
Mycorrhizas improve nitrogen nutrition of Trifolium repens after 8 yr of selection under elevated...
12
www.newphytologist.org 531 Research Blackwell Publishing, Ltd. Mycorrhizas improve nitrogen nutrition of Trifolium repens after 8 yr of selection under elevated atmospheric CO 2 partial pressure Hannes Gamper 1,3 , Ueli A. Hartwig 2 and Adrian Leuchtmann 1 1 Swiss Federal Institute of Technology (ETH Zürich), Geobotanical Institute, Zollikerstrasse 107, CH-8008 Zürich, Switzerland; 2 Academia Engiadina, Quadratscha 18, CH-7503 Samedan, Switzerland; 3 Present address: Department of Biology, PO Box 373, University of York, Heslington, York, YO10 5YW, UK Summary • Altered environmental conditions may change populations of arbuscular mycor- rhizal fungi and thereby affect mycorrhizal functioning. We investigated whether 8 yr of free-air CO 2 enrichment has selected fungi that differently influence the nutrition and growth of host plants. • In a controlled pot experiment, two sets of seven randomly picked single spore isolates, originating from field plots of elevated (60 Pa) or ambient CO 2 partial pres- sure (pCO 2 ), were inoculated on nodulated Trifolium repens (white clover) plants. Fungal isolates belonged to the Glomus claroideum or Glomus intraradices species complex, and host plants were clonal micropropagates derived from nine genets. • Total nitrogen (N) concentration was increased in leaves of plants inoculated with fungal isolates from elevated-pCO 2 plots. These isolates took up nearly twice as much N from the soil as isolates from ambient-pCO 2 plots and showed much greater stim- ulation of biological N 2 fixation. The morpho-species identity of isolates had a more pronounced effect on N 2 fixation and on root length colonized than isolate identity. • We conclude that rising atmospheric pCO 2 may select for fungal strains that will help their host plants to meet increased N demands. Key words: arbuscular mycorrhizal fungi (AMF), global environmental change, Glomus claroideum, Glomus intraradices, nitrogen acquisition, selection, symbiotic functioning, Trifolium repens (white clover). New Phytologist (2005) 167 : 531–542 © New Phytologist (2005) doi : 10.1111/j.1469-8137.2005.01440.x Author for correspondence: Hannes Gamper Tel: +44 (0) 1904 328553 Fax: +44 (0) 1904 328510 Email: [email protected] Received: 7 January 2005 Accepted: 8 March 2005 Introduction Arbuscular mycorrhizal fungi (AMF, Glomeromycota) are a ubiquitous group of soil fungi closely involved in the cycling of nutrients between plants and soil (Jakobsen et al., 1992; Smith & Read, 1997). They depend exclusively on organic carbon (C) sources provided by their host plants and thus are likely to respond to environmentally induced alteration in plant photosynthesis. Responses of mycorrhizas to conditions of elevated atmospheric CO 2 partial pressure (pCO 2 ) are variable and diverse (Staddon & Fitter, 1998; Rillig & Allen, 1999; Fitter et al., 2000; Rillig et al., 2002; Treseder, 2004). The most notable observed response to elevated pCO 2 is an enlargement of the extraradical hyphal network and thus the surfaces for nutrient absorption (Klironomos et al., 1998; Sanders et al., 1998; Staddon et al., 2004). Previous research on the effects of elevated atmospheric CO 2 on AMF has mainly focused on increased allocation of photosynthetates to mycorrhizas. Whether host plants directly channel the flow of C to their symbiotic root fungi or whether the latter just profit from an increased C supply of a leaky root system is not known (Fitter et al., 2000). Moreover, plant mineral nutrient deficiency caused by stimulation of photo- synthesis under elevated atmospheric pCO 2 has rarely been considered. Host plants may be able to favour mycorrhizal fungal strains that are beneficial in terms of mineral nutrition.
Transcript of Mycorrhizas improve nitrogen nutrition of Trifolium repens after 8 yr of selection under elevated...

www.newphytologist.org
531
Research
Blackwell Publishing, Ltd.
Mycorrhizas improve nitrogen nutrition of
Trifolium repens
after 8 yr of selection under elevated atmospheric
CO
2
partial pressure
Hannes Gamper
1,3
, Ueli A. Hartwig
2
and Adrian Leuchtmann
1
1
Swiss Federal Institute of Technology (ETH Zürich), Geobotanical Institute, Zollikerstrasse 107, CH-8008 Zürich, Switzerland;
2
Academia Engiadina,
Quadratscha 18, CH-7503 Samedan, Switzerland;
3
Present address: Department of Biology, PO Box 373, University of York, Heslington, York, YO10 5YW, UK
Summary
• Altered environmental conditions may change populations of arbuscular mycor-rhizal fungi and thereby affect mycorrhizal functioning. We investigated whether8 yr of free-air CO
2
enrichment has selected fungi that differently influence thenutrition and growth of host plants.• In a controlled pot experiment, two sets of seven randomly picked single sporeisolates, originating from field plots of elevated (60 Pa) or ambient CO
2
partial pres-sure (pCO
2
), were inoculated on nodulated
Trifolium repens
(white clover) plants.Fungal isolates belonged to the
Glomus claroideum
or
Glomus intraradices
speciescomplex, and host plants were clonal micropropagates derived from nine genets.• Total nitrogen (N) concentration was increased in leaves of plants inoculated withfungal isolates from elevated-pCO
2
plots. These isolates took up nearly twice as muchN from the soil as isolates from ambient-pCO
2
plots and showed much greater stim-ulation of biological N
2
fixation. The morpho-species identity of isolates had a morepronounced effect on N
2
fixation and on root length colonized than isolate identity.• We conclude that rising atmospheric pCO
2
may select for fungal strains that willhelp their host plants to meet increased N demands.
Key words:
arbuscular mycorrhizal fungi (AMF), global environmental change,
Glomus claroideum
,
Glomus intraradices
, nitrogen acquisition, selection, symbioticfunctioning,
Trifolium repens
(white clover).
New Phytologist
(2005)
167
: 531–542
©
New Phytologist
(2005)
doi
: 10.1111/j.1469-8137.2005.01440.x
Author for correspondence:
Hannes Gamper Tel: +44 (0) 1904 328553 Fax: +44 (0) 1904 328510 Email: [email protected]
Received:
7 January 2005
Accepted:
8 March 2005
Introduction
Arbuscular mycorrhizal fungi (AMF, Glomeromycota) are aubiquitous group of soil fungi closely involved in the cyclingof nutrients between plants and soil ( Jakobsen
et al
., 1992;Smith & Read, 1997). They depend exclusively on organiccarbon (C) sources provided by their host plants and thusare likely to respond to environmentally induced alteration inplant photosynthesis. Responses of mycorrhizas to conditionsof elevated atmospheric CO
2
partial pressure (pCO
2
) arevariable and diverse (Staddon & Fitter, 1998; Rillig & Allen,1999; Fitter
et al
., 2000; Rillig
et al
., 2002; Treseder, 2004).The most notable observed response to elevated pCO
2
is an
enlargement of the extraradical hyphal network and thusthe surfaces for nutrient absorption (Klironomos
et al
., 1998;Sanders
et al
., 1998; Staddon
et al
., 2004).Previous research on the effects of elevated atmospheric
CO
2
on AMF has mainly focused on increased allocation ofphotosynthetates to mycorrhizas. Whether host plants directlychannel the flow of C to their symbiotic root fungi or whetherthe latter just profit from an increased C supply of a leaky rootsystem is not known (Fitter
et al
., 2000). Moreover, plantmineral nutrient deficiency caused by stimulation of photo-synthesis under elevated atmospheric pCO
2
has rarely beenconsidered. Host plants may be able to favour mycorrhizalfungal strains that are beneficial in terms of mineral nutrition.

