
Regulatory T cells and dendritic cells in transplantation tolerance: molecular markers and...
16
Regulatory T cells and dendritic cells in transplantation tolerance: molecular markers and mechanisms Stephen P. Cobbold Kathleen F. Nolan Luis Graca Raquel Castejon Alain Le Moine Mark Frewin Susan Humm Elizabeth Adams Sara Thompson Diana Zelenika Alison Paterson Stephen Yates Paul J. Fairchild Herman Waldmann Authors’ addresses Stephen P. Cobbold 1 , Kathleen F. Nolan 1 , Luis Graca 1 , Raquel Castejon 2 , Alain Le Moine 3 , Mark Frewin 1 , Susan Humm 1 , Elizabeth Adams 1 , Sara Thompson 4 , Diana Zelenika 5 , Alison Paterson 1 , Stephen Yates 1 , Paul J. Fairchild 1 , Herman Waldmann 1 , 1 Sir William Dunn School of Pathology, University of Oxford, Oxford, UK. 2 Laboratorio de Medicina Internal, Clı ´nica Puerta de Hierro, Madrid, Spain. 3 Faculte de Medecine, Hospital Erasme, Bruxelles, Belgium. 4 University of Cambridge Neurology Unit, Addenbrookes Hospital, Cambridge, UK. 5 Center National de Ge ´notypage, Evry, France. Correspondence to: Stephen P. Cobbold Therapeutic Immunology Group Sir William Dunn School of Pathology South Parks Road Oxford OX1 3RE, UK Tel.: þ44 (0) 1865 275504 Fax: þ44 (0) 1865 275501 E-mail: [email protected] Acknowledgements This work was supported by a program grant from the Medical Research Council, UK, and by TolerRx Inc., Boston, MA, USA. Summary: Transplantation tolerance can be induced in adult rodents using monoclonal antibodies against coreceptor or costimulation molecules on the surface of T cells. There are currently two well- characterized populations of T cells, demonstrating regulatory capacity: the ‘natural’ CD4 þ CD25 þ T cells and the interleukin (IL)-10-producing Tr1 cells. Although both types of regulatory T cells can induce transplantation tolerance under appropriate conditions, it is not clear whether either one plays any role in drug-induced dominant tolerance, primarily due to a lack of clear-cut molecular or functional markers. Similarly, although dendritic cells (DCs) can be pharmacologically manipulated to promote tolerance, the phenotype of such populations remains poorly defined. We have used serial analysis of gene expression (SAGE) with 29 different T-cell and antigen-presenting cell libraries to identify gene-expression signatures associated with immune regulation. We found that independ- ently derived, regulatory Tr1-like clones were highly concordant in their patterns of gene expression but were quite distinct from CD4 þ CD25 þ regulatory T cells from the spleen. DCs that were treated with the tolerance-enhancing agents IL-10 or vitamin D3 expressed a gene signature reflecting a functional specification in common with the most immature DCs derived from embryonic stem cells. Introduction The availability of a diverse array of immunosuppressive drugs has enabled the transplantation of cells and organs to become a rapidly evolving therapeutic modality. These drugs usually require lifelong administration and patient compliance, and they risk side-effects such as susceptibility to infections, can- cer, and a host of other adverse events. Clearly, the field of transplantation would benefit if one could harness tolerance mechanisms normally used by the immune system, reducing the need for these drugs. The mechanisms exploited need not reflect the same hierarchy as those used by the normal immune system to ensure self-tolerance, but rather, those best suited to the circumstances of the patient could be util- ized. CD4 þ regulatory T cells mediating a suppression of graft rejection were discovered as a result of somewhat empirical Immunological Reviews 2003 Vol. 196: 109–124 Printed in Denmark. All rights reserved Copyright ß Blackwell Munksgaard 2003 Immunological Reviews 0105-2896 109
Transcript of Regulatory T cells and dendritic cells in transplantation tolerance: molecular markers and...

Regulatory T cells and dendritic cells
in transplantation tolerance:
molecular markers and mechanisms
Stephen P. Cobbold
Kathleen F. Nolan
Luis Graca
Raquel Castejon
Alain Le Moine
Mark Frewin
Susan Humm
Elizabeth Adams
Sara Thompson
Diana Zelenika
Alison Paterson
Stephen Yates
Paul J. Fairchild
Herman Waldmann
Authors’ addresses
Stephen P. Cobbold1, Kathleen F. Nolan1, Luis Graca1,
Raquel Castejon2, Alain Le Moine3, Mark Frewin1, Susan
Humm1, Elizabeth Adams1, Sara Thompson4, Diana
Zelenika5, Alison Paterson1, Stephen Yates1, Paul
J. Fairchild1, Herman Waldmann1,1Sir William Dunn School of Pathology,
University of Oxford, Oxford, UK.2Laboratorio de Medicina Internal, Clınica
Puerta de Hierro, Madrid, Spain.3Faculte de Medecine, Hospital Erasme,
Bruxelles, Belgium.4University of Cambridge Neurology Unit,
Addenbrookes Hospital, Cambridge, UK.5Center National de Genotypage, Evry, France.
Correspondence to:
Stephen P. Cobbold
Therapeutic Immunology Group
Sir William Dunn School of Pathology
South Parks Road
Oxford OX1 3RE, UK
Tel.: þ44 (0) 1865 275504
Fax: þ44 (0) 1865 275501
E-mail: [email protected]
Acknowledgements
This work was supported by a program grant from the
Medical Research Council, UK, and by TolerRx Inc.,
Boston, MA, USA.
Summary: Transplantation tolerance can be induced in adult rodentsusing monoclonal antibodies against coreceptor or costimulationmolecules on the surface of T cells. There are currently two well-characterized populations of T cells, demonstrating regulatory capacity: the‘natural’ CD4þCD25þ T cells and the interleukin (IL)-10-producing Tr1cells. Although both types of regulatory T cells can induce transplantationtolerance under appropriate conditions, it is not clear whether either oneplays any role in drug-induced dominant tolerance, primarily due to alack of clear-cut molecular or functional markers. Similarly, althoughdendritic cells (DCs) can be pharmacologically manipulated to promotetolerance, the phenotype of such populations remains poorly defined. Wehave used serial analysis of gene expression (SAGE) with 29 differentT-cell and antigen-presenting cell libraries to identify gene-expressionsignatures associated with immune regulation. We found that independ-ently derived, regulatory Tr1-like clones were highly concordant in theirpatterns of gene expression but were quite distinct from CD4þCD25þ
regulatory T cells from the spleen. DCs that were treated with thetolerance-enhancing agents IL-10 or vitamin D3 expressed a genesignature reflecting a functional specification in common with the mostimmature DCs derived from embryonic stem cells.
Introduction
The availability of a diverse array of immunosuppressive drugs
has enabled the transplantation of cells and organs to become
a rapidly evolving therapeutic modality. These drugs usually
require lifelong administration and patient compliance, and
they risk side-effects such as susceptibility to infections, can-
cer, and a host of other adverse events. Clearly, the field of
transplantation would benefit if one could harness tolerance
mechanisms normally used by the immune system, reducing
the need for these drugs. The mechanisms exploited need not
reflect the same hierarchy as those used by the normal
immune system to ensure self-tolerance, but rather, those
best suited to the circumstances of the patient could be util-
ized. CD4þ regulatory T cells mediating a suppression of graft
rejection were discovered as a result of somewhat empirical
Immunological Reviews 2003
Vol. 196: 109–124
Printed in Denmark. All rights reserved
Copyright � Blackwell Munksgaard 2003
Immunological Reviews0105-2896
109

attempts in rodents to induce transplantation tolerance in a
mature immune system (1, 2). As dominant effectors of tolerance
processes, they would seem eminently suitable for exploitation.
This utilization requires that we understand exactly what they are
and how they work. However, their full characterization has been
hampered by a lack of appropriate surface markers and suitable
in vitro and in vivo assays. The parallel discovery of so-called natural
(CD4þCD25þ) regulatory T cells that prevent autoimmune
disease and chronic immunopathology (3, 4) has led many to
assume that these same T cells were responsible for all forms of
regulation by CD4þ T cells. That assumption, however, has not
yet been formally demonstrated. Indeed, evidence for hetero-
geneity of regulatory T cells already exists with the description
of Tr1 cells, which can also prevent immunopathology (5).
Transplantation tolerance maintained by CD4+
regulatory T cells
There are many ways of generating tolerance, and it is not yet
clear whether the same regulatory T cells are generated or
whether they function through identical mechanisms in dif-
ferent circumstances. In general, CD4þ regulatory T cells seem
to appear in any circumstance where tolerance has been
induced to transplanted tissues without the presence of
donor-derived hematopoietic macrochimerism. Regulatory
T cells have been demonstrated where tolerance is induced
with a diversity of antibodies (CD4, CD3, CD2, and CD154)
and a range of different drugs (6). More recently, a number of
reports have demonstrated that regulatory T cells can be
elicited where antigens are presented by dendritic cells (DCs)
that are pharmacologically treated to be decommissioned from
full maturation (7–12). Following any intervention that may
generate tolerance, it may be many weeks before regulatory
T cells become detectable in functional readouts (13, 14). They
are also highly dependent on a continuous supply of antigen;
tolerant cells, parked in T-cell deficient recipients, rapidly lose
the ability to suppress unless re-exposed to antigen (13, 15, 16).
The question of antigen specificity
Regulation, once induced, shows specificity for the eliciting
antigen, yet it can extend to other alloantigens located in
the same tissue (linked suppression). For example, an A-type
animal tolerant of B will often accept grafts carrying third-
party antigens from (B� C)F1 donors (16–20). Over time,
animals then become tolerant to C-type grafts. Tolerance is
‘infectious’ (21); the use of genetically marked cell popula-
tions has demonstrated that CD4þ regulatory cells not only
suppress naıve T cells but also guide them into a state of
tolerance. Such second-generation tolerance is also of the
dominant type, i.e. it is also dependent on regulatory CD4þ
T cells (2, 16, 19, 22). This process can be reiterated over
several generations of naıve recipients. The observation of
linked suppression further supports the notion that the
antigen-presenting cell (APC) is a key component in the
regulatory process. In the same way that CD4þ T cells recog-
nize foreign proteins presented on APCs, regulatory T cells
recognize their target graft-derived antigens after they have
been processed/reprocessed by host-type APCs, i.e. through
indirect presentation (23). Regulatory T cells can be found
colonizing tolerated tissues, such as accepted skin grafts (24),
and T cells from such tolerated tissue can repopulate the
peripheral immune system of immunodeficient recipients so
as to prevent graft rejection by naıve T cells. T cells from
tolerated kidneys, for example, proved exceptionally potent
compared to those from spleen, suggesting enrichment within
the tolerated tissue (25).
Identifying regulatory T-cell populations
A number of key properties of the dominant tolerant state have
been defined, but it is not yet clear whether the emerging
functions of either CD4þCD25þ or Tr1-like regulatory cells are
sufficient to explain all of these properties. Adoptive transfer
studies in lymphopenic mice have suggested that, in the case
of grafts mismatched at multiple minor histocompatibility
antigens, both CD4þCD25þ and CD4þCD25– T cells can
suppress rejection by naıve T cells. Similar adoptive transfer
studies, with T cells from animals tolerant across major histo-
compatibility complex (MHC) barriers, suggest that only
CD4þCD25þ T cells can be shown to block rejection (26,
27). Contrary to published reports (28), we have found no
evidence that CD4þCD25þ T cells carry the specificity for the
tolerizing antigen, as T cells from strain A mice tolerant of
B-type grafts are capable of preventing rejection of grafts
from strain C. Even naıve CD4þCD25þ T cells are able to do
the same (LG and ALM, unpublished data) (Fig. 1).
This finding suggests that our suppression readouts in
lymphopenic hosts may only represent one aspect of regulatory
T-cell activity. In such circumstances, these cells may be acting
to regulate homeostatic expansion irrespective of antigen, and
they may also be more effective competitors in repopulation
(29). In light of these data, the surface phenotype of regulatory
T cells in transplantation tolerance remains unresolved.
CD4þ T-cell populations from tolerant mice are able to
suppress rejection by both naıve and primed CD4þ or CD8þ
Cobbold et al �Regulatory T cells and APCs in tolerance
110 Immunological Reviews 196/2003

T cells (30) on adoptive transfer to lymphopenic recipients.
Tr1-like clones can also prevent graft rejection by fully differ-
entiated effector T-helper (Th)-1 and Th2 clones (Fig. 2) in
such lymphopenic models (31). Consequently, it would be
important to establish the relationship, if any, between
CD4þCD25þ T cells and Tr1-like cells and to determine
whether both may play a part in dominant tolerance. Unfortun-
ately, there are too few discriminating surface markers
described for this study to be done at present.
Naıve T cells introduced into a mouse exhibiting dominant
tolerance can still proliferate and accumulate in response to
graft antigens, but the cells may fail to become competent
effectors (32, 33). There is no a priori theoretical reason or
evidence to suggest that suppression blocks necessarily the
initial T-cell proliferative response to antigen. Indeed, there
has never been a strong correlation between tolerance and any
loss of donor antigen-driven T-cell proliferation, whether
measured in vitro or in vivo, in the dominant transplantation
tolerance models (6, 15, 32). Therefore, we have to remain
very cautious in any functional interpretations based on the
suppression of proliferation in vitro or on the homeostatic
expansion of T cells in vivo. It may be that many of the
molecules so far implicated in suppressive mechanisms, such
as cytotoxic T-lymphocyte antigen-4 (CTLA-4), interleukin
(IL)-10 (26, 34), and transforming growth factor (TGF)-b(35), are only major contributors under experimental condi-
tions where an inhibition of proliferation is a required com-
ponent.
