
Mycorrhizas and nutrient cycling in ecosystems - a journey towards relevance
18
© New Phytologist (2003) 157: 475 – 492 www.newphytologist.com 475 Review Blackwell Science, Ltd Research review Mycorrhizas and nutrient cycling in ecosystems – a journey towards relevance? D. J. Read 1 and J. Perez-Moreno 2 1 Department of Animal & Plant Science, University of Sheffield, Sheffield, S10 2TN, UK; 2 Colegio de Postgraduados, Microbiologia Edafologia-a-Irenat, Montecillo, Texcoco, CP 56320, Mexico Summary Progress towards understanding the extent to which mycorrhizal fungi are involved in the mobilization of nitrogen (N) and phosphorus (P) from natural substrates is reviewed here. While mycorrhiza research has emphasized the role of the symbiosis in facilitation of capture of these nutrients in ionic form, attention has shifted since the mid-1980s to analysing the mycorrhizal fungal abilities to release N and P from the detrital materials of microbial faunal and plant origins, which are the primary sources of these elements in terrestrial ecosystems. Ericoid, and some ectomycorrhizal fungi have the potential to be directly involved in attack both on structural polymers, which may render nutrients inaccessible, and in mobilization of N and P from the organic polymers in which they are sequestered. The advantages to the plant of achieving intervention in the microbial mobilization–immobilization cycles are stressed. While the new approaches may initially lack the precision achieved in studies of readily characterized ionic forms of N and P, they do provide insights of greater ecological relevance. The results support the hypothesis that selection has favoured ericoid and ectomycorrhizal systems with well developed saprotrophic capabilities in those ecosystems characterized by retention of N and P as organic complexes in the soil. The need for further investigation of the abilities of arbuscular mycorrhizal fungi to intervene in nutrient mobilization processes is stressed. © New Phytologist (2003) 157 : 475–492 Author for correspondence: D J Read Tel: +44(0)114 2224318 Fax: +44(0)114 2220002 Email: d.j.read@sheffield.ac.uk Received: 3 September 2002 Accepted: 9 December 2002 Key words: ericoid mycorrhiza, ectomycorrhiza, arbuscular mycorrhiza, nitrogen nutrition, phosphorus nutrition, ecosystem. Introduction More than 10 years have elapsed since it was proposed (Read, 1991) that because plants with ericoid (ERM), ecto-(ECM) and arbuscular (AM) mycorrhizas typically predominate under distinctive soil conditions, their respective fungal associates might, in each case, be making contrasting types of contribution to plant nutrition. At the heart of the original proposition was the notion that at the global scale, shifts from lower to higher latitudes or altitudes, through their effects directly upon climate and indirectly upon decomposition, led to fundamental changes in the nature of the soil as a nutrient source for plants. On the basis that these gradients involved progressive increases in the extent to which the major nutrients nitrogen (N) and phosphorus (P) were sequestered in organic forms which were unavailable to autotrophs, it was
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Mycorrhizas and nutrient cycling in ecosystems - a journey towards relevance

©
New Phytologist
(2003)
157
: 475–492
www.newphytologist.com
475
Review
Blackwell Science, Ltd
Research review
Mycorrhizas and nutrient cycling in ecosystems – a journey towards
relevance?
D. J. Read
1
and J. Perez-Moreno
2
1
Department of Animal & Plant Science, University of Sheffield, Sheffield, S10 2TN, UK;
2
Colegio de Postgraduados, Microbiologia Edafologia-a-Irenat, Montecillo, Texcoco, CP 56320, Mexico
Summary
Progress towards understanding the extent to which mycorrhizal fungi are involvedin the mobilization of nitrogen (N) and phosphorus (P) from natural substrates isreviewed here. While mycorrhiza research has emphasized the role of the symbiosisin facilitation of capture of these nutrients in ionic form, attention has shifted sincethe mid-1980s to analysing the mycorrhizal fungal abilities to release N and P fromthe detrital materials of microbial faunal and plant origins, which are the primarysources of these elements in terrestrial ecosystems. Ericoid, and some ectomycorrhizalfungi have the potential to be directly involved in attack both on structural polymers,which may render nutrients inaccessible, and in mobilization of N and P from theorganic polymers in which they are sequestered. The advantages to the plant ofachieving intervention in the microbial mobilization–immobilization cycles are stressed.While the new approaches may initially lack the precision achieved in studies ofreadily characterized ionic forms of N and P, they do provide insights of greaterecological relevance. The results support the hypothesis that selection has favouredericoid and ectomycorrhizal systems with well developed saprotrophic capabilities inthose ecosystems characterized by retention of N and P as organic complexes in thesoil. The need for further investigation of the abilities of arbuscular mycorrhizal fungito intervene in nutrient mobilization processes is stressed.
©
New Phytologist
(2003)
157
: 475–492
Author for correspondence:
D J Read
Tel: +44(0)114 2224318
Fax: +44(0)114 2220002
Email: [email protected]
Received:
3 September 2002
Accepted:
9 December 2002
Key words:
ericoid mycorrhiza, ectomycorrhiza, arbuscular mycorrhiza, nitrogen nutrition, phosphorus nutrition, ecosystem.
Introduction
More than 10 years have elapsed since it was proposed (Read,1991) that because plants with ericoid (ERM), ecto-(ECM)and arbuscular (AM) mycorrhizas typically predominateunder distinctive soil conditions, their respective fungalassociates might, in each case, be making contrasting types ofcontribution to plant nutrition. At the heart of the original
proposition was the notion that at the global scale, shifts fromlower to higher latitudes or altitudes, through their effectsdirectly upon climate and indirectly upon decomposition, ledto fundamental changes in the nature of the soil as a nutrientsource for plants. On the basis that these gradients involvedprogressive increases in the extent to which the majornutrients nitrogen (N) and phosphorus (P) were sequesteredin organic forms which were unavailable to autotrophs, it was

Research review
www.newphytologist.com
©
New Phytologist
(2003)
157
: 475–492
Review476
hypothesised that at any point along its length, naturalselection would have favoured associations between plantsand those fungal mutualists which were capable of unlockingthe two key growth limiting resources.
Soil scientists acknowledge that, even in agricultural soils,mineralisation of N and P involves a sequence of processes forwhich the living microbial biomass provides the enzymes anddead microbial material much of the substrate (Mengel, 1996;Appel & Mengel, 1998). In general, however, they have notappreciated that a large proportion both of the microbialbiomass and necromass of soils in most terrestrial ecosystemsconsists of the mycelium of mycorrhizal fungi (Finlay &Söderström, 1992; Read, 1992; Högberg & Högberg, 2002)or that access to photosynthate from their autotrophic part-ners releases this component of the microbial biomass fromcarbon limitation so providing it with the potential to play amajor role in nutrient mobilisation. The consequences of anability of mycorrhizal fungi to intercede in the processesotherwise leading to release of N and P in ionic form wouldbe considerable, not only in terms of provision for their hostplants of preferential access to the nutrients but also becausethe early intervention in events normally leading to minerali-sation would reduce the downstream supply of these elementsin the ionic forms required by nonmycorrhizal competitors.
Unfortunately, and probably under the influence of theextensive agricultural literature, research carried out throughmuch of the last century emphasised the roles of mycorrhizasin the capture of mineral nutrient ions (see Harley & Smith,1983). The possibility that fungal symbionts might be them-selves involved in direct attack upon nutrient containingorganic polymers, with the exception of the visionary specu-lation of Frank who proposed the ‘Organic Nitrogen Theory’in 1894 (Frank, 1894), was largely ignored until the mid-1980s. As a result a view has emerged which sees two entirelyseparate functional groups of fungi in soil, one, the decom-posers, being alone involved in destruction of organicsubstrates and release of any nutrients contained in them andthe other, the fungal mutualists, which absorb mineral ionsreleased by the decomposition processes.
Research over the last two decades has begun to throw con-siderable doubt both on the validity of any such functionalseparation between decomposers and mutualists, and uponthe view that mycorrhizal fungi have access only to mineralnutrient ions. In so doing it has lent further support to thehypothesis that the different mycorrhizal types are selectedaccording to their abilities to mobilise essential nutrients frompolymeric sources. The main distinguishing feature of thisperiod of research is the move away from reductionistapproaches which consider only simple mineral ions as pos-sible sources of the major elements, to an emphasis upon theabilities of ERM, ECM and AM fungi to exploit N and P con-tained in substrates representative of those occurring in theirdistinctive natural environments. Where model compoundshave the potential to provide insights these have been used in
the new generation of experiments, but increasingly materialsfrom the rooting zones of the plants themselves have beenemployed. To the extent that some these of have not receivedfull physico-chemical characterisation, the novel approachesmay initially lack some of the precision obtainable whenemploying simple mineral ions, but it is the contention here(Fig. 1), that the gains in terms of ecological relevance willoutweigh the disadvantages.
In this paper the nutritional status of the three major typesof mycorrhizal symbiosis is reassessed and factors determiningtheir separate occurrence or coexistence are considered in thelight of the new observational and experimental evidence.
Nutrient cycling by ericoid mycorrhizal fungi in heathland ecosystems
Heathlands are characterised on the basis of the pre-dominance of dwarf shrubs of the order Ericales and of theprevalence of acidic raw humus soils in which mineralisationprocesses are inhibited to the extent that the essential nutrientsN and P are present almost exclusively in combination withorganic residues. While a number of structural attributesof above-ground tissues, in particular sclerophylly and leafretention, contribute to nutrient conservation in ericaceousplants, there is a accumulating evidence that it is the ericoidmycorrhizal symbiosis which provides the key to releasingsuch resources from detrital materials. The epidermal cellsof the ‘hair roots’ of the dominant members of these com-munities are extensively penetrated by hyphae of a smallnumber of distinctive ascomycetous fungi. Amongst these themost widely occurring and most extensively investigated arestrains of the
Hymenoscyphus ericae-Scytalidium vaccinii
complex (Helotiales) (Egger & Sigler, 1993; Read, 1996) and of
Oidiodendron maius
(Onygenales) (Hambleton
et al
., 1998).The mycorrhizal cells, and their emanating hyphae, occupy aninterface between the potential nutrient resources of the rawhumus within which the roots proliferate and the body of theplant. The fungi forming ericoid mycorrhiza can be readilycultured enabling experimental studies of their abilities todegrade organic polymers. These have confirmed that ericoidfungi have considerable saprotrophic capabilities, which, ifexpressed in soil, would be expected to facilitate nutrientmobilisation from the residues exploited by the mycorrhizalroots and mycelium. For the most part, such studies have usedmodel compounds and so describe only the potential of themycelium for
in situ
mobilisation. They are none the lessimportant because they emphasise the biochemical versatilitywhich is accessible to the ericaceous plant as a result of itsselective association with this group of fungal symbionts.
Mobilisation of N and P from ‘model’ compounds
Two classes of polymer degrading enzymes can be recognised,one which provides the potential to cleave those organic

Research review
©
New Phytologist
(2003)
157
: 475–492
www.newphytologist.com
Review 477
molecules such as lignins, polyphenols and tannins whichmay protect or precipitate essential nutrients (Table 1) andanother which facilitates direct attack upon the nutrient-containing molecules themselves. The former containslignases and polyphenol oxidases which are of particularinterest because they would be expected to contribute in amajor way to decomposition as well as to plant nutrition.Despite the demonstration (Haselwandter
et al
., 1990) usingrespirometric methods, that
H. ericae
was capable of ring-cleavage of aromatic constituents of lignin, subsequentsearches (Bending & Read, 1996b, 1997; Burke & Cairney,1998) have failed to find evidence for the production of theenzymes lignin (LiP) and manganese (MnP) peroxidase which
would be required to enable direct attack, of the kind achievedby ‘white rot’ fungi, upon the aromatic structures which makeup the polymer. However, recognition that
H. ericae
releaseshydrogen peroxide (Bending & Read, 1997) and hydroxylradicals (Burke & Cairney, 1998) into culture media confirmsthat the fungus may contribute to a form of lignin degrad-ation, similar to that seen in ‘brown rot’ fungi, in which theradicals facilitate fragmentation of the lignin polymer bydemethoxylation and side chain oxidation (Burke & Cairney,1998; Cairney & Burke, 1998). It is of interest that inaddition to their impacts upon lignin, hydroxyl radicals havebeen implicated in the degradation of cellulose by ‘brown rot’fungi ( Joseleau
et al
., 1994).
