (1945)Moscovian Cantabrian Mountains (N
Spain)Carbonate 31/18 0 Rodríguez and Kullmann (1999)
Late Moscovian South Urals (Russia) Warm water carbonate 7/6 0 Kossovaya (2007)Late Moscovian (top) South Urals (Russia) Mixed carbonate–siliciclastic 7/5 1 Kossovaya (2007)Moscovian-Gzhelian Central USA Carbonate 24/3 0 Jeffords (1947)Cisuralian Subpolar-Northern Urals
(Russia)Shalow cool water siliciclastic 19/10 0 Kossovaya (2007)
Cisuralian Central Urals (Russia) Deep cool water siliciclastic 8/7 0 Kossovaya (2007)Cisuralian South Urals (Russia) Warm water reefal carbonate 8/6 0 Kossovaya (2007)Guadalupian Arctic Canada Carbonate 8/6 0 Fedorowski and Bamber (2001)
J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357 355
for the Aladag Unit of the Taurides, the Central Iran belongsto the Kueichouphyllym Zone which corresponds in part tothe Minor Asian, Middle Asian and Chinese Provinces ofSando (1990).
(4) The Badakhshan (NE Afghanistan). Von Schouppé (1970)gives a short description of the upper Tournaisian fauna.The most common corals are Caninophyllum, together withZaphrentites, Amygdalophyllum and Proheterelasma. Thefacies are mostly shallow-water carbonates. The fauna asso-ciation is poor (4 genera, 4 families) but questionably incom-plete (preliminary data).
The Turkish Taurides, Iran and Afghanistan areas are easilycomparable from a faunal point of view, composed of both non-dissepimented and dissepimented solitary rugose corals. The twofirst area are characterized by the occurrence of the genus Kueic-houphyllum. The Sinai and Hakkari assemblages show a strong sim-ilarity, both being dominated by small solitary non-dissepimentedcorals. Further comparison is delicate because the faunal associa-tions are strongly facies-related. Any palaeobiogeographic inter-pretation is thus unrepresentative.
5. Conclusions
In the Hakkari area (SE Turkey), the Early Carboniferous, repre-sented by the Köprülü Formation, crops out in the S�ort Dere sectionin the Zap river valley. The lower part of the formation yields anabundant but poorly diversified coral fauna. The coral assemblageis mainly composed of small non-dissepimented solitary coralsbelonging to the genera Rotiphyllum, Zaphrentites, cf. Gorizdronia,gen. et sp. indet. and Amplexizaphrentis, including a new species,A. zapense – and the dissepimented coral Caninia. The coral assem-blage is similar to that described from the Um Bogma Formation of
the Sinai Peninsula (NE Egypt). The Sinai and Hakkari area werepart, during the Early Carboniferous, of the northern margin ofthe Gondwana, and edged northward by a carbonate platform, to-day dismembered and incorporated in the Alpino-Himalayan oro-gen within the Turkish Taurides, northern Iran and Afghanistan.Those areas contain coral assemblages sharing several taxa withthe Hakkari and Sinai but have a higher diversity. The Köprülü For-mation gives an idea of the coral fauna that lived in the southernArabian mixed carbonate–siliciclastic platform.
Acknowledgements
The authors acknoledge Edouard Poty and Markus Aretz whoprovided constructive remarks on the early version of the manu-script and stimulating discussions on systematics and stratigraphy.Olga Kossovaya and an anonymous reviewer are thanked for theirhelpfull comments and advice. This paper is part of the PhD Thesisof the first author who benefited finantial supports from the Bel-gian national fund for scientific research (FNRS-FRIA) and the Uni-versity of Liège.
References
Aretz, M., 2010. Habitats of colonial rugose corals: the Mississippian of westernEurope as example for a general classification. Lethaia 43 (4), 558–572.
Aretz, M., 2011. Corals from the Carboniferous of central Sahara (Algeria): thecollection ‘‘Marie Legrand-Blain’’. Geodiversitas 33, 581–624.
Ball, J., 1916. The Geography and Geology of West-central Sinai. Egyptian SurveyDepartment, Cairo, pp. 151–163.
Berkowski, B., 2002. Famennian Rugosa and Heterocorallia from Southern Poland.Palaeontologia Polonica 61, 1–87.
