
Synthesis, purification, and characterization of human ciliary neuronotrophic factor fromE. coli
Upload
independentCategory
view
2download
0
Therapeutic Benefit of Ciliary NeurotrophicFactor in Progressive Motor Neuronopathy
Depends on the Route of DeliveryGeorg Haase, MD,* Brigitte Pettmann, PhD,† Thierry Bordet, MSc,* Pascal Villa, PhD,†
Emmanuelle Vigne, PhD,‡ Henning Schmalbruch, MD,§ and Axel Kahn, MD*
Ciliary neurotrophic factor (CNTF) has demonstrated therapeutic effects in several mouse mutants with motoneuronaldegeneration. However, the poor bioavailability and toxic side effects of recombinant CNTF protein have complicated itsuse in patients with amyotrophic lateral sclerosis. CNTF gene transfer strategies were developed but faced the questionof whether CNTF should be delivered to motoneuron cell bodies or to their axons or muscle targets. To address thisissue, we have used an adenoviral vector (AdCNTF) coding for a secretable form of CNTF and compared different routesof its administration in the mouse mutant progressive motor neuronopathy (pmn). Intramuscular, intravenous, andintracerebroventricular injections of AdCNTF or the control vector AdlacZ resulted in transgene expression in skeletalmuscle fibers, hepatocytes, and ependymal cells, respectively, as determined by histochemistry and reverse transcription–polymerase chain reaction. AdCNTF intramuscularly treated and intravenously treated pmn mice showed a 25% increasein mean life span and a reduced degeneration of phrenic myelinated nerve fibers, which correlated with elevated CNTFserum bioactivities. In contrast, intracerebroventricular AdCNTF administration did not affect the mean life span ormotor axonal degeneration of pmn mice. The differential efficacy of peripheral and central CNTF vector administrationsmight be of interest for future studies in human motor neuron diseases.
Haase G, Pettmann B, Bordet T, Villa P, Vigne E, Schmalbruch H, Kahn A. Therapeutic benefitof ciliary neurotrophic factor in progressive motor neuronopathy depends on the route
of delivery. Ann Neurol 1999;45:296–304
Amyotrophic lateral sclerosis (ALS) and spinal muscu-lar atrophies are most often fatal diseases characterizedby degeneration of lower (a) motoneurons in spinalcord and brainstem and, in ALS, also upper (cortical)motoneurons. The therapeutic use of neurotrophic fac-tors has been suggested,1 but their subcutaneous ad-ministration as recombinant proteins was complicatedby toxicity or limited bioavailability (for review, seeHenderson2 and Yuen and Mobley3). Moreover, it re-mains unknown whether neurotrophic factors shouldbe delivered to motoneuron cell bodies in the centralnervous system (CNS) or to their axons, nerve endings,or target muscles in the body periphery.
Ciliary neurotrophic factor (CNTF), a 200-aminoacid protein member of the interleukin-6/leukemia in-hibitory factor (LIF)/cardiotrophin-1 (CT-1) neurocy-tokine family, was the first neurotrophic factor shownto support the survival of motoneurons in vitro4 and invivo.5–8 CNTF actions are mediated by a receptor
complex composed of gp130 and LIF receptor-b(LIFRb), which are also involved in LIF and CT-1 sig-nal transduction, and the CNTF specificity conferringsubunit CNTF receptor-a (CNTFRa), which is ex-pressed by upper and lower motoneurons9,10 and alsoin muscle.11
Several observations have provided a rationale for pe-ripheral CNTF delivery in motoneuronal degeneration.Recombinant CNTF protein applied to the proximalnerve stump delayed the death of axotomized mo-toneurons in newborn rats,5 and repeated subcutaneousinjections of CNTF provided therapeutic benefit towobbler mice.8,12 Intraperitoneally and subcutaneouslyimplanted cells engineered to produce CNTF increasedlife span, improved muscle performance, and reducedneuronal degeneration in pmn (progressive motor neu-ronopathy) mice.7,13 The systemic administration ofCNTF protein, however, is complicated by its serumhalf-life of less than 3 minutes14 and cytokine-like side
From *INSERM U129, Institut Cochin de Genetique Moleculaire,Paris, †INSERM U382, Institut de Biologie du Developpement deMarseille, Marseille, and ‡CNRS-PR2 URA 1301, Institut GustaveRoussy, Villejuif, France; and §Institute of Medical Physiology, Pa-num Institute, University of Copenhagen, Copenhagen, Denmark.
Received May 17, 1998, and in revised form Sep 16. Accepted forpublication Sep 17, 1998.
Address correspondence to Dr Kahn, INSERM U129, Institut Co-chin de Genetique Moleculaire, 24 Rue du Fb St Jacques, 75014Paris, France.
296 Copyright © 1999 by the American Neurological Association

effects.14,15 Clinical trials with repeated subcutaneousinjections in ALS patients failed and were complicatedby side effects such as anorexia, cough, and fever.16–18
CNTF does not efficiently cross the blood–brainbarrier,14 although it may reach the cell bodies ofa-motoneurons by retrograde axonal transport.19 Di-rect delivery of CNTF to the CNS has the advantageof reducing systemic side effects and might also influ-ence cortical motoneurons. Aebischer and colleagues20
measured elevated CNTF concentrations in the cere-brospinal fluid (CSF) of ALS patients after intrathecalimplantation of microcapsules with genetically engi-neered fibroblasts; however, data on therapeutic effi-cacy have not yet been published. In mice or rats, thisapproach is not feasible and adenovirus-mediated genetransfer into the CNS represents an interesting alterna-tive. After intracerebroventricular injection, adenovirusvectors easily spread in the CSF and can transducenumerous ependymal cells lining the ventricles,21,22
leading to secretion of recombinant proteins intothe CSF.23
We have now tested the efficacy of CNTF in pmnmice24 depending on the route of administration. Asingle high dose of an adenoviral vector coding for asecretable form of CNTF was administered in the ne-onate by intravenous, intramuscular, or intracerebro-ventricular injection. Therapeutic efficacy and potentialside effects of these delivery modes were assessedthrough analysis of weight gain, survival, and motoraxonal degeneration.