Fig. 1 Until recently almost all studies of the role of mycorrhizas in plant nutrition examined simple mineral ions as the sources of the key elements nitrogen (N) and phosphorus (P) (thick arrow lower left). While the processes of uptake of such elements can be observed in experimental systems with great precision, their primary repositories in most ecosystems are the organic residues of the soil microflora and fauna and of the plants themselves. It has been the convention to assume that N and P locked into organic macromolecules of these kinds was accessible only to specialist decomposers. However, new research using model polymers and natural organic substrates (thin arrows centre and top right) increasingly suggests that some classes of mycorrhizal fungi can mobilise these elements from the primary sources or their intermediates. By breaking into the mineralisation pathway and providing their autotrophic partners with access to the nutrients before they are re-immobilised in the microflora, these fungi would enable considerable increases in the efficiency with which plants are able to acquire and recycle N and P. The chemistry of primary nutrient sources in soils is often poorly characterised and as a consequence there may initially be a loss of precision in the new research, but it is argued that the advantages gained in terms of ecological relevance (bottom right) outweigh the disadvantages. Ultimately, identification of those compounds and processes which play key roles in the trophic cascades occurring in the ‘black box’ should provide the impetus necessary to ensure their chemical evaluation. This will enable the scientifically desirable balance between precision and relevance to be restored. Mycorrhizal fungi are increasingly seen to have the potential to be the drivers of nutrient mobilisation processes in some ecosystems.

Research review
www.newphytologist.com
©
New Phytologist
(2003)
157
: 475–492
Review478
Polyphenolic compounds also characteristically reach highconcentrations in the heathland soils ( Jalal & Read, 1983).Enrichment of their surface horizons by residues rich inphenolic compounds which can reach over 30% of tissue dryweight in plants such as
Calluna vulgaris
( Jalal
et al
., 1982), isan inevitable consequence of the accumulation of these terti-ary metabolites in ericaceous plants. It has implications bothfor pedogenic processes and for the plants which forage fornutrients within such soils. A key to understanding the nutri-tional contribution of ericoid mycorrhizas in these phenolrich environments may lie in the now proven ability of theirfungal partners not only to use many monomeric phenoliccompounds as carbon sources (Leake & Read, 1989) but alsoto release the enzymes laccase and catechol oxidase (Bending& Read, 1996a,b, 1997) that are involved in the degradationof hydrolysable polyphenols. These attributes, which aredeveloped far more extensively in ericoid than in ectomycor-rhizal fungi so far examined (Bending & Read, 1996a,b), arenow thought to be of significance from the perspectives bothof raw humus formation and plant nutrition. When thefungus is grown on culture media containing, as sole carbonsource, the model polyphenol tannic acid (Bending & Read,1996a), it utilises the carbon contained in the substrate, andreleases as a residue, dark-coloured reactive quinone-likecompounds which are thought to be the precursors of therecalcitrant humic and fulvic acid polymers that form the bulkof raw humus (Stevenson, 1982).
From the nutritional standpoint it is the propensity ofphenolic compounds to form complexes with substrates con-taining the major elements nitrogen and phosphorus which islikely to be of greatest significance. Here, the abilities of
H.ericae
to assimilate individual phenolic monomers thus reduc-ing the extent of complexation and, by cleaving polymers, tosecure release of nutrients coprecipitated with polyphenols
must both be considered. Bending & Read (1996b) demon-strated that, in addition to degrading tannic acid-protein pre-cipitates by releasing polyphenol oxidase,
H. ericae
was able toacquire nitrogen from these complexes through the expressionof extracellular acid protease activity. This property, which thefungus shares with
Oidiodendron
, was not shown by any offive ectomycorrhizal fungi examined. Since it became recog-nised (Bajwa
et al
., 1985) that ERM fungi could mobilise theN contained in model proteins and assimilate the releasedamino-acids, it has been revealed that representatives of otherfunctional types of plants which are often associated withericoid shrubs in tundra and boreal forest environments,notably in the Cyperaceae (Chapin
et al
., 1993; Kielland, 1994;Raab
et al
., 1996) and Gramineae (Näsholm
et al
., 1998;Näsholm & Persson, 2001) could readily assimilate amino-acid N. Such observations indicate that the ability to absorbamino acids and hence to bypass the N mineralisation processmay be widespread in organic soils. However, they should notbe allowed to obscure the essential ecophysiological distinc-tion between the capacity for amino-acid scavenging whichis an attribute of some nonmycorrhizal as well as mycorrhizalplants, and the ability conferred upon ERM (and someECM – see below) plants by their mycorrhizal symbionts,to carry out direct attack upon the sources of soil amino-Ncompounds, which are the proteins themselves. It has beenproposed (Read, 1996) that the overwhelming predominanceof the ericoid mycorrhizal functional group in heathlandecosystems may provide some indication of the relativeeffectiveness of the two strategies for N acquisition.
Mobilisation of N and P from ‘natural’ substrates
While experiments using model compounds as surrogate sub-strates can be enlightening there is clearly a need to consider
Table 1 Extra cellular enzymes, known to be produced by the ericoid mycorrhizal fungus Hymenoscyp us ericae, which would be expected to provide the ability to degrade structural components of plant litter, thereby contributing to decomposition processes and to ‘unmasking’ of nutrients
References
Plant Cell Wall degradation Pectin Polygalacturonase Perotto et al. (1997), Peretto et al. (1993)Cellulose Cellulase Varma & Bonfante (1994), Burke & Cairney (1997a)Cellobiose Cellobiohydrolase Bending & Read (1986a), Burke & Cairney (1997a),
Burke & Cairney (1998) Cairney & Burke, 1998Hemicellulose Xylanase Bending & Read (1996a), Burke & Cairney (1997a,b)
β Xylosidase Burke & Cairney (1997a)β D-mannosidase “β D-galactosidase “β L-arabinosidase “β 1;3 glucanase Varma & Bonfante (1994)
Oxidation of Phenolic Polyphenols Polyphenol oxidase Bending & Read (1996b), Bending &Read (1997)Acids and Tannins Laccase
Catechol oxidase Burke & Cairney (1998)Hydrolysis of Lignin Lignin Lignase Haselwandter et al. (1990)
Italics indicate results based upon indirect methods of observation. For older literature see Leake & Read (1997).

Research review
©
New Phytologist
(2003)
157
: 475–492
www.newphytologist.com
Review 479
naturally occurring materials. In species-poor heathlandsystems the residues reaching the soil are derived largely fromthe ericaceous plants themselves and their fungal symbionts.The shoot residues form a distinct litter (L) layer at the surfacewhereas the fine root and fungal residues are concentratedimmediately below this in the fermentation (F) layer in whichthey selectively proliferate while alive. Ideally, these materialswould be harvested and used as substrates in comparative ana-lyses of the growth of mycorrhizal and nonmycorrhizal plantsbut because they carry inoculum of ERM fungi, sterilisationis required in order to maintain control plants in thenonmycorrhizal condition. Since all forms of sterilisation sofar employed on these organic materials have produced changesin their quality such approaches have little ecological appeal.
Faced with these difficulties advances towards relevancehave been made by a multifaceted approach using the micro-bial and plant tissues themselves, produced aseptically, andkilled naturally by air drying, to provide ‘necromass’, this thenbeing supplied as the sole course of these elements to axeni-cally grown M and NM plants (Fig. 1).
Analysis of the nutritional role of the fungal cell wallpolymer chitin, which is known to be an important potentialN source in raw humus soils (Kerley & Read, 1998) has beenachieved using a combination of these approaches. It was firstdemonstrated (Leake & Read, 1990a; Mitchell
et al
., 1992),using crustacean chitin as a model compound, that
H. ericae
could cleave the polymer to its constitutent subunits N-acetylglucosamine and N-acetyl galactosamine, both of which werereadily assimilated by the fungus. A significant proportion ofthe N so acquired was subsequently shown (Kerley & Read,1995) to be transferred to aseptically grown mycorrhizalplants of
Vaccinium macrocarpon
. Natural sources of thepolymer, in the form of mycelial necromass of
H. ericae
itself,were later used as substrates (Kerley & Read, 1997). Hyphaeof the fungus were supplied to M and NM plants either aspurified cell wall fractions or in the unfractionated form assole sources of N. In the M condition plants gained access tothe N contained in both pure and unfractionated wall frac-tions, their yields and N contents on the former being almostas high as those on the latter. The ability to effectively recycleN originally sequestered in the fungus will contribute to a less-ening of competition between the partners in the symbiosis.Similar mycorrhiza-dependent pathways for nitrogen transferwere demonstrated (Kerley & Read, 1998) using asepticallyproduced shoot and root necromass of the ericaceous plantsthemselves.
Clearly, caution must be exercised when interpreting resultsobtained from use of aseptically produced necromass as nutri-tional substrates. As they have not been subject to the biolog-ical and physico-chemical processes associated with normalsenescence and decomposition, these materials are not trulyrepresentative of those found in the litter layer. However, assurrogates for such materials they can contribute new insightswhen used as part of a broader screening of natural substrates.
Relatively little attention has been paid to the acquisition ofP from polymeric sources by ericoid plants. However, bothDNA in purified form (Leake & Miles, 1996) and entirenuclei (Myers & Leake, 1996) have been shown to constituteuseable sources of the element for
V. macrocarpon
providingthe plants are in the mycorrhizal condition. These polymerswere selected as surrogates for the organic P sources whichmust be continuously released into soil as the tissues ofmicrobial and plant populations senesce. In these casesmobilisation of P is dependent upon production by
H. ericae
of phosphodiesterases (Leake & Read, 1997).Collectively, these studies confirm, using model compounds,
naturally occurring components of heathland ecosystems, ortheir intermediates, that ERM colonisation will enable inter-vention in the immobilisation processes of soil N and P cyclesand facilitate a direct pathway for transfer of these key growthlimiting elements to the autotrophs.
Nutrient cycling by ectomycorrhizal fungi in boreal and temperate forest ecosystems
It is a prerequisite of any treatment of the nutritionalcapabilities of ECM’s to recognise that over 5000 species offungi are probably capable of forming symbioses of this type(Molina
et al
., 1992), and that only a fraction of this numberare readily culturable and physiologically characterised.Hence if we generalise we do so on the basis of knowledge ofa small number of unfastidious species that can be easilymanipulated under laboratory conditions. Since even these,which are likely to be ‘ruderal’ strategists, have been shown toexpress a range of ‘decomposer’ capabilities (Table 2) there isthe likelihood that the full potential of this functional groupof organisms is greatly underestimated. Awareness of thislimitation has increased in recent times in particular becauseDNA fingerprinting methods (White
et al
., 1990; Gardes &Bruns, 1993, 1996; Gardes
et al
., 1991) have revealed theimportance of groups of fungi which have been hithertooverlooked because they produce inconspicuous hypogeousor resupinate fruit bodies. These can occupy a large propor-tion of the ECM root population in boreo-temerate forests(Baar
et al
., 1999; Erland & Taylor, 1999; Kõljalg
et al
., 2000;Dahlberg, 2001). Amongst the basidiomycetes, members ofthe genera
Tomentella
(Kõljalg, 1996),
Tomentellopsis
(Thelephoraceae) (Kõljalg
et al
., 2000) and
Tylospora
(Corticiaceae) (Taylor
et al
., 2000) are prominent. The newfindings are challenging, nor least because many of the fungiinvolved have hitherto been regarded as decomposers of woodydebris. If these organisms expressed the ability to degradestructural and nutritional polymers when in the mycorrhizalcondition, the conventional distinction between decomposersand symbionts as identifiable functional groups would be asblurred in ECM as they are in the ERM. Some molecularphylogenetic studies of ECM fungi are already pointing inthis direction (Hibbett
et al
., 2000). Clearly there is an urgent

Research review
www.newphytologist.com
©
New Phytologist
(2003)
157
: 475–492
Review480
need to identify the roles played by these recalcitrant fungi inthe nutrient economies of forest systems. It is to be expectedthat in addition to discovery of distinctive hydrolyticcapabilities in some of these organisms, there may be otherswhich are found to lack these attributes, so conforming moreclosely to the conventional view of ECM fungi as being largelynonsaprotrophic.
