
CD8+CD122+ regulatory T cells recognize activated T cells via conventional MHC class I- TCR...
11
International Immunology, Vol. 20, No. 7, pp. 937–947 doi:10.1093/intimm/dxn052 ª The Japanese Society for Immunology. 2008. All rights reserved. For permissions, please e-mail: [email protected] CD8 1 CD122 1 regulatory T cells recognize activated T cells via conventional MHC class I–abTCR interaction and become IL-10-producing active regulatory cells Muhaimin Rifa’i 1,2, *, Zhe Shi 1, *, Shu-Yun Zhang 3 , Young Ho Lee 1 , Hiroshi Shiku 4,5 , Ken-ichi Isobe 1 and Haruhiko Suzuki 1 1 Department of Immunology, Nagoya University Graduate School of Medicine, Nagoya 466-8550, Japan 2 Department of Biology, Faculty of Science, Brawijaya University, Malang 65145, East-Java, Indonesia 3 Department of Occupational and Environmental Health, Nagoya University Graduate School of Medicine, Nagoya 466-8550, Japan 4 Department of Cancer Vaccine and 5 Department of Immuno-Gene Therapy, Mie University Graduate School of Medicine, Tsu 514-8507, Japan Keywords: in vitro assay, mouse, suppression, Treg Abstract CD8 1 CD122 1 regulatory T cells (CD8 1 CD122 1 Treg) are naturally occurring Treg that effectively suppress the proliferation and IFN-g production of both CD8 1 and CD4 1 target cells. This study investigated the molecular mechanisms of the recognition of target cells by CD8 1 CD122 1 Treg using an in vitro culture system that reconstitutes the regulatory action of these cells. Naive CD8 1 CD122 1 Treg co-cultured with pre-activated T cells became active Treg that produced IL-10 and suppressed IFN-g production from the target T cells. CD8 1 CD122 1 Treg effectively suppressed the IFN-g production of the target cells of syngeneic mouse strains but not of allogeneic mouse strains with incompatible MHC. By using MHC-congeneic mouse strains, MHC-restricted suppression by CD8 1 CD122 1 Treg was further confirmed. The blockade of cell surface molecules either on the Treg or on the target cells by specific blocking antibodies indicated that H-2K, H-2D, abTCR and CD8 were involved in the regulatory action but I-A and Qa-1 were not. These results indicate that CD8 1 CD122 1 Treg recognize already-activated T cells via the interaction of conventional MHC class I–abTCR and become active regulatory cells that produce IL-10 and suppress the target cells. Introduction Regulation of the immune system is of paramount importance for the suppression of autoimmune reactions and for the regula- tion of the immune response against foreign pathogens. T cells are the key players that control the overall immune response and there are two major sub-populations of T cells, distin- guished by their cell surface markers: CD4 + and CD8 + . T cells are also divided by their functions into ‘helper’, ‘cytotoxic’ and ‘regulatory’ cells, which roughly correspond to cell types of CD4 + , CD8 + and both, respectively. Immunoregulatory T cells were originally described as ‘suppressor’ T cells, the majority of which were CD8 + (1–3). After the discovery of CD4 + CD25 + (Foxp3 + ) immunoregulatory T cells, however, such CD4 + T cells became the main object of regulatory T cells (Treg) and were called Treg (4–6). CD4 + CD25 + Treg are naturally oc- curring Treg. Before inducing an immune reaction in vivo or in vitro, such Treg can be distinguished from other T cells by specific markers such as CD25 and Foxp3. The finding of nat- urally occurring Treg enabled prospective studies of T cell reg- ulation. On the other hand, many induced-type Treg have also been identified, as represented by Tr1 and T h 3 cells (7–9). So far, studies of Treg in CD4 + cells have greatly advanced, com- pared with those of Treg in CD8 + cells. Although many CD8 + Treg with different markers have also been reported (10–16), most are categorized into induced-type Treg. CD8 + CD122 + Treg (CD8 + CD122 + Treg) are newly identified Treg (17). Among various CD8 + Treg, CD8 + CD122 + Treg *These authors contributed equally to this work. Correspondence to: H. Suzuki; E-mail: [email protected] Received 4 March 2008, accepted 25 April 2008 Transmitting editor: K. Okumura Advance Access publication 20 May 2008 by guest on July 11, 2016 http://intimm.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of CD8+CD122+ regulatory T cells recognize activated T cells via conventional MHC class I- TCR...

International Immunology, Vol. 20, No. 7, pp. 937–947doi:10.1093/intimm/dxn052
ª The Japanese Society for Immunology. 2008. All rights reserved.For permissions, please e-mail: [email protected]
CD81CD1221 regulatory T cells recognize activatedT cells via conventional MHC class I–abTCRinteraction and become IL-10-producing activeregulatory cells
Muhaimin Rifa’i1,2,*, Zhe Shi1,*, Shu-Yun Zhang3, Young Ho Lee1, Hiroshi Shiku4,5, Ken-ichi Isobe1
and Haruhiko Suzuki1
1Department of Immunology, Nagoya University Graduate School of Medicine, Nagoya 466-8550, Japan2Department of Biology, Faculty of Science, Brawijaya University, Malang 65145, East-Java, Indonesia3Department of Occupational and Environmental Health, Nagoya University Graduate School of Medicine, Nagoya 466-8550,Japan4Department of Cancer Vaccine and 5Department of Immuno-Gene Therapy, Mie University Graduate School of Medicine,Tsu 514-8507, Japan
Keywords: in vitro assay, mouse, suppression, Treg
Abstract
CD81CD1221 regulatory T cells (CD81CD1221 Treg) are naturally occurring Treg that effectivelysuppress the proliferation and IFN-g production of both CD81 and CD41 target cells. This studyinvestigated the molecular mechanisms of the recognition of target cells by CD81CD1221 Treg usingan in vitro culture system that reconstitutes the regulatory action of these cells. Naive CD81CD1221
Treg co-cultured with pre-activated T cells became active Treg that produced IL-10 and suppressedIFN-g production from the target T cells. CD81CD1221 Treg effectively suppressed the IFN-gproduction of the target cells of syngeneic mouse strains but not of allogeneic mouse strains withincompatible MHC. By using MHC-congeneic mouse strains, MHC-restricted suppression byCD81CD1221 Treg was further confirmed. The blockade of cell surface molecules either on the Treg oron the target cells by specific blocking antibodies indicated that H-2K, H-2D, abTCR and CD8 wereinvolved in the regulatory action but I-A and Qa-1 were not. These results indicate that CD81CD1221
Treg recognize already-activated T cells via the interaction of conventional MHC class I–abTCR andbecome active regulatory cells that produce IL-10 and suppress the target cells.
Introduction
Regulation of the immune system is of paramount importancefor the suppression of autoimmune reactions and for the regula-tion of the immune response against foreign pathogens. T cellsare the key players that control the overall immune responseand there are two major sub-populations of T cells, distin-guished by their cell surface markers: CD4+ and CD8+. T cellsare also divided by their functions into ‘helper’, ‘cytotoxic’ and‘regulatory’ cells, which roughly correspond to cell types ofCD4+, CD8+ and both, respectively. Immunoregulatory T cellswere originally described as ‘suppressor’ T cells, the majorityof which were CD8+ (1–3). After the discovery of CD4+CD25+
(Foxp3+) immunoregulatory T cells, however, such CD4+
T cells became the main object of regulatory T cells (Treg)
and were called Treg (4–6). CD4+CD25+ Treg are naturally oc-curring Treg. Before inducing an immune reaction in vivo orin vitro, such Treg can be distinguished from other T cells byspecific markers such as CD25 and Foxp3. The finding of nat-urally occurring Treg enabled prospective studies of T cell reg-ulation. On the other hand, many induced-type Treg have alsobeen identified, as represented by Tr1 and Th3 cells (7–9). Sofar, studies of Treg in CD4+ cells have greatly advanced, com-pared with those of Treg in CD8+ cells. Although many CD8+
Treg with different markers have also been reported (10–16),most are categorized into induced-type Treg.
CD8+CD122+ Treg (CD8+CD122+ Treg) are newly identifiedTreg (17). Among various CD8+ Treg, CD8+CD122+ Treg
*These authors contributed equally to this work.
