
Intertextuality and hybrid discourses: The infusion of pop culture in educational discourse - 2004.
Upload
independentCategory
view
2download
0
Ameliorative Effects of a Neuroprotective Agent, T-817MA, on PlaceLearning Deficits Induced by Continuous Infusion of Amyloid-b
Peptide (1–40) in Rats
Phuong Thi Hong Nguyen,1,2 Tatsuo Kimura,2,3 Son Anh Ho,1,2 Anh Hai Tran,2,3
Taketoshi Ono,2,3 and Hisao Nishijo1,2*
ABSTRACT: Alzheimer’s disease (AD) is a neurodegenerative diseasecharacterized by cognitive decline due to neuronal loss and neural net-work dysfunction. It has been postulated that progressive neuronal lossin AD is consequence of the neurotoxic properties of the amyloid-b pep-tide (Ab). In the present study, we investigated the effect of T-817MA(1-{3-[2-(1-benzothiophen-5-yl)ethoxy] propyl}-3-azetidinol maleate), anewly synthesized neurotrophic compound, on place learning deficits inrats with hippocampal damages. To induce granule cell loss in thedentate gyrus (DG) of the hippocampus, Ab (1–40) was continuouslyinfused (300 pmol/day) into the cerebral ventricle using a mini-osmoticpump for 5 weeks. Three weeks after the Ab infusion, the rats weretested in a place learning task, which required them to alternatively visittwo diametrically opposed areas in an open field to obtain intracranialself-stimulation reward. The results indicated that the Ab-infused ratswithout treatment of T-817MA displayed learning impairment in thetask; their performance level was significantly inferior to that of thevehicle rats. Treatment of T-817MA (8.4 mg/kg/day, p.o.) significantlyimproved the task performance of the Ab-infused rats. Furthermore,T-817MA prevented granule cell loss due to Ab-infusion, which was cor-related to task performance of the rats. However, other cognitiveenhancer, an acetylcholinesterase inhibitor, had no such effects. Theresults demonstrated that T-817MA ameliorated learning deficitsinduced by Ab infusion, which might be attributed to neuroprotectionin the hippocampus. VVC 2007 Wiley-Liss, Inc.
KEY WORDS: dentate gyrus; hippocampus; memory; Alzheimer’sdisease; dementia
INTRODUCTION
Alzheimer’s disease (AD) is the most common cause of dementia, andthe number of AD patients is increasing year by year. In the brain ofAD patients as well as AD rodent model, large numbers of amyloid-bpeptides (Ab) containing neuritic plaques are found in neocortex andhippocampus, along with progressive cognitive impairment (Hsiao,2001). Several lines of evidence have implicated the hippocampal forma-tion (HF) as a possible locus for spatial and nonspatial memory deficits
in aged animals (Barnes and McNaughton, 1985;Ward et al., 1999; Stoelzel et al., 2002). The HF isthe most susceptible to senescence-related pathologicalchanges, such as those in senile dementia and humanAD (Marchbanks, 1982; Hyman et al., 1984; Ottet al., 1993; Carr et al., 1997).
It was reported that abnormal extracellular accumu-lation of Ab protein in the brain is one of the mainfeatures of AD (Nitta et al., 1994; Behl, 1997; Varadar-ajan et al., 2000; Bayer et al., 2001; Suh and Cheler,2002; Racchi and Govoni, 2003). Ab is a 4-kDa poly-peptide derived by the proteolytic cleavage of b-amy-loid precursor protein (APP) (Selkoe, 1998). Based onthese findings, animal models of AD have been pro-posed to delineate the mechanisms that cause braindysfunction, and to identify a variety of new therapeu-tic target and novel treatment. Continuous intracere-bral ventricular (i.c.v.) infusion of Ab (1–40) (300pmol/day) in rats resulted in learning and memory def-icits, suggesting that accumulation of Ab in the brainis related to cognitive impairments in AD (Nabeshimaand Nitta, 1994; Nitta et al., 1994; Yamada et al.,1998). The other studies showed that chronic infusionof Ab (1–40) into the rat brain induced memory defi-cits, which became evident from 7th day and continuedat least up to 14th day of the infusion (Yamada et al.,1999a,b; Yamada and Nabeshima, 2000).
Currently, many studies on various kinds of drugsare being conducted to treat AD using animal models ofAD. In the present study, 1-{3-[2-(1-Benzothiophen-5-yl)ethoxy]propyl}-3-azetidinol maleate (T-817MA) wastested as a therapeutic agent for neurodegenerative dis-orders such as AD (Fig. 1). It has been reported thatT-817MA protected neurons against Ab and hydrogenperoxide-induced neurotoxicity, and promoted neuriteoutgrowth in HF slice cultures and reaggregation cul-tures of rat cortical neurons (Hirata et al., 2005).These results suggest that T-817MA works as a neuro-trophic factor such as insulin-like growth factor-1(IGF-1) that has neuroprotective effects by promotingneurogenesis, development, differentiation, synapseformation, and glucose utilization throughout thebrain (Aberg et al., 2000; Trejo et al., 2001; Venters,2001). In the present study, we investigated effect ofT-817MA on performance of rats with Ab (1–40)
1 System Emotional Science, Graduate School of Medicine, University ofToyama, Sugitani 2630, Toyama 930-0194, Japan; 2CREST, JST, Tokyo,Japan; 3Molecular and Integrative Emotional Neuroscience, GraduateSchool of Medicine, University of Toyama, Sugitani 2630, Toyama 930-0194, Japan*Correspondence to: Dr. Hisao Nishijo, System Emotional Science, Grad-uate School of Medicine, University of Toyama, Sugitani 2630, Toyama930-0194, Japan. E-mail: [email protected] for publication 8 February 2007DOI 10.1002/hipo.20281Published online 30 March 2007 in Wiley InterScience (www.interscience.wiley.com).
HIPPOCAMPUS 17:443–455 (2007)
VVC 2007 WILEY-LISS, INC.

infusion to clarify its neuroprotective effect on central nervoussystem (CNS). The rats with and without Ab (1–40) infusionand with and without T-817MA treatment were tested withplace learning paradigms using intracranial self-stimulation(ICSS) as reward, in which performance of aged rats and youngrats with selective HF lesions were disturbed (Zhong et al.,2000; Hori et al., 2002; Kurimoto et al., 2004).
MATERIALS AND METHODS
Subjects
Male Crj:Wistar rats (7 weeks, n 5 49) were used. All ratswere given food and water ad libitum in a clear cage andhandled on three consecutive days before start of the experi-ments. The housing area was provided a temperature-controlledenvironment under a 12/12 h light circle (on at 08:00 h, offat 20:00 h). These rats were divided into five groups: vehicle(n 5 9), Ab-infused control (n 5 9), Ab-infusion 1 high-doseT-817MA (8.4 mg/kg) (n 5 11), Ab-infusion 1 low-doseT-817MA (0.84 mg/kg) (n 5 11), Ab-infusion 1 Donepezil(0.5 mg/kg) (n 5 9).
All experimental protocols were performed in accordancewith the guide for care and use of laboratory animals at Uni-versity of Toyama and the National Institutes of Health Guidefor the Care and Use of Laboratory Animals.
Surgery for Continuous Ab Infusion andIntracranial Self-Stimulation
Rats were anesthetized with pentobarbital (40 mg/kg, i.p.).An infusion cannula for Ab delivery and monopolar stimulat-ing electrodes (polyurethane insulated, 0.1-mm diameter,stainless steel) for ICSS reward were implanted into the leftventricle (A: 20.3 mm, L: 1.2 mm, V: 3.6 mm) and into themedial forebrain bundle (MFB) at the level of the lateral hypo-thalamus (LHA) (4.3 mm, 1.6 mm, and 8.7–8.9 mm) respec-tively, according to the atlas of Paxinos and Watson (1986). Ab(1–40) (AnaSpec, San Jose, CA) was dissolved in 30% acetoni-trile/ 0.1% trifluoroacetic acid. Continuous infusion of Ab(1–40) (300 pmol/day) was maintained for 5 weeks by a mini-osmotic pump (Alzet2002) connecting to an infusion cannula(Nabeshima and Nitta, 1994; Nitta et al., 1994; Yamada et al.,1998). The vehicle-treated rats were only infused with 30%acetonitrile/ 0.1% trifluoroacetic acid.
ICSS Training
After 1 week of recovery from electrode implantation, therats were screened for self-stimulation in an operant chamberequipped with a lever on one wall. Each lever press triggeredthe delivery of a 0.5 s train of 0.3 ms negative square wavepulses at 100 Hz. The current intensity for ICSS was deter-mined to produce 40–70 lever presses/min in the operantchamber. All rats used in the present study met this criterion.The rats were trained for ICSS for 7 days (15–30 min/day).
The use of ICSS as a reward provided the following advan-tages over natural reinforces such as food and water: (i) rapidlearning of task, (ii) lack of satiation, and (iii) absence of exter-nal sensory information (visual, auditory, olfactory, etc.)(Fukuda et al., 1992).