An emerging possible solution to the problem of uncultur-ability is to extract and sequence DNA from dried basidiomesin the search for the genes which encode key enzymes. Usingthis approach, Chen
et al
. (2001) screened for the presence ofLiP and MnP genes in more than 40 ECM fungi. Around68% of the species examined were found to possess at least 1LiP gene and several species including
Piloderma croceum
,
Cortinarius rotundisporus
and
Tylospora fibrillosa
yielded theMnP gene, which had previously been identified in
T. fibril-losa
(Chambers
et al
., 1999). On the basis of such findingsChen
et al
. are justified in drawing attention to the potentialof these fungi for decomposition of lignin in temperate for-ests, but it is only a potential! The need to establish and quan-tify the extent of gene expression is paramount. Timonen &Sen (1998) examined gene expression in identified functionalcomponents of entire mycorrhizal systems associated with
Pinus
sylvestris. They observed differential expression ofisozymes, the activities of some of which, notably polyphenoloxidase and acid phosphatase, were markedly increased in thehyphal fronts of
P. involutus
and
S. bovinus
mycelial systems asthey advanced into forest humus. Observations made underthese more ecologically realistic circumstances have much intheir favour. There is the potential to extend them to recalci-trant fungi some of which can successfully be grown withplants in microcosms even though they are not readily cul-tured
in vitro
(Kõljalg
et al
., 2002). Some caution will still berequired when interpreting the results of such studies since
up-regulation of genes is not necessarily an indication thattheir products will be expressed in the external environment.
Of even greater importance than the essentially qualitativequestions of gene or enzyme presence, are the quantitativeissues relating to the extent of activity of the enzymes in theenvironment. When comparisons are made between levels ofexpression of those exoenzymes likely to be deployed indecomposition of complex organic polymers by ECM and bysaprotrophic fungi, the latter group invariably show greateractivity when the fungi are grown under identical conditions(Maijala
et al
., 1991; Bending & Read, 1996a, 1997; Colpaert& Van Laere, 1996; Colpaert & Van Tichelen, 1996). Thissuggests that in competitive circumstances ECM fungi will berelatively poorly equipped to exploit such substrates. Theirprevalence in specific parts of the soil profile, notably thefermentation horizon (FH) horizon, is likely to reflectpreferential exploitation of substrates of a particular quality asreflected by their C : N ratio (Read, 1991). Substrate special-isation would lead to avoidance of competitive relationshipswith specialist decomposers. The nutrient foraging inter-actions between one such group, the white-root fungi, andECMF are for the first time now being investigated underseminatural circumstances (Lindahl
et al
., 1999; Leake
et al
.,2001, 2002) and more work is required before generalisationsabout their outcomes can be made.
Mobilisation of N and P from ‘model’ compounds
As with studies of ERM, research on the roles of ECM inplant nutrition, have increasingly focussed in recent times onthe ability to mobilise resources from model polymers.Recognition of the proteolytic capabilities of ERMF in themid 1980s led to a search for these properties in ECM systems(Smith & Read, 1997; Chalot & Brun, 1998). Model proteins
Table 2 Extra cellular enzymes, known to be produced by selected ectomycorrhizal fungi, which would be expected to provide some abilities to degrade structural components of plant litter thereby contributing to decomposition processes and to ‘unmasking’ of nutrients. Italics indicate observations based upon gene presence rather than enzyme expression. For older literature see Leake & Read (1997)
Process Substrate Enzymes Reference
Cuticle Degradation Cutin, Lipid, Waxes Fatty Acid Esterase Hutchison (1990b), Caldwell et al. (1991)Plant Cell Wall degradation Pectin Polygalacturonase Hutchison (1990a)
Cellulose Cellulase Maijala et al. (1991), Colpaert & van Laere (1996)Cellobiose Cellobiohydrolase Burke & Cairney (1998)Hemicellulose Xylanase Cao & Crawford (1993), Terashita et al. (1995),
Cairney & Burke (1996b)Oxidation of Phenolic Monophenols Tyrosinase Hutchison (1990b)Acids and Tannins Polyphenols Polyphenol oxidase Bending & Read (1997), Colpaert & van Laere
(1996), Günther et al. (1998)Peroxidase Bending & Read (1997), Cairney & Burke
(1994), Griffiths & Caldwell (1992)Laccase Hutchison (1990b), Kanunfre & Zancan (1998)
Hydrolysis of Lignin Lignin Manganese peroxidase Chambers et al. (1999), Chen et al. (2001)Lignin Peroxidase Chen et al. (2001)
Italics indicate results based upon indirect methods of observation.
Research review
©
New Phytologist
(2003)
157
: 475–492
www.newphytologist.com
Review 481
have since been extensively used (Leake & Read, 1990b,Finlay
et al
., 1992; Ryan & Alexander, 1992; Turnbull
et al
.,1995) as substrates to determine the potential of ECM fungiand their plant associates to mobilise N from polymeric macro-molecules. The main extracellular acid carboxy proteinaseinvolved in the mobilisation was purified and characterised (El-Badaoui & Botton, 1989; Zhu
et al
., 1990). While the ECMfungi so far examined have generally demonstrated lower levelsof proteolytic activity than those seen in ERM fungi, andsome, the so-called ‘nonprotein’ fungi express no such activity,many have been shown readily to cleave the protein and tofacilitate transfer of the N contained in it, to mycorrhizal plants.Some are also able to gain access to N contained in proteinwhich had been precipitated with phenolic acid (Bending &Read, 1996a) but again not to the extent seen in ERM.
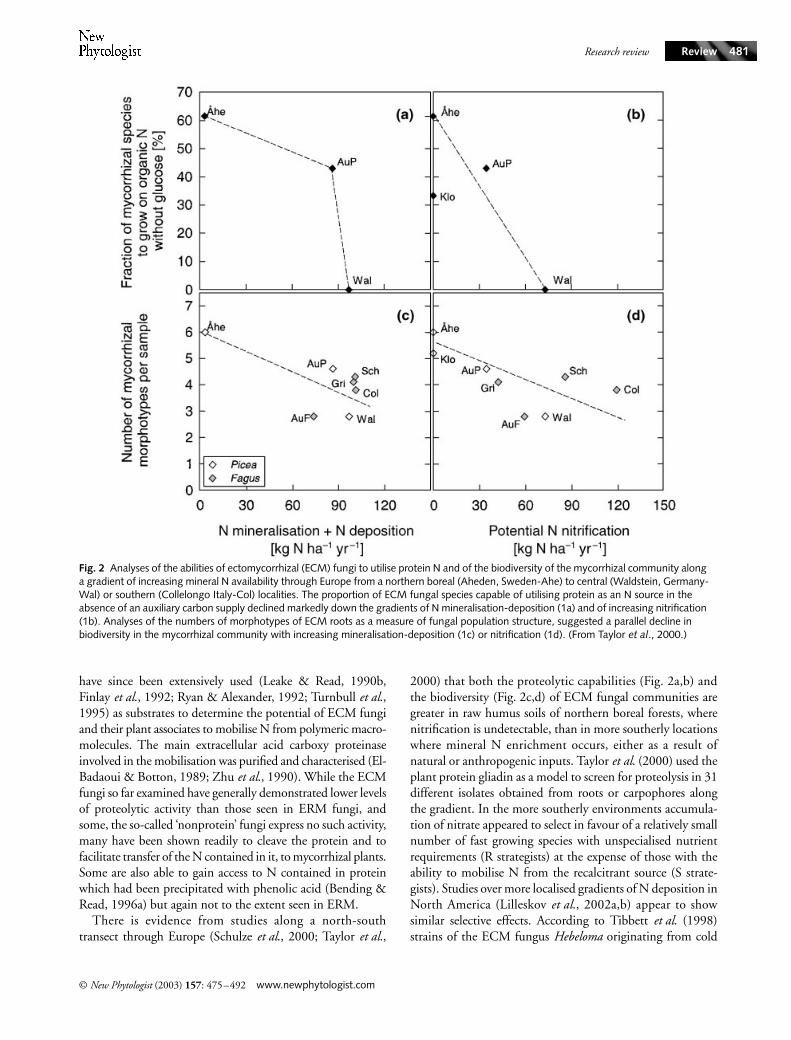
There is evidence from studies along a north-southtransect through Europe (Schulze
et al
., 2000; Taylor et al.,
2000) that both the proteolytic capabilities (Fig. 2a,b) andthe biodiversity (Fig. 2c,d) of ECM fungal communities aregreater in raw humus soils of northern boreal forests, wherenitrification is undetectable, than in more southerly locationswhere mineral N enrichment occurs, either as a result ofnatural or anthropogenic inputs. Taylor et al. (2000) used theplant protein gliadin as a model to screen for proteolysis in 31different isolates obtained from roots or carpophores alongthe gradient. In the more southerly environments accumula-tion of nitrate appeared to select in favour of a relatively smallnumber of fast growing species with unspecialised nutrientrequirements (R strategists) at the expense of those with theability to mobilise N from the recalcitrant source (S strate-gists). Studies over more localised gradients of N deposition inNorth America (Lilleskov et al., 2002a,b) appear to showsimilar selective effects. According to Tibbett et al. (1998)strains of the ECM fungus Hebeloma originating from cold
Fig. 2 Analyses of the abilities of ectomycorrhizal (ECM) fungi to utilise protein N and of the biodiversity of the mycorrhizal community along a gradient of increasing mineral N availability through Europe from a northern boreal (Aheden, Sweden-Ahe) to central (Waldstein, Germany-Wal) or southern (Collelongo Italy-Col) localities. The proportion of ECM fungal species capable of utilising protein as an N source in the absence of an auxiliary carbon supply declined markedly down the gradients of N mineralisation-deposition (1a) and of increasing nitrification (1b). Analyses of the numbers of morphotypes of ECM roots as a measure of fungal population structure, suggested a parallel decline in biodiversity in the mycorrhizal community with increasing mineralisation-deposition (1c) or nitrification (1d). (From Taylor et al., 2000.)

Research review
www.newphytologist.com © New Phytologist (2003) 157: 475–492
Review482
Fig. 3 (a–d) Selective exploitation of different natural substrates by mycelium of Paxillus involutus growing from its mycorrhizal associations with Betula pendula. (a) Entire microcosm showing the low nutrient basal substrate of peat plus inert alkathane beads, and the trays of litter collected from the fermentation horizon (FH) horizons of Fagus (left) Pinus (centre) and Betula (right) forests and supplied as the only major potential source of N and P. Note heavy mycorrhizal colonisation of Betula plant by P. involutus and proliferation of the mycelium of this fungus in all trays. Bar, 1 cm. (b) Close up of tray in microcosm of same design as in (a) but with trays supplied with Pinus pollen as potential nutrient

Research review
© New Phytologist (2003) 157: 475–492 www.newphytologist.com
Review 483
environments have inherently greater proteolytic potentialthan those from temperate regions. Such observations,coupled with others (Tibbett et al., 1999) indicating that inthe same genus a thermal optimum for activity of proteasemay be as low as 0–6°C, suggest that selection favours ‘proteinfungi’ in arctic and boreal environments where low tempera-tures, acidity and poor resource quality combine to inhibit Nmineralisation.
In theory, the natural abundance of the heavier isotope ofN, δ15N, in plant and fungal tissues should reflect that of thenitrogenous substrates on which the organisms have beengrown. Some reports (Michelsen et al., 1996) have indeedsuggested that the N signatures of field grown plants withERM, ECM and AM colonisation demonstrate the hypo-thesised preferences for organic vs mineral N sources in naturalsoil. However analyses using amino acids and protein of pre-determined δ15N signatures as model substrates for fungi andplants representative of these mycorrhizal groups (Emmertonet al., 2001a,b) have dampened earlier enthusiasm for thisindirect approach to identification of substrate exploitation. Itwas shown that extensive fractionations of substrate N occurin the course of assimilation by the organisms and that theseare of sufficient magnitude to distort the signatures seen insink tissues.
In view of the fact that, as in the case of N, most of the soilP in boreo-temperate systems is in organic combination, thereis a lamentable ignorance of the extent to which, and themechanisms whereby, these sources are accessible to plants.Inositol hexaphosphate (IHP) used as a model polymericsource of P, has been shown to be accessible to a number ofECM fungi (Antibus et al., 1992). Since IHP is believed to beone of the major repositories of P in soil organic matter, albeitnot in pure form, use of this polymer provides information ofdirect ecological relevance.
Mobilisation of N and P from ‘natural’ substrates
In recent times, attention has been turned to assessment ofthe abilities of ECM systems to access N and P sequesteredin naturally occurring rather than ‘model’ substrates. Recog-nition of the propensity of these fungi to form dense mycelialmats when encountering particular types of soil organicmatter both in the field (Hintikka & Naykki, 1967; Cromacket al., 1979; Griffiths et al., 1991) and in microcosms (Read,1991; Bending & Read, 1995a,b) has led to analyses of themicrobial dynamics of these areas of enhanced fungal growth.Ingham et al. (1991) showed that mat soils had higher levelsof microbial biomass while Cromack et al. (1988) observed
that populations of protozoans, nematodes and micro-arthropods in mat and nonmat soil were qualitatively andquantitatively different. Using microcosms it has also beenshown that the patterns of distribution and the populationstructures of bacterial communities associated with ECMmycelium are distinctive (Nurmiaho-Lassila et al., 1997).Recognition that diverse communities of organisms areapparently selectively present in association with ECMmycelia represents an important conceptual advance butmuch remains to be learned about the contribution of thedifferent trophic groups to nutritional dynamics of thesesystems. Assays of enzyme activity have confirmed enhancedlevels of cellulase, peroxidase, proteinase and phosphatase inmat relative to nonmat soils (Griffiths et al., 1991) while inmicrocosms, Bending & Read (1995b) observed higher levelsof protease and polyphenol oxidase together with short-termelevation of phosphatase activity in patches of soil occupied bydense growth of Paxillus mycelium, relative to that seen inadjacent unoccupied soil.