New Phytologist
(2005)
167
: 531–542
www.newphytologist.org
©
New Phytologist
(2005)
Research532
A recent meta-analysis of field studies revealed thatmycorrhizal fungal abundance is increased disproportion-ately relative to root length under elevated pCO
2
conditions(Treseder, 2004). In the same study, Treseder put forward theplant investment hypothesis to explain changes in mycor-rhizal fungal colonization rates upon either phosphorus (P)and nitrogen (N) fertilization or atmospheric CO
2
fumiga-tion. Under this hypothesis, plants adjust allocation of C tomycorrhizal fungi according to the degree to which plantgrowth is nutrient limited (Treseder, 2004). If plant controlover C flows exists, as suggested by the term ‘investment’,plants may preferentially stimulate growth of particularsymbionts and thus change the composition of fungal strainassemblages according to their needs. Plant-mediated selec-tion may be hypothesized to have happened as a result ofprolonged exposure to free-air CO
2
enrichment (FACE).Conditions of elevated atmospheric pCO
2
under FACEare coupled with plant nutrient deficiency (Bazzaz, 1990;Fischer
et al
., 1997; Suter
et al
., 2002; Pendall
et al
., 2004) andshould favour mycorrhizal fungal strains that are more effi-cient in nutrient acquisition and thus more beneficial. Con-versely, under mineral nutrient fertilization or anthropogenicnitrogen (N) deposition, an increase in the abundance ofless beneficial AMF strains has been found (Johnson, 1993;Egerton-Warburton & Allen, 2000).
Adaptive changes in AMF assemblages in response toaltered nutritional conditions may take place via preferentialproliferation and competition. There is considerable variabil-ity among AMF strains or taxa with respect to nutrientacquisition and transfer capabilities [for phosphorus (P),e.g. Jakobsen
et al
., 1992; Munkvold
et al
., 2004; Smith
et al
.,2004; for N, e.g. Hawkins
et al
., 2000; Azcon
et al
., 2001],which is a precondition for selection. Nutrition-mediatedselection was observed in a nonrelated plant pathogen inresponse to elevated pCO
2
(Chakraborty & Datta, 2003) andin a natural yeast population in response to C or N limitation(Goddard & Bradford, 2003).
The long-term response of fertile grasslands is thought todepend heavily on legumes and their associated root symbi-oses (Soussana
et al
., 1996; Zanetti
et al
., 1997; Gamper
et al
., 2004). This group of plants usually forms tripartitenutritional symbioses with bacteria, collectively referred to asrhizobia, and AMF. In this super-symbiotic system, the nod-ule symbiosis provides biologically fixed N, whereas AMFenhance mainly the acquisition of poorly available P. Inexchange, the legume hosts supply their bacterial and fungalroot symbionts with photosynthates. Therefore, under nutrient-poor soil conditions, a true reciprocal exchange of limitingresources results in mutual benefit for all three symbiotic part-ners (Olesniewicz & Thomas, 1999; Schulze, 2004).
Maintenance of both the mycorrhizal and nodule symbi-oses is energetically costly. Each root symbiosis can accountfor up to 20% of net photosynthesis, which is diverted fromhost plant metabolism (Bethlenfalvay
et al
., 1985; Murphy,
1986; Jakobsen & Rosendahl, 1990; Schulze, 2004). Therefore,legumes harbouring both symbioses may acquire symbiontsthat are particularly efficient in carbohydrate use, because ofcompetition between the two root symbionts, whereas grassesunder conditions of elevated pCO
2
may preferentially choosesymbionts conferring N nutritional benefit, because of severeN deficiency. In either case, selection for more beneficialfungal strains under exposure to contrasting pCO
2
conditionsshould be strong. In a model grassland under FACE, thefractional root length colonized by AMF and the relativecontribution of biological nitrogen fixation to total plant Nwere found to be significantly higher (Zanetti
et al
., 1997;Gamper
et al
., 2004; M. Richter, ETH, Zürich, Switzerland,pers. comm.), suggesting an increased contribution ofsymbionts to plant nutrition. From plots of this modelgrassland, which has been exposed to either elevated (60 Pa)or ambient atmospheric pCO
2
for 8 yr, we isolated represent-ative AMF strains with the aim of testing them for theirsymbiotic abilities.
The present study focused on whether growth historiesunder different atmospheric pCO
2
conditions may haveselected for AMF strains with altered symbiotic functioning.We specifically tested whether a set of isolates grown in theFACE plots under elevated CO
2
would (i) improve host plantmineral nutrition and (ii) more heavily colonize roots, com-pared with a set of isolates grown under ambient conditions.In a pot experiment,
Trifolium repens
(white clover) plantswere inoculated with the two sets of AMF strains andgrown under identical and strictly controlled conditions. Wehypothesized that severe N and perhaps P limitations underelevated pCO
2
(Fischer
et al
., 1997; Richter
et al
., 2003) haveinduced host plant-mediated selection for nutritionally morebeneficial strains. The abundance of beneficial strains wasexpected to be increased in the FACE plots under elevatedpCO
2
, increasing the likelihood of isolating strains that willimprove the nutrition of experimental host plants. All AMFsingle spore inocula used in the present study were producedon the same host plant species and under identical glasshouseand soil conditions after isolation from trap cultures. Thus,two growth cycles under identical conditions should havelimited potential maternal effects. Seven isolates each weresampled at random to be included in the experimental setsof AMF isolates from the two pools with contrasting CO
2
selection histories. Uniform and functional nodulation wasensured by inoculating the experimental one-to-one fungus–plant pairs with a common rhizobial strain. Harvest took placeafter 72 d of growth under identical, ambient pCO
2
conditions.Because the AMF isolates belonged to two distinct groups of
spore morphotypes, we were able to further assess the effectof fungal taxonomic grouping on effect size and significanceof difference in measured parameters. Thus we investigatedwhether the identity of morpho-species or AMF strain wasmore important for symbiotic measures. To our knowledge,this is the first experiment under controlled conditions and

©
New Phytologist
(2005)
www.newphytologist.org
New Phytologist
(2005)
167
: 531–542
Research 533
using multiple single spore isolates that has assessed thephysiological effects of past selection under experimentallyelevated atmospheric CO
2
on AMF in the field.
Materials and Methods
AMF inocula
AMF inocula were prepared from open-pot trapping cultures(Gilmore, 1968) with soil samples taken from the Swiss-FACEexperiment, located 20 km north-east of Zürich, Switzerland.This experiment consisted of permanent monoculture plots of
Lolium perenne
L. cv. Bastion (perennial ryegrass) and
Trifoliumrepens
L. cv. Milkanova (white clover) and the mixture of bothplant species in ambient and elevated pCO
2
(60 Pa) treatments,and in an additional cross-factorial N-fertilization treatment (14and 56 g m
−
2
yr
−
1
) at the subplot level. At the time of samplingin autumn 2000, half of the plots had been fumigated withCO
2
for 8 yr. The soil at the site of the FACE experimentis a clay loam, classified as fertile, eutric cambisol [Food andAgriculture Organization (FAO) Classification System]. Forfurther details see Zanetti
et al
. (1996) or Suter
et al
. (2002).After 5 months of trap culturing, pure AMF isolates
were established from single spores on
Plantago lanceolata
L.(ribwort plantain), using a potting substrate which consistedof 1 : 2 : 2 volumes of pasteurized field soil from theupper horizon [clay loam, pH (H
2
O) 5.8, available P (Olsen)52 mg P kg
−
1
, re-inoculated with a filtrate of the naturalmicroflora], silica sand (0.7–1.2 mm) and an expandedattapulgite clay (Oil Dry Chem-Sorb WR 24/18, BrenntagMediterranée Export, Vitrolles Cedex, France). All success-fully isolated single spore cultures (approx. 10%) belonged tothe genus
Glomus
and were propagated in bi-species cultures
of
Zea mays
cv. Corso and
P. lanceolata
over a period of7.5 months. Identical soil conditions and host plants duringthe two growth cycles of single spore isolation and inoculumproduction were expected to have minimized potential mater-nal effects caused by host–plant differences in trap cultures(Table 1).
Fourteen fungal isolates were recovered from plots sub-jected to ambient pCO
2
(ambCO
2
isolates), and 44 fromplots subjected to elevated pCO
2
(elevCO
2
isolates). Fromeach of these pools, seven isolates were randomly chosen irre-spective of morpho-species identity, previous field conditionsor trap-culture origin (Table 1). This strategy of randomlychoosing AMF isolates avoided any bias caused by
a priori
expectations. Isolates belonged to the
Glomus claroideum
Schenck & Smith (Gc) and the
Glomus intraradices
(Gi)Schenck & Smith species complexes. Gi isolates were initiallysubclassified as
G. intraradices
and
G. aggregatum
Schenck &Smith, but this became untenable after single spore and inoc-ulum propagation [for a similar finding see Johnson (1993),p. 752]. Certain inocula of Gi isolates contained spore aggre-gates of up to macroscopic size and root cortices of
Z. mays
and
P. lanceolata
that were densely packed with completelyhyaline vesicles. Both AMF species were found in considerableabundance in the root systems of
T. repens
in the originalFACE plots (H. Gamper, U. A. Hartwig & A. Leuchtmann,unpublished).