Berkowski, B., 2004. Monospecific rugosan assemblage from the Emsianhydrothermal vents of Morocco. Acta Palaeontologica Polonica 49 (1), 75–84.
Berkowski, B., 2006. Vent and mound rugose coral associations from the middleDevonian of Hamar Laghdad (Anti-Atlas, Morocco). Geobios 39, 155–170.

356 J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357
Berkowski, B., 2008. Emsian deep-water Rugosa assemblages of Hamar Laghdad(Devonian, anti-Atlas, Morocco). Palaeontographica, Abteilung A 284 (1–3), 17–68.
Brenckle, P.L., Marchant, T.R., 1987. Calcareous microfossils, depositionalenvironments and correlation of the lower Carboniferous Um Bogmaformation at Gebel Nukhul, Sinai, Egypt. Journal of Foraminiferal Research 17(1), 74–91.
Carruthers, R.G., 1910. On the evolution of Zaphrentites delanouei in lowerCarboniferous times. Geological Society of London, Quarterly Journal 66, 523–538.
Cater, J.M.L., Tunbridge, I.P., 1992. Palaeozoic tectonic history of SE Turkey. Journalof Petroleum Geology 15 (1), 35–50.
Chwieduk, E., 2005. Late Devonian and early Carboniferous Rugosa from westernPomerania, northern Poland. Acta Geologica Polonica 55, 393–443.
Cotton, G., 1973. The Rugose Coral Genera. Elsevier, p. 358.Dean, W.T., Zhou, Z., 1988. Upper Ordovician trilobites from the Zap Valley, south-
east Turkey. Palaeontology 31, 621–649.Denayer, J., Poty, E., Aretz, M., 2011. Uppermost Devonian and Dinantian rugose
corals from Southern Belgium and surrounding areas. In: Aretz, M., Poty, E.(Eds.), 11th International Symposium on Fossil Cnidaria and Porifera, Liège,2011, Field guides, vol. 20. Kölner Forum für Palaöntologie und Geologie, pp.151–201.
Denayer, J., Poty, E., Marion, J.-M., Mottequin, B., 2012. Lower and middleFamennian (upper Devonian) rugose corals from southern Belgium andnorthern France. In: Denayer, J., Aretz, M., Poty, E. (Eds), Proceedings of the11th International Symposium on Fossil Cnidaria and Porifera, vol. 15/4. Liège2011. Geologica Belgica, pp. 273–283.
Dobroljubova, T.A., Kabakovich, N.V., Sayutina, T.A., 1966. Chetyrekhluchevyekorally nizhnego karbona Kuznetskoy kotloviny. Akademiya Nauk SSSR,Paleontologicheskii Institut, Trudy 111, 5–198.
Dybowski, W.N., 1873. Monographie des Zoantharia Sclerodermata Rugosa aus derSilurformation Estlands, Nord-Livlands und der Insel Gotland. Archiv fürNaturkund Liv-, Ehst- und Kurlands 5, 257–414.
Easton, W.H., 1944. Corals of the Chouteau and related formations of the MississippiValley region. Illinois State Geological Survey 97, 1–93.
Fan, Y., Yu, X., He, Y., Pan, Y., Li, X., Wang, F., Tang, D., Chen, S., Zhao, P., Liu, J., 2003.The Late Palaeozoic Rugose Corals of Xizang (Tibet) and Adjacent Regions andtheir Palaeobiogeography. Hunan Science and Technology Press, p. 679.
Fedorowski, F., 2009. Early Bashkirian Rugosa (Anthozoa) from the Donets Basin,Ukraine. Part 1. Introductory considerations and the genus RotiphyllumHudson, 1942. Acta Geologica Polonica 29, 1–37.
Fedorowski, J., 2012. The new Upper Serpukhovian genus Zaphrufimia andhomeomorphism in some rugose corals. Palaeontographica Beiträge zurNaturgeschichte der vorzeit, Abteilung A: Paläozoologie-Stratigraphie, 109–161.
Fedorowski, F., Bamber, E.W., 2001. Guadlupian (middle Permian) solitary rugosecorals from the Degerböls and Trol Fiord formations, Ellesmere and Melvilleislands, Canadian Arctic Archipelago. Acta Geologica Polonica 51, 31–79.
