
Central β-amyloid peptide-induced peripheral interleukin-6 responses in mice: β-Amyloid peptide...
10
Journal of Neurochemistry, 2001, 76, 1326–1335 Central b-amyloid peptide-induced peripheral interleukin-6 responses in mice Dong-Keun Song, Yeong-Bin Im, Jun-Sub Jung, Jaeyoung Cho, Hong-Won Suh and Yung-Hi Kim Department of Pharmacology, College of Medicine, Institute of Natural Medicine, Hallym University, Chunchon, South Korea Abstract b-Amyloid peptides (Abs) share with lipopolysaccharide, a potent pro-inflammatory agent, the property of stimulating glial cells or macrophages to induce various inflammatory media- tors. We recently reported that central administration of lipopolysaccharide induces peripheral interleukin-6 responses via both the central and peripheral norepinephrine system. In this study, the effect of intracerebroventricular injection of various synthetic Abs on plasma interleukin-6 levels was examined in mice. Ab 1242 dose-dependently increased plasma interleukin-6 levels: ‘aged’ Ab 1242 was more effective than fresh, whereas Ab 4221 had no effect. ‘Aged’ Ab 1242 (205 pmol/mouse i.c.v.)-induced plasma interleukin-6 peaked at 2 h post injection, which is earlier than the peak time of the Ab 1242 -induced brain interleukin-6, tumor necrosis factor-a and interleukin-1b levels, which was 4, 4 and 24 h, respec- tively. Among various peripheral organs, Ab 1242 (205 pmol/ mouse i.c.v.) significantly increased interleukin-6 mRNA expression in lymph nodes and liver. Ab 1242 (205 pmol/ mouse i.c.v.) significantly increased norepinephrine turnover in both hypothalamus and spleen. Either central or peripheral norepinephrine depletion effectively inhibited the Ab 1242 - induced peripheral interleukin-6 response. Pretreatment with prazosin (a 1 -adrenergic antagonist), yohimbine (a 2 -adrener- gic antagonist), and ICI-118,551 (b 2 -adrenergic antagonist), but not with betaxolol (b 1 -adrenergic antagonist), inhibited Ab 1242 -induced plasma interleukin-6 levels. These results demonstrate that centrally administered Ab 1242 effectively induces the systemic interleukin-6 response which is mediated, in part, by central Ab 1242 -induced activation of the central and the peripheral norepinephrine systems. Keywords: Alzheimer’s disease, 6-hydroxydopamine, intra- cerebroventricular injection, norepinephrine, sympathetic nervous system. J. Neurochem. (2001) 76, 1326–1335. Accumulating evidence demonstrates that the inflammatory process plays an important role in Alzheimer’s disease (AD) (Aisen 1997; Hull et al. 1999; McGeer and McGeer 1999; Combs et al. 2000). b-Amyloid peptide (Ab), a major component of senile plaque, is postulated to play a pivotal role in inducing the inflammatory process in the AD brain; Ab can activate glial cells (astrocytes and microglia) in culture to induce or potentiate the production of a variety of inflammatory products, including interleukin (IL)-1b, tumor necrosis factor (TNF)-a, IL-6, IL-8, nitric oxide (NO) and reactive oxygen species (Araujo and Cotman 1992; Gitter et al. 1995; Meda et al. 1995; Walker et al. 1995; Ii et al. 1996; McDonald et al. 1997). Furthermore, the glial/ macrophage-stimulating properties of Abs and some of their signal transduction pathways are shared with lipo- polysaccharide (LPS), a potent mediator of inflammation; both stimuli similarly activate tyrosine kinases of src family, such as Syk, and Lyn (Boulet et al. 1992; Crowley et al. 1996; McDonald et al. 1997; Combs et al. 1999), and mitogen-activated protein kinases (MAPK) (McDonald et al. 1998; Pyo et al. 1998; Combs et al. 1999) in glia/ macrophages. Interleukin-6 is a multifunctional cytokine involved in the regulation of immune responses, hematopoiesis, and acute- phase reactions (Hirano 1998). Head injury (Kossmann et al. 1995), and various neurological diseases, including ischemic stroke (Fassbender et al. 1994), multiple sclerosis (Frei et al. 1326 q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326–1335 Received May 19, 2000; revised manuscript received October 2, 2000; accepted October 6, 2000. Address correspondence and reprints requests to D.-K. Song, Department of Pharmacology and Institute of Natural Medicine, College of Medicine, Hallym University, Chunchon, Kangwon-Do, 200-702, South Korea. E-mail: [email protected] Abbreviations used: Ab, b-amyloid peptide; AD, Alzheimer’s disease; IL, interleukin; i.c.v., intracerebroventricular; i.p., intra- peritoneal; LPS, lipopolysaccharide; MAPK, mitogen-activated protein kinases; NE, norepinephrine; NO, nitric oxide; 6-OHDA, 6-hydroxy- dopamine; TNF-a, tumor necrosis factor-a.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Central β-amyloid peptide-induced peripheral interleukin-6 responses in mice: β-Amyloid peptide...

Journal of Neurochemistry, 2001, 76, 1326±1335
Central b-amyloid peptide-induced peripheral interleukin-6
responses in mice
Dong-Keun Song, Yeong-Bin Im, Jun-Sub Jung, Jaeyoung Cho, Hong-Won Suh and Yung-Hi Kim
Department of Pharmacology, College of Medicine, Institute of Natural Medicine, Hallym University, Chunchon, South Korea
Abstract
b-Amyloid peptides (Abs) share with lipopolysaccharide, a
potent pro-in¯ammatory agent, the property of stimulating glial
cells or macrophages to induce various in¯ammatory media-
tors. We recently reported that central administration of
lipopolysaccharide induces peripheral interleukin-6 responses
via both the central and peripheral norepinephrine system. In
this study, the effect of intracerebroventricular injection of
various synthetic Abs on plasma interleukin-6 levels was
examined in mice. Ab1242 dose-dependently increased plasma
interleukin-6 levels: `aged' Ab1242 was more effective than
fresh, whereas Ab4221 had no effect. `Aged' Ab1242
(205 pmol/mouse i.c.v.)-induced plasma interleukin-6 peaked
at 2 h post injection, which is earlier than the peak time of the
Ab1242-induced brain interleukin-6, tumor necrosis factor-a
and interleukin-1b levels, which was 4, 4 and 24 h, respec-
tively. Among various peripheral organs, Ab1242 (205 pmol/
mouse i.c.v.) signi®cantly increased interleukin-6 mRNA
expression in lymph nodes and liver. Ab1242 (205 pmol/
mouse i.c.v.) signi®cantly increased norepinephrine turnover
in both hypothalamus and spleen. Either central or peripheral
norepinephrine depletion effectively inhibited the Ab1242-
induced peripheral interleukin-6 response. Pretreatment with
prazosin (a1-adrenergic antagonist), yohimbine (a2-adrener-
gic antagonist), and ICI-118,551 (b2-adrenergic antagonist),
but not with betaxolol (b1-adrenergic antagonist), inhibited
Ab1242-induced plasma interleukin-6 levels. These results
demonstrate that centrally administered Ab1242 effectively
induces the systemic interleukin-6 response which is
mediated, in part, by central Ab1242-induced activation of
the central and the peripheral norepinephrine systems.
Keywords: Alzheimer's disease, 6-hydroxydopamine, intra-
cerebroventricular injection, norepinephrine, sympathetic
nervous system.
J. Neurochem. (2001) 76, 1326±1335.
Accumulating evidence demonstrates that the in¯ammatory
process plays an important role in Alzheimer's disease (AD)
(Aisen 1997; Hull et al. 1999; McGeer and McGeer 1999;
Combs et al. 2000). b-Amyloid peptide (Ab), a major
component of senile plaque, is postulated to play a pivotal
role in inducing the in¯ammatory process in the AD brain;
Ab can activate glial cells (astrocytes and microglia) in
culture to induce or potentiate the production of a variety of
in¯ammatory products, including interleukin (IL)-1b, tumor
necrosis factor (TNF)-a, IL-6, IL-8, nitric oxide (NO) and
reactive oxygen species (Araujo and Cotman 1992; Gitter
et al. 1995; Meda et al. 1995; Walker et al. 1995; Ii et al.
1996; McDonald et al. 1997). Furthermore, the glial/
macrophage-stimulating properties of Abs and some of
their signal transduction pathways are shared with lipo-
polysaccharide (LPS), a potent mediator of in¯ammation;
both stimuli similarly activate tyrosine kinases of src family,
such as Syk, and Lyn (Boulet et al. 1992; Crowley et al.