Plant material
Clonal material was produced from uniform single nodecuttings of different genets of
T. repens
L. cv. Milkanova(Dansk Planteforaedling, Research Division, Store Heddinge,Denmark). Leaf nodes were surface-disinfected and precultured
pCO2 (Pa) Morpho-species Field host(s)
N fertilization (g m−2 yr−1) Trap hosts Isolate number
35 Gc Tr 14 Df & La 412Tr 56 Df & La 52
Gi Tr 56 Df & La 57Tr 56 Df & La 588Lp & Tr 14 Df & La 538Lp & Tr 14 Df & La 539Lp & Tr 14 Df & La 540
60 Gc Lp & Tr 14 Ob & Tp 283Lp & Tr 14 Ob & Tp 284Lp & Tr 56 Lp & Tr 562
Gi Lp & Tr 56 Pl & Zm 484Lp & Tr 56 Pl & Zm 492Lp & Tr 56 Pl & Zm 494Lp & Tr 56 Pl & Zm 498
Fungal isolates belonged to the Glomus claroideum (Gc) or Glomus intraradices (Gi) species complex.Df, Dipsacus fullonum L.; La, Leontodon autumnalis L.; Lp, Lolium perenne L.; Ob, Oenothera biennis L.; Pl, Plantago lanceolata L.; Tp, Tagetes patula L.; Tr, Trifolium repens L.; Zm, Zea mays L. cv. Corso.
Table 1 Identity and histories of host plant species cover, atmospheric CO2 partial pressure (pCO2) and nitrogen (N) fertilization in the field of origin of the 14 single spore isolates of arbuscular mycorrhizal fungi (AMF) used in this experiment.

New Phytologist (2005) 167: 531–542 www.newphytologist.org © New Phytologist (2005)
Research534
on cellulose filter paper on top of 1 × Hoagland (Hoagland& Arnon, 1938) nutrition solution agar (0.7%, weight/volume). After 10 d, micropropagates were transplanted tofinely ground Perlite, and after an additional 14 d plantlets ofthe nine most vigorous and uniform genets (labelled A, B, andF–L) were selected for the current inoculation experiment.
Experimental set-up and growth conditions
The inoculation experiment used nine replicate 1 l pots(14 × 10 cm) for each of the 14 AMF inocula. Replicate potswere planted each with a different micropropagate of the nineT. repens genets, resulting in an orthogonal design with eachplant genet/AMF isolate combination represented once (126pots). As control, six additional sets of pots with the nine plantgenets lacking AMF inoculum were used (54 pots).
Inocula, stored at 4°C for 8 months before use, were thor-oughly mixed into AMF-free potting substrate (compositionas described above) in a volumetric ratio of 2 : 5. The inoculaconsisted of dried pot culture material, which containedspores, colonized root fragments, and extraradical hyphae.Nonmycorrhizal control pots (NM-control) received anAMF-free mock-inoculum, but were otherwise identical to theinoculated pots. Additional NM-control pots, also replicatednine times, were established for five different P-fertilizationtreatments. These pots received cumulated amounts of 50,100, 150, 200 and 250 mg readily available phosphorus(KH2PO4) per kg of dry potting substrate in eight weeklyapplications (50 ml) from the third week onwards. Substratesurfaces of all pots were covered with a layer of coarse silicasand to prevent the formation of adventive roots from leafnodes of stolons which otherwise could have given plantsunequal access to soil nutrients.
To balance the non-AMF microbial community qualita-tively (Koide & Li, 1989), 50 ml of a suspension of a filtrateprepared from the fungal and mock inocula was applied toeach pot. To determine the proportion of biologically fixedN (%bfN2), 10 g Na15NO3 (10% enriched) and 5 g Ca(15NO3)2(5% enriched) were added as soil 15N tracer in the samesuspension. As reference plants, L. perenne seedlings weregrown in two extra pots to which the same amount of 15Nwas added (McAuliffe et al., 1958). To promote uniformnodulation of T. repens plants, dense plate-washes of Rhizobiumleguminosarum bv. trifolii (strain RBL 5020; Leiden, theNetherlands) were added.
Two identical climate chambers (Conviron, PGV-36,Winnipeg, Manitoba, Canada) were required to carry all pots.Sets of four and five pots of each AMF isolate or P fertilizationtreatment were randomly chosen and assigned to one of theclimate chambers, in which they were positioned randomly.Growth conditions consisted of a light phase of 16 h [450–550 µmol m−2 s−1 photosynthetic fluence rate (PPFR) overthe waveband 400–700 nm at plant height], including dawnphases of 30 min twice a day, a day/night temperature profile
of 23/15°C at plant height and 70–80% relative air humidity.Cool white fluorescent lamps and incandescent bulbs (OsramSylvania Inc, St. Marys, PA, USA) provided the high photo-synthetic active irradiance. Automatic drip-irrigation withdeionized water from a tensiometer-type system (TropfBlumat, Weninger GmbH & Co. KG, Telfs, Austria)maintained substrate humidity at about 40%, or 50–60% ofits water-holding capacity, preventing any leaching frompots. Twenty-four, 32 and 43 d after transplantation, 50 mlof 0.1 × Hoagland nutrient solution (Hoagland & Arnon,1938), lacking P, was applied to each pot. On day 43 anadditional 50 ml of a dense suspension of rhizobia was addedto each pot.
Plant growth measurements
Seventy-three days after inoculation, plants of T. repenswere destructively harvested and dry masses determined afterdrying to constant weight for at least 48 h at 65°C. The wholeplant leaf area was determined by passing all leaf laminaethrough a Li-Cor 3000A leaf area meter (Li-Cor Inc., Lincoln,NE, USA).
From each washed root system, a random subsample(≤ 1 g) was preserved in 70% ethanol for root length determi-nation using the grid line intersect method (Tennant, 1975)adapted for computer scanner-assisted estimation. Lengthestimates for the whole root systems were made based on dryweights of the subsamples and the whole root systems. Thedry weight of whole root systems was corrected for the twosubsamples, and the root : shoot dry weight ratio, specificroot length and specific leaf area were calculated using the dryweights of the respective plant fractions.
Fungal growth measurements
Root colonization was determined in the root fragments(∼1.5 cm in length) of a random subsample (≤ 0.3 g) takenfrom the middle zone of root systems. Root fragments werecleared and stained using a modified method of Phillips &Hayman (1970) in which lacto-phenol was substituted bylacto-glycerol and methyl blue was used in addition to trypanblue. Proportions (%) of root length colonized by hyphae,arbuscules and vesicles were determined by enumerating100 intersections of at least 20 root fragments per sample,according to the magnified intersections method at ×200magnification (McGonigle et al., 1990). The total root lengthcolonized by AMF (RLC) was calculated by multiplyingthe absolute root length by the proportion of root lengthcolonized by hyphae.
The density of extraradical hyphal length was estimatedfor duplicate samples of 5 g of fresh material from three cores(diameter 1 cm) of potting substrate, which were takenbetween the plant and the pot edge and thoroughly mixed. Amodified membrane filter technique based on the formula

© New Phytologist (2005) www.newphytologist.org New Phytologist (2005) 167: 531–542
Research 535
of Newman (1966) was used (Jakobsen et al., 1992; Sylvia,1992) to determine hyphal length. Briefly, after an aqueousextraction, intersections of hyphal fragments were enumeratedon 0.45-µm gridded nitrocellulose filters of 25 mm diameter(HAWG, Millipore, Bedford, MA, USA), while calculatedlength estimates were corrected for the dilution of suspen-sions and water contents of each individual sample of pottingsubstrate.
Nutrient concentrations in plant tissue
Chemical analyses of P and N were carried out on subsamplesof the ground leaves (without petioles) and roots. These plantfractions were chosen because they represent the physiol-ogically most active organs.
For measuring P concentrations, samples of approximately500 mg of dry powder (40°C) were incinerated at 500°C for6 h, dissolved in 10% HCl, filtered (0.5 µm) and analysedby the method of Murphy & Riley (1962). Dilutions to therange required were measured in a continuous flow Skalarapparatus (Skalar Analytic GmbH, Erkelenz, Germany) at880 nm, where the absorbance by the ascorbic acid-reducedantimony-phospho-molybdate complex is maximal.