Flügel, H., 1963. Korallen aus der oberen Visé-Stufe (Kueichouphyllum-Zone) Nord-Irans. Jahrbuch des Geologischen Bundesanstalt 106, 365–404.
Fomichev, W.D., 1953. Korally Rugosa I stratigrapfija sredne Iverkhnekamennougol’nykh I permskikh otlozhenij Donetskogo bassejna.Trudy VSEGEI, Moscow 1, 1–622.
Frech, F., 1916. Geologie Kleinasiens im Bereich der Baghdadbahn, Ergebnisseeigener Reisen, vergleichender Studien und paläontologischer Untersuchungen.Sonder-Abdruck aus der Zeitschrift der Deutschen Geologischen Gesellschaft68, 1–322.
Garwood, E.J., 1913. The lower Carboniferous succession in the north-west ofEngland. Quarterly Journal of the Geological Society, London 68, 449–586.
Gourvennec, R., Hos�gör, I., 2012. New record of a middle Devonian brachiopodfauna from the northern Arabian Plate, Zap Anticline, Çukurca-Hakkari,southeastern Turkey. Bulletin of Geosciences (87/2), 347–358.
Göncüoglu, M.C., Dirik, K., Kozlu, H., 1997. General characteristics of pre-Alpine andAlpine Terranes in Turkey: explanatory notes to the terrane map of Turkey.Annales Géologiques des Pays Hélléniques 37, 515–536.
Grabau, A.W., 1928. Palaeozoic corals of China: Part 1, Tetraseptata II, secondcontribution. Palaeontologica Sinica, Serie B 2/2, 1–175.
Guo, S., 1990. Frasnian–Famennian extinction and late Devonian rugose corals formGreat Xing’an Ranges, NE China. Acta Palaeontologica Sinica 10, 431–445.
Heravi, M.A., Khaksar, K., 1999. An assemblage of corals of Iran. Geological Survey ofIran, 357.
Higgs, K.T., Finucane, D., Turnbridge, I.P., 2002. Late Devonian and earlyCarboniferous microfloras from the Hakkari province of southeastern Turkey.Review of Palaeobotany and Palynology 118, 141–156.
Hill, D., 1938. A monograph of the Carboniferous rugose corals of Scotland.Palaeontographical Society, London, 213.
Hill, D., 1981. Coelenterata; Supplement 1, Rugosa and Tabulata, In: Teichert, C.(Ed.), Treatise on Invertebrate Paleontology, Part F (2 vols.), The GeologicalSociety of America and the University of Kansas. Boulder, Colorado, Lawrence,Kansas, 762 p.
Hos�gör, I., Okan, Y., Göncüoglu, M.C., 2012. Posidonia becheri Bronn, 1828 from theTournaisian of SE Turkey. A palaeobiogeographic enigma. Comptes RendusPalevol (11/1), 13–20.
Hos�gör, I., Okan, Y., 2012. Early Carboniferous ‘non Paleozoic-Type’ Monotaxic shellbeds from the northern Arabian Plate (Hakkari-Çukurca area, SE Turkey): a newGondwanan occurrence. In: Yalçın, M.N., Çorbacıoglu, H., Aksu, Ö., Bozdogan, N.
(Eds.), Paleozoic of Northern Gondwana and its Petroleum Potential a fieldWorkshop, vol. 6, 9–14 September. Turkish Association of Petroleum Geology,Special Publication, pp. 41–45.
Hudson, R.G.S., 1941. On the Carboniferous corals: Zaphrentis carruthersi sp. nov.from the Mirk fell beds and its relation to the Z. delanouei species-group.Proceedings of the Yorkshire Geological Society 24, 290–311.
Hudson, R.G.S., 1942. Fasciculophyllum Thomson and other genera of the‘‘Zaphrentis’’ omaliusi group of Carboniferous corals. Geological Magazine(79/5), 257–263.
Hudson, R.G.S., 1944. On the lower Carboniferous corals: Zaphrentites crassus and Z.tenuis, spp. n. Annal and Magazine of Natural History, 145–154.
Husseini, M.I., 1992. Upper Palaeozoic tectono-sedimentary evolution of theArabian and adjoining plates. Journal of the Geological Society, London 149,419–429.