1996; McDonald et al. 1997; Combs et al. 1999), and
mitogen-activated protein kinases (MAPK) (McDonald et al.
1998; Pyo et al. 1998; Combs et al. 1999) in glia/
macrophages.
Interleukin-6 is a multifunctional cytokine involved in the
regulation of immune responses, hematopoiesis, and acute-
phase reactions (Hirano 1998). Head injury (Kossmann et al.
1995), and various neurological diseases, including ischemic
stroke (Fassbender et al. 1994), multiple sclerosis (Frei et al.
1326 q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326±1335
Received May 19, 2000; revised manuscript received October 2, 2000;
accepted October 6, 2000.
Address correspondence and reprints requests to D.-K. Song,
Department of Pharmacology and Institute of Natural Medicine,
College of Medicine, Hallym University, Chunchon, Kangwon-Do,
200-702, South Korea. E-mail: [email protected]
Abbreviations used: Ab, b-amyloid peptide; AD, Alzheimer's
disease; IL, interleukin; i.c.v., intracerebroventricular; i.p., intra-
peritoneal; LPS, lipopolysaccharide; MAPK, mitogen-activated protein
kinases; NE, norepinephrine; NO, nitric oxide; 6-OHDA, 6-hydroxy-
dopamine; TNF-a, tumor necrosis factor-a.

1991), and AD (Kalman et al. 1997; Singh and Guthikonda
1997; Bonaccorso et al. 1998), have been associated with
increased plasma IL-6 levels. Recently, it has been shown
that the central administration of several potent mediators of
in¯ammation, such as IL-1b (De Simoni et al. 1990, 1993;
Romero et al. 1996; Kitamura et al. 1998), TNF (Benigni
et al. 1996) or LPS (Gottschall et al. 1992; Muramami et al.
1993; De Simoni et al. 1995), increases plasma levels of IL-
6 and IL-6 mRNA expression in peripheral organs,
demonstrating that the in¯ammatory process in the CNS
induces systemic IL-6 responses (Terreni and De Simoni
1998).
We hypothesized that central administration of Abs may
induce systemic IL-6 responses. In this study we examined
the effect of intracerebroventricular (i.c.v.) injection of Abs
on the plasma IL-6 levels and IL-6 mRNA expression in
various peripheral tissues in mice.
Materials and methods
Animals and i.c.v. injection
Male ICR mice weighing 25±30 g (Myung-Jin, Inc., Seoul, South
Korea) were used in all experiments. The animals were housed ®ve
per cage in a room maintained at 22 ^ 18C with an alternating 12-h
light/dark cycle. Food and water were available ad libitum. All
animal procedures were carried out as approved by the Animal
Care and Use Committee at Hallym University College of
Medicine. The i.c.v. administration was performed following the
procedure established by Laursen and Belknap (1986), which was
modi®ed from the method of Haley and McCormick (1957). The
animal was injected at bregma with a 50-mL Hamilton syringe
®tted with a 26-gauge needle, which was adjusted to be inserted
2.4 mm deep. The i.c.v. injection volume was 5 mL, and injection
sites were veri®ed by injecting the same volume of 1% methylene
blue into the site and then observing the distribution of the injected
dye in the ventricular space.
Reagents
Ab1242, Ab1240, Ab25235, Ab1228 (RBI, Natick, MA, USA) and
Ab4221 (Bachem, Bubendorf, Switzerland) were prepared as stock
solutions in sterile deionized water, and aliquots were stored at
2208C. Ab1240 and Ab1242 were used either fresh or after the
`aging' process, i.e. incubation of Abs in saline at ®nal concen-
tration at 378C for 3 days (Pike et al. 1993). Ab25235 and Ab1228
were used only in the `aged' state. Phentolamine mesylate, pro-
pranolol HCl, prazosin HCl, and ICI-118,551 HCl were purchased
from RBI, yohimbine HCl, and methyl ester of a-methyl-p-tyrosine
were from Sigma Chemical Co. (St Louis, MO, USA), and
betaxolol HCl was from Tocris Cookson Ltd. (Bristol, UK).
Prazosin HCl and yohimbine HCl were dissolved in saline con-
taining 10 and 20% dimethylsulfoxide, respectively. Other drugs
were dissolved in sterile saline.
Plasma and brain cytokine assay
Plasma was separated by centrifugation of the freshly drawn blood
from the retro-orbital venous plexus and stored at 2708C until
assayed. The plasma levels of IL-6 as well as IL-1b and TNF-a were
determined 1.5 h after an i.c.v. injection of Ab1242 with an ELISA
kit (Genzyme, Cambridge, MA, USA). Whole brain at various time
points after `aged' Ab1242 injection was homogenized with 10 vol.
of phosphate-buffered saline (pH 7.2) and centrifuged at 10 000 g
for 20 min twice, and the supernatant was used for cytokine assay.
Assays were performed exactly as described by the manufacturers.
Reverse transcription (RT) PCR
In the experiment on the tissue distribution of central Ab1242-
induced IL-6 mRNA expression various peripheral tissues [pituit-
ary, adrenals, kidney, heart, liver, spleen, and lymph nodes (iliac)]
were collected at 1 h after Ab1242 administration. The number of
animals used for each experiment was seven for pituitary, ®ve for
adrenals, and three for all other tissues. Total cellular RNAs were
extracted from tissues using a rapid guanidinium thiocyanate±
water saturated phenol±chloroform extraction and subsequent
precipitation with acidic sodium acetate (Chomczynski and Sacchi
1987). Total cellular RNAs in the aqueous phase were precipitated
using cold isopropyl alcohol. Isolated RNA samples were subjected
to spectrophotometric analysis at 260 and 280 nm, and samples
were stored at 2708C until use. RNA was denatured by incubating
at 708C for 5 min and then chilled quickly to 48C. cDNA synthesis
was conducted on 1 mg total RNA. The reaction mixture for the
synthesis of cDNA by RT reaction included the following: 15 mm
MgCl2; 5� reaction buffer containing 375 mm KCl and 250 mm
Tris±HCl (pH 8.3); 100 mm each dATP, dCTP, dGTP, and dTTP
(Pharmacia); ribonuclease inhibitor (RNasin, 40 U/mL, Promega);
and Moloney murine leukemia virus reverse transcriptase (200 U/mL,
GibcoBRL). One microgram of sample RNA was added to 20 mm
oligo(dT)16 primer, RT master mix, which contained 10 mm of
each dNTP, and 1 U of RNasin. The RT reaction mixture was
incubated in a Techine-PHCZ thermal cycler at 258C for 10 min,
378C for 60 min, 998C for 5 min, 48C for 5 min, and stored at
2208C. The PCR mixture contained the following: 15 mm MgCl2;
10� reaction buffer containing 500 mm KCl, 100 mm Tris±
HCl (pH 8.3), and 0.01% (w/v) gelatin; and Taq DNA polymerase
(5 U/mL; Perkin-Elmer). Primers for IL-6 and b-actin were
synthesized at Bohan Biomedical Inc. (Seoul, South Korea). The
sequences of these primers were as described previously (Faulkner
et al. 1995); b-actin, 5 0-TGGAATCCTGTGGCATCCATGAAAC-
3 0, 5 0-TAAAACGCAGCTCAGTAACAGTCCG-3 0 (348 bp); IL-6,
5 0-TGGAGTCACAGAAGGAGTGGCTAAG-3 0, 5 0-TCTGACCA-
CAGTGAGGAATGTCCAC-3 0 (155 bp). For each reaction, 30 mL
of master mix containing 10� reaction buffer, 0.5 U of Taq DNA
polymerase, and 20 mm of each primer was added to a tube
containing 3 mL of the cDNA synthesized in the RT reaction. The
tubes were incubated in a thermal cycler at 958C for 2 min (once),
948C for 45 s, 678C for 2 min, and 728C for 3 min (28±31 cycles),
and 728C for 10 min, and then held at 48C. PCR products were
visualized by ethidium bromide staining after agarose (1.2%) gel
electrophoresis. The density of each spot was quanti®ed using a
BIO-1D image analyzer. The unregulated mRNA encoding b-actin
was used as an internal loading control.
Depletion of either central or peripheral norepinephrine by
either central or peripheral injection of 6-hydroxydopamine
6-Hydroxydopamine (6-OHDA) hydrobromide (Sigma), dissolved
in sterile 0.1% ascorbic acid, was injected either i.c.v. or i.p. to
b-Amyloid peptide and interleukin-6 1327
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326±1335

deplete either central or peripheral stores of norepinephrine (NE).