Correspondence to: H. Suzuki; E-mail: [email protected] Received 4 March 2008, accepted 25 April 2008
Transmitting editor: K. Okumura Advance Access publication 20 May 2008
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from

should be distinguished from others based on the followingtwo points. First, CD8+CD122+ Treg are naturally occurringTreg. They are identified in immunologically naive youngmice. Second, their essential role is proved by in vivo experi-ments. CD8+CD122+ Treg are indispensable in the mainte-nance of immune homeostasis because activated T cellsexpand without regulation and cause lethal disorders in micein the absence of CD8+CD122+ Treg. CD122-knockout (KO)mice that lack CD8+CD122+ Treg and T cell-deficient miceadoptively supplemented with T cells depleted ofCD8+CD122+ cells show similar lethal phenotypes with un-controlled activation of T cells (17, 18). In vitro experimentsproved that CD8+CD122+ Treg effectively suppress cell pro-liferation and IFN-c production from CD8+ and CD4+ cells(17). CD8+CD122+ Treg directly suppress target cells withoutthe intervention of special antigen-presenting cells (APCs),a feature that distinguishes these cells from CD4+CD25+
Treg (17, 19–21).Although the role of CD8+CD122+ Treg in the incidence
and/or progression of certain clinical disorders, such as au-toimmune diseases and transplantation-associated immunereactions, has not been proven yet, the strong immune-suppressing power of the CD8+CD122+ Treg promises futureapplications of these Treg, possibly in co-ordination withCD4+CD25+ Treg, to immunity-based intractable diseases.Understanding the molecular actions of regulatory cells isa necessary step for the application of these cells to dis-eases. The cellular and molecular mechanisms of thesuppressing action of CD8+CD122+ Treg have been investi-gated, and the most important effect-transmitting factor hasbeen identified as IL-10 (22). Currently, other regulatorymechanisms of the target cells of CD8+CD122+ Treg andthe recognition molecules of CD8+CD122+ Treg are largelyunknown. In this study, we investigated the molecular mech-anisms of CD8+CD122+ Treg in the recognition of targetcells. CD8+CD122+ T cells recognize previously activatedT cells by interacting with cell surface molecules, includingconventional MHC class I–abTCR. CD8+CD122+ T cellsthat have recognized the target cells become active regula-tory cells that produce effect-transmitting factors such asIL-10.
Methods
Mice
C57BL/6, BALB/c, C3H/He, A/J, C57BL/10, B10.A/Sg,B10.BR/Sg and B10.D2/n mice were purchased from SLCJapan (Hamamatsu, Japan). C57BL/6CD45.1 mice and OT-1TCR transgenic mice with a C57BL/6 recombination-activating gene (RAG-1�/�) genetic background were origi-nally purchased from Taconic (Hudson, NY, USA) and main-tained in our animal facility. b2-microglobulin-KO mice andIL-10-KO mice with C57BL/6 genetic background were origi-nally purchased from the Jackson Laboratory (Bar Harbor,ME, USA) and maintained in our animal facility. All miceused in this study were maintained under specific patho-gen-free conditions. The animal care was conducted in ac-cordance with the guidelines of Nagoya University (Nagoya,Japan) and of Mie University (Tsu, Japan).
Antibodies
FITC- or PE-conjugated anti-mouse CD8a (clone: 53-6.7),FITC-conjugated anti-mouse CD25 (clone: PC61.5), PE-conjugated anti-mouse CD4 (clone: GK1.5), biotin-conjugatedanti-mouse CD122 (clone: 5H4), biotin-conjugated anti-mouseIFN-c (clone: XMG1.2), biotin-conjugated anti-mouse IL-10(clone: JES5-2A5) and FITC- or biotin-conjugated anti-mouseCD45.1 (clone: A20) antibodies and the streptavidin–PE–Cy5conjugate were purchased from eBioscience (San Diego, CA,USA). Blocking anti-H-2Kb (clone: AF6-88.5), anti-H-2Db
(clone: 28-14-8), anti-I-Ab (clone: AF6-120.1), anti-mouseTCRb (clone: H57-597) and anti-mouse CD8a (clone: Lyt2.2)antibodies were gifts from E. Nakayama (Okayama Univer-sity, Okayama, Japan). The neutralizing anti-mouse IL-10(clone: JES5-2A5) and anti-mouse Qa-1b antibodies (clone:6A8.6F10.1A6) were purchased from BD Biosciences(San Jose, CA, USA). The concentrations of anti-H-2Kb, anti-H-2Db, anti-TCRb and anti-CD8a antibodies for the use ofblocking regulatory activity were determined by test experi-ments in which the addition of antibodies blocked allogeneicCTL activity.
Cell preparation
Spleen cells obtained from 6- to 8-week old mice were incu-bated with anti-mouse CD8-coated magnetic beads (MACS,Miltenyi Biotec, Bergisch Gladbach, Germany), and theCD8+ cell-enriched fraction was recovered from the mag-netic column according to the manufacturer’s protocol.Then, cells were stained with FITC-conjugated anti-CD8 andbiotin-conjugated anti-CD122 followed by staining withstreptavidin–PE–Cy5. Stained cells were subjected to theFACSVantage cell sorter (BD Biosciences), and CD8+CD122+
and CD8+CD122� cells were obtained. APCs were preparedby treating C57BL/6 spleen cells with anti-Thy1.2 antibodyfollowed by a guinea pig complement (MP Biomedicals,Solon, OH, USA) and extensive washing with PBS. To pre-pare activated T cells, cells were cultured in anti-CD3-coatedplates (BD BioCoatTM, BD Biosciences) for 24 h. For the cul-ture of OT-1 TCR transgenic T cells, 2 3 105 OT-1 T cellswere cultured with 2 3 105 APCs in a medium containing100 lg ml�1 of antigenic ovalbumin (OVA) peptide 257–264.LPS-stimulated B cells were prepared by culturing purifiedB220+ splenic cells in 25 lg ml�1 LPS for 24 h. In every caseof co-culture of Treg and pre-activated T or B cells, pre-activated cells were extensively washed with PBS beforetransferring fresh culture wells containing the Treg.
In vitro suppression assay using pre-activated T cells
Pre-activated T cells were prepared as described in the formersection. Freshly isolated CD8+CD122+ Treg (1 3 105) were co-cultured with 4 3 105 pre-activated T cells for 60 h in a mediumcontaining recombinant human IL-2 (10 ng ml�1) in a round-bottom 96-well plate. Then the cells were subjected to theanalysis of intracellular cytokine staining or the culture su-pernatant was assessed using ELISA assay kit (QuantikineImmunoassay, R&D Systems, Minneapolis, MN, USA). Todistinguish CD8+CD122+ Treg from pre-activated T cells, ei-ther one of the cell populations was obtained from C57BL/6CD45.1 mice or labeled with carboxy fluorescein diacetatesuccinimide ester before starting the co-culture.
938 Molecular mechanism of suppression by CD8+CD122+ Treg
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from

In vitro suppression assay using diffusion chambers that passmedium-soluble factors
The typical culture using diffusion chambers was performedas follows: A mixture of 1 3 105 Treg and 1 3 105 pre-activated T cells was cultured inside the diffusion chamber(Chemotaxicell 96 3 m, Kurabo, Osaka, Japan) and 2 3 105
APCs plus 2 3 105 OT-1 T cells or 4 3 105 CD8+CD122�
cells stimulated by plate-bound anti-CD3 antibody (BDBioCoatTM, BD Biosciences) were cultured outside the diffu-sion chamber. Cells either inside or outside of the diffusionchamber were harvested and analyzed after 60 h of culture.
In vitro suppression assay using OT-1 TCR-transgenic T cells
CD8+CD122� OT-1 T cells (2 3 105) and 2 3 105 APCs werecultured in a medium containing 100 lg ml�1 OVA peptide.Twenty-four hours later, 1 3 105 CD8+CD122+ cells isolatedfrom C57BL/6CD45.1 mice were added to the cultures. After60 h of co-culture, cells were subjected to intracellular IL-10staining or IFN-c staining.
Detection of intracellular cytokine production
In order to detect intracellular cytokines, we did not re-stimulatecells with PMA and ionomycin, which are commonly usedfor enhancing the production of cytokines. When we usesuch strong stimulating reagents, it is impossible to detectthe difference between positive suppression and non-suppression, as demonstrated in Supplementary Figure S1(available at International Immunology Online). For the samereason, we also did not use reagents such as GolgiPlug(BD Biosciences) that inhibit secretion of cytokines and allowthem to accumulate intracellularly. Without using re-stimulatingreagents and secretion inhibitors, we could detect intra-cellular cytokine production in the case of FACS-purifiedCD8+CD122+ cells or CD8+CD122� cells. Harvested cellswere first stained with fluorescence-conjugated antibodiesfor cell surface molecules and then subjected to the detec-tion of intracellular cytokines according to the protocols us-ing Cytofix/cytoperm (BD Biosciences) and biotin-conjugated anti-mouse IL-10 or anti-mouse IFN-c antibodyfollowed by the streptavidin–PE–Cy5 conjugate. Cytokine ex-pression was analyzed with a FACSCalibur Flow Cytometer(BD Biosciences).
Statistical analysis and ethical considerations
Data were expressed as mean 6 standard deviation. Differ-ences between groups were examined for statistical signifi-cance using the Student’s t test. A P value <0.01 denotedthe presence of a statistically significant difference. All ani-mal experiments conducted in this study were approved bythe Ethics Review Committee for Animal Experimentation ofNagoya University Graduate School of Medicine.