Experimental Set Up for Behavioral Testing
The apparatus used for the behavioral testing in the presentstudy was substantially the same as described previously(Fukuda et al., 1992; Kobayashi et al., 1997; Zhong et al.,2000; Tamura et al., 2001; Hori et al., 2002; Kurimoto et al.,2004). Schematic diagram of the apparatus was illustrated inFigure 2A.
For the three spatial tasks noted below, an open field (150-cm diameter) was used. The open field was enclosed by a blackcurtain. The ceiling of the enclosure contained four smallspeakers mounted near the circumference, spaced 908 apart,four light bulbs individually mounted near the inner edge ofeach speaker, and a charge coupled device (CCD) video cameraat the center (Fig. 2A). Usually a light bulb at the 3 o’clockposition and a white noise speaker at the 9 o’clock position onthe ceiling of the enclosure were turned on. A CCD cameraviewed the open field containing a rat from top center, to sig-nal the rat’s position in Cartesian coordinates. The video signalwas sent to a conventional TV monitor and a digital interface,which sent the X and Y coordinates of a miniature light bulbattached to the head of the rat to a microcomputer. The micro-computer plotted the trail of the rat, compared the rat’s behav-ior with preset criteria, and gated ICSS delivery as reward tothe MFB at the level of the LHA from a stimulator when thecriteria were met.
Behavioral Tests
After ICSS training, the rats were first tested in the openfield to measure spontaneous motor activity (SMA), then testedin the three kinds of spatial tasks: distance movement task(DMT), random reward place search task (RRPST), and placelearning task (PLT) in the open field. In each of these threespatial tasks, the small electric bulb on the head of the rat waslit at the start of a trial, and a train of ICSS current was deliv-ered to activate the rat. Each trial was terminated after 50rewards had been delivered or 10 min had elapsed, whicheveroccurred first. The rats were given training of five trials/day[see Fukuda et al. (1992) for details of this procedure].
FIGURE 1. Chemical structure of T-817MA.
444 NGUYEN ET AL.
Hippocampus DOI 10.1002/hipo

SMA
SMA was measured in the open field on the first train-ing day in the DMT. A rat was put on the center of the openfield. SMA of the rat was monitored by the CCD camera,
and distance traveled by the rat for 10 min was com-puted. There was no ICSS reward given to the rats duringthis activity. Then, the rat was subsequently trained with theDMT.
FIGURE 2. Scheme of experimental setup (A), behavioral par-adigm (B) and experimental schedule (C). (A) An open field (150-cm diameter) containing a rat was viewed from top center by aCCD video camera with brightness tracking interface that signaledthe rat’s position in Cartesian coordinates. The open field wasenclosed by a black curtain. The incandescent lamp at the 3o’clock position and a white noise speaker at the 9 o’clock posi-tion on the ceiling of the enclosure were turned on. The video sig-nal was sent to a conventional TV monitor and a digital interfacesent the X and Y coordinates of a miniature light bulb attached tothe head of the rat to a microcomputer. The microcomputer plot-ted the trail of the rat, compared the rat’s behavior with preset cri-teria, and gated rewarding intracranial self-stimulation (ICSS)from a stimulator to the lateral hypothalamic area (LHA) whenthe criteria were met. No distinctive local cues marked the rewardarea for the rat. (B) Behavioral paradigm. In DMT(a), a computer
program computed distance traveled from a trail. The rat acquiredICSS rewards if it moved the fixed distance (i.e., 120, 160, 200,240 cm). In RRPST (b), a computer program delimited a circularreward place (thick line circle; 70-cm diameter) at some randomlyselected coordinate. The rat was rewarded with ICSS when itentered the reward place, which was then made inactive (changedto thin line circle). After a 5 s interval, the reward place wasmoved to a different location randomly and reactivated. In PLT(c), the rat received rewards in two target areas (two thick linecircles; 40-cm diameter) when it returned to one reward place aftera visit to the other reward place. Large circle, open field; Start,location of the rat at start of a trial; line, trail of the rat; eachsmall open circle, the location of reward delivery. (C) After 1 weekfrom the start of Ab infusion and ICSS surgery, the rats werescreened for ICSS in an operant box. After the ICSS training, ratswere trained to perform spatial tasks in the open field.
NEUROPROTECTIVE AGENT AGAINST AMYLOID-b PATHOLOGY 445
Hippocampus DOI 10.1002/hipo

Distance movement task
The rats learned to move in the open field on this task (Fig.2Ba). A computer program computed the cumulative distancetraveled by the rat from the rats’ trail. The rat acquired ICSSrewards if it had moved the fixed distance (i.e., 120, 160, 200,or 240 cm). The predetermined fixed distance to acquire ICSSreward was progressively increased by 40 cm from 120 to 240 cmwhen the rat acquired 50 ICSS rewards within 10 min of onetrial. If the rat could not acquire 50 rewards, the trial was alsoterminated at the end of the 10 min. If the rat could acquire50 rewards within a trial, the distance was also increased by40 cm from the next trial. When the rat passed the criterion inthe DMT (i.e., distance of 160 cm), training in the DMT wascompleted. Thus, rats learned association of ICSS rewards withmovements in the DMT.
RRPST
In this protocol, rats must learn to navigate randomly in theopen field. A computer program delimited a circular rewardplace (70 cm-diameter, thick line circle); its center was chosenrandomly within a square circumscribed around the open field(Fig. 2Bb). The rat was rewarded with ICSS when it enteredthe reward place, which was then made inactive (changed tothin line circle). After a 5 s-interval, the reward place wasmoved to a different location and reactivated. The rat wastrained until it could achieve a criterion of 25 rewards acquisi-tion over three trials in a day.
Place learning task
Two 40 cm diameter reward places were located diametri-cally opposite to one another in the open field. The rat wasrewarded in both reward places, when it returned to one ofthem after a visit to the other one (Fig. 2Bc). Furthermore, therats were required to stay in the reward place for more than 1 sin order to acquire ICSS reward; otherwise, they could notreceive the reward even though they had entered the rewardplace. So, in this task the rat was required to learn and memo-rize the places where ICSS rewards were delivered. The taskwas terminated when the rat had acquired 50 rewards or when10 min had elapsed, whichever came first. The rats were testedfor 14 days with this protocol.
Administration of the Drugs
T-817MA was provided by the Research Laboratory ofToyama Chemical (Toyama, Japan). Donepezil is acetylcholine(ACh) esterase inhibitor, the most popular drug for AD treat-ment in the world market. T-817MA (high-dose: 8.4 mg/kgand low-dose: 0.84 mg/kg) and Donepezil (0.5 mg/kg) weredissolved in distilled water. Suspension of these drugs or waterwas administrated p.o. (forced-fed via a gastric tube) and daily(5 ml/kg/day) 1 h before the training/testing time from the firstday of Ab infusion until the end of the experiment (Fig. 2C).
Histology
After completion of the PLT, the rats were deeply anesthe-tized with pentobarbital sodium (50 mg/kg, i.p.). They weresacrificed by transcardial perfusion with 0.9% saline containingheparin followed by 10% buffered formalin. The whole brainswere then removed and immersed in the same fixative for1 day. After fixation, the brains were dehydrated with gradedethanol, passed through xylene, and embedded in paraffin.Serial 5-lm thick sections (coronal sections), which includedthe dorsal HF, were made. The sections were stained withhematoxylin and eosin (H&E), and then examined under alight microscope. Finally, the numbers of normal granule cellsand pyramidal cells were counted in the DG, CA1, and CA3areas.
Data Analysis
To evaluate the influence of Ab-infusion and/or T-817MAtreatment on motivational level and spontaneous activity, thedata of (i) ICSS current intensity, (ii) SMA for 10 min, and(iii) number of rewards acquired and the navigation distancetraveled in the last day of the RRPST training, were analyzedby one-way ANOVA.
Then, the behavioral data in the PLT were analyzed in termsof the following four parameters: (i) navigation pattern, (ii)mean number of rewards acquired in a trial, (iii) mean trialduration taken for a rat to complete the PLT, and (iv) meannavigation distance between the two reward places. A two-wayanalysis of variance (ANOVA) was performed to compare per-formance in the PLT among the all groups. Bonferroni posthoc test was employed for pair wise comparisons.
Number of normal granule cells was separately counted inthe whole granular cell layer from four regions in the left andright sides of the DG at the AP level 4.0 and 5.0 mm posteriorfrom the bregma. Histological evaluation was performed by dif-ferent experimenters in order to avoid the experimenter biasesin the selection of animals. Granule cells were defined as thosedisplaying cell profiles with nuclei pale and a granular back-ground, surrounded by a ring of visible cytoplasm and display-ing one or more clearly visible nucleolus (Mignini et al.,2004). The granule cell density was defined as the number ofnormal granule cells per 1-mm strip along the granular celllayer, and granule cell densities in both the sides were averagedin each rat. In the CA1 and CA3 subfields, the numbers of sur-viving cells in a 0.2-mm wide and 0.8-mm long strip of tissuefrom four regions in the left and right sides of the DG at theAP level 4.0 and 5.0 mm posterior from the bregma werecounted under a light microscope according to the previousstudy (Hori et al., 2002). These data from the different animalgroups were compared by one-way ANOVA. Bonferroni testwas used for post hoc comparisons.