Three days after the 6-OHDA injection, NE contents were
measured in hypothalamus and spleen, as a representative tissue
for evaluating central and peripheral NE depletion, respectively.
Hypothalamus and spleen were homogenized in 0.1 m perchloric
acid containing 0.1 m sodium metabisul®te. After these samples
were centrifuged using a refrigerating microcentrifuge for 10 min,
the supernatant was ®ltered through a 0.45-mm Millipore HV-4
®lter. Ten microliters of sample was injected onto a C18 m-
Bondapak column (Waters, Milford, MA, USA). As a mobile
phase, 0.1 m KH2PO4 (adjusted to pH 3.8) containing sodium
octanesulfonic acid (0.25 mm), disodium EDTA (0.1 mm) and
acetonitrile (9%, v/v) was used. The ¯ow rate was 1 mL/min, and
the oxidation potential was 1 V. Previously, an i.c.v. injection of
6-OHDA at the dose of 50 mg/mouse was shown to cause a 73.1%
decrease in hypothalamic NE content without affecting dopamine
(DA) and serotonin content when measured 3 days after injection
(Suaudeau et al. 1995). We showed that the same dose of 6-OHDA
induced an 85.6% decrease in hypothalamic NE content without
affecting DA content 3 days after injection (Song et al. 1999a). In
contrast, an intraperitoneal injection of 6-OHDA (100 mg/kg)
induced an 86.6% decrease in NE content without changes in DA
content in spleen, when measured 3 days after injection (Song et al.
1999a). This treatment did not affect the hypothalamic NE and DA
content (Song et al. 1999a).
NE turnover
NE turnover was assessed by a decrease in tissue NE concentration
after the inhibition of NE biosynthesis with a-methyl-p-tyrosine.
Mice were injected i.c.v. with saline, Ab4221 (205 pmol/mouse), or
Ab1242 (205 pmol/mouse) and then i.p. with methyl ester of
a-methyl-p-tyrosine (300 mg/kg). Before (time 0) and 4 h after
a-methyl-p-tyrosine injection, NE levels in the hypothalamus and
spleen were determined.
Endotoxin assay
Levels of endotoxin in Abs solution were measured using an
E-Toxate kit (Sigma) and were not detected.
Statistical analysis
Statistical analysis was carried out by one-way (Fig. 1a) or two-
way (Figs 1b, 3, 4, 5b and c) analysis of variance (anova).
Bennett's test was used for post hoc comparisons. Student's t-test
was used for Table 1 and Figs 1(c) and 5(a). p-Values , 0.05 were
considered to indicate statistical signi®cance.
Results
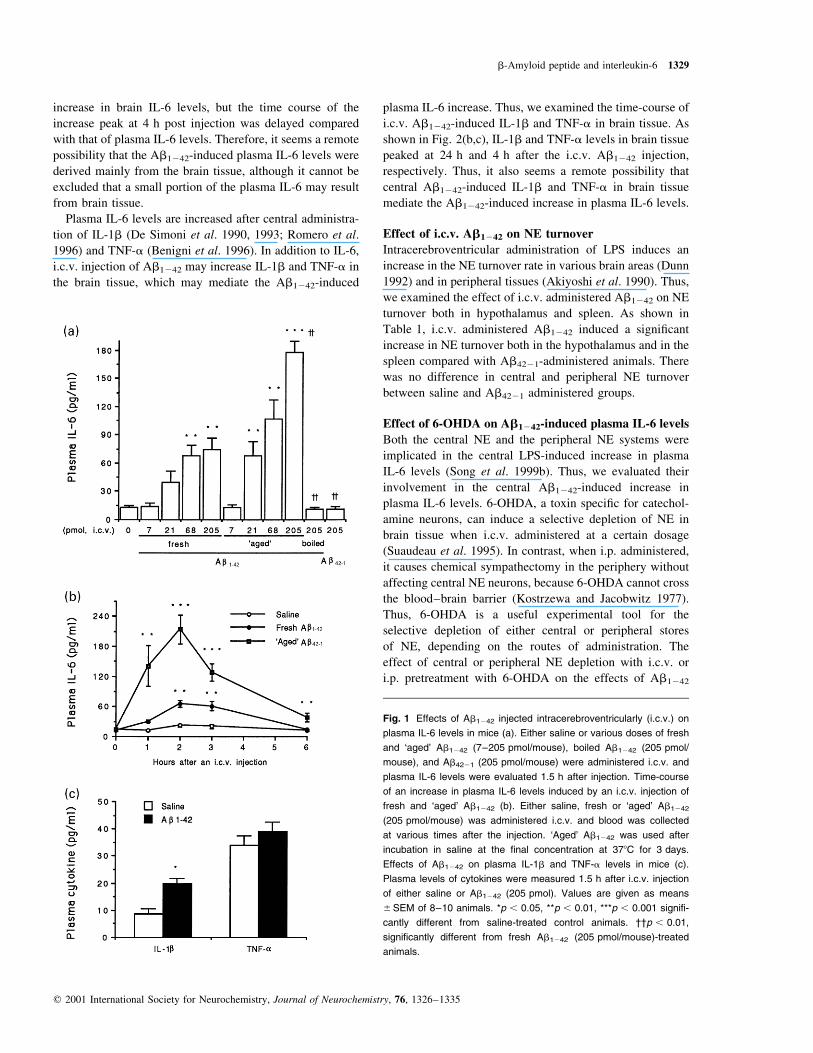
Effect of i.c.v. Abs on plasma IL-6 levels
Various synthetic Abs, including Ab1242, Ab1240, Ab25235,
and Ab1228 were administered i.c.v., and plasma IL-6 levels
were measured at 1.5 h after injection. The native full-
length fresh Ab1242 induced a dose-dependent increase in
plasma IL-6 levels (F � 9.97, p , 0.001; Fig. 1a). Ab1242-
induced plasma IL-6 levels returned to a basal level by 6 h
after injection (Fig. 1b). Ab4221 (205 pmol) and boiled
Ab1242 (205 pmol) had no effect. `Aged' Ab1242 was more
effective than fresh (Fig. 1a); plasma IL-6 levels induced
by `aged' Ab1242 peaked at 2 h, remained increased at
6 h (Fig. 1b), and returned to basal levels 24 h after injec-
tion (data not shown). Because `fresh' Ab1242 effectively
induced an increase in plasma IL-6 in this study, we used
`fresh' Ab1242 in subsequent experiments. `Aged', but not
fresh, Ab1240 increased plasma IL-6 levels slightly at a
dose of 410 pmol/mouse i.c.v. (13 ^ 2 and 30 ^ 4 pg/mL
for saline-treated and Ab1240-treated animals, respectively;
p , 0.05). Intracerebroventricular Ab1228 and Ab25235 had
no effect at a dose of 205 pmol/mouse (data not shown).
IL-6, IL-1b and TNF-a are major pro-in¯ammatory
cytokines. Thus, it was of interest to study the effects of
i.c.v. administered Ab1242 on the plasma levels of IL-1b
and TNF-a. As shown in Fig. 1(c), Ab1242 (205 pmol/
mouse) induced a slight, but signi®cant, increase in plasma
IL-1b but not TNF-a levels. `Aged' Ab1240 (410 pmol/
mouse) also induced a slight but signi®cant increase in
plasma IL-1b (9 ^ 2 and 22 ^ 4 pg/mL for saline-,
Ab1240-treated animals, respectively; p , 0.05) but not
TNF-a levels.
