
The Insect Frontal Ganglion and Stomatogastric Pattern Generator Networks

European Journal of Neuroscience, Vol. 8, pp. 521-529, 1996 0 European Neuroscience Association
Ciliary Neurotrophic Factor Constitutively Expressed in the Nervous System of Transgenic Mice Protects Embryonic Dorsal Root Ganglion Neurons from Apoptosis
Emanuela Tolosano, Miguel Angel Cutufia, Emilio Hirsch, Guglielmo Stefanuto, Samuele Voyron’ , Aldo Fasolo’, Lorenzo Silengo and Fiorella Altruda Department of Genetics, Biology and Medical Chemistry, University of Turin, Via Santena Sbis, 10126 Turin, Italy ’Department of Animal Biology, University of Turin, Italy
Keywords: CNTF, transgenic mice, apoptosis
Abstract Ciliary neurotrophic factor (CNTF) is a potent survival factor for several neuronal populations. It is expressed postnatally by Schwann cells in the peripheral nervous system and by some glial and neuronal cells in the central nervous system. We used the promoter of the neurofilament light chain gene to produce transgenic mice that express CNTF in neurons from the beginning of neuronal differentiation. These transgenic animals may represent a suitable model to identify neuronal cell types responsive to CNTF in vivo and to study the mechanism of action of this neurotrophic factor. We show that dorsal root ganglion neurons of transgenic mice expressing CNTF in neurons are protected from apoptosis during embryonic development: 40% of these cells undergo apoptosis between embryonic day 12.5 and postnatal day 5 in transgenic mice whereas 60% do so in control animals. However, protection from apoptosis does not result in an increase in the total number of neurons at the end of development. We discuss our results with regard to CNTF potentialities in vivo and the significance of programmed cell death during development.
Introduction Ciliary neurotrophic factor (CNTF) was originally identified as a target-derived factor supporting the survival of parasympathetic ciliary ganglion neurons (Barbin et al., 1984). The pleiotropic nature of CNTF action has since been widely established. In vitro CNTF acts as a survival-promoting factor for sympathetic (Blottner et al., 1989; Emsberger et al., 1989), sensory (Skaper and Varon, 1986), hippocampal (Ip et al., 1991). retinal (Hoffmann, 1988), spinal (Arakawa et al., 1990) and cortical motor (Magal et al., 1993) neurons. Moreover, CNTF acts as a phenotype-modulating factor on cultured sympathetic neurons (Saadat et al., 1989; Rao et al., 1992) and 0-2A glial precursor cells (Lillien and Raff, 1990). In vivo, CNTF is able to protect motoneurons from naturally occumng and injury-induced cell death (Sendtner et al., 1990; Oppenheim et al., 1991). In addition it can prevent axotomy-induced degeneration of septa1 (Hagg et al., 1992). thalamic (Clatterbuck et al., 1993). retinal (Mey and Thanos, 1993) and substantia nigra (Hagg and Varon, 1993) neurons.
CNTF is prevalently expressed by myelinating Schwann cells in the peripheral nervous system (PNS) and by selected glial and neuronal populations in the central nervous system (CNS) (Stockli et al., 1991; Dobrea et al., 1992; Rende et al., 1992; Henderson et al., 1994). The protein has been detected only during postnatal development and in adulthood. During embryonic development a transcript for CNTF is detectable as early as embryonic day (E) 11
in the rat (Ip et al., 1993), but the protein has not been revealed so far. Gene-targeted knockout mice that lack CNTF develop normally, but their motoneurons atrophy 4 weeks after birth, resulting in a significant reduction in muscle strength (Masu et al., 1993). Taken together, these data have suggested that CNTF is not a target-derived neurotrophic factor involved in the establishment of the nervous system, but is important for the postnatal maintenance of some neuronal populations, particularly motoneurons (Davies, 1993). Nevertheless the wide range of neuronal populations responsive in vitro to CNTF, its potency as a neurotrophic factor and the wide distribution of its receptor have led to the suggestion that it may have therapeutic effectiveness in the treatment of several neurological diseases. In this respect it is important to identify which neuronal populations are able to respond to CNTF in vivo and to know how CNTF exerts its action.
The mechanism of action of neurotrophic factors has been investi- gated in vitro; sympathetic neurons of the superior cervical ganglion maintained in culture in the presence of nerve growth factor (NGF) undergo cell death after removing the growth factor (Martin et al., 1988). NGF deprivation-induced death is morphologically apoptotic and is accompanied by DNA cleavage into oligonucleosome-sized fragments. Apoptosis can be prevented by inhibitors of RNA or protein synthesis, indicating that cell death is due to the activation of a specific genetic programme. Similar results have been obtained
Correspondence to: Fiorella Altruda, as above
Received 19 June 1995, revised I September 1995, accepted I0 October 1995

522 CNTF protection from apoptosis in transgenic mice
with proprioceptive neurons maintained in culture by brain-derived neurotrophic factor, sensory neurons by NGF and parasympathetic neurons by CNTF (Scott and Davies, 1990). Therefore it seems that neurotrophic factors promote neuronal survival not simply by stimulating metabolic processes of growth, but by inhibiting the activation of an intrinsic 'suicide programme'.
We used the transgenic approach to evaluate in vivo potentialities of CNTF. Transgenic mice expressing CNTF in neurons from the beginning of neuronal differentiation were produced. Neuron cell loss through apoptosis during the development of dorsal root ganglia (DRG) in transgenic and control animals was evaluated. Here we show that DRG neurons of the transgenic mice are prevented from undergoing apoptosis during embryonic development: the reduction of the apoptotic rate is -20%. This reduction does not result, however, in an increase in the total number of neurons at the end of development.
Materials and methods
Construction of NF/CNTF-A+ transgene Plasmid pUC19, containing a 2.3 kb EcoRI-Hind111 fragment of the human neurofilament light-chain gene promoter (pNF-L; kindly provided by J.-P. Julien), was linearized by digestion with HindIII, which cleaves at a sequence at the 3' end of the promoter, and the protruding sequences were filled in with the Klenow fragment of DNA polymerase I (Biolabs, Beverly, MA). The plasmid was ligated to a blunted 2400 bp PvuII-htuII fragment excised from the phage clone CNF8 (Negro et ul., 1991) containing the genomic coding region of the human CNTF gene without its own polyadenylation sites. This clone was designated pNF/CNTF. Subsequently the 1350 bp HindIIISulI fragment of the pNF/CNTF clone corresponding to the 3' end of the CNTF gene was substituted with a 1800 bp HindIII- Sun fragment excised from another plasmid clone (Negro et ul., 1991); this fragment contains the coding sequence in the second exon of the CNTF gene with its own polyadenylation sites. The resulting clone was called pNF/CNTF-A'.