Materials and MethodsConstruction of the Adenoviral Vector AdCNTFThe first-generation (DE1, DE3) replication-deficient adeno-viral vector AdCNTF vector contains a 150-bp fragmentwith the presequence of mouse nerve growth factor(preNGF) signal sequence followed by a 1.5-kb fragmentcoding for a chimeric rat/mouse CNTF (rat exon 1–mouseintron–rat exon 2) and is derived from plasmid pChim5, akind gift of Dr Sendtner.7 Transcription of the preNGF/CNTF gene is driven by the Rous sarcoma virus (RSV) long-terminal repeat (LTR) promoter and is terminated by twopoly(A) signals in the adenoviral pIX gene. Details on vectorconstruction have been reported previously.22 Viral titerswere determined on human embryonic kidney 293 cells ac-cording to Hitt and co-workers25 and ranged from 1 3 1011
to 3 3 1012 plaque-forming units (pfu)/ml.
Fibroblast Culture and Viral InfectionHuman fibroblasts were grown to a density of 1 3 106 cells/25-cm2 dish and infected in duplicate with AdCNTF or thecontrol vector AdHEXA26 encoding the a-subunit of humanhexosaminidase A. Adenoviral infections were performed for1 hour in 500 ml of culture medium (Dulbecco’s modifiedEagle medium (MEM) with Glutamax-I; Gibco, Cergy Pon-toise, France, containing 2% [vol/vol] fetal calf serum). Afterseveral rinses, the cultures were incubated with fresh culture
medium (2 ml), which was replaced every 24 hours. Condi-tioned media were sampled from 48 to 72 hours after infec-tion, filtered (0.22 mm; Millipore, Bedford, MA), and storedat 220°C.
Ciliary Ganglionic Neuron Cultures and CellSurvival DeterminationNeuronal cultures (CG8) were obtained from 8-day-oldchicken embryos following, with some modifications, theprocedure described by Manthorpe and Varon.27 The neu-ronal cell suspension was seeded in chemically defined me-dium (CDM) (0.5 mg/ml bovine serum albumin, 5 mg/mlinsulin, 50 mg/ml transferrin, 6 ng/ml progesterone, 15mg/ml putrescine, and 5 ng/ml sodium selenite in Dulbec-co’s MEM with Glutamax-I) on polyornithine laminin–treated 96-well culture plastic plates at a final concentrationof 2,000 cells per well. Samples to be tested for their neuro-trophic activity were diluted in CDM and distributed in thecoated 96-well plates before cell seeding. Recombinant ratand human CNTF protein and polyclonal anti-rat CNTFantibody were from R&D (Abingdon, UK). Cell survival wasmeasured at 48 hours. A solution of the vital dye MTT inCDM was added to the wells, at a final concentration of 0.5mg/ml, after 47 hours of culture. One hour later, the cellswere fixed for 20 minutes by addition of an equal volume ofa 4% (vol/vol) paraformaldehyde solution. After rinsing withphosphate-buffered saline, the blue cells having metabolizedthe MTT were counted under the microscope.
Adenoviral Vector Injections in pmn MiceHomozygous Xt1 pmn/Xt1 pmn mice (“pmn mice”) were ob-tained by crossing double heterozygous Xt pmn1/Xt1 pmnmice (“Xt mice,” 28) and identified by the absence of ex-tranumerous toes. Before vector injection, pmn mice wereanesthetized briefly with ether and in hypothermia. Intra-muscular vector injections were performed at age 3 to 5 daysas described.29 For intravenous injection, 2- to 3-day-oldpmn mice were injected with 2 3 109 pfu (volume, 20 ml) ofAdCNTF into the right temporal vein. Intracerebroventric-ular injections were performed under transillumination in 1-to 2-day-old pmn mice. Using magnification lenses (Zeiss,Oberkochen, Germany), 2 3 109 pfu (2 ml) of vector wasinjected with a microsyringe (Hamilton, Bonaduz, Switzer-land) into the right cerebral ventricle. As controls, pmn miceand also Xt mice of the same litters were not treated or in-jected with AdlacZ, an adenoviral vector encoding Esche-richia coli b-galactosidase targeted to the nucleus.30 Vectorinjections and animal care were performed under L3 bio-security conditions and according to institutional guidelines.Mortality due to the AdlacZ or AdCNTF vector injectionswas low (intramuscularly, 0; intravenously, 5 of 59; and in-tracerebroventricularly, 4 of 53).
RNA and Protein AnalysesFor reverse transcription–polymerase chain reaction (RT-PCR), RNAs were isolated by the RNazol B method (Bio-probe, Montreuil, France) and 0.5 mg reverse transcribedinto cDNA. Adenoviral NGF/CNTF cDNAs were amplifiedby 35 cycles of PCR (30 seconds at 95°C, 30 seconds at55°C, and 1 minute at 72°C), using primers corresponding
Haase et al: Adenoviral CNTF Gene Transfer in pmn Mice 297
to preNGF (59-AGCTCACCTCAGTGTCTGGG) andCNTF exon 2 (59-ACCATCCACTGAGTCAAGGC). Con-trol RT-PCR reactions were performed with primers specificto b-actin. E. coli b-galactosidase enzymatic activity in pro-tein extracts was determined with a chemiluminescent assay(Tropix, Bedford, MA) according to the manufacturer’sinstructions.