The 15N isotope dilution method (McAuliffe et al., 1958)was used to determine partitioning of N in T. repens betweensymbiotic N2 fixation and uptake from the potting substrate.Total N and percentage 15N were determined in samples of2 ± 0.1 mg dry powder (40°C) of T. repens (and L. perenne asa reference plant) at the Stable Isotope Facility of the Universityof California, Davis, using a continuous flow isotope ratio massspectrometer (cf-IRMS, Europa Integra; PDZ-Europa Scientific,Sandbach, UK). Atom per cent 15N excess (i.e. 15N enrich-ment) was calculated and applied to the formula of McAuliffeet al. (1958) to estimate %bfN2, the remainder being assumedto represent N that was taken up from the substrate.
Statistical analyses
The experimental designs for the separate AMF inoculationand NM-control experiments were each cross-factorialwith respect to the treatment T. repens genet. There was noreplication of the treatment combinations of AMF isolate (14levels) × plant genet (nine levels) and P fertilization (six levels)× plant genet (nine levels) in the two separate experiments.The AMF isolate treatment (for the inoculation experiment)and P-fertilization level (for the NM-control experiment)were replicated by the nine genets of T. repens. This experi-mental design provided control over the variability withinAMF inocula and among genets of T. repens. It allowed detectionof the effects of AMF isolates and genets of T. repens, but notof the AMF isolate × plant genet interaction.
Linear mixed analysis of variance (ANOVA) modelswith nested factors were used to assess the effects of the pre-experimental CO2 history of AMF isolates (two levels), the
morpho-species identity of AMF isolates (two levels), theisolate identity (14 levels), the plant genet identity (nine levels)and the technical factor climate chamber (two levels), in SPSSv.11.0.2 for MacOSX (SPSS Inc., Chicago, IL, USA). Analysesof covariance (ANCOVAs) were applied to the data to assessthe importance of root and hyphal length for mineral nutritionof the plants. Similarly, the potential effects of differences inspore numbers in the inocula on all dependent variables wereassessed. AMF isolate and plant genet were treated as randomfactors, except where inter-isolate variability was examined inFigs 2 and 3 (see later). CO2 history, morpho-species andclimate chamber were defined as fixed factors. Morpho-speciesidentity was nested within CO2 history and the interactionterm (CO2 history × morpho-species) within AMF isolates.NM-control treatments were excluded from ANOVA, becausewe were particularly interested in effects caused by the twosets of AMF isolates with different CO2 histories and not ineffects caused by the colonization status of plants.
The initial statistical model was simplified throughelimination of the interaction term, in cases where it didnot significantly contribute to variance. Schwarz’s BayesianInformation Criterion (BIC) under maximum likelihood(ML) variance estimation was used to control improvementin model goodness. Final significances, reported in the Resultssection, were determined via restricted maximum likelihood(REML) variance estimation.
In two additional analyses of variance, morpho-specieswas replaced by either the pre-experimental N-fertilizationtreatment or the pre-experimental host plant species treat-ment of the FACE plots. These analyses were used to checkfor potential interference of these treatments as selectionfactors, in addition to the CO2 history. All reported statisticalresults are from mixed ANOVA, which included the factormorpho-species identity of AMF isolates, if not statedotherwise. Significance values for the factor CO2 history werealmost identical in the three separate ANOVAs on the samedependent variable.
Phosphorus fertilization and plant genet effects onNM-controls were analysed by two-way ANOVA, followedby multiple pairwise comparisons of the means of factor levelsusing Tukey–Kramer’s honestly significant difference test of v.5.1 (SAS Institute Inc., Cary, NC, USA). Dunnett’s test,as implemented in v.5.1, was used for multiple pairwisecomparisons between the mock-inoculated NM-controltreatment and all the AMF inoculation and P-fertilizationtreatments.
Pearson pairwise correlation coefficients were calculatedand their significance determined in two-tailed t-tests asindicators of the direction of the relationship between mea-sured plant and fungal parameters. Guidelines in Zar (1999)were followed when data had to be transformed to meetassumptions in ANOVA. Weights and concentrations wereln(x + 1), counts were √(x + 3/8) and ratios were arcsin√(x)-transformed.
New Phytologist (2005) 167: 531–542 www.newphytologist.org © New Phytologist (2005)
Research536
Results
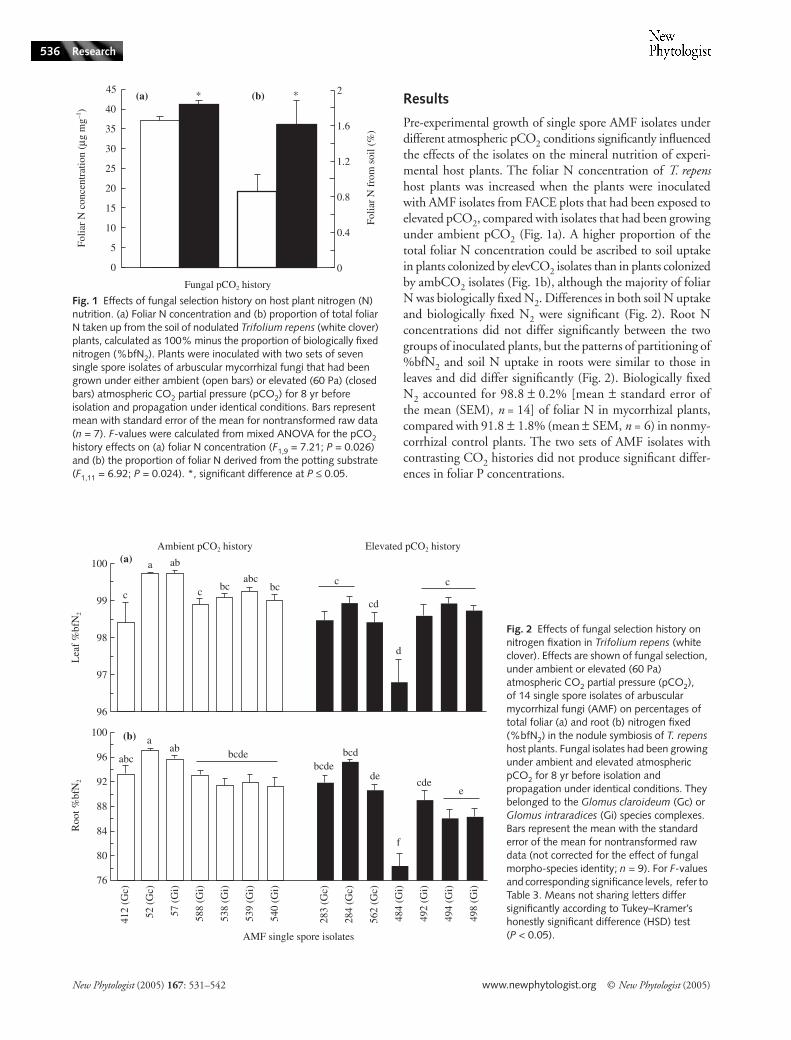
Pre-experimental growth of single spore AMF isolates underdifferent atmospheric pCO2 conditions significantly influencedthe effects of the isolates on the mineral nutrition of experi-mental host plants. The foliar N concentration of T. repenshost plants was increased when the plants were inoculatedwith AMF isolates from FACE plots that had been exposed toelevated pCO2, compared with isolates that had been growingunder ambient pCO2 (Fig. 1a). A higher proportion of thetotal foliar N concentration could be ascribed to soil uptakein plants colonized by elevCO2 isolates than in plants colonizedby ambCO2 isolates (Fig. 1b), although the majority of foliarN was biologically fixed N2. Differences in both soil N uptakeand biologically fixed N2 were significant (Fig. 2). Root Nconcentrations did not differ significantly between the twogroups of inoculated plants, but the patterns of partitioning of%bfN2 and soil N uptake in roots were similar to those inleaves and did differ significantly (Fig. 2). Biologically fixedN2 accounted for 98.8 ± 0.2% [mean ± standard error ofthe mean (SEM), n = 14] of foliar N in mycorrhizal plants,compared with 91.8 ± 1.8% (mean ± SEM, n = 6) in nonmy-corrhizal control plants. The two sets of AMF isolates withcontrasting CO2 histories did not produce significant differ-ences in foliar P concentrations.
Fig. 2 Effects of fungal selection history on nitrogen fixation in Trifolium repens (white clover). Effects are shown of fungal selection, under ambient or elevated (60 Pa) atmospheric CO2 partial pressure (pCO2), of 14 single spore isolates of arbuscular mycorrhizal fungi (AMF) on percentages of total foliar (a) and root (b) nitrogen fixed (%bfN2) in the nodule symbiosis of T. repens host plants. Fungal isolates had been growing under ambient and elevated atmospheric pCO2 for 8 yr before isolation and propagation under identical conditions. They belonged to the Glomus claroideum (Gc) or Glomus intraradices (Gi) species complexes. Bars represent the mean with the standard error of the mean for nontransformed raw data (not corrected for the effect of fungal morpho-species identity; n = 9). For F-values and corresponding significance levels, refer to Table 3. Means not sharing letters differ significantly according to Tukey–Kramer’s honestly significant difference (HSD) test (P < 0.05).