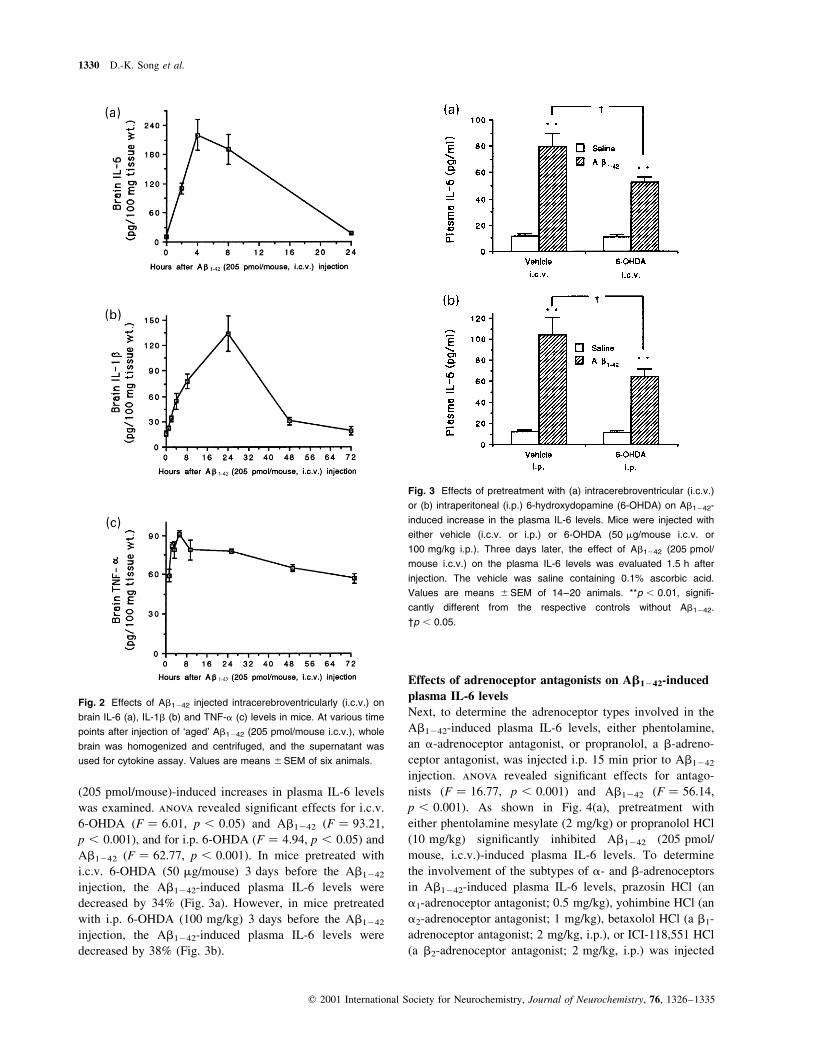
Effect of i.c.v. Ab1242 on brain IL-6, IL-1b and
TNF-a levels
Intracerebroventricular injection of Ab1242 may increase
IL-6 in the brain tissue, which may leak into the systemic
circulation. Thus we examined the time-course of the
Ab1242-induced increase in IL-6 in the brain. As shown in
Fig. 2(a), Ab1242 (205 pmol/mouse) induced a marked
Table 1. Effects of intracerebroventricular injection of Ab1242 on norepinephrine turnover in the hypothalamus and spleen
4 h after a-methyl-p-tyrosine
Time 0 Saline Ab4221 Ab1242
Hypothalamus 2185� ^ 100 1123� ^ 98 1167� ^ 71 823� ^ 47*
Spleen 274� ^ 24 154� ^ 8 155� ^ 10 128� ^ 6*
Mice were injected intracerebroventricularly with Ab4221 (205 pmol/mouse) or Ab1242 (205 pmol/mouse) and then intraperitoneally with the methyl
ester of a-methyl-p-tyrosine (300 mg/kg) 4 h before norepinephrine assay. The data are means ^SE in ng/g tissue weight of six animals.
*p , 0.05, signi®cantly different from the saline-or Ab4221-treated controls.
1328 D.-K. Song et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326±1335
increase in brain IL-6 levels, but the time course of the
increase peak at 4 h post injection was delayed compared
with that of plasma IL-6 levels. Therefore, it seems a remote
possibility that the Ab1242-induced plasma IL-6 levels were
derived mainly from the brain tissue, although it cannot be
excluded that a small portion of the plasma IL-6 may result
from brain tissue.
Plasma IL-6 levels are increased after central administra-
tion of IL-1b (De Simoni et al. 1990, 1993; Romero et al.
1996) and TNF-a (Benigni et al. 1996). In addition to IL-6,
i.c.v. injection of Ab1242 may increase IL-1b and TNF-a in
the brain tissue, which may mediate the Ab1242-induced
plasma IL-6 increase. Thus, we examined the time-course of
i.c.v. Ab1242-induced IL-1b and TNF-a in brain tissue. As
shown in Fig. 2(b,c), IL-1b and TNF-a levels in brain tissue
peaked at 24 h and 4 h after the i.c.v. Ab1242 injection,
respectively. Thus, it also seems a remote possibility that
central Ab1242-induced IL-1b and TNF-a in brain tissue
mediate the Ab1242-induced increase in plasma IL-6 levels.
Effect of i.c.v. Ab1242 on NE turnover
Intracerebroventricular administration of LPS induces an
increase in the NE turnover rate in various brain areas (Dunn
1992) and in peripheral tissues (Akiyoshi et al. 1990). Thus,
we examined the effect of i.c.v. administered Ab1242 on NE
turnover both in hypothalamus and spleen. As shown in
Table 1, i.c.v. administered Ab1242 induced a signi®cant
increase in NE turnover both in the hypothalamus and in the
spleen compared with Ab4221-administered animals. There
was no difference in central and peripheral NE turnover
between saline and Ab4221 administered groups.
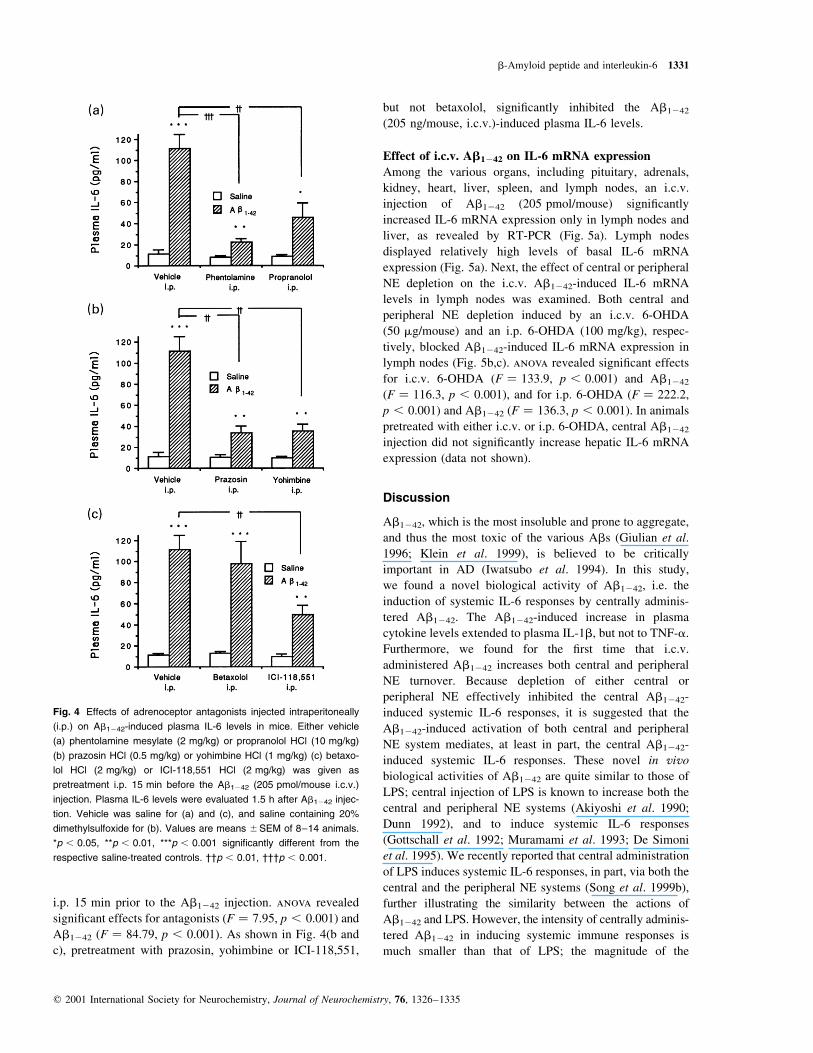
Effect of 6-OHDA on Ab1242-induced plasma IL-6 levels
Both the central NE and the peripheral NE systems were
implicated in the central LPS-induced increase in plasma
IL-6 levels (Song et al. 1999b). Thus, we evaluated their
involvement in the central Ab1242-induced increase in
plasma IL-6 levels. 6-OHDA, a toxin speci®c for catechol-
amine neurons, can induce a selective depletion of NE in
brain tissue when i.c.v. administered at a certain dosage
(Suaudeau et al. 1995). In contrast, when i.p. administered,
it causes chemical sympathectomy in the periphery without
affecting central NE neurons, because 6-OHDA cannot cross
the blood±brain barrier (Kostrzewa and Jacobwitz 1977).