Histological ExaminationsAnimals were killed with chloroform or deep ether anesthe-sia. For histochemical analysis of lacZ expression in muscleand liver, organs were dissected out, cryoprotected withCryomatrix (Shandon, Pittsburgh, PA), and frozen in liquidN2–cooled isopentane. Five- to 10-mm-thick cryosectionswere stained with 5-chromo-4-chloro-3-indolyl-b-galactoside(X-Gal) at 28°C overnight. Brains and spinal cords wereobtained after transcardiac perfusion with heparinizedphosphate-buffered saline followed by 4% (wt/vol) parafor-maldehyde, postfixed for 12 hours in paraformaldehyde andin toto stained with X-Gal. After cryoprotection with 30%sucrose for 48 hours at 4°C, they were frozen in isopentanecooled to 240°C. Serial frozen sections (20 mm) were pre-pared and counterstained with Nuclear Fast Red. For mor-phological analysis, mice were transcardially perfused withglutaraldehyde; nerve samples were postfixed with osmic te-troxide and embedded in epoxy resin. The phrenic nerve wascut close to the diaphragm. Sections, 3 mm thick, werestained with p-phenylenediamine and investigated by lightmicroscopy. The investigator (H.S.) was masked with respectto the treatment of the animals.
Statistical AnalysesDifferences between results of treated and control pmn micewere analyzed by means of Student’s t test, unpaired samples,two-tailed. The number of myelinated phrenic nerve fiberswas correlated to CNTF serum bioactivities according toFisher and Yates.
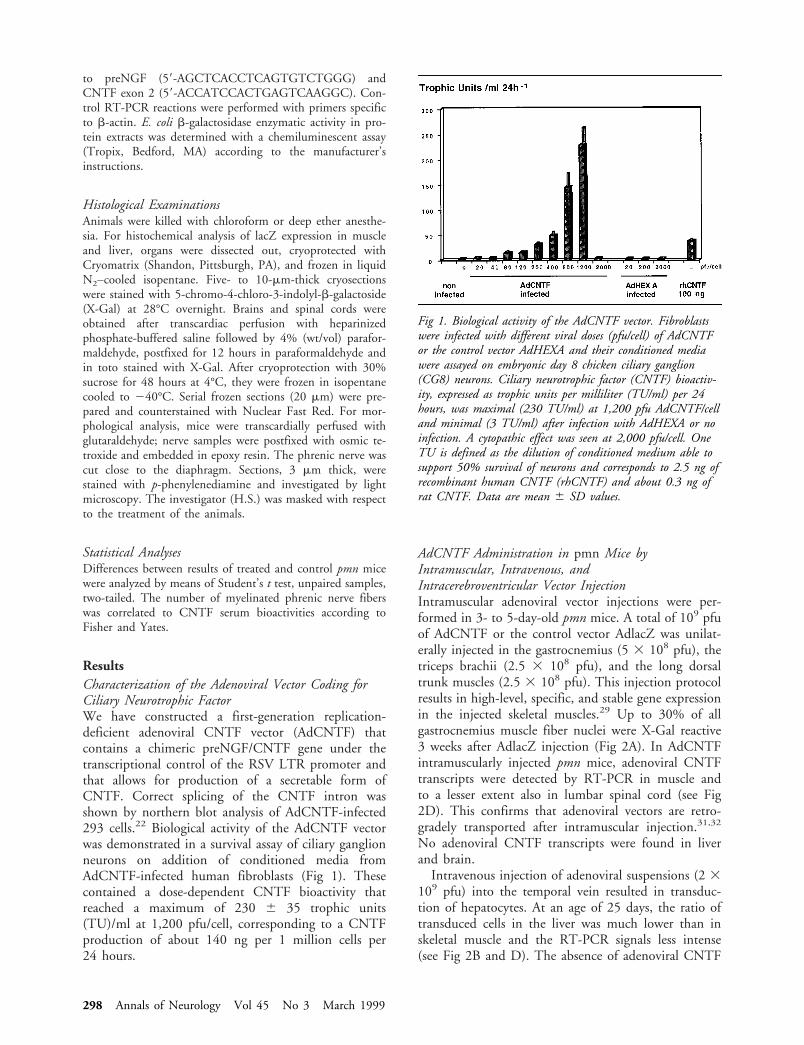
ResultsCharacterization of the Adenoviral Vector Coding forCiliary Neurotrophic FactorWe have constructed a first-generation replication-deficient adenoviral CNTF vector (AdCNTF) thatcontains a chimeric preNGF/CNTF gene under thetranscriptional control of the RSV LTR promoter andthat allows for production of a secretable form ofCNTF. Correct splicing of the CNTF intron wasshown by northern blot analysis of AdCNTF-infected293 cells.22 Biological activity of the AdCNTF vectorwas demonstrated in a survival assay of ciliary ganglionneurons on addition of conditioned media fromAdCNTF-infected human fibroblasts (Fig 1). Thesecontained a dose-dependent CNTF bioactivity thatreached a maximum of 230 6 35 trophic units(TU)/ml at 1,200 pfu/cell, corresponding to a CNTFproduction of about 140 ng per 1 million cells per24 hours.