Production of transgenic mice The 5.9 kb DNA fragment produced by EcoRI-Sun digestion of the NF/CNTF-A' clone described above was separated on a 0.8% agarose gel, extracted from the gel using Geneclean (BIO 101) and microinjected into fertilized CBNC57 mouse embryos. Injections and implantations were carried out using standard procedures. Mice were screened for the transgene using Southem hybridization analysis of DNA extracted from the tail. Ten micrograms of DNA was digested with HindIII or BumHI, separated on a 0.8% agarose gel and transferred to a nylon membrane (N-Hybond, Amersham). The filter was incubated with a random-primed [32P]dCTP-labelled probe corresponding to the insert used for microinjection. Hybridization was carried out for 16 h at 65°C in 6 X salt sodium citrate (SSC), 5 X Denhardt's solution, 0.1% SDS, 100 pg/ml salmon sperm DNA. The filter was washed successively in 2 X SSC, 0.1 % SDS for 30 min at 65°C and in 0.2 X SSC, 0.1% SDS for 30 min at 65°C and exposed overnight at -80°C with intensifying screens.
Reverse transcription-polymerase chain reaction Total RNA was extracted according to Chomczynski and Sacchi (1987). %o micrograms of RNA was reverse-transcribed for 30 min at 42°C using an 18-base poly(dT) oligomer as primer and 100 units of M-MLV reverse transcriptase (BRL). cDNAs were analysed by the polymerase chain reaction (PCR) using the following primers: (A)
5'-GCTAGCAAGGAAGATTC-3'; (B) 5'-GGAACTACGCATATTC- C-3'; (C) 5'-TCCTGCGTCTGGACCTGG-3'; (D) 5'-CCATCTCTT- GCTCGAAGT-3'. Primers A and B are located on the first exon and on the 3' non-coding region of the human CNTF gene respectively. Primers C and D are located on the mouse cytoplasmic p-actin cDNA. Amplification conditions for CNTF were as follows: the annealing temperature of the reaction was decreased by 2°C every second cycle from 60°C to a touchdown at 50"C, at which temperature 20 cycles were carried out. Denaturing was carried out at 90°C. extension at 72°C. Amplification conditions for p-actin were 90°C for 1 min, followed by two cycles at 65°C for 30 s, 72°C for 30 s and 18 cycles at 55°C for 30 s and 72°C for 30 s. The use of equal amounts of RNA for each reverse transcription (RT)-PCR was verified by comparing the expression of the housekeeping gene p-actin between all preparations. As controls, PCR amplifications were also performed without a preceding RT reaction to rule out contamination by genomic DNA. The products of amplification were analysed by electrophoresis on a 1.2% agarose gel, Southern blotting and hybridization with a 32P-labelled probe to the second exon of the human CNTF gene. Quantitation of the band intensity was performed by densitometry using a scanner connected to a personal computer (Biomed Instru- ments, Fullerton, CA).
lmmunoprecipitation and Western blot analysis Tissue samples were prepared according to Rohrer (1992) and protein concentration was determined using the Bio-Rad Protein Assay. Three hundred micrograms of extract was precleared by incubation with Ig-Sepharose beads for 1 h at 4°C and subsequently incubated with the 8E2 antibody (Cazzola etul., 1993) and protein A-Sepharose beads (Pharmacia, Uppsala, Sweden) overnight at 4°C. The precipitates were washed extensively with 20 mM Tris-HC1 (pH 7.4), 150 mM NaCI, Tris-buffered saline (TBS)/O.5% Triton X- 100 (BDH Chemicals) and resuspended in Laemli buffer. Immunoprecipitated proteins were electrophoresed on a 15% polyacrylamide gel under non-reducing conditions and electroblotted onto nitrocellulose membranes. The membranes were blocked with 5% bovine serum albumin (BSA) in TBS for 1 h at 42°C and probed with the antibody 8E2 diluted 1:400 in TBS/I% BSA overnight at 4°C. Blots were then incubated with horseradish peroxidase-conjugated anti-mouse Ig (Sigma) and developed with the chemiluminescence detection method (ECL; Amersham) according to the supplier's instructions.
Tissue processing Adult mice were anaesthetized with ketamine and perfused intra- cardially with phosphate-buffered saline (PBS) followed by 4% paraformaldehyde in 0.1 M phosphate buffer (PB), pH 7.4. Fixed tissues were dissected out and immersed for 6 h in the same fixative at 4°C. After rinsing with 7.5-15-30% sucrose in PB for 2 days, Vibratome sections (50-100 pm thick) through the coronal plane of the whole brain were collected in PB and processed for immunocyto- chemistry as described below.
Animals at different embryonic and early postnatal developmental stages were anaesthetized by hypothermia, killed by decapitation and fixed in 4% paraformaldehyde in 0.1 M PB (pH 7.4). Specimens were dehydrated, embedded in paraffin wax and cut at a thickness of 7 ym (EIO, E12, E14) or 12 pm (E16, E l 8 and postnatal). Sections were mounted onto albumin-coated glass slides.
lmmunocytochemistry Immunostaining was carried out by incubating free-floating sections on a shaking platform sequentially for: (i) 1 h at room temperature

CNTF protection from apoptosis in transgenic mice 523
with 10% normal goat serum in PBS/0.2% Triton X-100; (ii) 2 days at 4°C with the monoclonal antibody 8E2 diluted 1:400 in PBS/0.2% Triton X-100; (iii) 2 h at room temperature with biotinylated horse anti-rabbit IgG (Vectastain, Vectors Laboratories, CA) diluted 1:250 in PBS/0.2% Triton X-100; (iv) 90 min at room temperature with ABC Elite complex (Vectastain) 1:lOO in 0.05 M Tris-HCI, pH 7.6. The sections were then incubated for 10 min in 0.05 M Tris-HCI, pH 7.6, containing 0.01 5% 3,3’-diaminobenzidine and 0.002% H202. air-dried and coverslipped in DPX. Between successive steps sections were washed three times for 10 min in PBS. Specificity controls were performed by omitting the primary antibody.