Thus, 6-OHDA is a useful experimental tool for the
selective depletion of either central or peripheral stores
of NE, depending on the routes of administration. The
effect of central or peripheral NE depletion with i.c.v. or
i.p. pretreatment with 6-OHDA on the effects of Ab1242
Fig. 1 Effects of Ab1242 injected intracerebroventricularly (i.c.v.) on
plasma IL-6 levels in mice (a). Either saline or various doses of fresh
and `aged' Ab1242 (7±205 pmol/mouse), boiled Ab1242 (205 pmol/
mouse), and Ab4221 (205 pmol/mouse) were administered i.c.v. and
plasma IL-6 levels were evaluated 1.5 h after injection. Time-course
of an increase in plasma IL-6 levels induced by an i.c.v. injection of
fresh and `aged' Ab1242 (b). Either saline, fresh or `aged' Ab1242
(205 pmol/mouse) was administered i.c.v. and blood was collected
at various times after the injection. `Aged' Ab1242 was used after
incubation in saline at the ®nal concentration at 378C for 3 days.
Effects of Ab1242 on plasma IL-1b and TNF-a levels in mice (c).
Plasma levels of cytokines were measured 1.5 h after i.c.v. injection
of either saline or Ab1242 (205 pmol). Values are given as means
^SEM of 8±10 animals. *p , 0.05, **p , 0.01, ***p , 0.001 signi®-
cantly different from saline-treated control animals. ²²p , 0.01,
signi®cantly different from fresh Ab1242 (205 pmol/mouse)-treated
animals.
b-Amyloid peptide and interleukin-6 1329
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326±1335
(205 pmol/mouse)-induced increases in plasma IL-6 levels
was examined. anova revealed signi®cant effects for i.c.v.
6-OHDA (F � 6.01, p , 0.05) and Ab1242 (F � 93.21,
p , 0.001), and for i.p. 6-OHDA (F � 4.94, p , 0.05) and
Ab1242 (F � 62.77, p , 0.001). In mice pretreated with
i.c.v. 6-OHDA (50 mg/mouse) 3 days before the Ab1242
injection, the Ab1242-induced plasma IL-6 levels were
decreased by 34% (Fig. 3a). However, in mice pretreated
with i.p. 6-OHDA (100 mg/kg) 3 days before the Ab1242
injection, the Ab1242-induced plasma IL-6 levels were
decreased by 38% (Fig. 3b).
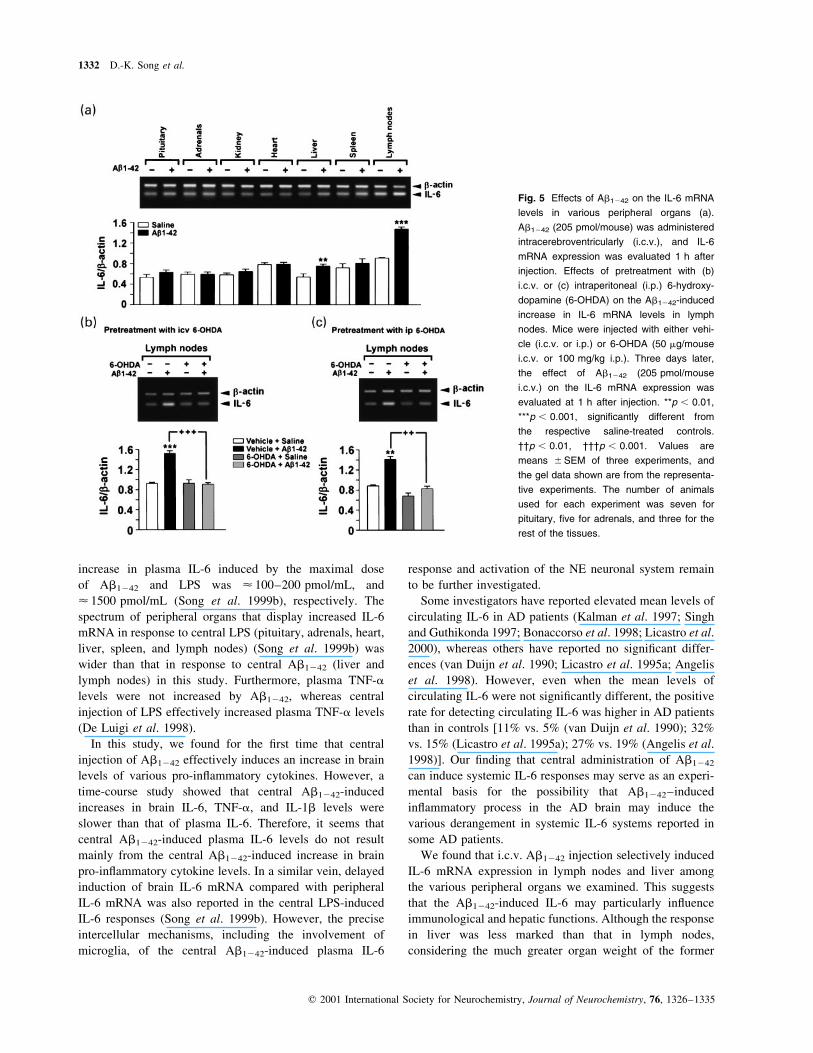
Effects of adrenoceptor antagonists on Ab1242-induced
plasma IL-6 levels
Next, to determine the adrenoceptor types involved in the
Ab1242-induced plasma IL-6 levels, either phentolamine,
an a-adrenoceptor antagonist, or propranolol, a b-adreno-
ceptor antagonist, was injected i.p. 15 min prior to Ab1242
injection. anova revealed signi®cant effects for antago-
nists (F � 16.77, p , 0.001) and Ab1242 (F � 56.14,
p , 0.001). As shown in Fig. 4(a), pretreatment with
either phentolamine mesylate (2 mg/kg) or propranolol HCl
(10 mg/kg) signi®cantly inhibited Ab1242 (205 pmol/
mouse, i.c.v.)-induced plasma IL-6 levels. To determine
the involvement of the subtypes of a- and b-adrenoceptors
in Ab1242-induced plasma IL-6 levels, prazosin HCl (an
a1-adrenoceptor antagonist; 0.5 mg/kg), yohimbine HCl (an
a2-adrenoceptor antagonist; 1 mg/kg), betaxolol HCl (a b1-
adrenoceptor antagonist; 2 mg/kg, i.p.), or ICI-118,551 HCl
(a b2-adrenoceptor antagonist; 2 mg/kg, i.p.) was injected
Fig. 2 Effects of Ab1242 injected intracerebroventricularly (i.c.v.) on
brain IL-6 (a), IL-1b (b) and TNF-a (c) levels in mice. At various time
points after injection of `aged' Ab1242 (205 pmol/mouse i.c.v.), whole
brain was homogenized and centrifuged, and the supernatant was
used for cytokine assay. Values are means ^SEM of six animals.
Fig. 3 Effects of pretreatment with (a) intracerebroventricular (i.c.v.)
or (b) intraperitoneal (i.p.) 6-hydroxydopamine (6-OHDA) on Ab1242-
induced increase in the plasma IL-6 levels. Mice were injected with
either vehicle (i.c.v. or i.p.) or 6-OHDA (50 mg/mouse i.c.v. or
100 mg/kg i.p.). Three days later, the effect of Ab1242 (205 pmol/
mouse i.c.v.) on the plasma IL-6 levels was evaluated 1.5 h after
injection. The vehicle was saline containing 0.1% ascorbic acid.
Values are means ^SEM of 14±20 animals. **p , 0.01, signi®-
cantly different from the respective controls without Ab1242.
²p , 0.05.
1330 D.-K. Song et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326±1335
i.p. 15 min prior to the Ab1242 injection. anova revealed
signi®cant effects for antagonists (F � 7.95, p , 0.001) and
Ab1242 (F � 84.79, p , 0.001). As shown in Fig. 4(b and
c), pretreatment with prazosin, yohimbine or ICI-118,551,
but not betaxolol, signi®cantly inhibited the Ab1242
(205 ng/mouse, i.c.v.)-induced plasma IL-6 levels.