We have interpreted the fundamental observations above as
indicating that the generation of regulatory T cells is a normal
evoked response of the immune system where immunity has
been compromised by antibodies or drugs. The data also
CD25+
CD25+
H-2dTolerant
H-2dTolerant
B10BRTolerant
CBA (H-2k) RAG-KO recipients
CD25–CD4+ T cells
CD4+ T cells
CD4+ T cells
CD4+ T cells
CD4+ T cells
CD25–
CD25–
Naïve
Skin graft
Rejection
Rejection
Non-specificsuppression
Non-specificsuppression
Specificsuppression
Skin grafts
12 weeks
12 weeks
12 weeks
12 weeks
12 weeks
Naïve
H-2d H-2d H-2b
H-2d H-2b
H-2d H-2b
H-2d H-2b
H-2b
H-2d
H-2d
H-2d
B10BR B10BR
Fig. 1. A comparison of CD25þ and CD25–
regulatory T cells in transplantation models. Arecognized assay for regulatory T-cell activity is tocotransfer the population of interest intoimmunodeficient recipient mice together withnormal naıve spleen cells capable of rejecting a graft,unless they are suppressed by the test population.Spleen cells from either tolerant mice (given a skingraft and non-depleting anti-CD4, anti-CD8, andanti-CD40L mAbs at least 4 weeks previously) ornormal mice can be sorted (magnetically or in aflow cytometer) into CD4þ populations that areeither CD25þ or CD25–, and cotransferred withnaıve CD4þ T cells into immunodeficient[recombination-activating gene (RAG)-1 knockout]recipient mice. Such mice receiving CD25– cells areunable to suppress the rejection of majorhistocompatibility complex (MHC)-mismatchedskin grafts, regardless of whether the cells camefrom tolerant or naıve donors, and in fact, CD25–
cells from tolerant mice can cause rejection on theirown (data not shown). Mice given CD25þ cells,however, accept skin grafts from MHC incompatibledonors, and this acceptance does not require the Tcells to have previously experienced the donorantigens, as naıve or tolerant CD25þ cells are equallyeffective. Furthermore, if recipient mice arerechallenged with fresh donor plus third partygrafts, the suppression observed is not specific to theoriginal donor-type MHC antigens. In contrast,when skin grafts differ only in non-MHC antigens(i.e. minor antigens), the CD25– cells from tolerantmice (but not naıve mice) are not only suppressivebut also are able to transfer antigen-specificregulation and infectious tolerance. CD25þ cellsfrom either tolerant or naıve mice are able tosuppress the rejection of non-MHC skin grafts, butas above, the effect is found to be non-antigenspecific.
Cobbold et al �Regulatory T cells and APCs in tolerance
Immunological Reviews 196/2003 111

suggest that the influence of regulatory T cells is local rather
than systemic. This effect could result from their action on
inductive events within the microenvironment of the APC, at
the effector stage in the transplant itself, or both. Infectious
tolerance, the process by which one set of regulatory T cells
encourages development of further cohorts of regulatory
T cells, could be explained as a consequence of graft accept-
ance (6). An accepted healed graft would provide a steady
source of antigens to immature DCs, so enabling these DCs to
elicit further regulatory T-cell function from the naıve T-cell
population that can ‘chronically’ stimulate new T cells, which
are prevented from rejecting the graft. In other words, the
process of generating and maintaining further cohorts of
regulatory T cells appears to be self-sustaining once the graft
is accepted. Removal of the graft would, as a corollary, remove
the drive for this process.
Drugs or blocking therapeutic antibodies that enable regulatory
T cells to be generated
We have, in a previous review, proposed the following
explanation for how drugs and antibodies may bring about
dominant tolerance (36). During the induction phase of
tolerance, effective agents block rejection by preventing
T cells from generating effector function. Consequently,
the inflammation in the graft settles down and the graft
heals. Graft antigens constitutively released by the
transplant organ will be processed and presented to
T cells by host APCs. Such APCs would remain decom-
missioned, as they would have no ‘danger’ signals (37)
to activate them. Instead, we have proposed, they would
generate chronic, albeit incomplete, signals to T cells
that recognize them. Sustained but incomplete signaling,
we propose, is what drives T cells to regulatory activity.
All T cells could be vulnerable to such sustained incom-
plete signaling irrespective of their prior history, i.e. Th1,
Th2, or Th0. The result would be that some T cells
become regulatory, resetting their threshold for activation
so as to appear functionally anergic. Aware of the
presence of anergic T cells in recipients showing dom-
inant tolerance (38, 39), we speculated that such T cells
could include regulatory T cells. The ‘Civil Service’
analogy (6, 40, 41) was coined to reflect the ability of
compromised ineffectual T cells to obstruct the function
Th1
4 weeks
4 weeks
4 weeks
4 weeks
Skin grafts Skin graft
Rejection
Rejection
Suppression
Suppression
Male MaleFemale
Female RAG-KO recipients
Th2
Anti-HY
Anti-HY
Anti-HY
Anti-HY
Tr1
Tr1
Th1
Th2
Th1
Th2
Fig. 2. Tr1-like clones can suppress rejection byboth Th1 and Th2 clones with identical
specificity. T-cell clones with identical specificity forthe male antigen H-Y as presented by H-2Ek weregenerated from female A1�RAG-1–/– T-cellreceptor-transgenic mice. It was found that both Th1and Th2 clones were able to rapidly reject male, butnot control female, skin grafts given to recipientRAG-1 knockout mice. Such mice could be givenfresh male skin grafts and further injections of theTh1 or Th2 clones and were still able to causerejection as expected. In contrast, Tr1-like cloneswere unable to cause skin graft rejection. If theserecipients were later given fresh male skin grafts andsufficient Th1 or Th2 clones to cause rejection incontrol mice that had not previously received Tr1cells, rejection was abolished, demonstrating theability of Tr1-like cells to act as regulatory cells in atransplantation setting.
Cobbold et al �Regulatory T cells and APCs in tolerance
112 Immunological Reviews 196/2003

of otherwise competent T cells. This hypothesis predicted
that any agent enabling chronic, but incomplete signaling,
of T cells would achieve the same outcome. Drugs that
decommission DCs or those that interfere with effector
functions of T cells might all enable graft acceptance and
consequently guarantee a continuous source of antigen to
sustain the incomplete signals to T cells. Regulatory T cells
would, by virtue of their appropriate T-cell receptor (TCR),
localize to sites of maximal antigen exposure and act there to
subvert the immune (rejection) response.
Characterizing the regulatory T cells and the dendritic cells that
drive them
From the discussion above, it is clear that further charac-
terization of regulatory T cells, the DCs that drive them,
and any heterogeneity of such cells requires us to establish
what patterns of gene products they might be expressing,
both as diagnostic markers and as plausible explanations for
their regulatory function. The following sections represent
our efforts in this direction, and we not only provide the
reader with a list of gene products of interest but also a
resource that we hope others will utilize to compare the
phenotypes of regulatory T cells and tolerogenic DCs in
their own laboratories.
Gene expression analysis of regulatory T cells and
dendritic cells
It is clear from the discussion above that we are in desperate
need of molecular markers that can distinguish between dif-
ferent regulatory T cell and dendritic cell populations. It even
remains a fundamental question whether there are specific
T cells with a ‘professional’ regulatory function or more global
mechanisms of immune regulation, shared across a range of
cells. One way to approach these problems is to correlate gene
expression with functional subsets in a broad and unbiased
way, without any preconceptions about which gene products
are likely to be discriminatory.
Proteomics is still insufficiently advanced for this purpose.
Alternatively, one can analyze mRNA transcripts as an indirect
measure of potential protein expression. The two main
methods are based either on hybridization to a microarray of
known cDNAs or sampling and sequencing of short tags
reflecting the entire population of mRNA from a given cell
type. While microarrays have some advantages in conveni-
ence, they are limited to known gene sequences and are
difficult to standardize, due to variability of sequence
dependent hybridization. Serial analysis of gene expression
(SAGE) (42) works by extracting tags and concatenating
them for efficient sequencing, and the relative abundance of
each tag is a measure of the frequency of the associated
transcript. Such data then, in principle, can be directly
compared between libraries, even if they derive from different
laboratories.
We focused our analysis on the question of how the
immune system is regulated and whether there are gene
signatures that correlate with pro-tolerogenic compared to
immunogenic phenotypes. We used SAGE libraries of well-
characterized resting Th1, Th2, and Tr1-like clones (43, 44) as
a starting point, and then we included a range of purified
ex vivo T cells (27) and APCs (dendritic and B cells) at different
stages of maturation and after treatment with agents thought
to bias antigen presentation for tolerance (10, 45). We have
been able to identify gene signatures of regulatory T cells,
which suggest that these cells reflect an altered state of
known T-cell types, and of pro-tolerogenic DCs, which reflect
expression of a defined gene set. In addition, this study
establishes the limits of utility of current SAGE methodology
for this purpose.
Reliability and limitations of SAGE analysis
The SAGE analysis presented here is based on 29 different
libraries (Table 1) and is made up of 811 804 tags, containing
24259 unique tags represented three or more times in total. Of
these, 682 tags were mapped to ribosomal proteins (excluded
from further analysis), and 625 tags were hand annotated for
known genes. Four thousand seven hundred and sixty two were
computationally assigned to known mRNAs, 4117 to Unigene
clusters with a putative poly A signal sequence, 1853 to
expressed sequence tags (ESTs) from Unigene clusters without
a poly A signal, and 13 420 were unassigned.
The high number of unassigned tags was of considerable
interest. This occurrence has been noted in previous SAGE
analyses (46–48), where the unassigned tags were considered
novel transcripts (48). Closer inspection of these tags in our
analysis suggested that approximately 60% of them could be
explained by non-random accumulation of sequence errors in
highly abundant gene tags or linker sequences. Using the
highly abundant ferritin heavy chain tag as an example, 21
tags differing by a single base were found to occur >6 times
within the data. The most abundant putative error tag repre-
sented approximately 3%, and similar tags made up approxi-
mately 12% of the total. In addition, tags from incompletely
Cobbold et al �Regulatory T cells and APCs in tolerance
Immunological Reviews 196/2003 113

Table 1. Details of SAGE Libraries
Library name Details of cell type Number of tags Preamplified method NCBI GEO accession References
Th1 Th1 anti-HY cloneR2.2: derived fromprimed A1�RAG-1–/–
female (resting)
10498 SMART GSM3677 43
Th2 Th2 anti-HY cloneR2.4: derived fromprimed A1�RAG-1–/–
female (resting)
11159 SMART GSM3678 43
Tr1D1(Mast) Treg anti-HY cloneTr1D1 derived fromnaıve A1�RAG-1–/–
female (resting): contains�2% mast cells
30246 SMART GSM3679 43
A1RAG_nodes Draining lymph nodesfrom male skingrafted A1xRAG-1–/–
mice (day 7)
11525 None GSM3680 43
CBA_nodes Draining lymph nodesfrom B10.BR skingrafted CBA/Camice (day 7)
5597 None GSM3687 43
CD4þCD25-Act Splenic CD4þCD25–
T cells (MACS sortedfrom CBA/Ca):CD3 activation (16 h)
21446 SMART GSM3685 27
CD4þCD25– Splenic CD4þ
CD25– T cells(MACS sorted fromCBA/Ca): resting
23802 SMART GSM3683 27
CD4þCD25þ Splenic CD4þCD25þ
T cells (MACS sortedfrom CBA/Ca):resting
21243 SMART GSM3686 27
CD4þCD25þAct Splenic CD4þCD25þ
T cells (MACS sortedfrom CBA/Ca):CD3 activation (16 h)
20926 SMART GSM3684 27
Tr1D1_CD4 Treg clone Tr1D1 (resting):APC depleted,CD4þ (MACS sorted)
12127 SMART GSM3681 43
Skin_LineA Treg line grown fromtolerated male skingraft on A1(M) female mouse
22135 SMART GSM3682 43
Skin_LineB Treg line grown in anti-CTLA4mAb, from toleratedmale skin on A1(M) female
20219 SMART GSM3824
Tr1D1_Act Treg clone Tr1D1 (resting):APC depleted,CD4þ (MACS sorted):CD3 activation (16 h)
21336 SMART GSM3825
Th2_Act Th2 clone R2.4:CD3 activation (16 h)
11316 SMART GSM3826
Th1_Act Th1 clone R2.2:CD3 activation (16 h)
20848 SMART GSM3827
BW5147 Thymoma cell linefrom AKR mice
19750 None GSM3828
ESF_116 Embryonic stem cell line(CBA/Ca derived): multiplepassages in LIF without feeders
15711 None GSM3829
ESDC Embryonic stem cell(ESF116) derived dendritic cells
31723 None GSM3830
Cobbold et al �Regulatory T cells and APCs in tolerance
114 Immunological Reviews 196/2003

NlaIII-digested mRNA were present at up to 5%, consistent
with previous observations (49). The occurrence of these
artefactual tags tended to mirror that of the most abundant
correct tag. Non-random sequencing errors were also found in
many other Unigene ESTs. It has been suggested that these
errors might represent true polymorphisms in human SAGE
analyses (48). In the mouse, most ESTs derive from a few
inbred strains, so they are probably sequence errors.