Results
CD8+CD122+ Treg co-cultured with previously activatedT cells produce IL-10
We have already proved that CD8+CD122+ Treg producemedium-soluble materials, including IL-10, as suppressiveeffect-transmitting factors that work at the terminal stage of
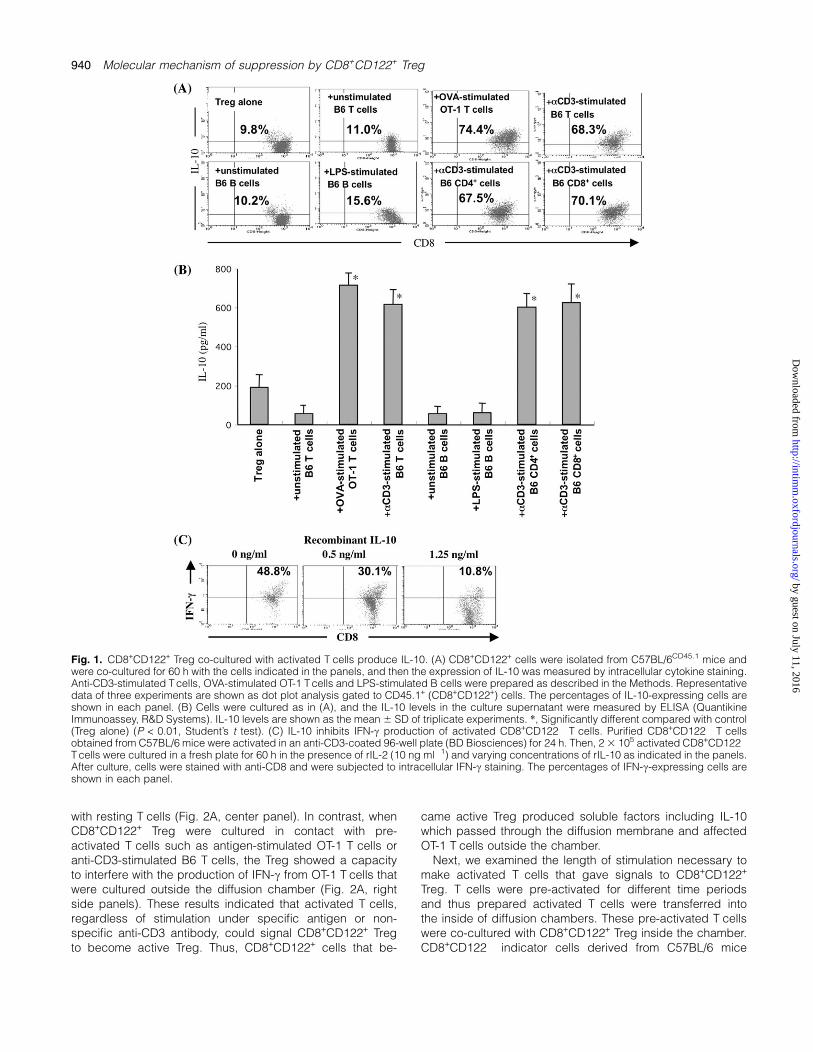
suppression with CD8+CD122+ Treg (22). In this study, weexamined the mechanism by which naive CD8+CD122+ Tregbecome active Treg that produce IL-10. First, we looked forthe type of cells that give signals to CD8+CD122+ Treg to be-come active Treg. We measured the production of IL-10 inCD8+CD122+ Treg that had been co-cultured with varioustypes of cells either in resting or in activated states.CD8+CD122+ cells were isolated from C57BL/6CD45.1 miceand were co-cultured for 60 h. Here, CD8+CD122+ cells werenot directly stimulated by anti-CD3 antibody but were simplyco-cultured with variously pre-treated cells. Production ofIL-10 was first measured by intracellular cytokine stainingusing a specific anti-mouse IL-10 antibody. This methodenables identification of the responsible IL-10-producingcells in the mixture of Treg and other cells by staining of mul-tiple cell surface markers, including CD45.1 which distin-guishes CD8+CD122+ Treg from co-cultured cells. Asa result, CD8+CD122+ Treg co-cultured with pre-activatedT cells produced more IL-10 than those co-cultured withresting T cells (Fig. 1A). In this case, an increase of IL-10-producing cells was the same whether the pre-activatedcells were created by antigen-specific stimulation with OVApeptide antigen or by non-specific stimulation with anti-CD3antibody. We next measured the IL-10 level in culture super-natant by ELISA assay and found that the level of IL-10 inculture supernatant was almost parallel with the number ofIL-10-producing cells in CD8+CD122+ Treg, suggestingthat IL-10 production in the culture supernatant was largelydependent on CD8+CD122+ Treg but not on other cells co-cultured (Fig. 1B).
To confirm that the level of IL-10 in the culture supernatantcontrols the production of IFN-c in the target cells, wecultured the activated CD8+CD122� cells in varying concen-trations of recombinant IL-10 (Fig. 1C). Suppression of IFN-cproduction was dependent on the concentration of rIL-10.This experiment also confirmed the reliability of our experi-ment using rIL-2 (10 ng ml�1), which allowed us to maintainthe viability of purified CD8+ T cells in culture.
CD8+CD122+ Treg co-cultured with pre-activated T cellssuppress IFN-c production from target cells
Next, we examined whether CD8+CD122+ Treg that becameactive Treg by interacting with pre-activated T cells reallyacquired the regulatory activity to suppress the target cells.We designed an experiment involving the culture of test cellsand indicator cells separated by a diffusion chamberthat passes medium-soluble effect-transmitting factors. Anti-CD3-stimulated T cells or OVA-stimulated T cells were pre-pared and were co-cultured with CD8+CD122+ Treg insidethe chamber and OT-1 T cells were cultured outside thechamber. Sixty hours later, OT-1 T cells were harvested andsubjected to staining of cell surface CD8 and intracellularIFN-c. When indicator OT-1 T cells outside the chamber werecultured under the stimulation of the OVA antigenic peptidepresent on the APCs, these cells produced IFN-c (Fig. 2A,left end panel). Test cells were cultured inside the chamber,and their suppressive activity was measured by IFN-c pro-duction from the indicator OT-1 T cells outside the chamber.CD8+CD122+ Treg were not able to suppress IFN-c produc-tion from OT-1 indicator cells when the Treg were co-cultured
Molecular mechanism of suppression by CD8+CD122+ Treg 939
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from
with resting T cells (Fig. 2A, center panel). In contrast, whenCD8+CD122+ Treg were cultured in contact with pre-activated T cells such as antigen-stimulated OT-1 T cells oranti-CD3-stimulated B6 T cells, the Treg showed a capacityto interfere with the production of IFN-c from OT-1 T cells thatwere cultured outside the diffusion chamber (Fig. 2A, rightside panels). These results indicated that activated T cells,regardless of stimulation under specific antigen or non-specific anti-CD3 antibody, could signal CD8+CD122+ Tregto become active Treg. Thus, CD8+CD122+ cells that be-
came active Treg produced soluble factors including IL-10which passed through the diffusion membrane and affectedOT-1 T cells outside the chamber.
Next, we examined the length of stimulation necessary tomake activated T cells that gave signals to CD8+CD122+
Treg. T cells were pre-activated for different time periodsand thus prepared activated T cells were transferred intothe inside of diffusion chambers. These pre-activated T cellswere co-cultured with CD8+CD122+ Treg inside the chamber.CD8+CD122� indicator cells derived from C57BL/6 mice
Fig. 1. CD8+CD122+ Treg co-cultured with activated T cells produce IL-10. (A) CD8+CD122+ cells were isolated from C57BL/6CD45.1 mice andwere co-cultured for 60 h with the cells indicated in the panels, and then the expression of IL-10 was measured by intracellular cytokine staining.Anti-CD3-stimulated T cells, OVA-stimulated OT-1 T cells and LPS-stimulated B cells were prepared as described in the Methods. Representativedata of three experiments are shown as dot plot analysis gated to CD45.1+ (CD8+CD122+) cells. The percentages of IL-10-expressing cells areshown in each panel. (B) Cells were cultured as in (A), and the IL-10 levels in the culture supernatant were measured by ELISA (QuantikineImmunoassey, R&D Systems). IL-10 levels are shown as the mean 6 SD of triplicate experiments. *, Significantly different compared with control(Treg alone) (P < 0.01, Student’s t test). (C) IL-10 inhibits IFN-c production of activated CD8+CD122� T cells. Purified CD8+CD122� T cellsobtained from C57BL/6 mice were activated in an anti-CD3-coated 96-well plate (BD Biosciences) for 24 h. Then, 2 3 105 activated CD8+CD122�
T cells were cultured in a fresh plate for 60 h in the presence of rIL-2 (10 ng ml�1) and varying concentrations of rIL-10 as indicated in the panels.After culture, cells were stained with anti-CD8 and were subjected to intracellular IFN-c staining. The percentages of IFN-c-expressing cells areshown in each panel.