AdCNTF Administration in pmn Mice byIntramuscular, Intravenous, andIntracerebroventricular Vector InjectionIntramuscular adenoviral vector injections were per-formed in 3- to 5-day-old pmn mice. A total of 109 pfuof AdCNTF or the control vector AdlacZ was unilat-erally injected in the gastrocnemius (5 3 108 pfu), thetriceps brachii (2.5 3 108 pfu), and the long dorsaltrunk muscles (2.5 3 108 pfu). This injection protocolresults in high-level, specific, and stable gene expressionin the injected skeletal muscles.29 Up to 30% of allgastrocnemius muscle fiber nuclei were X-Gal reactive3 weeks after AdlacZ injection (Fig 2A). In AdCNTFintramuscularly injected pmn mice, adenoviral CNTFtranscripts were detected by RT-PCR in muscle andto a lesser extent also in lumbar spinal cord (see Fig2D). This confirms that adenoviral vectors are retro-gradely transported after intramuscular injection.31,32
No adenoviral CNTF transcripts were found in liverand brain.
Intravenous injection of adenoviral suspensions (2 3109 pfu) into the temporal vein resulted in transduc-tion of hepatocytes. At an age of 25 days, the ratio oftransduced cells in the liver was much lower than inskeletal muscle and the RT-PCR signals less intense(see Fig 2B and D). The absence of adenoviral CNTF
Fig 1. Biological activity of the AdCNTF vector. Fibroblastswere infected with different viral doses (pfu/cell) of AdCNTFor the control vector AdHEXA and their conditioned mediawere assayed on embryonic day 8 chicken ciliary ganglion(CG8) neurons. Ciliary neurotrophic factor (CNTF) bioactiv-ity, expressed as trophic units per milliliter (TU/ml) per 24hours, was maximal (230 TU/ml) at 1,200 pfu AdCNTF/celland minimal (3 TU/ml) after infection with AdHEXA or noinfection. A cytopathic effect was seen at 2,000 pfu/cell. OneTU is defined as the dilution of conditioned medium able tosupport 50% survival of neurons and corresponds to 2.5 ng ofrecombinant human CNTF (rhCNTF) and about 0.3 ng ofrat CNTF. Data are mean 6 SD values.
298 Annals of Neurology Vol 45 No 3 March 1999

transcripts in skeletal muscle, brain, and spinal cord ofAdCNTF intravenously injected animals indicated therelative specificity of the intravenous route of vectoradministration for the liver.
For intracerebroventricular adenoviral vector admin-istration, 1- to 2-day old pmn mice were injected intothe right lateral ventricle with 2 3 109 pfu of vectorsuspension. At an age of 25 days, all AdlacZ-injectedpmn mice showed numerous intensely labeled ependy-mal cells in the entire cerebroventricular system (seeFig 2C); the brain parenchyma was only stained adja-cent to the injection site. Dilatation of the right ven-tricle was seen in 2 of 4 mice and was attributed to therelatively large volume injected (2 ml). In AdCNTF in-tracerebroventricularly injected pmn mice, adenoviralCNTF expression was detected by RT-PCR in brain
and lumbar spinal cord but not in liver or muscle (seeFig 2D).
The overall efficacy of the three routes of vector ad-ministration was quantitated; total b-galactosidase ac-tivities (mU/organ) in 25-day-old AdlacZ-injected micewere 1,845 6 307 mU in the injected gastrocnemiusmuscle, 386 6 101 mU in liver, and 228 6 82 mU inbrain after intravenous and intracerebroventricular in-jection, respectively (mean 6 SEM; n 5 3–4).
Weight Gain and Survival of AdCNTF-Treatedpmn MiceAll treated pmn mice were weighed twice weekly, todetect CNTF-induced weight losses.15,16,18 Comparedwith that after AdlacZ injection, the weight gain wassignificantly reduced after AdCNTF intramuscular in-
Fig 2. Adenoviral transgene expression in 25-day-old pmn (progressive motor neuronopathy) mice after different routes of vectoradministration as analyzed by X-Gal (5-chromo-4-chloro-3-indolyl-b-galactoside) histochemistry (A–C) and reverse transcription–polymerase chain reaction (RT-PCR) (D). (A) After AdlacZ intramuscular injection, numerous X-Gal–stained myofiber nuclei wereobserved in the gastrocnemius muscle. (B) Livers of AdlacZ intravenously injected pmn mice contained X-Gal–stained hepatocytes.(C) AdlacZ intracerebroventricular injection resulted in transduction of ependymal cells in the injected right lateral ventricle andalso the third and fourth ventricles (not shown). Bars: 50 mm (A); 250 mm (B); and 500 mm (C). (D) Adenoviral ciliary neuro-trophic factor (CNTF) transcripts could be detected in gastrocnemius muscles after AdCNTF intramuscular injection (left panels,lanes 2–4), livers after AdCNTF intravenous injection (middle panels, 2–4), and brains after AdCNTF intracerebroventricularinjection (right panels, 2–4). RT-PCR signals in the spinal cords of some intramuscular and intracerebroventricularly injected pmnmice (lanes 2 and 3) reflect retrograde axonal transport of vector in neurons connected to the injected muscle (intramuscular) andvector spread in the cerebrospinal fluid (intracerebroventricular), respectively. Controls: RNAs from AdCNTF-infected 293 cells (1),AdlacZ-injected (5), and noninjected (6, 7) pmn mice; no template (0). Markers M1 and M2: at 500 mg to 1 mg/lane.
Haase et al: Adenoviral CNTF Gene Transfer in pmn Mice 299

jection, and it tended to be reduced after intravenousinjection. No such effect occurred after AdCNTF in-tracerebroventricular injection (Fig 3).
Untreated pmn mice survived for 41.3 6 1.9 days(n 5 23) with a maximum of 57 days. The mean lifespan was not modified by AdlacZ intramuscular, intra-venous, or intracerebroventricular injection (see Fig 3).Peripheral AdCNTF administration enhanced meanand maximum survival; the mean life span was 52.9 64.4 days (n 5 13; p , 0.02) after intramuscular injec-tion with a maximum survival of 92 days, and it was52.7 6 3.9 days (n 5 7; p , 0.05) after intravenousinjection. At day 50, 55% (11 of 20) of AdCNTF in-tramuscularly or intravenously treated pmn mice werestill alive, in comparison with 12% (6 of 51) of the
AdlacZ-injected or nontreated animals. AdCNTF in-tracerebroventricular administration did not prolongthe mean life span (48.5 6 4.9 days; n 5 13; p 50.22), although 2 pmn mice (15%) survived for 84and 88 days.