Many published SAGE libraries claim to approach the entire
transcriptome (50) of a given cell type, often exceeding
100 000 tags (51). The limitations shown above suggest that
artefacts accumulate faster than novel gene tags once the
library reaches a depth equivalent to 0.1–1% of the most
abundant transcripts (in practice between 10 000 and 30 000
total tags), and so most of the libraries described here are
limited to this size. Tags were excluded if they remained
unidentified and differed in sequence by only one base from
a more abundant (�10) tag, linker, or primer sequence. In the
future, the new long SAGE method (52) should overcome this
limitation by providing sequence redundancy.
ESDC_LPS Embryonic stem cell (ESF116)derived dendritic cellsþ LPS
31861 SMART GSM3831
BMDC_LPS Bone marrow-deriveddendritic cells (day7)þ LPS
13518 None GSM3832
BMDC Bone marrow-deriveddendritic cells (day7)
22126 None GSM3833
BMDC_IL10 Bone marrow-deriveddendritic cellsþ IL-10 (days 6–9)
31561 None GSM3834
BMDC_IL10LPS Bone marrow-deriveddendritic cellsþ IL-10(days 6–9)þ LPS
31531 None GSM3835
B-cells Splenic B cells(MACS sorted): resting
10148 None GSM3837
3T3_Fibro Untransformed3T3 fibroblasts
28531 None SAGENet* Veculescu, V.(unpublished data)
R1_ES Embryonic stemcell line (B6 derived): R1
137906 None GSM580 53
Brain Normal mouse braingranular precursor cells
61256 None GSM767 Wechsler-Reya, R.,et al.(unpublished data)
abIEL a/b intraepitheliallymphocytes (CD8þ)
91000 None See URLy 54
Grand Total All of the above libraries 811804
Legend to SAGE libraries (43): Total RNA was isolated using thiocyanate buffer and pelleted through a 5.7 M CsCl cushion. The poly(A)þ fraction waspurified using oligo(dT)25 dynabeads (Dynal, Oslo, Norway), and cDNAs generated using a kit from Roche (Lewes, UK). In the case of SMART libraries,total RNA was isolated using the SV total RNA isolation system (Promega). First strand cDNAs were prepared from approximately 1 mg of total RNAusing the anchoring primer: 50-GACTCGAGTTGACATCGAGG(T)20V-30 with the SMART PCR cDNA synthesis kit (Clontech, Palo Alto, CA, USA),and preamplified using the forward SMART PCR and reverse 50-GACTCGAGTTGACATCGAG-30 primers and the advantage-GC cDNA PCRpolymerase mix (Clontech), with 1 M of GC-Melt, for 16–25 cycles of 94 �C for 30 s, 68 �C for 7 min (43). All SAGE used NlaIII as anchoring enzyme,BsmF1 as tagging enzyme, and SphI as cloning enzyme, as described (42, 80). DNA sequencing used either the 377 (ABI) or MegaBACE (MolecularDynamics, Piscataway, NJ, USA) automated sequencers.Generation of Th1, Th2, Treg, and Tskin CD4þ T-cell clones: The production of Th1, Th2, and Tr1D1 clones, and lines from tolerated skin grafts, havebeen described elsewhere (31, 43). Short-term polarized lines were generated from A1(M)�RAG-1–/– (43) mice after Barret et al. (81).MACS enrichment of CD4þ T-cell clones: T-cell cultures were stimulated for 7–14 days with mitomycin-treated male spleen cells from CBK (Kbþ)transgenic mice, and the Kbþ cells depleted by AutoMACSTM (Miltenyi Biotec, Auburn, CA, USA) as described (43). Purity of all fractions was >97%CD4þKb– cells. Activation was overnight with 1mg/ml of plate bound anti-CD3 (KT3).MACS enrichment of spleen CD4þCD25þ cells (27): CBA/Ca mice were depleted of CD8þ T cells and MHC-IIþ and Igþ cells depleted with Dynabeads(Dynal). CD25þ cells were positively selected using biotin antimouse CD25 (Pharmingen, San Diego, CA, USA) and streptavidin MACSTM microbeads.This CD4þCD25þ fraction was >90% pure by fluorescence-associated cell sorter (FACS) analysis. CD4þ cells were positively selected from the CD25–
fraction and were >98% CD4þCD25– cells. Activation was overnight with 1 mg/ml of plate bound anti-CD3 (KT3).Splenic B cells: The B-cell untouched kit (Miltenyi Biotec) and AutoMACSTM was used to obtain a population >93% CD3–B220þ cells from T-cell-depleted CBA/Ca mice.Bone marrow derived DCs: Day 7 bmDCs were generated from CBA/Ca mice (82). 1a,25-dihydroxycholecalciferol(10�7 M day 3–7; Sigma),recombinant mouse IL-10 (20 ng/ml day 6–9, R&D Systems), or lipopolysaccharide (LPS) (1mg/ml for the final 18–20 h; Sigma) were added asappropriate. Each population was >90% CD11cþ, MHC-IIþ and was tested in an allogeneic mixed lymphocyte reaction.ES-derived DCs: ESF116 was maintained and differentiated to dendritic cells (esDC) as described (83), and the esDC matured using LPS (84). ESF116cells for SAGE analysis were passaged six times on gelatinized flasks in 1000 U/mL of recombinant leukemia-inhibitory factor.Database accession numbers: All SAGE libraries from our laboratory have been deposited at GenBank under the NCBI Gene Expression Omnibus SAGEaccession numbers GSM 3677–3687 and GSM 3824–3837 inclusive.*http://www.sagenet.org/SAGEData/3T3.htmyhttp://www.kcl.ac.uk/ip/jeremycridland/research/sage/SAGE_IEL_Master_Database.xls
Cobbold et al �Regulatory T cells and APCs in tolerance
Immunological Reviews 196/2003 115

An important aspect of the SAGE approach is the reliability
of interlibrary comparisons. A comparison of two libraries
from independent regulatory T-cell clones (Fig. 3A) demon-
strated very few differences. Further, two independent
embryonic stem (ES) cell lines [one library from the literature
(53) and one generated in house] were also similar (Fig. 3B),
with differences being compatible with residual fibroblasts
(used to maintain ES cell lines) in the R1 library, as indicated
by overexpression of genes such as procollagen type Ia2 and
fibronectin 1. This finding confirms that useful comparisons
can be made between SAGE data from our own and other
laboratories, and therefore four published datasets [one a/bintraepithelial lymphocyte (IEL) (54) and three non-immune
cell types, see Table 1] have been included.
Relationships between cell types defined by gene expression
patterns
Cluster analysis was performed using different tag abundance
thresholds (from 100 to 4000 of the most abundant, but
excluding all ribosomal derived tags) and using a variety of
methods. Most analyses generated similar clusters, and a repre-
sentative dendrogram is shown in Fig. 4A. The relationships
between cell types was broadly compatible with expectations:
CD4þ T-cell clones clustered together, with the three inde-
pendent Tr1-like regulatory T cell (Treg) clones most closely
related [Fig. 4A(iv)], but separate from the ex vivo purified
naıve T cells [Fig. 4A(vii)]. In vitro generated DCs clustered
together [Fig. 4A(ii)], whether derived from bone marrow
(bmDC) or ES cells (esDC), while the two ES cell lines were
most similar to each other [Fig. 4A(i)]. Ex vivo derived resting
B cells clustered with the lymph node populations from nor-
mal or TCR-transgenic mice [Fig. 4A(iii)]. Surprisingly, CD3-
activated Th1, Th2, and Tr1-like clones were generally more
similar to each other [Fig. 4A(v)] than to their resting partners,
but the cells did not particularly resemble CD3-stimulated
naıve CD4þ T cells, whether CD25þ or CD25–.
Distinct clusters of gene tags were associated with each cell
cluster (Fig. 4B). The three genes most strongly correlated
with ES cells were pH 34 (ES cell specific gene 1), Unigene
cluster Mm.30502 and Oct3/4, the latter being a transcrip-
tion factor associated with pluripotency (55). Potential
markers for DCs included CCL9, C-type lectin (calcium
dependent) member 10, and cathepsin H, although these
genes are more widely expressed on myeloid cells. The
B-cell and lymph node cluster included components of the
B-cell receptor [immunoglobulin (Ig) M heavy and J chains,
CD79a,b], although most other tags in this cluster were more
widely distributed (e.g. Igk). The resting Th1 and Th2 clones
clustered together with the Tr1D1 culture dominated by mast
cell products (44). This cluster expressed an unusually high
number of unknown tags as well as CD4 and the g chain of
the common cytokine receptor. The genes most closely
0
0.7
2.1
4.6
9
16.
30.
Ski
n tr
eg li
ne B
Skin treg line B ES cell line ESF116
ES
cel
l lin
e R
155.
99
176
>320 A B
0.7 2.1 4.6 9 16. 30. 55. 99 176 >3200
0.7
2.1
4.6
9
16.
30.
55.Over expressed
Key:99
176
>320
0.7 2.1 4.6 9 16. 30. 55. 99 176 >320
(Nor
mal
ized
tag
freq
uenc
y ×1
04)
(Normalized tag frequency ×104)
Uniquelyexpressed
Non-differential
Common(house keep)Excluded tags
Fig. 3. Reproducibility of serial analysis of gene expression (SAGE)
library data. Scatter plots of SAGE libraries from two independent, skin-derived, Tr1-like lines (A) and two unrelated ES cell lines generated bydifferent laboratories (B). The normalized frequency of each tag is plottedwith green representing non-differential tags and gray representing‘housekeeping’ transcripts, defined as tags with a standard error less thanthe mean across all the libraries. The marked cone encloses an area of>95% confidence of at least 1.5-fold difference, and overexpressed tagsare color-coded as indicated. Tags uniquely overexpressed in only the SAGElibrary shown are marked in red. SAGE data analysis: SAGE 3.04 beta (KWKinzler, Johns Hopkins Oncology Center, Baltimore, MD, USA) was usedto extract ditags and unique tags before export into Access (Microsoft,Seattle, WA, USA). Automated unique tag to gene assignments used the
Unigene full mapping files and a search algorithm giving priority to hand-annotated genes, followed by cDNAs with a polyA signal in the correctorientation, then EST clusters with a probable polyA signal, cDNAs with nopolyA signal, and finally other matching ESTs. The tabulated data wasimported into custom written software (!SAGEClus) for cluster analysis andto generate scatter plots and charts. Tags were excluded if they matched 9/10 bases of any other tag that occurred at a >10-fold frequency within theentire dataset or any linkers or primers used. A conservative estimate of thedifferential upregulation of each was calculated using a Bayesian statisticsmodel (43, 79). An upregulation of 1.5-fold with 95% confidence wasgenerally considered significant. The !SAGEClus software and collated SAGEdata tables can be obtained from the website at http://www.molbiol.ox.ac.uk/pathology/tig/software/softlist.html.
Cobbold et al �Regulatory T cells and APCs in tolerance
116 Immunological Reviews 196/2003

correlating with the cluster of three Tr1-like cells included
some already published as potential Treg markers, such as
glucocorticoid-induced tumor necrosis factor receptor family-
related gene (GITR) (56) and CD137 (57).Most of the transcripts,
however, were highly expressed only on resting Tr1-like cells and
were downregulated after activation with anti-CD3 monoclonal
antibody (mAb). After activation, Tr1D1 clustered with CD3-
activated Th1 and Th2 clones, partly due to coexpression of the
chemokine CCL4 and other unassigned tags.
The analysis of splenic CD4þ T cells was based on a fractionation
into CD25þ and CD25– cells, as the former are thought to include
naturally occurring ‘professional’ Treg cells (58).Most of the SAGE
tags overexpressed in these ex vivo T cells remain uncharacterized
(Fig. 4B). Surprisingly, no splenic subpopulation was closely
related in its pattern of transcripts to any of the T-cell lines or
clones maintained in vitro, even after CD3 activation. a/b IELs and
the BW5147 thymoma were not associated with any of the other
T-cell libraries, although there were no other representatives of
non-CD4þ subsets.
The expression pattern of the few restricted genes (13–30
per cluster and <150 in total) was sufficient to generate
a similar dendrogram, but their removal from the database
did not abolish the clusters. This finding suggests that subtle
patterns of gene expression define genetic and therefore, by
implication, functional relationships between different
immune cell types. The evidence for truly lineage- or subset-
specific markers is relatively weak.
Global patterns of gene expression are primarily associated
with context
Very few genes were found to associate with conventional
lineage relationships. The only genes expressed on all T cells
A BDistance between clusters of cell types (%)
10 20 300
α/β IEL (CD8+)
Brain (PGCP)
R1 (ES cell line)
3T3 (Fibroblast)
ESF-116
BMDC + IL-10 + LPS
ESDC
BMDC + VitD3
BMDC
BMDC + IL10
BMDC + LPS
ESDC + LPS
Th1 (CD3 activated)
Th2 (CD3 activated)
Tr1D1 (CD3 activated)
Skin Treg Line B
Skin Treg Line A
Tr1D1 clone (CD4+)
Splenic B cells
CBA nodes
A1 × RAG nodes
Tr1D1(inc. mast cells)
Th2 clone (R2.4)
Th1 clone (R2.2)
CD4+CD25+ (CD3 act.)