These statistical tests were performed using StatView 5.0 J(Abacus Concepts, Berkeley, CA). Significance level employedfor all tests was P < 0.05.
446 NGUYEN ET AL.
Hippocampus DOI 10.1002/hipo
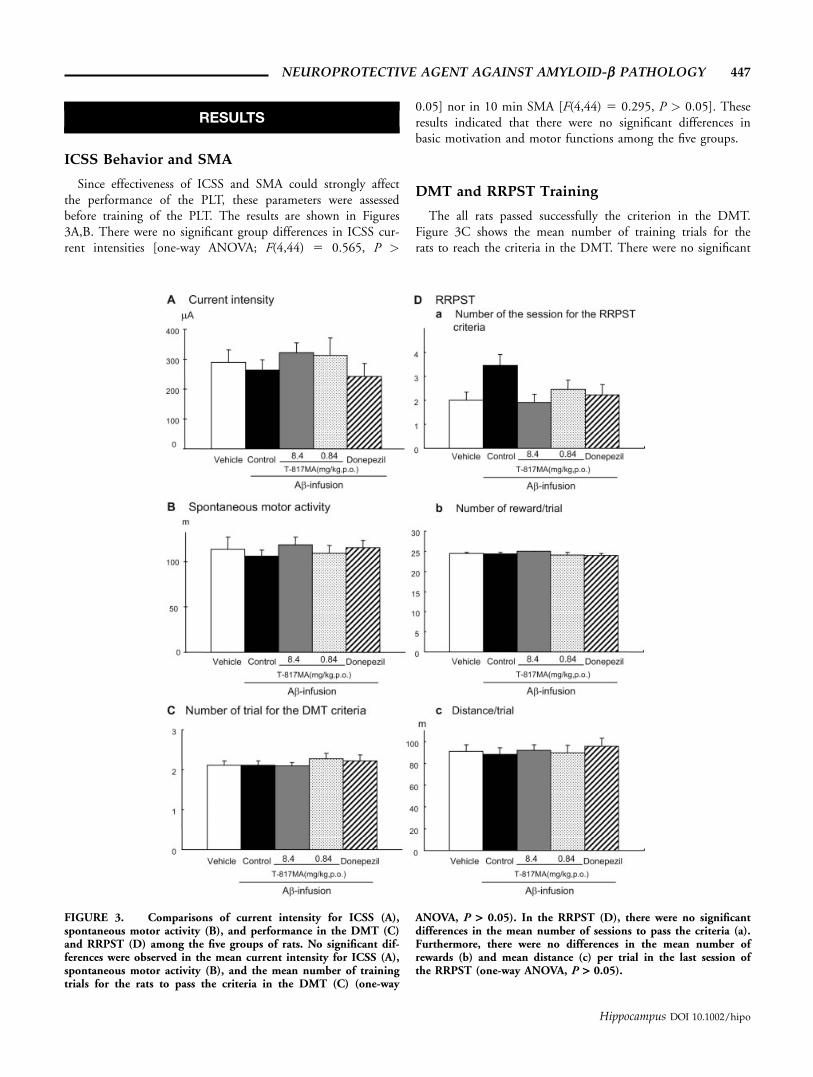
RESULTS
ICSS Behavior and SMA
Since effectiveness of ICSS and SMA could strongly affectthe performance of the PLT, these parameters were assessedbefore training of the PLT. The results are shown in Figures3A,B. There were no significant group differences in ICSS cur-rent intensities [one-way ANOVA; F(4,44) 5 0.565, P >
0.05] nor in 10 min SMA [F(4,44) 5 0.295, P > 0.05]. Theseresults indicated that there were no significant differences inbasic motivation and motor functions among the five groups.
DMT and RRPST Training
The all rats passed successfully the criterion in the DMT.Figure 3C shows the mean number of training trials for therats to reach the criteria in the DMT. There were no significant
FIGURE 3. Comparisons of current intensity for ICSS (A),spontaneous motor activity (B), and performance in the DMT (C)and RRPST (D) among the five groups of rats. No significant dif-ferences were observed in the mean current intensity for ICSS (A),spontaneous motor activity (B), and the mean number of trainingtrials for the rats to pass the criteria in the DMT (C) (one-way
ANOVA, P > 0.05). In the RRPST (D), there were no significantdifferences in the mean number of sessions to pass the criteria (a).Furthermore, there were no differences in the mean number ofrewards (b) and mean distance (c) per trial in the last session ofthe RRPST (one-way ANOVA, P > 0.05).
NEUROPROTECTIVE AGENT AGAINST AMYLOID-b PATHOLOGY 447
Hippocampus DOI 10.1002/hipo
differences in the mean number of training trials among thesefive groups [one-way ANOVA: F(4,44) 5 0.462, P > 0.05].
Figure 3Da shows the mean number of training sessions forthe rats to reach the criteria in the RRPST. There were no sig-nificant differences in the training sessions among the threegroups of the rats [one-way ANOVA: F(4,44) 5 2.320, P >0.05]. On the final day of RRPST training, as shown in Figure4a, each group rat, which passed the criterion, moved randomlyin the open field. There were no significant differences amongthe five groups in mean number of rewards per trial (Fig. 3Db)[one-way ANOVA; F(4,44) 5 0.833, P > 0.05], nor inthe mean navigation distance traveled per trial (Fig. 3Dc)[F(4,44) 5 0.174, P > 0.05].
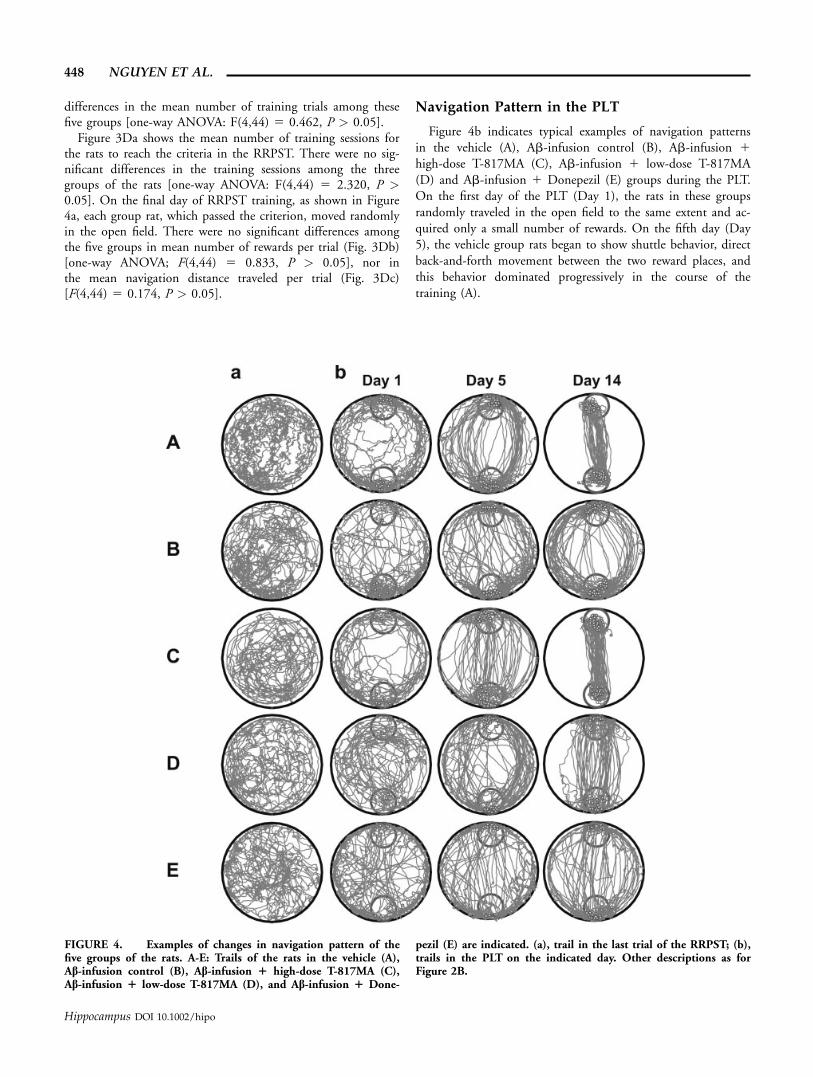
Navigation Pattern in the PLT
Figure 4b indicates typical examples of navigation patternsin the vehicle (A), Ab-infusion control (B), Ab-infusion 1high-dose T-817MA (C), Ab-infusion 1 low-dose T-817MA(D) and Ab-infusion 1 Donepezil (E) groups during the PLT.On the first day of the PLT (Day 1), the rats in these groupsrandomly traveled in the open field to the same extent and ac-quired only a small number of rewards. On the fifth day (Day5), the vehicle group rats began to show shuttle behavior, directback-and-forth movement between the two reward places, andthis behavior dominated progressively in the course of thetraining (A).