Motoneuronal Degeneration in AdCNTF-Treatedpmn MiceMotoneuronal degeneration in pmn mice starts at theendplates and results in a massive loss of motor axonsthat is not accompanied by significant collateral sprout-ing of surviving motor axons.24 A significant loss ofanterior horn cells does not occur until end-stage dis-ease.24,29 Death of pmn mice is most probably due to
Fig 3. Weight gain and survival of pmn (progressive motor neuronopathy) mice injected by different routes with AdCNTF. New-born pmn mice received a single intramuscular, intravenous, or intracerebroventricular injection of AdCNTF (boldfaced line) orAdlacZ (dotted line) or were nontreated. (Left) Body weights are in grams and are represented as mean 6 SEM values. AdCNTFintramuscularly treated pmn mice showed a reduced weight gain, which was significantly lower than in the AdlacZ-injected littermice from day 18 on (*p , 0.05). A trend toward lower body weight was also observed after AdCNTF intravenous administra-tion but not after AdCNTF intracerebroventricular administration. (Right) Survival of pmn mice is expressed as the percentage ofinvestigated animals and as mean life span (‚). AdCNTF intramuscularly and intravenously treated pmn mice showed a prolongedmean life span when compared with pmn mice injected with AdlacZ intramuscularly (**p , 0.02), intravenously (*p , 0.05),or nontreated. After AdCNTF intracerebroventricular administration, 2 of 13 pmn mice survived for more than 50 days, but themean life span was not significantly improved (p 5 0.22, NS).
300 Annals of Neurology Vol 45 No 3 March 1999
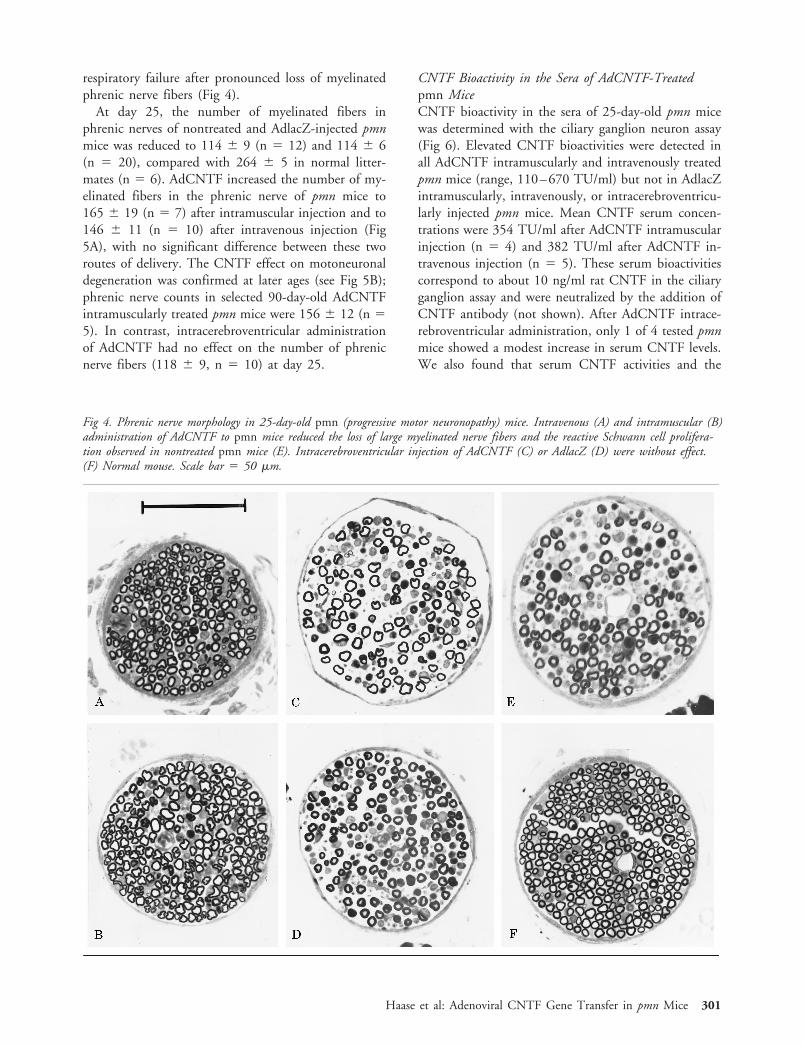
respiratory failure after pronounced loss of myelinatedphrenic nerve fibers (Fig 4).
At day 25, the number of myelinated fibers inphrenic nerves of nontreated and AdlacZ-injected pmnmice was reduced to 114 6 9 (n 5 12) and 114 6 6(n 5 20), compared with 264 6 5 in normal litter-mates (n 5 6). AdCNTF increased the number of my-elinated fibers in the phrenic nerve of pmn mice to165 6 19 (n 5 7) after intramuscular injection and to146 6 11 (n 5 10) after intravenous injection (Fig5A), with no significant difference between these tworoutes of delivery. The CNTF effect on motoneuronaldegeneration was confirmed at later ages (see Fig 5B);phrenic nerve counts in selected 90-day-old AdCNTFintramuscularly treated pmn mice were 156 6 12 (n 55). In contrast, intracerebroventricular administrationof AdCNTF had no effect on the number of phrenicnerve fibers (118 6 9, n 5 10) at day 25.