Occupation by dense mats of mycorrhizal mycelium,whether of soil in nature (Entry et al., 1991) or materialcollected from the FH and placed as discrete blocks inmicrocosms (Bending & Read, 1995a Perez-Moreno & Read,2000) is associated with reductions in N and P contents ofthese substrates. Entry et al. (1991) reported reductions ofboth these elements of around 33% while in adjacent nonmatsoils over the same period losses were around 17%. Using themicrocosm approach Bending & Read (1995a) observed thatthe extent of depletion of N and P in FH material differedbetween fungal symbionts. When growing in association withPinus sylvestris, dense patches of Suillus bovinus myceliumproduced reductions of c23% in both N and P concentrationsof pine litter, while Thelephora terrestris reduced N contents byonly 13% and failed to provide a significant reduction of P. Inthese cases analyses of control litter samples incubated free ofECM fungal colonisation showed that mineralisation wasinsufficient to account for gains of N or P seen in the plants.
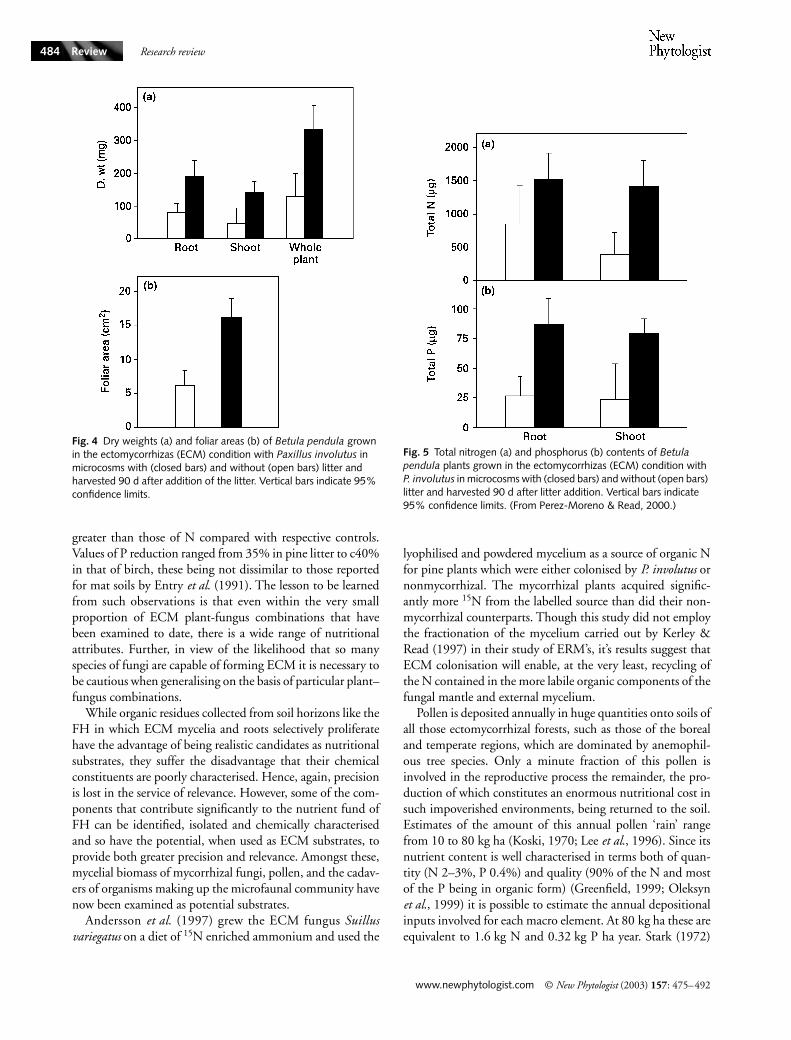
The importance of differences between combinations ofhost plants and fungi is further emphasised by the study ofPerez-Moreno & Read (2000). In this case a similar experi-mental design was employed but with a wider range of littertypes and Betula pendula-Paxillus involutus as the photo andmycobionts (Fig. 3a,c). As in the earlier study (Bending &Read, 1995a) exploitation of the FH materials by the mycor-rhizal fungi facilitated significant increases of yield (Fig. 4a),photosynthetic area (Fig. 4b), N and P acquisition (Fig. 5) incolonised relative to nonmycorrhizal plants. However, in thiscase amounts of P extracted from the litters were considerably
Fig. 3 (continued) source. Photograph taken 115 d after addition of pollen to microcosms. Note intensive exploitation of the tray by mycelium of P. involutus. Bar, 3.3 cm. (c) Close up of Fagus litter tray from (a) showing selective proliferation of Paxillus mycelium. Bar, 3.5 cm. (d) Lower left portion of microcosm of same design as (a) but with nematode necromass added as substrate to right hand try. Note intensive mycelial exploitation of necromass by P. involutus. The left hand tray received no nematode addition. Some increase in mycelial density is visible in this tray which results from ‘downstream’ invigoration of mycelium enabled by nutrient transfer from the nematode-enriched trays. Photograph taken 100 d after addition of necromass to microcosm. Bar, 1.3 cm.
Research review
www.newphytologist.com © New Phytologist (2003) 157: 475–492
Review484
greater than those of N compared with respective controls.Values of P reduction ranged from 35% in pine litter to c40%in that of birch, these being not dissimilar to those reportedfor mat soils by Entry et al. (1991). The lesson to be learnedfrom such observations is that even within the very smallproportion of ECM plant-fungus combinations that havebeen examined to date, there is a wide range of nutritionalattributes. Further, in view of the likelihood that so manyspecies of fungi are capable of forming ECM it is necessary tobe cautious when generalising on the basis of particular plant–fungus combinations.
While organic residues collected from soil horizons like theFH in which ECM mycelia and roots selectively proliferatehave the advantage of being realistic candidates as nutritionalsubstrates, they suffer the disadvantage that their chemicalconstituents are poorly characterised. Hence, again, precisionis lost in the service of relevance. However, some of the com-ponents that contribute significantly to the nutrient fund ofFH can be identified, isolated and chemically characterisedand so have the potential, when used as ECM substrates, toprovide both greater precision and relevance. Amongst these,mycelial biomass of mycorrhizal fungi, pollen, and the cadav-ers of organisms making up the microfaunal community havenow been examined as potential substrates.
Andersson et al. (1997) grew the ECM fungus Suillusvariegatus on a diet of 15N enriched ammonium and used the
lyophilised and powdered mycelium as a source of organic Nfor pine plants which were either colonised by P. involutus ornonmycorrhizal. The mycorrhizal plants acquired signific-antly more 15N from the labelled source than did their non-mycorrhizal counterparts. Though this study did not employthe fractionation of the mycelium carried out by Kerley &Read (1997) in their study of ERM’s, it’s results suggest thatECM colonisation will enable, at the very least, recycling ofthe N contained in the more labile organic components of thefungal mantle and external mycelium.
Pollen is deposited annually in huge quantities onto soils ofall those ectomycorrhizal forests, such as those of the borealand temperate regions, which are dominated by anemophil-ous tree species. Only a minute fraction of this pollen isinvolved in the reproductive process the remainder, the pro-duction of which constitutes an enormous nutritional cost insuch impoverished environments, being returned to the soil.Estimates of the amount of this annual pollen ‘rain’ rangefrom 10 to 80 kg ha (Koski, 1970; Lee et al., 1996). Since itsnutrient content is well characterised in terms both of quan-tity (N 2–3%, P 0.4%) and quality (90% of the N and mostof the P being in organic form) (Greenfield, 1999; Oleksynet al., 1999) it is possible to estimate the annual depositionalinputs involved for each macro element. At 80 kg ha these areequivalent to 1.6 kg N and 0.32 kg P ha year. Stark (1972)
Fig. 4 Dry weights (a) and foliar areas (b) of Betula pendula grown in the ectomycorrhizas (ECM) condition with Paxillus involutus in microcosms with (closed bars) and without (open bars) litter and harvested 90 d after addition of the litter. Vertical bars indicate 95% confidence limits.
Fig. 5 Total nitrogen (a) and phosphorus (b) contents of Betula pendula plants grown in the ectomycorrhizas (ECM) condition with P. involutus in microcosms with (closed bars) and without (open bars) litter and harvested 90 d after litter addition. Vertical bars indicate 95% confidence limits. (From Perez-Moreno & Read, 2000.)
Research review
© New Phytologist (2003) 157: 475–492 www.newphytologist.com
Review 485
hypothesised that the pollen rain would constitute a sign-ificant seasonal source of N and P for litter decay fungi andHutchison & Barron (1997) observed that 41 of 147 fungalsaprotrophs could use pollen as a nutrient resource, but untilrecently the possible contribution of ECM fungi to theprocesses of nutrient recovery by the trees had not beenconsidered.
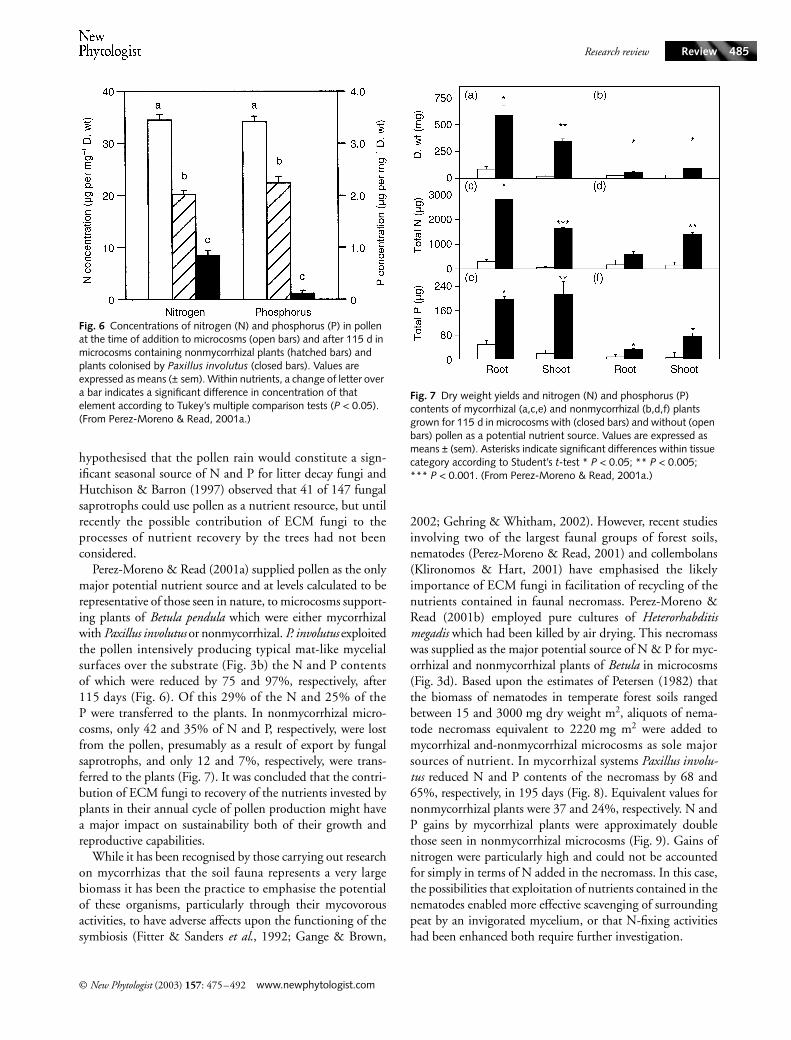
Perez-Moreno & Read (2001a) supplied pollen as the onlymajor potential nutrient source and at levels calculated to berepresentative of those seen in nature, to microcosms support-ing plants of Betula pendula which were either mycorrhizalwith Paxillus involutus or nonmycorrhizal. P. involutus exploitedthe pollen intensively producing typical mat-like mycelialsurfaces over the substrate (Fig. 3b) the N and P contentsof which were reduced by 75 and 97%, respectively, after115 days (Fig. 6). Of this 29% of the N and 25% of theP were transferred to the plants. In nonmycorrhizal micro-cosms, only 42 and 35% of N and P, respectively, were lostfrom the pollen, presumably as a result of export by fungalsaprotrophs, and only 12 and 7%, respectively, were trans-ferred to the plants (Fig. 7). It was concluded that the contri-bution of ECM fungi to recovery of the nutrients invested byplants in their annual cycle of pollen production might havea major impact on sustainability both of their growth andreproductive capabilities.