FIGURE 4. Examples of changes in navigation pattern of thefive groups of the rats. A-E: Trails of the rats in the vehicle (A),Ab-infusion control (B), Ab-infusion 1 high-dose T-817MA (C),Ab-infusion 1 low-dose T-817MA (D), and Ab-infusion 1 Done-
pezil (E) are indicated. (a), trail in the last trial of the RRPST; (b),trails in the PLT on the indicated day. Other descriptions as forFigure 2B.
448 NGUYEN ET AL.
Hippocampus DOI 10.1002/hipo
In contrast to the vehicle group, the rats in the Ab-infusioncontrol group did not show shuttle behavior during the 14-daytest period (B). Rather, it developed a navigation pattern ofmoving along the wall of the open field, although it showedweak tendency of shortcut later in the training period. Simi-larly, the rats in the Ab-infused 1 Donepezil treated groupshowed such a behavioral pattern (E).
The rats in the Ab-infusion 1 high-dose T-817MA groupshowed a similar pattern to that in the vehicle rats (C). Therats began to show shuttle behavior around the fifth day andestablished it during the rest of the training period. The rats inthe Ab-infused 1 low-dose T-817MA treated group were stillefficient but were not as fast as those rats (D).
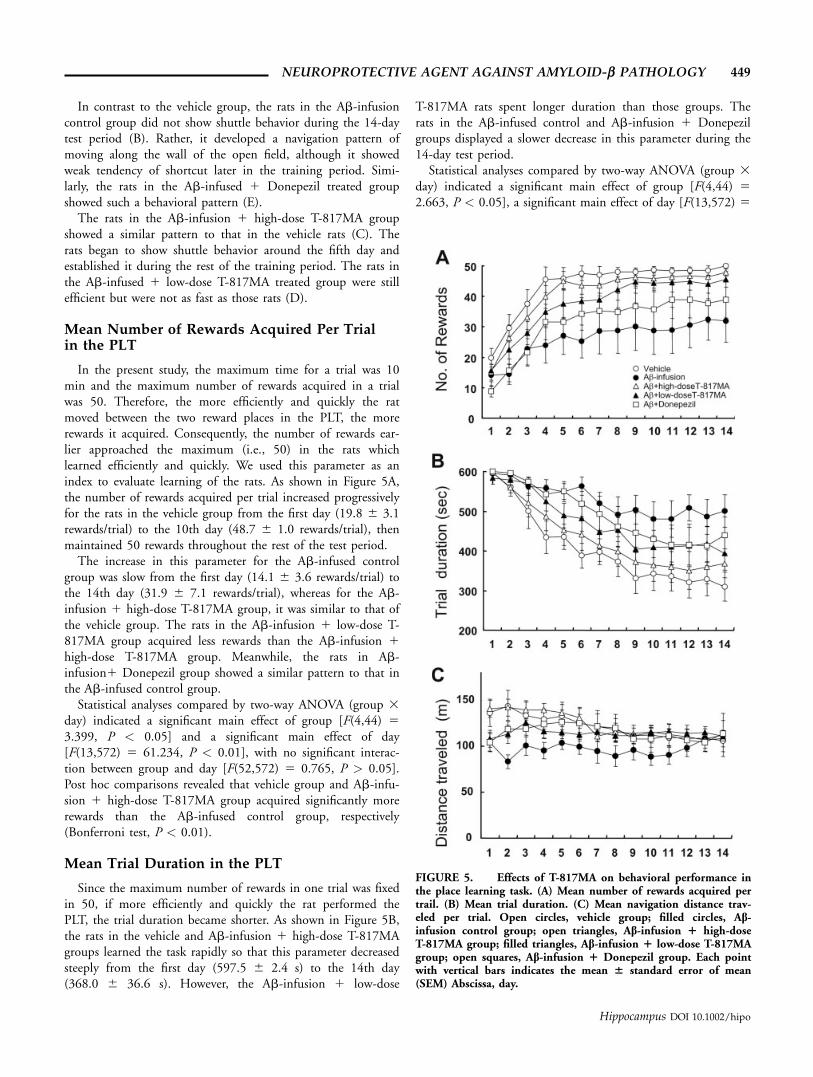
Mean Number of Rewards Acquired Per Trialin the PLT
In the present study, the maximum time for a trial was 10min and the maximum number of rewards acquired in a trialwas 50. Therefore, the more efficiently and quickly the ratmoved between the two reward places in the PLT, the morerewards it acquired. Consequently, the number of rewards ear-lier approached the maximum (i.e., 50) in the rats whichlearned efficiently and quickly. We used this parameter as anindex to evaluate learning of the rats. As shown in Figure 5A,the number of rewards acquired per trial increased progressivelyfor the rats in the vehicle group from the first day (19.8 6 3.1rewards/trial) to the 10th day (48.7 6 1.0 rewards/trial), thenmaintained 50 rewards throughout the rest of the test period.
The increase in this parameter for the Ab-infused controlgroup was slow from the first day (14.1 6 3.6 rewards/trial) tothe 14th day (31.9 6 7.1 rewards/trial), whereas for the Ab-infusion 1 high-dose T-817MA group, it was similar to that ofthe vehicle group. The rats in the Ab-infusion 1 low-dose T-817MA group acquired less rewards than the Ab-infusion 1high-dose T-817MA group. Meanwhile, the rats in Ab-infusion1 Donepezil group showed a similar pattern to that inthe Ab-infused control group.
Statistical analyses compared by two-way ANOVA (group 3day) indicated a significant main effect of group [F(4,44) 53.399, P < 0.05] and a significant main effect of day[F(13,572) 5 61.234, P < 0.01], with no significant interac-tion between group and day [F(52,572) 5 0.765, P > 0.05].Post hoc comparisons revealed that vehicle group and Ab-infu-sion 1 high-dose T-817MA group acquired significantly morerewards than the Ab-infused control group, respectively(Bonferroni test, P < 0.01).
Mean Trial Duration in the PLT
Since the maximum number of rewards in one trial was fixedin 50, if more efficiently and quickly the rat performed thePLT, the trial duration became shorter. As shown in Figure 5B,the rats in the vehicle and Ab-infusion 1 high-dose T-817MAgroups learned the task rapidly so that this parameter decreasedsteeply from the first day (597.5 6 2.4 s) to the 14th day(368.0 6 36.6 s). However, the Ab-infusion 1 low-dose
T-817MA rats spent longer duration than those groups. Therats in the Ab-infused control and Ab-infusion 1 Donepezilgroups displayed a slower decrease in this parameter during the14-day test period.
Statistical analyses compared by two-way ANOVA (group 3day) indicated a significant main effect of group [F(4,44) 52.663, P < 0.05], a significant main effect of day [F(13,572) 5
FIGURE 5. Effects of T-817MA on behavioral performance inthe place learning task. (A) Mean number of rewards acquired pertrail. (B) Mean trial duration. (C) Mean navigation distance trav-eled per trial. Open circles, vehicle group; filled circles, Ab-infusion control group; open triangles, Ab-infusion 1 high-doseT-817MA group; filled triangles, Ab-infusion 1 low-dose T-817MAgroup; open squares, Ab-infusion 1 Donepezil group. Each pointwith vertical bars indicates the mean 6 standard error of mean(SEM) Abscissa, day.
NEUROPROTECTIVE AGENT AGAINST AMYLOID-b PATHOLOGY 449
Hippocampus DOI 10.1002/hipo

49.199, P < 0.01]. There was no significant interaction betweengroup and day [F(52,572) 5 1.127, P > 0.05]. Post hoc com-parisons revealed that trial duration in the Ab-infused controlgroup was significantly longer than those in the vehicle groupand Ab-infusion 1 high-dose T-817MA group (Bonferroni test,P < 0.01 and P < 0.05, respectively).
Mean Navigation Distance Traveled Per Trialin the PLT
Figure 5C shows mean distance traveled during the 14 daysof navigation learning. The mean distance gradually decreasedfrom the 5th day (132.7 6 14.8 m) to the 14th day (106.0 611.6 m) in the vehicle group. In the Ab-infused control, themean navigation distance increased from the 5th day (103.9 67.7 m) to the 14th day (112.3 6 23.6 m). On the otherhand, this value gradually decreased throughout the testing pe-riod from (140.8 6 9.3 m) to (108.5 6 3.7 m) in the Ab-infusion 1 high-dose T-817MA group. In the Ab-infusion 1low-dose T-817MA and Ab-infusion 1 Donepezil groups, dis-tance also decreased from 5th day to the 14th day, but not soremarkably. The navigation distance tended to be shorter in thevehicle group and Ab-infusion 1 high-dose T-817MA groupthan in the Ab-infusion control group.
Statistical analyses by two-way ANOVA (group 3 day) indi-cated that there was no significant main effect of group [F(4,44)5 1.519, P > 0.05], while there were a significant main effectof day [F(13,572) 5 4.248, P < 0.01] and a significant interac-tion between group and day [F(52,572) 5 1.593, P < 0.01].