CNTF Bioactivity in the Sera of AdCNTF-Treatedpmn MiceCNTF bioactivity in the sera of 25-day-old pmn micewas determined with the ciliary ganglion neuron assay(Fig 6). Elevated CNTF bioactivities were detected inall AdCNTF intramuscularly and intravenously treatedpmn mice (range, 110–670 TU/ml) but not in AdlacZintramuscularly, intravenously, or intracerebroventricu-larly injected pmn mice. Mean CNTF serum concen-trations were 354 TU/ml after AdCNTF intramuscularinjection (n 5 4) and 382 TU/ml after AdCNTF in-travenous injection (n 5 5). These serum bioactivitiescorrespond to about 10 ng/ml rat CNTF in the ciliaryganglion assay and were neutralized by the addition ofCNTF antibody (not shown). After AdCNTF intrace-rebroventricular administration, only 1 of 4 tested pmnmice showed a modest increase in serum CNTF levels.We also found that serum CNTF activities and the
Fig 4. Phrenic nerve morphology in 25-day-old pmn (progressive motor neuronopathy) mice. Intravenous (A) and intramuscular (B)administration of AdCNTF to pmn mice reduced the loss of large myelinated nerve fibers and the reactive Schwann cell prolifera-tion observed in nontreated pmn mice (E). Intracerebroventricular injection of AdCNTF (C) or AdlacZ (D) were without effect.(F) Normal mouse. Scale bar 5 50 mm.
Haase et al: Adenoviral CNTF Gene Transfer in pmn Mice 301

number of myelinated phrenic nerve fibers were corre-lated in the 12 mice in which both parameters hadbeen determined (see Fig 6). It is interesting that the
unique AdCNTF intracerebroventricularly injectedpmn mouse with detectable circulating CNTF also dis-played an increased number of phrenic nerve fibers.
DiscussionAdenovirus vectors represent an interesting means todeliver neurotrophic factors in motoneuronal degener-ation.29,33 Indeed, they can infect all components ofthe motor unit, motoneurons,21 muscle cells,30,34 and
Fig 5. Effect of AdCNTF on the loss of myelinated phrenicnerve fibers in pmn (progressive motor neuronopathy) mice.(A) At day 25, the mean number of myelinated fibers inphrenic nerves was 264 6 5 (n 5 6) in normal mice and114 6 9 (n 5 12) in nontreated pmn mice. A significantlyhigher number of myelinated fibers was observed in pmn micetreated by AdCNTF intramuscular (i.m.) and intravenous(i.v.) administration (165 6 19, n 5 7; and 145 6 11,n 5 8, respectively) but not in AdCNTF intracerebroventricu-larly (i.c.v.) injected pmn mice (118 6 9, n 5 10). AdlacZintramuscular, intravenous, and intracerebroventricular injec-tions were without notable effect on the fiber counts in pmnmice (intramuscular: 121 6 11, n 5 6; intravenous: 111 611, n 5 8; intracerebroventricular: 113 6 7, n 5 6). Sta-tistical significance: AdCNTF intramuscular (**p , 0.02, vsnontreated; #p 5 0.074, borderline, vs AdlacZ intramuscu-lar), AdCNTF intravenous (*p , 0.05, vs nontreated and vsAdlacZ intravenous), AdCNTF intracerebroventricular (ns:nonsignificant vs nontreated and AdlacZ intracerebroventricu-lar). (B) Time course of myelinated phrenic nerve fiber degen-eration in pmn mice intramuscularly injected with AdCNTF(n), AdlacZ (▫), or noninjected (e). Normal controls (x).Data are mean 6 SEM values.
Fig 6. Ciliary neurotrophic factor (CNTF) bioactivities in seraof 25-day-old pmn (progressive motor neuronopathy) mice,measured in a ciliary neuron survival assay as trophic unitsper milliliter (TU/ml). (A) Elevated serum CNTF bioactivitieswere detected in all AdCNTF intramuscularly (i.m.) (n; n 54) and AdCNTF intravenously (i.v.) (Œ; n 5 5) treatedpmn mice, but in only 1 of 4 AdCNTF intracerebroventricu-larly (i.c.v.) injected (●) and in none of the AdlacZ-injectedpmn mice (open symbols). Detection limit 5 25 TU/ml.(B) Serum CNTF bioactivities correlated with the number ofmyelinated phrenic nerve fibers in intravenously and intracere-broventricularly treated animals (n 5 12). Note the reductionin nerve fiber degeneration in the unique AdCNTF intracere-broventricularly treated pmn mouse with detectable CNTFserum levels (arrow).
302 Annals of Neurology Vol 45 No 3 March 1999

Schwann cells,35 and also remote cells with the capac-ity to release these factors into blood or CSF.
In this study, intramuscular and intravenous injec-tions of the AdCNTF vector were performed in new-born pmn mice, to achieve long-term transgene expres-sion by muscle fibers and hepatocytes, respectively.The mean life span was increased by 25%, and the lossof phrenic nerve fibers was 20 to 35% less than inuntreated pmn mice (see also Sendtner and co-workers,7 and Sagot and associates13). CNTF bioactiv-ities at greater than 350 TU/ml of serum were reached,comparable with those in pmn mice with intraperito-neally implanted CNTF-secreting tumor cells.7 Adeno-viral CNTF expression was also detected in the lumbarspinal cord after intramuscular but not after intrave-nous injection, which suggests retrograde axonal trans-port from the injected gastrocnemius muscle to neuro-nal somata.31,32 Nevertheless, as the therapeutic benefitwas similar after both routes of delivery, it is probablydue to systemic CNTF release from infected musclefibers and hepatocytes rather than to retrograde axonaltransport of the vector.