CD4+CD25+ spleen
CD4+CD25– spleen
CD4+CD25– (CD3 act.)
BW5147 (thymoma)
i
ii
iii
iv
v
vi
viiT
h1 (
clon
e R
2.2)
Th2
(cl
one
R2.
4)T
r1D
1 (in
c. m
ast)
A1
× R
AG
–/– n
odes
C
BA
nod
esC
D4+
CD
25+ (
CD
3)C
D4+
CD
25+
CD
4+C
D25
–
CD
4+C
D25
– (C
D3)
Tr1
D1
clon
e (C
D4)
Ski
n tr
eg li
ne A
S
kin
treg
line
B
Tr1
D1
clon
e (C
D3)
Th2
clo
ne (
CD
3)T
h1 c
lone
(C
D3)
BW
5147
thym
oma
ES
F-1
16 (
ES
cel
ls)
ES
DC
ES
DC
+ L
PS
BM
DC
+ L
PS
BM
DC
BM
DC
+ IL
-10
BM
DC
+ IL
-10
+ L
PS
BM
DC
+ V
itD3
B c
ells
3T3
fibro
blas
tR
1 (E
S c
ells
)B
rain
(P
GC
P)
IEL
(α/β
CD
8+)
Unknown, GTAACAACGCUnknown, TCCTACAGTGESTs, TGGTAACAAC, 35653Unknown, TCCTAAAGTGUnknown, GAAAATGATAIL-17/poly Ig R, ATTTTCAGAT, 5419Unknown, GAATATGGCAUnknown, CCAGGTATGAUnknown, TCTAAGTACGRIKEN 1110013G13, ATCGGTTCCA, 212Unknown, GCCGTTCTTAUnknown, CCGACGGGCGUnknown, GGATGTCTCTUnknown, GTCAAGCGAGtrans. reg. SIN3A, CACACACACA, 15755
GITR (TNFRsf18); CTCTGCACCC, 3180CD137 (4-1BB) alt sp1, GGAAACAACT, 3720RGS-1 truncated, CCAAGGTTGA, 103701GA3PDH seq error?, GCTCCAAGGA, 5289kallikrein 8, GACAACAGGG, 5193Unknown, AGGAAGCGGCUnknown, ACCGCCCACARIKEN 2200003J09, ACACCTTCCT, 21160Unknown, ACCAATGAACUnknown, AGTCGGGTGGprotein tyr. phosphatase, GCTGACACTT, 361RIKEN 1110001H19, GGAGCACTGG, 732histone gc 2, GCGGCGGTGC,154564RIKEN 3110001N18, GCGCCGCAGA, 21724Unknown, TTGCTGCCCAUnknown, GCCTTCAGTGtumor stroma, TAGACCAAGC, 116687Unknown, CAAGTGACAGtransgelin 2, CTCTGGGGCT, 22632serine (or C) proteinase, AAGCTACAGT, 2044Ly6A, TATGCCTGTC, 1583TAR (HIV) RNA binding, GTGAGCTTGT, 176Unknown, GCGCCCAGCCUnknown, CCACTTTGGCpituitary tumor-transf. 1,GGAATCTGAT,6856Unknown, CACTACACGGexpr. seq. AU022351, AGGGTGAGGA, 13774RIKEN 2410003F05, CATCCGATCG, 60230CD137 (4-1BB) alt sp2, ACTCCTGGAC,3720Unknown, GACGGAGTGG
Key to Gene expression chart:Abundantly expressed gene (>1%)Abundantly expressed gene (>0.3%)Moderately expressed gene (>0.1%)Significant expression (≥7 tags)
Positive expression (≥3 tags)Positive (not significant <3 tags)Negative (undetected or no tags)Negative (but library <10 000 tags)
Fig. 4. Cluster analysis of serial analysis of gene expression (SAGE)
libraries. A representative dendrogram is shown (A) illustrating therelationships between different SAGE libraries. Pairwise similaritycomparisons were here calculated using the 4000 most abundant tags(excluding ribosomal proteins and artefactual tags) and a root meansquared distance method for logarithmically transformed data with a five-
fold weighting to points with >95% confidence of >1.5-fold change.Clusters of cell types are highlighted and charts that depict expressionpatterns of gene tags most restricted to the Tr1-like Treg (iv) and theCD4þCD25þ (vii) clusters are shown (B), with colors relating to tagfrequency as indicated. Each tag is identified by a description (may betruncated), the 10 bp tag sequence and Unigene cluster number.
Cobbold et al �Regulatory T cells and APCs in tolerance
Immunological Reviews 196/2003 117
were components of the TCR: CD3g, CD3d, TCRa, TCRb, andCD4 (all but one of the T-cell libraries derived from CD4þ
cells). Similarly, the only transcripts unique to B cells were
those related to the Ig receptor (as above), and some Ig genes
were even expressed in Th2 cells, probably germline
transcripts driven from the enhancer in response to Th2 cyto-
kines (59). A pairwise comparison of different T cells before
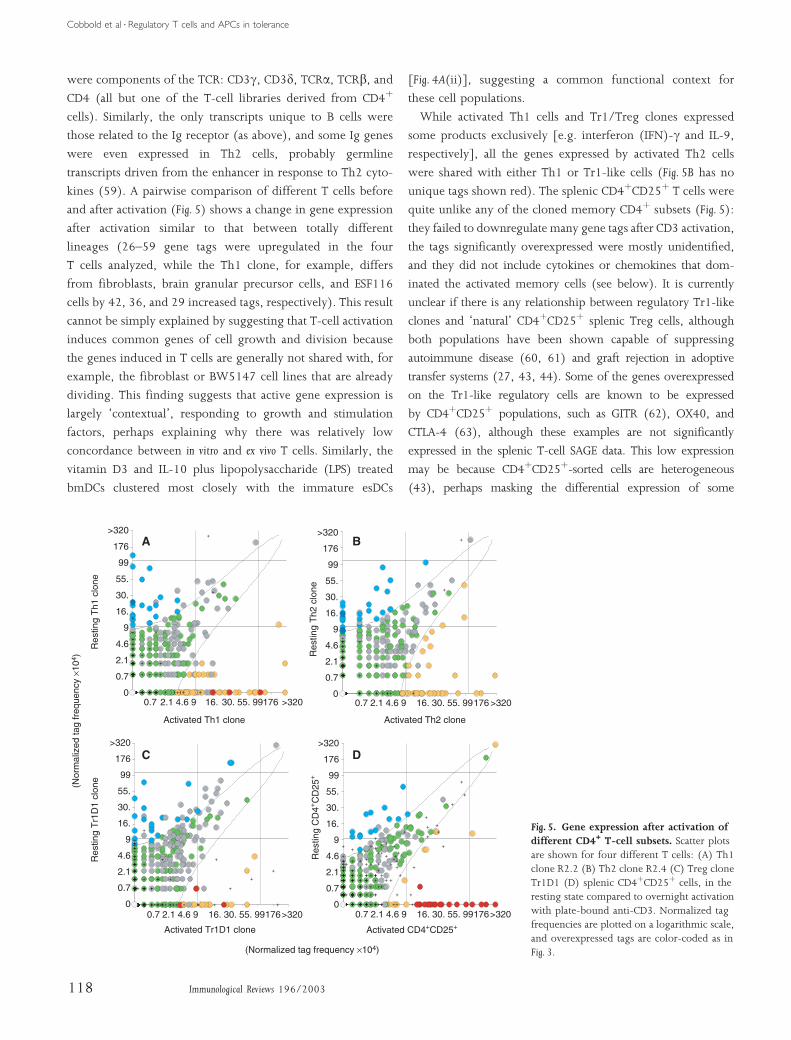
and after activation (Fig. 5) shows a change in gene expression
after activation similar to that between totally different
lineages (26–59 gene tags were upregulated in the four
T cells analyzed, while the Th1 clone, for example, differs
from fibroblasts, brain granular precursor cells, and ESF116
cells by 42, 36, and 29 increased tags, respectively). This result
cannot be simply explained by suggesting that T-cell activation
induces common genes of cell growth and division because
the genes induced in T cells are generally not shared with, for
example, the fibroblast or BW5147 cell lines that are already
dividing. This finding suggests that active gene expression is
largely ‘contextual’, responding to growth and stimulation
factors, perhaps explaining why there was relatively low
concordance between in vitro and ex vivo T cells. Similarly, the
vitamin D3 and IL-10 plus lipopolysaccharide (LPS) treated
bmDCs clustered most closely with the immature esDCs
[Fig. 4A(ii)], suggesting a common functional context for
these cell populations.
While activated Th1 cells and Tr1/Treg clones expressed
some products exclusively [e.g. interferon (IFN)-g and IL-9,
respectively], all the genes expressed by activated Th2 cells
were shared with either Th1 or Tr1-like cells (Fig. 5B has no
unique tags shown red). The splenic CD4þCD25þ T cells were
quite unlike any of the cloned memory CD4þ subsets (Fig. 5):
they failed to downregulate many gene tags after CD3 activation,
the tags significantly overexpressed were mostly unidentified,
and they did not include cytokines or chemokines that dom-
inated the activated memory cells (see below). It is currently
unclear if there is any relationship between regulatory Tr1-like
clones and ‘natural’ CD4þCD25þ splenic Treg cells, although
both populations have been shown capable of suppressing
autoimmune disease (60, 61) and graft rejection in adoptive
transfer systems (27, 43, 44). Some of the genes overexpressed
on the Tr1-like regulatory cells are known to be expressed
by CD4þCD25þ populations, such as GITR (62), OX40, and
CTLA-4 (63), although these examples are not significantly
expressed in the splenic T-cell SAGE data. This low expression
may be because CD4þCD25þ-sorted cells are heterogeneous
(43), perhaps masking the differential expression of some
Activated Th1 clone
0
0.7
2.1
4.6
9
16.
30.
55.
99
176
>320
0.7 2.1 4.6 9 16. 30. 55. 99176 >320
+
+
+++
+
++ +
+
+++ ++ ++++ +
+
++++ +++++ +++++++++
+
++ ++++
+
+++
+
+
+
+++++
+
+
+++++++++
+
++ ++++
+
++++++++++++++++++++++++++++++++++++
+
+++
+
++++++++++
+
+++
+
++++++++++++ ++++++ ++++++
+
+++++ +++++++
+
+++++++++
+
+++++++++++++++++++++++
+
++++++++++++ +++++++++++++++++++++++++++++++++++++++++++++++++++
+
+
+
++++++ ++++++++ +++++
+
++++++
+
++++++
+
+++
+
+++++++++++++++++++++++++++++++++++
+
++++++++
+
+++++++++++++++++++++++++++++++++++++
+
++
+
+++
+
+++++++
+
+++++++++
+
++
+
++++++++++++++++++ +++++++++++++++++++++++++++++++
+
++++++++++++
+
+++++++ ++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++ ++++++
+
++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++ +++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++
+
+++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++ +++++
+
++++++
+
++++++++
+
++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++
+
+++
+
+++++++++++++++++++++++++++
+
+
+
++++++++++++++++
+
+++++++++++++++
+
++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++ +++++++++++++++++++++++++++++
+
+
+
+++++++++++++++++ ++
+
++++++++++++++++++++++++++++++++++
+
++++++ ++++++
+
+++++++++++++++++++++++++++++++++++++++++++
+
+++
+
+++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++
+
++++++++++
++
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++
+
+++++++++++
+
++++++
+
+++++++++++++++
+
++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
+++++++++ +++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++ ++++++++++++++++++++++++++++++
+
++++++++
+
+++++++++++++++++++++++++++++++ ++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++
+
+++++
+
++++
+
+++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++
+
++
+
+++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++ ++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++
++
++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++
+
+++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++
+
+++++++++++++++++++++++++++
+
+++++++++++++++++++++++++
+
+++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++
+
+++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
Res
ting
Th1
clo
ne
0
0.7
2.1
4.6
9
16.
30.
55.
99
176
>320
0.7 2.1 4.6 9 16. 30. 55. 99176 >320
+
+
+ ++
+
+ + +
+
+++ +++++++
+
+++ +++++++++++ ++++ +++++++
+
+
+
+
+
++++++
+
+++++
+
+++++
+
+
+++++
+
++++++++++++++++++++++++++++++++++++++++
+
+
+
+++
+
++++
+
+++
+
+++
+
+++++++++++++++
+
+++++++++++++++++
+
+++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++
+
++++++
+
++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++
++
+
+
++++++++
+
++++++++
+
+
+
+++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++
+
+++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++
+
++++
+
++++++
+
++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+
+++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++
+
++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++
+
+++++++++++
+
++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++
+
+++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++
+
++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++
+
+++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++
+
+++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++
+
+++++++++++++++++++++++++++++++++++++++
++
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++
+
+++++++++++++
+
++++++
+
+
+
++++++++++++++++++++++++++++
+
++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++
++
+++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++
+
++++
+
++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++
+
++++++++++++++++
+
+++++++++++++++++++
+
+++
+
+++++++
+
++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++
+
++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++
+
++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++
+
++++++++++++++++++++++++++++++++++
Activated Th2 clone
Res
ting
Th2
clo
ne
0
0.7
2.1
4.6
9
16.
30.
55.