Histology
Histological changes in the HF are shown in Figure 6. Atthe lower magnification, the DG granule cell layer was clearlyobserved in the all groups (Fig. 6, left panels). At the highermagnification, many abnormal (densely stained) granule cellswere observed in the DG in the Ab-infusion control, Ab-infu-sion 1 low-dose T-817MA and Ab-infusion 1 Donepezilgroups (Fig. 6B, right; Fig. 6D, right and Fig. 6E, right).
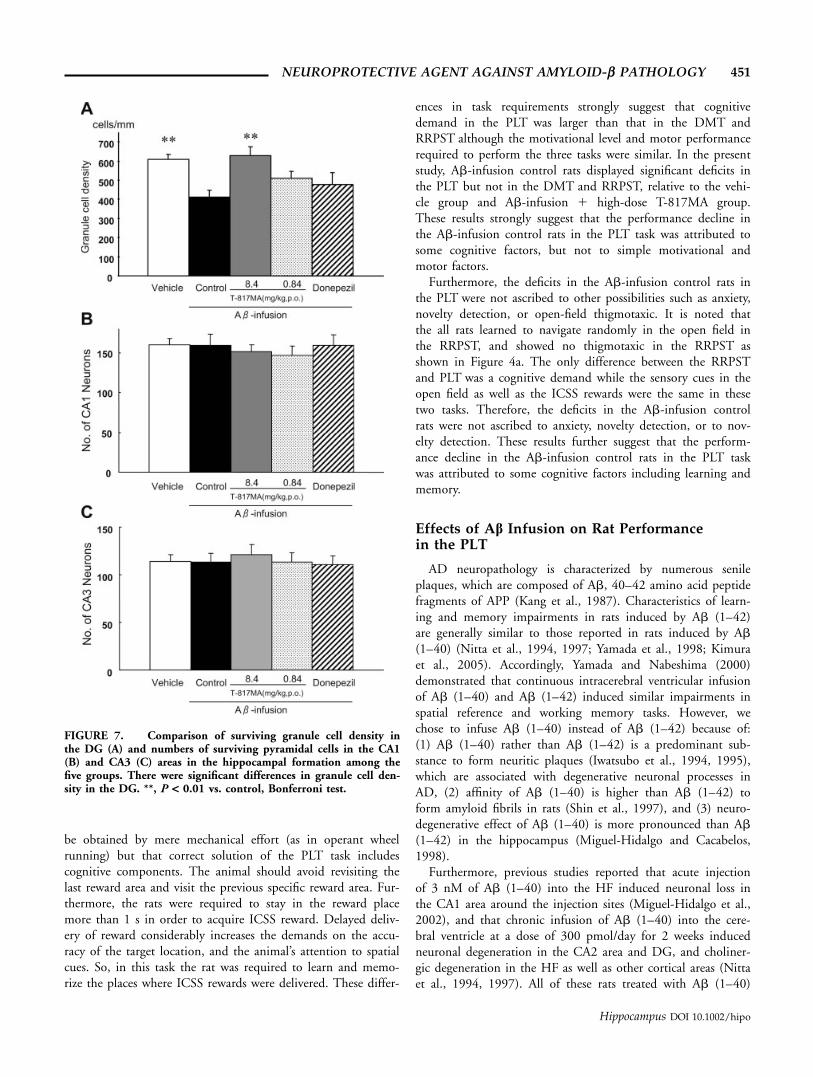
The density of normal granule cells in the DG is shown inFigure 7. The statistical analysis by one-way ANOVA indicatedthat there were significant differences in granule cell density amongthe five groups [F(4,43) 5 5.251, P < 0.01]. The post hoc testindicated that density of granule cell was significantly larger in thevehicle than Ab-infusion control groups (Bonferroni test, P <0.01), and larger in the Ab-infusion high-dose T-817MA than Ab-infusion control groups (Bonferroni test, P < 0.01). On the otherhand, treatment with Donepezil did not increase the density ofgranule cells; there was no significant difference in the granule celldensity between the Ab-infusion control and Ab-infusion 1Donepezil groups (Bonferroni test, P > 0.05).
However, after the cessation of the Ab-infusion, significantcell loss was not observed in the CA1 area [One-way ANOVA;F(4,43) 5 0.291, P > 0.05] (Fig. 7B), nor in the CA3 area[One-way ANOVA; F(4,43) 5 0.186, P > 0.05] (Fig. 7C).
DISCUSSION
Characteristics of the three Spatial Tasks
The rats were required to navigate in the same open fieldto acquire same ICSS rewards in the all three spatial tasks.However, in the DMT and RRPST tasks, the rats did not nec-essarily recognize spatial cues (i.e., a lamp and speaker) in con-trast to the PLT in which the rats had to recognize the rewardareas in reference to those spatial cues. That is, reward cannot
FIGURE 6. Photomicrographs of the DG in vehicle (A), Ab-infusion control (B), Ab-infusion 1 high-dose T-817MA (C), Ab-infusion 1 low-dose T-817MA (D) and Ab-infusion 1 Donepezil(E) groups. Left column, whole DG taken at low magnification;right column, a part of the DG indicated by the bracket in the leftphotomicrograph taken at high magnification. Horizontal linesbelow the photomicrographs under (E), scale bars.
450 NGUYEN ET AL.
Hippocampus DOI 10.1002/hipo
be obtained by mere mechanical effort (as in operant wheelrunning) but that correct solution of the PLT task includescognitive components. The animal should avoid revisiting thelast reward area and visit the previous specific reward area. Fur-thermore, the rats were required to stay in the reward placemore than 1 s in order to acquire ICSS reward. Delayed deliv-ery of reward considerably increases the demands on the accu-racy of the target location, and the animal’s attention to spatialcues. So, in this task the rat was required to learn and memo-rize the places where ICSS rewards were delivered. These differ-
ences in task requirements strongly suggest that cognitivedemand in the PLT was larger than that in the DMT andRRPST although the motivational level and motor performancerequired to perform the three tasks were similar. In the presentstudy, Ab-infusion control rats displayed significant deficits inthe PLT but not in the DMT and RRPST, relative to the vehi-cle group and Ab-infusion 1 high-dose T-817MA group.These results strongly suggest that the performance decline inthe Ab-infusion control rats in the PLT task was attributed tosome cognitive factors, but not to simple motivational andmotor factors.
Furthermore, the deficits in the Ab-infusion control rats inthe PLT were not ascribed to other possibilities such as anxiety,novelty detection, or open-field thigmotaxic. It is noted thatthe all rats learned to navigate randomly in the open field inthe RRPST, and showed no thigmotaxic in the RRPST asshown in Figure 4a. The only difference between the RRPSTand PLT was a cognitive demand while the sensory cues in theopen field as well as the ICSS rewards were the same in thesetwo tasks. Therefore, the deficits in the Ab-infusion controlrats were not ascribed to anxiety, novelty detection, or to nov-elty detection. These results further suggest that the perform-ance decline in the Ab-infusion control rats in the PLT taskwas attributed to some cognitive factors including learning andmemory.
Effects of Ab Infusion on Rat Performancein the PLT
AD neuropathology is characterized by numerous senileplaques, which are composed of Ab, 40–42 amino acid peptidefragments of APP (Kang et al., 1987). Characteristics of learn-ing and memory impairments in rats induced by Ab (1–42)are generally similar to those reported in rats induced by Ab(1–40) (Nitta et al., 1994, 1997; Yamada et al., 1998; Kimuraet al., 2005). Accordingly, Yamada and Nabeshima (2000)demonstrated that continuous intracerebral ventricular infusionof Ab (1–40) and Ab (1–42) induced similar impairments inspatial reference and working memory tasks. However, wechose to infuse Ab (1–40) instead of Ab (1–42) because of:(1) Ab (1–40) rather than Ab (1–42) is a predominant sub-stance to form neuritic plaques (Iwatsubo et al., 1994, 1995),which are associated with degenerative neuronal processes inAD, (2) affinity of Ab (1–40) is higher than Ab (1–42) toform amyloid fibrils in rats (Shin et al., 1997), and (3) neuro-degenerative effect of Ab (1–40) is more pronounced than Ab(1–42) in the hippocampus (Miguel-Hidalgo and Cacabelos,1998).
Furthermore, previous studies reported that acute injectionof 3 nM of Ab (1–40) into the HF induced neuronal loss inthe CA1 area around the injection sites (Miguel-Hidalgo et al.,2002), and that chronic infusion of Ab (1–40) into the cere-bral ventricle at a dose of 300 pmol/day for 2 weeks inducedneuronal degeneration in the CA2 area and DG, and choliner-gic degeneration in the HF as well as other cortical areas (Nittaet al., 1994, 1997). All of these rats treated with Ab (1–40)
FIGURE 7. Comparison of surviving granule cell density inthe DG (A) and numbers of surviving pyramidal cells in the CA1(B) and CA3 (C) areas in the hippocampal formation among thefive groups. There were significant differences in granule cell den-sity in the DG. **, P < 0.01 vs. control, Bonferroni test.