Fig. 1 Effects of fungal selection history on host plant nitrogen (N) nutrition. (a) Foliar N concentration and (b) proportion of total foliar N taken up from the soil of nodulated Trifolium repens (white clover) plants, calculated as 100% minus the proportion of biologically fixed nitrogen (%bfN2). Plants were inoculated with two sets of seven single spore isolates of arbuscular mycorrhizal fungi that had been grown under either ambient (open bars) or elevated (60 Pa) (closed bars) atmospheric CO2 partial pressure (pCO2) for 8 yr before isolation and propagation under identical conditions. Bars represent mean with standard error of the mean for nontransformed raw data (n = 7). F-values were calculated from mixed ANOVA for the pCO2 history effects on (a) foliar N concentration (F1,9 = 7.21; P = 0.026) and (b) the proportion of foliar N derived from the potting substrate (F1,11 = 6.92; P = 0.024). *, significant difference at P ≤ 0.05.

© New Phytologist (2005) www.newphytologist.org New Phytologist (2005) 167: 531–542
Research 537
The CO2 histories of the two sets of AMF isolates affectedthe percentage of root length colonized by hyphae and vesiclesand the total root length colonized by hyphae (RLC, Fig. 3).Other morphometric plant parameters, including total plantdry weight, shoot : root dry weight ratio, total and specific leafareas, and total and specific root lengths did not differ betweenthe plants colonized by AMF isolates with different CO2histories. Similarly, there was no difference between the extra-radical hyphal length densities of the two sets of AMF isolates[7.40 ± 0.65 m g−1 (mean ± SEM, n = 14)].
Pre-experimental N fertilization and host plant, two otherpotential selecting factors, had no effect on the plant and fungalparameters studied, with two exceptions: there was a signifi-cant main effect of the host plant on percentage of root lengthcolonized by hyphae (F1,11 = 10.76, P = 0.007) and vesicles(F1,11 = 8.18, P = 0.016); this effect did not, however, interactwith the effect of CO2 history.
Extraradical hyphal length density and root length werenot significant covariates for foliar N and P concentrations. Theproportions of root length colonized by the different AMF
Fig. 3 Effects of fungal selection history on root colonization in Trifolium repens (white clover). Effects are shown of fungal selection, under ambient and elevated (60 Pa) atmospheric CO2 partial pressure (pCO2), of 14 single spore isolates of arbuscular mycorrhizal fungi (AMF) on the percentage of root length colonized by hyphae (a), arbuscules (b) and vesicles (c) and on the total colonized root length (d) of experimental T. repens host plants. Fungal isolates had been growing under ambient and elevated atmospheric pCO2 for 8 yr before isolation and propagation under identical conditions. They belonged to the Glomus claroideum (Gc) or Glomus intraradices (Gi) species complexes. Bars represent the mean with the standard error of the mean for nontransformed raw data (not corrected for the effect of fungal morpho-species identity; n = 9). For F-values and corresponding significance levels, refer to Table 3. Means not sharing letters differ significantly according to Tukey–Kramer’s HSD test (P < 0.05).

New Phytologist (2005) 167: 531–542 www.newphytologist.org © New Phytologist (2005)
Research538
structures or the total colonized root length did not explainfoliar N concentration either. The lack of a contribution toplant nutrition by surrogates of root and hyphal surfaces isalso evident from the significant negative pairwise correlationslisted in Table 2, and from negative correlations (P < 0.01)between the root length colonized by hyphae and %bfN2 ofleaves (r2 = −0.264) and roots (r2 = −0.564).
The 14 AMF single spore isolates did not significantlycontribute to either foliar [2.00 ± 0.05 mg g−1 (mean ± SEM,n = 14)] or root [1.66 ± 0.03 mg g−1 (SEM, n = 14)] Pconcentrations of the T. repens host plants, when comparedwith the mock-inoculated NM-control [2.02 ± 0.18 and 1.75 ±0.15 mg g−1 (SEM, n = 9), respectively], although increasingthe P fertilization of the NM-controls significantly increasedboth these values. P concentrations differed significantly (Tukey’sHSD test at P < 0.05: Q = 3.61) among the NM-control plantsreceiving cumulated amounts of 50 and 150, 100 and 200, and150 and 250 mg readily available P (data not shown). FoliarN concentrations did not significantly differ between the mock-inoculated NM-control and the AMF inoculation treatments[35.99 ± 2.24 (mean ± SEM, n = 9) and 39.14 ± 0.91 mg g−1
(mean ± SEM, n = 14), respectively; Dunnett’s test: d = 2.93,
P > 0.241]. Proportions of symbiotically fixed nitrogen in theleaves and roots did not differ between the mock-inoculatedNM-control and the AMF inoculated plants either, except forthe fungal inoculation treatments 52 and 57, which showedsignificantly lower values (Dunnett’s test: d = 2.93,P < 0.05). These values were comparable to those of the fiveNM-controls with addition of P fertilizer (data not shown).Moreover, only two AMF inoculation treatments (one Gc iso-late from each of the CO2 fumigation histories) reduced totalhost plant biomass [Dunnett’s test: d = 2.93; isolate 283:8.27 ± 1.08 g (mean ± SEM, n = 9), P = 0.027; isolate 412:8.65 ± 1.40 g (mean ± SEM, n = 9), P = 0.032; mock-inocu-lation control: 16.33 ± 1.63 g (mean ± SEM, n = 9)], mainlyas a result of reductions in shoot dry weight. None of theNM-control plants was colonized by AMF. Differences inspore numbers of inocula did not contribute to any of the dif-ferences in the measured parameters, as evidenced byANCOVA (data not shown).
Effect sizes explained by morpho-species identity of AMFisolates for the parameters presented in Figs 2 and 3 were largerthan those explained by isolate identity, although both effectswere significant (Table 3). Indeed, the values of F-ratios
P concentration N concentration
Root Foliar Root Foliar
Root length −0.502*** −0.469*** −0.569*** −0.593***Extraradical hyphal length −0.292*** −0.246** −0.298*** −0.293***RLC −0.391*** −0.387*** −0.438*** −0.353***
Fungal isolates belonged to the Glomus claroideum (Gc) or Glomus intraradices (Gi) species complexes.Significance levels derived from two-tailed t-tests: **, P < 0.01; ***, P < 0.001.
Table 2 Pairwise Pearson correlations for root and foliar phosphorus (P) and nitrogen (N) concentrations, and root, extraradical hyphal and total colonized root length (RLC) in Trifolium repens plants colonized by 14 different single spore isolates of arbuscular mycorrhizal fungi (n = 126).
Table 3 Summary table for mixed ANOVA testing the parameters foliar and root nitrogen fixed in the nodule symbiosis (%bfN2), percentage of root length colonized by hyphae, arbuscules, and vesicles, and total colonized root length for effects of factors CO2 history, arbuscular mycorrhizal fungi (AMF) morpho-species identity (Glomus claroideum or G. intraradices), AMF isolate identity, Trifolium repens genet identity and climate chamber.
Parameter
Factor
CO2 history (morpho-species) (df = 2)
Morpho-species identity (df = 1)
Isolate identity (CO2 history × morpho-species) (df = 10)
Plant genet identity (Z-value)
Climate chamber(df = 1)
Foliar %bfN2 21.31*** 0.59ns 6.71*** 0.28ns 0.06ns
Root %bfN2 48.57*** 58.77*** 6.48*** 1.29ns 0.98ns
Hyphae (%) 32.21*** 65.79*** 14.03*** 1.29ns 0.26ns
Arbuscules (%) 1.36ns 16.48*** 3.30** 0.30ns 0.22ns
Vesicles (%) 49.18*** 306.15*** 14.45*** 1.10ns 0.64ns
Total colonization (m) 14.72*** 53.82*** 6.50*** 1.53ns 0.93ns
F-values and corresponding significance levels are given for all fixed factors, and the Wald Z statistic for the random factor plant genet identity.Significance levels: ns, P > 0.05; **, P < 0.01; ***, P < 0.001.Denominator degrees of freedom (df) were 103 for all factors, except for the factor climate chamber for which 80 < df < 105.