In situ nick translation of tissue sections In siru nick translation (ISNT) was performed according to Gold et al. (1993). Briefly, sections were deparaffinized, washed in 10 mM Tris-HC1, 1 mM EDTA (pH 8), equilibrated in 1 X DNA polymerase I buffer (Biolabs) and incubated for 1 h at 37°C in a reaction mixture containing 1 X DNA polymerase I buffer, dNTPs [50 X digoxigenin DNA labelling mixture (Boehringer): 1 mh4 dATP, dCTP, dGTP, 0.65 mM dTTP, 0.35 mM digoxigenin-dUTP] and 2.5 Ulslide DNA polymerase I (Biolabs). The reaction was stopped in 5 mM EDTA. Sections were then washed in TBS, saturated in TBS/2% BSA/0.3% Triton X-100 and incubated for 1 h at room temperature with an alkaline phosphatase-labelled anti-digoxigenin antibody (Boehringer) diluted 1:250 in TBS/0.3% Triton X-100. After washing in TBS the colour reaction was visualized by alkaline phosphatase histochemistry using 0.3 mg/ml nitro-blue tetrazolium (Sigma) and 0.2 mg/ml 5-bromo-4-chloro-3-indolyl phosphate (Sigma). Sections were counterstained with nuclear fast red (Fluka) or 4’,6-diamidino-2-phenylindole dihydrochloride (DAPI; Fluka) and mounted in glycerol. As a positive control sections were preincubated with 100 ng/ml DNase I (Boehringer) for 10 min at 37°C. As a negative control adjacent sections were processed by the standard procedure but the DNA polymerase I was omitted.
Quantitative analysis Sections stained as described above were used for the quantitation of apoptosis. Because of the low fraction of apoptotic cells, apoptosis was expressed as an apoptotic index as reported by Wijsman et ul. (1993). This index is the ratio between the number of apoptotic nuclei and the total number of nuclei. Apoptotic nuclei were counted in every fifth section on both ganglia, on ISNT-stained sections and total nuclei on consecutive DAPI-stained sections. For each developmental stage at least three transgenic mice of each line and three control mice were analysed.
Counts of the total number of neurons of the ganglia at the end of development were performed on sections stained with cresyl violet (Fluka). Neurons having a clear nucleolus were counted in every fifth section. Counts were not corrected for split nucleoli. Measures of neuronal cell size were performed using an IBAS 20 system (Kontrod Zeiss) run on a Kontron 386 personal computer.
Differences were tested using the one-tailed Student’s t-test.
Results
Production of transgenic mice In order to obtain animals that express high levels of CNTF at the time when developmental cell death occurs, we produced transgenic mice in which the transcription of the human CNTF gene was driven by the human neurofilament light-chain (NF-L) gene promoter (Fig.
FIG. 1 . (A) Diagram showing the EcoRISan fragment of the clone pNF/ CNTF-A+ used to generate transgenic mice. The HindIII restriction site used for the screening of transgenic founder mice is also indicated. (B) Southem blot analysis of DNA from NF/CNTF-A+ transgenic founder mice. Ten micrograms of each DNA sample was digested with HindIII, separated on a 0.8% agarose gel, transferred to a nylon filter and hybridized with a ’*P- labelled pNF/CNTF-A+ probe. Lanes containing DNA from positive animals are indicated by the original number of the founder mice; lanes marked with an asterisk contain DNA from non-transgenic littermates. Arrows to the right indicate size standards: 6557, 4361 bp.
1A). NF-L promoter has already been used to obtain neuronal expression both in vitm and in vivo (Pepin et al., 1992). We used the genomic fragment of the human CNTF gene (Negro et al., 1991), which is -3.5 kb long and consists of two exons, an intron and a 3’ flanking region containing two polyadenylation signals. The first exon is 114 bp long, the second 486 bp, the intron is -1 kb and the 3’ untranslated region is -2 kb.
Mice were screened for integration of the transgene by Southern hybridization analysis on DNA extracted from the tail, digested with HindIII and probed with the insert used for microinjection. As shown in Figure lB, four founder mice were isolated that have integrated in their genome multiple copies of the NF/CNTF-A+ construct with a head-to-tail orientation, with the expected 5.9 kb fragment. They were called N1, N4, N10 and N15. Southern analysis of the same DNA digested with BumHI, which cuts several times into the construct, confirmed the correct integration of the transgene (not shown). Founder NiO, probably a chimera, did not generate transgenic offspring, though -60 offspring were screened. Three independ- ent transgenic lines were established from the other founders.
524 CNTF protection from apoptosis in transgenic mice
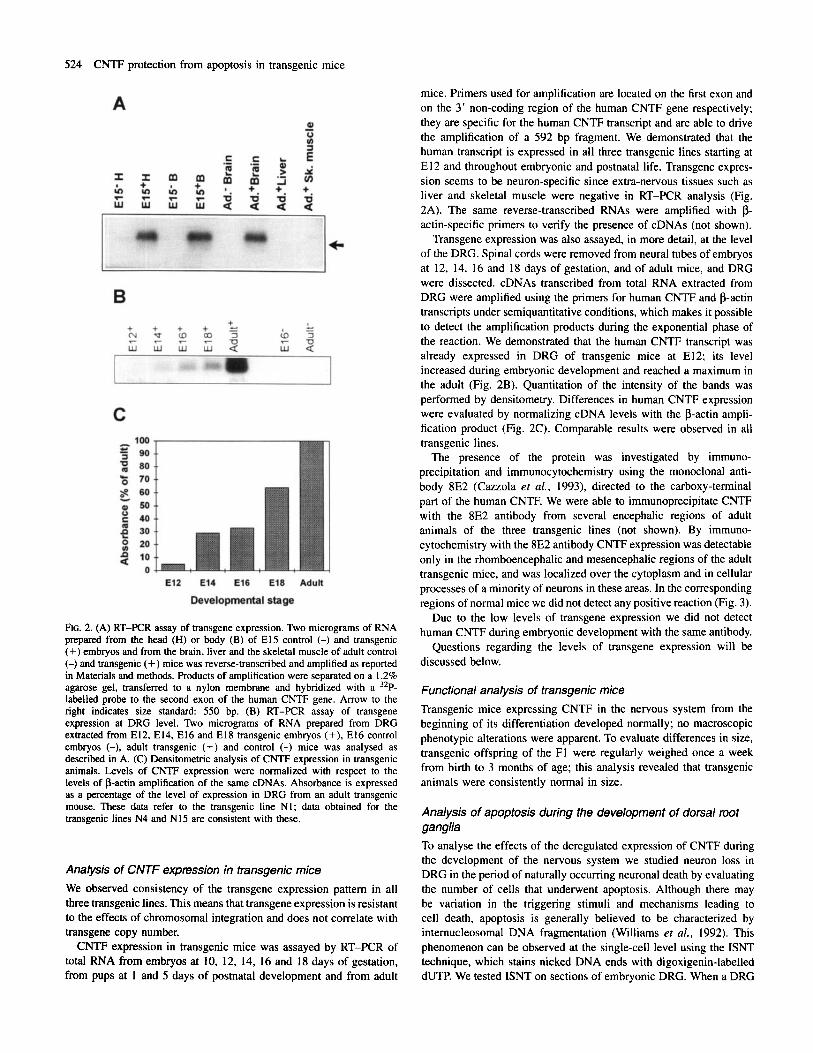
FIG. 2. (A) RT-F'CR assay of transgene expression. Two micrograms of RNA prepared from the head (H) or body (B) of El5 control (-) and transgenic (+) embryos and from the brain, liver and the skeletal muscle of adult control (-) and transgenic (+) mice was reverse-transcribed and amplified as reported in Materials and methods. hoducts of amplification were separated on a 1.2% agarose gel, transferred to a nylon membrane and hybridized with a 32P- labelled probe to the second exon of the human CNTF gene. Arrow to the right indicates size standard: 550 bp. (B) RT-PCR assay of transgene expression at DRG level. Two micrograms of RNA prepared from DRG extracted from E12, E14, El6 and El8 transgenic embryos (+), El6 control embryos (-), adult transgenic (+) and control (-) mice was analysed as described in A. (C) Densitometric analysis of CNTF expression in transgenic animals. Levels of CNTF expression were normalized with respect to the levels of p-actin amplification of the same cDNAs. Absorbance is expressed as a percentage of the level of expression in DRG from an adult transgenic mouse. These data refer to the transgenic line NI ; data obtained for the transgenic lines N4 and N15 are consistent with these.