940 Molecular mechanism of suppression by CD8+CD122+ Treg
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from
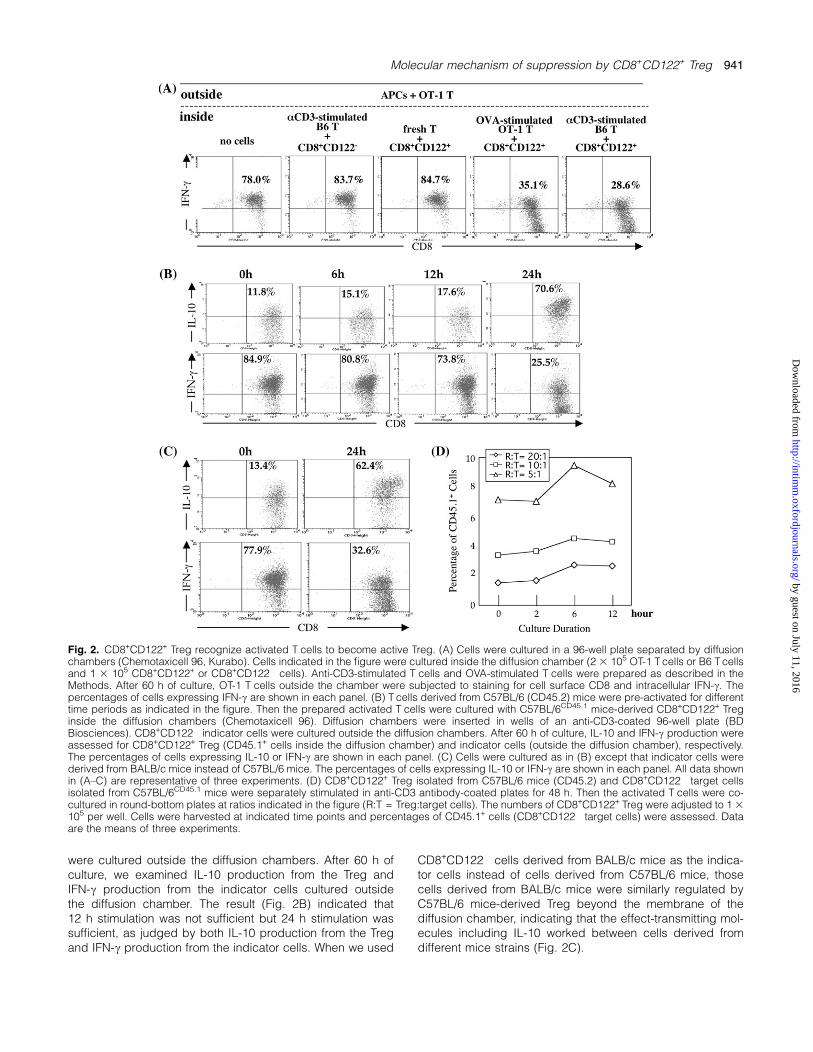
were cultured outside the diffusion chambers. After 60 h ofculture, we examined IL-10 production from the Treg andIFN-c production from the indicator cells cultured outsidethe diffusion chamber. The result (Fig. 2B) indicated that12 h stimulation was not sufficient but 24 h stimulation wassufficient, as judged by both IL-10 production from the Tregand IFN-c production from the indicator cells. When we used
CD8+CD122� cells derived from BALB/c mice as the indica-tor cells instead of cells derived from C57BL/6 mice, thosecells derived from BALB/c mice were similarly regulated byC57BL/6 mice-derived Treg beyond the membrane of thediffusion chamber, indicating that the effect-transmitting mol-ecules including IL-10 worked between cells derived fromdifferent mice strains (Fig. 2C).
Fig. 2. CD8+CD122+ Treg recognize activated T cells to become active Treg. (A) Cells were cultured in a 96-well plate separated by diffusionchambers (Chemotaxicell 96, Kurabo). Cells indicated in the figure were cultured inside the diffusion chamber (2 3 105 OT-1 T cells or B6 T cellsand 1 3 105 CD8+CD122+ or CD8+CD122� cells). Anti-CD3-stimulated T cells and OVA-stimulated T cells were prepared as described in theMethods. After 60 h of culture, OT-1 T cells outside the chamber were subjected to staining for cell surface CD8 and intracellular IFN-c. Thepercentages of cells expressing IFN-c are shown in each panel. (B) T cells derived from C57BL/6 (CD45.2) mice were pre-activated for differenttime periods as indicated in the figure. Then the prepared activated T cells were cultured with C57BL/6CD45.1 mice-derived CD8+CD122+ Treginside the diffusion chambers (Chemotaxicell 96). Diffusion chambers were inserted in wells of an anti-CD3-coated 96-well plate (BDBiosciences). CD8+CD122� indicator cells were cultured outside the diffusion chambers. After 60 h of culture, IL-10 and IFN-c production wereassessed for CD8+CD122+ Treg (CD45.1+ cells inside the diffusion chamber) and indicator cells (outside the diffusion chamber), respectively.The percentages of cells expressing IL-10 or IFN-c are shown in each panel. (C) Cells were cultured as in (B) except that indicator cells werederived from BALB/c mice instead of C57BL/6 mice. The percentages of cells expressing IL-10 or IFN-c are shown in each panel. All data shownin (A–C) are representative of three experiments. (D) CD8+CD122+ Treg isolated from C57BL/6 mice (CD45.2) and CD8+CD122� target cellsisolated from C57BL/6CD45.1 mice were separately stimulated in anti-CD3 antibody-coated plates for 48 h. Then the activated T cells were co-cultured in round-bottom plates at ratios indicated in the figure (R:T = Treg:target cells). The numbers of CD8+CD122+ Treg were adjusted to 1 3105 per well. Cells were harvested at indicated time points and percentages of CD45.1+ cells (CD8+CD122� target cells) were assessed. Dataare the means of three experiments.
Molecular mechanism of suppression by CD8+CD122+ Treg 941
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from

We also examined whether the suppressive actions ofCD8+CD122+ Treg were related to killing activity against theregulated target cells. When we co-cultured CD8+CD122+
Treg with CD8+CD122� target cells, the ratio of Treg andtarget cells did not change for the initial 12 h, suggestingthat CD8+CD122+ Treg did not actively kill the target cells(Fig. 2D). The suppressive mechanism of CD8+CD122+ Tregseems to be different from that of Qa-1-restricted CD8+ Tregthat possess cytotoxic activity against target cells (23–25).
Allogeneic target cells are not regulated by CD8+CD122+
Treg
From the result of Fig. 2(C), we understand that the finalsuppressing effect-transmitting soluble factors includingIL-10 work between CD8+CD122+ Treg and their target cellsof different mouse strains. However, does such strain-independent action occur in the previous step in which naiveCD8+CD122+ Treg become active IL-10-producing Treg? Weexamined whether CD8+CD122+ Treg have the ability to reg-ulate allogeneic target cells. First, IL-10 production in theco-culture of CD8+CD122+ Treg and pre-activated cells ofsyngeneic or allogeneic mouse strain was measured.CD8+CD122� T cells isolated from C57BL/6, BALB/c orC3H/He mice were stimulated in anti-CD3-coated plates for24 h. Then activated T cells were transferred to a new cul-ture plate without anti-CD3 antibody coating and wereco-cultured with fresh CD8+CD122+ T cells isolated fromC57BL/6, BALB/c or C3H/He mice. After 60 h, the culturesupernatant was subjected to ELISA assay to detect the pro-duction of IL-10. As we have confirmed that CD8+CD122�
cells never produce IL-10 after any kind of stimulation, theIL-10 in the culture supernatant must be produced fromCD8+CD122+ cells (Fig. 3A, left end bars, and other datanot shown here). Apparently, CD8+CD122+ Treg co-culturedwith syngeneic target cells produced IL-10 but those co-cultured with allogeneic target cells did not (Fig. 3A). Al-though the absolute amount of IL-10 in this experiment lookssmaller than that in Fig. 1, the real effect of IL-10 may bemore dependent on the concentration of IL-10 in the localarea surrounding cells than that in total culture supernatant.When we examined whether CD8+CD122+ Treg suppressedthe IFN-c production from T cells of syngeneic or allogeneicstrains, CD8+CD122+ Treg could only suppress the IFN-cproduction from T cells of syngeneic strain but could notsuppress that of MHC-mismatched allogeneic mouse strains(Fig. 3B). In these experiments, anti-CD3 antibody did notdirectly stimulate CD8+CD122+ Treg. Instead, pre-activatedT cells provided signals to CD8+CD122+ Treg. Therefore,these results are reasonably explained if cell-to-cell interac-tion involving MHC–TCR interaction is critical forCD8+CD122+ Treg to become active Treg that produce IL-10.