© New Phytologist (2005) www.newphytologist.org New Phytologist (2005) 167: 531–542
Research 539
from ANOVA for the factor morpho-species identity wereconsistently higher than those for the factor isolate identityfor all dependent variables, foliar %bfN2, root %bfN2, hyphal,arbuscular, and vesicular root colonization, and RLC (Table 3).In general, %bfN2 and percentage of root length colonizedby arbuscules were higher in T. repens host plants colonized byAMF isolates of G. claroideum than in those colonized byisolates belonging to the G. intraradices species complex. Theopposite was found for hyphal and vesicular root colonizationrates and RLC.
Discussion
Effects of fungal selection history
In this pot experiment, we showed that 8 yr of FACE selectedfor more plant-beneficial AMF strains in a permanent,agricultural model grassland. Experimental T. repens plantshad 10% higher foliar N concentrations when inoculatedwith AMF isolates from field plots with elevated pCO2conditions (elevCO2 isolates), compared with plantsassociated with isolates from plots with ambient pCO2conditions (ambCO2 isolates, Fig. 1a). ElevCO2 isolatesstimulated both biological N2 fixation in the nodulesymbiosis and nitrate uptake from the growth substrate,as measured by the 15N dilution method. Nitrogen uptakefrom soil was nearly twice as high in plants colonized byelevCO2 isolates as in plants colonized by ambCO2 isolates(Fig. 1b). Synergism between the arbuscular mycorrhizaland nodule symbioses (e.g. Olesniewicz & Thomas, 1999)and the contribution of AMF to plant N nutrition (Tobaret al., 1994; Hawkins et al., 2000; Azcon et al., 2001) havebeen previously observed. Recently, Bago et al. (2004) reportedon the ability of an G. intraradices isolate to take up nitrateefficiently through a change in extraradical hyphal growth underin vitro conditions. The new finding of our study is that only8 yr of CO2 fumigation had favoured AMF strains with improvedN nutritional capabilities, which is particularly noteworthyfor asexual, slow-growing organisms where adaptation toenvironmental change is thought to be slow (Sanders, 2002).
The observed selective change in the AMF field assemblagewas probably plant-mediated. In this FACE experiment,mineral nutrient deficiency and growth limitation of hostplants occurred under elevated atmospheric pCO2 (Fischeret al., 1997; Suter et al., 2002). Moreover, accumulation ofsoil organic matter under elevated pCO2 may have increasedmicrobial immobilization of mineral nutrients (Zak et al.,2000; Schneider et al., 2004). Nutrient retention in soils andincreased plant demand under elevated pCO2 both makenutritional symbioses more important. An immediate uptakeof nutrients released from decaying organic material bymycorrhizal fungi may be crucial under elevated pCO2. Infact, acquisition of N from organic sources by plants in theFACE experiment became more important as the experiment
went on (Schneider et al., 2004). It is possible that theglutamine synthetase activities involved in N acquisitiondiffered in plants inoculated with the two sets of AMF, ashas been observed by Smith et al. (1985) and Breuninger et al.(2004). The shift in mycorrhizal fungal properties towardsmore beneficial strains under long-term atmospheric CO2fumigation is consistent with the concept of a mutualism–parasitism continuum (Johnson et al., 1997) and concurswith the evolutionary predictions deduced from the plantinvestment hypothesis (Treseder, 2004).
Another, although less likely, explanation for the observeddifferences in N concentration of plant tissues is unequalenergetic costs (C costs) of AMF isolates. Increased availabilityof C under elevated atmospheric pCO2 could have favouredmore C-demanding AMF strains, leaving fewer C resources toexperimental host plants for building up biomass. This mayhave increased N concentration in hosts of the elevCO2 isolates(Jarrell & Beverly, 1981). Significantly smaller biomasses oftwo of the 14 AMF inoculation treatments and no growthresponse in the NM-controls to P fertilization (data notshown) suggest that P was not limiting and thus C costs mightindeed have been present. Moreover, more intense hyphal andvesicular root colonization observed in plants with elevCO2isolates may indicate that C investments into biomass ofAMF were enhanced (Fig. 3). In addition, limited energeticresources may have shifted the partitioning of N acquisitionfrom the energetically more expensive symbiotic N2 fixationto nitrate uptake from soil.
The past N-fertilization regime and field hosts in FACEplots did not interfere with the observed effects of the CO2history on the plant nutritional parameters. There were nostatistically significant main or interaction effects of hostplant, N fertilization, and CO2 history. Therefore, the observeddifferences in the symbiotic traits affecting plant nutritionwere the result of selection resulting from differences inatmospheric pCO2. Only hyphal and vesicular root colonizationwas affected by the field-host history, suggesting that thesetraits may have been influenced by pre-experimental CO2conditions and/or host plant.
The observed effect of field hosts on hyphal and vesicularroot colonization rates suggests a selective effect of hostplant species in FACE plots. In the pot experiment, we foundhigher hyphal and vesicular root colonization rates for AMFstrains originating from mixed T. repens and L. perenne plots,compared with those originating from monocultures ofT. repens. Differences in C and N metabolism of grasses andlegumes may have affected the AMF assemblages of the twohosts with respect to the relative abundance of strains withdifferent colonization abilities. On the one hand, L. perennecould have selected for more C-demanding AMF strains,because as a non-N2-fixing plant it has been shown to sufferparticularly from N deficiency and C-sink limitation underelevated atmospheric pCO2 (Fischer et al., 1997; Suter et al.,2002). On the other hand, the assemblage of AMF strains

New Phytologist (2005) 167: 531–542 www.newphytologist.org © New Phytologist (2005)
Research540
from T. repens monocultures may have adapted to their hosts,such that fungal strains that require less intense root coloniza-tion for the same symbiotic benefit became more abundant.
The tripartite symbiosis among legume host plants, AMF,and rhizobial strains involves a close interdependence of theplant C and mineral nutrient metabolisms. Regulation of theN status in legumes is complex and cannot yet be explainedby a unifying concept (Schulze, 2004). Feedback mechanismsvia hormones as well as C costs, N status and O2 permeabilityin nodules were invoked to explain patterns in plant Nnutrition (Bethlenfalvay et al., 1985; Murphy, 1986; Hartwiget al., 1994; Hartwig, 1998; Catford et al., 2003). Thus, giventhe regulatory complexity of legume physiology, furtherstudies would be needed to uncover the mechanisms that haveled to the observed N nutritional patterns in the experimentalT. repens plants. Nevertheless, it is fairly clear from the resultsof ANCOVA that the improved plant N nutrition conferredby the elevCO2 isolates was not a result of better soil exploi-tation by AMF hyphae or their host plant roots. Enlarging thepotential surfaces for absorption would not have been aneconomic scavenging strategy for nitrate, an element that ishighly mobile and evenly distributed in soils. The negativecorrelations between the plant tissue P and N concentrationsand the plant and fungal organs involved in nutrient uptake(Table 2) indicate that plants appear to regulate their invest-ment into the root system and mycorrhizal partners accordingto their actual requirements. Conversely, from a fungal point ofview, extraradical hyphal growth of AMF seems to be diminishedwhen host plants are better supplied with mineral nutrients,perhaps because of limitation in available C resources.
Effects of mycorrhizal inoculation and P fertilization
In general, dry weights of plants did not differ among theAMF inoculation treatments and the mock-inoculation control,except for the dry weights of host plants inoculated with twoGc isolates, which were significantly lower (see Results andDiscussion sections above). This may reflect the high photo-synthetic compensation ability of T. repens towards mycorrhizalC costs (Wright et al., 1998). Moreover, mycorrhizal plants,compared to the unfertilized mock-inoculated plants, did notdiffer in their foliar and root P and N concentrations, indicatingthat the AMF isolates did not contribute, or only marginallycontributed, to plant mineral nutrition under the presentexperimental conditions. An explanation for this limitednutritional function of AMF might be the nutrient-rich fieldsoil contained in our potting substrate. Therefore, it is evenmore striking that, in plants inoculated with the two sets ofAMF isolates, significant differences in N nutrition were found.
P fertilization applied to NM-control plants of T. repensneither increased biomass of plants nor decreased root : shootdry weight ratio and specific root length, although it increasedfoliar and root P concentrations. The lack of symptoms ofmineral nutrient deficiency and the absence of a growth
response to P fertilization clearly indicate that P was availablein sufficient quantity. In fact, nonlimiting soil P shouldexclude significant, additional plant P nutritional benefitsfrom the symbiosis, and thus C costs imposed by AMF shouldbe particularly manifested (Peng et al., 1993; Graham et al.,1996). Our experimental fungal isolates may have been selectedby the original soil conditions, which were rich in available P,and thus should be adapted to such growth conditions. Theseconditions may have enhanced fungal C costs and made Nmore limiting, and thus the present type of potting substratewas suitable for detecting N-nutritional AMF effects. Moreover,the experimental outcome may have been influenced by thechoice of the rhizobial strain, as strong interactions betweenthe bacterial and fungal root symbioses are well established(Ianson & Linderman, 1993; Xavier & Germida, 2002).