NEUROPROTECTIVE AGENT AGAINST AMYLOID-b PATHOLOGY 451
Hippocampus DOI 10.1002/hipo

displayed memory deficits (Nitta et al., 1994, 1997; Miguel-Hidalgo et al., 2002). In the present study, Ab (1–40) waschronically infused for 5 weeks, and granule cells in the DGwere degenerated. These results in the previous studies as wellas the present results were consistent in that Ab-infusioninduced neuronal degeneration in the HF as well as memorydisturbance.
In the PLT the vehicle rats altered the behavioral strategyfrom inefficient (random movement) to the efficient (directshuttle movement) patterns in the early stage of the PLT, whilemost Ab-infusion rats did not display such a behavioral alter-nation. These rats with Ab-infusion tended to show a differentbehavior such as moving along the wall of the open field. Con-sequently, time duration for the Ab-infusion rats to performthe PLT was longer than the vehicle rats. These navigation pat-terns were similar to those with bilateral HF lesions in theMorris water maze (Block et al., 1997; Jeltsch et al., 2001) andthose in the same task (i.e., PLT) (Hori et al., 2002). Theseresults suggest that the Ab-infused rats with neuronal loss inthe DG showed deficits in learning the locations of the rewardplaces. The other behavioral parameter in the PLT (no. ofrewards per trial) also demonstrated impaired performance ofthe Ab-infused rats.
Dentate Gyrus and Cognitive Deficits Due toAb Pathology
In the present study, we used Ab-infusion rat model for cog-nitive impairment in AD, and neuronal loss and degenerationwere confined in the DG. Previous studies reported that tran-sient cerebral ischemia produces neuronal cell loss confined tothe CA1 subfield in gerbils (Kirino, 1982), rats (Pulsinelliet al., 1982), monkeys (Nishijo et al., 1992, 1995; Tabuchiet al., 1992, 1995; Zola-Morgan et al., 1992; Yamashita et al.,1996) and humans (Volpe et al., 1986; Zola-Morgan et al.,1986; Petito et al., 1987). These animals with damages in theCA1 subfield after transient cerebral ischemia were used asanimal models of cerebrovascular dementia, and showedimpairment in spatial learning and memory (Morris et al.,1982; Jaspers et al., 1990; Nunn et al., 1994; Iwasaki et al.,1996; Ogura et al., 2000; Hori et al., 2002). On the otherhand, recent studies using transgenic mouse models of ADover-expressing APP reported that the DG was affected in anearly stage of Ab pathology (Redwine et al., 2003; Reilly et al.,2003; Chin et al., 2005; Jacobsen et al., 2006). Furthermore,direct infusion of Ab into the DG induced deficits in workingmemory in a radial maze task and impairment in long-termpotentiation (LTP) in the DG (Stephan et al., 2001). Theseresults along with the present results suggest that the DG,which receives inputs from the entorhinal cortex (EC), plays akey role in age- and AD-associated cognitive deficits. This sug-gestion is consistent with previous lesion studies, in whichlesions in the DG resulted in deficits in spatial tasks (Xavieret al., 1999; Gilbert et al., 2001; Jeltsch et al., 2001).McNaughton et al. (1989) also claimed that destruction of
more than 75% of the dentate granule cells severely disruptedperformance in the three different spatial tasks. These resultsstrongly suggest that HF dentate granule cells play an impor-tant role in spatial learning and memory, and one of key struc-tures in the AD.
It is noted that place learning deficits seem to be moreprominent in the present results with Ab infusion than thoseexpected from the previous study with intradentate colchicinesinjection (Jeltsch et al., 2001). A previous study reported thatneuronal dysfunction occurred before accumulation of Abplaques and neurodegeneration in transgenic mice over-express-ing APP (Jacobsen et al., 2006). In these mice, spine densitieswere decreased in the DG before memory deficits occurred,suggesting that memory deficits were ascribed to the reductionof spine densities in the DG (Jacobsen et al., 2006). This sug-gests that chronic Ab infusion also reduced spine densities ofsurviving granule cells in the DG in the present study, whichmight induce more prominent memory deficits in the PLTthan those expected from the previous lesion studies. Furtherstudies are required to prove this possibility.
Effects of Donepezil
Donepezil is an acetylcholinesterase inhibitor used for treatmentof AD. Most previous studies reported neuroprotective effects ofDonepezil using in vitro set up in N-methyl-D-aspartate toxicitymodel (Akasofu et al., 2006), oxygen–glucose deprivationmodel (Akasofu et al., 2003), and glutamate insult model(Takada et al., 2003). Some in vivo studies reported that Done-pezil ameliorated behavioral deficits (Csernansky et al., 2005;Dong et al., 2005; Yamada et al., 2005; Iwasaki et al., 2006).Accordingly, Dong et al. (2005) showed that daily administra-tion of Donepezil at a dose of 0.3 mg/kg/day improved thecontextual and cued memory in Tg(1) mice. Similarly, singleadministration of Donepezil at 0.1, 0.3, or 1.0 mg/kg alsoameliorated MK-801-induced deficits in spatial reversal learn-ing and in contextual and cued memory in a dose-dependentmanner (Csernansky et al., 2005). On the other hand, Iwasakiet al. (2006) and Yamada et al. (2005) have demonstrated thatadministration of Donepezil at a relatively higher dose (3 mg/kg and 2.5 mg/kg, p.o., respectively) ameliorated memoryimpairment in Ab-infused rats combined with cerebral ische-mia. However, Donepezil was administered only once or oncein a week in these studies (Yamada et al., 2005; Iwasaki et al.,2006). It is noted that the most effective dose of Donepezil hasbeen reported to be around 0.5 mg/kg/day, which is same asthat in the present study, when Donepezil was consecutivelyadministered to ameliorate some memory deficits in rodents(Ogura et al., 2000; Tohda et al., 2003; Meunier et al., 2006).In the present study using Ab-infusion model, daily adminis-tration of Donepezil at a dose of 0.5 mg/kg/day did not showsignificant ameliorative effects on behavioral deficits in the PLTnor neurodegeneration in the DG. The discrepancy betweenthe previous and present studies may be due to the differencesin the amount of Ab (1–40) and Ab (1–42) injected into the
452 NGUYEN ET AL.
Hippocampus DOI 10.1002/hipo

hippocampus, injection method or especially timing and diffi-culty of behavioral tasks. Furthermore, it is supposed thatDonepezil exerts its neuroprotective effects at least in partthrough nicotinic ACh receptors (nAChRs). These findingssuggest that pharmacological activation of nAChRs may not beeffective in Ab infusion model.
Effects of T-817MA on Performance of theAb-Infused Rats
In the present study, the Ab-infused rats with neuronal lossin the DG showed slow spatial learning and low navigationefficacy, which was similar to those in the previous studiesusing rats with CA1 lesions (Nakada et al., 2001; Tamuraet al., 2001; Hori et al., 2002). Furthermore, there were signifi-cant differences between the Ab-infusion control and Ab-infu-sion 1 high-dose T-817MA groups, while there were no signif-icant differences between the vehicle and Ab-infusion 1 high-dose T-817MA groups in any of the parameters. These findingsindicated that the Ab-infusion 1 high-dose T-817MA rats per-formed as efficiently as the vehicle rats in the PLT. Thus, thepresent study demonstrated that administration of high-dose T-817MA significantly ameliorated impairment of task perform-ance in the Ab-infused rats.
Neuroprotective Effects of T-817MA
The present results indicated that there were significant dif-ferences in granule cell density in the DG among the threegroups. Granule cell density in the Ab-infusion 1 high-doseT-817MA group was comparable to that in the vehicle group,indicating that administration of T-817MA immediately afterAb infusion alleviated neuronal damages in the DG. A previ-ous study in vitro reported that T-817MA exerted its neuropro-tective effects through modulation of endogenous antioxidativemechanisms rather than scavenging of reactive oxygen radicals,through which T-817MA protected cultured neurons from Abtoxicity (Hirata et al., 2005). Furthermore, T-817MA works asa neurotrophic factor such as insulin-like growth factor-1 (IGF-1) to promote neurite outgrowth (Hirata et al., 2005). It hasbeen reported that dendritic spine density decreased and LTPwas impaired in the DG in transgenic mice over-expressinghuman APP in an early stage before forming amyloid plaquesand neuronal degeneration (Jacobsen et al., 2006). This sug-gests that neurotrophic effects of T-817MA might be beneficialto protect neurons from such pathological changes in dendriticspines. In addition, T-817MA was easily transferred into thebrain across the blood–brain barrier after oral administration;brain level of T-817MA was �10 times higher than that inblood level (unpublished in house data). These facts may sug-gest a therapeutic potential of T-817MA for AD therapy.
In conclusion, treatment of T-817MA (8.4 mg/kg/day, p.o.),significantly improved the PLT performance of the Ab-infusedrats, which was attributed to hippocampal neuroprotection byT-817MA.
REFERENCES
Aberg MA, Aberg ND, Hedbacker H, Oscarsson J, Eriksson PS.2000. Peripheral infusion of IGF-I selectively induces neurogenesisin the adult rat hippocampus. J Neurosci 20:2896–2903.