Effect of i.c.v. Ab1242 on IL-6 mRNA expression
Among the various organs, including pituitary, adrenals,
kidney, heart, liver, spleen, and lymph nodes, an i.c.v.
injection of Ab1242 (205 pmol/mouse) signi®cantly
increased IL-6 mRNA expression only in lymph nodes and
liver, as revealed by RT-PCR (Fig. 5a). Lymph nodes
displayed relatively high levels of basal IL-6 mRNA
expression (Fig. 5a). Next, the effect of central or peripheral
NE depletion on the i.c.v. Ab1242-induced IL-6 mRNA
levels in lymph nodes was examined. Both central and
peripheral NE depletion induced by an i.c.v. 6-OHDA
(50 mg/mouse) and an i.p. 6-OHDA (100 mg/kg), respec-
tively, blocked Ab1242-induced IL-6 mRNA expression in
lymph nodes (Fig. 5b,c). anova revealed signi®cant effects
for i.c.v. 6-OHDA (F � 133.9, p , 0.001) and Ab1242
(F � 116.3, p , 0.001), and for i.p. 6-OHDA (F � 222.2,
p , 0.001) and Ab1242 (F � 136.3, p , 0.001). In animals
pretreated with either i.c.v. or i.p. 6-OHDA, central Ab1242
injection did not signi®cantly increase hepatic IL-6 mRNA
expression (data not shown).
Discussion
Ab1242, which is the most insoluble and prone to aggregate,
and thus the most toxic of the various Abs (Giulian et al.
1996; Klein et al. 1999), is believed to be critically
important in AD (Iwatsubo et al. 1994). In this study,
we found a novel biological activity of Ab1242, i.e. the
induction of systemic IL-6 responses by centrally adminis-
tered Ab1242. The Ab1242-induced increase in plasma
cytokine levels extended to plasma IL-1b, but not to TNF-a.
Furthermore, we found for the ®rst time that i.c.v.
administered Ab1242 increases both central and peripheral
NE turnover. Because depletion of either central or
peripheral NE effectively inhibited the central Ab1242-
induced systemic IL-6 responses, it is suggested that the
Ab1242-induced activation of both central and peripheral
NE system mediates, at least in part, the central Ab1242-
induced systemic IL-6 responses. These novel in vivo
biological activities of Ab1242 are quite similar to those of
LPS; central injection of LPS is known to increase both the
central and peripheral NE systems (Akiyoshi et al. 1990;
Dunn 1992), and to induce systemic IL-6 responses
(Gottschall et al. 1992; Muramami et al. 1993; De Simoni
et al. 1995). We recently reported that central administration
of LPS induces systemic IL-6 responses, in part, via both the
central and the peripheral NE systems (Song et al. 1999b),
further illustrating the similarity between the actions of
Ab1242 and LPS. However, the intensity of centrally adminis-
tered Ab1242 in inducing systemic immune responses is
much smaller than that of LPS; the magnitude of the
Fig. 4 Effects of adrenoceptor antagonists injected intraperitoneally
(i.p.) on Ab1242-induced plasma IL-6 levels in mice. Either vehicle
(a) phentolamine mesylate (2 mg/kg) or propranolol HCl (10 mg/kg)
(b) prazosin HCl (0.5 mg/kg) or yohimbine HCl (1 mg/kg) (c) betaxo-
lol HCl (2 mg/kg) or ICI-118,551 HCl (2 mg/kg) was given as
pretreatment i.p. 15 min before the Ab1242 (205 pmol/mouse i.c.v.)
injection. Plasma IL-6 levels were evaluated 1.5 h after Ab1242 injec-
tion. Vehicle was saline for (a) and (c), and saline containing 20%
dimethylsulfoxide for (b). Values are means ^SEM of 8±14 animals.
*p , 0.05, **p , 0.01, ***p , 0.001 signi®cantly different from the
respective saline-treated controls. ²²p , 0.01, ²²²p , 0.001.
b-Amyloid peptide and interleukin-6 1331
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326±1335
increase in plasma IL-6 induced by the maximal dose
of Ab1242 and LPS was < 100±200 pmol/mL, and
< 1500 pmol/mL (Song et al. 1999b), respectively. The
spectrum of peripheral organs that display increased IL-6
mRNA in response to central LPS (pituitary, adrenals, heart,
liver, spleen, and lymph nodes) (Song et al. 1999b) was
wider than that in response to central Ab1242 (liver and
lymph nodes) in this study. Furthermore, plasma TNF-a
levels were not increased by Ab1242, whereas central
injection of LPS effectively increased plasma TNF-a levels
(De Luigi et al. 1998).
In this study, we found for the ®rst time that central
injection of Ab1242 effectively induces an increase in brain
levels of various pro-in¯ammatory cytokines. However, a
time-course study showed that central Ab1242-induced
increases in brain IL-6, TNF-a, and IL-1b levels were
slower than that of plasma IL-6. Therefore, it seems that
central Ab1242-induced plasma IL-6 levels do not result
mainly from the central Ab1242-induced increase in brain
pro-in¯ammatory cytokine levels. In a similar vein, delayed
induction of brain IL-6 mRNA compared with peripheral
IL-6 mRNA was also reported in the central LPS-induced
IL-6 responses (Song et al. 1999b). However, the precise
intercellular mechanisms, including the involvement of
microglia, of the central Ab1242-induced plasma IL-6
response and activation of the NE neuronal system remain
to be further investigated.
Some investigators have reported elevated mean levels of
circulating IL-6 in AD patients (Kalman et al. 1997; Singh
and Guthikonda 1997; Bonaccorso et al. 1998; Licastro et al.
2000), whereas others have reported no signi®cant differ-
ences (van Duijn et al. 1990; Licastro et al. 1995a; Angelis
et al. 1998). However, even when the mean levels of
circulating IL-6 were not signi®cantly different, the positive
rate for detecting circulating IL-6 was higher in AD patients
than in controls [11% vs. 5% (van Duijn et al. 1990); 32%
vs. 15% (Licastro et al. 1995a); 27% vs. 19% (Angelis et al.
1998)]. Our ®nding that central administration of Ab1242
can induce systemic IL-6 responses may serve as an experi-
mental basis for the possibility that Ab1242±induced
in¯ammatory process in the AD brain may induce the
various derangement in systemic IL-6 systems reported in
some AD patients.
We found that i.c.v. Ab1242 injection selectively induced
IL-6 mRNA expression in lymph nodes and liver among
the various peripheral organs we examined. This suggests
that the Ab1242-induced IL-6 may particularly in¯uence
immunological and hepatic functions. Although the response
in liver was less marked than that in lymph nodes,
considering the much greater organ weight of the former
Fig. 5 Effects of Ab1242 on the IL-6 mRNA
levels in various peripheral organs (a).
Ab1242 (205 pmol/mouse) was administered
intracerebroventricularly (i.c.v.), and IL-6
mRNA expression was evaluated 1 h after
injection. Effects of pretreatment with (b)
i.c.v. or (c) intraperitoneal (i.p.) 6-hydroxy-
dopamine (6-OHDA) on the Ab1242-induced
increase in IL-6 mRNA levels in lymph
nodes. Mice were injected with either vehi-
cle (i.c.v. or i.p.) or 6-OHDA (50 mg/mouse
i.c.v. or 100 mg/kg i.p.). Three days later,
the effect of Ab1242 (205 pmol/mouse
i.c.v.) on the IL-6 mRNA expression was
evaluated at 1 h after injection. **p , 0.01,
***p , 0.001, signi®cantly different from
the respective saline-treated controls.
²²p , 0.01, ²²²p , 0.001. Values are
means ^SEM of three experiments, and
the gel data shown are from the representa-
tive experiments. The number of animals
used for each experiment was seven for
pituitary, ®ve for adrenals, and three for the
rest of the tissues.
1332 D.-K. Song et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326±1335

compared with the latter, the contribution of liver to central
Ab1242-induced plasma IL-6 levels may not be less, or may
even be more, than that of lymph nodes. Because IL-6 is a
major cytokine involved in hepatic acute phase protein
synthesis, the Ab1242-induced increase in hepatic IL-6
mRNA in this study may be related to previous reports of
increased serum levels of a-antichymotrypsin, an acute-
phase protein, in some AD patients (Matsubara et al. 1990;
Brugge et al. 1992; Hinds et al. 1994; Altstiel et al. 1995;
Licastro et al. 1995a, b, 1997, 2000; Lieberman et al. 1995).
Among the various Abs with differing peptide lengths,
Ab1242 was the most effective; it increased plasma IL-6
levels even in the fresh state, whereas Ab1240 was margin-
ally effective only after the `aging' process, i.e. formation of
®brillar peptide, and other synthetic Abs were without
effect. Interestingly, this pattern of activity is almost iden-
tical to the neurotoxicity pro®le of various Abs via
stimulation of microglia (Giulian et al. 1996); in that report
Ab1242 is more potent than Ab1240 in activating microglia
to kill neurons, whereas other Abs, including Ab1228 and
Ab25235, have no effect. Similarly, non®brillar Ab1242 and
®brillar, but not non®brillar, Ab1240 have been shown to
increase LPS-induced IL-1b release from monocytes
(Lorton et al. 1996). The result of this study, which is in
line with the well-known phenomenon that the `aging'
process, i.e. the formation of ®brillar peptide, induces or
increases neurotoxicity (Pike et al. 1993) and the glial/
monocyte stimulating activity (Gitter et al. 1995; Lorton
et al. 1996; McDonald et al. 1997) of various Abs, suggests
that the activity of centrally administered Abs in inducing a
peripheral IL-6 response is also dependent on the degree of
aggregation or ®brillar formation. Furthermore, the lack of
effect of Ab4221 on NE turnover and plasma IL-6 levels
indicates the necessity for the speci®c conformation of
Ab1242 in activating NE system and in inducing peripheral
IL-6 responses.