While it has been recognised by those carrying out researchon mycorrhizas that the soil fauna represents a very largebiomass it has been the practice to emphasise the potentialof these organisms, particularly through their mycovorousactivities, to have adverse affects upon the functioning of thesymbiosis (Fitter & Sanders et al., 1992; Gange & Brown,
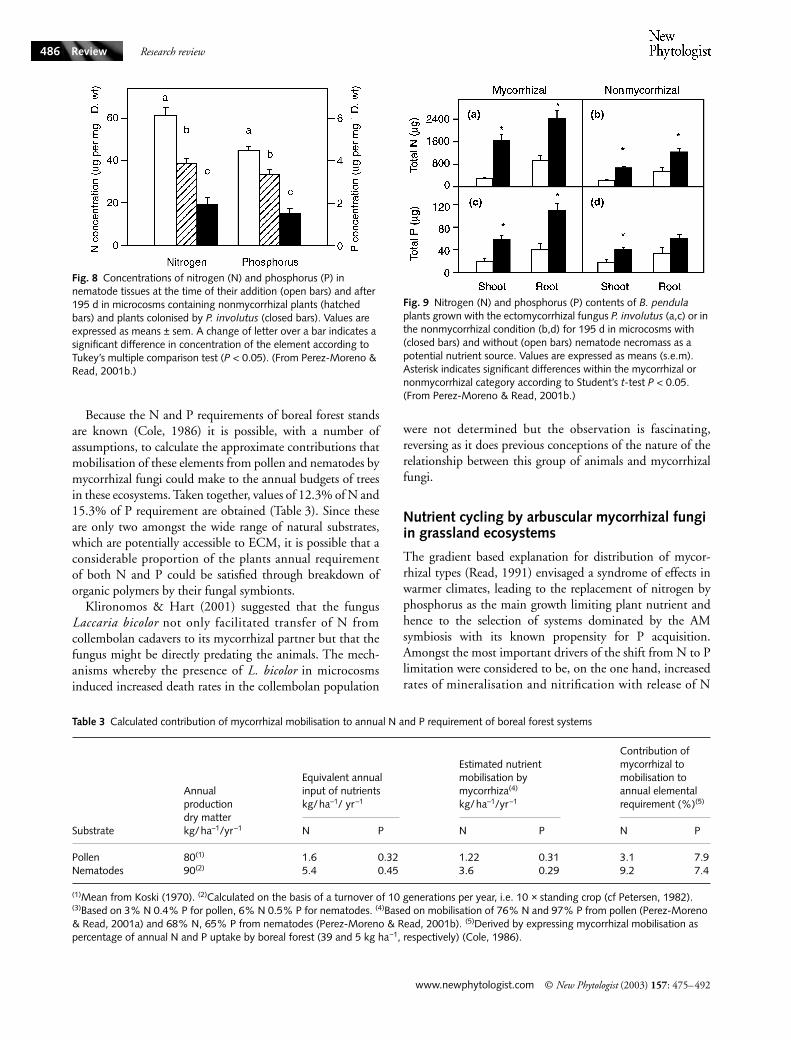
2002; Gehring & Whitham, 2002). However, recent studiesinvolving two of the largest faunal groups of forest soils,nematodes (Perez-Moreno & Read, 2001) and collembolans(Klironomos & Hart, 2001) have emphasised the likelyimportance of ECM fungi in facilitation of recycling of thenutrients contained in faunal necromass. Perez-Moreno &Read (2001b) employed pure cultures of Heterorhabditismegadis which had been killed by air drying. This necromasswas supplied as the major potential source of N & P for myc-orrhizal and nonmycorrhizal plants of Betula in microcosms(Fig. 3d). Based upon the estimates of Petersen (1982) thatthe biomass of nematodes in temperate forest soils rangedbetween 15 and 3000 mg dry weight m2, aliquots of nema-tode necromass equivalent to 2220 mg m2 were added tomycorrhizal and-nonmycorrhizal microcosms as sole majorsources of nutrient. In mycorrhizal systems Paxillus involu-tus reduced N and P contents of the necromass by 68 and65%, respectively, in 195 days (Fig. 8). Equivalent values fornonmycorrhizal plants were 37 and 24%, respectively. N andP gains by mycorrhizal plants were approximately doublethose seen in nonmycorrhizal microcosms (Fig. 9). Gains ofnitrogen were particularly high and could not be accountedfor simply in terms of N added in the necromass. In this case,the possibilities that exploitation of nutrients contained in thenematodes enabled more effective scavenging of surroundingpeat by an invigorated mycelium, or that N-fixing activitieshad been enhanced both require further investigation.
Fig. 6 Concentrations of nitrogen (N) and phosphorus (P) in pollen at the time of addition to microcosms (open bars) and after 115 d in microcosms containing nonmycorrhizal plants (hatched bars) and plants colonised by Paxillus involutus (closed bars). Values are expressed as means (± sem). Within nutrients, a change of letter over a bar indicates a significant difference in concentration of that element according to Tukey’s multiple comparison tests (P < 0.05). (From Perez-Moreno & Read, 2001a.)
Fig. 7 Dry weight yields and nitrogen (N) and phosphorus (P) contents of mycorrhizal (a,c,e) and nonmycorrhizal (b,d,f) plants grown for 115 d in microcosms with (closed bars) and without (open bars) pollen as a potential nutrient source. Values are expressed as means ± (sem). Asterisks indicate significant differences within tissue category according to Student’s t-test * P < 0.05; ** P < 0.005; *** P < 0.001. (From Perez-Moreno & Read, 2001a.)
Research review
www.newphytologist.com © New Phytologist (2003) 157: 475–492
Review486
Because the N and P requirements of boreal forest standsare known (Cole, 1986) it is possible, with a number ofassumptions, to calculate the approximate contributions thatmobilisation of these elements from pollen and nematodes bymycorrhizal fungi could make to the annual budgets of treesin these ecosystems. Taken together, values of 12.3% of N and15.3% of P requirement are obtained (Table 3). Since theseare only two amongst the wide range of natural substrates,which are potentially accessible to ECM, it is possible that aconsiderable proportion of the plants annual requirementof both N and P could be satisfied through breakdown oforganic polymers by their fungal symbionts.
Klironomos & Hart (2001) suggested that the fungusLaccaria bicolor not only facilitated transfer of N fromcollembolan cadavers to its mycorrhizal partner but that thefungus might be directly predating the animals. The mech-anisms whereby the presence of L. bicolor in microcosmsinduced increased death rates in the collembolan population
were not determined but the observation is fascinating,reversing as it does previous conceptions of the nature of therelationship between this group of animals and mycorrhizalfungi.
Nutrient cycling by arbuscular mycorrhizal fungi in grassland ecosystems
The gradient based explanation for distribution of mycor-rhizal types (Read, 1991) envisaged a syndrome of effects inwarmer climates, leading to the replacement of nitrogen byphosphorus as the main growth limiting plant nutrient andhence to the selection of systems dominated by the AMsymbiosis with its known propensity for P acquisition.Amongst the most important drivers of the shift from N to Plimitation were considered to be, on the one hand, increasedrates of mineralisation and nitrification with release of N
Table 3 Calculated contribution of mycorrhizal mobilisation to annual N and P requirement of boreal forest systems
Substrate
Annual productiondry matter kg/ha–1/yr–1
Equivalent annual input of nutrientskg/ha–1/ yr–1
Estimated nutrient mobilisation by mycorrhiza(4)
kg/ha–1/yr–1
Contribution ofmycorrhizal to mobilisation to annual elemental requirement (%)(5)
N P N P N P
Pollen 80(1) 1.6 0.32 1.22 0.31 3.1 7.9Nematodes 90(2) 5.4 0.45 3.6 0.29 9.2 7.4
(1)Mean from Koski (1970). (2)Calculated on the basis of a turnover of 10 generations per year, i.e. 10 × standing crop (cf Petersen, 1982). (3)Based on 3% N 0.4% P for pollen, 6% N 0.5% P for nematodes. (4)Based on mobilisation of 76% N and 97% P from pollen (Perez-Moreno & Read, 2001a) and 68% N, 65% P from nematodes (Perez-Moreno & Read, 2001b). (5)Derived by expressing mycorrhizal mobilisation as percentage of annual N and P uptake by boreal forest (39 and 5 kg ha−1, respectively) (Cole, 1986).
Fig. 8 Concentrations of nitrogen (N) and phosphorus (P) in nematode tissues at the time of their addition (open bars) and after 195 d in microcosms containing nonmycorrhizal plants (hatched bars) and plants colonised by P. involutus (closed bars). Values are expressed as means ± sem. A change of letter over a bar indicates a significant difference in concentration of the element according to Tukey’s multiple comparison test (P < 0.05). (From Perez-Moreno & Read, 2001b.)
Fig. 9 Nitrogen (N) and phosphorus (P) contents of B. pendula plants grown with the ectomycorrhizal fungus P. involutus (a,c) or in the nonmycorrhizal condition (b,d) for 195 d in microcosms with (closed bars) and without (open bars) nematode necromass as a potential nutrient source. Values are expressed as means (s.e.m). Asterisk indicates significant differences within the mycorrhizal or nonmycorrhizal category according to Student’s t-test P < 0.05. (From Perez-Moreno & Read, 2001b.)

Research review
© New Phytologist (2003) 157: 475–492 www.newphytologist.com
Review 487
in the form of mobile nitrate ions, and, on the other, thesequestration of phosphate ions with salts, associated withgreater rates of evapotranspiration and/or raised pH. Thisview was in accordance with the perception that AM fungi,in contrast to their ECM and ERM counterparts, lackedthe saprotrophic capability to enable N mineralisation butpossessed effective P-scavenging attributes. While little hasemerged over the past 10 years to radically change this view, arefreshing new awareness has emerged of the possibility that,under some circumstances, AMF may be involved both indecomposition processes and in the capture of the less mobileamino-acids or ammonium ions. Hodge (2001) added 15N/13C labelled glycine as a localised organic N source to mineralsoil and grew plants with 3 different AMF species or in thenonmycorrhizal condition. None of the fungi responded tothe presence of the glycine by hyphal proliferation and neitherN nor C capture were increased in the M relative to NM plants.By contrast, experimental analysis of ammonium uptakeindicates that both capture and transport of this ion to theplant can be enhanced by AMF (Mäder et al., 2000). Provisionof access to ammonium ions may well be of ecologicalimportance for plants in biomes with acidic organic soils, suchas those prevailing in some tropical environments, which cansupport abundant AM colonisation (Moyersoen et al., 2001).
There is a single report (Hodge et al., 2001) of enhanceddecomposition and increased N capture by AMF fromorganic necromass. Dual 15N/13C labelled grass leaves weresupplied as an organic residue which could be colonised byfungal mycelium extending through a mesh barrier fromcolonised roots. AM hyphae were shown to facilitate enhance-ment of N capture from the litter, the N gain in the plantsbeing linearly related to hyphal density in the organic matter.Enhanced release of 13C in the AMF compartment was alsoindicative of increased decomposition rates. Since analysis ofthe microbial community structure in the organic patchesshowed no qualitative differences between AMF colonisedand uncolonised material it was concluded that the fungalsymbionts may be directly involved in the decompositionprocesses. The possibility remains, however, that allocation ofphotosynthate by these fungi might facilitate enhancementof the activity of microbial generalists in the AMF compart-ments. Since AM mycelia provide extensive conduits forallocation of carbon to the soil system (Johnson et al., 2001,2002) the extent to which they act directly, rather than asfacilitators, of decomposition and nutrient release urgentlyneeds to be determined. Here, as in the case of ERM systems(Kerley & Read, 1998) studies under monoxenic conditionsmay offer a fruitful way forward.