Iljina, T.G., 1970. Nekotorye novye permskie rugozy yugovstochnogo Pamira. In:Astrova, G.G., Chudinova, I.I. (Eds.), Novye vidy paleozoyskikh mshanok ikorallov. Nauka, Moscow, pp. 146–151.
Ivanowski, A.B., 1967. Etyudy o rannekamennougolnykh rugozakh. Nauka, Moscow,p. 97.
Janvier, P., Lethiers, F., Monod, O., Balkas�, Ö., 1984. Discovery of a vertebrate faunaat the Devonian–Carboniferous boundary in SE Turkey (Hakkari province).Journal of Petroleum Geology 7, 147–168.
Jeffords, RM., 1947. Pennsylvanian Lophophyllidid corals. University of Kansas,Publications, Paleontological Contributions Number 1, 1–28.
Jux, U., Omara, S., 1983. Pachypteria sinaitica n. sp. – eine aufgewachsene,austerähnliche Muschel aus dem Unterkarbon Ägyptens. PaläontologischeZeitschrift, 79–91.
Klebelsberg, R., 1911. Ein Beitrag zur Kenntnis des Sinai-Karbons. Zeitschrift derDeutschen Geologischen Gesellschaft 63, 594–603.
Kato, M., 1979. Some upper Palaeozoic corals from Turkey. Journal of the Faculty ofScience, Hokkaido University, 137–148.
Kora, M., 1989. Lower Carboniferous (Viséan) fauna from Wadi Budra, west-centralSinai, Egypt. Neues Jahrbuch für Geologie und Paläontologie Monatshefte 9,523–538.
Kora, M., 1992. Carboniferous macrofauna from Wadi Khaboba, West-Central Sinai(Egypt). Geologica et Palaeontologica 26, 13–27.
Kora, M., 1995. Carboniferous macrofauna from Sinai, Egypt: biostratigraphy andpalaeogeography. Journal of African Earth Sciences 20, 37–51.
Kora, M., Jux, U., 1986. On the early Carboniferous macrofauna from the Um Bogmaformation, Sinai. Neues Jahrbuch für Geologie und Paläontologie Monatshefte 2,85–98.
Korn, D., Weyer, D., 2003. High resolution stratigraphy of the Devonian–Carboniferous transitional beds in the Rhenish mountains. Mitteilungen ausdem Museum für Naturkunde Berlin, Geowissenschaftliche Reihe 6, 79–124.
Kossovaya, O., 2007. Ecological aspect of upper Carboniferous–lower Permian‘Cyathaxonia Fauna’ taxonomical diversity (the Urals). In: Hubmann, B., Piller,W.E. (Eds), Fossil Corals and Sponges. Proceedings of the 9th InternationalSymposium on Fossil Cnidaria and Porifera. Österrichischen Akademie derWissenschaften, vol. 17, pp. 383–405.
Köylüoglu, M., Altıner, D., 1989. Micropaléontologie (Foraminifères) etbiostratigraphie du Permien supérieur de la région d’Hakkari (SE Turquie).Revue de Paléobiologie 8, 467–503.
Kullmann, J., 1973. Goniatite-coral associations from the Devonian of Istanbul. In:Kaya, O. (Ed.), Paleozoic of Istanbul, vol. 40. Ege Üniversitesi Fen FakültesiKitaplar Serisi, Turkey, pp. 7–116.
Kullman, J., Liao, W.-H., 1985. Hornförmige Einzelkorallen (Rugosa) aus demUnterkarbon von Süd-China. Palaeontographica 189, 125–157.
Lang, W.D., Smith, S., Thomas, H.D., 1940. Index of Palaeozoic coral genera. BritishMuseum of Natural History, London, p. 231.
Liao, W., Cai, T., 1987. Sequence of Devonian rugose coral assemblages fromnorthern Xinjiang. Acta Palaeontologica Sinica 26, 690–707.
Michelin, H., 1840. Astrée, Astrea. In: Gervais, P. (Ed.), Dictionnaire des SciencesNaturelles, Paris, supplement 1, pp. 481–487.
Milne-Edwards, H., Haime, J., 1850–1855. A monograph of the British fossil corals.Palaeontographical Society, London, Monography, pp. 1–71 (1850), 147–210(1851), 211–244 (1853), 245–299 (1855).
Minato, M., 1955. Japanese Carboniferous and Permian corals. Journal of the Facultyof Science, Hokkaido University, Series IV: Geology and Mineralogy 9 (2), 1–202.