Analysis of CNTF expression in transgenic mice We observed consistency of the transgene expression pattern in all three transgenic lines. This means that transgene expression is resistant to the effects of chromosomal integration and does not correlate with transgene copy number. CNTF expression in transgenic mice was assayed by RT-PCR of
total RNA from embryos at 10, 12, 14, 16 and 18 days of gestation, from pups at 1 and 5 days of postnatal development and from adult
mice. Primers used for amplification are located on the first exon and on the 3' non-coding region of the human CNTF gene respectively; they are specific for the human CNTF transcript and are able to drive the amplification of a 592 bp fragment. We demonstrated that the human transcript is expressed in all three transgenic lines starting at El 2 and throughout embryonic and postnatal life. Transgene expres- sion seems to be neuron-speci fic since extra-nervous tissues such as liver and skeletal muscle were negative in RT-PCR analysis (Fig. 2A). The same reverse-transcribed RNAs were amplified with p- actin-specific primers to verify the presence of cDNAs (not shown).
Transgene expression was also assayed, in more detail, at the level of the DRG. Spinal cords were removed from neural tubes of embryos at 12, 14, 16 and 18 days of gestation, and of adult mice, and DRG were dissected. cDNAs transcribed from total RNA extracted from DRG were amplified using the primers for human CNTF and p-actin transcripts under semiquantitative conditions, which makes it possible to detect the amplification products during the exponential phase of the reaction. We demonstrated that the human CNTF transcript was already expressed in DRG of transgenic mice at E12; its level increased during embryonic development and reached a maximum in the adult (Fig. 2B). Quantitation of the intensity of the bands was performed by densitometry. Differences in human CNTF expression were evaluated by normalizing cDNA levels with the p-actin ampli- fication product (Fig. 2C). Comparable results were observed in all transgenic lines.
The presence of the protein was investigated by immuno- precipitation and immunocytochemistry using the monoclonal anti- body 8E2 (Cazzola et al., 1993), directed to the carboxy-terminal part of the human CNTF. We were able to immunoprecipitate CNTF with the 8E2 antibody from several encephalic regions of adult animals of the three transgenic lines (not shown). By immuno- cytochemistry with the 8E2 antibody CNTF expression was detectable only in the rhomboencephalic and mesencephalic regions of the adult transgenic mice, and was localized over the cytoplasm and in cellular processes of a minority of neurons in these areas. In the corresponding regions of normal mice we did not detect any positive reaction (Fig. 3).
Due to the low levels of transgene expression we did not detect human CNTF during embryonic development with the same antibody.
Questions regarding the levels of transgene expression will be discussed below.
Functional analysis of transgenic mice Transgenic mice expressing CNTF in the nervous system from the beginning of its differentiation developed normally; no macroscopic phenotypic alterations were apparent. To evaluate differences in size, transgenic offspring of the F1 were regularly weighed once a week from birth to 3 months of age; this analysis revealed that transgenic animals were consistently normal in size.
Analysis of apoptosis during the development of dorsal root ganglia To analyse the effects of the deregulated expression of CNTF during the development of the nervous system we studied neuron loss in DRG in the period of naturally occurring neuronal death by evaluating the number of cells that underwent apoptosis. Although there may be variation in the triggering stimuli and mechanisms leading to cell death, apoptosis is generally believed to be characterized by internucleosomal DNA fragmentation (Williams et al., 1992). This phenomenon can be observed at the single-cell level using the ISNT technique, which stains nicked DNA ends with digoxigenin-labelled dUTP. We tested ISNT on sections of embryonic DRG. When a DRG

CNTF protection from apoptosis in transgenic mice 525
FIG. 3. Immunocytochemistry with the antibody 8E2 on the mesencephalic region of a transgenic adult mouse of the line N1 (a) and of a control (b) adult mouse. In (a) are clearly visible three labelled neurons that express the transgene at detectable levels with the 8E2 antibody. Bar, 30 pm.
section was pretreated with DNase I to create artificial nicks, all nuclei in the ganglion were stained, thus proving that each nucleus is capable of being stained. Moreover, by comparing consecutive sections stained with ISNT and cresyl violet respectively we demon- strated that nuclei labelled by ISNT have morphological characteristics of dying nuclei (not shown). ISNT analysis was performed from El0 to P5, the period of naturally occurring neuron death; DRG of the lumbar region were analysed. The same analysis was performed on transgenic embryos of the three independent lines. Figure 4 shows a DRG from transgenic and control mice at E13. At every stage of development at least three transgenic mice of each line and three normal mice were analysed and the apoptotic index was calculated. The first evidence of apoptotic nuclei occurred at E12; the major proportion of them could be detected between E l 4 and E16. At E l 8 few apoptotic nuclei were present. At PI and P5 no labelled cells
FIG. 4. ISNT on a lumbar DRG section from (a) a normal mouse embryo and (b) a transgenic mouse embryo. Both embryos were at stage E13. Bar, 30 pm.