In view of the preoccupation with phosphorus captureshown in studies of the AM symbiosis over the years therehave been remarkably few analyses of the ability of AM fungito mobilise P from the organic residues which, even in themineral soils that frequently support AM communities canbe the main potential source of the element for plants. The
deficiency is compounded by the fact that so many studies ofAM systems do not provide adequate discrimination betweennutrient mobilising activities of the roots and their fungalsymbionts. Tarafdar & Marschner (1994) separated the twoactivities by allowing an AM fungus to grow from a host plant(Triticum aestivum) through a root-excluding mesh. Through-out the hyphal compartment so obtained, acid phosphataseactivities were much higher in the mycelial compartment anda strong correlation between this activity and AM hyphallength provided circumstantial evidence that the enzyme wasproduced by the fungal symbiont rather than by the back-ground microflora. In this study, of the total P uptake by theplant, the mycorrhizal contribution accounted for 22–33%when P was supplied in inorganic form and 48–59% with Pin organic form. It should be said, however, that when Joneret al. (1995) repeated this type of experiment using a similardesign they could find no influence of the presence of AMhyphae on soil phosphatase activity in spite of high hyphaldensities in the mycelial compartments of their microcosms.A failure to detect increases in phosphatase activity in AMmycorrhizal systems is in some ways surprising since it hasbeen shown (Koide & Kabir, 2000) under axenic conditionsthat the extra radical hyphae of one such fungus, Glomusintraradices, was readily capable of hydrolysing exogenouslysupplied organic P sources. Similarly, Joner et al. (2000),again under circumstances where other microbes wereexcluded from the assay system, showed that an AM funguswas capable of mobilisation and transfer of 32P from theorganic compound AMP to a plant.
Many more studies of this kind are required before we willbe in a position adequately to discriminate between direct andfacilitated attack on soil organic resources by AM fungi. Ofparticular value would be analyses of P starved tropical rainforest ecosystems in which AM roots, probably colonised byso-far undescribed fungal symbionts, are known to be inti-mately associated with decaying organic matter (Newberyet al., 1997; Moyersoen et al., 1998).
Overview
The conventional view that all plants are dependent fortheir supplies of N and P upon the mineralising activitiesof microbial generalists is increasingly challenged by theemerging evidence. Rather, it appears that in environmentswhere rates of mineralisation are sufficiently slow to threatenthe fitness of autotrophs, selection has favoured associationswith fungal symbionts that are physiologically equipped tofacilitate capture of these elements from their locally pre-dominant organic sources. These nutrient mobilising attributesare selectively deployed largely in superficial soil environmentswhere attacks upon substrates of particular quality can pre-empt both the immobilisation of N and P by saprotrophs, andtheir incorporation into the heterocyclic humic compoundswhich are the long-term repositories of these elements if they

Research review
www.newphytologist.com © New Phytologist (2003) 157: 475–492
Review488
reach the lower parts of the profile. The mechanism is one inwhich plants invest carbon, a currency not in short supply formost autotrophs, to enable mobilisation, by mycorrhizalfungi, of the fitness threatening resource.
The broader implications of this relatively recent apprecia-tion of nutrient cycling processes in ecosystems requireemphasis. Though the proportion of plant species associatedwith the ERM or ECM fungi known to be capable of theseactivities is relatively small, perhaps around 10% of thoseexamined to date (Trappe, 1987), as dominants of the tundra,taiga, and boreo-temperate forest zones, they cover up to70% of the terrestrial surface of the northern hemisphere andexploit soils containing the largest global capital of organiccarbon (Post et al., 1982). By occupying the interface betweenthe plants which are the sites of carbon fixation, and the soilin which the element is stored, these fungi have the potentialprofoundly to influence the carbon source-sink relationshipsupon which global climate systems ultimately depend. Inparticular through their involvement in the removal of Nand P from organic polymers they inevitably increase theC : N and C : P ratios of the residual materials and thus willcontribute to C retention in soil.
The major potential sources of N and P are the residues ofthe locally dominant symbiotic and free-living microflora,of the micro and meso-fauna, and of the plants that character-ise the particular ecosytem or biome. It is clear that ERM andmany ECM fungi have the abilities to degrade some of thestructural units within such residues which might otherwiserestrict access to these elements. They then can attack manyof the nutrient containing polymers themselves. There is little,in contrast, to indicate that mycorrhizal fungi, even if some ofthem possess modest ligninolytic capabilities, have the poten-tial to compete with ‘white rot’ fungi in the essentially N andP-poor environments provided by large woody debris. Sincetheir energy requirements are satisfied by autotrophs therewould, indeed, seem to be little advantage for them tocompete in such carbon-rich habitats.
The results obtained over the last decade from experimen-tal analysis of the physiological capabilities of mycorrhizalfungi have provided much support for the hypothesis (Read,1991) that on a global scale these heterotrophs may be makingsignificant contributions which are distinctive at latitudinalor altitudinal scales to ecosystem nutrient cycling (Fig. 10).Some recent studies at a local scale have added refinement
Fig. 10. The proposed relationships, on a northern hemisphere based global scale, between the distribution of biomes along environmental gradients and the roles of the prevailing mycorrhizal association in facilitation of N and P capture by the characteristic functional groups of plant.

Research review
© New Phytologist (2003) 157: 475–492 www.newphytologist.com
Review 489
to the perspective. Giesler et al. (1998) examined plant–microbe–nutrient interactions across an extreme gradientwithin the northern boreal zone in which soil conditionschanged, over a distance of only 90 m, from extreme acidity(pH 3.5) and an absence of nitrification at one end, to nearneutrality (pH 6.4) with high levels of NO3 in a localisedzone of ground water discharge at the other. Because levels ofavailable P were consistently low across this pH-N gradientthe transition, in effect, was one of decreasing P : N ratio.Along the gradient the ground flora was seen to change froman exclusive dominance of ERM shrubs in the absence ofnitrification in the acidic region to a dominance of potentiallyAM colonised tall herbs at the other. Although, as would beexpected at this northerly latitude an overstorey of sprucewas maintained across the gradient, quantitative analyses ofmicrobial fatty acid signatures indicated that the extent ofECM fungal mycelial development was greatly reduced asproductivity increased in the nitrogen enriched part of thesystem. Giesler et al. (1998) propose that the success of dis-tinctive types of understory plant at particular points alongthis short gradient are likely to be determined by the abilitiesof their mycorrhizal symbionts to access the locally limitingmacro-element. It remains to be demonstrated experimentallythat AM fungi contribute to enabling the gradual increase inrepresentation of herbs along such gradients of decreasingP : N ratio. In particular, analyses of the responsiveness ofputatively AM herbs to colonisation by their naturallyselected fungal endophytes under realistic nutrient regimesare long overdue.
At a scale intermediate between the global (Read, 1991)and local (Giesler et al., 1998), the occurrence of distinctiveunderstory associations dominated by ericacaeous, mixedshrub-herb or herb systems has long been recognised at theregional level in boreal biomes, where it has underpinnedplant community (Cajander, 1926) or nutrient-based (Dahlet al., 1967; Lahti & Vaisanen, 1987) descriptions of foresttypes. More recently, Cornellissen et al. (2001) used 83British plants species of known functional and mycorrhizalcategories to test the hypothesis that classification of the plantfunctional types according to mycorrhizal association couldhelp to explain nutritional feedbacks between plant produc-tivity and litter turnover. They observed within this particularsubset of the temperate flora, that ericoid and ectomycorrhizalstrategies were linked to low and arbuscular mycorrhizalspecies to high ecosystem turnover of carbon and nutrients.This study provides a complementary link between the globaland local scales and largely confirms their predictions. Theemerging appreciation of the full physiological potential ofeach of the major mycorrhizal types should help to improveour mechanistic understanding of the factors enablingdominance of distinctive plant communities at local, regionaland global scales.
While Giesler et al. (1998) described a pristine borealenvironment which was likely to be in long-term nutritional
equilibrium, decreases in P : N ratio are occurring acrossmuch of the northern hemisphere as a result of anthropogenicN enrichment. These changes should also favour the domi-nance of AM-based communities (Aerts, 2002). Clearly, thereis a need to examine the extent of involvement of mycorrhizalfungi in the changing patterns of plant distribution which arearising globally in response to the nutritional disturbancesinduced by human activities.
ReferencesAerts R. 2002. The role of various types of mycorrhizal fungi in nutrient
cycling and plant competition. In: van der Heijden MGA, Sanders I, eds. Mycorrhizal ecology. Ecological Studies, Vol. 157. Berlin, Germany: Springer-Verlag, 117–133.
Andersson S, Ek H, Söderström B. 1997. Effects of liming on the uptake of organic and inorganic nitrogen by mycorrhizal (Paxillus involutus) and non mycorrhizal Pinus sylvestris plants. New Phytologist 135: 763–771.
Antibus RK, Sinsabaugh RL, Linkins AE. 1992. Phosphatase activities and phosphorus uptake from insositol phosphate by ectomycorrhizal fungi. Canadian Journal of Botany 70: 794–801.
Appel T, Mengel K. 1998. Prediction of mineralizable nitrogen in soils on the basis of an analysis of extractable organic N. Zeitschrift Fur Planzenernährung und Bodenkunde 161: 433–452.
Baar J, Horton TR, Kretzer AM, Bruns TD. 1999. Mycorrhizal colonisation of Pinus muricata from resistant propagules after a stand replacing wildfire. New Phytologist 143: 409–418.
Bajwa R, Abuarghub S, Read DJ. 1985. The Biology of Mycorrhiza in the Ericaceae. X. The utilisation of proteins and the production of proteolytic enzymes by the mycorrhizal endophyte and by mycorrhizal plants. New Phytologist 101: 469–486.
Bending GD, Read DJ. 1995a. The structure and function of the vegetative mycelium of ectomycorrhizal plants. V. Foraging behaviour and translocation of nutrients from exploited organic matter. New Phytologist. 130: 401–409.
Bending GD, Read DJ. 1995b. The structure and function of the vegetative mycelium of ectomycorrhizal plants. VI. Activities of nutrient mobilising enzymes in birch litter colonised by Paxillus involutus (Fr.) Er. New Phytologist. 130: 411–417.
Bending GD, Read DJ. 1996a. Effects of the soluble polyphenol tannic acid on the activities of ericoid and ectomycorrhizal fungi. Soil Biology and Biochemistry 28: 1595–1602.
Bending GD, Read DJ. 1996b. Nitrogen mobilization from protein-polyphenol complex by ericoid and ectomycorrhizal fungi. Soil Biology and Biochemistry 28: 1603–1612.
Bending GD, Read DJ. 1997. Lignin and soluble-phenolic degradation by ectomycorrhizal and ericoid mycorrhizal fungi. Mycological Research 101: 1348–1354.
Burke RM, Cairney JWG. 1997a. Carbohydrolase production by the ericoid mycorrhizal fungus Hymenoscyhus ericae under solid-state fermentation conditions. Mycological Research 101: 1135–1139.
Burke RM, Cairney JWG. 1997b. Purification and characterization of a β-1,4-endoxylanase from the ericoid mycorrhizal fungus Hymenoscyphus ericae. New Phytologist 135: 345–352.
Burke RM, Cairney JWG. 1998. Carbohydrate oxidases in ericoid and ectomycorrhizal fungi: a possible source of Fenton radicals during degradation of lignocellulose. New Phytologist 139: 637–645.
Cairney JWG, Burke RM. 1994. Fungal enzymes degrading plant cell walls: their possible significance in the ectomycorrhizal symbiosis. Mycological Research 98: 1345–1346.
Cairney JWG, Burke RM. 1996a. Physiological heterogeneity within fungal mycelia: an important concept for a functional understanding of the ectomycorrhizal symbiosis. New Phytologist 134: 685–695.

Research review
www.newphytologist.com © New Phytologist (2003) 157: 475–492
Review490
Cairney JWG, Burke RM. 1996b. Plant cell wall-degrading enzymes in ericoid and ectomycorrhizal fungi. In: Azcon-Aguilar C, Barea JM, eds. Mycorrhizas in integrated systems. Brussels, Belgium: European Commission, 218–221.
Cairney JWG, Burke RM. 1998. Extracellular enzyme activities of the ericoid mycorrhizal endophyte Hymenoscyphus ericae (Read) Korf & Kernan: their likely roles in decomposition of dead plant tissues in soil. Plant and Soil 205: 181–192.
Cajander AK. 1926. The theory of forest types. Acta Forestalia Fennica 29: 1–108.
Caldwell BA, Castellano MA, Griffiths RP. 1991. Fatty acid esterase production by ectomycorrhizal fungi. Mycologia 83: 233–236.
Cao W, Crawford DL. 1993. Carbon nutrition and hydrolytic and cellololytic activities in the ectomycorrhizal fungus Pisolithus tinctorius. Canadian Journal of Microbiology 39: 529–535.
Chalot M, Brun A. 1998. Physiology of organic nitrogen acquisition by ectomycorrhizal fungi and ectomycorrhizas. FEMS Microbiology Reviews 22: 21–44.