Akasofu S, Kosasa T, Kimura M, Kubota A. 2003. Protective effect ofdonepezil in a primary culture of rat cortical neurons exposed tooxygen–glucose deprivation. Eur J Pharmacol 472:57–63.
Akasofu S, Kimura M, Kosasa T, Ogura H, Sawada K. 2006. Protec-tive effect of donepezil in primary-cultured rat cortical neuronsexposed to N-methyl-D-aspartate (NMDA) toxicity. Eur J Pharma-col 530:215–222.
Barnes CA, McNaughton BL. 1985. An age comparison of the ratesof acquisition and forgetting of spatial information in relation tolong-term enhancement of hippocampal synapses. Behav Neurosci99:1040–1048.
Bayer TA, Wirths O, Majtenyi K, Hartmann T, Multhaup G, Beyr-euther K, Czech C. 2001. Key factors in Alzheimer’s disease:b-Amyloid precursor protein processing, metabolism and intra-neuronal transport. Brain Pathol 11:1–11.
Behl C. 1997. Amyloid b-protein toxicity and oxidative stress inAlzheimer’s disease. Cell Tissue Res 290:471–480.
Block F, Pergande G, Schwarz M. 1997. Flupirtine reduces functionaldeficits and neuronal damage after global ischemia in rats. BrainRes 754:279–284.
Carr DB, Goate A, Phild D, Morris JC. 1997. Current concepts inthe pathogenesis of Alzheimer’s disease. Am J Med 103:3S–10S.
Chin J, Palop JJ, Puolivali J, Massaro C, Bien-Ly N, Gerstein H,Scearce-Levie K, Masliah E, Mucke L. 2005. Fyn kinase inducessynaptic and cognitive impairments in a transgenic mouse modelof Alzheimer’s disease. J Neurosci 25:9694–9703.
Csernansky JG, Martin M, Shah R, Bertchume A, Colvin J, Dong H.2005. Cholinesterase inhibitors ameliorate behavioral deficitsinduced by MK-801 in mice. Neuropsychopharmacology 30:2135–2143.
Dong H, Csernansky CA, Martin MV, Bertchume A, Vallera D, Cser-nansky JG. 2005. Acetylcholinesterase inhibitors ameliorate behav-ioral deficits in the Tg2576 mouse model of Alzheimer’s disease.Psychopharmacology 181:145–152.
Fukuda M, Kobayashi T, Bures J, Ono T. 1992. Rat exploratorybehavior controlled by intracranial self-stimulation improves thestudy of place cell activity. J Neurosci Methods 44:121–131.
Gilbert PE, Kesner RP, Lee I. 2001. Dissociating hippocampal sub-regions: A double dissociation between dentate gyrus and CA1.Hippocampus 11:626–636.
Hirata K, Yamaguchi H, Takamura Y, Takagi A, Fukushima T,Iwakami N, Saitoh A, Nakagawa M, Yamada T. 2005. A novelneurotrophic agent, T-817MA [1-{3-[2-(1-benzothiophen-5-yl)ethoxy] propyl}-3-azetidinol maleate], attenuates amyloid-b-induced neurotoxicity and promotes neurite outgrowth in rat cul-tured central nervous system neurons. J Pharmacol Exp Ther314:252–259.
Hori E, Uwano T, Tamura R, Miyake N, Nishijo H, Ono T. 2002.Effects of a novel arginine–vasopressin derivative, NC-1900, on thespatial memory impairment of rats with transient forebrain ische-mia. Brain Res Cogn Brain Res 13:1–15.
Hsiao KA. 2001. Learning and memory in transgenic mice modelingAlzheimer’s disease. Learn Mem 8:301–308.
Hyman BT, Van Hoesen GW, Damasio AR, Barnes CL. 1984. Alzhei-mer’s disease: Cell-specific pathology isolates the hippocampalformation. Science 225:1168–1170.
Iwasaki K, Kitamura Y, Ohgami Y, Mishima K, Fujiwara M. 1996.The disruption of spatial cognition and changes in brain aminoacid, monoamine and acetylcholine in rats with transient cerebralischemia. Brain Res 709:163–172.
NEUROPROTECTIVE AGENT AGAINST AMYLOID-b PATHOLOGY 453
Hippocampus DOI 10.1002/hipo

Iwasaki K, Egashira N, Hatip-Al-Khatib I, Akiyoshi Y, Arai T,Takagaki Y, Watanabe T, Mishima K, Fujiwara M. 2006. Cerebralischemia combined with b-amyloid impairs spatial memory in theeight-arm radial maze task in rats. Brain Res 1097:216–223.
Iwatsubo T, Odaka A, Suzuki N, Mizusawa H, Nukina N, Ihara Y.1994. Visualization of A b 42(43) and A b 40 in senile plaqueswith end-specific A b monoclonals: Evidence that an initially de-posited species is A b 42(43). Neuron 13:45–53.
Iwatsubo T, Mann DM, Odaka A, Suzuki N, Ihara Y. 1995. Amyloidb protein (A b) deposition: A b 42(43) precedes A b 40 in Downsyndrome. Ann Neurol 37:294–299.
Jacobsen JS, Wu CC, Redwine JM, Comery TA, Arias R, Bowlby M,Martone R, Morrison JH, Pangalos MN, Reinhart PH, Bloom FE.2006. Early-onset behavioral and synaptic deficits in a mousemodel of Alzheimer’s disease. Neuroscience 103:5161–5166.
Jaspers RM, Block F, Heim C, Sontag KH. 1990. Spatial learning isaffected by transient occlusion of common carotid arteries (2VO):Comparison of behavioural and histopathological changes after‘2VO’ and ‘four-vessel-occlusion’ in rats. Neurosci Lett 117:149–153.
Jeltsch H, Bertrand F, Lazarus C, Cassel JC. 2001. Cognitive perform-ances and locomotor activity following dentate granule cell damagein rats: Role of lesion extent and type of memory tested. NeurobiolLearn Mem 76:81–105.
Kang J, Lemaire HG, Unterbeck A, Salbaum JM, Masters CL,Grzeschik KH, Multhaup G, Beyreuther K, Muller H. 1987. Theprecursor of Alzheimer’s disease amyloid A4 protein resembles acell-surface receptor. Nature 325:733–736.
Kimura M, Komatsu H, Ogura H, Sawada K. 2005. Comparison ofdonepezil and memantine for protective effect against amyloid-b(1-42) toxicity in rat septal neurons. Neurosci Lett 391:17–21.
Kirino T. 1982. Delayed neuronal death in the gerbil hippocampusfollowing ischemia. Brain Res 239:57–69.
Kobayashi T, Nishijo H, Fukuda M, Bures J, Ono T. 1997. Taskdependent representations in rat hippocampal place neurons.J Physiol 78:597–613.
Kurimoto H, Nishijo H, Uwano T, Yamaguchi H, Zhong YM, Kawa-nishi K, Ono T. 2004. Effects of nonsaponin fraction of red gin-seng on learning deficits in aged rats. Physiol Behav 82:345–355.
Marchbanks RM. 1982. Biochemistry of Alzheimer’s dementia. J Neu-rochem 39:9–15.
McNaughton BL, Barnes CA, Meltzer J, Sutherland RJ. 1989. Hippo-campal granule cells are necessary for normal spatial learning butnot for spatially-selective pyramidal cell discharge. Exp Brain Res76:485–496.
Meunier J, Ieni J, Maurice T. 2006. The anti-amnesic and neuropro-tective effects of donepezil against amyloid b (25–35) peptide-induced toxicity in mice involve an interaction with the sigma(1)receptor. Br J Pharmacol 149:998–1012.
Mignini F, Vitaioli L, Sabbatini M, Tomassoni D, Amenta F. 2004.The cerebral cortex of spontaneously hypertensive rats: A quantita-tive microanatomical study. Clin Exp Hypertens 26:287–303.
Miguel-Hidalgo JJ, Cacabelos R. 1998. b-amyloid (1–40)-inducedneurodegeneration in the rat hippocampal neurons of the CA1 sub-field. Acta Neuropathol 95:455–465.
Miguel-Hidalgo JJ, Alvarez XA, Cacabelos R, Quack G. 2002. Neuro-protection by memantine against neurodegeneration induced by b-amyloid (1–40). Brain Res 958:210–221.
Morris RG, Garrud P, Rawlins JN, O’Keefe J. 1982. Place navigationimpaired in rats with hippocampal lesions. Nature 297:681–683.
Nabeshima T, Nitta A. 1994. Memory impairment and neuronal dys-function induced by b-amyloid protein in rats. Tohoku J ExpPharmacol 174:24–29.
Nakada Y, Tamura R, Kuriwaki JI, Kimura T, Uwano T, Nishijo H,Ono T. 2001. Ameliorative effects of a cognitive inhancer, T-588,on place learning impairment induced by transient forebrain ische-mia in rats. Physiol Behav 74:227–235.