99
176
>320
0.7 2.1 4.6 9 16. 30. 55. 99176>320
+
+
+ +
+
+
+
+
+
++ ++
+
+
++ +
+
+
+
+++
+
++
+
+
+
+
+
+
+
+++++++
+
+
+
+++
+
+
+
+
+
++++++++++++
+
+++ +
+
+
+
+
+
+
+
+++++ +++++++
+
++++++++ +++++++++
+
++++++++++
+
+++++
++
++++
+
+
+
+
+
++++++++
+
+++++++++
+
++++++++
+
+++++++++++
+
++++++++++++++++++++
+
++++++
+
++
+
+
+++++++++
+
+++
+
+
+++
+
++++++++++++
+
+++++
+
+++
+
++++++
+
+++++++++++++++++++
+
+++++++++++++++
+
++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
+++++
+
+++++++++++++++
+
+++
+
+++++++++
+
+++++++++++++++++++++++++++++++++++++++++++
+
+++++++ ++++++++++++++
+
++++++++++ +++++++++++++++++++++++++++++
+
+++++++
+
+++++
+
+++++++++++++++++++++++++++++++++++++++++++++
+++
+++++
+
++++
+
++
+
+++++++++++++++++++++++++++++++++
+
+++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++
+
+
++
+
+++++++++++++++++++++ ++++++++++
+
+++++++++++++++++++++++++++++
+
+++++++++++
+
+++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ +++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++
+
++++++
+
++++++++++++++++++++++
+
+++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++
+
+++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++
+
+++
+
++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++
+
+++++
+
++++++++++++++++++++++++++
+
+++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++
+
++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++ ++++++++++++++++++++++++++
++
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++
+
++++++++++++++++++++++++
+
++++++++
+
++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++
+
++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ +++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ +++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++ +++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++ ++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
Activated Tr1D1 clone
Res
ting
Tr1
D1
clon
e
0
0.7
2.1
4.6
9
16.
30.
55.
99
176
>320
0.7 2.1 4.6 9 16. 30. 55. 99176>320
+
+
+
+
+
+
+
+
+
+
+
+
+++
+
+
+
+ +
+
++ +
+
+
+
+
++
++ +
+
+
+
+++++++++
+
++
+
+
+
+++
+
+
+++
++
+++
+
++
+
+++++++++ ++++++++++++++
+
++++ ++++
+
+++
++
+++++++
+
+++++++++++++++++++
+
+
+++++
+
+
+
++++
+
++++++
+
++++++
+
+
++
+
+
+
+
+++++
+
+
+
+++++
+
++
+
+++++
+
+++++++
+
++
+
+++++
+
+
+
+
+
+
++++++++
+
+++++
+
++
+
++++
+
+++
+
+++++++++
+
+++
+
++++++
+
+
+
++++++++++++++++++
+
++++++
+
+++++++++++++++
+
+++++++++++++++++++++++++++++
+
+++++++++ +++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++
+
++++++
+
++++++++++++++++++++++++
+
++++++++++++
+
+++++++++++++++++++++
+
+++++++
+
++++++++++++++
+
++++++++++++++++++++++++++++++
+
+
+++++
+
+++++
+
+
+
+++++++
+
+++++++++
+
+++
+
++++++++++++++
+
+++
+
+
+
+++++
+
++++++
+
++++++++++++++++++++ ++++
+
++++++++++++
++
+++++++
+
+++++++++++
+
++++++++++++++
+
+++++++++++++
+
+++++++
+
+++++++++++++++++
+
+
+
+++++
+
+++++
+
+
+
+
++++
+
++
+
+++++++++++++++++++++++++
+
+++++++++
+
+++++++++++++++
+
+++++
+
+
+
+++++++
+
++++++++++++
+
+
+++
+
+
++
++
++
++
++++++++++++++++
+
+
+
++++++
+
+++++
+
+++++++++++++++++++++++++++++++++++++++++
+
++++
+
+++++++++
+
+++++++++++++++++++++++
+
+++++++++
+
+++++++++++++
+
+
+
++++++++++++++
+
++++++++
+
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++
+
++++++++++
+
+++++++++++++++++++
+
+
+
+++++++++++++++++++++
+
++++++
+
++
+
++++++++++++
+
++++++++++++++++++
+
++++
+
++
+
+++++++++++++++++++++++++
+
+
+
+++++++++++
+
+++++
+
++++++++++++
+
+++++++++++++++++++++++
+
+
+
+++
+
++++++++++++++++++++++++
+
++++
+
++
+
++++++++++++
+
++++++++++++++
+
+++++++++++++++++++++++++++
+
++++++++++++++++++++
+
++
+
++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++
+
+++
+
+++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++
+
++
+
++++++
+
+++++++++++
+
++++++++
+
++++++++++++
+
+++++++
++
+++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++
+
+
++++
+
+++++
+
+++
+
++++++
+
+++++
+
+++++++++++
+
+
+
+
++
+
++++
+
+++++++ +++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++
+
+++++++
+
+
+
+++++++++++++++++++++++
+
++++++++++++++++++++++++++++++
+
++++++++++
+
+++++++++++++++++
+
+++++
+
++
++
+++++++++
+
++++++++++
+
+++++
+
+++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++
+
++++++
+
+++
+
+++++++
+
+++++++
+
+++++++++++++++++++
+
++++++++++++
+
+++++
+
+++++++++
+
++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++
+
++
+
+
++
++++++++++++++++++++
+
++++++++
+
+
+++++
+
++++++++++++++++
+
++++++++++++++++
+
++++++
+
+++++++
+
++++++++++++++++
+
+
+
+
+++++++++++++++++++++++++++++++++++++++++++++
+
+
+
+++++++++++++
+
++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++
+
+++++++++++
+
++++++++++++++++++++++++++
+
++++++
+
+++++++
+
++++++++++++++
+
++
+
+++++++++++
+
++++++++++++++++
+
+
+
++++++++++ +++++++
+
+++++++++
+
+++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++
+
++++++
+
++
++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ ++
+
+++
+
++++++++++
+
+++++++++++++++++++++++++++++++++++++++
+
+++++++
+
+
+
+++++++++++++++++
+
+++++
+
+++++++++++++++
+
++++++++++++++++
+
++++++
+
++++++++++++++++++++++++++
+
+++++++++++
+
+
+
+++
+
+++++++++++++++++++++++++++++++++++++++++++++
+
++++
+
++
+
+++++++
+
+++++++++++++
+
+++++++++
+
++++++++++++++++++++++++++++++++++++++++
+
+++
+
++++++
+
++++++++++++++++++++++++++
+
+++
+
++++++
+
+
++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++
+
+
+
+
++++++++++++++++++++++++
+
+++++++++++++++++++++++++
+
+++++
+
+++++++++++++
+
+
+++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++
+
+
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++
+
+
++++++++++++++++++++++++++++++++++++
+
+++++
+
++++++++++++++
+
++++++++++++
+
++++++++++++++
+
+
+
++++++
+
++
+
++ ++++++++++++++++++++++++++++++
+
+++++++++
+
++++++++
+
++++++++++
+
+
+++++
+
++++++++++++++++++++++++++++++++++
+
++++++++++++++
+
+++++++++
+
++++++ ++++++++++++++++
++
+
+
+++
++
+
++
+
+++++++++++++++++++ ++++++++++++++
+
++++++++++++
+
+++++
+
++++++++++++++++++++++++
+
++++++++++++++++++++
+
+
+
++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++
+
++++++++++
+
++++++++++++
++
+++++++++
+
+++++++++++
+
+++++++
+
++++++++++++++++++++++++++++
+
++++++
+
++++++++++++
+
++++++
+
+
+
+++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++
+
+++++
+
++++
+
++++ ++++
+
+++
+
+++
+
++++++
+
+++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++
+
+++++++++++++++++++
+
+
++++++++++
+
++
+
++
+
++++++++++++++++++
+
+++++++++++++++
+
++++++
+
+++++++++++
+
+
+
+++++++++++++++
+
+++++++++++++++++++++
+
+++++++++++++
+
+++++++
+
++++++++++++++++
+
++++++++++++
+
++++++++++
+
+
+
+
+
++
+
+++++
+
+
+
+++++++++++++++++++++++++++++
+
+
+
+
+
++
+++++++++
+
+++
+
++
+
+++++
+
++++++++++
+
+++++
+
++++++++++++++
+
++++++
+
+++++++++
+
++++++++++++++
+
+++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++
+
++++++++++
+
++++++++
++
++
+
+++++++
+
++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++
+
+++++++++++++
+
+++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++
+
++++++++++++++++++++++++++++++++
++
++++
+
++
+++++++++++
+
+++++++++++++
+
+
+
++
+
+++++
+
+
+
+
+++++++++++++++++++
+
++++++++++++++++ +++++
+
+
+
++++++++++++++++++++++++++++++
+
++++
+
++++++++
+
+++++++
+
++++++++++++++++
+
+++++
+
++++++++++++++++++++++++++++
+
+++++
+
+++++
+
++++
+
+
+
+++++++++++
+
+++
+
+++++++++++++++++++++++++
+
++++++++++++++++++ ++++++++++++++++++++++
++
++++++++++++++++++
+
+++++++++++++++++++++++ +++++++++
+
+++++++++++++++++++++++++++
+
++++++++++++
+
+++++++++++++++++
+
+++++++++++++++++++++++++++++
++
+++
+
+
++++++++++++++++++++
++
+++++++++++
+
+++++++++++++++++++++
+
++++++++++++++
+
+++++
+
+++++++++
+
++++++
+
++++
+
++++
+
+++++++++++++++++++
+
++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++
+
+++++++
+
++++++++++++++++
+
++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++
+
++++
+
+++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++
+
+++++++
Activated CD4+CD25+
Res
ting
CD
4+C
D25
+
(Nor
mal
ized
tag
freq
uenc
y ×1
04)
(Normalized tag frequency ×104)
A B
DC
Fig. 5. Gene expression after activation ofdifferent CD4+ T-cell subsets. Scatter plotsare shown for four different T cells: (A) Th1clone R2.2 (B) Th2 clone R2.4 (C) Treg cloneTr1D1 (D) splenic CD4þCD25þ cells, in theresting state compared to overnight activationwith plate-bound anti-CD3. Normalized tagfrequencies are plotted on a logarithmic scale,and overexpressed tags are color-coded as inFig. 3.
Cobbold et al �Regulatory T cells and APCs in tolerance
118 Immunological Reviews 196/2003

Treg-associated markers. For example, the tag for FoxP3
(CCTAAGCGTG), a recently described transcription factor
associated with CD25þ Treg cells (64) appeared only once in
this library. We are now in the process of generating SAGE
libraries from more highly purified CD4þCD25þ subsets,
although the phenotype of the most active Treg cell within
this population remains unclear (63, 65).
Gene transcripts that distinguish Th2 and Tr1-like Treg clones
Resting Tr1-like Treg clones have a similar pattern of gene
expression to Th2 cells, but they overexpress transcripts such as
ppENK and Granzyme A with a reduction in the transcription
factors GATA-3 and early growth response gene-1 (Egr-1) (43).
GATA-3 promotes transcription of a number of genes involved in
Th2 effector function (66, 67), such as IL-4, IL-13 and ST2L, and is
itself regulated by the repressor of GATA (ROG) (68). The tags for
these two genes in the SAGE data suggested that GATA-3 was
highly expressed in resting Th2 cells, while ROG was expressed
in the Tr1-like cells. Quantitative reverse transcriptase-polymerase
chain reaction (RT-PCR) confirmed these observations (Fig. 6A),
particularly after CD3 activation, suggesting that ROG may be a
useful marker for Tr1-like regulatory cells. ROG may act in these
cells to repress effector and cytokine function in a manner
analogous to FoxP3 in CD25þ Treg cells (64), as the latter is not
expressed in Tr1-like cells by quantitative RT-PCR (data not
shown).
A gene previously found in both Tr1-like and CD4þCD25þ
cells is the regulator of G protein signaling, RGS-1 (69). Two
different tags mapping to this gene were found in the SAGE
data, one derived from the 30 UTR and the other from the
beginning of the first exon (data not shown). Alternative
polyadenylation signals were found within the third and
fourth introns, which could generate truncated transcripts,
and the presence of these, particularly in the Tr1-like cell
mRNA, was confirmed by specific RT-PCR (Fig. 6B). The trun-
cated forms of RGS-1 no longer encode the regulatory domain
but retain the N-terminal region interacting with target G
proteins and may compete with the full length protein (70).
Regulatory T cells expressing excess truncated isoforms might
be hyper-responsive to certain chemokines.
Abundant transcripts with differential expression across the
immune system
Overall, cytokines, chemokines, and their receptors were the
most abundant and differentially expressed transcripts. A
summary of their expression is shown Fig. 7. Th2 cells have
almost no unique transcripts; they are nearly all shared either
by Th1 or Tr1-like Treg cells. While Th1 cells overexpress a
number of chemokine ligands and receptors, especially after
activation, Tr1-like Treg cells mostly expressed transcripts in
common with Th2 cells, as previously published (43), with
the exception of IL-9 (confirmed by enzyme-linked immuno-
sorbent assay, data not shown). IL-9 is known to be regulated
independently of the other Th2 cytokines (71) and may play a
role in the previously observed association of Tr1-like clones
and mast cells (44). CCR1 and CCR5 (expressed on unstimu-
lated and IL-10-treated bmDCs; Fig. 7) mediate recruitment of
immature DCs to inflamed sites, while a switch to CCR7
expression directs homing to lymphoid organs, where
CCL17 and CCL21 serve to promote DC–T-cell interactions
(as seen in LPS-treated DCs; Fig. 7). Also of note is the receptor
for programmed cell death (PD-1) on both Th2 and Treg
clones, while the PD-1 ligand (PD-1L) is expressed on both
mature and IL-10-treated DCs, where it has been implicated in
the regulation of T-cell responses (72).