Chambers SM, Burke RM, Brooks PR, Cairney JWG. 1999. Molecular and biochemical evidence for manganese-dependent peroxidase activity in Tylospora fibrillose. Mycological Research 103: 1098–1102.
Chapin FS III, Moilanen L, Kielland K. 1993. Preferential use of organic nitrogen for growth by a non-mycorrhizal arctic sedge. Nature 361: 150–153.
Chen DM, Taylor AFS, Burke RM, Cairney JWG. 2001. Identification of genes for lignin peroxidases and manganese peroxidases in ectomycorrhizal fungi. New Phytologist 152: 151–158.
Cole DW. 1986. Nutrient cycling in world forests. In: Gessel SP, ed. Forest site and productivity. Dordrecht, The Netherlands: Martinus-Nijhoff, 103–115.
Colpaert JV, Van Laere A. 1996. A comparison of the extracellular enzyme activities of two ectomycorrhizal and a leaf-saprotrophic basidiomycete colonising beech litter. New Phytologist 134: 133–141.
Colpaert JV, van Tichelen KK. 1996. Decomposition, nitrogen and phosphorus mineralization from beech leaf litter colonized by ectomycorrhizal or litter-decomposing basidiomycetes. New Phytologist 134: 123–132.
Cornelissen JHC, Aerts R, Cerabolini B, Werger MJA, van der Heijden MGA. 2001. Carbon cycling traits of plant species are linked with mycorrhizal strategy. Oecologia 129: 611–619.
Cromack K Jr, Fichter BL, Moldenke AM, Entry JA, Ingham ER. 1988. Interactions between soil animals and ectomycorrhizal fungal mats. Agriculture, Ecosystems and Environment 24: 161–168.
Cromack K, Sollins P, Graustein WC, Speidel K, Todd AW, Spycher G, Li CT, Todd RL. 1979. Calcium oxalate accumulation and soil weathering in mats of the hypogeous fungus Hysterangium crassum. Soil Biology and Biochemistry 11: 463.
Dahl E, Gjems O, Kjelland-Lund J Jr. 1967. On the vegetation of Norwegian conifer forest in relation to the chemical properties of the humus layer. Meddelende I Norske Skogsforsøksvesen 85: 501–531.
Dahlberg A. 2001. Community ecology of ectomycorrhizal fungi: an advancing interdisciplinary field. New Phytologist 150: 555–562.
Egger KN, Sigler L. 1993. Relatedness of the ericoid endophytes Scytalidium vaccinii and Hymenoscyphus ericae inferred from analysis of ribosomal DNA. Mycologia 85: 219–230.
El-Badaoui K, Botton B. 1989. Production and characterization of exocellular proteases in ectomycorrhizal fungi. Annales Des Sciences Forestieres 46: 728s–730s.
Emmerton KS, Callaghan TV, Jones HE, Leake JR, Michelsen A, Read DJ. 2001a. Assimilation and isotopic fractionation of nitrogen by mycorrhizal and nonmycorrhizal subarctic plant. New Phytologist 151: 513–524.
Emmerton KS, Callaghan TV, Jones HE, Leake JR, Michelsen A, Read DJ. 2001b. Assimilation and isotopic fractionation of nitrogen by mycorrhizal fungi. New Phytologist 151: 503–511.
Entry J, Rose CL, Cromack K. 1991. Litter decomposition and nutrient release in ectomycorrhizal mat soils of a Douglas-fir ecosystems. Soil Biology and Biochemistry 23: 285–290.
Erland S, Taylor AFS. 1999. Resupinate ectomycorrhizal fungal genera. In: Cairney JWG, Chambers SM, eds. Ectomycorrhizal fungi: key genera in profile. Berlin, Germany: Springer-Verlag, 347–363.
Finlay RD, Frosegärd Å, Sonnerfeldt AM. 1992. Utilization of organic and inorganic nitrogen sources by ectomycorrhizal fungi in pure culture and in symbiosis with Pinus contorta Dougl. Ex Loud. New Phytologist 120: 105–115.
Finlay RD, Söderström B. 1992. Mycorrhiza and carbon flow to the soil. In: Allen M, ed. Mycorrhiza functioning. London, UK: Chapman & Hall, 134–160.
Fitter AH, Sanders 1. 1992. Interactions with the soil fauna. In: Allen, MF, ed. Mycorrhizal functioning. New York, USA: Chapman & Hall, 333–354.
Frank AB. 1894. Die Bedeutung der Mykorrhizapilze für die gemeine Kiefer. Forstwissenschaftliche Centralblat 16: 1852–1890.
Gange AC, Brown VK. 2002. Actions and interactions of soil invertebrates and arbuscular mycorrhizal fungi in affecting the structure of plant communities. In: van der Heijden MGA, Sanders I, eds. Mycorrhizal ecology. Ecological Studies, Vol. 157. Berlin, Germany: Springer-Verlag, 321–344.
Gardes M, Bruns TD. 1993. Its primers with enhanced specificity for basidiomycetes: application to the identification of mycorrhiza and rusts. Molecular Ecology 2: 113–118.
Gardes M, Bruns TD. 1996. Community structure of ectomycorrhizal fungi in a Pinus muricata forest: Above- and below-ground views. Canadian Journal of Botany 74: 1572–1583.
Gardes M, White TJ, Fortin JA, Bruns TD, Taylor JW. 1991. Identification of indigenous and introduced symbiotic fungi in ectomycorrhizae by amplification of nuclear and mitochondrial ribosomal DNA. Canadian Journal of Botany 69: 80–190.
Gehring CA, Whitham TG. 2002. Mycorrhizae–herbivore interactions: Population and community consequences. In: van der Heijden MGA, Sanders I, eds. Mycorrhizal ecology. Ecological Studies, Vol. 157. Berlin, Germany: Springer-Verlag, 295–320.
Giesler R, Högberg M, Högberg P. 1998. Soil chemistry and plants in fennoscandian boreal forest as exemplified by a local gradient. Ecology 79: 119–137.
Greenfield LG. 1999. Weight loss and release of mineral nitrogen from decomposing pollen. Soil Biology and Biochemistry 31: 353–361.
Griffiths RP, Caldwell BA. 1992. Mycorrhizal mat communities in forest soils. In: Read DJ, Lewis DH, Fitter AH, Alexander IJ, eds. Mycorrhizas in ecosystems. Wallingford, UK: CAB International, 98–105.
Griffiths RP, Castellano MA, Caldwell BA. 1991. Ectomycorrhizal mats formal by Gautieria monticola and Hysterangium setchellii and their association with Douglas-fir seedlings, a case study. Plant and Soil 134: 255–259.
Günther T, Perner B, Gramms G. 1998. Activities of phenol oxidising enzymes of ectomycorrhizal fungi in axenic culture and in symbiosis with Scots pine (Pinus sylvestris L.). Journal of Basic Microbiology 38: 197–206.
Hambleton S, Egger KN, Currah RS. 1998. The genus Oidiodendron: species delimitation and phylogenetics relationships based on nuclear ribosomal DNA analysis. Mycologia 90: 854–868.
Harley JL, Smith SE. 1983. Mycorrhizal symbiosis, 1st edn. London, UK: Academic Press.
Haselwandter K, Bobleter O, Read DJ. 1990. Degradation of 14C-labelled lignin and dehydropolymer of coniferyl alcohol by ericoid and ectomycorrhizal fungi. Archives of Microbiology 153: 352–354.
Hibbett DS, Gilbert L-B, Donaghue MJ. 2000. Evolutionary instability of ectomycorrhizal symbioses in basidiomycetes. Nature 407: 506–508.
Hintikka V, Naykki O. 1967. on the effects of the fungus Hydnellum ferrugineum (Fr.) Karst. On forest soil and vegetation. Communications Instituti Forestalis Fenniae 62: 1–23.

Research review
© New Phytologist (2003) 157: 475–492 www.newphytologist.com
Review 491
Hodge A. 2001. Arbuscular mycorrhizal fungi influence decomposition of, but not plant nutrient capture from, glycine patches in soil. New Phytologist 151: 725–734.
Hodge A, Campbell CD, Fitter AH. 2001. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 413: 297–299.
Högberg MN, Högberg P. 2002. Extramatrical ectomycorrhizal mycelium contributes half the microbial biomass and produces, together with associated roots, half the dissolved organic carbon in a forest soil. New Phytologist. 154: 791–795.
Hutchison L. 1990a. Studies on the systematics of ectomycorrhizal fungi in axenic culture. II. The enzymatic degradation of selected carbon and nitrogen compounds. Canadian Journal of Botany 68: 1522–1530.
Hutchison L. 1990b. Studies on the systematics of ectomycorrhizal fungi in axenic culture. III. Patterns of polyphenol oxidase activity. Mycologia 82: 424–435.
Hutchison LJ, Barron GL. 1997. Parasitism of pollen as a nutritional source for lignicolous Basidiomycota and other fungi. Mycological Research 101: 191–194.
Ingham ER, Griffiths RP, Cromack K Jr, Entry JA. 1991. Comparison of direct versus fumigation-flush microbial biomass estimates from ectomycorrhizal mat and non-mat soils. Soil Biology and Biochemistry 23: 465–471.
Jalal MAF, Read DJ. 1983. The organic acid composition of Calluna heathland soil with special reference to phyto- and fungi-toxicity. I. Isolation and identification of organic acids. Plant and Soil. 70: 257–272.
Jalal MAF, Read DJ, Haslam E. 1982. Phenolic composition and its seasonal variation in Calluna vulgaris. Phytochemistry 21: 1397–1401.
Johnson D, Leake JR, Ostle N, Ineson P, Read DJ. 2002. In situ 13CO2 pulse-labelling of upland grassland demonstrates a rapid pathway of carbon flux from arbuscular mycorrhizal mycelia to the soil. New Phytologist 153: 327–334.
Johnson D, Leake JR, Read DJ. 2001. Novel in-growth core system enables functional studies of grassland mycorrhizal mycelial networks. New Phytologist 152: 555–562.
Joner EJ, Magid J, Gahoonia TS, Jakobsen I. 1995. P Depletion and activity of phosphatases in the rhizosphere of mycorrhizal and non-mycorrhizal cucumber (Cucumis sativus L.). Soil and Biology and Biochemistry 27: 1145–1151.
Joner EJ, Ravnskov S, Jakobsen I. 2000. Arbuscular mycorrhizal phosphate transport under monoxenic conditions using radio-labelled inorganic and organic phosphate. Biotechnology Letters : 1705–1708.
Joseleau J-P, Gharibian S, Comtat J, Lefebrve A, Ruel K. 1994. Indirect involvement of ligninolytic enzyme systems in cell wall degredation. FEMS Microbiology Letters 13: 255–264.
Kanunfre CC, Zancan GT. 1998. Physiology of exolaccase production by Thelephora terrestris. FEMS Microbiology Letters 161: 151–156.
Kerley SJ, Read DJ. 1995. The biology of mycorrhiza in the Ericaceae, XV Chitin degradation by Hymenoscyphus ericae and transfer of chitin-nitrogen to the host plant. New Phytologist 131: 369–375.
Kerley SJ, Read DJ. 1997. The biology of mycorrhiza in the Ericaceae XIX. Fungal mycelium as a nitrogen source for the ericoid mycorrhizal fungas Hymenoscyphus ericae (Read) Korf & Kernan and its host plants. New Phytologist 136: 691–701.
Kerley SJ, Read DJ. 1998. The biology of mycorrhiza in the Ericaceae XX plant and mycorrhizal necromass as nitrogenous substrates for the ericoid mycorrhizal fungus and its host. New Phytologist 139: 353–360.
Kielland K. 1994. Amino acid absorption by arctic plants: implications for plant nutrition and nitrogen cycling. Ecology 75: 2373–2383.
Klironomos JN, Hart MM. 2001. Animal nitrogen swap for plant carbon. Nature 410: 651–652.
Koide RT, Kabir Z. 2000. Extraradical hyphae of the mycorrhizal fungus Glomus intraradices can hydrolyse organic phosphate. New Phytologist 148: 511–517.
Kõljalg U. 1996. Tomentella (Basidiomycota) and related genera in temperate Eurasia. Synopsis Fungorum, Fungiflora, Oslo 9: 1–213.
Kõljalg U, Dahlberg A, Taylor AFS, Larsson E, Hallenberg N, Stenlid J, Larsson K-H, Fransson PM, Kåren O, Jonsson L. 2000. Diversity and abundance of resupinate thelephoroid fungi as ectomycorrhizal symbionts in Swedish boreal forests. Molecular Ecology 9: 1985–1996.