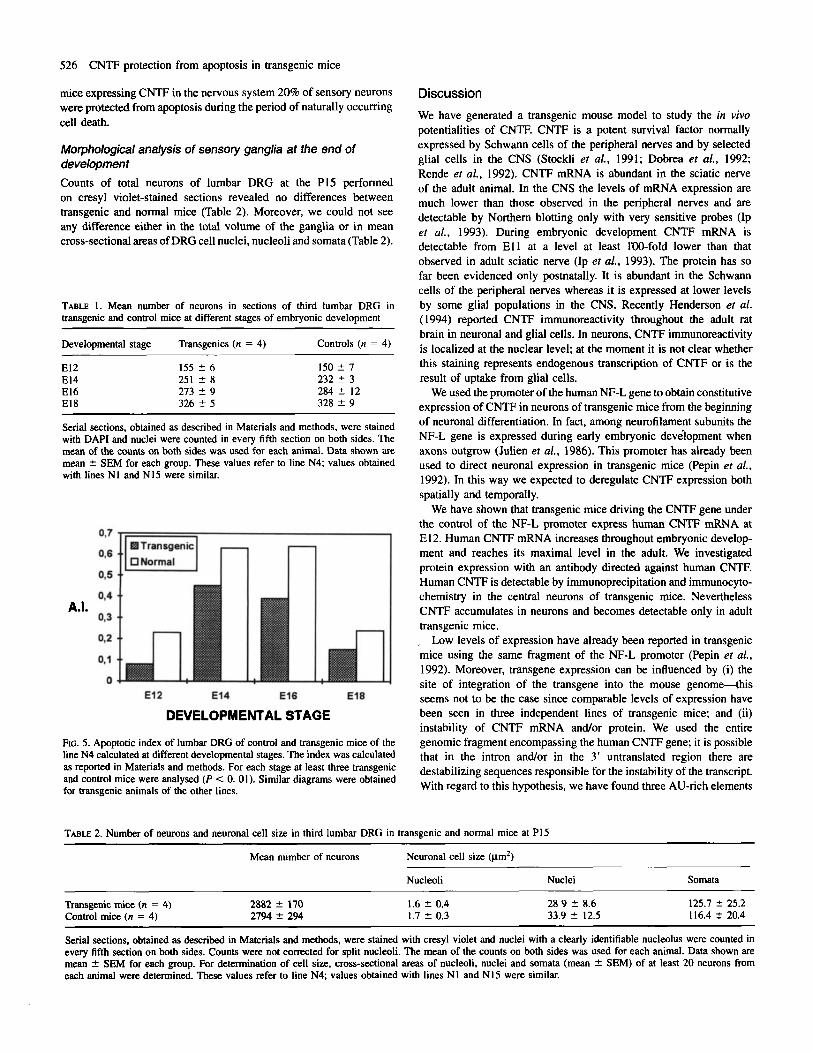
were observed. The mean number of total cells at every stage was similar in transgenic and normal mice (Table l), as was the time course of apoptosis. On the contrary, the apoptotic index was significantly lower in transgenic mice compared to normal ones (Fig. 5). To calculate the proportion of cells that died @tween E l 2 and El 8 we considered a clearance time for apoptotic cells of -1 h once a pyknotic cell was seen, as reported by Barres et al. (1992). In this way we calculated that in normal mice between E l 2 and El8 -60% of the cells died through apoptosis, whereas in the three lines of transgenic mice this value fell to 40%. In conclusion, in transgenic
526 CNTF protection from apoptosis in transgenic mice
mice expressing CNTF in the nervous system 20% of sensory neurons were protected from apoptosis during the period of naturally occurring cell death.
Morphological analysis of sensory ganglia at the end of development Counts of total neurons of lumbar DRG at the P15 performed on cresyl violet-stained sections revealed no differences between transgenic and normal mice (Table 2). Moreover, we could not see any difference either in the total volume of the ganglia or in mean cross-sectional areas of DRG cell nuclei, nucleoli and somata (Table 2).
TABLE 1. Mean number of neurons in sections of third lumbar DRG in transgenic and control mice at different stages of embryonic development
Developmental stage Transgenics (n = 4) Controls (n = 4) ~
El2 155 2 6 150 5 7 El4 251 ? 8 232 ? 3 El6 273 5 9 284 5 12 El8 326 5 5 328 5 9
Serial sections, obtained as described in Materials and methods, were stained with DAPI and nuclei were counted in every fifth section on both sides. The mean of the counts on both sides was used for each animal. Data shown are mean 2 SEM for each group. These values refer to line N4; values obtained with lines N1 and N15 were similar.
A.I.
DEVELOPMENTAL STAGE
FIG. 5. Apoptotic index of lumbar DRG of control and transgenic mice of the line N4 calculated at different developmental stages. The index was calculated as reported in Materials and methods. For each stage at least three transgenic and control mice were analysed (P < 0. 01). Similar diagrams were obtained for transgenic animals of the other lines.
Discussion We have generated a transgenic mouse model to study the in vivo potentialities of CNTF. CNTF is a potent survival factor normally expressed by Schwann cells of the peripheral nerves and by selected glial cells in the CNS (Stockli er af., 1991; Dobrea et al., 1992; Rende et af., 1992). CNTF mRNA is abundant in the sciatic nerve of the adult animal. In the CNS the levels of mRNA expression are much lower than those observed in the peripheral nerves and are detectable by Northern blotting only with very sensitive probes (Ip er af., 1993). During embryonic development CNTF mRNA is detectable from E l l at a level at least IW-fold lower than that observed in adult sciatic nerve (Ip et al., 1993). The protein has so far been evidenced only postnatally. It is abundant in the Schwann cells of the peripheral nerves whereas it is expressed at lower levels by some glial populations in the CNS. Recently Henderson et af. (1994) reported CNTF immunoreactivity throughout the adult rat brain in neuronal and glial cells. In neurons, CNTF immunoreactivity is localized at the nuclear level; at the moment it is not clear whether this staining represents endogenous transcription of CNTF or is the result of uptake from glial cells.
We used the promoter of the human NF-L gene to obtain constitutive expression of CNTF in neurons of transgenic mice from the beginning of neuronal differentiation. In fact, among neurofilament subunits the NF-L gene is expressed during early embryonic devilopment when axons outgrow (Julien et af., 1986). This promoter has already been used to direct neuronal expression in transgenic mice (Pepin et al., 1992). In this way we expected to deregulate CNTF expression both spatially and temporally.