Weight loss is an undesirable side effect of systemi-cally administered CNTF and also occurred in thisstudy after peripheral CNTF vector injection. SystemicCNTF toxicity might be reduced by combining severalfactors29,36 or by directly delivering CNTF to the CNSand the affected neurons. Our study provides the firstexample of intracerebral neurotrophic factor delivery ina mouse model of progressive motoneuronal degenera-tion. Intracerebroventricular adenovirus vector injec-tions in newborn pmn mice were well tolerated andresulted in transgene expression in numerous ependy-mal cells. The size of the mice did not allow measuringCNTF levels in the CSF, but our previous findings inrats indicate that ependymal cells produce CNTF afterAdCNTF vector injection.22 AdCNTF intracerebro-ventricular administration, however, failed to improvemean life span and motor axonal degeneration of pmnmice. It is noteworthy that the only AdCNTF intra-cerebroventricularly injected mouse with an increasednumber of phrenic nerve fibers also displayed elevatedserum CNTF levels. We speculate that in this animal,and also in the 2 AdCNTF intracerebroventricularlyinjected long-term surviving pmn mice, some CNTFvector leakage or blood–brain barrier damage had oc-curred, leading to CNTF gene expression in the bodyperiphery.
The following several hypotheses could explain thedifferential effects of central and peripheral AdCNTFadministrations: (1) CNTF from the CSF might notbecome available to motoneuron cell bodies, either dueto limited diffusion in the brain parenchyma or ineffi-cient binding. The latter is unlikely because CNTF Rareceptors have been demonstrated by immunolocaliza-tion in motoneuronal perikarya and dendrites.10 (2)
One may speculate that CNTF must reach motoneu-ron cell bodies via the axon rather than directly; aneffect of CNTF on retrograde axonal transport in pmnmice has indeed been observed.37 (3) The CNTF effectitself might be peripheral and involve axons38 or mus-cle fibers.39 (4) The lack of central CNTF efficacymight reflect a particular feature of the pmn dying-backmotor neuronopathy, starting at the endplates and thusrequiring peripheral trophic support.
In summary, adenoviral gene transfer offers uniquepossibilities to study the effects of long-term neurotro-phic factor administration in animal models of mo-toneuronal degeneration. Different neurotrophic fac-tors should also be tested in other mouse models ofmotoneuronal degeneration before conclusions with re-spect to routes of delivery in patients can be made.Defining better routes of administration for neurotro-phic factors, alone or in combination, will be crucialfor the design of new clinical trials.
This study was supported by grants from INSERM, NATO, RPRGenCell, Danish Medical Research Council, and Association Fran-caise Centre les Myopathies. Dr Haase acknowledges support froman RPR GenCell fellowship.
We thank Dr S. Nicolaidis for advice concerning intracerebroven-tricular injections, Dr P. Briand for help with initial intravenousvector injections, and Mrs Bjaerg for excellent technical help.
References1. Thoenen H, Hughes RA, Sendtner M. Trophic support of
motoneurons: physiological, pathophysiological, and therapeuticimplications. Exp Neurol 1993;124:47–55
2. Henderson CE. Neurotrophic factors as therapeutic agents inamyotrophic lateral sclerosis—potential and pitfalls. In: Serra-trice GT, Munsat TL, eds. Pathogenesis and therapy of amyo-trophic lateral sclerosis, vol 68. Philadelphia: Lippincott-Raven,1995:235–240
3. Yuen EC, Mobley WC. Therapeutic potential of neurotrophicfactors for neurological disorders. Ann Neurol 1996;40:346–354
4. Arakawa Y, Sendtner M, Thoenen H. Survival effect of ciliaryneurotrophic factor (CNTF) on chick embryonic motoneuronsin culture: comparison with other neurotrophic factors and cy-tokines. J Neurosci 1990;10:3507–3515
5. Sendtner M, Kreutzberg GW, Thoenen H. Ciliary neurotro-phic factor prevents the degeneration of motor neurons afteraxotomy. Nature 1990;345:440–441
6. Oppenheim RW, Prevette D, Yin QW, et al. Control of em-bryonic motoneuron survival in vivo by ciliary neurotrophicfactor. Science 1991;251:1616–1618
7. Sendtner M, Schmalbruch H, Stockli KA, et al. Ciliary neuro-trophic factor prevents degeneration of motor neurons in mousemutant progressive motor neuronopathy. Nature 1992;358:502–504
8. Mitsumoto H, Ikeda K, Holmlund T, et al. The effects of cil-iary neurotrophic factor on motor neuron dysfunction in wob-bler mouse motor neuron disease. Ann Neurol 1994;36:142–148
9. Ip NY, McClain J, Barrezueta NX, et al. The alpha componentof the CNTF receptor is required for signaling and defines po-
Haase et al: Adenoviral CNTF Gene Transfer in pmn Mice 303

tential CNTF targets in the adult and during development.Neuron 1993;10:89–102
10. MacLennan AJ, Vinson EN, Marks L, et al. Immunohisto-chemical localization of ciliary neurotrophic factor receptor al-pha expression in the rat nervous system. J Neurosci 1996;16:621–630
11. Davis S, Aldrich TH, Valenzuela DM, et al. The receptor forciliary neurotrophic factor. Science 1991;253:59–63