MHC-restricted regulation by CD8+CD122+ Treg
To confirm the MHC-restricted suppression by CD8+CD122+
Treg, we used an MHC-congeneic mouse system of C57BL/10 background. The results clearly indicated that there weremore IL-10-producing cells in the CD8+CD122+ Treg co-cultured with target cells of MHC-matched mouse strainsthan in the CD8+CD122+ Treg co-cultured with cells of MHC-
mismatched strains (Fig. 4A). This difference was clearly ob-served independent of genetic backgrounds other than theMHC haplotype (Fig. 4A). CD8+CD122+ Treg could suppressIFN-c production from MHC-matched target cells even if theother genetic background was totally different (Fig. 4B). In
Fig. 3. Regulatory action is not performed between cells of MHC-incompatible allogeneic mouse strains. (A) CD8+CD122� T cells(target) isolated from C57BL/6, BALB/c or C3H/He mice werestimulated in anti-CD3-coated plates for 24 h. Then in vitrosuppression assay as described in the Methods was performedusing CD8+CD122+ cells (Treg) isolated from C57BL/6, BALB/c orC3H/He mice. Culture supernatant was assessed for production of IL-10 by ELISA assay. Data are shown as the mean 6 SD of triplicateexperiments. *, Significantly different (P < 0.01, Student’s t test);y, not significantly different (P > 0.5, Student’s t test), compared withthose without Treg (-). (B) CD8+CD122� cells were isolated from threeMHC-incompatible strains: C57BL/6, BALB/c or C3H/He and acti-vated cells were prepared as in (A). In every combination of strains,co-culture was performed as in (A). Cells were analyzed forintracellular IFN-c. Data are shown as the histogram analysis inCD8+CD122� cells (filled histogram lines). The percentages of cellsexpressing IFN-c are shown in each panel. Open lines are data ofisotype control antibody.
942 Molecular mechanism of suppression by CD8+CD122+ Treg
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from
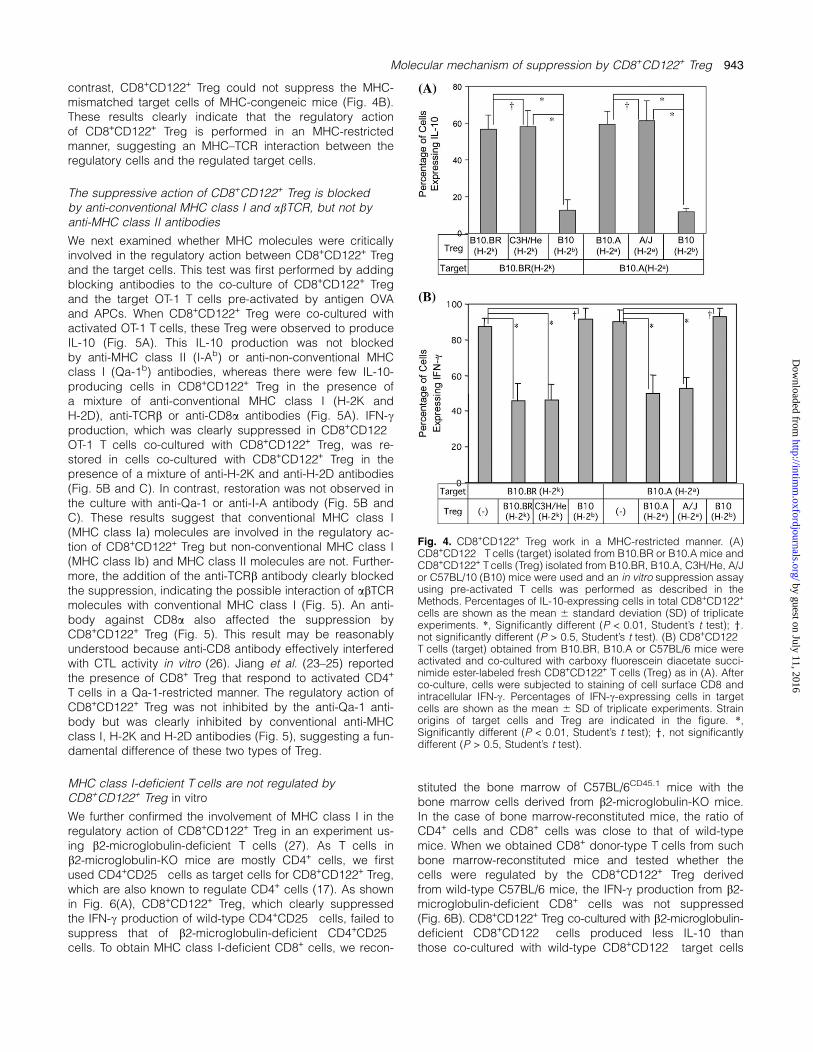
contrast, CD8+CD122+ Treg could not suppress the MHC-mismatched target cells of MHC-congeneic mice (Fig. 4B).These results clearly indicate that the regulatory actionof CD8+CD122+ Treg is performed in an MHC-restrictedmanner, suggesting an MHC–TCR interaction between theregulatory cells and the regulated target cells.
The suppressive action of CD8+CD122+ Treg is blockedby anti-conventional MHC class I and abTCR, but not byanti-MHC class II antibodies
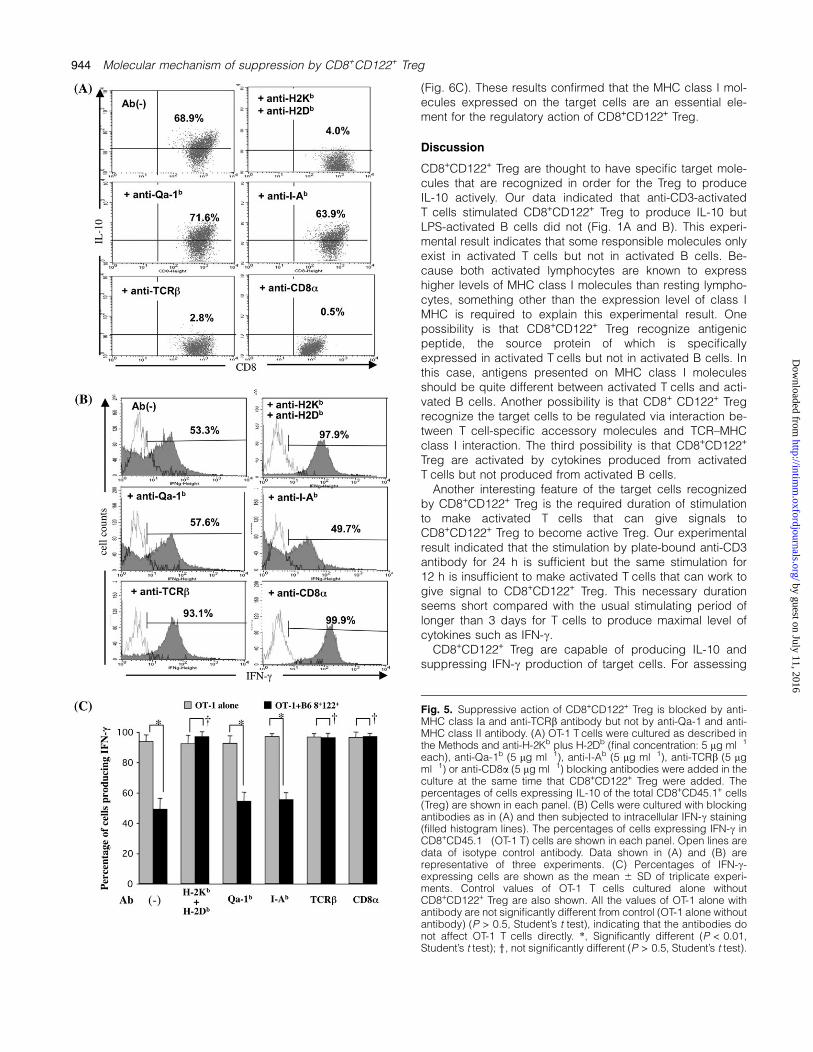
We next examined whether MHC molecules were criticallyinvolved in the regulatory action between CD8+CD122+ Tregand the target cells. This test was first performed by addingblocking antibodies to the co-culture of CD8+CD122+ Tregand the target OT-1 T cells pre-activated by antigen OVAand APCs. When CD8+CD122+ Treg were co-cultured withactivated OT-1 T cells, these Treg were observed to produceIL-10 (Fig. 5A). This IL-10 production was not blockedby anti-MHC class II (I-Ab) or anti-non-conventional MHCclass I (Qa-1b) antibodies, whereas there were few IL-10-producing cells in CD8+CD122+ Treg in the presence ofa mixture of anti-conventional MHC class I (H-2K andH-2D), anti-TCRb or anti-CD8a antibodies (Fig. 5A). IFN-cproduction, which was clearly suppressed in CD8+CD122�
OT-1 T cells co-cultured with CD8+CD122+ Treg, was re-stored in cells co-cultured with CD8+CD122+ Treg in thepresence of a mixture of anti-H-2K and anti-H-2D antibodies(Fig. 5B and C). In contrast, restoration was not observed inthe culture with anti-Qa-1 or anti-I-A antibody (Fig. 5B andC). These results suggest that conventional MHC class I(MHC class Ia) molecules are involved in the regulatory ac-tion of CD8+CD122+ Treg but non-conventional MHC class I(MHC class Ib) and MHC class II molecules are not. Further-more, the addition of the anti-TCRb antibody clearly blockedthe suppression, indicating the possible interaction of abTCRmolecules with conventional MHC class I (Fig. 5). An anti-body against CD8a also affected the suppression byCD8+CD122+ Treg (Fig. 5). This result may be reasonablyunderstood because anti-CD8 antibody effectively interferedwith CTL activity in vitro (26). Jiang et al. (23–25) reportedthe presence of CD8+ Treg that respond to activated CD4+
T cells in a Qa-1-restricted manner. The regulatory action ofCD8+CD122+ Treg was not inhibited by the anti-Qa-1 anti-body but was clearly inhibited by conventional anti-MHCclass I, H-2K and H-2D antibodies (Fig. 5), suggesting a fun-damental difference of these two types of Treg.