Mitchell, W.I., 1989. Biostratigraphy of Viséan (Dinantian) rugose coral faunas ofBritain. Proceedings of the Yorkshire Geological Society 47, 233–247.
Moore, C.M., Jeffords, R.M., 1945. Description of lower Pennsylvanian corals fromTexas and adjacent states. The University of Texas Publication 4401, 77–208.
Oliver Jr., W.A., 1992. Corals form the Turkey Creek limestone (lower Devonian),southern Oklahoma. Oklahoma Geological Survey Bulletin 145, 137–159.
Omara, S., 1967. Contribution to the stratigraphy of the Egyptian Carboniferousexposures. 6th Arabian Petroleum Congress, Baghdad 44, 1–7.
Omara, S., 1971. Early Carboniferous tabulate corals from Um Bogma area, Sinai,Egypt. Rivista Italiana de Paleontologia (77/2), 141–154.
Omara, S., Conil, R., 1965. Lower Carboniferous foraminifera from southwesternSinai, Egypt. Annales de la Société géologique de Belgique (88/5), 221–245.
Perınçek, D., 1990. Stratigraphy of the Hakkari Province, Southeast Turkey. TurkishAssociation of Petroleum Geologists Bulletin 2, 21–68.
Perınçek, D., Duran, O., Bozdogan, N., Çoruh, T., 1991. Stratigraphy andpaleogeographical evolution of the autochthonous sedimentary rocks inSoutheast Turkey, In: Turgut, S., (Ed.), Tectonics and hydrocarbon potential ofAnatolia and surrounding regions. Ozan Sungurlu Symposium, Proceedings of

J. Denayer, I. Hos�gör / Journal of Asian Earth Sciences 79 (2014) 345–357 357
the Turkish Petroleum Corporation, Turkish Association of PetroleumGeologists, 274–305.
Poty, E., 1981. Recherches sur les Tétracoralliaires et les Hétérocoralliaires duViséen de la Belgique. Mededelingen Rijks Geologische Dienst (35/1), 1–161.
Poty, E., Devuyst, F.-X., Hance, L., 2006. Upper Devonian and Mississippianforaminiferal and rugose coral zonations of Belgium and northern France: atool for Eurasian correlations. Geological Magazine 143, 829–857.
Poty, E., Onoprienko, Y., 1984. Rugose corals. In: Shilo, N.A., Bouckaert, J., Afanasieva,M.J.M., Bless, M.J.M., Conil, R., Erlanger, O.A., Gagiev, M.H., Lazarev, S.S.,Onoprienko, Y., Poty, E., Razina, T.P., Simakov, K.V., Smirnova, L.V., Streel, M.,Swennen, R. (Eds.), Sedimentological and Palaeontological Atlas of the LateFamennian and Tournaisian Deposits in the Omolon Region (NE USSR). Annalesde la Société géologique de Belgique 107, 200–213.
Rodríguez, S., Falces, S., 1992. Corales Rugosos. In: Rodríguez, S., Arribas, M.E.,Comas-Rengifo, M., de la Peña, J., Falces, S., Kullmann, J., Gegundez, P., Legrand-Blain, M., Martinez-Chacon, M.L., Moreno-Eiris, Y.E., Perejon, A., Sanchez, J.L.,Sanchez-Chico, F., Sarmiento, G. (Eds.), Análisis paleontológico ysedimentológico de la cuenca carbonífera de Los Santos de Maimona(Bajadoz). Coloquios de Paleontología 44, 49–90.
Rodríguez, S., Falces, S., 1994. Coral distribution patterns in the Los Santos deMaimona Lower Carboniferous Basin (Badajoz, SW Spain). CourrierForschunginstitut Seckenberg 172, 193–202.
Rodríguez, S., Kullmann, J., 1999. Rugose corals from the upper member of the Picosde Europa Formation (Moscovian, Cantabrian Mountains, NW Spain).Palaeontographica, Abteilung A 284 (1–3), 23–92.
Ró _zkowska, M., 1969. Famennian tetracoralloid and heterocoralloid fauna from theHoly Cross mountains (Poland). Acta Palaeontologica Polonica 24, 1–187.
Sando, W.J., 1990. Global Mississippian coral zonation. Courrier Forschungs-InstitutSenckenberg 130, 173–187.