Effects of morpho-species and isolate identity of AMF
Both morpho-species and AMF isolate identity contributedsignificantly to the observed patterns in AMF root colon-ization and proportion of N fixed in the nodule symbiosis(Figs 2 and 3). Yet, effect sizes expressed in the form ofF-values were consistently higher for morpho-species(G. claroideum or G. intraradices) than for isolate identity(Table 3). However, our finding that morpho-species identityat the species level is relevant to functional differences is newfor arbuscular mycorrhizal research and might challengethe report by Munkvold et al. (2004), which showed thatfunctional variability among AMF isolates may override thevariability found among morpho-species and phylo-species.Moreover, Hart & Klironomos (2002) compared thecoefficients of variance of the mycorrhizal plant growthresponse at several levels of biological integration of AMF andfound that sizes in the coefficients of variance were comparableat the morpho-species and isolate levels. By contrast, variationsat the isolate level in our data are consistent with the geneticvariability found among isolates of G. intraradices also from asingle field site (Koch et al., 2004).
Based on the hierarchy of biological integration, largereffect sizes at higher taxonomic levels should generally beexpected. Our data provide evidence for hierarchical structur-ing of effect sizes at the species level, and supplement know-ledge of even more pronounced variation in fungal growthpatterns at the level of AMF families (Hart & Reader, 2002).Consequently, future experiments should include adequatereplication at each level of taxonomic integration to morecomprehensively resolve the sources of variance of measuredparameters caused by experimental treatments or environ-mental selection.
Conclusions
Knowledge about the impact of rising atmospheric pCO2on mycorrhizal symbioses is crucial for predicting potential

© New Phytologist (2005) www.newphytologist.org New Phytologist (2005) 167: 531–542
Research 541
changes in ecosystems. We have shown that elevatedatmospheric pCO2 for only 8 yr in model grasslands selectedAMF strains that can improve the N nutrition of T. repenshost plants, a common species of temperate grasslands.Improved N nutrition was found to result from bothstimulation of biological N2 fixation in the nodule symbiosisand soil N uptake. Alleviation of mineral nutrient deficiencyby AMF strains could be particularly relevant for nonlegumes,which are known to experience severe N-growth limitationunder elevated atmospheric pCO2.
Intraspecific variability observed among isolates of theG. claroideum and G. intraradices species complexes, as well asinterspecific variability between these two taxonomic groups,suggests that future inoculation experiments should considerboth intra- and interspecific variation. Further experimentswould be needed to scale up our findings to the ecosystemlevel. In particular, an experiment with a nonlegume hostplant lacking the buffering effect of biological N2 fixationwould be of interest, as in such a system the N-nutritionaleffects of AMF might be even more pronounced. This is thefirst experiment conducted under controlled conditions thatdemonstrates that altered atmospheric pCO2 may induceshifts in AMF composition with consequences for the mycor-rhizal symbiosis.
Acknowledgements
We would like to thank Sonja Müller, Lucinda Palma,Yolanda Callau and Ionut Popa for technical assistanceand Prof. Josef Nösberger, Prof. Nina Buchmann and Prof.Emmanuel Frossard from the Institute of Plant Sciences(ETH) for kind permission to use their research facilities.Dr Hans-Rudolf Roth and Mr Bruno Tona from the statisticalconsulting service at ETH Zürich assisted in designing theexperiment, and Dr Dieter Ramseier, PD Dr Andreas Lüscherand Dr Matthias Suter helped with statistical analyses.Comments by Prof. Ian R. Sanders, Prof. Alistair H. Fitter, theEditor, and Dr Jan Jansa helped to improve the presentationof our findings and are gratefully acknowledged. Financialsupport for this research was provided by a research grantfrom the Federal Institute of Technology (ETH) Zürich andby funds supporting the Swiss FACE experiment.
References
Azcon R, Ruiz-Lozano J, Rodriguez R. 2001. Differential contribution of arbuscular mycorrhizal fungi to plant nitrate uptake (15N) under increasing N supply to the soil. Canadian Journal of Botany 79: 1175–1180.
Bago B, Cano C, Azcon-Aguilar C, Samson J, Coughlan AP, Piche Y. 2004. Differential morphogenesis of the extraradical mycelium of an arbuscular mycorrhizal fungus grown monoxenically on spatially heterogeneous culture media. Mycologia 96: 452–462.
Bazzaz FA. 1990. The response of natural ecosystems to the rising global CO2 levels. Annual Review of Ecology and Systematics 21: 167–196.
Bethlenfalvay GJ, Brown MS, Stafford AE. 1985. Glycine–Glomus–Rhizobium symbiosis. 2. Antagonistic effects between mycorrhizal colonization and nodulation. Plant Physiology 79: 1054–1058.
Breuninger M, Trujillo CG, Serrano E, Fischer R, Requena N. 2004. Different nitrogen sources modulate activity but not expression of glutamine synthetase in arbuscular mycorrhizal fungi. Fungal Genetics and Biology 41: 542–552.
Catford JG, Staehelin C, Lerat S, Piche Y, Vierheilig H. 2003. Suppression of arbuscular mycorrhizal colonization and nodulation in split-root systems of alfalfa after pre- inoculation and treatment with Nod factors. Journal of Experimental Botany 54: 1481–1487.
Chakraborty S, Datta S. 2003. How will plant pathogens adapt to host plant resistance at elevated CO2 under a changing climate? New Phytologist 159: 733–742.
Egerton-Warburton LM, Allen EB. 2000. Shifts in arbuscular mycorrhizal communities along an anthropogenic nitrogen deposition gradient. Ecological Applications 10: 484–496.
Fischer BU, Frehner M, Hebeisen T, Zanetti S, Stadelmann F, Lüscher A, Hartwig UA, Hendrey GR, Blum H, Nösberger J. 1997. Source–sink relations in Lolium perenne L. as reflected by carbohydrate concentrations in leaves and pseudo-stems during regrowth in a free air carbon dioxide enrichment (FACE) experiment. Plant, Cell & Environment 20: 945–952.
Fitter AH, Heinemeyer A, Staddon PL. 2000. The impact of elevated CO2 and global climate change on arbuscular mycorrhizas: a mycocentric approach. New Phytologist 147: 179–187.
Gamper H, Peter M, Jansa J, Lüscher A, Hartwig UA, Leuchtmann A. 2004. Arbuscular mycorrhizal fungi benefit from 7 years of free air CO2 enrichment in well-fertilized grass and legume monocultures. Global Change Biology 10: 189–199.
Gilmore AE. 1968. Phycomycetous mycorrhizal organisms collected by open-pot culture methods. Hilgardia 39: 87–105.
Goddard MR, Bradford MA. 2003. The adaptive response of a natural microbial population to carbon- and nitrogen-limitation. Ecology Letters 6: 594–598.
Graham JH, Drouillard DL, Hodge NC. 1996. Carbon economy of sour orange in response to different Glomus spp. Tree Physiology 16: 1023–1029.
Hart MM, Klironomos JN. 2002. Diversity of arbuscular mycorrhizal fungi and ecosystem functioning. In: van der Heijden MGA, Sanders IR, eds. Mycorrhizal ecology. Heidelberg, Germany: Springer, 225–239.
Hart MM, Reader RJ. 2002. Taxonomic basis for variation in the colonization strategy of arbuscular mycorrhizal fungi. New Phytologist 153: 335–344.
Hartwig UA. 1998. The regulation of symbiotic N2 fixation: a conceptual model of N feedback from the ecosystem to the gene expression level. Perspectives in Plant Ecology, Evolution and Systematics 1: 92–120.
Hartwig UA, Heim I, Lüscher A, Nösberger J. 1994. The nitrogen-sink is involved in the regulation of nitrogenase activity in white clover after defoliation. Physiologia Plantarum 92: 375–382.
Hawkins HJ, Johansen A, George E. 2000. Uptake and transport of organic and inorganic nitrogen by arbuscular mycorrhizal fungi. Plant and Soil 226: 275–285.
Hoagland DR, Arnon DI. 1938. The water-culture method for growing plants without soil. California Agriculture Experiment Station Circular 347: 1–39.