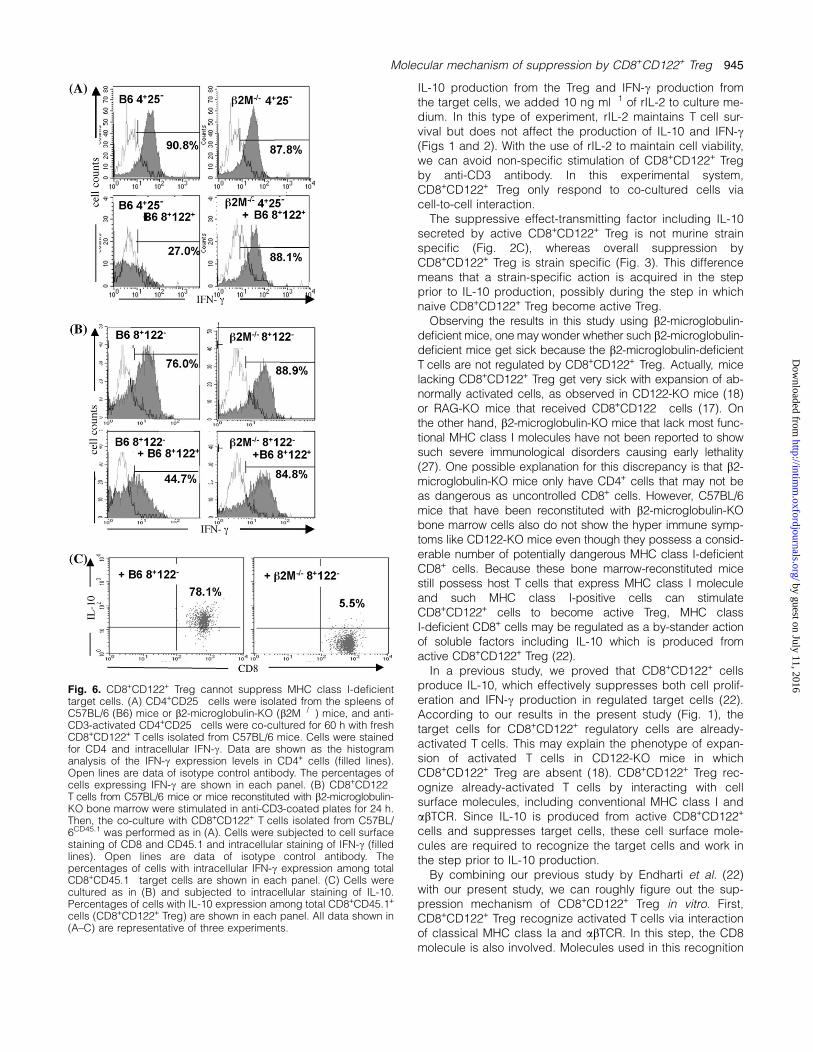
MHC class I-deficient T cells are not regulated byCD8+CD122+ Treg in vitro
We further confirmed the involvement of MHC class I in theregulatory action of CD8+CD122+ Treg in an experiment us-ing b2-microglobulin-deficient T cells (27). As T cells inb2-microglobulin-KO mice are mostly CD4+ cells, we firstused CD4+CD25� cells as target cells for CD8+CD122+ Treg,which are also known to regulate CD4+ cells (17). As shownin Fig. 6(A), CD8+CD122+ Treg, which clearly suppressedthe IFN-c production of wild-type CD4+CD25� cells, failed tosuppress that of b2-microglobulin-deficient CD4+CD25�
cells. To obtain MHC class I-deficient CD8+ cells, we recon-
stituted the bone marrow of C57BL/6CD45.1 mice with thebone marrow cells derived from b2-microglobulin-KO mice.In the case of bone marrow-reconstituted mice, the ratio ofCD4+ cells and CD8+ cells was close to that of wild-typemice. When we obtained CD8+ donor-type T cells from suchbone marrow-reconstituted mice and tested whether thecells were regulated by the CD8+CD122+ Treg derivedfrom wild-type C57BL/6 mice, the IFN-c production from b2-microglobulin-deficient CD8+ cells was not suppressed(Fig. 6B). CD8+CD122+ Treg co-cultured with b2-microglobulin-deficient CD8+CD122� cells produced less IL-10 thanthose co-cultured with wild-type CD8+CD122� target cells
Fig. 4. CD8+CD122+ Treg work in a MHC-restricted manner. (A)CD8+CD122� T cells (target) isolated from B10.BR or B10.A mice andCD8+CD122+ T cells (Treg) isolated from B10.BR, B10.A, C3H/He, A/Jor C57BL/10 (B10) mice were used and an in vitro suppression assayusing pre-activated T cells was performed as described in theMethods. Percentages of IL-10-expressing cells in total CD8+CD122+
cells are shown as the mean 6 standard deviation (SD) of triplicateexperiments. *, Significantly different (P < 0.01, Student’s t test); y.not significantly different (P > 0.5, Student’s t test). (B) CD8+CD122�
T cells (target) obtained from B10.BR, B10.A or C57BL/6 mice wereactivated and co-cultured with carboxy fluorescein diacetate succi-nimide ester-labeled fresh CD8+CD122+ T cells (Treg) as in (A). Afterco-culture, cells were subjected to staining of cell surface CD8 andintracellular IFN-c. Percentages of IFN-c-expressing cells in targetcells are shown as the mean 6 SD of triplicate experiments. Strainorigins of target cells and Treg are indicated in the figure. *,Significantly different (P < 0.01, Student’s t test); y, not significantlydifferent (P > 0.5, Student’s t test).
Molecular mechanism of suppression by CD8+CD122+ Treg 943
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from
(Fig. 6C). These results confirmed that the MHC class I mol-ecules expressed on the target cells are an essential ele-ment for the regulatory action of CD8+CD122+ Treg.
Discussion
CD8+CD122+ Treg are thought to have specific target mole-cules that are recognized in order for the Treg to produceIL-10 actively. Our data indicated that anti-CD3-activatedT cells stimulated CD8+CD122+ Treg to produce IL-10 butLPS-activated B cells did not (Fig. 1A and B). This experi-mental result indicates that some responsible molecules onlyexist in activated T cells but not in activated B cells. Be-cause both activated lymphocytes are known to expresshigher levels of MHC class I molecules than resting lympho-cytes, something other than the expression level of class IMHC is required to explain this experimental result. Onepossibility is that CD8+CD122+ Treg recognize antigenicpeptide, the source protein of which is specificallyexpressed in activated T cells but not in activated B cells. Inthis case, antigens presented on MHC class I moleculesshould be quite different between activated T cells and acti-vated B cells. Another possibility is that CD8+ CD122+ Tregrecognize the target cells to be regulated via interaction be-tween T cell-specific accessory molecules and TCR–MHCclass I interaction. The third possibility is that CD8+CD122+
Treg are activated by cytokines produced from activatedT cells but not produced from activated B cells.