Sando, W.J., Bamber, E.W., 1985. Coral zonation of the Mississippian system in theWestern interior province of North America. U.S. Geological Survey ProfessionalPaper 1334, 1–61.
Scrutton, C.T., 1998. The Palaeozoic corals, II: structure, variation andpalaeoecology. Proceedings of the Yorkshire Geological Society 52 (1), 1–57.
Semenoff-Tian-Chansky, P., 1974. Recherches sur les Tétracoralliaires duCarbonifère du Sahara Occidental. Mémoires du Muséum National d’HistoireNaturelle, série C Sciences de la Terre 30, 1–316.
Semenoff-Tian-Chansky, P., 1988. Corals from the Devonian–Carboniferousboundary at La Serre (Montagne Noire, France). Courrier Forschungs-InstitutSenckenberg 100, 129–138.
Smith, A.B., 1910. The faunal succession of the Upper Bernician. Transactions of theNatural History Society of Northumberland, Durham and Newcastle-Upon-Thyne 3, 591–644.
Sobhy, M., Ezaki, Y., 2006. First record of Heterocorallia (Hexaphyllia Stuckenberg,1904) from the lower Carboniferous (Viséan) of west-central Sinai, Egypt.Senckenbergiana lethaea 86 (1), 1–21.
Sorauf, J.E., 1992. Late Devonian (Famennian) rugose coral fauna of the Percha shaleof southwestern New Mexico. Journal of Paleontology 66, 730–749.
Soto, F., Liao, W.-H., 2002. Laccophyllidae Grabau 1928 (Rugosa) from theHongguleleng formation (Devonian: lower Famennian) at the northwesternmargin of Junggar basin (northern Xinjiang, NW China). Coral Research Bulletin7, 209–220.
Stampfli, G.M., 2000. Tethyan oceans. In: Bozkurt, E., Winchester, J.A., Piper, J.D.A.(Eds.), Tectonics and Magmatism in Turkey and Surrounding Area, vol. 173.Geological Society of London, Special Publication, pp. 1–23.
Stampfli, G., Borel, G.D., 2002. A plate tectonic model for the Palaeozoic andMesozoic constrained by dynamic plate boundaries and restored syntheticocean isochrons. Earth and Planetary Science Letters 169, 17–33.
Stampfli, G., Borel, G.D., 2004. The TRANSMED transects in space and time:constraints on the paleotectonic evolution of the Mediterranean domain. In:Cavazza, W., Roure, F., Spakman, W., Stampfli, G., Ziegler, P.A. (Eds.), TheTRANSMED Atlas: the Mediterranean Region from Crust to Mantle. Springer-Verlag, pp. 53–80.
Stampfli, G., Marcoux, J., Baud, A., 1991. Tethyan margins in space and time.Palaeogeography, Palaeoclimatology, Palaeoecology 87, 373–410.
Stampfli, G., Mosar, J., de Bono, A., Vavasis, I., 1998. Late Paleozoic, early Mesozoicplate tectonics of the western tethys. Bulletin of the Geological Society ofGreece 32, 113–120.
Stampfli, G., Von Raumer, J.F. & Borel, G.D., 2002. Paleozoic evolution of pre-Variscan terranes: From Gondwana to the Variscan collision. In: Martinez-Catalàn, J.R., Hatcher, D.R.J., Arenas, R. & Diaz-Garcia, F. (Eds), Variscan-Appalachian dynamics: the building of the Late Paleozoic basement.Geological Society of America, Special Paper, 364, pp. 263-280.
Stuckenberg, A., 1895. Korallen und Bryozoen der Steinkohlen ablagerungen desUral und des Timan. Mémoires du Comité de Géologie de Saint-Petersbourg 10,1–244.
Sutherland, P.K., 1958. Carboniferous stratigraphy and rugose coral faunas ofnortheastern British Columbia. Geological Survey of Canada, memoir 295, 1–108.
Tolmachev, I.P., 1931. Faune du Calcaire du Bassin du Kuznetsk. Comité géologiquede Leningrad, matériaux pour la géologie générale et appliquée 25, 1–663.