Kõljalg U, Tammi H, Timonen S, Agerer R, Sen R. 2002. ITS rDNA sequence-based phylogenetic analysis of Tomentellopsis species from boreal and temperate forests, and the identification of pink-type ectomycorrhizas. Mycological Progress 1: 81–92.
Koski V. 1970. A study of pollen dispersal as a mechanism of gene flow in conifers. Communicationes Instituti Forestalis Fenniae 70: 1–78.
Lahti T, Väisänen RA. 1987. Ecological gradients of boreal forests in south Finland: an ordination test of Cajander’s forest type theory. Vegetatio 68: 145–156.
Leake JR, Donnelly DP, Boddy L. 2002. Interactions between ectomycorrhizal and saprotrophic fungi. In: van der Heijden MGA, Sanders T, eds. Mycorrhizal ecology. Ecological Studies, Vol. 187. Berlin, Germany: Springer-Verlag, 346–372.
Leake JR, Donnelly DP, Saunders EM, Boddy L, Read DJ. 2001. Rates and quantities of carbon flux to ectomycorrhizal mycelium following 14C pulse labelling of Pinus sylvestris seedlings: effects of litter patches and interaction with wood-decomposer fungus. Tree Physiology. 21: 71–82.
Leake JR, Miles W. 1996. Phosphodiesters as mycorrhizal P sources I. Phosphodiesterase production and the utilization of DNA as a phosphorus source by the ericoid mycorrhizal fungus Hymenoscyphus ericae. New Phytologist 132: 435–443.
Leake JR, Read DJ. 1989. The effects of phenolic compounds on nitrogen mobilization by ericoid mycorrhizal systems. Agriculture, Ecosystems and Environment 29: 225–236.
Leake JR, Read DJ. 1990a. Chitin as a nitrogen source for mycorrhizal fungi. Mycological Research 94: 993–1008.
Leake JR, Read DJ. 1990b. Proteinase activity in mycorrhizal fungi. I. The effect of extracellular pH on the production and activity of proteinase by ericoid. New Phytologist 115: 243–250.
Leake JR, Read DJ. 1997. Mycorrhizal fungi in terrestrial habitats. In: Wicklow D, Söderström B, eds. The mycota IV environmental and microbial relationships. Berlin, Germany: Springer-Verlag, 281–301.
Lee J, Kenkel T, Booth T. 1996. Atmospheric deposition of macronutrients by pollen in the boreal forest. Ecoscience 3: 304–309.
Lilleskov EA, Fahey TJ, Horton TR, Lovett GM. 2002a. Belowground ectomycorrhizal fungal community change over a nitrogen deposition gradient in Alaska. Ecology 83: 104–115.
Lilleskov EA, Hobbie EA, Fahey TJ. 2002b. Ectomycorrhizal fungal taxa differing in response to nitrogen deposition also differ in pure culture organic nitrogen use and natural abundance of nitrogen isotopes. New Phytologist 154: 219–231.
Lindahl B, Stenlid J, Olsson S, Finlay RD. 1999. Translocation of 32P between interacting mycelia of a wood-decomposing and ectomycorrhizal fungi in microcosm systems. New Phytologist 144: 183–193.
Mäder P, Vierhailig H, Streitwolf-Engel R, Boller T, Frey B, Christie P, Wiemken A. 2000. Transport of 15N from soil compartment separated by a polytetrafluoroethylene membrane to plant roots via the hyphae of arbuscular mycorrhizal fungi. New Phytologist 146: 155–161.
Maijala P, Fagerstedt KV, Raudaskoski M. 1991. Detection of extracellular cellololytic and proteolytic activity in ectomycorrhizal fungi and Heterobasidion annosum (Fr.) Bref. New Phytologist 117: 643–648.
Mengel K. 1996. Turnover of organic nitrogen in soils and its availability to crops. Plant and Soil 181: 83–93.
Michelsen AS, Schmidt IK, Jonasson S, Quarmby C, Sleep S. 1996. Leaf 15N abundance of subarctic plants provides field evidence that ericoid, ectomycorrhizal and non- and arbuscular mycorrhizal species access different sources of soil nitrogen. Oecologia 105: 53–63.

Research review
www.newphytologist.com © New Phytologist (2003) 157: 475–492
Review492
Mitchell DT, Sweeney M, Kennedy A. 1992. Chitin degradation by Hymenoscyphus ericae and the influence of H. ericae on the growth of ectomycorrhizal fungi. In: Read DJ, Lewis DH, Fitter AH, Alexander IJ, eds. Mycorrhizas in ecosystems. Wallingford, UK: CAB International, 246–251.
Molina R, Massicotte H, Trappe JM. 1992. Specificity phenomena in mycorrhizal symbioses: Community-ecological consequences and practical implications. In: Allen MF, ed. Mycorrhizal functioning: an integrative plant–fungal process. New York, USA: Chapman & Hall, 357–423.
Moyersoen B, Becker P, Alexander IJ. 2001. Are ectomycorrhizas more abundant than arbuscular mycorrhizas in tropical heath forests? New Phytologist 150: 591–599.
Moyersoen B, Fitter AH, Alexander IJ. 1998. Spatial distribution of ectomycorrhizas and arbuscular mycorrhizas in Korup National Park rain forest, Cameroon, in relation to edaphic parameters. New Phytologist 139: 311–320.
Myers MD, Leake JR. 1996. Phosphodiesters as mycorrhizal P sources II. Ericoid mycorrhiza and the utilization of nuclei as a phosphorus source by Vaccinium macrocarpon. New Phytologist 132: 445–451.
Näsholm T, Ekbald A, Nordin A, Gieslr R, Högberg M, Högberg P. 1998. Boreal forest plants take up organic nitrogen. Nature 392: 914–916.
Näsholm T, Persson J. 2001. Plant acquisition of organic nitrogen in boreal forests. Physiologia Plantarum 111: 419–426.
Newbery DM, Alexander IJ, Rother JA. 1997. Phosphorus dynamics in a lowland Africian rain forest: the influence of ectomycorrhizal trees. Ecological Monographs 67: 367–409.
Nurmiaho-Lassila E-L, Timonen S, Haahtela K, Sen R. 1997. Bacterial colonization patterns of intact Pinus sylvestris mycorrhizospheres in dry pine forest soil: an electron microscopy study. Canadian Journal of Microbiology 43: 1017–1035.
Oleksyn J, Reich PB, Karolewski P, Tjoelker MG, Chalupka W. 1999. Nutritional status of pollen and needles of diverse Pinus sylvestris populations grown at sites with contrasting pollution. Water, Air and Soil Pollution 110: 195–212.
Peretto R, Bettini V, Bonfante P. 1993. Evidence of two polygalacturonases produced by a mycorrhizal ericoid fungus during its saprophytic growth. FEMS Microbiology Letters 114: 85–92.
Perez-Moreno J, Read DJ. 2000. Mobilization and transfer of nutrients from litter to tree seedlings via the vegetative mycelium of ectomycorrhizal plants. New Phytologist 145: 301–309.
Perez-Moreno J, Read DJ. 2001a. Exploitation of pollen by mycorrhizal mycelial systems with special reference to nutrient recycling in boreal forests. Proceedings of the Royal Society of London B 268: 1329–1335.
Perez-Moreno J, Read DJ. 2001b. Nutrient transfer from soil nematodes to plants: a direct pathway provided by the mycorrhizal mycelial network. Plant, Cell & Environment 24: 1219–1226.
Perotto S, Coisson JD, Perugini I, Cometti V, Bonfante P. 1997. Production of pectin-degrading enzymes by ericoid mycorrhizal fungi. New Phytologist 135: 151–162.
Petersen H. 1982. Structure and size of animal populations. Oikos 39: 306–329.
Post WM, Emanuel WR, Zinke PJ, Stangenberger AG. 1982. Soil carbon pools and world life zones. Nature 298: 156–159.
Raab TK, Lipson DA, Monson RK. 1996. Non-mycorrhizal uptake of amino acids by the roots of the alpine sedge Kobresia myosuroides: implications for the alpine nitrogen cycle. Oecologia 108: 488–494.
Read DJ. 1991. Mycorrhizas in ecosystems. Experientia 47: 376–391.Read DJ. 1992. The mycorrhizal mycelium. In: Allen MF, ed. Mycorrhizal.
London, UK: Chapman & Hall, 102–133.Read DJ. 1996. The structure and function of the ericoid mycorrhizal root.
Annals of Botany 77: 365–374.
Ryan EA, Alexander IJ. 1992. Mycorrhizal aspects of improved growth of spruce when grown in mixed stands on heathlands. In: Read DJ, Lewis DH, Fitter AH, Alexander IJ, eds. Mycorrhizas in ecosystems. Wallingford, UK: CAB International, 237–245.
Schulze E-D, Högberg P, Van Oene H, Persson T, Harrison AF, Read DJ, Kjøller A, Matteucci G. 2000. Interactions between the carbon and nitrogen cycles and the role of biodiversity: A synopsis of a study along a north–south transect through Europe. In: Schulze E-D, ed. Carbon Nitrogen Cycling in European forest ecosystems. Ecological Studies 142. Berlin, Germany: Springer-Verlag, 468–491.
Smith SE, Read DJ. 1997. Mycorrhizal Symbiosis, 2nd edn. San Diego, USA: Academic Press.
Stark N. 1972. Nutrient cycling pathways and litter fungi. Bioscience 22: 355–360.
Stevenson FJ. 1982. Humus chemistry. New York, USA: Wiley Interscience, 443.
Tarafdar JC, Marschner H. 1994. Phosphatase-activity in the rhizosphere and hyphosphere of VA mycorrhizal wheat supplied with inorganic and organic phosphorus. Soil Biology and Biochemistry Vol: 387–395.
Taylor AFS, Martin F, Read DJ. 2000. Fungal diversity in ectomycorrhizal communities of Norway spruce (Picea abies (L.) Karst.) and Beech (Fagus sylvatica L.) in forests along north–south transects in Europe. In: Schulze E-D, ed. Carbon nitrogen cycling in European forest ecosystems. Ecological Studies 142. Berlin, Germany: Springer-Verlag, 343–365.
Terashita T, Kono M, Yoshikawa K, Shishiyama J. 1995. Productivity of hydrolytic enzymes by mycorrhizal mushrooms. Mycoscience 36: 221–225.
Tibbett M, Sanders FE, Cairney JWG, Leake JR. 1999. Temperature regulation of extracellular proteases in ectomycorrhizal fungi (Hebeloma spp.) grown in axenic culture. Mycological Research 103: 707–714.
Tibbett M, Sanders FE, Minto SJ, Dowell M, Cairney JWG. 1998. Utilization of organic nitrogen by ectomycorrhizal fungi (Hebeloma spp.) of arctic and temperate origin. Mycological Research 102: 1525–1532.
Timonen S, Jørgensen KS, Haahtela K, Sen R. 1998. Bacterial community structure at defined locations of Pinus sylvestris-Suillus bovines and Pinus sylvestris-Paxillus involutus mycorrhizospheres in dry pine forest humus and nursery peat. Canadian Journal of Microbiology 44: 499–513.
Timonen S, Sen R. 1998. Heterogeneity of fungal and plant enzyme expression in intact Scots pine-Suillus bovines and -Paxillus involutus mycorrhizospheres developed in natural forest humus. New Phytologist 138: 355–366.
Trappe JM. 1987. Phylogenetic and ecologic aspects of mycotrophy in the angiosperms from an evolutionary standpoint. In: Safir GR, ed. Ecophysiology of va mycorrhizal plants. Boca Raton, FL, USA: CRC Press, 5–25.
Turnbull MH, Goodall R, Stewart GR. 1995. The impact of mycorrhizal colonisation upon nitrogen source utilisation and metabolism in seedlings of Eucalyptus grandis Hill ex Maiden and Eucalyptus maculata Hook. Plant, Cell & Environment 18: 1386–1394.
Varma A, Bonfante P. 1994. Utilization of cell-wall related carbohydrates by ericoid mycorrhizal endophytes. Symbiosis 16: 301–313.
White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfaud DH, Sninsky JJ, White TJ, eds. PCR protocols. A guide to methods and applications. San Diego, CA, USA: Academic Press, 315–322.
Zhu H, Dancik BP, Higginbotham KO. 1994. Regulation of extracellular proteinase production in an ectomycorrhizal fungus Hebeloma crustuliniforme. Mycologia 86: 227–234.
Zhu H, Guo D, Dancik BP. 1990. Purification and characterization of an extracellular acid proteinase from the ectomycorrhizal fungus. Hebeloma crustuliniforme. Applied and Environmental Microbiology 56: 837–743.




