IL-10-treated dendritic cells are modulated rather than blocked
from maturing
While mature DCs drive naıve T cells to immunity, immature
DCs have been implicated in mediating tolerance (7). In
addition, IL-10 (10) and vitamin D3 (45) are reported to
modulate DCs in favor of promoting tolerance, either by
inhibiting normal maturation (73) or by generating an alter-
native activation state (74). LPS-induced maturation of bmDCs
induced significant changes in 73 gene tags (Fig. 8A). In
response to IL-10 treatment (Fig. 8B), 31 of these same changes
were seen, suggesting IL-10 itself induces a partial maturation,
while 102 other tags were modulated. After LPS treatment of
these cells, 41 of the 73 changes associated with maturation
were still present, while 71 additional genes were different
from untreated bmDCs (data not shown). In general, the SAGE
data support the hypothesis that IL-10 limits inflammation
whilst enhancing phagocytosis (75) and antigen proces-
sing (cathepsins B, C, D, S, and L, lysozyme, CD16, CD64,
complement components C1q all increase, while MHC class
II a/b remain low). IL-10 treatment alone mediated
expression of CXCL7 (Fig. 7), the precursor forms of which
can be anti-inflammatory (76).
Cluster analysis revealed a signature of genes coexpressed in
IL-10-treated (�LPS) and vitamin D3-treated bmDCs and the
esDCs (Fig. 8C). The signature included anti-inflammatory
genes, such as IL-1R antagonist and hemoxygenase-1 (HO-1),
which are also implicated in the suppression of graft rejection
Cobbold et al �Regulatory T cells and APCs in tolerance
Immunological Reviews 196/2003 119
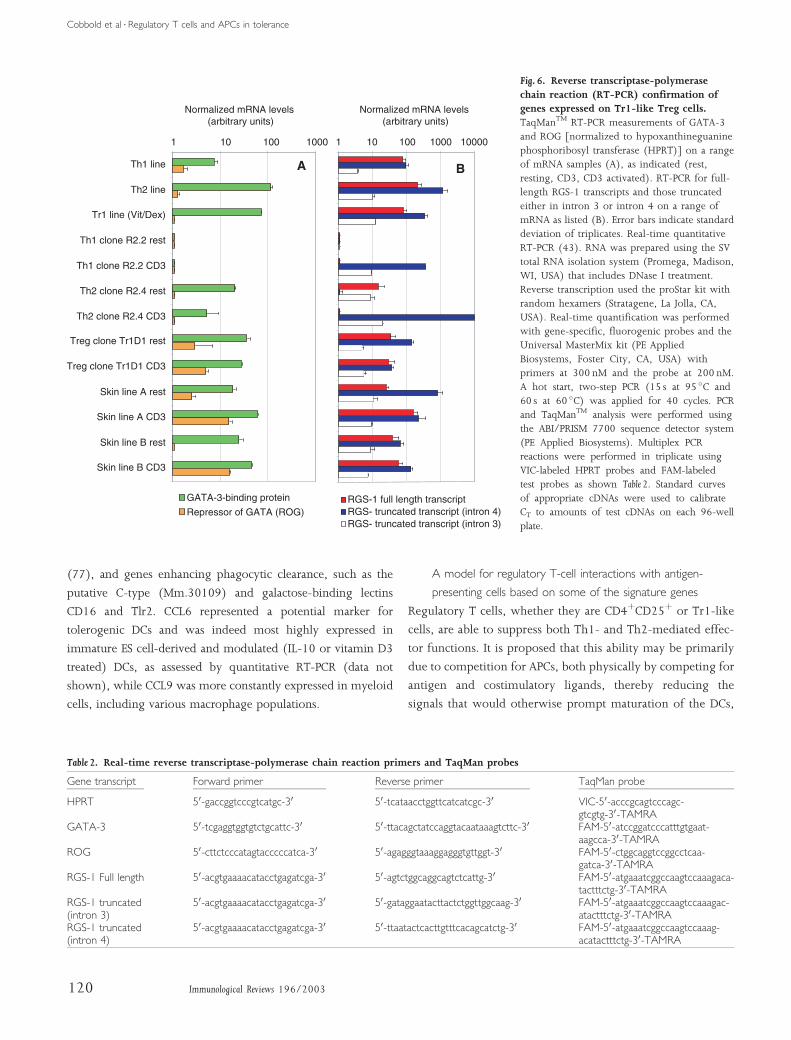
(77), and genes enhancing phagocytic clearance, such as the
putative C-type (Mm.30109) and galactose-binding lectins
CD16 and Tlr2. CCL6 represented a potential marker for
tolerogenic DCs and was indeed most highly expressed in
immature ES cell-derived and modulated (IL-10 or vitamin D3
treated) DCs, as assessed by quantitative RT-PCR (data not
shown), while CCL9 was more constantly expressed in myeloid
cells, including various macrophage populations.
A model for regulatory T-cell interactions with antigen-
presenting cells based on some of the signature genes
Regulatory T cells, whether they are CD4þCD25þ or Tr1-like
cells, are able to suppress both Th1- and Th2-mediated effec-
tor functions. It is proposed that this ability may be primarily
due to competition for APCs, both physically by competing for
antigen and costimulatory ligands, thereby reducing the
signals that would otherwise prompt maturation of the DCs,
RGS-1 full length transcriptRGS- truncated transcript (intron 4)RGS- truncated transcript (intron 3)
1 10 100 1000 10000
Treg clone Tr1D1 rest
Treg clone Tr1D1 CD3
Skin line A rest
Skin line A CD3
Skin line B rest
Skin line B CD3
Th1 line
Th2 line
Tr1 line (Vit/Dex)
Th1 clone R2.2 rest
Th1 clone R2.2 CD3
Th2 clone R2.4 rest
Th2 clone R2.4 CD3
Normalized mRNA levels (arbitrary units)
Normalized mRNA levels (arbitrary units)
1 10 100 1000
GATA-3-binding protein
Repressor of GATA (ROG)
A B
Fig. 6. Reverse transcriptase-polymerase
chain reaction (RT-PCR) confirmation ofgenes expressed on Tr1-like Treg cells.
TaqManTM RT-PCR measurements of GATA-3and ROG [normalized to hypoxanthineguaninephosphoribosyl transferase (HPRT)] on a rangeof mRNA samples (A), as indicated (rest,resting, CD3, CD3 activated). RT-PCR for full-length RGS-1 transcripts and those truncatedeither in intron 3 or intron 4 on a range ofmRNA as listed (B). Error bars indicate standarddeviation of triplicates. Real-time quantitativeRT-PCR (43). RNA was prepared using the SVtotal RNA isolation system (Promega, Madison,WI, USA) that includes DNase I treatment.Reverse transcription used the proStar kit withrandom hexamers (Stratagene, La Jolla, CA,USA). Real-time quantification was performedwith gene-specific, fluorogenic probes and theUniversal MasterMix kit (PE AppliedBiosystems, Foster City, CA, USA) withprimers at 300 nM and the probe at 200 nM.A hot start, two-step PCR (15 s at 95 �C and60 s at 60 �C) was applied for 40 cycles. PCRand TaqManTM analysis were performed usingthe ABI/PRISM 7700 sequence detector system(PE Applied Biosystems). Multiplex PCRreactions were performed in triplicate usingVIC-labeled HPRT probes and FAM-labeledtest probes as shown Table 2. Standard curvesof appropriate cDNAs were used to calibrateCT to amounts of test cDNAs on each 96-wellplate.
Table 2. Real-time reverse transcriptase-polymerase chain reaction primers and TaqMan probes
Gene transcript Forward primer Reverse primer TaqMan probe
HPRT 50-gaccggtcccgtcatgc-30 50-tcataacctggttcatcatcgc-30 VIC-50-acccgcagtcccagc-gtcgtg-30-TAMRA
GATA-3 50-tcgaggtggtgtctgcattc-30 50-ttacagctatccaggtacaataaagtcttc-30 FAM-50-atccggatcccatttgtgaat-aagcca-30-TAMRA
ROG 50-cttctcccatagtacccccatca-30 50-agagggtaaaggagggtgttggt-30 FAM-50-ctggcaggtccggcctcaa-gatca-30-TAMRA
RGS-1 Full length 50-acgtgaaaacatacctgagatcga-30 50-agtctggcaggcagtctcattg-30 FAM-50-atgaaatcggccaagtccaaagaca-tactttctg-30-TAMRA
RGS-1 truncated(intron 3)
50-acgtgaaaacatacctgagatcga-30 50-gataggaatacttactctggttggcaag-30 FAM-50-atgaaatcggccaagtccaaagac-atactttctg-30-TAMRA
RGS-1 truncated(intron 4)
50-acgtgaaaacatacctgagatcga-30 50-ttaatactcacttgtttcacagcatctg-30 FAM-50-atgaaatcggccaagtccaaag-acatactttctg-30-TAMRA
Cobbold et al �Regulatory T cells and APCs in tolerance
120 Immunological Reviews 196/2003

but also by modulating the APCs to express protective and
anti-inflammatory genes such as HO-1, IL-1 receptor antag-
onist (IL1RA), the chemokine CCL6, and the PD-1L. The SAGE
data suggest a close relationship between Th2 effector cells and
Tr1-like regulatory cells, although the latter seem to lack
effector functions driven by GATA-3, possibly as a conse-
quence of expressing the ROG and a truncated form of RGS-
1 that may modify chemokine responsiveness. CD4þCD25þ
regulatory cells may have an analogous relationship to Th1
effector cells, with FoxP3 acting to repress cytokine produc-
tion and responsiveness with the loss of the signal transducer
and activator of transcription (STAT) molecules, STAT4 and
STAT6. On the assumption that APCs are required to drive
immune responses through to Th1 and Th2 effector cells to
Th1
Th2
Upregulated after CD3 stimulation
Downregulated after CD3 stimulation
CCR2
IFN-γ
CCL5CXCR3
IL-25IL-2RγTRAF2IL-15RaCCL4
Granzyme B IL-9
CD25–
CD4+
CD25+
CD4+IL-17
IL-6Rα
CCL1
CCL3,4
CD134
GITR
Treg
Tr1-like
Shared with T-cell subset(s)
DC (unstimulated)
DC + IL-10
CCL17
CCL5
CCL21b
CCR7
IL-12p40
IL-1β
CCL9 CCR1,5HO-1 CCL6
Lymphotoxin B
CCL21a TNFRSF6 IL-1RII
CXCL7
B cells
IL-10R α
IL-3Rα
CD137
PD-1LCCL22
Lymphotoxin B
DC + LPS
CXCR6 IL-4RαPD-1 IL-1RII
CD137IL-4,5
Granzyme A
Fig. 7. The expression of abundant gene familieson T cells and antigen-presenting cells. Venndiagrams of gene expression according to serialanalysis of gene expression tag abundance forchemokines, cytokines, and their receptors, andother transcripts of note. All transcripts shown arestatistically differential (i.e. three or more tags intotal or a difference of seven or more tags betweenlibraries).
A
B
Isocitrate dehydrogenase 1,GGCAATAATG,9925Expressed seq. C88302,TTTATGGAAT,23968TGF beta-induced 68 kDa,GTGCATTTGT,14455Sterol O-acyltransferase 1,CCTGGCCAAG,28099Ninjurin 1,CTGGTACTTC,18503Aminolevulinic acid synth 1,AGCAAGATGG,19143Heme oxygenase 1,GCCACTTTGA,17980Liver glycogen phosph.,GAGCCTTCGG,30047α-N-acetylglucosaminidase,GCTGAGCTGG,6142C-type lectin???Mm.30109,AAAGCTGCAA,30109Hexosaminidase A,GTACGCCTGA,2284CCL 6 long form,TAGCCACAAA,137RIKEN 2700038C09,ATAGAAGAGA,182914Neutr. cytosolic factor 2,TGAATGAAGA,10729ESTs,CTGGACATCC,140215NIMA (never in mitosis),GACTATGCGA,143818CCL 6,TAGCCAAAAA,137CSF-1 receptor,AGGGGTCTGG,22574CCAAT binding protein,GCGGCCGGTT,4863ESTs,GGAAGGTGGG,29046Interleukin-1 RA,CTATGTATTG,882Lectin; galactose binding,CTGAGAGAAA,2970CD 16 (low affinity FcR),TATATTTTTA,22119CD107b LAMP-2,ACCATTATAA,486Expressed seq. AI840826,GCTCTGGCCT,38488RIKEN 1110012E06,CTGGACACTG,41331Syntaxin binding 2,TCGGAAATGA,7247Expressed seq. AI132321,GGAAGACAGC,203915Legumain,GACAAAGTGT,17185Toll-like receptor2,GTCCAGCAGC,87596Prog. cell death 1 ligand,GTGTTGGATT,168681
Th1
(cl
one
R2.