Another interesting feature of the target cells recognizedby CD8+CD122+ Treg is the required duration of stimulationto make activated T cells that can give signals toCD8+CD122+ Treg to become active Treg. Our experimentalresult indicated that the stimulation by plate-bound anti-CD3antibody for 24 h is sufficient but the same stimulation for12 h is insufficient to make activated T cells that can work togive signal to CD8+CD122+ Treg. This necessary durationseems short compared with the usual stimulating period oflonger than 3 days for T cells to produce maximal level ofcytokines such as IFN-c.
CD8+CD122+ Treg are capable of producing IL-10 andsuppressing IFN-c production of target cells. For assessing
Fig. 5. Suppressive action of CD8+CD122+ Treg is blocked by anti-MHC class Ia and anti-TCRb antibody but not by anti-Qa-1 and anti-MHC class II antibody. (A) OT-1 T cells were cultured as described inthe Methods and anti-H-2Kb plus H-2Db (final concentration: 5 lg ml�1
each), anti-Qa-1b (5 lg ml�1), anti-I-Ab (5 lg ml�1), anti-TCRb (5 lgml�1) or anti-CD8a (5 lg ml�1) blocking antibodies were added in theculture at the same time that CD8+CD122+ Treg were added. Thepercentages of cells expressing IL-10 of the total CD8+CD45.1+ cells(Treg) are shown in each panel. (B) Cells were cultured with blockingantibodies as in (A) and then subjected to intracellular IFN-c staining(filled histogram lines). The percentages of cells expressing IFN-c inCD8+CD45.1� (OT-1 T) cells are shown in each panel. Open lines aredata of isotype control antibody. Data shown in (A) and (B) arerepresentative of three experiments. (C) Percentages of IFN-c-expressing cells are shown as the mean 6 SD of triplicate experi-ments. Control values of OT-1 T cells cultured alone withoutCD8+CD122+ Treg are also shown. All the values of OT-1 alone withantibody are not significantly different from control (OT-1 alone withoutantibody) (P > 0.5, Student’s t test), indicating that the antibodies donot affect OT-1 T cells directly. *, Significantly different (P < 0.01,Student’s t test); y, not significantly different (P > 0.5, Student’s t test).
944 Molecular mechanism of suppression by CD8+CD122+ Treg
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from
IL-10 production from the Treg and IFN-c production fromthe target cells, we added 10 ng ml�1 of rIL-2 to culture me-dium. In this type of experiment, rIL-2 maintains T cell sur-vival but does not affect the production of IL-10 and IFN-c(Figs 1 and 2). With the use of rIL-2 to maintain cell viability,we can avoid non-specific stimulation of CD8+CD122+ Tregby anti-CD3 antibody. In this experimental system,CD8+CD122+ Treg only respond to co-cultured cells viacell-to-cell interaction.
The suppressive effect-transmitting factor including IL-10secreted by active CD8+CD122+ Treg is not murine strainspecific (Fig. 2C), whereas overall suppression byCD8+CD122+ Treg is strain specific (Fig. 3). This differencemeans that a strain-specific action is acquired in the stepprior to IL-10 production, possibly during the step in whichnaive CD8+CD122+ Treg become active Treg.
Observing the results in this study using b2-microglobulin-deficient mice, one may wonder whether such b2-microglobulin-deficient mice get sick because the b2-microglobulin-deficientT cells are not regulated by CD8+CD122+ Treg. Actually, micelacking CD8+CD122+ Treg get very sick with expansion of ab-normally activated cells, as observed in CD122-KO mice (18)or RAG-KO mice that received CD8+CD122� cells (17). Onthe other hand, b2-microglobulin-KO mice that lack most func-tional MHC class I molecules have not been reported to showsuch severe immunological disorders causing early lethality(27). One possible explanation for this discrepancy is that b2-microglobulin-KO mice only have CD4+ cells that may not beas dangerous as uncontrolled CD8+ cells. However, C57BL/6mice that have been reconstituted with b2-microglobulin-KObone marrow cells also do not show the hyper immune symp-toms like CD122-KO mice even though they possess a consid-erable number of potentially dangerous MHC class I-deficientCD8+ cells. Because these bone marrow-reconstituted micestill possess host T cells that express MHC class I moleculeand such MHC class I-positive cells can stimulateCD8+CD122+ cells to become active Treg, MHC classI-deficient CD8+ cells may be regulated as a by-stander actionof soluble factors including IL-10 which is produced fromactive CD8+CD122+ Treg (22).
In a previous study, we proved that CD8+CD122+ cellsproduce IL-10, which effectively suppresses both cell prolif-eration and IFN-c production in regulated target cells (22).According to our results in the present study (Fig. 1), thetarget cells for CD8+CD122+ regulatory cells are already-activated T cells. This may explain the phenotype of expan-sion of activated T cells in CD122-KO mice in whichCD8+CD122+ Treg are absent (18). CD8+CD122+ Treg rec-ognize already-activated T cells by interacting with cellsurface molecules, including conventional MHC class I andabTCR. Since IL-10 is produced from active CD8+CD122+
cells and suppresses target cells, these cell surface mole-cules are required to recognize the target cells and work inthe step prior to IL-10 production.
By combining our previous study by Endharti et al. (22)with our present study, we can roughly figure out the sup-pression mechanism of CD8+CD122+ Treg in vitro. First,CD8+CD122+ Treg recognize activated T cells via interactionof classical MHC class Ia and abTCR. In this step, the CD8molecule is also involved. Molecules used in this recognition
Fig. 6. CD8+CD122+ Treg cannot suppress MHC class I-deficienttarget cells. (A) CD4+CD25� cells were isolated from the spleens ofC57BL/6 (B6) mice or b2-microglobulin-KO (b2M�/�) mice, and anti-CD3-activated CD4+CD25� cells were co-cultured for 60 h with freshCD8+CD122+ T cells isolated from C57BL/6 mice. Cells were stainedfor CD4 and intracellular IFN-c. Data are shown as the histogramanalysis of the IFN-c expression levels in CD4+ cells (filled lines).Open lines are data of isotype control antibody. The percentages ofcells expressing IFN-c are shown in each panel. (B) CD8+CD122�
T cells from C57BL/6 mice or mice reconstituted with b2-microglobulin-KO bone marrow were stimulated in anti-CD3-coated plates for 24 h.Then, the co-culture with CD8+CD122+ T cells isolated from C57BL/6CD45.1 was performed as in (A). Cells were subjected to cell surfacestaining of CD8 and CD45.1 and intracellular staining of IFN-c (filledlines). Open lines are data of isotype control antibody. Thepercentages of cells with intracellular IFN-c expression among totalCD8+CD45.1� target cells are shown in each panel. (C) Cells werecultured as in (B) and subjected to intracellular staining of IL-10.Percentages of cells with IL-10 expression among total CD8+CD45.1+
cells (CD8+CD122+ Treg) are shown in each panel. All data shown in(A–C) are representative of three experiments.
Molecular mechanism of suppression by CD8+CD122+ Treg 945
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from

phase by CD8+CD122+ Treg are quite similar to those usedby conventional CD8+ effector T cells, compared with someother CD8+ Treg that use the non-classical MHC class Ibmolecule, Qa-1 (23–25, 28). If CD8+CD122+ Treg recognizethe target cells in the same way as conventional effectorT cells, they may also use other accessory molecules such asCD28 by interacting with CD80/CD86. CD8+CD122+ Treg ex-press the same level of CD28 as other conventional CD8+
T cells, which is different from previously reported CD8+CD28�
Treg (29, 30) and actually CD28 is involved in the regulatoryaction (31). CD8+CD122+ Treg that recognized activatedT cells as their target cells become active Treg and produceIL-10, which is thought to be the essential effect-transmittingmolecule. Although the suppressive mechanism in vitro is clar-ified to some extent, the real mechanism of in vivo suppres-sion by CD8+CD122+ Treg is still unclear. Actually, the role ofIL-10 in vitro is proved without any doubt, while that in vivocan be compensated by something other than IL-10 (22).
One may wonder why CD8+CD122+ Treg exist along withCD4+CD25+ Treg. We cannot answer this question confi-dently. It may be too early to figure out the relationship andshared role of these two Treg when the real in vivo role of thesingle Treg is not clarified yet. Both Treg are indispensableand may cooperate to maintain immune homeostasis. Ourresults revealed that CD8+CD122+ Treg suppress already-activated T cells, suggesting that CD8+CD122+ Treg generallywork in the late phase of the immune reaction to calm downthe rampant immune reaction rather than suppressing spe-cific immune responses including autoimmune reactions.