12. Ikeda K, Wong V, Holmlund TH, et al. Histometric effects ofciliary neurotrophic factor in wobbler mouse motor neuron dis-ease. Ann Neurol 1995;37:47–54
13. Sagot Y, Tan SA, Baetge E, et al. Polymer encapsulated celllines genetically engineered to release ciliary neurotrophic factorcan slow down progressive motor neuronopathy in the mouse.Eur J Neurosci 1995;7:1313–1322
14. Dittrich F, Thoenen H, Sendtner M. Ciliary neurotrophicfactor: pharmacokinetics and acute-phase response in rat. AnnNeurol 1994;35:151–163
15. Henderson JT, Seniuk NA, Richardson PM, et al. Systemic ad-ministration of ciliary neurotrophic factor induces cachexia inrodents. J Clin Invest 1994;93:2632–2638
16. Lotz B, Brooks B, Sanjak M, et al. A double-blind placebo-controlled clinical trial of subcutaneous recombinant humanciliary neurotrophic factor (rHCNTF) in amyotrophic lateralsclerosis. Neurology 1996;46:1244–1249
17. Cedarbaum JM, Chapman C, Charatan M, et al. A phase Istudy of recombinant human ciliary neurotrophic factor(rHCNTF) in patients with amyotrophic lateral sclerosis. ClinNeuropharmacol 1995;18:515–532
18. Miller RG, Petajan JH, Bryan WW, et al. A placebo-controlledtrial of recombinant human ciliary neurotrophic (rhCNTF) fac-tor in amyotrophic lateral sclerosis. Ann Neurol 1996;39:256–260
19. Curtis R, Adryan KM, Zhu Y, et al. Retrograde axonal trans-port of ciliary neurotrophic factor is increased by peripheralnerve injury. Nature 1993;365:253–255
20. Aebischer P, Schluep M, Deglon N, et al. Intrathecal deliveryof CNTF using encapsulated genetically modified xenogeneiccells in amyotrophic lateral sclerosis patients. Nat Med 1996;2:696–699
21. Akli S, Caillaud C, Vigne E, et al. Transfer of a foreign geneinto the brain using adenovirus vectors. Nat Genet 1993;3:224–228
22. Lisovoski F, Akli S, Peltekian E, et al. Phenotypic alteration ofastrocytes induced by ciliary neurotrophic factor in the intactadult brain, as revealed by adenovirus-mediated gene transfer.J Neurosci 1997;17:7228–7236
23. Bajocchi G, Feldman SH, Crystal RG, et al. Direct in vivo genetransfer to ependymal cells in the central nervous system usingrecombinant adenovirus vectors. Nat Genet 1993;3:229–234
24. Schmalbruch H, Jensen HS, Bjaerg M, et al. A new mousemutant with progressive motor neuronopathy. J NeuropatholExp Neurol 1991;50:192–204
25. Hitt M, Bett AJ, Prevec L, et al. Construction and propagationof human adenovirus vectors. In: Celis JE, ed. Cell biology—alaboratory handbook, vol 1. San Diego: Academic Press, 1994:479–490
26. Akli S, Guidotti JE, Vigne E, et al. Restoration of hexosamini-dase A activity in human Tay-Sachs fibroblasts via adenoviralvector-mediated gene transfer. Gene Ther 1996;3:769–774
27. Manthorpe M, Varon S. Use of chick embryo ciliary ganglionicneurons for the in vitro assay of neurotrophic factors. In: Sha-har A, de Vellis J, Vernadakis A, et al, eds. Dissection and tis-sue culture manual of the nervous system. New York: Alan R.Liss, 1989:317–321
28. Brunialti ALB, Poirier C, Schmalbruch H, et al. The mousemutation progressive motor neuronopathy (pmn) maps to chro-mosome 13. Genomics 1995;29:131–135
29. Haase G, Kennel P, Pettmann B, et al. Gene therapy of a mu-rine motor neuron disease using adenoviral vectors for neuro-trophic factors. Nat Med 1997;3:429–436
30. Stratford-Perricaudet LD, Makeh I, Perricaudet M, et al. Wide-spread long-term gene transfer to mouse skeletal muscles andheart. J Clin Invest 1992;90:626–630
31. Ghadge GD, Roos RP, Kang UJ, et al. CNS gene delivery byretrograde transport of recombinant replication-defective adeno-viruses. Gene Ther 1995;2:132–137
32. Finiels F, Ribotta MG, Barkats M, et al. Specific and efficientgene transfer strategy offers new potentialities for the treatmentof motor neurone diseases. Neuroreport 1995;6:2473–2478
33. Gravel C, Gotz R, Lorrain A, et al. Adenoviral gene transfer ofciliary neurotrophic factor and brain-derived neurotrophic fac-tor leads to long-term survival of axotomized motor neurons.Nat Med 1997;3:765–770
34. Quantin B, Perricaudet LD, Tajbakhsh S, et al. Adenovirus asan expression vector in muscle cells in vivo. Proc Natl Acad SciUSA 1992;89:2581–2584
35. Shy ME, Tani M, Shi Y, et al. An adenoviral vector can transferlacZ expression into Schwann cells in culture and in sciaticnerve. Ann Neurol 1995;38:429–436
36. Mitsumoto H, Ikeda K, Klinkosz B, et al. Arrest of motor neu-ron disease in wobbler mice cotreated with CNTF and BDNF.Science 1994;265:1107–1110
37. Sagot Y, Rosse T, Vejsada R, et al. Differential effects of neu-rotrophic factors on motoneuron retrograde labelling in a mu-rine model of motoneuron disease. J Neurosci 1998;18:1132–1141
38. Gurney ME, Yamamoto H, Kwon Y. Induction of motor neu-ron sprouting in vivo by ciliary neurotrophic factor and basicfibroblast growth factor. J Neurosci 1992;12:3241–3247
39. Helgren ME, Squinto SP, Davis HL, et al. Trophic effect ofciliary neurotrophic factor on denervated skeletal muscle. Cell1994;76:493–504
304 Annals of Neurology Vol 45 No 3 March 1999
Copyright © 2022 FDOKUMEN




