Ianson DC, Linderman RG. 1993. Variation in the response on nodulating pigeonpea (Cajanus cajan) to different isolates of mycorrhizal fungi. Symbiosis 15: 105–119.
Jakobsen I, Abbott LK, Robson AD. 1992. External hyphae of vesicular arbuscular mycorrhizal fungi associated with Trifolium subterraneum L. 1. Spread of hyphae and phosphorus inflow into roots. New Phytologist 120: 371–380.
Jakobsen I, Rosendahl L. 1990. Carbon flow into soil and external hyphae from roots of mycorrhizal cucumber plants. New Phytologist 115: 77–83.
Jarrell WM, Beverly RB. 1981. The dilution effect in plant nutrition studies. Advances in Agronomy 34: 197–224.

New Phytologist (2005) 167: 531–542 www.newphytologist.org © New Phytologist (2005)
Research542
Johnson NC. 1993. Can fertilization of soil select less mutualistic mycorrhizae? Ecological Application 3: 749–757.
Johnson NC, Graham JH, Smith FA. 1997. Functioning of mycorrhizal associations along the mutualism–parasitism continuum. New Phytologist 135: 575–586.
Klironomos JN, Ursic M, Rillig M, Allen MF. 1998. Interspecific differences in the response of arbuscular mycorrhizal fungi to Artemisia tridentata grown under elevated atmospheric CO2. New Phytologist 138: 599–605.
Koch AM, Kuhn G, Fontanillas P, Fumagalli L, Goudet I, Sanders IR. 2004. High genetic variability and low local diversity in a population of arbuscular mycorrhizal fungi. Proceedings of the National Academy of Sciences, USA 101: 2369–2374.
Koide RT, Li M. 1989. Appropriate controls for vesicular arbuscular mycorrhizal research. New Phytologist 111: 35–44.
McAuliffe C, Chamblee DS, Uribe-Arango H, Woodhouse WW. 1958. Influence of inorganic nitrogen on nitrogen fixation by legumes as revealed by 15N. Agronomy Journal 50: 334–337.
McGonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA. 1990. A new method which gives an objective measure of colonization of roots by vesicular arbuscular mycorrhizal fungi. New Phytologist 115: 495–501.
Munkvold L, Kjoller R, Vestberg M, Rosendahl S, Jakobsen I. 2004. High functional diversity within species of arbuscular mycorrhizal fungi. New Phytologist 164: 357–364.
Murphy J, Riley JP. 1962. A modified single solution method for the determination of phosphate in natural waters. Analytica Chimica Acta 27: 31–36.
Murphy PM. 1986. Effect of light and atmospheric carbon dioxide concentration on nitrogen fixation by herbage legumes. Plant and Soil 95: 399–409.
Newman EI. 1966. A method of estimating the total length of root in a sample. Journal of Applied Ecology 3: 139–145.
Olesniewicz KS, Thomas RB. 1999. Effects of mycorrhizal colonization on biomass production and nitrogen fixation of black locust (Robinia pseudoacacia) seedlings grown under elevated atmospheric carbon dioxide. New Phytologist 142: 133–140.
Pendall E, Mosier AR, Morgan JA. 2004. Rhizodeposition stimulated by elevated CO2 in a semiarid grassland. New Phytologist 162: 447–458.
Peng S, Eissenstat DM, Graham JH, Williams K, Hodge NC. 1993. Growth depression in mycorrhizal citrus at high phosphorus supply. Plant Physiology 101: 1063–1071.
Phillips JM, Hayman DS. 1970. Improved procedures for clearing roots and staining parasitic and vesicular arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society 55: 158–160.
Richter M, Hartwig UA, Frossard E, Nösberger J, Cadisch G. 2003. Gross fluxes of nitrogen in grassland soil exposed to elevated atmospheric pCO2 for seven years. Soil Biology and Biochemistry 35: 1325–1335.
Rillig M, Treseder KK, Allen MF. 2002. Global change and mycorrhizal fungi. In: van der Heijden MGA, Sanders IR, eds. Mycorrhizal ecology. Heidelberg, Germany: Springer, 135–153.
Rillig MC, Allen MF. 1999. What is the role of arbuscular mycorrhizal fungi in plant-to-ecosystem responses to elevated atmospheric CO2? Mycorrhiza 9: 1–8.
Sanders IR. 2002. Ecology and evolution of multigenomic arbuscular mycorrhizal fungi. American Naturalist 160: S128–S141.
Sanders IR, Streitwolf-Engel R, van der Heijden MGA, Boller T, Wiemken A. 1998. Increased allocation to external hyphae of arbuscular mycorrhizal fungi under CO2 enrichment. Oecologia 117: 496–503.
Schneider MK, Lüscher A, Richter M, Aeschlimann U, Hartwig UA, Blum H, Frossard E, Nösberger J. 2004. Ten years of free-air CO2 enrichment altered the mobilization of N from soil in Lolium perenne L. swards. Global Change Biology 10: 1377–1388.
Schulze J. 2004. How are nitrogen fixation rates regulated in legumes? Journal of Plant Nutrition and Soil Science 167: 125–137.
Smith SE, Read DJ. 1997. Mycorrhizal symbioses, 2nd edn. London, UK: Academic Press.
Smith SE, Smith FA, Jakobsen I. 2004. Functional diversity in arbuscular mycorrhizal (AM) symbioses: the contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake. New Phytologist 162: 511–524.
Smith SE, St John BJ, Smith FA, Nicholas DJD. 1985. Activity of glutamine synthetase and glutamate dehydrogenase in Trifolium subterraneum L. and Allium cepa L. – effects of mycorrhizal infection and phosphate nutrition. New Phytologist 99: 211–227.
Soussana JF, Casella E, Loiseau P. 1996. Long-term effects of CO2 enrichment and temperature increase on a temperate grass sward. 2. Plant nitrogen budgets and root fraction. Plant and Soil 182: 101–114.
Staddon PL, Fitter AH. 1998. Does elevated atmospheric carbon dioxide affect arbuscular mycorrhizas? Trends in Ecology and Evolution 13: 455–458.
Staddon PL, Gregersen R, Jakobsen I. 2004. The response of two Glomus mycorrhizal fungi and a fine endophyte to elevated atmospheric CO2, soil warming and drought. Global Change Biology 10: 1909–1921.
Suter D, Frehner M, Fischer BU, Nösberger J, Lüscher A. 2002. Elevated CO2 increases carbon allocation to the roots of Lolium perenne under free-air CO2 enrichment but not in a controlled environment. New Phytologist 154: 65–75.
Sylvia DM. 1992. Quantification of external hyphae of vesicular arbuscular mycorrhizal fungi. Methods in Microbiology 24: 53–65.
Tennant D. 1975. A test of a modified line intersect method of estimating root length. Journal of Ecology 63: 995–1001.
Tobar R, Azcon R, Barea JM. 1994. Improved nitrogen uptake and transport from 15N labeled nitrate by external hyphae of arbuscular mycorrhiza under water stressed conditions. New Phytologist 126: 119–122.
Treseder KK. 2004. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytologist 164: 347–355.
Wright DP, Scholes JD, Read DJ. 1998. Effects of VA mycorrhizal colonization on photosynthesis and biomass production of Trifolium repens L. Plant, Cell & Environment 21: 209–216.
Xavier LJC, Germida JJ. 2002. Response of lentil under controlled conditions to co-inoculation with arbuscular mycorrhizal fungi and rhizobia varying in efficacy. Soil Biology and Biochemistry 34: 181–188.
Zak DR, Pregitzer KS, King JS, Holmes WE. 2000. Elevated atmospheric CO2, fine roots and the response of soil microorganisms: a review and hypothesis. New Phytologist 147: 201–222.
Zanetti S, Hartwig UA, van Kessel C, Lüscher A, Hebeisen T, Frehner M, Fischer BU, Hendrey GR, Blum H, Nösberger J. 1997. Does nitrogen nutrition restrict the CO2 response of fertile grassland lacking legumes? Oecologia 112: 17–25.
Zanetti S, Hartwig UA, Lüscher A, Hebeisen T, Frehner M, Fischer BU, Hendrey GR, Blum H, Nösberger J. 1996. Stimulation of symbiotic N2 fixation in Trifolium repens L. under elevated atmospheric pCO2 in a grassland ecosystem. Plant Physiology 112: 575–583.
Zar JH. 1999. Biostatistical analysis, 4th edn. Upper Saddle River, NJ, USA: Prentice Hall.




