2)T
h2 (
clon
e R
2.4)
Tr1
D1
(inc.
mas
t)A
1 ×
RA
G–/
– nod
esC
BA
nod
esC
D4+
CD
25+ (
CD
3)C
D4+
CD
25+
CD
4+C
D25
–
CD
4+C
D25
– (C
D3)
Tr1
D1
clon
e (C
D4)
Ski
n tr
eg li
ne A
S
kin
treg
line
B
Tr1
D1
clon
e (C
D3)
Th2
clo
ne (
CD
3)T
h1 c
lone
(C
D3)
BW
5147
thym
oma
ES
F-1
16 (
ES
cel
ls)
ES
DC
ES
DC
+ L
PS
BM
DC
+ L
PS
BM
DC
BM
DC
+ IL
-10
BM
DC
+ IL
-10
+ L
PS
BM
DC
+ V
itD3
B c
ells
3T3
fibro
blas
tR
1 (E
S c
ells
)B
rain
(P
GC
P)
IEL
(α/β
CD
8+)
IL-10 treated bmDC
00.72.14.6
916.30.55.99
176>320
0.7 2.1 4.6 9 16. 30. 55.99 176 >320
+
+
++
+
+
+ ++
+
++ +++ +++ + +
+
++ +++ + ++
+
+++ ++++++ +++
+
+++++
+
++
+
+
+
++
+
+
++
++++++++
++
+++
+
+++
+
+
+
+++++
+
++++
+
+
+
++++
+
++++++++
+
++++++
+
++
+
+
++++
++++++
+
+++++++++++
+
++++++ ++++++
+
+++++++++++++++++
+
++++++++
+
+
+
++++++++++++
+
++++++++
+
++++++++++++++
+
++++++
+
++++++
+
+
+
++
+
++++
+
++++++++
+
+++
+
++++++++++++++++++++++++
+
++++++++
+
+++
+
++++++++++++++++++++++++++
+
+
++
+++++
+
+
+
+
+
++++++++++++++++++
+
++
+
++++++
+
++++++++++
+
+++++++++++
+
++++++++++++++++
+
+++++++++++++++++++++++
+
+++++++++++
+
+++++++++++
+
++++++++++++++
+
++++++
+
++++++++++++++++++++++++++++++++++++++++ ++++++++++
++
++++++
+
+++++++++++
+
++
+
+++++++++++++++++++++++++ ++++++
+
++++++++++++++++++++++++++++++++
+
+++
+
+++++++++++++++++
+
+++++++++++++++++++++++++++++
+
++
+
++++++
+
+++++
+
+++++++++++++++++++
+
+
+
++++++++++++++++++++++++
+
+++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++
+
++++++++++++++++++++
+
++++++++++++++
+
++++++++++++
+
+++++++++++++++++++++++
+
++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++ +++++
+
+++
+
++++++++++++++++++++++++
+
++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++
+
+++++++
+
+++++++
+
+++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++ ++++++
+
+++++
+
+++++++++++++++++
+
++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ ++++++++++++
+
++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
+++++++++++++
++
++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
+++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++
+
++++++++++++++++++
+
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++
+
++++
+
+++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++ ++++++
++
++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++
+
++++++++
+
++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++
+
+++++++++
+
+++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++
+
+++++++++
+
++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++
+
+++++++++++++++
+
++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
++
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++
+
++++++++
+
+++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++
+
++++++++++++++++++++
+
++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+
+
++++++++++++++++++++
+
+++++++++++++++
++
+++++++++++++++++++++++++++++++++++
+
++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++
Unt
reat
ed b
mD
C
(Normalized tag frequency ×104)
(Nor
mal
ized
tag
freq
uenc
y ×1
04 )
00.72.14.6
916.30.55.99
176>320
0.7 2.1 4.6 9 16. 30. 55. 99 176 >320
+
+
++
+
+
+++
+
+++++ +++++
+
++++++++
+
+++++++++ +++
+
+++++
+
++
+
+
+
++
+
+
++
++++++++
++
+++
+
+++
+
+
+
+++++
+
++++
+
+
+
++++
+
++++++++
+
++++++
+
++
+
+
++++
++++++
+
+++++++++++
+
++++++++++++
+
+++++++++++++++++
+
++++++++
+
+
+
++++++++++++
+
++++++++
+
++++++++++++++
+
++++++
+
++++++
+
+
+
++
+
++++
+
++++++++
+
+++
+
++++++++++++++++++++++++
+
++++++++
+
+++
+
++++++++++++++++++++++++++
+
+
++
+++++
+
+
+
+
+
++++++++++++++++++
+
++
+
++++++
+
++++++++++
+
+++++++++++
+
++++++++++++++++
+
+++++++++++++++++++++++
+
+++++++++++
+
+++++++++++
+
++++++++++++++
+
++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++
++
++++++
+
+++++++++++
+
++
+
+++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++
+
+++
+
+++++++++++++++++
+
+++++++++++++++++++++++++++++
+
++
+
++++++
+
+++++
+
+++++++++++++++++++
+
+
+
++++++++++++++++++++++++
+
+++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++
+
++++++++++++++++++++
+
++++++++++++++
+
++++++++++++
+
+++++++++++++++++++++++
+
++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++
+
++++++++++++++++++++++++
+
++++++
+
++++++++++++ +++++++++++++++++++++++++++++++++++++++++
+
++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++
+
+++++++
+
+++++++
+
+++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++
+
+++++
+
+++++++++++++++++
+
++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
+++++++++++++
++
++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
+++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++
+
++++++++++++++++++
+
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++
+
++++
+
+++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++
++
++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++
+
++++++++
+
++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++
+
+++++++++
+
+++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++
+
++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++
+
+++++++++
+
++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++
+
+++++++++++++++
+
++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
++
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
+++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
+
++++++++++++++++++
+
++++++++
+
+++++++++++
+
+++++++++++++++++++++++++++++++++++++++
LPS treated bmDC
Unt
reat
ed b
mD
C
C
Fig. 8. Gene expression signatures for modulated dendritic cells.
Scatter plots of untreated bmDCs compared to those treated withlipopolysaccharide (LPS) (A) or with interleukin-10 (IL-10) (B).Normalized tag frequencies are plotted on a logarithmic scale and
overexpressed tags are color-coded as in Fig. 3. The expression pattern of acluster of genes overexpressed in immature (esDC) and modulated (IL-10,IL-10þ LPS, or vitamin D3) bmDCs are shown in (C), color-coded as inFig. 4.
Cobbold et al �Regulatory T cells and APCs in tolerance
Immunological Reviews 196/2003 121

mediate graft rejection, infectious tolerance may then be
explained if the result of such competition at the APC leads
to a reduced or incomplete signal to the emerging effector cells
such that they are converted to regulatory cells. Such a
conversion can be achieved artificially by transducing FoxP3
into naıve T cells (64) that would otherwise become Th1 cells,
but it remains to be determined whether similar conversions
take place in physiological conditions (Fig. 9).
Conclusions
The two T-cell populations with known regulatory ability, the
CD4þCD25þ and Tr1-like Tregs, are apparently no more
related in their overall pattern of gene expression to each
other than they are to Th1 or Th2 cells. Both types of Treg
cells have lost some transcripts and, by implication, effector
functions associated with other memory cell phenotypes such
as Th1 or Th2 (43), but many of these functions are reduced
in all the T cells after activation. This finding may reflect
a chronic activation state of both Treg populations, which
would fit with their common expression of CD25 and
CTLA4 (43, 63). The recent discovery of a functional role
for FoxP3 that includes the repression of cytokines in CD25þ
Treg cells (64) and the finding here that ROG is expressed in
Tr1-like Treg cells suggest a mechanism whereby disabled
effector cells (32) are able to have regulatory activity. One
can speculate that CD25þ cells may compete with potential
effectors during homeostatic expansion (29) and control innate
immune responses (78), while Tr1-like cells may regulate in the
local vicinity of their specific antigen (i.e. the ‘Civil Service’
hypothesis) (6), such as in a tolerated skin graft (24).
On the APC side, two different treatments that claimed to
promote tolerance did not simply inhibit DC maturation, but
they resulted in expression of a cluster of genes associated with
the modulated phenotype. The gene expression data presented
here provides a valuable baseline to look for the expression of
novel genes associated with the physiological activation of
mixed populations of T cells and DCs in vitro or directly ex vivo
in tissues from tolerant mice. Such approaches should allow us
to identify and characterize the regulatory T cells and APCs that
maintain transplantation tolerance in the mouse models, with
a view to developing surrogate markers for tolerance in clinical
settings and new molecular targets for immunomodulatory
therapies.
Th2 cell
STAT4,6
Modulateddendritic
cell
PD1-L
HO-1
IL1RA?
CD40L
CCL6?
GATA3
ROGRGS1 ∆
Tr1-likeRegulatory
T cell
FoxP3
IFN-γ
IFN-γ
CD4+CD25+
RegulatoryT cell
IL-4
IL-4
CD28
Th1 cell
Fig. 9. A model of CD4þCD25þ and Tr1-likeimmune regulation. CD4þCD25þ and Tr1-likeregulatory T cells are shown competing with Th1and Th2 cells for the antigen-presentingdendritic cells (DCs), with the consequence thatthe DC is blocked from maturing and isadditionally modulated to produce anti-inflammatory and protective molecules. Thisgeneralized model may apply at both theinduction phase of the immune response,continuing through to the effector and memoryphases, and may take place in either the draininglymph nodes or directly within the grafted tissue.Example gene products expressed by each celltype, as determined by the SAGE analysis, areindicated.
References
1. Hall BM, Jelbart ME, Gurley KE, Dorsch SE.
Specific unresponsiveness in rats with
prolonged cardiac allograft survival after
treatment with cyclosporine. Mediation of
specific suppression by T helper/inducer
cells. J Exp Med 1985;162:1683–1694.
2. Qin S, et al. ‘‘Infectious’’ transplantation
tolerance. Science 1993;259:974–977.
3. Powrie F, Leach MW, Mauze S, Caddle LB,
Coffman RL. Phenotypically distinct subsets
of CD4þ T cells induce or protect from
chronic intestinal inflammation in C. B-17
scid mice. Int Immunol 1993;5:1461–1471.
4. Sakaguchi S, Sakaguchi N, Asano M, Itoh M,
Toda M. Immunologic self-tolerance
maintained by activated T cells expressing
IL-2 receptor alpha-chains (CD25).
Cobbold et al �Regulatory T cells and APCs in tolerance
122 Immunological Reviews 196/2003

Breakdown of a single mechanism of
self-tolerance causes various autoimmune
diseases. J Immunol 1995;155:1151–1164.
5. Groux H, et al. A CD4þ T-cell subset inhibits
antigen-specific T-cell responses and prevents
colitis. Nature 1997;389:737–742.
6. Waldmann H, Cobbold S. Regulating the
immune response to transplants. A role
for CD4þ regulatory cells? Immunity
2001;14:399–406.
7. Jonuleit H, Schmitt E, Schuler G, Knop J,
Enk AH. Induction of interleukin
10-producing, nonproliferating CD4(þ)
T cells with regulatory properties by
repetitive stimulation with allogeneic
immature human dendritic cells. J Exp Med
2000;192:1213–1222.
8. Jonuleit H, Schmitt E, Steinbrink K, Enk AH.
Dendritic cells as a tool to induce anergic
and regulatory T cells. Trends Immunol
2001;22:394–400.
9. Roncarolo MG, Levings MK, Traversari C.
Differentiation of T regulatory cells by
immature dendritic cells. J Exp Med
2001;193:F5–F9.
10. Steinbrink K, Graulich E, Kubsch S, Knop J,
Enk AH. CD4(þ) and CD8(þ) anergic T cells
induced by interleukin-10-treated human
dendritic cells display antigen-specific
suppressor activity. Blood 2002;99:
2468–2476.
11. Gregori S, Casorati M, Amuchastegui S,
Smiroldo S, Davalli AM, Adorini L.
Regulatory T cells induced by 1 alpha,
25-dihydroxyvitamin D3 and mycophenolate
mofetil treatment mediate transplantation
tolerance. J Immunol 2001;167:1945–1953.
12. Fairchild PJ, Waldmann H. Dendritic cells and
prospects for transplantation tolerance.
Curr Opin Immunol 2000;12:528–535.
13. Scully R, Qin S, Cobbold S, Waldmann H.
Mechanisms in CD4 antibody-mediated
transplantation tolerance: kinetics of
induction, antigen dependency and role
of regulatory T cells. Eur J Immunol
1994;24:2383–2392.
14. Saitovitch D, Bushell A, Mabbs DW, Morris PJ,
Wood KJ. Kinetics of induction of
transplantation tolerance with a nondepleting
anti-Cd4 monoclonal antibody and
donor-specific transfusion before
transplantation. A critical period of time is
required for development of immunological
unresponsiveness. Transplantation
1996;61:1642–1647.
15. Cobbold SP, Adams E, Marshall SE, Davies JD,
Waldmann H. Mechanisms of peripheral
tolerance and suppression induced by
monoclonal antibodies to CD4 and CD8.
Immunol Rev 1996;149:5–33.
16. Chen ZK, Cobbold SP,Waldmann H,Metcalfe S.
Amplification of natural regulatory immune
mechanisms for transplantation tolerance.
Transplantation 1996;62: 1200–1206.
17. Davies JD, Leong LY, Mellor A, Cobbold SP,
Waldmann H. T cell suppression in
transplantation tolerance through linked
recognition. J Immunol 1996;156:
3602–3607.