Tunbridge, I.P., 1988. The Upper Devonian clastic wedge in southeastern Turkey. In:McMillian, N.J., Embrey, A.F., Glass, D.J. (Eds.), Devonian of the World, vol. 14.Canadian Society of Petroleum Geologists, Memoirs, pp. 309–311.
Vaughan, A., 1906. The Carboniferous Limestone series (Avonian) of the AvonGorges. Proceedings of the Bristol Naturalists’ Society, 4th serie 1, 73–166.
Vaughan, A., 1908. An account of the faunal succession and correlation. In: Matley,C.A., Vaughan, A. (Eds.), The Carboniferous Rocks at Loughshinny (CountyDublin), vol. 6. Quarterly Journal of the Geological Society, London, pp. 36–472.
Volkova, M.S., 1941. Corals from the Carboniferous of central Kazakhstan. MaterGeoloji Polez i iskop vost Kazakhsatana 11, 1–119.
von Raumer, J.F., Stampfli, G., Busy, F., 2003. Gondwana-derived microcontinents –the constituents of the varsican and alpine collisional orogens. Tectonophysics365, 7–22.
Von Schouppé, A., 1970. Lower Carboniferous corals from Badakhshan (northeastAfghanistan). Italian expedition to the Karakorum (K2) and Hindu Kush.Scientific Reports – IV Palaeontology and Palaeobotany 2, 1–23.
Vuillemin, C., 1990. Les Tétracoralliaires (Rugosa) du Carbonifère inférieur duMassif Armoricain (France). Editions du CNRS, 167p.
Webb, G.E., 1987. The coral faunal of the Pitkin formation (Chesterian),northeastern Oklahoma and northwestern Arkansas. Journal of Paleontology61 (3), 462–493.
Webb, G.E., 1990. Lower Carboniferous coral fauna of the Rockhampton group, east-central Queensland. Memoirs of the Association of Australian Palaeontologists10, 1–167.
Webb, G.E., Sutherland, P.K., 1993. Coral fauna of the Imo formation, uppermostChesterian, north-central Arkansas. Journal of Paleontology 67 (2), 179–193.
Wedekind, R., 1927. Die Zoantharia Rugosa von Gotland. Svergies GeologiskaUndersökning, serie C 19, 1–94.
Weyer, D., 1991. Rugosa (Anthozoa) aus dem Unter-Famenne von Rübeland (Harz).Abhandlungen und Berichte für Naturkunde 15, 25–43.
Weyer, D., 1993. Korallen aus dem Obertournai und Untervisé der Inseln Hiddenseeund Rügen. Abhandlungen und Berichte für Naturkunde 16, 31–69.
Weyer, D., 1994. Korallen im Untertournai-profil von Drewer (RheinischesSchiefergebirge). Geologie and Paläontologie in Westfalen 29, 177–221.
Weyer, D., 2004. Neue Rugosa-Funde aus dem Unter-Famenne von Deutschland(Anthozoa, Oberdevon). Geologische Jahrbruch, Hessen 131, 203–223.
White, C.A., 1862. Description of new species of fossils from the Devonian andCarboniferous rocks of the Mississippi Valley. Boston Society of Natural History,Proceedings 9, 8–33.
Wu, W.-S., 1964. Lower Carboniferous corals in Central Hunan. Memoirs of theInstitute of Geology and Palaeontology 3, 76–162.
Wu, W.-S., Zhang, Y.S., 1980, Carboniferous Rugose Corals from Eastern Xizang andWestern Sichuan, Beijing, 166p.
Wu, W.-S., Zhao, J., 1989. Carboniferous and early Permian Rugosa from westernGuizhou and eastern Yunnan, SW, China. Palaeontologica Sinica 177, 1–224.
Xu, S., Poty, E., 1997. Rugose corals near the Tournaisian–Viséan boundary in SouthChina. Bolltino del Real Societad Espanola de historia natural (section Geologia)92 (1–4), 349–363.
Yalçın, N., Yılmaz, I., 2010. Devonian in Turkey, a review. Geologica Carpathica 61(3), 235–253.
Copyright © 2022 FDOKUMEN




















![MANGROVES. SEAGRASSES AND CORALS [A simple layman-type field guide]](https://static.fdokumen.com/doc/165x107/6321610a0c12e1161503c4a8/mangroves-seagrasses-and-corals-a-simple-layman-type-field-guide.jpg)