Nishijo H, Kuze S, Ono T, Tabuchi E, Endo S, Kogure K. 1992. Cal-cium entry blocker ameliorates ischemic neuronal damage in mon-key hippocampus. Brain Res Bull 29:519–524.
Nishijo H, Tabuchi E, Ono T, Fukuda M, Kuze S, Endo S. 1995.Physiological results of monkey brain ischemia, and protection bya calcium blocker. Brain Res Bull 37:89–101.
Nitta A, Itoh A, Hasegawa T, Nabeshima T. 1994. b-Amyloid protein-induced Alzheimer’s disease animal model. Neurosci Lett 170:63–66.
Nitta A, Fukuta T, Hasegawa T, Nabeshima T. 1997. Continuousinfusion of b-Amyloid protein into the rat cerebral ventricle indu-ces learning impairment and neuronal and morphological degenera-tion. Jpn J Pharmacol 73:51–57.
Nunn JA, LePeillet E, Netto CA, Hodges H, Gray JA, Meldrum BS.1994. Global ischaemia: Hippocampal pathology and spatial defi-cits in the water maze. Brain Res 62:41–54.
Ogura H, Kosasa T, Kuriya Y, Yamanishi Y. 2000. Donepezil, a cen-trally acting acetylcholinesterase inhibitor, alleviates learning deficitsin hypocholinergic models in rats. Methods Find Exp Clin Phar-macol 22:89–95.
Ott BR, Saver JL. 1993. Unilateral amnesic stroke. Six new cases anda review of the literature. Stroke 24:1033–1042.
Paxinos G, Watson C. 1986. The Rat Brain in the Stereotaxic Coordi-nates. New York: Academic Press.
Petito CK, Feldmann E, Pulsinelli WA, Plum F. 1987. Delayed hippo-campal damage in humans following cardiorespiratory arrest.Neurology 37:1281–1286.
Pulsinelli WA, Brierley JB, Plum F. 1982. Temporal profile of neuro-nal damage in a model of transient forebrain ischemia. Ann Neurol11:491–498.
Racchi M, Govoni S. 2003. The pharmacology of amyloid precursorprotein processing. Exp Gerontol 38:145–157.
Redwine JM, Kosofsky B, Jacobs RE, Games D, Reilly JF, MorrisonJH, Young WG, Bloom FE. 2003. Dentate gyrus volume isreduced before onset of plaque formation in PDAPP mice: A mag-netic resonance microscopy and stereologic analysis. Neurosci100:1381–1386.
Reilly JF, Games D, Rydel RE, Freedman S, Schenk D, Young WG,Morrison JH, Bloom FE. 2003. Amyloid deposition in the hippo-campus and entorhinal cortex: Quantitative analysis of a transgenicmouse model. Neuroscience 100:4837–4842.
Selkoe DJ. 1998. The cell biology of b-amyloid precursor protein andpresenilin in Alzheimer’s disease. Trends Cell Biol 8:447–453.
Shin RW, Ogino K, Kondo A, Saido TC, Trojanowski JQ, KitamotoT, Tateishi J. 1997. Amyloid b-protein (Ab) 1–40 but not Ab 1–42 contributes to the experimental formation of Alzheimer diseaseamyloid fibrils in rat brain. J Neurosci 17:8187–8193.
Stephan A, Laroche S, Davis S. 2001. Generation of aggregated b-amyloid in the rat hippocampus impairs synaptic transmission andplasticity and causes memory deficits. J Neurosci 21:5703–5714.
Stoelzel CR, Stavnezer AJ, Denenberg VH, Ward M, Markus EJ.2002. The effects of aging and dorsal hippocampal lesions: Per-formance on spatial and nonspatial comparable versions of thewater maze. Neurobiol Learn Mem 78:217–233.
Suh YH, Cheler F. 2002. Amyloid precursor protein, presenilins, anda-synuclein: Molecular pathogenesis and pharmacological applica-tions in Alzheimer’s disease. Pharmacol Rev 54:469–525.
Tabuchi E, Endo S, Ono T, Nishijo H, Kuze S, Kogure K. 1992. Hip-pocampal neuronal damage after transient forebrain ischemia inmonkeys. Brain Res Bull 29:685–690.
Tabuchi E, Ono T, Nishijo H, Endo S, Kuzec S. 1995. Ischemic neu-ronal damage specific to monkey hippocampus: Histological inves-tigation. Brain Res Bull 37:73–87.
Takada Y, Yonezawa A, Kume T, Katsuki H, Kaneko S, Sugimoto H,Akaike A. 2003. Nicotinic acetylcholine receptor-mediated neuro-protection by donepezil against glutamate neurotoxicity in rat corti-cal neurons. J Pharmacol Exp Ther 306:772–777.
454 NGUYEN ET AL.
Hippocampus DOI 10.1002/hipo

Tamura R, Nakada Y, Nishijo H, Miyake N, Ono T. 2001. Ameliora-tive effects of tamolarizine on place learning impairment inducedby transient forebrain ischemia in rats. Brain Res 853:81–92.
Tohda C, Tamura T, Komatsu K. 2003. Repair of amyloid b (25–35)-induced memory impairment and synaptic loss by a Kampoformula, Zokumei-to. Brain Res 990:141–147.
Trejo JL, Carro E, Torres-Aleman I. 2001. Circulating insulin-likegrowth factor I mediates exercise-induced increases in the numberof new neurons in the adult hippocampus. J Neurosci 21:1628–1634.
Varadarajan S, Yatin S, Aksenova M, Butterfield DA. 2000. Review:Alzheimer’s amyloid b-peptide-associated free radical oxidativestress and neurotoxicity. J Struct Biol 130:184–208.
Venters HD. 2001. Tumor necrosis factora and insulin-like growthfactor-I in the brain: Is the whole greater than the sum of its parts?J Neuroimmunol 119:151–165.
Volpe BT, Holtzman JD, Hirst W. 1986. Further characterization ofpatients with amnesia after cardiac arrest: Preserved recognitionmemory. Neurology 36:408–411.
Ward MT, Oler JA, Markus EJ. 1999. Hippocampal dysfunction dur-ing aging. I. Deficits in memory consolidation. Neurobiol Aging20:363–372.
Xavier GF, Oliveira-Filho FJ, Santos AM. 1999. Dentate gyrus-selec-tive colchicine lesion and disruption of performance in spatialtasks: Difficulties in \place strategy" because of a lack of flexibilityin the use of environmental cues? Hippocampus 9:668–681.
Yamada K, Nabeshima T. 2000. Animal models of Alzheimer’s diseaseand evaluation of anti-dementia drugs. Pharmacol Ther 88:93–113.
Yamada K, Tanaka T, Senzakia K, Kameyama T, Nabeshima T. 1998.Propentofylline improves learning and memory deficits in rats
induced by b-amyloid protein-(1–40). Eur J Pharmacol 349:15–22.
Yamada K, Tanaka T, Han D, Senzaki K, Kameyama T, Nabeshima T.1999a. Protective effects of idebenone and a-tocopherol on b-amy-loid (1–42)-induced learning and memory deficits in rats: Implica-tion of oxidative stress in b-amyloid-induced neurotoxicity in vivo.Eur J Neurosci 11:83–90.
Yamada K, Tanaka T, Zou LB, Senzaki K, Yano K, Osada T, OlariuA, Ren X, Kameyama T, Nabeshima T. 1999b. Long-term depriva-tion of oestrogens by ovariectomy potentiates b-amyloid-inducedworking memory deficits in rats. Br J Pharmacol 128:419–427.
Yamashita T, Saido TC, Takita M, Miyazawa A, Yamano J, MiyakawaA, Nishijyo H, Yamashita J, Kawashima S, Ono T, Yoshioka T.1996. Transient brain ischaemia provokes Ca21, PIP2 and calpainresponses prior to delayed neuronal death in monkeys. Eur J Neu-rosci 8:1932–1944.
Yamada K, Takayanagi M, Kamei H, Nagai T, Dohniwa M, Kobaya-shi K, Yoshida S, Ohhara T, Takuma K, Nabeshima T. 2005.Effects of memantine and donepezil on amyloid b-induced mem-ory impairment in a delayed-matching to position task in rats.Behav Brain Res 162:191–199.
Zhong YM, Nishijo H, Uwano T, Tamura R, Kawanishi K, Ono T.2000. Red ginseng ameliorated place navigation deficits in youngrats with hippocampal lesions and aged rats. Physiol Behav69:511–525.
Zola-Morgan S, Squire LR, Amaral DG. 1986. Human amnesia andthe medial temporal region: Enduring memory impairment follow-ing a bilateral lesion limited to field CA1 of the hippocampus. JNeurosci 6:2950–2967.
Zola-Morgan S, Squire LR, Rempel NL, Glower RP, Amaral DG.1992. Enduring memory impairment in monkeys after ischemicdamage to the hippocampus. J Neurosci 12:2582–2596.
NEUROPROTECTIVE AGENT AGAINST AMYLOID-b PATHOLOGY 455
Hippocampus DOI 10.1002/hipo
Copyright © 2022 FDOKUMEN




