In conclusion, this study demonstrates that centrally
administered Ab1242 effectively induces systemic IL-6
responses, which are mediated, at least in part, by central
Ab1242-induced activation of the central and the peripheral
NE systems. These activities of Ab1242, together with the
induction of pro-in¯ammatory cytokine levels in the brain
tissue, further illustrate the immune stimulating activity of
Ab1242 in vivo.
Acknowledgements
This work was supported by the Korea Ministry of Education
(Basic Medicine Research Fund 1997 (997-021-F0026) and
Brain Korea 21 Program), KOSEF through the Brain Disease
Research Center at Ajou University and the Korea Research
Foundation (1998-019-F00047). We thank Drs Yoo-Hun Suh
and Sung-Oh Huh for encouragement and kind advise,
Dr Choon-Gon Jang for statistical analysis and Mr Gordon
Slysz for revision of the English text.
References
Aisen P. S. (1997) In¯ammation and Alzheimer's disease: mechanisms
and therapeutic strategies. Gerontology 43, 143±149.
Akiyoshi M., Shimizu Y. and Saito M. (1990) Interleukin-1 increases
norepinephrine turnover in the spleen and lung in rats. Biochem.
Biophys. Res. Commun. 173, 1266±1270.
Altstiel L. D., Lawlor B. A., Mohs R. C., Schmeidler J., Dalbn A.,
Mehta P. and Davis K. L. (1995) Elevated alpha 1-antic-
hymotrypsin serum levels in a subset of nondemented ®rst-degree
relatives of Alzheimer disease patients. Dementia 6, 17±20.
Angelis P., Scharf S., Mander A., Vajda F. and Christophidis N. (1998)
Serum interleukin-6 and interleukin-6 soluble receptor in
Alzheimer's disease. Neurosci. Lett. 244, 106±108.
Araujo D. M. and Cotman C. W. (1992) b-Amyloid stimulates glial
cells in vitro to produce growth factors that accumulate in senile
plaques in Alzheimer's disease. Brain Res. 569, 141±145.
Benigni F., Faggioni R., Sironi M., Fantuzzi G., Vandenabeele P.,
Takahashi N., Sacco S., Fiers W., Buurman W. A. and Ghezzi P.
(1996) TNF receptor p55 plays a major role in centrally mediated
increases of serum IL-6 and corticosterone after intracerebroven-
tricular injection of TNF. J. Immunol. 157, 5563±5568.
Bonaccorso S., Lin A., Song C., Verkerk R., Kenis G., Bosmans E.,
Scharpe S., Vandewoude M., Dossche A. and Maes M. (1998)
Serotonin±immune interactions in elderly volunteers and in
patients with Alzheimer's disease (DAT): lower plasma trypto-
phan availability to the brain in the elderly and increased serum
interleukin-6 in DAT. Aging (Milano) 10, 316±323.
Boulet I., Ralph S., Stanley E., Lock P., Dunn A. R., Green S. P. and
Phillips W. A. (1992) Lipopolysaccharide- and interferon-gamma-
induced expression of hck and lyn tyrosine kinases in murine bone
marrow-derived macrophages. Oncogene 7, 703±710.
Brugge K., Katzman R., Hill L. R., Hansen L. A. and Saitoh T. (1992)
Serological alpha 1 antichymotrypsin in Down's syndrome and
Alzheimer's disease. Ann. Neurol. 32, 192±197.
Chomczynski P. and Sacchi N. (1987) Single-step method of RNA
isolation by acid guanidium thiocyanate±phenol±chloroform
extraction. Anal. Biochem. 162, 156±159.
Combs C. K., Johnson D. E., Cannady S. B., Lehman T. M. and
Landreth G. E. (1999) Identi®cation of microglial signal trans-
duction pathways mediating a neurotoxic response to amyloido-
genic fragments of beta-amyloid and prion proteins. J. Neurosci.
19, 928±939.
Combs C. K., Johnson D. E., Karlo J. C., Cannady S. B. and Landreth
G. E. (2000) In¯ammatory mechanisms in Alzheimer's disease:
inhibition of beta-amyloid-stimulated proin¯ammatory responses
and neurotoxicity by PPARgamma agonists. J. Neurosci. 20, 558±
567.
Crowley M. T., Harmer S. L. and DeFranco A. L. (1996) Identi®cation
of microglial signal transduction pathways mediating a neurotoxic
response to amyloidogenic fragments of beta-amyloid and prion
proteins. J. Biol. Chem. 271, 1145±1152.
De Luigi A., Terreni L., Sironi M. and De Simoni M. G. (1998) The
sympathetic nervous system tonically inhibits peripheral inter-
leukin-1b and interleukin-6 induction by central lipopolysaccharide.
Neuroscience 83, 1245±1250.
De Simoni M. G., De Sironi M., Luigi A., Manfridi A., Mantovani A.
and Ghezzi P. (1990) Intracerebroventricular injection of inter-
leukin-1 induces high circulating levels of interleukin-6. J. Exp.
Med. 171, 1773±1778.
b-Amyloid peptide and interleukin-6 1333
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326±1335

De Simoni M. G., Luigi A., Gemma L., Sironi M., Manfridi A. and
Ghezzi P. (1993) Modulation of systemic interleukin-6 induction
by central interleukin-1. Am. J. Physiol. 265 (Regul. Integ. Comp.
Physiol. 34,) R739±R742.
De Simoni M. G., del Bo R., Luigi A., Simard S. and Forloni G. (1995)
Central endotoxin induces different patterns of interleukin (IL)-1b
and IL-6 messenger ribonucleic acid expression and IL-6 secretion
in the brain and periphery. Endocrinology 136, 897±902.
van Duijn C. M., Hofman A. and Nagelkerken L. (1990) Serum levels
of interleukin-6 are not elevated in patients with Alzheimer's
disease. Neurosci. Lett. 108, 350±354.
Dunn A. J. (1992) Endotoxin-induced activation of cerebral catecho-
lamine and serotonin metabolism: comparison with interleukin-1.
J. Pharmacol. Exp. Ther .261, 964±949.
Fassbender K., Rossol S., Kammer T., Daffertshofer M., Wirth S.,
Dollman M. and Hennerici M. (1994) Proin¯ammatory cytokines
in serum of patients with acute cerebral ischemia: kinetics of
secretion and relation to the extent of brain damage and outcome
of disease. J. Neurol. Sci. 122, 135±139.
Faulkner C. B., Simecka J. W., Davidson M. K., Davis J. K., Schoeb
T. R., Lindsey J. R. and Everson M. P. (1995) Gene expression
and production of tumor necrosis alpha, interleukin 1, inter-
leukin 6, and gamma interferon in C3H/HeN and C57BL/6N mice
in acute Mycoplasma pulmonis disease. Infect. Immun. 63, 4084±
4090.
Frei K., Fredrikson S., Fontana A. and Link H. (1991) Interleukin-6 is
elevated in plasma in multiple sclerosis. J. Neuroimmunol. 31,
147±153.
Gitter B. D., Cox L. M., Rydel R. E. and May P. C. (1995) Amyloid b
peptide potentiates cytokine secretion by interleukin-1b-activated
human astrocytoma cells. Proc. Natl Acad. Sci. USA 92, 10738±
10741.
Giulian D., Haverkamp L. J., Yu J. H., Karshin W., Tom D., Li J.,
Kirkpatrick J., Kuo L. M. and Roher A. E. (1996) Speci®c
domains of beta-amyloid from Alzheimer plaque elicit neuron
killing in human microglia. J. Neurosci. 16, 6021±6037.
Gottschall P. E., Komaki G. and Arimura A. (1992) Increased
circulating interleukin-1 and interleukin-6 after intracerebroven-
tricular injection of lipopolysaccharide. Neuroendocrinology 56,
935±938.