18. Wong W, Morris PJ, Wood KJ. Pretransplant
administration of a single donor class I major
histocompatibility complex molecule is
sufficient for the indefinite survival of fully
allogeneic cardiac allografts: evidence for
linked epitope suppression. Transplantation
1997;63:1490–1494.
19. Bemelman F, Honey K, Adams E, Cobbold S,
Waldmann H. Bone marrow transplantation
induces either clonal deletion or infectious
tolerance depending on the dose. J Immunol
1998;160:2645–2648.
20. Honey K, Cobbold SP, Waldmann H. CD40
ligand blockade induces CD4þ T cell
tolerance and linked suppression. J Immunol
1999;163:4805–4810.
21. Cobbold S, Waldmann H. Infectious
tolerance. Curr Opin Immunol
1998;10:518–524.
22. Onodera K, Lehmann M, Akalin E, Volk HD,
Sayegh MH, Kupiec-Weglinski JW. Induction
of ‘‘infectious’’ tolerance to MHC-
incompatible cardiac allografts in CD4
monoclonal antibody-treated sensitized rat
recipients. J Immunol 1996;157:1944–1950.
23. WiseMP, Bemelman F, Cobbold SP,Waldmann
H. Linked suppression of skin graft rejection
can operate through indirect recognition.
J Immunol 1998;161:5813–5816.
24. Graca L, Cobbold SP, Waldmann H.
Identification of regulatory T cells in tolerated
allografts. J Exp Med 2002;195:1641–1646.
25. Zhai Y, Kupiec-Weglinski JW. What is the
role of regulatory T cells in transplantation
tolerance? Curr Opin Immunol
1999;11:497–503.
26. Kingsley CI, Karim M, Bushell AR, Wood KJ.
CD25þCD4þ regulatory T cells prevent graft
rejection. CTLA-4– and IL-10-dependent
immunoregulation of alloresponses.
J Immunol 2002;168:1080–1086.
27. Graca L, Thompson S, Lin CY, Adams E,
Cobbold SP, Waldmann H. Both CD4(þ)
CD25(þ) and CD4(þ) CD25(–) regulatory
cells mediate dominant transplantation
tolerance. J Immunol 2002;168:5558–5565.
28. Wood KJ, Sakaguchi S. Regulatory T cells in
transplantation tolerance. Nat Rev Immunol
2003;3:199–210.
29. Barthlott T, Kassiotis G, Stockinger B. T Cell
regulation as a side effect of homeostasis and
competition. J Exp Med 2003;197:451–460.
30. Marshall SE, Cobbold SP, Davies JD,MartinGM,
Phillips JM, Waldmann H. Tolerance and
suppression in a primed immune system.
Transplantation 1996;62:1614–1621.
31. Zelenika D, et al. Rejection of H-Y disparate
skin grafts by monospecific CD4þ Th1 and
Th2 cells: no requirement for CD8þ T cells or
B cells. J Immunol 1998;161:1868–1874.
32. Lin CY, Graca L, Cobbold SP, Waldmann H.
Dominant transplantation tolerance impairs
CD8þ T cell function but not expansion.
Nat Immunol 2002;3:1208–1213.
33. Curtsinger JM, Lins DC, Mescher MF. Signal 3
determines tolerance versus full activation of
naıve CD8 T cells: dissociating proliferation
and development of effector function. J Exp
Med 2003;197:1141–1151.
34. Takahashi T, et al. Immunologic self-
tolerance maintained by CD25(þ)CD4(þ)
regulatory T cells constitutively expressing
cytotoxic T lymphocyte-associated antigen 4.
J Exp Med 2000;192:303–310.
35. Zeller JC, et al. Induction of CD4þ T cell
alloantigen-specific hyporesponsiveness by
IL-10 and TGF-beta. J Immunol
1999;163:3684–3691.
36. Waldmann H. Reprogramming the immune
system. Immunol Rev 2002;185:227–235.
37. Gallucci S, Matzinger P. Danger signals: SOS
to the immune system. Curr Opin Immunol
2001;13:114–119.
38. Qin SX, Cobbold S, Benjamin R, Waldmann H.
Induction of classical transplantation tolerance
in the adult. J Exp Med 1989;169:779–794.
39. Qin SX, et al. Induction of tolerance in
peripheral T cells with monoclonal
antibodies. Eur J Immunol 1990;20:
2737–2745.
40. Waldmann H, Qin S, Cobbold S. Monoclonal
antibodies as agents to reinduce tolerance in
autoimmunity. J Autoimmun1992;5: 93–102.
41. Waldmann H, Cobbold S. How do
monoclonal antibodies induce tolerance?
A role for infectious tolerance? Annu Rev
Immunol 1998;16:619–644.
42. Velculescu VE, Zhang L, Vogelstein B,
Kinzler KW. Serial analysis of gene
expression. Science 1995;270:484–487.
43. Zelenika D, et al. Regulatory T cells
overexpress a subset of Th2 gene transcripts.
J Immunol 2002;168:1069–1079.
44. Zelenika D, Adams E, Humm S, Lin CY,
Waldmann H, Cobbold SP. The role of CD4þT-cell subsets in determining transplantation
rejection or tolerance. Immunol Rev
2001;182:164–179.
45. Adorini L, PennaG,GiarratanaN,UskokovicM.
Tolerogenic dendritic cells induced by
vitamin D receptor ligands enhance regulatory
T cells inhibiting allograft rejection and
autoimmune diseases. J Cell Biochem
2003;88:227–233.
46. Madden SL, Galella EA, Zhu J, Bertelsen AH,
Beaudry GA. SAGE transcript profiles for p53-
dependent growth regulation. Oncogene
1997;15:1079–1085.
Cobbold et al �Regulatory T cells and APCs in tolerance
Immunological Reviews 196/2003 123

47. Hashimoto SI, et al. Gene expression profile
in human leukocytes. Blood 2003;101:
3509–3513.
48. Chen J, Sun M, Lee S, Zhou G, Rowley JD,
Wang SM. Identifying novel transcripts and
novel genes in the human genome by using
novel SAGE tags. Proc Natl Acad Sci USA
2002;99:12257–12262.
49. Velculescu VE, et al. Characterization of the
yeast transcriptome. Cell 1997;88:243–251.
50. Caron H, et al. The human transcriptome
map: clustering of highly expressed genes in
chromosomal domains. Science
2001;291:1289–1292.
51. Lash AE, et al. SAGEmap: a public gene
expression resource. Genome Res
2000;10:1051–1060.
52. Saha S, et al. Using the transcriptome to
annotate the genome. Nat Biotechnol
2002;20:508–512.
53. Anisimov SV, Tarasov KV, Tweedie D,
Stern MD, Wobus AM, Boheler KR. SAGE
identification of gene transcripts with profiles
unique to pluripotent mouse R1 embryonic
stem cells. Genomics 2002;79:169–176.
54. Shires J, Theodoridis E, Hayday AC.
Biological insights into TCRgammadeltaþ and
TCRalphabetaþ intraepithelial lymphocytes
provided by serial analysis of gene expression
(SAGE). Immunity 2001;15:419–434.
55. Shimazaki T, Okazawa H, Fujii H, Ikeda M,
Tamai K, McKay RD, et al. Hybrid cell
extinction and re-expression of Oct-3
function correlates with differentiation
potential. EMBO J 1993;12:4489–4498.
56. Shimizu J, Yamazaki S, Takahashi T, Ishida Y,
Sakaguchi S. Stimulation of CD25(þ)CD4(þ)
regulatory T cells through GITR breaks
immunological self-tolerance. Nat Immunol
2002;3:135–142.
57. Wilcox RA, Tamada K, Strome SE, Chen L.
Signaling through NK cell-associated CD137
promotes both helper function for CD8þ
cytolytic T cells and responsiveness to IL-2
but not cytolytic activity. J Immunol
2002;169:4230–4236.
58. Thornton AM, Shevach EM. CD4þCD25þ
immunoregulatory T cells suppress
polyclonal T cell activation in vitro by
inhibiting interleukin 2 production. J ExpMed
1998;188:287–296.
59. Schaffer A, Cerutti A, Shah S, ZanH, Casali P. The
evolutionarily conserved sequence upstream of
the human Ig heavy chain S gamma 3 region is
an inducible promoter: synergistic activation by
CD40 ligand and IL-4 via cooperative NF-kappa
B and STAT-6 binding sites. J Immunol
1999;162: 5327–5336.
60. Singh B, et al. Control of intestinal
inflammation by regulatory T cells. Immunol
Rev 2001;182:190–200.
61. Sakaguchi S, et al. Immunologic tolerance
maintained by CD25þCD4þ regulatory T
cells: their common role in controlling
autoimmunity, tumor immunity, and
transplantation tolerance. Immunol Rev
2001;182:18–32.
62. McHugh RS, et al. CD4(þ)CD25(þ)
immunoregulatory T cells: gene expression
analysis reveals a functional role for the
glucocorticoid-induced TNF receptor.
Immunity 2002;16:311–323.
63. Read S, Malmstrom V, Powrie F. Cytotoxic
T lymphocyte-associated antigen 4 plays
an essential role in the function of
CD25(þ)CD4(þ) regulatory cells that
control intestinal inflammation. J Exp Med
2000;192:295–302.
64. Hori S, Nomura T, Sakaguchi S. Control
of regulatory T cell development by the
transcription factor foxp3. Science
2003;299:1057–1061.
65. Lehmann J, et al. Expression of the integrin
alpha Ebeta 7 identifies unique subsets of
CD25þ as well as CD25– regulatory T cells.
Proc Natl Acad Sci USA 2002;99:
13031–13036.
66. Zheng W, Flavell RA. The transcription factor
GATA-3 is necessary and sufficient for Th2
cytokine gene expression in CD4 T cells. Cell
1997;89:587–596.
67. Nawijn MC, et al. Enforced expression of
GATA-3 in transgenic mice inhibits Th1
differentiation and induces the formation of
a T1/ST2-expressing Th2-committed T cell
compartment in vivo. J Immunol
2001;167:724–732.
68. Miaw SC, Choi A, Yu E, Kishikawa H, Ho IC.
ROG, repressor of GATA, regulates the
expression of cytokine genes. Immunity
2000;12:323–333.
69. Moratz C, et al. Regulator of G protein
signaling 1 (RGS1) markedly impairs Gi alpha
signaling responses of B lymphocytes.
J Immunol 2000;164:1829–1838.
70. Reif K, Cyster JG. RGS molecule expression in
murine B lymphocytes and ability to down-
regulate chemotaxis to lymphoid chemokines.
J Immunol 2000;164: 4720–4729.
71. Monteyne P, Renauld JC, Van Broeck J,
Dunne DW, Brombacher F, Coutelier JP.
IL-4-independent regulation of in vivo IL-9
expression. J Immunol 1997;159:2616–2623.
72. Brown JA, et al. Blockade of programmed
death-1 ligands on dendritic cells enhances
T cell activation and cytokine production.
J Immunol 2003;170:1257–1266.
73. Langrish CL, Buddle JC, Thrasher AJ, Goldblatt
D. Neonatal dendritic cells are intrinsically
biased against Th-1 immune responses. Clin
Exp Immunol 2002;128:118–123.
74. Goerdt S, Orfanos CE. Other functions, other
genes: alternative activation of antigen-
presenting cells. Immunity 1999;10: 137–142.
75. Moore KW, de Waal Malefyt R, Coffman RL,
O’Garra A. Interleukin-10 and the
interleukin-10 receptor. Annu Rev Immunol
2001;19:683–765.
76. Ehlert JE, Ludwig A, Grimm TA, Lindner B,
Flad HD, Brandt E. Down-regulation of
neutrophil functions by the ELR(þ) CXC
chemokine platelet basic protein. Blood
2000;96:2965–2972.
77. Soares MP, Brouard S, Smith RN, Bach FH.
Heme oxygenase-1, a protective gene that
prevents the rejection of transplanted organs.
Immunol Rev 2001;184:275–285.
78. Maloy KJ, Salaun L, Cahill R, Dougan G,
Saunders NJ, Powrie F. CD4þCD25þ T(R)
cells suppress innate immune pathology
through cytokine-dependent mechanisms.
J Exp Med 2003;197:111–119.
79. Chen H, Centola M, Altschul SF, Metzger H.
Characterization of gene expression in resting
and activated mast cells. J Exp Med
1998;188:1657–1668.
80. Powell J. Enhanced concatemer cloning-
a modification to the SAGE (Serial Analysis
of Gene Expression) technique. Nucleic
Acids Res 1998;26:3445–3446.
81. Barrat FJ, et al. In vitro generation of
interleukin 10-producing regulatory CD4(þ)
T cells is induced by immunosuppressive
drugs and inhibited by T helper type 1 (Th1)-
and Th2-inducing cytokines. J Exp Med
2002;195:603–616.
82. Inaba K, et al. Generation of large numbers of
dendritic cells from mouse bone marrow
cultures supplemented with granulocyte/
macrophage colony-stimulating factor. J Exp
Med 1992;176:1693–1702.
83. Fairchild PJ, et al. Directed differentiation of
dendritic cells from mouse embryonic stem
cells. Curr Biol 2000;10:1515–1518.
84. Fairchild PJ, Nolan KF, Waldmann H. Probing
dendritic cell function by guiding the
differentiation of embryonic stem cells.
Methods Enzymol 2003;365:169–186.
Cobbold et al �Regulatory T cells and APCs in tolerance
124 Immunological Reviews 196/2003




