This study provided the first evidence of conventionalMHC class I–abTCR interactions in the suppressive functionof naturally occurring CD8+ Treg. CD8+CD122+ Treg thathave recognized the target cells become active regulatorycells that produce effect-transmitting factors represented byIL-10. Assuming that most studies concerning the role ofTreg may have a final goal to cure autoimmune disease, weare now examining the role of CD8+CD122+ Treg in the ex-perimental autoimmune encephalomyelitis (EAE) model. Ourdata suggest that CD8+CD122+ Treg appeared to improvesymptoms of EAE (32). Since CD8+CD122+ Treg were shownto down-regulate already-activated T cells, the use ofCD8+CD122+ Treg in combination with CD4+CD25+ cellsmay have a synergistic benefit in autoimmune diseases, par-ticularly when activated T cells cannot be regulated byCD4+CD25+ alone. By targeting the molecules involved inthe action of CD8+CD122+ Treg, our findings may provideapplications of CD8+CD122+ Treg for autoimmune diseaseand transplantation-associated immune disorders.
Supplementary data
Supplementary figure are available at International Immunol-ogy Online.
Funding
Grants-in-aid for Scientific Research from the Ministry of Ed-ucation, Culture, Sports, Science and Technology of Japan(17047017); the Japan Society for the Promotion of Science(17390142).
Acknowledgements
We thank H. Tsuboi and Y. Yamada for helpful discussions and E.Nakayama for providing antibodies.
Abbreviations
APC antigen-presenting cellEAE experimental autoimmune encephalomyelitisKO knockoutOVA ovalbuminRAG recombination-activating geneTreg regulatory T cells
References
1 Cantor, H., Shen, F. W. and Boyse, E. A. 1976. Separation of helperT cells from suppressor T cells expressing different Ly compo-nents. II. Activation by antigen: after immunization, antigen-specific suppressor and helper activities are mediated by distinctT-cell subclasses. J. Exp. Med. 143:1391.
2 Okumura, K., Takemori, T., Tokuhisa, T. and Tada, T. 1977. Specificenrichment of the suppressor T cell bearing I-J determinants:parallel functional and serological characterizations. J. Exp. Med146:1234.
3 Dorf, M. E. and Benacerraf, B. 1984. Suppressor cells andimmunoregulation. Annu. Rev. Immunol. 2:127.
4 Sakaguchi, S., Sakaguchi, N., Asano, M., Itoh, M. and Toda, M.1995. Immunologic self-tolerance maintained by activated T cellsexpressing IL-2 receptor a-chains (CD25). Breakdown of a singlemechanism of self-tolerance causes various autoimmune dis-eases. J. Immunol. 155:1151.
5 Shevach, E. M. 2000. Regulatory T cells in autoimmmunity. Annu.Rev. Immunol. 18:423.
6 Sakaguchi, S. 2004. Naturally arising CD4+ regulatory T cells forimmunologic self-tolerance and negative control of immuneresponses. Annu. Rev. Immunol. 22:531.
7 Groux, H., O’Garra, A., Bigler, M. et al. 1997. A CD4+ T-cell subsetinhibits antigen-specific T-cell responses and prevents colitis.Nature 389:737.
8 Roncarolo, M. G., Bacchetta, R., Bordignon, C., Narula, S. andLevings, M. K. 2001. Type 1 T regulatory cells. Immunol. Rev. 182:68.
9 Weiner, H. L. 2001. Induction and mechanism of action oftransforming growth factor-b-secreting Th3 regulatory cells.Immunol. Rev. 182:207.
10 Cosmi, L., Liotta, F., Lazzeri, E. et al. 2003. Human CD8+CD25+
thymocytes share phenotypic and functional features withCD4+CD25+ regulatory thymocytes. Blood 102:4107.
11 Xystrakis, E., Dejean, A. S., Bernard, I. et al. 2004. Identification ofa novel natural regulatory CD8 T-cell subset and analysis of itsmechanism of regulation. Blood 104:3294.
12 Filaci, G. and Suciu-Foca, N. 2002. CD8+ T suppressor cells areback to the game: are they players in autoimmunity? Autoimmun.Rev. 1:279.
13 Zimring, J. C., Levery, S. B., Kniep, B., Kapp, L. M., Fuller, M. andKapp, J. A. 2003. CD75s is a marker of murine CD8+ suppressorT cells. Int. Immunol. 15:1389.
14 Gilliet, M. and Liu, Y. G. 2002. Generation of human CD8T regulatory cells by CD40 ligand-activated plasmacytoid den-dritic cells. J. Exp. Med. 195:695.
15 Field, A. C., Caccavelli, L., Bloch, M. F. and Bellon, B. 2003.Regulatory CD8+ T cells control neonatal tolerance to a Th2-mediated autoimmunity. J. Immunol. 170:2508.
16 Noble, A., Giorgini, A. and Leggat, J. A. 2006. Cytokine-inducedIL-10-secreting CD8 T cells represent a phenotypically distinctsuppressor T-cell lineage. Blood 107:4475.
17 Rifa’i, M., Kawamoto, Y., Nakashima, I. and Suzuki, H. 2004.Essential roles of CD8+CD122+ regulatory T cells in the mainte-nance of T cell homeostasis. J. Exp. Med. 200:1123.
18 Suzuki, H., Kundig, T. M., Furlonger, C. et al. 1995. DeregulatedT cell activation and autoimmunity in mice lacking interleukin-2receptor b. Science 268:1472.
946 Molecular mechanism of suppression by CD8+CD122+ Treg
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from

19 Takahashi, T., Kuniyasu, Y., Toda, M. et al. 1998. Immunologic self-tolerance maintained by CD25+CD4+ naturally anergic andsuppressive T cells: induction of autoimmune disease by breakingtheir anergic/suppressive state. Int. Immunol. 10:1969.
20 Thornton, A. M. and Shevach, E. M. 1998. CD4+CD25+ immuno-regulatory T cells suppress polyclonal T cell activation in vitro byinhibiting interleukin 2 production. J. Exp. Med. 188:287.
21 Thornton, A. M. and Shevach, E. M. 2000. Suppressor effectorfunction of CD4+CD25+ immunoregulatory T cells is antigennonspecific. J. Immunol. 164:183.
22 Endharti, A. T., Rifa’i, M., Shi, Z. et al. 2005. Cutting edge:CD8+CD122+ regulatory T cells produce IL-10 to suppressIFN-c production and proliferation of CD8+ T cells. J. Immunol.175:7093.
23 Jiang, H., Ware, R., Stall, A., Flaherty, L., Chess, L. and Pernis, B.1995. Murine CD8+ T cells that specifically delete autologousCD4+ T cells expressing Vb8 TCR: a role of the Qa-1 molecule.Immunity 2:185.
24 Jiang, H., Kashleva, H., Xu, L. X. et al. 1998. T cell vaccinationinduces T cell receptor Vb-specific Qa-1-restricted regulatoryCD8+ T cells. Proc. Natl Acad. Sci. USA 95:4533.
25 Jiang, H. and Chess, L. 2000. The specific regulation of immuneresponses by CD8+ T cells restricted by the MHC class Ibmolecule, Qa-1. Annu. Rev. Immunol. 18:185.
26 Nakayama, E., Shiku, H., Stockert, E., Oettgen, H. F. and Old, L. J.1979. Cytotoxic T cells: Lyt phenotype and blocking of killingactivity by Lyt antisera. Proc. Natl Acad. Sci. USA 76:1977.
27 Zijlstra, M., Bix, M., Simister, N. E., Loring, J. M., Raulet, D. H. andJaenisch, R. 1990. b2-microglobulin deficient mice lack CD4-8+
cytolytic T cells. Nature 344:742.28 Tang, X., Maricic, I., Purohit, N. et al. 2006. Regulation of immunity
by a novel population of Qa-1-restricted CD8aa+ TCRab+ T cells.J. Immunol. 177:7645.
29 Cortesini, R., LeMaoult, J., Ciubotariu, R. and Cortesini, N. S.2001. CD8+CD28- T suppressor cells and the induction of antigen-specific, antigen-presenting cell-mediated suppression of Threactivity. Immunol. Rev. 182:201.
30 Menager-Marcq, I., Pomie, C., Romagnoli, P. and van Meerwijk, J. P.2006. CD8+CD28- regulatory T lymphocytes prevent experimentalinflammatory bowel disease in mice. Gastroenterology 131:1775.
31 Shi, Z., Rifa’i, M., Lee, Y. H., Shiku, H., Isobe, K.-I. and Suzuki, H.2008. Importance of CD80/CD86-CD28 interactions in therecognition of target cells by CD8+CD122+ regulatory T cells.Immunology 124:121.
32 Lee, Y. H., Ishida, Y., Rifa’i, M., Shi, Z., Isobe, K.-I. and Suzuki, H.2008. Essential role of CD8+CD122+ regulatory T cells in therecovery from experimental autoimmune encephalomyelitis.J. Immunol. 180:825.
Molecular mechanism of suppression by CD8+CD122+ Treg 947
by guest on July 11, 2016http://intim
m.oxfordjournals.org/
Dow
nloaded from




