We have shown that transgenic mice driving the CNTF gene under the control of the NF-L promoter express human CNTF mRNA at E12. Human CNTF mRNA increases throughout embryonic develop- ment and reaches its maximal level in the adult. We investigated protein expression with an antibody directed against human CNTF. Human CNTF is detectable by immunoprecipitation and immunocyto- chemistry in the central neurons of transgenic mice. Nevertheless CNTF accumulates in neurons and becomes detectable only in adult transgenic mice. , Low levels of expression have already been reported in transgenic mice using the same fragment of the NF-L promoter (Pepin er al., 1992). Moreover, transgene expression can be influenced by (i) the site of integration of the transgene into the mouse genome-this seems not to be the case since comparable levels of expression have been seen in three independent lines of transgenic mice; and (ii) instability of CNTF mRNA and/or protein. We used the entire genomic fragment encompassing the human CNTF gene; it is possible that in the intron andor in the 3' untranslated region there are destabilizing sequences responsible for the instability of the transcript. With regard to this hypothesis, we have found three AU-rich elements
TABLE 2. Number of neurons and neuronal cell size in third lumbar DRG in transgenic and normal mice at PI5
Mean number of neurons Neuronal cell size (p2)
Nucleoli Nuclei Somata
Transgenic mice (n = 4) 2882 2 170 1.6 2 0.4 28 9 2 8.6 125.7 2 25.2 Control mice (n = 4) 2794 2 294 1.7 ? 0.3 33.9 2 12.5 11 6.4 2 20.4
~
Serial sections. obtained as described in Materials and methods, were stained with cresyl violet and nuclei with a clearly identifiable nucleolus were counted in every fifth section on both sides. Counts were not corrected for split nucleoli. The mean of the counts on both sides was used for each animal. Data shown are mean 2 SEM for each group. For determination of cell size, cross-sectional areas of nucleoli, nuclei and somata (mean ? SEM) of at least 20 neurons from each animal were determined. These values refer to line N4; values obtained with lines N1 and N15 were similar.

CNTF protection from apoptosis in transgenic mice 527
in the 3‘ untranslated region of the human gene. It has been reported that these AU-rich elements are involved in mRNA destabilization (Sachs, 1993). The instability of CNTF mRNA could account for the low amounts of CNTF protein and consequently for the difficulty in localizing it in transgenic mice.
The same observations may be made concerning endogenous CNTF expression. In fact, despite the high number of antibodies that have so far been produced against recombinant CNTF and different peptides there are no conclusive data on the immunocytochemical localization of this protein in the CNS.
CNTF is an extremely potent neurotrophic factor (EC5o = 2 pM). This is true also for our transgenic mice, in which we demonstrated a biological effect, albeit a low level of transgene expression. We used transgenic mice to identify neuronal systems responsive in vivo to CNTF. Previously reported studies have demonstrated that CNTF supports the survival of cultured parasympathetic (Barbin et af., 1984), sympathetic (Blottner et al., 1989; Ernsberger et al., 1989), sensory (Skaper and Varon, 1986), hippocampal (Ip et al., 1991), retinal (Hoffmann, 1988) and motor neurons (Arakawa et al., 1990; Magal et af., 1993). In vivo CNTF administration after lesion is able to protect motor (Sendtner et al., 1990). thalamic (Hagg et af., 1992), septa1 (Clatterbuck ef al., 1993). retinal (Mey and Thanos, 1993) and substantia nigra neurons (Hagg and Varon, 1993) from degeneration. CNTF is also able to protect motoneurons from developmental cell death in the chick embryo (Oppenheim et af., 1991). The mechanism of action of CNTF has been investigated in cultures of parasympathetic neurons; in this system Scott and Davies (1990) have demonstrated that CNTF promotes neuron survival by protecting neurons from apoptosis.
We started the analysis of transgenic mice by studying neuron loss during the development of the DRG. We decided to analyse DRG neurons because (i) embryonic DRG neurons are maintained in culture by CNTF (Skaper and Varon, 1986); (ii) DRG neurons express CNTF receptor a from the beginning of neuron differentiation (Ip et af., 1993); and (iii) Schwann cells of the sensory fibres running through the neuronal cells of DRG express CNTF postnatally (Dobrea et af., 1992). So DRG represent a potential target for NFKNTF-A+ transgene action and consequently a good model in which to test the hypothesis that CNTF is able to protect in vivo neurons from apoptosis. We demonstrated that human CNTF is already expressed in DRG of transgenic mice at E l2 and increases during embryonic development. We used ISNT (Gold et af., 1993) to label apoptotic nuclei; this technique has already been successfully used to study neuron loss during development (Wood et al., 1993). Moreover, ISNT has also been applied to the study of sensory ganglia (Migheli et al., 1994).
Our results indicate that in transgenic mice expressing CNTF in neurons there is a reduction of -20% in apoptosis: we counted -40% of apoptotic nuclei in transgenic mice against 60% in normal mice. A cell loss of -60% in normal mice is in agreement with previously reported data on developmental cell death (Lance-Jones, 1982). The percentage of reduction (20%) correlates well with the level of protection due to CNTF in other systems (Sendtner et al., 1992). Nevertheless, both during embryogenesis and at the end of develop- ment the total number of neurons in control and transgenic mice is the same; the reduction in the apoptotic rate does not result in an increase in the total number of neurons. This can be due to the occurrence of cell death via different mechanisms insensitive to CNTF, or to differences in neurogenesis. With regard to the first of these mechanisms, Clarke (1990) described three types of cell death in the nervous system at the ultrastructural level: apoptotic, auto- phagic and non-lysosomal vesciculate. Only apoptotic cell death
would be characterized by DNA fragmentation. Recently Schwartz et af. (1994) suggested, on the basis of morphological and molecular studies, that more than one cell death mechanism is used during development. CNTF might protect sensory neurons from apoptosis but have no effect against other modalities of death. With regard to the mechanism involving differences in neurogenesis, this process continues in mice approximately until E l 5 (Lawson et af., 1974), when apoptosis also occurs. The timing of these developmental events presents the possibility that neuroblasts in this region are stimulated to divide by signals derived from dying cells. The possibility that dying cells control the mitosis of precursor cells has already been hypothesized with regard to neurogenesis and cell death in the dentate gyrus (Could and McEwen, 1993). Moreover, Homma ef af. (1994) have recently reported that debris from dead cells in the chick spinal cord are phagocytosed by healthy neighbouring neuroepithelial cells, suggesting that this could be a mechanism of communication between dead cells and developing ones. On the other hand, CNTF may have an antimitotic effect on sensory neuroblasts similar to the effect on sympathetic neurons described by Ernsberger et d.,( 1989).
Oppenheim et af. (1991) have previously shown that CNTF administered to the chick embryo during the period of naturally occurring cell death is able to protect motoneurons from death, but not sensory, sympathetic, nodose and ciliary neurons. This work is only apparently in contrast with ours. Oppenheim et af. counted neuronal nuclei at the end of the period of naturally occurring cell death and demonstrated no differences from controls. We obtained the same result. However, even if the final result is the same, we showed an effect of CNTF on a specific mechanism of cell death. Our preliminary results indicate that with ISNT it is possible to label only sporadic nuclei in the ventral horn of the spinal cord. This could be due to the existence of another modality of cell death different from apoptosis, or alternatively to an apoptotic process not characterized by DNA internucleosomal fragmentation.