Haley T. J. and McCormick W. G. (1957) Pharmacological effects
produced by intracerebral injection of drugs in the conscious
mouse. Br. J. Pharmacol. 12, 12±15.
Hinds T. R., Kukull W. A., Van Belle G., Schellenberg G. D., Villacres
E. C. and Larson E. B. (1994) The relationship between serum
alpha 1 antichymotrypsin and Alzheimer's disease. Neurobiol.
Aging 15, 21±27.
Hirano T. (1998) Interleukin-6, in The Cytokine Handbook (Thomson
A., ed. ), pp. 197±228. Academic Press, London.
Hull M., Fiebich B. L., Schumann G., Lieb K. and Bauer J. (1999)
Anti-in¯ammatory substances ± a new therapeutic option in
Alzheimer's disease. Drug Discov. Today 4, 275±282.
Ii M., Sunamoto M., Ohnishi K. and Ichimori Y. (1996) Beta-amyloid
protein-dependent nitric oxide production from microglial cells
and neurotoxicity. Brain Res. 720, 93±100.
Iwatsubo T., Odaka A., Suzuki N., Mizusawa H., Nukina N. and Ihara
Y. (1994) Visualization of A beta 42 (43) and A beta 40 in
senile plaques with end-speci®c A beta monoclonals: evidence
that an initially deposited species is A beta 42. Neuron 13, 45±
53.
Kalman J., Juhasz A., Laird G., Dickens P., Jardanhazy T., Rimanoczy
A., Boncz I., Parry-Jones W. L. and Janka Z. (1997) Serum
interleukin-6 levels correlate with the severity of dementia in
Down's syndrome and in Alzheimer's disease. Acta Neurol.
Scand. 96, 236±240.
Kitamura H., Okamoto S., Shimamoto Y., Morimatsu M., Terao A. and
Saito M. (1998) Central IL-1 differentially regulates peripheral
IL-6 and TNF synthesis. Cell. Mol. Life Sci. 54, 282±287.
Klein A. M., Kowall N. W. and Ferrante R. J. (1999) Neurotoxicity
and oxidative damage of beta amyloid 1±42 versus beta amyloid
1±40 in the mouse cerebral cortex. Ann. NY Acad. Sci. 893, 314±
320.
Kossmann T., Hans V. H., Imhof H. G., Stocker R., Grob P., Trentz O.
and Morganti-Kossmann C. (1995) Intrathecal and serum inter-
leukin-6 and the acute-phase response in patients with severe
traumatic brain injuries. Shock 4, 311±317.
Kostrzewa R. M. and Jacobwitz D. M. (1977) Pharmacological actions
of 6-hydroxydopamine. Pharmacol. Rev. 26, 199±288.
Laursen S. E. and Belknap J. K. (1986) Intracerebroventricular injec-
tions in mice. J. Pharmacol. Methods 16, 355±357.
Licastro F., Morini M. C., Polazzi E. and Davis L. J. (1995a) Increased
serum a1-antichymotrypsin in patients with probable Alzheimer's
disease: an acute phase reactant without the peripheral acute phase
response. J. Neuroimmunol. 57, 71±75.
Licastro F., Parnetti L., Morini M. C., Davis L. J., Cucinotta D., Gaiti A.
and Senin U. (1995b) Acute phase reactant a1-antichymotrypsin
is increased in cerebrospinal ¯uid and serum of patients with
probable Alzheimer's disease. Alzheimer Dis. Assoc. Dis. 9, 112±
118.
Licastro F., Sirri V., Trere D. and Davis L. J. (1997) Monomeric and
polymeric forms of a1-antichymotrypsin in sera from patients
with probable late onset Alzheimer's disease. Dement. Geriatr.
Cogn. Disord. 8, 337±342.
Licastro F., Pedrini S., Caputo L., Annoni G., Davis L. J., Ferri C.,
Casadei V. and Grimaldi L. M. (2000) Increased plasma levels of
interleukin-1, interleukin-6 and alpha-1-antichymotrypsin in
patients with Alzheimer's disease: peripheral in¯ammation or
signals from the brain? J. Neuroimmunol. 103, 97±102.
Lieberman J., Schleissner L., Tachiki K. H. and Kling A. S. (1995)
Serum a1-antichymotrypsin level as a marker for Alzheimer-type
dementia. Neurobiol. Aging 16, 747±753.
Lorton D., Kocsis J. M., King L., Madden K. and Brunden K. R. (1996)
Beta-amyloid induces increased release of interleukin-1 beta
from lipopolysaccharide-activated human monocytes. J. Neuro-
immunol. 67, 21±29.
Matsubara E., Hirai S., Amari M., Shoji M., Yamaguchi H., Okamoto
K., Ishiguro K., Harigawa Y. and Wakabayashi K. (1990) a1-
Antichymotrypsin as a possible biochemical marker for Alzhei-
mer-type dementia. Ann. Neurol. 28, 561±567.
McDonald D. R., Brunden K. R. and Landreth G. E. (1997) Amyloid
®brils activate tyrosine kinase-dependent signaling and superoxide
production in microglia. J. Neurosci. 17, 2284±2294.
McDonald D. R., Bamberger M. E., Combs C. K. and Landreth G. E.
(1998) b-Amyloid ®brils activate parallel mitogen-activated
protein kinase pathways in microglia and THP1 monocytes.
J. Neurosci. 18, 4451±4460.
McGeer P. L. and McGeer E. G. (1999) In¯ammation of the brain in
Alzheimer's disease: implications for therapy. J. Leukoc. Biol. 65,
409±415.
Meda L., Cassatella M. A., Szendrei G. I., Otvos L. Jr, Baron P.,
Villalba M., Ferrari D. and Rossi F. (1995) Activation of
microglial cells by beta-amyloid protein and interferon-gamma.
Nature 374, 647±650.
Muramami N., Fukata J., Tsukada T., Kobayashi H., Ebisui O., Segawa
H., Muro S., Imura H. and Nakao K. (1993) Bacterial lipo-
polysaccharide-induced expression of interleukin-6 messenger
1334 D.-K. Song et al.
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326±1335

ribonucleic acid in the rat hypothalamus, pituitary, adrenal gland,
and spleen. Endocrinology 133, 2574±2578.
Pike C. J., Burdick D., Walencewicz A. J., Glabe C. G. and Cotman C.
W. (1993) Neurodegeneration induced by beta-amyloid peptides
in vitro: the role of peptide assembly state. J. Neurosci. 13, 676±
687.
Pyo H., Jou I., Jung S., Hong S. and Joe E. H. (1998) Mitogen-activated
protein kinases activated by lipopolysaccharide and beta-amyloid
in cultured rat microglia. Neuroreport 9, 871±874.
Romero L. I., Kakucska I., Lechan R. M. and Reichlin S. (1996)
Interleukin-6 (IL-6) is secreted from the brain after intracerebro-
ventricular injection of IL-1b in rats. Am. J. Physiol. 270 (Regul.
Integ. Comp. Physiol. 39,) R518±R524.
Singh V. K. and Guthikonda P. (1997) Circulating cytokines in
Alzheimer's disease. J. Psychiatr. Res. 31, 657±660.
Song D.-K., Im Y.-B., Jung J.-S., Suh H.-W., Huh S.-O., Song J.-H. and
Kim Y.-H. (1999a) Central injection of nicotine increases hepatic
and splenic interleukin (IL)-6 mRNA expression and plasma IL-6
levels in mice: involvement of peripheral sympathetic nervous
system. FASEB J. 13, 1259±1267.
Song D.-K., Im Y.-B., Jung J.-S., Suh H.-W., Huh S.-O., Park S.-W.,
Wie M.-B. and Kim Y.-H. (1999b) Differential involvement of
central and peripheral norepinephrine in the central LPS-induced
IL-6 responses in mice. J. Neurochem. 72, 1625±1633.
Suaudeau C., Dourmap N. and Costentin J. (1995) Rapid and long
lasting reduction of the hypothalamic effect of a D2 dopamine
agonist after an intracerebroventricular injection of 6-hydroxydo-
pamine. Neuropharmacology 34, 101±105.
Terreni L. and De Simoni M. G. (1998) Role of the brain in interleukin-
6 modulation. Neuroimmunomodulation 5, 214±219.
Walker D. G., Kim S. U. and McGeer P. L. (1995) Complement and
cytokine gene expression in cultured microglial derived from
postmortem human brains. J. Neurosci. Res. 4, 478±493.
b-Amyloid peptide and interleukin-6 1335
q 2001 International Society for Neurochemistry, Journal of Neurochemistry, 76, 1326±1335




