How can CNTF produced by neurons exert its action on the same producing cells? This question arises naturally since CNTF neither has a signal peptide (Lin et af., 1989; Stockli el af., 1989; Lam et al., 1991; Negro et al., 1991) nor is it released in the culture medium of producing cells (Lin el af., 1989; Stockli et af., 1989; Negro et af., 1991). Nevertheless, it has been hypothesized that CNTF can be released through an unconventional mechanism of secretion or altern- atively through debris of the plasma membrane due to ageing or damage. We think that the same mechanisms can be hypothesized for the secretion of CNTF by neurons of transgenic mice. Recently Edwards and Tolkovsky (1994) reported that sympathetic neurons undergoing apoptosis after NGF withdrawal did not fragment into apoptotic bodies, but instead underwent plasma membrane break- down. If this is the case for DRG neurons one could think that CNTF released by dying cells acts on neighbouring cells. The issue of secretion of CNTF may be addressed by performing cultures of neurons from transgenic mice.
In conclusion, transgenic mice constitutively expressing CNTF in neurons represent a good model to test the in vivo potentialities of CNTF. Our results show that CNTF plays a positive role in the survival of DRG neurons by protecting them from apoptosis.
Acknowledgements We thank Dr Laura Salvetti for helping in the morphological analysis and Immacolata Carfora and Lauredana Cavarretta for technical assistance. This work was supported by grants from Telethon and CNR Progetti Finalizzati ‘Ingegneria Genetica’ e ‘Biotecnologie e Biostrumentazioni’.

528 CNTF protection from apoptosis in transgenic mice
Abbreviations embryo: a possible means of early phenotypic selection. J. Comp. Neurol., 345, 377-395.
BSA CNS CNTF DAPI DRG EDTA ISNT NF-L PB PBS FCR PNS RT-PCR TBS
bovine serum albumin central nervous system ciliary neurotrophic factor 4',6-diamidino-2-phenylindole dihydrochloride dorsal root ganglion (ganglia) ethylenediamine tetraacetic acid in situ nick translation neurofilament light chain phosphate buffer phosphate-buffered saline polymerase chain reaction peripheral nervous system reverse transcription-polymerase chain reaction Tris-buffered saline
References Arakawa, Y., Sendtner, M. and Thoenen, H. (1990) Survival effect of ciliary
neurotrophic factor (CNTF) on chick embryonic motoneurons in culture: comparison with other neurotrophic factors and cytokines. J. Neurosci., 10,
Barbin, G., Manthorpe, M. and Varon, S. J. (1984) Purification of the chick eye ciliary neuronotrophic factor. J. Neurochem., 43, 1468-1478.
Barres, B. A., Hart, I. K., Coles, H. S. R., Burne, J. F., Voyvodic, J. T., Richardson, W. D. and Raff, M. C. (1992) Cell death and control of cell survival in the oligodendrocyte lineage. Cell, 70, 3146.
Blottner, D., Bruggemann, W. and Unsicker, K. (1989) Ciliary neurotrophic factor supports target-derived preganglionic sympathetic spinal cord neurons. Neurosci. Lett., 105, 316-320.
Cazzola, F., Battiston, S., Fabris, M., Negro, A,, Soranzo, C., Corona, G. and Callegaro, L. (1993) Production and characterization of monoclonal antibodies to human ciliary neurotrophic factor with defined epitope recognition. Hybridoma, 12, 259-270.
Chomczynski, P. and Sacchi, N. (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenokhloroform extraction. Anal. Biochem., 162, 156-159.
Clarke, P. G. H. (1990) Developmental cell death: morphological diversity and multiple mechanisms. Anat. Embryol., 181, 195-21 3.
Clatterbuck, R. E., Price, D. L. and Koliatsos, V. E. (1993) Ciliary neurotrophic factor prevents retrograde neuronal death in the adult central nervous system. Proc. Natl Acad. Sci. USA, 90, 2222-2226.
Davies, A. M. (1993) Promoting motor neuron survival. Cur,: Opin. Neurobiol.,
Dobrea, G. M., Unnerstall, J. R. and Rao, M. S. (1992) The expression of CNTF message and immunoreactivity in the central and peripheral nervous system. Dev. Brain Res., 66, 209-219.
Edwards, S. N. and Tolkovsky, A. M. (1994) Characterization of apoptosis in cultured rat sympathetic neurons after nerve growth factor withdrawal. J. Cell Biol., 124, 537-546.
Ernsberger, U., Sendtner, M. and Rohrer, H. (1989) Proliferation and differentiation of embryonic chick sympathetic neurons: effects of ciliary neurotrophic factor. Neuron, 2, 1275-1284.
Gold, R., Schmied, M., Rothe, G., Zischler, H., Breitschopf, H., Wekerle, H. and Lassmann, H. (1993) Detection of DNA fragmentation in apoptosis: application of in situ nick translation to cell culture systems and tissue sections. J . Histochem. Cytochern., 41, 1023-1030.
Gould, E. and McEwen, B. S. (1993) Neuronal birth and death. Cur,: Opin. Neurobiol., 3, 676-682.
Hagg, T. and Varon, S. (1993) Ciliary neurotrophic factor (CNTF) prevents degeneration of adult rat substantia nigra dopaminergic neurons in vivo. Proc. Natl Acad. Sci. LISA, 90,6315-6319.
Hagg, T., Quon, D., Higaki, J. and Varon, S. (1992) Ciliary neurotrophic factor prevents neuronal degeneration and promotes low affinity NGF receptor expression in the adult rat CNS. Neuron, 8, 145-158.
Henderson, J. T., Seniuk, N. A. and Roder, J. C. (1994) Localization of CNTF immunoreactivity to neurons and astroglia in the CNS. Mol. Brain Res., 22, 151-165.
Hoffmann, H. D. (1988) Ciliary neuronotrophic factor stimulates choline acetyltransferase activity in cultured chicken retina neurons. J. Neurochem., 51, 109-113.
Homma, S., Yaginuma, H. and Oppenheim, R. W. (1994) Programmed cell death during the earliest stages of spinal cord development in the chick
3507-35 15.
3, 879-881.
Ip, N. Y., Li, Y., van de Stadt, I., Panayotatos, N., Alderson, R. F. and Lindsay, R. M. (1991) Ciliary neurotrophic factor enhances neuronal survival in embryonic rat hippocampal cultures. J. Neurosci., 11, 3124-3 134.
Ip, N. Y., McClain, J., Barrezueta, N. X., Aldrich, T. H., Pan, L., Li, Y., Wiegand, S. J., Frieman, B., Davis, S. and Yancopoulos, G. D. (1993) The a component of the CNTF receptor is required for signaling and defines potential CNTF targets in the adult and during development. Neuron, 10,
Julien, J.-P., Meijer, D., Hurst, J. and Grosveld, F. (1986) Cloning and developmental expression of the murine neurofilament gene family. Mol. Brain Res., 1, 243-250.
Lam, A., Fuller, F., Miller, J., Kloss, J., Manthorpe, M., Varon, S. and Cordell, B. (1991) Sequence and structural organization of the human gene encoding ciliary neurotrophic factor. Gene, 102, 27 1-276.
Lance-Jones, C. (1982) Motoneuron cell death in the developing lumbar spinal cord of the mouse. Dev. Brain Rex, 4, 473479.
Lawson, S. N., Caddy, K. W. T. and Biscoe, T. J. (1974) Development of rat dorsal root ganglion neurones. Studies of cell birthdays and changes in mean cell diameter. Cell Tissue Res., 153, 399413.
Lillien, L. E. and Raff, M. C. (1990) Differentiation signals in the CNS: type- 2 astrocyte development in vitro as a model system. Neuron, 5, 11 1-1 19.
Lin, L.-F. H., Mismer, D., Lile, J. D., Armes, L. G., Butler, E. T., 111, Vannice, J. L. and Collins, F. (1989) Purification, cloning and expression of ciliary neurotrophic factor (CNTF). Science, 246, 1023-1025.
Magal, E., Louis, J.-C., Oudega, M. and Varon, S. (1993) CNTF promotes the survival of neonatal rat corticospinal neurons in vitm. NeuroReport, 4,
Martin, D. P., Schmidt, R. E., DiStefano, P. S., Lowry, 0. H., Carter, J. G. and Johnson, E. M., Jr (1988) Inhibitors of protein synthesis and RNA synthesis prevent neuronal death caused by nerve growth factor deprivation. J . Cell Biol., 106, 829-844.
Maw, Y., Wolf, E., Holtmann, B., Sendtner, M., Brem, G . and Thoenen, H. (1993) Disruption of the CNTF gene results in motor neuron degeneration. Nature, 365, 27-32.
Mey, J. and Thanos, S. (1993) Intravitreal injections of neurotrophic factors support the survival of axotomized retinal ganglion cells in adult rats in vivo. Brain Res., 602, 304-317.
Migheli, A., Cavalla, P., Marino, S. and Schiffer, D. (1994) A study of apoptosis in normal and pathological nervous tissue after in situ end- labeling of DNA strand breaks. J. Neuropathol. Exp. Neurol.. 53, 606-616.
Negro, A,, Tolosano, E., Skaper, S. D., Martini, I., Callegaro, L., Silengo, L., Fiorini, F. and Altruda, F. (1991) Cloning and expression of human ciliary neurotrophic factor. Eu,: J. Biochem., 201, 289-294.
Oppenheim, R. W., Prevette, D., Qin-Wei, Y., Collins, F. and McDonald, J. (1991) Control of embryonic motoneuron survival in vivo by ciliary neurotrophic factor. Science, 251, 1616-1618.
Pepin, M.-C., Pothier, F. and Barden. N. (1992) Impaired type I1 glucocorticoid receptor function in mice bearing antisense RNA transgene. Nature, 355, 725-128.
Rao, M. S., Tyrrell, S., Landis, S. C. and Patterson, P. H. (1992) Effects of ciliary neurotrophic factor (CNTF) and depolarization on neuropeptide expression in cultured sympathetic neurons. Dev. Biol., 150, 28 1-293.
Rende, M., Muir, D., Rouslahti, E., Hagg, T., Varon, S. and Manthorpe, M. (1992) Immunolocalization of ciliary neurotrophic factor in adult rat sciatic nerve. Glia, 5, 25-32.
Rohrer, H. (1992) Cholinergic neuronal differentiation factors: evidence for the presence of both CNTF-like and non-CNTF-like factors in developing rat footpad. Development, 114, 689-698.
Saadat, S., Sendtner, M. and Rohrer, H. (1989) Ciliary neurotrophic factor induces cholinergic differentiation of rat sympathetic neurons in culture. J . Cell Biol., 108, 1807-1816.
Sachs, A. B. (1993) Messenger RNA degradation in eukaryotes. Cell, 74, 413421.
Schwartz, L. M., Smith, S. W., Jones, M. E. E. and Osborne, B. A. (1994) Do all programmed cell deaths occur via apoptosis? Proc. Natl Acad. Sci. USA, 90,980-984.
Scott, S. A. and Davies, A. M. (1990) Inhibition of protein synthesis prevents cell death in sensory and parasympathetic neurons deprived of neurotrophic factor in vitro. J. Neurobiol., 21, 6 3 W O .
Sendtner, M., Kreutzberg, G. W. and Thoenen, H. (1990) Ciliary neurotrophic factor prevents the degeneration of motor neurons after axotomy. Nature, 345,440441.
Sendtner, M., Schmalbruch, H., Stockli, K. A,, Carroll, P., Kreutzberg, G. W.
89-102.
779-182.

CNTF protection from apoptosis in transgenic mice 529
and Thoenen, H. (1992) Ciliary neurotrophic factor prevents degeneration of motor neurons in mouse mutant progressive motor neuronopathy. Nature, 358, 502-504.
Skaper, S. D. and Varon, S . (1986) Age-dependent control of dorsal root ganglion neuron survival by macromolecular and low-molecular weight trophic agents and substratum-bound laminins. Brain Rex, 389, 39-46.
Stockli, K. A,, Lottspeich, F., Sendtner, M., Masiakowski, P., Carroll, P., Gotz, R., Lindholm, D. and Thoenen, H. (1989) Molecular cloning, expression and regional distribution of rat ciliary neurotrophic factor. Nature, 342, 920-923.
Stockli, K. A,, Lillien, L. E., Naher-No&, M., Breitfeld, G., Hughes, R. A,, Raff, M. C., Thoenen, H. and Sendtner, M. (1991) Regional distribution,
developmental changes and cellular localization of CNTF-mRNA and protein in the rat brain. J. Cell Biol., 115, 447459.
Wijsman, J. H., Jonker, R. R., Keijzer, R., Van de Velde, C. J. H., Cornelisse, C. J. and Van Dierendonck, J. H. (1993) A new method to detect apoptosis in paraffin sections: in situ end labeling of fragmented DNA. J. Hisrochem. Cytochem., 41, 7-12.
Williams, G. T., Smith, C. A., McCarthy, N. J. and Grimes, E. A. (1992) Apoptosis: final control point in cell biology. Trends in Cell Biology, 2 ,
Wood, K. A,, DiPasquale, B. and Youle, R. J. (1993) In siru labeling of granule cells for apoptosis-associated DNA fragmentation reveals different mechanisms of cell loss in developing cerebellum. Neuron, 11, 621-632.
263-267.
Copyright © 2022 FDOKUMEN




















